Corresponding authors: *E-mail: 350971781@qq.com First author contact:?These authors contributed equally to this paper Received:2019-12-17Accepted:2020-02-26Online:2020-07-01
Abstract Low temperature, drought, high salt, hypoxia and other adverse environmental changes affect plant growth and development. Plants adapt to these adverse conditions through the development of complex regulatory mechanisms during long-term evolution. APETALA2/ethylene responsive factor (AP2/ERF) is a plant-specific transcription factor that plays a key regulatory role in various stress responses. In recent years, more and more studies have shown that plant hormone-mediated signaling is closely related to stress responses, and AP2/ERF transcription factor and hormone signal transduction form a cross-regulatory network. Many AP2/ERF transcription factors respond to plant hormones abscisic acid (ABA) and ethylene (ET), activating the expression of stress response genes that are dependent on and independent of ABA and ET. In addition, AP2/ERF transcription factors are also involved in gibberellin (GA), cytokinin (CTK) and brassinsteroid (BR) mediated growth and developmental processes and stress responses. This paper briefly reviews the research progress of AP2/ERF transcription factors in term of structure, transcriptional regulation, posttranslational modifications, binding sites and interacting proteins as well as its transduction pathways involved in hormone dependent- or independent- regulation of the abiotic stress responses, which will provide the basis for further understanding the roles of different AP2/ERF transcription factors in the regulation of hormone and stress response network in plants. Keywords:AP2/ERF transcription factor;hormone;abiotic stress;regulation
PDF (1076KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 洪林, 杨蕾, 杨海健, 王武. AP2/ERF转录因子调控植物非生物胁迫响应研究进展. 植物学报, 2020, 55(4): 481-496 doi:10.11983/CBB19243 Hong Lin, Yang Lei, Yang Haijian, Wang Wu. Research Advances in AP2/ERF Transcription Factors in Regulating Plant Responses to Abiotic Stress. Chinese Bulletin of Botany, 2020, 55(4): 481-496 doi:10.11983/CBB19243
植物在生长发育过程中经常受到生物或非生物胁迫的影响, 由此, 植物逐渐形成了复杂的调节机制感知胁迫信号, 对外部环境变化做出快速响应(郭倩倩和周文彬, 2019)。脱落酸(abscisic acid, ABA)、乙烯(ethylene, ET)及赤霉素(gibberellin, GA)等激素作为信号分子能够调节植物的生长发育、形态建成以及抗逆反应等生物学过程。该过程也受转录因子(transcription factor, TF)调控, 即转录因子通过依赖或不依赖于激素介导的信号通路参与调控植物对干旱、低温、缺氧、高盐和淹水等非生物胁迫的响应, 进而影响植物的逆境耐受能力(Gibbs et al., 2015; Verma et al., 2016; Li et al., 2018)。AP2/ERF (APETALA2/eth-ylene responsive factor)是植物特有的转录因子家族之一, 已在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、高粱(Sorghum bicolor)、大白菜(Brassica pekinensis)、毛竹(Phyllostachys edulis)、玉米(Zea mays)、大麦(Hordeum vulgare)和小麦(Triticum aestivum)等物种中被成功分离鉴定(Xu and Chua, 2011; Chandler, 2018)。通过突变体实验, 科学家发现了许多与AP2/ERF相关的非生物胁迫或激素信号应答基因。目前, AP2/ERF已成为研究非生物胁迫和激素互作的热点候选基因。本文从AP2/ERF转录因子的结构特征、转录调控、翻译后修饰、结合位点、协同互作蛋白以及其参与调控依赖或不依赖激素信号途径的非生物胁迫响应进行综述, 为进一步阐明AP2/ERF转录因子在非生物胁迫调控网络中的作用提供理论依据。
1 AP2/ERF转录因子的结构特征和分类
AP2/ERF转录因子具有独特的结构特征, 其由DNA结合域(DNA-binding domain)、转录调控域(transcription regulation domain)、寡聚化位点(oligomerization site)和核定位信号(nuclear localization signal, NLS) 4个主要功能区组成, 并以高度保守的AP2/ ERF结合域而得名(Sakuma et al., 2002)。通常情况下, AP2/ERF转录因子至少含有1个AP2结构域的DNA结合区, 该结构域包含60-70个氨基酸残基, 按照3个β折叠和1个α螺旋方式形成典型的三维结构(Nakano et al., 2006; Abiri et al., 2017)。根据包含的AP2/ERF结构域数量和结合序列特点, 可将AP2/ ERF家族分为AP2 (APETALA2)、RAV (related to ABI3/VP1)、脱水反应元件结合蛋白(dehydration- responsive element binding protein, DREB) (A1-A6亚组)、乙烯应答因子(ethylene-responsive factor, ERF) (V-X亚组)和Soloist五个亚族(Nakano et al., 2006; Licausi et al., 2013)。其中, 对AP2、DREB和ERF的研究较为广泛和深入, 而有关Soloist亚族的研究报道极少, 已知其核苷酸序列在多数植物中高度保守(Sun et al., 2016)。
2 AP2/ERF转录因子的结合位点
AP2/ERF转录因子通过特异性结合胁迫应答基因启动子区顺式元件, 参与调控植物的逆境防御反应。DREB亚族成员特异性识别并结合启动子区域的DRE/CRT (dehydration responsive element/C-re-peat)元件(核心序列A/GCCGAC), 诱导干旱、低温和盐胁迫相关应答基因的表达。ERF亚族成员则可通过与乙烯响应元件(ethylene response element, ERE) (核心序列AGCCGCC, 又称GCC-box)结合, 从而参与调控乙烯应答和非生物胁迫(Franco-Zorrilla et al., 2014)。此外, 研究发现许多AP2/ERF转录因子可同时与DRE/CRT和ERE两种元件结合, 如DREB亚族成员TINY、CBF1、ERF53、RAP2.4、TG/RAP2.4a以及ERF亚族成员ERF1、ERF4和ERF71 (Chen et al., 2012; Zhu et al., 2014; Lee et al., 2015), 但结合能力有差异。在拟南芥中, DREB2C和DEAR3与类GCC-box元件(GCCGCC)的结合能力远超DRE元件(Franco-Zorrilla et al., 2014)。AP2/ERF转录因子还能与DRE和ERE之外的顺式元件结合, 如偶联元件1 (CE1, TGCCACCG)、类偶联元件3 (CE-3like, CGCG)和缺氧响应启动子元件(hypoxia-responsive promoter element, HRPE), 以及ATCTA、CAACA、CATGCA、CAA/CA/CTG、ATCGAG和(NC/GT) CGNCCA (Bossi et al., 2009; Zhu et al., 2010; Chen et al., 2016; Gasch et al., 2016; Park et al., 2016)。近年来, 科学家通过蛋白质结合微阵列(protein binding microarray, PBM)技术发现结构特征高度相似的转录因子具有相似的DNA结合位点, 同时也具有相似的生物学功能。该研究结果合理地解释了转录因子功能冗余现象, 也为预测和挖掘未知AP2/ERF转录因子提供了高效可行的技术方法(Franco-Zorrilla et al., 2014)。
3 AP2/ERFs的转录调控、翻译后修饰及协同互作蛋白
3.1 AP2/ERFs的转录调控
正常环境下, AP2/ERF转录因子表达水平较低, 激素或逆境胁迫相关基因与AP2/ERF转录因子保守的顺式作用元件结合, 进而调控其表达(Owji et al., 2017)。研究表明, 拟南芥DREB1A/CBF3、DREB1C/CBF2、DREB2C、DREB2G和DEAR3的启动子中包含较多的热激元件(heat shock element, HSE)、低温响应(low temperature responsive, LTR)、脱落酸响应(ABA responsive element, ABRE)等顺式作用元件(Sazegari et al., 2015)。热激因子1 (heat shock factor 1, HSF1)、脱落酸响应结合蛋白(ABA responsive element binding protein/ABRE binding factors, AREB/ABF)可分别与DREB2A启动子中的热激应答和ABA响应元件结合, 调控其表达, 进而改变植物胁迫的耐受性(Kim et al., 2011; Liu et al., 2011)。蛋白磷酸酶2C (type 2C protein phosphatase, PP2C)包含2个以上的ABRE元件, AREB1、AREB2和ABF3协同正调控AHG1、AHG3、HAI1及HAI2等PP2Cs基因表达(Yoshida et al., 2010)。此外, 越来越多的研究证明ET信号通路参与拟南芥盐胁迫反应。进一步分析表明, ESE1 (ethylene and salt-inducible ERF1)在EIN3/EIL1下游调节ET响应基因。EIN3 (ethylene insensitive 3)可与ESE1启动子结合形成EIN3-ESE1转录复合体, 继而调控RD29A和COR15A等基因的表达(Zhang et al., 2011)。磷酸化、泛素化、甲基化和乙酰化等组蛋白修饰方式通过调控染色质开放或封闭的空间状态, 激活或抑制AP2/ERF转录(Pfluger and Wagner, 2007)。组蛋白去乙酰化酶(histone deacetylase, HDAC)和聚乙二醇(PEG)促进DREB1基因启动子区乙酰化, 增强DREB1基因的表达(Zhang et al., 2018)。非生物胁迫下, 组蛋白变体和组蛋白修饰状态也可发生改变, 并可遗传到下一代(Asensii-Fabado et al., 2017)。miRNA介导的RNA沉默和翻译抑制在AP2/ERF表达调控中也发挥重要作用。研究发现miRNA172靶向拟南芥AP2mRNA并抑制其翻译, 而miRNA156和miRNA838两者也具有类似调节AP2/ERF的功能(Kavas et al., 2015)。此外, 在拟南芥、水稻及玉米中还发现通过可变剪切产生的OsDREB2A/2B、ZmDREB2A、WDREB2以及HvDRF1等多种AP2/-ERF功能异构体(Matsukura et al., 2010)。
3.2 AP2/ERFs翻译后修饰
翻译后修饰(post-translational modification, PTM)是蛋白质合成途径的后期加工过程, 不同的修饰方式对AP2/ERF蛋白的活性、稳定性和丰度的影响有差异。磷酸化是转录因子的主要修饰途径之一, 磷酸化水平与下游靶基因的表达水平关系密切。拟南芥ABA信号通路的正调控因子蔗糖非酵解型蛋白激酶(Snf1-re-lated protein kinases, SnRKs)可磷酸化RAV1, 进而抑制RAV1的转录阻遏效应(Feng et al., 2014), 而ERF104和ERF6可被后MPK3/6 (mitogen-activated protein kinase 3/6)磷酸化修饰, 并在下游级联反应中发挥重要作用, 调控植物对病原体的免疫反应(Meng et al., 2013)。DREB2A是拟南芥的关键转录激活因子, 在其中央序列存在一个负调控域(negative regulatory domain, NRD)。热胁迫条件下, NRD中的Ser/Thr残基磷酸化水平降低, DREB2A蛋白稳定性提高, 敲除NRD后DREB2A呈组成型激活, 表明抑制NRD磷酸化可以稳定和激活DREB2A的表达, 从而增强植物的耐热性(Mizoi et al., 2019)。泛素介导的蛋白质降解(26S蛋白酶体途径)也参与调节AP2/ ERF蛋白的稳定性。非生物胁迫条件下, DREB2A和ERF75/RAP-2.2蛋白分别被RING家族E3泛素连接酶DRIP1/2 (DREB2A-interacting protein1/2)和SINAT2 (seven in absentia of Arabidopsis 2)直接泛素化(Cheng et al., 2012; Papdi et al., 2015)。研究发现CUL3-E3连接酶适配子BPMs (BTB/POZ and math domain)可与DREB2A蛋白的负调控域NRD在核内发生互作, 敲除BPM增加DREB2A蛋白的积累, 促进下游靶基因的表达, 提高植株的耐热和耐旱能力, 这也说明BPM对DREB2A蛋白的稳定性起负调控作用(Morimoto et al., 2017)。此外, 研究发现ERF-VII类蛋白可在质膜上与乙酰辅基A结合蛋白(Acyl-CoA binding protein, ACBP)互作。在缺氧条件下, RAP2.12与ACBP分离后进入细胞核, 在脱酰胺基和精氨酰基化的作用下通过氧依赖途径移除ERF72/RAP2.3、ERF74/RAP2.12蛋白的甲硫氨酸N末端, 将半胱氨酸氧化成半胱亚磺酸, 引发蛋白快速降解并暴露出非稳定N端的氨基酸残基(Gibbs et al., 2015; Abbas et al., 2015)。
3.3 AP2/ERFs的协同互作蛋白
许多研究表明, AP2/ERF转录因子可直接与靶基因启动子结合, 激活或抑制下游靶基因的表达, 其N端DNA结合域和C端转录激活域在胁迫相关靶基因的转录激活中起重要作用(Nakano et al., 2006)。进一步分析表明, AP2/ERF转录因子中含有EAR基序(LxLxL或DLNxxP)、TLLLFR基序和BRD结构域(B3 repression domain, 保守序列为R/KLFGV)等多个转录抑制结构域(Ikeda and Ohme-Takagi, 2009; Kagale and Rozwadowski, 2011; Deepika et al., 2016)。含EAR基序的转录因子是一种重要的抑制子, 在植物非生物胁迫应答基因的诱导表达中起负调控作用。Tiwari等(2012)从拟南芥ERF和AP2亚族中发现了一种转录激活关键元件EDLL, EDLL可与EAR基序(ERF-associated amphiphilic repression)发生互作, 拮抗后者介导的转录抑制作用。而不同的AP2/ ERF转录因子也可协同招募TPL (topless)和TPR (topless-related)等转录共抑制子(Causier et al., 2012)或组蛋白修饰因子, 进而抑制下游靶基因的表达(Song et al., 2005; Song and Galbraith, 2006)。Song等(2005)发现AtERF7特异性地结合GCC-box, 与抑制因子ATSIN3及组蛋白去乙酰化酶19 (histone deacetylase 19, HDA19)发生互作并形成转录因子复合物, 参与干旱胁迫和ABA响应调控。AtERF7过表达株系保卫细胞对ABA的敏感性减弱, 水分散失增加; 反之, AtERF7 RNA干扰系对ABA的敏感性增强。类似的, ERF3能与组蛋白去乙酰化酶复合物亚基SAP18 (SIN3 associated polypeptide P18)互作, 共同招募HDA19形成复合体, 进而抑制相关基因的表达(Song and Galbraith, 2006)。BRD结构域被认为是RAV1和RAV2转录因子行使抑制功能的关键元件, 其保守序列中第1位氨基酸残基多为疏水性亮氨酸、缬氨酸和蛋氨酸残基, 突变导致功能丧失(Ikeda and Ohme-Takagi, 2009)。研究显示, 与AP2/ERFs协同调控的基因同样可被AP2/ERFs靶基因富集, 分析此类基因的同源基序有助于识别转录因子的假定靶基因并预测其生物学功能(Franco-Zorrillaetal et al., 2014)。
CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员。近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018)。冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015)。ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017)。ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018)。而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011)。目前, 关于ICE-CBF-COR冷信号途径研究最为深入。低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a)。而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016)。CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013)。利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016)。在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015)。与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007)。
部分A5亚组DREB类转录因子则对上述调控网络具有负反馈调节作用。DEAR1 (DREB and EAR motif protein 1)可能既在CBF上游作用, 又与RAP2.1在CBF下游扮演冷胁迫响应的调节子, 调节RD29A/COR78、COR15A和KIN1等冷诱导相关基因的表达, 负调控植物的耐冷和耐旱性(Fowler and Thomashow, 2002; Tsutsui et al., 2009; Dong and Liu, 2010)。DEAR1过表达抑制冷诱导CBF的表达, 进而降低植物的耐冷能力(Tsutsui et al., 2009)。低温胁迫或组成型表达CBF可诱导RAP2.1的表达, CBF2比RAP2.1对低温更为敏感(Dong and Liu, 2010)。在CBF调控通路中, DEAR1和RAP2.1如何整合还需深入研究。RAP2.1是否与其它DEAR转录因子共同调节植物非生物胁迫响应仍不清楚。研究发现DREB-A5亚组有6种含EAR基序的转录抑制子可与DRE元件结合, 进而调控非生物胁迫响应基因的表达(Nakano et al., 2006)。
ERF亚家族转录因子是ET介导的胁迫应答信号通路下游的关键调控因子(Licausi et al., 2013; Gibbs et al., 2015; Kazan, 2015; Müller and Munné-Bosch, 2015)。冷胁迫下, 植物体内的ET主要起负调控作用, 过量ET或1-氨基环丙烷-1-羧酸降低植物的耐冷性, 而ET生物合成抑制剂氨基乙氧基甘氨酸或感知拮抗剂Ag+具有相反的作用(Shi et al., 2012)。etr1-1、ein4-1、ein2-5、ein3-1和ein3eil突变体表现出更强的耐冷性, 但组成型ET响应etr1-1和ein3过表达植株表现出耐冷性减弱; 冷胁迫诱导ETR1、EIN4、EBF1及EBF2快速上调表达, 而EIN2和EIL1下调表达。进一步分析表明, EIN3通过与启动子区特定元件结合, 抑制ARR5、ARR7、ARR15和CBF转录, ARRs作为冷信号通路的关键节点基因, 在ET和CTK介导的胁迫响应网络中起着关键作用, 过表达ARRs使植株的耐冷能力增强(Shi et al., 2012)。此外, EIN3可激活ERF1和ESEs, 进而参与调控下游胁迫相关基因, 提高植物的耐盐性(Kazan, 2015; Tao et al., 2015)。35S:AtERF4-GFP转基因植株中ABI2、RD29B和RAP18的表达受抑制, 进一步说明AtERF4是ET和ABA信号途径的负调控因子(Yang et al., 2011)。
此外, ET促进RAP2.3转录因子定位于细胞核内, 通过依赖于RAP2.3的方式激发ORA59介导的ET响应过程(Kim et al., 2018)。而ERF-VIIs通过ET不依赖途径参与调控缺氧响应。在ET不敏感突变体或生物合成抑制剂存在的情况下, 缺氧依然可以诱导ERF73/HRE1的表达, ERF-VIIs也可能通过反馈调节负调控ET信号(Hinz et al., 2010; Yang et al., 2011)。研究发现, ET合酶ACS在RAP2.2过表达植株中表达量降低, 而在rap2.2-2敲除突变体中表达上调(Hinz et al., 2010)。然而, 胁迫条件下ERF-VIIs如何通过负反馈机制调控ET生物合成有待进一步研究。
4.2.3 参与赤霉素介导的胁迫应答
赤霉素(GA)广泛分布在植物中并参与调控叶、芽以及合子发育等多种生物学过程。GA由GA20氧化酶(gibberellin 20-oxidases, GA20ox)和GA3氧化酶(GA3ox)等关键酶合成, 在GA2氧化酶(GA2ox)作用下被分解代谢(Rieu et al., 2008)。DELLA蛋白作为GA信号转导途径中的主要负调控因子, 也是整合生长素、ABA、ET和JA等植物激素或环境信号的枢纽蛋白。植物体接受GA信号后, 通过泛素-蛋白酶体途径降解DELLA蛋白, 解除其对生长的抑制作用(Zhang et al., 2009)。冷、盐和渗透胁迫下, GA含量降低, 导致植物生长减缓(Colebrook et al., 2014)。极低浓度GA下, DELLA蛋白抑制GA介导的应答调控, 外源GA处理促进DELLA蛋白降解(Claeys et al., 2012)。
AP2/ERFs参与调控植物GA生物合成相关基因的转录。盐胁迫下, DREB1E和DREB1F抑制GA20ox的表达, 使GA生物合成减少, 引起植株生长迟缓(Magome et al., 2004)。过表达ERF6抑制GA2ox的表达水平, 诱导赤霉素降解酶(gibberellin 2-oxidase 6, GA2ox6)的合成, 维持DELLA蛋白的稳定性。渗透胁迫下, erf6功能获得突变体植株矮小且对胁迫高度敏感, 而erf5/erf6缺失突变体生长受胁迫影响较小。研究发现ERF6还激活了STZ、MYB51和WRKY33等渗透胁迫响应基因的表达(Dubois et al., 2013)。冷胁迫下, CBF1转录因子激发GA2ox基因的表达而降低GA含量, 组成型表达CBF1抑制GA的积累, 植株表现矮化, 表明DELLA蛋白是CBF1介导的冷胁迫响应的关键组分(Achard et al., 2008)。相反, ERF11作为一种GA生物合成和信号转导的正调控因子, 抑制ET生物合成, 促进植株节间伸长。过表达ERF11上调GA3ox1和GA20ox基因的表达水平, 增加GA的积累量(Zhou et al., 2016)。ERF6作为转录激活因子诱导ERF11的表达, 反之, ERF11抑制GA2-OX6、STZ、MYB51和WRKY33等ERF6相关靶基因的表达。进一步实验表明, 4个靶基因在erf11/erf6过表达植株中也不能被诱导表达, ERF11过表达可消除ERF6过表达导致的极度矮化现象, 说明ERF11可能通过直接竞争目标基因启动子而在分子水平上拮抗ERF6 (Dubois et al., 2015)。
在拟南芥和水稻中, SUB1A、SK1和SK2参与GA信号转导通路的调控。淹水胁迫下, SUB1A激活SLR1 (slender rice 1)和SLRL1 (slender rice 1 like 1) (DELLA类似蛋白)转录, 同时SUB1A抑制受GA诱导的淀粉代谢相关基因的表达, 调控植株节间伸长和呼吸作用(Fukao and Bailey-Serres, 2008; Locke et al., 2018; Perata, 2018)。SK1和SK2两个转录因子中均含有EIN3结合位点, 电泳迁移率测定显示, 类EIN3基因EIL1b可与SK1和SK2启动子结合, 激活其转录活性。研究发现, 淹水状态下拟南芥中ET合成增加, 进而诱导SK1和SK2表达, 激发GA20oxs基因上调表达, 促进植株节间伸长, 减轻淹水对其生长的抑制效应(Hattori et al., 2009; Ayano et al., 2014)。
4.2.4 参与细胞分裂素介导的胁迫应答
细胞分裂素(CTK)在植物中具有广泛的生物学效应, 不仅可促进细胞分裂、花芽分化、打破种子休眠及调控营养物质运输, 而且在植物应对逆境胁迫中也起重要作用(Zwack and Rashotte, 2015)。在拟南芥中, CTK利用一种类似于细菌双元组分系统的途径传递信号至下游元件, 受体组氨酸激酶(Arabidopsis histidine kinase, AHK)与CTK结合后发生自磷酸化, 并由磷酸转运蛋白(Arabidopsis histidine-phosphotransfer protein, AHP)介导磷酸基团转移到A型和B型反应调节因子上, 进而调节下游的细胞分裂素响应基因(M?h?nen et al., 2006)。A型RR是目前发现的受细胞分裂素调控的主要蛋白家族, 且A型ARR通过抑制B型RR的活性负调控依赖性CTK信号途径。
细胞分裂素应答因子(cytokinin response factor, CRF)在植物胚、子叶和叶片发育过程中起关键调控作用(Rashotte et al., 2006)。拟南芥crf1/2/5和crf2/3/6突变体的转录组分析表明CRF参与CTK介导的调控网络。CRF基因启动子区含有多个B型ARR结合位点, 据此推测CRFs很可能是B型ARR的直接靶标, 两者之间存在协同作用, 约60%的CTK响应基因同时受CRF和B型ARR调控(Rashotte et al., 2006)。Zwack等(2013)认为CRF6是胁迫条件下AHK3和TCS介导的CTK信号通路下游的新组分, CRF6和ARR2可能通过一个部分重叠或相互作用的平行通路负调控胁迫诱导的叶片衰老过程, 这也表明CRF6和 CTK与非生物胁迫之间存在独特的关系(Zwack et al., 2013)。CRF6除正向调节CTK通路外, 还抑制CTK的生物合成、信号转导和胞内转运相关靶基因的表达, 减轻CTK对非生物胁迫的负调控作用(Zwack et al., 2016b)。CRF6和CTK在胁迫响应调控中的作用相反, 但在叶片衰老调控中的作用相似, 推测CRF6也可能通过2条途径调控CTK信号通路。目前有关CRF调控的具体机制仍不明确, 鉴定CRF靶基因和上游信号分子将有助于更好地理解CTK响应非生物胁迫的作用机制。
4.2.5 参与油菜素内酯介导的胁迫应答
油菜素内酯(BR)在植物细胞伸长、叶片发育、花粉管生长、木质部分化、衰老、光形态发生以及应激反应过程中起重要作用(Ye et al., 2017)。BR信号始于细胞膜上受体激酶(brassinosteroid insensitive, BRI), BKI1 (BRI kinase inhibitor 1)是BRI1的关键负调控蛋白。无BR时, BKI1和14-3-3蛋白分别抑制BR的受体BRI1和转录因子BES1 (bri1-ems-suppressor 1) (Wang et al., 2011)。而当BR被BRI1感知后, 即促使BKI1发生磷酸化, 激活BRI1和BES1以调控负调节因子BIN2 (brassinosteroid insensitive 2)的表达, 诱导BES1和BZR1 (brassinazole-resistant 1)积累并与转录因子互作, 影响参与植物生长和胁迫反应的BR应答基因的转录水平(Wang et al., 2011; Guo et al., 2013)。
BZR1可结合CBF1和CBF2的启动子区并促进两者在冷胁迫下的表达。bin2-3、bil1、bil2缺失突变体在持续的冷胁迫下可以诱导去磷酸化BZR1的积累, BZR1通过调控WKRY6、PYL6、SOC1、JMT和SAG21等不依赖CBF途径的COR基因而增强拟南芥的耐冷性(Li et al., 2017)。研究发现BR可与干旱诱导相关转录因子RD26互作, 拮抗干旱响应基因的表达, 进而负调控植物的抗旱性(Ye et al., 2017)。BR与ABA信号途径中的受体、转录因子之间也产生拮抗效应, 进而调节胁迫下植物体的生长平衡(Nolan et al., 2017)。目前已从BES1和BZR1靶基因中鉴定出一些AP2/ERF类转录因子, 暗示AP2/ERF具有整合BR信号通路与非生物胁迫响应的潜在功能(Sun et al., 2010; Yu et al., 2011)。ERF72可能是研究BR信号通路与胁迫响应交叉互作的候选基因。在拟南芥中, ERF72/RAP2.3拮抗BZR1和ARF6 (auxin responsive factor 6), 抑制下胚轴伸长, 此过程中BR信号途径如何与胁迫响应调控网络协同互作发挥作用尚不清楚(Liu et al., 2018b)。研究表明, SUB1A参与调节GA和BR的交叉网络互作(Schmitz et al., 2013)。GA和BR途径通过DELLA蛋白、BZR1等转录因子协同调控植物的生长(Oh et al., 2012)。在植物淹水过程中SUB1A差异调节BR合成相关基因的表达, 激活BR生物合成和信号转导, 诱导GA降解关键基因GA2ox7的表达, 进而控制水稻植株体内的GA水平, 促进DELLA蛋白的积累(Schmitz et al., 2013)。因此, 推测AP2/ERF可能通过不同的机制参与调控BR信号通路。
AbbasM, BerckhanS, RooneyDJ, GibbsDJ, CondeVJ, CorreiaCS, BasseGW, de la RosaMN, LeonJ, AlabadíD, BlázquezMA, HoldsworthMJ (2015). Oxygen sensing coordinates photomorphogenesis to facilitate seedling survival Curr Biol 25, 1483-1488. URLPMID:25981794 [本文引用: 1]
AbiriR, ShaharuddinNA, MaziahM, YusofBZN, AtabakiN, SahebiM, ValdianiA, KalhoriN, AziziP, HanafiMM (2017). Role of ethylene and the APETALA 2/ethylene response factor superfamily in rice under various abiotic and biotic stress conditions Environ Exp Bot 134, 33-44. [本文引用: 1]
AchardP, GongF, CheminantS, AliouaM, HeddenP, GenschikP (2008). The cold-inducible CBF1 factor-de-pendent signaling pathway modulates the accumulation of the growth-repressing DELLA proteins via its effect on gibberellin metabolism Plant Cell 20, 2117-2129. DOI:10.1105/tpc.108.058941URLPMID:18757556 [本文引用: 1] Plants have evolved robust mechanisms to respond and adapt to unfavorable environmental conditions, such as low temperature. The C-repeat/drought-responsive element binding factor CBF1/DREB1b gene encodes a transcriptional activator transiently induced by cold that controls the expression of a set of genes responding to low temperature (the CBF regulon). Constitutive expression of CBF1 confers freezing tolerance but also slows growth. Here, we propose that low temperature-induced CBF1 expression restrains growth at least in part by allowing the accumulation of DELLAs, a family of nuclear growth-repressing proteins, the degradation of which is stimulated by gibberellin (GA). We show that cold/CBF1 enhances the accumulation of a green fluorescent protein (GFP)-tagged DELLA protein (GFP-RGA) by reducing GA content through stimulating expression of GA-inactivating GA 2-oxidase genes. Accordingly, transgenic plants that constitutively express CBF1 accumulate less bioactive GA and as a consequence exhibit dwarfism and late flowering. Both phenotypes are suppressed when CBF1 is expressed in a line lacking two DELLA proteins, GA-INSENSITIVE and REPRESSOR OF GA1-3. In addition, we show that DELLAs contribute significantly to CBF1-induced cold acclimation and freezing tolerance by a mechanism that is distinct from the CBF regulon. We conclude that DELLAs are components of the CBF1-mediated cold stress response.
AgarwalM, HaoYJ, KapoorA, DongCH, FujiiH, ZhengXW, ZhuJK (2006). A R2R3 type MYB transcription factor is involved in the cold regulation of CBF genes and in acquired freezing tolerance J Biol Chem 281, 37636-37645. URLPMID:17015446 [本文引用: 1]
Asensi-FabadoMA, AmtmannA, PerrellaG (2017). Plant responses to abiotic stress: the chromatin context of transcriptional regulation Biochim Biophys Acta 1860, 106-122. [本文引用: 1]
AyanoM, KaniT, KojimaM, SakakibaraH, KitaokaT, KurohaT, Angeles-ShimRB, KitanoH, NagaiK, AshikariM (2014). Gibberellin biosynthesis and signal transduction is essential for internode elongation in deepwater rice Plant Cell Environ 37, 2313-2324. DOI:10.1111/pce.12377URLPMID:24891164 [本文引用: 1] Under flooded conditions, the leaves and internodes of deepwater rice can elongate above the water surface to capture oxygen and prevent drowning. Our previous studies showed that three major quantitative trait loci (QTL) regulate deepwater-dependent internode elongation in deepwater rice. In this study, we investigated the age-dependent internode elongation in deepwater rice. We also investigated the relationship between deepwater-dependent internode elongation and the phytohormone gibberellin (GA) by physiological and genetic approach using a QTL pyramiding line (NIL-1 + 3 + 12). Deepwater rice did not show internode elongation before the sixth leaf stage under deepwater condition. Additionally, deepwater-dependent internode elongation occurred on the sixth and seventh internodes during the sixth leaf stage. These results indicate that deepwater rice could not start internode elongation until the sixth leaf stage. Ultra-performance liquid chromatography tandem mass-spectrometry (UPLC-MS/MS) method for the phytohormone contents showed a deepwater-dependent GA1 and GA4 accumulation in deepwater rice. Additionally, a GA inhibitor abolished deepwater-dependent internode elongation in deepwater rice. On the contrary, GA feeding mimicked internode elongation under ordinary growth conditions. However, mutations in GA biosynthesis and signal transduction genes blocked deepwater-dependent internode elongation. These data suggested that GA biosynthesis and signal transduction are essential for deepwater-dependent internode elongation in deepwater rice.
BossiF, CordobaE, DupréP, MendozaMS, RománCS, LeónP (2009). The Arabidopsis ABA-INSENSITIVE (ABI) 4 factor acts as a central transcription activator of the expression of its own gene, and for the induction of ABI5 and SBE2.2 genes during sugar signaling Plant J 59, 359-374. DOI:10.1111/j.1365-313X.2009.03877.xURLPMID:19392689 [本文引用: 1] The transcription factor ABA INSENSITIVE 4 (ABI4), discovered nearly 10 years ago, plays a central role in a variety of functions in plants, including sugar responses. However, not until very recently has its mechanism of action begun to be elucidated. Modulating gene expression is one of the primary mechanisms of sugar regulation in plants. Nevertheless, the transcription factors involved in regulating sugar responses and their role(s) during the signal transduction cascade remain poorly defined. In this paper we analyzed the participation of ABI4, as it is one of the main transcription factors implicated in glucose signaling during early seedling development. Our studies show that ABI4 is an essential activator of its own expression during development, in ABA signaling and in sugar responses. It is also important for the glucose-mediated expression of the genes ABI5 and SBE2.2. We demonstrate that ABI4 binds directly to the promoter region of all three genes and activates their expression in vivo through at CE1-like element. Previous studies found that ABI4 also functions as a transcriptional repressor of sugar-regulated genes, therefore this transcription factor is a versatile protein with dual functions for modulating gene expression.
BuiLT, GiuntoliB, KosmaczM, ParlantiS, LicausiF (2015). Constitutively expressed ERF-VII transcription factors redundantly activate the core anaerobic response in Arabidopsis thaliana Plant Sci 236, 37-43. DOI:10.1016/j.plantsci.2015.03.008URLPMID:26025519 [本文引用: 1] Plant adaptation to hypoxic conditions is mediated by the transcriptional activation of genes involved in the metabolic reprogramming of plant cells to cope with reduced oxygen availability. Recent studies indicated that members of the group VII of the Ethylene Responsive Transcription Factor (ERFs) family act as positive regulators of this molecular response. In the current study, the five ERF-VII transcription factors of Arabidopsis thaliana were compared to infer a hierarchy in their role with respect to the anaerobic response. When the activity of each transcription factor was tested on a set of hypoxia-responsive promoters, RAP2.2, RAP2.3 and RAP2.12 appeared to be the most powerful activators. RAP2.12 was further dissected in transactivation assays in Arabidopsis protoplasts to identify responsible regions for transcriptional activation. An ultimate C-terminal motif was identified as sufficient to drive gene transcription. Finally, using realtime RT-PCR in single and double mutants for the corresponding genes, we confirmed that RAP2.2 and RAP2.12 exert major control upon the anaerobic response.
CausierB, AshworthM, GuoWJ, DaviesB (2012). The TOPLESS interactome: a framework for gene repression in Arabidopsis Plant Physiol 158, 423-438. DOI:10.1104/pp.111.186999URLPMID:22065421 [本文引用: 1] Transcription factors activate or repress target gene expression or switch between activation and repression. In animals and yeast, Groucho/Tup1 corepressor proteins are recruited by diverse transcription factors to induce context-specific transcriptional repression. Two groups of Groucho/Tup1-like corepressors have been described in plants. LEUNIG and LEUNIG_HOMOLOG constitute one group and TOPLESS (TPL) and the four TPL-related (TPR) corepressors form the other. To discover the processes in which TPL and the TPR corepressors operate, high-throughput yeast two-hybrid approaches were used to identify interacting proteins. We found that TPL/TPR corepressors predominantly interact directly with specific transcription factors, many of which were previously implicated in transcriptional repression. The interacting transcription factors reveal that the TPL/TPR family has been coopted multiple times to modulate gene expression in diverse processes, including hormone signaling, stress responses, and the control of flowering time, for which we also show biological validation. The interaction data suggest novel mechanisms for the involvement of TPL/TPR corepressors in auxin and jasmonic acid signaling. A number of short repression domain (RD) sequences have previously been identified in Arabidopsis (Arabidopsis thaliana) transcription factors. All known RD sequences were enriched among the TPL/TPR interactors, and novel TPL-RD interactions were identified. We show that the presence of RD sequences is essential for TPL/TPR recruitment. These data provide a framework for TPL/TPR-dependent transcriptional repression. They allow for predictions about new repressive transcription factors, corepressor interactions, and repression mechanisms and identify a wide range of plant processes that utilize TPL/TPR-mediated gene repression.
ChandlerJW (2018). Class VIIIb APETALA2 ethylene response factors in plant development Trends Plant Sci 23, 151-162. DOI:10.1016/j.tplants.2017.09.016URLPMID:29074232 [本文引用: 1] The APETALA2 (AP2) transcription factor superfamily in many plant species is extremely large. In addition to well-documented roles in stress responses, some AP2 members in arabidopsis, such as those of subgroup VIIIb, which includes DORNROSCHEN, DORNROSCHEN-LIKE, PUCHI, and LEAFY PETIOLE, are also important developmental regulators throughout the plant life cycle. Information is accumulating from orthologs of these proteins in important crop species that they influence key agronomic traits, such as the release of bud-burst in woody perennials and floral meristem identity and branching in cereals, and thereby represent potential for agronomic improvement. Given the increasing recognition of their developmental significance, this review highlights the function of these proteins and addresses their phylogenetic and evolutionary relationships.
ChenH, JeJ, SongC, HwangJE, LimCO (2012). A proximal promoter region of Arabidopsis DREB2C confers tissue-speci?c expression under heat stress J Integr Plant Biol 54, 640-651. URLPMID:22716647 [本文引用: 2]
ChenHY, HsiehEJ, ChengMC, ChenYC, HwangYS, LinTP (2016). ORA47 (octadecanoid-responsive AP2/ERF- domain transcription factor 47) regulates jasmonic acid and abscisic acid biosynthesis and signaling through binding to a novel cis-element New Phytol 211, 599-613. URLPMID:26974851 [本文引用: 2]
ChengMC, HsiehEJ, ChenJH, ChenHY, LinTP (2012). Arabidopsis RGLG2, functioning as a RING E3 ligase, interacts with AtERF53 and negatively regulates the plant drought stress response Plant Physiol 158, 363-375. DOI:10.1104/pp.111.189738URLPMID:22095047 [本文引用: 1] Transcriptional activities of plants play important roles in responses to environmental stresses. ETHYLENE RESPONSE FACTOR53 (AtERF53) is a drought-induced transcription factor that belongs to the AP2/ERF superfamily and has a highly conserved AP2 domain. It can regulate drought-responsive gene expression by binding to the GCC box and/or the dehydration-responsive element in the promoter of downstream genes. Overexpression of AtERF53 driven by the cauliflower mosaic virus 35S promoter resulted in an unstable drought-tolerant phenotype in T2 transgenic Arabidopsis (Arabidopsis thaliana) plants. Using a yeast two-hybrid screen, we identified a RING domain ubiquitin E3 ligase, RGLG2, which interacts with AtERF53 in the nucleus. The copine domain of RGLG2 exhibited the strongest interacting activity. We also demonstrated that RGLG2 could move from the plasma membrane to the nucleus under stress treatment. Using an in vitro ubiquitination assay, RGLG2 and its closest sequelog, RGLG1, were shown to have E3 ligase activity and mediated AtERF53 ubiquitination for proteasome degradation. The rglg1rglg2 double mutant but not the rglg2 or rglg1 single mutant exhibited a drought-tolerant phenotype when compared with wild-type plants. AtERF53-green fluorescent proteins expressed in the rglg1rglg2 double mutants were stable. The 35S:AtERF53-green fluorescent protein/rglg1rglg2 showed enhanced AtERF53-regulated gene expression and had greater tolerance to drought stress than the rglg1rglg2 double mutant. In conclusion, RGLG2 negatively regulates the drought stress response by mediating AtERF53 transcriptional activity in Arabidopsis.
ChinnusamyV, ZhuJH, ZhuJK (2007). Cold stress regulation of gene expression in plants Trends Plant Sci 12, 444-451. URLPMID:17855156 [本文引用: 1]
ClaeysH, SkiryczA, MaleuxK, InzéD (2012). DELLA signaling mediates stress-induced cell differentiation in Arabidopsis leaves through modulation of anaphasepromoting complex/cyclosome activity Plant Physiol 159, 739-747. DOI:10.1104/pp.112.195032URLPMID:22535421 [本文引用: 1] Drought is responsible for considerable yield losses in agriculture due to its detrimental effects on growth. Drought responses have been extensively studied, but mostly on the level of complete plants or mature tissues. However, stress responses were shown to be highly tissue and developmental stage specific, and dividing tissues have developed unique mechanisms to respond to stress. Previously, we studied the effects of osmotic stress on dividing leaf cells in Arabidopsis (Arabidopsis thaliana) and found that stress causes early mitotic exit, in which cells end their mitotic division and start endoreduplication earlier. In this study, we analyzed this phenomenon in more detail. Osmotic stress induces changes in gibberellin metabolism, resulting in the stabilization of DELLAs, which are responsible for mitotic exit and earlier onset of endoreduplication. Consequently, this response is absent in mutants with altered gibberellin levels or DELLA activity. Mitotic exit and onset of endoreduplication do not correlate with an up-regulation of known cell cycle inhibitors but are the result of reduced levels of DP-E2F-LIKE1/E2Fe and UV-B-INSENSITIVE4, both inhibitors of the developmental transition from mitosis to endoreduplication by modulating anaphase-promoting complex/cyclosome activity, which are down-regulated rapidly after DELLA stabilization. This work fits into an emerging view of DELLAs as regulators of cell division by regulating the transition to endoreduplication and differentiation.
ColebrookEH, ThomasSG, PhillipsAL, HeddenP (2014). The role of gibberellin signaling in plant responses to abiotic stress J Exp Biol 217, 67-75. DOI:10.1242/jeb.089938URLPMID:24353205 [本文引用: 2] Plant hormones are small molecules that regulate plant growth and development, as well as responses to changing environmental conditions. By modifying the production, distribution or signal transduction of these hormones, plants are able to regulate and coordinate both growth and/or stress tolerance to promote survival or escape from environmental stress. A central role for the gibberellin (GA) class of growth hormones in the response to abiotic stress is becoming increasingly evident. Reduction of GA levels and signalling has been shown to contribute to plant growth restriction on exposure to several stresses, including cold, salt and osmotic stress. Conversely, increased GA biosynthesis and signalling promote growth in plant escape responses to shading and submergence. In several cases, GA signalling has also been linked to stress tolerance. The transcriptional regulation of GA metabolism appears to be a major point of regulation of the GA pathway, while emerging evidence for interaction of the GA-signalling molecule DELLA with components of the signalling pathway for the stress hormone jasmonic acid suggests additional mechanisms by which GA signalling may integrate multiple hormone signalling pathways in the response to stress. Here, we review the evidence for the role of GA in these processes, and the regulation of the GA signalling pathway on exposure to abiotic stress. The potential mechanisms by which GA signalling modulates stress tolerance are also discussed.
DeepikaL, AshutoshP, YogeshwarVD, SumitKB, PrabodhKT, MeharHA (2016). Genome-wide analysis of the AP2/ERF family in Musa species reveals divergence and neofunctionalisation during evolution Sci Rep 6, 18878. URLPMID:26733055 [本文引用: 1]
DongCJ, LiuJY (2010). The Arabidopsis EAR-motif-containing protein RAP2.1 functions as an active transcriptional repressor to keep stress responses under tight control BMC Plant Biol 10, 47. DOI:10.1186/1471-2229-10-47URLPMID:20230648 [本文引用: 2] BACKGROUND: Plants respond to abiotic stress through complex regulation of transcription, including both transcriptional activation and repression. Dehydration-responsive-element binding protein (DREB)-type transcription factors are well known to play important roles in adaptation to abiotic stress. The mechanisms by which DREB-type transcription factors activate stress-induced gene expression have been relatively well studied. However, little is known about how DREB-type transcriptional repressors modulate plant stress responses. In this study, we report the functional analysis of RAP2.1, a DREB-type transcriptional repressor. RESULTS: RAP2.1 possesses an APETALA2 (AP2) domain that binds to dehydration-responsive elements (DREs) and an ERF-associated amphiphilic repression (EAR) motif, as the repression domain located at the C-terminus of the protein. Expression of RAP2.1 is strongly induced by drought and cold stress via an ABA-independent pathway. Arabidopsis plants overexpressing RAP2.1 show enhanced sensitivity to cold and drought stresses, while rap2.1-1 and rap2.1-2 T-DNA insertion alleles result in reduced sensitivity to these stresses. The reduced stress sensitivity of the plant containing the rap2.1 allele can be genetically complemented by the expression of RAP2.1, but not by the expression of EAR-motif-mutated RAP2.1. Furthermore, chromatin immunoprecipitation (ChIP) analysis has identified Responsive to desiccation/Cold-regulated (RD/COR) genes as downstream targets of RAP2.1 in vivo. Stress-induced expression of the RD/COR genes is repressed by overexpression of RAP2.1 and is increased in plants expressing the rap2.1 allele. In addition, RAP2.1 can negatively regulate its own expression by binding to DREs present in its own promoter. Our data suggest that RAP2.1 acts as a negative transcriptional regulator in defence responses to cold and drought stress in Arabidopsis. CONCLUSIONS: A hypothetical model for the role of RAP2.1 in modulating plant responses to cold and drought is proposed in this study. It appears that RAP2.1 acts as a negative
DouHO, XvKP, MengQW, LiG, YangXH (2015). Potato plants ectopically expressing Arabidopsis thaliana CBF3 exhibit enhanced tolerance to high-temperature stress Plant Cell Environ 38, 61-72. DOI:10.1111/pce.12366URLPMID:24811248 [本文引用: 1] CBF3, a known cold-inducible gene that encodes a transcription factor, was isolated from Arabidopsis thaliana and introduced into the potato (Solanum tuberosum cv. 'luyin NO.1') under the control of the CaMV35S promoter or the rd29A promoter. Our results revealed that temperature of 40 degrees C or higher can significantly induce AtCBF3 expression. After heat stress, the net photosynthetic rate (Pn ), the maximal photochemical efficiency of photosystem II (PSII) (Fv /Fm ) and the accumulation of the D1 protein were higher in the transgenic lines than in the wild-type (WT) line. Moreover, compared with the WT line, O2 (-) and H2 O2 accumulation in the transgenic lines were reduced. A Q-PCR assay of a subset of the genes involved in photosynthesis and antioxidant defence further verified the above results. Interestingly, under heat stress conditions, the accumulation of heat-shock protein 70 (HSP70) increased in the WT line but decreased in the transgenic lines. These results suggest that potato plants ectopically expressing AtCBF3 exhibited enhanced tolerance to high temperature, which is associated with improved photosynthesis and antioxidant defence via induction of the expression of many stress-inducible genes. However, this mechanism may not depend upon the regulatory pathways in which HSP70 is involved.
