 ,1,2,*
,1,2,*NLR and Its Regulation on Plant Disease Resistance
Chenghuizi Yang1,2, Xianyu Tang2, Wei Li1,2, Shitou Xia,1,2,*通讯作者:
责任编辑: 孙冬花
收稿日期:2019-10-23接受日期:2020-01-23网络出版日期:2020-07-01
| 基金资助: |
Corresponding authors:
Received:2019-10-23Accepted:2020-01-23Online:2020-07-01

摘要
关键词:
Abstract
Keywords:
PDF (914KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
杨程惠子, 唐先宇, 李威, 夏石头. NLR及其在植物抗病中的调控作用. 植物学报, 2020, 55(4): 497-504 doi:10.11983/CBB19207
Yang Chenghuizi, Tang Xianyu, Li Wei, Xia Shitou.
植物病害常造成大范围的粮食减产, 培育抗病品种是抵御作物病害最为经济且环保的手段之一。1994年, 植物核苷酸结合的富含亮氨酸重复蛋白(nucleotide-binding leucine-rich repeat, NLR)基因首次被克隆, 分别为烟草(Nicotiana tabacum)的N基因(Whitham et al., 1994)和拟南芥(Arabidopsis thaliana)的RPS2基因(Bent et al., 1994; Mindrinos et al., 1994); 其中, N对烟草花叶病毒具有特异抗性, 而RPS2赋予了植物对Pseudomonas syringae pv. glycinea (具有AvrRPS2)的特异抗性。过去20年中, 一系列植物R基因及其对应的Avr基因(无毒基因)被克隆。与动物NLRs不同, 植物NLRs可识别快速进化的效应子, 动物NLRs则更倾向于响应保守的病原相关分子模式(pathogen-associated molecule pattern, PAMPs)。
1 NLR的结构与特性
典型的NLR分为3部分, 即非保守的N端结构域、位于序列中间的核苷酸结合的寡聚结构域(nucleotide- binding and oligomerization domain, NOD)以及C端LRR (leucine-rich repeat)结构域。N端序列通常由TIR (toll/interleukin-1 receptor like)或CC (coiled-coil)组成。根据N端结构域的不同, NLR可分为TNL和CNL两大类。1.1 N端结构特性
与CC相比, TIR结构域更加保守, 且广泛存在于动、植物中(单子叶植物除外), 被认为与先天免疫系统的蛋白和蛋白互作相关。CC结构域的序列则较为多样化, 且常与其它结构域串联。例如, 番茄(Solanum lycopersicum)中的Sw-5b与SD序列相连(Peiró et al., 2014), 二者结合可在植物免疫中发挥更重要的作用。近年, 有关植物NLR蛋白N端的晶体结构解析表明, 其在整个免疫反应中具有重要作用。NLR蛋白可以发生自身二聚化或者多聚化。例如, 大麦(Hordeum vulgare) MLA10的CC结构域通过形成二聚体实现自激活, 阻止MLA10 CC结构域的二聚化可中断下游信号转导(El Kasmi and Nishimura, 2016)。与TNL被激活后才形成二聚体不同, 许多CNL在未被诱导的静息状态下, 同样处于二聚体形态, 如RPM1、RPS5、HRT、MLA10、Rp1-D和Sr50 (El Kasmi et al., 2017)。但与MLA10近源的Sr33及远源NLR Rx的CC端在静息状态时却仍为单体晶体结构(Casey et al., 2016; El Kasmi and Nishimura, 2016)。上述结果表明, N端二聚体的形成对于NLR的激活和下游的信号转导至关重要。此外, 植物NLR的N端也参与特异性识别病原菌效应子和引起细胞死亡的过程。动、植物体内NLR蛋白被病原体诱导后, TIR结构域的NAD+裂解活性被激活, 识别病原体并转化为细胞死亡信号, 从而引起免疫反应(Horsefield et al., 2019; Wan et al., 2019)
1.2 NB-ARC结构域特性
NB-ARC属于NLR蛋白保守结构域, 与哺乳动物的细胞凋亡蛋白酶激活因子Apaf-1 (apoptotic protease- activating factor-1)、植物R基因编码的抗病蛋白和秀丽线虫的细胞死亡蛋白CED-4 (caenorhabditis elegans death-4 protein)具有同源性(van der Biezen and Jones, 1998), 通常被认为具有“信号开关”功能。亚麻锈病抗病蛋白M的NB-ARC区域在结合ADP时处于静息状态, 结合ATP后才可被激活, 进而引发植物的超敏反应(Williams et al., 2011)。同为亚麻抗病蛋白的L6和L7, 因与ADP的结合程度不同诱发的免疫强度也不同。L7蛋白NB-ARC与ADP结合更紧密, 故其免疫反应稍弱于L6 (Bernoux et al., 2016)。近期, 我国科学家使用冷冻电镜技术揭示了拟南芥抗病蛋白ZAR1 (HopZ-activated resistance 1)的作用机制。他们发现, 当ZAR1-RKS1复合体与ADP结合时处于失活状态, 与被AvrAC诱导后尿苷酰化的PBL2UMP结合, 形成ZAR1-RKS1-PBL2UMP复合体时, 构象发生改变, 释放ADP而处于中间状态; 形成的复合体与dATP/ATP结合后诱发ZAR1寡聚结构域暴露, 导致ZAR1-RKS1-PBL2UMP形成轮状五聚体免疫抗病小体, 进而激活超敏反应和抗病性(Wang et al., 2019a, 2019b)。进一步的生化和功能分析表明, 该漏斗状结构使得激活状态的抗病小体与质膜(plasma membrane, PM)结合, 而这一功能对于细胞死亡和抗病性不可或缺, 暗示抗病小体很可能通过质膜穿孔或形成离子通道发挥作用(夏石头和李昕, 2019)。1.3 LRR结构域特性
LRR是整个蛋白结构中最为保守且简单的结构域, 处于NLR蛋白的C端, 由多个富含亮氨酸的重复序列组成。功能域互换实验表明, 即使蛋白的其它部分高度相似, 仅LRR不同, NLR蛋白识别的效应子也不同。例如, NAIP5 (neuronal apoptosis inhibitory protein 5)可通过其C末端的LRR结构域识别不同的细菌鞭毛蛋白(Yang et al., 2018)。但多数情况下, LRR并不直接与病原菌相互识别。例如, Prf的LRR区域就是通过与伴侣蛋白SGT1 (suppressor of the G2 allele of skp1)互作来感受病原菌, 进而引发一系列抗病反应(Kud et al., 2013)。也有研究表明, LRR不仅参与病原菌的相互识别, 还参与调节NLR蛋白的信号转导。拟南芥中RPS2、RPP1A和RPS5对信号转导具有抑制作用, 去除LRR结构域后表现出持续的激活状态, 引发抗病反应(Qi et al., 2012)。2 NLR的识别与信号转导
2.1 NLR对病原体的识别
植物通过对病原菌效应蛋白的特异识别感知病原体的入侵, 进而激活快速、精准的免疫反应。识别方式包括2类: 一类为相对简单的直接识别(图1A); 另一类为复杂的间接识别。直接识别即NLR蛋白直接与病原菌效应蛋白互作, 进行特异识别。例如, 拟南芥RBA1 (response to the bacterial type III effector protein HopBA1)与细菌III型分泌系统效应蛋白HopBA1直接识别, 以调节细胞死亡过程(Nishimura et al., 2017); 烟草Roq1 (recognition of XopQ 1)蛋白可特异识别黄单胞杆菌XopQ及假单孢杆菌HopQ1两种不同效应子, 且免疫共沉淀实验显示, Roq1可与二者互作(Schultink et al., 2017)。此外, 在直接识别模式下, 又衍生出微量识别模式(图1D)。例如, 番茄Sw-5b可直接识别NSm, 进而诱导免疫反应, 当其CC端串联SD序列时, SD序列先与微量NSm结合, 增强Sw-5b与NSm的结合能力, 从而引起更强的抗病反应(Li et al., 2019)。图1
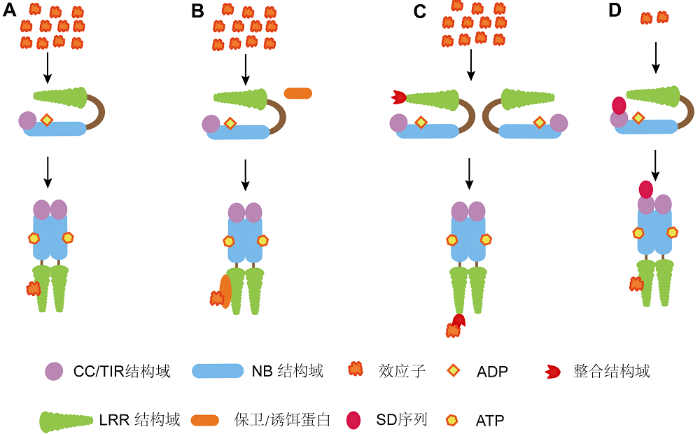 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1NLRs的识别和激活模型
(A) 直接识别模型; (B) 保卫/诱饵模型; (C) 整合激活模型; (D) 微量识别模型
Figure 1Models for recognition and activation of NLRs
(A) Direct interaction model; (B) Guard/decoy model; (C) Integrated activation model; (D) Microrecognition model
间接识别指NLR蛋白不直接与病原菌效应蛋白互作, 而是作用于某些目标蛋白, 通过感知目标蛋白(也称保卫蛋白)的变化引发免疫反应(图1B)。间接识别最初仅有保卫模式, 随后衍生出诱饵模式。在保卫模式下, 病原菌效应蛋白与保卫蛋白互作, 引起后者生理生化变化和结构改变, 被NLR蛋白感知后引发防御反应。例如, 拟南芥CRCK3 (calmodulin-binding receptor-like cytoplasmic kinase 3)作为磷酸化途径成员, 其磷酸化后被NLR蛋白SUMM2 (suppressor of mkk1 mkk2 2)识别, 进一步促进免疫反应发生(Zhang et al., 2017)。水稻(Oryza sativa)保卫蛋白APIP10 (AvrPiz-t and AvrPiz-t interacting protein 6)通过抑制NLR蛋白Piz-t维持植物的稳态。米曲霉入侵时, APIP10被效应蛋白AvrPiz-t降解, 从而解除对Piz-t的抑制, 造成Piz-t的大量积累, 继而引发ETI (effector-triggered immunity)反应(Park et al., 2016)。与保卫模式不同, 诱饵模式下的保卫蛋白会根据与其互作的NLR蛋白有无, 选择性地改变其特性。NLR存在时, 保卫蛋白以最佳状态增强与病原菌效应蛋白的互作, 以便被NLR蛋白识别; NLR不存在时, 保卫蛋白则会尽量避免与效应子结合, 以减轻其对植物的伤害。例如, 水稻中的Exo70作为诱饵蛋白, 被稻瘟菌效应蛋白Avr-Pii识别后形成大量复合物, 继而被Pi-i识别并激发ETI, 从而增强植株对病原的抗性(Fujisaki et al., 2015)。
