 ,2,*
,2,*Advances of LORELEI-like Glycosylphosphatidylinositol-anchor (LLG) Proteins in Plants
Sijia Li1, Yongxue Zhang1, Mingsheng Jia2, Ying Li1, Shaojun Dai,2,*通讯作者: E-mail:daishaojun@hotmail.com
责任编辑: 白羽红
收稿日期:2020-04-5接受日期:2020-05-20网络出版日期:2020-09-01
| 基金资助: |
Corresponding authors: E-mail:daishaojun@hotmail.com
Received:2020-04-5Accepted:2020-05-20Online:2020-09-01

摘要
类LORELEI糖基磷脂酰肌醇锚定蛋白(LLG)定位于细胞质膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与其转运和胞外信号转导, 从而调控植物生殖发育以及免疫与逆境应答等过程。LLG2/3与ANX和BUPS互作, 调控花粉管顶端生长与爆裂。LLG1与FER (FERONIA)互作, 调控下游的NADPH氧化酶产生活性氧(ROS), 促进根部细胞伸长和根毛生长。此外, LLG1作为FER的共受体, 与快速碱化因子(RALFs)互作, 调节G蛋白β亚基(AGB1)和质膜H +-ATPase功能、胞内ROS稳态以及Ca 2+瞬变, 引起根部和气孔的盐应答反应。LLG1与FLS2和EFR互作激活下游RbohD, 调节ROS产生, 调控植物免疫应答。该文综述了植物LLG的相关研究进展, 可为深入理解LLG的生物学功能提供重要信息。
关键词:
Abstract
The outer surface of plasma membrane (PM)-localized LORELEI-like glycosylphosphatidylinositol-anchor (LLG) proteins, as the molecular chaperone of CrRLK1Ls family of receptor-like kinase, are involved in the transport of CrRLKs and extracellular signal transduction, regulating plant reproduction, development, as well as immune and stress responses. LLG2/3 interacting with ANX and BUPS regulates pollen tube growth and rupture. LLG1 interacted with FER activates the ROPGEF1-ROP2-NADPH oxidase pathway for ROS production, and then promotes root cell elongation and root hair growth. Besides, LLG1, as co-receptor of FER, interacts with RALFs, and then regulates G protein β (AGB1), PM H +-ATPase activity, as well as the homeostasis of intracellular ROS and Ca 2+, for modulating stomata and roots in response to salinity. For immune response, LLG1 interacts with FLS2 and EFR, activating the downstream RbohD for ROS production. This review provides important information for understanding LLG biological functions.
Keywords:
PDF (2373KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
李思佳, 张咏雪, 贾明生, 李莹, 戴绍军. 植物类LORELEI糖基磷脂酰肌醇锚定蛋白研究进展. 植物学报, 2020, 55(5): 541-550 doi:10.11983/CBB20058
Li Sijia, Zhang Yongxue, Jia Mingsheng, Li Ying, Dai Shaojun.

植物糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol-anchored protein, GPI-AP)定位于细胞膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与调节细胞生殖、生长以及免疫与盐碱胁迫应答等多种生物学过程(Yu et al., 2013; Cheung et al., 2014; Zhang et al., 2020)。植物GPI-AP家族包括COBRA、ENODL和LRE (LORELEI)三个亚家族, 分别调节细胞壁纤维素生物合成(Liu et al., 2013)、花粉管接受(Hou et al., 2016)和双受精作用(Feng et al., 2019)。拟南芥(Arabidopsis thaliana)中有3个类LORELEI糖基磷脂酰肌醇锚定蛋白(LORELEI-like GPI-AP, LLG), 其中LLG1在根和叶片等多个器官中表达, LLG2和LLG3在花粉粒和花粉管中表达(Ge et al., 2019)。LLG参与FER (FERONIA)等多种类受体激酶(receptor-like kinase, RLK)从内质网向质膜的转运及其质膜定位, 并作为这些RLK的共受体感知外部信号和快速碱化因子(rapid alkalinization factor, RALF)等配体(Li et al., 2015), 调节RLK的功能, 参与花粉管生长与爆裂、根与根毛生长以及免疫与盐胁迫应答等过程(Li et al., 2015; Shen et al., 2017; Feng et al., 2018; Yu et al., 2018; Zhao et al., 2018; Ge et al., 2019)。
1 LLG参与RLKs的转运与定位
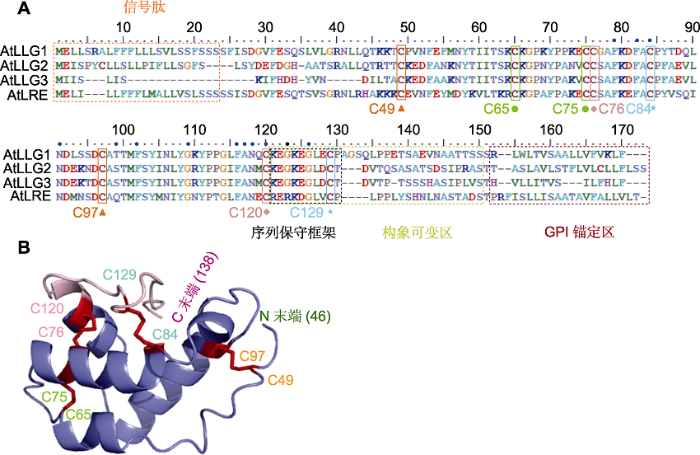
LLG的氨基酸序列包括N端信号肽(含23个高度保守的氨基酸残基)、中央区域、C端构象可变区以及C端GPI锚(约20个氨基酸残基) (图1A)。LLG有8个保守的半胱氨酸, 形成4对二硫键, 参与蛋白质3D结构形成(图1B) (José-Estanyol et al., 2004; Liu et al., 2016; Shen et al., 2017)。LLG形成过程中, GPI-AP前体在内质网中添加预组装的GPI锚, C末端被脂质化修饰, 然后通过高尔基体转运, 分泌到细胞膜外小叶, 主要分布于富含鞘脂和胆固醇的膜微区(Zurzolo and Simons, 2016)。LLG与FER等RLKs跨膜结构域N末端的胞外近膜区(juxtamenbrane region, exJM)结合, 并将这些RLKs从内质网转运到质膜上(Li et al., 2015)。LLG作为RLKs的共同受体, 感知细胞外部信号或配体, 调节下游各种信号通路。在拟南芥llg1突变体中, FER-GFP滞留在内质网和细胞质, 而在llg1突变体中回补表达LLG1可以减少FER-GFP在细胞质中的滞留, 恢复FER的质膜定位。llg1突变体早期生长发育表型与fer的表型相似, 如对RALF1的敏感性降低、表皮细胞形状改变以及根毛生长缺陷(Li et al., 2015)。LLG1/2/3与其配体RALFs保守的N端(4-17位的14个氨基酸)结合, 并与FER胞外结构域(FERECD)互作, 形成LLG-RALF-FER复合物, 调控下游信号通路(Xiao et al., 2019)。图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1拟南芥LLG氨基酸序列与3D结构模型
(A) LLG氨基酸序列(彩色框标注的分别是N端信号肽、序列保守框架、C端构象可变区及C端GPI锚定区; 三角形、绿色圆点、菱形和五角星表示可形成4对二硫键的8个Cys位点, 蓝色圆点表示保守氨基酸位点); (B) LLG1的46-138位氨基酸的3D结构(α螺旋、β折叠、N末端(46)、C末端(138)及8个保守Cys位点形成4对二硫键)
Figure 1Arabidopsis LLG amino acid sequences and 3D structure model
(A) LLG amino acid sequence (The colored boxes indicate the N-terminal signal peptide, sequence conservative frame, C-terminal conformational flexible region, and C-terminal GPI anchor region, respectively; the triangles, green dots, diamonds and pentagonal stars represent 8 Cys sites that can form 4 pairs of disulfide bonds, and the blue dots represent conservative amino acid sites); (B) The 3D structure of amino acids 46-138 of LLG1 (α-helix, β-sheet, N terminal (46), C terminal (138), and conserved 8 Cys sites formed 4 pairs of disulfide bonds)
2 LLG2/3调节花粉管顶端生长与爆裂
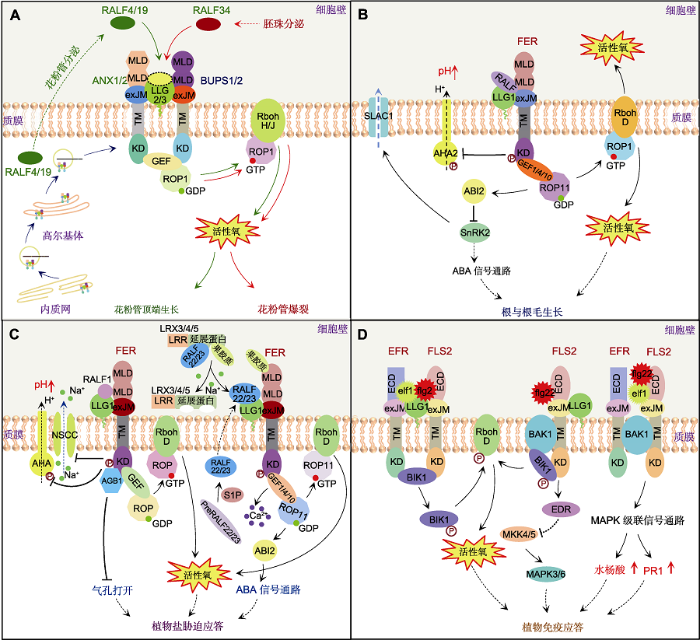
植物花粉管快速生长进入胚珠的过程受到精细调控(Johnson et al., 2019)。定位在花粉管顶端的多种RLKs感知外部信号, 调控花粉管生长(Li and Yang, 2016; Zhong and Qu, 2019)。ANX1/2 (ANXUR 1/2)和BUPS1/2 (Buddha’s paper seal 1/2)是花粉管顶端质膜定位的RLKs, 两者形成受体激酶复合物, 通过响应RALF小肽信号调节花粉管生长和精细胞释放(Ge et al., 2017, 2019; Franck et al., 2018; Li and Yang, 2018)。花粉特异表达的LLG2/3可以与ANX和BUPS的exJM区互作, 调节其质膜定位, 缺失exJM区的ANX/BUPS滞留在胞内(Ge et al., 2019)。LLG2/3作为分子伴侣, 协助ANX和BUPS从内质网分泌到花粉管顶端的质膜, 形成受体-共受体复合物, 共同感受胞外RALF信号(图2A); 通过感受花粉管自分泌的RALF4和RALF19小肽, 维持花粉管的完整性和顶端生长(Mecchia et al., 2017; Feng et al., 2019); 通过识别胚珠分泌的RALF34小肽, 控制花粉管爆裂并释放精细胞(Ge et al., 2017)。RALF4和RALF19显著增强LLG2/3与ANX/BUPS的相互作用, RALF4的C端与LLG结合, 其N端(包括YISY motif)与LLG微弱互作, 而与ANX/BUPS强烈互作(Ge et al., 2019)。Xiao等(2019)解析了FERECD、ANX1、ANX2、LLG1以及RALF23-LLG2-FERECD复合体的晶体结构(Xiao et al., 2019)。结果表明, FERECD与ANX1ECD和ANX2ECD的晶体结构非常相似, RALF23的N端α螺旋与LLG2表面大沟结合, 且FERECD与RALF23和LLG2互作后, 其构象无明显变化(Xiao et al., 2019)。以上研究结果为认识RALF4/19/34与LLG2/3和ANX/ BUPS之间的互作关系提供了参考。图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2LLG与RLK家族蛋白不同成员互作调控花粉与根发育以及盐胁迫与免疫应答过程
(A) LLG2/3与ANX1/2和BUPS1/2互作调控花粉管生长与爆裂; (B) LLG1与FER互作调控根与根毛生长; (C) LLG1与FER互作调控盐胁迫应答; (D) LLG1与FLS2和EFR互作调控免疫应答。ABA: 脱落酸; ABI2: A型 PP2Cs磷酸酶; AGB1: 异源三聚体G蛋白β亚基; AHA2: 质膜H+-ATPase 2; ANX1/2: ANXUR1/2; BAK1: 油菜素受体激酶; BIK1: Botrytis诱导激酶1; BUPS1/2: 佛祖之金字亚贴1/2; ECD: 胞外结构域; EDR1: 负调控抗病蛋白1; EFR: 延伸因子Tu受体; elf18: 细菌延伸因子Tu N端18个氨基酸小肽; exJM: 胞外近膜结构域; FER: FERONIA; flg22: 细菌鞭毛蛋白N端22个氨基酸小肽保守基序; FLS2: 鞭毛蛋白传感蛋白2; GDP: 鸟苷二磷酸; GEF1/4/10: 鸟嘌呤核苷酸交换因子1/4/10; GTP: 鸟苷三磷酸; KD: 激酶结构域; LLG1/2/3: 类LORELEI糖基磷脂酰肌醇锚定蛋白1/2/3; LRR: 富含亮氨酸重复序列; LRX: 富含亮氨酸重复序列的延展蛋白; MAPK: 丝裂原活化蛋白激酶; MKK: 丝裂原激活的蛋白激酶激酶; MLD: 类Malectin结构域; NSCC: 非选择性阳离子通道; PR1: 病程相关因子1; RALF: 快速碱化因子; Rboh: 呼吸爆发氧化酶; ROP1/11: 植物Rho相关小G蛋白1/11; S1P: 位点-1蛋白酶; SLAC1: 慢阴离子通道蛋白1; SnRK2: 丝氨酸/苏氨酸蛋白激酶SnRK2D; TM: 跨膜结构域。实线表示直接调控过程, 虚线表示间接调控过程或物质转运。箭头表示促进, T表示抑制。
Figure 2Interaction between different members of the LLG and RLK family proteins regulates pollen and root development and salt and immune response processes
(A) LLG2/3 interacts with ANX1/2 and BUPS1/2 to regulate pollen tube growth and burst; (B) LLG1 interacts with FER to regulate root and root hair growth; (C) LLG1 interacts with FER to regulate salt stress response; (D) LLG1 interacts with FLS2 and EFR to regulate the immune response. ABA: Abscisic acid; ABI2: ABA insensitive 2; AGB1: Heterotrimeric G-protein β-subunit; AHA2: Plasma membrane H+-ATPase 2; ANX1/2: ANXUR1/2; BAK1: Brassinosteroid insensitive 1-associated receptor kinase 1; BIK1: Botrytis-induced kinase 1; BUPS1/2: Buddha’s paper seal1/2; ECD: Extracellular domain; EDR1: Enhanced disease resistance 1; EFR: EF-Tu receptor; elf18: 18 amino acid peptide of EF-Tu N-terminus; eXJM: Extracellular membrane domain; FER: FERONIA; flg22: 22 amino acid peptide of bacterial flagellin N-terminus; FLS2: Flagellin sensing 2; GDP: Guanosine diphosphate; GEF1/4/10: Guanine nucleotide exchange factor1/4/10; GTP: Guanosine triphosphate; KD: Kinase domain; LLG1/2/3: LORELEI-like GPI-anchored protein1/2/3; LRR: Leucine-rich repeat; LRX: Leucine-rich repeat extensin-like protein; MAPK: Mitogen-activated protein kinase; MKK: Mitogen-activated protein kinase kinase; MLD: Malectin-like domain; NSCC: Non-selective cation channels; PR1: Pathogenesis-related factor 1; RALF: Rapid alkalinization factor; Rboh: Respiratory burst oxidase homolog; ROP1/11: Rho-related GTPase1/11 from plants; S1P: Site-1 protease; SLAC1: Slow anion channel 1; SnRK2: Serine/threonine-protein kinase SnRK2D; TM: Transmembrane domain. The solid line represents the direct regulation process, the dotted line represents the indirect regulation process or material transport. The arrow represents promotion, and the T represents inhibition.