DuboisM, SkiryczA, ClaeysH, MaleuxK, DhondtS, De BodtS, BosscheRV, De MildeL, YoshizumiT, MatsuiM, InzéD (2013). Ethylene Response Factor 6 acts as a central regulator of leaf growth under water-limiting conditions in Arabidopsis Plant Physiol 162, 319-332. DOI:10.1104/pp.113.216341URLPMID:23553636 [本文引用: 1] Leaf growth is a complex developmental process that is continuously fine-tuned by the environment. Various abiotic stresses, including mild drought stress, have been shown to inhibit leaf growth in Arabidopsis (Arabidopsis thaliana), but the underlying mechanisms remain largely unknown. Here, we identify the redundant Arabidopsis transcription factors ETHYLENE RESPONSE FACTOR5 (ERF5) and ERF6 as master regulators that adapt leaf growth to environmental changes. ERF5 and ERF6 gene expression is induced very rapidly and specifically in actively growing leaves after sudden exposure to osmotic stress that mimics mild drought. Subsequently, enhanced ERF6 expression inhibits cell proliferation and leaf growth by a process involving gibberellin and DELLA signaling. Using an ERF6-inducible overexpression line, we demonstrate that the gibberellin-degrading enzyme GIBBERELLIN 2-OXIDASE6 is transcriptionally induced by ERF6 and that, consequently, DELLA proteins are stabilized. As a result, ERF6 gain-of-function lines are dwarfed and hypersensitive to osmotic stress, while the growth of erf5erf6 loss-of-function mutants is less affected by stress. Besides its role in plant growth under stress, ERF6 also activates the expression of a plethora of osmotic stress-responsive genes, including the well-known stress tolerance genes STZ, MYB51, and WRKY33. Interestingly, activation of the stress tolerance genes by ERF6 occurs independently from the ERF6-mediated growth inhibition. Together, these data fit into a leaf growth regulatory model in which ERF5 and ERF6 form a missing link between the previously observed stress-induced 1-aminocyclopropane-1-carboxylic acid accumulation and DELLA-mediated cell cycle exit and execute a dual role by regulating both stress tolerance and growth inhibition.
DuboisM, Van den BroeckL, ClaeysH, Van VlierbergheK, MatsuiM, InzéD (2015). The ETHYLENE RESPONSE FACTORs ERF6 and ERF11 antagonistically regulate mannitol-induced growth inhibition in Arabidopsis Plant Physiol 169, 166-179. DOI:10.1104/pp.15.00335URLPMID:25995327 [本文引用: 1] Leaf growth is a tightly regulated and complex process, which responds in a dynamic manner to changing environmental conditions, but the mechanisms that reduce growth under adverse conditions are rather poorly understood. We previously identified a growth inhibitory pathway regulating leaf growth upon exposure to a low concentration of mannitol and characterized the ETHYLENE RESPONSE FACTOR (ERF)/APETALA2 transcription factor ERF6 as a central activator of both leaf growth inhibition and induction of stress tolerance genes. Here, we describe the role of the transcriptional repressor ERF11 in relation to the ERF6-mediated stress response in Arabidopsis (Arabidopsis thaliana). Using inducible overexpression lines, we show that ERF6 induces the expression of ERF11. ERF11 in turn molecularly counteracts the action of ERF6 and represses at least some of the ERF6-induced genes by directly competing for the target gene promoters. As a phenotypical consequence of the ERF6-ERF11 antagonism, the extreme dwarfism caused by ERF6 overexpression is suppressed by overexpression of ERF11. Together, our data demonstrate that dynamic mechanisms exist to fine-tune the stress response and that ERF11 counteracts ERF6 to maintain a balance between plant growth and stress defense.
FengCZ, ChenY, WangC, KongYH, WuWH, ChenYF (2014). Arabidopsis RAV1 transcription factor, phosphorylated by SnRK2 kinases, regulates the expression of ABI3, ABI4, and ABI5 during seed germination and early seedling development Plant J 80, 654-668. DOI:10.1111/tpj.12670URL [本文引用: 2] The phytohormone abscisic acid (ABA) modulates a number of processes during plant growth and development. In this study, the molecular mechanism of Arabidopsis RAV (Related to ABI3/VP1) transcription factor RAV1 involving ABA signaling was investigated. RAV1-underexpressing lines were more sensitive to ABA than wild-type plants during seed germination and early seedling development, whereas RAV1-overexpressing lines showed strong ABA-insensitive phenotypes. Overexpression of RAV1 repressed ABI3, ABI4, and ABI5 expression, and RAV1 bound to the ABI3, ABI4, and ABI5 promoters in vitro and in vivo, indicating that RAV1 directly down-regulates the expression of ABI3, ABI4, and ABI5. The interruption of ABI5 function in RAV1-U abi5 plants abolished the ABA-hypersensitive phenotype of RAV1-U plants, demonstrating that ABI5 is epistatic to RAV1. RAV1 interacted with SNF1-RELATED PROTEIN KINASE SnRK2.2, SnRK2.3 and SnRK2.6 in the nucleus. In vitro kinase assays showed that SnRK2.2, SnRK2.3 and SnRK2.6 phosphorylated RAV1. Transient expression assays revealed that SnRK2.2, SnRK2.3 and SnRK2.6 reduced the RAV1-dependent repression of ABI5, and the ABA-insensitive phenotype of the RAV1-overexpressing line was impaired by overexpression of SnRK2.3 in the RAV1 OE3 plants. Together, these results demonstrated that the Arabidopsis RAV1 transcription factor plays an important role in ABA signaling by modulating the expression of ABI3, ABI4, and ABI5, and that its activity is negatively affected by SnRK2s.
FinkelsteinR (2013). Abscisic acid synthesis and response Arabidopsis Book 11, e0166. URLPMID:24273463 [本文引用: 1]
FowlerS, ThomashowMF (2002). Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway Plant Cell 14, 1675-1690. URLPMID:12172015 [本文引用: 1]
Franco-ZorrillaJM, López-VidrieroI, CarrascoJL, GodoyM, VeraP, SolanoR (2014). DNA-binding specificities of plant transcription factors and their potential to define target genes Proc Natl Acad Sci USA 111, 2367-2372. DOI:10.1073/pnas.1316278111URLPMID:24477691 [本文引用: 4] Transcription factors (TFs) regulate gene expression through binding to cis-regulatory specific sequences in the promoters of their target genes. In contrast to the genetic code, the transcriptional regulatory code is far from being deciphered and is determined by sequence specificity of TFs, combinatorial cooperation between TFs and chromatin competence. Here we addressed one of these determinants by characterizing the target sequence specificity of 63 plant TFs representing 25 families, using protein-binding microarrays. Remarkably, almost half of these TFs recognized secondary motifs, which in some cases were completely unrelated to the primary element. Analyses of coregulated genes and transcriptomic data from TFs mutants showed the functional significance of over 80% of all identified sequences and of at least one target sequence per TF. Moreover, combining the target sequence information with coexpression analysis we could predict the function of a TF as activator or repressor through a particular DNA sequence. Our data support the correlation between cis-regulatory elements and the sequence determined in vitro using the protein-binding microarray and provides a framework to explore regulatory networks in plants.
FuMJ, KangHK, SonSH, KimSK, NamKH (2014). A subset of Arabidopsis RAV transcription factors modulates drought and salt stress responses independent of ABA Plant Cell Physiol 55, 1892-1904. DOI:10.1093/pcp/pcu118URLPMID:25189341 [本文引用: 2] Arabidopsis RAV1, RAV1L and RAV2/TEM2 are Related to ABI3/VP1 (RAV) transcription factors that contain both plant-specific B3 and AP2 domains. RAV1 was known to be a negative regulator of growth and its transcript level was repressed by brassinolide (BL). In this study, we found that the expressions of RAV1, and its closest homologs RAV1L and RAV2 were also regulated by other plant hormones, and especially repressed significantly by BL and abscisic acid (ABA), which mediate various abiotic stress responses in plants. Therefore, to further investigate the physiological functions of RAV1, RAV1L and RAV2 in abiotic stress responses, we isolated T-DNA insertional knockout mutants of each gene and produced transgenic plants overexpressing the RAVs. Under normal conditions, each single mutant showed slightly promoted growth patterns only at an early stage of development. In comparison, the RAV1-overexpressing plants exhibited strong growth retardation with semi-dwarfed stature. In drought conditions, RAVs-overexpressing transgenic plants exhibited higher transpirational water loss than the wild type. In salt conditions, seed germination of the RAVs-overexpressing transgenic plants was more inhibited than that of the wild type, while ravs mutants showed promoted seed germination. We also found that RAVs expressions were reduced by dryness and salt. RAV1-overexpressing plants showed the same patterns of increased expression as stress-inducible genes such as RD29A, RD29B and the genes encoding ABA biosynthetic enzymes, as did the wild type and rav1 mutant. However, the RAV1-overexpressing transgenic plants were insensitive to ABA, regardless of the higher accumulation of ABA even in normal conditions. Taken together, these results suggest that RAVs are versatile negative regulators for growth and abiotic stresses, drought and salt, and that negative regulatory effects of RAVs on abiotic stresses are likely to be operated independently of ABA.
FukaoT, Bailey-SerresJ (2008). Submergence tolerance conferred by Sub1A is mediated by SLR1 and SLRL1 restriction of gibberellin responses in rice Proc Natl Acad Sci USA 105, 16814-16819. DOI:10.1073/pnas.0807821105URLPMID:18936491 [本文引用: 1] Submergence-tolerant rice maintains viability during complete submergence by limiting underwater elongation until floodwaters recede. Acclimation responses to submergence are coordinated by the submergence-inducible Sub1A, which encodes an ethylene-responsive factor-type transcription factor (ERF). Sub1A is limited to tolerant genotypes and sufficient to confer submergence tolerance to intolerant accessions. Here we evaluated the role of Sub1A in the integration of ethylene, abscisic acid (ABA), and gibberellin (GA) signaling during submergence. The submergence-stimulated decrease in ABA content was Sub1A-independent, whereas GA-mediated underwater elongation was significantly restricted by Sub1A. Transgenics that ectopically express Sub1A displayed classical GA-insensitive phenotypes, leading to the hypothesis that Sub1A limits the response to GA. Notably Sub1A increased the accumulation of the GA signaling repressors Slender Rice-1 (SLR1) and SLR1 Like-1 (SLRL1) and concomitantly diminished GA-inducible gene expression under submerged conditions. In the Sub1A overexpression line, SLR1 protein levels declined under prolonged submergence but were accompanied by an increase in accumulation of SLRL1, which lacks the DELLA domain. In the presence of Sub1A, the increase in these GA signaling repressors and decrease in GA responsiveness were stimulated by ethylene, which promotes Sub1A expression. Conversely, ethylene promoted GA responsiveness and shoot elongation in submergence-intolerant lines. Together, these results demonstrate that Sub1A limits ethylene-promoted GA responsiveness during submergence by augmenting accumulation of the GA signaling repressors SLR1 and SLRL1.
FursovaOV, PogorelkoGV, TarasovVA (2009). Identification of ICE2, a gene involved in cold acclimation which determines freezing tolerance in Arabidopsis thaliana Gene 429, 98-103. URLPMID:19026725
GaschP, FundingerM, MüllerJT, LeeT, Bailey-SerresJ, MustrophA (2016). Redundant ERF-VII transcription factors bind to an evolutionarily conserved cis-motif to regulate hypoxia-responsive gene expression in Arabidopsis Plant Cell 28, 160-180. URLPMID:26668304 [本文引用: 1]
GibbsDJ, CondeJV, BerckhanS, PrasadG, MendiondoGM, HoldsworthMJ (2015). Group VII ethylene response factors coordinate oxygen and nitric oxide signal transduction and stress responses in plants Plant Physiol 169, 23-31. URLPMID:25944828 [本文引用: 5]
HattoriY, NagaiK, FurukawaS, SongXJ, KawanoR, SakakibaraH, WuJZ, MatsumotoT, YoshimuraA, KitanoH, MatsuokaM, MoriH, AshikariM (2009). The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water Nature 460, 1026-1030. URLPMID:19693083 [本文引用: 2]
HinzM, WilsonIW, YangJ, BuerstenbinderK, LlewellynD, DennisES, SauterM, DolferusR (2010). Arabidopsis RAP2.2: an ethylene response transcription factor that is important for hypoxia survival Plant Physiol 153, 757-772. DOI:10.1104/pp.110.155077URLPMID:20357136 [本文引用: 2] Arabidopsis (Arabidopsis thaliana) RAP2.2 (At3g14230) is an APETALA2/ethylene response factor-type transcription factor that belongs to the same subfamily as the rice (Oryza sativa) submergence tolerance gene SUB1A. RAP2.2 is expressed at constitutively high levels in the roots and at lower levels in the shoots, where it is induced by darkness. Effector studies and analysis of ethylene signal transduction mutants indicate that RAP2.2 is induced in shoots by ethylene and functions in an ethylene-controlled signal transduction pathway. Overexpression of RAP2.2 resulted in improved plant survival under hypoxia (low-oxygen) stress, whereas lines containing T-DNA knockouts of the gene had poorer survival rates than the wild type. This indicates that RAP2.2 is important in a plant's ability to resist hypoxia stress. Observation of the expression pattern of 32 low-oxygen and ethylene-associated genes showed that RAP2.2 affects only part of the low-oxygen response, particularly the induction of genes encoding sugar metabolism and fermentation pathway enzymes, as well as ethylene biosynthesis genes. Our results provide a new insight on the regulation of gene expression under low-oxygen conditions. Lighting plays an important regulatory role and is intertwined with hypoxia conditions; both stimuli may act collaboratively to regulate the hypoxic response.
HorstmanA, WillemsenV, BoutilierK, HeidstraR (2014). AINTEGUMENTA-LIKE proteins: hubs in a plethora of networks Trends Plant Sci 19, 146-157. DOI:10.1016/j.tplants.2013.10.010URLPMID:24280109 [本文引用: 1] Members of the AINTEGUMENTA-LIKE (AIL) family of APETALA 2/ETHYLENE RESPONSE FACTOR (AP2/ERF) domain transcription factors are expressed in all dividing tissues in the plant, where they have central roles in developmental processes such as embryogenesis, stem cell niche specification, meristem maintenance, organ positioning, and growth. When overexpressed, AIL proteins induce adventitious growth, including somatic embryogenesis and ectopic organ formation. The Arabidopsis (Arabidopsis thaliana) genome contains eight AIL genes, including AINTEGUMENTA, BABY BOOM, and the PLETHORA genes. Studies on these transcription factors have revealed their intricate relationship with auxin as well as their involvement in an increasing number of gene regulatory networks, in which extensive crosstalk and feedback loops have a major role.
HsiehEJ, ChengMC, LinTP (2013). Functional characterization of an abiotic stress-inducible transcription factor AtERF53 in Arabidopsis thaliana. Plant Mol Biol 82, 223-237. DOI:10.1007/s11103-013-0054-zURLPMID:23625358 [本文引用: 1] AP2/ERF proteins play crucial roles in plant growth and development and in responses to biotic and abiotic stresses. ETHYLENE RESPONSE FACTOR 53 (AtERF53) belongs to group 1 in the ERF family and is induced in the early hours of dehydration and salt treatment. The functional study of AtERF53 is hampered because its protein expression in Arabidopsis is vulnerable to degradation in overexpressed transgenic lines. Taking advantage of the RING domain ligase1/RING domain ligase2 (rglg1rglg2) double mutant in which the AtERF53 can express stably, we investigate the physiological function of AtERF53. In this study, we demonstrate that expression of AtERF53 in wild-type Arabidopsis was responsive to heat and abscisic acid (ABA) treatment. From results of the cotransfection experiment, we concluded that AtERF53 has positive transactivation activity. Overexpression of AtERF53 in the rglg1rglg2 double mutant conferred better heat-stress tolerance and had resulted in higher endogenous ABA and proline levels compared to rglg1rglg2 double mutants. AtERF53 also has a function to regulate guard-cell movement because the stomatal aperture of AtERF53 overexpressed in rglg1rglg2 double mutant was smaller than that in the rglg1rglg2 double mutant under ABA treatment. In a global gene expression study, we found higher expressions of many stress-related genes, such as DREB1A, COR15A, COR15B, PLC, P5CS1, cpHSC70 s and proline and ABA metabolic-related genes. Furthermore, we identified several downstream target genes of AtERF53 by chromatin immunoprecipitation assay. In conclusion, the genetic, molecular and biochemical result might explain how AtERF53 serving as a transcription factor contributes to abiotic stress tolerance in Arabidopsis.
IkedaM, Ohme-TakagiM (2009). A novel group of transcriptional repressors in Arabidopsis Plant Cell Physiol 50, 970-975. DOI:10.1093/pcp/pcp048URLPMID:19324928 [本文引用: 2] We showed previously that the ERF-associated amphiphilic repression (EAR) motif is a plant-specific repression domain that contains the conserved amino acid sequence LXLXL. In this report, we describe the identification of a novel repression domain, L/VR/KLFGVXM/V/L, which is different from known EAR motifs, in B3 DNA-binding domain transcription factors in Arabidopsis. Database analysis revealed that 29 Arabidopsis transcription factors, which included members of the RAV, ARF, Hsf and MYB families, contain the R/KLFGV conserved motif found in the novel repression domain. We demonstrated that factors that contain the R/KLFGV motif, namely, RAV1, RAV2, HsfB1 and HsfB2b, exhibited the repressive activity.
JeonJ, ChoC, LeeMR, van BinhN, KimJ (2016). CYTOKININ RESPONSE FACTOR 2 (CRF2) and CRF3 regulate lateral root development in response to cold stress in Arabidopsis Plant Cell 28, 1828-1843. URLPMID:27432872 [本文引用: 3]
JiaYX, DingYL, ShiYT, ZhangXY, GongZZ, YangSH (2016). The cbfs triple mutants reveal the essential functions of CBFs in cold acclimation and allow the definition of CBF regulons in Arabidopsis New Phytol 212, 345-353. URLPMID:27353960 [本文引用: 2]
KagaleS, RozwadowskiK (2011). EAR motif-mediated transcriptional repression in plants: an underlying mechanism for epigenetic regulation of gene expression Epigenetics 6, 141-146. DOI:10.4161/epi.6.2.13627URLPMID:20935498 [本文引用: 1] Ethylene-responsive element binding factor-associated Amphiphilic Repression (EAR) motif-mediated transcriptional repression is emerging as one of the principal mechanisms of plant gene regulation. The EAR motif, defined by the consensus sequence patterns of either LxLxL or DLNxxP, is the most predominant form of transcriptional repression motif so far identified in plants. Additionally, this active repression motif is highly conserved in transcriptional regulators known to function as negative regulators in a broad range of developmental and physiological processes across evolutionarily diverse plant species. Recent discoveries of co-repressors interacting with EAR motifs, such as TOPLESS (TPL) and AtSAP18, have begun to unravel the mechanisms of EAR motif-mediated repression. The demonstration of genetic interaction between mutants of TPL and AtHDA19, co-complex formation between TPL-related 1 (TPR1) and AtHDA19, as well as direct physical interaction between AtSAP18 and AtHDA19 support a model where EAR repressors, via recruitment of chromatin remodeling factors, facilitate epigenetic regulation of gene expression. Here, we discuss the biological significance of EAR-mediated gene regulation in the broader context of plant biology and present literature evidence in support of a model for EAR motif-mediated repression via the recruitment and action of chromatin modifiers. Additionally, we discuss the possible influences of phosphorylation and ubiquitination on the function and turnover of EAR repressors.
KarabaA, DixitS, GrecoR, AharoniA, TrijatmikoKR, Marsch-MartinezN, KrishnanA, NatarajaKN, UdayakumarM, PereiraA (2007). Improvement of water use efficiency in rice by expression of HARDY, an Arabidopsis drought and salt tolerance gene Proc Natl Acad Sci USA 104, 15270-15275. DOI:10.1073/pnas.0707294104URLPMID:17881564 [本文引用: 2] Freshwater is a limited and dwindling global resource; therefore, efficient water use is required for food crops that have high water demands, such as rice, or for the production of sustainable energy biomass. We show here that expression of the Arabidopsis HARDY (HRD) gene in rice improves water use efficiency, the ratio of biomass produced to the water used, by enhancing photosynthetic assimilation and reducing transpiration. These drought-tolerant, low-water-consuming rice plants exhibit increased shoot biomass under well irrigated conditions and an adaptive increase in root biomass under drought stress. The HRD gene, an AP2/ERF-like transcription factor, identified by a gain-of-function Arabidopsis mutant hrd-D having roots with enhanced strength, branching, and cortical cells, exhibits drought resistance and salt tolerance, accompanied by an enhancement in the expression of abiotic stress associated genes. HRD overexpression in Arabidopsis produces thicker leaves with more chloroplast-bearing mesophyll cells, and in rice, there is an increase in leaf biomass and bundle sheath cells that probably contributes to the enhanced photosynthesis assimilation and efficiency. The results exemplify application of a gene identified from the model plant Arabidopsis for the improvement of water use efficiency coincident with drought resistance in the crop plant rice.
KavasM, KizildoganA, G?kdemirG, BalogluMC (2015). Genome-wide investigation and expression analysis of AP2-ERF gene family in salt tolerant common bean EXCLI J 14, 1187-1206. URLPMID:27152109 [本文引用: 1]
KazanK (2015). Diverse roles of jasmonates and ethylene in abiotic stress tolerance Trends Plant Sci 20, 219-229. DOI:10.1016/j.tplants.2015.02.001URLPMID:25731753 [本文引用: 4] Jasmonates (JAs) and ethylene (ET), often acting cooperatively, play essential roles in regulating plant defense against pests and pathogens. Recent research reviewed here has revealed mechanistic new insights into the mode of action of these hormones in plant abiotic stress tolerance. During cold stress, JAs and ET differentially regulate the C-repeat binding factor (CBF) pathway. Major JA and ET signaling hubs such as JAZ proteins, CTR1, MYC2, components of the mediator complex, EIN2, EIN3, and several members of the AP2/ERF transcription factor gene family all have complex regulatory roles during abiotic stress adaptation. Better understanding the roles of these phytohormones in plant abiotic stress tolerance will contribute to the development of crop plants tolerant to a wide range of stressful environments.
KimJ (2016). CYTOKININ RESPONSE FACTORs gating environmental signals and hormones Trends Plant Sci 21, 993-996. DOI:10.1016/j.tplants.2016.10.004URLPMID:27773669 [本文引用: 2] CYTOKININ RESPONSE FACTORs (CRFs) encode transcription factors belonging to a small family within the APETALA2/ETHYLENE RESPONSIVE FACTOR (AP2/ERF) superfamily. Recent studies have revealed the biological functions of some arabidopsis CRFs, providing insight into the role of these plant transcription factors in integrating environmental and hormonal signals for plant adaptation.
KimJS, MizoiJ, YoshidaT, FujitaY, NakajimaJ, OhoriT, TodakaD, NakashimaK, HirayamaT, ShinozakiK, Yamaguchi-ShinozakiK (2011). An ABRE promoter sequence is involved in osmotic stress-responsive expression of the DREB2A gene, which encodes a transcription factor regulating drought-inducible genes in Arabidopsis Plant Cell Physiol 52, 2136-2146. DOI:10.1093/pcp/pcr143URLPMID:22025559 [本文引用: 1] In plants, osmotic stress-responsive transcriptional regulation depends mainly on two major classes of cis-acting elements found in the promoter regions of stress-inducible genes: ABA-responsive elements (ABREs) and dehydration-responsive elements (DREs). ABRE has been shown to perceive ABA-mediated osmotic stress signals, whereas DRE is known to be involved in an ABA-independent pathway. Previously, we reported that the transcription factor DRE-BINDING PROTEIN 2A (DREB2A) regulates DRE-mediated transcription of target genes under osmotic stress conditions in Arabidopsis (Arabidopsis thaliana). However, the transcriptional regulation of DREB2A itself remains largely uncharacterized. To elucidate the transcriptional mechanism associated with the DREB2A gene under osmotic stress conditions, we generated a series of truncated and base-substituted variants of the DREB2A promoter and evaluated their transcriptional activities individually. We found that both ABRE and coupling element 3 (CE3)-like sequences located approximately -100 bp from the transcriptional initiation site are necessary for the dehydration-responsive expression of DREB2A. Coupling our transient expression analyses with yeast one-hybrid and chromatin immunoprecipitation (ChIP) assays indicated that the ABRE-BINDING PROTEIN 1 (AREB1), AREB2 and ABRE-BINDING FACTOR 3 (ABF3) bZIP transcription factors can bind to and activate the DREB2A promoter in an ABRE-dependent manner. Exogenous ABA application induced only a modest accumulation of the DREB2A transcript when compared with the osmotic stress treatment. However, the osmotic stress-induced DREB2A expression was found to be markedly impaired in several ABA-deficient and ABA-insensitive mutants. These results suggest that in addition to an ABA-independent pathway, the ABA-dependent pathway plays a positive role in the osmotic stress-responsive expression of DREB2A.
KimNY, JangYJ, ParkOK (2018). AP2/ERF family transcription factors ORA59 and RAP2.3 interact in the nucleus and function together in ethylene responses Front Plant Sci 9, 1675. URLPMID:30510560 [本文引用: 1]
LeeSY, BoonNJ, WebbAAR, TanakaRJ (2016). Synergistic activation of RD29A via integration of salinity stress and abscisic acid in Arabidopsis thaliana Plant Cell Physiol 57, 2147-2160. DOI:10.1093/pcp/pcw132URLPMID:27497445 [本文引用: 1] Plants perceive information from the surroundings and elicit appropriate molecular responses. How plants dynamically respond to combinations of external inputs is yet to be revealed, despite the detailed current knowledge of intracellular signaling pathways. We measured dynamics of Response-to-Dehydration 29A (RD29A) expression induced by single or combined NaCl and ABA treatments in Arabidopsis thaliana. RD29A expression in response to a combination of NaCl and ABA leads to unique dynamic behavior that cannot be explained by the sum of responses to individual NaCl and ABA. To explore the potential mechanisms responsible for the observed synergistic response, we developed a mathematical model of the DREB2 and AREB pathways based on existing knowledge, where NaCl and ABA act as the cognate inputs, respectively, and examined various system structures with cross-input modulation, where non-cognate input affects expression of the genes involved in adjacent signaling pathways. The results from the analysis of system structures, combined with the insights from microarray expression profiles and model-guided experiments, predicted that synergistic activation of RD29A originates from enhancement of DREB2 activity by ABA. Our analysis of RD29A expression profiles demonstrates that a simple mathematical model can be used to extract information from temporal dynamics induced by combinatorial stimuli and produce experimentally testable hypotheses.
LeeSY, HwangEY, SeokHY, TarteVN, JeongMS, JangSB, MoonYH (2015). Arabidopsis AtERF71/HRE2 functions as transcriptional activator via cis-acting GCC box or DRE/CRT element and is involved in root development through regulation of root cell expansion Plant Cell Rep 34, 223-231. DOI:10.1007/s00299-014-1701-9URLPMID:25344007 [本文引用: 1] KEY MESSAGE: AtERF71/HRE2 binds to GCC box or DRE/CRT as transcription activator and plays an important role in root development via root cell expansion regulation. AtERF71/HRE2 transcription factor, a member of the AP2/ERF family, plays a key role in the stress response. GCC box and DRE/CRT, both essential cis-acting elements, have been shown to be recognized by AP2/ERF family transcription factors. However, it remains unclear whether or not AtERF71/HRE2 directly interacts with GCC box and/or DRE/CRT. Here, we showed that AtERF71/HRE2 binds to GCC box and DRE/CRT by electrophoretic mobility shift assay (EMSA). Binding of AtERF71/HRE2 to GCC box and DRE/CRT was also detected by fluorescence measurement and surface plasmon resonance spectroscopy (BIAcore) experiments. Folding properties of AtERF71/HRE2 proteins were characterized by CD spectroscopy, and AtERF71/HRE2 showed thermal stability as evidenced by two endothermic peaks (T d) at 53 and 65 degrees C. In addition, AtERF71/HRE2 showed transcriptional activation activity via GCC box and DRE/CRT in Arabidopsis protoplasts. Interestingly, AtERF71/HRE2 OXs showed increased primary root length due to elevated root cell expansion. Our data indicate that AtERF71/HRE2 binds to both GCC box and DRE/CRT, transactivates expression of genes downstream via GCC box or DRE/CRT, and plays an important role in root development through regulation of root cell expansion.
LiH, YeKY, ShiYT, ChengJK, ZhangXY, YangSH (2017). BZR1 positively regulates freezing tolerance via CBF-dependent and CBF-independent pathways in Arabidopsis Mol Plant 10, 545-559. DOI:10.1016/j.molp.2017.01.004URLPMID:28089951 [本文引用: 1] Cold stress is a major environmental factor that adversely affects plant growth and development. The C-repeat binding factor/DRE binding factor 1 (CBF/DREB1) transcriptional regulatory cascade has been shown to play important roles in plant response to cold. Here we demonstrate that two key components of brassinosteroid (BR) signaling modulate freezing tolerance of Arabidopsis plants. The loss-of-function mutant of the GSK3-like kinases involved in BR signaling, bin2-3 bil1 bil2, showed increased freezing tolerance, whereas overexpression of BIN2 resulted in hypersensitivity to freezing stress under both non-acclimated and acclimated conditions. By contrast, gain-of-function mutants of the transcription factors BZR1 and BES1 displayed enhanced freezing tolerance, and consistently cold treatment could induce the accumulation of dephosphorylated BZR1. Biochemical and genetic analyses showed that BZR1 acts upstream of CBF1 and CBF2 to directly regulate their expression. Moreover, we found that BZR1 also regulated other COR genes uncoupled with CBFs, such as WKRY6, PYL6, SOC1, JMT, and SAG21, to modulate plant response to cold stress. Consistently, wrky6 mutants showed decreased freezing tolerance. Taken together, our results indicate that BZR1 positively modulates plant freezing tolerance through CBF-dependent and CBF-independent pathways.
LiPC, HuangJG, YuSW, LiYY, SunP, WuCA, ZhengCC (2016). Arabidopsis YL1/BPG2 is involved in seedling shoot response to salt stress through ABI4 Sci Rep 6, 30163. DOI:10.1038/srep30163URLPMID:27444988 [本文引用: 1] The chloroplast-localized proteins play roles in plant salt stress response, but their mechanisms remain largely unknown. In this study, we screened a yellow leaf mutant, yl1-1, whose shoots exhibited hypersensitivity to salt stress. We mapped YL1 to AT3G57180, which encodes a YqeH-type GTPase. YL1, as a chloroplast stroma-localized protein, could be markedly reduced by high salinity. Upon exposure to high salinity, seedling shoots of yl1-1 and yl1-2 accumulated significantly higher levels of Na(+) than wild type. Expression analysis of factors involved in plant salt stress response showed that the expression of ABI4 was increased and HKT1 was evidently suppressed in mutant shoots compared with the wild type under normal growth conditions. Moreover, salinity effects on ABI4 and HKT1 were clearly weakened in the mutant shoots, suggesting that the loss of YL1 function impairs ABI4 and HKT1 expression. Notably, the shoots of yl1-2 abi4 double mutant exhibited stronger resistance to salt stress and accumulated less Na(+) levels after salt treatment compared with the yl1-2 single mutant, suggesting the salt-sensitive phenotype of yl1-2 seedlings could be rescued via loss of ABI4 function. These results reveal that YL1 is involved in the salt stress response of seedling shoots through ABI4.
LicausiF, Ohme-TakagiM, PerataP (2013). APETALA2/ Ethylene Responsive Factor (AP2/ERF) transcription factors: mediators of stress responses and developmental programs New Phytol 199, 639-649. URLPMID:24010138 [本文引用: 4]
LinRC, ParkHJ, WangHY (2008). Role of Arabidopsis RAP2.4 in regulating light and ethylene-mediated developmental processes and drought stress tolerance Mol Plant 1, 42-57. DOI:10.1093/mp/ssm004URLPMID:20031913 [本文引用: 1] Light and the plant hormone ethylene regulate many aspects of plant growth and development in an overlapping and interdependent fashion. Little is known regarding how their signal transduction pathways cross-talk to regulate plant development in a coordinated manner. Here, we report functional characterization of an AP2/DREB-type transcription factor, Arabidopsis RAP2.4, in mediating light and ethylene signaling. Expression of the RAP2.4 gene is down-regulated by light but up-regulated by salt and drought stresses. RAP2.4 protein is constitutively targeted to the nucleus and it can bind to both the ethylene-responsive GCC-box and the dehydration-responsive element (DRE). We show that RAP2.4 protein possesses an intrinsic transcriptional activation activity in yeast cells and that it can activate a reporter gene driven by the DRE cis-element in Arabidopsis protoplasts. Overexpression of RAP2.4 or mutation in RAP2.4 cause altered expression of representative light-, ethylene-, and drought-responsive genes. Although no salient phenotype was observed with a rap2.4 loss-of-function mutant, constitutive overexpression of RAP2.4 results in defects in multiple developmental processes regulated by light and ethylene, including hypocotyl elongation and gravitropism, apical hook formation and cotyledon expansion, flowering time, root elongation, root hair formation, and drought tolerance. Based on these observations, we propose that RAP2.4 acts at or downstream of a converging point of light and ethylene signaling pathways to coordinately regulate multiple developmental processes and stress responses.
LiuHC, LiaoHT, CharngYY (2011). The role of class A1 heat shock factors (HSFA1s) in response to heat and other stresses in Arabidopsis Plant Cell Environ 34, 738-751. DOI:10.1111/j.1365-3040.2011.02278.xURLPMID:21241330 [本文引用: 1] In Arabidopsis, there are four homologs of class A1 heat shock factor (HSFA1) genes, which likely encode the master regulators of heat shock response (HSR). However, previous studies with double knockout (KO) mutants were unable to confirm this point probably due to functional redundancy. Here, we generated a quadruple KO (QK) and four triple KO mutants to dissect their functions. Our data show that members of the HSFA1 group not only play a pivotal role in HSR but also are involved in growth and development. Alterations in morphology and retardation in growth were observed in the quadruple but not in triple KO mutants. The basal and acquired thermotolerance capacity was dramatically decreased in the QK mutant but varied in triple KO mutants at different developmental stages. The transcriptomics profiles suggested that more than 65% of the heat stress (HS)-up-regulated genes were HSFA1 dependent. HSFA1s were also involved in the expression of several HS genes induced by H(2) O(2) , salt and mannitol, which is consistent with the increased sensitive phenotype of the QK mutant to the stress factors. In conclusion, the Arabidopsis HSFA1s function as the master regulators of HSR and participate as important components in other abiotic stress responses as well.
LiuJY, ShiYT, YangSH (2018a). Insights into the regulation of C-repeat binding factors in plant cold signaling J Integr Plant Biol 60, 780-795. URLPMID:29667328 [本文引用: 1]
LiuK, LiYH, ChenXN, LiLJ, LiuK, ZhaoHP, WangYD, HanSC (2018b). ERF72 interacts with ARF6 and BZR1 to regulate hypocotyl elongation in Arabidopsis J Exp Bot 69, 3933-3947. DOI:10.1093/jxb/ery220URL [本文引用: 1]
LiuPQ, SunF, GaoR, DongHS (2012). RAP2.6L overexpression delays waterlogging induced premature senescence by increasing stomatal closure more than antioxidant enzyme activity Plant Mol Biol 79, 609-622. DOI:10.1007/s11103-012-9936-8URLPMID:22661072 [本文引用: 1] Waterlogging usually results from overuse or poor management of irrigation water and is a serious constraint due to its damaging effects. RAP2.6L (At5g13330) overexpression enhances plant resistance to jasmonic acid, salicylic acid, abscisic acid (ABA) and ethylene in Arabidopsis thaliana. However, it is not known whether RAP2.6L overexpression in vivo improves plant tolerance to waterlogging stress. In this study, the RAP2.6L transcript was induced by waterlogging or an ABA treatment, which was reduced after pretreatment with an ABA biosynthesis inhibitor tungstate. Water loss and membrane leakage were reduced in RAP2.6L overexpression plants under waterlogging stress. Time course analyses of ABA content and production of hydrogen peroxide (H(2)O(2)) showed that increased ABA precedes the increase of H(2)O(2). It is also followed by a marked increase in the antioxidant enzyme activities. Increased ABA promoted stomatal closure and made leaves exhibit a delayed waterlogging induced premature senescence. Furthermore, RAP2.6L overexpression caused significant increases in the transcripts of antioxidant enzyme genes APX1 (ascorbate peroxidase 1) and FSD1 (Fe-superoxide dismutase 1), the ABA biosynthesis gene ABA1 (ABA deficient 1) and signaling gene ABH1 (ABA-hypersensitive 1) and the waterlogging responsive gene ADH1 (alcohol dehydrogenase 1), while the transcript of ABI1 (ABA insensitive 1) was decreased. ABA inhibits seed germination and seedling growth and phenotype analysis showed that the integration of abi1-1 mutation into the RAP2.6L overexpression lines reduces ABA sensitivity. These suggest that RAP2.6L overexpression delays waterlogging induced premature senescence and might function through ABI1-mediated ABA signaling pathway.
LockeAM, Barding GAJr, SathnurS, LariveCK, Bailey-SerresJ (2018). Rice SUB1A constrains remodelling of the transcriptome and metabolome during submergence to facilitate post-submergence recovery Plant Cell Environ 41, 721-736. DOI:10.1111/pce.13094URLPMID:29094353 [本文引用: 2] The rice (Oryza sativa L.) ethylene-responsive transcription factor gene SUB1A-1 confers tolerance to prolonged, complete submergence by limiting underwater elongation growth. Upon desubmergence, SUB1A-1 genotypes rapidly recover photosynthetic function and recommence development towards flowering. The underpinnings of the transition from stress amelioration to the return to homeostasis are not well known. Here, transcriptomic and metabolomic analyses were conducted to identify mechanisms by which SUB1A improves physiological function over the 24 hr following a sublethal submergence event. Evaluation of near-isogenic genotypes after submergence and over a day of reaeration demonstrated that SUB1A transiently constrains the remodelling of cellular activities associated with growth. SUB1A influenced the abundance of ca. 1,400 transcripts and had a continued impact on metabolite content, particularly free amino acids, glucose, and sucrose, throughout the recovery period. SUB1A promoted recovery of metabolic homeostasis but had limited influence on mRNAs associated with growth processes and photosynthesis. The involvement of low energy sensing during submergence and recovery was supported by dynamics in trehalose-6-phosphate and mRNAs encoding key enzymes and signalling proteins, which were modulated by SUB1A. This study provides new evidence of convergent signalling pathways critical to the rapidly reversible management of carbon and nitrogen metabolism in submergence resilient rice.
MagomeH, YamaguchiS, HanadaA, KamiyaYJ, OdaKJ (2004). Dwarf and delayed-flowering 1, a novel Arabidopsis mutant deficient in gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor Plant J 37, 720-729. URLPMID:14871311 [本文引用: 1]
M?h?nenAP, HiguchiM, T?rm?kangasK, MiyawakiK, PischkeMS, SussmanMR, HelariuttaY, KakimotoT (2006). Cytokinins regulate a bidirectional phosphorelay network in Arabidopsis Curr Biol 16, 1116-1122. DOI:10.1016/j.cub.2006.04.030URLPMID:16753566 [本文引用: 1] The cytokinin class of plant hormones plays key roles in regulating diverse developmental and physiological processes. Arabidopsis perceives cytokinins with three related and partially redundant receptor histidine kinases (HKs): CRE1 (the same protein as WOL and AHK4), AHK2, and AHK3 (CRE-family receptors). It is suggested that binding of cytokinins induces autophosphorylation of these HKs and subsequent transfer of the phosphoryl group to a histidine phosphotransfer protein (HPt) and then to a response regulator (RR), ultimately regulating downstream signaling events. Here we demonstrate that, in vitro and in a yeast system, CRE1 is not only a kinase that phosphorylates HPts in the presence of cytokinin but is also a phosphatase that dephosphorylates HPts in the absence of cytokinin. To explore the roles of these activities in planta, we replaced CRE1 with mutant versions of the gene or with AHK2. Replacing CRE1 with CRE1(T278I), which lacks cytokinin binding activity and is locked in the phosphatase form, decreased cytokinin sensitivity. Conversely, replacing CRE1 with AHK2, which favors kinase activity, increased cytokinin sensitivity. These results indicate that in the presence of cytokinins, cytokinin receptors feed phosphate to phosphorelay-integrating HPt proteins. In the absence of cytokinins, CRE1 removes phosphate from HPt proteins, decreasing the system phosphoload.
MaruyamaK, TakedaM, KidokoroS, YamadaK, SakumaY, UranoK, FujitaM, YoshiwaraK, MatsukuraS, MorishitaY, SasakiR, SuzukiH, SaitoK, ShibataD, ShinozakiK, Yamaguchi-ShinozakiK (2009). Metabolic pathways involved in cold acclimation identified by integrated analysis of metabolites and transcripts regulated by DREB1A and DREB2A Plant Physiol 150, 1972-1980. DOI:10.1104/pp.109.135327URLPMID:19502356 [本文引用: 1] DREB1A/CBF3 and DREB2A are transcription factors that specifically interact with a cis-acting dehydration-responsive element (DRE), which is involved in cold- and dehydration-responsive gene expression in Arabidopsis (Arabidopsis thaliana). Overexpression of DREB1A improves stress tolerance to both freezing and dehydration in transgenic plants. In contrast, overexpression of an active form of DREB2A results in significant stress tolerance to dehydration but only slight tolerance to freezing in transgenic plants. The downstream gene products for DREB1A and DREB2A are reported to have similar putative functions, but downstream genes encoding enzymes for carbohydrate metabolism are very different between DREB1A and DREB2A. We demonstrate that under cold and dehydration conditions, the expression of many genes encoding starch-degrading enzymes, sucrose metabolism enzymes, and sugar alcohol synthases changes dynamically; consequently, many kinds of monosaccharides, disaccharides, trisaccharides, and sugar alcohols accumulate in Arabidopsis. We also show that DREB1A overexpression can cause almost the same changes in these metabolic processes and that these changes seem to improve freezing and dehydration stress tolerance in transgenic plants. In contrast, DREB2A overexpression did not increase the level of any of these metabolites in transgenic plants. Strong freezing stress tolerance of the transgenic plants overexpressing DREB1A may depend on accumulation of these metabolites.
