Abstract SUMOylation, a post-translational modification, is essential for plant normal growth and development. To date, thousands of possible SUMO substrates have been identified, but due to the relatively low SUMOylation level, biological significance of the SUMOylation remains largely unknown. Here, we summarized the SUMOylation detection methods, including in vitro and in vivo SUMOylation assays, which help to understand the functions of SUMOylation in plants. Keywords:Arabidopsis;SUMO;SUMOylation;detection methods
PDF (1533KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 曲高平, 金京波. 植物蛋白SUMO化修饰检测方法. 植物学报, 2020, 55(1): 83-89 doi:10.11983/CBB19213 Qu Gaoping, Jin Jingbo. Detection of SUMOylation in Plants. Chinese Bulletin of Botany, 2020, 55(1): 83-89 doi:10.11983/CBB19213
拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用。SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007)。SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015)。拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21)。其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018)。SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018)。此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014)。它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域。拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017)。目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016)。
SUMO1和SUMO2的N端含有多个赖氨酸, 可形成SUMO链(polySUMOylation)。polySUMOylation或多位点SUMO化修饰(mutiSUMOylation or mutiple monoSUMOylation)可以招募含有SIM基序(SUMO interacting motif)的蛋白, 介导蛋白-蛋白的非共价结合。SUMO化修饰除了影响蛋白-蛋白互作外, 还调控底物蛋白的亚细胞定位、活性或稳定性等。截至目前, 通过酵母双杂交(yeast two-hybrid)和质谱分析(mass spectrometry)等方法, 已鉴定出上千个可能的SUMO底物(Elrouby and Coupland, 2010; Miller et al., 2010; Rytz et al., 2018)。但由于体内SUMO化修饰水平很低,且处于动态变化中(Sadanandom et al., 2015; Lin et al., 2016), 鉴定特定底物的SUMO化修饰较为困难。本文仅介绍检测植物蛋白SUMO化修饰的常用方法。若想了解更多关于SUMO化修饰的研究方法, 可参阅文献Rodriguez (2016)。
Table 1 表1 表1SUMO反应相关蛋白的诱导条件 Table 1Expression of SUMO reaction related proteins
Proteins
IPTG concentration (mmol·L-1)
Induction time
His-SUMO E1
1
16-20 h (16°C)
His-SUMO E2
1
3-5 h (28°C)
His-SUMO1GG
1
16-20 h (16°C)
His-SUMO1AA
1
16-20 h (16°C)
GST-protein X-Myc
1
3-5 h (28°C)
To enhance the western blot detection, 6×Myc tag was fused to the C-terminus of the target protein. IPTG: Isopropyl-β-D-thiogalactopyranoside 为提高western blot检测特异性, 可在目的条带的C端加1个6×Myc标签。IPTG: 异丙基-β-D-硫代半乳糖苷
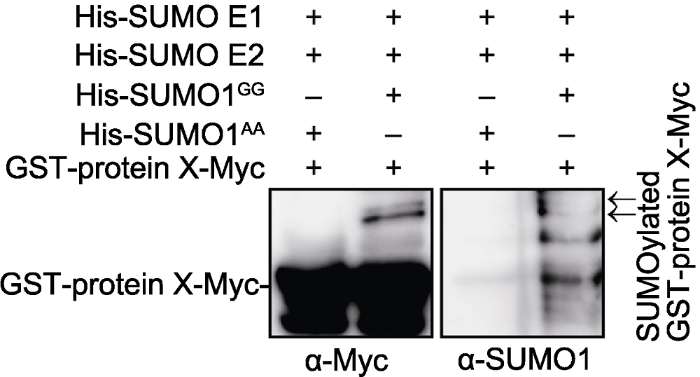
GST-protein X-Myc分别与His-SUMO E1、His-SUMO E2和His- SUMO1GG (His-SUMO1AA作为阴性对照), 进行体外SUMO化反应。反应后用Glutathione beads进一步纯化。Western blot, 分别用anti-Myc和anti-SUMO1抗体进行检测。箭头指示SUMO化修饰的GST-protein X-Myc。 Figure 1Protein X can be SUMOylated in vitro
GST-protein X-Myc was incubated with His-SUMO E1, His- SUMO E2 and His-SUMO1GG (His-SUMO1AA was used as a negative control) at 30°C for 3 h. After reaction, GST-protein X-Myc was purified with Glutathione beads and detected with anti-Myc and anti-SUMO1 antibodies. Arrows represent SUMOylated GST-protein X-Myc.
GFP-protein X分别与FLAG-SUMO1GG或FLAG-SUMO1AA在烟草叶片中共表达, IP (anti-GFP)产物分别用anti-GFP和anti- FLAG抗体检测。 Figure 2SUMOylation of GFP-protein X in Nicotiana benthamiana
GFP-protein X was transiently co-expressed with FLAG-SUMO1GG or FLAG-SUMO1AA in Nicotiana benthamiana leaves. GFP-protein X was immunoprecipitated with anti-GFP antibody, and IP products were detected with anti-FLAG antibody.
Myc-protein X转基因材料和野生型(WT)在对照条件(control, C)或处理条件(treatment, T)下生长3天。Myc-protein X和SUMO化修饰的Myc-protein X分别用anti-protein X和anti- SUMO1抗体检测。竖线指示SUMO化修饰的Myc-protein X。 Figure 3SUMOylation of Myc-protein X in transgenic plants
Myc-protein X transgenic plants and wild-type (WT) were grown in control (C) or treatment (T) conditions for 3 days. Myc-protein X and SUMOylated Myc-protein X were detected with anti-protein X and anti-SUMO1 antibodies, respectively. Vertical line indicates SUMOylated Myc-protein X bands.
