 ,*山西师范大学生命科学学院, 植物分子与环境胁迫响应山西省高校重点实验室, 临汾 041000
,*山西师范大学生命科学学院, 植物分子与环境胁迫响应山西省高校重点实验室, 临汾 041000Response of Arabidopsis Cohesin RAD21 to Cell Division after Enhanced UV-B Radiation
Fangfang He, Huize Chen, Jinlin Feng, Lin Gao, Jiao Niu, Rong Han,*Higher Education Key Laboratory of Plant Molecular and Environmental Stress Response in Shanxi Province, College of Life Sciences, Shanxi Normal University, Linfen 041000, China通讯作者:
责任编辑: 孙冬花
收稿日期:2020-01-15接受日期:2020-05-15网络出版日期:2020-07-01
| 基金资助: |
Corresponding authors:
Received:2020-01-15Accepted:2020-05-15Online:2020-07-01

摘要
关键词:
Abstract
Keywords:
PDF (4225KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
贺芳芳, 陈慧泽, 冯金林, 高琳, 牛娇, 韩榕. 拟南芥黏连蛋白RAD21对增强UV-B辐射后细胞分裂的响应. 植物学报, 2020, 55(4): 407-420 doi:10.11983/CBB20009
He Fangfang, Chen Huize, Feng Jinlin, Gao Lin, Niu Jiao, Han Rong.
紫外线辐射是太阳电磁辐射的一部分, 到达地球表面的太阳辐射约有7%是紫外线(Frohnmeyer and Staiger, 2003)。紫外线分为UV-A (315-400 nm)、UV-B (280-315 nm)和UV-C (200-280 nm)。最具活力的UV-C被臭氧(O3)层完全吸收; UV-A未被臭氧层衰减, 但破坏最小且参与植物的光形态建成(Bj?rn, 1996)。UV-B属于中长波段, 约95%的UV-B辐射被臭氧层吸收, 到达地表的UV-B平均强度为1 W·m-2 (?ejka et al., 2011), 可引起多种破坏效应, 属于强效应波(Sugimoto-Shirasu and Roberts, 2003)。近年的研究表明, UV-B辐射影响植物的生长发育和生理生化过程(Bj?rn et al., 1997), 导致其形态结构、生理代谢、遗传特性和生长周期等发生改变(Caldwell et al., 1995; Nogués et al., 1998), 包括植物的生物量减少(Vandenbussche et al., 2018)、表皮变化、异常生长(Searles et al., 2001)以及光系统受损害(Liu et al., 2013)等。因此, 揭示UV-B辐射对植物的影响及作用机理尤为重要。
控制细胞分裂和扩张是调节器官大小的核心(方荧等, 2018)。细胞水平上, UV-B诱导细胞分裂、伸长和分化变化等异常现象的发生(Robson et al., 2015)。韩榕(2002)在小麦(Triticum aestivum)根尖中发现UV-B辐射导致植物DNA损伤, 造成有丝分裂异常, 出现翘根、分束分裂和染色体畸变等现象, 并抑制细胞的分裂, 最终在生理水平上表现出生物量的减小。拟南芥(Arabidopsis thaliana)是一种优良的模式植物(陈建权等, 2018), 目前已有一些UV-B辐射对其影响的研究。UV-B辐射抑制拟南芥细胞周期的进程(Jiang et al., 2011), DNA受损使拟南芥根尖细胞G1/S期的转变延缓, 可能是其抑制植物生长的原因之一(王静等, 2009)。延缓G1/S期的转变, 推测可能是由于切除CPDs后才可进行DNA复制, 细胞周期的抑制促进DNA修复, 但也会导致细胞数量减少或者核内复制发生(陈慧泽和韩榕, 2015)。UV-B辐射减小拟南芥莲座叶直径及面积, 主要原因是细胞分裂和扩张受到抑制(Hectors et al., 2010), 进而抑制叶片生长(Casadevall et al., 2013)。
细胞分裂是生物体生长繁殖的基础, 真核生物的细胞分裂包括有丝分裂和减数分裂(Suzuki et al., 2013)。染色体黏着在有丝分裂和减数分裂过程中扮演着重要角色, 是保证姐妹(或同源)染色体正确分离并分配到子细胞中的关键调控环节之一(张亮然, 2006)。黏着素(cohesin)是一种将姐妹染色单体紧密连接在一起的蛋白质复合物, 由SMC1、SMC3、SCC1/RAD21和SCC3组成, 参与有丝分裂期间姐妹染色单体的凝聚(Losada, 2007; Skibbens, 2009), 其不正常的黏着与分离可导致染色体分离出现错误。芽殖酵母(Saccharomyces cerevisiae)姐妹染色单体的内聚性在G1晚期和S期建立, 在中期/后期转变中被消除, 以允许有丝分裂姐妹染色单体正确且及时分离(da Costa-Nunes et al., 2014)。免疫荧光定位分析表明, OsRad21-1和OsRad21-3特异地定位于有丝分裂染色体上, 可能参与有丝分裂姐妹染色体之间的黏着(张亮然, 2006)。拟南芥复合体核心蛋白亚基REC8/RAD21有3个同源基因: AtRAD21.1/SYN2、AtRAD21.2/SYN3和AtRAD21.3/SYN4 (Dong et al., 2001; Da Costa-Nunes, 2006)。AtRAD21.1 (At5g40840)和AtRAD21.3 (At5g16270)位于V号染色体上, AtRAD21.2 (At3g59550)位于III号染色体上。RAD21.3编码一个SCC1/REC8同源物, 可能参与有丝分裂。由于AtRAD21.3与核基质或姐妹染色单体的内聚性有关(Sadano et al., 2000), AtRAD21.1与AtRAD21.3可能代表有丝分裂凝聚蛋白, 但在有丝分裂中功能冗余(Da Costa-Nunes et al., 2006; Jiang et al., 2007)。有丝分裂过程中, 染色体S期复制产生的姐妹染色单体通过黏着素紧密联系, 直到后期起始时才分开(Xu et al., 2004)。凝聚力缺陷会导致中期染色体错位(Hoque and Ishikawa, 2002), 从而阻止进入后期; RAD21缺陷则导致有丝分裂停滞和非整倍体产生(Hirano, 2000; Nasmyth et al., 2000)。γ射线对atrad21.3突变体的作用已有报道(da Costa-Nunes et al., 2014), 关于UV-B辐射对atrad21.3突变体有丝分裂的影响目前尚未有研究。
我们通过构建过表达载体, 检测RAD21.3的亚细胞定位, 设置对照与UV-B辐射处理组, 初步比较野生型(WT)、atrad21.3突变体和过表达株系形态及生理生化指标的变化, 并进一步观察atrad21.3的有丝分裂现象, 统计畸变率, 研究RAD21.3对UV-B辐射诱导的异常有丝分裂的响应。
1 材料与方法
1.1 材料及培养方法
哥伦比亚生态型拟南芥(Arabidopsis thaliana (L.) Heynh.) (Col-0)和本氏烟草(Nicotiana benthamiana)种子由植物分子与环境胁迫响应山西省高等学校重点实验室保存。atrad21.3突变体(salk_076116C)购自拟南芥资源中心(Nottingham Arabidopsis Stock Centre, NASC)。植物表达载体pSuper1300-GFP、大肠杆菌(Escherichia coli) DH5α、农杆菌(Agrobacterium tumefaciens) GV3101均由本课题组保存。过表达载体pSuper1300-RAD21.3-GFP为本实验室构建。培养方法: 将种子在4°C下春化, 之后用1.5%次氯酸钠溶液消毒10分钟, 灭菌水清洗3-5次, 点种到MS或1/2MS培养基上。移栽后密封托盘, 放置在温度为(22±2)°C、相对湿度为60%-80%、光周期为16小时光照/8小时黑暗的培养室中培养。
1.2 AtRAD21.3的克隆及亚细胞定位
1.2.1 AtRAD21.3的克隆从拟南芥基因组数据库(TAIR)中查找RAD21.3基因序列并设计引物。LP: 5°-GCGTCGACATGTTTTATTCGCAGTTTATATTAGCTAA-3°; RP: 5°-GGGGTACCGAAGATGGATTTGGTGAGCTT-3°。以反转录cDNA为模板, 使用Supermix酶扩增。扩增程序: 95°C预变性1分钟; 95°C20秒, 53.5°C20秒, 72°C90秒, 35个循环; 72°C延伸5分钟。PCR产物使用1%琼脂糖凝胶电泳检测, 使用DNA纯化回收试剂盒回收约3 096 bp的AtRAD21.3目的片段。用SalI以及KpnI双酶切载体与目的基因, 切胶回收。载体与目的基因按照等摩尔比例混合, 用T4-DNA连接酶4°C过夜连接, 连接产物用热激法转入DH5α, 将转化成功的大肠杆菌接种到LB固体培养基(含卡那霉素)上, 于37°C恒温培养箱中培养12-16小时。挑取单菌落于LB液体培养基中(含卡那霉素), 通过菌液PCR验证阳性菌落。提取质粒, 进行双酶切验证后, 送北京华大基因公司测序。
1.2.2 AtRAD21.3的亚细胞定位
将构建成功的载体转化GV3101, 涂板, 于28°C培养箱中培养。挑取单菌落摇菌, OD600为0.8-1集菌。重悬菌液, OD600为0.5-0.6, 暗培养3小时, 注射烟草叶片。避光处理30小时后对叶片注射DAPI (4′,6-diamidino-2-phenylindole), 用激光共聚焦扫描显微镜观察烟草叶片的瞬时表达; 并进一步观察转基因拟南芥根尖、AtRAD21.3的亚细胞定位及UV-B辐射后的变化。
1.3 UV-B辐射对AtRAD21表达量的影响
采用本课题组前期筛选出的UV-B剂量(5 kJ·m-2)处理拟南芥幼苗, 提取整株幼苗的RNA, 反转录成cDNA, 以ACTIN为内参, 通过RT-PCR检测AtRAD21.1和AtRAD21.3在UV-B辐射前后表达量的变化, 使用Image J软件分析。1.4 UV-B辐射对拟南芥形态及生理生化指标的影响
植物在胁迫条件下会表现出形态和生理生化变化。本研究统计了UV-B辐射后, WT、atrad21.3和过表达株系的种子萌发率(李晓阳等, 2013), 幼苗根长、株高和抽薹时间, 叶绿素和可溶性糖含量, 及丙二醛(malon-dialdehyde, MDA)浓度和超氧化物歧化酶(superoxi-de dismutase, SOD)活性的变化。叶绿素含量、MDA浓度和SOD活性的测定及可溶性糖的提取参考张志良等(2009)所述方法, 并稍作修改。1.5 UV-B辐射对atrad21.3有丝分裂的影响
