Phylogenetic Study of Amaranthaceae sensu lato Based on Multiple Plastid DNA Fragments
Jiuxiang Huang1, Wenna Chen2, Yuling Li1, Gang Yao,1,*1South China Limestone Plants Research Center, College of Forestry and Landscape Architecture, South China Agricultural University, Guangzhou 510642, China 2College of Landscape and Ecological Engineering, Hebei University of Engineering, Handan 056038, China
Abstract Amaranthaceae sensu lato, including Amaranthaceae sensu stricto and Chenopodiaceae, is the second largest family in Caryophyllales. However, the family status of Chenopodiaceae is disputable and phylogenetic relationships among all of the subfamilies circumscribed within Amaranthaceae s.l. have not been well resolved to date. In the present study, phylogeny of Amaranthaceae s.l. was reconstructed based on a comprehensive taxonomic sampling of all 13 subfamilies (59 species) circumscribed using eight plastid DNA fragments, and a molecular dating analysis of the family was also conducted. Results revealed that, the monophyly of Amaranthaceae both in broad and narrow sense was strongly supported, but the monophyly of Chenopodiaceae was rejected. Thus the concept of Amaranthaceae s.l. is accepted here. Phylogenetic positions of all subfamilies were all resolved with strong support values, except that the position of the subfamily Polycnemoideae was weakly supported. Additionally, the crown age of Amaranthaceae s.l. was estimated at ca. 69.9 million years ago (Ma) in the late Cretaceous, and a period of rapid divergence may have occurred near the Cretaceous-Paleogene (K-Pg) boundary (ca. 66.0 Ma). Keywords:Amaranthaceae;Caryophyllales;molecular dating;phylogeny
PDF (1913KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 黄久香, 陈文娜, 李玉玲, 姚纲. 基于多个叶绿体基因序列片段重建广义苋科系统发育关系. 植物学报, 2020, 55(4): 457-467 doi:10.11983/CBB19228 Huang Jiuxiang, Chen Wenna, Li Yuling, Yao Gang. Phylogenetic Study of Amaranthaceae sensu lato Based on Multiple Plastid DNA Fragments. Chinese Bulletin of Botany, 2020, 55(4): 457-467 doi:10.11983/CBB19228
黑色圆点表示时间键准点所在位置。空心箭头示广义苋科冠群节点。Ach: 牛膝亚科; Aer: 白花苋亚科; Ama: 苋亚科; Bet: 甜菜亚科; Cam: 樟味藜亚科; Cel: 青葙亚科; Che: 藜亚科; Cor: 虫实亚科; Gom: 千日红亚科; Pol: 多节草亚科; Sali: 盐角草亚科; Sals: 猪毛菜亚科; Sua: 碱蓬亚科 Figure 2Chronogram based on the BEAST analysis of the combined matrix of eight plastid DNA regions and four calibrations
Calibration points are depicted with black circles. The crown node of Amaranthaceae sensu lato is shown by a hollow arrowhead. Ach: Achyranthoideae; Aer: Aervoideae; Ama: Amaranthoideae; Bet: Betoideae; Cam: Camphorosmoideae; Cel: Celosioideae; Che: Chenopodioideae; Cor: Corispermoideae; Gom: Gomphrenoideae; Pol: Polycnemoideae; Sali: Salicornioideae; Sals: Salsoloideae; Sua: Suaedoideae
在Walker等(2018)基于转录组数据构建的石竹目系统发育关系中, 传统界定的藜科(含多节草亚科)形成一个单系分支, 且与狭义苋科成姐妹群关系。该研究还表明, 在藜科系统关系拓扑结构的主干靠基部连续3个节点, 均有约75%的基因树并不支持所呈现的拓扑结构, 这说明该研究获取的转录组数据中相关基因之间系统发育信号冲突明显, 因此所呈现的拓扑结构可能并未揭示出相关类群真实的进化关系。此外, 在Yang等(2018)基于转录组数据所构建的石竹目系统发育关系中, 猪笼草科与茅膏菜科关系较近, 而与露松科关系较远, 且相关节点均获得很高的支持率, 这与基于叶绿体基因组数据所得结果一致(Yao et al., 2019)。但Walker等(2018)的研究结果则支持猪笼草科与露松科关系更近, 与茅膏菜科关系相对较远, 相关节点支持率同样很高。在基于转录组数据的系统发育分析中, 猪笼草科、茅膏菜科及露松科之间的系统关系在不同研究中出现明显冲突, 可能与相关研究在转录组数据分析背景下所筛选出来用于系统发育树构建的“直系同源基因”存在差异有关, 这意味着基于转录组数据筛选出的大量“直系同源基因”所构建的系统发育关系仍然值得商榷。另一方面, 在系统发育分析中, 基于叶绿体基因序列数据所得结果与核基因序列数据所得结果存在冲突的先例在被子植物类群中普遍存在, 这一现象通常被认为可能与杂交、叶绿体捕获以及不完全的谱系分选等进化事件有关(Gurushidze et al., 2010; Yi et al., 2015; Guo et al., 2018)。因此, 阐释广义苋科中基于叶绿体数据与核基因数据所得结果存在冲突的原因, 将是后期研究中需要关注和探讨的重要问题。
在传统藜科的分类处理中, Scott (1977a, 1977b)还曾建议将猪毛菜科(Salsolaceae Moq.)和盐角草科(Salicorniaceae J. Agardh)等类群提升至科的水平, 但并未被后来****承认, 相关类群在本研究中被证实仍然属于广义苋科的范畴, 且嵌套在该科内部。此外, 曾被置于藜科的肉叶刺茎藜属(Sarcobatus Nees)以及滨藜叶属(Halophytum Speg.)均从藜科中分出并独立成科, 且与广义苋科系统位置较远。其中肉叶刺茎藜科(Sarcobataceae)隶属石竹目商陆分支(Phytolaccoid clade), 与萝卜藤科(Agdestidaceae)成姐妹群关系(Walker et al., 2018; Yao et al., 2019); 而滨藜叶科(Halophytaceae)则隶属石竹目马齿苋亚目分支(Portulacineae clade), 与龙树科-落葵科分支(Basellaceae-Didiereaceae clade)成姐妹群关系(Yao et al., 2019)。
赤珠苋属共3种, 主要分布于密克罗尼西亚群岛、塞浦路斯及西喜马拉雅地区; 炬苋树属共6种, 主要分布于夏威夷及土布艾群岛(Stevens, 2001)。然而在亚科水平位置的归属方面, 这2属还有待确定。本研究得出的系统发育关系表明, 赤珠苋属与炬苋树属在广义苋科之中均占据独立的系统位置并在狭义苋科基部依次分出, 且相关节点均获得很高的支持(图1)。从生活型上, 赤珠苋属与炬苋树属均为木本类型, 而狭义苋科则以多年生草本为主, 仅有极少数类群为木本(如青葙族(Celosieae)及鸽苋属(Chamissoa Kunth)类群之中的大部分物种) (Müller and Borsch, 2005)。赤珠苋属果实为浆果, 这与狭义苋科浆果苋属(Deeringia R. Br.)和多脉苋属(Pleuropetalum Hook. f.)等类群相似, 而狭义苋科其它类群及传统藜科类群则以胞果为主(Müller and Borsch, 2005)。因此, 基于本研究得出的系统关系, 结合生活型及形态特征等多方面证据, 我们认为赤珠苋属与炬苋树属都有必要提升为独立的亚科。
2.4.3 苋科及其主要分支的起源与分化
对于广义苋科的起源时间, 前人多项研究均有涉及。Wikstr?m等(2001)研究表明, 该科干群节点起源可能发生在约28、38或40 Ma; 而Kadereit等(2012)认为该科干群起源时间发生在87-47 Ma; Magallón等(2015)则认为该科干群节点可能发生在64.2 Ma (81.8-43.6 Ma)或76.4 Ma (80.5-72.0 Ma)。本研究表明, 该科干群节点起源可能发生在白垩纪晚期约80.5 Ma (82.5-78.3 Ma)。在广义苋科中, 目前已知最早的可信化石记录为Polyporina cribraria Srivastava, 该化石物种为发现于白垩纪晚期马斯里奇特阶(Maestrichtian, 72.1-66.0 Ma)的花粉化石。在形态特征上, 该化石与传统藜科类群(尤其是现存物种Salicornia australis Sol. ex F. Muell.)的花粉化石非常相似。该化石发现于加拿大海洋环境向陆地环境的过渡地带, 与现存众多藜科类群的生境也非常相似。此外, 自马斯里奇特阶之后在相似生境中也发现过一系列类似的花粉化石(Muller, 1981; Kadereit et al., 2003)。基于形态特征及其发现地的生境特征, 该花粉化石被广泛接受为广义苋科类群并多次用于该科类群分化时间校准分析(Kadereit et al., 2005; Yao et al., 2019; Li et al., 2019)。因此, 从化石证据角度看, Wikstr?m等(2001)所得结果在时间尺度上明显偏早。而本研究结果在时间区间上与Kadereit等(2012)及Magallón等(2015)所得结果存在很大重叠, 较为一致。另外, 本研究得出广义苋科类群冠群节点分化时间(约69.9 Ma (73.3-67.0 Ma))也与该科中可信的最早化石记录所在地层时间较为一致。
本研究表明, 在广义苋科中, 分支A早期在白垩纪-古近纪边界附近时期(约66 Ma)可能发生过明显的快速分化。该分支于67.3 Ma (71.6-63.5 Ma)开始分化出分支A1与A2, 而分支A1中紧接着在65.3 Ma (70.1-60.7 Ma)分化出多节草亚科及其姐妹群。从显示支长的系统树(图1)上能明显看出, 相关节点存在明显的长短支现象, 意味着此处可能发生过快速辐射演化。在白垩纪-古近纪边界附近时期, 小行星撞击地球使地球环境发生巨大变化, 导致众多生物类群走向灭绝(Schulte et al., 2010), 但同时也为其它幸存者提供了广阔的生存空间, 为众多新的生物物种爆发提供了重要的生态机会。前人研究结果表明众多被子植物类群(如防己科(Menispermaceae) (Wang et al., 2012)、兰科(Orchidaceae) (Givnish et al., 2015)和楝科(Meliaceae) (Koenen et al., 2015))在此时可能都经历过明显的分化。苋科相关分支类群的起源与分化极有可能也与这一事件有关。
附录1
本研究所用类群名称及DNA序列信息
Appendix 1
Appendix table 2 Molecular dating results of Amaranthaceae sensu lato derived from the BEAST analysis
中国科学院中国植物志编辑委员会 (1979). 中国植物志, 第25卷第2分册. 北京: 科学出版社. pp. 241. [本文引用: 4]
APGIV (2016). An update of the angiosperm phylogeny group classi?cation for the orders and families of ?owering plants: APG IV Bot J Linn Soc 181, 1-20. [本文引用: 4]
BenthamG, HookerJD (1880). Genera Plantarum Vol. 3. London: Lovell Reeve. pp. 1258. [本文引用: 1]
BrockingtonSF, AlexandreR, RamdialJ, MooreMJ, CrawleyS, DhingraA, HiluK, SoltisDE, SoltisPS (2009). Phylogeny of the Caryophyllales sensu lato: revisiting hypotheses on pollination biology and perianth differentiation in the core Caryophyllales Int J Plant Sci 170, 627-643. [本文引用: 1]
ChristenhuszMJM, ByngJW (2016). The number of known plants species in the world and its annual increase Phytotaxa 261, 201-217. [本文引用: 1]
CronquistA (1988). Caryophyllidae. In: The Evolution and Classification of Flowering Plants, 2nd edn New York: The New York Botanical Garden. pp. 309-320. [本文引用: 1]
CuénoudP, SavolainenV, ChatrouLW, PowellM, GrayerRJ, ChaseMW (2002). Molecular phylogenetics of Caryophyllales based on nuclear 18S rDNA and plastid rbcL, atpB, and matK DNA sequences Am J Bot 89, 132-144. [本文引用: 1]
DrummondAJ, SuchardMA, XieD, RambautA (2012). Bayesian phylogenetics with BEAUTi and the BEAST 1.7 Mol Biol Evol 29, 1967-1973. [本文引用: 1]
GivnishTJ, SpalinkD, AmesM, LyonSP, HunterSJ, ZuluagaA, IlesWJD, ClementsMA, ArroyoMTK, Leebens-MackJ, EndaraL, KriebelR, NeubigKM, WhittenWM, WilliamsNH, CameronKM (2015). Orchid phylogenomics and multiple drivers of their extraordinary diversi?cation Proc R Soc B Biol Sci 282, 20151553. [本文引用: 1]
GuoX, ThomasDC, SaundersRMK (2018). Gene tree discordance and coalescent methods support ancient intergeneric hybridisation between Dasymaschalon and Friesodielsia(Annonaceae) Mol Phylogenet Evol 127, 14-29. [本文引用: 1]
Hernández-LedesmaP, BerendsohnWG, BorschT, von MeringS, AkhaniH, AriasS, Casta?eda-NoaI, EggliU, ErikssonR, Flores-OlveraH, Fuentes-BazánS, KadereitG, KlakC, KorotkovaN, NyffelerR, OcampoG, OchoterenaH, OxelmanB, RabelerRK, SanchezA, SchlumpbergerBO, UotilaP (2015). A taxonomic backbone for the global synthesis of species diversity in the angiosperm order Caryophyllales Willdenowia 45, 281-383. [本文引用: 4]
KadereitG, AckerlyD, PirieMD (2012). A broader model for C4 photosynthesis evolution in plants inferred from the goosefoot family (Chenopodiaceae s.s.) Proc R Soc B Biol Sci 279, 3304-3311. [本文引用: 2]
KadereitG, BorschT, WeisingK, FreitagH (2003). Phylogeny of Amaranthaceae and Chenopodiaceae and the evolution of C4 photosynthesis Int J Plant Sci 164, 959-986. DOI:10.1086/378649URL [本文引用: 8]
KadereitG, GotzekD, JacobsS, FreitagH (2005). Origin and age of Australian Chenopodiaceae Org Divers Evol 5, 59-80. [本文引用: 3]
KatohK, StandleyDM (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability Mol Biol Evol 30, 772-780. URLPMID:23329690 [本文引用: 1]
KoenenEJM, ClarksonJJ, PenningtonTD, ChatrouLW (2015). Recently evolved diversity and convergent radiations of rainforest mahoganies (Meliaceae) shed new light on the origins of rainforest hyperdiversity New Phytol 207, 327-339. [本文引用: 1]
KühnU, BittrichV, CarolinR, FreitagH, HedgeIC, UotilaP, WilsonPG (1993). Chenopodiaceae. In: Kubitzki K, ed. Families and Genera of Vascular Plants, Vol. 2 Berlin: Springer. pp. 253-281. [本文引用: 2]
LiHT, YiTS, GaoLM, MaPF, ZhangT, YangJB, GitzendannerMA, FritschPW, CaiJ, LuoY, WangH, van der BankM, ZhangSD, WangQF, WangJ, ZhangZR, FuCN, YangJ, HollingsworthPM, ChaseMW, SoltisDE, SoltisPS, LiDZ (2019). Origin of angiosperms and the puzzle of the Jurassic gap Nat Plants 5, 461-470. URLPMID:31061536 [本文引用: 2]
MagallónS, Gómez-AcevedoS, Sánchez-ReyesLL, Hernández-HernándezT (2015). A metacalibrated time-tree documents the early rise of ?owering plant phylogenetic diversity New Phytol 207, 437-453. URLPMID:25615647 [本文引用: 2]
MassonR, KadereitG (2013). Phylogeny of Polycnemoideae (Amaranthaceae): implications for biogeography, character evolution and taxonomy Taxon 62, 100-111. [本文引用: 3]
MillerMA, PfeifferW, SchwartzT (2010). Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In: Proceedings of the Gateway Computing Environments Workshop (GCE) New Orleans, LA: IEEE. pp. 1-8. [本文引用: 1]
Moquin-TandonA (1837). Note sur le genre Polycnemum et sur une nouvelle tribu de la famille des Paronychees Ann Sci Nat 7, 33-42. [本文引用: 1]
MullerJ (1981). Fossil pollen records of extant angiosperms Bot Rev 47, 1-142. [本文引用: 1]
MülerK, BorschT (2005). Phylogenetics of Amaranthaceae based on matK/trnK sequence data—evidence from parsimony, likelihood, and Bayesian analyses Ann Missouri Bot Gard 92, 66-102. [本文引用: 5]
PosadaD (2008). jModelTest: phylogenetic model averaging Mol Biol Evol 25, 1253-1256. DOI:10.1093/molbev/msn083URLPMID:18397919 [本文引用: 1] jModelTest is a new program for the statistical selection of models of nucleotide substitution based on
PrattDB (2003). Phylogeny and Morphological Evolution of the Chenopodiaceae-Amaranthaceae Alliance. Ph.D. thesis Ames: Iowa State University. pp. 116. [本文引用: 2]
RambautA (2012). FigTree version 1.4.0. Available from: http://tree.bio.ed.ac.uk/software/figtree/. URL [本文引用: 2]
SchulteP, AlegretL, ArenillasI, ArzJA, BartonPJ, BownPR, BralowerTJ, ChristesonGL, ClaeysP, CockellCS, CollinsGS, DeutschA, GoldinTJ, GotoK, Grajales-NishimuraJM, GrieveRAF, GulickSPS, JohnsonKR, KiesslingW, KoeberlC, KringDA, MacLeodKG, MatsuiT, MeloshJ, MontanariA, MorganJV, NealCR, NicholsDJ, NorrisRD, PierazzoE, RavizzaG, Rebolledo-VieyraM, ReimoldWU, RobinE, SalgeT, SpeijerRP, SweetAR, Urrutia-FucugauchiJ, VajdaV, WhalenMT, WillumsenPS (2010). The chicxulub asteroid impact and mass extinction at the Cretaceous-Paleogene boundary Science 327, 1214-1218. DOI:10.1126/science.1177265URLPMID:20203042 [本文引用: 1] The Cretaceous-Paleogene boundary approximately 65.5 million years ago marks one of the three largest mass extinctions in the past 500 million years. The extinction event coincided with a large asteroid impact at Chicxulub, Mexico, and occurred within the time of Deccan flood basalt volcanism in India. Here, we synthesize records of the global stratigraphy across this boundary to assess the proposed causes of the mass extinction. Notably, a single ejecta-rich deposit compositionally linked to the Chicxulub impact is globally distributed at the Cretaceous-Paleogene boundary. The temporal match between the ejecta layer and the onset of the extinctions and the agreement of ecological patterns in the fossil record with modeled environmental perturbations (for example, darkness and cooling) lead us to conclude that the Chicxulub impact triggered the mass extinction.
