赖先军
1 , 张义正
2 , 古英洪
2 , 颜朗
, 1 , * 1 西昌学院农业科学学院, 四川马铃薯高等学校重点实验室, 凉山 6150132 四川大学生命科学学院, 四川省分子生物学及生物技术重点实验室, 成都 610064Transformation of Insect Derived Antifreeze Gene into Sweet Potato (Ipomoea batatas ) and Enhanced Its Freeze-tolerance Xianjun Lai
1 , Yizheng Zhang
2 , Yinghong Gu
2 , Lang Yan
, 1 , * 1 Sichuan Key Laboratory of Potatoes, College of Agriculture Science, Xichang College, Liangshan 615013, China 2 Sichuan Key Laboratory of Molecular Biology & Biotechnology, College of Life Science, Sichuan University, Chengdu 610064, China 通讯作者: *E-mail: langyan0807@hotmail.com 责任编辑: 朱亚娜
收稿日期: 2019-07-5
接受日期: 2019-09-24
网络出版日期: 2020-01-01
基金资助: 国家科技支撑计划 (2007BAD78B03 )四川省“十一五”重点科技攻关项目 (No.07SG111-003-1 )
Corresponding authors: *E-mail: langyan0807@hotmail.com Received: 2019-07-5
Accepted: 2019-09-24
Online: 2020-01-01
摘要 为明确昆虫抗冻蛋白基因转入甘薯(Ipomoea batatas )后是否能提升其抗冻能力, 进而为培育甘薯抗冻育种材料奠定基础, 将黄粉虫(Tenebrio molitor )抗冻蛋白基因TmAFP 导入植物基因表达质粒, 经农杆菌介导的遗传转化获得抗冻甘薯新材料。以甘薯品种Huachano为受体材料建立甘薯植株高效再生体系, 并采用不同成分的体细胞胚成熟培养基培养胚性悬浮细胞。胚性愈伤组织对除草剂的敏感性测试结果表明, 转基因阳性植株筛选的最适培养基为MS+0.2 mg·L -1 2,4-D+0.8 mg·L -1 GAP+100 mg·L -1 Carb。将表达质粒分别转化Huachano后共获得7个胚性愈伤团并最终获得42株再生抗性植株, 其中转pSUIBEV3-AFP有23个株系, 转pCAMBIA-AFP有19个株系, 经PCR、Southern杂交和RT-PCR检测后证实TmAFP 基因已整合至甘薯基因组中并获得表达。将转基因甘薯及对照植株在-1°C下处理15小时后转移至室温, 结果表明, 转基因甘薯植株的抗冻能力显著提升。 关键词: 甘薯 ;
抗冻蛋白 ;
转基因 ;
甘薯植株再生 ;
分子育种 Abstract To explore whether the gene encoding antifreeze protein from insect can enhance the freezing tolerance of sweet potato through gene transformation, and to prepare freeze-tolerance materials for breeding purposes, we constructed a plant gene expression vector harboring an antifreeze gene TmAFP from yellow mealworms (Tenebrio molitor ) and obtained transgenic freeze-tolerance sweet potato lines using Agrobacterium -mediated transformation method. A high-frequency regeneration system of sweet potato was established using the variety Huachano as the recipient material, and the embryogenic suspension cells were cultured in the somatic embryo maturation medium. The sensitivity test of embryogenic cells to herbicides indicated that the combination of MS+0.2 mg·L -1 2,4-D+0.8 mg·L -1 GAP+100 mg·L -1 Carb is the most effective medium for screening the transgenic positive plants. Seven embryogenic calli were obtained and 42 resistant seedlings were regenerated, among which 23 harbored pSUIBEV3-AFP and 19 had pCAMBIA-AFP. All resistant seedlings were examined by PCR, Southern hybridization and RT-PCR, and the results showed that the TmAFP gene was integrated into the plant genome and expressed. The transgenic and non-transgenic plants were treated at -1°C for 15 hours, and then transferred to room temperature. The results demonstrated that the freeze-tolerance of the transgenic plants was greatly improved. Keywords: sweet potato ;
antifreeze protein ;
transgenic ;
plant regeneration of sweet potato ;
molecular breeding PDF (14013KB) 摘要页面 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 赖先军, 张义正, 古英洪, 颜朗 . 转昆虫抗冻蛋白基因增强甘薯抗冻能力. 植物学报, 2020,
55 (1): 9-20 doi:10.11983/CBB19133
Lai Xianjun, Zhang Yizheng, Gu Yinghong, Yan Lang .
Transformation of Insect Derived Antifreeze Gene into Sweet Potato (Ipomoea batatas ) and Enhanced Its Freeze-tolerance .
Chinese Bulletin of Botany , 2020,
55 (1): 9-20 doi:10.11983/CBB19133
甘薯(
Ipomoea batatas )是旋花科一年生植物, 也是世界上重要的块根类粮食作物, 广泛应用于食品、饲料、工业及生物能源领域(
Liu, 2011 )。甘薯生长和产量提升对于解决世界粮食资源安全等问题尤为重要(
Mwanga et al., 2017 )。然而, 受遗传因素制约, 甘薯对低温敏感, 极不耐霜冻, 在10°C以下, 甘薯细胞的原生质活动停滞, 生理机能受到影响, 导致冷害发生; 当温度低至-2°C时, 甘薯细胞间隙结冰, 其组织被破坏, 从而发生冻害。冷害或冻害的发生造成甘薯新陈代谢异常, 贮藏性能和抗病性降低, 贮藏期甘薯容易受到病菌侵染, 导致烂窖的发生(
瓜谷郁三, 2004 )。据统计, 每年约有30%的甘薯由于发生烂窖而失去利用价值, 损失约几十亿元人民币(
马代夫等, 2005 )。此外, 甘薯的生长适宜温度比水稻(
Oryza sativa )、玉米(
Zea mays )和棉花(
Gossypium spp.)等作物高5-10°C, 在春季幼苗期常由于倒春寒突然降温导致甘薯冻害的发生, 对甘薯生产造成重大损失。全球每年因低温冻害造成的农作物损失高达数千亿元, 因此, 培育抗冻甘薯新种质具有重要意义(
Zang et al., 2007 )。
由于甘薯具有杂交不亲和性及结实率低等特点, 严重限制了甘薯育种中的资源利用, 因此基因工程是改良甘薯品种的一种有效手段(
Yang et al., 2009 )。抗冻蛋白(antifreeze protein, AFP)又称热滞蛋白(thermal hysteresis protein, THP), 具有多个能够直接作用于冰晶的亲水性冰结合位点, 可阻止低温下冰晶在细胞间隙中的形成和再结晶, 以维持生物体正常的新陈代谢(
Wang et al., 2017 )。自De Vrie等(1969)从一种大西洋冷水鱼血液中发现抗冻蛋白以来, 其在细菌、昆虫、鱼类和植物中也相继被发现, 尤其是昆虫抗冻蛋白具有抗冻活性高和结构规律等特点, 其滞热活性与其它生物的抗冻蛋白相比高10-100倍, 这使得昆虫抗冻蛋白在植物抗冻遗传转化中具有巨大的优势和潜力(
Nada and Furukawa, 2011 )。Davies等(1987)首次将美洲拟鲽(
Pseudopleuronectes americanus )抗冻蛋白基因整合到Ti质粒中, 通过叶圆片法转化郁金香(
Tulipa gesneriana )和油菜(
Brassica campestris )等, 使转基因植株获得了一定的抗冻能力(
Zhang and Davies, 1987 )。
Cutler等(1989) 用真空透析法将冬比目鱼(
Pleuronectiformes heterosomata )抗冻蛋白基因导入马铃薯(
Solanum tuberosum )、拟南芥(
Arabidopsis thaliana )和欧洲油菜(
B. napus ), 使此类植物的自然结冰温度降低1.8°C, 证实转动物源抗冻蛋白基因可提高植物的抗寒性(
Cutler et al., 1989 )。
Wang等(2009) 用准噶尔小胸鳖甲(
Microdera punctipennis )抗冻蛋白基因
MPAFP149 转化烟草(
Nicotiana tabacum ), 并对T
0 代转基因烟草在-1°C处理48小时后, 发现转基因烟草相对电导率和表型明显优于野生型。室温恢复实验证实转基因烟草可存活并恢复生长, 而野生型烟草则遭受了不可逆的低温冻害。该研究表明, 转化后携带昆虫抗冻蛋白基因的烟草相比野生型具有明显的抗寒能力。
黄粉虫(
Tenebrio molitor )抗冻蛋白(TmAFP)二聚体的抗冻活性是已知抗冻蛋白中最高的, 可达鱼类抗冻蛋白活性的10-100倍。黄粉虫抗冻蛋白为右手β螺旋结构, 每一螺旋由12个保守的氨基酸组成(CT- XSXXCXXAXT, X为任一氨基酸残基), 依据12个氨基酸保守结构域重复次数可将
TmAFP 基因克隆分为3类, 分别编码84、96和120个氨基酸。TmAFP富含苏氨酸和半胱氨酸, 沿着蛋白折叠的一侧有两行苏氨酸残基与冰晶表面的棱柱和基面匹配结合(
Yue and Zhang, 2009 )。当接近冰点温度时, 抗冻蛋白表面冰结合位点上的苏氨酸侧链形成更优化的旋转异构体, 这种特有的冰结合构造使抗冻蛋白-冰分界面与冰晶相匹配, 从而阻止冰晶增长(
Ramya and Ramakrishnan, 2016 )。研究者将克隆到的
TmAFP 基因进行原核表达, 其表达蛋白经纯化后表现出冰结合活性和体外抗冻活性, 进一步将TmAFP蛋白代替传统抗冻保护剂DMSO制备大肠杆菌感受态细胞, 获得了转化高效的感受态细胞(
刘忠渊等, 2006 )。在提高植物抗冻能力方面, 有研究者将胡萝卜(
Daucus carota )抗冻蛋白基因(
DcAFP )和黄粉虫抗冻蛋白基因(
TmAFP )串联构建融合基因表达载体, 通过农杆菌介导转化拟南芥, 证实
TmAFP 的转基因拟南芥抗冻能力增强(
张振华等, 2012 )。然而, 至今未见有
TmAFP 基因在其它模式植物和经济作物中的转基因研究报道。因此, 通过基因工程技术将黄粉虫抗冻蛋白基因转入甘薯体内使其产生抗冻蛋白, 有望提高甘薯抗冻耐寒能力。
本研究将前期克隆到的黄粉虫抗冻蛋白基因编码区序列构建甘薯块根特异性表达载体, 将抗冻蛋白基因转化至甘薯基因组中, 并使其在低温下诱导表达, 获得抗冻甘薯新材料。研究结果有助于解决甘薯贮藏和苗期生长过程中易发生的冻害问题, 延长甘薯的生长季节和栽种范围, 同时为该基因的应用提供参考。
1 材料与方法 1.1 实验材料和试剂 供试材料为秘鲁甘薯(
Ipomoea batatas L.)品种Huachano, 由四川大学生命科学学院分子生物学实验室保存。植物表达质粒pCAMBIA1302和根癌农杆菌菌株EHA105 (自带利福平/链霉素抗性)由本实验室保存。pCAMBIA3300由山东农业大学提供。
Taq DNA聚合酶、T4 DNA连接酶以及DNA maker购自宝生物工程(大连)有限公司。KOD-FX高保真DNA聚合酶购自东洋纺(上海)生物科技有限公司。限制性内切酶购自立陶宛Fermentas公司。RNA提取试剂以及蛋白质电泳用试剂为Sigma公司产品。α-
32P (100 μCi, 3000 Ci·mmol
-1 )购自美国PerkinElmer公司。
1.2 甘薯体细胞胚的诱导及植株再生 取收获的甘薯块根于26°C温室发芽, 使其长出健壮的植株。待其长至5-6片叶时, 取其茎尖(长约10 cm)用流水冲洗30分钟, 75%乙醇浸泡30秒, 无菌水冲洗3次, 转入0.1%氯化汞中消毒8-10分钟, 再用无菌水冲洗4-5次。修整伤口并将其切成带1个茎节的茎段, 然后接种于含3%蔗糖和1%琼脂的MS基本培养基上培养, 培养温度为(26±2)°C, 光照强度为54 μmol·m