通过设置对照和UV-B处理组, 对WT和atrad21.3进行转录组高通量测序(RNA-seq), 筛选出部分与细胞周期相关的基因, 通过RT-qPCR观察UV-B辐射前后细胞周期相关基因的相对表达量变化。切取不同处理组的根尖, 用1 mol·mL-1盐酸, 60°C下解离5分钟。碱性品红染色后在荧光显微镜下观察有丝分裂现象, 并统计染色体畸变率。1.6 数据分析
所有检测指标均重复测定3次。使用Microsoft Excel 2010软件进行实验数据统计。使用SPSS Statistics 17.0软件中的Duncan法进行差异显著性分析。用SigmaPlot 12.5软件绘图。2 结果与讨论
2.1 atrad21.3突变体的鉴定与表型观察
从TAIR数据库中查阅atrad21.3突变体的基因信息。测序结果显示, atrad21.3纯合突变体的T-DNA插在基因的第11个外显子(图1A)。atrad21.3的表达分析表明, T-DNA插入阻止了全长AtRAD21.3转录物的表达。利用三引物法(LP、RP和BP)鉴定突变体, 用LP+RP和RP+BP引物进行DNA水平的鉴定(图1B)。通过检测突变体中RAD21.3基因的转录本情况鉴定RNA水平。提取生长7天的WT和atrad21.3突变体的RNA, 用LP+RP引物进行PCR扩增(图1C), 发现WT扩增出特异性条带, atrad21.3突变体相应位置无条带, 而内参基因ACTIN在WT和atrad21.3突变体中得到很好的扩增。由此可知, atrad21.3突变体中RAD-21.3基因的转录本水平受到影响。图1
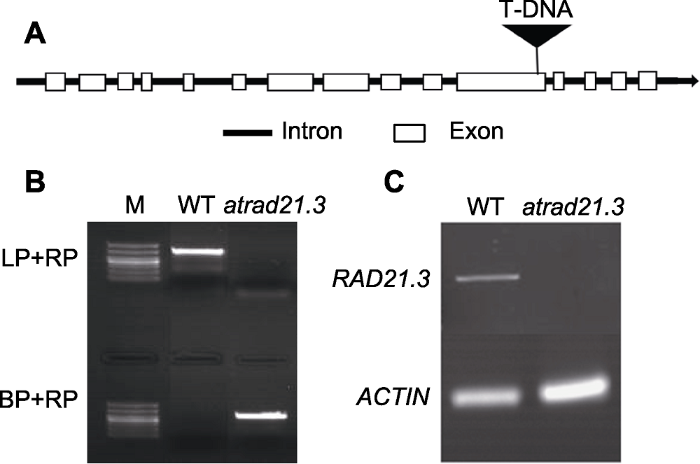 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1atrad21.3的T-DNA插入示意图及鉴定
(A) atrad21.3基因结构以及T-DNA插入位点示意图; (B) atrad21.3 DNA水平鉴定(M: 分子量标记); (C) atrad21.3中RAD 21.3基因的转录本水平。WT: 野生型
Figure 1T-DNA insertions and identification of atrad21.3
(A) Gene structure and T-DNA insertions of atrad21.3; (B) Identification of DNA level in atrad21.3(M: Marker); (C) Transcriptional level of RAD21.3 gene in atrad21.3. WT: Wild type
有研究表明, AtRAD21.3在染色体结构维持中的作用可以解释拟南芥发育延迟现象, 特别是atrad21.3突变体延迟抽薹表型(Da Costa-Nunes et al., 2006)。故我们首先对突变体的表型进行了观察。使用体式显微镜观察WT和atrad21.3的种子(图2A, D), 发现二者形态无明显差异。之后, 观察第12天的WT与atrad- 21.3的莲座叶数目, 发现也无区别(图2B, E); 但第32天的WT抽薹, atrad21.3尚未抽薹(图2C, F)。比较WT与atrad21.3突变体的株高, 结果表明atrad21.3突变体较矮(图2G)。综上, 在营养生长阶段WT与atrad- 21.3无明显差异, 但在生殖阶段atrad21.3抽薹延迟且株高较矮。说明atrad21.3突变体在营养阶段无明显的发育表型, 可育且可收获有活力的种子, 并经历正常的营养生长。
图2
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2野生型(WT)和atrad21.3的表型分析
(A) WT种子; (B) 第12天的WT莲座叶; (C) WT抽薹; (D) atrad21.3种子; (E) 第12天的atrad21.3莲座叶; (F) atrad21.3未抽薹; (G) WT和atrad21.3的株高. (A), (D) Bars=10 μm; (B), (C), (E)-(G) Bars=1.0 cm
Figure 2Phenotypic analysis of wild type (WT) and atrad21.3
(A) Seed of WT; (B) Rosette leaf (day 12) of WT; (C) Bolting of WT; (D) Seed of atrad21.3; (E) Rosette leaf (day 12) of atrad21.3; (F) Bolting of atrad21.3; (G) Plant height of WT and atrad21.3. (A), (D) Bars=10 μm; (B), (C), (E)-(G) Bars=1.0 cm
2.2 AtRAD21.3的克隆及亚细胞定位分析
2.2.1 AtRAD21.3的克隆构建AtRAD21.3过表达载体。以WT的cDNA为模板进行PCR扩增, 经琼脂糖凝胶电泳检测到长度约为3 096 bp的单一条带(图3A), 且条带大小与预期相符。切胶纯化, 条带如图3B所示, 位置正确, 条带单一, 浓度可用; 双酶切载体与目的基因, 并切胶回收。将AtRAD21.3基因片段连接在pSuper1300-GFP载体上; 转化DH5α, 挑取单菌落进行RT-PCR检测, 筛选出2、3、4和6号管, 其条带明显(图3C), 说明转化成功。提取质粒, 用限制性内切酶KpnI和SalI进行双酶切验证, 电泳结果显示出现2条产物, 切出大小约为3 096 bp的条带(图3D)。初步认定重组质粒构建成功, 送北京华大基因公司测序, 验证正确。
图3
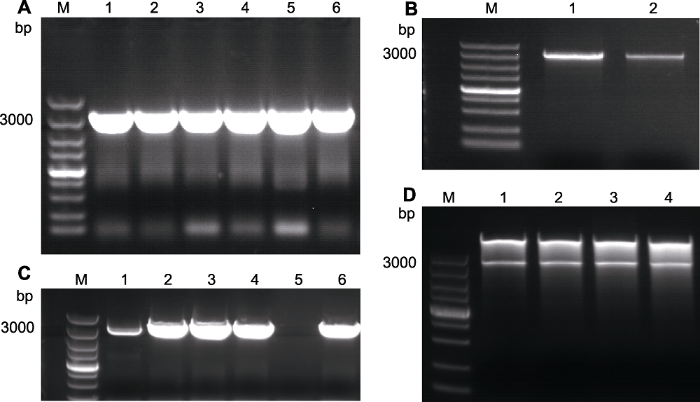 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3AtRAD21.3的克隆
(A) AtRAD21.3的扩增产物(1-6: 产物); (B) AtRAD21.3切胶回收电泳结果(1, 2: 切胶回收产物); (C) 菌液PCR结果(1-4, 6: 阳性单菌落; 5: 阴性对照); (D) 质粒双酶切验证(1-4: 双酶切结果)。M: 分子量标记
Figure 3Cloning of AtRAD21.3
(A) PCR products of AtRAD21.3 (1-6: Products); (B) Gel cutting recovery result of AtRAD21.3 (1, 2: Gel cutting recovery products); (C) Bacterial PCR result (1-4, 6: Positive single colony; 5: Negative control products); (D) Dual-restriction digestion of plasmid (1-4: Dual-restriction result). M: DNA marker
2.2.2 AtRAD21.3的亚细胞定位分析
(1) 过表达载体在烟草中的瞬时表达 将测序成功的pSuper1300-RAD21.3-GFP质粒转入GV3101, 摇菌, 重悬菌液; 对4片叶的烟草叶表皮细胞进行注射, 黑暗培养30小时后向叶片注射DAPI (染细胞核), 取烟草下表皮, 在激光共聚焦显微镜下观察, 发现DAPI和GFP均在细胞核表达(图4), 因此推测AtRAD21.3可能定位于细胞核。
图4
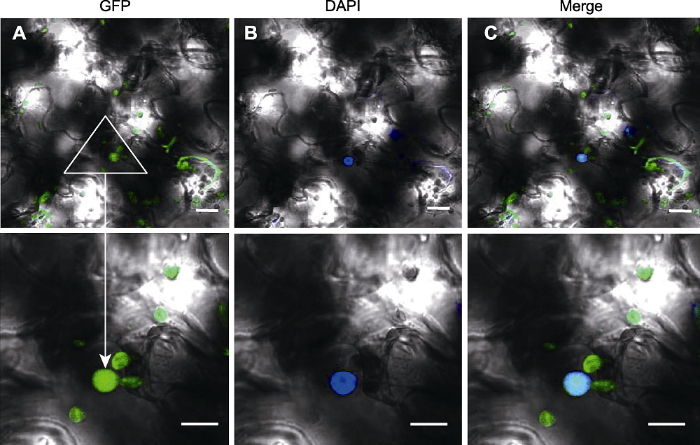 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4pSuper1300-RAD21.3-GFP在烟草叶片的瞬时表达
(A) pSuper1300-RAD21.3-GFP的GFP荧光信号图像; (B) pSuper1300-RAD21.3-GFP在烟草叶片的DAPI染色图像; (C) A和B叠加的图像。Bars=10 μm
Figure 4Transient expression of pSuper1300-RAD21.3-GFP in leaves of Nicotiana benthamiana
(A) GFP signal of pSuper1300-RAD21.3-GFP; (B) DAPI staining of pSuper1300-RAD21.3-GFP in leaves of Nicotiana benthamiana; (C) Merged image of A and B. Bars=10 μm
(2) pSuper1300-RAD21.3-GFP过表达植株根尖的亚细胞定位观察 生物信息学预测AtRAD21.3定位在细胞核, 并显示AtRAD21.3中间区域含有潜在的核定位信号, 及PEST序列等。本研究以pSuper1300-RAD21.3- GFP转基因植株的根尖为材料, 进行DAPI染色, 用激光共聚焦扫描显微镜观察。UV-B处理前后AtRAD21.3蛋白的荧光信号集中在细胞核上; CK组细胞核边缘比较整齐, 而UV-B处理后细胞核整体形态发生变化(图5)。
图5
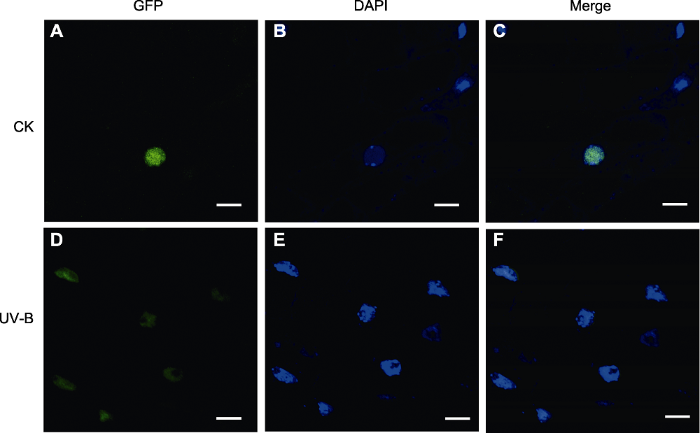 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5过表达载体pSuper1300-RAD21.3-GFP在UV-B处理前后的定位(Bars=10 μm)
Figure 5Localization of pSuper1300-RAD21.3-GFP before and after UV-B treatment (Bars=10 μm)
2.3 UV-B辐射后AtRAD21表达量的检测
将WT用5 kJ·m-2的UV-B辐射7天, 迅速提取整株RNA, 并反转录成cDNA; 设计引物, 采用半定量RT-PCR检测AtRAD21.1和AtRAD21.3在UV-B辐射前后表达量的变化。结果表明, UV-B辐射后, AtRAD- 21.1和AtRAD21.3的表达量均降低, 但AtRAD21.3在UV-B辐射后表达量降低更明显(图6)。使用Image J软件进行灰度值分析, 发现AtRAD21.3的表达量在UV-B辐射前后差异显著(P<0.05) (图6B)。说明UV-B辐射对AtRAD21.3表达量的影响更大。图6
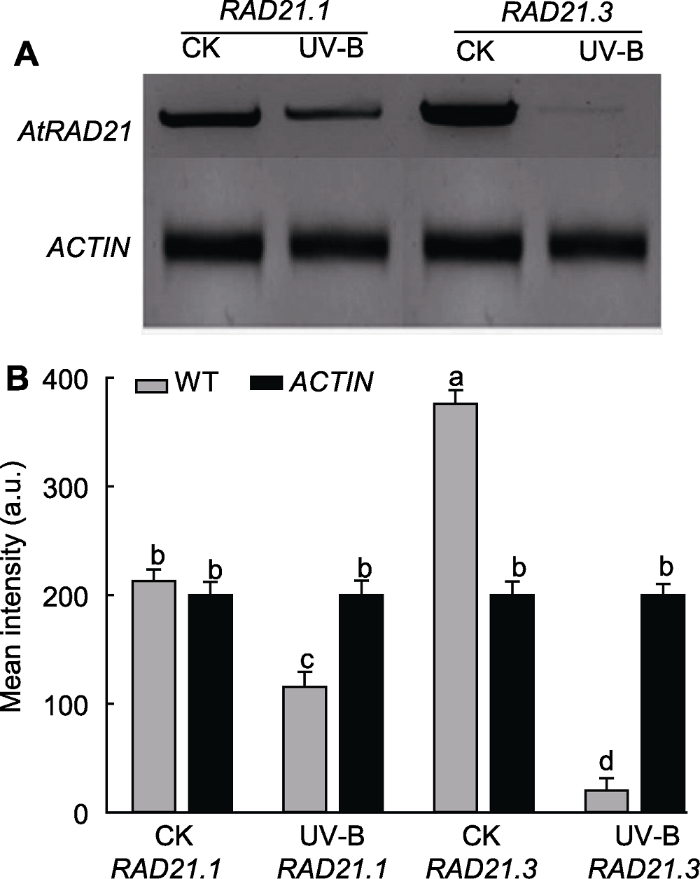 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6UV-B辐射后AtRAD21基因表达量检测
(A) 半定量RT-PCR电泳图; (B) 灰度值分析。WT: 野生型。不同小写字母表示差异显著(Duncan法, P<0.05)。
Figure 6Detection of UV-B radiation on AtRAD21 expres-sion quantity
(A) Result of RT-PCR; (B) Gray value analysis. WT: Wild type. Different lowercase letters show significant differences (Duncan method, P<0.05).