MatsukuraS, MizoiJ, YoshidaT, TodakaD, ItoY, MaruyamaK, ShinozakiK, Yamaguchi-ShinozakiK (2010). Comprehensive analysis of rice DREB2-type genes that encode transcription factors involved in the expression of abiotic stress-responsive genes Mol Genet Genomics 283, 185-196. URLPMID:20049613 [本文引用: 2]
MengLS, WangZB, YaoSQ, LiuAZ (b). 2015 The ARF2- ANT-COR15A gene cascade regulates ABA-signaling- mediated resistance of large seeds to drought in Arabidopsis J Cell Sci 128, 3922-3932. [本文引用: 1]
MengXZ, XuJ, HeYX, YangKY, MordorskiB, LiuYD, ZhangSQ (2013). Phosphorylation of an ERF transcription factor by Arabidopsis MPK3/MPK6 regulates plant defense gene induction and fungal resistance Plant Cell 25, 1126-1142. [本文引用: 1]
MizoiJ, KanazawaN, KidokoroS, TakahashiF, QinF, MorimotoK, ShinozakiK, Yamaguchi-ShinozakiK (2019). Heat-induced inhibition of phosphorylation of the stress-protective transcription factor DREB2A promotes thermotolerance of Arabidopsis thaliana J Biol Chem 294, 902-917. [本文引用: 1]
MizoiJ, ShinozakiK, Yamaguchi-ShinozakiK (2012). AP2/ERF family transcription factors in plant abiotic stress responses Biochim Biophys Acta 1819, 86-96. [本文引用: 2]
MorimotoK, OhamaN, KidokoroS, MizoiJ, TakahashiF, TodakaD, MogamiJ, SatoH, QinF, KimJS, FukaoY, FujiwaraM, ShinozakiK, Yamaguchi-ShinozakiK (2017). BPM-CUL3 E3 ligase modulates thermotolerance by facilitating negative regulatory domain-mediated degradation of DREB2A in Arabidopsis Proc Natl Acad Sci USA 114, E8528-E8536. [本文引用: 1]
MüllerM, Munné-BoschS (2015). Ethylene response factors: a key regulatory hub in hormone and stress signaling Plant Physiol 169, 32-41. [本文引用: 2]
NakanoT, SuzukiK, FujimuraT, ShinshiH (2006). Genome-wide analysis of the ERF gene family in Arabidopsis and rice Plant Physiol 140, 411-432. [本文引用: 4]
NolanTM, BrennanB, YangMR, ChenJN, ZhangMC, LiZH, WangXL, BasshamDC, WalleyJ, YinYH (2017). Selective autophagy of BES1 mediated by DSK2 balances plant growth and survival Dev Cell 41, 33-46. [本文引用: 2]
NovilloF, MedinaJ, SalinasJ (2007). Arabidopsis CBF1 and CBF3 have a different function than CBF2 in cold acclimation and define different gene classes in the CBF regulon Proc Natl Acad Sci USA 104, 21002-21007. [本文引用: 1]
OhE, ZhuJY, WangZY (2012). Interaction between BZR1 and PIF4 integrates brassinosteroid and environmental responses Nat Cell Biol 14, 802-809. [本文引用: 1]
OsnatoM, CastillejoC, Matías-HernándezL, PelazS (2012). TEMPRANILLO genes link photoperiod and gibberellin pathways to control flowering in Arabidopsis Nat Commun 3, 808. [本文引用: 1]
OwjiH, HajiebrahimiA, SeradjH, HemmatiS (2017). Identification and functional prediction of stress responsive AP2/ERF transcription factors in Brassica napus by genome-wide analysis Comput Biol Chem 71, 32-56. [本文引用: 1]
PapdiC, Pérez-SalamóI, JosephMP, GiuntoliB, B?greL, KonczC, SzabadosL (2015). The low oxygen, oxidative and osmotic stress responses synergistically act through the ethylene response factor VII genes RAP2.12, RAP2.2 and RAP2.3 Plant J 82, 772-784. [本文引用: 2]
ParkCS, GoYS, SuhMC (2016). Cuticular wax biosynthesis is positively regulated by WRINKLED 4, an AP2/ERF- type transcription factor, in Arabidopsis stems Plant J 88, 257-270. [本文引用: 1]
ParkSC, LeeCM, DohertyCJ, GilmourSJ, KimY, ThomashowMF (2015). Regulation of the Arabidopsis CBF regulon by a complex low-temperature regulatory network Plant J 82, 193-207. [本文引用: 3]
PerataP (2018). The rice SUB1A gene: making adaptation to submergence and post-submergence possible Plant Cell Environ 41, 717-720. [本文引用: 1]
PflugerJ, WagnerD (2007). Histone modifications and dynamic regulation of genome accessibility in plants Curr Opin Plant Biol 10, 645-652. [本文引用: 1]
QiJS, SongCP, WangBS, ZhouJM, Kangasj?rviJ, ZhuJK, GongZZ (2018). Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack J Integr Plant Biol 60, 805-826. [本文引用: 1]
QiaoH, ShenZX, HuangSSC, SchmitzRJ, UrichMA, BriggsSP, EckerJR (2012). Processing and subcellular trafficking of ER-tethered EIN2 control response to ethylene gas Science 338, 390-393. [本文引用: 1]
QinF, ShinozakiK, Yamaguchi-ShinozakiK (2011). Achievements and challenges in understanding plant abiotic stress responses and tolerance Plant Cell Physiol 52, 1569-1582. [本文引用: 1]
RashotteAM, MasonMG, HutchisonCE, FerreiraFJ, SchallerEG, KieberJJ (2006). A subset of Arabidopsis AP2 transcription factors mediates cytokinin responses in concert with a two-component pathway Proc Natl Acad Sci USA 103, 11081-11085. [本文引用: 3]
RieuI, ErikssonS, PowersSJ, GongF, GriffithsJ, WoolleyL, BenllochR, NilssonO, ThomasSG, HeddenP, PhillipsAL (2008). Genetic analysis reveals that C19-GA 2-oxidation is a major gibberellin inactivation pathway in Arabidopsis Plant Cell 20, 2420-2436. [本文引用: 1]
RudnikR, BulchaJT, ReifschneiderE, EllersiekU, BaierM (2017). Specificity versus redundancy in the RAP2.4 transcription factor family of Arabidopsis thaliana: transcriptional regulation of genes for chloroplast peroxidases BMC Plant Biol 17, 144. [本文引用: 1]
SahSK, ReddyKR, LiJX (2016). Abscisic acid and abiotic stress tolerance in crop plants Front Plant Sci 7, 571. [本文引用: 3]
SaitoS, HiraiN, MatsumotoC, OhigashiH, OhtaD, SakataK, MizutaniM (2004). Arabidopsis CYP707As encode(+)-abscisic acid 8°-hydroxylase, a key enzyme in the oxidative catabolism of abscisic acid Plant Physiol 134, 1439-1449. [本文引用: 1]
SakumaY, LiuQ, DubouzetJG, AbeH, ShinozakiK, Yamaguchi-ShinozakiaK (2002). DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression Biochem Biophys Res Commun 290, 998-1009. [本文引用: 1]
SazegariS, NiaziA, AhmadiFS (2015). A study on the regulatory network with promoter analysis for Arabidopsis DREB-genes Bioinformation 11, 101-106. [本文引用: 1]
SchmitzAJ, FolsomJJ, JikamaruY, RonaldP, WaliaH (2013). SUB1A-mediated submergence tolerance response in rice involves differential regulation of the brassinosteroid pathway New Phytol 198, 1060-1070. [本文引用: 2]
ShiYT, DingYL, YangSH (2018). Molecular regulation of CBF signaling in cold acclimation Trends Plant Sci 23, 623-637. [本文引用: 2]
ShiYT, TianSW, HouLY, HuangXZ, ZhangXY, GuoHW, YangSH (2012). Ethylene signaling negatively regulates freezing tolerance by repressing expression of CBF and type-A ARR genes in Arabidopsis Plant Cell 24, 2578-2595. [本文引用: 2]
SongCP, AgarwalM, OhtaM, GuoY, HalfterU, WangPC, ZhuJK (2005). Role of an Arabidopsis AP2/EREBP- type transcriptional repressor in abscisic acid and drought stress responses Plant Cell 17, 2384-2396. [本文引用: 2]
SongCP, GalbraithDW (2006). AtSAP18, an orthologue of human SAP18, is involved in the regulation of salt stress and mediates transcriptional repression in Arabidopsis Plant Mol Biol 60, 241-257. [本文引用: 2]
SunY, FanXY, CaoDM, TangWQ, HeK, ZhuJY, HeJX, BaiMY, ZhuSW, OhE, PatilS, KimTW, JiHK, WongWH, RheeSY, WangZY (2010). Integration of brassinosteroid signal transduction with the transcription network for plant growth regulation in Arabidopsis Dev Cell 19, 765-777. [本文引用: 1]
SunZM, ZhouML, WangD, TangYX, LinM, WuYM (2016). Overexpression of the lotus corniculatus soloist gene LcAP2/ERF107 enhances tolerance to salt stress Protein Peptide Lett 23, 442-449. [本文引用: 1]
TaoJJ, ChenHW, MaB, ZhangWK, ChenSY, ZhangJS (2015). The role of ethylene in plants under salinity stress Front Plant Sci 6, 1059. [本文引用: 2]
TsutsuiT, KatoW, AsadaY, SakoK, SatoT, SonodaY, KidokoroS, Yamaguchi-ShinozakiK, TamaokiM, ArakawaK, IchikawaT, NakazawaM, SekiM, ShinozakiK, MatsuiM, IkedaA, YamaguchiJ (2009). DEAR1, a transcriptional repressor of DREB protein that mediates plant defense and freezing stress responses in Arabidopsis J Plant Res 122, 633-643. [本文引用: 2]
Van den BroeckL, DuboisM, VermeerschM, StormeV, MatsuiM, InzéD (2017). From network to phenotype: the dynamic wiring of an Arabidopsis transcriptional network induced by osmotic stress Mol Syst Biol 13, 961. [本文引用: 1]
WangHJ, YangCJ, ZhangC, WangNY, LuDH, WangJ, ZhangSS, WangZX, MaH, WangXL (2011). Dual role of BKI1 and 14-3-3s in brassinosteroid signaling to link receptor with transcription factors Dev Cell 21, 825-834. [本文引用: 2]
XiaoHG, SiddiquaM, BraybrookS, NassuthA (2006). Three grape CBF/DREB1 genes respond to low temperature, drought and abscisic acid Plant Cell Environ 29, 1410-1421. [本文引用: 1]
YangCY, HsuFC, LiJP, WangNN, ShihMC (2011). The AP2/ERF transcription factor AtERF73/HRE1 modulates ethylene responses during hypoxia in Arabidopsis Plant Physiol 156, 202-212. [本文引用: 2]
YaoY, HeRJ, XieQL, ZhaoXH, DengXM, HeJB, SongLL, HeJ, MarchantA, ChenXY, WuAM (2017). ETHYLENE RESPONSE FACTOR 74 (ERF74) plays an essential role in controlling a respiratory burst oxidase homolog D (RbohD)-dependent mechanism in response to different stresses in Arabidopsis New Phytol 213, 1667-1681. [本文引用: 3]
YiJ, ZhaoDM, ChuJF, YanJJ, LiuJS, WuMJ, ChengJF, JiangHY, ZengYJ, LiuD (2019). AtDPG1 is involved in the salt stress response of Arabidopsis seedling through ABI4 Plant Sci 287, 110180. [本文引用: 1]
YoshidaT, FujitaY, SayamaH, KidokoroS, MaruyamaK, MizoiJ, ShinozakiK, Yamaguchi-ShinozakiK (2010). AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation Plant J 61, 672-685. [本文引用: 1]
YuXF, LiL, ZolaJ, AluruM, YeHX, FoudreeA, GuoHQ, AndersonS, AluruS, LiuP, RodermelS, YinYH (2011). A brassinosteroid transcriptional network revealed by genome-wide identification of BESI target genes in Arabidopsis thaliana Plant J 65, 634-646. [本文引用: 1]
ZhangF, WangLK, KoEE, ShaoK, QiaoH (2018). Histone deacetylases SRT1 and SRT2 interact with ENAP1 to mediate ethylene-induced transcriptional repression Plant Cell 30, 153-166. [本文引用: 1]
ZhangGY, ChenM, LiLC, XuZS, ChenXP, GuoJM, MaYZ (2009). Overexpression of the soybean GmERF3 gene, an AP2/ERF type transcription factor for increased tolerances to salt, drought, and diseases in transgenic tobacco J Exp Bot 60, 3781-3796. [本文引用: 1]
ZhangLX, LiZF, QuanRD, LiGJ, WangRG, HuangRF (2011). An AP2 domain-containing gene,ESE1, targeted by the ethylene signaling component EIN3 is important for the salt response in Arabidopsis Plant Physiol 157, 854-865. [本文引用: 1]
ZhangZW, FengLY, ChengJ, TangH, XuF, ZhuF, ZhaoZY, YuanM, ChenYE, WangJH, YuanS, LinHH (2013). The roles of two transcription factors, ABI4 and CBFA, in ABA and plastid signaling and stress responses Plant Mol Biol 83, 445-458. [本文引用: 1]
ZhaoCZ, ZhangZJ, XieSJ, SiT, LiYY, ZhuJK (2016). Mutational evidence for the critical role of CBF transcription factors in cold acclimation in Arabidopsis Plant Physiol 171, 2744-2759. [本文引用: 1]
ZhaoCZ, ZhuJK (2016). The broad roles of CBF genes: from development to abiotic stress Plant Signal Behav 11, e1215794. [本文引用: 1]
ZhouMQ, ShenC, WuLH, TangKX, LinJ (2011). CBF-dependent signaling pathway: a key responder to low temperature stress in plants Crit Rev Biotechnol 31, 186-192. [本文引用: 3]
ZhuDL, WuZ, CaoGY, LiJG, WeiJ, TsugeT, GuHY, AoyamaT, QuLJ (2014). TRANSLUCENT GREEN, an ERF family transcription factor, controls water balance in Arabidopsis by activating the expression of aquaporin genes Mol Plant 7, 601-615. [本文引用: 3]
ZhuQ, ZhangJT, GaoXS, TongJH, XiaoLT, LiWB, ZhangHX (2010). The Arabidopsis AP2/ERF transcription factor RAP2.6 participates in ABA, salt and osmotic stress responses Gene 457, 1-12. [本文引用: 2]
ZwackPJ, ComptonMA, AdamsCI, RashotteAM (2016a). Cytokinin response factor 4 (CRF4) is induced by cold and involved in freezing tolerance Plant Cell Rep 35, 573-584. [本文引用: 1]
ZwackPJ, De ClercqI, HowtonTC, HallmarkHT, HurnyA, KeshishianEA, ParishAM, BenkovaE, MukhtarMS, van BreusegemF, RashotteAM (2016b). Cytokinin response factor 6 represses cytokinin-associated genes during oxidative stress Plant Physiol 172, 1249-1258. [本文引用: 3]
ZwackPJ, RashotteAM (2015). Interactions between cytokinin signaling and abiotic stress responses J Exp Bot 66, 4863-4871. [本文引用: 1]
ZwackPJ, RobinsonBR, RisleyMG, RashotteAM (2013). Cytokinin response factor 6 negatively regulates leaf senescence and is induced in response to cytokinin and numerous abiotic stresses Plant Cell Physiol 54, 971-981. [本文引用: 2]
植物响应联合胁迫机制的研究进展 1 2019
... 植物在生长发育过程中经常受到生物或非生物胁迫的影响, 由此, 植物逐渐形成了复杂的调节机制感知胁迫信号, 对外部环境变化做出快速响应(郭倩倩和周文彬, 2019).脱落酸(abscisic acid, ABA)、乙烯(ethylene, ET)及赤霉素(gibberellin, GA)等激素作为信号分子能够调节植物的生长发育、形态建成以及抗逆反应等生物学过程.该过程也受转录因子(transcription factor, TF)调控, 即转录因子通过依赖或不依赖于激素介导的信号通路参与调控植物对干旱、低温、缺氧、高盐和淹水等非生物胁迫的响应, 进而影响植物的逆境耐受能力(Gibbs et al., 2015; Verma et al., 2016; Li et al., 2018).AP2/ERF (APETALA2/eth-ylene responsive factor)是植物特有的转录因子家族之一, 已在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、高粱(Sorghum bicolor)、大白菜(Brassica pekinensis)、毛竹(Phyllostachys edulis)、玉米(Zea mays)、大麦(Hordeum vulgare)和小麦(Triticum aestivum)等物种中被成功分离鉴定(Xu and Chua, 2011; Chandler, 2018).通过突变体实验, 科学家发现了许多与AP2/ERF相关的非生物胁迫或激素信号应答基因.目前, AP2/ERF已成为研究非生物胁迫和激素互作的热点候选基因.本文从AP2/ERF转录因子的结构特征、转录调控、翻译后修饰、结合位点、协同互作蛋白以及其参与调控依赖或不依赖激素信号途径的非生物胁迫响应进行综述, 为进一步阐明AP2/ERF转录因子在非生物胁迫调控网络中的作用提供理论依据. ...
CBF: 平衡植物低温应答与生长发育的关键 1 2017
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
Oxygen sensing coordinates photomorphogenesis to facilitate seedling survival 1 2015
... 翻译后修饰(post-translational modification, PTM)是蛋白质合成途径的后期加工过程, 不同的修饰方式对AP2/ERF蛋白的活性、稳定性和丰度的影响有差异.磷酸化是转录因子的主要修饰途径之一, 磷酸化水平与下游靶基因的表达水平关系密切.拟南芥ABA信号通路的正调控因子蔗糖非酵解型蛋白激酶(Snf1-re-lated protein kinases, SnRKs)可磷酸化RAV1, 进而抑制RAV1的转录阻遏效应(Feng et al., 2014), 而ERF104和ERF6可被后MPK3/6 (mitogen-activated protein kinase 3/6)磷酸化修饰, 并在下游级联反应中发挥重要作用, 调控植物对病原体的免疫反应(Meng et al., 2013).DREB2A是拟南芥的关键转录激活因子, 在其中央序列存在一个负调控域(negative regulatory domain, NRD).热胁迫条件下, NRD中的Ser/Thr残基磷酸化水平降低, DREB2A蛋白稳定性提高, 敲除NRD后DREB2A呈组成型激活, 表明抑制NRD磷酸化可以稳定和激活DREB2A的表达, 从而增强植物的耐热性(Mizoi et al., 2019).泛素介导的蛋白质降解(26S蛋白酶体途径)也参与调节AP2/ ERF蛋白的稳定性.非生物胁迫条件下, DREB2A和ERF75/RAP-2.2蛋白分别被RING家族E3泛素连接酶DRIP1/2 (DREB2A-interacting protein1/2)和SINAT2 (seven in absentia of Arabidopsis 2)直接泛素化(Cheng et al., 2012; Papdi et al., 2015).研究发现CUL3-E3连接酶适配子BPMs (BTB/POZ and math domain)可与DREB2A蛋白的负调控域NRD在核内发生互作, 敲除BPM增加DREB2A蛋白的积累, 促进下游靶基因的表达, 提高植株的耐热和耐旱能力, 这也说明BPM对DREB2A蛋白的稳定性起负调控作用(Morimoto et al., 2017).此外, 研究发现ERF-VII类蛋白可在质膜上与乙酰辅基A结合蛋白(Acyl-CoA binding protein, ACBP)互作.在缺氧条件下, RAP2.12与ACBP分离后进入细胞核, 在脱酰胺基和精氨酰基化的作用下通过氧依赖途径移除ERF72/RAP2.3、ERF74/RAP2.12蛋白的甲硫氨酸N末端, 将半胱氨酸氧化成半胱亚磺酸, 引发蛋白快速降解并暴露出非稳定N端的氨基酸残基(Gibbs et al., 2015; Abbas et al., 2015). ...
Role of ethylene and the APETALA 2/ethylene response factor superfamily in rice under various abiotic and biotic stress conditions 1 2017
... AP2/ERF转录因子具有独特的结构特征, 其由DNA结合域(DNA-binding domain)、转录调控域(transcription regulation domain)、寡聚化位点(oligomerization site)和核定位信号(nuclear localization signal, NLS) 4个主要功能区组成, 并以高度保守的AP2/ ERF结合域而得名(Sakuma et al., 2002).通常情况下, AP2/ERF转录因子至少含有1个AP2结构域的DNA结合区, 该结构域包含60-70个氨基酸残基, 按照3个β折叠和1个α螺旋方式形成典型的三维结构(Nakano et al., 2006; Abiri et al., 2017).根据包含的AP2/ERF结构域数量和结合序列特点, 可将AP2/ ERF家族分为AP2 (APETALA2)、RAV (related to ABI3/VP1)、脱水反应元件结合蛋白(dehydration- responsive element binding protein, DREB) (A1-A6亚组)、乙烯应答因子(ethylene-responsive factor, ERF) (V-X亚组)和Soloist五个亚族(Nakano et al., 2006; Licausi et al., 2013).其中, 对AP2、DREB和ERF的研究较为广泛和深入, 而有关Soloist亚族的研究报道极少, 已知其核苷酸序列在多数植物中高度保守(Sun et al., 2016). ...
The cold-inducible CBF1 factor-de-pendent signaling pathway modulates the accumulation of the growth-repressing DELLA proteins via its effect on gibberellin metabolism 1 2008
... AP2/ERFs参与调控植物GA生物合成相关基因的转录.盐胁迫下, DREB1E和DREB1F抑制GA20ox的表达, 使GA生物合成减少, 引起植株生长迟缓(Magome et al., 2004).过表达ERF6抑制GA2ox的表达水平, 诱导赤霉素降解酶(gibberellin 2-oxidase 6, GA2ox6)的合成, 维持DELLA蛋白的稳定性.渗透胁迫下, erf6功能获得突变体植株矮小且对胁迫高度敏感, 而erf5/erf6缺失突变体生长受胁迫影响较小.研究发现ERF6还激活了STZ、MYB51和WRKY33等渗透胁迫响应基因的表达(Dubois et al., 2013).冷胁迫下, CBF1转录因子激发GA2ox基因的表达而降低GA含量, 组成型表达CBF1抑制GA的积累, 植株表现矮化, 表明DELLA蛋白是CBF1介导的冷胁迫响应的关键组分(Achard et al., 2008).相反, ERF11作为一种GA生物合成和信号转导的正调控因子, 抑制ET生物合成, 促进植株节间伸长.过表达ERF11上调GA3ox1和GA20ox基因的表达水平, 增加GA的积累量(Zhou et al., 2016).ERF6作为转录激活因子诱导ERF11的表达, 反之, ERF11抑制GA2-OX6、STZ、MYB51和WRKY33等ERF6相关靶基因的表达.进一步实验表明, 4个靶基因在erf11/erf6过表达植株中也不能被诱导表达, ERF11过表达可消除ERF6过表达导致的极度矮化现象, 说明ERF11可能通过直接竞争目标基因启动子而在分子水平上拮抗ERF6 (Dubois et al., 2015). ...
A R2R3 type MYB transcription factor is involved in the cold regulation of CBF genes and in acquired freezing tolerance 1 2006
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
Plant responses to abiotic stress: the chromatin context of transcriptional regulation 1 2017
... 正常环境下, AP2/ERF转录因子表达水平较低, 激素或逆境胁迫相关基因与AP2/ERF转录因子保守的顺式作用元件结合, 进而调控其表达(Owji et al., 2017).研究表明, 拟南芥DREB1A/CBF3、DREB1C/CBF2、DREB2C、DREB2G和DEAR3的启动子中包含较多的热激元件(heat shock element, HSE)、低温响应(low temperature responsive, LTR)、脱落酸响应(ABA responsive element, ABRE)等顺式作用元件(Sazegari et al., 2015).热激因子1 (heat shock factor 1, HSF1)、脱落酸响应结合蛋白(ABA responsive element binding protein/ABRE binding factors, AREB/ABF)可分别与DREB2A启动子中的热激应答和ABA响应元件结合, 调控其表达, 进而改变植物胁迫的耐受性(Kim et al., 2011; Liu et al., 2011).蛋白磷酸酶2C (type 2C protein phosphatase, PP2C)包含2个以上的ABRE元件, AREB1、AREB2和ABF3协同正调控AHG1、AHG3、HAI1及HAI2等PP2Cs基因表达(Yoshida et al., 2010).此外, 越来越多的研究证明ET信号通路参与拟南芥盐胁迫反应.进一步分析表明, ESE1 (ethylene and salt-inducible ERF1)在EIN3/EIL1下游调节ET响应基因.EIN3 (ethylene insensitive 3)可与ESE1启动子结合形成EIN3-ESE1转录复合体, 继而调控RD29A和COR15A等基因的表达(Zhang et al., 2011).磷酸化、泛素化、甲基化和乙酰化等组蛋白修饰方式通过调控染色质开放或封闭的空间状态, 激活或抑制AP2/ERF转录(Pfluger and Wagner, 2007).组蛋白去乙酰化酶(histone deacetylase, HDAC)和聚乙二醇(PEG)促进DREB1基因启动子区乙酰化, 增强DREB1基因的表达(Zhang et al., 2018).非生物胁迫下, 组蛋白变体和组蛋白修饰状态也可发生改变, 并可遗传到下一代(Asensii-Fabado et al., 2017).miRNA介导的RNA沉默和翻译抑制在AP2/ERF表达调控中也发挥重要作用.研究发现miRNA172靶向拟南芥AP2mRNA并抑制其翻译, 而miRNA156和miRNA838两者也具有类似调节AP2/ERF的功能(Kavas et al., 2015).此外, 在拟南芥、水稻及玉米中还发现通过可变剪切产生的OsDREB2A/2B、ZmDREB2A、WDREB2以及HvDRF1等多种AP2/-ERF功能异构体(Matsukura et al., 2010). ...
Gibberellin biosynthesis and signal transduction is essential for internode elongation in deepwater rice 1 2014
... 在拟南芥和水稻中, SUB1A、SK1和SK2参与GA信号转导通路的调控.淹水胁迫下, SUB1A激活SLR1 (slender rice 1)和SLRL1 (slender rice 1 like 1) (DELLA类似蛋白)转录, 同时SUB1A抑制受GA诱导的淀粉代谢相关基因的表达, 调控植株节间伸长和呼吸作用(Fukao and Bailey-Serres, 2008; Locke et al., 2018; Perata, 2018).SK1和SK2两个转录因子中均含有EIN3结合位点, 电泳迁移率测定显示, 类EIN3基因EIL1b可与SK1和SK2启动子结合, 激活其转录活性.研究发现, 淹水状态下拟南芥中ET合成增加, 进而诱导SK1和SK2表达, 激发GA20oxs基因上调表达, 促进植株节间伸长, 减轻淹水对其生长的抑制效应(Hattori et al., 2009; Ayano et al., 2014). ...
The Arabidopsis ABA-INSENSITIVE (ABI) 4 factor acts as a central transcription activator of the expression of its own gene, and for the induction of ABI5 and SBE2.2 genes during sugar signaling 1 2009
... AP2/ERF转录因子通过特异性结合胁迫应答基因启动子区顺式元件, 参与调控植物的逆境防御反应.DREB亚族成员特异性识别并结合启动子区域的DRE/CRT (dehydration responsive element/C-re-peat)元件(核心序列A/GCCGAC), 诱导干旱、低温和盐胁迫相关应答基因的表达.ERF亚族成员则可通过与乙烯响应元件(ethylene response element, ERE) (核心序列AGCCGCC, 又称GCC-box)结合, 从而参与调控乙烯应答和非生物胁迫(Franco-Zorrilla et al., 2014).此外, 研究发现许多AP2/ERF转录因子可同时与DRE/CRT和ERE两种元件结合, 如DREB亚族成员TINY、CBF1、ERF53、RAP2.4、TG/RAP2.4a以及ERF亚族成员ERF1、ERF4和ERF71 (Chen et al., 2012; Zhu et al., 2014; Lee et al., 2015), 但结合能力有差异.在拟南芥中, DREB2C和DEAR3与类GCC-box元件(GCCGCC)的结合能力远超DRE元件(Franco-Zorrilla et al., 2014).AP2/ERF转录因子还能与DRE和ERE之外的顺式元件结合, 如偶联元件1 (CE1, TGCCACCG)、类偶联元件3 (CE-3like, CGCG)和缺氧响应启动子元件(hypoxia-responsive promoter element, HRPE), 以及ATCTA、CAACA、CATGCA、CAA/CA/CTG、ATCGAG和(NC/GT) CGNCCA (Bossi et al., 2009; Zhu et al., 2010; Chen et al., 2016; Gasch et al., 2016; Park et al., 2016).近年来, 科学家通过蛋白质结合微阵列(protein binding microarray, PBM)技术发现结构特征高度相似的转录因子具有相似的DNA结合位点, 同时也具有相似的生物学功能.该研究结果合理地解释了转录因子功能冗余现象, 也为预测和挖掘未知AP2/ERF转录因子提供了高效可行的技术方法(Franco-Zorrilla et al., 2014). ...
Constitutively expressed ERF-VII transcription factors redundantly activate the core anaerobic response in Arabidopsis thaliana 1 2015
... ERF亚族成员在低氧和淹水胁迫响应过程中也发挥重要作用(Bui et al., 2015; Gibbs et al., 2015).SUB1A (submergence 1A)是ERF亚族B2亚组成员之一, Sub1A-1超表达的同时促进乙醇脱氢酶基因(alcohol dehydrogenase 1, Adh1)上调表达, 抑制Sub1C的转录水平, 说明Sub1A-1是耐淹性的主要调节因子, 在淹水胁迫应答调控中起关键作用(Xu et al., 2006).淹水状态下, 低水平海藻糖-6-磷酸合酶(trehalose-6-phosphate, T6P)与CIPK15 (CBL-inter-acting protein kinase 15)协同激活SnRK1, SnRK1介导SUB1A抑制幼苗胚轴生长和呼吸相关基因的表达, 直接或间接调控碳同化过程.而SK1/SK2 (snorkel1/2)激活参与叶柄、茎和导管发育相关基因, 促进植株快速生长(Hattori et al., 2009; Locke et al., 2018).缺氧胁迫下, 拟南芥ERF-VIIs可以通过氧依赖的N端规则通路降解(Gibbs et al., 2015).RING (really interesting gene)类蛋白SINAT2可通过其N端保守的C3H4型RING结构域调控RAP2.12, 沉默SINAT1/2可上调RAP2.12的表达水平(Papdi et al., 2015).此外, ERF71/HRE2、ERF72/RAP2.3、ERF73/HRE1、ERF74/RAP2.12及ERF75/RAP2.2d也受缺氧胁迫诱导, 进而正向调控糖代谢和激素信号相关的缺氧响应基因, 提高植株的缺氧耐受能力.过表达RAP2.2、RAP2.3和RAP2.12可提高植株在低氧和渗透胁迫下的存活率, 而rap2.12-2/rap2.3-1双突变体对淹水和渗透胁迫表现出较高的敏感性(Yao et al., 2017). ...
The TOPLESS interactome: a framework for gene repression in Arabidopsis 1 2012
... 许多研究表明, AP2/ERF转录因子可直接与靶基因启动子结合, 激活或抑制下游靶基因的表达, 其N端DNA结合域和C端转录激活域在胁迫相关靶基因的转录激活中起重要作用(Nakano et al., 2006).进一步分析表明, AP2/ERF转录因子中含有EAR基序(LxLxL或DLNxxP)、TLLLFR基序和BRD结构域(B3 repression domain, 保守序列为R/KLFGV)等多个转录抑制结构域(Ikeda and Ohme-Takagi, 2009; Kagale and Rozwadowski, 2011; Deepika et al., 2016).含EAR基序的转录因子是一种重要的抑制子, 在植物非生物胁迫应答基因的诱导表达中起负调控作用.Tiwari等(2012)从拟南芥ERF和AP2亚族中发现了一种转录激活关键元件EDLL, EDLL可与EAR基序(ERF-associated amphiphilic repression)发生互作, 拮抗后者介导的转录抑制作用.而不同的AP2/ ERF转录因子也可协同招募TPL (topless)和TPR (topless-related)等转录共抑制子(Causier et al., 2012)或组蛋白修饰因子, 进而抑制下游靶基因的表达(Song et al., 2005; Song and Galbraith, 2006).Song等(2005)发现AtERF7特异性地结合GCC-box, 与抑制因子ATSIN3及组蛋白去乙酰化酶19 (histone deacetylase 19, HDA19)发生互作并形成转录因子复合物, 参与干旱胁迫和ABA响应调控.AtERF7过表达株系保卫细胞对ABA的敏感性减弱, 水分散失增加; 反之, AtERF7 RNA干扰系对ABA的敏感性增强.类似的, ERF3能与组蛋白去乙酰化酶复合物亚基SAP18 (SIN3 associated polypeptide P18)互作, 共同招募HDA19形成复合体, 进而抑制相关基因的表达(Song and Galbraith, 2006).BRD结构域被认为是RAV1和RAV2转录因子行使抑制功能的关键元件, 其保守序列中第1位氨基酸残基多为疏水性亮氨酸、缬氨酸和蛋氨酸残基, 突变导致功能丧失(Ikeda and Ohme-Takagi, 2009).研究显示, 与AP2/ERFs协同调控的基因同样可被AP2/ERFs靶基因富集, 分析此类基因的同源基序有助于识别转录因子的假定靶基因并预测其生物学功能(Franco-Zorrillaetal et al., 2014). ...
Class VIIIb APETALA2 ethylene response factors in plant development 1 2018
... 植物在生长发育过程中经常受到生物或非生物胁迫的影响, 由此, 植物逐渐形成了复杂的调节机制感知胁迫信号, 对外部环境变化做出快速响应(郭倩倩和周文彬, 2019).脱落酸(abscisic acid, ABA)、乙烯(ethylene, ET)及赤霉素(gibberellin, GA)等激素作为信号分子能够调节植物的生长发育、形态建成以及抗逆反应等生物学过程.该过程也受转录因子(transcription factor, TF)调控, 即转录因子通过依赖或不依赖于激素介导的信号通路参与调控植物对干旱、低温、缺氧、高盐和淹水等非生物胁迫的响应, 进而影响植物的逆境耐受能力(Gibbs et al., 2015; Verma et al., 2016; Li et al., 2018).AP2/ERF (APETALA2/eth-ylene responsive factor)是植物特有的转录因子家族之一, 已在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、高粱(Sorghum bicolor)、大白菜(Brassica pekinensis)、毛竹(Phyllostachys edulis)、玉米(Zea mays)、大麦(Hordeum vulgare)和小麦(Triticum aestivum)等物种中被成功分离鉴定(Xu and Chua, 2011; Chandler, 2018).通过突变体实验, 科学家发现了许多与AP2/ERF相关的非生物胁迫或激素信号应答基因.目前, AP2/ERF已成为研究非生物胁迫和激素互作的热点候选基因.本文从AP2/ERF转录因子的结构特征、转录调控、翻译后修饰、结合位点、协同互作蛋白以及其参与调控依赖或不依赖激素信号途径的非生物胁迫响应进行综述, 为进一步阐明AP2/ERF转录因子在非生物胁迫调控网络中的作用提供理论依据. ...
A proximal promoter region of Arabidopsis DREB2C confers tissue-speci?c expression under heat stress 2 2012
... AP2/ERF转录因子通过特异性结合胁迫应答基因启动子区顺式元件, 参与调控植物的逆境防御反应.DREB亚族成员特异性识别并结合启动子区域的DRE/CRT (dehydration responsive element/C-re-peat)元件(核心序列A/GCCGAC), 诱导干旱、低温和盐胁迫相关应答基因的表达.ERF亚族成员则可通过与乙烯响应元件(ethylene response element, ERE) (核心序列AGCCGCC, 又称GCC-box)结合, 从而参与调控乙烯应答和非生物胁迫(Franco-Zorrilla et al., 2014).此外, 研究发现许多AP2/ERF转录因子可同时与DRE/CRT和ERE两种元件结合, 如DREB亚族成员TINY、CBF1、ERF53、RAP2.4、TG/RAP2.4a以及ERF亚族成员ERF1、ERF4和ERF71 (Chen et al., 2012; Zhu et al., 2014; Lee et al., 2015), 但结合能力有差异.在拟南芥中, DREB2C和DEAR3与类GCC-box元件(GCCGCC)的结合能力远超DRE元件(Franco-Zorrilla et al., 2014).AP2/ERF转录因子还能与DRE和ERE之外的顺式元件结合, 如偶联元件1 (CE1, TGCCACCG)、类偶联元件3 (CE-3like, CGCG)和缺氧响应启动子元件(hypoxia-responsive promoter element, HRPE), 以及ATCTA、CAACA、CATGCA、CAA/CA/CTG、ATCGAG和(NC/GT) CGNCCA (Bossi et al., 2009; Zhu et al., 2010; Chen et al., 2016; Gasch et al., 2016; Park et al., 2016).近年来, 科学家通过蛋白质结合微阵列(protein binding microarray, PBM)技术发现结构特征高度相似的转录因子具有相似的DNA结合位点, 同时也具有相似的生物学功能.该研究结果合理地解释了转录因子功能冗余现象, 也为预测和挖掘未知AP2/ERF转录因子提供了高效可行的技术方法(Franco-Zorrilla et al., 2014). ...
... DREB-A2亚组转录因子DREB2主要参与调节植物的抗旱和耐热性(Mizoi et al., 2012).干旱和高温可诱导DREB2基因的表达, 进而正向调控下游干旱或热胁迫响应基因(Maruyama et al., 2009).研究表明DREB-A4家族成员HRD (hardy)和DREB-A6家族成员(ERF53、RAP2.4和TG/RAP2.4a)也在干旱和耐盐性调控中发挥关键作用(Karaba et al., 2007; Lin et al., 2008; Chen et al., 2012; Zhu et al., 2014).过表达HRD能显著增强拟南芥和水稻植株的耐旱和耐盐性(Karaba et al., 2007).同样, 过表达TG (translucent green)导致植株叶片玻璃化, 叶片细胞含水量增加(Zhu et al., 2014).TG也参与调控抗坏血酸过氧化物酶基因APX (ascorbate peroxidase)的表达, 抵御活性氧(reactive oxygen species, ROS)引起的光氧化胁迫(Rudnik et al., 2017).除拟南芥外, 人们对水稻、玉米、番茄(Lycopersicon esculentum)、葡萄(Vitis vinifera)等作物的DREB1和DREB2转录因子也进行了广泛的研究(Xiao et al., 2006; Matsukura et al., 2010; Dou et al., 2014). ...
ORA47 (octadecanoid-responsive AP2/ERF- domain transcription factor 47) regulates jasmonic acid and abscisic acid biosynthesis and signaling through binding to a novel cis-element 2 2016
... AP2/ERF转录因子通过特异性结合胁迫应答基因启动子区顺式元件, 参与调控植物的逆境防御反应.DREB亚族成员特异性识别并结合启动子区域的DRE/CRT (dehydration responsive element/C-re-peat)元件(核心序列A/GCCGAC), 诱导干旱、低温和盐胁迫相关应答基因的表达.ERF亚族成员则可通过与乙烯响应元件(ethylene response element, ERE) (核心序列AGCCGCC, 又称GCC-box)结合, 从而参与调控乙烯应答和非生物胁迫(Franco-Zorrilla et al., 2014).此外, 研究发现许多AP2/ERF转录因子可同时与DRE/CRT和ERE两种元件结合, 如DREB亚族成员TINY、CBF1、ERF53、RAP2.4、TG/RAP2.4a以及ERF亚族成员ERF1、ERF4和ERF71 (Chen et al., 2012; Zhu et al., 2014; Lee et al., 2015), 但结合能力有差异.在拟南芥中, DREB2C和DEAR3与类GCC-box元件(GCCGCC)的结合能力远超DRE元件(Franco-Zorrilla et al., 2014).AP2/ERF转录因子还能与DRE和ERE之外的顺式元件结合, 如偶联元件1 (CE1, TGCCACCG)、类偶联元件3 (CE-3like, CGCG)和缺氧响应启动子元件(hypoxia-responsive promoter element, HRPE), 以及ATCTA、CAACA、CATGCA、CAA/CA/CTG、ATCGAG和(NC/GT) CGNCCA (Bossi et al., 2009; Zhu et al., 2010; Chen et al., 2016; Gasch et al., 2016; Park et al., 2016).近年来, 科学家通过蛋白质结合微阵列(protein binding microarray, PBM)技术发现结构特征高度相似的转录因子具有相似的DNA结合位点, 同时也具有相似的生物学功能.该研究结果合理地解释了转录因子功能冗余现象, 也为预测和挖掘未知AP2/ERF转录因子提供了高效可行的技术方法(Franco-Zorrilla et al., 2014). ...
... DREB-A3亚组成员ABI4 (ABA insensitive 4)是ABA信号通路的重要组分.植株受到胁迫后, 体内积累ABA和ROS, ABI4可被ABA、ROS及糖质体等多种信号通路靶向调控.CCAAT结合因子A (CCAAT binding factor A, CBFA)是四吡咯化合物HAP (heme activator protein)三聚体转录复合物的亚基, 当产生过量ROS时, ABI4可与其它转录因子竞争性地结合CCACGT元件, 抑制CBFA的表达, 进而调控质体反向信号通路中相关基因的表达(Zhang et al., 2013).拟南芥突变体实验表明, ABI4在YL1 (yellow leaf 1) (Li et al., 2016)和DPG1 (delayed pale-greening 1) (Yi et al., 2019)等基因参与盐胁迫响应的调控网络中也发挥重要作用.逆境胁迫下, ORA47在JA (jasmo-nate acid)和ABA信号转导途径中靶向调节ABA和JA诱导的下游应答基因, ERF18/ORA-47识别启动子顺式元件(NC/GT)CGNCCA (O-box), 激活PP2C家族ABI2转录因子的表达.ABI1则在ORA-47上游起作用, 从而形成ABI1-ORA47-ABI2正反馈基因表达回路, 调控ABA信号转导和干旱胁迫响应(Chen et al., 2016).此外, RAV1转录因子在ABA信号转导途径中也发挥关键作用, RAV1能与ABI3、ABI4和ABI5启动子结合, 其过表达抑制ABI3、ABI4和ABI5的转录活性, 导致根系生长发育过程中对ABA不敏感.同时, 蔗糖非酵解型蛋白激酶(SnRK2.2、SnRK2.3和SnRK2.6)在核内磷酸化RAV1, 降低ABI5对RAV1的依赖性, 并负调控RAV1对靶基因的转录抑制作用(Feng et al., 2014). ...
Arabidopsis RGLG2, functioning as a RING E3 ligase, interacts with AtERF53 and negatively regulates the plant drought stress response 1 2012
... 翻译后修饰(post-translational modification, PTM)是蛋白质合成途径的后期加工过程, 不同的修饰方式对AP2/ERF蛋白的活性、稳定性和丰度的影响有差异.磷酸化是转录因子的主要修饰途径之一, 磷酸化水平与下游靶基因的表达水平关系密切.拟南芥ABA信号通路的正调控因子蔗糖非酵解型蛋白激酶(Snf1-re-lated protein kinases, SnRKs)可磷酸化RAV1, 进而抑制RAV1的转录阻遏效应(Feng et al., 2014), 而ERF104和ERF6可被后MPK3/6 (mitogen-activated protein kinase 3/6)磷酸化修饰, 并在下游级联反应中发挥重要作用, 调控植物对病原体的免疫反应(Meng et al., 2013).DREB2A是拟南芥的关键转录激活因子, 在其中央序列存在一个负调控域(negative regulatory domain, NRD).热胁迫条件下, NRD中的Ser/Thr残基磷酸化水平降低, DREB2A蛋白稳定性提高, 敲除NRD后DREB2A呈组成型激活, 表明抑制NRD磷酸化可以稳定和激活DREB2A的表达, 从而增强植物的耐热性(Mizoi et al., 2019).泛素介导的蛋白质降解(26S蛋白酶体途径)也参与调节AP2/ ERF蛋白的稳定性.非生物胁迫条件下, DREB2A和ERF75/RAP-2.2蛋白分别被RING家族E3泛素连接酶DRIP1/2 (DREB2A-interacting protein1/2)和SINAT2 (seven in absentia of Arabidopsis 2)直接泛素化(Cheng et al., 2012; Papdi et al., 2015).研究发现CUL3-E3连接酶适配子BPMs (BTB/POZ and math domain)可与DREB2A蛋白的负调控域NRD在核内发生互作, 敲除BPM增加DREB2A蛋白的积累, 促进下游靶基因的表达, 提高植株的耐热和耐旱能力, 这也说明BPM对DREB2A蛋白的稳定性起负调控作用(Morimoto et al., 2017).此外, 研究发现ERF-VII类蛋白可在质膜上与乙酰辅基A结合蛋白(Acyl-CoA binding protein, ACBP)互作.在缺氧条件下, RAP2.12与ACBP分离后进入细胞核, 在脱酰胺基和精氨酰基化的作用下通过氧依赖途径移除ERF72/RAP2.3、ERF74/RAP2.12蛋白的甲硫氨酸N末端, 将半胱氨酸氧化成半胱亚磺酸, 引发蛋白快速降解并暴露出非稳定N端的氨基酸残基(Gibbs et al., 2015; Abbas et al., 2015). ...
Cold stress regulation of gene expression in plants 1 2007
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
DELLA signaling mediates stress-induced cell differentiation in Arabidopsis leaves through modulation of anaphasepromoting complex/cyclosome activity 1 2012
... 赤霉素(GA)广泛分布在植物中并参与调控叶、芽以及合子发育等多种生物学过程.GA由GA20氧化酶(gibberellin 20-oxidases, GA20ox)和GA3氧化酶(GA3ox)等关键酶合成, 在GA2氧化酶(GA2ox)作用下被分解代谢(Rieu et al., 2008).DELLA蛋白作为GA信号转导途径中的主要负调控因子, 也是整合生长素、ABA、ET和JA等植物激素或环境信号的枢纽蛋白.植物体接受GA信号后, 通过泛素-蛋白酶体途径降解DELLA蛋白, 解除其对生长的抑制作用(Zhang et al., 2009).冷、盐和渗透胁迫下, GA含量降低, 导致植物生长减缓(Colebrook et al., 2014).极低浓度GA下, DELLA蛋白抑制GA介导的应答调控, 外源GA处理促进DELLA蛋白降解(Claeys et al., 2012). ...