-2 ·s
-1 , 光周期为16小时光照/8小时黑暗, 使茎节上腋芽萌发形成无性系, 无性系每28天继代1次。
将试管苗去腋芽茎段切成长约5 mm的外植体, 接种于愈伤组织诱导培养基, 每个培养皿接种10个左右外植体, 置于(26±2)°C黑暗下培养, 然后接种于体细胞胚诱导培养基, 诱导获得体细胞胚。选取正常萌发的体细胞胚(具完整的上、下胚轴)接种于再生培养基中再生植株。每瓶培养基接种4个外植体, 培养温度为(26±2)°C, 光照强度为54 μmol·m
-2 ·s
-1 , 光周期为16小时光照/8小时黑暗。4周后统计分析成苗与植株的生长情况。
1.3 抗冻蛋白基因表达载体的构建 由于不同筛选标记对转基因甘薯幼苗植株存活率的影响较大, 因此我们以pCAMBIA3300质粒上携带的除草剂抗性基因(
bar 基因)替换pCAMBIA1302质粒上的潮霉素抗性基因(
hpt II), 构建新的组成型植物双元表达质粒pCAMBIA3302。该质粒以
bar 基因作为植物标记基因, 以CaMV35S启动子调控的
GFP 基因作为报告基因, 其阳性克隆经质粒PCR鉴定并经
Xho I、
Kpn I、
Sal I、
Hin dIII、
Nco I和
Bst EII酶切验证。为使目的基因在甘薯块根中特异表达, 以本实验室克隆的甘薯贮藏蛋白基因启动子(SP)替换pCAMBIA3302上的启动子CaMV35S, 从而构建甘薯块根特异表达载体, 命名为pSUIBEV3。对该质粒进行PCR和酶切并测序验证。
通过
Pfu DNA聚合酶扩增
TmAFP 目的基因, 引物序列为TmAFP1-F (5'-CATGCCATGGCATTCAAAACGTGTG-3')和TmAFP1-R (5'-TGGTCACCTTAATGTCCGGGACATCCTG-3')。扩增体系为25 μL, 其中2.5 μL 10×Pfu缓冲液, 1.5 μL MgCl
2 (25 mmol·L
-1 ), 2.0 μL dNTP (2.5 mmol·L
-1 ), 0.5 μL引物(10 μmol·L
-1 ), 1 μL模板, 0.24 μL
Pfu DNA聚合酶(2.5 U·μL
-1 ), 16.76 μL PCR级无菌水。PCR产物纯化加A后在T4 DNA连接酶的作用下与pMD 18-T质粒连接, 后转化大肠杆菌JM109感受态细胞。LB平板(氨苄青霉素抗性)中挑取阳性克隆, 经菌落PCR和质粒酶切验证后将其命名为pMD-AFP质粒。pMD-AFP质粒经
Nco I和
Bst EII双酶切后, 与用
Hin dIII和
Bst EII双酶切pCAMBIA3302后回收的大片段, 以及用
Hin dIII和
Nco I双酶切pUCSP后回收的甘薯贮藏蛋白基因启动子(SP)片段连接, 构建
TmAFP 基因的甘薯块根特异表达载体pSUIBEV3-AFP; 用
Nco I和
Bst EII双酶切pCAMBIA3302以及pMD-AFP质粒, 分别回收大小片段并连接, 构建
TmAFP 基因的组成型表达载体pCAMBIA-AFP。
1.4 甘薯转基因植株的获得 采用液氮冻融法将质粒转化至根癌农杆菌EHA105感受态细胞。大致过程为: 将1 μg重组质粒加入200 μL感受态细胞中, 混匀后冰上放置30分钟, 然后于液氮中冻5分钟, 37°C水浴中融化5分钟; 将其转入含2 mL YEB (不含抗生素)中, 25°C振荡培养2-4小时, 最大转速离心1分钟, 弃上清, 重悬于1 mL YEB培养基(含10 mg·L
-1 利福平(Rif), 100 mg·L
-1 链霉素(Strep), 50 mg·L
-1 卡那霉素(Kan))并涂布于含相同抗生素的YEB固体培养基上, 25°C倒置培养2天后挑取单克隆进行鉴定。
采用根癌农杆菌介导法进行植物的遗传转化。取振荡培养26-32周的胚性悬浮细胞浸入农杆菌工作液中5分钟, 吸弃菌液, 将侵染后的胚性细胞团转至MS+0.2 mg·L
-1 2,4-D固体培养基中, 28°C黑暗条件下共培养3天; 共培养后的胚性细胞团用MS+0.2 mg·L
-1 2,4-D+500 mg·L
-1 Carb液体培养基洗涤5-6次, 转至MS+0.2 mg·L
-1 2,4-D+100 mg·L
-1 Carb液体培养基中, 28°C黑暗条件下振荡培养1周; 将胚性细胞团转至MS+0.2 mg·L
-1 2,4-D+100 mg·L
-1 Carb+ 0.8 mg·L
-1 GAP (草铵膦, PESTANAL®)固体选择培养基中, 28°C黑暗条件下选择培养数周(每2周继代1次); 将抗性愈伤组织转移到MS+100 mg·L
-1 Carb+ 1.0 mg·L
-1 GAP固体培养基上(培养条件为28°C, 13小时光照/11小时黑暗), 诱导体细胞胚的形成和萌发, 得到抗性芽苗, 并进一步将其转移至MS+1.0 mg·L
-1 GAP固体培养基使其发育成完整植株。
1.5 转基因甘薯总DNA的提取和PCR验证 采用CTAB法提取DNA。取10 g甘薯组织经液氮研磨后转入50 mL离心管中, 加入20 mL预热至90°C的2× CTAB提取缓冲液并于65°C水浴中孵育90分钟; 将混合物冷却至室温后加入等体积的氯仿/异戊醇(24:1), 室温下5 000 ×
g 离心10分钟, 转移上清后加入1/100体积的RNA酶贮液, 颠倒混匀后于37°C保温30分钟; 将丝状DNA勾出后转移至装有70%乙醇的离心管中清洗2次, 空气中干燥后溶于200 μL TE缓冲液。利用琼脂糖凝胶电泳和分光光度计检测DNA完整性和质量。
采用KOD-FX高保真聚合酶扩增
GFP 编码序列、
RPB 基因和
AFP 基因, 引物序列分别为GFP2-F (5'- CGCGGATCCAGCAAAGGAGAAGAACTTTTCACTG-3'), GFP2-R (5'-CCCAAGCTTCTCATCCATGCCATGTGTAATCCCA-3'), RPB2-F (5'-TAGGGATGAAGAGAAAAAGATGGG-3'), RPB2-R (5'-CAATGGTCATCCGAGAAGGAATAG-3'), TmAFP1-F和TmAFP1-R的序列与前一致。PCR反应体系为20 μL, 其中包括10 μL KOD-FX缓冲液, 4 μL dNTP (2 mmol·L
-1 ), 0.5 μL引物(10 μmol·L
-1 ), 3.6 μL PCR级无菌水, 0.4 μL KOD-FX聚合酶(1 U·μL
-1 )以及1 μL DNA模板。反应流程遵循KOD-FX高保真DNA聚合酶标准反应流程。
1.6 Southern杂交验证 用
Eco RI对甘薯总DNA进行酶切, 反应体系40 µL, 其中20 µL总DNA (约10 µg), 5 µL 10×
Eco RI酶切缓冲液, 1 µL RNase (10 mg·mL
-1 ), 12 µL dH
2 O, 2 µL
Eco RI内切酶(10 U·µL
-1 )。轻柔混匀后置于37°C孵育过夜。酶切反应产物经0.8%琼脂糖凝胶电泳(0.5% TBE, 电压25-50 V) 4-6小时, 然后将DNA由琼脂糖凝胶转移至Hybond-N+尼龙膜。转膜过程大致为: 在磁盘中倒入适量的转膜缓冲液, 在玻璃板上架2层长滤纸搭成盐桥, 依胶大小裁好尼龙膜后紧贴于胶上, 防止有气泡产生, 再压2张同样大小的滤纸, 堆积吸水纸, 上方再放一小玻板, 板上加500 g左右的重物。转膜16-20小时后将尼龙膜取下于2× SSC溶液中浸泡20分钟, 滤纸吸干后于烘箱中烘干2小时(80°C)。通过PCR方法将
AFP 基因片段中dCTP进行[α-
32 p]同位素标记制备探针, 反应体系为50 μL, 其中包括5 μL 10×缓冲液, 1 μL dNTP (10 mmol·L
-1 ), 1 μL dCTP (0.1 mmol·L
-1 ), 5 μL引物(10 μmol·L
-1 ), 5 μL α-
32 P dCTP, 22 μL PCR级无菌水, 1 μL
Taq DNA聚合酶(1 U·μL
-1 )以及5 μL DNA模板。将膜浸入5× SSC溶液中2分钟, 再浸入65°C预热的预杂交液中预杂交1小时, 随后将膜浸入5 mL杂交液及5 μL同位素探针中, 65°C杂交过夜。膜的洗涤过程为经过20 mL的2× SSC, 0.1% SDS溶液中洗涤2次, 0.1× SSC, 0.1% SDS溶液中洗涤2次, 再将膜转入20 mL洗涤缓冲液中振荡洗涤5分钟。-70°C条件下将膜于X-ray胶片(柯达)曝光24小时。
1.7 RNA的提取及RT-PCR反应 采用植物RNA提取试剂盒(Omega, Cat No.R6834- 01)提取甘薯组织总RNA, 按试剂盒说明书进行操作。总RNA经琼脂糖凝胶电泳和分光光度计检测质量后进行反转录(Takara, Cat No.RR047A)。为了在转录水平检测转基因甘薯中抗冻蛋白基因的表达, 对
AFP 和
GFP 基因进行RT-PCR扩增, 引物分别为TmAFP1-F和TmAFP1-R、GFP2F和GFP2R (序列与前一致), 以
actin β 基因作为内参, 引物为IBActinF (5'-TTCCCCGGTATTGCGGATAGAATG-3')和IBActinR (5'-CGGACCGGACTCATCATACTCTG-3')。
1.8 转AFP 基因甘薯抗冻能力检测 将6株转基因甘薯植株及4株对照植株放入光照培养箱中, 在-1°C条件下低温处理, 光照强度为54 μmol·m
-2 ·s
-1 , 光周期为16小时光照/8小时黑暗, 处理24小时后测定叶片相对电导率。同时, 将另外3株转基因甘薯植株及3个对照植株置于相同条件下的光照培养箱中, 低温处理15小时后转移至室温条件观察并记录表型变化。
采用浸泡法测定转基因甘薯叶片相对电导率(低温胁迫下反映膜系统状态的重要生化指标): 取大小一致的叶片组织, 用蒸馏水冲洗3次后, 滤纸吸干表面水分, 将叶片剪成适宜长度的细条(避开主脉), 快速称取鲜样3份, 每份0.1 g, 分别置于装有10 mL去离子水的刻度试管中, 置于室温下浸泡处理12小时。用电导仪测定浸提液电导(R1), 然后沸水浴加热30分钟, 冷却至室温后摇匀, 再次测定浸提液电导(R2)。计算相对电导率(%)=(R1/R2)×100%。重复测试3次。
1.9 显著性分析 采用SPSS软件进行数据统计分析, 采用新复极差法(SSR法)进行差异显著性分析。
2 结果与讨论 2.1 甘薯高效再生体系的建立 为建立甘薯转基因体系, 我们将经液体振荡培养且生长良好的胚性悬浮细胞接种于MS+0.2 mg·L
-1 2,4-D液体培养基, 28°C暗培养8周后(每2周继代1次), 再转到体细胞胚成熟培养基中培养2周(28°C, 13小时光照/11小时黑暗), 然后再转到MS培养基上培养6周(28°C, 13小时光照/11小时黑暗), 使体细胞胚萌发并生成完整小植株(
图1A -D)。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1甘薯遗传转化体系 (A), (B) 分别在MS+0.2 mg·L
-1 2,4-D液体培养基中继代18和22周后的甘薯胚性悬浮细胞; (C) 在MS+0.2 mg·L
-1 2,4-D固体培养基上培养8周后的甘薯胚性悬浮细胞; (D) 甘薯体细胞胚再生的小植株。(A), (B), (D) Bars=1 cm; (C) Bar=4 cm
Figure 1Transformation system of sweet potato (A), (B) Sweet potato embryogenic suspension cells at 18 and 22 weeks cultivated in MS+0.2 mg·L
-1 2,4-D liquid medium, respectively; (C) Sweet potato embryogenic suspension cells at 8 weeks cultivated in MS+0.2 mg·L
-1 2,4-D solid medium; (D) Sweet potato seedlings regenerated from somatic embryo. (A), (B), (D) Bars=1 cm; (C) Bar=4 cm
为了在体细胞胚成熟培养过程中建立甘薯高效再生体系, 我们采用不同成分的体细胞胚成熟培养基培养胚性悬浮细胞, 并统计每个悬浮培养胚性愈伤团再生芽苗数(
表1 )。结果表明, 再生芽苗数受培养基成分的影响。MS培养基的再生效果最好, 平均每个胚性愈伤团可再生44.8个芽苗, 而MS+4.0 mg·L
-1 ABA+1.0 mg·L
-1 GA3的再生效果最差, 平均每个胚性愈伤团可再生21.8个芽苗。在5种体细胞胚成熟培养基中, MS培养基的再生效果显著优于其它培养基, 且极显著优于第3-5号培养基。一般而言, 在培养基中添加ABA和(或) GA3有利于胚细胞再生。而对于本实验选用的甘薯品种Huachano, 培养基中添加ABA或GA3均显著降低胚细胞再生芽苗株数, 其中GA3对胚细胞再生的影响达极显著水平。
Table 1 表1 表1 不同体细胞胚成熟培养基对甘薯芽苗再生的影响
Table 1
Effect of different somatic embryo maturation medium on the regenerated seedlings Treatment Components-1 ) Number of seedlings in average * ** 1 MS 44.8±5.22 a A 2 MS+ABA1.0 33.6±5.18 b AB 3 MS+GA3 1.0 30.2±4.73 bc B 4 MS+ABA1.0+GA3 1.0 23.2±6.29 c B 5 MS+ABA4.0+GA3 1.0 21.8±6.58 c B
* indicate significant differences (P <0.05), ** indicate extremely significant differences (P <0.01). * 表示显著差异(P <0.05), ** 表示极显著差异(P <0.01)。 新窗口打开 |
下载CSV 2.2 胚性愈伤组织对除草剂(GAP)的敏感性 为了筛选转基因甘薯阳性植株, 确定理想的抗生素和除草剂用量, 我们测试了胚性愈伤组织对除草剂(GAP)的敏感性。首先取液体培养12周后的胚性悬浮细胞接种于MS+0.2 mg·L