2.4 UV-B辐射对不同株系拟南芥形态及生理生化的影响
2.4.1 UV-B辐射对不同株系拟南芥种子萌发、根长、株高及抽薹的影响为进一步验证观察到的表型差异, 我们设置了对照与UV-B处理组, 统计分析了不同处理对拟南芥WT、atrad21.3突变体以及RAD21.3的过表达植株(35S:RAD21.3-GFP1和35S:RAD21.3-GFP2)的种子萌发率、幼苗根长、株高和抽薹时间的影响。5 kJ·m-2 的UV-B处理后, 所有株系种子的萌发率与CK组相比均降低。与WT CK相比, atrad21.3突变体在UV-B处理后种子的萌发率降低了44%, 并且达显著水平(P<0.05); WT UV-B处理和atrad21.3 CK分别降低了28.46%和22.4% (图7A, E); 而35S:RAD21.3-GFP的萌发率比WT CK稍有增加。由此可知, RAD21.3基因缺失与WT UV-B处理的种子萌发率相近, UV-B处理atrad21.3突变体后萌发率降低更加明显, 过表达植株减缓了UV-B辐射导致的萌发率降低现象。
图7
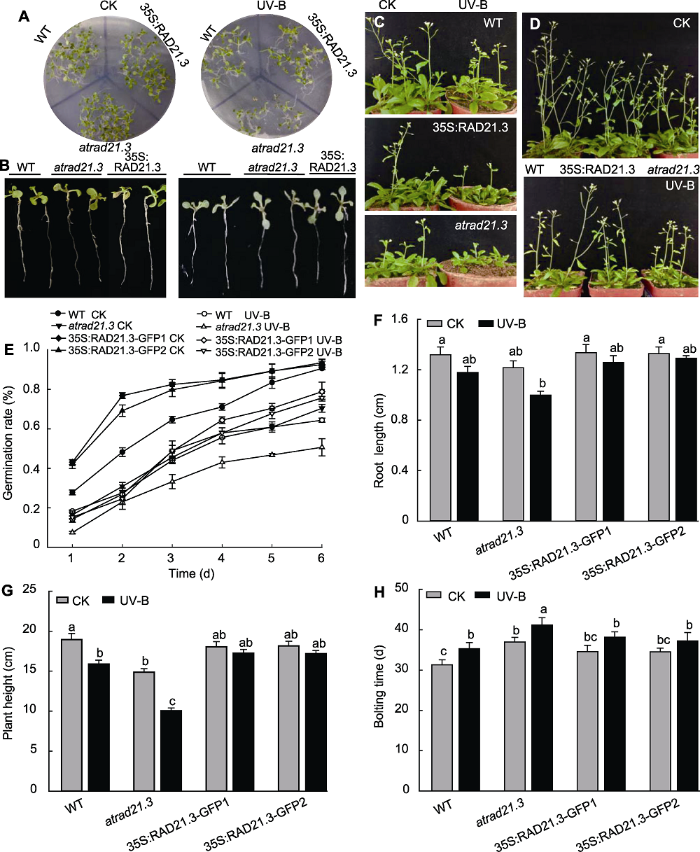 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图7UV-B处理对拟南芥种子萌发率及幼苗根长、株高和抽薹时间的影响
(A), (E) UV-B处理对拟南芥种子萌发率的影响; (B), (F) UV-B处理对拟南芥幼苗根长的影响; (C), (H) UV-B处理对拟南芥抽薹时间的影响; (D), (G) UV-B处理对拟南芥株高的影响。WT: 野生型。不同小写字母表示差异显著(Duncan法, P<0.05)。
Figure 7The seed germination rate, root length, plant height and bolting time of Arabidopsis thaliana with UV-B treatment
(A), (E) The seed germination rate of Arabidopsis thaliana with UV-B treatment; (B), (F) The root length of Arabidopsis thaliana seedling with UV-B treatment; (C), (H) The bolting time of Arabidopsis thaliana with UV-B treatment; (D), (G) The plant height of Arabidopsis thaliana with UV-B treatment. WT: Wild type. Different lowercase letters show significant differences (Duncan method, P<0.05).
此外, UV-B处理后所有株系幼苗的根长与CK组相比均变短(图7B, F)。与WT CK相比, atrad21.3突变体UV-B处理后根长明显变短, 并且达显著水平(P< 0.05); WT UV-B处理和atrad21.3 CK的根长分别为1.178 4和1.216 7 cm; 35S:RAD21.3-GFP CK的根长则较WT CK的长。由此可知, RAD21.3基因缺失与UV-B处理WT后的根长无显著性差异, UV-B处理atrad21.3后加剧了根长变短, 严重阻碍了拟南芥根的生长, 然而过表达株系减缓了这种变化。
此外, UV-B处理后所有株系的株高与CK组相比均变矮(图7D, G)。与WT CK相比, atrad 21.3突变体在UV-B处理后株高降低了46.9%, 且达显著水平(P< 0.05); WT UV-B处理和atrad21.3 CK的株高分别降低了18.83%以及21.5%; 35S:RAD21.3-GFP CK的株高则高于WT CK。由此可知, UV-B处理WT与RAD21.3基因缺失的拟南芥株高相近, atrad21.3 UV-B处理的株高变矮更明显, 但过表达株系的株高变矮现象有所减缓。
有文献报道atrad21.3突变体抽薹延迟(Da Costa- Nunes et al., 2006)。本研究对拟南芥各株系的抽薹情况进行了观察与统计(图7C, H), 表明UV-B处理后所有株系的抽薹与CK 组相比均延迟。与WT CK相比, atrad21.3突变体UV-B处理后抽薹时间延迟了10天, 达显著水平(P<0.05); atrad21.3 CK和WT UV-B处理的抽薹时间分别为36.9和35.2天; 35S:RAD21.3-GFP CK的抽薹时间比WT CK延迟3天。由此可知, UV-B处理WT与RAD21.3基因缺失的拟南芥抽薹时间相近, UV-B处理atrad21.3后其抽薹延迟现象更明显, 过表达株系则减缓了抽薹延迟现象。
2.4.2 UV-B辐射对拟南芥各项生理生化指标的影响
为进一步探究atrad21.3突变体抽薹延迟和株高变矮现象的内在机制, 我们从生理生化水平(包括丙二醛浓度、超氧化物歧化酶活性、叶绿素和可溶性糖含量)探讨了不同处理对拟南芥各生理生化指标的影响。结果表明, UV-B处理后所有株系的MDA浓度与CK组相比均增加。与WT CK相比, atrad21.3突变体UV-B处理后MDA浓度增加了24.5%, 达显著水平(P<0.05); atrad21.3 CK和WT UV-B处理的MDA浓度分别增加了10.3%和6.64%; 35S:RAD21.3-GFP CK降低了10.2% (图8A)。由此可知, RAD21.3基因缺失与WT UV-B处理的膜脂过氧化增加程度相近, 说明RAD21.3基因缺失与WT UV-B处理均导致植株生长受影响, UV-B处理atrad21.3后加剧了膜脂过氧化程度, 使植物生长受影响更加严重, 而过表达株系的膜脂过氧化程度降低。
图8
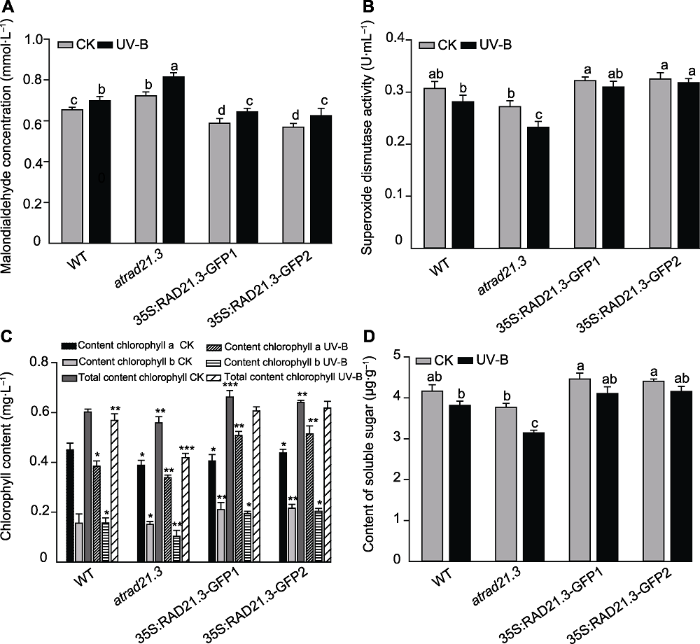 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图8UV-B辐射对拟南芥植株生理生化指标的影响
(A) UV-B辐射对拟南芥丙二醛(MDA)浓度的影响; (B) UV-B辐射对拟南芥超氧化物歧化酶(SOD)活性的影响; (C) UV-B辐射对拟南芥叶绿素含量的影响; (D) UV-B辐射对拟南芥叶片可溶性糖含量的影响。WT: 野生型。不同小写字母(或*)表示差异显著(Duncan法, P<0.05)。
Figure 8The influence of physiological and biochemical in Arabidopsis thaliana with UV-B treatment
(A) Effect of UV-B radiation on Arabidopsis thaliana malondialdehyde (MDA) concentration; (B) Effect of UV-B radiation on Arabidopsis thaliana superoxide dismutase (SOD) activity; (C) Effect of UV-B radiation on chlorophyll content in Arabidopsis thaliana; (D) Effect of UV-B radiation on content of soluble sugar of Arabidopsis thaliana. WT: Wild type. Different lowercase letters (or * ) show significant differences (Duncan method, P<0.05).