NLR本身也可成为诱饵蛋白, 此类NLR含有特殊的整合结构域, 可对病原微生物进行特异性识别(图1C)。水稻中RNG4/RNG5形成复合物时可特异识别稻瘟菌Avr-CO39和Avr-Pia两种效应子, 进而引发下游的抗病反应, 但二者单独存在时, 却不诱发ETI (Ortiz et al., 2017)。拟南芥中, CHS3 (chilling sensitive 3)和CSA1 (constitutive shade-avoidance 1)是两个近源NLR, 其中CHS3具有识别功能, CSA1则提供抗性, 二者结合后才能激活完整的抗病反应(Xu et al., 2015)。高等植物中普遍存在类似两个NLR蛋白联合行使功能的抗病方式(Kroj et al., 2016)。Xie等(2019)研究发现, 有些NLR在植物体内以辅助蛋白的形式存在, 如最新克隆的水稻Pizh-2, 其单独存在时不具有抗病功能, 与Pizh-1结合后可增强后者对稻瘟菌的抗性。
2.2 NLR的激活与信号转导
NLR通常需要形成二聚体或者多聚体才被激活(El Kasmi and Nishimura, 2016; Wang et al., 2019a)。二聚体可以由NLR自身不同结构域之间结合形成(以N、SNC1和L6为代表的TNLs或以Prf、RPS5和MLA10为代表的CNLs都可自身结合形成二聚体而激活) (Qi and Innes, 2013); 也可是NLR两两配对形成, 如马铃薯(S. tuberosum), 其SINRC4a与LeEIX2相互结合形成二聚体而激活表达(Leibman-Markus et al., 2018)。这些NLRs在染色体上的定位通常相邻(大部分前后相连), 但DM1 (DANGEROUS MIX 1)和DM2d (DANGEROUS MIX 2d)较为特别, 虽然分别存在于不同的拟南芥生态型中, 但仍可在体外烟草中共表达后形成二聚体(Tran et al., 2017)。ZAR1则需要形成五聚体后才可以被完全激活(Wang et al., 2019b)。由此推测, 植物中可能还存在其它的NLR激活方式。许多CNLs (包括RPS5、RPS2和RPM1)定位于质膜(Qi and Innes, 2013), 其膜定位对于它们的活性是必需的, 可能与其相应效应子(或保卫/诱饵蛋白)的质膜定位相关。大部分NLRs被激活后亚细胞定位发生改变。不同定位的同一蛋白行使的功能可能不同。SNC1和Rx1均存在于细胞核和细胞质中, 但仅定位于细胞核中的SNC1才具活性, 仅定位于细胞质中的Rx1 (resistance to Potato virus X)才发挥作用(Slootweg et al., 2010; Xu et al., 2014a)。定位于细胞质的NLRs可能主要与病原菌的识别相关, 或与引发MAPK磷酸途径和诱导ROS积累相关; 细胞核中的NLRs则可能更多与转录因子相互作用。
2.3 NLR与转录因子互作调控下游免疫信号转导
研究表明, NLRs可通过与转录因子直接互作来调控下游免疫信号转导。例如, 大麦CNL MLA10通过其CC结构域与两种相互拮抗的转录因子WRKY1和MYB6 (v-myb avian myeloblastosis viral oncogene homolog 6)直接互作, 进而调控植物免疫(Chang et al., 2013)。而SNC1可与两种(如TPR1和bHLH84)及以上不同的转录因子相互作用。TPR1是一种转录抑制因子, 抑制多种已知免疫负调节因子的表达; tpr三重突变体中snc1的自身免疫被完全抑制(Zhu et al., 2010)。bHLH84是一种转录激活因子, 可作为免疫正调节因子与SNC1和RPS4互作。bHLH84过表达可增强免疫力, 敲除bHLH84及其2个近缘旁系同源蛋白则抑制RPS4和SNC1活性(Xu et al., 2014b)。此外, 烟草转录因子SPL6 (squamosa promoter-binding protein like6)激活后可与细胞核中的N蛋白相互作用, SPL6调节一部分防御基因, 为N蛋白下游ETI所必需。与此类似, 拟南芥同源基因AtSPL6为RPS4下游ETI所必需(Padmanabhan et al., 2013)。近期, Zhai等(2019)研究发现, 水稻转录因子RRM (RNA-recognition motif)可直接与NLR PigmR互作以激活其防御, 进而特异抵抗稻瘟菌的入侵。3 NLR的调节方式
3.1 表观遗传调控
植物具备多种表观遗传调控机制, 主要包括染色质重塑、DNA甲基化、组蛋白修饰和非编码RNA调控4类。染色质重塑因子SPLAYED (在筛选MUSE过程中得到)属于SWI/SNF家族, 其通过影响SNC1转录抑制由SNC1介导的免疫反应(Johnson et al., 2015)。nrpc7-1 (RNA聚合酶III的亚基缺失突变体)也在MUSE的正向筛选过程中得到, 该突变在增强植物抗性的同时引起了一系列表型缺陷, 原因可能是突变体在小RNA水平发生了修饰(Johnson et al., 2016)。染色质重塑因子CHR5 (chromatin-remodeling factor 5) (与组蛋白单泛素酶HUB1 (HISTONE MONOUBIQUITINATION 1)互作)上调细胞内SNC1的表达, 并可与另一种染色质重塑基因DDM1产生拮抗(Zou et al., 2017)。此外, DNA甲基化也调节SNC1的转录。Espinas等(2016)研究发现, mos1和ddm1均负调控胞嘧啶甲基化, 但二者在snc1调节中相互拮抗。通常情况下, DNA甲基化缺陷导致防御反应增强, DNA甲基化程度上升则增强易感性。有研究表明, 在转录因子RPP7的第1个内含子中插入反转录转座子COPIAR7会导致转录提前终止, 对RPP7等转录因子的调节可影响组蛋白甲基化, 并且各种组蛋白甲基化突变体中RPP7的表达也受到影响(Tsuchiya and Eulgem, 2013; Le et al., 2015)。与此类似, SNC1以及RPP4的表达依赖于组蛋白赖氨酸甲基转移酶ATXR7 (Arabidopsis trithorax-related 7)及其相关蛋白MOS9 (modifier of snc1,9) (Xia et al., 2013)。Halter和Navarro (2015)研究表明, miRNAs的产生对植物的抗病过程有显著影响, RNA的调节与植物免疫之间关系密切。NLR超量表达会对植物的生长发育造成不利影响, miRNAs通常作为NLR的负调控因子维持其在植物体内的稳态。从进化角度看, miRNAs高度靶向NLR保守序列, 以此减少其在植物体内积累造成的影响(Zhang et al., 2016)。
番茄miR482/2118超级家族与拟南芥miR472- RDR6为同源microRNA, 它们通过RNA诱导沉默方式在转录后水平调节多个CNLs (Boccara et al., 2014)。miR482/2118家族靶向调控NLR在番茄体内的积累, 从而调控其对卵菌和细菌病原体的抗性(Canto-Pastor et al., 2019)。miR9863家族通过特异调节大麦CNL MLA进而调控大麦的免疫反应及细胞死亡信号转导(Liu et al., 2014)。烟草miR6019以及miR6020靶向调节TNL N, 影响N蛋白在烟草中的表达及其介导的抗烟草花叶病毒反应(Deng et al., 2018)。大豆(Glycine max) miR1510a/b靶向调节特异性防御大豆疫霉所需的TNL, 但在大豆疫霉菌感染期间其效应被抑制(Liu et al., 2014)。gma-miR151则靶向调节大豆中NB-LRR免疫受体基因Glyma.16G135500, 在调节对大豆疫霉的抗性中起关键作用(Cui et al., 2017)。
3.2 可变剪接调控
可变剪接在调控NLR (特别是TLR)的过程中起重要作用, 其通过产生异常转录本/蛋白(或通过引发NMD (nonsense-mediated decay))的方式行使功能。转录组分析显示, 可变剪接广泛存在于具有多个内含子的NLR (特别是TNL)中。Xu等(2012)在MOS筛选过程中鉴定出多个突变体, 这些突变体大多属于MOS4相关复合物(MAC), 可调节NLR的拼接。若SNC1和RPS4的可变剪接发生改变, 将影响植物的基础免疫抗性(Xu et al., 2012)。但需要指出的是, MAC相关蛋白MOS2、CDC5和PRL1也参与miRNA的生物合成, 表明这两种系统间可能存在某种联系(Zhang et al., 2014)。与此不同的是, 参与miRNA剪接的小核糖核蛋白所需的甲基体复合物的突变可增强植物的免疫力, 这与其它剪接突变体的表型相反(Huang et al., 2016a)。Zhang等(2014)研究发现, NMD因子突变体表现出依赖于PAD4 (pHYTOALEXIN DEFICIENT4)和EDS1 (enhanced disease susceptibility 1)的自身免疫, 表明NMD与可变剪接密切配合, 在转录后水平调节NLR, 防止自身免疫并调节免疫应答。3.3 翻译后修饰
翻译后修饰主要包括泛素化、磷酸化和糖基化等几类, 其中泛素化修饰的应用最为广泛, 可以在蛋白水平调节植物的免疫。Huang等(2014, 2016b)在MUSE突变体筛选过程中, 得到拟南芥唯一的与E4酶和2个E3酶相关的TRAF (TNF receptor associated factor)蛋白(MUSE13以及MUSE14)。Gou等(2012)研究发现, SNC1和RPS2通过F-Box E3 SCFCPR1经由泛素-蛋白酶体途径直接调节植物的免疫反应。Wang等(2016)则发现RING型E3 MIR1调节大麦中的多种MLA NLRs; 其它E3s靶向非NLR免疫调节因子或影响对效应子的识别(Copeland et al., 2016a, 2016b; Tong et al., 2017); 免疫正调节因子E3泛素连接酶SAUL1 (senescence-associated E3 ubiquitin ligase 1)由TNL SOC3保卫(Tong et al., 2017)。已有研究表明, RAR1-SGT1b-HSP90伴侣复合体各组分对由各种NLR介导的ETI都很重要(例如, RpiBLB2信号转导需要SGT1 (suppressor of the G2 allele of skp1), 但无须HSP90 (heat shock protein 90)或RAR1 (required for Mla12 resistance) (Oh et al., 2014)), 但作用机制相当复杂, 尚需要深入探究(Kadota and Shirasu, 2012)。
4 总结与展望
NLR作为植物抗病蛋白中最重要的一类, 自发现之日起便受到研究者的广泛关注。随着NLR基因的不断被克隆, 相关研究也在不断深入。冷冻电镜技术的应用, 尽管可使我们更加直观地观察到NLR的结构组成及激活方式, 但对其整个信号通路目前仍不完全清楚。此外, CRISPR技术的日渐成熟为培育抗病新品种提供了新思路。因此, 在未来可预见的一段时间内, 对NLR的探究仍然是植物抗性研究领域的热点。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
URLPMID:26744216 [本文引用: 1]
Arabidopsis miR472- RDR6 silencing pathway modulates PAMP- and effector- triggered immunity through the post-transcriptional control of disease resistance genes
[本文引用: 1]
URLPMID:30679269 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:27340941 [本文引用: 1]