-1 2,4-D培养基上, 并添加不同浓度(0、0.2、0.4、0.6和0.8 mg·L
-1 ) GAP进行暗培养, 培养8周后观察组织生长情况(每2周继代1次)。结果表明, 随着GAP浓度的增加, 胚性愈伤组织的增殖发育均被明显抑制。但即使GAP浓度达0.8 mg·L
-1 时, 还有极少量抗性愈伤组织存活(
图2A )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2不同浓度除草剂和抗生素对甘薯胚性愈伤组织和组培苗的影响 (A) 选择培养基上培养8周的甘薯胚性悬浮细胞。从左到右培养基依次为MS+0.2 mg·L
-1 2,4-D分别添加0、0.2、0.4、0.6和0.8 mg·L
-1 GAP; (B) 培养6周(从左至右第1, 2)和8周(3, 4)的胚性悬浮细胞, 培养基分别为MS+0.2 mg·L
-1 2,4-D (1, 3)和MS+0.2 mg·L
-1 2,4-D+100 mg·L
-1 Carb (2, 4); (C) 培养8周的胚性悬浮细胞, 培养基为MS+0.2 mg·L
-1 2,4-D+100 mg·L
-1 Carb分别添加0.8、1.0、1.2和1.4 mg·L
-1 GAP; (D) 分别为MS+0.2 mg·L
-1 2,4-D+0.8 mg·L
-1 GAP+100 mg·L
-1 Carb培养基上接种0天和培养3周的Huachano茎尖。(A), (B), (C) Bars=1 cm; (D) Bar=2 cm
Figure 2Effects of herbicide and antibiotic in different concentrations on embryogenic callus and regenerated seedlings of sweet potato (A) Sweet potato embryogenic suspension cells at 8 weeks cultivated in selective medium. The medium from left to right was MS+0.2 mg·L
-1 2,4-D with 0, 0.2, 0.4, 0.6, 0.8 mg·L
-1 GAP, respectively; (B) Embryogenic suspension cells at 6 weeks (the first and second from left to right) and 8 weeks (the third and fourth). The medium were MS+0.2 mg·L
-1 2,4-D (the first and third) MS+0.2 mg·L
-1 2,4-D+100 mg·L
-1 Carb (the second and fourth); (C) Embryogenic suspension cells at 8 weeks, the medium are MS+0.2 mg·L
-1 2,4-D+100 mg·L
-1 Carb with 0.8, 1.0, 1.2, 1.4 mg·L
-1 GAP, respectively; (D) Huachano stem tips cultivated 0 day and 3 weeks on medium of MS+0.2 mg·L
-1 2,4-D+0.8 mg·L
-1 GAP+100 mg·L
-1 Carb, respectively. (A), (B), (C) Bars=1 cm; (D) Bar=2 cm
取液体培养20周的胚性悬浮细胞分别接种于MS+ 0.2 mg·L
-1 2,4-D以及MS+0.2 mg·L
-1 2,4-D+100 mg·L
-1 Carb并添加不同浓度(0.8、1.0、1.2和1.4 mg·L
-1 ) GAP的培养基中暗培养, 每2周转移1次, 培养8周后观察。结果表明, 在添加100 mg·L
-1 Carb的情况下, GAP浓度为0.8 mg·L
-1 时, 能够完全抑制Huachano愈伤组织的生长, 且100 mg·L
-1 Carb对愈伤组织的生长影响较小(
图2B , C)。因此, 我们认为转基因阳性甘薯株系筛选的最适培养基为MS+0.2 mg·L
-1 2,4-D+0.8 mg·L
-1 GAP+100 mg·L
-1 Carb。为了验证该培养基对甘薯试管苗的生长同样具有抑制效果, 将试管苗的茎尖接种于该培养基中, 培养3周后组培苗全部死亡(
图2D )。上述结果表明, 该浓度组合的培养基完全能抑制Huachano非转基因苗的生长。
2.3 甘薯胚性悬浮细胞遗传转化体系的建立与TmAFP 转基因甘薯植株的获得 将低温保存的工程菌pCAMBIA-AFP/EHA105、pCAMBIA/EHA105、pSUIBEV3-AFP/EHA105和pSUI-BEV3/EHA105活化后, 用MS+0.2 mg·L
-1 2,4-D液体培养基洗涤1次, 重悬于25 mL的MS+0.2 mg·L
-1 2,4-D液体培养基中备用(
图3A )。取出振荡培养32周的胚性悬浮细胞浸入上述农杆菌工作液中5分钟, 转至MS+0.2 mg·L
-1 2,4-D固体培养基, 28°C黑暗条件下培养3天。共培养后的胚性细胞团用MS+0.2 mg·L
-1 2,4-D+500 mg·L
-1 Carb液体培养基洗涤5次, 然后转到MS+0.2 mg·L
-1 2,4-D+100 mg·L
-1 Carb液体培养基中, 28°C黑暗条件下振荡培养1周。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3甘薯抗性体胚及其转基因植株再生过程 (A) 用于转化的甘薯胚性悬浮细胞在MS+0.2 mg·L
-1 2,4-D液体培养基中28°C黑暗振荡培养32周; (B) 选择与非选择培养4周的胚性愈伤组织, a-c为选择性培养基(MS+0.2 mg·L
-1 2,4-D+100 mg·L
-1 Carb+0.8 mg·L
-1 GAP), 箭头所指为抗性愈伤组织; d为未加除草剂的对照; (C) 转基因植株再生过程, a: 再生组织; b: 生芽; c: 生叶; d: 成株; e: 成苗。Bars=1 cm
Figure 3Sweet potato resistant somatic embryo and the regeneration of transgenic plants (A) Sweet potato embryogenic suspension cells cultivated in MS+0.2 mg·L
-1 2,4-D liquid medium at 28°C for 32 weeks; (B) Embryogenic callus cultivated in selective and non-selective medium for 4 weeks, a-c: Embryogenic callus cultivated in selective medium (MS+0.2 mg·L
-1 2,4-D+100 mg·L
-1 Carb+0.8 mg·L
-1 GAP), resistant callus was marked by arrows; d: Embryogenic callus cultivated in control medium without herbicide; (C) The processes of transgenic plant regeneration, a: Reproductive tissue; b: Bud; c: Leaf; d: Seedling; e: Reproductive plant. Bars=1 cm
将延迟选择培养1周后的胚性细胞团转至MS+ 0.2 mg·L
-1 2,4-D+100 mg·L
-1 Carb+0.8 mg·L
-1 GAP固体选择培养基中, 28°C黑暗条件下选择培养数周即得到抗性愈伤组织(
图3B ), 而培养基中未添加除草剂(对照)的胚性愈伤组织则增殖迅速, 且体胚正在发育。将抗性愈伤组织转移到MS+100 mg·L
-1 Carb+0.8 mg·L
-1 GAP固体培养基上诱导体细胞胚的形成和萌发, 得到抗性芽苗, 并进一步将其转移到MS+1.0 mg·L
-1 GAP固体培养基上使其发育成完整植株(
图3C )。也可将抗性愈伤组织直接转移到MS固体培养基再生后, 将试管苗再用MS+1.0 mg·L
-1 GAP固体培养基进一步筛选, 从而得到抗性植株(结果未展示)。
2.4 TmAFP 转基因甘薯的鉴定与抗冻能力检测 我们用除草剂对再生植株进行再次筛选, 最终获得42株再生抗性植株, 其中转pSUIBEV3-AFP的23个, 转pCAMBIA-AFP的19个(
图4A )。快速PCR鉴定结果表明, 非抗除草剂植株和未转化对照植株均无
TmAFP 基因扩增条带, 而抗性植株均能扩增出
TmAFP 基因条带(
图4B )。同时, 提取转基因甘薯叶片DNA, 分别以转化质粒和未转化植株DNA作为正、负对照, 对目标
TmAFP 基因和内参RNA聚合酶II基因(
RPB2 )进行扩增(
图4C ), 结果表明转基因植株中能扩增出目标基因条带。我们进一步对转基因甘薯植株基因组DNA进行southern blotting分析, 以含
TmAFP 基因的质粒为正对照, 以非转基因植株基因组DNA为负对照, 利用同位素标记
TmAFP 基因为探针进行杂交。结果表明, 与正对照一致, 转
TmAFP 基因甘薯阳性植株基因组DNA能够与探针进行杂交, 而非转基因对照、转pSUIBEV3和pCAMBIA3302空质粒甘薯基因组DNA则未能和探针结合(
图4D )。Southern杂交结果还表明, 目标
TmAFP 基因在转基因甘薯基因组上的整合位点均只有1个, 但整合位置并不相同, 其中株系31-2、44-1、44-4和54-5的拷贝数高于其它株系。为了检测转基因甘薯的基因表达水平, 提取转基因植株总RNA, 以反转录获得的cDNA为模板, 对转pCAMBIA空载及pCAMBIA-AFP中的报告基因
GFP 和目标基因
TmAFP 进行RT-PCR扩增, 结果表明转基因甘薯中
GFP 和
TmAFP 基因均获得表达(
图4E )。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4TmAFP 转基因甘薯植株的鉴定 (A) 采用除草剂(1.0 mg·L
-1 GAP)对再生植株进行筛选(4-12、4-11、4-10、4-9: 抗性植株; 47-1、47-2: 非抗性植株; CK: 非转基因对照); (B) PCR快速鉴定转基因植株(
AFP 片段长度353 bp) (M: D2000 DNA marker (下同); +CK: pSUIBEV3-AFP质粒正对照; -CK: 未转化对照植株; 4-9、4-10、4-12: GAP抗性植株; 47-2: 非抗性植株); (C) 转基因植株的PCR检测(+CK: pSUIBEV3-AFP质粒作正对照扩增
AFP 基因; -CK: 未转化对照植株扩增
AFP 基因; 4-9-AFP: 转基因植株扩增
AFP 基因; 4-9-RPB2: 转基因植株扩增
RPB2 基因; -CK-RPB2: 未转化对照植株扩增
RPB2 基因); (D) Southern杂交(+CK: pCAMBIA-AFP质粒正对照; -CK: 未转化对照植株; 其余为不同转基因株系); (E) 转基因植株RT-PCR检测(RP1、RP2: 转pCAMBIA空载植株; 4-12、4-11、44-1、54-5: 转pCAMBIA-AFP株系)。
Figure 4Detection of TmAFP in the transgenic sweet potato plants (A) Screening with 1.0 mg·L
-1 GAP (Line 4-12, 4-11, 4-10, 4-9: Resistant seedlings; Line 47-1, 47-2: Non-resistant seedlings; CK: Non-transgenic control); (B) Amplified 353-bp fragment of
AFP gene (M: D2000 molecular weight marker; +CK: pSUIBEV3-AFP vector as positive control; -CK: Non-transgenic seedlings as negative control; Line 4-9, 4-10, 4-12: GAP resistant seedlings; Line 47-2: Non-resistant seedlings); (C) PCR detection of transgenic seedlings (+CK: Amplifying
AFP gene using pSUIBEV3-AFP as template; -CK: Amplifying
AFP gene using non-transgenic seedling; 4-9-AFP: Amplifying
AFP gene in transgenic seedling; 4-9-RPB2: Amplifying
RPB2 gene in transgenic seedling; -CK-RPB2: Amplifying
RPB2 gene in non-transgenic seedling); (D) Southern blotting analysis (+CK: pCAMBIA-AFP vector as positive control; -CK: Non-transgenic seedlings as negative control; The others represent different transgenic seedling lines); (E) RT-PCR detection of transgenic seedlings (RP1, RP2: Transgenic seedlings with empty pCAMBIA vector; Line 4-12, 4-11, 44-1, 54-5: Transgenic seedlings with pCAMBIA-AFP vector).