UV-B处理后, 所有株系的SOD活性与CK组相比均降低(图8B)。与WT CK相比, atrad21.3突变体UV-B处理后SOD活性降低了24.4%, 达显著水平(P<0.05)。atrad21.3 CK与WT UV-B处理的SOD活性相近, 分别降低了11.4%和8.36%; 35S:RAD21.3-GFP CK的SOD活性比WT CK稍有增加(图8B)。上述结果表明, RAD21.3基因缺失与UV-B处理WT的SOD活性无显著差异, UV-B处理atrad21.3后其SOD活性降低更明显, 过表达植株则减缓此现象。
叶绿素含量表征植物的生长状况。从图8C可以看出, UV-B处理后所有株系的叶绿素含量与CK组相比均下降。与WT CK相比, atrad21.3突变体UV-B处理后叶绿素含量降低了30.5%, 达显著水平(P<0.05); atrad21.3 CK与WT UV-B处理的叶绿素含量相近; 35S:RAD21.3-GFP CK的叶绿素含量比WT CK增加了6.3%, 达显著水平(P<0.05) (图8C)。以上表明RAD21.3基因缺失与UV-B处理WT后的叶绿素含量无差异; UV-B处理atrad21.3后加剧了叶绿素含量的降低, 植株长势弱, 过表达植株则缓解了叶绿素降低现象, 促进了植物的生长。
可溶性糖可以反映植物的代谢程度。从图8D可以看出, UV-B处理后所有株系的可溶性糖含量与CK组相比均降低。与WT CK相比, atrad21.3突变体UV-B辐射后降低了24.6%, 达显著水平(P<0.05); atrad21.3 CK和WT UV-B处理的可溶性糖含量分别降低了9.6%和8.5%; 35S:RAD21.3-GFP CK的可溶性糖含量比WT CK稍有增加(图8D)。由此可知, RAD21.3基因缺失和UV-B处理WT后可溶性糖含量均降低, UV-B处理atrad21.3后可溶性糖含量降低更加明显, 但过表达植株增强了植物的代谢程度。
2.5 UV-B辐射对拟南芥有丝分裂的影响
2.5.1 UV-B辐射对拟南芥细胞周期相关基因表达量的影响通过对对照和UV-B处理7天的WT和atrad21.3突变体进行RNA-seq, 我们筛选出部分细胞有丝分裂的重要调控基因, 分别为CDKA1;1、CDKB2;1、CYCB1;1和CDC5。CDKA1;1主要活跃在细胞有丝分裂的G1和S期, CYCB1;1控制G2至M期的转换, CDKB2;1为植物所特有, 在S期转换至M期时表达(马兰等, 2018)。AtCDC5属于一种细胞周期蛋白, 为生物体进行正常生命活动所必需, 其突变可导致细胞分裂无法正常进行, 植物发育受到抑制(徐金龙等, 2019)。我们将提取的RNA反转录成cDNA, 对细胞周期相关基因进行RT-qPCR检测。结果表明, UV-B处理后细胞周期相关基因的相对表达量与CK组相比均降低。与WT CK的CDKA1;1、CDKB2;1、CYCB1;1和CDC5的相对表达量相比, WT UV-B处理和atrad21.3 CK的基因表达量均降低, 但atrad21.3 UV-B处理组降低更明显, 达显著水平(P<0.05); WT UV-B处理的表达量比atrad21.3 UV-B处理的高。综上, 与WT CK相比, atrad21.3 CK与WT UV-B处理的基因表达量无显著差异, 但atrad21.3 UV-B处理组明显降低, 35S:RAD- 21.3则变化不大。
2.5.2 UV-B辐射对atrad21.3突变体有丝分裂的影响
有丝分裂是亲代细胞的染色体经过复制以后, 精确、平均分配到2个子细胞中去。通过对拟南芥各时期的有丝分裂进行观察, 我们发现正常有丝分裂的间期可见细胞核, 看不到染色体; 前期, 高度螺旋化变成光镜可见的染色体形态, 散乱排列在纺锤体内; 中期, 染色体排列规律; 后期, 染色体均等分裂(图10)。而不同处理的拟南芥体细胞均出现异常有丝分裂现象, 如前期散乱、落后染色体、游离染色体和染色体桥现象(图11)。
图9
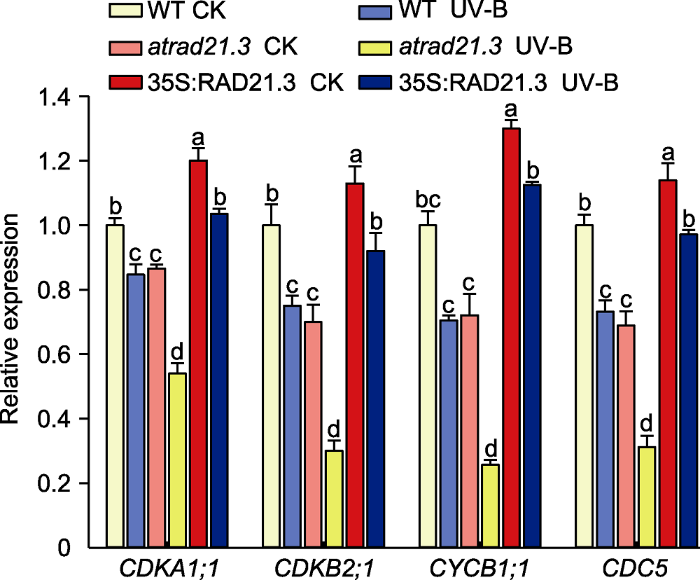 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图9拟南芥细胞周期相关基因的相对表达量
WT: 野生型。不同小写字母表示差异显著(Duncan法, P<0.05)。
Figure 9Relative expression of cell cycle related genes in Arabidopsis thaliana
WT: Wild type. Different lowercase letters show significant differences (Duncan method, P<0.05).
图10
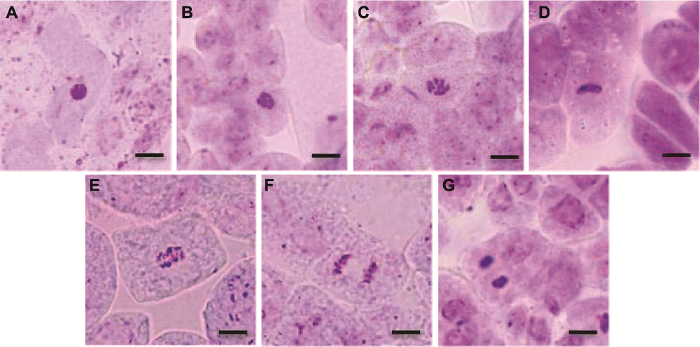 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图10拟南芥根尖正常有丝分裂各时期图
(A) 间期; (B), (C) 前期; (D) 中期; (E) 中后期; (F) 后期; (G) 末期。Bars=20 μm
Figure 10Normal phase types of mitosis in root tip of Arabidopsis thaliana
(A) Interphase; (B), (C) Prophase; (D) Metaphase; (E) Meta-anaphase; (F) Anaphase; (G) Telephase. Bars=20 μm
图11
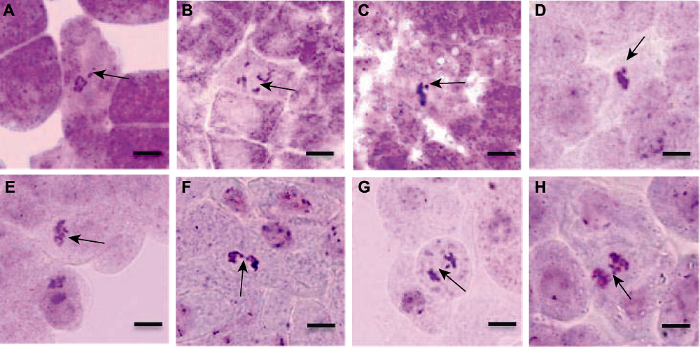 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图11拟南芥根尖染色体畸变的不同类型
(A) 前期不定向(箭头); (B) 前期散乱(箭头); (C) 游离染色体(箭头); (D) 游离染色体(箭头); (E) 后期不对称(箭头); (F) 染色体桥(箭头); (G) 落后染色体(箭头); (H) 落后染色体(箭头)。Bars=20 μm
Figure 11Different types of chromosome aberration in root tip of Arabidopsis thaliana
(A) Unorientation at prophase (arrow); (B) Cofusion at prophase (arrow); (C) Fragments chromosomes (arrow); (D) Fragments chromosomes (arrow); (E) Asymmetric at anaphase (arrow); (F) Chromosome bridge (arrow); (G) Lagging chromosome (arrow); (H) Lagging chromosome (arrow). Bars=20 μm
我们对处于分裂期的细胞进行了统计(表1), 结果表明, 与WT CK相比, WT UV-B处理和atrad21.3 CK均抑制了细胞分裂并且染色体畸变率增加, atrad21.3 UV-B处理的分裂期细胞比率为2.75%, 染色体畸变率为0.638%, 且差异显著(P<0.05)。WT UV-B处理的分裂期细胞比率比atrad21.3 UV-B处理的高, 但是畸变率比atrad21.3 UV-B处理的低。35S:RAD21.3 UV-B处理的畸变率比WT UV-B处理的低。说明UV-B处理或RAD21.3基因缺失均抑制拟南芥细胞的分裂, atrad21.3 UV-B处理加剧了对根尖细胞分裂的抑制且畸变率增加, 但35S:RAD21.3 UV-B处理的畸变率变化不大, 推测RAD21.3对细胞分裂有一定的作用可能与UV-B辐射后细胞出现异常分裂有关。
Table1
表1
表1UV-B辐射对拟南芥有丝分裂的影响
Table1
| Treatment | Total of observing cells | Total of dividing cells | Total of aberration cells | Percentage of dividing cells (%) | Percentage of chromosomal aberration (%) |
|---|---|---|---|---|---|
| WT CK | 5390 | 195 | 8 | 3.6±0.202 a | 0.148±0.102 c |
| WT UV-B | 5376 | 166 | 25 | 3.08±0.258 c | 0.465±0.056 b |
| atrad21.3 CK | 6998 | 215 | 32 | 3.07±0.195 c | 0.457±0.085 b |
| atrad21.3 UV-B | 7044 | 194 | 45 | 2.75±0.182 d | 0.638±0.105 a |
| 35S:RAD21.3 CK | 5603 | 186 | 7 | 3.3±0.132 ab | 0.125±0.136 c |
| 35S:RAD21.3 UV-B | 5536 | 193 | 21 | 3.48±0.129 ab | 0.379±0.125 bc |
新窗口打开|下载CSV
2.6 讨论
真核细胞分裂过程中染色体的准确分离依赖姐妹染色单体内聚力的建立与维持(Chen et al., 2002)。黏着素复合物的主要功能是将DNA拷贝连接在一起并使其不过早分离。RAD21作为黏着素的一个亚基在有丝分裂和修复双链DNA断裂时将姐妹染色单体结合在一起, 在中期至后期转变时被后期类似Caspase的ESP1分离酶(一种半胱氨酸蛋白酶, 与Caspase有较远的亲缘关系)特异切割, 从而调节姐妹染色单体的分离, 使姐妹染色单体分离到纺锤体的两极(Nasmyth et al., 2000), 并确保染色体与子细胞准确分离(Jallepalli et al., 2001)。有丝分裂前期, 黏着素通过不依赖于切割的机制从染色体臂上去除, 导致DNA凝聚成离散的染色单体臂, 并留下少量黏着蛋白与着丝粒染色单体结合(Uhlmann et al., 2000; Warren et al., 2000)。中期, RAD21被ESP1裂解导致复合体解聚, 单个姐妹染色体分开并均匀分配到子细胞中, 由此保证每个子细胞均继承1份基因组的复制本(Uhlmann et al.,1999; Hauf et al., 2001)。后期, 着丝粒上的黏着素被ESP1切割, 触发姐妹染色单体分离(Waizenegger et al., 2000; Hauf et al., 2001)。染色体分离缺陷(Hauf et al., 2001; Rao et al., 2001)以及RAD21分离酶的断裂被多种途径破坏, 均会导致异倍体发生。因此, RAD21作为黏着素蛋白复合体结构完整性的核心,在染色体对称及平均分配到子细胞中具有关键作用(Xu et al., 2014)。