在感受胞外RALF4/19后, LLG2/3-ANX1/2-BUPS 1/2复合体通过与GDP-ROP1相互作用, 将其激活为GTP-ROP1, 进而激活下游花粉特异表达的NADPH氧化酶RbohH/J, 产生活性氧(ROS), 导致ROS在花粉管顶端积累, 调节花粉管生长, 降低花粉管的爆裂率(图2A) (Kaya et al., 2014; Mangano et al., 2016; Feng et al., 2019)。拟南芥LLG2/3 RNAi植株的花粉管ROS含量降低, 生长迟滞, 爆裂率较高。LLG2/3的缺乏引起花粉管细胞壁成分改变, 甲酯化果胶质在花粉管顶端区域积累, 去甲酯化果胶质在花粉管亚顶端和shank区域积累, 花粉管中的胼胝质含量降低, 引起花粉管生长变缓(Feng et al., 2019)。外源施加H2O2能够以剂量依赖的方式恢复LLG2 RNAi植株花粉管的长度, 降低花粉管爆裂率, 这与anx1?/?/anx2+/? 突变体花粉管表型能被外源施加H2O2恢复相似(Duan et al., 2010; Feng et al., 2019)。由此可见, ROS稳态对于RALF-LLG-ANX/BUPS调节花粉管生长与爆裂十分重要。
3 LLG1调控根与根毛生长
LLG1通过与FER的exJM结合, 协助FER定位于根部细胞质膜, 进而共同调控根与根毛生长(图2B) (Duan et al., 2010; Li et al., 2015)。拟南芥llg1突变体与fer-4突变体有相似的根毛缺陷表型(Li et al., 2015)。fer突变体的营养生长(Keinath et al., 2010)、根毛生长(Duan et al., 2010; Huang et al., 2013)以及下胚轴伸长均受到抑制(Guo et al., 2009; Deslauriers and Larsen, 2010), 根毛出现卷曲或异常分枝(Duan et al., 2010; Li et al., 2015)。在llg1与fer突变体中分别转入LLG1和FER, 可以恢复其野生型表型(Duan et al., 2010; Li et al., 2015)。LLG1与FER相互作用共同感受RALF1, 形成LLG1-RALF1-FER复合物并激活FER, 进而调控根生长。外源施加RALF1抑制野生型幼苗根的生长, 但并不影响llg1与fer-4突变体的根长。激活的FER与RopGEFs互作, 将RAC/ROP从与GDP结合的无活性状态转变为与GTP结合的活化状态。fer突变体与几种rac/rop突变体表型相似, 表明fer与rac/rop之间有调控关系(图2B) (Li et al., 2015)。活化的RAC/ROP调节NADPH氧化酶产生ROS, 进而调控根生长(Duan et al., 2010, 2014; Li et al., 2015)。llg1与fer-4突变体根中的ROS水平显著降低(Swanson and Gilroy, 2010), 导致细胞壁丧失完整性、细胞质外渗及细胞塌陷(Li et al., 2015)。同时, 激活的FERKD引起AHA2磷酸化并失活, 导致质外体pH值升高, 引起细胞壁硬化, 抑制根部细胞伸长和根毛生长(图2B) (Haruta et al., 2014; Li et al., 2015; Xiao et al., 2019)。反之, 低pH值会导致细胞因膨胀紊乱而爆裂(Monshausen et al., 2007)。由此表明, LLG1和FER复合体通过与胞外小肽RALF1互作, 动态调节质外体ROS与pH稳态, 从而调控根与根毛生长。
LLG1可能与FER共同参与脱落酸(ABA)、生长素、乙烯及油菜素内酯(BR)等激素信号转导过程, 从而调控根与根毛生长(Guo et al., 2009; Duan et al., 2010; Deslauriers and Larsen, 2010; Huang et al., 2013)。llg1与fer-4突变体对生长素和ABA有相似的响应。fer突变体根毛伸长对生长素不敏感(Duan et al., 2010; Li et al., 2015), 根伸长对ABA的抑制敏感(Yu et al., 2012; Li et al., 2015), 下胚轴伸长对乙烯和BR敏感(Deslauriers and Larsen, 2010)。当ABA存在时, FER通过与GEF1/4/10和GTP-ROP11互作, 激活A-type PP2C磷酸酶ABI2 (ABA insensitive 2)的活性, 抑制SnRK2活性, 从而抑制ABA信号通路, 同时抑制SLAC1通道和NADPH氧化酶介导的ROS产生, 负调控根生长(Yu et al., 2012)。
4 LLG1调控盐胁迫应答
盐离子干扰植物根部细胞壁结构, 导致细胞壁完整性降低, 抑制细胞极性生长, 引起细胞爆裂(Dinneny et al., 2008; Feng et al., 2018)。盐胁迫引起的细胞壁果胶质交联变化可以被FER胞外2个串联的MLD (malectin-like domain) A和B感知, 引起胞内激酶活性变化, 激发胞质Ca2+浓度瞬变, 启动盐应答信号通路(图2C) (Feng et al., 2018)。LLG1作为FER的共受体参与盐应答过程。拟南芥fer2和fer4突变体根的生长存在缺陷, 并且对Na+敏感, 但对甘露醇和山梨醇引起的渗透胁迫不敏感(Feng et al., 2018)。与之相似, llg1突变体的根也对Na+敏感。用浓度高于100 mmol·L-1 NaCl处理可引起llg1和fer4突变体根部伸长区细胞丧失活力; 而对于sos1和sos2突变体而言, 50 mmol·L-1 NaCl即可导致其根部从伸长区至根尖大范围的细胞死亡。这表明LLG1和FER可能参与调控与SOS通路不同的盐胁迫应答途径(Feng et al., 2018)。
LLG1通过与RALFs和FER互作形成三元复合物, 调控下游信号通路。三者的关键氨基酸位点可能影响其互作关系(Xiao et al., 2019)。LLG1通过其C端构象变化调控其与RALFs保守的N端结合, 进而与FERECD互作(Xiao et al., 2019)。LLG1/2/3和LRE中12个保守氨基酸在其与RALF23互作中发挥关键作用(图1A)。与RALF23结合时, LLG1的129-138位氨基酸区域的构象发生变化。同时, LLG1/2/3的121KEGKEGLD128区域非常保守, 其中G123R突变会减弱LLG1与RALF23互作(Xiao et al., 2019)。llg1-3突变体发生的G114R替换也降低了LLG1与RALF23的体外互作, 减弱了RALF23对幼苗生长的抑制作用(Xiao et al., 2019)。但是, LLG1有些位点的突变(如N91A、T99R、A117Y和N118Y)并未影响其与RALF23和FER的互作(Xiao et al., 2019)。此外, RALF23关键位点突变(I6A、I6Y、L11Y和N14A)会影响其与LLG1的互作, 进而消除其对elf18诱导的ROS产生与幼苗生长的抑制作用(Xiao et al., 2019)。FER氨基酸位点突变也会影响RALF23对其的调节作用。与野生型相比, 在fer-4背景下回补表达点突变的FER (G257A)和FER (N303Y), 幼苗生长的抑制明显受到RALF23诱导(Xiao et al., 2019)。
在盐胁迫应答过程中, RALF22/23诱导FER通过胞吞途径内化, 从而负调控FER的功能(图2C) (Zhao et al., 2018)。正常状态下, 细胞壁富含亮氨酸重复序列延展蛋白(leucine-rich repeat extensin, LRX) 3/4/5 N端的富含亮氨酸重复序列(leucine-rich repeat, LRR)与配体RALF22/23结合, C端高度糖基化的延展蛋白与细胞壁成分(果胶质)交联; 而在盐胁迫条件下, 盐诱导的细胞壁交联变化被LRX3/4/5感知, 释放出与其结合的RALF22/23。同时, 盐胁迫也诱导依赖于SITE-1肽酶(site-1 protease, S1P)催化的成熟RALF22的积累。盐诱导的RALF22和RALF23的增加, 促进了其与FER的互作, 导致FER内化(图2C)。拟南芥llg1突变体、lrx3/4/5三突变体、fer4突变体以及RALF22和RALF23过表达植株都表现出相似的生长迟滞和盐敏感表型(Zhao et al., 2018)。也有研究表明, FERECD的malectin基序可以与细胞壁中的果胶质相互作用, 感知盐引起的细胞壁软化, 触发FER介导的Ca2+瞬变, 调节细胞壁完整性, 以防盐胁迫下根细胞在生长过程中发生爆裂(Feng et al., 2018)。
在叶片中, FER与G蛋白β亚基(heterotrimeric G-protein β, AGB1)相互作用, 抑制气孔开放(图2C) (Yu et al., 2018)。agb1和fer均表现出盐敏感表型, 而agb1的表型相对温和(Yu and Assmann, 2015; Yu et al., 2018)。有研究表明, AGB1通路和FER通路协同作用, 参与盐胁迫应答, 且这种作用可能受到蒸腾条件的影响(Yu et al., 2018)。在蒸腾条件下, agb1突变体气孔导度较大, 而fer突变体大量气孔关闭, 这导致两者蒸腾拉力有较大差异; 两者Na+从根向地上部转运的效率不同, agb1表现为全株盐敏感, 且Na+在地上部大量积累, 但fer突变体无显著的Na+积累(Yu et al., 2018)。在非蒸腾条件下, 植株细胞内Na+含量主要取决于Na+通过非选择性阳离子通道(non-selective cation channels, NSCC)内流和Na+/H+反转运子外排。fer突变体的AHA活性较高, 导致质外体酸化驱动的Na+/H+反转运子活跃, 促进Na+外排, 相对增强耐盐性(Yang et al., 2010)。然而, fer的盐敏感表型可能是由于fer不能持续激活AHA (Yu et al., 2018)。fer突变体中大量Na+积累可能主要由于根部表皮和皮层细胞完整性丧失所致(Feng et al., 2018)。
RALF1通过诱导FER磷酸化将其激活(Haruta et al., 2014; Chen et al., 2016), 进而通过磷酸化AHA的Ser899, 抑制其质子转运能力(Haruta et al., 2014), 降低质外体酸化和Na+外排, 阻止细胞延长和根生长, 且对盐敏感(图2C) (Yu et al., 2018)。此外, RALF1处理会引起根部FER依赖的胞质Ca2+增加(Haruta et al., 2008, 2014)。外源施加1 μmol·L-1 RALF1可以抑制拟南芥野生型和agb1突变体根的生长, 但不影响fer2、fer4和agb1-2/fer2的表型, 这表明RALF1通过FER对根生长发挥抑制作用(Yu et al., 2018)。
盐诱导质膜定位的NADPH氧化酶RbohD产生ROS, 并受到AGB1和FER的协同调控(图2C) (Yu et al., 2018)。盐胁迫影响agb1和fer的ROS稳态, agb1/ fer双突变体表现出比各自单突变体更严重的ROS失衡表型, 且这种ROS失衡在不同浓度和时间的盐处理以及不同器官(叶片和根)中存在差异(Yu et al., 2018)。
盐胁迫引起ABA含量升高进而激活FER, 调控下游通路; 同时, FER也通过GEF1/4/10-ROP11途径增强ABI2的活性, ABI2直接与FER互作将其去磷酸化而失活, 从而抑制ABA响应。虽然fer突变体对ABA和盐胁迫非常敏感, 但是FER调控并非全部依赖ABA, 其它信号通路也可能参与其中, 具体机制尚不清楚(Chen et al., 2016)。
对盐生牧草小花碱茅(Puccinellia tenuiflora)盐碱应答转录组的分析表明, FER及其相关基因HERK1、NOTIA和MARIS, 以及FLS2及其互作蛋白BAK1、EFR、BIK1、SCD1、BSK1和GRP7协同作用, 调节G蛋白介导的Ca2+信号通路、ABA信号通路以及MAPK级联信号通路和ROS稳态, 从而调控盐碱应答过程(Zhang et al., 2020)。
5 LLG1调控免疫应答
在植物应答病原菌侵染过程中, 质膜模式识别受体(pattern recognition receptors, PRRs)参与激活免疫系统。多数植物PRRs是具有胞外配体识别结构域、跨膜结构域和胞质激酶结构域的类受体激酶, 如富含亮氨酸重复序列受体激酶FLS2 (flagellin sensing 2)和EFR (EF-Tu receptor)。LLG1作为分子伴侣, 与EFR和FLS2的exJM区互作调节其质膜定位, 进而调控植物免疫应答过程(Shen et al., 2017)。当植物受到病原菌侵害时, FLS2和EFR迅速与其共受体BAK1 (brassinosteroid insensitive 1-associated receptor kinase 1)形成复合体, 激活下游MAPK级联信号通路, 调控防御基因的表达, 促进水杨酸(SA)等防御激素的积累, 提高植株的抗病性(Sun et al., 2013)。MAPK级联信号通路中的EDR1 (enhanced disease resistance 1)是一种Raf-like MAP3K, 通过调节MKK4/5-MPK3/6通路负调控植物免疫, 且拟南芥edr1突变体具有抗病性(Frye et al., 2001)。Shen等(2017)在edr1背景下将LLG1的Gly114突变为Arg114获得双突变体edr1/llg1-3, 并将T-DNA插入获得的llg1-2突变体与edr1杂交获得双突变体edr1/llg1-2。研究表明, edr1/llg1-3和edr1/llg1-2都对白粉病菌(Golovinomyces cichoracearum UCSC1)敏感。llg1-3通过抑制edr1突变体中免疫标记基因PR1的表达和SA的积累削弱edr1对白粉病的抗性。edr1/llg1-3对活体营养型卵菌(Hyaloperonospora arabidopsidis Noco2)和植物病原细菌(Pseudomonas syringae pv tomato (Pto) DC3000)等多种病原体敏感。llg1-2和llg1-3突变体对上述3种病原体都敏感, 且突变体体内SA和PR1表达水平都受到G. cichoracearum侵染的抑制。在llg1-2和llg1-3突变体中转入LLG1可以恢复其野生型表型, 表明LLG1参与对多种病原体的免疫应答过程(图2D) (Shen et al., 2017)。
LLG1不同位点突变对其与FER、FLS2和EFR互作的调控存在差异。llg1-2具有明显的生长缺陷表型(Li et al., 2015), 而llg1-3 (LLG1G114R)中LLG1与FER的互作及质膜定位不受影响, 无生长发育缺陷表型, 这表明LLG1G114R点突变仅失去免疫功能, 对植物生长发育无影响(Shen et al., 2017)。酵母双杂交和CO-IP实验证明LLG1与EDR1不能互作, 这暗示LLG1对免疫的调节可能与EDR1信号通路无关。LLG1与FLS2和EFR的互作不受flg22处理及LLG1G114R点突变的影响, 并且在flg22诱导下可以与BAK1互作(图2D) (Shen et al., 2017)。与野生型相比, FLS2的积累、质膜定位以及flg22诱导的降解在llg1-2中均降低, 但在llg1-3中无变化。这表明LLG1具有的分子伴侣与信号转导功能可能是相互独立的, LLG1G114R点突变并未影响其与FLS2互作, 但可能影响LLG1和FLS2的构象, 从而导致FLS2调控的下游信号通路改变(图2D) (Shen et al., 2017)。LLG1与FLS2和EFR形成的复合体调控胞内BIK1 (botrytis-induced kinase 1)磷酸化, 激活下游的RbohD产生ROS, 参与免疫应答反应(图2D) (Shi et al., 2013; Li et al., 2014)。llg突变体中flg22诱导的FLS2和BAK1互作未受影响, 但llg1-2和llg1-3原生质体中flg22诱导的BIK1磷酸化水平降低, llg1-2和llg1-3突变体中受flg22和elf18诱导的ROS水平低于野生型(Shen et al., 2017)。
6 结论与展望
LLG作为质膜定位的RLKs (如FER和FLS2)的分子伴侣, 负责将其从内质网运送到质膜, 继而在RALFs等配体和胞外信号的调节下, 动态调控RLKs的质膜定位与活性。拟南芥llg1和fer突变体的多样化发育与逆境应答表型, 暗示LLG与FER等RLKs之间的互作可能不是简单的开关, 而是存在精细的调节机制。对拟南芥LLG构象的解析及其与RALFs和FER互作位点的研究表明, LLG可能通过构象变化调节三者的互作关系, 从而精细调控胞外信号感知, 招募不同的RLKs完成胞内信号转导(Xiao et al., 2019)。然而, 在特定发育(如叶表皮细胞形状、花粉管生长与爆裂、根部伸长区与根毛细胞生长)与逆境(如病原体、盐碱、温度、低氮、低磷以及碳水化合物)应答过程中, LLG如何感受质外体信号(如ROS水平及不同配体)调整自身构象, 如何调节其与不同配体(如RALFs)结合, 如何招募不同的RLKs并激活其下游信号通路(Huang et al., 2013; Yang et al., 2015; Yeats et al., 2016; Yin et al., 2018; Xu et al., 2019), 以及LLG的4对保守的半胱氨酸位点在ROS信号感知和蛋白质构象调节方面是否具有功能(Duan et al., 2020; Yu et al., 2020)等, 这些科学问题都有待深入研究。(责任编辑: 白羽红)
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1073/pnas.1608449113URLPMID:27566404 [本文引用: 2]

Receptor-like kinase FERONIA (FER) plays a crucial role in plant response to small molecule hormones [e.g., auxin and abscisic acid (ABA)] and peptide signals [e.g., rapid alkalinization factor (RALF)]. It remains unknown how FER integrates these different signaling events in the control of cell growth and stress responses. Under stress conditions, increased levels of ABA will inhibit cell elongation in the roots. In our previous work, we have shown that FER, through activation of the guanine nucleotide exchange factor 1 (GEF1)/4/10-Rho of Plant 11 (ROP11) pathway, enhances the activity of the phosphatase ABA Insensitive 2 (ABI2), a negative regulator of ABA signaling, thereby inhibiting ABA response. In this study, we found that both RALF and ABA activated FER by increasing the phosphorylation level of FER. The FER loss-of-function mutant displayed strong hypersensitivity to both ABA and abiotic stresses such as salt and cold conditions, indicating that FER plays a key role in ABA and stress responses. We further showed that ABI2 directly interacted with and dephosphorylated FER, leading to inhibition of FER activity. Several other ABI2-like phosphatases also function in this pathway, and ABA-dependent FER activation required PYRABACTIN RESISTANCE (PYR)/PYR1-LIKE (PYL)/REGULATORY COMPONENTS OF ABA RECEPTORS (RCAR)-A-type protein phosphatase type 2C (PP2CA) modules. Furthermore, suppression of RALF1 gene expression, similar to disruption of the FER gene, rendered plants hypersensitive to ABA. These results formulated a mechanism for ABA activation of FER and for cross-talk between ABA and peptide hormone RALF in the control of plant growth and responses to stress signals.