[本文引用: 1]
URLPMID:29293695 [本文引用: 1]
URLPMID:28808003 [本文引用: 1]
DOI:10.1073/pnas.1615933113URLPMID:27803318 [本文引用: 3]
URLPMID:27563304 [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URLPMID:31439792 [本文引用: 1]
URLPMID:27756575 [本文引用: 1]
URLPMID:26867179 [本文引用: 1]
[本文引用: 1]
URLPMID:26063389 [本文引用: 1]
DOI:10.1093/jxb/erw020URLPMID:26865731 [本文引用: 1]

Plants employ five DNA-dependent RNA polymerases (Pols) in transcription. One of these polymerases, Pol III, has previously been reported to transcribe 5S rRNA, tRNAs, and a number of small RNAs. However, in-depth functional analysis is complicated by the fact that knockout mutations in Pol subunits are typically lethal. Here, we report the characterization of the first known viable Pol III subunit mutant,nrpc7-1 This mutant was originally isolated from a forward genetic screen designed to identify enhancers of the autoimmune mutantsnc1, which contains a gain-of-function mutation in a nucleotide-binding leucine-rich repeat (NLR) immune receptor-encoding gene. Thenrpc7-1mutation occurs in an intron-exon splice site and results in intron retention in someNRPC7transcripts. There is a global disruption in RNA equilibrium innrpc7-1, exemplified by the altered expression of a number of RNA molecules, some of which are not reported to be transcribed by Pol III. There are developmental defects associated with the mutation, as homozygous mutant plants are dwarf, have stunted roots and siliques, and possess serrated leaves. These defects are possibly due to altered small RNA stability or activity. Additionally, thenrpc7-1mutation confers anNLR-specific alternative splicing defect that correlates with enhanced disease resistance, highlighting the importance of alternative splicing in regulating NLR activity. Altogether, these results reveal novel roles for Pol III in maintaining RNA homeostasis, adjusting the expression of a diverse suite of genes, and indirectly modulating gene splicing. Future analyses using thenrpc7-1mutant will be instrumental in examining other unknown Pol III functions.
[本文引用: 1]
DOI:10.1111/nph.13869URLPMID:26848538 [本文引用: 1]
Plant immune receptors of the class of nucleotide-binding and leucine-rich repeat domain (NLR) proteins can contain additional domains besides canonical NB-ARC (nucleotide-binding adaptor shared by APAF-1, R proteins, and CED-4 (NB-ARC)) and leucine-rich repeat (LRR) domains. Recent research suggests that these additional domains act as integrated decoys recognizing effectors from pathogens. Proteins homologous to integrated decoys are suspected to be effector targets and involved in disease or resistance. Here, we scrutinized 31 entire plant genomes to identify putative integrated decoy domains in NLR proteins using the Interpro search. The involvement of the Zinc Finger-BED type (ZBED) protein containing a putative decoy domain, called BED, in rice (Oryza sativa) resistance was investigated by evaluating susceptibility to the blast fungus Magnaporthe oryzae in rice over-expression and knock-out mutants. This analysis showed that all plants tested had integrated various atypical protein domains into their NLR proteins (on average 3.5% of all NLR proteins). We also demonstrated that modifying the expression of the ZBED gene modified disease susceptibility. This study suggests that integration of decoy domains in NLR immune receptors is widespread and frequent in plants. The integrated decoy model is therefore a powerful concept to identify new proteins involved in disease resistance. Further in-depth examination of additional domains in NLR proteins promises to unravel many new proteins of the plant immune system.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/pce.13347URLPMID:29790585 [本文引用: 1]
Plant recognition and defence against pathogens employs a two-tiered perception system. Surface-localized pattern recognition receptors (PRRs) act to recognize microbial features, whereas intracellular nucleotide-binding leucine-rich repeat receptors (NLRs) directly or indirectly recognize pathogen effectors inside host cells. Employing the tomato PRR LeEIX2/EIX model system, we explored the molecular mechanism of signalling pathways. We identified an NLR that can associate with LeEIX2, termed SlNRC4a (NB-LRR required for hypersensitive response-associated cell death-4). Co-immunoprecipitation demonstrates that SlNRC4a is able to associate with different PRRs. Physiological assays with specific elicitors revealed that SlNRC4a generally alters PRR-mediated responses. SlNRC4a overexpression enhances defence responses, whereas silencing SlNRC4 reduces plant immunity. Moreover, the coiled-coil domain of SlNRC4a is able to associate with LeEIX2 and is sufficient to enhance responses upon EIX perception. On the basis of these findings, we propose that SlNRC4a acts as a noncanonical positive regulator of immunity mediated by diverse PRRs. Thus, SlNRC4a could link both intracellular and extracellular immune perceptions.