本研究构建了RAD21.3过表达载体, 并通过烟草叶片的瞬时表达和拟南芥转基因植株根尖的观察发现RAD21.3集中在细胞核。设置对照与UV-B处理组, 比较分析了WT UV-B处理与atrad21.3 CK在形态、生理生化及细胞水平的变化。结果表明, WT UV-B处理与atrad21.3 CK的种子萌发率、抽薹时间、株高、MDA浓度、SOD活性、叶绿素和可溶性糖含量均无显著差异, 但atrad21.3 UV-B处理受影响严重, 说明atrad21.3对UV-B辐射敏感, 35S:RAD21.3对UV-B辐射不敏感。WT UV-B处理和atrad21.3 CK株系在生理生化水平上出现差异, 说明植株的生长受到影响。细胞分裂(包括有丝分裂和减数分裂)是生物体生长繁殖的基础, 推测缺少RAD21.3基因及WT在UV-B处理后, 可能影响了体细胞的有丝分裂。我们进一步在细胞水平进行了观察和统计, 使用RNA测序筛选出部分与细胞周期相关的基因, 进行RT-qPCR检测。结果表明, WT UV-B处理与atrad21.3 CK的CDKA1;1、CDKB2;1、CYCB1;1以及CDC5基因的表达量无显著差异, atrad21.3突变体在UV-B处理后的基因表达量明显降低。碱性品红染色并观察拟南芥根尖的有丝分裂, 统计畸变率, 结果表明RAD21.3基因缺失和UV-B处理均导致拟南芥有丝分裂异常。UV-B处理atrad21.3后的有丝分裂异常现象更加严重。推测可能是RAD21.3基因缺失后, 染色体黏连蛋白缺失, 无法使染色体黏连在一起, 致使染色体过早分离, 且未能正确平均分配到子细胞中, 但具体机制有待进一步探索。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
DOI:10.11983/CBB17129URL [本文引用: 1]

通过转基因烟草(Nicotiana tabacum)验证天山雪莲(Saussurea involucrata) Δ9硬脂酰-ACP脱饱和酶基因SiSAD与拟南芥(Arabidopsis thaliana)中同源基因AtFAB2的抗寒性功能。利用农杆菌介导法将植物表达载体PSiSAD:AtFAB2和PSiSAD:SiSAD导入烟草, 然后将2种转基因和野生型烟草分别置于20°C、10°C、5°C、0°C及-2°C下处理2小时, 检测其相对电导率、丙二醛(MDA)含量、叶绿素荧光参数(Fv/Fm)及脂肪酸含量。将-2°C处理2小时后的植株置于25°C培养1周进行生长恢复实验。结果表明, 生长恢复实验中转SiSAD基因烟草的恢复效果显著优于转AtFAB2基因和野生型烟草。在0°C和-2°C处理2小时后, 转SiSAD、AtFAB2基因型和野生型烟草的相对电导率和丙二醛含量呈现显著递增趋势; 转SiSAD、AtFAB2基因型烟草的Fv/Fm显著高于野生型烟草, 其中, 转SiSAD基因烟草的Fv/Fm显著高于转AtFAB2基因烟草。转AtFAB2基因型和野生型烟草的油酸(C18:1)含量随着温度的降低逐渐升高后降低并在0°C时达到最高值; 而转SiSAD基因型烟草C18:1含量持续升高, 并在-2°C时达到最高值, 其含量分别是转AtFAB2基因型和野生型烟草的1.58倍和1.7倍。以上结果表明, 天山雪莲Δ9硬脂酰-ACP脱饱和酶基因SiSAD与拟南芥中同源基因AtFAB2均可以显著增强非低温驯化烟草的抗寒性, 但是SiSAD基因效果显著优于AtFAB2。
[本文引用: 1]
博士论文.
[本文引用: 1]
DOI:10.3724/SP.J.1259.2013.00052URL [本文引用: 1]
以哥伦比亚生态型(Columbia-0)拟南芥(Arabidopsis thaliana)为实验材料, 人工模拟UV-B辐照处理拟南芥种子, 统计其发芽势和发芽率, 测定根长、株高、叶绿素、可溶性糖、可溶性蛋白以及丙二醛(MDA)含量, 研究UV-B辐照处理对拟南芥种子萌发和幼苗生长的影响。研究结果表明, 低剂量的UV-B辐照可以促进拟南芥种子萌发和幼苗生长, 并且最佳辐照剂量为1.0 kJ·m–2(P<0.05), 此时的发芽势、发芽率、根长、株高、叶绿素、可溶性糖及可溶性蛋白的含量均达到最大, 而丙二醛的含量变化则不明显(P>0.05)。当辐照剂量大于1.0 kJ·m–2时, 促进作用逐渐变小, 并且随着辐照剂量的增加, 表现出了抑制作用。实验结果表明, 适当剂量的UV-B辐照在一定程度上可以促进拟南芥种子萌发和幼苗生长, 而过高剂量的辐照则对其产生抑制或损伤作用。
[本文引用: 1]
DOI:10.3969/j.issn.1674-3466.2009.04.003URL [本文引用: 1]
以同步化的拟南芥(Arabidopsis thaliana)根尖细胞为材料, 研究了UV-B辐射对拟南芥细胞周期G1/S期转变的影响。细胞周期荧光显微图像分析表明, UV-B辐射延缓了拟南芥根尖细胞G1/S期的转变。基因表达的RT-PCR检测表明, 在UV-B辐射下G1/S期转变的标志基因Histone H4和E2Fa的表达受抑制, 而G1/S期转变的抑制因子KRP2基因表达则受诱导上调。单细胞凝胶电泳检测结果表明, UV-B辐射引起拟南芥根尖细胞内积累大量的环丁烷嘧啶二聚体。以上结果表明, UV-B辐射抑制植物的生长可能受细胞周期和DNA损伤调控。
[本文引用: 1]
博士论文.
[本文引用: 2]
[本文引用: 1]
DOI:10.1023/A:1009782207376URL [本文引用: 1]
DOI:10.1080/00207239608711082URL [本文引用: 1]
[本文引用: 1]
DOI:10.1105/tpc.113.117473URL [本文引用: 1]
Because of their sessile lifestyle, plants are continuously exposed to solar UV-B radiation. Inhibition of leaf growth is one of the most consistent responses of plants upon exposure to UV-B radiation. In this work, we investigated the role of GROWTH-REGULATING FACTORs (GRFs) and of microRNA miR396 in UV-B-mediated inhibition of leaf growth in Arabidopsis thaliana plants. We demonstrate that miRNA396 is upregulated by UV-B radiation in proliferating tissues and that this induction is correlated with a decrease in GRF1, GRF2, and GRF3 transcripts. Induction of miR396 results in inhibition of cell proliferation, and this outcome is independent of the UV-B photoreceptor UV resistance locus 8, as well as ATM AND RAD3-RELATED and the mitogen-activated protein kinase MPK6, but is dependent on MPK3. Transgenic plants expressing an artificial target mimic directed against miR396 (MIM396) with a decrease in the endogenous microRNA activity or plants expressing miR396-resistant copies of several GRFs are less sensitive to this inhibition. Consequently, at intensities that can induce DNA damage in Arabidopsis plants, UV-B radiation limits leaf growth by inhibiting cell division in proliferating tissues, a process mediated by miR396 and GRFs.
DOI:10.3109/02713683.2011.574332URL [本文引用: 1]
Purpose: Exposure of the cornea to UV radiation from sunlight evokes intraocular inflammation, photokeratitis. Photokeratitis is caused by UVB radiation. It is accompanied by changes of corneal hydration and light absorption. The aim of this study was to examine the effect of two UVB doses on corneal optics in rabbits and to compare these UVB doses with the equivalent exposure of UVB radiation reaching the human cornea from sunlight.
Materials and Methods: Rabbit corneas were irradiated with a daily UVB dose of 0.25 J/cm(2) or 0.5 J/cm(2) for 4 days. One day after finishing the irradiations the rabbits were sacrificed and corneal light absorption measured using our spectrophotometrical method. Corneal hydration was examined using an ultrasonic Pachymeter every experimental day before the irradiation procedure and the last day before sacrificing the animals.