为了检测转
TmAFP 基因甘薯的抗冻能力, 我们将转基因甘薯及对照植株在-1°C条件下处理24小时后测定叶片电导率, 结果发现转基因甘薯叶片电导率上升9.5%, 而对照甘薯叶片电导率上升23%, 两者差异显著(
P =2.45×10
-6 ); 处理48小时后, 转基因甘薯叶片电导率上升28%, 而对照甘薯叶片电导率上升达53%, 两者差异显著(
P =1.33×10
-4 ) (
图5A )。此外, 我们将转基因甘薯植株及对照在-1°C条件下处理15小时后转移至室温条件并观察表型变化。结果表明, 转基因甘薯12片叶中只有2片老叶出现萎蔫现象, 而对照11片叶全部萎蔫。室温生长数天后, 转基因甘薯已恢复正常生长, 而对照植株全部死亡(
图5B )。上述结果表明, 本研究中转基因甘薯的抗冻能力大幅提升。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5TmAFP 转基因甘薯植株抗冻能力的检测 (A) 不同冷处理条件下植株电导率变化(CK1: 非转基因对照; CK2-4: 转空载的非抗冻植株; 4-9、4-10、4-11、4-12、44-1和54-5:
TmAFP 转基因株系; ** 表示差异极显著(
P <0.01)); (B) -1°C条件下处理15小时后转移至室温条件, 转基因甘薯植株及对照的表型变化(Overall: 盆中同时栽种CK1、4-9、4-10、4-11、4-12; Non-transgenic: CK1放大图; Transgenic: 转基因株系4-9放大图)。Bars=5 cm
Figure 5Detection of freezing-tolerance ability of TmAFP transgenic sweet potato plants (A) Conductivity assay under different freeze-treatments (CK1: Non-transgenic control; CK2-4: Transgenic plants with empty vector; 4-9, 4-10, 4-11, 4-12, 44-1, 54-5:
TmAFP transgenic lines; ** indicate extremely significant differences (
P <0.01)); (B) Phenotypic changes of transgenic sweet potato plants and controls after 15 h treatment at -1°C (Overall: CK1, 4-9, 4-10, 4-11, 4-12 planted in the same pot; Non-transgenic: Zoomed in CK1; Transgenic: Zoomed in transgenic line 4-9). Bars=5 cm
2.5 讨论 近年来, 随着分子生物学的快速发展, 利用基因工程对植物性状进行遗传改良, 以获得具有特定优异性状新育种材料的技术体系已日趋成熟(
Wang et al., 2003 )。由于植物长期暴露于盐碱、干旱和寒冷等非生物逆境, 其本身会触发许多共同的防御机制, 如ABA含量增加、可溶性糖和保护性蛋白质的积累以及抗氧化水平的提升。因此将非生物胁迫相关的植物源基因导入可提高甘薯对盐碱、干旱、低温和氧化逆境等的抗性(
翟红等, 2017 )。例如, 将
Cu /
Zn SOD 和
APX 基因导入甘薯, 可增强转基因甘薯的耐盐性(
王欣等, 2011 ); 将菠菜 (
Spinacia oleracea )
SoBADH 基因导入甘薯可增强转基因甘薯对盐、低温及氧化胁迫的抗性(
Fan et al., 2012 )。同时, 可通过克隆甘薯自身的抗性基因, 然后增强其表达的方式创制转基因甘薯新材料。例如, 分别从甘薯耐盐株系ND98中克隆了
IbNFU1 、
IbP5CR 、
IbMas 、
IbSIMT1 和
IbNHX2 基因, 并在甘薯中过表达这些基因, 可显著增强转基因甘薯植株的耐盐性(
Wang et al., 2013 ,
2016 ;
Liu et al., 2014a ,
2014b ,
2015 )。另有研究发现, 在盐及干旱胁迫下, 将甘薯
IbMIP1 过表达后可显著上调肌醇生物合成、磷脂酰肌醇(PI)信号途径、ABA信号途径及胁迫响应等相关基因的表达, 进而显著提高转基因甘薯的耐盐性和抗旱性(
Zhai et al., 2016 )。虽然甘薯转基因体系构建和相关技术趋于成熟, 但与甘薯抗旱、耐盐和抗氧化转基因研究相比, 甘薯抗冻基因及转基因植株培育进展相对缓慢, 目前尚未见甘薯抗冻基因遗传转化的研究报道。
本研究利用基因工程技术将黄粉虫抗冻蛋白基因
TmAFP 导入甘薯中, 创制抗冻甘薯新材料且抗冻效果显著。在甘薯遗传转化体系中, 绝大部分报道都是利用卡那霉素作为选择标记。也有研究以潮霉素作为选择标记。然而, 这2种抗生素对转基因甘薯的筛选效果均不理想, 越来越多的研究者以除草剂作为筛选标记, 且筛选效果较好(
阮龙等, 2010 )。因此, 本研究通过对现有质粒进行改造, 利用除草剂抗性基因(
bar 基因)作为选择标记, 以甘薯品种Huachano胚性悬浮细胞为受体, 用根癌农杆菌EH105为转化菌株, 成功建立了甘薯遗传转化体系, 且转基因甘薯结薯正常, 表明该转化体系是一个高效的甘薯遗传转化体系。同时, 本研究以
bar 基因作为筛选标记, 将目标基因导入甘薯植株的同时, 也将除草剂抗性一并导入, 赋予了转基因植株很好的除草剂抗性, 使甘薯生产过程中的除草作业更加方便和经济。
此外, 目前已发现的抗冻蛋白主要为动物来源, 其中以美洲拟蝶、绒杜父鱼(
Hemitripterus villosus )、美洲大绵(
Macrozoarces americanus )和南极鳕鱼(
Dissostichus eleginoides )来源为4大代表类型(
蔺忠龙等, 2009 )。虽然抗冻蛋白在包括鱼类、昆虫和植物在内的不同生物体中广泛存在, 但鱼类和昆虫来源的AFP研究起步较早, 目前已通过转基因技术将鱼类和昆虫来源的抗冻蛋白转入拟南芥、烟草和番茄(
Solanum lycopersicum )等模式植物中, 并在低温处理下获得了较好的抗冻效果(
Perl et al., 1993 ;
Pearce, 1999 )。但是植物来源的
AFP 基因研究起步较晚, 且已有研究的绝大多数植物材料中,
AFP 基因活性远低于鱼类和昆虫。虽然所研究的植物种类已有几十种, 但真正被分离纯化的植物源抗冻蛋白却非常少。因此, 如何从植物中分离出更多的、活性更高的AFP, 并通过遗传转化将其导入抗冻性弱或不抗冻的植物中, 进而获得抗冻性强的转基因植株, 将是今后植物抗冻基因工程研究的主要内容之一。
[1] 蔺忠龙 , 李维薇 , 白现广 , 吕广磊 , 程在全 (2009 ). 植物抗冻基因最新研究进展北方园艺 ( 1 ), 119 -123 . [本文引用: 1] [2] 刘忠渊 , 王芸 , 吕国栋 , 王贤磊 , 张富春 , 马纪 (2006 ). Tenebriomolitor抗冻蛋白基因家族cDNA片段的克隆、序列分析及原核表达遗传 28, 1532 -1540 . [本文引用: 1] [3] 马代夫 , 李洪民 , 李秀英 , 谢逸平 , 李强 (2005 ). 甘薯育种与甘薯产业发展成都: 中国作物学会 . pp. 3 -10 . [本文引用: 1] [4] 阮龙 , 高正良 , 陈义红 , 张玮 , 张云华 , 吴跃进 , 邵希文 (2010 ). 干旱耐逆基因(HS1 )转化甘薯获得转基因植株激光生物学报 19, 552 -556 . [本文引用: 1] [5] 王欣 , 过晓明 , 李强 , 唐忠厚 , 郭尚洙 , 马代夫 (2011 ). 转逆境诱导型启动子SWPA2驱动Cu/ZnSOD和APX基因甘薯(Ipomoea batatas (L.) Lam.)耐盐性分子植物育种 9, 754 -759 . [本文引用: 1] [6] 王艳 , 马纪 , 黄薇 , 邱立明 , 叶锋 , 张富春 (2009 ). 叶绿体型转昆虫抗冻蛋白基因烟草的耐寒性作物学报 35, 1253 -1260 . [本文引用: 1] [7] 臧宁 , 翟红 , 王玉萍 , 于波 , 何绍贞 , 刘庆昌 (2007 ). 表达bar 基因的抗除草剂转基因甘薯的获得分子植物育种 5, 475 -479 . [本文引用: 1] [8] 翟红 , 何绍贞 , 赵宁 , 刘庆昌 (2017 ). 甘薯生物技术育种研究进展江苏师范大学学报(自然科学版) 35, 25 -29 . [本文引用: 1] [9] 张振华 , 陈介南 , 卢孟柱 , 章怀云 , 刘伯斌 (2012 ). 胡萝卜与黄粉虫抗冻融合基因在拟南芥中的表达与抗冻性分析中国农学通报 28(31 ), 146 -152 . [本文引用: 1] [10] 瓜谷郁三 ( 谢国生 , 李合生译 ) (2004 ). 植物逆境生物化学及分子生物学: 着重热带薯类 . 北京: 中国农业出版社. pp. 202 -204 . [本文引用: 1] [11] Cutler AJ Saleem M Kendall E Gusta LV Georges F Fletcher GL 1989 ). Winter flounder antifreeze protein improves the cold hardiness of plant tissuesJ Plant Physiol 351 -354 . [本文引用: 2] [12] Fan WJ Zhang M Zhang HX Zhang P 2012 ). Improved tolerance to various abiotic stresses in transgenic sweet potato ( Ipomoea batatas ) expressing spinach betaine aldehyde dehydrogenasePLoS One e37344 . [本文引用: 1] [13] Liu DG He SZ Song XJ Zhai H Liu N Zhang DD Ren ZT Liu QC 2015 ). IbSIMT1 , a novel salt-induced methyltransferase gene from Ipomoea batatas , is involved in salt tolerancePlant Cell Tissue Organ Cult 701 -715 . [本文引用: 1] [14] Liu DG He SZ Zhai H Wang LJ Zhao Y Wang B Li RJ Liu QC 2014a ). Overexpression of IbP5CR enhances salt tolerance in transgenic sweetpotatoPlant Cell Tissue Organ Cult 1 -16 . [本文引用: 1] [15] Liu DG Wang LJ Zhai H Song XJ He SZ Liu QC 2014b ). A novel α/β-hydrolase gene IbMas enhances salt tolerance in transgenic sweetpotatoPLoS One e115128 . [本文引用: 1] [16] Liu QC 2011 ). Sweet potato omics and biotechnology in ChinaPlant Omics 295 -301 . [本文引用: 1] [17] Mwanga ROM Andrade MI Carey EE Low JW Yencho GC Grüneberg WJ 2017 ). Sweetpotato (Ipomoea batatas L.)Genetic Improvement of Tropical Crops. Cham: Springer 181 -218 . [本文引用: 1] [18] Nada H Furukawa Y 2011 ). Growth inhibition at the ice prismatic plane induced by a spruce budworm antifreeze protein: a molecular dynamics simulation studyPhys Chem Chem Phys 19936 -19942 . [本文引用: 1] [19] Pearce RS 1999 ). Molecular analysis of acclimation to coldPlant Growth Regul 47 -76 . [本文引用: 1] [20] Perl A Perl-Treves R Galili S Aviv D Shalgi E Malkin S Galun E 1993 ). Enhanced oxidative-stress defense in transgenic potato expressing tomato Cu, Zn superoxide dismutasesTheor Appl Genet 568 -576 . [本文引用: 1] [21] Ramya L Ramakrishnan V 2016 ). Interaction of tenebrio molitor antifreeze protein with ice crystal: insights from molecular dynamics simulationsMol Inform 268 -277 . [本文引用: 1] [22] Wang B Zhai H He SZ Zhang H Ren ZT Zhang DD Liu QC 2016 ). A vacuolar Na+ /H+ antiporter gene, IbNHX2 , enhances salt and drought tolerance in transgenic sweetpotatoSci Hortic 153 -166 . [本文引用: 1] [23] Wang C Pakhomova S Newcomer ME Christner BC Luo BH 2017 ). Structural basis of antifreeze activity of a bacterial multi-domain antifreeze proteinPLoS One e0187169 . [本文引用: 1] [24] Wang LJ He SZ Zhai H Liu DG Wang YN Liu QC 2013 ). Molecular cloning and functional characterization of a salt tolerance-associated gene IbNFU1 from sweetpotatoJ Integr Agric 27 -35 . [本文引用: 1] [25] Wang WX Vinocur B Altman A 2003 ). Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerancePlanta 1 -14 . [本文引用: 1] [26] Yang Y Guan S Zhai H He S Liu Q 2009 ). Development and evaluation of a storage root-bearing sweetpotato somatic hybrid between Ipomoea batatas(L.) Lam. and I. triloba LPlant Cell Tissue Organ Cult 83 -89 . [本文引用: 1] [27] Yue CW Zhang YZ 2009 ). Cloning and expression of Tenebrio molitor antifreeze protein in Escherichia coli Mol Biol Rep 529 -536 . [本文引用: 1] [28] Zhai H Wang FB Si ZZ Huo JX Xing L An YY He SZ Liu QC 2016 ). A myo -inositol-1-phosphate synthase gene, IbMIPS1 , enhances salt and drought tolerance and stem nematode resistance in transgenic sweet potatoPlant Biotechnol J 592 -602 . [本文引用: 1] [29] Zhang JH Davies WJ 1987 ). Increased synthesis of ABA in partially dehydrated root tips and ABA transport from roots to leavesJ Exp Bot 2015 -2023 . [本文引用: 1] 植物抗冻基因最新研究进展
1
2009
... 此外, 目前已发现的抗冻蛋白主要为动物来源, 其中以美洲拟蝶、绒杜父鱼(
Hemitripterus villosus )、美洲大绵(
Macrozoarces americanus )和南极鳕鱼(
Dissostichus eleginoides )来源为4大代表类型(
蔺忠龙等, 2009 ).虽然抗冻蛋白在包括鱼类、昆虫和植物在内的不同生物体中广泛存在, 但鱼类和昆虫来源的AFP研究起步较早, 目前已通过转基因技术将鱼类和昆虫来源的抗冻蛋白转入拟南芥、烟草和番茄(
Solanum lycopersicum )等模式植物中, 并在低温处理下获得了较好的抗冻效果(
Perl et al., 1993 ;
Pearce, 1999 ).但是植物来源的
AFP 基因研究起步较晚, 且已有研究的绝大多数植物材料中,
AFP 基因活性远低于鱼类和昆虫.虽然所研究的植物种类已有几十种, 但真正被分离纯化的植物源抗冻蛋白却非常少.因此, 如何从植物中分离出更多的、活性更高的AFP, 并通过遗传转化将其导入抗冻性弱或不抗冻的植物中, 进而获得抗冻性强的转基因植株, 将是今后植物抗冻基因工程研究的主要内容之一. ...
Tenebriomolitor抗冻蛋白基因家族cDNA片段的克隆、序列分析及原核表达
1
2006
... 黄粉虫(
Tenebrio molitor )抗冻蛋白(TmAFP)二聚体的抗冻活性是已知抗冻蛋白中最高的, 可达鱼类抗冻蛋白活性的10-100倍.黄粉虫抗冻蛋白为右手β螺旋结构, 每一螺旋由12个保守的氨基酸组成(CT- XSXXCXXAXT, X为任一氨基酸残基), 依据12个氨基酸保守结构域重复次数可将
TmAFP 基因克隆分为3类, 分别编码84、96和120个氨基酸.TmAFP富含苏氨酸和半胱氨酸, 沿着蛋白折叠的一侧有两行苏氨酸残基与冰晶表面的棱柱和基面匹配结合(
Yue and Zhang, 2009 ).当接近冰点温度时, 抗冻蛋白表面冰结合位点上的苏氨酸侧链形成更优化的旋转异构体, 这种特有的冰结合构造使抗冻蛋白-冰分界面与冰晶相匹配, 从而阻止冰晶增长(
Ramya and Ramakrishnan, 2016 ).研究者将克隆到的
TmAFP 基因进行原核表达, 其表达蛋白经纯化后表现出冰结合活性和体外抗冻活性, 进一步将TmAFP蛋白代替传统抗冻保护剂DMSO制备大肠杆菌感受态细胞, 获得了转化高效的感受态细胞(
刘忠渊等, 2006 ).在提高植物抗冻能力方面, 有研究者将胡萝卜(
Daucus carota )抗冻蛋白基因(
DcAFP )和黄粉虫抗冻蛋白基因(
TmAFP )串联构建融合基因表达载体, 通过农杆菌介导转化拟南芥, 证实
TmAFP 的转基因拟南芥抗冻能力增强(
张振华等, 2012 ).然而, 至今未见有
TmAFP 基因在其它模式植物和经济作物中的转基因研究报道.因此, 通过基因工程技术将黄粉虫抗冻蛋白基因转入甘薯体内使其产生抗冻蛋白, 有望提高甘薯抗冻耐寒能力. ...
甘薯育种与甘薯产业发展
1
2005
... 甘薯(
Ipomoea batatas )是旋花科一年生植物, 也是世界上重要的块根类粮食作物, 广泛应用于食品、饲料、工业及生物能源领域(
Liu, 2011 ).甘薯生长和产量提升对于解决世界粮食资源安全等问题尤为重要(
Mwanga et al., 2017 ).然而, 受遗传因素制约, 甘薯对低温敏感, 极不耐霜冻, 在10°C以下, 甘薯细胞的原生质活动停滞, 生理机能受到影响, 导致冷害发生; 当温度低至-2°C时, 甘薯细胞间隙结冰, 其组织被破坏, 从而发生冻害.冷害或冻害的发生造成甘薯新陈代谢异常, 贮藏性能和抗病性降低, 贮藏期甘薯容易受到病菌侵染, 导致烂窖的发生(
瓜谷郁三, 2004 ).据统计, 每年约有30%的甘薯由于发生烂窖而失去利用价值, 损失约几十亿元人民币(
马代夫等, 2005 ).此外, 甘薯的生长适宜温度比水稻(
Oryza sativa )、玉米(
Zea mays )和棉花(
Gossypium spp.)等作物高5-10°C, 在春季幼苗期常由于倒春寒突然降温导致甘薯冻害的发生, 对甘薯生产造成重大损失.全球每年因低温冻害造成的农作物损失高达数千亿元, 因此, 培育抗冻甘薯新种质具有重要意义(
Zang et al., 2007 ). ...