URLPMID:30639751 [本文引用: 1]
[本文引用: 2]
URLPMID:7923358 [本文引用: 1]
URLPMID:28137883 [本文引用: 1]
URLPMID:25289011 [本文引用: 1]
URLPMID:28087830 [本文引用: 1]
URLPMID:23516366 [本文引用: 1]
DOI:10.1371/journal.ppat.1005529URLPMID:27031246 [本文引用: 1]
Although nucleotide-binding domain, leucine-rich repeat (NLR) proteins are the major immune receptors in plants, the mechanism that controls their activation and immune signaling remains elusive. Here, we report that the avirulence effector AvrPiz-t from Magnaporthe oryzae targets the rice E3 ligase APIP10 for degradation, but that APIP10, in return, ubiquitinates AvrPiz-t and thereby causes its degradation. Silencing of APIP10 in the non-Piz-t background compromises the basal defense against M. oryzae. Conversely, silencing of APIP10 in the Piz-t background causes cell death, significant accumulation of Piz-t, and enhanced resistance to M. oryzae, suggesting that APIP10 is a negative regulator of Piz-t. We show that APIP10 promotes degradation of Piz-t via the 26S proteasome system. Furthermore, we demonstrate that AvrPiz-t stabilizes Piz-t during M. oryzae infection. Together, our results show that APIP10 is a novel E3 ligase that functionally connects the fungal effector AvrPiz-t to its NLR receptor Piz-t in rice.
URLPMID:24690181 [本文引用: 1]
DOI:10.1104/pp.112.194035URLPMID:22331412 [本文引用: 1]
The Arabidopsis (Arabidopsis thaliana) RESISTANCE TO PSEUDOMONAS SYRINGAE5 (RPS5) disease resistance protein mediates recognition of the Pseudomonas syringae effector protein AvrPphB. RPS5 belongs to the coiled-coil-nucleotide-binding site-leucine-rich repeat (CC-NBS-LRR) family and is activated by AvrPphB-mediated cleavage of the protein kinase PBS1. Here, we present a structure-function analysis of the CC and LRR domains of RPS5 using transient expression assays in Nicotiana benthamiana. We found that substituting the CC domain of RPS2 for the RPS5 CC domain did not alter RPS5 specificity and only moderately reduced its ability to activate programmed cell death, suggesting that the CC domain does not play a direct role in the recognition of PBS1 cleavage. Analysis of an RPS5-super Yellow Fluorescent Protein fusion revealed that RPS5 localizes to the plasma membrane (PM). Alanine substitutions of predicted myristoylation (glycine-2) and palmitoylation (cysteine-4) residues affected RPS5 PM localization, protein stability, and function in an additive manner, indicating that PM localization is essential to RPS5 function. The first 20 amino acids of RPS5 were sufficient for directing super Yellow Fluorescent Protein to the PM. C-terminal truncations of RPS5 revealed that the first four LRR repeats are sufficient for inhibiting RPS5 autoactivation; however, the complete LRR domain was required for the recognition of PBS1 cleavage. Substitution of the RPS2 LRR domain resulted in the autoactivation of RPS5, indicating that the LRR domain must coevolve with the NBS domain. We conclude that the RPS5 LRR domain functions to suppress RPS5 activation in the absence of PBS1 cleavage and promotes RPS5 activation in its presence.
URLPMID:24155748 [本文引用: 2]
DOI:10.1111/tpj.13715URLPMID:28891100 [本文引用: 1]
Xanthomonas spp. are phytopathogenic bacteria that can cause disease on a wide variety of plant species resulting in significant impacts on crop yields. Limited genetic resistance is available in most crop species and current control methods are often inadequate, particularly when environmental conditions favor disease. The plant Nicotiana benthamiana has been shown to be resistant to Xanthomonas and Pseudomonas due to an immune response triggered by the bacterial effector proteins XopQ and HopQ1, respectively. We used a reverse genetic screen to identify Recognition of XopQ 1 (Roq1), a nucleotide-binding leucine-rich repeat (NLR) protein with a Toll-like interleukin-1 receptor (TIR) domain, which mediates XopQ recognition in N. benthamiana. Roq1 orthologs appear to be present only in the Nicotiana genus. Expression of Roq1 was found to be sufficient for XopQ recognition in both the closely-related Nicotiana sylvestris and the distantly-related beet plant (Beta vulgaris). Roq1 was found to co-immunoprecipitate with XopQ, suggesting a physical association between the two proteins. Roq1 is able to recognize XopQ alleles from various Xanthomonas species, as well as HopQ1 from Pseudomonas, demonstrating widespread potential application in protecting crop plants from these pathogens.
[本文引用: 1]
URLPMID:28691210 [本文引用: 2]
URLPMID:28416116 [本文引用: 1]
URLPMID:23940361 [本文引用: 1]
DOI:10.1016/s0968-0004(98)01311-5URLPMID:9868361 [本文引用: 1]
URLPMID:31439793 [本文引用: 1]
URLPMID:30948527 [本文引用: 2]
[本文引用: 2]
DOI:10.1104/pp.16.01520URLPMID:27780896 [本文引用: 1]
Following the detection of pathogen cognate effectors, plant Nod-like receptors (NLRs) trigger isolate-specific immunity that is generally associated with cell death. The regulation of NLR stability is important to ensure effective immunity. In barley (Hordeum vulgare), the allelic Mildew locus A (MLA) receptors mediate isolate-specific disease resistance against powdery mildew fungus (Blumeria graminis f. sp. hordei). Currently, how MLA stability is controlled remains unknown. Here, we identified an MLA-interacting RING-type E3 ligase, MIR1, that interacts with several MLAs. We showed that the carboxyl-terminal TPR domain of MIR1 mediates the interaction with the coiled-coil domain-containing region of functional MLAs, such as MLA1, MLA6, and MLA10, but not with that of the nonfunctional MLA18-1. MIR1 can ubiquitinate the amino-terminal region of MLAs in vitro and promotes the proteasomal degradation of MLAs in vitro and in planta. Both proteasome inhibitor treatment and virus-induced gene silencing-mediated MIR1 silencing significantly increased MLA abundance in barley transgenic lines. Furthermore, overexpression of MIR1 specifically compromised MLA-mediated disease resistance in barley, while coexpression of MIR1 and MLA10 attenuated MLA10-induced cell death signaling in Nicotiana benthamiana Together, our data reveal a mechanism for the control of the stability of MLA immune receptors and for the attenuation of MLA-triggered defense signaling by a RING-type E3 ligase via the ubiquitin proteasome system.
DOI:10.1016/0092-8674(94)90283-6URLPMID:7923359 [本文引用: 1]
The products of plant disease resistance genes are postulated to recognize invading pathogens and rapidly trigger host defense responses. Here we describe isolation of the resistance gene N of tobacco that mediates resistance to the viral pathogen tobacco mosaic virus (TMV). The N gene was isolated by transposon tagging using the maize Activator transposon. A genomic DNA fragment containing the N gene conferred TMV resistance to TMV susceptible tobacco. Sequence analysis of the N gene shows that it encodes a protein of 131.4 kDa with an amino-terminal domain similar to that of the cytoplasmic domain of the Drosophila Toll protein and the interleukin-1 receptor (IL-1R) in mammals, a nucleotide-binding site (NBS), and 14 [corrected] imperfect leucine-rich repeats (LRR). The sequence similarity of N, Toll, and IL-1R suggests that N mediates rapid gene induction and TMV resistance through a Toll-IL-1-like pathway.