ScottAJ (1977a). Nomina conservanda proposita Taxon 26, 246. [本文引用: 1]
ScottAJ (1977b). Reinstatement and revision of Salicorniaceae J. Agardh (Caryophyllales) Bot J Linn Soc 75, 357-374. [本文引用: 1]
SorianoA (1944). El género Nitrophila en la Argentina y su posiciónsistemática Rev Argent Agron 11, 302. [本文引用: 1]
StamatakisA (2006). RAxML-VI-HPC: maximum likelihood- based phylogenetic analyses with thousands of taxa and mixed models Bioinformatics 22, 2688-2690. DOI:10.1093/bioinformatics/btl446URLPMID:16928733 [本文引用: 4] UNLABELLED: RAxML-VI-HPC (randomized axelerated maximum likelihood for high performance computing) is a sequential and parallel program for inference of large phylogenies with maximum likelihood (ML). Low-level technical optimizations, a modification of the search algorithm, and the use of the GTR+CAT approximation as replacement for GTR+Gamma yield a program that is between 2.7 and 52 times faster than the previous version of RAxML. A large-scale performance comparison with GARLI, PHYML, IQPNNI and MrBayes on real data containing 1000 up to 6722 taxa shows that RAxML requires at least 5.6 times less main memory and yields better trees in similar times than the best competing program (GARLI) on datasets up to 2500 taxa. On datasets > or =4000 taxa it also runs 2-3 times faster than GARLI. RAxML has been parallelized with MPI to conduct parallel multiple bootstraps and inferences on distinct starting trees. The program has been used to compute ML trees on two of the largest alignments to date containing 25,057 (1463 bp) and 2182 (51,089 bp) taxa, respectively. AVAILABILITY: icwww.epfl.ch/~stamatak
StevensPF (2001). Angiosperm Phylogeny Website. Version 14, July 2017 [and more or less continuously updated since]. Available from: http://www.mobot.org/MOBOT/research/APweb/. URL [本文引用: 5]
SukhorukovAP, MavrodievEV, StruwigM, NilovaMV, DzhalilovaKK, BalandinSA, ErstA, KrinitsynaAA (2015). One-seeded fruits in the core Caryophyllales: their origin and structural diversity PLoS One 10, e0117974. URLPMID:25710481 [本文引用: 3]
TakhtajanA (1997). Diversity and Classification of Flowering Plants. New York: Columbia University Press. pp. 1-643. [本文引用: 1]
UlbrichE (1934). Chenopodiaceae. In: Engler A, Prantl K, eds. Die Naturlichen P?anzenfamilien, Vol. 16c Leipzig: Engelmann. pp. 379-584. [本文引用: 2]
WalkerJF, YangY, FengT, TimonedaA, MikenasJ, HutchisonV, EdwardsC, WangN, AhluwaliaS, OlivieriJ, Walker-HaleN, MajureLC, PuenteR, KadereitG, LauterbachM, EggliU, Flores-OlveraH, OchoterenaH, BrockingtonSF, MooreMJ, SmithSA (2018). From cacti to carnivores: improved phylotranscriptomic sampling and hierarchical homology inference provide further insight into the evolution of Caryophyllales Am J Bot 105, 446-462. DOI:10.1002/ajb2.1069URLPMID:29738076 [本文引用: 4] PREMISE OF THE STUDY: The Caryophyllales contain ~12,500 species and are known for their cosmopolitan distribution, convergence of trait evolution, and extreme adaptations. Some relationships within the Caryophyllales, like those of many large plant clades, remain unclear, and phylogenetic studies often recover alternative hypotheses. We explore the utility of broad and dense transcriptome sampling across the order for resolving evolutionary relationships in Caryophyllales. METHODS: We generated 84 transcriptomes and combined these with 224 publicly available transcriptomes to perform a phylogenomic analysis of Caryophyllales. To overcome the computational challenge of ortholog detection in such a large data set, we developed an approach for clustering gene families that allowed us to analyze >300 transcriptomes and genomes. We then inferred the species relationships using multiple methods and performed gene-tree conflict analyses. KEY RESULTS: Our phylogenetic analyses resolved many clades with strong support, but also showed significant gene-tree discordance. This discordance is not only a common feature of phylogenomic studies, but also represents an opportunity to understand processes that have structured phylogenies. We also found taxon sampling influences species-tree inference, highlighting the importance of more focused studies with additional taxon sampling. CONCLUSIONS: Transcriptomes are useful both for species-tree inference and for uncovering evolutionary complexity within lineages. Through analyses of gene-tree conflict and multiple methods of species-tree inference, we demonstrate that phylogenomic data can provide unparalleled insight into the evolutionary history of Caryophyllales. We also discuss a method for overcoming computational challenges associated with homolog clustering in large data sets.
WangW, OrtizRDC, JacquesFMB, XiangXG, LiHL, LinL, LiRQ, LiuY, SoltisPS, SoltisDE, ChenZD (2012). Menispermaceae and the diversification of tropical rainforests near the Cretaceous-Paleogene boundary New Phytol 195, 470-478. [本文引用: 1]
Wikstr?mN, SavolainenV, ChaseMW (2001). Evolution of the angiosperms: calibrating the family tree Proc R Soc B Biol Sci 268, 2211-2220. [本文引用: 5]
YangY, MooreMJ, BrockingtonSF, MikenasJ, OlivieriJ, WalkerJF, SmithSA (2018). Improved transcriptome sampling pinpoints 26 ancient and more recent polyploidy events in Caryophyllales, including two allopolyploidy events New Phytol 217, 855-870. DOI:10.1111/nph.14812URLPMID:28944472 [本文引用: 2] Studies of the macroevolutionary legacy of polyploidy are limited by an incomplete sampling of these events across the tree of life. To better locate and understand these events, we need comprehensive taxonomic sampling as well as homology inference methods that accurately reconstruct the frequency and location of gene duplications. We assembled a data set of transcriptomes and genomes from 168 species in Caryophyllales, of which 43 transcriptomes were newly generated for this study, representing one of the most densely sampled genomic-scale data sets available. We carried out phylogenomic analyses using a modified phylome strategy to reconstruct the species tree. We mapped the phylogenetic distribution of polyploidy events by both tree-based and distance-based methods, and explicitly tested scenarios for allopolyploidy. We identified 26 ancient and more recent polyploidy events distributed throughout Caryophyllales. Two of these events were inferred to be allopolyploidy. Through dense phylogenomic sampling, we show the propensity of polyploidy throughout the evolutionary history of Caryophyllales. We also provide a framework for utilizing transcriptome data to detect allopolyploidy, which is important as it may have different macroevolutionary implications compared with autopolyploidy.
YaoG, JinJJ, LiHT, YangJB, MandalaVS, CroleyM, MostowR, DouglasNA, ChaseMW, ChristenhuszMJM, SoltisDE, SoltisPS, SmithSA, BrockingtonSF, MooreMJ, YiTS, LiDZ (2019). Plastid phylogenomic insights into the evolution of Caryophyllales Mol Phylogenet Evol 134, 74-86. DOI:10.1016/j.ympev.2018.12.023URLPMID:30735725 [本文引用: 15] The Caryophyllales includes 40 families and 12,500 species, representing a large and diverse clade of angiosperms. Collectively, members of the clade grow on all continents and in all terrestrial biomes and often occupy extreme habitats (e.g., xeric, salty). The order is characterized by many taxa with unusual adaptations including carnivory, halophytism, and multiple origins of C4 photosynthesis. However, deep phylogenetic relationships within the order have long been problematic due to putative rapid divergence. To resolve the deep-level relationships of Caryophyllales, we performed phylogenomic analyses of all 40 families of Caryophyllales. We time-calibrated the molecular phylogeny of this clade, and evaluated putative correlations among plastid structural changes and rates of molecular substitution. We recovered a well-resolved and well-supported phylogeny of the Caryophyllales that was largely congruent with previous estimates of this order. Our results provide improved support for the phylogenetic position of several key families within this clade. The crown age of Caryophyllales was estimated at ca. 114.4million years ago (Ma), with periods of rapid divergence in the mid-Cretaceous. A strong, positive correlation between nucleotide substitution rate and plastid structural changes was detected. Our study highlights the importance of broad taxon sampling in phylogenomic inference and provides a firm basis for future investigations of molecular, morphological, and ecophysiological evolution in Caryophyllales.
YiTS, JinGH, WenJ (2015). Chloroplast capture and intra- and inter-continental biogeographic diversification in the Asian—New World disjunct plant genus Osmorhiza(Apiaceae) Mol Phylogenet Evol 85, 10-21. URLPMID:25585153 [本文引用: 1]
4 1979
... 苋科(Amaranthaceae sensu lato)隶属石竹目核心分支(core Caryophyllales), 其与玛瑙果科(Achatocarpaceae)是姐妹群(Yao et al., 2019).目前所接受的苋科为其广义概念(Stevens, 2001; APG IV, 2016; Yao et al., 2019), 包含传统界定的狭义苋科(Amaranthaceae sensu stricto)和藜科(Chenopodiaceae)两大类群, 共有约165属2 040种, 是石竹目中仅次于石竹科(Caryophyllaceae)的第二大科(Christenhusz and Byng, 2016).苋科是包含众多作物及蔬菜等具有重要经济价值的植物类群, 如苋菜(Amaranthus tricolor L.)、甜菜(Beta vulgaris L.)、藜麦(Chenopodium quinoa Willd.)、菠菜(Spinacia oleracea L.) (中国科学院中国植物志编辑委员会, 1979; Stevens, 2001; Hernández-Ledesma et al., 2015).在传统分类系统中, 狭义苋科与藜科通常基于苞片、花被及雄蕊的特征被处理为2个独立的科(中国科学院中国植物志编辑委员会, 1979; Cronquist, 1988; Takhtajan, 1997), 其中藜科苞叶及花被常为草质或肉质, 雄蕊常分离; 而苋科苞叶及花被常为干膜质, 雄蕊基部常合生(中国科学院中国植物志编辑委员会, 1979).在地理分布上, 苋科主要分布于热带、亚热带地区; 而藜科则主要分布于温带至亚热带地区, 且常见于干旱、半干旱甚至盐碱地等生境(中国科学院中国植物志编辑委员会, 1979; Hernández-Ledesma et al., 2015). ...
An update of the angiosperm phylogeny group classi?cation for the orders and families of ?owering plants: APG IV 4 2016
... 苋科(Amaranthaceae sensu lato)隶属石竹目核心分支(core Caryophyllales), 其与玛瑙果科(Achatocarpaceae)是姐妹群(Yao et al., 2019).目前所接受的苋科为其广义概念(Stevens, 2001; APG IV, 2016; Yao et al., 2019), 包含传统界定的狭义苋科(Amaranthaceae sensu stricto)和藜科(Chenopodiaceae)两大类群, 共有约165属2 040种, 是石竹目中仅次于石竹科(Caryophyllaceae)的第二大科(Christenhusz and Byng, 2016).苋科是包含众多作物及蔬菜等具有重要经济价值的植物类群, 如苋菜(Amaranthus tricolor L.)、甜菜(Beta vulgaris L.)、藜麦(Chenopodium quinoa Willd.)、菠菜(Spinacia oleracea L.) (中国科学院中国植物志编辑委员会, 1979; Stevens, 2001; Hernández-Ledesma et al., 2015).在传统分类系统中, 狭义苋科与藜科通常基于苞片、花被及雄蕊的特征被处理为2个独立的科(中国科学院中国植物志编辑委员会, 1979; Cronquist, 1988; Takhtajan, 1997), 其中藜科苞叶及花被常为草质或肉质, 雄蕊常分离; 而苋科苞叶及花被常为干膜质, 雄蕊基部常合生(中国科学院中国植物志编辑委员会, 1979).在地理分布上, 苋科主要分布于热带、亚热带地区; 而藜科则主要分布于温带至亚热带地区, 且常见于干旱、半干旱甚至盐碱地等生境(中国科学院中国植物志编辑委员会, 1979; Hernández-Ledesma et al., 2015). ...
... 在分子系统学研究中, 狭义苋科的单系性获得了很好的支持, 然而藜科被认为并非单系(Kadereit et al., 2003; Sukhorukov et al., 2015; Yao et al., 2019).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... 分子系统学的应用对苋科、藜科系统关系的理解起到了极大的促进作用, 亚科、族、属等类群范围的界定不断完善(Kadereit et al., 2003; Müller and Borsch, 2005; Masson and Kadereit, 2013; Sukhorukov et al., 2015; Walker et al., 2018; Yao et al., 2019).对于苋科与藜科的系统学归属, 目前主要有两派观点.一派以APG系统为主, 即支持广义苋科的观点(APG IV, 2016; Yao et al., 2019); 另一派则支持将传统藜科中的多节草亚科并入狭义苋科, 而将藜科核心类群依然接受为藜科(Hernández-Ledesma et al., 2015; Walker et al., 2018).本研究基于8个叶绿体基因序列分析结果表明, 藜科核心类群中的藜亚科、虫实亚科、甜菜亚科、多节草亚科与狭义苋科共同形成一支, 而藜科核心类群中其它4个亚科共同形成另一支, 且这2个分支均获得很高的支持率.这意味着藜科核心类群并非单系, 因此将其依然接受为藜科的观点在本研究中未能获得支持.基于此, 我们支持APG系统将狭义苋科与藜科合并为广义苋科的分类处理. ...
Genera Plantarum 1 1880
... 多节草亚科的系统位置在传统分类处理中历来争议很大, 其系统学位置的确定对于广义苋科系统发育关系的理解非常重要.在传统处理中, 该类群曾被置于传统藜科(Ulbrich, 1934; Kühn et al., 1993)或狭义苋科(Soriano, 1944), 甚至石竹科(Moquin-Tandon, 1837)之中.从形态特征上, 该亚科类群花被常干膜质或纸质, 花丝基部合生成管状, 这与狭义苋科类群相似而与藜科不同, 但其花常单生于枝上部的叶腋处又与传统藜科及狭义苋科其它类群均不同(Masson and Kadereit, 2013).该类群常分布于亚热带至暖温带地区的干旱或盐碱地等开阔生境, 与传统藜科类群极为相似(Masson and Kadereit, 2013).因此, 多节草亚科曾被认为是联系传统藜科与狭义苋科的中间过渡类群(Bentham and Hooker, 1880).有时也被认为是联系石竹科指甲草族(Paronychieae)与苋科-藜科联盟的中间过渡类群(Ulbrich, 1934; Kühn et al., 1993).在分子系统学研究中, 多节草亚科的系统位置也存在争议(见前言部分), 且相关研究揭示的该类群系统位置的支持率通常较低(Kadereit et al., 2003; Pratt, 2003; Müller and Borsch, 2005; Sukhorukov et al., 2015).在本研究中, 多节草亚科与狭义苋科以及传统藜科中的藜亚科、虫实亚科、甜菜亚科共同形成一个支持率很高的分支(图1, 分支A), 并与后3个亚科成姐妹群关系(图1, 分支A1), 但该姐妹群关系的支持率较低(MLBS=70%; PP=0.86).多节草亚科系统关系未能得到很好地解决可能与相关节点经历过快速辐射分化有关, 这不仅从系统发育树所呈现出的该亚科干群节点处存在明显的长短分支情况可以看出(图1), 而且从分子钟估算结果中也不难发现(图2).建议今后增加取样类群并从基因组层面开展广义苋科类群的系统发育关系研究, 可能有助于确立多节草亚科的系统位置. ...