DOI:10.1104/pp.114.246926URLPMID:25288636 [本文引用: 1]
URLPMID:20400488 [本文引用: 3]
DOI:10.1126/science.1153795URLPMID:18436742 [本文引用: 1]
Little is known about the way developmental cues affect how cells interpret their environment. We characterized the transcriptional response to high salinity of different cell layers and developmental stages of the Arabidopsis root and found that transcriptional responses are highly constrained by developmental parameters. These transcriptional changes lead to the differential regulation of specific biological functions in subsets of cell layers, several of which correspond to observable physiological changes. We showed that known stress pathways primarily control semiubiquitous responses and used mutants that disrupt epidermal patterning to reveal cell-layer-specific and inter-cell-layer effects. By performing a similar analysis using iron deprivation, we identified common cell-type-specific stress responses and revealed the crucial role the environment plays in defining the transcriptional outcome of cell-fate decisions.
DOI:10.1038/ncomms4129URLPMID:24451849 [本文引用: 1]
In flowering plants, sperm are transported inside pollen tubes to the female gametophyte for fertilization. The female gametophyte induces rupture of the penetrating pollen tube, resulting in sperm release and rendering them available for fertilization. Here we utilize the Arabidopsis FERONIA (FER) receptor kinase mutants, whose female gametophytes fail to induce pollen tube rupture, to decipher the molecular mechanism of this critical male-female interactive step. We show that FER controls the production of high levels of reactive oxygen species at the entrance to the female gametophyte to induce pollen tube rupture and sperm release. Pollen tube growth assays in vitro and in the pistil demonstrate that hydroxyl free radicals are likely the most reactive oxygen molecules, and they induce pollen tube rupture in a Ca(2+)-dependent process involving Ca(2+) channel activation. Our results provide evidence for a RHO GTPase-based signalling mechanism to mediate sperm release for fertilization in plants.
DOI:10.1073/pnas.1005366107URLPMID:20876100 [本文引用: 8]
Plant RHO GTPases (RAC/ROPs) mediate multiple extracellular signals ranging from hormone to stress and regulate diverse cellular processes important for polarized cell growth, differentiation, development, reproduction, and responses to the environment. They shuttle between the GDP-bound inactive state and the GTP-bound activated state and their activation is predominantly mediated by a family of guanine nucleotide exchange factors (GEFs) referred to as ROPGEFs. Using the Arabidopsis ROPGEF1 as bait, we identified members of a receptor-like kinase (RLK) family as potential upstream regulators for RAC/ROP signaling. NADPH oxidase-derived reactive oxygen species (ROS) are emerging as important regulators for growth and development and play a crucial role in mediating RAC/ROP-regulated root hair development, a polarized cell growth process. We therefore screened T-DNA insertion mutants in these RLKs for root hair defects and found that mutations in one of them, At3g51550 encoding the FERONIA (FER) receptor-like kinase, induced severe root hair defects. We show that the fer phenotypes correlated with reduced levels of active RAC/ROPs and NADPH oxidase-dependent, auxin-regulated ROS accumulation in roots and root hairs and that up-regulating RAC/ROP signaling in fer countered the mutant phenotypes. Taken together, these observations strongly support FER as an upstream regulator for the RAC/ROP-signaled pathway that controls ROS-mediated root hair development. Moreover, FER was pulled down by ROP2 GTPase in a guanine nucleotide-regulated manner implying a dynamic signaling complex involving FER, a ROPGEF, and a RAC/ROP.
URLPMID:32214247 [本文引用: 1]
DOI:10.1016/j.molp.2019.09.004URLPMID:31541739 [本文引用: 5]
Pollen tube growth is crucial for successful fertilization. In Arabidopsis thaliana, the ANXUR (ANX)/BUPS receptor kinase complex controls and maintains pollen tube growth in response to autocrine rapid alkalinization factor 4/19 (RALF4/19) signaling; however, the molecular and cellular mechanisms underlying the ANX/BUPS-mediated regulation of pollen tube growth remain unclear. In this study, we found that pollen-specific LORELEI-like GPI-anchored proteins 2 and 3 (LLG2/3) promote pollen tube growth in vitro and in vivo. LLG2/3 interacte with ANX/BUPS in a RALF4-concentration-dependent manner, suggesting that ANX/BUPS and LLG2/3 might form a receptor-coreceptor complex for perceiving RALF peptide signals. Disruption of the ANX/BUPS-LLG2/3 interaction led to the cytoplasmic retention of ANX1/2, in either llg2/3 knockdown mutants or in anx1/2 mutants lacking the J region, which mediates the ANX/BUPS-LLG2/3 interaction. Moreover, we found that RALF4 induced the production of reactive oxygen species (ROS), which stimulate pollen tube growth and reduce pollen burst rate. ROS levels are reduced in the pollen tubes of LLG2/3 RNAi lines, and application of exogenous H2O2 could partially rescue the defective pollen tube growth of LLG2/3 RNAi lines. Taken together, our study identifies LLG2/3 as novel regulatory components of pollen tube growth, and shows that they chaperone ANX/BUPS for secretion to the apical plasma membrane of pollen tube and act as coreceptors of ANX/BUPS in the activation of ROS production for promoting pollen tube growth.
DOI:10.1016/j.cub.2018.01.023URLPMID:29456142 [本文引用: 7]
Cells maintain integrity despite changes in their mechanical properties elicited during growth and environmental stress. How cells sense their physical state and compensate for cell-wall damage is poorly understood, particularly in plants. Here we report that FERONIA (FER), a plasma-membrane-localized receptor kinase from Arabidopsis, is necessary for the recovery of root growth after exposure to high salinity, a widespread soil stress. The extracellular domain of FER displays tandem regions of homology with malectin, an animal protein known to bind di-glucose in vitro and important for protein quality control in the endoplasmic reticulum. The presence of malectin-like domains in FER and related receptor kinases has led to widespread speculation that they interact with cell-wall polysaccharides and can potentially serve a wall-sensing function. Results reported here show that salinity causes softening of the cell wall and that FER is necessary to sense these defects. When this function is disrupted in the fer mutant, root cells explode dramatically during growth recovery. Similar defects are observed in the mur1 mutant, which disrupts pectin cross-linking. Furthermore, fer cell-wall integrity defects can be rescued by treatment with calcium and borate, which also facilitate pectin cross-linking. Sensing of these salinity-induced wall defects might therefore be a direct consequence of physical interaction between the extracellular domain of FER and pectin. FER-dependent signaling elicits cell-specific calcium transients that maintain cell-wall integrity during salt stress. These results reveal a novel extracellular toxicity of salinity, and identify FER as a sensor of damage to the pectin-associated wall.
DOI:10.1146/annurev-arplant-042817-040557URLPMID:29539271 [本文引用: 1]
Plant cells are surrounded by cell walls protecting them from a myriad of environmental challenges. For successful habitat adaptation, extracellular cues are perceived at the cell wall and relayed to downstream signaling constituents to mediate dynamic cell wall remodeling and adapted intracellular responses. Plant malectin-like receptor kinases, also known as Catharanthus roseus receptor-like kinase 1-like proteins (CrRLK1Ls), take part in these perception and relay processes. CrRLK1Ls are involved in many different plant functions. Their ligands, interactors, and downstream signaling partners are being unraveled, and studies about CrRLK1Ls' roles in plant species other than the plant model Arabidopsis thaliana are beginning to flourish. This review focuses on recent CrRLK1L-related advances in cell growth, reproduction, hormone signaling, abiotic stress responses, and, particularly, immunity. We also give an overview of the comparative genomics and evolution of CrRLK1Ls, and present a brief outlook for future research.