URLPMID:21539434 [本文引用: 1]
DOI:10.1104/pp.113.214551URLPMID:23690534 [本文引用: 1]
Plant nucleotide-binding leucine-rich repeat (NB-LRR) proteins serve as intracellular sensors to detect pathogen effectors and trigger immune responses. Transcription of the NB-LRR-encoding Resistance (R) genes needs to be tightly controlled to avoid inappropriate defense activation. How the expression of the NB-LRR R genes is regulated is poorly understood. The Arabidopsis (Arabidopsis thaliana) suppressor of npr1-1, constitutive 1 (snc1) mutant carries a gain-of-function mutation in a Toll/Interleukin1 receptor-like (TIR)-NB-LRR-encoding gene, resulting in the constitutive activation of plant defense responses. A snc1 suppressor screen identified modifier of snc1,9 (mos9), which partially suppresses the autoimmune phenotypes of snc1. Positional cloning revealed that MOS9 encodes a plant-specific protein of unknown function. Expression analysis showed that MOS9 is required for the full expression of TIR-NB-LRR protein-encoding RECOGNITION OF PERONOSPORA PARASITICA 4 (RPP4) and SNC1, both of which reside in the RPP4 cluster. Coimmunoprecipitation and mass spectrometry analyses revealed that MOS9 associates with the Set1 class lysine 4 of histone 3 (H3K4) methyltransferase Arabidopsis Trithorax-Related7 (ATXR7). Like MOS9, ATXR7 is also required for the full expression of SNC1 and the autoimmune phenotypes in the snc1 mutant. In atxr7 mutant plants, the expression of RPP4 is similarly reduced, and resistance against Hyaloperonospora arabidopsidis Emwa1 is compromised. Consistent with the attenuated expression of SNC1 and RPP4, trimethylated H3K4 marks are reduced around the promoters of SNC1 and RPP4 in mos9 plants. Our data suggest that MOS9 functions together with ATXR7 to regulate the expression of SNC1 and RPP4 through H3K4 methylation, which plays an important role in fine-tuning their transcription levels and functions in plant defense.
DOI:10.1098/rstb.2018.0308URLPMID:30967012 [本文引用: 1]
Rice blast caused by Magnaporthe oryzae is the most destructive fungal disease in crops, greatly threatening rice production and food security worldwide. The identification and utilization of broad-spectrum resistance genes are considered to be the most economic and effective method to control the disease. In the past decade, many blast resistance ( R) genes have been identified, which mainly encode nucleotide-binding leucine-rich repeat (NLR) receptor family and confer limited race-specific resistance to the fungal pathogen. Resistance genes conferring broad-spectrum blast resistance are still largely lacking. In this study, we carried out a map-based cloning of the new blast R locus Pizh in variety ZH11. A bacterial artificial chromosome (BAC) clone of 165 kb spanning the Pizh locus was sequenced and identified 9 NLR genes, among which only Pizh-1 and Pizh-2 were expressed. Genetic complementation experiments indicated that Pizh-1 but not Pizh-2 alone could confer blast resistance. Intriguingly, both mutations on Pizh-1 and Pizh-2 by CRISPR-Cas9 abolished the Pizh-mediated resistance. We also observed that Pizh-1-mediated resistance was partially dependent on Pizh-2. Pizh-1 and Pizh-2 form a complex of NLRs through direct interaction. This suggests that Pizh-1 may function as the executor NLR and Pizh-2 as a 'helper' NLR that shares functional redundancy with other NLRs. Our current study provides not only a good tool for rice disease resistance breeding but also deep insight into NLR association and function in plant immunity. This article is part of the theme issue 'Biotic signalling sheds light on smart pest management'.
URLPMID:25237053 [本文引用: 1]
DOI:10.1371/journal.ppat.1004312URLPMID:25144198 [本文引用: 1]
In plants and animals, nucleotide-binding and leucine-rich repeat domain containing (NLR) immune receptors are utilized to detect the presence or activities of pathogen-derived molecules. However, the mechanisms by which NLR proteins induce defense responses remain unclear. Here, we report the characterization of one basic Helix-loop-Helix (bHLH) type transcription factor (TF), bHLH84, identified from a reverse genetic screen. It functions as a transcriptional activator that enhances the autoimmunity of NLR mutant snc1 (suppressor of npr1-1, constitutive 1) and confers enhanced immunity in wild-type backgrounds when overexpressed. Simultaneously knocking out three closely related bHLH paralogs attenuates RPS4-mediated immunity and partially suppresses the autoimmune phenotypes of snc1, while overexpression of the other two close paralogs also renders strong autoimmunity, suggesting functional redundancy in the gene family. Intriguingly, the autoimmunity conferred by bHLH84 overexpression can be largely suppressed by the loss-of-function snc1-r1 mutation, suggesting that SNC1 is required for its proper function. In planta co-immunoprecipitation revealed interactions between not only bHLH84 and SNC1, but also bHLH84 and RPS4, indicating that bHLH84 associates with these NLRs. Together with previous finding that SNC1 associates with repressor TPR1 to repress negative regulators, we hypothesize that nuclear NLR proteins may interact with both transcriptional repressors and activators during immune responses, enabling potentially faster and more robust transcriptional reprogramming upon pathogen recognition.
[本文引用: 2]
DOI:10.1038/srep08792URLPMID:25740259 [本文引用: 1]
Plant innate immunity depends on the function of a large number of intracellular immune receptor proteins, the majority of which are structurally similar to mammalian nucleotide-binding oligomerization domain (NOD)-like receptor (NLR) proteins. CHILLING SENSITIVE 3 (CHS3) encodes an atypical Toll/Interleukin 1 Receptor (TIR)-type NLR protein with an additional Lin-11, Isl-1 and Mec-3 (LIM) domain at its C-terminus. The gain-of-function mutant allele chs3-2D exhibits severe dwarfism and constitutively activated defense responses, including enhanced resistance to virulent pathogens, high defence marker gene expression, and salicylic acid accumulation. To search for novel regulators involved in CHS3-mediated immune signaling, we conducted suppressor screens in the chs3-2D and chs3-2D pad4-1 genetic backgrounds. Alleles of sag101 and eds1-90 were isolated as complete suppressors of chs3-2D, and alleles of sgt1b were isolated as partial suppressors of chs3-2D pad4-1. These mutants suggest that SAG101, EDS1-90, and SGT1b are all positive regulators of CHS3-mediated defense signaling. Additionally, the TIR-type NLR-encoding CSA1 locus located genomically adjacent to CHS3 was found to be fully required for chs3-2D-mediated autoimmunity. CSA1 is located 3.9 kb upstream of CHS3 and is transcribed in the opposite direction. Altogether, these data illustrate the distinct genetic requirements for CHS3-mediated defense signaling.
[本文引用: 1]
DOI:10.1016/j.molcel.2019.03.013URLPMID:30975460 [本文引用: 1]
Nucleotide-binding site leucine-rich repeat (NLR) receptors perceive pathogen effectors and trigger plant immunity. However, the mechanisms underlying NLR-triggered defense responses remain obscure. The recently discovered Pigm locus in rice encodes a cluster of NLRs, including PigmR, which confers broad-spectrum resistance to blast fungus. Here, we identify PIBP1 (PigmR-INTERACTING and BLAST RESISTANCE PROTEIN 1), an RRM (RNA-recognition motif) protein that specifically interacts with PigmR and other similar NLRs to trigger blast resistance. PigmR-promoted nuclear accumulation of PIBP1 ensures full blast resistance. We find that PIBP1 and a homolog, Os06 g02240, bind DNA and function as unconventional transcription factors at the promoters of the defense genes OsWAK14 and OsPAL1, activating their expression. Knockout of PIBP1 and Os06 g02240 greatly attenuated blast resistance. Collectively, our study discovers previously unappreciated RRM transcription factors that directly interact with NLRs to activate plant defense, establishing a direct link between transcriptional activation of immune responses with NLR-mediated pathogen perception.