The role of gibberellin signaling in plant responses to abiotic stress 2 2014
... AP2/ERF转录因子在参与调控植物应对冷、干旱、高温、高盐及缺氧等多种非生物胁迫过程中具有重要功能(Licausi et al., 2013).其中, DREB和ERF亚家族成员在植物非生物逆境应答中起重要调控作用.研究表明其参与调控的植物环境胁迫应答信号网络比较复杂.胁迫条件下, 部分AP2/ERFs可被快速持续地诱导表达, 而其它进程的响应较为缓慢, 说明可能存在依赖或不依赖于激素信号途径2种胁迫响应模式, 且两者之间存在一定的交叉互作(Van den Broeck et al., 2017).前人的研究也证明, AP2/ERFs广泛参与调控ABA、ET、GA、细胞分裂素(cytokinin, CTK)和油菜素内酯(brassinolide, BR)等植物激素介导的胁迫响应(Colebrook et al., 2014; Kazan et al., 2015; Tao et al., 2015; Sah et al., 2016; Nolan et al., 2017) (图1). ...
... 赤霉素(GA)广泛分布在植物中并参与调控叶、芽以及合子发育等多种生物学过程.GA由GA20氧化酶(gibberellin 20-oxidases, GA20ox)和GA3氧化酶(GA3ox)等关键酶合成, 在GA2氧化酶(GA2ox)作用下被分解代谢(Rieu et al., 2008).DELLA蛋白作为GA信号转导途径中的主要负调控因子, 也是整合生长素、ABA、ET和JA等植物激素或环境信号的枢纽蛋白.植物体接受GA信号后, 通过泛素-蛋白酶体途径降解DELLA蛋白, 解除其对生长的抑制作用(Zhang et al., 2009).冷、盐和渗透胁迫下, GA含量降低, 导致植物生长减缓(Colebrook et al., 2014).极低浓度GA下, DELLA蛋白抑制GA介导的应答调控, 外源GA处理促进DELLA蛋白降解(Claeys et al., 2012). ...
Genome-wide analysis of the AP2/ERF family in Musa species reveals divergence and neofunctionalisation during evolution 1 2016
... 许多研究表明, AP2/ERF转录因子可直接与靶基因启动子结合, 激活或抑制下游靶基因的表达, 其N端DNA结合域和C端转录激活域在胁迫相关靶基因的转录激活中起重要作用(Nakano et al., 2006).进一步分析表明, AP2/ERF转录因子中含有EAR基序(LxLxL或DLNxxP)、TLLLFR基序和BRD结构域(B3 repression domain, 保守序列为R/KLFGV)等多个转录抑制结构域(Ikeda and Ohme-Takagi, 2009; Kagale and Rozwadowski, 2011; Deepika et al., 2016).含EAR基序的转录因子是一种重要的抑制子, 在植物非生物胁迫应答基因的诱导表达中起负调控作用.Tiwari等(2012)从拟南芥ERF和AP2亚族中发现了一种转录激活关键元件EDLL, EDLL可与EAR基序(ERF-associated amphiphilic repression)发生互作, 拮抗后者介导的转录抑制作用.而不同的AP2/ ERF转录因子也可协同招募TPL (topless)和TPR (topless-related)等转录共抑制子(Causier et al., 2012)或组蛋白修饰因子, 进而抑制下游靶基因的表达(Song et al., 2005; Song and Galbraith, 2006).Song等(2005)发现AtERF7特异性地结合GCC-box, 与抑制因子ATSIN3及组蛋白去乙酰化酶19 (histone deacetylase 19, HDA19)发生互作并形成转录因子复合物, 参与干旱胁迫和ABA响应调控.AtERF7过表达株系保卫细胞对ABA的敏感性减弱, 水分散失增加; 反之, AtERF7 RNA干扰系对ABA的敏感性增强.类似的, ERF3能与组蛋白去乙酰化酶复合物亚基SAP18 (SIN3 associated polypeptide P18)互作, 共同招募HDA19形成复合体, 进而抑制相关基因的表达(Song and Galbraith, 2006).BRD结构域被认为是RAV1和RAV2转录因子行使抑制功能的关键元件, 其保守序列中第1位氨基酸残基多为疏水性亮氨酸、缬氨酸和蛋氨酸残基, 突变导致功能丧失(Ikeda and Ohme-Takagi, 2009).研究显示, 与AP2/ERFs协同调控的基因同样可被AP2/ERFs靶基因富集, 分析此类基因的同源基序有助于识别转录因子的假定靶基因并预测其生物学功能(Franco-Zorrillaetal et al., 2014). ...
The Arabidopsis EAR-motif-containing protein RAP2.1 functions as an active transcriptional repressor to keep stress responses under tight control 2 2010
... 部分A5亚组DREB类转录因子则对上述调控网络具有负反馈调节作用.DEAR1 (DREB and EAR motif protein 1)可能既在CBF上游作用, 又与RAP2.1在CBF下游扮演冷胁迫响应的调节子, 调节RD29A/COR78、COR15A和KIN1等冷诱导相关基因的表达, 负调控植物的耐冷和耐旱性(Fowler and Thomashow, 2002; Tsutsui et al., 2009; Dong and Liu, 2010).DEAR1过表达抑制冷诱导CBF的表达, 进而降低植物的耐冷能力(Tsutsui et al., 2009).低温胁迫或组成型表达CBF可诱导RAP2.1的表达, CBF2比RAP2.1对低温更为敏感(Dong and Liu, 2010).在CBF调控通路中, DEAR1和RAP2.1如何整合还需深入研究.RAP2.1是否与其它DEAR转录因子共同调节植物非生物胁迫响应仍不清楚.研究发现DREB-A5亚组有6种含EAR基序的转录抑制子可与DRE元件结合, 进而调控非生物胁迫响应基因的表达(Nakano et al., 2006). ...
... 对低温更为敏感(Dong and Liu, 2010).在CBF调控通路中, DEAR1和RAP2.1如何整合还需深入研究.RAP2.1是否与其它DEAR转录因子共同调节植物非生物胁迫响应仍不清楚.研究发现DREB-A5亚组有6种含EAR基序的转录抑制子可与DRE元件结合, 进而调控非生物胁迫响应基因的表达(Nakano et al., 2006). ...
... DREB-A2亚组转录因子DREB2主要参与调节植物的抗旱和耐热性(Mizoi et al., 2012).干旱和高温可诱导DREB2基因的表达, 进而正向调控下游干旱或热胁迫响应基因(Maruyama et al., 2009).研究表明DREB-A4家族成员HRD (hardy)和DREB-A6家族成员(ERF53、RAP2.4和TG/RAP2.4a)也在干旱和耐盐性调控中发挥关键作用(Karaba et al., 2007; Lin et al., 2008; Chen et al., 2012; Zhu et al., 2014).过表达HRD能显著增强拟南芥和水稻植株的耐旱和耐盐性(Karaba et al., 2007).同样, 过表达TG (translucent green)导致植株叶片玻璃化, 叶片细胞含水量增加(Zhu et al., 2014).TG也参与调控抗坏血酸过氧化物酶基因APX (ascorbate peroxidase)的表达, 抵御活性氧(reactive oxygen species, ROS)引起的光氧化胁迫(Rudnik et al., 2017).除拟南芥外, 人们对水稻、玉米、番茄(Lycopersicon esculentum)、葡萄(Vitis vinifera)等作物的DREB1和DREB2转录因子也进行了广泛的研究(Xiao et al., 2006; Matsukura et al., 2010; Dou et al., 2014). ...
Ethylene Response Factor 6 acts as a central regulator of leaf growth under water-limiting conditions in Arabidopsis 1 2013
... AP2/ERFs参与调控植物GA生物合成相关基因的转录.盐胁迫下, DREB1E和DREB1F抑制GA20ox的表达, 使GA生物合成减少, 引起植株生长迟缓(Magome et al., 2004).过表达ERF6抑制GA2ox的表达水平, 诱导赤霉素降解酶(gibberellin 2-oxidase 6, GA2ox6)的合成, 维持DELLA蛋白的稳定性.渗透胁迫下, erf6功能获得突变体植株矮小且对胁迫高度敏感, 而erf5/erf6缺失突变体生长受胁迫影响较小.研究发现ERF6还激活了STZ、MYB51和WRKY33等渗透胁迫响应基因的表达(Dubois et al., 2013).冷胁迫下, CBF1转录因子激发GA2ox基因的表达而降低GA含量, 组成型表达CBF1抑制GA的积累, 植株表现矮化, 表明DELLA蛋白是CBF1介导的冷胁迫响应的关键组分(Achard et al., 2008).相反, ERF11作为一种GA生物合成和信号转导的正调控因子, 抑制ET生物合成, 促进植株节间伸长.过表达ERF11上调GA3ox1和GA20ox基因的表达水平, 增加GA的积累量(Zhou et al., 2016).ERF6作为转录激活因子诱导ERF11的表达, 反之, ERF11抑制GA2-OX6、STZ、MYB51和WRKY33等ERF6相关靶基因的表达.进一步实验表明, 4个靶基因在erf11/erf6过表达植株中也不能被诱导表达, ERF11过表达可消除ERF6过表达导致的极度矮化现象, 说明ERF11可能通过直接竞争目标基因启动子而在分子水平上拮抗ERF6 (Dubois et al., 2015). ...
The ETHYLENE RESPONSE FACTORs ERF6 and ERF11 antagonistically regulate mannitol-induced growth inhibition in Arabidopsis 1 2015
... AP2/ERFs参与调控植物GA生物合成相关基因的转录.盐胁迫下, DREB1E和DREB1F抑制GA20ox的表达, 使GA生物合成减少, 引起植株生长迟缓(Magome et al., 2004).过表达ERF6抑制GA2ox的表达水平, 诱导赤霉素降解酶(gibberellin 2-oxidase 6, GA2ox6)的合成, 维持DELLA蛋白的稳定性.渗透胁迫下, erf6功能获得突变体植株矮小且对胁迫高度敏感, 而erf5/erf6缺失突变体生长受胁迫影响较小.研究发现ERF6还激活了STZ、MYB51和WRKY33等渗透胁迫响应基因的表达(Dubois et al., 2013).冷胁迫下, CBF1转录因子激发GA2ox基因的表达而降低GA含量, 组成型表达CBF1抑制GA的积累, 植株表现矮化, 表明DELLA蛋白是CBF1介导的冷胁迫响应的关键组分(Achard et al., 2008).相反, ERF11作为一种GA生物合成和信号转导的正调控因子, 抑制ET生物合成, 促进植株节间伸长.过表达ERF11上调GA3ox1和GA20ox基因的表达水平, 增加GA的积累量(Zhou et al., 2016).ERF6作为转录激活因子诱导ERF11的表达, 反之, ERF11抑制GA2-OX6、STZ、MYB51和WRKY33等ERF6相关靶基因的表达.进一步实验表明, 4个靶基因在erf11/erf6过表达植株中也不能被诱导表达, ERF11过表达可消除ERF6过表达导致的极度矮化现象, 说明ERF11可能通过直接竞争目标基因启动子而在分子水平上拮抗ERF6 (Dubois et al., 2015). ...
Arabidopsis RAV1 transcription factor, phosphorylated by SnRK2 kinases, regulates the expression of ABI3, ABI4, and ABI5 during seed germination and early seedling development 2 2014
... 翻译后修饰(post-translational modification, PTM)是蛋白质合成途径的后期加工过程, 不同的修饰方式对AP2/ERF蛋白的活性、稳定性和丰度的影响有差异.磷酸化是转录因子的主要修饰途径之一, 磷酸化水平与下游靶基因的表达水平关系密切.拟南芥ABA信号通路的正调控因子蔗糖非酵解型蛋白激酶(Snf1-re-lated protein kinases, SnRKs)可磷酸化RAV1, 进而抑制RAV1的转录阻遏效应(Feng et al., 2014), 而ERF104和ERF6可被后MPK3/6 (mitogen-activated protein kinase 3/6)磷酸化修饰, 并在下游级联反应中发挥重要作用, 调控植物对病原体的免疫反应(Meng et al., 2013).DREB2A是拟南芥的关键转录激活因子, 在其中央序列存在一个负调控域(negative regulatory domain, NRD).热胁迫条件下, NRD中的Ser/Thr残基磷酸化水平降低, DREB2A蛋白稳定性提高, 敲除NRD后DREB2A呈组成型激活, 表明抑制NRD磷酸化可以稳定和激活DREB2A的表达, 从而增强植物的耐热性(Mizoi et al., 2019).泛素介导的蛋白质降解(26S蛋白酶体途径)也参与调节AP2/ ERF蛋白的稳定性.非生物胁迫条件下, DREB2A和ERF75/RAP-2.2蛋白分别被RING家族E3泛素连接酶DRIP1/2 (DREB2A-interacting protein1/2)和SINAT2 (seven in absentia of Arabidopsis 2)直接泛素化(Cheng et al., 2012; Papdi et al., 2015).研究发现CUL3-E3连接酶适配子BPMs (BTB/POZ and math domain)可与DREB2A蛋白的负调控域NRD在核内发生互作, 敲除BPM增加DREB2A蛋白的积累, 促进下游靶基因的表达, 提高植株的耐热和耐旱能力, 这也说明BPM对DREB2A蛋白的稳定性起负调控作用(Morimoto et al., 2017).此外, 研究发现ERF-VII类蛋白可在质膜上与乙酰辅基A结合蛋白(Acyl-CoA binding protein, ACBP)互作.在缺氧条件下, RAP2.12与ACBP分离后进入细胞核, 在脱酰胺基和精氨酰基化的作用下通过氧依赖途径移除ERF72/RAP2.3、ERF74/RAP2.12蛋白的甲硫氨酸N末端, 将半胱氨酸氧化成半胱亚磺酸, 引发蛋白快速降解并暴露出非稳定N端的氨基酸残基(Gibbs et al., 2015; Abbas et al., 2015). ...
... DREB-A3亚组成员ABI4 (ABA insensitive 4)是ABA信号通路的重要组分.植株受到胁迫后, 体内积累ABA和ROS, ABI4可被ABA、ROS及糖质体等多种信号通路靶向调控.CCAAT结合因子A (CCAAT binding factor A, CBFA)是四吡咯化合物HAP (heme activator protein)三聚体转录复合物的亚基, 当产生过量ROS时, ABI4可与其它转录因子竞争性地结合CCACGT元件, 抑制CBFA的表达, 进而调控质体反向信号通路中相关基因的表达(Zhang et al., 2013).拟南芥突变体实验表明, ABI4在YL1 (yellow leaf 1) (Li et al., 2016)和DPG1 (delayed pale-greening 1) (Yi et al., 2019)等基因参与盐胁迫响应的调控网络中也发挥重要作用.逆境胁迫下, ORA47在JA (jasmo-nate acid)和ABA信号转导途径中靶向调节ABA和JA诱导的下游应答基因, ERF18/ORA-47识别启动子顺式元件(NC/GT)CGNCCA (O-box), 激活PP2C家族ABI2转录因子的表达.ABI1则在ORA-47上游起作用, 从而形成ABI1-ORA47-ABI2正反馈基因表达回路, 调控ABA信号转导和干旱胁迫响应(Chen et al., 2016).此外, RAV1转录因子在ABA信号转导途径中也发挥关键作用, RAV1能与ABI3、ABI4和ABI5启动子结合, 其过表达抑制ABI3、ABI4和ABI5的转录活性, 导致根系生长发育过程中对ABA不敏感.同时, 蔗糖非酵解型蛋白激酶(SnRK2.2、SnRK2.3和SnRK2.6)在核内磷酸化RAV1, 降低ABI5对RAV1的依赖性, 并负调控RAV1对靶基因的转录抑制作用(Feng et al., 2014). ...
Abscisic acid synthesis and response 1 2013
... 植物激素是一类调控植物生长发育以及抵御不良环境影响的重要小分子.脱落酸(ABA)是应答干旱、盐、冷、热等非生物胁迫的关键激素之一.ABA可诱导气孔关闭, 调节根系结构, 促进渗透物质的合成, 进而抵御外部逆境(Sah et al., 2016).NCED是ABA合成的限速酶, 干旱和渗透胁迫诱导NCED基因快速上调表达, 促进ABA的生物合成.ABA与其受体PYR/PLY/ RCAR结合, 再与PP2C作用形成复合物, 解除PP2C对SnRK2激酶活性的抑制作用.活性形式的SnRK2磷酸化AREB/ABF、离子通道蛋白和NADPH氧化酶等下游底物, 进而诱导ABA响应基因的表达(Finkelstein, 2013; Sah et al., 2016). ...
Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF cold response pathway 1 2002
... 部分A5亚组DREB类转录因子则对上述调控网络具有负反馈调节作用.DEAR1 (DREB and EAR motif protein 1)可能既在CBF上游作用, 又与RAP2.1在CBF下游扮演冷胁迫响应的调节子, 调节RD29A/COR78、COR15A和KIN1等冷诱导相关基因的表达, 负调控植物的耐冷和耐旱性(Fowler and Thomashow, 2002; Tsutsui et al., 2009; Dong and Liu, 2010).DEAR1过表达抑制冷诱导CBF的表达, 进而降低植物的耐冷能力(Tsutsui et al., 2009).低温胁迫或组成型表达CBF可诱导RAP2.1的表达, CBF2比RAP2.1对低温更为敏感(Dong and Liu, 2010).在CBF调控通路中, DEAR1和RAP2.1如何整合还需深入研究.RAP2.1是否与其它DEAR转录因子共同调节植物非生物胁迫响应仍不清楚.研究发现DREB-A5亚组有6种含EAR基序的转录抑制子可与DRE元件结合, 进而调控非生物胁迫响应基因的表达(Nakano et al., 2006). ...
DNA-binding specificities of plant transcription factors and their potential to define target genes 4 2014
... AP2/ERF转录因子通过特异性结合胁迫应答基因启动子区顺式元件, 参与调控植物的逆境防御反应.DREB亚族成员特异性识别并结合启动子区域的DRE/CRT (dehydration responsive element/C-re-peat)元件(核心序列A/GCCGAC), 诱导干旱、低温和盐胁迫相关应答基因的表达.ERF亚族成员则可通过与乙烯响应元件(ethylene response element, ERE) (核心序列AGCCGCC, 又称GCC-box)结合, 从而参与调控乙烯应答和非生物胁迫(Franco-Zorrilla et al., 2014).此外, 研究发现许多AP2/ERF转录因子可同时与DRE/CRT和ERE两种元件结合, 如DREB亚族成员TINY、CBF1、ERF53、RAP2.4、TG/RAP2.4a以及ERF亚族成员ERF1、ERF4和ERF71 (Chen et al., 2012; Zhu et al., 2014; Lee et al., 2015), 但结合能力有差异.在拟南芥中, DREB2C和DEAR3与类GCC-box元件(GCCGCC)的结合能力远超DRE元件(Franco-Zorrilla et al., 2014).AP2/ERF转录因子还能与DRE和ERE之外的顺式元件结合, 如偶联元件1 (CE1, TGCCACCG)、类偶联元件3 (CE-3like, CGCG)和缺氧响应启动子元件(hypoxia-responsive promoter element, HRPE), 以及ATCTA、CAACA、CATGCA、CAA/CA/CTG、ATCGAG和(NC/GT) CGNCCA (Bossi et al., 2009; Zhu et al., 2010; Chen et al., 2016; Gasch et al., 2016; Park et al., 2016).近年来, 科学家通过蛋白质结合微阵列(protein binding microarray, PBM)技术发现结构特征高度相似的转录因子具有相似的DNA结合位点, 同时也具有相似的生物学功能.该研究结果合理地解释了转录因子功能冗余现象, 也为预测和挖掘未知AP2/ERF转录因子提供了高效可行的技术方法(Franco-Zorrilla et al., 2014). ...
... ), 但结合能力有差异.在拟南芥中, DREB2C和DEAR3与类GCC-box元件(GCCGCC)的结合能力远超DRE元件(Franco-Zorrilla et al., 2014).AP2/ERF转录因子还能与DRE和ERE之外的顺式元件结合, 如偶联元件1 (CE1, TGCCACCG)、类偶联元件3 (CE-3like, CGCG)和缺氧响应启动子元件(hypoxia-responsive promoter element, HRPE), 以及ATCTA、CAACA、CATGCA、CAA/CA/CTG、ATCGAG和(NC/GT) CGNCCA (Bossi et al., 2009; Zhu et al., 2010; Chen et al., 2016; Gasch et al., 2016; Park et al., 2016).近年来, 科学家通过蛋白质结合微阵列(protein binding microarray, PBM)技术发现结构特征高度相似的转录因子具有相似的DNA结合位点, 同时也具有相似的生物学功能.该研究结果合理地解释了转录因子功能冗余现象, 也为预测和挖掘未知AP2/ERF转录因子提供了高效可行的技术方法(Franco-Zorrilla et al., 2014). ...
... 许多研究表明, AP2/ERF转录因子可直接与靶基因启动子结合, 激活或抑制下游靶基因的表达, 其N端DNA结合域和C端转录激活域在胁迫相关靶基因的转录激活中起重要作用(Nakano et al., 2006).进一步分析表明, AP2/ERF转录因子中含有EAR基序(LxLxL或DLNxxP)、TLLLFR基序和BRD结构域(B3 repression domain, 保守序列为R/KLFGV)等多个转录抑制结构域(Ikeda and Ohme-Takagi, 2009; Kagale and Rozwadowski, 2011; Deepika et al., 2016).含EAR基序的转录因子是一种重要的抑制子, 在植物非生物胁迫应答基因的诱导表达中起负调控作用.Tiwari等(2012)从拟南芥ERF和AP2亚族中发现了一种转录激活关键元件EDLL, EDLL可与EAR基序(ERF-associated amphiphilic repression)发生互作, 拮抗后者介导的转录抑制作用.而不同的AP2/ ERF转录因子也可协同招募TPL (topless)和TPR (topless-related)等转录共抑制子(Causier et al., 2012)或组蛋白修饰因子, 进而抑制下游靶基因的表达(Song et al., 2005; Song and Galbraith, 2006).Song等(2005)发现AtERF7特异性地结合GCC-box, 与抑制因子ATSIN3及组蛋白去乙酰化酶19 (histone deacetylase 19, HDA19)发生互作并形成转录因子复合物, 参与干旱胁迫和ABA响应调控.AtERF7过表达株系保卫细胞对ABA的敏感性减弱, 水分散失增加; 反之, AtERF7 RNA干扰系对ABA的敏感性增强.类似的, ERF3能与组蛋白去乙酰化酶复合物亚基SAP18 (SIN3 associated polypeptide P18)互作, 共同招募HDA19形成复合体, 进而抑制相关基因的表达(Song and Galbraith, 2006).BRD结构域被认为是RAV1和RAV2转录因子行使抑制功能的关键元件, 其保守序列中第1位氨基酸残基多为疏水性亮氨酸、缬氨酸和蛋氨酸残基, 突变导致功能丧失(Ikeda and Ohme-Takagi, 2009).研究显示, 与AP2/ERFs协同调控的基因同样可被AP2/ERFs靶基因富集, 分析此类基因的同源基序有助于识别转录因子的假定靶基因并预测其生物学功能(Franco-Zorrillaetal et al., 2014). ...
A subset of Arabidopsis RAV transcription factors modulates drought and salt stress responses independent of ABA 2 2014
... RAVs和AP2s除调节茎与根细胞有丝分裂以及开花等发育过程(Osnato et al., 2012; Horstman et al., 2014)外, 还参与调控非生物胁迫响应.干旱和盐胁迫下, 拟南芥RAV1、RAV1L和RAV2的表达水平下降, 但三者对ABA的敏感性不同.进一步研究发现, 野生型、rav1突变体和RAV1超表达植株中RD29A、RD29B以及9-顺式环氧类胡萝卜素双加氧酶基因NCED9 (nine-cis-epxycarotenoid dioxygenase)和NCED3表现出相同的胁迫响应模式(Fu et al., 2014).Saito等(2004)发现过表达RAV1抑制ABA降解基因(CYP707A1和CYP707A2)的活性.正常条件下, RAV1过表达提高了NCED9和NCED3基因的表达水平, 而受胁迫后植株体内的ABA水平未发生明显变化(Fu et al., 2014).以上说明RAV1可通过不依赖ABA的途径负调控植物的非生物胁迫响应.此外, 研究发现ANT (AINTEGUMENTA)控制发育中的根细胞数量和大小, 同时也可抑制类SOS3钙结合蛋白8 (sos3-like calcium binding protein 8, SCABP8)转录, 负调控耐盐性, 而ANT功能缺失增强了拟南芥幼苗的耐盐能力, 并维持离子稳态(Meng et al., 2015a). ...
... 基因的表达水平, 而受胁迫后植株体内的ABA水平未发生明显变化(Fu et al., 2014).以上说明RAV1可通过不依赖ABA的途径负调控植物的非生物胁迫响应.此外, 研究发现ANT (AINTEGUMENTA)控制发育中的根细胞数量和大小, 同时也可抑制类SOS3钙结合蛋白8 (sos3-like calcium binding protein 8, SCABP8)转录, 负调控耐盐性, 而ANT功能缺失增强了拟南芥幼苗的耐盐能力, 并维持离子稳态(Meng et al., 2015a). ...
Submergence tolerance conferred by Sub1A is mediated by SLR1 and SLRL1 restriction of gibberellin responses in rice 1 2008
... 在拟南芥和水稻中, SUB1A、SK1和SK2参与GA信号转导通路的调控.淹水胁迫下, SUB1A激活SLR1 (slender rice 1)和SLRL1 (slender rice 1 like 1) (DELLA类似蛋白)转录, 同时SUB1A抑制受GA诱导的淀粉代谢相关基因的表达, 调控植株节间伸长和呼吸作用(Fukao and Bailey-Serres, 2008; Locke et al., 2018; Perata, 2018).SK1和SK2两个转录因子中均含有EIN3结合位点, 电泳迁移率测定显示, 类EIN3基因EIL1b可与SK1和SK2启动子结合, 激活其转录活性.研究发现, 淹水状态下拟南芥中ET合成增加, 进而诱导SK1和SK2表达, 激发GA20oxs基因上调表达, 促进植株节间伸长, 减轻淹水对其生长的抑制效应(Hattori et al., 2009; Ayano et al., 2014). ...
Identification of ICE2, a gene involved in cold acclimation which determines freezing tolerance in Arabidopsis thaliana 0 2009
Redundant ERF-VII transcription factors bind to an evolutionarily conserved cis-motif to regulate hypoxia-responsive gene expression in Arabidopsis 1 2016
... AP2/ERF转录因子通过特异性结合胁迫应答基因启动子区顺式元件, 参与调控植物的逆境防御反应.DREB亚族成员特异性识别并结合启动子区域的DRE/CRT (dehydration responsive element/C-re-peat)元件(核心序列A/GCCGAC), 诱导干旱、低温和盐胁迫相关应答基因的表达.ERF亚族成员则可通过与乙烯响应元件(ethylene response element, ERE) (核心序列AGCCGCC, 又称GCC-box)结合, 从而参与调控乙烯应答和非生物胁迫(Franco-Zorrilla et al., 2014).此外, 研究发现许多AP2/ERF转录因子可同时与DRE/CRT和ERE两种元件结合, 如DREB亚族成员TINY、CBF1、ERF53、RAP2.4、TG/RAP2.4a以及ERF亚族成员ERF1、ERF4和ERF71 (Chen et al., 2012; Zhu et al., 2014; Lee et al., 2015), 但结合能力有差异.在拟南芥中, DREB2C和DEAR3与类GCC-box元件(GCCGCC)的结合能力远超DRE元件(Franco-Zorrilla et al., 2014).AP2/ERF转录因子还能与DRE和ERE之外的顺式元件结合, 如偶联元件1 (CE1, TGCCACCG)、类偶联元件3 (CE-3like, CGCG)和缺氧响应启动子元件(hypoxia-responsive promoter element, HRPE), 以及ATCTA、CAACA、CATGCA、CAA/CA/CTG、ATCGAG和(NC/GT) CGNCCA (Bossi et al., 2009; Zhu et al., 2010; Chen et al., 2016; Gasch et al., 2016; Park et al., 2016).近年来, 科学家通过蛋白质结合微阵列(protein binding microarray, PBM)技术发现结构特征高度相似的转录因子具有相似的DNA结合位点, 同时也具有相似的生物学功能.该研究结果合理地解释了转录因子功能冗余现象, 也为预测和挖掘未知AP2/ERF转录因子提供了高效可行的技术方法(Franco-Zorrilla et al., 2014). ...
Group VII ethylene response factors coordinate oxygen and nitric oxide signal transduction and stress responses in plants 5 2015
... 植物在生长发育过程中经常受到生物或非生物胁迫的影响, 由此, 植物逐渐形成了复杂的调节机制感知胁迫信号, 对外部环境变化做出快速响应(郭倩倩和周文彬, 2019).脱落酸(abscisic acid, ABA)、乙烯(ethylene, ET)及赤霉素(gibberellin, GA)等激素作为信号分子能够调节植物的生长发育、形态建成以及抗逆反应等生物学过程.该过程也受转录因子(transcription factor, TF)调控, 即转录因子通过依赖或不依赖于激素介导的信号通路参与调控植物对干旱、低温、缺氧、高盐和淹水等非生物胁迫的响应, 进而影响植物的逆境耐受能力(Gibbs et al., 2015; Verma et al., 2016; Li et al., 2018).AP2/ERF (APETALA2/eth-ylene responsive factor)是植物特有的转录因子家族之一, 已在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、高粱(Sorghum bicolor)、大白菜(Brassica pekinensis)、毛竹(Phyllostachys edulis)、玉米(Zea mays)、大麦(Hordeum vulgare)和小麦(Triticum aestivum)等物种中被成功分离鉴定(Xu and Chua, 2011; Chandler, 2018).通过突变体实验, 科学家发现了许多与AP2/ERF相关的非生物胁迫或激素信号应答基因.目前, AP2/ERF已成为研究非生物胁迫和激素互作的热点候选基因.本文从AP2/ERF转录因子的结构特征、转录调控、翻译后修饰、结合位点、协同互作蛋白以及其参与调控依赖或不依赖激素信号途径的非生物胁迫响应进行综述, 为进一步阐明AP2/ERF转录因子在非生物胁迫调控网络中的作用提供理论依据. ...
... 翻译后修饰(post-translational modification, PTM)是蛋白质合成途径的后期加工过程, 不同的修饰方式对AP2/ERF蛋白的活性、稳定性和丰度的影响有差异.磷酸化是转录因子的主要修饰途径之一, 磷酸化水平与下游靶基因的表达水平关系密切.拟南芥ABA信号通路的正调控因子蔗糖非酵解型蛋白激酶(Snf1-re-lated protein kinases, SnRKs)可磷酸化RAV1, 进而抑制RAV1的转录阻遏效应(Feng et al., 2014), 而ERF104和ERF6可被后MPK3/6 (mitogen-activated protein kinase 3/6)磷酸化修饰, 并在下游级联反应中发挥重要作用, 调控植物对病原体的免疫反应(Meng et al., 2013).DREB2A是拟南芥的关键转录激活因子, 在其中央序列存在一个负调控域(negative regulatory domain, NRD).热胁迫条件下, NRD中的Ser/Thr残基磷酸化水平降低, DREB2A蛋白稳定性提高, 敲除NRD后DREB2A呈组成型激活, 表明抑制NRD磷酸化可以稳定和激活DREB2A的表达, 从而增强植物的耐热性(Mizoi et al., 2019).泛素介导的蛋白质降解(26S蛋白酶体途径)也参与调节AP2/ ERF蛋白的稳定性.非生物胁迫条件下, DREB2A和ERF75/RAP-2.2蛋白分别被RING家族E3泛素连接酶DRIP1/2 (DREB2A-interacting protein1/2)和SINAT2 (seven in absentia of Arabidopsis 2)直接泛素化(Cheng et al., 2012; Papdi et al., 2015).研究发现CUL3-E3连接酶适配子BPMs (BTB/POZ and math domain)可与DREB2A蛋白的负调控域NRD在核内发生互作, 敲除BPM增加DREB2A蛋白的积累, 促进下游靶基因的表达, 提高植株的耐热和耐旱能力, 这也说明BPM对DREB2A蛋白的稳定性起负调控作用(Morimoto et al., 2017).此外, 研究发现ERF-VII类蛋白可在质膜上与乙酰辅基A结合蛋白(Acyl-CoA binding protein, ACBP)互作.在缺氧条件下, RAP2.12与ACBP分离后进入细胞核, 在脱酰胺基和精氨酰基化的作用下通过氧依赖途径移除ERF72/RAP2.3、ERF74/RAP2.12蛋白的甲硫氨酸N末端, 将半胱氨酸氧化成半胱亚磺酸, 引发蛋白快速降解并暴露出非稳定N端的氨基酸残基(Gibbs et al., 2015; Abbas et al., 2015). ...
... ERF亚族成员在低氧和淹水胁迫响应过程中也发挥重要作用(Bui et al., 2015; Gibbs et al., 2015).SUB1A (submergence 1A)是ERF亚族B2亚组成员之一, Sub1A-1超表达的同时促进乙醇脱氢酶基因(alcohol dehydrogenase 1, Adh1)上调表达, 抑制Sub1C的转录水平, 说明Sub1A-1是耐淹性的主要调节因子, 在淹水胁迫应答调控中起关键作用(Xu et al., 2006).淹水状态下, 低水平海藻糖-6-磷酸合酶(trehalose-6-phosphate, T6P)与CIPK15 (CBL-inter-acting protein kinase 15)协同激活SnRK1, SnRK1介导SUB1A抑制幼苗胚轴生长和呼吸相关基因的表达, 直接或间接调控碳同化过程.而SK1/SK2 (snorkel1/2)激活参与叶柄、茎和导管发育相关基因, 促进植株快速生长(Hattori et al., 2009; Locke et al., 2018).缺氧胁迫下, 拟南芥ERF-VIIs可以通过氧依赖的N端规则通路降解(Gibbs et al., 2015).RING (really interesting gene)类蛋白SINAT2可通过其N端保守的C3H4型RING结构域调控RAP2.12, 沉默SINAT1/2可上调RAP2.12的表达水平(Papdi et al., 2015).此外, ERF71/HRE2、ERF72/RAP2.3、ERF73/HRE1、ERF74/RAP2.12及ERF75/RAP2.2d也受缺氧胁迫诱导, 进而正向调控糖代谢和激素信号相关的缺氧响应基因, 提高植株的缺氧耐受能力.过表达RAP2.2、RAP2.3和RAP2.12可提高植株在低氧和渗透胁迫下的存活率, 而rap2.12-2/rap2.3-1双突变体对淹水和渗透胁迫表现出较高的敏感性(Yao et al., 2017). ...
... ).缺氧胁迫下, 拟南芥ERF-VIIs可以通过氧依赖的N端规则通路降解(Gibbs et al., 2015).RING (really interesting gene)类蛋白SINAT2可通过其N端保守的C3H4型RING结构域调控RAP2.12, 沉默SINAT1/2可上调RAP2.12的表达水平(Papdi et al., 2015).此外, ERF71/HRE2、ERF72/RAP2.3、ERF73/HRE1、ERF74/RAP2.12及ERF75/RAP2.2d也受缺氧胁迫诱导, 进而正向调控糖代谢和激素信号相关的缺氧响应基因, 提高植株的缺氧耐受能力.过表达RAP2.2、RAP2.3和RAP2.12可提高植株在低氧和渗透胁迫下的存活率, 而rap2.12-2/rap2.3-1双突变体对淹水和渗透胁迫表现出较高的敏感性(Yao et al., 2017). ...
... ERF亚家族转录因子是ET介导的胁迫应答信号通路下游的关键调控因子(Licausi et al., 2013; Gibbs et al., 2015; Kazan, 2015; Müller and Munné-Bosch, 2015).冷胁迫下, 植物体内的ET主要起负调控作用, 过量ET或1-氨基环丙烷-1-羧酸降低植物的耐冷性, 而ET生物合成抑制剂氨基乙氧基甘氨酸或感知拮抗剂Ag+具有相反的作用(Shi et al., 2012).etr1-1、ein4-1、ein2-5、ein3-1和ein3eil突变体表现出更强的耐冷性, 但组成型ET响应etr1-1和ein3过表达植株表现出耐冷性减弱; 冷胁迫诱导ETR1、EIN4、EBF1及EBF2快速上调表达, 而EIN2和EIL1下调表达.进一步分析表明, EIN3通过与启动子区特定元件结合, 抑制ARR5、ARR7、ARR15和CBF转录, ARRs作为冷信号通路的关键节点基因, 在ET和CTK介导的胁迫响应网络中起着关键作用, 过表达ARRs使植株的耐冷能力增强(Shi et al., 2012).此外, EIN3可激活ERF1和ESEs, 进而参与调控下游胁迫相关基因, 提高植物的耐盐性(Kazan, 2015; Tao et al., 2015).35S:AtERF4-GFP转基因植株中ABI2、RD29B和RAP18的表达受抑制, 进一步说明AtERF4是ET和ABA信号途径的负调控因子(Yang et al., 2011). ...
Mechanisms and networks for brassinosteroid regulated gene expression 1 2013
... 油菜素内酯(BR)在植物细胞伸长、叶片发育、花粉管生长、木质部分化、衰老、光形态发生以及应激反应过程中起重要作用(Ye et al., 2017).BR信号始于细胞膜上受体激酶(brassinosteroid insensitive, BRI), BKI1 (BRI kinase inhibitor 1)是BRI1的关键负调控蛋白.无BR时, BKI1和14-3-3蛋白分别抑制BR的受体BRI1和转录因子BES1 (bri1-ems-suppressor 1) (Wang et al., 2011).而当BR被BRI1感知后, 即促使BKI1发生磷酸化, 激活BRI1和BES1以调控负调节因子BIN2 (brassinosteroid insensitive 2)的表达, 诱导BES1和BZR1 (brassinazole-resistant 1)积累并与转录因子互作, 影响参与植物生长和胁迫反应的BR应答基因的转录水平(Wang et al., 2011; Guo et al., 2013). ...
The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water 2 2009
... ERF亚族成员在低氧和淹水胁迫响应过程中也发挥重要作用(Bui et al., 2015; Gibbs et al., 2015).SUB1A (submergence 1A)是ERF亚族B2亚组成员之一, Sub1A-1超表达的同时促进乙醇脱氢酶基因(alcohol dehydrogenase 1, Adh1)上调表达, 抑制Sub1C的转录水平, 说明Sub1A-1是耐淹性的主要调节因子, 在淹水胁迫应答调控中起关键作用(Xu et al., 2006).淹水状态下, 低水平海藻糖-6-磷酸合酶(trehalose-6-phosphate, T6P)与CIPK15 (CBL-inter-acting protein kinase 15)协同激活SnRK1, SnRK1介导SUB1A抑制幼苗胚轴生长和呼吸相关基因的表达, 直接或间接调控碳同化过程.而SK1/SK2 (snorkel1/2)激活参与叶柄、茎和导管发育相关基因, 促进植株快速生长(Hattori et al., 2009; Locke et al., 2018).缺氧胁迫下, 拟南芥ERF-VIIs可以通过氧依赖的N端规则通路降解(Gibbs et al., 2015).RING (really interesting gene)类蛋白SINAT2可通过其N端保守的C3H4型RING结构域调控RAP2.12, 沉默SINAT1/2可上调RAP2.12的表达水平(Papdi et al., 2015).此外, ERF71/HRE2、ERF72/RAP2.3、ERF73/HRE1、ERF74/RAP2.12及ERF75/RAP2.2d也受缺氧胁迫诱导, 进而正向调控糖代谢和激素信号相关的缺氧响应基因, 提高植株的缺氧耐受能力.过表达RAP2.2、RAP2.3和RAP2.12可提高植株在低氧和渗透胁迫下的存活率, 而rap2.12-2/rap2.3-1双突变体对淹水和渗透胁迫表现出较高的敏感性(Yao et al., 2017). ...
... 在拟南芥和水稻中, SUB1A、SK1和SK2参与GA信号转导通路的调控.淹水胁迫下, SUB1A激活SLR1 (slender rice 1)和SLRL1 (slender rice 1 like 1) (DELLA类似蛋白)转录, 同时SUB1A抑制受GA诱导的淀粉代谢相关基因的表达, 调控植株节间伸长和呼吸作用(Fukao and Bailey-Serres, 2008; Locke et al., 2018; Perata, 2018).SK1和SK2两个转录因子中均含有EIN3结合位点, 电泳迁移率测定显示, 类EIN3基因EIL1b可与SK1和SK2启动子结合, 激活其转录活性.研究发现, 淹水状态下拟南芥中ET合成增加, 进而诱导SK1和SK2表达, 激发GA20oxs基因上调表达, 促进植株节间伸长, 减轻淹水对其生长的抑制效应(Hattori et al., 2009; Ayano et al., 2014). ...
Arabidopsis RAP2.2: an ethylene response transcription factor that is important for hypoxia survival 2 2010
... 此外, ET促进RAP2.3转录因子定位于细胞核内, 通过依赖于RAP2.3的方式激发ORA59介导的ET响应过程(Kim et al., 2018).而ERF-VIIs通过ET不依赖途径参与调控缺氧响应.在ET不敏感突变体或生物合成抑制剂存在的情况下, 缺氧依然可以诱导ERF73/HRE1的表达, ERF-VIIs也可能通过反馈调节负调控ET信号(Hinz et al., 2010; Yang et al., 2011).研究发现, ET合酶ACS在RAP2.2过表达植株中表达量降低, 而在rap2.2-2敲除突变体中表达上调(Hinz et al., 2010).然而, 胁迫条件下ERF-VIIs如何通过负反馈机制调控ET生物合成有待进一步研究. ...
... 敲除突变体中表达上调(Hinz et al., 2010).然而, 胁迫条件下ERF-VIIs如何通过负反馈机制调控ET生物合成有待进一步研究. ...
AINTEGUMENTA-LIKE proteins: hubs in a plethora of networks 1 2014
... RAVs和AP2s除调节茎与根细胞有丝分裂以及开花等发育过程(Osnato et al., 2012; Horstman et al., 2014)外, 还参与调控非生物胁迫响应.干旱和盐胁迫下, 拟南芥RAV1、RAV1L和RAV2的表达水平下降, 但三者对ABA的敏感性不同.进一步研究发现, 野生型、rav1突变体和RAV1超表达植株中RD29A、RD29B以及9-顺式环氧类胡萝卜素双加氧酶基因NCED9 (nine-cis-epxycarotenoid dioxygenase)和NCED3表现出相同的胁迫响应模式(Fu et al., 2014).Saito等(2004)发现过表达RAV1抑制ABA降解基因(CYP707A1和CYP707A2)的活性.正常条件下, RAV1过表达提高了NCED9和NCED3基因的表达水平, 而受胁迫后植株体内的ABA水平未发生明显变化(Fu et al., 2014).以上说明RAV1可通过不依赖ABA的途径负调控植物的非生物胁迫响应.此外, 研究发现ANT (AINTEGUMENTA)控制发育中的根细胞数量和大小, 同时也可抑制类SOS3钙结合蛋白8 (sos3-like calcium binding protein 8, SCABP8)转录, 负调控耐盐性, 而ANT功能缺失增强了拟南芥幼苗的耐盐能力, 并维持离子稳态(Meng et al., 2015a). ...
Functional characterization of an abiotic stress-inducible transcription factor AtERF53 in 1 2013
... 研究表明, 非生物胁迫下ABA可促进ANT (Meng et al., 2015b)、ERF53 (Hsieh et al., 2013)、RAP2.6L (Liu et al., 2012)和RAP2.6 (Zhu et al., 2010)转录, 上述转录因子进而结合胁迫应答顺式作用元件DRE/ABRE, 调控下游相关靶基因的表达, 增强植物对非生物胁迫的耐受能力.在水稻中ERF71正向调控ABA信号转导, 过表达ERF71可促进根中ABI5、PP2C68、RAB16C及RAB16D等基因上调表达.此外, ERF71可直接调控木质素生物合成基因CCR1、CCR10及C4H转录, 诱导根系径向生长, 改变根系构型, 提高耐旱能力(Lee et al., 2017; Li et al., 2018).盐胁迫和ABA双重诱导下, ABA增强DREB2的转录活性, 进而促进Rd29A协同表达(Lee et al., 2016). ...
A novel group of transcriptional repressors in Arabidopsis 2 2009
... 许多研究表明, AP2/ERF转录因子可直接与靶基因启动子结合, 激活或抑制下游靶基因的表达, 其N端DNA结合域和C端转录激活域在胁迫相关靶基因的转录激活中起重要作用(Nakano et al., 2006).进一步分析表明, AP2/ERF转录因子中含有EAR基序(LxLxL或DLNxxP)、TLLLFR基序和BRD结构域(B3 repression domain, 保守序列为R/KLFGV)等多个转录抑制结构域(Ikeda and Ohme-Takagi, 2009; Kagale and Rozwadowski, 2011; Deepika et al., 2016).含EAR基序的转录因子是一种重要的抑制子, 在植物非生物胁迫应答基因的诱导表达中起负调控作用.Tiwari等(2012)从拟南芥ERF和AP2亚族中发现了一种转录激活关键元件EDLL, EDLL可与EAR基序(ERF-associated amphiphilic repression)发生互作, 拮抗后者介导的转录抑制作用.而不同的AP2/ ERF转录因子也可协同招募TPL (topless)和TPR (topless-related)等转录共抑制子(Causier et al., 2012)或组蛋白修饰因子, 进而抑制下游靶基因的表达(Song et al., 2005; Song and Galbraith, 2006).Song等(2005)发现AtERF7特异性地结合GCC-box, 与抑制因子ATSIN3及组蛋白去乙酰化酶19 (histone deacetylase 19, HDA19)发生互作并形成转录因子复合物, 参与干旱胁迫和ABA响应调控.AtERF7过表达株系保卫细胞对ABA的敏感性减弱, 水分散失增加; 反之, AtERF7 RNA干扰系对ABA的敏感性增强.类似的, ERF3能与组蛋白去乙酰化酶复合物亚基SAP18 (SIN3 associated polypeptide P18)互作, 共同招募HDA19形成复合体, 进而抑制相关基因的表达(Song and Galbraith, 2006).BRD结构域被认为是RAV1和RAV2转录因子行使抑制功能的关键元件, 其保守序列中第1位氨基酸残基多为疏水性亮氨酸、缬氨酸和蛋氨酸残基, 突变导致功能丧失(Ikeda and Ohme-Takagi, 2009).研究显示, 与AP2/ERFs协同调控的基因同样可被AP2/ERFs靶基因富集, 分析此类基因的同源基序有助于识别转录因子的假定靶基因并预测其生物学功能(Franco-Zorrillaetal et al., 2014). ...