DOI:10.1073/pnas.011405198URLPMID:11114160 [本文引用: 1]
The enhanced disease resistance 1 (edr1) mutation of Arabidopsis confers resistance to powdery mildew disease caused by the fungus Erysiphe cichoracearum. Resistance mediated by the edr1 mutation is correlated with induction of several defense responses, including host cell death. Double mutant analysis revealed that all edr1-associated phenotypes are suppressed by mutations that block salicylic acid (SA) perception (nim1) or reduce SA production (pad4 and eds1). The NahG transgene, which lowers endogenous SA levels, also suppressed edr1. In contrast, the ein2 mutation did not suppress edr1-mediated resistance and associated phenotypes, indicating that ethylene and jasmonic acid-induced responses are not required for edr1 resistance. The EDR1 gene was isolated by positional cloning and was found to encode a putative MAP kinase kinase kinase similar to CTR1, a negative regulator of ethylene responses in Arabidopsis. Taken together, these data suggest that EDR1 functions at the top of a MAP kinase cascade that negatively regulates SA-inducible defense responses. Putative orthologs of EDR1 are present in monocots such as rice and barley, indicating that EDR1 may regulate defense responses in a wide range of crop species.
DOI:10.1126/science.aao3642URLPMID:29242234 [本文引用: 2]
In flowering plants, fertilization requires complex cell-to-cell communication events between the pollen tube and the female reproductive tissues, which are controlled by extracellular signaling molecules interacting with receptors at the pollen tube surface. We found that two such receptors in Arabidopsis, BUPS1 and BUPS2, and their peptide ligands, RALF4 and RALF19, are pollen tube-expressed and are required to maintain pollen tube integrity. BUPS1 and BUPS2 interact with receptors ANXUR1 and ANXUR2 via their ectodomains, and both sets of receptors bind RALF4 and RALF19. These receptor-ligand interactions are in competition with the female-derived ligand RALF34, which induces pollen tube bursting at nanomolar concentrations. We propose that RALF34 replaces RALF4 and RALF19 at the interface of pollen tube-female gametophyte contact, thereby deregulating BUPS-ANXUR signaling and in turn leading to pollen tube rupture and sperm release.
DOI:10.1111/nph.15645URLPMID:30556141 [本文引用: 5]
Contents Summary 687 I. Introduction 687 II. Pollen tube membrane-localized receptors coordinate cell integrity and sperm release 689 III. RALF peptides mediate autocrine and paracrine signaling 689 IV. ROS and ion channel signaling mediate intracellular response 690 V. Involvements from pollen tube cell wall components 690 VI. Concluding remarks 691 Acknowledgements 692 Author contributions 692 References 692 SUMMARY: Unlike in animals, sperm in flowering plants are immotile and they are embraced as passive cargoes by a pollen tube which embarks on a long journey in the pistil to deliver them to the female gametophyte for fertilization. How the pollen tube switches from a rapid polarized growth towards its target to an abrupt disintegration for sperm cell release inside the female gametophyte is puzzling. Recent studies have shown that members of the Catharanthus roseus RLK1-like (CrRLK1L) receptor kinase family and their ligands, 5-kDa cysteine-rich peptide rapid alkalinization factors (RALFs), engage in an intricate balancing act involving autocrine and paracrine signaling to maintain pollen tube growth and induce timely tube rupture at the spatially confined pollen tube-female gametophyte interface. Here, we review recent progress related to pollen tube integrity control, mainly focusing on the molecular understanding of signaling as well as intracellular signaling nodes in Arabidopsis. Some missing links and future perspectives are also discussed.
DOI:10.1073/pnas.0812346106URLPMID:19383785 [本文引用: 2]
Cell elongation in plants is controlled by environmental cues such as light and internal growth regulators including plant steroid hormones, brassinosteroids (BRs). In this study, we found that 3 related receptor-like kinases (RLKs), HERCULES1, THESEUS1, and FERONIA, are transcriptionally induced by BRs and are down-regulated in the loss-of-function BR mutant bri1 and up-regulated in the constitutive BR-response mutant bes1-D. These RLKs belong to the CrRLK family that has 17 members in Arabidopsis. We hypothesize that these RLKs are involved in BR-regulated processes. Although 2 of the RLKs were recently found to mediate male-female interaction during pollen tube reception (FERONIA) and to sense cell wall integrity (THESEUS1), our genetic studies demonstrated that they are required for cell elongation during vegetative growth as herk1 the1 double and fer RNAi mutants displayed striking dwarf phenotypes. The herk1 the1 double mutant enhances the dwarf phenotype of bri1 and partially suppresses bes1-D phenotype, supporting a role of HERK1/THE1 in BR-mediated cell elongation. Microarray experiments demonstrated that these RLKs control the expression of a unique set of genes including those implicated in cell elongation and 16% of the genes affected in herk1 the1 are regulated by BRs. Our results, therefore, identify a previously unknown pathway that functions cooperatively with, but largely independent of the BR pathway to regulate cell elongation. The work establishes a platform to identify other signaling components in this important pathway for plant growth and provides a paradigm to study the coordination of independent pathways in the regulation of a common biological process.
DOI:10.1021/bi8001488URLPMID:18494498 [本文引用: 1]
Transient increases in the cytoplasmic Ca(2+) concentration are key events that initiate many cellular signaling pathways in response to developmental and environmental cues in plants; however, only a few extracellular mediators regulating cytoplasmic Ca(2+) singling are known to date. To identify endogenous cell signaling peptides regulating cytoplasmic Ca(2+) signaling, Arabidopsis seedlings expressing aequorin were used for an in vivo luminescence assay for Ca(2+) changes. These seedlings were challenged with fractions derived from plant extracts. Multiple heat-stable, protease-sensitive peaks of calcium elevating activity were observed after fractionation of these extracts by high-performance liquid chromatography. Tandem mass spectrometry identified the predominant active molecule isolated by a series of such chromatographic separations as a 49-amino acid polypeptide, AtRALF1 (the rapid alkalinization factor protein family). Within 40 s of treatment with nanomolar concentrations of the natural or synthetic version of the peptides, the cytoplasmic Ca(2+) level increased and reached its maximum. Prior treatment with a Ca(2+) chelator or inhibitor of IP 3-dependent signaling partially suppressed the AtRALF1-induced Ca(2+) concentration increase, indicating the likely involvement of Ca(2+) influx across the plasma membrane as well as release of Ca(2+) from intracellular reserves. Ca(2+) imaging using seedlings expressing the FRET-based Ca(2+) sensor yellow cameleon (YC) 3.6 showed that AtRALF1 could induce an elevation in Ca(2+) concentration in the surface cells of the root consistent with the very rapid effects of addition of AtRALF1 on Ca(2+) levels as reported by aequorin. Our data support a model in which the RALF peptide mediates Ca(2+)-dependent signaling events through a cell surface receptor, where it may play a role in eliciting events linked to stress responses or the modulation of growth.
DOI:10.1126/science.1244454URLPMID:24458638 [本文引用: 4]
Plant cells are immobile; thus, plant growth and development depend on cell expansion rather than cell migration. The molecular mechanism by which the plasma membrane initiates changes in the cell expansion rate remains elusive. We found that a secreted peptide, RALF (rapid alkalinization factor), suppresses cell elongation of the primary root by activating the cell surface receptor FERONIA in Arabidopsis thaliana. A direct peptide-receptor interaction is supported by specific binding of RALF to FERONIA and reduced binding and insensitivity to RALF-induced growth inhibition in feronia mutants. Phosphoproteome measurements demonstrate that the RALF-FERONIA interaction causes phosphorylation of plasma membrane H(+)-adenosine triphosphatase 2 at Ser(899), mediating the inhibition of proton transport. The results reveal a molecular mechanism for RALF-induced extracellular alkalinization and a signaling pathway that regulates cell expansion.
DOI:10.1016/j.cub.2016.06.053URLPMID:27524487 [本文引用: 1]
During the angiosperm (flowering-plant) life cycle, double fertilization represents the hallmark between diploid and haploid generations [1]. The success of double fertilization largely depends on compatible communication between the male gametophyte (pollen tube) and the maternal tissues of the flower, culminating in precise pollen tube guidance to the female gametophyte (embryo sac) and its rupture to release sperm cells. Several important factors involved in the pollen tube reception have been identified recently [2-6], but the underlying signaling pathways are far from being understood. Here, we report that a group of female-specific small proteins, early nodulin-like proteins (ENODLs, or ENs), are required for pollen tube reception. ENs are featured with a plastocyanin-like (PCNL) domain, an arabinogalactan (AG) glycomodule, and a predicted glycosylphosphatidylinositol (GPI) anchor motif. We show that ENs are asymmetrically distributed at the plasma membrane of the synergid cells and accumulate at the filiform apparatus, where arriving pollen tubes communicate with the embryo sac. EN14 strongly and specifically interacts with the extracellular domain of the receptor-like kinase FERONIA, localized at the synergid cell surface and known to critically control pollen tube reception [6]. Wild-type pollen tubes failed to arrest growth and to rupture after entering the ovules of quintuple loss-of-function EN mutants, indicating a central role of ENs in male-female communication and pollen tube reception. Moreover, overexpression of EN15 by the endogenous promoter caused disturbed pollen tube guidance and reduced fertility. These data suggest that female-derived GPI-anchored ENODLs play an essential role in male-female communication and fertilization.
DOI:10.1111/nph.12432URLPMID:23915272 [本文引用: 3]
We investigated a genetic pathway in root hair development in Arabidopsis thaliana, involving the receptor-like kinase FERONIA (FER), two guanine nucleotide exchange factors for ROPs (RopGEF4 and RopGEF10), and the small GTPase Rho of plants (ROPs). Loss- and gain-of-function analyses demonstrated distinct roles of RopGEF4 and RopGEF10 such that RopGEF4 is only important for root hair elongation, while RopGEF10 mainly contributes to root hair initiation. Domain dissection by truncation and domain-swapping experiments indicated that their functional distinctions were mainly contributed by the noncatalytic domains. Using fluorescent ratio imaging, we showed that functional loss of RopGEF4 and RopGEF10 additively reduced reactive oxygen species (ROS) production. Bimolecular fluorescence complementation experiments demonstrated that RopGEF4 and RopGEF10 had the same interaction specificity as ROPs, suggesting common downstream components. We further showed that the promoting effects of environmental cues such as exogenous auxin and phosphate limitation on root hair development depended on FER. However, although functional loss of RopGEF4 and RopGEF10 largely abolished FER-induced ROS production, it did not compromise the responses to FER-mediated environmental cues on root hair development. Our results demonstrated that RopGEF4 and RopGEF10 are genetic components in FER-mediated, developmentally (but not environmentally) regulated, root hair growth.
URLPMID:30822112 [本文引用: 1]
DOI:10.1016/j.plaphy.2004.03.009URLPMID:15191737 [本文引用: 1]
A number of protein sequences deduced from the molecular analysis of plant cDNA or genomic libraries can be grouped in relation to a defined number of cysteine residues located in distinct positions of their sequences. This is the case for a group of around 500 polypeptides from different species that contain a small domain (less than 100 amino acids residues) displaying a pattern of eight-cysteines in a specific order. The plant sequences containing this motif belong to proteins having different functions, ranging from storage, protection, enzyme inhibition and lipid transfer, to cell wall structure. The eight-cysteine motif (8CM) appears to be a structural scaffold of conserved helical regions connected by variable loops, as observed by three-dimensional structure analysis. It is proposed that the cysteine residues would form a network of disulfide bridges necessary, for the maintenance of the tertiary structure of the molecule together with the central helical core, while the variable loops would provide the sequences required for the specific functions of the proteins.
DOI:10.1105/tpc.113.120642URLPMID:24610725 [本文引用: 1]
In flowering plants, pollen germinates on the stigma and pollen tubes grow through the style to fertilize the ovules. Enzymatic production of reactive oxygen species (ROS) has been suggested to be involved in pollen tube tip growth. Here, we characterized the function and regulation of the NADPH oxidases RbohH and RbohJ (Respiratory burst oxidase homolog H and J) in pollen tubes in Arabidopsis thaliana. In the rbohH and rbohJ single mutants, pollen tube tip growth was comparable to that of the wild type; however, tip growth was severely impaired in the double mutant. In vivo imaging showed that ROS accumulation in the pollen tube was impaired in the double mutant. Both RbohH and RbohJ, which contain Ca(2+) binding EF-hand motifs, possessed Ca(2+)-induced ROS-producing activity and localized at the plasma membrane of the pollen tube tip. Point mutations in the EF-hand motifs impaired Ca(2+)-induced ROS production and complementation of the double mutant phenotype. We also showed that a protein phosphatase inhibitor enhanced the Ca(2+)-induced ROS-producing activity of RbohH and RbohJ, suggesting their synergistic activation by protein phosphorylation and Ca(2+). Our results suggest that ROS production by RbohH and RbohJ is essential for proper pollen tube tip growth, and furthermore, that Ca(2+)-induced ROS positive feedback regulation is conserved in the polarized cell growth to shape the long tubular cell.
DOI:10.1074/jbc.M110.160531URLPMID:20843791 [本文引用: 1]
Plasma membrane compartmentalization spatiotemporally regulates cell-autonomous immune signaling in animal cells. To elucidate immediate early protein dynamics at the plant plasma membrane in response to the bacterial pathogen-associated molecular pattern (PAMP) flagellin (flg22) we employed quantitative mass spectrometric analysis on detergent-resistant membranes (DRMs) of Arabidopsis thaliana suspension cells. This approach revealed rapid and profound changes in DRM protein composition following PAMP treatment, prominently affecting proton ATPases and receptor-like kinases, including the flagellin receptor FLS2. We employed reverse genetics to address a potential contribution of a subset of these proteins in flg22-triggered cellular responses. Mutants of three candidates (DET3, AHA1, FER) exhibited a conspicuous defect in the PAMP-triggered accumulation of reactive oxygen species. In addition, these mutants showed altered mitogen-activated protein kinase (MAPK) activation, a defect in PAMP-triggered stomatal closure as well as altered bacterial infection phenotypes, which revealed three novel players in elicitor-dependent oxidative burst control and innate immunity. Our data provide evidence for dynamic elicitor-induced changes in the membrane compartmentalization of PAMP signaling components.