Phylogeny of the Caryophyllales sensu lato: revisiting hypotheses on pollination biology and perianth differentiation in the core Caryophyllales 1 2009
... 在分子系统学研究中, 狭义苋科的单系性获得了很好的支持, 然而藜科被认为并非单系(Kadereit et al., 2003; Sukhorukov et al., 2015; Yao et al., 2019).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
The number of known plants species in the world and its annual increase 1 2016
... 苋科(Amaranthaceae sensu lato)隶属石竹目核心分支(core Caryophyllales), 其与玛瑙果科(Achatocarpaceae)是姐妹群(Yao et al., 2019).目前所接受的苋科为其广义概念(Stevens, 2001; APG IV, 2016; Yao et al., 2019), 包含传统界定的狭义苋科(Amaranthaceae sensu stricto)和藜科(Chenopodiaceae)两大类群, 共有约165属2 040种, 是石竹目中仅次于石竹科(Caryophyllaceae)的第二大科(Christenhusz and Byng, 2016).苋科是包含众多作物及蔬菜等具有重要经济价值的植物类群, 如苋菜(Amaranthus tricolor L.)、甜菜(Beta vulgaris L.)、藜麦(Chenopodium quinoa Willd.)、菠菜(Spinacia oleracea L.) (中国科学院中国植物志编辑委员会, 1979; Stevens, 2001; Hernández-Ledesma et al., 2015).在传统分类系统中, 狭义苋科与藜科通常基于苞片、花被及雄蕊的特征被处理为2个独立的科(中国科学院中国植物志编辑委员会, 1979; Cronquist, 1988; Takhtajan, 1997), 其中藜科苞叶及花被常为草质或肉质, 雄蕊常分离; 而苋科苞叶及花被常为干膜质, 雄蕊基部常合生(中国科学院中国植物志编辑委员会, 1979).在地理分布上, 苋科主要分布于热带、亚热带地区; 而藜科则主要分布于温带至亚热带地区, 且常见于干旱、半干旱甚至盐碱地等生境(中国科学院中国植物志编辑委员会, 1979; Hernández-Ledesma et al., 2015). ...
Caryophyllidae. In: The Evolution and Classification of Flowering Plants, 2nd edn 1 1988
... 苋科(Amaranthaceae sensu lato)隶属石竹目核心分支(core Caryophyllales), 其与玛瑙果科(Achatocarpaceae)是姐妹群(Yao et al., 2019).目前所接受的苋科为其广义概念(Stevens, 2001; APG IV, 2016; Yao et al., 2019), 包含传统界定的狭义苋科(Amaranthaceae sensu stricto)和藜科(Chenopodiaceae)两大类群, 共有约165属2 040种, 是石竹目中仅次于石竹科(Caryophyllaceae)的第二大科(Christenhusz and Byng, 2016).苋科是包含众多作物及蔬菜等具有重要经济价值的植物类群, 如苋菜(Amaranthus tricolor L.)、甜菜(Beta vulgaris L.)、藜麦(Chenopodium quinoa Willd.)、菠菜(Spinacia oleracea L.) (中国科学院中国植物志编辑委员会, 1979; Stevens, 2001; Hernández-Ledesma et al., 2015).在传统分类系统中, 狭义苋科与藜科通常基于苞片、花被及雄蕊的特征被处理为2个独立的科(中国科学院中国植物志编辑委员会, 1979; Cronquist, 1988; Takhtajan, 1997), 其中藜科苞叶及花被常为草质或肉质, 雄蕊常分离; 而苋科苞叶及花被常为干膜质, 雄蕊基部常合生(中国科学院中国植物志编辑委员会, 1979).在地理分布上, 苋科主要分布于热带、亚热带地区; 而藜科则主要分布于温带至亚热带地区, 且常见于干旱、半干旱甚至盐碱地等生境(中国科学院中国植物志编辑委员会, 1979; Hernández-Ledesma et al., 2015). ...
Molecular phylogenetics of Caryophyllales based on nuclear 18S rDNA and plastid rbcL, atpB, and matK DNA sequences 1 2002
... 在分子系统学研究中, 狭义苋科的单系性获得了很好的支持, 然而藜科被认为并非单系(Kadereit et al., 2003; Sukhorukov et al., 2015; Yao et al., 2019).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
Bayesian phylogenetics with BEAUTi and the BEAST 1.7 1 2012
... 为揭示广义苋科及其主要分支的起源分化时间, 采用BEAST v. 1.8.4软件(Drummond et al., 2012)对其进行分化时间估算.基于广义苋科中所报道的化石记录以及前期研究所得苋科与其近缘类群间的分化时间对相关节点进行时间校准.具体如下: (1) 参考Kadereit等(2003, 2005)用化石物种Parvangula randeckensis Hiltermann & Schmitz的中新世早期种子化石对藜属(Chenopodium)所代表的藜族第1分支(Chenopodieae I clade)干群(stem node)的起源时间进行校准, 化石时间的使用采用对数正态分布模型(lognormal distribution model), 最小时间限定在16.0百万年前(offset=16.0 Million-years ago (Ma)), 平均值限定为1.0 (mean=1.0), 标准差限定为0.5 (standard deviation (SD)=0.5); (2) 参考Kadereit等(2003, 2005)、Yao等(2019)及Li等(2019)的研究, 用化石物种Polyporina cribaria Srivastava的白垩纪晚期花粉化石对广义苋科冠群(crown node)起源时间进行校准, 化石时间同样采用对数正态分布模型, 最小时间限定为66.0 Ma, 平均值限定为1.0, 标准差限定为0.5; (3) 参考Yao等(2019)所得结果, 对苋科与玛瑙果科分化时间采用均匀分布模型(uniform distribution model), 时间上限限定为82.5 Ma, 下限限定为77.9 Ma; (4) 参考Yao等(2019)所得结果, 对石竹科与苋科-玛瑙果科分支分化时间采用均匀分布模型, 时间上限限定为92.2 Ma, 下限限定为85.7 Ma.分化时间在“出生-灭绝”物种形成模型(birth-death speciation process)之下, 基于对数正态分布的松散分子钟(lognormal relaxed molecular clock)进行估算; 各序列碱基替代模型参考1.2节序列最佳碱基替代模型筛选结果; 马尔科夫链蒙特卡洛(Markov chain Monte Carlo, MCMC)运算300 000 000代, 每5 000代取样1次, 该运算步骤重复3次独立运算; 基于Tracer v. 1.6软件(Rambaut et al., 2014)对所得参数进行检查, 确认所有参数的有效居群规模(effective sample sizes, ESS)数值均超过200, 结果可靠; 基于BEAST软件包中TreeAnotator v. 1.8.4软件, 将最初所得15 000棵系统树作为老化样品舍弃, 对剩下的所有系统树进行总结并计算其一致性树, 统计相关节点的分化时间.在Figtree v1.4.2软件(Rambaut, 2012)中查看所得结果. ...
Orchid phylogenomics and multiple drivers of their extraordinary diversi?cation 1 2015
... 本研究表明, 在广义苋科中, 分支A早期在白垩纪-古近纪边界附近时期(约66 Ma)可能发生过明显的快速分化.该分支于67.3 Ma (71.6-63.5 Ma)开始分化出分支A1与A2, 而分支A1中紧接着在65.3 Ma (70.1-60.7 Ma)分化出多节草亚科及其姐妹群.从显示支长的系统树(图1)上能明显看出, 相关节点存在明显的长短支现象, 意味着此处可能发生过快速辐射演化.在白垩纪-古近纪边界附近时期, 小行星撞击地球使地球环境发生巨大变化, 导致众多生物类群走向灭绝(Schulte et al., 2010), 但同时也为其它幸存者提供了广阔的生存空间, 为众多新的生物物种爆发提供了重要的生态机会.前人研究结果表明众多被子植物类群(如防己科(Menispermaceae) (Wang et al., 2012)、兰科(Orchidaceae) (Givnish et al., 2015)和楝科(Meliaceae) (Koenen et al., 2015))在此时可能都经历过明显的分化.苋科相关分支类群的起源与分化极有可能也与这一事件有关. ...
Gene tree discordance and coalescent methods support ancient intergeneric hybridisation between Dasymaschalon and Friesodielsia(Annonaceae) 1 2018
... 在Walker等(2018)基于转录组数据构建的石竹目系统发育关系中, 传统界定的藜科(含多节草亚科)形成一个单系分支, 且与狭义苋科成姐妹群关系.该研究还表明, 在藜科系统关系拓扑结构的主干靠基部连续3个节点, 均有约75%的基因树并不支持所呈现的拓扑结构, 这说明该研究获取的转录组数据中相关基因之间系统发育信号冲突明显, 因此所呈现的拓扑结构可能并未揭示出相关类群真实的进化关系.此外, 在Yang等(2018)基于转录组数据所构建的石竹目系统发育关系中, 猪笼草科与茅膏菜科关系较近, 而与露松科关系较远, 且相关节点均获得很高的支持率, 这与基于叶绿体基因组数据所得结果一致(Yao et al., 2019).但Walker等(2018)的研究结果则支持猪笼草科与露松科关系更近, 与茅膏菜科关系相对较远, 相关节点支持率同样很高.在基于转录组数据的系统发育分析中, 猪笼草科、茅膏菜科及露松科之间的系统关系在不同研究中出现明显冲突, 可能与相关研究在转录组数据分析背景下所筛选出来用于系统发育树构建的“直系同源基因”存在差异有关, 这意味着基于转录组数据筛选出的大量“直系同源基因”所构建的系统发育关系仍然值得商榷.另一方面, 在系统发育分析中, 基于叶绿体基因序列数据所得结果与核基因序列数据所得结果存在冲突的先例在被子植物类群中普遍存在, 这一现象通常被认为可能与杂交、叶绿体捕获以及不完全的谱系分选等进化事件有关(Gurushidze et al., 2010; Yi et al., 2015; Guo et al., 2018).因此, 阐释广义苋科中基于叶绿体数据与核基因数据所得结果存在冲突的原因, 将是后期研究中需要关注和探讨的重要问题. ...
Species-level phylogeny of Allium subgenus Melanocrommyum: incomplete lineage sorting, hybridization and trnF gene duplication 1 2010
... 在Walker等(2018)基于转录组数据构建的石竹目系统发育关系中, 传统界定的藜科(含多节草亚科)形成一个单系分支, 且与狭义苋科成姐妹群关系.该研究还表明, 在藜科系统关系拓扑结构的主干靠基部连续3个节点, 均有约75%的基因树并不支持所呈现的拓扑结构, 这说明该研究获取的转录组数据中相关基因之间系统发育信号冲突明显, 因此所呈现的拓扑结构可能并未揭示出相关类群真实的进化关系.此外, 在Yang等(2018)基于转录组数据所构建的石竹目系统发育关系中, 猪笼草科与茅膏菜科关系较近, 而与露松科关系较远, 且相关节点均获得很高的支持率, 这与基于叶绿体基因组数据所得结果一致(Yao et al., 2019).但Walker等(2018)的研究结果则支持猪笼草科与露松科关系更近, 与茅膏菜科关系相对较远, 相关节点支持率同样很高.在基于转录组数据的系统发育分析中, 猪笼草科、茅膏菜科及露松科之间的系统关系在不同研究中出现明显冲突, 可能与相关研究在转录组数据分析背景下所筛选出来用于系统发育树构建的“直系同源基因”存在差异有关, 这意味着基于转录组数据筛选出的大量“直系同源基因”所构建的系统发育关系仍然值得商榷.另一方面, 在系统发育分析中, 基于叶绿体基因序列数据所得结果与核基因序列数据所得结果存在冲突的先例在被子植物类群中普遍存在, 这一现象通常被认为可能与杂交、叶绿体捕获以及不完全的谱系分选等进化事件有关(Gurushidze et al., 2010; Yi et al., 2015; Guo et al., 2018).因此, 阐释广义苋科中基于叶绿体数据与核基因数据所得结果存在冲突的原因, 将是后期研究中需要关注和探讨的重要问题. ...
A taxonomic backbone for the global synthesis of species diversity in the angiosperm order Caryophyllales 4 2015
... 苋科(Amaranthaceae sensu lato)隶属石竹目核心分支(core Caryophyllales), 其与玛瑙果科(Achatocarpaceae)是姐妹群(Yao et al., 2019).目前所接受的苋科为其广义概念(Stevens, 2001; APG IV, 2016; Yao et al., 2019), 包含传统界定的狭义苋科(Amaranthaceae sensu stricto)和藜科(Chenopodiaceae)两大类群, 共有约165属2 040种, 是石竹目中仅次于石竹科(Caryophyllaceae)的第二大科(Christenhusz and Byng, 2016).苋科是包含众多作物及蔬菜等具有重要经济价值的植物类群, 如苋菜(Amaranthus tricolor L.)、甜菜(Beta vulgaris L.)、藜麦(Chenopodium quinoa Willd.)、菠菜(Spinacia oleracea L.) (中国科学院中国植物志编辑委员会, 1979; Stevens, 2001; Hernández-Ledesma et al., 2015).在传统分类系统中, 狭义苋科与藜科通常基于苞片、花被及雄蕊的特征被处理为2个独立的科(中国科学院中国植物志编辑委员会, 1979; Cronquist, 1988; Takhtajan, 1997), 其中藜科苞叶及花被常为草质或肉质, 雄蕊常分离; 而苋科苞叶及花被常为干膜质, 雄蕊基部常合生(中国科学院中国植物志编辑委员会, 1979).在地理分布上, 苋科主要分布于热带、亚热带地区; 而藜科则主要分布于温带至亚热带地区, 且常见于干旱、半干旱甚至盐碱地等生境(中国科学院中国植物志编辑委员会, 1979; Hernández-Ledesma et al., 2015). ...
... ; Hernández-Ledesma et al., 2015). ...
... 在分子系统学研究中, 狭义苋科的单系性获得了很好的支持, 然而藜科被认为并非单系(Kadereit et al., 2003; Sukhorukov et al., 2015; Yao et al., 2019).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... 分子系统学的应用对苋科、藜科系统关系的理解起到了极大的促进作用, 亚科、族、属等类群范围的界定不断完善(Kadereit et al., 2003; Müller and Borsch, 2005; Masson and Kadereit, 2013; Sukhorukov et al., 2015; Walker et al., 2018; Yao et al., 2019).对于苋科与藜科的系统学归属, 目前主要有两派观点.一派以APG系统为主, 即支持广义苋科的观点(APG IV, 2016; Yao et al., 2019); 另一派则支持将传统藜科中的多节草亚科并入狭义苋科, 而将藜科核心类群依然接受为藜科(Hernández-Ledesma et al., 2015; Walker et al., 2018).本研究基于8个叶绿体基因序列分析结果表明, 藜科核心类群中的藜亚科、虫实亚科、甜菜亚科、多节草亚科与狭义苋科共同形成一支, 而藜科核心类群中其它4个亚科共同形成另一支, 且这2个分支均获得很高的支持率.这意味着藜科核心类群并非单系, 因此将其依然接受为藜科的观点在本研究中未能获得支持.基于此, 我们支持APG系统将狭义苋科与藜科合并为广义苋科的分类处理. ...
A broader model for C4 photosynthesis evolution in plants inferred from the goosefoot family (Chenopodiaceae s.s.) 2 2012
... 对于广义苋科的起源时间, 前人多项研究均有涉及.Wikstr?m等(2001)研究表明, 该科干群节点起源可能发生在约28、38或40 Ma; 而Kadereit等(2012)认为该科干群起源时间发生在87-47 Ma; Magallón等(2015)则认为该科干群节点可能发生在64.2 Ma (81.8-43.6 Ma)或76.4 Ma (80.5-72.0 Ma).本研究表明, 该科干群节点起源可能发生在白垩纪晚期约80.5 Ma (82.5-78.3 Ma).在广义苋科中, 目前已知最早的可信化石记录为Polyporina cribraria Srivastava, 该化石物种为发现于白垩纪晚期马斯里奇特阶(Maestrichtian, 72.1-66.0 Ma)的花粉化石.在形态特征上, 该化石与传统藜科类群(尤其是现存物种Salicornia australis Sol. ex F. Muell.)的花粉化石非常相似.该化石发现于加拿大海洋环境向陆地环境的过渡地带, 与现存众多藜科类群的生境也非常相似.此外, 自马斯里奇特阶之后在相似生境中也发现过一系列类似的花粉化石(Muller, 1981; Kadereit et al., 2003).基于形态特征及其发现地的生境特征, 该花粉化石被广泛接受为广义苋科类群并多次用于该科类群分化时间校准分析(Kadereit et al., 2005; Yao et al., 2019; Li et al., 2019).因此, 从化石证据角度看, Wikstr?m等(2001)所得结果在时间尺度上明显偏早.而本研究结果在时间区间上与Kadereit等(2012)及Magallón等(2015)所得结果存在很大重叠, 较为一致.另外, 本研究得出广义苋科类群冠群节点分化时间(约69.9 Ma (73.3-67.0 Ma))也与该科中可信的最早化石记录所在地层时间较为一致. ...