... ).BRD结构域被认为是RAV1和RAV2转录因子行使抑制功能的关键元件, 其保守序列中第1位氨基酸残基多为疏水性亮氨酸、缬氨酸和蛋氨酸残基, 突变导致功能丧失(Ikeda and Ohme-Takagi, 2009).研究显示, 与AP2/ERFs协同调控的基因同样可被AP2/ERFs靶基因富集, 分析此类基因的同源基序有助于识别转录因子的假定靶基因并预测其生物学功能(Franco-Zorrillaetal et al., 2014). ...
CYTOKININ RESPONSE FACTOR 2 (CRF2) and CRF3 regulate lateral root development in response to cold stress in Arabidopsis 3 2016
... 近年来, 有关ERF亚族参与非生物胁迫响应的研究越来越多.在拟南芥中发现多种非生物胁迫可诱导细胞分裂素应答因子(cytokinin response factor, CRF)的表达, CRF是ERF-VI亚族成员之一, 具有正向调节植物渗透胁迫耐受性和抗冻性作用(Rashotte et al., 2006).CRF2和CRF6可与生长素外输载体PIN (PIN- FORMED)基因启动子区的特定元件结合, 正向调控PIN基因的表达, 这在CRF超表达和缺失突变体研究中得到了验证(Kim, 2016).CRF2和CRF3均参与调控冷胁迫下拟南芥侧根(lateral root, LR)的发育, crf2、crf3、crf2/crf3突变体在冷胁迫下侧根密度比野生型植株更低, CRF2或CRF3过表达均可促进植株侧根密度的增加(Jeon et al., 2016).冷胁迫可诱导CRF2的表达, 因此CRF2可能通过诱导PIN1和PIN7的表达以弥补冷胁迫对侧根发育的影响; 而在缺失突变体crf2中, CRF3超表达可减弱冷胁迫对侧根生长的抑制效应, 推测CRF3可能通过其它途径参与侧根的发育调控(Jeon et al., 2016; Kim, 2016).进一步分析显示, CRF2和CRF3分别通过依赖和不依赖于TCS (two-component signaling)的途径响应冷胁迫(Joen et al., 2016).CRF4是少数不受CTK调控的CRF转录因子之一, 可促进CORs的表达并正向调控植物的冷/冻耐受性(Zwack et al., 2016a).多种非生物胁迫会诱导植物积累活性氧(ROS), 导致细胞损伤, 因此植物维持最低水平的活性氧对其生存至关重要.多种胁迫刺激均可诱导CRF6的表达, CRF6可能通过抑制细胞分裂素相关基因的表达介导细胞分裂素与氧化应激反应之间的联系(Zwack et al., 2016b).CRF6在H2O2诱导的氧化胁迫应答中直接或间接抑制ARR6 (type-A response regulator 6)、ARR9、ARR11、LOG7及ABCG14基因的表达.酵母单杂交实验显示, CRF6直接结合到ARR6启动子区, 激活ARR6的表达, 而对ARR9、LOG7的表达调控可能需要其它植物特异因子的参与(Zwack et al., 2016b). ...
... 可能通过其它途径参与侧根的发育调控(Jeon et al., 2016; Kim, 2016).进一步分析显示, CRF2和CRF3分别通过依赖和不依赖于TCS (two-component signaling)的途径响应冷胁迫(Joen et al., 2016).CRF4是少数不受CTK调控的CRF转录因子之一, 可促进CORs的表达并正向调控植物的冷/冻耐受性(Zwack et al., 2016a).多种非生物胁迫会诱导植物积累活性氧(ROS), 导致细胞损伤, 因此植物维持最低水平的活性氧对其生存至关重要.多种胁迫刺激均可诱导CRF6的表达, CRF6可能通过抑制细胞分裂素相关基因的表达介导细胞分裂素与氧化应激反应之间的联系(Zwack et al., 2016b).CRF6在H2O2诱导的氧化胁迫应答中直接或间接抑制ARR6 (type-A response regulator 6)、ARR9、ARR11、LOG7及ABCG14基因的表达.酵母单杂交实验显示, CRF6直接结合到ARR6启动子区, 激活ARR6的表达, 而对ARR9、LOG7的表达调控可能需要其它植物特异因子的参与(Zwack et al., 2016b). ...
... 分别通过依赖和不依赖于TCS (two-component signaling)的途径响应冷胁迫(Joen et al., 2016).CRF4是少数不受CTK调控的CRF转录因子之一, 可促进CORs的表达并正向调控植物的冷/冻耐受性(Zwack et al., 2016a).多种非生物胁迫会诱导植物积累活性氧(ROS), 导致细胞损伤, 因此植物维持最低水平的活性氧对其生存至关重要.多种胁迫刺激均可诱导CRF6的表达, CRF6可能通过抑制细胞分裂素相关基因的表达介导细胞分裂素与氧化应激反应之间的联系(Zwack et al., 2016b).CRF6在H2O2诱导的氧化胁迫应答中直接或间接抑制ARR6 (type-A response regulator 6)、ARR9、ARR11、LOG7及ABCG14基因的表达.酵母单杂交实验显示, CRF6直接结合到ARR6启动子区, 激活ARR6的表达, 而对ARR9、LOG7的表达调控可能需要其它植物特异因子的参与(Zwack et al., 2016b). ...
The cbfs triple mutants reveal the essential functions of CBFs in cold acclimation and allow the definition of CBF regulons in Arabidopsis 2 2016
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
... 基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
EAR motif-mediated transcriptional repression in plants: an underlying mechanism for epigenetic regulation of gene expression 1 2011
... 许多研究表明, AP2/ERF转录因子可直接与靶基因启动子结合, 激活或抑制下游靶基因的表达, 其N端DNA结合域和C端转录激活域在胁迫相关靶基因的转录激活中起重要作用(Nakano et al., 2006).进一步分析表明, AP2/ERF转录因子中含有EAR基序(LxLxL或DLNxxP)、TLLLFR基序和BRD结构域(B3 repression domain, 保守序列为R/KLFGV)等多个转录抑制结构域(Ikeda and Ohme-Takagi, 2009; Kagale and Rozwadowski, 2011; Deepika et al., 2016).含EAR基序的转录因子是一种重要的抑制子, 在植物非生物胁迫应答基因的诱导表达中起负调控作用.Tiwari等(2012)从拟南芥ERF和AP2亚族中发现了一种转录激活关键元件EDLL, EDLL可与EAR基序(ERF-associated amphiphilic repression)发生互作, 拮抗后者介导的转录抑制作用.而不同的AP2/ ERF转录因子也可协同招募TPL (topless)和TPR (topless-related)等转录共抑制子(Causier et al., 2012)或组蛋白修饰因子, 进而抑制下游靶基因的表达(Song et al., 2005; Song and Galbraith, 2006).Song等(2005)发现AtERF7特异性地结合GCC-box, 与抑制因子ATSIN3及组蛋白去乙酰化酶19 (histone deacetylase 19, HDA19)发生互作并形成转录因子复合物, 参与干旱胁迫和ABA响应调控.AtERF7过表达株系保卫细胞对ABA的敏感性减弱, 水分散失增加; 反之, AtERF7 RNA干扰系对ABA的敏感性增强.类似的, ERF3能与组蛋白去乙酰化酶复合物亚基SAP18 (SIN3 associated polypeptide P18)互作, 共同招募HDA19形成复合体, 进而抑制相关基因的表达(Song and Galbraith, 2006).BRD结构域被认为是RAV1和RAV2转录因子行使抑制功能的关键元件, 其保守序列中第1位氨基酸残基多为疏水性亮氨酸、缬氨酸和蛋氨酸残基, 突变导致功能丧失(Ikeda and Ohme-Takagi, 2009).研究显示, 与AP2/ERFs协同调控的基因同样可被AP2/ERFs靶基因富集, 分析此类基因的同源基序有助于识别转录因子的假定靶基因并预测其生物学功能(Franco-Zorrillaetal et al., 2014). ...
Improvement of water use efficiency in rice by expression of HARDY, an Arabidopsis drought and salt tolerance gene 2 2007
... DREB-A2亚组转录因子DREB2主要参与调节植物的抗旱和耐热性(Mizoi et al., 2012).干旱和高温可诱导DREB2基因的表达, 进而正向调控下游干旱或热胁迫响应基因(Maruyama et al., 2009).研究表明DREB-A4家族成员HRD (hardy)和DREB-A6家族成员(ERF53、RAP2.4和TG/RAP2.4a)也在干旱和耐盐性调控中发挥关键作用(Karaba et al., 2007; Lin et al., 2008; Chen et al., 2012; Zhu et al., 2014).过表达HRD能显著增强拟南芥和水稻植株的耐旱和耐盐性(Karaba et al., 2007).同样, 过表达TG (translucent green)导致植株叶片玻璃化, 叶片细胞含水量增加(Zhu et al., 2014).TG也参与调控抗坏血酸过氧化物酶基因APX (ascorbate peroxidase)的表达, 抵御活性氧(reactive oxygen species, ROS)引起的光氧化胁迫(Rudnik et al., 2017).除拟南芥外, 人们对水稻、玉米、番茄(Lycopersicon esculentum)、葡萄(Vitis vinifera)等作物的DREB1和DREB2转录因子也进行了广泛的研究(Xiao et al., 2006; Matsukura et al., 2010; Dou et al., 2014). ...
... 能显著增强拟南芥和水稻植株的耐旱和耐盐性(Karaba et al., 2007).同样, 过表达TG (translucent green)导致植株叶片玻璃化, 叶片细胞含水量增加(Zhu et al., 2014).TG也参与调控抗坏血酸过氧化物酶基因APX (ascorbate peroxidase)的表达, 抵御活性氧(reactive oxygen species, ROS)引起的光氧化胁迫(Rudnik et al., 2017).除拟南芥外, 人们对水稻、玉米、番茄(Lycopersicon esculentum)、葡萄(Vitis vinifera)等作物的DREB1和DREB2转录因子也进行了广泛的研究(Xiao et al., 2006; Matsukura et al., 2010; Dou et al., 2014). ...
Genome-wide investigation and expression analysis of AP2-ERF gene family in salt tolerant common bean 1 2015
... 正常环境下, AP2/ERF转录因子表达水平较低, 激素或逆境胁迫相关基因与AP2/ERF转录因子保守的顺式作用元件结合, 进而调控其表达(Owji et al., 2017).研究表明, 拟南芥DREB1A/CBF3、DREB1C/CBF2、DREB2C、DREB2G和DEAR3的启动子中包含较多的热激元件(heat shock element, HSE)、低温响应(low temperature responsive, LTR)、脱落酸响应(ABA responsive element, ABRE)等顺式作用元件(Sazegari et al., 2015).热激因子1 (heat shock factor 1, HSF1)、脱落酸响应结合蛋白(ABA responsive element binding protein/ABRE binding factors, AREB/ABF)可分别与DREB2A启动子中的热激应答和ABA响应元件结合, 调控其表达, 进而改变植物胁迫的耐受性(Kim et al., 2011; Liu et al., 2011).蛋白磷酸酶2C (type 2C protein phosphatase, PP2C)包含2个以上的ABRE元件, AREB1、AREB2和ABF3协同正调控AHG1、AHG3、HAI1及HAI2等PP2Cs基因表达(Yoshida et al., 2010).此外, 越来越多的研究证明ET信号通路参与拟南芥盐胁迫反应.进一步分析表明, ESE1 (ethylene and salt-inducible ERF1)在EIN3/EIL1下游调节ET响应基因.EIN3 (ethylene insensitive 3)可与ESE1启动子结合形成EIN3-ESE1转录复合体, 继而调控RD29A和COR15A等基因的表达(Zhang et al., 2011).磷酸化、泛素化、甲基化和乙酰化等组蛋白修饰方式通过调控染色质开放或封闭的空间状态, 激活或抑制AP2/ERF转录(Pfluger and Wagner, 2007).组蛋白去乙酰化酶(histone deacetylase, HDAC)和聚乙二醇(PEG)促进DREB1基因启动子区乙酰化, 增强DREB1基因的表达(Zhang et al., 2018).非生物胁迫下, 组蛋白变体和组蛋白修饰状态也可发生改变, 并可遗传到下一代(Asensii-Fabado et al., 2017).miRNA介导的RNA沉默和翻译抑制在AP2/ERF表达调控中也发挥重要作用.研究发现miRNA172靶向拟南芥AP2mRNA并抑制其翻译, 而miRNA156和miRNA838两者也具有类似调节AP2/ERF的功能(Kavas et al., 2015).此外, 在拟南芥、水稻及玉米中还发现通过可变剪切产生的OsDREB2A/2B、ZmDREB2A、WDREB2以及HvDRF1等多种AP2/-ERF功能异构体(Matsukura et al., 2010). ...
Diverse roles of jasmonates and ethylene in abiotic stress tolerance 4 2015
... AP2/ERF转录因子在参与调控植物应对冷、干旱、高温、高盐及缺氧等多种非生物胁迫过程中具有重要功能(Licausi et al., 2013).其中, DREB和ERF亚家族成员在植物非生物逆境应答中起重要调控作用.研究表明其参与调控的植物环境胁迫应答信号网络比较复杂.胁迫条件下, 部分AP2/ERFs可被快速持续地诱导表达, 而其它进程的响应较为缓慢, 说明可能存在依赖或不依赖于激素信号途径2种胁迫响应模式, 且两者之间存在一定的交叉互作(Van den Broeck et al., 2017).前人的研究也证明, AP2/ERFs广泛参与调控ABA、ET、GA、细胞分裂素(cytokinin, CTK)和油菜素内酯(brassinolide, BR)等植物激素介导的胁迫响应(Colebrook et al., 2014; Kazan et al., 2015; Tao et al., 2015; Sah et al., 2016; Nolan et al., 2017) (图1). ...
... ERF亚家族转录因子是ET介导的胁迫应答信号通路下游的关键调控因子(Licausi et al., 2013; Gibbs et al., 2015; Kazan, 2015; Müller and Munné-Bosch, 2015).冷胁迫下, 植物体内的ET主要起负调控作用, 过量ET或1-氨基环丙烷-1-羧酸降低植物的耐冷性, 而ET生物合成抑制剂氨基乙氧基甘氨酸或感知拮抗剂Ag+具有相反的作用(Shi et al., 2012).etr1-1、ein4-1、ein2-5、ein3-1和ein3eil突变体表现出更强的耐冷性, 但组成型ET响应etr1-1和ein3过表达植株表现出耐冷性减弱; 冷胁迫诱导ETR1、EIN4、EBF1及EBF2快速上调表达, 而EIN2和EIL1下调表达.进一步分析表明, EIN3通过与启动子区特定元件结合, 抑制ARR5、ARR7、ARR15和CBF转录, ARRs作为冷信号通路的关键节点基因, 在ET和CTK介导的胁迫响应网络中起着关键作用, 过表达ARRs使植株的耐冷能力增强(Shi et al., 2012).此外, EIN3可激活ERF1和ESEs, 进而参与调控下游胁迫相关基因, 提高植物的耐盐性(Kazan, 2015; Tao et al., 2015).35S:AtERF4-GFP转基因植株中ABI2、RD29B和RAP18的表达受抑制, 进一步说明AtERF4是ET和ABA信号途径的负调控因子(Yang et al., 2011). ...
... ).此外, EIN3可激活ERF1和ESEs, 进而参与调控下游胁迫相关基因, 提高植物的耐盐性(Kazan, 2015; Tao et al., 2015).35S:AtERF4-GFP转基因植株中ABI2、RD29B和RAP18的表达受抑制, 进一步说明AtERF4是ET和ABA信号途径的负调控因子(Yang et al., 2011). ...
CYTOKININ RESPONSE FACTORs gating environmental signals and hormones 2 2016
... 近年来, 有关ERF亚族参与非生物胁迫响应的研究越来越多.在拟南芥中发现多种非生物胁迫可诱导细胞分裂素应答因子(cytokinin response factor, CRF)的表达, CRF是ERF-VI亚族成员之一, 具有正向调节植物渗透胁迫耐受性和抗冻性作用(Rashotte et al., 2006).CRF2和CRF6可与生长素外输载体PIN (PIN- FORMED)基因启动子区的特定元件结合, 正向调控PIN基因的表达, 这在CRF超表达和缺失突变体研究中得到了验证(Kim, 2016).CRF2和CRF3均参与调控冷胁迫下拟南芥侧根(lateral root, LR)的发育, crf2、crf3、crf2/crf3突变体在冷胁迫下侧根密度比野生型植株更低, CRF2或CRF3过表达均可促进植株侧根密度的增加(Jeon et al., 2016).冷胁迫可诱导CRF2的表达, 因此CRF2可能通过诱导PIN1和PIN7的表达以弥补冷胁迫对侧根发育的影响; 而在缺失突变体crf2中, CRF3超表达可减弱冷胁迫对侧根生长的抑制效应, 推测CRF3可能通过其它途径参与侧根的发育调控(Jeon et al., 2016; Kim, 2016).进一步分析显示, CRF2和CRF3分别通过依赖和不依赖于TCS (two-component signaling)的途径响应冷胁迫(Joen et al., 2016).CRF4是少数不受CTK调控的CRF转录因子之一, 可促进CORs的表达并正向调控植物的冷/冻耐受性(Zwack et al., 2016a).多种非生物胁迫会诱导植物积累活性氧(ROS), 导致细胞损伤, 因此植物维持最低水平的活性氧对其生存至关重要.多种胁迫刺激均可诱导CRF6的表达, CRF6可能通过抑制细胞分裂素相关基因的表达介导细胞分裂素与氧化应激反应之间的联系(Zwack et al., 2016b).CRF6在H2O2诱导的氧化胁迫应答中直接或间接抑制ARR6 (type-A response regulator 6)、ARR9、ARR11、LOG7及ABCG14基因的表达.酵母单杂交实验显示, CRF6直接结合到ARR6启动子区, 激活ARR6的表达, 而对ARR9、LOG7的表达调控可能需要其它植物特异因子的参与(Zwack et al., 2016b). ...
... ; Kim, 2016).进一步分析显示, CRF2和CRF3分别通过依赖和不依赖于TCS (two-component signaling)的途径响应冷胁迫(Joen et al., 2016).CRF4是少数不受CTK调控的CRF转录因子之一, 可促进CORs的表达并正向调控植物的冷/冻耐受性(Zwack et al., 2016a).多种非生物胁迫会诱导植物积累活性氧(ROS), 导致细胞损伤, 因此植物维持最低水平的活性氧对其生存至关重要.多种胁迫刺激均可诱导CRF6的表达, CRF6可能通过抑制细胞分裂素相关基因的表达介导细胞分裂素与氧化应激反应之间的联系(Zwack et al., 2016b).CRF6在H2O2诱导的氧化胁迫应答中直接或间接抑制ARR6 (type-A response regulator 6)、ARR9、ARR11、LOG7及ABCG14基因的表达.酵母单杂交实验显示, CRF6直接结合到ARR6启动子区, 激活ARR6的表达, 而对ARR9、LOG7的表达调控可能需要其它植物特异因子的参与(Zwack et al., 2016b). ...
An ABRE promoter sequence is involved in osmotic stress-responsive expression of the DREB2A gene, which encodes a transcription factor regulating drought-inducible genes in Arabidopsis 1 2011
... 正常环境下, AP2/ERF转录因子表达水平较低, 激素或逆境胁迫相关基因与AP2/ERF转录因子保守的顺式作用元件结合, 进而调控其表达(Owji et al., 2017).研究表明, 拟南芥DREB1A/CBF3、DREB1C/CBF2、DREB2C、DREB2G和DEAR3的启动子中包含较多的热激元件(heat shock element, HSE)、低温响应(low temperature responsive, LTR)、脱落酸响应(ABA responsive element, ABRE)等顺式作用元件(Sazegari et al., 2015).热激因子1 (heat shock factor 1, HSF1)、脱落酸响应结合蛋白(ABA responsive element binding protein/ABRE binding factors, AREB/ABF)可分别与DREB2A启动子中的热激应答和ABA响应元件结合, 调控其表达, 进而改变植物胁迫的耐受性(Kim et al., 2011; Liu et al., 2011).蛋白磷酸酶2C (type 2C protein phosphatase, PP2C)包含2个以上的ABRE元件, AREB1、AREB2和ABF3协同正调控AHG1、AHG3、HAI1及HAI2等PP2Cs基因表达(Yoshida et al., 2010).此外, 越来越多的研究证明ET信号通路参与拟南芥盐胁迫反应.进一步分析表明, ESE1 (ethylene and salt-inducible ERF1)在EIN3/EIL1下游调节ET响应基因.EIN3 (ethylene insensitive 3)可与ESE1启动子结合形成EIN3-ESE1转录复合体, 继而调控RD29A和COR15A等基因的表达(Zhang et al., 2011).磷酸化、泛素化、甲基化和乙酰化等组蛋白修饰方式通过调控染色质开放或封闭的空间状态, 激活或抑制AP2/ERF转录(Pfluger and Wagner, 2007).组蛋白去乙酰化酶(histone deacetylase, HDAC)和聚乙二醇(PEG)促进DREB1基因启动子区乙酰化, 增强DREB1基因的表达(Zhang et al., 2018).非生物胁迫下, 组蛋白变体和组蛋白修饰状态也可发生改变, 并可遗传到下一代(Asensii-Fabado et al., 2017).miRNA介导的RNA沉默和翻译抑制在AP2/ERF表达调控中也发挥重要作用.研究发现miRNA172靶向拟南芥AP2mRNA并抑制其翻译, 而miRNA156和miRNA838两者也具有类似调节AP2/ERF的功能(Kavas et al., 2015).此外, 在拟南芥、水稻及玉米中还发现通过可变剪切产生的OsDREB2A/2B、ZmDREB2A、WDREB2以及HvDRF1等多种AP2/-ERF功能异构体(Matsukura et al., 2010). ...
AP2/ERF family transcription factors ORA59 and RAP2.3 interact in the nucleus and function together in ethylene responses 1 2018
... 此外, ET促进RAP2.3转录因子定位于细胞核内, 通过依赖于RAP2.3的方式激发ORA59介导的ET响应过程(Kim et al., 2018).而ERF-VIIs通过ET不依赖途径参与调控缺氧响应.在ET不敏感突变体或生物合成抑制剂存在的情况下, 缺氧依然可以诱导ERF73/HRE1的表达, ERF-VIIs也可能通过反馈调节负调控ET信号(Hinz et al., 2010; Yang et al., 2011).研究发现, ET合酶ACS在RAP2.2过表达植株中表达量降低, 而在rap2.2-2敲除突变体中表达上调(Hinz et al., 2010).然而, 胁迫条件下ERF-VIIs如何通过负反馈机制调控ET生物合成有待进一步研究. ...
... 研究表明, 非生物胁迫下ABA可促进ANT (Meng et al., 2015b)、ERF53 (Hsieh et al., 2013)、RAP2.6L (Liu et al., 2012)和RAP2.6 (Zhu et al., 2010)转录, 上述转录因子进而结合胁迫应答顺式作用元件DRE/ABRE, 调控下游相关靶基因的表达, 增强植物对非生物胁迫的耐受能力.在水稻中ERF71正向调控ABA信号转导, 过表达ERF71可促进根中ABI5、PP2C68、RAB16C及RAB16D等基因上调表达.此外, ERF71可直接调控木质素生物合成基因CCR1、CCR10及C4H转录, 诱导根系径向生长, 改变根系构型, 提高耐旱能力(Lee et al., 2017; Li et al., 2018).盐胁迫和ABA双重诱导下, ABA增强DREB2的转录活性, 进而促进Rd29A协同表达(Lee et al., 2016). ...
Synergistic activation of RD29A via integration of salinity stress and abscisic acid in Arabidopsis thaliana 1 2016
... 研究表明, 非生物胁迫下ABA可促进ANT (Meng et al., 2015b)、ERF53 (Hsieh et al., 2013)、RAP2.6L (Liu et al., 2012)和RAP2.6 (Zhu et al., 2010)转录, 上述转录因子进而结合胁迫应答顺式作用元件DRE/ABRE, 调控下游相关靶基因的表达, 增强植物对非生物胁迫的耐受能力.在水稻中ERF71正向调控ABA信号转导, 过表达ERF71可促进根中ABI5、PP2C68、RAB16C及RAB16D等基因上调表达.此外, ERF71可直接调控木质素生物合成基因CCR1、CCR10及C4H转录, 诱导根系径向生长, 改变根系构型, 提高耐旱能力(Lee et al., 2017; Li et al., 2018).盐胁迫和ABA双重诱导下, ABA增强DREB2的转录活性, 进而促进Rd29A协同表达(Lee et al., 2016). ...
Arabidopsis AtERF71/HRE2 functions as transcriptional activator via cis-acting GCC box or DRE/CRT element and is involved in root development through regulation of root cell expansion 1 2015
... AP2/ERF转录因子通过特异性结合胁迫应答基因启动子区顺式元件, 参与调控植物的逆境防御反应.DREB亚族成员特异性识别并结合启动子区域的DRE/CRT (dehydration responsive element/C-re-peat)元件(核心序列A/GCCGAC), 诱导干旱、低温和盐胁迫相关应答基因的表达.ERF亚族成员则可通过与乙烯响应元件(ethylene response element, ERE) (核心序列AGCCGCC, 又称GCC-box)结合, 从而参与调控乙烯应答和非生物胁迫(Franco-Zorrilla et al., 2014).此外, 研究发现许多AP2/ERF转录因子可同时与DRE/CRT和ERE两种元件结合, 如DREB亚族成员TINY、CBF1、ERF53、RAP2.4、TG/RAP2.4a以及ERF亚族成员ERF1、ERF4和ERF71 (Chen et al., 2012; Zhu et al., 2014; Lee et al., 2015), 但结合能力有差异.在拟南芥中, DREB2C和DEAR3与类GCC-box元件(GCCGCC)的结合能力远超DRE元件(Franco-Zorrilla et al., 2014).AP2/ERF转录因子还能与DRE和ERE之外的顺式元件结合, 如偶联元件1 (CE1, TGCCACCG)、类偶联元件3 (CE-3like, CGCG)和缺氧响应启动子元件(hypoxia-responsive promoter element, HRPE), 以及ATCTA、CAACA、CATGCA、CAA/CA/CTG、ATCGAG和(NC/GT) CGNCCA (Bossi et al., 2009; Zhu et al., 2010; Chen et al., 2016; Gasch et al., 2016; Park et al., 2016).近年来, 科学家通过蛋白质结合微阵列(protein binding microarray, PBM)技术发现结构特征高度相似的转录因子具有相似的DNA结合位点, 同时也具有相似的生物学功能.该研究结果合理地解释了转录因子功能冗余现象, 也为预测和挖掘未知AP2/ERF转录因子提供了高效可行的技术方法(Franco-Zorrilla et al., 2014). ...
BZR1 positively regulates freezing tolerance via CBF-dependent and CBF-independent pathways in Arabidopsis 1 2017
... BZR1可结合CBF1和CBF2的启动子区并促进两者在冷胁迫下的表达.bin2-3、bil1、bil2缺失突变体在持续的冷胁迫下可以诱导去磷酸化BZR1的积累, BZR1通过调控WKRY6、PYL6、SOC1、JMT和SAG21等不依赖CBF途径的COR基因而增强拟南芥的耐冷性(Li et al., 2017).研究发现BR可与干旱诱导相关转录因子RD26互作, 拮抗干旱响应基因的表达, 进而负调控植物的抗旱性(Ye et al., 2017).BR与ABA信号途径中的受体、转录因子之间也产生拮抗效应, 进而调节胁迫下植物体的生长平衡(Nolan et al., 2017).目前已从BES1和BZR1靶基因中鉴定出一些AP2/ERF类转录因子, 暗示AP2/ERF具有整合BR信号通路与非生物胁迫响应的潜在功能(Sun et al., 2010; Yu et al., 2011).ERF72可能是研究BR信号通路与胁迫响应交叉互作的候选基因.在拟南芥中, ERF72/RAP2.3拮抗BZR1和ARF6 (auxin responsive factor 6), 抑制下胚轴伸长, 此过程中BR信号途径如何与胁迫响应调控网络协同互作发挥作用尚不清楚(Liu et al., 2018b).研究表明, SUB1A参与调节GA和BR的交叉网络互作(Schmitz et al., 2013).GA和BR途径通过DELLA蛋白、BZR1等转录因子协同调控植物的生长(Oh et al., 2012).在植物淹水过程中SUB1A差异调节BR合成相关基因的表达, 激活BR生物合成和信号转导, 诱导GA降解关键基因GA2ox7的表达, 进而控制水稻植株体内的GA水平, 促进DELLA蛋白的积累(Schmitz et al., 2013).因此, 推测AP2/ERF可能通过不同的机制参与调控BR信号通路. ...
OsERF71 confers drought tolerance via modulating ABA signaling and proline biosynthesis 2 2018
... 植物在生长发育过程中经常受到生物或非生物胁迫的影响, 由此, 植物逐渐形成了复杂的调节机制感知胁迫信号, 对外部环境变化做出快速响应(郭倩倩和周文彬, 2019).脱落酸(abscisic acid, ABA)、乙烯(ethylene, ET)及赤霉素(gibberellin, GA)等激素作为信号分子能够调节植物的生长发育、形态建成以及抗逆反应等生物学过程.该过程也受转录因子(transcription factor, TF)调控, 即转录因子通过依赖或不依赖于激素介导的信号通路参与调控植物对干旱、低温、缺氧、高盐和淹水等非生物胁迫的响应, 进而影响植物的逆境耐受能力(Gibbs et al., 2015; Verma et al., 2016; Li et al., 2018).AP2/ERF (APETALA2/eth-ylene responsive factor)是植物特有的转录因子家族之一, 已在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、高粱(Sorghum bicolor)、大白菜(Brassica pekinensis)、毛竹(Phyllostachys edulis)、玉米(Zea mays)、大麦(Hordeum vulgare)和小麦(Triticum aestivum)等物种中被成功分离鉴定(Xu and Chua, 2011; Chandler, 2018).通过突变体实验, 科学家发现了许多与AP2/ERF相关的非生物胁迫或激素信号应答基因.目前, AP2/ERF已成为研究非生物胁迫和激素互作的热点候选基因.本文从AP2/ERF转录因子的结构特征、转录调控、翻译后修饰、结合位点、协同互作蛋白以及其参与调控依赖或不依赖激素信号途径的非生物胁迫响应进行综述, 为进一步阐明AP2/ERF转录因子在非生物胁迫调控网络中的作用提供理论依据. ...
... 研究表明, 非生物胁迫下ABA可促进ANT (Meng et al., 2015b)、ERF53 (Hsieh et al., 2013)、RAP2.6L (Liu et al., 2012)和RAP2.6 (Zhu et al., 2010)转录, 上述转录因子进而结合胁迫应答顺式作用元件DRE/ABRE, 调控下游相关靶基因的表达, 增强植物对非生物胁迫的耐受能力.在水稻中ERF71正向调控ABA信号转导, 过表达ERF71可促进根中ABI5、PP2C68、RAB16C及RAB16D等基因上调表达.此外, ERF71可直接调控木质素生物合成基因CCR1、CCR10及C4H转录, 诱导根系径向生长, 改变根系构型, 提高耐旱能力(Lee et al., 2017; Li et al., 2018).盐胁迫和ABA双重诱导下, ABA增强DREB2的转录活性, 进而促进Rd29A协同表达(Lee et al., 2016). ...
Arabidopsis YL1/BPG2 is involved in seedling shoot response to salt stress through ABI4 1 2016
... DREB-A3亚组成员ABI4 (ABA insensitive 4)是ABA信号通路的重要组分.植株受到胁迫后, 体内积累ABA和ROS, ABI4可被ABA、ROS及糖质体等多种信号通路靶向调控.CCAAT结合因子A (CCAAT binding factor A, CBFA)是四吡咯化合物HAP (heme activator protein)三聚体转录复合物的亚基, 当产生过量ROS时, ABI4可与其它转录因子竞争性地结合CCACGT元件, 抑制CBFA的表达, 进而调控质体反向信号通路中相关基因的表达(Zhang et al., 2013).拟南芥突变体实验表明, ABI4在YL1 (yellow leaf 1) (Li et al., 2016)和DPG1 (delayed pale-greening 1) (Yi et al., 2019)等基因参与盐胁迫响应的调控网络中也发挥重要作用.逆境胁迫下, ORA47在JA (jasmo-nate acid)和ABA信号转导途径中靶向调节ABA和JA诱导的下游应答基因, ERF18/ORA-47识别启动子顺式元件(NC/GT)CGNCCA (O-box), 激活PP2C家族ABI2转录因子的表达.ABI1则在ORA-47上游起作用, 从而形成ABI1-ORA47-ABI2正反馈基因表达回路, 调控ABA信号转导和干旱胁迫响应(Chen et al., 2016).此外, RAV1转录因子在ABA信号转导途径中也发挥关键作用, RAV1能与ABI3、ABI4和ABI5启动子结合, 其过表达抑制ABI3、ABI4和ABI5的转录活性, 导致根系生长发育过程中对ABA不敏感.同时, 蔗糖非酵解型蛋白激酶(SnRK2.2、SnRK2.3和SnRK2.6)在核内磷酸化RAV1, 降低ABI5对RAV1的依赖性, 并负调控RAV1对靶基因的转录抑制作用(Feng et al., 2014). ...
APETALA2/ Ethylene Responsive Factor (AP2/ERF) transcription factors: mediators of stress responses and developmental programs 4 2013
... AP2/ERF转录因子具有独特的结构特征, 其由DNA结合域(DNA-binding domain)、转录调控域(transcription regulation domain)、寡聚化位点(oligomerization site)和核定位信号(nuclear localization signal, NLS) 4个主要功能区组成, 并以高度保守的AP2/ ERF结合域而得名(Sakuma et al., 2002).通常情况下, AP2/ERF转录因子至少含有1个AP2结构域的DNA结合区, 该结构域包含60-70个氨基酸残基, 按照3个β折叠和1个α螺旋方式形成典型的三维结构(Nakano et al., 2006; Abiri et al., 2017).根据包含的AP2/ERF结构域数量和结合序列特点, 可将AP2/ ERF家族分为AP2 (APETALA2)、RAV (related to ABI3/VP1)、脱水反应元件结合蛋白(dehydration- responsive element binding protein, DREB) (A1-A6亚组)、乙烯应答因子(ethylene-responsive factor, ERF) (V-X亚组)和Soloist五个亚族(Nakano et al., 2006; Licausi et al., 2013).其中, 对AP2、DREB和ERF的研究较为广泛和深入, 而有关Soloist亚族的研究报道极少, 已知其核苷酸序列在多数植物中高度保守(Sun et al., 2016). ...
... AP2/ERF转录因子在参与调控植物应对冷、干旱、高温、高盐及缺氧等多种非生物胁迫过程中具有重要功能(Licausi et al., 2013).其中, DREB和ERF亚家族成员在植物非生物逆境应答中起重要调控作用.研究表明其参与调控的植物环境胁迫应答信号网络比较复杂.胁迫条件下, 部分AP2/ERFs可被快速持续地诱导表达, 而其它进程的响应较为缓慢, 说明可能存在依赖或不依赖于激素信号途径2种胁迫响应模式, 且两者之间存在一定的交叉互作(Van den Broeck et al., 2017).前人的研究也证明, AP2/ERFs广泛参与调控ABA、ET、GA、细胞分裂素(cytokinin, CTK)和油菜素内酯(brassinolide, BR)等植物激素介导的胁迫响应(Colebrook et al., 2014; Kazan et al., 2015; Tao et al., 2015; Sah et al., 2016; Nolan et al., 2017) (图1). ...
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
... ERF亚家族转录因子是ET介导的胁迫应答信号通路下游的关键调控因子(Licausi et al., 2013; Gibbs et al., 2015; Kazan, 2015; Müller and Munné-Bosch, 2015).冷胁迫下, 植物体内的ET主要起负调控作用, 过量ET或1-氨基环丙烷-1-羧酸降低植物的耐冷性, 而ET生物合成抑制剂氨基乙氧基甘氨酸或感知拮抗剂Ag+具有相反的作用(Shi et al., 2012).etr1-1、ein4-1、ein2-5、ein3-1和ein3eil突变体表现出更强的耐冷性, 但组成型ET响应etr1-1和ein3过表达植株表现出耐冷性减弱; 冷胁迫诱导ETR1、EIN4、EBF1及EBF2快速上调表达, 而EIN2和EIL1下调表达.进一步分析表明, EIN3通过与启动子区特定元件结合, 抑制ARR5、ARR7、ARR15和CBF转录, ARRs作为冷信号通路的关键节点基因, 在ET和CTK介导的胁迫响应网络中起着关键作用, 过表达ARRs使植株的耐冷能力增强(Shi et al., 2012).此外, EIN3可激活ERF1和ESEs, 进而参与调控下游胁迫相关基因, 提高植物的耐盐性(Kazan, 2015; Tao et al., 2015).35S:AtERF4-GFP转基因植株中ABI2、RD29B和RAP18的表达受抑制, 进一步说明AtERF4是ET和ABA信号途径的负调控因子(Yang et al., 2011). ...
Role of Arabidopsis RAP2.4 in regulating light and ethylene-mediated developmental processes and drought stress tolerance 1 2008
... DREB-A2亚组转录因子DREB2主要参与调节植物的抗旱和耐热性(Mizoi et al., 2012).干旱和高温可诱导DREB2基因的表达, 进而正向调控下游干旱或热胁迫响应基因(Maruyama et al., 2009).研究表明DREB-A4家族成员HRD (hardy)和DREB-A6家族成员(ERF53、RAP2.4和TG/RAP2.4a)也在干旱和耐盐性调控中发挥关键作用(Karaba et al., 2007; Lin et al., 2008; Chen et al., 2012; Zhu et al., 2014).过表达HRD能显著增强拟南芥和水稻植株的耐旱和耐盐性(Karaba et al., 2007).同样, 过表达TG (translucent green)导致植株叶片玻璃化, 叶片细胞含水量增加(Zhu et al., 2014).TG也参与调控抗坏血酸过氧化物酶基因APX (ascorbate peroxidase)的表达, 抵御活性氧(reactive oxygen species, ROS)引起的光氧化胁迫(Rudnik et al., 2017).除拟南芥外, 人们对水稻、玉米、番茄(Lycopersicon esculentum)、葡萄(Vitis vinifera)等作物的DREB1和DREB2转录因子也进行了广泛的研究(Xiao et al., 2006; Matsukura et al., 2010; Dou et al., 2014). ...
The role of class A1 heat shock factors (HSFA1s) in response to heat and other stresses in Arabidopsis 1 2011
... 正常环境下, AP2/ERF转录因子表达水平较低, 激素或逆境胁迫相关基因与AP2/ERF转录因子保守的顺式作用元件结合, 进而调控其表达(Owji et al., 2017).研究表明, 拟南芥DREB1A/CBF3、DREB1C/CBF2、DREB2C、DREB2G和DEAR3的启动子中包含较多的热激元件(heat shock element, HSE)、低温响应(low temperature responsive, LTR)、脱落酸响应(ABA responsive element, ABRE)等顺式作用元件(Sazegari et al., 2015).热激因子1 (heat shock factor 1, HSF1)、脱落酸响应结合蛋白(ABA responsive element binding protein/ABRE binding factors, AREB/ABF)可分别与DREB2A启动子中的热激应答和ABA响应元件结合, 调控其表达, 进而改变植物胁迫的耐受性(Kim et al., 2011; Liu et al., 2011).蛋白磷酸酶2C (type 2C protein phosphatase, PP2C)包含2个以上的ABRE元件, AREB1、AREB2和ABF3协同正调控AHG1、AHG3、HAI1及HAI2等PP2Cs基因表达(Yoshida et al., 2010).此外, 越来越多的研究证明ET信号通路参与拟南芥盐胁迫反应.进一步分析表明, ESE1 (ethylene and salt-inducible ERF1)在EIN3/EIL1下游调节ET响应基因.EIN3 (ethylene insensitive 3)可与ESE1启动子结合形成EIN3-ESE1转录复合体, 继而调控RD29A和COR15A等基因的表达(Zhang et al., 2011).磷酸化、泛素化、甲基化和乙酰化等组蛋白修饰方式通过调控染色质开放或封闭的空间状态, 激活或抑制AP2/ERF转录(Pfluger and Wagner, 2007).组蛋白去乙酰化酶(histone deacetylase, HDAC)和聚乙二醇(PEG)促进DREB1基因启动子区乙酰化, 增强DREB1基因的表达(Zhang et al., 2018).非生物胁迫下, 组蛋白变体和组蛋白修饰状态也可发生改变, 并可遗传到下一代(Asensii-Fabado et al., 2017).miRNA介导的RNA沉默和翻译抑制在AP2/ERF表达调控中也发挥重要作用.研究发现miRNA172靶向拟南芥AP2mRNA并抑制其翻译, 而miRNA156和miRNA838两者也具有类似调节AP2/ERF的功能(Kavas et al., 2015).此外, 在拟南芥、水稻及玉米中还发现通过可变剪切产生的OsDREB2A/2B、ZmDREB2A、WDREB2以及HvDRF1等多种AP2/-ERF功能异构体(Matsukura et al., 2010). ...
Insights into the regulation of C-repeat binding factors in plant cold signaling 1 2018
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
ERF72 interacts with ARF6 and BZR1 to regulate hypocotyl elongation in Arabidopsis 1 2018
... BZR1可结合CBF1和CBF2的启动子区并促进两者在冷胁迫下的表达.bin2-3、bil1、bil2缺失突变体在持续的冷胁迫下可以诱导去磷酸化BZR1的积累, BZR1通过调控WKRY6、PYL6、SOC1、JMT和SAG21等不依赖CBF途径的COR基因而增强拟南芥的耐冷性(Li et al., 2017).研究发现BR可与干旱诱导相关转录因子RD26互作, 拮抗干旱响应基因的表达, 进而负调控植物的抗旱性(Ye et al., 2017).BR与ABA信号途径中的受体、转录因子之间也产生拮抗效应, 进而调节胁迫下植物体的生长平衡(Nolan et al., 2017).目前已从BES1和BZR1靶基因中鉴定出一些AP2/ERF类转录因子, 暗示AP2/ERF具有整合BR信号通路与非生物胁迫响应的潜在功能(Sun et al., 2010; Yu et al., 2011).ERF72可能是研究BR信号通路与胁迫响应交叉互作的候选基因.在拟南芥中, ERF72/RAP2.3拮抗BZR1和ARF6 (auxin responsive factor 6), 抑制下胚轴伸长, 此过程中BR信号途径如何与胁迫响应调控网络协同互作发挥作用尚不清楚(Liu et al., 2018b).研究表明, SUB1A参与调节GA和BR的交叉网络互作(Schmitz et al., 2013).GA和BR途径通过DELLA蛋白、BZR1等转录因子协同调控植物的生长(Oh et al., 2012).在植物淹水过程中SUB1A差异调节BR合成相关基因的表达, 激活BR生物合成和信号转导, 诱导GA降解关键基因GA2ox7的表达, 进而控制水稻植株体内的GA水平, 促进DELLA蛋白的积累(Schmitz et al., 2013).因此, 推测AP2/ERF可能通过不同的机制参与调控BR信号通路. ...
RAP2.6L overexpression delays waterlogging induced premature senescence by increasing stomatal closure more than antioxidant enzyme activity 1 2012
... 研究表明, 非生物胁迫下ABA可促进ANT (Meng et al., 2015b)、ERF53 (Hsieh et al., 2013)、RAP2.6L (Liu et al., 2012)和RAP2.6 (Zhu et al., 2010)转录, 上述转录因子进而结合胁迫应答顺式作用元件DRE/ABRE, 调控下游相关靶基因的表达, 增强植物对非生物胁迫的耐受能力.在水稻中ERF71正向调控ABA信号转导, 过表达ERF71可促进根中ABI5、PP2C68、RAB16C及RAB16D等基因上调表达.此外, ERF71可直接调控木质素生物合成基因CCR1、CCR10及C4H转录, 诱导根系径向生长, 改变根系构型, 提高耐旱能力(Lee et al., 2017; Li et al., 2018).盐胁迫和ABA双重诱导下, ABA增强DREB2的转录活性, 进而促进Rd29A协同表达(Lee et al., 2016). ...