DOI:10.7554/eLife.06587URL [本文引用: 15]
DOI:10.1007/s11427-016-0118-xURLPMID:27525988 [本文引用: 1]
Different from animals, sessile plants are equipped with a large receptor-like kinase (RLK) superfamily. RLKs are a family of single trans-membrane proteins with divergent N-terminal extracellular domains capped by a signal peptide and C-terminal intracellular kinase. Researches in the last two decades have uncovered an increasing number of RLKs that regulate plant development, stress response and sexual reproduction, highlighting a dominant role of RLK signaling in cell-to-cell communications. Sexual reproduction in flowering plants is featured by interactions between the male gametophyte and the female tissues to facilitate sperm delivery and fertilization. Emerging evidences suggest that RLKs regulate almost every aspect of plant reproductive process, especially during pollination. Therefore, in this review we will focus mainly on the function and signaling of RLKs in plant male-female interaction and discuss the future prospects on these topics.
URLPMID:29571836 [本文引用: 1]
DOI:10.1016/j.chom.2014.02.009URLPMID:24629339 [本文引用: 1]
The Arabidopsis immune receptor FLS2 senses the bacterial flagellin epitope flg22 to activate transient elevation of cytosolic calcium ions, production of reactive oxygen species (ROS), and other signaling events to coordinate antimicrobial defenses, such as stomatal closure that limits bacterial invasion. However, how FLS2 regulates these signaling events remains largely unknown. Here we show that the receptor-like cytoplasmic kinase BIK1, a component of the FLS2 immune receptor complex, not only positively regulates flg22-triggered calcium influx but also directly phosphorylates the NADPH oxidase RbohD at specific sites in a calcium-independent manner to enhance ROS generation. Furthermore, BIK1 and RbohD form a pathway that controls stomatal movement in response to flg22, thereby restricting bacterial entry into leaf tissues. These findings highlight a direct role of the FLS2 complex in the regulation of RbohD-mediated ROS production and stomatal defense.
DOI:10.1371/journal.pgen.1003704URLPMID:23990797 [本文引用: 1]
Cellulose represents the most abundant biopolymer in nature and has great economic importance. Cellulose chains pack laterally into crystalline forms, stacking into a complicated crystallographic structure. However, the mechanism of cellulose crystallization is poorly understood. Here, via functional characterization, we report that Brittle Culm1 (BC1), a COBRA-like protein in rice, modifies cellulose crystallinity. BC1 was demonstrated to be a glycosylphosphatidylinositol (GPI) anchored protein and can be released into cell walls by removal of the GPI anchor. BC1 possesses a carbohydrate-binding module (CBM) at its N-terminus. In vitro binding assays showed that this CBM interacts specifically with crystalline cellulose, and several aromatic residues in this domain are essential for binding. It was further demonstrated that cell wall-localized BC1 via the CBM and GPI anchor is one functional form of BC1. X-ray diffraction (XRD) assays revealed that mutations in BC1 and knockdown of BC1 expression decrease the crystallite width of cellulose; overexpression of BC1 and the CBM-mutated BC1s caused varied crystallinity with results that were consistent with the in vitro binding assay. Moreover, interaction between the CBM and cellulose microfibrils was largely repressed when the cell wall residues were pre-stained with two cellulose dyes. Treating wild-type and bc1 seedlings with the dyes resulted in insensitive root growth responses in bc1 plants. Combined with the evidence that BC1 and three secondary wall cellulose synthases (CESAs) function in different steps of cellulose production as revealed by genetic analysis, we conclude that BC1 modulates cellulose assembly by interacting with cellulose and affecting microfibril crystallinity.
URLPMID:27081182 [本文引用: 1]
DOI:10.1104/pp.16.00191URLPMID:27208283 [本文引用: 1]
Root hair cells and pollen tubes, like fungal hyphae, possess a typical tip or polar cell expansion with growth limited to the apical dome. Cell expansion needs to be carefully regulated to produce a correct shape and size. Polar cell growth is sustained by oscillatory feedback loops comprising three main components that together play an important role regulating this process. One of the main components are reactive oxygen species (ROS) that, together with calcium ions (Ca(2+)) and pH, sustain polar growth over time. Apoplastic ROS homeostasis controlled by NADPH oxidases as well as by secreted type III peroxidases has a great impact on cell wall properties during cell expansion. Polar growth needs to balance a focused secretion of new materials in an extending but still rigid cell wall in order to contain turgor pressure. In this review, we discuss the gaps in our understanding of how ROS impact on the oscillatory Ca(2+) and pH signatures that, coordinately, allow root hair cells and pollen tubes to expand in a controlled manner to several hundred times their original size toward specific signals.
DOI:10.1126/science.aao5467URLPMID:29242232 [本文引用: 1]
The communication of changes in the extracellular matrix to the interior of the cell is crucial for a cell's function. The extracellular peptides of the RAPID ALKALINIZATION FACTOR (RALF) family have been identified as ligands of receptor-like kinases of the CrRLK1L subclass, but the exact mechanism of their perception is unclear. We found that Arabidopsis RALF4 and RALF19 redundantly regulate pollen tube integrity and growth, and that their function depends on pollen-expressed proteins of the LEUCINE-RICH REPEAT EXTENSIN (LRX) family, which play a role in cell wall development but whose mode of action is not understood. The LRX proteins interact with RALFs, monitoring cell wall changes, which are communicated to the interior of the pollen tube via the CrRLK1L pathway to sustain normal growth.
DOI:10.1073/pnas.0708586104URLPMID:18079291 [本文引用: 1]
Root hairs show highly localized cell expansion focused to their growing tips. This growth pattern is accomplished through restriction of secretion to the elongating apex and modulation of cell wall properties, with the wall just behind the tip becoming rigidified to resist the lateral expansive forces of turgor. In this report we show that root hairs exhibit oscillating growth that is associated with oscillating increases in extracellular pH and reactive oxygen species (ROS), which lag growth by approximately 7 s. Consistent with a role for these changes in growth control, artificially increasing extracellular pH arrested root hair elongation, whereas decreasing pH elicited bursting at the tip. Similarly, application of exogenous ROS arrested elongation, whereas scavenging of ROS led to root hair bursting. Roots hairs of the root hair-defective rhd2-1 mutant, which lack a functional version of the NADPH oxidase ATRBOH C, burst at the transition to tip growth. This phenotype could be rescued by elevating the pH of the growth medium to >/=6.0. Such rescued root hairs showed reduced cytoplasmic ROS levels and a lack of the oscillatory production of ROS at the tip. However, they exhibited apparently normal tip growth, including generation of the tip-focused Ca(2+) gradient thought to drive apical growth, indicating that ATRBOH C is not absolutely required to sustain tip growth. These observations indicate that root hair elongation is coupled to spatially distinct regulation of extracellular pH and ROS production that likely affect wall properties associated with the polarized expansion of the cell.
URLPMID:28507137 [本文引用: 9]
DOI:10.1105/tpc.112.107904URLPMID:23532072 [本文引用: 1]
Pathogen-associated molecular pattern (PAMP)-trigged immunity (PTI) is the first defensive line of plant innate immunity and is mediated by pattern recognition receptors. Here, we show that a mutation in BR-SIGNALING KINASE1 (BSK1), a substrate of the brassinosteroid (BR) receptor BRASSINOSTEROID INSENSITIVE1, suppressed the powdery mildew resistance caused by a mutation in ENHANCED DISEASE RESISTANCE2, which negatively regulates powdery mildew resistance and programmed cell death, in Arabidopsis thaliana. A loss-of-function bsk1 mutant displayed enhanced susceptibility to virulent and avirulent pathogens, including Golovinomyces cichoracearum, Pseudomonas syringae, and Hyaloperonospora arabidopsidis. The bsk1 mutant also accumulated lower levels of salicylic acid upon infection with G. cichoracearum and P. syringae. BSK1 belongs to a receptor-like cytoplasmic kinase family and displays kinase activity in vitro; this kinase activity is required for its function. BSK1 physically associates with the PAMP receptor FLAGELLIN SENSING2 and is required for a subset of flg22-induced responses, including the reactive oxygen burst, but not for mitogen-activated protein kinase activation. Our data demonstrate that BSK1 is involved in positive regulation of PTI. Together with previous findings, our work indicates that BSK1 represents a key component directly involved in both BR signaling and plant immunity.
DOI:10.1126/science.1243825URLPMID:24114786 [本文引用: 1]
Flagellin perception in Arabidopsis is through recognition of its highly conserved N-terminal epitope (flg22) by flagellin-sensitive 2 (FLS2). Flg22 binding induces FLS2 heteromerization with BRASSINOSTEROID INSENSITIVE 1-associated kinase 1 (BAK1) and their reciprocal activation followed by plant immunity. Here, we report the crystal structure of FLS2 and BAK1 ectodomains complexed with flg22 at 3.06 angstroms. A conserved and a nonconserved site from the inner surface of the FLS2 solenoid recognize the C- and N-terminal segment of flg22, respectively, without oligomerization or conformational changes in the FLS2 ectodomain. Besides directly interacting with FLS2, BAK1 acts as a co-receptor by recognizing the C terminus of the FLS2-bound flg22. Our data reveal the molecular mechanisms underlying FLS2-BAK1 complex recognition of flg22 and provide insight into the immune receptor complex activation.
DOI:10.1111/j.1399-3054.2009.01313.xURLPMID:19947976 [本文引用: 1]
Reactive oxygen species (ROS) are now recognized as important regulators of plant developmental programs and recent work on tip-growing systems has revealed a central role for the NADPH oxidases in generating such developmentally important ROS. Tip-growing cells have also shown that the functions of cytosolic ROS, acting as regulators of activities such as ion channel gating, are closely linked to those of ROS produced to the apoplast, where they act to modulate cell wall properties. Thus, coordination of ROS production and their activities between compartments is emerging as an important theme in our understanding of how growth and developmental programs are integrated.
DOI:10.1038/s41586-019-1409-7URLPMID:31291642 [本文引用: 13]
Receptor kinases of the Catharanthus roseus RLK1-like (CrRLK1L) family have emerged as important regulators of plant reproduction, growth and responses to the environment(1). Endogenous RAPID ALKALINIZATION FACTOR (RALF) peptides(2) have previously been proposed as ligands for several members of the CrRLK1L family(1). However, the mechanistic basis of this perception is unknown. Here we report that RALF23 induces a complex between the CrRLK1L FERONIA (FER) and LORELEI (LRE)-LIKE GLYCOSYLPHOSPHATIDYLINOSITOL (GPI)-ANCHORED PROTEIN 1 (LLG1) to regulate immune signalling. Structural and biochemical data indicate that LLG1 (which is genetically important for RALF23 responses) and the related LLG2 directly bind RALF23 to nucleate the assembly of RALF23-LLG1-FER and RALF23-LLG2-FER heterocomplexes, respectively. A conserved N-terminal region of RALF23 is sufficient for the biochemical recognition of RALF23 by LLG1, LLG2 or LLG3, and binding assays suggest that other RALF peptides that share this conserved N-terminal region may be perceived by LLG proteins in a similar manner. Structural data also show that RALF23 recognition is governed by the conformationally flexible C-terminal sides of LLG1, LLG2 and LLG3. Our work reveals a mechanism of peptide perception in plants by GPI-anchored proteins that act together with a phylogenetically unrelated receptor kinase. This provides a molecular framework for understanding how diverse RALF peptides may regulate multiple processes, through perception by distinct heterocomplexes of CrRLK1L receptor kinases and GPI-anchored proteins of the LRE and LLG family.
DOI:10.1093/jxb/erz378URLPMID:31433471 [本文引用: 1]
The ratio between carbon (C) and nitrogen (N) utilization must be precisely coordinated to enable plant growth. Although numerous physiological studies have examined carbon/nitrogen (C/N) ratios, the mechanisms of sensing the C/N balance and C/N signaling remain elusive. Here, we report that a mutation of FERONIA (FER), a receptor kinase that plays versatile roles in plant cell growth and stress responses, caused hypersensitivity to a high C/N ratio in Arabidopsis. In contrast, FER-overexpressing plants displayed more resistant phenotypes. FER can interact with and phosphorylate ATL6, an E3 ubiquitin ligase that has been shown to regulate plant C/N responses. FER-mediated ATL6 phosphorylation enhanced the interaction between ATL6 and its previously identified target 14-3-3 proteins, thus decreasing 14-3-3 protein levels, leading to an increased insensitivity to high C/N ratios. Further analyses showed that the rapid alkalinization factor peptide (RALF1), which is a ligand of FER, also influenced the stability of 14-3-3 proteins via a FER-ATL6-mediated pathway. These findings reveal a novel regulatory mechanism that links the RALF1/FER-ATL6 pathway to whole-plant C/N responses and growth.
DOI:10.1016/j.bbrc.2015.07.132URLPMID:26232644 [本文引用: 1]
Cell expansion is coordinated by several cues, but available energy is the major factor determining growth. Receptor protein kinase FERONIA (FER) is a master regulator of cell expansion, but the details of its control mechanisms are not clear. Here we show that FER interacts with cytosolic enzyme glyceraldehyde-3-phosphate dehydrogenase (GAPDH, GAPC1 and GAPC2), that catalyzes a key reaction in glycolysis, which contributes to energy production. When there is an FER deficiency, there are corresponding decreases in the enzyme activity of GAPDH and increased amounts of starch. More importantly, gapc1/2 mutants mimic fer4 mutants. These data indicate that FER regulated starch content is an evolutionarily conserved function in plants that connects the cell expansion and energy metabolism pathways.