... 所得结果在时间尺度上明显偏早.而本研究结果在时间区间上与Kadereit等(2012)及Magallón等(2015)所得结果存在很大重叠, 较为一致.另外, 本研究得出广义苋科类群冠群节点分化时间(约69.9 Ma (73.3-67.0 Ma))也与该科中可信的最早化石记录所在地层时间较为一致. ...
Phylogeny of Amaranthaceae and Chenopodiaceae and the evolution of C4 photosynthesis 8 2003
... 在分子系统学研究中, 狭义苋科的单系性获得了很好的支持, 然而藜科被认为并非单系(Kadereit et al., 2003; Sukhorukov et al., 2015; Yao et al., 2019).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... ).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... ).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... 为揭示广义苋科及其主要分支的起源分化时间, 采用BEAST v. 1.8.4软件(Drummond et al., 2012)对其进行分化时间估算.基于广义苋科中所报道的化石记录以及前期研究所得苋科与其近缘类群间的分化时间对相关节点进行时间校准.具体如下: (1) 参考Kadereit等(2003, 2005)用化石物种Parvangula randeckensis Hiltermann & Schmitz的中新世早期种子化石对藜属(Chenopodium)所代表的藜族第1分支(Chenopodieae I clade)干群(stem node)的起源时间进行校准, 化石时间的使用采用对数正态分布模型(lognormal distribution model), 最小时间限定在16.0百万年前(offset=16.0 Million-years ago (Ma)), 平均值限定为1.0 (mean=1.0), 标准差限定为0.5 (standard deviation (SD)=0.5); (2) 参考Kadereit等(2003, 2005)、Yao等(2019)及Li等(2019)的研究, 用化石物种Polyporina cribaria Srivastava的白垩纪晚期花粉化石对广义苋科冠群(crown node)起源时间进行校准, 化石时间同样采用对数正态分布模型, 最小时间限定为66.0 Ma, 平均值限定为1.0, 标准差限定为0.5; (3) 参考Yao等(2019)所得结果, 对苋科与玛瑙果科分化时间采用均匀分布模型(uniform distribution model), 时间上限限定为82.5 Ma, 下限限定为77.9 Ma; (4) 参考Yao等(2019)所得结果, 对石竹科与苋科-玛瑙果科分支分化时间采用均匀分布模型, 时间上限限定为92.2 Ma, 下限限定为85.7 Ma.分化时间在“出生-灭绝”物种形成模型(birth-death speciation process)之下, 基于对数正态分布的松散分子钟(lognormal relaxed molecular clock)进行估算; 各序列碱基替代模型参考1.2节序列最佳碱基替代模型筛选结果; 马尔科夫链蒙特卡洛(Markov chain Monte Carlo, MCMC)运算300 000 000代, 每5 000代取样1次, 该运算步骤重复3次独立运算; 基于Tracer v. 1.6软件(Rambaut et al., 2014)对所得参数进行检查, 确认所有参数的有效居群规模(effective sample sizes, ESS)数值均超过200, 结果可靠; 基于BEAST软件包中TreeAnotator v. 1.8.4软件, 将最初所得15 000棵系统树作为老化样品舍弃, 对剩下的所有系统树进行总结并计算其一致性树, 统计相关节点的分化时间.在Figtree v1.4.2软件(Rambaut, 2012)中查看所得结果. ...
... 分子系统学的应用对苋科、藜科系统关系的理解起到了极大的促进作用, 亚科、族、属等类群范围的界定不断完善(Kadereit et al., 2003; Müller and Borsch, 2005; Masson and Kadereit, 2013; Sukhorukov et al., 2015; Walker et al., 2018; Yao et al., 2019).对于苋科与藜科的系统学归属, 目前主要有两派观点.一派以APG系统为主, 即支持广义苋科的观点(APG IV, 2016; Yao et al., 2019); 另一派则支持将传统藜科中的多节草亚科并入狭义苋科, 而将藜科核心类群依然接受为藜科(Hernández-Ledesma et al., 2015; Walker et al., 2018).本研究基于8个叶绿体基因序列分析结果表明, 藜科核心类群中的藜亚科、虫实亚科、甜菜亚科、多节草亚科与狭义苋科共同形成一支, 而藜科核心类群中其它4个亚科共同形成另一支, 且这2个分支均获得很高的支持率.这意味着藜科核心类群并非单系, 因此将其依然接受为藜科的观点在本研究中未能获得支持.基于此, 我们支持APG系统将狭义苋科与藜科合并为广义苋科的分类处理. ...
... 多节草亚科的系统位置在传统分类处理中历来争议很大, 其系统学位置的确定对于广义苋科系统发育关系的理解非常重要.在传统处理中, 该类群曾被置于传统藜科(Ulbrich, 1934; Kühn et al., 1993)或狭义苋科(Soriano, 1944), 甚至石竹科(Moquin-Tandon, 1837)之中.从形态特征上, 该亚科类群花被常干膜质或纸质, 花丝基部合生成管状, 这与狭义苋科类群相似而与藜科不同, 但其花常单生于枝上部的叶腋处又与传统藜科及狭义苋科其它类群均不同(Masson and Kadereit, 2013).该类群常分布于亚热带至暖温带地区的干旱或盐碱地等开阔生境, 与传统藜科类群极为相似(Masson and Kadereit, 2013).因此, 多节草亚科曾被认为是联系传统藜科与狭义苋科的中间过渡类群(Bentham and Hooker, 1880).有时也被认为是联系石竹科指甲草族(Paronychieae)与苋科-藜科联盟的中间过渡类群(Ulbrich, 1934; Kühn et al., 1993).在分子系统学研究中, 多节草亚科的系统位置也存在争议(见前言部分), 且相关研究揭示的该类群系统位置的支持率通常较低(Kadereit et al., 2003; Pratt, 2003; Müller and Borsch, 2005; Sukhorukov et al., 2015).在本研究中, 多节草亚科与狭义苋科以及传统藜科中的藜亚科、虫实亚科、甜菜亚科共同形成一个支持率很高的分支(图1, 分支A), 并与后3个亚科成姐妹群关系(图1, 分支A1), 但该姐妹群关系的支持率较低(MLBS=70%; PP=0.86).多节草亚科系统关系未能得到很好地解决可能与相关节点经历过快速辐射分化有关, 这不仅从系统发育树所呈现出的该亚科干群节点处存在明显的长短分支情况可以看出(图1), 而且从分子钟估算结果中也不难发现(图2).建议今后增加取样类群并从基因组层面开展广义苋科类群的系统发育关系研究, 可能有助于确立多节草亚科的系统位置. ...
... 对于广义苋科的起源时间, 前人多项研究均有涉及.Wikstr?m等(2001)研究表明, 该科干群节点起源可能发生在约28、38或40 Ma; 而Kadereit等(2012)认为该科干群起源时间发生在87-47 Ma; Magallón等(2015)则认为该科干群节点可能发生在64.2 Ma (81.8-43.6 Ma)或76.4 Ma (80.5-72.0 Ma).本研究表明, 该科干群节点起源可能发生在白垩纪晚期约80.5 Ma (82.5-78.3 Ma).在广义苋科中, 目前已知最早的可信化石记录为Polyporina cribraria Srivastava, 该化石物种为发现于白垩纪晚期马斯里奇特阶(Maestrichtian, 72.1-66.0 Ma)的花粉化石.在形态特征上, 该化石与传统藜科类群(尤其是现存物种Salicornia australis Sol. ex F. Muell.)的花粉化石非常相似.该化石发现于加拿大海洋环境向陆地环境的过渡地带, 与现存众多藜科类群的生境也非常相似.此外, 自马斯里奇特阶之后在相似生境中也发现过一系列类似的花粉化石(Muller, 1981; Kadereit et al., 2003).基于形态特征及其发现地的生境特征, 该花粉化石被广泛接受为广义苋科类群并多次用于该科类群分化时间校准分析(Kadereit et al., 2005; Yao et al., 2019; Li et al., 2019).因此, 从化石证据角度看, Wikstr?m等(2001)所得结果在时间尺度上明显偏早.而本研究结果在时间区间上与Kadereit等(2012)及Magallón等(2015)所得结果存在很大重叠, 较为一致.另外, 本研究得出广义苋科类群冠群节点分化时间(约69.9 Ma (73.3-67.0 Ma))也与该科中可信的最早化石记录所在地层时间较为一致. ...
Origin and age of Australian Chenopodiaceae 3 2005
... 为揭示广义苋科及其主要分支的起源分化时间, 采用BEAST v. 1.8.4软件(Drummond et al., 2012)对其进行分化时间估算.基于广义苋科中所报道的化石记录以及前期研究所得苋科与其近缘类群间的分化时间对相关节点进行时间校准.具体如下: (1) 参考Kadereit等(2003, 2005)用化石物种Parvangula randeckensis Hiltermann & Schmitz的中新世早期种子化石对藜属(Chenopodium)所代表的藜族第1分支(Chenopodieae I clade)干群(stem node)的起源时间进行校准, 化石时间的使用采用对数正态分布模型(lognormal distribution model), 最小时间限定在16.0百万年前(offset=16.0 Million-years ago (Ma)), 平均值限定为1.0 (mean=1.0), 标准差限定为0.5 (standard deviation (SD)=0.5); (2) 参考Kadereit等(2003, 2005)、Yao等(2019)及Li等(2019)的研究, 用化石物种Polyporina cribaria Srivastava的白垩纪晚期花粉化石对广义苋科冠群(crown node)起源时间进行校准, 化石时间同样采用对数正态分布模型, 最小时间限定为66.0 Ma, 平均值限定为1.0, 标准差限定为0.5; (3) 参考Yao等(2019)所得结果, 对苋科与玛瑙果科分化时间采用均匀分布模型(uniform distribution model), 时间上限限定为82.5 Ma, 下限限定为77.9 Ma; (4) 参考Yao等(2019)所得结果, 对石竹科与苋科-玛瑙果科分支分化时间采用均匀分布模型, 时间上限限定为92.2 Ma, 下限限定为85.7 Ma.分化时间在“出生-灭绝”物种形成模型(birth-death speciation process)之下, 基于对数正态分布的松散分子钟(lognormal relaxed molecular clock)进行估算; 各序列碱基替代模型参考1.2节序列最佳碱基替代模型筛选结果; 马尔科夫链蒙特卡洛(Markov chain Monte Carlo, MCMC)运算300 000 000代, 每5 000代取样1次, 该运算步骤重复3次独立运算; 基于Tracer v. 1.6软件(Rambaut et al., 2014)对所得参数进行检查, 确认所有参数的有效居群规模(effective sample sizes, ESS)数值均超过200, 结果可靠; 基于BEAST软件包中TreeAnotator v. 1.8.4软件, 将最初所得15 000棵系统树作为老化样品舍弃, 对剩下的所有系统树进行总结并计算其一致性树, 统计相关节点的分化时间.在Figtree v1.4.2软件(Rambaut, 2012)中查看所得结果. ...
... 对于广义苋科的起源时间, 前人多项研究均有涉及.Wikstr?m等(2001)研究表明, 该科干群节点起源可能发生在约28、38或40 Ma; 而Kadereit等(2012)认为该科干群起源时间发生在87-47 Ma; Magallón等(2015)则认为该科干群节点可能发生在64.2 Ma (81.8-43.6 Ma)或76.4 Ma (80.5-72.0 Ma).本研究表明, 该科干群节点起源可能发生在白垩纪晚期约80.5 Ma (82.5-78.3 Ma).在广义苋科中, 目前已知最早的可信化石记录为Polyporina cribraria Srivastava, 该化石物种为发现于白垩纪晚期马斯里奇特阶(Maestrichtian, 72.1-66.0 Ma)的花粉化石.在形态特征上, 该化石与传统藜科类群(尤其是现存物种Salicornia australis Sol. ex F. Muell.)的花粉化石非常相似.该化石发现于加拿大海洋环境向陆地环境的过渡地带, 与现存众多藜科类群的生境也非常相似.此外, 自马斯里奇特阶之后在相似生境中也发现过一系列类似的花粉化石(Muller, 1981; Kadereit et al., 2003).基于形态特征及其发现地的生境特征, 该花粉化石被广泛接受为广义苋科类群并多次用于该科类群分化时间校准分析(Kadereit et al., 2005; Yao et al., 2019; Li et al., 2019).因此, 从化石证据角度看, Wikstr?m等(2001)所得结果在时间尺度上明显偏早.而本研究结果在时间区间上与Kadereit等(2012)及Magallón等(2015)所得结果存在很大重叠, 较为一致.另外, 本研究得出广义苋科类群冠群节点分化时间(约69.9 Ma (73.3-67.0 Ma))也与该科中可信的最早化石记录所在地层时间较为一致. ...
MAFFT multiple sequence alignment software version 7: improvements in performance and usability 1 2013
Recently evolved diversity and convergent radiations of rainforest mahoganies (Meliaceae) shed new light on the origins of rainforest hyperdiversity 1 2015
... 本研究表明, 在广义苋科中, 分支A早期在白垩纪-古近纪边界附近时期(约66 Ma)可能发生过明显的快速分化.该分支于67.3 Ma (71.6-63.5 Ma)开始分化出分支A1与A2, 而分支A1中紧接着在65.3 Ma (70.1-60.7 Ma)分化出多节草亚科及其姐妹群.从显示支长的系统树(图1)上能明显看出, 相关节点存在明显的长短支现象, 意味着此处可能发生过快速辐射演化.在白垩纪-古近纪边界附近时期, 小行星撞击地球使地球环境发生巨大变化, 导致众多生物类群走向灭绝(Schulte et al., 2010), 但同时也为其它幸存者提供了广阔的生存空间, 为众多新的生物物种爆发提供了重要的生态机会.前人研究结果表明众多被子植物类群(如防己科(Menispermaceae) (Wang et al., 2012)、兰科(Orchidaceae) (Givnish et al., 2015)和楝科(Meliaceae) (Koenen et al., 2015))在此时可能都经历过明显的分化.苋科相关分支类群的起源与分化极有可能也与这一事件有关. ...
Chenopodiaceae. In: Kubitzki K, ed. Families and Genera of Vascular Plants, Vol. 2 2 1993
... 多节草亚科的系统位置在传统分类处理中历来争议很大, 其系统学位置的确定对于广义苋科系统发育关系的理解非常重要.在传统处理中, 该类群曾被置于传统藜科(Ulbrich, 1934; Kühn et al., 1993)或狭义苋科(Soriano, 1944), 甚至石竹科(Moquin-Tandon, 1837)之中.从形态特征上, 该亚科类群花被常干膜质或纸质, 花丝基部合生成管状, 这与狭义苋科类群相似而与藜科不同, 但其花常单生于枝上部的叶腋处又与传统藜科及狭义苋科其它类群均不同(Masson and Kadereit, 2013).该类群常分布于亚热带至暖温带地区的干旱或盐碱地等开阔生境, 与传统藜科类群极为相似(Masson and Kadereit, 2013).因此, 多节草亚科曾被认为是联系传统藜科与狭义苋科的中间过渡类群(Bentham and Hooker, 1880).有时也被认为是联系石竹科指甲草族(Paronychieae)与苋科-藜科联盟的中间过渡类群(Ulbrich, 1934; Kühn et al., 1993).在分子系统学研究中, 多节草亚科的系统位置也存在争议(见前言部分), 且相关研究揭示的该类群系统位置的支持率通常较低(Kadereit et al., 2003; Pratt, 2003; Müller and Borsch, 2005; Sukhorukov et al., 2015).在本研究中, 多节草亚科与狭义苋科以及传统藜科中的藜亚科、虫实亚科、甜菜亚科共同形成一个支持率很高的分支(图1, 分支A), 并与后3个亚科成姐妹群关系(图1, 分支A1), 但该姐妹群关系的支持率较低(MLBS=70%; PP=0.86).多节草亚科系统关系未能得到很好地解决可能与相关节点经历过快速辐射分化有关, 这不仅从系统发育树所呈现出的该亚科干群节点处存在明显的长短分支情况可以看出(图1), 而且从分子钟估算结果中也不难发现(图2).建议今后增加取样类群并从基因组层面开展广义苋科类群的系统发育关系研究, 可能有助于确立多节草亚科的系统位置. ...