[本文引用: 2]
URLPMID:27512116 [本文引用: 1]
DOI:10.15252/embr.201642704URLPMID:27986791 [本文引用: 1]
MAP kinase signaling is an integral part of plant immunity. Disruption of the MEKK1-MKK1/2-MPK4 kinase cascade results in constitutive immune responses mediated by the NLR protein SUMM2, but the molecular mechanism is so far poorly characterized. Here, we report that SUMM2 monitors a substrate protein of MPK4, CALMODULIN-BINDING RECEPTOR-LIKE CYTOPLASMIC KINASE 3 (CRCK3). Similar to SUMM2, CRCK3 was isolated from a suppressor screen of mkk1 mkk2 and is required for the autoimmunity phenotypes in mekk1, mkk1 mkk2, and mpk4 mutants. In wild-type plants, CRCK3 is mostly phosphorylated. MPK4 interacts with CRCK3 and can phosphorylate CRCK3 in vitro In mpk4 mutant plants, phosphorylation of CRCK3 is substantially reduced, suggesting that MPK4 phosphorylates CRCK3 in vivo Further, CRCK3 associates with SUMM2 in planta, suggesting SUMM2 senses the disruption of the MEKK1-MKK1/2-MPK4 kinase cascade through CRCK3. Our study suggests that a MAP kinase substrate is used as a guardee or decoy for monitoring the integrity of MAP kinase signaling.
URLPMID:20647385 [本文引用: 1]
Arabidopsis chromatin-remodeling factor CHR5 regulates plant immune responses and nucleosome occupancy
DOI:10.1093/pcp/pcx155URLPMID:29048607 [本文引用: 1]
ATP-dependent chromatin-remodeling factors use the energy of ATP hydrolysis to alter the structure of chromatin and are important regulators of eukaryotic gene expression. One such factor encoded by CHR5 (Chromatin-Remodeling Factor 5) in Arabidopsis (Arabidopsis thaliana) was previously found to be involved in regulation of growth and development. Here we show that CHR5 is required for the up-regulation of the intracellular immune receptor gene SNC1 (SUPPRESSOR OF npr1-1, CONSTITUTIVE1) and consequently the autoimmunity induced by SNC1 up-regulation. CHR5 functions antagonistically with another chromatin-remodeling gene DDM1 (DECREASED DNA METHYLATION 1) and independently with a histone mono-ubiquitinase HUB1 (HISTONE MONOUBIQUITINATION 1) in SNC1 regulation. In addition, CHR5 is a positive regulator of SNC1-independent plant immunity against the bacterial pathogen Pseudomonas syringae. Furthermore, the chr5 mutant has increased nucleosome occupancy in the promoter region relative to the gene body region at the whole-genome level, suggesting a global role for CHR5 in remodeling nucleosome occupancy. Our study thus establishes CHR5 as a positive regulator of plant immune responses including the expression of SNC1 and reveals a role for CHR5 in nucleosome occupancy which probably impacts gene expression genome wide.
开启防御之门: 植物抗病小体
1
2019
... NB-ARC属于NLR蛋白保守结构域, 与哺乳动物的细胞凋亡蛋白酶激活因子Apaf-1 (apoptotic protease- activating factor-1)、植物R基因编码的抗病蛋白和秀丽线虫的细胞死亡蛋白CED-4 (caenorhabditis elegans death-4 protein)具有同源性(
RPS2 of Arabidopsis thaliana: a leucine-rich repeat class of plant disease resistance genes
1
1994
... 植物病害常造成大范围的粮食减产, 培育抗病品种是抵御作物病害最为经济且环保的手段之一.1994年, 植物核苷酸结合的富含亮氨酸重复蛋白(nucleotide-binding leucine-rich repeat, NLR)基因首次被克隆, 分别为烟草(Nicotiana tabacum)的N基因(
Comparative analysis of the flax immune receptors L6 and L7 suggests an equilibrium- based switch activation model
1
2016
... NB-ARC属于NLR蛋白保守结构域, 与哺乳动物的细胞凋亡蛋白酶激活因子Apaf-1 (apoptotic protease- activating factor-1)、植物R基因编码的抗病蛋白和秀丽线虫的细胞死亡蛋白CED-4 (caenorhabditis elegans death-4 protein)具有同源性(
Arabidopsis miR472- RDR6 silencing pathway modulates PAMP- and effector- triggered immunity through the post-transcriptional control of disease resistance genes
1
2014
... 番茄miR482/2118超级家族与拟南芥miR472- RDR6为同源microRNA, 它们通过RNA诱导沉默方式在转录后水平调节多个CNLs (
Enhanced resistance to bacterial and oomycete pathogens by short tandem target mimic RNAs in tomato
1
2019
... 番茄miR482/2118超级家族与拟南芥miR472- RDR6为同源microRNA, 它们通过RNA诱导沉默方式在转录后水平调节多个CNLs (
The CC domain structure from the wheat stem rust resistance protein Sr33 challenges paradigms for dimerization in plant NLR proteins
1
2016
... 与CC相比, TIR结构域更加保守, 且广泛存在于动、植物中(单子叶植物除外), 被认为与先天免疫系统的蛋白和蛋白互作相关.CC结构域的序列则较为多样化, 且常与其它结构域串联.例如, 番茄(Solanum lycopersicum)中的Sw-5b与SD序列相连(
Barley MLA immune receptors directly interfere with antagonistically acting transcription factors to initiate disease resistance signaling
1
2013
... 研究表明, NLRs可通过与转录因子直接互作来调控下游免疫信号转导.例如, 大麦CNL MLA10通过其CC结构域与两种相互拮抗的转录因子WRKY1和MYB6 (v-myb avian myeloblastosis viral oncogene homolog 6)直接互作, 进而调控植物免疫(
The evolutionarily conserved E3 ubiquitin ligase AtCHIP contributes to plant immunity
1
2016
... 翻译后修饰主要包括泛素化、磷酸化和糖基化等几类, 其中泛素化修饰的应用最为广泛, 可以在蛋白水平调节植物的免疫.
AtCDC48A is involved in the turnover of an NLR immune receptor
1
2016
... 翻译后修饰主要包括泛素化、磷酸化和糖基化等几类, 其中泛素化修饰的应用最为广泛, 可以在蛋白水平调节植物的免疫.
Overexpression of gma-miR1510a/b suppresses the expression of a NB-LRR domain gene and reduces resistance to Phytophthora sojae
1
2017
... 番茄miR482/2118超级家族与拟南芥miR472- RDR6为同源microRNA, 它们通过RNA诱导沉默方式在转录后水平调节多个CNLs (
A role for small RNA in regulating innate immunity during plant growth
1
2018
... 番茄miR482/2118超级家族与拟南芥miR472- RDR6为同源microRNA, 它们通过RNA诱导沉默方式在转录后水平调节多个CNLs (
Signaling from the plasma-membrane localized plant immune receptor RP- M1 requires self-association of the full-length protein
1
2017
... 与CC相比, TIR结构域更加保守, 且广泛存在于动、植物中(单子叶植物除外), 被认为与先天免疫系统的蛋白和蛋白互作相关.CC结构域的序列则较为多样化, 且常与其它结构域串联.例如, 番茄(Solanum lycopersicum)中的Sw-5b与SD序列相连(
Structural insights into plant NLR immune receptor function
3
2016
... 与CC相比, TIR结构域更加保守, 且广泛存在于动、植物中(单子叶植物除外), 被认为与先天免疫系统的蛋白和蛋白互作相关.CC结构域的序列则较为多样化, 且常与其它结构域串联.例如, 番茄(Solanum lycopersicum)中的Sw-5b与SD序列相连(
... ;
... NLR通常需要形成二聚体或者多聚体才被激活(
Epigenetic control of defense signaling and priming in plants
1
2016
... 植物具备多种表观遗传调控机制, 主要包括染色质重塑、DNA甲基化、组蛋白修饰和非编码RNA调控4类.染色质重塑因子SPLAYED (在筛选MUSE过程中得到)属于SWI/SNF家族, 其通过影响SNC1转录抑制由SNC1介导的免疫反应(
Rice Exo70 interacts with a fungal effector, AVR-Pii, and is required for AVR-Pii-triggered immunity
1
2015
... 间接识别指NLR蛋白不直接与病原菌效应蛋白互作, 而是作用于某些目标蛋白, 通过感知目标蛋白(也称保卫蛋白)的变化引发免疫反应(
The F-box protein CPR1/CPR30 negatively regulates R protein SNC1 accumulation
1
2012
... 翻译后修饰主要包括泛素化、磷酸化和糖基化等几类, 其中泛素化修饰的应用最为广泛, 可以在蛋白水平调节植物的免疫.
Multilayer and interconnected post-transcriptional and co-transcriptional control of plant NLRs
1
2015
...
NAD+ cleavage activity by animal and plant TIR domains in cell death pathways
1
2019
... 此外, 植物NLR的N端也参与特异性识别病原菌效应子和引起细胞死亡的过程.动、植物体内NLR蛋白被病原体诱导后, TIR结构域的NAD+裂解活性被激活, 识别病原体并转化为细胞死亡信号, 从而引起免疫反应(
Identification of methylosome components as negative regulators of plant immunity using chemical genetics
1
2016
... 可变剪接在调控NLR (特别是TLR)的过程中起重要作用, 其通过产生异常转录本/蛋白(或通过引发NMD (nonsense-mediated decay))的方式行使功能.转录组分析显示, 可变剪接广泛存在于具有多个内含子的NLR (特别是TNL)中.