DOI:10.1105/tpc.109.069609URLPMID:20418496 [本文引用: 1]
The plasma membrane H(+)-ATPase (PM H(+)-ATPase) plays an important role in the regulation of ion and metabolite transport and is involved in physiological processes that include cell growth, intracellular pH, and stomatal regulation. PM H(+)-ATPase activity is controlled by many factors, including hormones, calcium, light, and environmental stresses like increased soil salinity. We have previously shown that the Arabidopsis thaliana Salt Overly Sensitive2-Like Protein Kinase5 (PKS5) negatively regulates the PM H(+)-ATPase. Here, we report that a chaperone, J3 (DnaJ homolog 3; heat shock protein 40-like), activates PM H(+)-ATPase activity by physically interacting with and repressing PKS5 kinase activity. Plants lacking J3 are hypersensitive to salt at high external pH and exhibit decreased PM H(+)-ATPase activity. J3 functions upstream of PKS5 as double mutants generated using j3-1 and several pks5 mutant alleles with altered kinase activity have levels of PM H(+)-ATPase activity and responses to salt at alkaline pH similar to their corresponding pks5 mutant. Taken together, our results demonstrate that regulation of PM H(+)-ATPase activity by J3 takes place via inactivation of the PKS5 kinase.
DOI:10.1104/pp.16.00302URLPMID:27013021 [本文引用: 1]
In order to understand factors controlling the synthesis and deposition of cellulose, we have studied the Arabidopsis (Arabidopsis thaliana) double mutant shaven3 shaven3-like1 (shv3svl1), which was shown previously to exhibit a marked cellulose deficiency. We discovered that exogenous sucrose (Suc) in growth medium greatly enhances the reduction in hypocotyl elongation and cellulose content of shv3svl1 This effect was specific to Suc and was not observed with other sugars or osmoticum. Live-cell imaging of fluorescently labeled cellulose synthase complexes revealed a slowing of cellulose synthase complexes in shv3svl1 compared with the wild type that is enhanced in a Suc-conditional manner. Solid-state nuclear magnetic resonance confirmed a cellulose deficiency of shv3svl1 but indicated that cellulose crystallinity was unaffected in the mutant. A genetic suppressor screen identified mutants of the plasma membrane Suc/H(+) symporter SUC1, indicating that the accumulation of Suc underlies the Suc-dependent enhancement of shv3svl1 phenotypes. While other cellulose-deficient mutants were not specifically sensitive to exogenous Suc, the feronia (fer) receptor kinase mutant partially phenocopied shv3svl1 and exhibited a similar Suc-conditional cellulose defect. We demonstrate that shv3svl1, like fer, exhibits a hyperpolarized plasma membrane H(+) gradient that likely underlies the enhanced accumulation of Suc via Suc/H(+) symporters. Enhanced intracellular Suc abundance appears to favor the partitioning of carbon to starch rather than cellulose in both mutants. We conclude that SHV3-like proteins may be involved in signaling during cell expansion that coordinates proton pumping and cellulose synthesis.
DOI:10.1093/pcp/pcy146URLPMID:30107607 [本文引用: 1]
BRASSINAZOLE RESISTANT 1 (BZR1), the critical regulator of brassinosteroid (BR) response, participates in various BR-mediated developmental processes. However, the roles of BZR1 in stress tolerance are less clear. Here, we found that BZR1-like protein in tomato controls BR response and is involved in thermotolerance by regulating the FERONIA (FER) homologs. The CRISPR-bzr1 mutant showed reduced growth and was not responsive to 24-epibrassinolide (EBR) with regard to the promotion of plant growth. Mutation in BZR1 impaired the induction of RESPIRATORY BURST OXIDASE HOMOLOG1 (RBOH1), production of H2O2 in the apoplast and heat tolerance. Exogenous H2O2 recovered the heat tolerance of the tomato bzr1 mutant. Overexpression of BZR1 enhanced the production of apoplastic H2O2 and heat stress responses. However, silencing of RBOH1 abolished the BZR1-mediated heat tolerance. Further analysis showed that BZR1 bound to the promoters of FERONIA2 (FER2) and FER3 and induced their expression. Silencing of FER2/3 suppressed BZR1-dependent BR signaling for the induction of RBOH1 transcripts, accumulation of apoplastic H2O2 and heat tolerance. These results indicate that BZR1 regulates heat stress responses in tomato through RBOH1-dependent reactive oxygen species (ROS) signaling, which is at least partially mediated by FER2 and FER3.
DOI:10.1073/pnas.1212547109URLPMID:22908257 [本文引用: 2]
Plant growth and development are controlled by a delicate balance of hormonal cues. Growth-promoting hormones and growth-inhibiting counterparts often antagonize each other in their action, but the molecular mechanisms underlying these events remain largely unknown. Here, we report a cross-talk mechanism that enables a receptor-like kinase, FERONIA (FER), a positive regulator of auxin-promoted growth, to suppress the abscisic acid (ABA) response through activation of ABI2, a negative regulator of ABA signaling. The FER pathway consists of a FER kinase interacting with guanine exchange factors GEF1, GEF4, and GEF10 that, in turn, activate GTPase ROP11/ARAC10. Arabidopsis mutants disrupted in any step of the FER pathway, including fer, gef1gef4gef10, or rop11/arac10, all displayed an ABA-hypersensitive response, implicating the FER pathway in the suppression mechanism. In search of the target for the FER pathway, we found that the ROP11/ARAC10 protein physically interacted with the ABI2 phosphatase and enhanced its activity, thereby linking the FER pathway with the inhibition of ABA signaling.
DOI:10.1089/ars.2019.7823URLPMID:31989831 [本文引用: 1]
Significance: Plant chloroplasts generate reactive oxygen species (ROS) during photosynthesis, especially under stresses. The sulfhydryl groups of protein cysteine residues are susceptible to redox modifications, which regulate protein structure and function, and thus different signaling and metabolic processes. The ROS-governed protein thiol redox switches play important roles in chloroplasts. Recent Advances: Various high-throughput thiol redox proteomic approaches have been developed, and they have enabled the improved understanding of redox regulatory mechanisms in chloroplasts. For example, the thioredoxin-modulated antioxidant enzymes help to maintain cellular ROS homeostasis. The light- and dark-dependent redox regulation of photosynthetic electron transport, the Calvin/Benson cycle, and starch biosynthesis ensures metabolic coordination and efficient energy utilization. In addition, redox cascades link the light with the dynamic changes of metabolites in nitrate and sulfur assimilation, shikimate pathway, and biosynthesis of fatty acid hormone as well as purine, pyrimidine, and thiamine. Importantly, redox regulation of tetrapyrrole and chlorophyll biosynthesis is critical to balance the photodynamic tetrapyrrole intermediates and prevent oxidative damage. Moreover, redox regulation of diverse elongation factors, chaperones, and kinases plays an important role in the modulation of gene expression, protein conformation, and posttranslational modification that contribute to photosystem II (PSII) repair, state transition, and signaling in chloroplasts. Critical Issues: This review focuses on recent advances in plant thiol redox proteomics and redox protein networks toward understanding plant chloroplast signaling, metabolism, and stress responses. Future Directions: Using redox proteomics integrated with biochemical and molecular genetic approaches, detailed studies of cysteine residues, their redox states, cross talk with other modifications, and the functional implications will yield a holistic understanding of chloroplast stress responses.
DOI:10.1016/j.cbpa.2013.09.016URLPMID:24128440 [本文引用: 1]
Covalent attachment of glycosylphosphatidylinositols (GPIs) to the protein C-terminus is one of the most common posttranslational modifications in eukaryotic cells. In addition to anchoring surface proteins to the cell membrane, GPIs also have many other important biological functions, determined by their unique structure and property. This account has reviewed the recent progress made in disclosing GPI and GPI-anchored protein biosynthesis, in the chemical and chemoenzymatic synthesis of GPIs and GPI-anchored proteins, and in understanding the conformation, organization, and distribution of GPIs in the lipid membrane.
DOI:10.1111/pce.12542URLPMID:25808946 [本文引用: 1]
Salinity stress includes both osmotic and ionic toxicity. Sodium homeostasis is influenced by Na(+) uptake and extrusion, vacuolar Na(+) compartmentation and root to shoot Na(+) translocation via transpiration. The knockout mutant of the Arabidopsis heterotrimeric G-protein Gbeta subunit, agb1, is hypersensitive to salt, exhibiting a leaf bleaching phenotype. We show that AGB1 is mainly involved in the ionic toxicity component of salinity stress and plays roles in multiple processes of Na(+) homeostasis. agb1 mutants accumulate more Na(+) and less K(+) in both shoots and roots of hydroponically grown plants, as measured by inductively coupled plasma atomic emission spectrometry. agb1 plants have higher root to shoot translocation rates of radiolabelled (24) Na(+) under transpiring conditions, as a result of larger stomatal apertures and increased stomatal conductance. (24) Na(+) tracer experiments also show that (24) Na(+) uptake rates by excised roots of agb1 and wild type are initially equal, but that agb1 has higher net Na(+) uptake at 90 min, implicating possible involvement of AGB1 in the regulation of Na(+) efflux. Calcium alleviates the salt hypersensitivity of agb1 by reducing Na(+) accumulation to below the toxicity threshold. Our results provide new insights into the regulatory pathways underlying plant responses to salinity stress, an important agricultural problem.
DOI:10.1104/pp.17.01277URLPMID:29301953 [本文引用: 10]
Heterotrimeric guanine nucleotide-binding (G) proteins are composed of Galpha, Gbeta, and Ggamma subunits and function as molecular switches in signal transduction. In Arabidopsis (Arabidopsis thaliana), there are one canonical Galpha (GPA1), three extra-large Galpha (XLG1, XLG2, and XLG3), one Gbeta (AGB1), and three Ggamma (AGG1, AGG2, and AGG3) subunits. To elucidate AGB1 molecular signaling, we performed immunoprecipitation using plasma membrane-enriched proteins followed by mass spectrometry to identify the protein interactors of AGB1. After eliminating proteins present in the control immunoprecipitation, commonly identified contaminants, and organellar proteins, a total of 103 candidate AGB1-associated proteins were confidently identified. We identified all of the G protein subunits except XLG1, receptor-like kinases, Ca(2+) signaling-related proteins, and 14-3-3-like proteins, all of which may couple with or modulate G protein signaling. We confirmed physical interaction between AGB1 and the receptor-like kinase FERONIA (FER) using bimolecular fluorescence complementation. The Rapid Alkalinization Factor (RALF) family of polypeptides have been shown to be ligands of FER. In this study, we demonstrate that RALF1 regulates stomatal apertures and does so in a G protein-dependent manner, inhibiting stomatal opening and promoting stomatal closure in Columbia but not in agb1 mutants. We further show that AGGs and XLGs, but not GPA1, participate in RALF1-mediated stomatal signaling. Our results suggest that FER acts as a G protein-coupled receptor for plant heterotrimeric G proteins.
[本文引用: 2]
DOI:10.1073/pnas.1816991115URLPMID:30514814 [本文引用: 3]
The perception and relay of cell-wall signals are critical for plants to regulate growth and stress responses, but the underlying mechanisms are poorly understood. We found that the cell-wall leucine-rich repeat extensins (LRX) 3/4/5 are critical for plant salt tolerance in Arabidopsis The LRXs physically associate with the RAPID ALKALINIZATION FACTOR (RALF) peptides RALF22/23, which in turn interact with the plasma membrane-localized receptor-like protein kinase FERONIA (FER). The lrx345 triple mutant as well as fer mutant plants display retarded growth and salt hypersensitivity, which are mimicked by overexpression of RALF22/23 Salt stress promotes S1P protease-dependent release of mature RALF22 peptides. Treatment of roots with mature RALF22/23 peptides or salt stress causes the internalization of FER. Our results suggest that the LRXs, RALFs, and FER function as a module to transduce cell-wall signals to regulate plant growth and salt stress tolerance.
DOI:10.1016/j.pbi.2019.03.004URLPMID:30999163 [本文引用: 1]
In flowering plants, extensive male-female interactions during pollen germination on the stigma, pollen tube growth and guidance in the transmitting tract, and pollen tube reception by the female gametophyte are required for successful double fertilization in which various signaling cascades are involved. Peptide/receptor-like kinase-mediated signaling has been found playing important roles in these male-female interactions. Here, we mainly summarized the progress made on the regulatory roles of peptide/receptor-like kinase-mediated signaling pathways in four critical stages during reproduction in higher plants.
DOI:10.1016/j.bbamem.2015.12.018URLPMID:26706096 [本文引用: 1]
Glycosylphosphatidylinositol (GPI)-anchored proteins (GPI-APs) are a class of membrane proteins containing a soluble protein attached by a conserved glycolipid anchor to the external leaflet of the plasma membrane. In polarized epithelial cells, GPI-APs are predominantly sorted to the apical surface in the trans-Golgi network (TGN) by clustering in sphingolipid- and cholesterol-dependent microdomains (or rafts), which have been proposed to act as apical sorting platforms. Recent data indicate that the mechanisms of GPI-AP sorting, occurring in the Golgi, control both the membrane transport of GPI-APs and their specific activity at the apical surface of fully polarized epithelial cells. Here, we discuss the most recent findings and the factors regulating apical sorting of GPI-APs at the Golgi in polarized epithelial cells. We also underline the differences in the plasma membrane organization of GPI-APs between polarized and non-polarized cells supporting the existence of various mechanisms that control GPI-AP organization in different cell types.
FERONIA interacts with ABI2-type phosphatases to facilitate signaling cross-talk between abscisic acid and RALF peptide in Arabidopsismodification
2
2016
... RALF1通过诱导FER磷酸化将其激活(
... 盐胁迫引起ABA含量升高进而激活FER, 调控下游通路; 同时, FER也通过GEF1/4/10-ROP11途径增强ABI2的活性, ABI2直接与FER互作将其去磷酸化而失活, 从而抑制ABA响应.虽然fer突变体对ABA和盐胁迫非常敏感, 但是FER调控并非全部依赖ABA, 其它信号通路也可能参与其中, 具体机制尚不清楚(
Glycosylphosphatidylinositol anchoring: control through modification
1
2014
... 植物糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol-anchored protein, GPI-AP)定位于细胞膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与调节细胞生殖、生长以及免疫与盐碱胁迫应答等多种生物学过程(
FERONIA is a key modulator of brassinosteroid and ethylene responsiveness in Arabidopsis hypocotyls.