Results: Changes in corneal optics appeared after the repeated exposure of the cornea to a UVB dose of 0.25 J/cm(2) and massively increased after the repeated exposure of the cornea to a UVB dose of 0.5 J/cm(2). The first significant changes in corneal hydration appeared after a single exposure of the cornea to a UVB dose of 0.25 J/cm(2).
Conclusions: Changes in corneal hydration appeared after the exposure of the rabbit cornea to a single UVB dose equivalent to 2.6 hours of solar UVB radiation reaching the human cornea, as measured by UVB sensors embedded in the eyes of mannequin heads facing the sun on a beach at noon in July. Repeated exposure of the rabbit cornea to the same UVB dose evoked profound changes in corneal optics. Although comparison of experimental and outdoor conditions are only approximate, the results in rabbits point to the danger for the human eye from UVB radiation when short stays in sunlight are repeated for several consecutive days without UV protection.
DOI:10.1074/jbc.M201322200URLPMID:11875078 [本文引用: 1]
Caspases are a conserved family of proteases that play a critical role in the execution of apoptosis by cleaving key cellular proteins at Asp residues and modifying their function. Using an expression cloning strategy we recently developed, we isolated human RAD21/SCC1/MCD1 as a novel caspase substrate. RAD21 is a component of the cohesin complex that holds sister chromatids together during mitosis and repairs double-strand DNA breaks. Interestingly, RAD21 is cleaved by a caspase-like Esp1/separase at the onset of anaphase to trigger sister chromatid separation. Here, we demonstrate that human RAD21 is preferentially cleaved at Asp(279) by caspases-3 and -7 in vitro to generate two major proteolytic products of approximately 65 and 48 kDa. Moreover, we show that RAD21 is specifically proteolyzed by caspases into a similarly sized 65-kDa carboxyl-terminal product in cells undergoing apoptosis in response to diverse stimuli. We also demonstrate that caspase proteolysis of RAD21 precedes apoptotic chromatin condensation and has important functional consequences, viz. the partial removal of RAD21 from chromatin and the production of a proapoptotic carboxyl-terminal cleavage product that amplifies the cell death signal. Taken together, these findings point to an entirely novel function of RAD21 in the execution of apoptosis.
URLPMID:16488915 [本文引用: 4]
DOI:10.1186/s12870-014-0353-9URLPMID:25511710 [本文引用: 2]
BACKGROUND: The RAD21 cohesin plays, besides its well-recognised role in chromatid cohesion, a role in DNA double strand break (dsb) repair. In Arabidopsis there are three RAD21 paralog genes (AtRAD21.1, AtRAD21.2 and AtRAD21.3), yet only AtRAD21.1 has been shown to be required for DNA dsb damage repair. Further investigation of the role of cohesins in DNA dsb repair was carried out and is here reported. RESULTS: We show for the first time that not only AtRAD21.1 but also AtRAD21.3 play a role in somatic DNA dsb repair. Comet data shows that the lack of either cohesins induces a similar high basal level of DNA dsb in the nuclei and a slower DNA dsb repair kinetics in both cohesin mutants. The observed AtRAD21.3 transcriptional response to DNA dsb induction reinforces further the role of this cohesin in DNA dsb repair. The importance of AtRAD21.3 in DNA dsb damage repair, after exposure to DNA dsb damage inducing agents, is notorious and recognisably evident at the phenotypical level, particularly when the AtRAD21.1 gene is also disrupted. CONCLUSIONS: Our data demonstrates that both Arabidopsis cohesin (AtRAD21.1 and AtRAD21.3) play a role in somatic DNA dsb repair. Furthermore, the phenotypical data from the atrad21.1 atrad21.3 double mutant indicates that these two cohesins function synergistically in DNA dsb repair. The implications of this data are discussed.
DOI:10.1104/pp.103.030049URLPMID:14681524 [本文引用: 1]
DOI:10.1126/science.1061376URLPMID:11509732 [本文引用: 3]
Cell division depends on the separation of sister chromatids in anaphase. In yeast, sister separation is initiated by cleavage of cohesin by the protease separase. In vertebrates, most cohesin is removed from chromosome arms by a cleavage-independent mechanism. Only residual amounts of cohesin are cleaved at the onset of anaphase, coinciding with its disappearance from centromeres. We have identified two separase cleavage sites in the human cohesin subunit SCC1 and have conditionally expressed noncleavable SCC1 mutants in human cells. Our results indicate that cohesin cleavage by separase is essential for sister chromatid separation and for the completion of cytokinesis.
DOI:10.1093/jxb/erq235URLPMID:20702567 [本文引用: 1]
Plants have evolved a broad spectrum of mechanisms to ensure survival under changing and suboptimal environmental conditions. Alterations of plant architecture are commonly observed following exposure to abiotic stressors. The mechanisms behind these environmentally controlled morphogenic traits are, however, poorly understood. In this report, the effects of a low dose of chronic ultraviolet (UV) radiation on leaf development are detailed. Arabidopsis rosette leaves exposed for 7, 12, or 19 d to supplemental UV radiation expanded less compared with non-UV controls. The UV-mediated decrease in leaf expansion is associated with a decrease in adaxial pavement cell expansion. Elevated UV does not affect the number and shape of adaxial pavement cells, nor the stomatal index. Cell expansion in young Arabidopsis leaves is asynchronous along a top-to-base gradient whereas, later in development, cells localized at both the proximal and distal half expand synchronously. The prominent, UV-mediated inhibition of cell expansion in young leaves comprises effects on the early asynchronous growing stage. Subsequent cell expansion during the synchronous phase cannot nullify the UV impact established during the asynchronous phase. The developmental stage of the leaf at the onset of UV treatment determines whether UV alters cell expansion during the synchronous and/or asynchronous stage. The effect of UV radiation on adaxial epidermal cell size appears permanent, whereas leaf shape is transiently altered with a reduced length/width ratio in young leaves. The data show that UV-altered morphogenesis is a temporal- and spatial-dependent process, implying that common single time point or single leaf zone analyses are inadequate.
DOI:10.1146/annurev.biochem.69.1.115URLPMID:10966455 [本文引用: 1]
The faithful segregation of genetic information requires highly orchestrated changes of chromosome structure during the mitotic cell cycle. The linkage between duplicated sister DNAs is established during S phase and maintained throughout G2 phase (cohesion). In early mitosis, dramatic structural changes occur to produce metaphase chromosomes, each consisting of a pair of compacted sister chromatids (condensation). At anaphase onset, a signal is produced to disrupt the linkage between sister chromatids (separation), allowing them to be pulled apart to opposite poles of the cell. This review discusses our current understanding of the three stages of large-scale structural changes of chromosomes in eukaryotic cells. Recent genetic and biochemical studies have identified key components involved in these processes and started to uncover hitherto unexpected functional links between mitotic chromosome dynamics and other important chromosome functions.
DOI:10.1074/jbc.M206836200URLPMID:12200439 [本文引用: 1]
Scc1/Mcd1 is a component of the cohesin complex that plays an essential role in sister chromatid cohesion in eukaryote cells. Knockout experiments of this gene have been described in budding yeast, fission yeast, and chicken cells, but no study has been reported on human Scc1 thus far. In this study, we found that an N-terminally truncated human Scc1 shows a dominant-negative effect, and we examined the phenotypes of human cells defective in Scc1 function. Scc1 defects led to failure of sister chromatid cohesion in both interphase and mitotic cells. Interestingly, four chromatids derived from two homologues occupied four distinct territories in the nucleus in chromosome painting experiments. In mitotic Scc1-defective cells, chromatids were disjoined with normal condensation, and the spindle-assembly checkpoint was activated. We also found that, although the disjoined kinetochore (half-kinetochore) in Scc1-defective cells contains CENP-A, -B, -C, and -E normally, it apparently does not establish the kinetochore-microtubule association. These results indicate that Scc1 is essential for the association of kinetochores with microtubules.
URLPMID:11371342 [本文引用: 1]
DOI:10.1007/s00425-010-1340-5URL [本文引用: 1]
Even though a number of studies have shown that UV-B radiation inhibits plant growth and regulates the cell cycle progress, little is known about the molecular and cellular mechanisms. Here, we developed a synchronous root-tip cell system to investigate expression changes of cell cycle marker genes and DNA damage under UV-B radiation. Expression analysis of cell cycle marker genes revealed that G1-to-S transition in root-tip cells was accomplished within 6 h. In the in vivo synchronous root-tip cells, high level of UV-B radiation (0.45 W m(-2)) induced expression changes of the cell cycle regulatory genes. Genes involved in G1-to-S transition, Histone H4 and E2Fa, were down-regulated by UV-B radiation during 2-6 h; whereas transcripts for KRP2, a negative regulator of G1-to-S transition, were up-regulated by UV-B at 2 h. The peak time for transcript level of CYCD3;1, a positive factor in G1-to-S transition, was delayed by UV-B radiation. Interestingly, a medium level of UV-B radiation (0.25 W m(-2)) did not change the expression of these genes in root tip cells from wild type. However, cell cycle regulatory genes were greatly affected in uvh1 mutant, which exhibited higher content of cyclobutane pyrimidine dimers (CPDs). Ascorbic acid treatment did not change the expression pattern of cell cycle regulatory genes that were affected by high-level UV-B. Our results implied that UV-B-induced DNA damage results in the delay of G1-to-S transition of plant cell cycle. UV-B-induced G1-to-S arrest may be a protective mechanism that prevents cells with damaged DNA from dividing and may explain the plant growth inhibition under increased solar UV-B radiation.
DOI:10.1111/j.1365-313X.2007.03106.xURLPMID:17488242 [本文引用: 1]
Alpha-kleisins are core components of meiotic and mitotic cohesin complexes. Arabidopsis contains genes for four alpha-kleisin proteins encoded by SYN genes. SYN1, a REC8 ortholog, is essential for meiosis, while SYN2 and SYN4 appear to be SCC1 orthologs and function in mitosis. Our analysis of AtSYN3 shows that it localizes primarily in the nucleolus of both meiotic and mitotic cells. Furthermore, analysis of plants containing an AtSYN3 T-DNA knockout mutation demonstrated that it is essential for megagametogenesis and plays an important role in pollen. These results suggest that SYN3 may not function as part of a typical cohesin complex; rather it may have evolved a specialized role in controlling rDNA structure, transcription or rRNA processing.
[本文引用: 1]
DOI:10.1007/s00412-007-0104-xURL [本文引用: 1]
Cohesin is a multiprotein complex, conserved from yeast to humans, that mediates sister chromatid cohesion. Its ring-shaped structure first suggested that it may perform its task by embracing the sister chromatids. The interaction of cohesin with chromatin is tightly regulated throughout the cell cycle, and several proteins contribute to cohesin loading and mobilization along DNA, establishment of cohesin-mediated cohesion, and removal of cohesin during mitosis. Recent studies suggest that distinct cohesin populations exist in different chromosomal regions and have particular requirements in their dynamic interaction with chromatin. In this review, I briefly summarize these studies and discuss their implications for current and future models of cohesin behavior.