Plant TRAF proteins regulate NLR immune receptor turnover
1
2016
... 翻译后修饰主要包括泛素化、磷酸化和糖基化等几类, 其中泛素化修饰的应用最为广泛, 可以在蛋白水平调节植物的免疫.
An E4 ligase facilitates poly- ubiquitination of plant immune receptor resistance pro- teins in Arabidopsis
1
2014
... 翻译后修饰主要包括泛素化、磷酸化和糖基化等几类, 其中泛素化修饰的应用最为广泛, 可以在蛋白水平调节植物的免疫.
The chromatin remodeler SPLAYED negatively regulates SNC1- mediated immunity
1
2015
... 植物具备多种表观遗传调控机制, 主要包括染色质重塑、DNA甲基化、组蛋白修饰和非编码RNA调控4类.染色质重塑因子SPLAYED (在筛选MUSE过程中得到)属于SWI/SNF家族, 其通过影响SNC1转录抑制由SNC1介导的免疫反应(
A partial loss-of-function mutation in an Arabidopsis RNA polymerase III subunit leads to pleiotropic defects
1
2016
... 植物具备多种表观遗传调控机制, 主要包括染色质重塑、DNA甲基化、组蛋白修饰和非编码RNA调控4类.染色质重塑因子SPLAYED (在筛选MUSE过程中得到)属于SWI/SNF家族, 其通过影响SNC1转录抑制由SNC1介导的免疫反应(
The HSP90 complex of plants
1
2012
... 已有研究表明, RAR1-SGT1b-HSP90伴侣复合体各组分对由各种NLR介导的ETI都很重要(例如, RpiBLB2信号转导需要SGT1 (suppressor of the G2 allele of skp1), 但无须HSP90 (heat shock protein 90)或RAR1 (required for Mla12 resistance) (
Integration of decoy domains derived from protein targets of pathogen effectors into plant immune receptors is widespread
1
2016
... NLR本身也可成为诱饵蛋白, 此类NLR含有特殊的整合结构域, 可对病原微生物进行特异性识别(
SGT1 interacts with the Prf resistance protein and is required for Prf accumulation and Prf-mediated defense signaling
1
2013
... LRR是整个蛋白结构中最为保守且简单的结构域, 处于NLR蛋白的C端, 由多个富含亮氨酸的重复序列组成.功能域互换实验表明, 即使蛋白的其它部分高度相似, 仅LRR不同, NLR蛋白识别的效应子也不同.例如, NAIP5 (neuronal apoptosis inhibitory protein 5)可通过其C末端的LRR结构域识别不同的细菌鞭毛蛋白(
Epigenetic regulation of intragenic transposable elements impacts gene transcription in Arabidopsis thaliana
1
2015
... 植物具备多种表观遗传调控机制, 主要包括染色质重塑、DNA甲基化、组蛋白修饰和非编码RNA调控4类.染色质重塑因子SPLAYED (在筛选MUSE过程中得到)属于SWI/SNF家族, 其通过影响SNC1转录抑制由SNC1介导的免疫反应(
The intracellular nucleotide-binding leucine-rich repeat receptor (SlNRC4a) enhances immune signaling elicited by extra- cellular perception
1
2018
... NLR通常需要形成二聚体或者多聚体才被激活(
A plant immune receptor adopts a two-step recognition mechanism to enhance viral effector perception
1
2019
... 植物通过对病原菌效应蛋白的特异识别感知病原体的入侵, 进而激活快速、精准的免疫反应.识别方式包括2类: 一类为相对简单的直接识别(
The miR9863 family regulates distinct Mla alleles in barley to attenuate NLR receptor-triggered disease resistance and cell-death signaling
2
2014
... 番茄miR482/2118超级家族与拟南芥miR472- RDR6为同源microRNA, 它们通过RNA诱导沉默方式在转录后水平调节多个CNLs (
... ) miR1510a/b靶向调节特异性防御大豆疫霉所需的TNL, 但在大豆疫霉菌感染期间其效应被抑制(
The A. thaliana disease resistance gene RPS2 encodes a protein containing a nucleotide-binding site and leucine- rich repeats
1
1994
... 植物病害常造成大范围的粮食减产, 培育抗病品种是抵御作物病害最为经济且环保的手段之一.1994年, 植物核苷酸结合的富含亮氨酸重复蛋白(nucleotide-binding leucine-rich repeat, NLR)基因首次被克隆, 分别为烟草(Nicotiana tabacum)的N基因(
TIR-only protein RBA1 recognizes a pathogen effector to regulate cell death in Arabidopsis
1
2017
... 植物通过对病原菌效应蛋白的特异识别感知病原体的入侵, 进而激活快速、精准的免疫反应.识别方式包括2类: 一类为相对简单的直接识别(
Rpi-blb2-mediated hypersensitive cell death caused by phytophthora infestans AVRblb2 requires SGT1, but not EDS1, NDR1, salicylic acid-, jasmonic acid-, or ethylene-mediated signaling
1
2014
... 已有研究表明, RAR1-SGT1b-HSP90伴侣复合体各组分对由各种NLR介导的ETI都很重要(例如, RpiBLB2信号转导需要SGT1 (suppressor of the G2 allele of skp1), 但无须HSP90 (heat shock protein 90)或RAR1 (required for Mla12 resistance) (
Recognition of the Magnaporthe oryzae effector AVR-Pia by the decoy domain of the rice NLR immune receptor RGA5
1
2017
... NLR本身也可成为诱饵蛋白, 此类NLR含有特殊的整合结构域, 可对病原微生物进行特异性识别(
Novel positive regulatory role for the SPL6 transcription factor in the N TIR-NB-LRR receptor-mediated plant innate immunity
1
2013
... 研究表明, NLRs可通过与转录因子直接互作来调控下游免疫信号转导.例如, 大麦CNL MLA10通过其CC结构域与两种相互拮抗的转录因子WRKY1和MYB6 (v-myb avian myeloblastosis viral oncogene homolog 6)直接互作, 进而调控植物免疫(
The E3 ligase APIP10 connects the effector AvrPiz-t to the NLR receptor Piz-t in rice
1
2016
... 间接识别指NLR蛋白不直接与病原菌效应蛋白互作, 而是作用于某些目标蛋白, 通过感知目标蛋白(也称保卫蛋白)的变化引发免疫反应(
The movement protein (NSm) of tomato spotted wilt virus is the avirulence determinant in the tomato Sw-5 gene-based resistance
1
2014
... 与CC相比, TIR结构域更加保守, 且广泛存在于动、植物中(单子叶植物除外), 被认为与先天免疫系统的蛋白和蛋白互作相关.CC结构域的序列则较为多样化, 且常与其它结构域串联.例如, 番茄(Solanum lycopersicum)中的Sw-5b与SD序列相连(
Structure-function analysis of the coiled-coil and leucine-rich repeat domains of the RPS5 disease resistance protein
1
2012
... LRR是整个蛋白结构中最为保守且简单的结构域, 处于NLR蛋白的C端, 由多个富含亮氨酸的重复序列组成.功能域互换实验表明, 即使蛋白的其它部分高度相似, 仅LRR不同, NLR蛋白识别的效应子也不同.例如, NAIP5 (neuronal apoptosis inhibitory protein 5)可通过其C末端的LRR结构域识别不同的细菌鞭毛蛋白(
Recent advances in plant NLR structure, function, localization, and signaling
2
2013
... NLR通常需要形成二聚体或者多聚体才被激活(
... 许多CNLs (包括RPS5、RPS2和RPM1)定位于质膜(
Roq1 mediates recognition of the Xanthomonas and Pseudomonas effector proteins XopQ and HopQ1
1
2017
... 植物通过对病原菌效应蛋白的特异识别感知病原体的入侵, 进而激活快速、精准的免疫反应.识别方式包括2类: 一类为相对简单的直接识别(
Nucleocytoplasmic distribution is required for activation of resistance by the potato NB-LRR receptor Rx1 and is balanced by its functional domains
1
2010
... 许多CNLs (包括RPS5、RPS2和RPM1)定位于质膜(
E3 ligase SAUL1 serves as a positive regulator of PAMP-triggered immunity and its homeostasis is monitored by immune receptor SOC3
2
2017
... 翻译后修饰主要包括泛素化、磷酸化和糖基化等几类, 其中泛素化修饰的应用最为广泛, 可以在蛋白水平调节植物的免疫.