3
2010
... LLG1通过与FER的exJM结合, 协助FER定位于根部细胞质膜, 进而共同调控根与根毛生长(
... LLG1可能与FER共同参与脱落酸(ABA)、生长素、乙烯及油菜素内酯(BR)等激素信号转导过程, 从而调控根与根毛生长(
... ), 下胚轴伸长对乙烯和BR敏感(
Cell identity mediates the response of Arabidopsis roots to abiotic stress.
1
2008
... 盐离子干扰植物根部细胞壁结构, 导致细胞壁完整性降低, 抑制细胞极性生长, 引起细胞爆裂(
Reactive oxygen species mediate pollen tube rupture to release sperm for fertilization in Arabidopsis.
1
2014
... LLG1与FER相互作用共同感受RALF1, 形成LLG1-RALF1-FER复合物并激活FER, 进而调控根生长.外源施加RALF1抑制野生型幼苗根的生长, 但并不影响llg1与fer-4突变体的根长.激活的FER与RopGEFs互作, 将RAC/ROP从与GDP结合的无活性状态转变为与GTP结合的活化状态.fer突变体与几种rac/rop突变体表型相似, 表明fer与rac/rop之间有调控关系(
FERONIA receptor-like kinase regulates RHO GTPase signaling of root hair development
8
2010
... 在感受胞外RALF4/19后, LLG2/3-ANX1/2-BUPS 1/2复合体通过与GDP-ROP1相互作用, 将其激活为GTP-ROP1, 进而激活下游花粉特异表达的NADPH氧化酶RbohH/J, 产生活性氧(ROS), 导致ROS在花粉管顶端积累, 调节花粉管生长, 降低花粉管的爆裂率(
... LLG1通过与FER的exJM结合, 协助FER定位于根部细胞质膜, 进而共同调控根与根毛生长(
... )、根毛生长(
... ), 根毛出现卷曲或异常分枝(
... , 可以恢复其野生型表型(
... LLG1与FER相互作用共同感受RALF1, 形成LLG1-RALF1-FER复合物并激活FER, 进而调控根生长.外源施加RALF1抑制野生型幼苗根的生长, 但并不影响llg1与fer-4突变体的根长.激活的FER与RopGEFs互作, 将RAC/ROP从与GDP结合的无活性状态转变为与GTP结合的活化状态.fer突变体与几种rac/rop突变体表型相似, 表明fer与rac/rop之间有调控关系(
... LLG1可能与FER共同参与脱落酸(ABA)、生长素、乙烯及油菜素内酯(BR)等激素信号转导过程, 从而调控根与根毛生长(
... 突变体根毛伸长对生长素不敏感(
FERONIA controls pectin- and nitric oxide-mediated male-female interaction
1
2020
... LLG作为质膜定位的RLKs (如FER和FLS2)的分子伴侣, 负责将其从内质网运送到质膜, 继而在RALFs等配体和胞外信号的调节下, 动态调控RLKs的质膜定位与活性.拟南芥llg1和fer突变体的多样化发育与逆境应答表型, 暗示LLG与FER等RLKs之间的互作可能不是简单的开关, 而是存在精细的调节机制.对拟南芥LLG构象的解析及其与RALFs和FER互作位点的研究表明, LLG可能通过构象变化调节三者的互作关系, 从而精细调控胞外信号感知, 招募不同的RLKs完成胞内信号转导(
LORELEI-LIKE GPI- ANCHORED PROTEINS 2/3 regulate pollen tube growth as chaperones and coreceptors for ANXUR/BUPS receptor kinases in Arabidopsis.
5
2019
... 植物糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol-anchored protein, GPI-AP)定位于细胞膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与调节细胞生殖、生长以及免疫与盐碱胁迫应答等多种生物学过程(
... 植物花粉管快速生长进入胚珠的过程受到精细调控(
... 在感受胞外RALF4/19后, LLG2/3-ANX1/2-BUPS 1/2复合体通过与GDP-ROP1相互作用, 将其激活为GTP-ROP1, 进而激活下游花粉特异表达的NADPH氧化酶RbohH/J, 产生活性氧(ROS), 导致ROS在花粉管顶端积累, 调节花粉管生长, 降低花粉管的爆裂率(
... 的缺乏引起花粉管细胞壁成分改变, 甲酯化果胶质在花粉管顶端区域积累, 去甲酯化果胶质在花粉管亚顶端和shank区域积累, 花粉管中的胼胝质含量降低, 引起花粉管生长变缓(
... ;
The FERONIA receptor kinase maintains cell- wall integrity during salt stress through Ca 2+ signaling
7
2018
... 植物糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol-anchored protein, GPI-AP)定位于细胞膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与调节细胞生殖、生长以及免疫与盐碱胁迫应答等多种生物学过程(
... 盐离子干扰植物根部细胞壁结构, 导致细胞壁完整性降低, 抑制细胞极性生长, 引起细胞爆裂(
... ) (
... LLG1作为FER的共受体参与盐应答过程.拟南芥fer2和fer4突变体根的生长存在缺陷, 并且对Na+敏感, 但对甘露醇和山梨醇引起的渗透胁迫不敏感(
... NaCl即可导致其根部从伸长区至根尖大范围的细胞死亡.这表明LLG1和FER可能参与调控与SOS通路不同的盐胁迫应答途径(
... 在盐胁迫应答过程中, RALF22/23诱导FER通过胞吞途径内化, 从而负调控FER的功能(
... 在叶片中, FER与G蛋白β亚基(heterotrimeric G-protein β, AGB1)相互作用, 抑制气孔开放(
Plant malectin-like receptor kinases: from cell wall integrity to immunity and beyond
1
2018
... 植物花粉管快速生长进入胚珠的过程受到精细调控(
Negative regulation of defense responses in plants by a conserved MAPKK kinase
1
2001
... 当植物受到病原菌侵害时, FLS2和EFR迅速与其共受体BAK1 (brassinosteroid insensitive 1-associated receptor kinase 1)形成复合体, 激活下游MAPK级联信号通路, 调控防御基因的表达, 促进水杨酸(SA)等防御激素的积累, 提高植株的抗病性(
Arabidopsis pollen tube integrity and sperm release are regulated by RALF-mediated signaling
2
2017
... 植物花粉管快速生长进入胚珠的过程受到精细调控(
... ); 通过识别胚珠分泌的RALF34小肽, 控制花粉管爆裂并释放精细胞(
Pollen tube integrity regulation in flowering plants: insights from molecular assemblies on the pollen tube surface
5
2019
... 植物糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol-anchored protein, GPI-AP)定位于细胞膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与调节细胞生殖、生长以及免疫与盐碱胁迫应答等多种生物学过程(
... ;
... 植物花粉管快速生长进入胚珠的过程受到精细调控(
... ).花粉特异表达的LLG2/3可以与ANX和BUPS的exJM区互作, 调节其质膜定位, 缺失exJM区的ANX/BUPS滞留在胞内(
... ).RALF4和RALF19显著增强LLG2/3与ANX/BUPS的相互作用, RALF4的C端与LLG结合, 其N端(包括YISY motif)与LLG微弱互作, 而与ANX/BUPS强烈互作(
Three related receptor-like kinases are required for optimal cell elongation in Arabidopsis thaliana.
2
2009
... LLG1通过与FER的exJM结合, 协助FER定位于根部细胞质膜, 进而共同调控根与根毛生长(
... LLG1可能与FER共同参与脱落酸(ABA)、生长素、乙烯及油菜素内酯(BR)等激素信号转导过程, 从而调控根与根毛生长(
A cytoplasmic Ca 2+ functional assay for identifying and purifying endogenous cell signaling peptides in Arabidopsis seedlings: identification of AtRALF1 peptide.
1
2008
... RALF1通过诱导FER磷酸化将其激活(
A peptide hormone and its receptor protein kinase regulate plant cell expansion
4
2014
... LLG1与FER相互作用共同感受RALF1, 形成LLG1-RALF1-FER复合物并激活FER, 进而调控根生长.外源施加RALF1抑制野生型幼苗根的生长, 但并不影响llg1与fer-4突变体的根长.激活的FER与RopGEFs互作, 将RAC/ROP从与GDP结合的无活性状态转变为与GTP结合的活化状态.fer突变体与几种rac/rop突变体表型相似, 表明fer与rac/rop之间有调控关系(
... RALF1通过诱导FER磷酸化将其激活(
... , 抑制其质子转运能力(
... ,
Maternal ENODLs are required for pollen tube reception in Arabidopsis.
1
2016
... 植物糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol-anchored protein, GPI-AP)定位于细胞膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与调节细胞生殖、生长以及免疫与盐碱胁迫应答等多种生物学过程(
Arabidopsis RopGEF4 and RopGEF10 are important for FERONIA-mediated developmental but not environmental regulation of root hair growth
3
2013
... LLG1通过与FER的exJM结合, 协助FER定位于根部细胞质膜, 进而共同调控根与根毛生长(
... LLG1可能与FER共同参与脱落酸(ABA)、生长素、乙烯及油菜素内酯(BR)等激素信号转导过程, 从而调控根与根毛生长(
... LLG作为质膜定位的RLKs (如FER和FLS2)的分子伴侣, 负责将其从内质网运送到质膜, 继而在RALFs等配体和胞外信号的调节下, 动态调控RLKs的质膜定位与活性.拟南芥llg1和fer突变体的多样化发育与逆境应答表型, 暗示LLG与FER等RLKs之间的互作可能不是简单的开关, 而是存在精细的调节机制.对拟南芥LLG构象的解析及其与RALFs和FER互作位点的研究表明, LLG可能通过构象变化调节三者的互作关系, 从而精细调控胞外信号感知, 招募不同的RLKs完成胞内信号转导(
A fruitful journey: pollen tube navigation from germination to fertilization
1
2019
... 植物花粉管快速生长进入胚珠的过程受到精细调控(
The eight-cysteine motif, a versatile structure in plant proteins
1
2004
... LLG的氨基酸序列包括N端信号肽(含23个高度保守的氨基酸残基)、中央区域、C端构象可变区以及C端GPI锚(约20个氨基酸残基) (
Ca2+-activated reactive oxygen species production by Arabidopsis RbohH and RbohJ is essential for proper pollen tube tip growth
1
2014
... 在感受胞外RALF4/19后, LLG2/3-ANX1/2-BUPS 1/2复合体通过与GDP-ROP1相互作用, 将其激活为GTP-ROP1, 进而激活下游花粉特异表达的NADPH氧化酶RbohH/J, 产生活性氧(ROS), 导致ROS在花粉管顶端积累, 调节花粉管生长, 降低花粉管的爆裂率(
PAMP (pathogen-associated molecular pattern)-induced changes in plasma membrane compartmentalization reveal novel components of plant immunity
1
2010
... LLG1通过与FER的exJM结合, 协助FER定位于根部细胞质膜, 进而共同调控根与根毛生长(
Glycosylphosphatidylinositol-anchored proteins as chaperones and co-receptors for FERONIA receptor kinase signaling in Arabidopsis
15
2015
... 植物糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol-anchored protein, GPI-AP)定位于细胞膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与调节细胞生殖、生长以及免疫与盐碱胁迫应答等多种生物学过程(
... ), 调节RLK的功能, 参与花粉管生长与爆裂、根与根毛生长以及免疫与盐胁迫应答等过程(
... LLG的氨基酸序列包括N端信号肽(含23个高度保守的氨基酸残基)、中央区域、C端构象可变区以及C端GPI锚(约20个氨基酸残基) (
... 的表型相似, 如对RALF1的敏感性降低、表皮细胞形状改变以及根毛生长缺陷(
... LLG1通过与FER的exJM结合, 协助FER定位于根部细胞质膜, 进而共同调控根与根毛生长(
... 突变体有相似的根毛缺陷表型(
... ;
... ;
... LLG1与FER相互作用共同感受RALF1, 形成LLG1-RALF1-FER复合物并激活FER, 进而调控根生长.外源施加RALF1抑制野生型幼苗根的生长, 但并不影响llg1与fer-4突变体的根长.激活的FER与RopGEFs互作, 将RAC/ROP从与GDP结合的无活性状态转变为与GTP结合的活化状态.fer突变体与几种rac/rop突变体表型相似, 表明fer与rac/rop之间有调控关系(
... ;
... ), 导致细胞壁丧失完整性、细胞质外渗及细胞塌陷(
... ;
... LLG1可能与FER共同参与脱落酸(ABA)、生长素、乙烯及油菜素内酯(BR)等激素信号转导过程, 从而调控根与根毛生长(
... ;
... LLG1不同位点突变对其与FER、FLS2和EFR互作的调控存在差异.llg1-2具有明显的生长缺陷表型(
RLKs orchestrate the signaling in plant male-female interaction
1
2016
... 植物花粉管快速生长进入胚珠的过程受到精细调控(
Ligands switch model for pollen- tube integrity and burst
1
2018
... 植物花粉管快速生长进入胚珠的过程受到精细调控(
The FLS2-associated kinase BIK1 directly phosphorylates the NADPH oxidase RbohD to control plant immunity
1
2014
... LLG1不同位点突变对其与FER、FLS2和EFR互作的调控存在差异.llg1-2具有明显的生长缺陷表型(
Brittle Culm1, a COBRA-like protein, functions in cellulose assembly through binding cellulose microfibrils
1
2013
... 植物糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol-anchored protein, GPI-AP)定位于细胞膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与调节细胞生殖、生长以及免疫与盐碱胁迫应答等多种生物学过程(
The role of LORELEI in pollen tube reception at the interface of the synergid cell and pollen tube requires the modified eight- cysteine motif and the receptor-like kinase FERONIA
1
2016