DOI:10.1126/science.288.5470.1379URLPMID:10827941 [本文引用: 2]
In eukaryotic cells, sister DNA molecules remain physically connected from their production at S phase until their separation during anaphase. This cohesion is essential for the separation of sister chromatids to opposite poles of the cell at mitosis. It also permits chromosome segregation to take place long after duplication has been completed. Recent work has identified a multisubunit complex called cohesin that is essential for connecting sisters. Proteolytic cleavage of one of cohesin's subunits may trigger sister separation at the onset of anaphase.
URLPMID:9576786 [本文引用: 1]
DOI:10.1038/35073627URLPMID:11309624 [本文引用: 1]
Cohesion between sister chromatids is established during DNA replication and depends on a protein complex called cohesin. At the metaphase-anaphase transition in the yeast Saccharomyces cerevisiae, the ESP1-encoded protease separin cleaves SCC1, a subunit of cohesin with a relative molecular mass of 63,000 (Mr 63K). The resulting 33K carboxy-terminal fragment of SCC1 bears an amino-terminal arginine-a destabilizing residue in the N-end rule. Here we show that the SCC1 fragment is short-lived (t1/2 approximately 2 min), being degraded by the ubiquitin/proteasome-dependent N-end rule pathway. Overexpression of a long-lived derivative of the SCC1 fragment is lethal. In ubr1Delta cells, which lack the N-end rule pathway, we found a highly increased frequency of chromosome loss. The bulk of increased chromosome loss in ubr1Delta cells is caused by metabolic stabilization of the ESP1-produced SCC1 fragment. This fragment is the first physiological substrate of the N-end rule pathway that is targeted through its N-terminal residue. A number of yeast proteins bear putative cleavage sites for the ESP1 separin, suggesting other physiological substrates and functions of the N-end rule pathway.
DOI:10.1111/pce.12374URLPMID:24890713 [本文引用: 1]
There is a need to reappraise the effects of UV-B radiation on plant morphology in light of improved mechanistic understanding of UV-B effects, particularly elucidation of the UV RESISTANCE LOCUS 8 (UVR8) photoreceptor. We review responses at cell and organismal levels, and explore their underlying regulatory mechanisms, function in UV protection and consequences for plant fitness. UV-induced morphological changes include thicker leaves, shorter petioles, shorter stems, increased axillary branching and altered root:shoot ratios. At the cellular level, UV-B morphogenesis comprises changes in cell division, elongation and/or differentiation. However, notwithstanding substantial new knowledge of molecular, cellular and organismal UV-B responses, there remains a clear gap in our understanding of the interactions between these organizational levels, and how they control plant architecture. Furthermore, despite a broad consensus that UV-B induces relatively compact architecture, we note substantial diversity in reported phenotypes. This may relate to UV-induced morphological changes being underpinned by different mechanisms at high and low UV-B doses. It remains unproven whether UV-induced morphological changes have a protective function involving shading and decreased leaf penetration of UV-B, counterbalancing trade-offs such as decreased photosynthetic light capture and plant-competitive abilities. Future research will need to disentangle seemingly contradictory interactions occurring at the threshold UV dose where regulation and stress-induced morphogenesis overlap.
URLPMID:10623634 [本文引用: 1]
DOI:10.1007/s004420000592URLPMID:28547159 [本文引用: 1]
The potential effects of increased ultraviolet-B radiation (UV-B, 280-320 nm) simulating stratospheric ozone depletion in field studies with vascular plants have previously been summarized only in narrative literature reviews. In this quantitative synthesis, we have assessed the significance of solar UV-B enhancement for ten commonly measured variables involving leaf pigmentation, plant growth and morphology, and photosynthesis using meta-analytic statistical methods. Of 103 papers published between 1976 and mid-1999 from field studies, more than 450 reports from 62 papers were included in the database. Effects of UV-B were most apparent for the case of UV-B-absorbing compounds with an average increase of approximately 10% across all studies when comparing the ambient solar UV-B control to the treatment (involving ambient UV-B plus a UV-B supplement from special UV lamps). Some morphological parameters such as plant height and leaf mass per area showed little or no response to enhanced UV-B. Leaf photosynthetic processes (leaf gas exchange and chlorophyll fluorescence) and the concentration of photosynthetic pigments (total chlorophylls and carotenoids) were also not affected. Shoot biomass and leaf area showed modest decreases under UV-B enhancement. The reduction in shoot biomass occurred only under very high levels of simulated ozone depletion and leaf area was affected only when studies inappropriately used the plant (i.e., the subreplicate) rather than the plot as the experimental replicate. To the best of our knowledge, this review provides the first quantitative estimates of UV-B effects in field-based studies using all suitable published studies as a database.
DOI:10.1016/j.cub.2009.10.067URLPMID:20064425 [本文引用: 1]
DOI:10.1016/j.pbi.2003.09.009URLPMID:14611952 [本文引用: 1]
Cells undergoing endoreduplication replicate chromosomal DNA without intervening mitoses. The resulting larger, higher-ploidy nucleus is often associated with an increase in cell size, but the molecular basis for this correlation remains poorly understood. Recent advances in characterising various mutants and transgenic plants are beginning to unravel how this unique type of cell cycling is regulated and how it contributes to cell-size control. Both cell growth (i.e. increase in cytoplasmic macromolecular mass) and cell expansion (i.e. increase in cell volume through vacuolation) contribute independently to increases in cell size in plants. A total organ-size checkpoint may also help to coordinate cell size and cell number within an organ, and can contribute to final cell-size determination in plants.
DOI:10.1016/j.gene.2012.11.012URL [本文引用: 1]
Onion can be used in experimental observation of mitotic cell division in plant science because its chromosome is large and easy to observe. However, molecular genetic studies are difficult in onion because of its large genome size, and only limited information of onion genes has been available to date. Here we cloned and characterized an onion homologue of mitotic RAD21 gene, AcRAD21-1, to develop a molecular marker of mitosis. The N-terminal, middle, and C-terminal regions of deduced AcRAD21-1 protein sequence were conserved with Arabidopsis SYN4/AtRAD213 and rice OsRAD21-1, whereas three characteristic types of repetitive motifs (Repeat-1, Repeat-2/2', and Repeat-3) were observed between the conserved regions. Such inserted repetitive amino acid sequences enlarge the AcRAD21-1 protein into almost 200 kDa, which belongs to the largest class of plant proteins. Genomic organization of the AcRAD21-1 locus was also determined, and the possibility of tandem exon duplication in Repeat-2 was revealed. Subsequently, the polyclonal antiserum was raised against the N-terminal region of AcRAD21-1, and purified by affinity chromatography. Immunohistochemical analysis with the purified antibody successfully showed localization of AcRAD21-1 in onion mitosis, suggesting that it can be used as a molecular marker visualizing dynamic movement of cohesin. (C) 2012 Elsevier B.V.
DOI:10.1038/21831URLPMID:10403247 [本文引用: 1]
Cohesion between sister chromatids is established during DNA replication and depends on a multiprotein complex called cohesin. Attachment of sister kinetochores to the mitotic spindle during mitosis generates forces that would immediately split sister chromatids were it not opposed by cohesion. Cohesion is essential for the alignment of chromosomes in metaphase but must be abolished for sister separation to start during anaphase. In the budding yeast Saccharomyces cerevisiae, loss of sister-chromatid cohesion depends on a separating protein (separin) called Esp1 and is accompanied by dissociation from the chromosomes of the cohesion subunit Scc1. Here we show that Esp1 causes the dissociation of Scc1 from chromosomes by stimulating its cleavage by proteolysis. A mutant Scc1 is described that is resistant to Esp1-dependent cleavage and which blocks both sister-chromatid separation and the dissociation of Scc1 from chromosomes. The evolutionary conservation of separins indicates that the proteolytic cleavage of cohesion proteins might be a general mechanism for triggering anaphase.
DOI:10.1016/s0092-8674(00)00130-6URLPMID:11081625 [本文引用: 1]
In eukaryotic cells, replicated DNA strands remain physically connected until their segregation to opposite poles of the cell during anaphase. This
DOI:10.1016/j.plantsci.2017.12.005URLPMID:29362084 [本文引用: 1]
Ultraviolet B light (UV-B, 280-315nm) is the shortest wavelength of the solar spectrum reaching the surface of the Earth. It has profound effects on plants, ranging from growth regulation to severe metabolic changes. Low level UV-B mainly causes photomorphogenic effects while higher levels can induce stress, yet these effects tend to overlap. Here we identified a condition that allows growth reduction without obvious detrimental stress in wild type Arabidopsis rosette plants. This condition was used to study the effects of a daily UV-B dose on plant characteristics of UV-B adapted plants in detail. Exploration of the transcriptome of developing leaves indicated downregulation of genes involved in stomata formation by UV-B, while at the same time genes involved in photoprotective pigment biosynthesis were upregulated. These findings correspond with a decreased stomatal density and increased UV-B absorbing pigments. Gene ontology analysis revealed upregulation of defense related genes and meta-analysis showed substantial overlap of the UV-B regulated transcriptome with transcriptomes of salicylate and jasmonate treated as well as herbivore exposed plants. Feeding experiments showed that caterpillars of Spodoptera littoralis are directly affected by UV-B, while performance of the aphid Myzus persicae is diminished by a plant mediated process.
DOI:10.1016/s0092-8674(00)00132-xURLPMID:11081627 [本文引用: 1]
In yeast, anaphase depends on cohesin cleavage. How anaphase is controlled in vertebrates is unknown because their cohesins dissociate from chromosomes before anaphase. We show that residual amounts of the cohesin SCC1 remain associated with human centromeres until the onset of anaphase when a similarly small amount of SCC1 is cleaved. In Xenopus extracts, SCC1 cleavage depends on the anaphase-promoting complex and separin. Separin immunoprecipitates are sufficient to cleave SCC1, indicating that separin is associated with a protease activity. Separin activation coincides with securin destruction and partial separin cleavage, suggesting that several mechanisms regulate separin activity. We propose that in vertebrates, a cleavage-independent pathway removes cohesin from chromosome arms during prophase, whereas a separin-dependent pathway cleaves centromeric cohesin at the metaphase-anaphase transition.
URLPMID:11102811 [本文引用: 1]
DOI:10.1038/sj.embor.7400121URLPMID:15031719 [本文引用: 1]
The evolutionarily conserved cohesin complex is required for the establishment and maintenance of sister chromatid cohesion, in turn essential for proper chromosome segregation. RAD21/SCC1 is a regulatory subunit of the mitotic cohesin complex, as it links together all other subunits of the complex. The destruction of RAD21/SCC1 along chromosomal arms and later at centromeres results in the dissociation of the cohesin complex, facilitating chromosome segregation. Here, we report for the first time that mammalian RAD21/SCC1 associates with the axial/lateral elements of the synaptonemal complex along chromosome arms and on centromeres of mouse spermatocytes. Importantly, RAD21/SCC1 is lost from chromosome arms in late prophase I but persists on centromeres. The loss of centromeric RAD21/SCC1 coincides with the separation of sister chromatids at anaphase II. These findings support a role for mammalian RAD21/SCC1 in maintaining sister chromatid cohesion in meiosis.