干旱耐逆基因(
HS1 )转化甘薯获得转基因植株
1
2010
... 本研究利用基因工程技术将黄粉虫抗冻蛋白基因
TmAFP 导入甘薯中, 创制抗冻甘薯新材料且抗冻效果显著.在甘薯遗传转化体系中, 绝大部分报道都是利用卡那霉素作为选择标记.也有研究以潮霉素作为选择标记.然而, 这2种抗生素对转基因甘薯的筛选效果均不理想, 越来越多的研究者以除草剂作为筛选标记, 且筛选效果较好(
阮龙等, 2010 ).因此, 本研究通过对现有质粒进行改造, 利用除草剂抗性基因(
bar 基因)作为选择标记, 以甘薯品种Huachano胚性悬浮细胞为受体, 用根癌农杆菌EH105为转化菌株, 成功建立了甘薯遗传转化体系, 且转基因甘薯结薯正常, 表明该转化体系是一个高效的甘薯遗传转化体系.同时, 本研究以
bar 基因作为筛选标记, 将目标基因导入甘薯植株的同时, 也将除草剂抗性一并导入, 赋予了转基因植株很好的除草剂抗性, 使甘薯生产过程中的除草作业更加方便和经济. ...
转逆境诱导型启动子SWPA2驱动Cu/ZnSOD和APX基因甘薯(
Ipomoea batatas (L.) Lam.)耐盐性
1
2011
... 近年来, 随着分子生物学的快速发展, 利用基因工程对植物性状进行遗传改良, 以获得具有特定优异性状新育种材料的技术体系已日趋成熟(
Wang et al., 2003 ).由于植物长期暴露于盐碱、干旱和寒冷等非生物逆境, 其本身会触发许多共同的防御机制, 如ABA含量增加、可溶性糖和保护性蛋白质的积累以及抗氧化水平的提升.因此将非生物胁迫相关的植物源基因导入可提高甘薯对盐碱、干旱、低温和氧化逆境等的抗性(
翟红等, 2017 ).例如, 将
Cu /
Zn SOD 和
APX 基因导入甘薯, 可增强转基因甘薯的耐盐性(
王欣等, 2011 ); 将菠菜 (
Spinacia oleracea )
SoBADH 基因导入甘薯可增强转基因甘薯对盐、低温及氧化胁迫的抗性(
Fan et al., 2012 ).同时, 可通过克隆甘薯自身的抗性基因, 然后增强其表达的方式创制转基因甘薯新材料.例如, 分别从甘薯耐盐株系ND98中克隆了
IbNFU1 、
IbP5CR 、
IbMas 、
IbSIMT1 和
IbNHX2 基因, 并在甘薯中过表达这些基因, 可显著增强转基因甘薯植株的耐盐性(
Wang et al., 2013 ,
2016 ;
Liu et al., 2014a ,
2014b ,
2015 ).另有研究发现, 在盐及干旱胁迫下, 将甘薯
IbMIP1 过表达后可显著上调肌醇生物合成、磷脂酰肌醇(PI)信号途径、ABA信号途径及胁迫响应等相关基因的表达, 进而显著提高转基因甘薯的耐盐性和抗旱性(
Zhai et al., 2016 ).虽然甘薯转基因体系构建和相关技术趋于成熟, 但与甘薯抗旱、耐盐和抗氧化转基因研究相比, 甘薯抗冻基因及转基因植株培育进展相对缓慢, 目前尚未见甘薯抗冻基因遗传转化的研究报道. ...
叶绿体型转昆虫抗冻蛋白基因烟草的耐寒性
1
2009
... 由于甘薯具有杂交不亲和性及结实率低等特点, 严重限制了甘薯育种中的资源利用, 因此基因工程是改良甘薯品种的一种有效手段(
Yang et al., 2009 ).抗冻蛋白(antifreeze protein, AFP)又称热滞蛋白(thermal hysteresis protein, THP), 具有多个能够直接作用于冰晶的亲水性冰结合位点, 可阻止低温下冰晶在细胞间隙中的形成和再结晶, 以维持生物体正常的新陈代谢(
Wang et al., 2017 ).自De Vrie等(1969)从一种大西洋冷水鱼血液中发现抗冻蛋白以来, 其在细菌、昆虫、鱼类和植物中也相继被发现, 尤其是昆虫抗冻蛋白具有抗冻活性高和结构规律等特点, 其滞热活性与其它生物的抗冻蛋白相比高10-100倍, 这使得昆虫抗冻蛋白在植物抗冻遗传转化中具有巨大的优势和潜力(
Nada and Furukawa, 2011 ).Davies等(1987)首次将美洲拟鲽(
Pseudopleuronectes americanus )抗冻蛋白基因整合到Ti质粒中, 通过叶圆片法转化郁金香(
Tulipa gesneriana )和油菜(
Brassica campestris )等, 使转基因植株获得了一定的抗冻能力(
Zhang and Davies, 1987 ).
Cutler等(1989) 用真空透析法将冬比目鱼(
Pleuronectiformes heterosomata )抗冻蛋白基因导入马铃薯(
Solanum tuberosum )、拟南芥(
Arabidopsis thaliana )和欧洲油菜(
B. napus ), 使此类植物的自然结冰温度降低1.8°C, 证实转动物源抗冻蛋白基因可提高植物的抗寒性(
Cutler et al., 1989 ).
Wang等(2009) 用准噶尔小胸鳖甲(
Microdera punctipennis )抗冻蛋白基因
MPAFP149 转化烟草(
Nicotiana tabacum ), 并对T
0 代转基因烟草在-1°C处理48小时后, 发现转基因烟草相对电导率和表型明显优于野生型.室温恢复实验证实转基因烟草可存活并恢复生长, 而野生型烟草则遭受了不可逆的低温冻害.该研究表明, 转化后携带昆虫抗冻蛋白基因的烟草相比野生型具有明显的抗寒能力. ...
表达
bar 基因的抗除草剂转基因甘薯的获得
1
2007
... 甘薯(
Ipomoea batatas )是旋花科一年生植物, 也是世界上重要的块根类粮食作物, 广泛应用于食品、饲料、工业及生物能源领域(
Liu, 2011 ).甘薯生长和产量提升对于解决世界粮食资源安全等问题尤为重要(
Mwanga et al., 2017 ).然而, 受遗传因素制约, 甘薯对低温敏感, 极不耐霜冻, 在10°C以下, 甘薯细胞的原生质活动停滞, 生理机能受到影响, 导致冷害发生; 当温度低至-2°C时, 甘薯细胞间隙结冰, 其组织被破坏, 从而发生冻害.冷害或冻害的发生造成甘薯新陈代谢异常, 贮藏性能和抗病性降低, 贮藏期甘薯容易受到病菌侵染, 导致烂窖的发生(
瓜谷郁三, 2004 ).据统计, 每年约有30%的甘薯由于发生烂窖而失去利用价值, 损失约几十亿元人民币(
马代夫等, 2005 ).此外, 甘薯的生长适宜温度比水稻(
Oryza sativa )、玉米(
Zea mays )和棉花(
Gossypium spp.)等作物高5-10°C, 在春季幼苗期常由于倒春寒突然降温导致甘薯冻害的发生, 对甘薯生产造成重大损失.全球每年因低温冻害造成的农作物损失高达数千亿元, 因此, 培育抗冻甘薯新种质具有重要意义(
Zang et al., 2007 ). ...
甘薯生物技术育种研究进展
1
2017
... 近年来, 随着分子生物学的快速发展, 利用基因工程对植物性状进行遗传改良, 以获得具有特定优异性状新育种材料的技术体系已日趋成熟(
Wang et al., 2003 ).由于植物长期暴露于盐碱、干旱和寒冷等非生物逆境, 其本身会触发许多共同的防御机制, 如ABA含量增加、可溶性糖和保护性蛋白质的积累以及抗氧化水平的提升.因此将非生物胁迫相关的植物源基因导入可提高甘薯对盐碱、干旱、低温和氧化逆境等的抗性(
翟红等, 2017 ).例如, 将
Cu /
Zn SOD 和
APX 基因导入甘薯, 可增强转基因甘薯的耐盐性(
王欣等, 2011 ); 将菠菜 (
Spinacia oleracea )
SoBADH 基因导入甘薯可增强转基因甘薯对盐、低温及氧化胁迫的抗性(
Fan et al., 2012 ).同时, 可通过克隆甘薯自身的抗性基因, 然后增强其表达的方式创制转基因甘薯新材料.例如, 分别从甘薯耐盐株系ND98中克隆了
IbNFU1 、
IbP5CR 、
IbMas 、
IbSIMT1 和
IbNHX2 基因, 并在甘薯中过表达这些基因, 可显著增强转基因甘薯植株的耐盐性(
Wang et al., 2013 ,
2016 ;
Liu et al., 2014a ,
2014b ,
2015 ).另有研究发现, 在盐及干旱胁迫下, 将甘薯
IbMIP1 过表达后可显著上调肌醇生物合成、磷脂酰肌醇(PI)信号途径、ABA信号途径及胁迫响应等相关基因的表达, 进而显著提高转基因甘薯的耐盐性和抗旱性(
Zhai et al., 2016 ).虽然甘薯转基因体系构建和相关技术趋于成熟, 但与甘薯抗旱、耐盐和抗氧化转基因研究相比, 甘薯抗冻基因及转基因植株培育进展相对缓慢, 目前尚未见甘薯抗冻基因遗传转化的研究报道. ...
胡萝卜与黄粉虫抗冻融合基因在拟南芥中的表达与抗冻性分析
1
2012
... 黄粉虫(
Tenebrio molitor )抗冻蛋白(TmAFP)二聚体的抗冻活性是已知抗冻蛋白中最高的, 可达鱼类抗冻蛋白活性的10-100倍.黄粉虫抗冻蛋白为右手β螺旋结构, 每一螺旋由12个保守的氨基酸组成(CT- XSXXCXXAXT, X为任一氨基酸残基), 依据12个氨基酸保守结构域重复次数可将
TmAFP 基因克隆分为3类, 分别编码84、96和120个氨基酸.TmAFP富含苏氨酸和半胱氨酸, 沿着蛋白折叠的一侧有两行苏氨酸残基与冰晶表面的棱柱和基面匹配结合(
Yue and Zhang, 2009 ).当接近冰点温度时, 抗冻蛋白表面冰结合位点上的苏氨酸侧链形成更优化的旋转异构体, 这种特有的冰结合构造使抗冻蛋白-冰分界面与冰晶相匹配, 从而阻止冰晶增长(
Ramya and Ramakrishnan, 2016 ).研究者将克隆到的
TmAFP 基因进行原核表达, 其表达蛋白经纯化后表现出冰结合活性和体外抗冻活性, 进一步将TmAFP蛋白代替传统抗冻保护剂DMSO制备大肠杆菌感受态细胞, 获得了转化高效的感受态细胞(
刘忠渊等, 2006 ).在提高植物抗冻能力方面, 有研究者将胡萝卜(
Daucus carota )抗冻蛋白基因(
DcAFP )和黄粉虫抗冻蛋白基因(
TmAFP )串联构建融合基因表达载体, 通过农杆菌介导转化拟南芥, 证实
TmAFP 的转基因拟南芥抗冻能力增强(
张振华等, 2012 ).然而, 至今未见有
TmAFP 基因在其它模式植物和经济作物中的转基因研究报道.因此, 通过基因工程技术将黄粉虫抗冻蛋白基因转入甘薯体内使其产生抗冻蛋白, 有望提高甘薯抗冻耐寒能力. ...
1
2004
... 甘薯(
Ipomoea batatas )是旋花科一年生植物, 也是世界上重要的块根类粮食作物, 广泛应用于食品、饲料、工业及生物能源领域(
Liu, 2011 ).甘薯生长和产量提升对于解决世界粮食资源安全等问题尤为重要(
Mwanga et al., 2017 ).然而, 受遗传因素制约, 甘薯对低温敏感, 极不耐霜冻, 在10°C以下, 甘薯细胞的原生质活动停滞, 生理机能受到影响, 导致冷害发生; 当温度低至-2°C时, 甘薯细胞间隙结冰, 其组织被破坏, 从而发生冻害.冷害或冻害的发生造成甘薯新陈代谢异常, 贮藏性能和抗病性降低, 贮藏期甘薯容易受到病菌侵染, 导致烂窖的发生(
瓜谷郁三, 2004 ).据统计, 每年约有30%的甘薯由于发生烂窖而失去利用价值, 损失约几十亿元人民币(
马代夫等, 2005 ).此外, 甘薯的生长适宜温度比水稻(
Oryza sativa )、玉米(
Zea mays )和棉花(
Gossypium spp.)等作物高5-10°C, 在春季幼苗期常由于倒春寒突然降温导致甘薯冻害的发生, 对甘薯生产造成重大损失.全球每年因低温冻害造成的农作物损失高达数千亿元, 因此, 培育抗冻甘薯新种质具有重要意义(
Zang et al., 2007 ). ...