... ; Kühn et al., 1993).在分子系统学研究中, 多节草亚科的系统位置也存在争议(见前言部分), 且相关研究揭示的该类群系统位置的支持率通常较低(Kadereit et al., 2003; Pratt, 2003; Müller and Borsch, 2005; Sukhorukov et al., 2015).在本研究中, 多节草亚科与狭义苋科以及传统藜科中的藜亚科、虫实亚科、甜菜亚科共同形成一个支持率很高的分支(图1, 分支A), 并与后3个亚科成姐妹群关系(图1, 分支A1), 但该姐妹群关系的支持率较低(MLBS=70%; PP=0.86).多节草亚科系统关系未能得到很好地解决可能与相关节点经历过快速辐射分化有关, 这不仅从系统发育树所呈现出的该亚科干群节点处存在明显的长短分支情况可以看出(图1), 而且从分子钟估算结果中也不难发现(图2).建议今后增加取样类群并从基因组层面开展广义苋科类群的系统发育关系研究, 可能有助于确立多节草亚科的系统位置. ...
Origin of angiosperms and the puzzle of the Jurassic gap 2 2019
... 为揭示广义苋科及其主要分支的起源分化时间, 采用BEAST v. 1.8.4软件(Drummond et al., 2012)对其进行分化时间估算.基于广义苋科中所报道的化石记录以及前期研究所得苋科与其近缘类群间的分化时间对相关节点进行时间校准.具体如下: (1) 参考Kadereit等(2003, 2005)用化石物种Parvangula randeckensis Hiltermann & Schmitz的中新世早期种子化石对藜属(Chenopodium)所代表的藜族第1分支(Chenopodieae I clade)干群(stem node)的起源时间进行校准, 化石时间的使用采用对数正态分布模型(lognormal distribution model), 最小时间限定在16.0百万年前(offset=16.0 Million-years ago (Ma)), 平均值限定为1.0 (mean=1.0), 标准差限定为0.5 (standard deviation (SD)=0.5); (2) 参考Kadereit等(2003, 2005)、Yao等(2019)及Li等(2019)的研究, 用化石物种Polyporina cribaria Srivastava的白垩纪晚期花粉化石对广义苋科冠群(crown node)起源时间进行校准, 化石时间同样采用对数正态分布模型, 最小时间限定为66.0 Ma, 平均值限定为1.0, 标准差限定为0.5; (3) 参考Yao等(2019)所得结果, 对苋科与玛瑙果科分化时间采用均匀分布模型(uniform distribution model), 时间上限限定为82.5 Ma, 下限限定为77.9 Ma; (4) 参考Yao等(2019)所得结果, 对石竹科与苋科-玛瑙果科分支分化时间采用均匀分布模型, 时间上限限定为92.2 Ma, 下限限定为85.7 Ma.分化时间在“出生-灭绝”物种形成模型(birth-death speciation process)之下, 基于对数正态分布的松散分子钟(lognormal relaxed molecular clock)进行估算; 各序列碱基替代模型参考1.2节序列最佳碱基替代模型筛选结果; 马尔科夫链蒙特卡洛(Markov chain Monte Carlo, MCMC)运算300 000 000代, 每5 000代取样1次, 该运算步骤重复3次独立运算; 基于Tracer v. 1.6软件(Rambaut et al., 2014)对所得参数进行检查, 确认所有参数的有效居群规模(effective sample sizes, ESS)数值均超过200, 结果可靠; 基于BEAST软件包中TreeAnotator v. 1.8.4软件, 将最初所得15 000棵系统树作为老化样品舍弃, 对剩下的所有系统树进行总结并计算其一致性树, 统计相关节点的分化时间.在Figtree v1.4.2软件(Rambaut, 2012)中查看所得结果. ...
... 对于广义苋科的起源时间, 前人多项研究均有涉及.Wikstr?m等(2001)研究表明, 该科干群节点起源可能发生在约28、38或40 Ma; 而Kadereit等(2012)认为该科干群起源时间发生在87-47 Ma; Magallón等(2015)则认为该科干群节点可能发生在64.2 Ma (81.8-43.6 Ma)或76.4 Ma (80.5-72.0 Ma).本研究表明, 该科干群节点起源可能发生在白垩纪晚期约80.5 Ma (82.5-78.3 Ma).在广义苋科中, 目前已知最早的可信化石记录为Polyporina cribraria Srivastava, 该化石物种为发现于白垩纪晚期马斯里奇特阶(Maestrichtian, 72.1-66.0 Ma)的花粉化石.在形态特征上, 该化石与传统藜科类群(尤其是现存物种Salicornia australis Sol. ex F. Muell.)的花粉化石非常相似.该化石发现于加拿大海洋环境向陆地环境的过渡地带, 与现存众多藜科类群的生境也非常相似.此外, 自马斯里奇特阶之后在相似生境中也发现过一系列类似的花粉化石(Muller, 1981; Kadereit et al., 2003).基于形态特征及其发现地的生境特征, 该花粉化石被广泛接受为广义苋科类群并多次用于该科类群分化时间校准分析(Kadereit et al., 2005; Yao et al., 2019; Li et al., 2019).因此, 从化石证据角度看, Wikstr?m等(2001)所得结果在时间尺度上明显偏早.而本研究结果在时间区间上与Kadereit等(2012)及Magallón等(2015)所得结果存在很大重叠, 较为一致.另外, 本研究得出广义苋科类群冠群节点分化时间(约69.9 Ma (73.3-67.0 Ma))也与该科中可信的最早化石记录所在地层时间较为一致. ...
A metacalibrated time-tree documents the early rise of ?owering plant phylogenetic diversity 2 2015
... 对于广义苋科的起源时间, 前人多项研究均有涉及.Wikstr?m等(2001)研究表明, 该科干群节点起源可能发生在约28、38或40 Ma; 而Kadereit等(2012)认为该科干群起源时间发生在87-47 Ma; Magallón等(2015)则认为该科干群节点可能发生在64.2 Ma (81.8-43.6 Ma)或76.4 Ma (80.5-72.0 Ma).本研究表明, 该科干群节点起源可能发生在白垩纪晚期约80.5 Ma (82.5-78.3 Ma).在广义苋科中, 目前已知最早的可信化石记录为Polyporina cribraria Srivastava, 该化石物种为发现于白垩纪晚期马斯里奇特阶(Maestrichtian, 72.1-66.0 Ma)的花粉化石.在形态特征上, 该化石与传统藜科类群(尤其是现存物种Salicornia australis Sol. ex F. Muell.)的花粉化石非常相似.该化石发现于加拿大海洋环境向陆地环境的过渡地带, 与现存众多藜科类群的生境也非常相似.此外, 自马斯里奇特阶之后在相似生境中也发现过一系列类似的花粉化石(Muller, 1981; Kadereit et al., 2003).基于形态特征及其发现地的生境特征, 该花粉化石被广泛接受为广义苋科类群并多次用于该科类群分化时间校准分析(Kadereit et al., 2005; Yao et al., 2019; Li et al., 2019).因此, 从化石证据角度看, Wikstr?m等(2001)所得结果在时间尺度上明显偏早.而本研究结果在时间区间上与Kadereit等(2012)及Magallón等(2015)所得结果存在很大重叠, 较为一致.另外, 本研究得出广义苋科类群冠群节点分化时间(约69.9 Ma (73.3-67.0 Ma))也与该科中可信的最早化石记录所在地层时间较为一致. ...
... 及Magallón等(2015)所得结果存在很大重叠, 较为一致.另外, 本研究得出广义苋科类群冠群节点分化时间(约69.9 Ma (73.3-67.0 Ma))也与该科中可信的最早化石记录所在地层时间较为一致. ...
Phylogeny of Polycnemoideae (Amaranthaceae): implications for biogeography, character evolution and taxonomy 3 2013
... 分子系统学的应用对苋科、藜科系统关系的理解起到了极大的促进作用, 亚科、族、属等类群范围的界定不断完善(Kadereit et al., 2003; Müller and Borsch, 2005; Masson and Kadereit, 2013; Sukhorukov et al., 2015; Walker et al., 2018; Yao et al., 2019).对于苋科与藜科的系统学归属, 目前主要有两派观点.一派以APG系统为主, 即支持广义苋科的观点(APG IV, 2016; Yao et al., 2019); 另一派则支持将传统藜科中的多节草亚科并入狭义苋科, 而将藜科核心类群依然接受为藜科(Hernández-Ledesma et al., 2015; Walker et al., 2018).本研究基于8个叶绿体基因序列分析结果表明, 藜科核心类群中的藜亚科、虫实亚科、甜菜亚科、多节草亚科与狭义苋科共同形成一支, 而藜科核心类群中其它4个亚科共同形成另一支, 且这2个分支均获得很高的支持率.这意味着藜科核心类群并非单系, 因此将其依然接受为藜科的观点在本研究中未能获得支持.基于此, 我们支持APG系统将狭义苋科与藜科合并为广义苋科的分类处理. ...
... 多节草亚科的系统位置在传统分类处理中历来争议很大, 其系统学位置的确定对于广义苋科系统发育关系的理解非常重要.在传统处理中, 该类群曾被置于传统藜科(Ulbrich, 1934; Kühn et al., 1993)或狭义苋科(Soriano, 1944), 甚至石竹科(Moquin-Tandon, 1837)之中.从形态特征上, 该亚科类群花被常干膜质或纸质, 花丝基部合生成管状, 这与狭义苋科类群相似而与藜科不同, 但其花常单生于枝上部的叶腋处又与传统藜科及狭义苋科其它类群均不同(Masson and Kadereit, 2013).该类群常分布于亚热带至暖温带地区的干旱或盐碱地等开阔生境, 与传统藜科类群极为相似(Masson and Kadereit, 2013).因此, 多节草亚科曾被认为是联系传统藜科与狭义苋科的中间过渡类群(Bentham and Hooker, 1880).有时也被认为是联系石竹科指甲草族(Paronychieae)与苋科-藜科联盟的中间过渡类群(Ulbrich, 1934; Kühn et al., 1993).在分子系统学研究中, 多节草亚科的系统位置也存在争议(见前言部分), 且相关研究揭示的该类群系统位置的支持率通常较低(Kadereit et al., 2003; Pratt, 2003; Müller and Borsch, 2005; Sukhorukov et al., 2015).在本研究中, 多节草亚科与狭义苋科以及传统藜科中的藜亚科、虫实亚科、甜菜亚科共同形成一个支持率很高的分支(图1, 分支A), 并与后3个亚科成姐妹群关系(图1, 分支A1), 但该姐妹群关系的支持率较低(MLBS=70%; PP=0.86).多节草亚科系统关系未能得到很好地解决可能与相关节点经历过快速辐射分化有关, 这不仅从系统发育树所呈现出的该亚科干群节点处存在明显的长短分支情况可以看出(图1), 而且从分子钟估算结果中也不难发现(图2).建议今后增加取样类群并从基因组层面开展广义苋科类群的系统发育关系研究, 可能有助于确立多节草亚科的系统位置. ...
... ).该类群常分布于亚热带至暖温带地区的干旱或盐碱地等开阔生境, 与传统藜科类群极为相似(Masson and Kadereit, 2013).因此, 多节草亚科曾被认为是联系传统藜科与狭义苋科的中间过渡类群(Bentham and Hooker, 1880).有时也被认为是联系石竹科指甲草族(Paronychieae)与苋科-藜科联盟的中间过渡类群(Ulbrich, 1934; Kühn et al., 1993).在分子系统学研究中, 多节草亚科的系统位置也存在争议(见前言部分), 且相关研究揭示的该类群系统位置的支持率通常较低(Kadereit et al., 2003; Pratt, 2003; Müller and Borsch, 2005; Sukhorukov et al., 2015).在本研究中, 多节草亚科与狭义苋科以及传统藜科中的藜亚科、虫实亚科、甜菜亚科共同形成一个支持率很高的分支(图1, 分支A), 并与后3个亚科成姐妹群关系(图1, 分支A1), 但该姐妹群关系的支持率较低(MLBS=70%; PP=0.86).多节草亚科系统关系未能得到很好地解决可能与相关节点经历过快速辐射分化有关, 这不仅从系统发育树所呈现出的该亚科干群节点处存在明显的长短分支情况可以看出(图1), 而且从分子钟估算结果中也不难发现(图2).建议今后增加取样类群并从基因组层面开展广义苋科类群的系统发育关系研究, 可能有助于确立多节草亚科的系统位置. ...
Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In: Proceedings of the Gateway Computing Environments Workshop (GCE) 1 2010
Note sur le genre Polycnemum et sur une nouvelle tribu de la famille des Paronychees 1 1837
... 多节草亚科的系统位置在传统分类处理中历来争议很大, 其系统学位置的确定对于广义苋科系统发育关系的理解非常重要.在传统处理中, 该类群曾被置于传统藜科(Ulbrich, 1934; Kühn et al., 1993)或狭义苋科(Soriano, 1944), 甚至石竹科(Moquin-Tandon, 1837)之中.从形态特征上, 该亚科类群花被常干膜质或纸质, 花丝基部合生成管状, 这与狭义苋科类群相似而与藜科不同, 但其花常单生于枝上部的叶腋处又与传统藜科及狭义苋科其它类群均不同(Masson and Kadereit, 2013).该类群常分布于亚热带至暖温带地区的干旱或盐碱地等开阔生境, 与传统藜科类群极为相似(Masson and Kadereit, 2013).因此, 多节草亚科曾被认为是联系传统藜科与狭义苋科的中间过渡类群(Bentham and Hooker, 1880).有时也被认为是联系石竹科指甲草族(Paronychieae)与苋科-藜科联盟的中间过渡类群(Ulbrich, 1934; Kühn et al., 1993).在分子系统学研究中, 多节草亚科的系统位置也存在争议(见前言部分), 且相关研究揭示的该类群系统位置的支持率通常较低(Kadereit et al., 2003; Pratt, 2003; Müller and Borsch, 2005; Sukhorukov et al., 2015).在本研究中, 多节草亚科与狭义苋科以及传统藜科中的藜亚科、虫实亚科、甜菜亚科共同形成一个支持率很高的分支(图1, 分支A), 并与后3个亚科成姐妹群关系(图1, 分支A1), 但该姐妹群关系的支持率较低(MLBS=70%; PP=0.86).多节草亚科系统关系未能得到很好地解决可能与相关节点经历过快速辐射分化有关, 这不仅从系统发育树所呈现出的该亚科干群节点处存在明显的长短分支情况可以看出(图1), 而且从分子钟估算结果中也不难发现(图2).建议今后增加取样类群并从基因组层面开展广义苋科类群的系统发育关系研究, 可能有助于确立多节草亚科的系统位置. ...
Fossil pollen records of extant angiosperms 1 1981
... 对于广义苋科的起源时间, 前人多项研究均有涉及.Wikstr?m等(2001)研究表明, 该科干群节点起源可能发生在约28、38或40 Ma; 而Kadereit等(2012)认为该科干群起源时间发生在87-47 Ma; Magallón等(2015)则认为该科干群节点可能发生在64.2 Ma (81.8-43.6 Ma)或76.4 Ma (80.5-72.0 Ma).本研究表明, 该科干群节点起源可能发生在白垩纪晚期约80.5 Ma (82.5-78.3 Ma).在广义苋科中, 目前已知最早的可信化石记录为Polyporina cribraria Srivastava, 该化石物种为发现于白垩纪晚期马斯里奇特阶(Maestrichtian, 72.1-66.0 Ma)的花粉化石.在形态特征上, 该化石与传统藜科类群(尤其是现存物种Salicornia australis Sol. ex F. Muell.)的花粉化石非常相似.该化石发现于加拿大海洋环境向陆地环境的过渡地带, 与现存众多藜科类群的生境也非常相似.此外, 自马斯里奇特阶之后在相似生境中也发现过一系列类似的花粉化石(Muller, 1981; Kadereit et al., 2003).基于形态特征及其发现地的生境特征, 该花粉化石被广泛接受为广义苋科类群并多次用于该科类群分化时间校准分析(Kadereit et al., 2005; Yao et al., 2019; Li et al., 2019).因此, 从化石证据角度看, Wikstr?m等(2001)所得结果在时间尺度上明显偏早.而本研究结果在时间区间上与Kadereit等(2012)及Magallón等(2015)所得结果存在很大重叠, 较为一致.另外, 本研究得出广义苋科类群冠群节点分化时间(约69.9 Ma (73.3-67.0 Ma))也与该科中可信的最早化石记录所在地层时间较为一致. ...