URLPMID:25464844 [本文引用: 1]
植物响应UV-B辐射的研究进展
1
2015
... 控制细胞分裂和扩张是调节器官大小的核心(
天山雪莲SiSAD基因与拟南芥AtFAB2基因转化烟草的抗寒性分析
1
2018
... 控制细胞分裂和扩张是调节器官大小的核心(
UV-B辐射增强影响作物生长发育的研究进展
1
2018
... 控制细胞分裂和扩张是调节器官大小的核心(
He-Ne激光对小麦增强UV-B辐射损伤的修复效应及机理
1
2002
... 控制细胞分裂和扩张是调节器官大小的核心(
UV-B辐射对拟南芥种子萌发和幼苗生长的影响
1
2013
... 植物在胁迫条件下会表现出形态和生理生化变化.本研究统计了UV-B辐射后, WT、atrad21.3和过表达株系的种子萌发率(
拟南芥突变体L1.3的表型分析及遗传定位
1
2018
... 通过对对照和UV-B处理7天的WT和atrad21.3突变体进行RNA-seq, 我们筛选出部分细胞有丝分裂的重要调控基因, 分别为CDKA1;1、CDKB2;1、CYCB1;1和CDC5.CDKA1;1主要活跃在细胞有丝分裂的G1和S期, CYCB1;1控制G2至M期的转换, CDKB2;1为植物所特有, 在S期转换至M期时表达(
UV-B辐射对拟南芥细胞周期G1/S期转变的影响
1
2009
... 控制细胞分裂和扩张是调节器官大小的核心(
拟南芥细胞周期基因AtCDC5的功能研究及抗体制备
1
2019
... 通过对对照和UV-B处理7天的WT和atrad21.3突变体进行RNA-seq, 我们筛选出部分细胞有丝分裂的重要调控基因, 分别为CDKA1;1、CDKB2;1、CYCB1;1和CDC5.CDKA1;1主要活跃在细胞有丝分裂的G1和S期, CYCB1;1控制G2至M期的转换, CDKB2;1为植物所特有, 在S期转换至M期时表达(
水稻RAD21/REC8家族基因的分离与功能分析
2
2006
... 细胞分裂是生物体生长繁殖的基础, 真核生物的细胞分裂包括有丝分裂和减数分裂(
... ).免疫荧光定位分析表明, OsRad21-1和OsRad21-3特异地定位于有丝分裂染色体上, 可能参与有丝分裂姐妹染色体之间的黏着(
1
2009
... 植物在胁迫条件下会表现出形态和生理生化变化.本研究统计了UV-B辐射后, WT、atrad21.3和过表达株系的种子萌发率(
The effects of UV-B radiation on European heathland species
1
1997
... 紫外线辐射是太阳电磁辐射的一部分, 到达地球表面的太阳辐射约有7%是紫外线(
Effects of ozone depletion and increased UV-B on terrestrial ecosystems
1
1996
... 紫外线辐射是太阳电磁辐射的一部分, 到达地球表面的太阳辐射约有7%是紫外线(
Effects of increased solar ultraviolet radiation on terrestrial plants
1
1995
... 紫外线辐射是太阳电磁辐射的一部分, 到达地球表面的太阳辐射约有7%是紫外线(
Repression of growth regulating factors by the microRNA396 inhibits cell proliferation by UV-B radiation in Arabidopsis leaves
1
2013
... 控制细胞分裂和扩张是调节器官大小的核心(
Hydration and transparency of the rabbit cornea irradiated with UVB-doses of 0.25 J/cm2 and 0.5 J/cm2 compared with equivalent UVB radiation exposure reaching the human cornea from sunlight
1
2011
... 紫外线辐射是太阳电磁辐射的一部分, 到达地球表面的太阳辐射约有7%是紫外线(
Caspase proteolysis of the cohesin component RAD21 promotes apoptosis
1
2002
... 真核细胞分裂过程中染色体的准确分离依赖姐妹染色单体内聚力的建立与维持(
Characterization of the three Arabidopsis thaliana RAD21 cohesins reveals differential responses to ionizing radiation
4
2006
... 细胞分裂是生物体生长繁殖的基础, 真核生物的细胞分裂包括有丝分裂和减数分裂(
... ), AtRAD21.1与AtRAD21.3可能代表有丝分裂凝聚蛋白, 但在有丝分裂中功能冗余(
... 有研究表明, AtRAD21.3在染色体结构维持中的作用可以解释拟南芥发育延迟现象, 特别是atrad21.3突变体延迟抽薹表型(
... 有文献报道atrad21.3突变体抽薹延迟(
The At- RAD21.1 and AtRAD21.3 Arabidopsis cohesins play a synergistic role in somatic DNA double strand break damage repair
2
2014
... 细胞分裂是生物体生长繁殖的基础, 真核生物的细胞分裂包括有丝分裂和减数分裂(
... 突变体的作用已有报道(
Ultraviolet-B radiation- mediated responses in plants. Balancing damage and protection
1
2003
... 紫外线辐射是太阳电磁辐射的一部分, 到达地球表面的太阳辐射约有7%是紫外线(
Cohesin cleavage by separase required for anaphase and cytokinesis in human cells
3
2001
... 有丝分裂前期, 黏着素通过不依赖于切割的机制从染色体臂上去除, 导致DNA凝聚成离散的染色单体臂, 并留下少量黏着蛋白与着丝粒染色单体结合(
... ;
... ).染色体分离缺陷(
UV radiation reduces epidermal cell expansion in leaves of Arabidopsis thaliana
1
2010
... 控制细胞分裂和扩张是调节器官大小的核心(
Chromosome cohesion, condensation, and separation
1
2000
... 细胞分裂是生物体生长繁殖的基础, 真核生物的细胞分裂包括有丝分裂和减数分裂(
Cohesin defects lead to premature sister chromatid separation, kinetochore dysfunction, and spindle-assembly checkpoint activation
1
2002
... 细胞分裂是生物体生长繁殖的基础, 真核生物的细胞分裂包括有丝分裂和减数分裂(
Securin is required for chromosomal stability in human cells
1
2001
... 真核细胞分裂过程中染色体的准确分离依赖姐妹染色单体内聚力的建立与维持(
UV-B-induced DNA damage mediates expression changes of cell cycle regulatory genes in Arabidopsis root tips
1
2011
... 控制细胞分裂和扩张是调节器官大小的核心(
The Arabidopsis cohesin protein SYN3 localizes to the nucleolus and is essential for gametogenesis
1
2007
... 细胞分裂是生物体生长繁殖的基础, 真核生物的细胞分裂包括有丝分裂和减数分裂(
Effects of ultraviolet-B radiation on primary photophysical process in photosystem II: a fluorescence spectrum analysis. In: Kuang TY, Lu CM, Zhang LX, eds. Photosynthesis Research for Food, Fuel and the Future
1
... 紫外线辐射是太阳电磁辐射的一部分, 到达地球表面的太阳辐射约有7%是紫外线(
Cohesin regulation: fashionable ways to wear a ring
1
2007
... 细胞分裂是生物体生长繁殖的基础, 真核生物的细胞分裂包括有丝分裂和减数分裂(
Splitting the chromosome: cutting the ties that bind sister chromatids
2
2000
... 细胞分裂是生物体生长繁殖的基础, 真核生物的细胞分裂包括有丝分裂和减数分裂(
... 真核细胞分裂过程中染色体的准确分离依赖姐妹染色单体内聚力的建立与维持(
Ultraviolet-B radiation effects on water relations, leaf development, and photosynthesis in droughted pea plants
1
1998
... 紫外线辐射是太阳电磁辐射的一部分, 到达地球表面的太阳辐射约有7%是紫外线(
Degradation of a cohesin subunit by the N-end rule pathway is essential for chromosome stability
1
2001
... 有丝分裂前期, 黏着素通过不依赖于切割的机制从染色体臂上去除, 导致DNA凝聚成离散的染色单体臂, 并留下少量黏着蛋白与着丝粒染色单体结合(
Re-interpreting plant morphological responses to UV-B radiation
1
2015
... 控制细胞分裂和扩张是调节器官大小的核心(
NXP-1, a human protein related to Rad21/Scc1/ Mcd1, is a component of the nuclear matrix
1
2000
... 细胞分裂是生物体生长繁殖的基础, 真核生物的细胞分裂包括有丝分裂和减数分裂(
A meta-analysis of plant field studies simulating stratospheric ozone depletion
1
2001
... 紫外线辐射是太阳电磁辐射的一部分, 到达地球表面的太阳辐射约有7%是紫外线(
Establishment of sister chromatid cohesion
1
2009
... 细胞分裂是生物体生长繁殖的基础, 真核生物的细胞分裂包括有丝分裂和减数分裂(
‘Big it up’: endoreduplication and cell-size control in plants
1
2003
... 紫外线辐射是太阳电磁辐射的一部分, 到达地球表面的太阳辐射约有7%是紫外线(
Cellular localization of mitotic RAD21 with repetitive amino acid motifs in Allium cepa
1
2013
... 细胞分裂是生物体生长繁殖的基础, 真核生物的细胞分裂包括有丝分裂和减数分裂(
Sister- chromatid separation at anaphase onset is promoted by cleavage of the cohesin subunit Scc1
1
1999
... 有丝分裂前期, 黏着素通过不依赖于切割的机制从染色体臂上去除, 导致DNA凝聚成离散的染色单体臂, 并留下少量黏着蛋白与着丝粒染色单体结合(
Cleavage of cohesin by the CD clan protease separin triggers anaphase in yeast
1
2000
... 有丝分裂前期, 黏着素通过不依赖于切割的机制从染色体臂上去除, 导致DNA凝聚成离散的染色单体臂, 并留下少量黏着蛋白与着丝粒染色单体结合(
An ultraviolet B condition that affects growth and defense in Arabidopsis
1
2018
... 紫外线辐射是太阳电磁辐射的一部分, 到达地球表面的太阳辐射约有7%是紫外线(
Two distinct pathways remove mammalian cohesin from chromosome arms in prophase and from centromeres in ana- phase
1
2000
... 有丝分裂前期, 黏着素通过不依赖于切割的机制从染色体臂上去除, 导致DNA凝聚成离散的染色单体臂, 并留下少量黏着蛋白与着丝粒染色单体结合(
The Drosophila RAD21 cohesin persists at the centromere region in mitosis
1
2000
... 有丝分裂前期, 黏着素通过不依赖于切割的机制从染色体臂上去除, 导致DNA凝聚成离散的染色单体臂, 并留下少量黏着蛋白与着丝粒染色单体结合(
A new role for the mitotic RAD21/ SCC1 cohesin in meiotic chromosome cohesion and segregation in the mouse
1
2004
... 细胞分裂是生物体生长繁殖的基础, 真核生物的细胞分裂包括有丝分裂和减数分裂(
Cohesin rad21 mediates loss of heterozygosity and is upregulated via Wnt promoting transcriptional dysregulation in gastrointestinal tumors
1
2014
... 有丝分裂前期, 黏着素通过不依赖于切割的机制从染色体臂上去除, 导致DNA凝聚成离散的染色单体臂, 并留下少量黏着蛋白与着丝粒染色单体结合(
备案号: 京ICP备16067583号-21
版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由北京玛格泰克科技发展有限公司设计开发

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}