... LLG的氨基酸序列包括N端信号肽(含23个高度保守的氨基酸残基)、中央区域、C端构象可变区以及C端GPI锚(约20个氨基酸残基) (
ROS regulation of polar growth in plant cells
1
2016
... 在感受胞外RALF4/19后, LLG2/3-ANX1/2-BUPS 1/2复合体通过与GDP-ROP1相互作用, 将其激活为GTP-ROP1, 进而激活下游花粉特异表达的NADPH氧化酶RbohH/J, 产生活性氧(ROS), 导致ROS在花粉管顶端积累, 调节花粉管生长, 降低花粉管的爆裂率(
RALF4/19 peptides interact with LRX proteins to control pollen tube growth in Arabidopsis
1
2017
... 植物花粉管快速生长进入胚珠的过程受到精细调控(
Oscillations in extracellular pH and reactive oxygen species modulate tip growth of Arabidopsis root hairs
1
2007
... LLG1与FER相互作用共同感受RALF1, 形成LLG1-RALF1-FER复合物并激活FER, 进而调控根生长.外源施加RALF1抑制野生型幼苗根的生长, 但并不影响llg1与fer-4突变体的根长.激活的FER与RopGEFs互作, 将RAC/ROP从与GDP结合的无活性状态转变为与GTP结合的活化状态.fer突变体与几种rac/rop突变体表型相似, 表明fer与rac/rop之间有调控关系(
Arabidopsis glycosylphosphatidylinositol-anchored protein LLG1 associates with and modulates FLS2 to regulate innate immunity
9
2017
... 植物糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol-anchored protein, GPI-AP)定位于细胞膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与调节细胞生殖、生长以及免疫与盐碱胁迫应答等多种生物学过程(
... LLG的氨基酸序列包括N端信号肽(含23个高度保守的氨基酸残基)、中央区域、C端构象可变区以及C端GPI锚(约20个氨基酸残基) (
... 在植物应答病原菌侵染过程中, 质膜模式识别受体(pattern recognition receptors, PRRs)参与激活免疫系统.多数植物PRRs是具有胞外配体识别结构域、跨膜结构域和胞质激酶结构域的类受体激酶, 如富含亮氨酸重复序列受体激酶FLS2 (flagellin sensing 2)和EFR (EF-Tu receptor).LLG1作为分子伴侣, 与EFR和FLS2的exJM区互作调节其质膜定位, 进而调控植物免疫应答过程(
... 当植物受到病原菌侵害时, FLS2和EFR迅速与其共受体BAK1 (brassinosteroid insensitive 1-associated receptor kinase 1)形成复合体, 激活下游MAPK级联信号通路, 调控防御基因的表达, 促进水杨酸(SA)等防御激素的积累, 提高植株的抗病性(
... ) (
... LLG1不同位点突变对其与FER、FLS2和EFR互作的调控存在差异.llg1-2具有明显的生长缺陷表型(
... ) (
... ) (
... 突变体中受flg22和elf18诱导的ROS水平低于野生型(
BR-SIGNALING KINASE 1 physically associates with FLAGELLIN SENSING 2 and regulates plant innate immunity in Arabidopsis
1
2013
... LLG1不同位点突变对其与FER、FLS2和EFR互作的调控存在差异.llg1-2具有明显的生长缺陷表型(
Structural basis for flg22-induced activation of the Arabidopsis FLS2-BAK1 immune complex
1
2013
... 当植物受到病原菌侵害时, FLS2和EFR迅速与其共受体BAK1 (brassinosteroid insensitive 1-associated receptor kinase 1)形成复合体, 激活下游MAPK级联信号通路, 调控防御基因的表达, 促进水杨酸(SA)等防御激素的积累, 提高植株的抗病性(
ROS in plant development
1
2010
... LLG1与FER相互作用共同感受RALF1, 形成LLG1-RALF1-FER复合物并激活FER, 进而调控根生长.外源施加RALF1抑制野生型幼苗根的生长, 但并不影响llg1与fer-4突变体的根长.激活的FER与RopGEFs互作, 将RAC/ROP从与GDP结合的无活性状态转变为与GTP结合的活化状态.fer突变体与几种rac/rop突变体表型相似, 表明fer与rac/rop之间有调控关系(
Mechanisms of RALF peptide perception by a heterotypic receptor complex
13
2019
... LLG的氨基酸序列包括N端信号肽(含23个高度保守的氨基酸残基)、中央区域、C端构象可变区以及C端GPI锚(约20个氨基酸残基) (
... 植物花粉管快速生长进入胚珠的过程受到精细调控(
... 复合体的晶体结构(
... 与RALF23和LLG2互作后, 其构象无明显变化(
... LLG1与FER相互作用共同感受RALF1, 形成LLG1-RALF1-FER复合物并激活FER, 进而调控根生长.外源施加RALF1抑制野生型幼苗根的生长, 但并不影响llg1与fer-4突变体的根长.激活的FER与RopGEFs互作, 将RAC/ROP从与GDP结合的无活性状态转变为与GTP结合的活化状态.fer突变体与几种rac/rop突变体表型相似, 表明fer与rac/rop之间有调控关系(
... LLG1通过与RALFs和FER互作形成三元复合物, 调控下游信号通路.三者的关键氨基酸位点可能影响其互作关系(
... 互作(
... 区域非常保守, 其中G123R突变会减弱LLG1与RALF23互作(
... 突变体发生的G114R替换也降低了LLG1与RALF23的体外互作, 减弱了RALF23对幼苗生长的抑制作用(
... ).但是, LLG1有些位点的突变(如N91A、T99R、A117Y和N118Y)并未影响其与RALF23和FER的互作(
... ).此外, RALF23关键位点突变(I6A、I6Y、L11Y和N14A)会影响其与LLG1的互作, 进而消除其对elf18诱导的ROS产生与幼苗生长的抑制作用(
... 背景下回补表达点突变的FER (G257A)和FER (N303Y), 幼苗生长的抑制明显受到RALF23诱导(
... LLG作为质膜定位的RLKs (如FER和FLS2)的分子伴侣, 负责将其从内质网运送到质膜, 继而在RALFs等配体和胞外信号的调节下, 动态调控RLKs的质膜定位与活性.拟南芥llg1和fer突变体的多样化发育与逆境应答表型, 暗示LLG与FER等RLKs之间的互作可能不是简单的开关, 而是存在精细的调节机制.对拟南芥LLG构象的解析及其与RALFs和FER互作位点的研究表明, LLG可能通过构象变化调节三者的互作关系, 从而精细调控胞外信号感知, 招募不同的RLKs完成胞内信号转导(
FERONIA phosphorylates E3 ubiquitin ligase ATL6 to modulate the stability of 14-3-3 proteins in response to the carbon/nitrogen ratio
1
2019
... LLG作为质膜定位的RLKs (如FER和FLS2)的分子伴侣, 负责将其从内质网运送到质膜, 继而在RALFs等配体和胞外信号的调节下, 动态调控RLKs的质膜定位与活性.拟南芥llg1和fer突变体的多样化发育与逆境应答表型, 暗示LLG与FER等RLKs之间的互作可能不是简单的开关, 而是存在精细的调节机制.对拟南芥LLG构象的解析及其与RALFs和FER互作位点的研究表明, LLG可能通过构象变化调节三者的互作关系, 从而精细调控胞外信号感知, 招募不同的RLKs完成胞内信号转导(
Receptor protein kinase FERO-NIA controls leaf starch accumulation by interacting with glyceraldehyde-3-phosphate dehydrogenase
1
2015
... LLG作为质膜定位的RLKs (如FER和FLS2)的分子伴侣, 负责将其从内质网运送到质膜, 继而在RALFs等配体和胞外信号的调节下, 动态调控RLKs的质膜定位与活性.拟南芥llg1和fer突变体的多样化发育与逆境应答表型, 暗示LLG与FER等RLKs之间的互作可能不是简单的开关, 而是存在精细的调节机制.对拟南芥LLG构象的解析及其与RALFs和FER互作位点的研究表明, LLG可能通过构象变化调节三者的互作关系, 从而精细调控胞外信号感知, 招募不同的RLKs完成胞内信号转导(
The Arabidopsis chaperone J3 regulates the plasma membrane H+-ATPase through interaction with the PKS5 kinase
1
2010
... 在叶片中, FER与G蛋白β亚基(heterotrimeric G-protein β, AGB1)相互作用, 抑制气孔开放(
Cellulose deficiency is enhanced on hyper accumulation of sucrose by a H+-coupled sucrose symporter
1
2016
... LLG作为质膜定位的RLKs (如FER和FLS2)的分子伴侣, 负责将其从内质网运送到质膜, 继而在RALFs等配体和胞外信号的调节下, 动态调控RLKs的质膜定位与活性.拟南芥llg1和fer突变体的多样化发育与逆境应答表型, 暗示LLG与FER等RLKs之间的互作可能不是简单的开关, 而是存在精细的调节机制.对拟南芥LLG构象的解析及其与RALFs和FER互作位点的研究表明, LLG可能通过构象变化调节三者的互作关系, 从而精细调控胞外信号感知, 招募不同的RLKs完成胞内信号转导(
BZR1 transcription factor regulates heat stress tolerance through FERONIA receptor-like kinase- mediated reactive oxygen species signaling in tomato
1
2018
... LLG作为质膜定位的RLKs (如FER和FLS2)的分子伴侣, 负责将其从内质网运送到质膜, 继而在RALFs等配体和胞外信号的调节下, 动态调控RLKs的质膜定位与活性.拟南芥llg1和fer突变体的多样化发育与逆境应答表型, 暗示LLG与FER等RLKs之间的互作可能不是简单的开关, 而是存在精细的调节机制.对拟南芥LLG构象的解析及其与RALFs和FER互作位点的研究表明, LLG可能通过构象变化调节三者的互作关系, 从而精细调控胞外信号感知, 招募不同的RLKs完成胞内信号转导(
FERONIA receptor kinase pathway suppresses abscisic acid signaling in Arabidopsis by activating ABI2 phosphatase
2
2012
... LLG1可能与FER共同参与脱落酸(ABA)、生长素、乙烯及油菜素内酯(BR)等激素信号转导过程, 从而调控根与根毛生长(
... ).当ABA存在时, FER通过与GEF1/4/10和GTP-ROP11互作, 激活A-type PP2C磷酸酶ABI2 (ABA insensitive 2)的活性, 抑制SnRK2活性, 从而抑制ABA信号通路, 同时抑制SLAC1通道和NADPH氧化酶介导的ROS产生, 负调控根生长(
Plant chloroplast stress response: insights from thiol redox proteomics
1
2020
... LLG作为质膜定位的RLKs (如FER和FLS2)的分子伴侣, 负责将其从内质网运送到质膜, 继而在RALFs等配体和胞外信号的调节下, 动态调控RLKs的质膜定位与活性.拟南芥llg1和fer突变体的多样化发育与逆境应答表型, 暗示LLG与FER等RLKs之间的互作可能不是简单的开关, 而是存在精细的调节机制.对拟南芥LLG构象的解析及其与RALFs和FER互作位点的研究表明, LLG可能通过构象变化调节三者的互作关系, 从而精细调控胞外信号感知, 招募不同的RLKs完成胞内信号转导(
Recent progress in synthetic and biological studies of GPI anchors and GPI-anchored proteins
1
2013
... 植物糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol-anchored protein, GPI-AP)定位于细胞膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与调节细胞生殖、生长以及免疫与盐碱胁迫应答等多种生物学过程(
The heterotrimeric G-protein β subunit, AGB1, plays multiple roles in the Arabidopsis salinity response
1
2015
... 在叶片中, FER与G蛋白β亚基(heterotrimeric G-protein β, AGB1)相互作用, 抑制气孔开放(
The G protein β-subunit, AGB1, interacts with FERONIA in RALF1- regulated stomatal movement
10
2018
... 植物糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol-anchored protein, GPI-AP)定位于细胞膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与调节细胞生殖、生长以及免疫与盐碱胁迫应答等多种生物学过程(
... 在叶片中, FER与G蛋白β亚基(heterotrimeric G-protein β, AGB1)相互作用, 抑制气孔开放(
... ;
... ).有研究表明, AGB1通路和FER通路协同作用, 参与盐胁迫应答, 且这种作用可能受到蒸腾条件的影响(
... 积累(
... 不能持续激活AHA (
... RALF1通过诱导FER磷酸化将其激活(
... 的表型, 这表明RALF1通过FER对根生长发挥抑制作用(
... 盐诱导质膜定位的NADPH氧化酶RbohD产生ROS, 并受到AGB1和FER的协同调控(
... 双突变体表现出比各自单突变体更严重的ROS失衡表型, 且这种ROS失衡在不同浓度和时间的盐处理以及不同器官(叶片和根)中存在差异(
A high-quality genome sequence of alkaligrass provides insights into halophyte stress tolerance
2
2020
... 植物糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol-anchored protein, GPI-AP)定位于细胞膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与调节细胞生殖、生长以及免疫与盐碱胁迫应答等多种生物学过程(
... 对盐生牧草小花碱茅(Puccinellia tenuiflora)盐碱应答转录组的分析表明, FER及其相关基因HERK1、NOTIA和MARIS, 以及FLS2及其互作蛋白BAK1、EFR、BIK1、SCD1、BSK1和GRP7协同作用, 调节G蛋白介导的Ca2+信号通路、ABA信号通路以及MAPK级联信号通路和ROS稳态, 从而调控盐碱应答过程(
Leucine-rich repeat extensin proteins regulate plant salt tolerance in Arabidopsis
3
2018
... 植物糖基磷脂酰肌醇锚定蛋白(glycosylphosphatidylinositol-anchored protein, GPI-AP)定位于细胞膜外表面, 作为CrRLK1L家族类受体激酶的分子伴侣, 参与调节细胞生殖、生长以及免疫与盐碱胁迫应答等多种生物学过程(
... 在盐胁迫应答过程中, RALF22/23诱导FER通过胞吞途径内化, 从而负调控FER的功能(
... 过表达植株都表现出相似的生长迟滞和盐敏感表型(
Peptide/receptor-like kinase-mediated signaling involved in male-female interactions
1
2019
... 植物花粉管快速生长进入胚珠的过程受到精细调控(
Glycosylphosphatidylinositol-anchored proteins: membrane organization and transport
1
2016
... LLG的氨基酸序列包括N端信号肽(含23个高度保守的氨基酸残基)、中央区域、C端构象可变区以及C端GPI锚(约20个氨基酸残基) (

{kind=link}
{kind=link}
{kind=link}
{kind=link}