Phylogenetics of Amaranthaceae based on matK/trnK sequence data—evidence from parsimony, likelihood, and Bayesian analyses 5 2005
... 在分子系统学研究中, 狭义苋科的单系性获得了很好的支持, 然而藜科被认为并非单系(Kadereit et al., 2003; Sukhorukov et al., 2015; Yao et al., 2019).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... 分子系统学的应用对苋科、藜科系统关系的理解起到了极大的促进作用, 亚科、族、属等类群范围的界定不断完善(Kadereit et al., 2003; Müller and Borsch, 2005; Masson and Kadereit, 2013; Sukhorukov et al., 2015; Walker et al., 2018; Yao et al., 2019).对于苋科与藜科的系统学归属, 目前主要有两派观点.一派以APG系统为主, 即支持广义苋科的观点(APG IV, 2016; Yao et al., 2019); 另一派则支持将传统藜科中的多节草亚科并入狭义苋科, 而将藜科核心类群依然接受为藜科(Hernández-Ledesma et al., 2015; Walker et al., 2018).本研究基于8个叶绿体基因序列分析结果表明, 藜科核心类群中的藜亚科、虫实亚科、甜菜亚科、多节草亚科与狭义苋科共同形成一支, 而藜科核心类群中其它4个亚科共同形成另一支, 且这2个分支均获得很高的支持率.这意味着藜科核心类群并非单系, 因此将其依然接受为藜科的观点在本研究中未能获得支持.基于此, 我们支持APG系统将狭义苋科与藜科合并为广义苋科的分类处理. ...
... 多节草亚科的系统位置在传统分类处理中历来争议很大, 其系统学位置的确定对于广义苋科系统发育关系的理解非常重要.在传统处理中, 该类群曾被置于传统藜科(Ulbrich, 1934; Kühn et al., 1993)或狭义苋科(Soriano, 1944), 甚至石竹科(Moquin-Tandon, 1837)之中.从形态特征上, 该亚科类群花被常干膜质或纸质, 花丝基部合生成管状, 这与狭义苋科类群相似而与藜科不同, 但其花常单生于枝上部的叶腋处又与传统藜科及狭义苋科其它类群均不同(Masson and Kadereit, 2013).该类群常分布于亚热带至暖温带地区的干旱或盐碱地等开阔生境, 与传统藜科类群极为相似(Masson and Kadereit, 2013).因此, 多节草亚科曾被认为是联系传统藜科与狭义苋科的中间过渡类群(Bentham and Hooker, 1880).有时也被认为是联系石竹科指甲草族(Paronychieae)与苋科-藜科联盟的中间过渡类群(Ulbrich, 1934; Kühn et al., 1993).在分子系统学研究中, 多节草亚科的系统位置也存在争议(见前言部分), 且相关研究揭示的该类群系统位置的支持率通常较低(Kadereit et al., 2003; Pratt, 2003; Müller and Borsch, 2005; Sukhorukov et al., 2015).在本研究中, 多节草亚科与狭义苋科以及传统藜科中的藜亚科、虫实亚科、甜菜亚科共同形成一个支持率很高的分支(图1, 分支A), 并与后3个亚科成姐妹群关系(图1, 分支A1), 但该姐妹群关系的支持率较低(MLBS=70%; PP=0.86).多节草亚科系统关系未能得到很好地解决可能与相关节点经历过快速辐射分化有关, 这不仅从系统发育树所呈现出的该亚科干群节点处存在明显的长短分支情况可以看出(图1), 而且从分子钟估算结果中也不难发现(图2).建议今后增加取样类群并从基因组层面开展广义苋科类群的系统发育关系研究, 可能有助于确立多节草亚科的系统位置. ...
... 赤珠苋属共3种, 主要分布于密克罗尼西亚群岛、塞浦路斯及西喜马拉雅地区; 炬苋树属共6种, 主要分布于夏威夷及土布艾群岛(Stevens, 2001).然而在亚科水平位置的归属方面, 这2属还有待确定.本研究得出的系统发育关系表明, 赤珠苋属与炬苋树属在广义苋科之中均占据独立的系统位置并在狭义苋科基部依次分出, 且相关节点均获得很高的支持(图1).从生活型上, 赤珠苋属与炬苋树属均为木本类型, 而狭义苋科则以多年生草本为主, 仅有极少数类群为木本(如青葙族(Celosieae)及鸽苋属(Chamissoa Kunth)类群之中的大部分物种) (Müller and Borsch, 2005).赤珠苋属果实为浆果, 这与狭义苋科浆果苋属(Deeringia R. Br.)和多脉苋属(Pleuropetalum Hook. f.)等类群相似, 而狭义苋科其它类群及传统藜科类群则以胞果为主(Müller and Borsch, 2005).因此, 基于本研究得出的系统关系, 结合生活型及形态特征等多方面证据, 我们认为赤珠苋属与炬苋树属都有必要提升为独立的亚科. ...
Phylogeny and Morphological Evolution of the Chenopodiaceae-Amaranthaceae Alliance. Ph.D. thesis 2 2003
... 在分子系统学研究中, 狭义苋科的单系性获得了很好的支持, 然而藜科被认为并非单系(Kadereit et al., 2003; Sukhorukov et al., 2015; Yao et al., 2019).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... 多节草亚科的系统位置在传统分类处理中历来争议很大, 其系统学位置的确定对于广义苋科系统发育关系的理解非常重要.在传统处理中, 该类群曾被置于传统藜科(Ulbrich, 1934; Kühn et al., 1993)或狭义苋科(Soriano, 1944), 甚至石竹科(Moquin-Tandon, 1837)之中.从形态特征上, 该亚科类群花被常干膜质或纸质, 花丝基部合生成管状, 这与狭义苋科类群相似而与藜科不同, 但其花常单生于枝上部的叶腋处又与传统藜科及狭义苋科其它类群均不同(Masson and Kadereit, 2013).该类群常分布于亚热带至暖温带地区的干旱或盐碱地等开阔生境, 与传统藜科类群极为相似(Masson and Kadereit, 2013).因此, 多节草亚科曾被认为是联系传统藜科与狭义苋科的中间过渡类群(Bentham and Hooker, 1880).有时也被认为是联系石竹科指甲草族(Paronychieae)与苋科-藜科联盟的中间过渡类群(Ulbrich, 1934; Kühn et al., 1993).在分子系统学研究中, 多节草亚科的系统位置也存在争议(见前言部分), 且相关研究揭示的该类群系统位置的支持率通常较低(Kadereit et al., 2003; Pratt, 2003; Müller and Borsch, 2005; Sukhorukov et al., 2015).在本研究中, 多节草亚科与狭义苋科以及传统藜科中的藜亚科、虫实亚科、甜菜亚科共同形成一个支持率很高的分支(图1, 分支A), 并与后3个亚科成姐妹群关系(图1, 分支A1), 但该姐妹群关系的支持率较低(MLBS=70%; PP=0.86).多节草亚科系统关系未能得到很好地解决可能与相关节点经历过快速辐射分化有关, 这不仅从系统发育树所呈现出的该亚科干群节点处存在明显的长短分支情况可以看出(图1), 而且从分子钟估算结果中也不难发现(图2).建议今后增加取样类群并从基因组层面开展广义苋科类群的系统发育关系研究, 可能有助于确立多节草亚科的系统位置. ...
The chicxulub asteroid impact and mass extinction at the Cretaceous-Paleogene boundary 1 2010
... 本研究表明, 在广义苋科中, 分支A早期在白垩纪-古近纪边界附近时期(约66 Ma)可能发生过明显的快速分化.该分支于67.3 Ma (71.6-63.5 Ma)开始分化出分支A1与A2, 而分支A1中紧接着在65.3 Ma (70.1-60.7 Ma)分化出多节草亚科及其姐妹群.从显示支长的系统树(图1)上能明显看出, 相关节点存在明显的长短支现象, 意味着此处可能发生过快速辐射演化.在白垩纪-古近纪边界附近时期, 小行星撞击地球使地球环境发生巨大变化, 导致众多生物类群走向灭绝(Schulte et al., 2010), 但同时也为其它幸存者提供了广阔的生存空间, 为众多新的生物物种爆发提供了重要的生态机会.前人研究结果表明众多被子植物类群(如防己科(Menispermaceae) (Wang et al., 2012)、兰科(Orchidaceae) (Givnish et al., 2015)和楝科(Meliaceae) (Koenen et al., 2015))在此时可能都经历过明显的分化.苋科相关分支类群的起源与分化极有可能也与这一事件有关. ...
Nomina conservanda proposita 1 1977
... 在传统藜科的分类处理中, Scott (1977a, 1977b)还曾建议将猪毛菜科(Salsolaceae Moq.)和盐角草科(Salicorniaceae J. Agardh)等类群提升至科的水平, 但并未被后来****承认, 相关类群在本研究中被证实仍然属于广义苋科的范畴, 且嵌套在该科内部.此外, 曾被置于藜科的肉叶刺茎藜属(Sarcobatus Nees)以及滨藜叶属(Halophytum Speg.)均从藜科中分出并独立成科, 且与广义苋科系统位置较远.其中肉叶刺茎藜科(Sarcobataceae)隶属石竹目商陆分支(Phytolaccoid clade), 与萝卜藤科(Agdestidaceae)成姐妹群关系(Walker et al., 2018; Yao et al., 2019); 而滨藜叶科(Halophytaceae)则隶属石竹目马齿苋亚目分支(Portulacineae clade), 与龙树科-落葵科分支(Basellaceae-Didiereaceae clade)成姐妹群关系(Yao et al., 2019). ...
Reinstatement and revision of Salicorniaceae J. Agardh (Caryophyllales) 1 1977
... 在传统藜科的分类处理中, Scott (1977a, 1977b)还曾建议将猪毛菜科(Salsolaceae Moq.)和盐角草科(Salicorniaceae J. Agardh)等类群提升至科的水平, 但并未被后来****承认, 相关类群在本研究中被证实仍然属于广义苋科的范畴, 且嵌套在该科内部.此外, 曾被置于藜科的肉叶刺茎藜属(Sarcobatus Nees)以及滨藜叶属(Halophytum Speg.)均从藜科中分出并独立成科, 且与广义苋科系统位置较远.其中肉叶刺茎藜科(Sarcobataceae)隶属石竹目商陆分支(Phytolaccoid clade), 与萝卜藤科(Agdestidaceae)成姐妹群关系(Walker et al., 2018; Yao et al., 2019); 而滨藜叶科(Halophytaceae)则隶属石竹目马齿苋亚目分支(Portulacineae clade), 与龙树科-落葵科分支(Basellaceae-Didiereaceae clade)成姐妹群关系(Yao et al., 2019). ...
Angiosperm phylogeny: 17 genes, 640 taxa 1 2011
... 在分子系统学研究中, 狭义苋科的单系性获得了很好的支持, 然而藜科被认为并非单系(Kadereit et al., 2003; Sukhorukov et al., 2015; Yao et al., 2019).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
El género Nitrophila en la Argentina y su posiciónsistemática 1 1944
... 多节草亚科的系统位置在传统分类处理中历来争议很大, 其系统学位置的确定对于广义苋科系统发育关系的理解非常重要.在传统处理中, 该类群曾被置于传统藜科(Ulbrich, 1934; Kühn et al., 1993)或狭义苋科(Soriano, 1944), 甚至石竹科(Moquin-Tandon, 1837)之中.从形态特征上, 该亚科类群花被常干膜质或纸质, 花丝基部合生成管状, 这与狭义苋科类群相似而与藜科不同, 但其花常单生于枝上部的叶腋处又与传统藜科及狭义苋科其它类群均不同(Masson and Kadereit, 2013).该类群常分布于亚热带至暖温带地区的干旱或盐碱地等开阔生境, 与传统藜科类群极为相似(Masson and Kadereit, 2013).因此, 多节草亚科曾被认为是联系传统藜科与狭义苋科的中间过渡类群(Bentham and Hooker, 1880).有时也被认为是联系石竹科指甲草族(Paronychieae)与苋科-藜科联盟的中间过渡类群(Ulbrich, 1934; Kühn et al., 1993).在分子系统学研究中, 多节草亚科的系统位置也存在争议(见前言部分), 且相关研究揭示的该类群系统位置的支持率通常较低(Kadereit et al., 2003; Pratt, 2003; Müller and Borsch, 2005; Sukhorukov et al., 2015).在本研究中, 多节草亚科与狭义苋科以及传统藜科中的藜亚科、虫实亚科、甜菜亚科共同形成一个支持率很高的分支(图1, 分支A), 并与后3个亚科成姐妹群关系(图1, 分支A1), 但该姐妹群关系的支持率较低(MLBS=70%; PP=0.86).多节草亚科系统关系未能得到很好地解决可能与相关节点经历过快速辐射分化有关, 这不仅从系统发育树所呈现出的该亚科干群节点处存在明显的长短分支情况可以看出(图1), 而且从分子钟估算结果中也不难发现(图2).建议今后增加取样类群并从基因组层面开展广义苋科类群的系统发育关系研究, 可能有助于确立多节草亚科的系统位置. ...
RAxML-VI-HPC: maximum likelihood- based phylogenetic analyses with thousands of taxa and mixed models 4 2006
... 分子系统学的应用对苋科、藜科系统关系的理解起到了极大的促进作用, 亚科、族、属等类群范围的界定不断完善(Kadereit et al., 2003; Müller and Borsch, 2005; Masson and Kadereit, 2013; Sukhorukov et al., 2015; Walker et al., 2018; Yao et al., 2019).对于苋科与藜科的系统学归属, 目前主要有两派观点.一派以APG系统为主, 即支持广义苋科的观点(APG IV, 2016; Yao et al., 2019); 另一派则支持将传统藜科中的多节草亚科并入狭义苋科, 而将藜科核心类群依然接受为藜科(Hernández-Ledesma et al., 2015; Walker et al., 2018).本研究基于8个叶绿体基因序列分析结果表明, 藜科核心类群中的藜亚科、虫实亚科、甜菜亚科、多节草亚科与狭义苋科共同形成一支, 而藜科核心类群中其它4个亚科共同形成另一支, 且这2个分支均获得很高的支持率.这意味着藜科核心类群并非单系, 因此将其依然接受为藜科的观点在本研究中未能获得支持.基于此, 我们支持APG系统将狭义苋科与藜科合并为广义苋科的分类处理. ...
... 在分子系统学研究中, 狭义苋科的单系性获得了很好的支持, 然而藜科被认为并非单系(Kadereit et al., 2003; Sukhorukov et al., 2015; Yao et al., 2019).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... 赤珠苋属共3种, 主要分布于密克罗尼西亚群岛、塞浦路斯及西喜马拉雅地区; 炬苋树属共6种, 主要分布于夏威夷及土布艾群岛(Stevens, 2001).然而在亚科水平位置的归属方面, 这2属还有待确定.本研究得出的系统发育关系表明, 赤珠苋属与炬苋树属在广义苋科之中均占据独立的系统位置并在狭义苋科基部依次分出, 且相关节点均获得很高的支持(图1).从生活型上, 赤珠苋属与炬苋树属均为木本类型, 而狭义苋科则以多年生草本为主, 仅有极少数类群为木本(如青葙族(Celosieae)及鸽苋属(Chamissoa Kunth)类群之中的大部分物种) (Müller and Borsch, 2005).赤珠苋属果实为浆果, 这与狭义苋科浆果苋属(Deeringia R. Br.)和多脉苋属(Pleuropetalum Hook. f.)等类群相似, 而狭义苋科其它类群及传统藜科类群则以胞果为主(Müller and Borsch, 2005).因此, 基于本研究得出的系统关系, 结合生活型及形态特征等多方面证据, 我们认为赤珠苋属与炬苋树属都有必要提升为独立的亚科. ...
One-seeded fruits in the core Caryophyllales: their origin and structural diversity 3 2015
... 在分子系统学研究中, 狭义苋科的单系性获得了很好的支持, 然而藜科被认为并非单系(Kadereit et al., 2003; Sukhorukov et al., 2015; Yao et al., 2019).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... ; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... ; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
1 1997
... 苋科(Amaranthaceae sensu lato)隶属石竹目核心分支(core Caryophyllales), 其与玛瑙果科(Achatocarpaceae)是姐妹群(Yao et al., 2019).目前所接受的苋科为其广义概念(Stevens, 2001; APG IV, 2016; Yao et al., 2019), 包含传统界定的狭义苋科(Amaranthaceae sensu stricto)和藜科(Chenopodiaceae)两大类群, 共有约165属2 040种, 是石竹目中仅次于石竹科(Caryophyllaceae)的第二大科(Christenhusz and Byng, 2016).苋科是包含众多作物及蔬菜等具有重要经济价值的植物类群, 如苋菜(Amaranthus tricolor L.)、甜菜(Beta vulgaris L.)、藜麦(Chenopodium quinoa Willd.)、菠菜(Spinacia oleracea L.) (中国科学院中国植物志编辑委员会, 1979; Stevens, 2001; Hernández-Ledesma et al., 2015).在传统分类系统中, 狭义苋科与藜科通常基于苞片、花被及雄蕊的特征被处理为2个独立的科(中国科学院中国植物志编辑委员会, 1979; Cronquist, 1988; Takhtajan, 1997), 其中藜科苞叶及花被常为草质或肉质, 雄蕊常分离; 而苋科苞叶及花被常为干膜质, 雄蕊基部常合生(中国科学院中国植物志编辑委员会, 1979).在地理分布上, 苋科主要分布于热带、亚热带地区; 而藜科则主要分布于温带至亚热带地区, 且常见于干旱、半干旱甚至盐碱地等生境(中国科学院中国植物志编辑委员会, 1979; Hernández-Ledesma et al., 2015). ...