... ); 免疫正调节因子E3泛素连接酶SAUL1 (senescence-associated E3 ubiquitin ligase 1)由TNL SOC3保卫(
Activation of a plant NLR complex through heteromeric association with an autoimmune risk variant of another NLR
1
2017
... NLR通常需要形成二聚体或者多聚体才被激活(
An alternative polyadenylation mechanism coopted to the Arabidopsis RPP7 gene through intronic retrotransposon domestication
1
2013
... 植物具备多种表观遗传调控机制, 主要包括染色质重塑、DNA甲基化、组蛋白修饰和非编码RNA调控4类.染色质重塑因子SPLAYED (在筛选MUSE过程中得到)属于SWI/SNF家族, 其通过影响SNC1转录抑制由SNC1介导的免疫反应(
Plant disease- resistance proteins and the gene-for-gene concept
1
1998
... NB-ARC属于NLR蛋白保守结构域, 与哺乳动物的细胞凋亡蛋白酶激活因子Apaf-1 (apoptotic protease- activating factor-1)、植物R基因编码的抗病蛋白和秀丽线虫的细胞死亡蛋白CED-4 (caenorhabditis elegans death-4 protein)具有同源性(
TIR domains of plant immune receptors are NAD+-cleaving enzymes that promote cell death
1
2019
... 此外, 植物NLR的N端也参与特异性识别病原菌效应子和引起细胞死亡的过程.动、植物体内NLR蛋白被病原体诱导后, TIR结构域的NAD+裂解活性被激活, 识别病原体并转化为细胞死亡信号, 从而引起免疫反应(
Reconstitution and structure of a plant NLR resistosome conferring immunity
2
2019
... NB-ARC属于NLR蛋白保守结构域, 与哺乳动物的细胞凋亡蛋白酶激活因子Apaf-1 (apoptotic protease- activating factor-1)、植物R基因编码的抗病蛋白和秀丽线虫的细胞死亡蛋白CED-4 (caenorhabditis elegans death-4 protein)具有同源性(
... NLR通常需要形成二聚体或者多聚体才被激活(
Ligand-triggered allosteric ADP release primes a plant NLR complex
2
2019
... NB-ARC属于NLR蛋白保守结构域, 与哺乳动物的细胞凋亡蛋白酶激活因子Apaf-1 (apoptotic protease- activating factor-1)、植物R基因编码的抗病蛋白和秀丽线虫的细胞死亡蛋白CED-4 (caenorhabditis elegans death-4 protein)具有同源性(
... NLR通常需要形成二聚体或者多聚体才被激活(
An E3 ligase affects the NLR receptor stability and immunity to powdery mildew
1
2016
... 翻译后修饰主要包括泛素化、磷酸化和糖基化等几类, 其中泛素化修饰的应用最为广泛, 可以在蛋白水平调节植物的免疫.
The product of the tobacco mosaic virus resistance gene N: similarity to toll and the interleukin-1 receptor
1
1994
... 植物病害常造成大范围的粮食减产, 培育抗病品种是抵御作物病害最为经济且环保的手段之一.1994年, 植物核苷酸结合的富含亮氨酸重复蛋白(nucleotide-binding leucine-rich repeat, NLR)基因首次被克隆, 分别为烟草(Nicotiana tabacum)的N基因(
An autoactive mutant of the M flax rust resistance protein has a preference for binding ATP, whereas wild-type M protein binds ADP
1
2011
... NB-ARC属于NLR蛋白保守结构域, 与哺乳动物的细胞凋亡蛋白酶激活因子Apaf-1 (apoptotic protease- activating factor-1)、植物R基因编码的抗病蛋白和秀丽线虫的细胞死亡蛋白CED-4 (caenorhabditis elegans death-4 protein)具有同源性(
Regulation of transcription of nucleotide-binding leucine- rich repeat-encoding genes SNC1 and RPP4 via H3K4 trimethylation
1
2013
... 植物具备多种表观遗传调控机制, 主要包括染色质重塑、DNA甲基化、组蛋白修饰和非编码RNA调控4类.染色质重塑因子SPLAYED (在筛选MUSE过程中得到)属于SWI/SNF家族, 其通过影响SNC1转录抑制由SNC1介导的免疫反应(
A nucleotide-binding site-leucine-rich repeat receptor pair confers broad-spectrum disease resistance through physical association in rice
1
2019
... NLR本身也可成为诱饵蛋白, 此类NLR含有特殊的整合结构域, 可对病原微生物进行特异性识别(
P-loop-dependent NLR SNC1 can oligomerize and activate immunity in the nucleus
1
2014
... 许多CNLs (包括RPS5、RPS2和RPM1)定位于质膜(
NLR-associating transcription factor bHLH84 and its paralogs function redundantly in plant immunity
1
2014
... 研究表明, NLRs可通过与转录因子直接互作来调控下游免疫信号转导.例如, 大麦CNL MLA10通过其CC结构域与两种相互拮抗的转录因子WRKY1和MYB6 (v-myb avian myeloblastosis viral oncogene homolog 6)直接互作, 进而调控植物免疫(
The cyclin L homolog MOS12 and the MOS4-associated complex are required for the proper splicing of plant resistance genes
2
2012
... 可变剪接在调控NLR (特别是TLR)的过程中起重要作用, 其通过产生异常转录本/蛋白(或通过引发NMD (nonsense-mediated decay))的方式行使功能.转录组分析显示, 可变剪接广泛存在于具有多个内含子的NLR (特别是TNL)中.
... 的可变剪接发生改变, 将影响植物的基础免疫抗性(
Autoimmunity conferred by chs3-2D relies on CSA1, its adjacent TNL-encoding neighbour
1
2015
... NLR本身也可成为诱饵蛋白, 此类NLR含有特殊的整合结构域, 可对病原微生物进行特异性识别(
Structural basis for specific flagellin recognition by the NLR protein NAIP5
1
2018
... LRR是整个蛋白结构中最为保守且简单的结构域, 处于NLR蛋白的C端, 由多个富含亮氨酸的重复序列组成.功能域互换实验表明, 即使蛋白的其它部分高度相似, 仅LRR不同, NLR蛋白识别的效应子也不同.例如, NAIP5 (neuronal apoptosis inhibitory protein 5)可通过其C末端的LRR结构域识别不同的细菌鞭毛蛋白(
RRM transcription factors interact with NLRs and regulate broad-spectrum blast resistance in rice
1
2019
... 研究表明, NLRs可通过与转录因子直接互作来调控下游免疫信号转导.例如, 大麦CNL MLA10通过其CC结构域与两种相互拮抗的转录因子WRKY1和MYB6 (v-myb avian myeloblastosis viral oncogene homolog 6)直接互作, 进而调控植物免疫(
PRL1, an RNA-binding protein, positively regulates the accumulation of miRNAs and siRNAs in Arabidopsis
2
2014
... 可变剪接在调控NLR (特别是TLR)的过程中起重要作用, 其通过产生异常转录本/蛋白(或通过引发NMD (nonsense-mediated decay))的方式行使功能.转录组分析显示, 可变剪接广泛存在于具有多个内含子的NLR (特别是TNL)中.
... ).
The diversification of plant NBS-LRR defense genes directs the evolution of microRNAs that target them
1
2016
...
The NLR protein SUMM2 senses the disruption of an immune signaling MAP kinase cascade via CRCK3
1
2017
... 间接识别指NLR蛋白不直接与病原菌效应蛋白互作, 而是作用于某些目标蛋白, 通过感知目标蛋白(也称保卫蛋白)的变化引发免疫反应(
Arabidopsis resistance protein SNC1 activates immune responses through association with a transcriptional corepressor
1
2010
... 研究表明, NLRs可通过与转录因子直接互作来调控下游免疫信号转导.例如, 大麦CNL MLA10通过其CC结构域与两种相互拮抗的转录因子WRKY1和MYB6 (v-myb avian myeloblastosis viral oncogene homolog 6)直接互作, 进而调控植物免疫(
Arabidopsis chromatin-remodeling factor CHR5 regulates plant immune responses and nucleosome occupancy
1
2017
... 植物具备多种表观遗传调控机制, 主要包括染色质重塑、DNA甲基化、组蛋白修饰和非编码RNA调控4类.染色质重塑因子SPLAYED (在筛选MUSE过程中得到)属于SWI/SNF家族, 其通过影响SNC1转录抑制由SNC1介导的免疫反应(
备案号: 京ICP备16067583号-21
版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由北京玛格泰克科技发展有限公司设计开发

{kind=link}
{kind=link}