Winter flounder antifreeze protein improves the cold hardiness of plant tissues
2
1989
... 由于甘薯具有杂交不亲和性及结实率低等特点, 严重限制了甘薯育种中的资源利用, 因此基因工程是改良甘薯品种的一种有效手段(
Yang et al., 2009 ).抗冻蛋白(antifreeze protein, AFP)又称热滞蛋白(thermal hysteresis protein, THP), 具有多个能够直接作用于冰晶的亲水性冰结合位点, 可阻止低温下冰晶在细胞间隙中的形成和再结晶, 以维持生物体正常的新陈代谢(
Wang et al., 2017 ).自De Vrie等(1969)从一种大西洋冷水鱼血液中发现抗冻蛋白以来, 其在细菌、昆虫、鱼类和植物中也相继被发现, 尤其是昆虫抗冻蛋白具有抗冻活性高和结构规律等特点, 其滞热活性与其它生物的抗冻蛋白相比高10-100倍, 这使得昆虫抗冻蛋白在植物抗冻遗传转化中具有巨大的优势和潜力(
Nada and Furukawa, 2011 ).Davies等(1987)首次将美洲拟鲽(
Pseudopleuronectes americanus )抗冻蛋白基因整合到Ti质粒中, 通过叶圆片法转化郁金香(
Tulipa gesneriana )和油菜(
Brassica campestris )等, 使转基因植株获得了一定的抗冻能力(
Zhang and Davies, 1987 ).
Cutler等(1989) 用真空透析法将冬比目鱼(
Pleuronectiformes heterosomata )抗冻蛋白基因导入马铃薯(
Solanum tuberosum )、拟南芥(
Arabidopsis thaliana )和欧洲油菜(
B. napus ), 使此类植物的自然结冰温度降低1.8°C, 证实转动物源抗冻蛋白基因可提高植物的抗寒性(
Cutler et al., 1989 ).
Wang等(2009) 用准噶尔小胸鳖甲(
Microdera punctipennis )抗冻蛋白基因
MPAFP149 转化烟草(
Nicotiana tabacum ), 并对T
0 代转基因烟草在-1°C处理48小时后, 发现转基因烟草相对电导率和表型明显优于野生型.室温恢复实验证实转基因烟草可存活并恢复生长, 而野生型烟草则遭受了不可逆的低温冻害.该研究表明, 转化后携带昆虫抗冻蛋白基因的烟草相比野生型具有明显的抗寒能力. ...
... ), 使此类植物的自然结冰温度降低1.8°C, 证实转动物源抗冻蛋白基因可提高植物的抗寒性(
Cutler et al., 1989 ).
Wang等(2009) 用准噶尔小胸鳖甲(
Microdera punctipennis )抗冻蛋白基因
MPAFP149 转化烟草(
Nicotiana tabacum ), 并对T
0 代转基因烟草在-1°C处理48小时后, 发现转基因烟草相对电导率和表型明显优于野生型.室温恢复实验证实转基因烟草可存活并恢复生长, 而野生型烟草则遭受了不可逆的低温冻害.该研究表明, 转化后携带昆虫抗冻蛋白基因的烟草相比野生型具有明显的抗寒能力. ...
Improved tolerance to various abiotic stresses in transgenic sweet potato (
Ipomoea batatas ) expressing spinach betaine aldehyde dehydrogenase
1
2012
... 近年来, 随着分子生物学的快速发展, 利用基因工程对植物性状进行遗传改良, 以获得具有特定优异性状新育种材料的技术体系已日趋成熟(
Wang et al., 2003 ).由于植物长期暴露于盐碱、干旱和寒冷等非生物逆境, 其本身会触发许多共同的防御机制, 如ABA含量增加、可溶性糖和保护性蛋白质的积累以及抗氧化水平的提升.因此将非生物胁迫相关的植物源基因导入可提高甘薯对盐碱、干旱、低温和氧化逆境等的抗性(
翟红等, 2017 ).例如, 将
Cu /
Zn SOD 和
APX 基因导入甘薯, 可增强转基因甘薯的耐盐性(
王欣等, 2011 ); 将菠菜 (
Spinacia oleracea )
SoBADH 基因导入甘薯可增强转基因甘薯对盐、低温及氧化胁迫的抗性(
Fan et al., 2012 ).同时, 可通过克隆甘薯自身的抗性基因, 然后增强其表达的方式创制转基因甘薯新材料.例如, 分别从甘薯耐盐株系ND98中克隆了
IbNFU1 、
IbP5CR 、
IbMas 、
IbSIMT1 和
IbNHX2 基因, 并在甘薯中过表达这些基因, 可显著增强转基因甘薯植株的耐盐性(
Wang et al., 2013 ,
2016 ;
Liu et al., 2014a ,
2014b ,
2015 ).另有研究发现, 在盐及干旱胁迫下, 将甘薯
IbMIP1 过表达后可显著上调肌醇生物合成、磷脂酰肌醇(PI)信号途径、ABA信号途径及胁迫响应等相关基因的表达, 进而显著提高转基因甘薯的耐盐性和抗旱性(
Zhai et al., 2016 ).虽然甘薯转基因体系构建和相关技术趋于成熟, 但与甘薯抗旱、耐盐和抗氧化转基因研究相比, 甘薯抗冻基因及转基因植株培育进展相对缓慢, 目前尚未见甘薯抗冻基因遗传转化的研究报道. ...
IbSIMT1 , a novel salt-induced methyltransferase gene from
Ipomoea batatas , is involved in salt tolerance
1
2015
... 近年来, 随着分子生物学的快速发展, 利用基因工程对植物性状进行遗传改良, 以获得具有特定优异性状新育种材料的技术体系已日趋成熟(
Wang et al., 2003 ).由于植物长期暴露于盐碱、干旱和寒冷等非生物逆境, 其本身会触发许多共同的防御机制, 如ABA含量增加、可溶性糖和保护性蛋白质的积累以及抗氧化水平的提升.因此将非生物胁迫相关的植物源基因导入可提高甘薯对盐碱、干旱、低温和氧化逆境等的抗性(
翟红等, 2017 ).例如, 将
Cu /
Zn SOD 和
APX 基因导入甘薯, 可增强转基因甘薯的耐盐性(
王欣等, 2011 ); 将菠菜 (
Spinacia oleracea )
SoBADH 基因导入甘薯可增强转基因甘薯对盐、低温及氧化胁迫的抗性(
Fan et al., 2012 ).同时, 可通过克隆甘薯自身的抗性基因, 然后增强其表达的方式创制转基因甘薯新材料.例如, 分别从甘薯耐盐株系ND98中克隆了
IbNFU1 、
IbP5CR 、
IbMas 、
IbSIMT1 和
IbNHX2 基因, 并在甘薯中过表达这些基因, 可显著增强转基因甘薯植株的耐盐性(
Wang et al., 2013 ,
2016 ;
Liu et al., 2014a ,
2014b ,
2015 ).另有研究发现, 在盐及干旱胁迫下, 将甘薯
IbMIP1 过表达后可显著上调肌醇生物合成、磷脂酰肌醇(PI)信号途径、ABA信号途径及胁迫响应等相关基因的表达, 进而显著提高转基因甘薯的耐盐性和抗旱性(
Zhai et al., 2016 ).虽然甘薯转基因体系构建和相关技术趋于成熟, 但与甘薯抗旱、耐盐和抗氧化转基因研究相比, 甘薯抗冻基因及转基因植株培育进展相对缓慢, 目前尚未见甘薯抗冻基因遗传转化的研究报道. ...
Overexpression of
IbP5CR enhances salt tolerance in transgenic sweetpotato
1
2014a
... 近年来, 随着分子生物学的快速发展, 利用基因工程对植物性状进行遗传改良, 以获得具有特定优异性状新育种材料的技术体系已日趋成熟(
Wang et al., 2003 ).由于植物长期暴露于盐碱、干旱和寒冷等非生物逆境, 其本身会触发许多共同的防御机制, 如ABA含量增加、可溶性糖和保护性蛋白质的积累以及抗氧化水平的提升.因此将非生物胁迫相关的植物源基因导入可提高甘薯对盐碱、干旱、低温和氧化逆境等的抗性(
翟红等, 2017 ).例如, 将
Cu /
Zn SOD 和
APX 基因导入甘薯, 可增强转基因甘薯的耐盐性(
王欣等, 2011 ); 将菠菜 (
Spinacia oleracea )
SoBADH 基因导入甘薯可增强转基因甘薯对盐、低温及氧化胁迫的抗性(
Fan et al., 2012 ).同时, 可通过克隆甘薯自身的抗性基因, 然后增强其表达的方式创制转基因甘薯新材料.例如, 分别从甘薯耐盐株系ND98中克隆了
IbNFU1 、
IbP5CR 、
IbMas 、
IbSIMT1 和
IbNHX2 基因, 并在甘薯中过表达这些基因, 可显著增强转基因甘薯植株的耐盐性(
Wang et al., 2013 ,
2016 ;
Liu et al., 2014a ,
2014b ,
2015 ).另有研究发现, 在盐及干旱胁迫下, 将甘薯
IbMIP1 过表达后可显著上调肌醇生物合成、磷脂酰肌醇(PI)信号途径、ABA信号途径及胁迫响应等相关基因的表达, 进而显著提高转基因甘薯的耐盐性和抗旱性(
Zhai et al., 2016 ).虽然甘薯转基因体系构建和相关技术趋于成熟, 但与甘薯抗旱、耐盐和抗氧化转基因研究相比, 甘薯抗冻基因及转基因植株培育进展相对缓慢, 目前尚未见甘薯抗冻基因遗传转化的研究报道. ...
A novel α/β-hydrolase gene
IbMas enhances salt tolerance in transgenic sweetpotato
1
2014b
... 近年来, 随着分子生物学的快速发展, 利用基因工程对植物性状进行遗传改良, 以获得具有特定优异性状新育种材料的技术体系已日趋成熟(
Wang et al., 2003 ).由于植物长期暴露于盐碱、干旱和寒冷等非生物逆境, 其本身会触发许多共同的防御机制, 如ABA含量增加、可溶性糖和保护性蛋白质的积累以及抗氧化水平的提升.因此将非生物胁迫相关的植物源基因导入可提高甘薯对盐碱、干旱、低温和氧化逆境等的抗性(
翟红等, 2017 ).例如, 将
Cu /
Zn SOD 和
APX 基因导入甘薯, 可增强转基因甘薯的耐盐性(
王欣等, 2011 ); 将菠菜 (
Spinacia oleracea )
SoBADH 基因导入甘薯可增强转基因甘薯对盐、低温及氧化胁迫的抗性(
Fan et al., 2012 ).同时, 可通过克隆甘薯自身的抗性基因, 然后增强其表达的方式创制转基因甘薯新材料.例如, 分别从甘薯耐盐株系ND98中克隆了
IbNFU1 、
IbP5CR 、
IbMas 、
IbSIMT1 和
IbNHX2 基因, 并在甘薯中过表达这些基因, 可显著增强转基因甘薯植株的耐盐性(
Wang et al., 2013 ,
2016 ;
Liu et al., 2014a ,
2014b ,
2015 ).另有研究发现, 在盐及干旱胁迫下, 将甘薯
IbMIP1 过表达后可显著上调肌醇生物合成、磷脂酰肌醇(PI)信号途径、ABA信号途径及胁迫响应等相关基因的表达, 进而显著提高转基因甘薯的耐盐性和抗旱性(
Zhai et al., 2016 ).虽然甘薯转基因体系构建和相关技术趋于成熟, 但与甘薯抗旱、耐盐和抗氧化转基因研究相比, 甘薯抗冻基因及转基因植株培育进展相对缓慢, 目前尚未见甘薯抗冻基因遗传转化的研究报道. ...
Sweet potato omics and biotechnology in China
1
2011
... 甘薯(
Ipomoea batatas )是旋花科一年生植物, 也是世界上重要的块根类粮食作物, 广泛应用于食品、饲料、工业及生物能源领域(
Liu, 2011 ).甘薯生长和产量提升对于解决世界粮食资源安全等问题尤为重要(
Mwanga et al., 2017 ).然而, 受遗传因素制约, 甘薯对低温敏感, 极不耐霜冻, 在10°C以下, 甘薯细胞的原生质活动停滞, 生理机能受到影响, 导致冷害发生; 当温度低至-2°C时, 甘薯细胞间隙结冰, 其组织被破坏, 从而发生冻害.冷害或冻害的发生造成甘薯新陈代谢异常, 贮藏性能和抗病性降低, 贮藏期甘薯容易受到病菌侵染, 导致烂窖的发生(
瓜谷郁三, 2004 ).据统计, 每年约有30%的甘薯由于发生烂窖而失去利用价值, 损失约几十亿元人民币(
马代夫等, 2005 ).此外, 甘薯的生长适宜温度比水稻(
Oryza sativa )、玉米(
Zea mays )和棉花(
Gossypium spp.)等作物高5-10°C, 在春季幼苗期常由于倒春寒突然降温导致甘薯冻害的发生, 对甘薯生产造成重大损失.全球每年因低温冻害造成的农作物损失高达数千亿元, 因此, 培育抗冻甘薯新种质具有重要意义(
Zang et al., 2007 ). ...
Sweetpotato (Ipomoea batatas L.)
1
2017
... 甘薯(
Ipomoea batatas )是旋花科一年生植物, 也是世界上重要的块根类粮食作物, 广泛应用于食品、饲料、工业及生物能源领域(
Liu, 2011 ).甘薯生长和产量提升对于解决世界粮食资源安全等问题尤为重要(
Mwanga et al., 2017 ).然而, 受遗传因素制约, 甘薯对低温敏感, 极不耐霜冻, 在10°C以下, 甘薯细胞的原生质活动停滞, 生理机能受到影响, 导致冷害发生; 当温度低至-2°C时, 甘薯细胞间隙结冰, 其组织被破坏, 从而发生冻害.冷害或冻害的发生造成甘薯新陈代谢异常, 贮藏性能和抗病性降低, 贮藏期甘薯容易受到病菌侵染, 导致烂窖的发生(
瓜谷郁三, 2004 ).据统计, 每年约有30%的甘薯由于发生烂窖而失去利用价值, 损失约几十亿元人民币(
马代夫等, 2005 ).此外, 甘薯的生长适宜温度比水稻(
Oryza sativa )、玉米(
Zea mays )和棉花(
Gossypium spp.)等作物高5-10°C, 在春季幼苗期常由于倒春寒突然降温导致甘薯冻害的发生, 对甘薯生产造成重大损失.全球每年因低温冻害造成的农作物损失高达数千亿元, 因此, 培育抗冻甘薯新种质具有重要意义(
Zang et al., 2007 ). ...