Chenopodiaceae. In: Engler A, Prantl K, eds. Die Naturlichen P?anzenfamilien, Vol. 16c 2 1934
... 多节草亚科的系统位置在传统分类处理中历来争议很大, 其系统学位置的确定对于广义苋科系统发育关系的理解非常重要.在传统处理中, 该类群曾被置于传统藜科(Ulbrich, 1934; Kühn et al., 1993)或狭义苋科(Soriano, 1944), 甚至石竹科(Moquin-Tandon, 1837)之中.从形态特征上, 该亚科类群花被常干膜质或纸质, 花丝基部合生成管状, 这与狭义苋科类群相似而与藜科不同, 但其花常单生于枝上部的叶腋处又与传统藜科及狭义苋科其它类群均不同(Masson and Kadereit, 2013).该类群常分布于亚热带至暖温带地区的干旱或盐碱地等开阔生境, 与传统藜科类群极为相似(Masson and Kadereit, 2013).因此, 多节草亚科曾被认为是联系传统藜科与狭义苋科的中间过渡类群(Bentham and Hooker, 1880).有时也被认为是联系石竹科指甲草族(Paronychieae)与苋科-藜科联盟的中间过渡类群(Ulbrich, 1934; Kühn et al., 1993).在分子系统学研究中, 多节草亚科的系统位置也存在争议(见前言部分), 且相关研究揭示的该类群系统位置的支持率通常较低(Kadereit et al., 2003; Pratt, 2003; Müller and Borsch, 2005; Sukhorukov et al., 2015).在本研究中, 多节草亚科与狭义苋科以及传统藜科中的藜亚科、虫实亚科、甜菜亚科共同形成一个支持率很高的分支(图1, 分支A), 并与后3个亚科成姐妹群关系(图1, 分支A1), 但该姐妹群关系的支持率较低(MLBS=70%; PP=0.86).多节草亚科系统关系未能得到很好地解决可能与相关节点经历过快速辐射分化有关, 这不仅从系统发育树所呈现出的该亚科干群节点处存在明显的长短分支情况可以看出(图1), 而且从分子钟估算结果中也不难发现(图2).建议今后增加取样类群并从基因组层面开展广义苋科类群的系统发育关系研究, 可能有助于确立多节草亚科的系统位置. ...
... ).有时也被认为是联系石竹科指甲草族(Paronychieae)与苋科-藜科联盟的中间过渡类群(Ulbrich, 1934; Kühn et al., 1993).在分子系统学研究中, 多节草亚科的系统位置也存在争议(见前言部分), 且相关研究揭示的该类群系统位置的支持率通常较低(Kadereit et al., 2003; Pratt, 2003; Müller and Borsch, 2005; Sukhorukov et al., 2015).在本研究中, 多节草亚科与狭义苋科以及传统藜科中的藜亚科、虫实亚科、甜菜亚科共同形成一个支持率很高的分支(图1, 分支A), 并与后3个亚科成姐妹群关系(图1, 分支A1), 但该姐妹群关系的支持率较低(MLBS=70%; PP=0.86).多节草亚科系统关系未能得到很好地解决可能与相关节点经历过快速辐射分化有关, 这不仅从系统发育树所呈现出的该亚科干群节点处存在明显的长短分支情况可以看出(图1), 而且从分子钟估算结果中也不难发现(图2).建议今后增加取样类群并从基因组层面开展广义苋科类群的系统发育关系研究, 可能有助于确立多节草亚科的系统位置. ...
From cacti to carnivores: improved phylotranscriptomic sampling and hierarchical homology inference provide further insight into the evolution of Caryophyllales 4 2018
... 分子系统学的应用对苋科、藜科系统关系的理解起到了极大的促进作用, 亚科、族、属等类群范围的界定不断完善(Kadereit et al., 2003; Müller and Borsch, 2005; Masson and Kadereit, 2013; Sukhorukov et al., 2015; Walker et al., 2018; Yao et al., 2019).对于苋科与藜科的系统学归属, 目前主要有两派观点.一派以APG系统为主, 即支持广义苋科的观点(APG IV, 2016; Yao et al., 2019); 另一派则支持将传统藜科中的多节草亚科并入狭义苋科, 而将藜科核心类群依然接受为藜科(Hernández-Ledesma et al., 2015; Walker et al., 2018).本研究基于8个叶绿体基因序列分析结果表明, 藜科核心类群中的藜亚科、虫实亚科、甜菜亚科、多节草亚科与狭义苋科共同形成一支, 而藜科核心类群中其它4个亚科共同形成另一支, 且这2个分支均获得很高的支持率.这意味着藜科核心类群并非单系, 因此将其依然接受为藜科的观点在本研究中未能获得支持.基于此, 我们支持APG系统将狭义苋科与藜科合并为广义苋科的分类处理. ...
... 在Walker等(2018)基于转录组数据构建的石竹目系统发育关系中, 传统界定的藜科(含多节草亚科)形成一个单系分支, 且与狭义苋科成姐妹群关系.该研究还表明, 在藜科系统关系拓扑结构的主干靠基部连续3个节点, 均有约75%的基因树并不支持所呈现的拓扑结构, 这说明该研究获取的转录组数据中相关基因之间系统发育信号冲突明显, 因此所呈现的拓扑结构可能并未揭示出相关类群真实的进化关系.此外, 在Yang等(2018)基于转录组数据所构建的石竹目系统发育关系中, 猪笼草科与茅膏菜科关系较近, 而与露松科关系较远, 且相关节点均获得很高的支持率, 这与基于叶绿体基因组数据所得结果一致(Yao et al., 2019).但Walker等(2018)的研究结果则支持猪笼草科与露松科关系更近, 与茅膏菜科关系相对较远, 相关节点支持率同样很高.在基于转录组数据的系统发育分析中, 猪笼草科、茅膏菜科及露松科之间的系统关系在不同研究中出现明显冲突, 可能与相关研究在转录组数据分析背景下所筛选出来用于系统发育树构建的“直系同源基因”存在差异有关, 这意味着基于转录组数据筛选出的大量“直系同源基因”所构建的系统发育关系仍然值得商榷.另一方面, 在系统发育分析中, 基于叶绿体基因序列数据所得结果与核基因序列数据所得结果存在冲突的先例在被子植物类群中普遍存在, 这一现象通常被认为可能与杂交、叶绿体捕获以及不完全的谱系分选等进化事件有关(Gurushidze et al., 2010; Yi et al., 2015; Guo et al., 2018).因此, 阐释广义苋科中基于叶绿体数据与核基因数据所得结果存在冲突的原因, 将是后期研究中需要关注和探讨的重要问题. ...
... ).但Walker等(2018)的研究结果则支持猪笼草科与露松科关系更近, 与茅膏菜科关系相对较远, 相关节点支持率同样很高.在基于转录组数据的系统发育分析中, 猪笼草科、茅膏菜科及露松科之间的系统关系在不同研究中出现明显冲突, 可能与相关研究在转录组数据分析背景下所筛选出来用于系统发育树构建的“直系同源基因”存在差异有关, 这意味着基于转录组数据筛选出的大量“直系同源基因”所构建的系统发育关系仍然值得商榷.另一方面, 在系统发育分析中, 基于叶绿体基因序列数据所得结果与核基因序列数据所得结果存在冲突的先例在被子植物类群中普遍存在, 这一现象通常被认为可能与杂交、叶绿体捕获以及不完全的谱系分选等进化事件有关(Gurushidze et al., 2010; Yi et al., 2015; Guo et al., 2018).因此, 阐释广义苋科中基于叶绿体数据与核基因数据所得结果存在冲突的原因, 将是后期研究中需要关注和探讨的重要问题. ...
Menispermaceae and the diversification of tropical rainforests near the Cretaceous-Paleogene boundary 1 2012
... 本研究表明, 在广义苋科中, 分支A早期在白垩纪-古近纪边界附近时期(约66 Ma)可能发生过明显的快速分化.该分支于67.3 Ma (71.6-63.5 Ma)开始分化出分支A1与A2, 而分支A1中紧接着在65.3 Ma (70.1-60.7 Ma)分化出多节草亚科及其姐妹群.从显示支长的系统树(图1)上能明显看出, 相关节点存在明显的长短支现象, 意味着此处可能发生过快速辐射演化.在白垩纪-古近纪边界附近时期, 小行星撞击地球使地球环境发生巨大变化, 导致众多生物类群走向灭绝(Schulte et al., 2010), 但同时也为其它幸存者提供了广阔的生存空间, 为众多新的生物物种爆发提供了重要的生态机会.前人研究结果表明众多被子植物类群(如防己科(Menispermaceae) (Wang et al., 2012)、兰科(Orchidaceae) (Givnish et al., 2015)和楝科(Meliaceae) (Koenen et al., 2015))在此时可能都经历过明显的分化.苋科相关分支类群的起源与分化极有可能也与这一事件有关. ...
Evolution of the angiosperms: calibrating the family tree 5 2001
... 在分子系统学研究中, 狭义苋科的单系性获得了很好的支持, 然而藜科被认为并非单系(Kadereit et al., 2003; Sukhorukov et al., 2015; Yao et al., 2019).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... 分子系统学的应用对苋科、藜科系统关系的理解起到了极大的促进作用, 亚科、族、属等类群范围的界定不断完善(Kadereit et al., 2003; Müller and Borsch, 2005; Masson and Kadereit, 2013; Sukhorukov et al., 2015; Walker et al., 2018; Yao et al., 2019).对于苋科与藜科的系统学归属, 目前主要有两派观点.一派以APG系统为主, 即支持广义苋科的观点(APG IV, 2016; Yao et al., 2019); 另一派则支持将传统藜科中的多节草亚科并入狭义苋科, 而将藜科核心类群依然接受为藜科(Hernández-Ledesma et al., 2015; Walker et al., 2018).本研究基于8个叶绿体基因序列分析结果表明, 藜科核心类群中的藜亚科、虫实亚科、甜菜亚科、多节草亚科与狭义苋科共同形成一支, 而藜科核心类群中其它4个亚科共同形成另一支, 且这2个分支均获得很高的支持率.这意味着藜科核心类群并非单系, 因此将其依然接受为藜科的观点在本研究中未能获得支持.基于此, 我们支持APG系统将狭义苋科与藜科合并为广义苋科的分类处理. ...
... ; Yao et al., 2019); 另一派则支持将传统藜科中的多节草亚科并入狭义苋科, 而将藜科核心类群依然接受为藜科(Hernández-Ledesma et al., 2015; Walker et al., 2018).本研究基于8个叶绿体基因序列分析结果表明, 藜科核心类群中的藜亚科、虫实亚科、甜菜亚科、多节草亚科与狭义苋科共同形成一支, 而藜科核心类群中其它4个亚科共同形成另一支, 且这2个分支均获得很高的支持率.这意味着藜科核心类群并非单系, 因此将其依然接受为藜科的观点在本研究中未能获得支持.基于此, 我们支持APG系统将狭义苋科与藜科合并为广义苋科的分类处理. ...
... 对于广义苋科的起源时间, 前人多项研究均有涉及.Wikstr?m等(2001)研究表明, 该科干群节点起源可能发生在约28、38或40 Ma; 而Kadereit等(2012)认为该科干群起源时间发生在87-47 Ma; Magallón等(2015)则认为该科干群节点可能发生在64.2 Ma (81.8-43.6 Ma)或76.4 Ma (80.5-72.0 Ma).本研究表明, 该科干群节点起源可能发生在白垩纪晚期约80.5 Ma (82.5-78.3 Ma).在广义苋科中, 目前已知最早的可信化石记录为Polyporina cribraria Srivastava, 该化石物种为发现于白垩纪晚期马斯里奇特阶(Maestrichtian, 72.1-66.0 Ma)的花粉化石.在形态特征上, 该化石与传统藜科类群(尤其是现存物种Salicornia australis Sol. ex F. Muell.)的花粉化石非常相似.该化石发现于加拿大海洋环境向陆地环境的过渡地带, 与现存众多藜科类群的生境也非常相似.此外, 自马斯里奇特阶之后在相似生境中也发现过一系列类似的花粉化石(Muller, 1981; Kadereit et al., 2003).基于形态特征及其发现地的生境特征, 该花粉化石被广泛接受为广义苋科类群并多次用于该科类群分化时间校准分析(Kadereit et al., 2005; Yao et al., 2019; Li et al., 2019).因此, 从化石证据角度看, Wikstr?m等(2001)所得结果在时间尺度上明显偏早.而本研究结果在时间区间上与Kadereit等(2012)及Magallón等(2015)所得结果存在很大重叠, 较为一致.另外, 本研究得出广义苋科类群冠群节点分化时间(约69.9 Ma (73.3-67.0 Ma))也与该科中可信的最早化石记录所在地层时间较为一致. ...
... ).因此, 从化石证据角度看, Wikstr?m等(2001)所得结果在时间尺度上明显偏早.而本研究结果在时间区间上与Kadereit等(2012)及Magallón等(2015)所得结果存在很大重叠, 较为一致.另外, 本研究得出广义苋科类群冠群节点分化时间(约69.9 Ma (73.3-67.0 Ma))也与该科中可信的最早化石记录所在地层时间较为一致. ...
Improved transcriptome sampling pinpoints 26 ancient and more recent polyploidy events in Caryophyllales, including two allopolyploidy events 2 2018
... 在分子系统学研究中, 狭义苋科的单系性获得了很好的支持, 然而藜科被认为并非单系(Kadereit et al., 2003; Sukhorukov et al., 2015; Yao et al., 2019).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... 在Walker等(2018)基于转录组数据构建的石竹目系统发育关系中, 传统界定的藜科(含多节草亚科)形成一个单系分支, 且与狭义苋科成姐妹群关系.该研究还表明, 在藜科系统关系拓扑结构的主干靠基部连续3个节点, 均有约75%的基因树并不支持所呈现的拓扑结构, 这说明该研究获取的转录组数据中相关基因之间系统发育信号冲突明显, 因此所呈现的拓扑结构可能并未揭示出相关类群真实的进化关系.此外, 在Yang等(2018)基于转录组数据所构建的石竹目系统发育关系中, 猪笼草科与茅膏菜科关系较近, 而与露松科关系较远, 且相关节点均获得很高的支持率, 这与基于叶绿体基因组数据所得结果一致(Yao et al., 2019).但Walker等(2018)的研究结果则支持猪笼草科与露松科关系更近, 与茅膏菜科关系相对较远, 相关节点支持率同样很高.在基于转录组数据的系统发育分析中, 猪笼草科、茅膏菜科及露松科之间的系统关系在不同研究中出现明显冲突, 可能与相关研究在转录组数据分析背景下所筛选出来用于系统发育树构建的“直系同源基因”存在差异有关, 这意味着基于转录组数据筛选出的大量“直系同源基因”所构建的系统发育关系仍然值得商榷.另一方面, 在系统发育分析中, 基于叶绿体基因序列数据所得结果与核基因序列数据所得结果存在冲突的先例在被子植物类群中普遍存在, 这一现象通常被认为可能与杂交、叶绿体捕获以及不完全的谱系分选等进化事件有关(Gurushidze et al., 2010; Yi et al., 2015; Guo et al., 2018).因此, 阐释广义苋科中基于叶绿体数据与核基因数据所得结果存在冲突的原因, 将是后期研究中需要关注和探讨的重要问题. ...