Rice SUB1A constrains remodelling of the transcriptome and metabolome during submergence to facilitate post-submergence recovery 2 2018
... ERF亚族成员在低氧和淹水胁迫响应过程中也发挥重要作用(Bui et al., 2015; Gibbs et al., 2015).SUB1A (submergence 1A)是ERF亚族B2亚组成员之一, Sub1A-1超表达的同时促进乙醇脱氢酶基因(alcohol dehydrogenase 1, Adh1)上调表达, 抑制Sub1C的转录水平, 说明Sub1A-1是耐淹性的主要调节因子, 在淹水胁迫应答调控中起关键作用(Xu et al., 2006).淹水状态下, 低水平海藻糖-6-磷酸合酶(trehalose-6-phosphate, T6P)与CIPK15 (CBL-inter-acting protein kinase 15)协同激活SnRK1, SnRK1介导SUB1A抑制幼苗胚轴生长和呼吸相关基因的表达, 直接或间接调控碳同化过程.而SK1/SK2 (snorkel1/2)激活参与叶柄、茎和导管发育相关基因, 促进植株快速生长(Hattori et al., 2009; Locke et al., 2018).缺氧胁迫下, 拟南芥ERF-VIIs可以通过氧依赖的N端规则通路降解(Gibbs et al., 2015).RING (really interesting gene)类蛋白SINAT2可通过其N端保守的C3H4型RING结构域调控RAP2.12, 沉默SINAT1/2可上调RAP2.12的表达水平(Papdi et al., 2015).此外, ERF71/HRE2、ERF72/RAP2.3、ERF73/HRE1、ERF74/RAP2.12及ERF75/RAP2.2d也受缺氧胁迫诱导, 进而正向调控糖代谢和激素信号相关的缺氧响应基因, 提高植株的缺氧耐受能力.过表达RAP2.2、RAP2.3和RAP2.12可提高植株在低氧和渗透胁迫下的存活率, 而rap2.12-2/rap2.3-1双突变体对淹水和渗透胁迫表现出较高的敏感性(Yao et al., 2017). ...
... 在拟南芥和水稻中, SUB1A、SK1和SK2参与GA信号转导通路的调控.淹水胁迫下, SUB1A激活SLR1 (slender rice 1)和SLRL1 (slender rice 1 like 1) (DELLA类似蛋白)转录, 同时SUB1A抑制受GA诱导的淀粉代谢相关基因的表达, 调控植株节间伸长和呼吸作用(Fukao and Bailey-Serres, 2008; Locke et al., 2018; Perata, 2018).SK1和SK2两个转录因子中均含有EIN3结合位点, 电泳迁移率测定显示, 类EIN3基因EIL1b可与SK1和SK2启动子结合, 激活其转录活性.研究发现, 淹水状态下拟南芥中ET合成增加, 进而诱导SK1和SK2表达, 激发GA20oxs基因上调表达, 促进植株节间伸长, 减轻淹水对其生长的抑制效应(Hattori et al., 2009; Ayano et al., 2014). ...
Dwarf and delayed-flowering 1, a novel Arabidopsis mutant deficient in gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor 1 2004
... AP2/ERFs参与调控植物GA生物合成相关基因的转录.盐胁迫下, DREB1E和DREB1F抑制GA20ox的表达, 使GA生物合成减少, 引起植株生长迟缓(Magome et al., 2004).过表达ERF6抑制GA2ox的表达水平, 诱导赤霉素降解酶(gibberellin 2-oxidase 6, GA2ox6)的合成, 维持DELLA蛋白的稳定性.渗透胁迫下, erf6功能获得突变体植株矮小且对胁迫高度敏感, 而erf5/erf6缺失突变体生长受胁迫影响较小.研究发现ERF6还激活了STZ、MYB51和WRKY33等渗透胁迫响应基因的表达(Dubois et al., 2013).冷胁迫下, CBF1转录因子激发GA2ox基因的表达而降低GA含量, 组成型表达CBF1抑制GA的积累, 植株表现矮化, 表明DELLA蛋白是CBF1介导的冷胁迫响应的关键组分(Achard et al., 2008).相反, ERF11作为一种GA生物合成和信号转导的正调控因子, 抑制ET生物合成, 促进植株节间伸长.过表达ERF11上调GA3ox1和GA20ox基因的表达水平, 增加GA的积累量(Zhou et al., 2016).ERF6作为转录激活因子诱导ERF11的表达, 反之, ERF11抑制GA2-OX6、STZ、MYB51和WRKY33等ERF6相关靶基因的表达.进一步实验表明, 4个靶基因在erf11/erf6过表达植株中也不能被诱导表达, ERF11过表达可消除ERF6过表达导致的极度矮化现象, 说明ERF11可能通过直接竞争目标基因启动子而在分子水平上拮抗ERF6 (Dubois et al., 2015). ...
Cytokinins regulate a bidirectional phosphorelay network in Arabidopsis 1 2006
Metabolic pathways involved in cold acclimation identified by integrated analysis of metabolites and transcripts regulated by DREB1A and DREB2A 1 2009
... DREB-A2亚组转录因子DREB2主要参与调节植物的抗旱和耐热性(Mizoi et al., 2012).干旱和高温可诱导DREB2基因的表达, 进而正向调控下游干旱或热胁迫响应基因(Maruyama et al., 2009).研究表明DREB-A4家族成员HRD (hardy)和DREB-A6家族成员(ERF53、RAP2.4和TG/RAP2.4a)也在干旱和耐盐性调控中发挥关键作用(Karaba et al., 2007; Lin et al., 2008; Chen et al., 2012; Zhu et al., 2014).过表达HRD能显著增强拟南芥和水稻植株的耐旱和耐盐性(Karaba et al., 2007).同样, 过表达TG (translucent green)导致植株叶片玻璃化, 叶片细胞含水量增加(Zhu et al., 2014).TG也参与调控抗坏血酸过氧化物酶基因APX (ascorbate peroxidase)的表达, 抵御活性氧(reactive oxygen species, ROS)引起的光氧化胁迫(Rudnik et al., 2017).除拟南芥外, 人们对水稻、玉米、番茄(Lycopersicon esculentum)、葡萄(Vitis vinifera)等作物的DREB1和DREB2转录因子也进行了广泛的研究(Xiao et al., 2006; Matsukura et al., 2010; Dou et al., 2014). ...
Comprehensive analysis of rice DREB2-type genes that encode transcription factors involved in the expression of abiotic stress-responsive genes 2 2010
... 正常环境下, AP2/ERF转录因子表达水平较低, 激素或逆境胁迫相关基因与AP2/ERF转录因子保守的顺式作用元件结合, 进而调控其表达(Owji et al., 2017).研究表明, 拟南芥DREB1A/CBF3、DREB1C/CBF2、DREB2C、DREB2G和DEAR3的启动子中包含较多的热激元件(heat shock element, HSE)、低温响应(low temperature responsive, LTR)、脱落酸响应(ABA responsive element, ABRE)等顺式作用元件(Sazegari et al., 2015).热激因子1 (heat shock factor 1, HSF1)、脱落酸响应结合蛋白(ABA responsive element binding protein/ABRE binding factors, AREB/ABF)可分别与DREB2A启动子中的热激应答和ABA响应元件结合, 调控其表达, 进而改变植物胁迫的耐受性(Kim et al., 2011; Liu et al., 2011).蛋白磷酸酶2C (type 2C protein phosphatase, PP2C)包含2个以上的ABRE元件, AREB1、AREB2和ABF3协同正调控AHG1、AHG3、HAI1及HAI2等PP2Cs基因表达(Yoshida et al., 2010).此外, 越来越多的研究证明ET信号通路参与拟南芥盐胁迫反应.进一步分析表明, ESE1 (ethylene and salt-inducible ERF1)在EIN3/EIL1下游调节ET响应基因.EIN3 (ethylene insensitive 3)可与ESE1启动子结合形成EIN3-ESE1转录复合体, 继而调控RD29A和COR15A等基因的表达(Zhang et al., 2011).磷酸化、泛素化、甲基化和乙酰化等组蛋白修饰方式通过调控染色质开放或封闭的空间状态, 激活或抑制AP2/ERF转录(Pfluger and Wagner, 2007).组蛋白去乙酰化酶(histone deacetylase, HDAC)和聚乙二醇(PEG)促进DREB1基因启动子区乙酰化, 增强DREB1基因的表达(Zhang et al., 2018).非生物胁迫下, 组蛋白变体和组蛋白修饰状态也可发生改变, 并可遗传到下一代(Asensii-Fabado et al., 2017).miRNA介导的RNA沉默和翻译抑制在AP2/ERF表达调控中也发挥重要作用.研究发现miRNA172靶向拟南芥AP2mRNA并抑制其翻译, 而miRNA156和miRNA838两者也具有类似调节AP2/ERF的功能(Kavas et al., 2015).此外, 在拟南芥、水稻及玉米中还发现通过可变剪切产生的OsDREB2A/2B、ZmDREB2A、WDREB2以及HvDRF1等多种AP2/-ERF功能异构体(Matsukura et al., 2010). ...
... DREB-A2亚组转录因子DREB2主要参与调节植物的抗旱和耐热性(Mizoi et al., 2012).干旱和高温可诱导DREB2基因的表达, 进而正向调控下游干旱或热胁迫响应基因(Maruyama et al., 2009).研究表明DREB-A4家族成员HRD (hardy)和DREB-A6家族成员(ERF53、RAP2.4和TG/RAP2.4a)也在干旱和耐盐性调控中发挥关键作用(Karaba et al., 2007; Lin et al., 2008; Chen et al., 2012; Zhu et al., 2014).过表达HRD能显著增强拟南芥和水稻植株的耐旱和耐盐性(Karaba et al., 2007).同样, 过表达TG (translucent green)导致植株叶片玻璃化, 叶片细胞含水量增加(Zhu et al., 2014).TG也参与调控抗坏血酸过氧化物酶基因APX (ascorbate peroxidase)的表达, 抵御活性氧(reactive oxygen species, ROS)引起的光氧化胁迫(Rudnik et al., 2017).除拟南芥外, 人们对水稻、玉米、番茄(Lycopersicon esculentum)、葡萄(Vitis vinifera)等作物的DREB1和DREB2转录因子也进行了广泛的研究(Xiao et al., 2006; Matsukura et al., 2010; Dou et al., 2014). ...
Arabidopsis AINTEGUMENTA mediates salt tolerance by trans-re-pressing SCABP8 1 2015
... RAVs和AP2s除调节茎与根细胞有丝分裂以及开花等发育过程(Osnato et al., 2012; Horstman et al., 2014)外, 还参与调控非生物胁迫响应.干旱和盐胁迫下, 拟南芥RAV1、RAV1L和RAV2的表达水平下降, 但三者对ABA的敏感性不同.进一步研究发现, 野生型、rav1突变体和RAV1超表达植株中RD29A、RD29B以及9-顺式环氧类胡萝卜素双加氧酶基因NCED9 (nine-cis-epxycarotenoid dioxygenase)和NCED3表现出相同的胁迫响应模式(Fu et al., 2014).Saito等(2004)发现过表达RAV1抑制ABA降解基因(CYP707A1和CYP707A2)的活性.正常条件下, RAV1过表达提高了NCED9和NCED3基因的表达水平, 而受胁迫后植株体内的ABA水平未发生明显变化(Fu et al., 2014).以上说明RAV1可通过不依赖ABA的途径负调控植物的非生物胁迫响应.此外, 研究发现ANT (AINTEGUMENTA)控制发育中的根细胞数量和大小, 同时也可抑制类SOS3钙结合蛋白8 (sos3-like calcium binding protein 8, SCABP8)转录, 负调控耐盐性, 而ANT功能缺失增强了拟南芥幼苗的耐盐能力, 并维持离子稳态(Meng et al., 2015a). ...
The ARF2- ANT-COR15A gene cascade regulates ABA-signaling- mediated resistance of large seeds to drought in Arabidopsis 1 2015
... 研究表明, 非生物胁迫下ABA可促进ANT (Meng et al., 2015b)、ERF53 (Hsieh et al., 2013)、RAP2.6L (Liu et al., 2012)和RAP2.6 (Zhu et al., 2010)转录, 上述转录因子进而结合胁迫应答顺式作用元件DRE/ABRE, 调控下游相关靶基因的表达, 增强植物对非生物胁迫的耐受能力.在水稻中ERF71正向调控ABA信号转导, 过表达ERF71可促进根中ABI5、PP2C68、RAB16C及RAB16D等基因上调表达.此外, ERF71可直接调控木质素生物合成基因CCR1、CCR10及C4H转录, 诱导根系径向生长, 改变根系构型, 提高耐旱能力(Lee et al., 2017; Li et al., 2018).盐胁迫和ABA双重诱导下, ABA增强DREB2的转录活性, 进而促进Rd29A协同表达(Lee et al., 2016). ...
Phosphorylation of an ERF transcription factor by Arabidopsis MPK3/MPK6 regulates plant defense gene induction and fungal resistance 1 2013
... 翻译后修饰(post-translational modification, PTM)是蛋白质合成途径的后期加工过程, 不同的修饰方式对AP2/ERF蛋白的活性、稳定性和丰度的影响有差异.磷酸化是转录因子的主要修饰途径之一, 磷酸化水平与下游靶基因的表达水平关系密切.拟南芥ABA信号通路的正调控因子蔗糖非酵解型蛋白激酶(Snf1-re-lated protein kinases, SnRKs)可磷酸化RAV1, 进而抑制RAV1的转录阻遏效应(Feng et al., 2014), 而ERF104和ERF6可被后MPK3/6 (mitogen-activated protein kinase 3/6)磷酸化修饰, 并在下游级联反应中发挥重要作用, 调控植物对病原体的免疫反应(Meng et al., 2013).DREB2A是拟南芥的关键转录激活因子, 在其中央序列存在一个负调控域(negative regulatory domain, NRD).热胁迫条件下, NRD中的Ser/Thr残基磷酸化水平降低, DREB2A蛋白稳定性提高, 敲除NRD后DREB2A呈组成型激活, 表明抑制NRD磷酸化可以稳定和激活DREB2A的表达, 从而增强植物的耐热性(Mizoi et al., 2019).泛素介导的蛋白质降解(26S蛋白酶体途径)也参与调节AP2/ ERF蛋白的稳定性.非生物胁迫条件下, DREB2A和ERF75/RAP-2.2蛋白分别被RING家族E3泛素连接酶DRIP1/2 (DREB2A-interacting protein1/2)和SINAT2 (seven in absentia of Arabidopsis 2)直接泛素化(Cheng et al., 2012; Papdi et al., 2015).研究发现CUL3-E3连接酶适配子BPMs (BTB/POZ and math domain)可与DREB2A蛋白的负调控域NRD在核内发生互作, 敲除BPM增加DREB2A蛋白的积累, 促进下游靶基因的表达, 提高植株的耐热和耐旱能力, 这也说明BPM对DREB2A蛋白的稳定性起负调控作用(Morimoto et al., 2017).此外, 研究发现ERF-VII类蛋白可在质膜上与乙酰辅基A结合蛋白(Acyl-CoA binding protein, ACBP)互作.在缺氧条件下, RAP2.12与ACBP分离后进入细胞核, 在脱酰胺基和精氨酰基化的作用下通过氧依赖途径移除ERF72/RAP2.3、ERF74/RAP2.12蛋白的甲硫氨酸N末端, 将半胱氨酸氧化成半胱亚磺酸, 引发蛋白快速降解并暴露出非稳定N端的氨基酸残基(Gibbs et al., 2015; Abbas et al., 2015). ...
Heat-induced inhibition of phosphorylation of the stress-protective transcription factor DREB2A promotes thermotolerance of Arabidopsis thaliana 1 2019
... 翻译后修饰(post-translational modification, PTM)是蛋白质合成途径的后期加工过程, 不同的修饰方式对AP2/ERF蛋白的活性、稳定性和丰度的影响有差异.磷酸化是转录因子的主要修饰途径之一, 磷酸化水平与下游靶基因的表达水平关系密切.拟南芥ABA信号通路的正调控因子蔗糖非酵解型蛋白激酶(Snf1-re-lated protein kinases, SnRKs)可磷酸化RAV1, 进而抑制RAV1的转录阻遏效应(Feng et al., 2014), 而ERF104和ERF6可被后MPK3/6 (mitogen-activated protein kinase 3/6)磷酸化修饰, 并在下游级联反应中发挥重要作用, 调控植物对病原体的免疫反应(Meng et al., 2013).DREB2A是拟南芥的关键转录激活因子, 在其中央序列存在一个负调控域(negative regulatory domain, NRD).热胁迫条件下, NRD中的Ser/Thr残基磷酸化水平降低, DREB2A蛋白稳定性提高, 敲除NRD后DREB2A呈组成型激活, 表明抑制NRD磷酸化可以稳定和激活DREB2A的表达, 从而增强植物的耐热性(Mizoi et al., 2019).泛素介导的蛋白质降解(26S蛋白酶体途径)也参与调节AP2/ ERF蛋白的稳定性.非生物胁迫条件下, DREB2A和ERF75/RAP-2.2蛋白分别被RING家族E3泛素连接酶DRIP1/2 (DREB2A-interacting protein1/2)和SINAT2 (seven in absentia of Arabidopsis 2)直接泛素化(Cheng et al., 2012; Papdi et al., 2015).研究发现CUL3-E3连接酶适配子BPMs (BTB/POZ and math domain)可与DREB2A蛋白的负调控域NRD在核内发生互作, 敲除BPM增加DREB2A蛋白的积累, 促进下游靶基因的表达, 提高植株的耐热和耐旱能力, 这也说明BPM对DREB2A蛋白的稳定性起负调控作用(Morimoto et al., 2017).此外, 研究发现ERF-VII类蛋白可在质膜上与乙酰辅基A结合蛋白(Acyl-CoA binding protein, ACBP)互作.在缺氧条件下, RAP2.12与ACBP分离后进入细胞核, 在脱酰胺基和精氨酰基化的作用下通过氧依赖途径移除ERF72/RAP2.3、ERF74/RAP2.12蛋白的甲硫氨酸N末端, 将半胱氨酸氧化成半胱亚磺酸, 引发蛋白快速降解并暴露出非稳定N端的氨基酸残基(Gibbs et al., 2015; Abbas et al., 2015). ...
AP2/ERF family transcription factors in plant abiotic stress responses 2 2012
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
... DREB-A2亚组转录因子DREB2主要参与调节植物的抗旱和耐热性(Mizoi et al., 2012).干旱和高温可诱导DREB2基因的表达, 进而正向调控下游干旱或热胁迫响应基因(Maruyama et al., 2009).研究表明DREB-A4家族成员HRD (hardy)和DREB-A6家族成员(ERF53、RAP2.4和TG/RAP2.4a)也在干旱和耐盐性调控中发挥关键作用(Karaba et al., 2007; Lin et al., 2008; Chen et al., 2012; Zhu et al., 2014).过表达HRD能显著增强拟南芥和水稻植株的耐旱和耐盐性(Karaba et al., 2007).同样, 过表达TG (translucent green)导致植株叶片玻璃化, 叶片细胞含水量增加(Zhu et al., 2014).TG也参与调控抗坏血酸过氧化物酶基因APX (ascorbate peroxidase)的表达, 抵御活性氧(reactive oxygen species, ROS)引起的光氧化胁迫(Rudnik et al., 2017).除拟南芥外, 人们对水稻、玉米、番茄(Lycopersicon esculentum)、葡萄(Vitis vinifera)等作物的DREB1和DREB2转录因子也进行了广泛的研究(Xiao et al., 2006; Matsukura et al., 2010; Dou et al., 2014). ...
BPM-CUL3 E3 ligase modulates thermotolerance by facilitating negative regulatory domain-mediated degradation of DREB2A in Arabidopsis 1 2017
... 翻译后修饰(post-translational modification, PTM)是蛋白质合成途径的后期加工过程, 不同的修饰方式对AP2/ERF蛋白的活性、稳定性和丰度的影响有差异.磷酸化是转录因子的主要修饰途径之一, 磷酸化水平与下游靶基因的表达水平关系密切.拟南芥ABA信号通路的正调控因子蔗糖非酵解型蛋白激酶(Snf1-re-lated protein kinases, SnRKs)可磷酸化RAV1, 进而抑制RAV1的转录阻遏效应(Feng et al., 2014), 而ERF104和ERF6可被后MPK3/6 (mitogen-activated protein kinase 3/6)磷酸化修饰, 并在下游级联反应中发挥重要作用, 调控植物对病原体的免疫反应(Meng et al., 2013).DREB2A是拟南芥的关键转录激活因子, 在其中央序列存在一个负调控域(negative regulatory domain, NRD).热胁迫条件下, NRD中的Ser/Thr残基磷酸化水平降低, DREB2A蛋白稳定性提高, 敲除NRD后DREB2A呈组成型激活, 表明抑制NRD磷酸化可以稳定和激活DREB2A的表达, 从而增强植物的耐热性(Mizoi et al., 2019).泛素介导的蛋白质降解(26S蛋白酶体途径)也参与调节AP2/ ERF蛋白的稳定性.非生物胁迫条件下, DREB2A和ERF75/RAP-2.2蛋白分别被RING家族E3泛素连接酶DRIP1/2 (DREB2A-interacting protein1/2)和SINAT2 (seven in absentia of Arabidopsis 2)直接泛素化(Cheng et al., 2012; Papdi et al., 2015).研究发现CUL3-E3连接酶适配子BPMs (BTB/POZ and math domain)可与DREB2A蛋白的负调控域NRD在核内发生互作, 敲除BPM增加DREB2A蛋白的积累, 促进下游靶基因的表达, 提高植株的耐热和耐旱能力, 这也说明BPM对DREB2A蛋白的稳定性起负调控作用(Morimoto et al., 2017).此外, 研究发现ERF-VII类蛋白可在质膜上与乙酰辅基A结合蛋白(Acyl-CoA binding protein, ACBP)互作.在缺氧条件下, RAP2.12与ACBP分离后进入细胞核, 在脱酰胺基和精氨酰基化的作用下通过氧依赖途径移除ERF72/RAP2.3、ERF74/RAP2.12蛋白的甲硫氨酸N末端, 将半胱氨酸氧化成半胱亚磺酸, 引发蛋白快速降解并暴露出非稳定N端的氨基酸残基(Gibbs et al., 2015; Abbas et al., 2015). ...
Ethylene response factors: a key regulatory hub in hormone and stress signaling 2 2015
... ERF亚家族转录因子是ET介导的胁迫应答信号通路下游的关键调控因子(Licausi et al., 2013; Gibbs et al., 2015; Kazan, 2015; Müller and Munné-Bosch, 2015).冷胁迫下, 植物体内的ET主要起负调控作用, 过量ET或1-氨基环丙烷-1-羧酸降低植物的耐冷性, 而ET生物合成抑制剂氨基乙氧基甘氨酸或感知拮抗剂Ag+具有相反的作用(Shi et al., 2012).etr1-1、ein4-1、ein2-5、ein3-1和ein3eil突变体表现出更强的耐冷性, 但组成型ET响应etr1-1和ein3过表达植株表现出耐冷性减弱; 冷胁迫诱导ETR1、EIN4、EBF1及EBF2快速上调表达, 而EIN2和EIL1下调表达.进一步分析表明, EIN3通过与启动子区特定元件结合, 抑制ARR5、ARR7、ARR15和CBF转录, ARRs作为冷信号通路的关键节点基因, 在ET和CTK介导的胁迫响应网络中起着关键作用, 过表达ARRs使植株的耐冷能力增强(Shi et al., 2012).此外, EIN3可激活ERF1和ESEs, 进而参与调控下游胁迫相关基因, 提高植物的耐盐性(Kazan, 2015; Tao et al., 2015).35S:AtERF4-GFP转基因植株中ABI2、RD29B和RAP18的表达受抑制, 进一步说明AtERF4是ET和ABA信号途径的负调控因子(Yang et al., 2011). ...
Genome-wide analysis of the ERF gene family in Arabidopsis and rice 4 2006
... AP2/ERF转录因子具有独特的结构特征, 其由DNA结合域(DNA-binding domain)、转录调控域(transcription regulation domain)、寡聚化位点(oligomerization site)和核定位信号(nuclear localization signal, NLS) 4个主要功能区组成, 并以高度保守的AP2/ ERF结合域而得名(Sakuma et al., 2002).通常情况下, AP2/ERF转录因子至少含有1个AP2结构域的DNA结合区, 该结构域包含60-70个氨基酸残基, 按照3个β折叠和1个α螺旋方式形成典型的三维结构(Nakano et al., 2006; Abiri et al., 2017).根据包含的AP2/ERF结构域数量和结合序列特点, 可将AP2/ ERF家族分为AP2 (APETALA2)、RAV (related to ABI3/VP1)、脱水反应元件结合蛋白(dehydration- responsive element binding protein, DREB) (A1-A6亚组)、乙烯应答因子(ethylene-responsive factor, ERF) (V-X亚组)和Soloist五个亚族(Nakano et al., 2006; Licausi et al., 2013).其中, 对AP2、DREB和ERF的研究较为广泛和深入, 而有关Soloist亚族的研究报道极少, 已知其核苷酸序列在多数植物中高度保守(Sun et al., 2016). ...
... ).根据包含的AP2/ERF结构域数量和结合序列特点, 可将AP2/ ERF家族分为AP2 (APETALA2)、RAV (related to ABI3/VP1)、脱水反应元件结合蛋白(dehydration- responsive element binding protein, DREB) (A1-A6亚组)、乙烯应答因子(ethylene-responsive factor, ERF) (V-X亚组)和Soloist五个亚族(Nakano et al., 2006; Licausi et al., 2013).其中, 对AP2、DREB和ERF的研究较为广泛和深入, 而有关Soloist亚族的研究报道极少, 已知其核苷酸序列在多数植物中高度保守(Sun et al., 2016). ...
... 许多研究表明, AP2/ERF转录因子可直接与靶基因启动子结合, 激活或抑制下游靶基因的表达, 其N端DNA结合域和C端转录激活域在胁迫相关靶基因的转录激活中起重要作用(Nakano et al., 2006).进一步分析表明, AP2/ERF转录因子中含有EAR基序(LxLxL或DLNxxP)、TLLLFR基序和BRD结构域(B3 repression domain, 保守序列为R/KLFGV)等多个转录抑制结构域(Ikeda and Ohme-Takagi, 2009; Kagale and Rozwadowski, 2011; Deepika et al., 2016).含EAR基序的转录因子是一种重要的抑制子, 在植物非生物胁迫应答基因的诱导表达中起负调控作用.Tiwari等(2012)从拟南芥ERF和AP2亚族中发现了一种转录激活关键元件EDLL, EDLL可与EAR基序(ERF-associated amphiphilic repression)发生互作, 拮抗后者介导的转录抑制作用.而不同的AP2/ ERF转录因子也可协同招募TPL (topless)和TPR (topless-related)等转录共抑制子(Causier et al., 2012)或组蛋白修饰因子, 进而抑制下游靶基因的表达(Song et al., 2005; Song and Galbraith, 2006).Song等(2005)发现AtERF7特异性地结合GCC-box, 与抑制因子ATSIN3及组蛋白去乙酰化酶19 (histone deacetylase 19, HDA19)发生互作并形成转录因子复合物, 参与干旱胁迫和ABA响应调控.AtERF7过表达株系保卫细胞对ABA的敏感性减弱, 水分散失增加; 反之, AtERF7 RNA干扰系对ABA的敏感性增强.类似的, ERF3能与组蛋白去乙酰化酶复合物亚基SAP18 (SIN3 associated polypeptide P18)互作, 共同招募HDA19形成复合体, 进而抑制相关基因的表达(Song and Galbraith, 2006).BRD结构域被认为是RAV1和RAV2转录因子行使抑制功能的关键元件, 其保守序列中第1位氨基酸残基多为疏水性亮氨酸、缬氨酸和蛋氨酸残基, 突变导致功能丧失(Ikeda and Ohme-Takagi, 2009).研究显示, 与AP2/ERFs协同调控的基因同样可被AP2/ERFs靶基因富集, 分析此类基因的同源基序有助于识别转录因子的假定靶基因并预测其生物学功能(Franco-Zorrillaetal et al., 2014). ...
... 部分A5亚组DREB类转录因子则对上述调控网络具有负反馈调节作用.DEAR1 (DREB and EAR motif protein 1)可能既在CBF上游作用, 又与RAP2.1在CBF下游扮演冷胁迫响应的调节子, 调节RD29A/COR78、COR15A和KIN1等冷诱导相关基因的表达, 负调控植物的耐冷和耐旱性(Fowler and Thomashow, 2002; Tsutsui et al., 2009; Dong and Liu, 2010).DEAR1过表达抑制冷诱导CBF的表达, 进而降低植物的耐冷能力(Tsutsui et al., 2009).低温胁迫或组成型表达CBF可诱导RAP2.1的表达, CBF2比RAP2.1对低温更为敏感(Dong and Liu, 2010).在CBF调控通路中, DEAR1和RAP2.1如何整合还需深入研究.RAP2.1是否与其它DEAR转录因子共同调节植物非生物胁迫响应仍不清楚.研究发现DREB-A5亚组有6种含EAR基序的转录抑制子可与DRE元件结合, 进而调控非生物胁迫响应基因的表达(Nakano et al., 2006). ...
Selective autophagy of BES1 mediated by DSK2 balances plant growth and survival 2 2017
... AP2/ERF转录因子在参与调控植物应对冷、干旱、高温、高盐及缺氧等多种非生物胁迫过程中具有重要功能(Licausi et al., 2013).其中, DREB和ERF亚家族成员在植物非生物逆境应答中起重要调控作用.研究表明其参与调控的植物环境胁迫应答信号网络比较复杂.胁迫条件下, 部分AP2/ERFs可被快速持续地诱导表达, 而其它进程的响应较为缓慢, 说明可能存在依赖或不依赖于激素信号途径2种胁迫响应模式, 且两者之间存在一定的交叉互作(Van den Broeck et al., 2017).前人的研究也证明, AP2/ERFs广泛参与调控ABA、ET、GA、细胞分裂素(cytokinin, CTK)和油菜素内酯(brassinolide, BR)等植物激素介导的胁迫响应(Colebrook et al., 2014; Kazan et al., 2015; Tao et al., 2015; Sah et al., 2016; Nolan et al., 2017) (图1). ...
... BZR1可结合CBF1和CBF2的启动子区并促进两者在冷胁迫下的表达.bin2-3、bil1、bil2缺失突变体在持续的冷胁迫下可以诱导去磷酸化BZR1的积累, BZR1通过调控WKRY6、PYL6、SOC1、JMT和SAG21等不依赖CBF途径的COR基因而增强拟南芥的耐冷性(Li et al., 2017).研究发现BR可与干旱诱导相关转录因子RD26互作, 拮抗干旱响应基因的表达, 进而负调控植物的抗旱性(Ye et al., 2017).BR与ABA信号途径中的受体、转录因子之间也产生拮抗效应, 进而调节胁迫下植物体的生长平衡(Nolan et al., 2017).目前已从BES1和BZR1靶基因中鉴定出一些AP2/ERF类转录因子, 暗示AP2/ERF具有整合BR信号通路与非生物胁迫响应的潜在功能(Sun et al., 2010; Yu et al., 2011).ERF72可能是研究BR信号通路与胁迫响应交叉互作的候选基因.在拟南芥中, ERF72/RAP2.3拮抗BZR1和ARF6 (auxin responsive factor 6), 抑制下胚轴伸长, 此过程中BR信号途径如何与胁迫响应调控网络协同互作发挥作用尚不清楚(Liu et al., 2018b).研究表明, SUB1A参与调节GA和BR的交叉网络互作(Schmitz et al., 2013).GA和BR途径通过DELLA蛋白、BZR1等转录因子协同调控植物的生长(Oh et al., 2012).在植物淹水过程中SUB1A差异调节BR合成相关基因的表达, 激活BR生物合成和信号转导, 诱导GA降解关键基因GA2ox7的表达, 进而控制水稻植株体内的GA水平, 促进DELLA蛋白的积累(Schmitz et al., 2013).因此, 推测AP2/ERF可能通过不同的机制参与调控BR信号通路. ...
Arabidopsis CBF1 and CBF3 have a different function than CBF2 in cold acclimation and define different gene classes in the CBF regulon 1 2007
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
Interaction between BZR1 and PIF4 integrates brassinosteroid and environmental responses 1 2012
... BZR1可结合CBF1和CBF2的启动子区并促进两者在冷胁迫下的表达.bin2-3、bil1、bil2缺失突变体在持续的冷胁迫下可以诱导去磷酸化BZR1的积累, BZR1通过调控WKRY6、PYL6、SOC1、JMT和SAG21等不依赖CBF途径的COR基因而增强拟南芥的耐冷性(Li et al., 2017).研究发现BR可与干旱诱导相关转录因子RD26互作, 拮抗干旱响应基因的表达, 进而负调控植物的抗旱性(Ye et al., 2017).BR与ABA信号途径中的受体、转录因子之间也产生拮抗效应, 进而调节胁迫下植物体的生长平衡(Nolan et al., 2017).目前已从BES1和BZR1靶基因中鉴定出一些AP2/ERF类转录因子, 暗示AP2/ERF具有整合BR信号通路与非生物胁迫响应的潜在功能(Sun et al., 2010; Yu et al., 2011).ERF72可能是研究BR信号通路与胁迫响应交叉互作的候选基因.在拟南芥中, ERF72/RAP2.3拮抗BZR1和ARF6 (auxin responsive factor 6), 抑制下胚轴伸长, 此过程中BR信号途径如何与胁迫响应调控网络协同互作发挥作用尚不清楚(Liu et al., 2018b).研究表明, SUB1A参与调节GA和BR的交叉网络互作(Schmitz et al., 2013).GA和BR途径通过DELLA蛋白、BZR1等转录因子协同调控植物的生长(Oh et al., 2012).在植物淹水过程中SUB1A差异调节BR合成相关基因的表达, 激活BR生物合成和信号转导, 诱导GA降解关键基因GA2ox7的表达, 进而控制水稻植株体内的GA水平, 促进DELLA蛋白的积累(Schmitz et al., 2013).因此, 推测AP2/ERF可能通过不同的机制参与调控BR信号通路. ...
TEMPRANILLO genes link photoperiod and gibberellin pathways to control flowering in Arabidopsis 1 2012
... RAVs和AP2s除调节茎与根细胞有丝分裂以及开花等发育过程(Osnato et al., 2012; Horstman et al., 2014)外, 还参与调控非生物胁迫响应.干旱和盐胁迫下, 拟南芥RAV1、RAV1L和RAV2的表达水平下降, 但三者对ABA的敏感性不同.进一步研究发现, 野生型、rav1突变体和RAV1超表达植株中RD29A、RD29B以及9-顺式环氧类胡萝卜素双加氧酶基因NCED9 (nine-cis-epxycarotenoid dioxygenase)和NCED3表现出相同的胁迫响应模式(Fu et al., 2014).Saito等(2004)发现过表达RAV1抑制ABA降解基因(CYP707A1和CYP707A2)的活性.正常条件下, RAV1过表达提高了NCED9和NCED3基因的表达水平, 而受胁迫后植株体内的ABA水平未发生明显变化(Fu et al., 2014).以上说明RAV1可通过不依赖ABA的途径负调控植物的非生物胁迫响应.此外, 研究发现ANT (AINTEGUMENTA)控制发育中的根细胞数量和大小, 同时也可抑制类SOS3钙结合蛋白8 (sos3-like calcium binding protein 8, SCABP8)转录, 负调控耐盐性, 而ANT功能缺失增强了拟南芥幼苗的耐盐能力, 并维持离子稳态(Meng et al., 2015a). ...
Identification and functional prediction of stress responsive AP2/ERF transcription factors in Brassica napus by genome-wide analysis 1 2017
... 正常环境下, AP2/ERF转录因子表达水平较低, 激素或逆境胁迫相关基因与AP2/ERF转录因子保守的顺式作用元件结合, 进而调控其表达(Owji et al., 2017).研究表明, 拟南芥DREB1A/CBF3、DREB1C/CBF2、DREB2C、DREB2G和DEAR3的启动子中包含较多的热激元件(heat shock element, HSE)、低温响应(low temperature responsive, LTR)、脱落酸响应(ABA responsive element, ABRE)等顺式作用元件(Sazegari et al., 2015).热激因子1 (heat shock factor 1, HSF1)、脱落酸响应结合蛋白(ABA responsive element binding protein/ABRE binding factors, AREB/ABF)可分别与DREB2A启动子中的热激应答和ABA响应元件结合, 调控其表达, 进而改变植物胁迫的耐受性(Kim et al., 2011; Liu et al., 2011).蛋白磷酸酶2C (type 2C protein phosphatase, PP2C)包含2个以上的ABRE元件, AREB1、AREB2和ABF3协同正调控AHG1、AHG3、HAI1及HAI2等PP2Cs基因表达(Yoshida et al., 2010).此外, 越来越多的研究证明ET信号通路参与拟南芥盐胁迫反应.进一步分析表明, ESE1 (ethylene and salt-inducible ERF1)在EIN3/EIL1下游调节ET响应基因.EIN3 (ethylene insensitive 3)可与ESE1启动子结合形成EIN3-ESE1转录复合体, 继而调控RD29A和COR15A等基因的表达(Zhang et al., 2011).磷酸化、泛素化、甲基化和乙酰化等组蛋白修饰方式通过调控染色质开放或封闭的空间状态, 激活或抑制AP2/ERF转录(Pfluger and Wagner, 2007).组蛋白去乙酰化酶(histone deacetylase, HDAC)和聚乙二醇(PEG)促进DREB1基因启动子区乙酰化, 增强DREB1基因的表达(Zhang et al., 2018).非生物胁迫下, 组蛋白变体和组蛋白修饰状态也可发生改变, 并可遗传到下一代(Asensii-Fabado et al., 2017).miRNA介导的RNA沉默和翻译抑制在AP2/ERF表达调控中也发挥重要作用.研究发现miRNA172靶向拟南芥AP2mRNA并抑制其翻译, 而miRNA156和miRNA838两者也具有类似调节AP2/ERF的功能(Kavas et al., 2015).此外, 在拟南芥、水稻及玉米中还发现通过可变剪切产生的OsDREB2A/2B、ZmDREB2A、WDREB2以及HvDRF1等多种AP2/-ERF功能异构体(Matsukura et al., 2010). ...
The low oxygen, oxidative and osmotic stress responses synergistically act through the ethylene response factor VII genes RAP2.12, RAP2.2 and RAP2.3 2 2015
... 翻译后修饰(post-translational modification, PTM)是蛋白质合成途径的后期加工过程, 不同的修饰方式对AP2/ERF蛋白的活性、稳定性和丰度的影响有差异.磷酸化是转录因子的主要修饰途径之一, 磷酸化水平与下游靶基因的表达水平关系密切.拟南芥ABA信号通路的正调控因子蔗糖非酵解型蛋白激酶(Snf1-re-lated protein kinases, SnRKs)可磷酸化RAV1, 进而抑制RAV1的转录阻遏效应(Feng et al., 2014), 而ERF104和ERF6可被后MPK3/6 (mitogen-activated protein kinase 3/6)磷酸化修饰, 并在下游级联反应中发挥重要作用, 调控植物对病原体的免疫反应(Meng et al., 2013).DREB2A是拟南芥的关键转录激活因子, 在其中央序列存在一个负调控域(negative regulatory domain, NRD).热胁迫条件下, NRD中的Ser/Thr残基磷酸化水平降低, DREB2A蛋白稳定性提高, 敲除NRD后DREB2A呈组成型激活, 表明抑制NRD磷酸化可以稳定和激活DREB2A的表达, 从而增强植物的耐热性(Mizoi et al., 2019).泛素介导的蛋白质降解(26S蛋白酶体途径)也参与调节AP2/ ERF蛋白的稳定性.非生物胁迫条件下, DREB2A和ERF75/RAP-2.2蛋白分别被RING家族E3泛素连接酶DRIP1/2 (DREB2A-interacting protein1/2)和SINAT2 (seven in absentia of Arabidopsis 2)直接泛素化(Cheng et al., 2012; Papdi et al., 2015).研究发现CUL3-E3连接酶适配子BPMs (BTB/POZ and math domain)可与DREB2A蛋白的负调控域NRD在核内发生互作, 敲除BPM增加DREB2A蛋白的积累, 促进下游靶基因的表达, 提高植株的耐热和耐旱能力, 这也说明BPM对DREB2A蛋白的稳定性起负调控作用(Morimoto et al., 2017).此外, 研究发现ERF-VII类蛋白可在质膜上与乙酰辅基A结合蛋白(Acyl-CoA binding protein, ACBP)互作.在缺氧条件下, RAP2.12与ACBP分离后进入细胞核, 在脱酰胺基和精氨酰基化的作用下通过氧依赖途径移除ERF72/RAP2.3、ERF74/RAP2.12蛋白的甲硫氨酸N末端, 将半胱氨酸氧化成半胱亚磺酸, 引发蛋白快速降解并暴露出非稳定N端的氨基酸残基(Gibbs et al., 2015; Abbas et al., 2015). ...
... ERF亚族成员在低氧和淹水胁迫响应过程中也发挥重要作用(Bui et al., 2015; Gibbs et al., 2015).SUB1A (submergence 1A)是ERF亚族B2亚组成员之一, Sub1A-1超表达的同时促进乙醇脱氢酶基因(alcohol dehydrogenase 1, Adh1)上调表达, 抑制Sub1C的转录水平, 说明Sub1A-1是耐淹性的主要调节因子, 在淹水胁迫应答调控中起关键作用(Xu et al., 2006).淹水状态下, 低水平海藻糖-6-磷酸合酶(trehalose-6-phosphate, T6P)与CIPK15 (CBL-inter-acting protein kinase 15)协同激活SnRK1, SnRK1介导SUB1A抑制幼苗胚轴生长和呼吸相关基因的表达, 直接或间接调控碳同化过程.而SK1/SK2 (snorkel1/2)激活参与叶柄、茎和导管发育相关基因, 促进植株快速生长(Hattori et al., 2009; Locke et al., 2018).缺氧胁迫下, 拟南芥ERF-VIIs可以通过氧依赖的N端规则通路降解(Gibbs et al., 2015).RING (really interesting gene)类蛋白SINAT2可通过其N端保守的C3H4型RING结构域调控RAP2.12, 沉默SINAT1/2可上调RAP2.12的表达水平(Papdi et al., 2015).此外, ERF71/HRE2、ERF72/RAP2.3、ERF73/HRE1、ERF74/RAP2.12及ERF75/RAP2.2d也受缺氧胁迫诱导, 进而正向调控糖代谢和激素信号相关的缺氧响应基因, 提高植株的缺氧耐受能力.过表达RAP2.2、RAP2.3和RAP2.12可提高植株在低氧和渗透胁迫下的存活率, 而rap2.12-2/rap2.3-1双突变体对淹水和渗透胁迫表现出较高的敏感性(Yao et al., 2017). ...
Cuticular wax biosynthesis is positively regulated by WRINKLED 4, an AP2/ERF- type transcription factor, in Arabidopsis stems 1 2016
... AP2/ERF转录因子通过特异性结合胁迫应答基因启动子区顺式元件, 参与调控植物的逆境防御反应.DREB亚族成员特异性识别并结合启动子区域的DRE/CRT (dehydration responsive element/C-re-peat)元件(核心序列A/GCCGAC), 诱导干旱、低温和盐胁迫相关应答基因的表达.ERF亚族成员则可通过与乙烯响应元件(ethylene response element, ERE) (核心序列AGCCGCC, 又称GCC-box)结合, 从而参与调控乙烯应答和非生物胁迫(Franco-Zorrilla et al., 2014).此外, 研究发现许多AP2/ERF转录因子可同时与DRE/CRT和ERE两种元件结合, 如DREB亚族成员TINY、CBF1、ERF53、RAP2.4、TG/RAP2.4a以及ERF亚族成员ERF1、ERF4和ERF71 (Chen et al., 2012; Zhu et al., 2014; Lee et al., 2015), 但结合能力有差异.在拟南芥中, DREB2C和DEAR3与类GCC-box元件(GCCGCC)的结合能力远超DRE元件(Franco-Zorrilla et al., 2014).AP2/ERF转录因子还能与DRE和ERE之外的顺式元件结合, 如偶联元件1 (CE1, TGCCACCG)、类偶联元件3 (CE-3like, CGCG)和缺氧响应启动子元件(hypoxia-responsive promoter element, HRPE), 以及ATCTA、CAACA、CATGCA、CAA/CA/CTG、ATCGAG和(NC/GT) CGNCCA (Bossi et al., 2009; Zhu et al., 2010; Chen et al., 2016; Gasch et al., 2016; Park et al., 2016).近年来, 科学家通过蛋白质结合微阵列(protein binding microarray, PBM)技术发现结构特征高度相似的转录因子具有相似的DNA结合位点, 同时也具有相似的生物学功能.该研究结果合理地解释了转录因子功能冗余现象, 也为预测和挖掘未知AP2/ERF转录因子提供了高效可行的技术方法(Franco-Zorrilla et al., 2014). ...