Growth inhibition at the ice prismatic plane induced by a spruce budworm antifreeze protein: a molecular dynamics simulation study
1
2011
... 由于甘薯具有杂交不亲和性及结实率低等特点, 严重限制了甘薯育种中的资源利用, 因此基因工程是改良甘薯品种的一种有效手段(
Yang et al., 2009 ).抗冻蛋白(antifreeze protein, AFP)又称热滞蛋白(thermal hysteresis protein, THP), 具有多个能够直接作用于冰晶的亲水性冰结合位点, 可阻止低温下冰晶在细胞间隙中的形成和再结晶, 以维持生物体正常的新陈代谢(
Wang et al., 2017 ).自De Vrie等(1969)从一种大西洋冷水鱼血液中发现抗冻蛋白以来, 其在细菌、昆虫、鱼类和植物中也相继被发现, 尤其是昆虫抗冻蛋白具有抗冻活性高和结构规律等特点, 其滞热活性与其它生物的抗冻蛋白相比高10-100倍, 这使得昆虫抗冻蛋白在植物抗冻遗传转化中具有巨大的优势和潜力(
Nada and Furukawa, 2011 ).Davies等(1987)首次将美洲拟鲽(
Pseudopleuronectes americanus )抗冻蛋白基因整合到Ti质粒中, 通过叶圆片法转化郁金香(
Tulipa gesneriana )和油菜(
Brassica campestris )等, 使转基因植株获得了一定的抗冻能力(
Zhang and Davies, 1987 ).
Cutler等(1989) 用真空透析法将冬比目鱼(
Pleuronectiformes heterosomata )抗冻蛋白基因导入马铃薯(
Solanum tuberosum )、拟南芥(
Arabidopsis thaliana )和欧洲油菜(
B. napus ), 使此类植物的自然结冰温度降低1.8°C, 证实转动物源抗冻蛋白基因可提高植物的抗寒性(
Cutler et al., 1989 ).
Wang等(2009) 用准噶尔小胸鳖甲(
Microdera punctipennis )抗冻蛋白基因
MPAFP149 转化烟草(
Nicotiana tabacum ), 并对T
0 代转基因烟草在-1°C处理48小时后, 发现转基因烟草相对电导率和表型明显优于野生型.室温恢复实验证实转基因烟草可存活并恢复生长, 而野生型烟草则遭受了不可逆的低温冻害.该研究表明, 转化后携带昆虫抗冻蛋白基因的烟草相比野生型具有明显的抗寒能力. ...
Molecular analysis of acclimation to cold
1
1999
... 此外, 目前已发现的抗冻蛋白主要为动物来源, 其中以美洲拟蝶、绒杜父鱼(
Hemitripterus villosus )、美洲大绵(
Macrozoarces americanus )和南极鳕鱼(
Dissostichus eleginoides )来源为4大代表类型(
蔺忠龙等, 2009 ).虽然抗冻蛋白在包括鱼类、昆虫和植物在内的不同生物体中广泛存在, 但鱼类和昆虫来源的AFP研究起步较早, 目前已通过转基因技术将鱼类和昆虫来源的抗冻蛋白转入拟南芥、烟草和番茄(
Solanum lycopersicum )等模式植物中, 并在低温处理下获得了较好的抗冻效果(
Perl et al., 1993 ;
Pearce, 1999 ).但是植物来源的
AFP 基因研究起步较晚, 且已有研究的绝大多数植物材料中,
AFP 基因活性远低于鱼类和昆虫.虽然所研究的植物种类已有几十种, 但真正被分离纯化的植物源抗冻蛋白却非常少.因此, 如何从植物中分离出更多的、活性更高的AFP, 并通过遗传转化将其导入抗冻性弱或不抗冻的植物中, 进而获得抗冻性强的转基因植株, 将是今后植物抗冻基因工程研究的主要内容之一. ...
Enhanced oxidative-stress defense in transgenic potato expressing tomato Cu, Zn superoxide dismutases
1
1993
... 此外, 目前已发现的抗冻蛋白主要为动物来源, 其中以美洲拟蝶、绒杜父鱼(
Hemitripterus villosus )、美洲大绵(
Macrozoarces americanus )和南极鳕鱼(
Dissostichus eleginoides )来源为4大代表类型(
蔺忠龙等, 2009 ).虽然抗冻蛋白在包括鱼类、昆虫和植物在内的不同生物体中广泛存在, 但鱼类和昆虫来源的AFP研究起步较早, 目前已通过转基因技术将鱼类和昆虫来源的抗冻蛋白转入拟南芥、烟草和番茄(
Solanum lycopersicum )等模式植物中, 并在低温处理下获得了较好的抗冻效果(
Perl et al., 1993 ;
Pearce, 1999 ).但是植物来源的
AFP 基因研究起步较晚, 且已有研究的绝大多数植物材料中,
AFP 基因活性远低于鱼类和昆虫.虽然所研究的植物种类已有几十种, 但真正被分离纯化的植物源抗冻蛋白却非常少.因此, 如何从植物中分离出更多的、活性更高的AFP, 并通过遗传转化将其导入抗冻性弱或不抗冻的植物中, 进而获得抗冻性强的转基因植株, 将是今后植物抗冻基因工程研究的主要内容之一. ...
Interaction of tenebrio molitor antifreeze protein with ice crystal: insights from molecular dynamics simulations
1
2016
... 黄粉虫(
Tenebrio molitor )抗冻蛋白(TmAFP)二聚体的抗冻活性是已知抗冻蛋白中最高的, 可达鱼类抗冻蛋白活性的10-100倍.黄粉虫抗冻蛋白为右手β螺旋结构, 每一螺旋由12个保守的氨基酸组成(CT- XSXXCXXAXT, X为任一氨基酸残基), 依据12个氨基酸保守结构域重复次数可将
TmAFP 基因克隆分为3类, 分别编码84、96和120个氨基酸.TmAFP富含苏氨酸和半胱氨酸, 沿着蛋白折叠的一侧有两行苏氨酸残基与冰晶表面的棱柱和基面匹配结合(
Yue and Zhang, 2009 ).当接近冰点温度时, 抗冻蛋白表面冰结合位点上的苏氨酸侧链形成更优化的旋转异构体, 这种特有的冰结合构造使抗冻蛋白-冰分界面与冰晶相匹配, 从而阻止冰晶增长(
Ramya and Ramakrishnan, 2016 ).研究者将克隆到的
TmAFP 基因进行原核表达, 其表达蛋白经纯化后表现出冰结合活性和体外抗冻活性, 进一步将TmAFP蛋白代替传统抗冻保护剂DMSO制备大肠杆菌感受态细胞, 获得了转化高效的感受态细胞(
刘忠渊等, 2006 ).在提高植物抗冻能力方面, 有研究者将胡萝卜(
Daucus carota )抗冻蛋白基因(
DcAFP )和黄粉虫抗冻蛋白基因(
TmAFP )串联构建融合基因表达载体, 通过农杆菌介导转化拟南芥, 证实
TmAFP 的转基因拟南芥抗冻能力增强(
张振华等, 2012 ).然而, 至今未见有
TmAFP 基因在其它模式植物和经济作物中的转基因研究报道.因此, 通过基因工程技术将黄粉虫抗冻蛋白基因转入甘薯体内使其产生抗冻蛋白, 有望提高甘薯抗冻耐寒能力. ...
A vacuolar Na
+ /H
+ antiporter gene,
IbNHX2 , enhances salt and drought tolerance in transgenic sweetpotato
1
2016
... 近年来, 随着分子生物学的快速发展, 利用基因工程对植物性状进行遗传改良, 以获得具有特定优异性状新育种材料的技术体系已日趋成熟(
Wang et al., 2003 ).由于植物长期暴露于盐碱、干旱和寒冷等非生物逆境, 其本身会触发许多共同的防御机制, 如ABA含量增加、可溶性糖和保护性蛋白质的积累以及抗氧化水平的提升.因此将非生物胁迫相关的植物源基因导入可提高甘薯对盐碱、干旱、低温和氧化逆境等的抗性(
翟红等, 2017 ).例如, 将
Cu /
Zn SOD 和
APX 基因导入甘薯, 可增强转基因甘薯的耐盐性(
王欣等, 2011 ); 将菠菜 (
Spinacia oleracea )
SoBADH 基因导入甘薯可增强转基因甘薯对盐、低温及氧化胁迫的抗性(
Fan et al., 2012 ).同时, 可通过克隆甘薯自身的抗性基因, 然后增强其表达的方式创制转基因甘薯新材料.例如, 分别从甘薯耐盐株系ND98中克隆了
IbNFU1 、
IbP5CR 、
IbMas 、
IbSIMT1 和
IbNHX2 基因, 并在甘薯中过表达这些基因, 可显著增强转基因甘薯植株的耐盐性(
Wang et al., 2013 ,
2016 ;
Liu et al., 2014a ,
2014b ,
2015 ).另有研究发现, 在盐及干旱胁迫下, 将甘薯
IbMIP1 过表达后可显著上调肌醇生物合成、磷脂酰肌醇(PI)信号途径、ABA信号途径及胁迫响应等相关基因的表达, 进而显著提高转基因甘薯的耐盐性和抗旱性(
Zhai et al., 2016 ).虽然甘薯转基因体系构建和相关技术趋于成熟, 但与甘薯抗旱、耐盐和抗氧化转基因研究相比, 甘薯抗冻基因及转基因植株培育进展相对缓慢, 目前尚未见甘薯抗冻基因遗传转化的研究报道. ...
Structural basis of antifreeze activity of a bacterial multi-domain antifreeze protein
1
2017
... 由于甘薯具有杂交不亲和性及结实率低等特点, 严重限制了甘薯育种中的资源利用, 因此基因工程是改良甘薯品种的一种有效手段(
Yang et al., 2009 ).抗冻蛋白(antifreeze protein, AFP)又称热滞蛋白(thermal hysteresis protein, THP), 具有多个能够直接作用于冰晶的亲水性冰结合位点, 可阻止低温下冰晶在细胞间隙中的形成和再结晶, 以维持生物体正常的新陈代谢(
Wang et al., 2017 ).自De Vrie等(1969)从一种大西洋冷水鱼血液中发现抗冻蛋白以来, 其在细菌、昆虫、鱼类和植物中也相继被发现, 尤其是昆虫抗冻蛋白具有抗冻活性高和结构规律等特点, 其滞热活性与其它生物的抗冻蛋白相比高10-100倍, 这使得昆虫抗冻蛋白在植物抗冻遗传转化中具有巨大的优势和潜力(
Nada and Furukawa, 2011 ).Davies等(1987)首次将美洲拟鲽(
Pseudopleuronectes americanus )抗冻蛋白基因整合到Ti质粒中, 通过叶圆片法转化郁金香(
Tulipa gesneriana )和油菜(
Brassica campestris )等, 使转基因植株获得了一定的抗冻能力(
Zhang and Davies, 1987 ).
Cutler等(1989) 用真空透析法将冬比目鱼(
Pleuronectiformes heterosomata )抗冻蛋白基因导入马铃薯(
Solanum tuberosum )、拟南芥(
Arabidopsis thaliana )和欧洲油菜(
B. napus ), 使此类植物的自然结冰温度降低1.8°C, 证实转动物源抗冻蛋白基因可提高植物的抗寒性(
Cutler et al., 1989 ).
Wang等(2009) 用准噶尔小胸鳖甲(
Microdera punctipennis )抗冻蛋白基因
MPAFP149 转化烟草(
Nicotiana tabacum ), 并对T
0 代转基因烟草在-1°C处理48小时后, 发现转基因烟草相对电导率和表型明显优于野生型.室温恢复实验证实转基因烟草可存活并恢复生长, 而野生型烟草则遭受了不可逆的低温冻害.该研究表明, 转化后携带昆虫抗冻蛋白基因的烟草相比野生型具有明显的抗寒能力. ...
Molecular cloning and functional characterization of a salt tolerance-associated gene
IbNFU1 from sweetpotato
1
2013
... 近年来, 随着分子生物学的快速发展, 利用基因工程对植物性状进行遗传改良, 以获得具有特定优异性状新育种材料的技术体系已日趋成熟(
Wang et al., 2003 ).由于植物长期暴露于盐碱、干旱和寒冷等非生物逆境, 其本身会触发许多共同的防御机制, 如ABA含量增加、可溶性糖和保护性蛋白质的积累以及抗氧化水平的提升.因此将非生物胁迫相关的植物源基因导入可提高甘薯对盐碱、干旱、低温和氧化逆境等的抗性(
翟红等, 2017 ).例如, 将
Cu /
Zn SOD 和
APX 基因导入甘薯, 可增强转基因甘薯的耐盐性(
王欣等, 2011 ); 将菠菜 (
Spinacia oleracea )
SoBADH 基因导入甘薯可增强转基因甘薯对盐、低温及氧化胁迫的抗性(
Fan et al., 2012 ).同时, 可通过克隆甘薯自身的抗性基因, 然后增强其表达的方式创制转基因甘薯新材料.例如, 分别从甘薯耐盐株系ND98中克隆了
IbNFU1 、
IbP5CR 、
IbMas 、
IbSIMT1 和
IbNHX2 基因, 并在甘薯中过表达这些基因, 可显著增强转基因甘薯植株的耐盐性(
Wang et al., 2013 ,
2016 ;
Liu et al., 2014a ,
2014b ,
2015 ).另有研究发现, 在盐及干旱胁迫下, 将甘薯
IbMIP1 过表达后可显著上调肌醇生物合成、磷脂酰肌醇(PI)信号途径、ABA信号途径及胁迫响应等相关基因的表达, 进而显著提高转基因甘薯的耐盐性和抗旱性(
Zhai et al., 2016 ).虽然甘薯转基因体系构建和相关技术趋于成熟, 但与甘薯抗旱、耐盐和抗氧化转基因研究相比, 甘薯抗冻基因及转基因植株培育进展相对缓慢, 目前尚未见甘薯抗冻基因遗传转化的研究报道. ...
Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance
1
2003
... 近年来, 随着分子生物学的快速发展, 利用基因工程对植物性状进行遗传改良, 以获得具有特定优异性状新育种材料的技术体系已日趋成熟(
Wang et al., 2003 ).由于植物长期暴露于盐碱、干旱和寒冷等非生物逆境, 其本身会触发许多共同的防御机制, 如ABA含量增加、可溶性糖和保护性蛋白质的积累以及抗氧化水平的提升.因此将非生物胁迫相关的植物源基因导入可提高甘薯对盐碱、干旱、低温和氧化逆境等的抗性(
翟红等, 2017 ).例如, 将
Cu /
Zn SOD 和
APX 基因导入甘薯, 可增强转基因甘薯的耐盐性(
王欣等, 2011 ); 将菠菜 (
Spinacia oleracea )
SoBADH 基因导入甘薯可增强转基因甘薯对盐、低温及氧化胁迫的抗性(
Fan et al., 2012 ).同时, 可通过克隆甘薯自身的抗性基因, 然后增强其表达的方式创制转基因甘薯新材料.例如, 分别从甘薯耐盐株系ND98中克隆了
IbNFU1 、
IbP5CR 、
IbMas 、
IbSIMT1 和
IbNHX2 基因, 并在甘薯中过表达这些基因, 可显著增强转基因甘薯植株的耐盐性(
Wang et al., 2013 ,
2016 ;
Liu et al., 2014a ,
2014b ,
2015 ).另有研究发现, 在盐及干旱胁迫下, 将甘薯
IbMIP1 过表达后可显著上调肌醇生物合成、磷脂酰肌醇(PI)信号途径、ABA信号途径及胁迫响应等相关基因的表达, 进而显著提高转基因甘薯的耐盐性和抗旱性(
Zhai et al., 2016 ).虽然甘薯转基因体系构建和相关技术趋于成熟, 但与甘薯抗旱、耐盐和抗氧化转基因研究相比, 甘薯抗冻基因及转基因植株培育进展相对缓慢, 目前尚未见甘薯抗冻基因遗传转化的研究报道. ...
Development and evaluation of a storage root-bearing sweetpotato somatic hybrid between Ipomoea batatas(L.) Lam. and
I. triloba L
1
2009
... 由于甘薯具有杂交不亲和性及结实率低等特点, 严重限制了甘薯育种中的资源利用, 因此基因工程是改良甘薯品种的一种有效手段(
Yang et al., 2009 ).抗冻蛋白(antifreeze protein, AFP)又称热滞蛋白(thermal hysteresis protein, THP), 具有多个能够直接作用于冰晶的亲水性冰结合位点, 可阻止低温下冰晶在细胞间隙中的形成和再结晶, 以维持生物体正常的新陈代谢(
Wang et al., 2017 ).自De Vrie等(1969)从一种大西洋冷水鱼血液中发现抗冻蛋白以来, 其在细菌、昆虫、鱼类和植物中也相继被发现, 尤其是昆虫抗冻蛋白具有抗冻活性高和结构规律等特点, 其滞热活性与其它生物的抗冻蛋白相比高10-100倍, 这使得昆虫抗冻蛋白在植物抗冻遗传转化中具有巨大的优势和潜力(
Nada and Furukawa, 2011 ).Davies等(1987)首次将美洲拟鲽(
Pseudopleuronectes americanus )抗冻蛋白基因整合到Ti质粒中, 通过叶圆片法转化郁金香(
Tulipa gesneriana )和油菜(
Brassica campestris )等, 使转基因植株获得了一定的抗冻能力(
Zhang and Davies, 1987 ).
Cutler等(1989) 用真空透析法将冬比目鱼(
Pleuronectiformes heterosomata )抗冻蛋白基因导入马铃薯(
Solanum tuberosum )、拟南芥(
Arabidopsis thaliana )和欧洲油菜(
B. napus ), 使此类植物的自然结冰温度降低1.8°C, 证实转动物源抗冻蛋白基因可提高植物的抗寒性(
Cutler et al., 1989 ).
Wang等(2009) 用准噶尔小胸鳖甲(
Microdera punctipennis )抗冻蛋白基因
MPAFP149 转化烟草(
Nicotiana tabacum ), 并对T
0 代转基因烟草在-1°C处理48小时后, 发现转基因烟草相对电导率和表型明显优于野生型.室温恢复实验证实转基因烟草可存活并恢复生长, 而野生型烟草则遭受了不可逆的低温冻害.该研究表明, 转化后携带昆虫抗冻蛋白基因的烟草相比野生型具有明显的抗寒能力. ...
Cloning and expression of Tenebrio molitor antifreeze protein in
Escherichia coli 1
2009
... 黄粉虫(
Tenebrio molitor )抗冻蛋白(TmAFP)二聚体的抗冻活性是已知抗冻蛋白中最高的, 可达鱼类抗冻蛋白活性的10-100倍.黄粉虫抗冻蛋白为右手β螺旋结构, 每一螺旋由12个保守的氨基酸组成(CT- XSXXCXXAXT, X为任一氨基酸残基), 依据12个氨基酸保守结构域重复次数可将
TmAFP 基因克隆分为3类, 分别编码84、96和120个氨基酸.TmAFP富含苏氨酸和半胱氨酸, 沿着蛋白折叠的一侧有两行苏氨酸残基与冰晶表面的棱柱和基面匹配结合(
Yue and Zhang, 2009 ).当接近冰点温度时, 抗冻蛋白表面冰结合位点上的苏氨酸侧链形成更优化的旋转异构体, 这种特有的冰结合构造使抗冻蛋白-冰分界面与冰晶相匹配, 从而阻止冰晶增长(
Ramya and Ramakrishnan, 2016 ).研究者将克隆到的
TmAFP 基因进行原核表达, 其表达蛋白经纯化后表现出冰结合活性和体外抗冻活性, 进一步将TmAFP蛋白代替传统抗冻保护剂DMSO制备大肠杆菌感受态细胞, 获得了转化高效的感受态细胞(
刘忠渊等, 2006 ).在提高植物抗冻能力方面, 有研究者将胡萝卜(
Daucus carota )抗冻蛋白基因(
DcAFP )和黄粉虫抗冻蛋白基因(
TmAFP )串联构建融合基因表达载体, 通过农杆菌介导转化拟南芥, 证实
TmAFP 的转基因拟南芥抗冻能力增强(
张振华等, 2012 ).然而, 至今未见有
TmAFP 基因在其它模式植物和经济作物中的转基因研究报道.因此, 通过基因工程技术将黄粉虫抗冻蛋白基因转入甘薯体内使其产生抗冻蛋白, 有望提高甘薯抗冻耐寒能力. ...
A
myo -inositol-1-phosphate synthase gene,
IbMIPS1 , enhances salt and drought tolerance and stem nematode resistance in transgenic sweet potato
1
2016
... 近年来, 随着分子生物学的快速发展, 利用基因工程对植物性状进行遗传改良, 以获得具有特定优异性状新育种材料的技术体系已日趋成熟(
Wang et al., 2003 ).由于植物长期暴露于盐碱、干旱和寒冷等非生物逆境, 其本身会触发许多共同的防御机制, 如ABA含量增加、可溶性糖和保护性蛋白质的积累以及抗氧化水平的提升.因此将非生物胁迫相关的植物源基因导入可提高甘薯对盐碱、干旱、低温和氧化逆境等的抗性(
翟红等, 2017 ).例如, 将
Cu /
Zn SOD 和
APX 基因导入甘薯, 可增强转基因甘薯的耐盐性(
王欣等, 2011 ); 将菠菜 (
Spinacia oleracea )
SoBADH 基因导入甘薯可增强转基因甘薯对盐、低温及氧化胁迫的抗性(
Fan et al., 2012 ).同时, 可通过克隆甘薯自身的抗性基因, 然后增强其表达的方式创制转基因甘薯新材料.例如, 分别从甘薯耐盐株系ND98中克隆了
IbNFU1 、
IbP5CR 、
IbMas 、
IbSIMT1 和
IbNHX2 基因, 并在甘薯中过表达这些基因, 可显著增强转基因甘薯植株的耐盐性(
Wang et al., 2013 ,
2016 ;
Liu et al., 2014a ,
2014b ,
2015 ).另有研究发现, 在盐及干旱胁迫下, 将甘薯
IbMIP1 过表达后可显著上调肌醇生物合成、磷脂酰肌醇(PI)信号途径、ABA信号途径及胁迫响应等相关基因的表达, 进而显著提高转基因甘薯的耐盐性和抗旱性(
Zhai et al., 2016 ).虽然甘薯转基因体系构建和相关技术趋于成熟, 但与甘薯抗旱、耐盐和抗氧化转基因研究相比, 甘薯抗冻基因及转基因植株培育进展相对缓慢, 目前尚未见甘薯抗冻基因遗传转化的研究报道. ...
Increased synthesis of ABA in partially dehydrated root tips and ABA transport from roots to leaves
1
1987
... 由于甘薯具有杂交不亲和性及结实率低等特点, 严重限制了甘薯育种中的资源利用, 因此基因工程是改良甘薯品种的一种有效手段(
Yang et al., 2009 ).抗冻蛋白(antifreeze protein, AFP)又称热滞蛋白(thermal hysteresis protein, THP), 具有多个能够直接作用于冰晶的亲水性冰结合位点, 可阻止低温下冰晶在细胞间隙中的形成和再结晶, 以维持生物体正常的新陈代谢(
Wang et al., 2017 ).自De Vrie等(1969)从一种大西洋冷水鱼血液中发现抗冻蛋白以来, 其在细菌、昆虫、鱼类和植物中也相继被发现, 尤其是昆虫抗冻蛋白具有抗冻活性高和结构规律等特点, 其滞热活性与其它生物的抗冻蛋白相比高10-100倍, 这使得昆虫抗冻蛋白在植物抗冻遗传转化中具有巨大的优势和潜力(
Nada and Furukawa, 2011 ).Davies等(1987)首次将美洲拟鲽(
Pseudopleuronectes americanus )抗冻蛋白基因整合到Ti质粒中, 通过叶圆片法转化郁金香(
Tulipa gesneriana )和油菜(
Brassica campestris )等, 使转基因植株获得了一定的抗冻能力(
Zhang and Davies, 1987 ).
Cutler等(1989) 用真空透析法将冬比目鱼(
Pleuronectiformes heterosomata )抗冻蛋白基因导入马铃薯(
Solanum tuberosum )、拟南芥(
Arabidopsis thaliana )和欧洲油菜(
B. napus ), 使此类植物的自然结冰温度降低1.8°C, 证实转动物源抗冻蛋白基因可提高植物的抗寒性(
Cutler et al., 1989 ).
Wang等(2009) 用准噶尔小胸鳖甲(
Microdera punctipennis )抗冻蛋白基因
MPAFP149 转化烟草(
Nicotiana tabacum ), 并对T
0 代转基因烟草在-1°C处理48小时后, 发现转基因烟草相对电导率和表型明显优于野生型.室温恢复实验证实转基因烟草可存活并恢复生长, 而野生型烟草则遭受了不可逆的低温冻害.该研究表明, 转化后携带昆虫抗冻蛋白基因的烟草相比野生型具有明显的抗寒能力. ...
备案号:
京ICP备16067583号-21 版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由
北京玛格泰克科技发展有限公司 设计开发
 ,1,*
,1,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT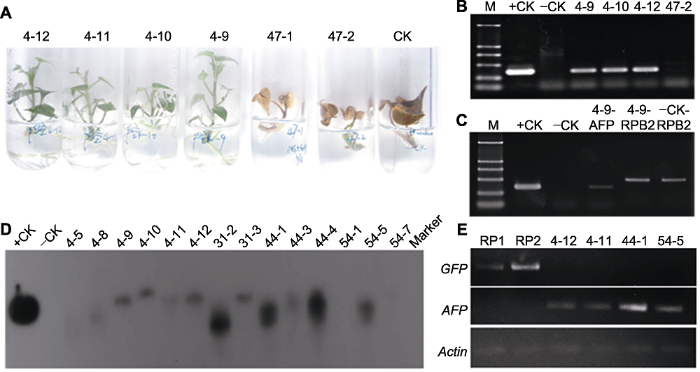 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT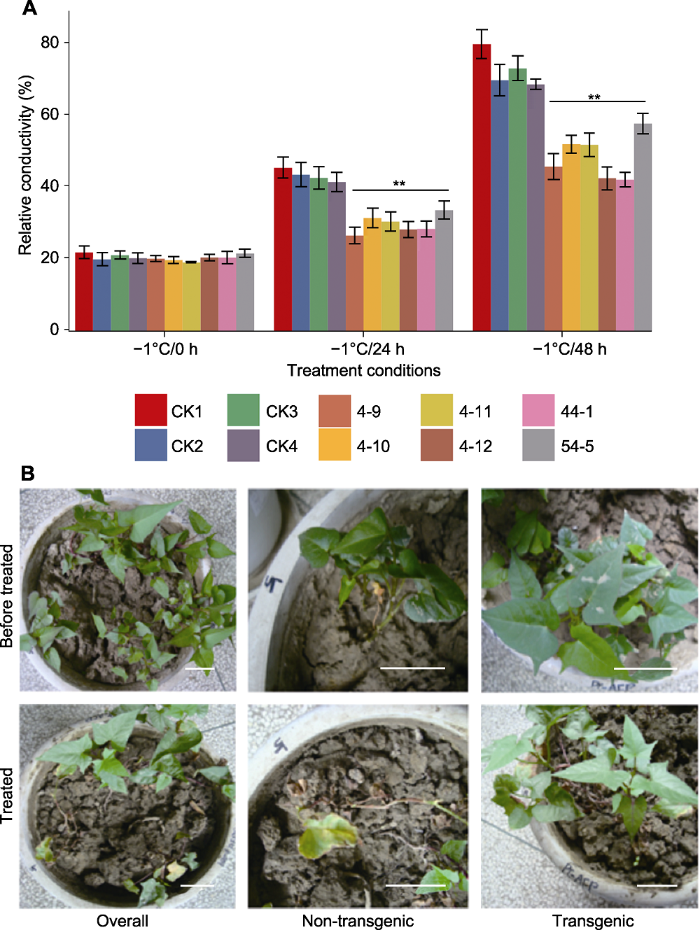 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}