Plastid phylogenomic insights into the evolution of Caryophyllales 15 2019
... 苋科(Amaranthaceae sensu lato)隶属石竹目核心分支(core Caryophyllales), 其与玛瑙果科(Achatocarpaceae)是姐妹群(Yao et al., 2019).目前所接受的苋科为其广义概念(Stevens, 2001; APG IV, 2016; Yao et al., 2019), 包含传统界定的狭义苋科(Amaranthaceae sensu stricto)和藜科(Chenopodiaceae)两大类群, 共有约165属2 040种, 是石竹目中仅次于石竹科(Caryophyllaceae)的第二大科(Christenhusz and Byng, 2016).苋科是包含众多作物及蔬菜等具有重要经济价值的植物类群, 如苋菜(Amaranthus tricolor L.)、甜菜(Beta vulgaris L.)、藜麦(Chenopodium quinoa Willd.)、菠菜(Spinacia oleracea L.) (中国科学院中国植物志编辑委员会, 1979; Stevens, 2001; Hernández-Ledesma et al., 2015).在传统分类系统中, 狭义苋科与藜科通常基于苞片、花被及雄蕊的特征被处理为2个独立的科(中国科学院中国植物志编辑委员会, 1979; Cronquist, 1988; Takhtajan, 1997), 其中藜科苞叶及花被常为草质或肉质, 雄蕊常分离; 而苋科苞叶及花被常为干膜质, 雄蕊基部常合生(中国科学院中国植物志编辑委员会, 1979).在地理分布上, 苋科主要分布于热带、亚热带地区; 而藜科则主要分布于温带至亚热带地区, 且常见于干旱、半干旱甚至盐碱地等生境(中国科学院中国植物志编辑委员会, 1979; Hernández-Ledesma et al., 2015). ...
... ; Yao et al., 2019), 包含传统界定的狭义苋科(Amaranthaceae sensu stricto)和藜科(Chenopodiaceae)两大类群, 共有约165属2 040种, 是石竹目中仅次于石竹科(Caryophyllaceae)的第二大科(Christenhusz and Byng, 2016).苋科是包含众多作物及蔬菜等具有重要经济价值的植物类群, 如苋菜(Amaranthus tricolor L.)、甜菜(Beta vulgaris L.)、藜麦(Chenopodium quinoa Willd.)、菠菜(Spinacia oleracea L.) (中国科学院中国植物志编辑委员会, 1979; Stevens, 2001; Hernández-Ledesma et al., 2015).在传统分类系统中, 狭义苋科与藜科通常基于苞片、花被及雄蕊的特征被处理为2个独立的科(中国科学院中国植物志编辑委员会, 1979; Cronquist, 1988; Takhtajan, 1997), 其中藜科苞叶及花被常为草质或肉质, 雄蕊常分离; 而苋科苞叶及花被常为干膜质, 雄蕊基部常合生(中国科学院中国植物志编辑委员会, 1979).在地理分布上, 苋科主要分布于热带、亚热带地区; 而藜科则主要分布于温带至亚热带地区, 且常见于干旱、半干旱甚至盐碱地等生境(中国科学院中国植物志编辑委员会, 1979; Hernández-Ledesma et al., 2015). ...
... 在分子系统学研究中, 狭义苋科的单系性获得了很好的支持, 然而藜科被认为并非单系(Kadereit et al., 2003; Sukhorukov et al., 2015; Yao et al., 2019).藜科多节草亚科(Polycnemoideae)与狭义苋科是姐妹群关系, 而藜科核心类群(core Chenopodiaceae) (藜科除多节草亚科以外的其它所有类群)形成1个单系, 并与多节草亚科-狭义苋科分支组成姐妹群, 但相关节点支持率很低或未能获得支持率(Kadereit et al., 2003; Sukhorukov et al., 2015).基于这一结果, 有****支持将多节草亚科归入狭义苋科, 而将藜科核心类群依然保留为藜科(Hernández- Ledesma et al., 2015; Sukhorukov et al., 2015).但也有分子系统学研究表明, 多节草亚科与狭义苋科-藜科核心类群分支组成支持率较低的姐妹群(Pratt, 2003; Müller and Borsch, 2005).Soltis等(2011)的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... 的研究甚至支持藜科核心类群也非单系, 但该研究对藜科取样较少(2种)且相关节点支持率很低.在Yao等(2019)开展的石竹目叶绿体基因组系统发育研究中, 基于83个叶绿体基因序列片段的分析结果也表明藜科核心类群并非单系, 但该研究以石竹目系统学研究为主, 所取藜科代表类群有限.在众多分子系统学研究中, 由狭义苋科与藜科共同形成的分支获得了很好的支持, 并被称为“苋科-藜科联盟”(Amaranthaceae- Chenopodiaceae alliance), 且被认为是石竹目中一个理想的进化分支(Cuénoud et al., 2002; APG IV, 2016; Yang et al., 2018; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... ; Yao et al., 2019).该分支具有一系列共源性状: 小花近无柄并排列成聚伞花序状; 花被片5, 单列, 覆瓦状排列; 胚珠常单个基生; 花粉具散孔; 筛管分子质体无中央内含晶体; 具有异黄酮类化合物(Kadereit et al., 2003).基于以上研究结果, 考虑到狭义苋科与藜科分支复杂的内部系统关系, 大多数****支持将二者合并从而建立广义苋科(Stevens, 2001; Brockington et al., 2009; Yao et al., 2019), 并被APG (Angiosperm Phylogeny Group)系统所采纳(APG IV, 2016).基于分子系统学结果并结合一系列形态特征, Stevens (2001)将目前所界定的广义苋科划分为13个亚科: 多节草亚科、苋亚科(Amaranthoideae)、白花苋亚科(Aervoideae)、牛膝亚科(Achyranthoideae)、青葙亚科(Celosioideae)、千日红亚科(Gomphrenoideae)、碱蓬亚科(Suaedoideae)、盐角草亚科(Salicornioideae)、樟味藜亚科(Camphorosmoideae)、猪毛菜亚科(Salsoloideae)、虫实亚科(Corispermoideae)、藜亚科(Chenopodioideae)和甜菜亚科(Betoideae).赤珠苋属(Bosea L.)与炬苋树属(Charpentiera Gaudichaud)在亚科水平的归属地位待定.在广义苋科内部, 部分亚科之间的系统关系, 尤其是多节草亚科的系统位置还存在争议. ...
... 对广义苋科在亚科水平进行广泛取样, 包含目前该科中所有亚科以及亚科系统位置归属待定的赤珠苋属与炬苋树属, 共涉及广义苋科44属59种(附表1).基于已发表的分子系统学研究结果(Yao et al., 2019), 选取玛瑙果科与石竹科为外类群(附表1).本研究选取atpB、ndhF、psbB、rbcL、rpoC2、rps4、rps16和matK/trnK共8个叶绿体基因序列片段构建苋科系统发育框架.其中部分序列数据为前期研究所得(Yao et al., 2019), 其它序列从NCBI网站(https://www.ncbi.nlm. nih.gov/)下载.序列详细信息见附表1; 部分个体序列不全, 其缺失的序列数据在相关序列矩阵中以缺失数据形式补齐. ...
... 共8个叶绿体基因序列片段构建苋科系统发育框架.其中部分序列数据为前期研究所得(Yao et al., 2019), 其它序列从NCBI网站(https://www.ncbi.nlm. nih.gov/)下载.序列详细信息见附表1; 部分个体序列不全, 其缺失的序列数据在相关序列矩阵中以缺失数据形式补齐. ...
... 为揭示广义苋科及其主要分支的起源分化时间, 采用BEAST v. 1.8.4软件(Drummond et al., 2012)对其进行分化时间估算.基于广义苋科中所报道的化石记录以及前期研究所得苋科与其近缘类群间的分化时间对相关节点进行时间校准.具体如下: (1) 参考Kadereit等(2003, 2005)用化石物种Parvangula randeckensis Hiltermann & Schmitz的中新世早期种子化石对藜属(Chenopodium)所代表的藜族第1分支(Chenopodieae I clade)干群(stem node)的起源时间进行校准, 化石时间的使用采用对数正态分布模型(lognormal distribution model), 最小时间限定在16.0百万年前(offset=16.0 Million-years ago (Ma)), 平均值限定为1.0 (mean=1.0), 标准差限定为0.5 (standard deviation (SD)=0.5); (2) 参考Kadereit等(2003, 2005)、Yao等(2019)及Li等(2019)的研究, 用化石物种Polyporina cribaria Srivastava的白垩纪晚期花粉化石对广义苋科冠群(crown node)起源时间进行校准, 化石时间同样采用对数正态分布模型, 最小时间限定为66.0 Ma, 平均值限定为1.0, 标准差限定为0.5; (3) 参考Yao等(2019)所得结果, 对苋科与玛瑙果科分化时间采用均匀分布模型(uniform distribution model), 时间上限限定为82.5 Ma, 下限限定为77.9 Ma; (4) 参考Yao等(2019)所得结果, 对石竹科与苋科-玛瑙果科分支分化时间采用均匀分布模型, 时间上限限定为92.2 Ma, 下限限定为85.7 Ma.分化时间在“出生-灭绝”物种形成模型(birth-death speciation process)之下, 基于对数正态分布的松散分子钟(lognormal relaxed molecular clock)进行估算; 各序列碱基替代模型参考1.2节序列最佳碱基替代模型筛选结果; 马尔科夫链蒙特卡洛(Markov chain Monte Carlo, MCMC)运算300 000 000代, 每5 000代取样1次, 该运算步骤重复3次独立运算; 基于Tracer v. 1.6软件(Rambaut et al., 2014)对所得参数进行检查, 确认所有参数的有效居群规模(effective sample sizes, ESS)数值均超过200, 结果可靠; 基于BEAST软件包中TreeAnotator v. 1.8.4软件, 将最初所得15 000棵系统树作为老化样品舍弃, 对剩下的所有系统树进行总结并计算其一致性树, 统计相关节点的分化时间.在Figtree v1.4.2软件(Rambaut, 2012)中查看所得结果. ...
... 对于广义苋科的起源时间, 前人多项研究均有涉及.Wikstr?m等(2001)研究表明, 该科干群节点起源可能发生在约28、38或40 Ma; 而Kadereit等(2012)认为该科干群起源时间发生在87-47 Ma; Magallón等(2015)则认为该科干群节点可能发生在64.2 Ma (81.8-43.6 Ma)或76.4 Ma (80.5-72.0 Ma).本研究表明, 该科干群节点起源可能发生在白垩纪晚期约80.5 Ma (82.5-78.3 Ma).在广义苋科中, 目前已知最早的可信化石记录为Polyporina cribraria Srivastava, 该化石物种为发现于白垩纪晚期马斯里奇特阶(Maestrichtian, 72.1-66.0 Ma)的花粉化石.在形态特征上, 该化石与传统藜科类群(尤其是现存物种Salicornia australis Sol. ex F. Muell.)的花粉化石非常相似.该化石发现于加拿大海洋环境向陆地环境的过渡地带, 与现存众多藜科类群的生境也非常相似.此外, 自马斯里奇特阶之后在相似生境中也发现过一系列类似的花粉化石(Muller, 1981; Kadereit et al., 2003).基于形态特征及其发现地的生境特征, 该花粉化石被广泛接受为广义苋科类群并多次用于该科类群分化时间校准分析(Kadereit et al., 2005; Yao et al., 2019; Li et al., 2019).因此, 从化石证据角度看, Wikstr?m等(2001)所得结果在时间尺度上明显偏早.而本研究结果在时间区间上与Kadereit等(2012)及Magallón等(2015)所得结果存在很大重叠, 较为一致.另外, 本研究得出广义苋科类群冠群节点分化时间(约69.9 Ma (73.3-67.0 Ma))也与该科中可信的最早化石记录所在地层时间较为一致. ...
Chloroplast capture and intra- and inter-continental biogeographic diversification in the Asian—New World disjunct plant genus Osmorhiza(Apiaceae) 1 2015
... 在Walker等(2018)基于转录组数据构建的石竹目系统发育关系中, 传统界定的藜科(含多节草亚科)形成一个单系分支, 且与狭义苋科成姐妹群关系.该研究还表明, 在藜科系统关系拓扑结构的主干靠基部连续3个节点, 均有约75%的基因树并不支持所呈现的拓扑结构, 这说明该研究获取的转录组数据中相关基因之间系统发育信号冲突明显, 因此所呈现的拓扑结构可能并未揭示出相关类群真实的进化关系.此外, 在Yang等(2018)基于转录组数据所构建的石竹目系统发育关系中, 猪笼草科与茅膏菜科关系较近, 而与露松科关系较远, 且相关节点均获得很高的支持率, 这与基于叶绿体基因组数据所得结果一致(Yao et al., 2019).但Walker等(2018)的研究结果则支持猪笼草科与露松科关系更近, 与茅膏菜科关系相对较远, 相关节点支持率同样很高.在基于转录组数据的系统发育分析中, 猪笼草科、茅膏菜科及露松科之间的系统关系在不同研究中出现明显冲突, 可能与相关研究在转录组数据分析背景下所筛选出来用于系统发育树构建的“直系同源基因”存在差异有关, 这意味着基于转录组数据筛选出的大量“直系同源基因”所构建的系统发育关系仍然值得商榷.另一方面, 在系统发育分析中, 基于叶绿体基因序列数据所得结果与核基因序列数据所得结果存在冲突的先例在被子植物类群中普遍存在, 这一现象通常被认为可能与杂交、叶绿体捕获以及不完全的谱系分选等进化事件有关(Gurushidze et al., 2010; Yi et al., 2015; Guo et al., 2018).因此, 阐释广义苋科中基于叶绿体数据与核基因数据所得结果存在冲突的原因, 将是后期研究中需要关注和探讨的重要问题. ...
 ,1,*
,1,*
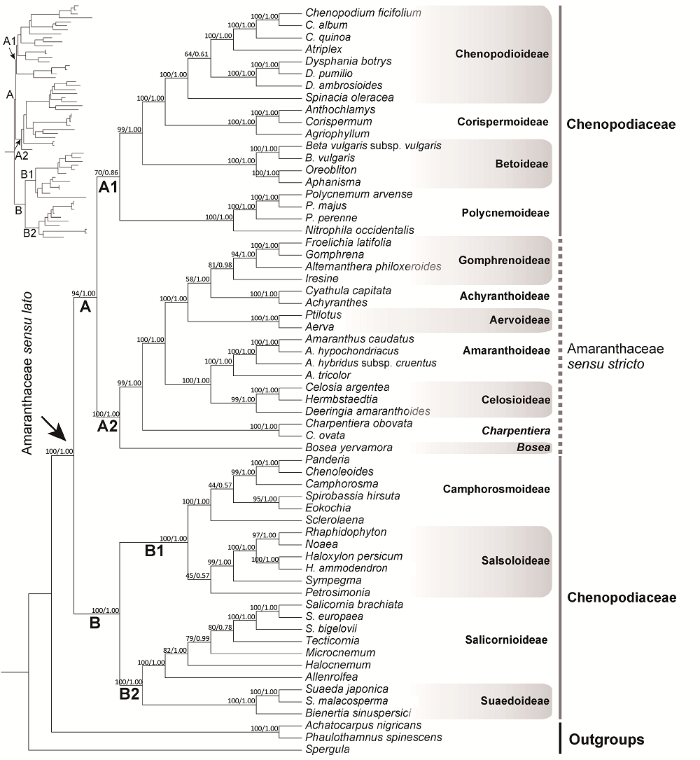 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT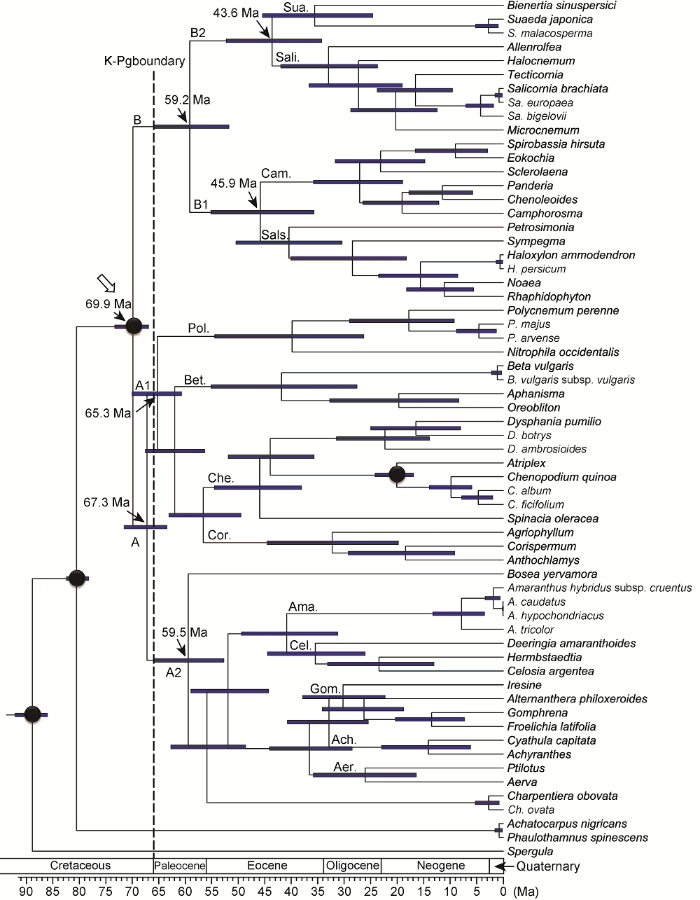 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}