Regulation of the Arabidopsis CBF regulon by a complex low-temperature regulatory network 3 2015
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
... 基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
... 的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
The rice SUB1A gene: making adaptation to submergence and post-submergence possible 1 2018
... 在拟南芥和水稻中, SUB1A、SK1和SK2参与GA信号转导通路的调控.淹水胁迫下, SUB1A激活SLR1 (slender rice 1)和SLRL1 (slender rice 1 like 1) (DELLA类似蛋白)转录, 同时SUB1A抑制受GA诱导的淀粉代谢相关基因的表达, 调控植株节间伸长和呼吸作用(Fukao and Bailey-Serres, 2008; Locke et al., 2018; Perata, 2018).SK1和SK2两个转录因子中均含有EIN3结合位点, 电泳迁移率测定显示, 类EIN3基因EIL1b可与SK1和SK2启动子结合, 激活其转录活性.研究发现, 淹水状态下拟南芥中ET合成增加, 进而诱导SK1和SK2表达, 激发GA20oxs基因上调表达, 促进植株节间伸长, 减轻淹水对其生长的抑制效应(Hattori et al., 2009; Ayano et al., 2014). ...
Histone modifications and dynamic regulation of genome accessibility in plants 1 2007
... 正常环境下, AP2/ERF转录因子表达水平较低, 激素或逆境胁迫相关基因与AP2/ERF转录因子保守的顺式作用元件结合, 进而调控其表达(Owji et al., 2017).研究表明, 拟南芥DREB1A/CBF3、DREB1C/CBF2、DREB2C、DREB2G和DEAR3的启动子中包含较多的热激元件(heat shock element, HSE)、低温响应(low temperature responsive, LTR)、脱落酸响应(ABA responsive element, ABRE)等顺式作用元件(Sazegari et al., 2015).热激因子1 (heat shock factor 1, HSF1)、脱落酸响应结合蛋白(ABA responsive element binding protein/ABRE binding factors, AREB/ABF)可分别与DREB2A启动子中的热激应答和ABA响应元件结合, 调控其表达, 进而改变植物胁迫的耐受性(Kim et al., 2011; Liu et al., 2011).蛋白磷酸酶2C (type 2C protein phosphatase, PP2C)包含2个以上的ABRE元件, AREB1、AREB2和ABF3协同正调控AHG1、AHG3、HAI1及HAI2等PP2Cs基因表达(Yoshida et al., 2010).此外, 越来越多的研究证明ET信号通路参与拟南芥盐胁迫反应.进一步分析表明, ESE1 (ethylene and salt-inducible ERF1)在EIN3/EIL1下游调节ET响应基因.EIN3 (ethylene insensitive 3)可与ESE1启动子结合形成EIN3-ESE1转录复合体, 继而调控RD29A和COR15A等基因的表达(Zhang et al., 2011).磷酸化、泛素化、甲基化和乙酰化等组蛋白修饰方式通过调控染色质开放或封闭的空间状态, 激活或抑制AP2/ERF转录(Pfluger and Wagner, 2007).组蛋白去乙酰化酶(histone deacetylase, HDAC)和聚乙二醇(PEG)促进DREB1基因启动子区乙酰化, 增强DREB1基因的表达(Zhang et al., 2018).非生物胁迫下, 组蛋白变体和组蛋白修饰状态也可发生改变, 并可遗传到下一代(Asensii-Fabado et al., 2017).miRNA介导的RNA沉默和翻译抑制在AP2/ERF表达调控中也发挥重要作用.研究发现miRNA172靶向拟南芥AP2mRNA并抑制其翻译, 而miRNA156和miRNA838两者也具有类似调节AP2/ERF的功能(Kavas et al., 2015).此外, 在拟南芥、水稻及玉米中还发现通过可变剪切产生的OsDREB2A/2B、ZmDREB2A、WDREB2以及HvDRF1等多种AP2/-ERF功能异构体(Matsukura et al., 2010). ...
Reactive oxygen species signaling and stomatal movement in plant responses to drought stress and pathogen attack 1 2018
... RbohD (respiratory burst oxidase homolog D)是一种NADPH氧化酶, 催化活性氧产生(Yao et al., 2017).ROS是细胞生化反应过程中产生的毒性物质, 可作为信号分子触发胁迫响应和信号转导, RAPs通过依赖于RbohD的途径调节非生物胁迫(Qi et al., 2018).erf74和erf74/erf75突变体中, ROS的产生和RbohD基因的表达均受抑制, ERF74和ERF75通过依赖于RbohD的ROS激活途径协同调控缺氧响应.其中, ERF74可结合RbohD基因的启动子并激活其表达, ERF74过表达促进后期ROS清除酶相关基因的表达, 说明在胁迫响应过程中ERF74作为分子开关控制RbohD依赖的ROS迸发(Yao et al., 2017).研究发现水稻中也存在类似的调控机制, OsLG3通过诱导活性氧清除相关基因APX1、APX4、APX8、CATB和POD1的表达进而正调控水稻的耐旱性(Xiong et al., 2018). ...
Processing and subcellular trafficking of ER-tethered EIN2 control response to ethylene gas 1 2012
Achievements and challenges in understanding plant abiotic stress responses and tolerance 1 2011
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
A subset of Arabidopsis AP2 transcription factors mediates cytokinin responses in concert with a two-component pathway 3 2006
... 近年来, 有关ERF亚族参与非生物胁迫响应的研究越来越多.在拟南芥中发现多种非生物胁迫可诱导细胞分裂素应答因子(cytokinin response factor, CRF)的表达, CRF是ERF-VI亚族成员之一, 具有正向调节植物渗透胁迫耐受性和抗冻性作用(Rashotte et al., 2006).CRF2和CRF6可与生长素外输载体PIN (PIN- FORMED)基因启动子区的特定元件结合, 正向调控PIN基因的表达, 这在CRF超表达和缺失突变体研究中得到了验证(Kim, 2016).CRF2和CRF3均参与调控冷胁迫下拟南芥侧根(lateral root, LR)的发育, crf2、crf3、crf2/crf3突变体在冷胁迫下侧根密度比野生型植株更低, CRF2或CRF3过表达均可促进植株侧根密度的增加(Jeon et al., 2016).冷胁迫可诱导CRF2的表达, 因此CRF2可能通过诱导PIN1和PIN7的表达以弥补冷胁迫对侧根发育的影响; 而在缺失突变体crf2中, CRF3超表达可减弱冷胁迫对侧根生长的抑制效应, 推测CRF3可能通过其它途径参与侧根的发育调控(Jeon et al., 2016; Kim, 2016).进一步分析显示, CRF2和CRF3分别通过依赖和不依赖于TCS (two-component signaling)的途径响应冷胁迫(Joen et al., 2016).CRF4是少数不受CTK调控的CRF转录因子之一, 可促进CORs的表达并正向调控植物的冷/冻耐受性(Zwack et al., 2016a).多种非生物胁迫会诱导植物积累活性氧(ROS), 导致细胞损伤, 因此植物维持最低水平的活性氧对其生存至关重要.多种胁迫刺激均可诱导CRF6的表达, CRF6可能通过抑制细胞分裂素相关基因的表达介导细胞分裂素与氧化应激反应之间的联系(Zwack et al., 2016b).CRF6在H2O2诱导的氧化胁迫应答中直接或间接抑制ARR6 (type-A response regulator 6)、ARR9、ARR11、LOG7及ABCG14基因的表达.酵母单杂交实验显示, CRF6直接结合到ARR6启动子区, 激活ARR6的表达, 而对ARR9、LOG7的表达调控可能需要其它植物特异因子的参与(Zwack et al., 2016b). ...
... 细胞分裂素应答因子(cytokinin response factor, CRF)在植物胚、子叶和叶片发育过程中起关键调控作用(Rashotte et al., 2006).拟南芥crf1/2/5和crf2/3/6突变体的转录组分析表明CRF参与CTK介导的调控网络.CRF基因启动子区含有多个B型ARR结合位点, 据此推测CRFs很可能是B型ARR的直接靶标, 两者之间存在协同作用, 约60%的CTK响应基因同时受CRF和B型ARR调控(Rashotte et al., 2006).Zwack等(2013)认为CRF6是胁迫条件下AHK3和TCS介导的CTK信号通路下游的新组分, CRF6和ARR2可能通过一个部分重叠或相互作用的平行通路负调控胁迫诱导的叶片衰老过程, 这也表明CRF6和 CTK与非生物胁迫之间存在独特的关系(Zwack et al., 2013).CRF6除正向调节CTK通路外, 还抑制CTK的生物合成、信号转导和胞内转运相关靶基因的表达, 减轻CTK对非生物胁迫的负调控作用(Zwack et al., 2016b).CRF6和CTK在胁迫响应调控中的作用相反, 但在叶片衰老调控中的作用相似, 推测CRF6也可能通过2条途径调控CTK信号通路.目前有关CRF调控的具体机制仍不明确, 鉴定CRF靶基因和上游信号分子将有助于更好地理解CTK响应非生物胁迫的作用机制. ...
... 突变体的转录组分析表明CRF参与CTK介导的调控网络.CRF基因启动子区含有多个B型ARR结合位点, 据此推测CRFs很可能是B型ARR的直接靶标, 两者之间存在协同作用, 约60%的CTK响应基因同时受CRF和B型ARR调控(Rashotte et al., 2006).Zwack等(2013)认为CRF6是胁迫条件下AHK3和TCS介导的CTK信号通路下游的新组分, CRF6和ARR2可能通过一个部分重叠或相互作用的平行通路负调控胁迫诱导的叶片衰老过程, 这也表明CRF6和 CTK与非生物胁迫之间存在独特的关系(Zwack et al., 2013).CRF6除正向调节CTK通路外, 还抑制CTK的生物合成、信号转导和胞内转运相关靶基因的表达, 减轻CTK对非生物胁迫的负调控作用(Zwack et al., 2016b).CRF6和CTK在胁迫响应调控中的作用相反, 但在叶片衰老调控中的作用相似, 推测CRF6也可能通过2条途径调控CTK信号通路.目前有关CRF调控的具体机制仍不明确, 鉴定CRF靶基因和上游信号分子将有助于更好地理解CTK响应非生物胁迫的作用机制. ...
Genetic analysis reveals that C19-GA 2-oxidation is a major gibberellin inactivation pathway in Arabidopsis 1 2008
... 赤霉素(GA)广泛分布在植物中并参与调控叶、芽以及合子发育等多种生物学过程.GA由GA20氧化酶(gibberellin 20-oxidases, GA20ox)和GA3氧化酶(GA3ox)等关键酶合成, 在GA2氧化酶(GA2ox)作用下被分解代谢(Rieu et al., 2008).DELLA蛋白作为GA信号转导途径中的主要负调控因子, 也是整合生长素、ABA、ET和JA等植物激素或环境信号的枢纽蛋白.植物体接受GA信号后, 通过泛素-蛋白酶体途径降解DELLA蛋白, 解除其对生长的抑制作用(Zhang et al., 2009).冷、盐和渗透胁迫下, GA含量降低, 导致植物生长减缓(Colebrook et al., 2014).极低浓度GA下, DELLA蛋白抑制GA介导的应答调控, 外源GA处理促进DELLA蛋白降解(Claeys et al., 2012). ...
Specificity versus redundancy in the RAP2.4 transcription factor family of Arabidopsis thaliana: transcriptional regulation of genes for chloroplast peroxidases 1 2017
... DREB-A2亚组转录因子DREB2主要参与调节植物的抗旱和耐热性(Mizoi et al., 2012).干旱和高温可诱导DREB2基因的表达, 进而正向调控下游干旱或热胁迫响应基因(Maruyama et al., 2009).研究表明DREB-A4家族成员HRD (hardy)和DREB-A6家族成员(ERF53、RAP2.4和TG/RAP2.4a)也在干旱和耐盐性调控中发挥关键作用(Karaba et al., 2007; Lin et al., 2008; Chen et al., 2012; Zhu et al., 2014).过表达HRD能显著增强拟南芥和水稻植株的耐旱和耐盐性(Karaba et al., 2007).同样, 过表达TG (translucent green)导致植株叶片玻璃化, 叶片细胞含水量增加(Zhu et al., 2014).TG也参与调控抗坏血酸过氧化物酶基因APX (ascorbate peroxidase)的表达, 抵御活性氧(reactive oxygen species, ROS)引起的光氧化胁迫(Rudnik et al., 2017).除拟南芥外, 人们对水稻、玉米、番茄(Lycopersicon esculentum)、葡萄(Vitis vinifera)等作物的DREB1和DREB2转录因子也进行了广泛的研究(Xiao et al., 2006; Matsukura et al., 2010; Dou et al., 2014). ...
Abscisic acid and abiotic stress tolerance in crop plants 3 2016
... AP2/ERF转录因子在参与调控植物应对冷、干旱、高温、高盐及缺氧等多种非生物胁迫过程中具有重要功能(Licausi et al., 2013).其中, DREB和ERF亚家族成员在植物非生物逆境应答中起重要调控作用.研究表明其参与调控的植物环境胁迫应答信号网络比较复杂.胁迫条件下, 部分AP2/ERFs可被快速持续地诱导表达, 而其它进程的响应较为缓慢, 说明可能存在依赖或不依赖于激素信号途径2种胁迫响应模式, 且两者之间存在一定的交叉互作(Van den Broeck et al., 2017).前人的研究也证明, AP2/ERFs广泛参与调控ABA、ET、GA、细胞分裂素(cytokinin, CTK)和油菜素内酯(brassinolide, BR)等植物激素介导的胁迫响应(Colebrook et al., 2014; Kazan et al., 2015; Tao et al., 2015; Sah et al., 2016; Nolan et al., 2017) (图1). ...
... 植物激素是一类调控植物生长发育以及抵御不良环境影响的重要小分子.脱落酸(ABA)是应答干旱、盐、冷、热等非生物胁迫的关键激素之一.ABA可诱导气孔关闭, 调节根系结构, 促进渗透物质的合成, 进而抵御外部逆境(Sah et al., 2016).NCED是ABA合成的限速酶, 干旱和渗透胁迫诱导NCED基因快速上调表达, 促进ABA的生物合成.ABA与其受体PYR/PLY/ RCAR结合, 再与PP2C作用形成复合物, 解除PP2C对SnRK2激酶活性的抑制作用.活性形式的SnRK2磷酸化AREB/ABF、离子通道蛋白和NADPH氧化酶等下游底物, 进而诱导ABA响应基因的表达(Finkelstein, 2013; Sah et al., 2016). ...
... ; Sah et al., 2016). ...
Arabidopsis CYP707As encode(+)-abscisic acid 8°-hydroxylase, a key enzyme in the oxidative catabolism of abscisic acid 1 2004
... RAVs和AP2s除调节茎与根细胞有丝分裂以及开花等发育过程(Osnato et al., 2012; Horstman et al., 2014)外, 还参与调控非生物胁迫响应.干旱和盐胁迫下, 拟南芥RAV1、RAV1L和RAV2的表达水平下降, 但三者对ABA的敏感性不同.进一步研究发现, 野生型、rav1突变体和RAV1超表达植株中RD29A、RD29B以及9-顺式环氧类胡萝卜素双加氧酶基因NCED9 (nine-cis-epxycarotenoid dioxygenase)和NCED3表现出相同的胁迫响应模式(Fu et al., 2014).Saito等(2004)发现过表达RAV1抑制ABA降解基因(CYP707A1和CYP707A2)的活性.正常条件下, RAV1过表达提高了NCED9和NCED3基因的表达水平, 而受胁迫后植株体内的ABA水平未发生明显变化(Fu et al., 2014).以上说明RAV1可通过不依赖ABA的途径负调控植物的非生物胁迫响应.此外, 研究发现ANT (AINTEGUMENTA)控制发育中的根细胞数量和大小, 同时也可抑制类SOS3钙结合蛋白8 (sos3-like calcium binding protein 8, SCABP8)转录, 负调控耐盐性, 而ANT功能缺失增强了拟南芥幼苗的耐盐能力, 并维持离子稳态(Meng et al., 2015a). ...
DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression 1 2002
... AP2/ERF转录因子具有独特的结构特征, 其由DNA结合域(DNA-binding domain)、转录调控域(transcription regulation domain)、寡聚化位点(oligomerization site)和核定位信号(nuclear localization signal, NLS) 4个主要功能区组成, 并以高度保守的AP2/ ERF结合域而得名(Sakuma et al., 2002).通常情况下, AP2/ERF转录因子至少含有1个AP2结构域的DNA结合区, 该结构域包含60-70个氨基酸残基, 按照3个β折叠和1个α螺旋方式形成典型的三维结构(Nakano et al., 2006; Abiri et al., 2017).根据包含的AP2/ERF结构域数量和结合序列特点, 可将AP2/ ERF家族分为AP2 (APETALA2)、RAV (related to ABI3/VP1)、脱水反应元件结合蛋白(dehydration- responsive element binding protein, DREB) (A1-A6亚组)、乙烯应答因子(ethylene-responsive factor, ERF) (V-X亚组)和Soloist五个亚族(Nakano et al., 2006; Licausi et al., 2013).其中, 对AP2、DREB和ERF的研究较为广泛和深入, 而有关Soloist亚族的研究报道极少, 已知其核苷酸序列在多数植物中高度保守(Sun et al., 2016). ...
A study on the regulatory network with promoter analysis for Arabidopsis DREB-genes 1 2015
... 正常环境下, AP2/ERF转录因子表达水平较低, 激素或逆境胁迫相关基因与AP2/ERF转录因子保守的顺式作用元件结合, 进而调控其表达(Owji et al., 2017).研究表明, 拟南芥DREB1A/CBF3、DREB1C/CBF2、DREB2C、DREB2G和DEAR3的启动子中包含较多的热激元件(heat shock element, HSE)、低温响应(low temperature responsive, LTR)、脱落酸响应(ABA responsive element, ABRE)等顺式作用元件(Sazegari et al., 2015).热激因子1 (heat shock factor 1, HSF1)、脱落酸响应结合蛋白(ABA responsive element binding protein/ABRE binding factors, AREB/ABF)可分别与DREB2A启动子中的热激应答和ABA响应元件结合, 调控其表达, 进而改变植物胁迫的耐受性(Kim et al., 2011; Liu et al., 2011).蛋白磷酸酶2C (type 2C protein phosphatase, PP2C)包含2个以上的ABRE元件, AREB1、AREB2和ABF3协同正调控AHG1、AHG3、HAI1及HAI2等PP2Cs基因表达(Yoshida et al., 2010).此外, 越来越多的研究证明ET信号通路参与拟南芥盐胁迫反应.进一步分析表明, ESE1 (ethylene and salt-inducible ERF1)在EIN3/EIL1下游调节ET响应基因.EIN3 (ethylene insensitive 3)可与ESE1启动子结合形成EIN3-ESE1转录复合体, 继而调控RD29A和COR15A等基因的表达(Zhang et al., 2011).磷酸化、泛素化、甲基化和乙酰化等组蛋白修饰方式通过调控染色质开放或封闭的空间状态, 激活或抑制AP2/ERF转录(Pfluger and Wagner, 2007).组蛋白去乙酰化酶(histone deacetylase, HDAC)和聚乙二醇(PEG)促进DREB1基因启动子区乙酰化, 增强DREB1基因的表达(Zhang et al., 2018).非生物胁迫下, 组蛋白变体和组蛋白修饰状态也可发生改变, 并可遗传到下一代(Asensii-Fabado et al., 2017).miRNA介导的RNA沉默和翻译抑制在AP2/ERF表达调控中也发挥重要作用.研究发现miRNA172靶向拟南芥AP2mRNA并抑制其翻译, 而miRNA156和miRNA838两者也具有类似调节AP2/ERF的功能(Kavas et al., 2015).此外, 在拟南芥、水稻及玉米中还发现通过可变剪切产生的OsDREB2A/2B、ZmDREB2A、WDREB2以及HvDRF1等多种AP2/-ERF功能异构体(Matsukura et al., 2010). ...
SUB1A-mediated submergence tolerance response in rice involves differential regulation of the brassinosteroid pathway 2 2013
... BZR1可结合CBF1和CBF2的启动子区并促进两者在冷胁迫下的表达.bin2-3、bil1、bil2缺失突变体在持续的冷胁迫下可以诱导去磷酸化BZR1的积累, BZR1通过调控WKRY6、PYL6、SOC1、JMT和SAG21等不依赖CBF途径的COR基因而增强拟南芥的耐冷性(Li et al., 2017).研究发现BR可与干旱诱导相关转录因子RD26互作, 拮抗干旱响应基因的表达, 进而负调控植物的抗旱性(Ye et al., 2017).BR与ABA信号途径中的受体、转录因子之间也产生拮抗效应, 进而调节胁迫下植物体的生长平衡(Nolan et al., 2017).目前已从BES1和BZR1靶基因中鉴定出一些AP2/ERF类转录因子, 暗示AP2/ERF具有整合BR信号通路与非生物胁迫响应的潜在功能(Sun et al., 2010; Yu et al., 2011).ERF72可能是研究BR信号通路与胁迫响应交叉互作的候选基因.在拟南芥中, ERF72/RAP2.3拮抗BZR1和ARF6 (auxin responsive factor 6), 抑制下胚轴伸长, 此过程中BR信号途径如何与胁迫响应调控网络协同互作发挥作用尚不清楚(Liu et al., 2018b).研究表明, SUB1A参与调节GA和BR的交叉网络互作(Schmitz et al., 2013).GA和BR途径通过DELLA蛋白、BZR1等转录因子协同调控植物的生长(Oh et al., 2012).在植物淹水过程中SUB1A差异调节BR合成相关基因的表达, 激活BR生物合成和信号转导, 诱导GA降解关键基因GA2ox7的表达, 进而控制水稻植株体内的GA水平, 促进DELLA蛋白的积累(Schmitz et al., 2013).因此, 推测AP2/ERF可能通过不同的机制参与调控BR信号通路. ...
... 的表达, 进而控制水稻植株体内的GA水平, 促进DELLA蛋白的积累(Schmitz et al., 2013).因此, 推测AP2/ERF可能通过不同的机制参与调控BR信号通路. ...
Molecular regulation of CBF signaling in cold acclimation 2 2018
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
... ; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
Ethylene signaling negatively regulates freezing tolerance by repressing expression of CBF and type-A ARR genes in Arabidopsis 2 2012
... ERF亚家族转录因子是ET介导的胁迫应答信号通路下游的关键调控因子(Licausi et al., 2013; Gibbs et al., 2015; Kazan, 2015; Müller and Munné-Bosch, 2015).冷胁迫下, 植物体内的ET主要起负调控作用, 过量ET或1-氨基环丙烷-1-羧酸降低植物的耐冷性, 而ET生物合成抑制剂氨基乙氧基甘氨酸或感知拮抗剂Ag+具有相反的作用(Shi et al., 2012).etr1-1、ein4-1、ein2-5、ein3-1和ein3eil突变体表现出更强的耐冷性, 但组成型ET响应etr1-1和ein3过表达植株表现出耐冷性减弱; 冷胁迫诱导ETR1、EIN4、EBF1及EBF2快速上调表达, 而EIN2和EIL1下调表达.进一步分析表明, EIN3通过与启动子区特定元件结合, 抑制ARR5、ARR7、ARR15和CBF转录, ARRs作为冷信号通路的关键节点基因, 在ET和CTK介导的胁迫响应网络中起着关键作用, 过表达ARRs使植株的耐冷能力增强(Shi et al., 2012).此外, EIN3可激活ERF1和ESEs, 进而参与调控下游胁迫相关基因, 提高植物的耐盐性(Kazan, 2015; Tao et al., 2015).35S:AtERF4-GFP转基因植株中ABI2、RD29B和RAP18的表达受抑制, 进一步说明AtERF4是ET和ABA信号途径的负调控因子(Yang et al., 2011). ...
... 使植株的耐冷能力增强(Shi et al., 2012).此外, EIN3可激活ERF1和ESEs, 进而参与调控下游胁迫相关基因, 提高植物的耐盐性(Kazan, 2015; Tao et al., 2015).35S:AtERF4-GFP转基因植株中ABI2、RD29B和RAP18的表达受抑制, 进一步说明AtERF4是ET和ABA信号途径的负调控因子(Yang et al., 2011). ...
Role of an Arabidopsis AP2/EREBP- type transcriptional repressor in abscisic acid and drought stress responses 2 2005
... 许多研究表明, AP2/ERF转录因子可直接与靶基因启动子结合, 激活或抑制下游靶基因的表达, 其N端DNA结合域和C端转录激活域在胁迫相关靶基因的转录激活中起重要作用(Nakano et al., 2006).进一步分析表明, AP2/ERF转录因子中含有EAR基序(LxLxL或DLNxxP)、TLLLFR基序和BRD结构域(B3 repression domain, 保守序列为R/KLFGV)等多个转录抑制结构域(Ikeda and Ohme-Takagi, 2009; Kagale and Rozwadowski, 2011; Deepika et al., 2016).含EAR基序的转录因子是一种重要的抑制子, 在植物非生物胁迫应答基因的诱导表达中起负调控作用.Tiwari等(2012)从拟南芥ERF和AP2亚族中发现了一种转录激活关键元件EDLL, EDLL可与EAR基序(ERF-associated amphiphilic repression)发生互作, 拮抗后者介导的转录抑制作用.而不同的AP2/ ERF转录因子也可协同招募TPL (topless)和TPR (topless-related)等转录共抑制子(Causier et al., 2012)或组蛋白修饰因子, 进而抑制下游靶基因的表达(Song et al., 2005; Song and Galbraith, 2006).Song等(2005)发现AtERF7特异性地结合GCC-box, 与抑制因子ATSIN3及组蛋白去乙酰化酶19 (histone deacetylase 19, HDA19)发生互作并形成转录因子复合物, 参与干旱胁迫和ABA响应调控.AtERF7过表达株系保卫细胞对ABA的敏感性减弱, 水分散失增加; 反之, AtERF7 RNA干扰系对ABA的敏感性增强.类似的, ERF3能与组蛋白去乙酰化酶复合物亚基SAP18 (SIN3 associated polypeptide P18)互作, 共同招募HDA19形成复合体, 进而抑制相关基因的表达(Song and Galbraith, 2006).BRD结构域被认为是RAV1和RAV2转录因子行使抑制功能的关键元件, 其保守序列中第1位氨基酸残基多为疏水性亮氨酸、缬氨酸和蛋氨酸残基, 突变导致功能丧失(Ikeda and Ohme-Takagi, 2009).研究显示, 与AP2/ERFs协同调控的基因同样可被AP2/ERFs靶基因富集, 分析此类基因的同源基序有助于识别转录因子的假定靶基因并预测其生物学功能(Franco-Zorrillaetal et al., 2014). ...
AtSAP18, an orthologue of human SAP18, is involved in the regulation of salt stress and mediates transcriptional repression in Arabidopsis 2 2006
... 许多研究表明, AP2/ERF转录因子可直接与靶基因启动子结合, 激活或抑制下游靶基因的表达, 其N端DNA结合域和C端转录激活域在胁迫相关靶基因的转录激活中起重要作用(Nakano et al., 2006).进一步分析表明, AP2/ERF转录因子中含有EAR基序(LxLxL或DLNxxP)、TLLLFR基序和BRD结构域(B3 repression domain, 保守序列为R/KLFGV)等多个转录抑制结构域(Ikeda and Ohme-Takagi, 2009; Kagale and Rozwadowski, 2011; Deepika et al., 2016).含EAR基序的转录因子是一种重要的抑制子, 在植物非生物胁迫应答基因的诱导表达中起负调控作用.Tiwari等(2012)从拟南芥ERF和AP2亚族中发现了一种转录激活关键元件EDLL, EDLL可与EAR基序(ERF-associated amphiphilic repression)发生互作, 拮抗后者介导的转录抑制作用.而不同的AP2/ ERF转录因子也可协同招募TPL (topless)和TPR (topless-related)等转录共抑制子(Causier et al., 2012)或组蛋白修饰因子, 进而抑制下游靶基因的表达(Song et al., 2005; Song and Galbraith, 2006).Song等(2005)发现AtERF7特异性地结合GCC-box, 与抑制因子ATSIN3及组蛋白去乙酰化酶19 (histone deacetylase 19, HDA19)发生互作并形成转录因子复合物, 参与干旱胁迫和ABA响应调控.AtERF7过表达株系保卫细胞对ABA的敏感性减弱, 水分散失增加; 反之, AtERF7 RNA干扰系对ABA的敏感性增强.类似的, ERF3能与组蛋白去乙酰化酶复合物亚基SAP18 (SIN3 associated polypeptide P18)互作, 共同招募HDA19形成复合体, 进而抑制相关基因的表达(Song and Galbraith, 2006).BRD结构域被认为是RAV1和RAV2转录因子行使抑制功能的关键元件, 其保守序列中第1位氨基酸残基多为疏水性亮氨酸、缬氨酸和蛋氨酸残基, 突变导致功能丧失(Ikeda and Ohme-Takagi, 2009).研究显示, 与AP2/ERFs协同调控的基因同样可被AP2/ERFs靶基因富集, 分析此类基因的同源基序有助于识别转录因子的假定靶基因并预测其生物学功能(Franco-Zorrillaetal et al., 2014). ...
... RNA干扰系对ABA的敏感性增强.类似的, ERF3能与组蛋白去乙酰化酶复合物亚基SAP18 (SIN3 associated polypeptide P18)互作, 共同招募HDA19形成复合体, 进而抑制相关基因的表达(Song and Galbraith, 2006).BRD结构域被认为是RAV1和RAV2转录因子行使抑制功能的关键元件, 其保守序列中第1位氨基酸残基多为疏水性亮氨酸、缬氨酸和蛋氨酸残基, 突变导致功能丧失(Ikeda and Ohme-Takagi, 2009).研究显示, 与AP2/ERFs协同调控的基因同样可被AP2/ERFs靶基因富集, 分析此类基因的同源基序有助于识别转录因子的假定靶基因并预测其生物学功能(Franco-Zorrillaetal et al., 2014). ...
Integration of brassinosteroid signal transduction with the transcription network for plant growth regulation in Arabidopsis 1 2010
... BZR1可结合CBF1和CBF2的启动子区并促进两者在冷胁迫下的表达.bin2-3、bil1、bil2缺失突变体在持续的冷胁迫下可以诱导去磷酸化BZR1的积累, BZR1通过调控WKRY6、PYL6、SOC1、JMT和SAG21等不依赖CBF途径的COR基因而增强拟南芥的耐冷性(Li et al., 2017).研究发现BR可与干旱诱导相关转录因子RD26互作, 拮抗干旱响应基因的表达, 进而负调控植物的抗旱性(Ye et al., 2017).BR与ABA信号途径中的受体、转录因子之间也产生拮抗效应, 进而调节胁迫下植物体的生长平衡(Nolan et al., 2017).目前已从BES1和BZR1靶基因中鉴定出一些AP2/ERF类转录因子, 暗示AP2/ERF具有整合BR信号通路与非生物胁迫响应的潜在功能(Sun et al., 2010; Yu et al., 2011).ERF72可能是研究BR信号通路与胁迫响应交叉互作的候选基因.在拟南芥中, ERF72/RAP2.3拮抗BZR1和ARF6 (auxin responsive factor 6), 抑制下胚轴伸长, 此过程中BR信号途径如何与胁迫响应调控网络协同互作发挥作用尚不清楚(Liu et al., 2018b).研究表明, SUB1A参与调节GA和BR的交叉网络互作(Schmitz et al., 2013).GA和BR途径通过DELLA蛋白、BZR1等转录因子协同调控植物的生长(Oh et al., 2012).在植物淹水过程中SUB1A差异调节BR合成相关基因的表达, 激活BR生物合成和信号转导, 诱导GA降解关键基因GA2ox7的表达, 进而控制水稻植株体内的GA水平, 促进DELLA蛋白的积累(Schmitz et al., 2013).因此, 推测AP2/ERF可能通过不同的机制参与调控BR信号通路. ...
Overexpression of the lotus corniculatus soloist gene LcAP2/ERF107 enhances tolerance to salt stress 1 2016
... AP2/ERF转录因子具有独特的结构特征, 其由DNA结合域(DNA-binding domain)、转录调控域(transcription regulation domain)、寡聚化位点(oligomerization site)和核定位信号(nuclear localization signal, NLS) 4个主要功能区组成, 并以高度保守的AP2/ ERF结合域而得名(Sakuma et al., 2002).通常情况下, AP2/ERF转录因子至少含有1个AP2结构域的DNA结合区, 该结构域包含60-70个氨基酸残基, 按照3个β折叠和1个α螺旋方式形成典型的三维结构(Nakano et al., 2006; Abiri et al., 2017).根据包含的AP2/ERF结构域数量和结合序列特点, 可将AP2/ ERF家族分为AP2 (APETALA2)、RAV (related to ABI3/VP1)、脱水反应元件结合蛋白(dehydration- responsive element binding protein, DREB) (A1-A6亚组)、乙烯应答因子(ethylene-responsive factor, ERF) (V-X亚组)和Soloist五个亚族(Nakano et al., 2006; Licausi et al., 2013).其中, 对AP2、DREB和ERF的研究较为广泛和深入, 而有关Soloist亚族的研究报道极少, 已知其核苷酸序列在多数植物中高度保守(Sun et al., 2016). ...
The role of ethylene in plants under salinity stress 2 2015
... AP2/ERF转录因子在参与调控植物应对冷、干旱、高温、高盐及缺氧等多种非生物胁迫过程中具有重要功能(Licausi et al., 2013).其中, DREB和ERF亚家族成员在植物非生物逆境应答中起重要调控作用.研究表明其参与调控的植物环境胁迫应答信号网络比较复杂.胁迫条件下, 部分AP2/ERFs可被快速持续地诱导表达, 而其它进程的响应较为缓慢, 说明可能存在依赖或不依赖于激素信号途径2种胁迫响应模式, 且两者之间存在一定的交叉互作(Van den Broeck et al., 2017).前人的研究也证明, AP2/ERFs广泛参与调控ABA、ET、GA、细胞分裂素(cytokinin, CTK)和油菜素内酯(brassinolide, BR)等植物激素介导的胁迫响应(Colebrook et al., 2014; Kazan et al., 2015; Tao et al., 2015; Sah et al., 2016; Nolan et al., 2017) (图1). ...
... ERF亚家族转录因子是ET介导的胁迫应答信号通路下游的关键调控因子(Licausi et al., 2013; Gibbs et al., 2015; Kazan, 2015; Müller and Munné-Bosch, 2015).冷胁迫下, 植物体内的ET主要起负调控作用, 过量ET或1-氨基环丙烷-1-羧酸降低植物的耐冷性, 而ET生物合成抑制剂氨基乙氧基甘氨酸或感知拮抗剂Ag+具有相反的作用(Shi et al., 2012).etr1-1、ein4-1、ein2-5、ein3-1和ein3eil突变体表现出更强的耐冷性, 但组成型ET响应etr1-1和ein3过表达植株表现出耐冷性减弱; 冷胁迫诱导ETR1、EIN4、EBF1及EBF2快速上调表达, 而EIN2和EIL1下调表达.进一步分析表明, EIN3通过与启动子区特定元件结合, 抑制ARR5、ARR7、ARR15和CBF转录, ARRs作为冷信号通路的关键节点基因, 在ET和CTK介导的胁迫响应网络中起着关键作用, 过表达ARRs使植株的耐冷能力增强(Shi et al., 2012).此外, EIN3可激活ERF1和ESEs, 进而参与调控下游胁迫相关基因, 提高植物的耐盐性(Kazan, 2015; Tao et al., 2015).35S:AtERF4-GFP转基因植株中ABI2、RD29B和RAP18的表达受抑制, 进一步说明AtERF4是ET和ABA信号途径的负调控因子(Yang et al., 2011). ...
The EDLL motif: a potent plant transcriptional activation domain from AP2/ERF transcription factors 1 2012
... 许多研究表明, AP2/ERF转录因子可直接与靶基因启动子结合, 激活或抑制下游靶基因的表达, 其N端DNA结合域和C端转录激活域在胁迫相关靶基因的转录激活中起重要作用(Nakano et al., 2006).进一步分析表明, AP2/ERF转录因子中含有EAR基序(LxLxL或DLNxxP)、TLLLFR基序和BRD结构域(B3 repression domain, 保守序列为R/KLFGV)等多个转录抑制结构域(Ikeda and Ohme-Takagi, 2009; Kagale and Rozwadowski, 2011; Deepika et al., 2016).含EAR基序的转录因子是一种重要的抑制子, 在植物非生物胁迫应答基因的诱导表达中起负调控作用.Tiwari等(2012)从拟南芥ERF和AP2亚族中发现了一种转录激活关键元件EDLL, EDLL可与EAR基序(ERF-associated amphiphilic repression)发生互作, 拮抗后者介导的转录抑制作用.而不同的AP2/ ERF转录因子也可协同招募TPL (topless)和TPR (topless-related)等转录共抑制子(Causier et al., 2012)或组蛋白修饰因子, 进而抑制下游靶基因的表达(Song et al., 2005; Song and Galbraith, 2006).Song等(2005)发现AtERF7特异性地结合GCC-box, 与抑制因子ATSIN3及组蛋白去乙酰化酶19 (histone deacetylase 19, HDA19)发生互作并形成转录因子复合物, 参与干旱胁迫和ABA响应调控.AtERF7过表达株系保卫细胞对ABA的敏感性减弱, 水分散失增加; 反之, AtERF7 RNA干扰系对ABA的敏感性增强.类似的, ERF3能与组蛋白去乙酰化酶复合物亚基SAP18 (SIN3 associated polypeptide P18)互作, 共同招募HDA19形成复合体, 进而抑制相关基因的表达(Song and Galbraith, 2006).BRD结构域被认为是RAV1和RAV2转录因子行使抑制功能的关键元件, 其保守序列中第1位氨基酸残基多为疏水性亮氨酸、缬氨酸和蛋氨酸残基, 突变导致功能丧失(Ikeda and Ohme-Takagi, 2009).研究显示, 与AP2/ERFs协同调控的基因同样可被AP2/ERFs靶基因富集, 分析此类基因的同源基序有助于识别转录因子的假定靶基因并预测其生物学功能(Franco-Zorrillaetal et al., 2014). ...
DEAR1, a transcriptional repressor of DREB protein that mediates plant defense and freezing stress responses in Arabidopsis 2 2009
... 部分A5亚组DREB类转录因子则对上述调控网络具有负反馈调节作用.DEAR1 (DREB and EAR motif protein 1)可能既在CBF上游作用, 又与RAP2.1在CBF下游扮演冷胁迫响应的调节子, 调节RD29A/COR78、COR15A和KIN1等冷诱导相关基因的表达, 负调控植物的耐冷和耐旱性(Fowler and Thomashow, 2002; Tsutsui et al., 2009; Dong and Liu, 2010).DEAR1过表达抑制冷诱导CBF的表达, 进而降低植物的耐冷能力(Tsutsui et al., 2009).低温胁迫或组成型表达CBF可诱导RAP2.1的表达, CBF2比RAP2.1对低温更为敏感(Dong and Liu, 2010).在CBF调控通路中, DEAR1和RAP2.1如何整合还需深入研究.RAP2.1是否与其它DEAR转录因子共同调节植物非生物胁迫响应仍不清楚.研究发现DREB-A5亚组有6种含EAR基序的转录抑制子可与DRE元件结合, 进而调控非生物胁迫响应基因的表达(Nakano et al., 2006). ...
... 的表达, 进而降低植物的耐冷能力(Tsutsui et al., 2009).低温胁迫或组成型表达CBF可诱导RAP2.1的表达, CBF2比RAP2.1对低温更为敏感(Dong and Liu, 2010).在CBF调控通路中, DEAR1和RAP2.1如何整合还需深入研究.RAP2.1是否与其它DEAR转录因子共同调节植物非生物胁迫响应仍不清楚.研究发现DREB-A5亚组有6种含EAR基序的转录抑制子可与DRE元件结合, 进而调控非生物胁迫响应基因的表达(Nakano et al., 2006). ...
From network to phenotype: the dynamic wiring of an Arabidopsis transcriptional network induced by osmotic stress 1 2017
... AP2/ERF转录因子在参与调控植物应对冷、干旱、高温、高盐及缺氧等多种非生物胁迫过程中具有重要功能(Licausi et al., 2013).其中, DREB和ERF亚家族成员在植物非生物逆境应答中起重要调控作用.研究表明其参与调控的植物环境胁迫应答信号网络比较复杂.胁迫条件下, 部分AP2/ERFs可被快速持续地诱导表达, 而其它进程的响应较为缓慢, 说明可能存在依赖或不依赖于激素信号途径2种胁迫响应模式, 且两者之间存在一定的交叉互作(Van den Broeck et al., 2017).前人的研究也证明, AP2/ERFs广泛参与调控ABA、ET、GA、细胞分裂素(cytokinin, CTK)和油菜素内酯(brassinolide, BR)等植物激素介导的胁迫响应(Colebrook et al., 2014; Kazan et al., 2015; Tao et al., 2015; Sah et al., 2016; Nolan et al., 2017) (图1). ...
Plant hormone- mediated regulation of stress responses 1 2016
... 植物在生长发育过程中经常受到生物或非生物胁迫的影响, 由此, 植物逐渐形成了复杂的调节机制感知胁迫信号, 对外部环境变化做出快速响应(郭倩倩和周文彬, 2019).脱落酸(abscisic acid, ABA)、乙烯(ethylene, ET)及赤霉素(gibberellin, GA)等激素作为信号分子能够调节植物的生长发育、形态建成以及抗逆反应等生物学过程.该过程也受转录因子(transcription factor, TF)调控, 即转录因子通过依赖或不依赖于激素介导的信号通路参与调控植物对干旱、低温、缺氧、高盐和淹水等非生物胁迫的响应, 进而影响植物的逆境耐受能力(Gibbs et al., 2015; Verma et al., 2016; Li et al., 2018).AP2/ERF (APETALA2/eth-ylene responsive factor)是植物特有的转录因子家族之一, 已在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、高粱(Sorghum bicolor)、大白菜(Brassica pekinensis)、毛竹(Phyllostachys edulis)、玉米(Zea mays)、大麦(Hordeum vulgare)和小麦(Triticum aestivum)等物种中被成功分离鉴定(Xu and Chua, 2011; Chandler, 2018).通过突变体实验, 科学家发现了许多与AP2/ERF相关的非生物胁迫或激素信号应答基因.目前, AP2/ERF已成为研究非生物胁迫和激素互作的热点候选基因.本文从AP2/ERF转录因子的结构特征、转录调控、翻译后修饰、结合位点、协同互作蛋白以及其参与调控依赖或不依赖激素信号途径的非生物胁迫响应进行综述, 为进一步阐明AP2/ERF转录因子在非生物胁迫调控网络中的作用提供理论依据. ...
Dual role of BKI1 and 14-3-3s in brassinosteroid signaling to link receptor with transcription factors 2 2011
... 油菜素内酯(BR)在植物细胞伸长、叶片发育、花粉管生长、木质部分化、衰老、光形态发生以及应激反应过程中起重要作用(Ye et al., 2017).BR信号始于细胞膜上受体激酶(brassinosteroid insensitive, BRI), BKI1 (BRI kinase inhibitor 1)是BRI1的关键负调控蛋白.无BR时, BKI1和14-3-3蛋白分别抑制BR的受体BRI1和转录因子BES1 (bri1-ems-suppressor 1) (Wang et al., 2011).而当BR被BRI1感知后, 即促使BKI1发生磷酸化, 激活BRI1和BES1以调控负调节因子BIN2 (brassinosteroid insensitive 2)的表达, 诱导BES1和BZR1 (brassinazole-resistant 1)积累并与转录因子互作, 影响参与植物生长和胁迫反应的BR应答基因的转录水平(Wang et al., 2011; Guo et al., 2013). ...
... ).而当BR被BRI1感知后, 即促使BKI1发生磷酸化, 激活BRI1和BES1以调控负调节因子BIN2 (brassinosteroid insensitive 2)的表达, 诱导BES1和BZR1 (brassinazole-resistant 1)积累并与转录因子互作, 影响参与植物生长和胁迫反应的BR应答基因的转录水平(Wang et al., 2011; Guo et al., 2013). ...
Three grape CBF/DREB1 genes respond to low temperature, drought and abscisic acid 1 2006
... DREB-A2亚组转录因子DREB2主要参与调节植物的抗旱和耐热性(Mizoi et al., 2012).干旱和高温可诱导DREB2基因的表达, 进而正向调控下游干旱或热胁迫响应基因(Maruyama et al., 2009).研究表明DREB-A4家族成员HRD (hardy)和DREB-A6家族成员(ERF53、RAP2.4和TG/RAP2.4a)也在干旱和耐盐性调控中发挥关键作用(Karaba et al., 2007; Lin et al., 2008; Chen et al., 2012; Zhu et al., 2014).过表达HRD能显著增强拟南芥和水稻植株的耐旱和耐盐性(Karaba et al., 2007).同样, 过表达TG (translucent green)导致植株叶片玻璃化, 叶片细胞含水量增加(Zhu et al., 2014).TG也参与调控抗坏血酸过氧化物酶基因APX (ascorbate peroxidase)的表达, 抵御活性氧(reactive oxygen species, ROS)引起的光氧化胁迫(Rudnik et al., 2017).除拟南芥外, 人们对水稻、玉米、番茄(Lycopersicon esculentum)、葡萄(Vitis vinifera)等作物的DREB1和DREB2转录因子也进行了广泛的研究(Xiao et al., 2006; Matsukura et al., 2010; Dou et al., 2014). ...
Natural variation in OsLG3 increases drought tolerance in rice by inducing ROS scavenging 1 2018
... RbohD (respiratory burst oxidase homolog D)是一种NADPH氧化酶, 催化活性氧产生(Yao et al., 2017).ROS是细胞生化反应过程中产生的毒性物质, 可作为信号分子触发胁迫响应和信号转导, RAPs通过依赖于RbohD的途径调节非生物胁迫(Qi et al., 2018).erf74和erf74/erf75突变体中, ROS的产生和RbohD基因的表达均受抑制, ERF74和ERF75通过依赖于RbohD的ROS激活途径协同调控缺氧响应.其中, ERF74可结合RbohD基因的启动子并激活其表达, ERF74过表达促进后期ROS清除酶相关基因的表达, 说明在胁迫响应过程中ERF74作为分子开关控制RbohD依赖的ROS迸发(Yao et al., 2017).研究发现水稻中也存在类似的调控机制, OsLG3通过诱导活性氧清除相关基因APX1、APX4、APX8、CATB和POD1的表达进而正调控水稻的耐旱性(Xiong et al., 2018). ...
Processing bodies and plant development 1 2011
... 植物在生长发育过程中经常受到生物或非生物胁迫的影响, 由此, 植物逐渐形成了复杂的调节机制感知胁迫信号, 对外部环境变化做出快速响应(郭倩倩和周文彬, 2019).脱落酸(abscisic acid, ABA)、乙烯(ethylene, ET)及赤霉素(gibberellin, GA)等激素作为信号分子能够调节植物的生长发育、形态建成以及抗逆反应等生物学过程.该过程也受转录因子(transcription factor, TF)调控, 即转录因子通过依赖或不依赖于激素介导的信号通路参与调控植物对干旱、低温、缺氧、高盐和淹水等非生物胁迫的响应, 进而影响植物的逆境耐受能力(Gibbs et al., 2015; Verma et al., 2016; Li et al., 2018).AP2/ERF (APETALA2/eth-ylene responsive factor)是植物特有的转录因子家族之一, 已在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、高粱(Sorghum bicolor)、大白菜(Brassica pekinensis)、毛竹(Phyllostachys edulis)、玉米(Zea mays)、大麦(Hordeum vulgare)和小麦(Triticum aestivum)等物种中被成功分离鉴定(Xu and Chua, 2011; Chandler, 2018).通过突变体实验, 科学家发现了许多与AP2/ERF相关的非生物胁迫或激素信号应答基因.目前, AP2/ERF已成为研究非生物胁迫和激素互作的热点候选基因.本文从AP2/ERF转录因子的结构特征、转录调控、翻译后修饰、结合位点、协同互作蛋白以及其参与调控依赖或不依赖激素信号途径的非生物胁迫响应进行综述, 为进一步阐明AP2/ERF转录因子在非生物胁迫调控网络中的作用提供理论依据. ...
Sub1A is an ethylene-response-factor- like gene that confers submergence tolerance to rice 1 2006
... ERF亚族成员在低氧和淹水胁迫响应过程中也发挥重要作用(Bui et al., 2015; Gibbs et al., 2015).SUB1A (submergence 1A)是ERF亚族B2亚组成员之一, Sub1A-1超表达的同时促进乙醇脱氢酶基因(alcohol dehydrogenase 1, Adh1)上调表达, 抑制Sub1C的转录水平, 说明Sub1A-1是耐淹性的主要调节因子, 在淹水胁迫应答调控中起关键作用(Xu et al., 2006).淹水状态下, 低水平海藻糖-6-磷酸合酶(trehalose-6-phosphate, T6P)与CIPK15 (CBL-inter-acting protein kinase 15)协同激活SnRK1, SnRK1介导SUB1A抑制幼苗胚轴生长和呼吸相关基因的表达, 直接或间接调控碳同化过程.而SK1/SK2 (snorkel1/2)激活参与叶柄、茎和导管发育相关基因, 促进植株快速生长(Hattori et al., 2009; Locke et al., 2018).缺氧胁迫下, 拟南芥ERF-VIIs可以通过氧依赖的N端规则通路降解(Gibbs et al., 2015).RING (really interesting gene)类蛋白SINAT2可通过其N端保守的C3H4型RING结构域调控RAP2.12, 沉默SINAT1/2可上调RAP2.12的表达水平(Papdi et al., 2015).此外, ERF71/HRE2、ERF72/RAP2.3、ERF73/HRE1、ERF74/RAP2.12及ERF75/RAP2.2d也受缺氧胁迫诱导, 进而正向调控糖代谢和激素信号相关的缺氧响应基因, 提高植株的缺氧耐受能力.过表达RAP2.2、RAP2.3和RAP2.12可提高植株在低氧和渗透胁迫下的存活率, 而rap2.12-2/rap2.3-1双突变体对淹水和渗透胁迫表现出较高的敏感性(Yao et al., 2017). ...
Functionality of soybean CBF/DREB1 transcription factors 1 2016
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
The AP2/ERF transcription factor AtERF73/HRE1 modulates ethylene responses during hypoxia in Arabidopsis 2 2011
... ERF亚家族转录因子是ET介导的胁迫应答信号通路下游的关键调控因子(Licausi et al., 2013; Gibbs et al., 2015; Kazan, 2015; Müller and Munné-Bosch, 2015).冷胁迫下, 植物体内的ET主要起负调控作用, 过量ET或1-氨基环丙烷-1-羧酸降低植物的耐冷性, 而ET生物合成抑制剂氨基乙氧基甘氨酸或感知拮抗剂Ag+具有相反的作用(Shi et al., 2012).etr1-1、ein4-1、ein2-5、ein3-1和ein3eil突变体表现出更强的耐冷性, 但组成型ET响应etr1-1和ein3过表达植株表现出耐冷性减弱; 冷胁迫诱导ETR1、EIN4、EBF1及EBF2快速上调表达, 而EIN2和EIL1下调表达.进一步分析表明, EIN3通过与启动子区特定元件结合, 抑制ARR5、ARR7、ARR15和CBF转录, ARRs作为冷信号通路的关键节点基因, 在ET和CTK介导的胁迫响应网络中起着关键作用, 过表达ARRs使植株的耐冷能力增强(Shi et al., 2012).此外, EIN3可激活ERF1和ESEs, 进而参与调控下游胁迫相关基因, 提高植物的耐盐性(Kazan, 2015; Tao et al., 2015).35S:AtERF4-GFP转基因植株中ABI2、RD29B和RAP18的表达受抑制, 进一步说明AtERF4是ET和ABA信号途径的负调控因子(Yang et al., 2011). ...
... 此外, ET促进RAP2.3转录因子定位于细胞核内, 通过依赖于RAP2.3的方式激发ORA59介导的ET响应过程(Kim et al., 2018).而ERF-VIIs通过ET不依赖途径参与调控缺氧响应.在ET不敏感突变体或生物合成抑制剂存在的情况下, 缺氧依然可以诱导ERF73/HRE1的表达, ERF-VIIs也可能通过反馈调节负调控ET信号(Hinz et al., 2010; Yang et al., 2011).研究发现, ET合酶ACS在RAP2.2过表达植株中表达量降低, 而在rap2.2-2敲除突变体中表达上调(Hinz et al., 2010).然而, 胁迫条件下ERF-VIIs如何通过负反馈机制调控ET生物合成有待进一步研究. ...
ETHYLENE RESPONSE FACTOR 74 (ERF74) plays an essential role in controlling a respiratory burst oxidase homolog D (RbohD)-dependent mechanism in response to different stresses in Arabidopsis 3 2017
... ERF亚族成员在低氧和淹水胁迫响应过程中也发挥重要作用(Bui et al., 2015; Gibbs et al., 2015).SUB1A (submergence 1A)是ERF亚族B2亚组成员之一, Sub1A-1超表达的同时促进乙醇脱氢酶基因(alcohol dehydrogenase 1, Adh1)上调表达, 抑制Sub1C的转录水平, 说明Sub1A-1是耐淹性的主要调节因子, 在淹水胁迫应答调控中起关键作用(Xu et al., 2006).淹水状态下, 低水平海藻糖-6-磷酸合酶(trehalose-6-phosphate, T6P)与CIPK15 (CBL-inter-acting protein kinase 15)协同激活SnRK1, SnRK1介导SUB1A抑制幼苗胚轴生长和呼吸相关基因的表达, 直接或间接调控碳同化过程.而SK1/SK2 (snorkel1/2)激活参与叶柄、茎和导管发育相关基因, 促进植株快速生长(Hattori et al., 2009; Locke et al., 2018).缺氧胁迫下, 拟南芥ERF-VIIs可以通过氧依赖的N端规则通路降解(Gibbs et al., 2015).RING (really interesting gene)类蛋白SINAT2可通过其N端保守的C3H4型RING结构域调控RAP2.12, 沉默SINAT1/2可上调RAP2.12的表达水平(Papdi et al., 2015).此外, ERF71/HRE2、ERF72/RAP2.3、ERF73/HRE1、ERF74/RAP2.12及ERF75/RAP2.2d也受缺氧胁迫诱导, 进而正向调控糖代谢和激素信号相关的缺氧响应基因, 提高植株的缺氧耐受能力.过表达RAP2.2、RAP2.3和RAP2.12可提高植株在低氧和渗透胁迫下的存活率, 而rap2.12-2/rap2.3-1双突变体对淹水和渗透胁迫表现出较高的敏感性(Yao et al., 2017). ...
... RbohD (respiratory burst oxidase homolog D)是一种NADPH氧化酶, 催化活性氧产生(Yao et al., 2017).ROS是细胞生化反应过程中产生的毒性物质, 可作为信号分子触发胁迫响应和信号转导, RAPs通过依赖于RbohD的途径调节非生物胁迫(Qi et al., 2018).erf74和erf74/erf75突变体中, ROS的产生和RbohD基因的表达均受抑制, ERF74和ERF75通过依赖于RbohD的ROS激活途径协同调控缺氧响应.其中, ERF74可结合RbohD基因的启动子并激活其表达, ERF74过表达促进后期ROS清除酶相关基因的表达, 说明在胁迫响应过程中ERF74作为分子开关控制RbohD依赖的ROS迸发(Yao et al., 2017).研究发现水稻中也存在类似的调控机制, OsLG3通过诱导活性氧清除相关基因APX1、APX4、APX8、CATB和POD1的表达进而正调控水稻的耐旱性(Xiong et al., 2018). ...
... 4过表达促进后期ROS清除酶相关基因的表达, 说明在胁迫响应过程中ERF74作为分子开关控制RbohD依赖的ROS迸发(Yao et al., 2017).研究发现水稻中也存在类似的调控机制, OsLG3通过诱导活性氧清除相关基因APX1、APX4、APX8、CATB和POD1的表达进而正调控水稻的耐旱性(Xiong et al., 2018). ...
RD26 mediates crosstalk between drought and brassinosteroid signaling pathways 2 2017
... 油菜素内酯(BR)在植物细胞伸长、叶片发育、花粉管生长、木质部分化、衰老、光形态发生以及应激反应过程中起重要作用(Ye et al., 2017).BR信号始于细胞膜上受体激酶(brassinosteroid insensitive, BRI), BKI1 (BRI kinase inhibitor 1)是BRI1的关键负调控蛋白.无BR时, BKI1和14-3-3蛋白分别抑制BR的受体BRI1和转录因子BES1 (bri1-ems-suppressor 1) (Wang et al., 2011).而当BR被BRI1感知后, 即促使BKI1发生磷酸化, 激活BRI1和BES1以调控负调节因子BIN2 (brassinosteroid insensitive 2)的表达, 诱导BES1和BZR1 (brassinazole-resistant 1)积累并与转录因子互作, 影响参与植物生长和胁迫反应的BR应答基因的转录水平(Wang et al., 2011; Guo et al., 2013). ...
... BZR1可结合CBF1和CBF2的启动子区并促进两者在冷胁迫下的表达.bin2-3、bil1、bil2缺失突变体在持续的冷胁迫下可以诱导去磷酸化BZR1的积累, BZR1通过调控WKRY6、PYL6、SOC1、JMT和SAG21等不依赖CBF途径的COR基因而增强拟南芥的耐冷性(Li et al., 2017).研究发现BR可与干旱诱导相关转录因子RD26互作, 拮抗干旱响应基因的表达, 进而负调控植物的抗旱性(Ye et al., 2017).BR与ABA信号途径中的受体、转录因子之间也产生拮抗效应, 进而调节胁迫下植物体的生长平衡(Nolan et al., 2017).目前已从BES1和BZR1靶基因中鉴定出一些AP2/ERF类转录因子, 暗示AP2/ERF具有整合BR信号通路与非生物胁迫响应的潜在功能(Sun et al., 2010; Yu et al., 2011).ERF72可能是研究BR信号通路与胁迫响应交叉互作的候选基因.在拟南芥中, ERF72/RAP2.3拮抗BZR1和ARF6 (auxin responsive factor 6), 抑制下胚轴伸长, 此过程中BR信号途径如何与胁迫响应调控网络协同互作发挥作用尚不清楚(Liu et al., 2018b).研究表明, SUB1A参与调节GA和BR的交叉网络互作(Schmitz et al., 2013).GA和BR途径通过DELLA蛋白、BZR1等转录因子协同调控植物的生长(Oh et al., 2012).在植物淹水过程中SUB1A差异调节BR合成相关基因的表达, 激活BR生物合成和信号转导, 诱导GA降解关键基因GA2ox7的表达, 进而控制水稻植株体内的GA水平, 促进DELLA蛋白的积累(Schmitz et al., 2013).因此, 推测AP2/ERF可能通过不同的机制参与调控BR信号通路. ...
AtDPG1 is involved in the salt stress response of Arabidopsis seedling through ABI4 1 2019
... DREB-A3亚组成员ABI4 (ABA insensitive 4)是ABA信号通路的重要组分.植株受到胁迫后, 体内积累ABA和ROS, ABI4可被ABA、ROS及糖质体等多种信号通路靶向调控.CCAAT结合因子A (CCAAT binding factor A, CBFA)是四吡咯化合物HAP (heme activator protein)三聚体转录复合物的亚基, 当产生过量ROS时, ABI4可与其它转录因子竞争性地结合CCACGT元件, 抑制CBFA的表达, 进而调控质体反向信号通路中相关基因的表达(Zhang et al., 2013).拟南芥突变体实验表明, ABI4在YL1 (yellow leaf 1) (Li et al., 2016)和DPG1 (delayed pale-greening 1) (Yi et al., 2019)等基因参与盐胁迫响应的调控网络中也发挥重要作用.逆境胁迫下, ORA47在JA (jasmo-nate acid)和ABA信号转导途径中靶向调节ABA和JA诱导的下游应答基因, ERF18/ORA-47识别启动子顺式元件(NC/GT)CGNCCA (O-box), 激活PP2C家族ABI2转录因子的表达.ABI1则在ORA-47上游起作用, 从而形成ABI1-ORA47-ABI2正反馈基因表达回路, 调控ABA信号转导和干旱胁迫响应(Chen et al., 2016).此外, RAV1转录因子在ABA信号转导途径中也发挥关键作用, RAV1能与ABI3、ABI4和ABI5启动子结合, 其过表达抑制ABI3、ABI4和ABI5的转录活性, 导致根系生长发育过程中对ABA不敏感.同时, 蔗糖非酵解型蛋白激酶(SnRK2.2、SnRK2.3和SnRK2.6)在核内磷酸化RAV1, 降低ABI5对RAV1的依赖性, 并负调控RAV1对靶基因的转录抑制作用(Feng et al., 2014). ...
AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation 1 2010
... 正常环境下, AP2/ERF转录因子表达水平较低, 激素或逆境胁迫相关基因与AP2/ERF转录因子保守的顺式作用元件结合, 进而调控其表达(Owji et al., 2017).研究表明, 拟南芥DREB1A/CBF3、DREB1C/CBF2、DREB2C、DREB2G和DEAR3的启动子中包含较多的热激元件(heat shock element, HSE)、低温响应(low temperature responsive, LTR)、脱落酸响应(ABA responsive element, ABRE)等顺式作用元件(Sazegari et al., 2015).热激因子1 (heat shock factor 1, HSF1)、脱落酸响应结合蛋白(ABA responsive element binding protein/ABRE binding factors, AREB/ABF)可分别与DREB2A启动子中的热激应答和ABA响应元件结合, 调控其表达, 进而改变植物胁迫的耐受性(Kim et al., 2011; Liu et al., 2011).蛋白磷酸酶2C (type 2C protein phosphatase, PP2C)包含2个以上的ABRE元件, AREB1、AREB2和ABF3协同正调控AHG1、AHG3、HAI1及HAI2等PP2Cs基因表达(Yoshida et al., 2010).此外, 越来越多的研究证明ET信号通路参与拟南芥盐胁迫反应.进一步分析表明, ESE1 (ethylene and salt-inducible ERF1)在EIN3/EIL1下游调节ET响应基因.EIN3 (ethylene insensitive 3)可与ESE1启动子结合形成EIN3-ESE1转录复合体, 继而调控RD29A和COR15A等基因的表达(Zhang et al., 2011).磷酸化、泛素化、甲基化和乙酰化等组蛋白修饰方式通过调控染色质开放或封闭的空间状态, 激活或抑制AP2/ERF转录(Pfluger and Wagner, 2007).组蛋白去乙酰化酶(histone deacetylase, HDAC)和聚乙二醇(PEG)促进DREB1基因启动子区乙酰化, 增强DREB1基因的表达(Zhang et al., 2018).非生物胁迫下, 组蛋白变体和组蛋白修饰状态也可发生改变, 并可遗传到下一代(Asensii-Fabado et al., 2017).miRNA介导的RNA沉默和翻译抑制在AP2/ERF表达调控中也发挥重要作用.研究发现miRNA172靶向拟南芥AP2mRNA并抑制其翻译, 而miRNA156和miRNA838两者也具有类似调节AP2/ERF的功能(Kavas et al., 2015).此外, 在拟南芥、水稻及玉米中还发现通过可变剪切产生的OsDREB2A/2B、ZmDREB2A、WDREB2以及HvDRF1等多种AP2/-ERF功能异构体(Matsukura et al., 2010). ...
A brassinosteroid transcriptional network revealed by genome-wide identification of BESI target genes in Arabidopsis thaliana 1 2011
... BZR1可结合CBF1和CBF2的启动子区并促进两者在冷胁迫下的表达.bin2-3、bil1、bil2缺失突变体在持续的冷胁迫下可以诱导去磷酸化BZR1的积累, BZR1通过调控WKRY6、PYL6、SOC1、JMT和SAG21等不依赖CBF途径的COR基因而增强拟南芥的耐冷性(Li et al., 2017).研究发现BR可与干旱诱导相关转录因子RD26互作, 拮抗干旱响应基因的表达, 进而负调控植物的抗旱性(Ye et al., 2017).BR与ABA信号途径中的受体、转录因子之间也产生拮抗效应, 进而调节胁迫下植物体的生长平衡(Nolan et al., 2017).目前已从BES1和BZR1靶基因中鉴定出一些AP2/ERF类转录因子, 暗示AP2/ERF具有整合BR信号通路与非生物胁迫响应的潜在功能(Sun et al., 2010; Yu et al., 2011).ERF72可能是研究BR信号通路与胁迫响应交叉互作的候选基因.在拟南芥中, ERF72/RAP2.3拮抗BZR1和ARF6 (auxin responsive factor 6), 抑制下胚轴伸长, 此过程中BR信号途径如何与胁迫响应调控网络协同互作发挥作用尚不清楚(Liu et al., 2018b).研究表明, SUB1A参与调节GA和BR的交叉网络互作(Schmitz et al., 2013).GA和BR途径通过DELLA蛋白、BZR1等转录因子协同调控植物的生长(Oh et al., 2012).在植物淹水过程中SUB1A差异调节BR合成相关基因的表达, 激活BR生物合成和信号转导, 诱导GA降解关键基因GA2ox7的表达, 进而控制水稻植株体内的GA水平, 促进DELLA蛋白的积累(Schmitz et al., 2013).因此, 推测AP2/ERF可能通过不同的机制参与调控BR信号通路. ...
Histone deacetylases SRT1 and SRT2 interact with ENAP1 to mediate ethylene-induced transcriptional repression 1 2018
... 正常环境下, AP2/ERF转录因子表达水平较低, 激素或逆境胁迫相关基因与AP2/ERF转录因子保守的顺式作用元件结合, 进而调控其表达(Owji et al., 2017).研究表明, 拟南芥DREB1A/CBF3、DREB1C/CBF2、DREB2C、DREB2G和DEAR3的启动子中包含较多的热激元件(heat shock element, HSE)、低温响应(low temperature responsive, LTR)、脱落酸响应(ABA responsive element, ABRE)等顺式作用元件(Sazegari et al., 2015).热激因子1 (heat shock factor 1, HSF1)、脱落酸响应结合蛋白(ABA responsive element binding protein/ABRE binding factors, AREB/ABF)可分别与DREB2A启动子中的热激应答和ABA响应元件结合, 调控其表达, 进而改变植物胁迫的耐受性(Kim et al., 2011; Liu et al., 2011).蛋白磷酸酶2C (type 2C protein phosphatase, PP2C)包含2个以上的ABRE元件, AREB1、AREB2和ABF3协同正调控AHG1、AHG3、HAI1及HAI2等PP2Cs基因表达(Yoshida et al., 2010).此外, 越来越多的研究证明ET信号通路参与拟南芥盐胁迫反应.进一步分析表明, ESE1 (ethylene and salt-inducible ERF1)在EIN3/EIL1下游调节ET响应基因.EIN3 (ethylene insensitive 3)可与ESE1启动子结合形成EIN3-ESE1转录复合体, 继而调控RD29A和COR15A等基因的表达(Zhang et al., 2011).磷酸化、泛素化、甲基化和乙酰化等组蛋白修饰方式通过调控染色质开放或封闭的空间状态, 激活或抑制AP2/ERF转录(Pfluger and Wagner, 2007).组蛋白去乙酰化酶(histone deacetylase, HDAC)和聚乙二醇(PEG)促进DREB1基因启动子区乙酰化, 增强DREB1基因的表达(Zhang et al., 2018).非生物胁迫下, 组蛋白变体和组蛋白修饰状态也可发生改变, 并可遗传到下一代(Asensii-Fabado et al., 2017).miRNA介导的RNA沉默和翻译抑制在AP2/ERF表达调控中也发挥重要作用.研究发现miRNA172靶向拟南芥AP2mRNA并抑制其翻译, 而miRNA156和miRNA838两者也具有类似调节AP2/ERF的功能(Kavas et al., 2015).此外, 在拟南芥、水稻及玉米中还发现通过可变剪切产生的OsDREB2A/2B、ZmDREB2A、WDREB2以及HvDRF1等多种AP2/-ERF功能异构体(Matsukura et al., 2010). ...
Overexpression of the soybean GmERF3 gene, an AP2/ERF type transcription factor for increased tolerances to salt, drought, and diseases in transgenic tobacco 1 2009
... 赤霉素(GA)广泛分布在植物中并参与调控叶、芽以及合子发育等多种生物学过程.GA由GA20氧化酶(gibberellin 20-oxidases, GA20ox)和GA3氧化酶(GA3ox)等关键酶合成, 在GA2氧化酶(GA2ox)作用下被分解代谢(Rieu et al., 2008).DELLA蛋白作为GA信号转导途径中的主要负调控因子, 也是整合生长素、ABA、ET和JA等植物激素或环境信号的枢纽蛋白.植物体接受GA信号后, 通过泛素-蛋白酶体途径降解DELLA蛋白, 解除其对生长的抑制作用(Zhang et al., 2009).冷、盐和渗透胁迫下, GA含量降低, 导致植物生长减缓(Colebrook et al., 2014).极低浓度GA下, DELLA蛋白抑制GA介导的应答调控, 外源GA处理促进DELLA蛋白降解(Claeys et al., 2012). ...
An AP2 domain-containing gene,ESE1, targeted by the ethylene signaling component EIN3 is important for the salt response in Arabidopsis 1 2011
... 正常环境下, AP2/ERF转录因子表达水平较低, 激素或逆境胁迫相关基因与AP2/ERF转录因子保守的顺式作用元件结合, 进而调控其表达(Owji et al., 2017).研究表明, 拟南芥DREB1A/CBF3、DREB1C/CBF2、DREB2C、DREB2G和DEAR3的启动子中包含较多的热激元件(heat shock element, HSE)、低温响应(low temperature responsive, LTR)、脱落酸响应(ABA responsive element, ABRE)等顺式作用元件(Sazegari et al., 2015).热激因子1 (heat shock factor 1, HSF1)、脱落酸响应结合蛋白(ABA responsive element binding protein/ABRE binding factors, AREB/ABF)可分别与DREB2A启动子中的热激应答和ABA响应元件结合, 调控其表达, 进而改变植物胁迫的耐受性(Kim et al., 2011; Liu et al., 2011).蛋白磷酸酶2C (type 2C protein phosphatase, PP2C)包含2个以上的ABRE元件, AREB1、AREB2和ABF3协同正调控AHG1、AHG3、HAI1及HAI2等PP2Cs基因表达(Yoshida et al., 2010).此外, 越来越多的研究证明ET信号通路参与拟南芥盐胁迫反应.进一步分析表明, ESE1 (ethylene and salt-inducible ERF1)在EIN3/EIL1下游调节ET响应基因.EIN3 (ethylene insensitive 3)可与ESE1启动子结合形成EIN3-ESE1转录复合体, 继而调控RD29A和COR15A等基因的表达(Zhang et al., 2011).磷酸化、泛素化、甲基化和乙酰化等组蛋白修饰方式通过调控染色质开放或封闭的空间状态, 激活或抑制AP2/ERF转录(Pfluger and Wagner, 2007).组蛋白去乙酰化酶(histone deacetylase, HDAC)和聚乙二醇(PEG)促进DREB1基因启动子区乙酰化, 增强DREB1基因的表达(Zhang et al., 2018).非生物胁迫下, 组蛋白变体和组蛋白修饰状态也可发生改变, 并可遗传到下一代(Asensii-Fabado et al., 2017).miRNA介导的RNA沉默和翻译抑制在AP2/ERF表达调控中也发挥重要作用.研究发现miRNA172靶向拟南芥AP2mRNA并抑制其翻译, 而miRNA156和miRNA838两者也具有类似调节AP2/ERF的功能(Kavas et al., 2015).此外, 在拟南芥、水稻及玉米中还发现通过可变剪切产生的OsDREB2A/2B、ZmDREB2A、WDREB2以及HvDRF1等多种AP2/-ERF功能异构体(Matsukura et al., 2010). ...
The roles of two transcription factors, ABI4 and CBFA, in ABA and plastid signaling and stress responses 1 2013
... DREB-A3亚组成员ABI4 (ABA insensitive 4)是ABA信号通路的重要组分.植株受到胁迫后, 体内积累ABA和ROS, ABI4可被ABA、ROS及糖质体等多种信号通路靶向调控.CCAAT结合因子A (CCAAT binding factor A, CBFA)是四吡咯化合物HAP (heme activator protein)三聚体转录复合物的亚基, 当产生过量ROS时, ABI4可与其它转录因子竞争性地结合CCACGT元件, 抑制CBFA的表达, 进而调控质体反向信号通路中相关基因的表达(Zhang et al., 2013).拟南芥突变体实验表明, ABI4在YL1 (yellow leaf 1) (Li et al., 2016)和DPG1 (delayed pale-greening 1) (Yi et al., 2019)等基因参与盐胁迫响应的调控网络中也发挥重要作用.逆境胁迫下, ORA47在JA (jasmo-nate acid)和ABA信号转导途径中靶向调节ABA和JA诱导的下游应答基因, ERF18/ORA-47识别启动子顺式元件(NC/GT)CGNCCA (O-box), 激活PP2C家族ABI2转录因子的表达.ABI1则在ORA-47上游起作用, 从而形成ABI1-ORA47-ABI2正反馈基因表达回路, 调控ABA信号转导和干旱胁迫响应(Chen et al., 2016).此外, RAV1转录因子在ABA信号转导途径中也发挥关键作用, RAV1能与ABI3、ABI4和ABI5启动子结合, 其过表达抑制ABI3、ABI4和ABI5的转录活性, 导致根系生长发育过程中对ABA不敏感.同时, 蔗糖非酵解型蛋白激酶(SnRK2.2、SnRK2.3和SnRK2.6)在核内磷酸化RAV1, 降低ABI5对RAV1的依赖性, 并负调控RAV1对靶基因的转录抑制作用(Feng et al., 2014). ...
Mutational evidence for the critical role of CBF transcription factors in cold acclimation in Arabidopsis 1 2016
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
The broad roles of CBF genes: from development to abiotic stress 1 2016
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
CBF-dependent signaling pathway: a key responder to low temperature stress in plants 3 2011
... CBFs (C-repeat binding factors)转录因子属于DREB亚族A1亚组成员.近年来, 科学家聚焦CBF参与植物冷信号途径的分子调控机制, 对其进行了深入系统的研究(Zhou et al., 2011; Yamasaki and Randall, 2016; Shi et al., 2018).冷胁迫下, 植物通过其它通路将冷信号间接传递给CBF基因(Park et al., 2015).ICE1/2 (inducer of CBF expression 1/2)、CAMTA1/3 (calmodulin-binding transcription activator 1/3)、MYB15 (myeloblastosis 15)、EIN3 (ethylene insensitive 3)、BZR1 (brassina-zole-resis- tant 1)、SOC1 (suppressor of overexpression of constans 1)、CCA1 (circadian clock associated 1)、LHY (late elongated hypocotyl)及PIF3/4/7 (phytochrome-interacting factor3/4/7)等关键转录调节因子则通过协同或拮抗作用参与CBF基因转录水平的调控(刘静妍等, 2017).ICE作为CBF冷信号途径中第1个被鉴定的正调控因子, 在CBF基因的表达调控网络中发挥重要作用(Park et al., 2015; Jia et al., 2016; Shi et al., 2018).而ICE1自身也受HOS1 (high osmotic expression 1)介导的泛素化、SIZ1 (SAP and Miz 1)介导的SUMO化和OST1 (open stomata 1)介导的磷酸化修饰, 以及MYB15、JAZ (jasmonate ZIM-domain)转录抑制子协同或拮抗作用的精细调控(Chinnusamy et al., 2007; Qin et al., 2011; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
... ; Zhou et al., 2011).目前, 关于ICE-CBF-COR冷信号途径研究最为深入.低温胁迫下, ICE与CBF共建冷胁迫调控通路, ICE特异性地结合CBF3启动子中的MYC结合位点CANNTG (Agarwal et al., 2006), 正调控CBF3基因的表达(Liu et al., 2018a).而CBF1的表达则受ICE1的同源基因ICE2调控, 超量表达ICE2促进CBF1的表达, 但CBF2的表达受CBF1和CBF3负调控, 其是否受ICE1或ICE2的调控仍未见实验证明(Zhao and Zhu, 2016).CBFs通过结合COR (cold regulated)、LTI (low-temperature induced)、RD (responsive to dehydration)及DHN (dehydrin)等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
... )等冷应答基因启动子区域的DRE/CRT顺式作用元件, 激活此类基因的表达, 进而增强植物的耐冷性(Zhou et al., 2011; Mizoi et al., 2012; Licausi et al., 2013).利用RNA-seq技术对cbf突变体进行分析, 结果显示CBF突变影响全转录组水平上约10%-25%的COR基因表达(Jia et al., 2016; Zhao et al., 2016).在拟南芥cbf突变体和过表达株系中, 低温均可快速激活CBF1、CBF2和CBF3的转录活性, 识别并结合下游基因启动子中的CRT/DRE顺式元件, 调控冷诱导相关基因的表达(Park et al., 2015).与野生型相比, 敲除CBF1和CBF3使拟南芥植株的抗冻能力降低60% (Novillo et al., 2007). ...
The ERF11 transcription factor promotes internode elongation by activating gibberellin biosynthesis and signaling 1 2016
... AP2/ERFs参与调控植物GA生物合成相关基因的转录.盐胁迫下, DREB1E和DREB1F抑制GA20ox的表达, 使GA生物合成减少, 引起植株生长迟缓(Magome et al., 2004).过表达ERF6抑制GA2ox的表达水平, 诱导赤霉素降解酶(gibberellin 2-oxidase 6, GA2ox6)的合成, 维持DELLA蛋白的稳定性.渗透胁迫下, erf6功能获得突变体植株矮小且对胁迫高度敏感, 而erf5/erf6缺失突变体生长受胁迫影响较小.研究发现ERF6还激活了STZ、MYB51和WRKY33等渗透胁迫响应基因的表达(Dubois et al., 2013).冷胁迫下, CBF1转录因子激发GA2ox基因的表达而降低GA含量, 组成型表达CBF1抑制GA的积累, 植株表现矮化, 表明DELLA蛋白是CBF1介导的冷胁迫响应的关键组分(Achard et al., 2008).相反, ERF11作为一种GA生物合成和信号转导的正调控因子, 抑制ET生物合成, 促进植株节间伸长.过表达ERF11上调GA3ox1和GA20ox基因的表达水平, 增加GA的积累量(Zhou et al., 2016).ERF6作为转录激活因子诱导ERF11的表达, 反之, ERF11抑制GA2-OX6、STZ、MYB51和WRKY33等ERF6相关靶基因的表达.进一步实验表明, 4个靶基因在erf11/erf6过表达植株中也不能被诱导表达, ERF11过表达可消除ERF6过表达导致的极度矮化现象, 说明ERF11可能通过直接竞争目标基因启动子而在分子水平上拮抗ERF6 (Dubois et al., 2015). ...
TRANSLUCENT GREEN, an ERF family transcription factor, controls water balance in Arabidopsis by activating the expression of aquaporin genes 3 2014
... AP2/ERF转录因子通过特异性结合胁迫应答基因启动子区顺式元件, 参与调控植物的逆境防御反应.DREB亚族成员特异性识别并结合启动子区域的DRE/CRT (dehydration responsive element/C-re-peat)元件(核心序列A/GCCGAC), 诱导干旱、低温和盐胁迫相关应答基因的表达.ERF亚族成员则可通过与乙烯响应元件(ethylene response element, ERE) (核心序列AGCCGCC, 又称GCC-box)结合, 从而参与调控乙烯应答和非生物胁迫(Franco-Zorrilla et al., 2014).此外, 研究发现许多AP2/ERF转录因子可同时与DRE/CRT和ERE两种元件结合, 如DREB亚族成员TINY、CBF1、ERF53、RAP2.4、TG/RAP2.4a以及ERF亚族成员ERF1、ERF4和ERF71 (Chen et al., 2012; Zhu et al., 2014; Lee et al., 2015), 但结合能力有差异.在拟南芥中, DREB2C和DEAR3与类GCC-box元件(GCCGCC)的结合能力远超DRE元件(Franco-Zorrilla et al., 2014).AP2/ERF转录因子还能与DRE和ERE之外的顺式元件结合, 如偶联元件1 (CE1, TGCCACCG)、类偶联元件3 (CE-3like, CGCG)和缺氧响应启动子元件(hypoxia-responsive promoter element, HRPE), 以及ATCTA、CAACA、CATGCA、CAA/CA/CTG、ATCGAG和(NC/GT) CGNCCA (Bossi et al., 2009; Zhu et al., 2010; Chen et al., 2016; Gasch et al., 2016; Park et al., 2016).近年来, 科学家通过蛋白质结合微阵列(protein binding microarray, PBM)技术发现结构特征高度相似的转录因子具有相似的DNA结合位点, 同时也具有相似的生物学功能.该研究结果合理地解释了转录因子功能冗余现象, 也为预测和挖掘未知AP2/ERF转录因子提供了高效可行的技术方法(Franco-Zorrilla et al., 2014). ...
... DREB-A2亚组转录因子DREB2主要参与调节植物的抗旱和耐热性(Mizoi et al., 2012).干旱和高温可诱导DREB2基因的表达, 进而正向调控下游干旱或热胁迫响应基因(Maruyama et al., 2009).研究表明DREB-A4家族成员HRD (hardy)和DREB-A6家族成员(ERF53、RAP2.4和TG/RAP2.4a)也在干旱和耐盐性调控中发挥关键作用(Karaba et al., 2007; Lin et al., 2008; Chen et al., 2012; Zhu et al., 2014).过表达HRD能显著增强拟南芥和水稻植株的耐旱和耐盐性(Karaba et al., 2007).同样, 过表达TG (translucent green)导致植株叶片玻璃化, 叶片细胞含水量增加(Zhu et al., 2014).TG也参与调控抗坏血酸过氧化物酶基因APX (ascorbate peroxidase)的表达, 抵御活性氧(reactive oxygen species, ROS)引起的光氧化胁迫(Rudnik et al., 2017).除拟南芥外, 人们对水稻、玉米、番茄(Lycopersicon esculentum)、葡萄(Vitis vinifera)等作物的DREB1和DREB2转录因子也进行了广泛的研究(Xiao et al., 2006; Matsukura et al., 2010; Dou et al., 2014). ...
... )导致植株叶片玻璃化, 叶片细胞含水量增加(Zhu et al., 2014).TG也参与调控抗坏血酸过氧化物酶基因APX (ascorbate peroxidase)的表达, 抵御活性氧(reactive oxygen species, ROS)引起的光氧化胁迫(Rudnik et al., 2017).除拟南芥外, 人们对水稻、玉米、番茄(Lycopersicon esculentum)、葡萄(Vitis vinifera)等作物的DREB1和DREB2转录因子也进行了广泛的研究(Xiao et al., 2006; Matsukura et al., 2010; Dou et al., 2014). ...
The Arabidopsis AP2/ERF transcription factor RAP2.6 participates in ABA, salt and osmotic stress responses 2 2010
... AP2/ERF转录因子通过特异性结合胁迫应答基因启动子区顺式元件, 参与调控植物的逆境防御反应.DREB亚族成员特异性识别并结合启动子区域的DRE/CRT (dehydration responsive element/C-re-peat)元件(核心序列A/GCCGAC), 诱导干旱、低温和盐胁迫相关应答基因的表达.ERF亚族成员则可通过与乙烯响应元件(ethylene response element, ERE) (核心序列AGCCGCC, 又称GCC-box)结合, 从而参与调控乙烯应答和非生物胁迫(Franco-Zorrilla et al., 2014).此外, 研究发现许多AP2/ERF转录因子可同时与DRE/CRT和ERE两种元件结合, 如DREB亚族成员TINY、CBF1、ERF53、RAP2.4、TG/RAP2.4a以及ERF亚族成员ERF1、ERF4和ERF71 (Chen et al., 2012; Zhu et al., 2014; Lee et al., 2015), 但结合能力有差异.在拟南芥中, DREB2C和DEAR3与类GCC-box元件(GCCGCC)的结合能力远超DRE元件(Franco-Zorrilla et al., 2014).AP2/ERF转录因子还能与DRE和ERE之外的顺式元件结合, 如偶联元件1 (CE1, TGCCACCG)、类偶联元件3 (CE-3like, CGCG)和缺氧响应启动子元件(hypoxia-responsive promoter element, HRPE), 以及ATCTA、CAACA、CATGCA、CAA/CA/CTG、ATCGAG和(NC/GT) CGNCCA (Bossi et al., 2009; Zhu et al., 2010; Chen et al., 2016; Gasch et al., 2016; Park et al., 2016).近年来, 科学家通过蛋白质结合微阵列(protein binding microarray, PBM)技术发现结构特征高度相似的转录因子具有相似的DNA结合位点, 同时也具有相似的生物学功能.该研究结果合理地解释了转录因子功能冗余现象, 也为预测和挖掘未知AP2/ERF转录因子提供了高效可行的技术方法(Franco-Zorrilla et al., 2014). ...
... 研究表明, 非生物胁迫下ABA可促进ANT (Meng et al., 2015b)、ERF53 (Hsieh et al., 2013)、RAP2.6L (Liu et al., 2012)和RAP2.6 (Zhu et al., 2010)转录, 上述转录因子进而结合胁迫应答顺式作用元件DRE/ABRE, 调控下游相关靶基因的表达, 增强植物对非生物胁迫的耐受能力.在水稻中ERF71正向调控ABA信号转导, 过表达ERF71可促进根中ABI5、PP2C68、RAB16C及RAB16D等基因上调表达.此外, ERF71可直接调控木质素生物合成基因CCR1、CCR10及C4H转录, 诱导根系径向生长, 改变根系构型, 提高耐旱能力(Lee et al., 2017; Li et al., 2018).盐胁迫和ABA双重诱导下, ABA增强DREB2的转录活性, 进而促进Rd29A协同表达(Lee et al., 2016). ...
Cytokinin response factor 4 (CRF4) is induced by cold and involved in freezing tolerance 1 2016
... 近年来, 有关ERF亚族参与非生物胁迫响应的研究越来越多.在拟南芥中发现多种非生物胁迫可诱导细胞分裂素应答因子(cytokinin response factor, CRF)的表达, CRF是ERF-VI亚族成员之一, 具有正向调节植物渗透胁迫耐受性和抗冻性作用(Rashotte et al., 2006).CRF2和CRF6可与生长素外输载体PIN (PIN- FORMED)基因启动子区的特定元件结合, 正向调控PIN基因的表达, 这在CRF超表达和缺失突变体研究中得到了验证(Kim, 2016).CRF2和CRF3均参与调控冷胁迫下拟南芥侧根(lateral root, LR)的发育, crf2、crf3、crf2/crf3突变体在冷胁迫下侧根密度比野生型植株更低, CRF2或CRF3过表达均可促进植株侧根密度的增加(Jeon et al., 2016).冷胁迫可诱导CRF2的表达, 因此CRF2可能通过诱导PIN1和PIN7的表达以弥补冷胁迫对侧根发育的影响; 而在缺失突变体crf2中, CRF3超表达可减弱冷胁迫对侧根生长的抑制效应, 推测CRF3可能通过其它途径参与侧根的发育调控(Jeon et al., 2016; Kim, 2016).进一步分析显示, CRF2和CRF3分别通过依赖和不依赖于TCS (two-component signaling)的途径响应冷胁迫(Joen et al., 2016).CRF4是少数不受CTK调控的CRF转录因子之一, 可促进CORs的表达并正向调控植物的冷/冻耐受性(Zwack et al., 2016a).多种非生物胁迫会诱导植物积累活性氧(ROS), 导致细胞损伤, 因此植物维持最低水平的活性氧对其生存至关重要.多种胁迫刺激均可诱导CRF6的表达, CRF6可能通过抑制细胞分裂素相关基因的表达介导细胞分裂素与氧化应激反应之间的联系(Zwack et al., 2016b).CRF6在H2O2诱导的氧化胁迫应答中直接或间接抑制ARR6 (type-A response regulator 6)、ARR9、ARR11、LOG7及ABCG14基因的表达.酵母单杂交实验显示, CRF6直接结合到ARR6启动子区, 激活ARR6的表达, 而对ARR9、LOG7的表达调控可能需要其它植物特异因子的参与(Zwack et al., 2016b). ...
 ,?,*, 杨蕾?, 杨海健, 王武重庆市农业科学院果树研究所, 重庆 401329
,?,*, 杨蕾?, 杨海健, 王武重庆市农业科学院果树研究所, 重庆 401329
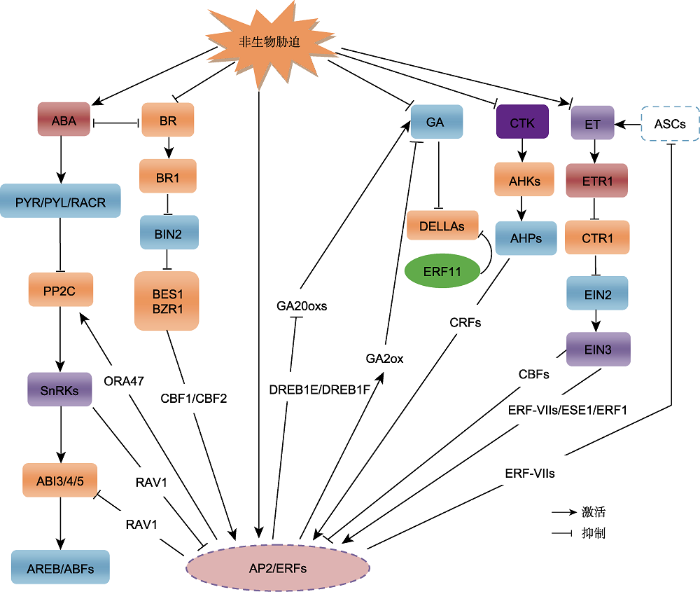 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}