5.3 SUMO化修饰位点的检测
SUMO底物通常含有保守的ΨKXD/E序列, 其中Ψ代表疏水性氨基酸, K是SUMO分子共价结合的赖氨酸, X为任意氨基酸, D/E为酸性氨基酸(天冬氨酸(D)或谷氨酸(E))。SUMO结合酶E2可识别ΨKXD/E基序, 介导底物SUMO化修饰(Bernier-Villamor et al., 2002)。常用的SUMO化修饰分析软件有GPS-SUMO (Zhao et al., 2014)、SUMOplot (http://www.abgent.com/umoplot)以及JASSA (http://www.jassa.fr/index.hp?jassa)。但由于SUMO化修饰位点并非均是保守的ΨKXD/E基序, 故还需要结合质谱检测进行分析。为提高质谱检测效率, 通常使用SUMO1H89R检测底物的SUMO化修饰位点(Miller et al., 2010)。
通过软件分析和质谱检测获得可能的SUMO化修饰结合位点, 然后通过SUMO化实验进行验证。目前, 判断SUMO化修饰结合位点的主要依据是SUMO结合位点赖氨酸(K)突变成精氨酸(R)能抑制底物SUMO化修饰(Bernier-Villamor et al., 2002)。如图4所示, Myc-protein X和FLAG-SUMO1GG在烟草中共表达, IP (anti-Myc)产物用anti-FALG抗体能检测到SUMO化修饰的Myc-protein X, 说明protein X是SUMO底物。当把protein X两个可能的SUMO化修饰结合位点K1和K2单独突变后, 仍能检测到SUMO化修饰条带, 但2个位点同时突变(2KR)后, 几乎检测不到, 说明K1和K2是protein X主要的SUMO化修饰结合位点。
Myc-protein X、Myc-protein XK1R、Myc-protein XK2R或Myc- protein X2KR分别与FLAG-SUMO1GG或FLAG-SUMO1AA在烟草中瞬时共表达。 Figure 4K1 and K2 are the primary SUMOylation sites of protein X
Myc-protein X, Myc-protein XK1R, Myc-protein XK2R or Myc- protein X2KR was transiently co-expressed with FLAG-SUMO1GG or FLAG-SUMO1AA in Nicotiana benthamiana leaves, respectively.
ColbyT, Matth?iA, BoeckelmannA, StuibleHP (2006). SUMO-conjugating and SUMO-deconjugating enzymes from Arabidopsis Plant Physiol 142, 318-332. [本文引用: 1]
ContiL, PriceG, O'DonnellE, SchwessingerB, DominyP, SadanandomA (2008). Small ubiquitin-like modifier proteases OVERLY TOLERANT TO SALT1 and -2 regulate salt stress responses in Arabidopsis Plant Cell 20, 2894-2908. [本文引用: 1]
ElroubyN, CouplandG (2010). Proteome-wide screens for small ubiquitin-like modifier (SUMO) substrates identify Arabidopsis proteins implicated in diverse biological processes Proc Natl Acad Sci USA 107, 17415-17420. [本文引用: 1]
HermkesR, FuYF, NürrenbergK, BudhirajaR, SchmelzerE, ElroubyN, DohmenRJ, BachmairA, CouplandG (2011). Distinct roles for Arabidopsis SUMO protease ESD4 and its closest homolog ELS1 Planta 233, 63-73. [本文引用: 1]
IshidaT, YoshimuraM, MiuraK, SugimotoK (2012). MMS21/HPY2 and SIZ1, two Arabidopsis SUMO E3 ligases, have distinct functions in development PLoS One 7, e46897. [本文引用: 1]
LiuLP, JiangY, ZhangXM, WangX, WangYB, HanYZ, CouplandG, JinJB, SearleI, FuYF, ChenFL (2017). Two SUMO proteases SUMO PROTEASE RELATED TO FERTILITY1 and 2 are required for fertility in Arabidopsis Plant Physiol 175, 1703-1719. [本文引用: 1]
MillerMJ, Barrett-WiltGA, HuaZH, VierstraRD (2010). Proteomic analyses identify a diverse array of nuclear processes affected by small ubiquitin-like modifier conjugation in Arabidopsis Proc Natl Acad Sci USA 107, 16512-16517. [本文引用: 2]
MurtasG, ReevesPH, FuYF, BancroftI, DeanC, CouplandG (2003). A nuclear protease required for flowering-time regulation in Arabidopsis reduces the abundance of SMALL UBIQUITIN-RELATED MODIFIER conjugates Plant Cell 15, 2308-2319. [本文引用: 1]
RodriguezMS (2016). SUMO: Methods and Protocols . New York: Humana Press. [本文引用: 1]
RytzTC, MillerMJ, McLoughlinF, AugustineRC, MarshallRS, JuanYT, CharngYY, ScalfM, SmithLM, VierstraRD (2018). SUMOylome profiling reveals a diverse array of nuclear targets modified by the SUMO ligase SIZ1 during heat stress Plant Cell 30, 1077-1099. [本文引用: 3]
SadanandomA, ádámé, OrosaB, VicziánA, KloseC, ZhangCJ, JosseEM, Kozma-BognárL, NagyF (2015). SUMOylation of phytochrome-B negatively regulates light- induced signaling in Arabidopsis thaliana Proc Natl Acad Sci USA 112, 11108-11113. [本文引用: 2]
SalehA, WithersJ, MohanR, MarquésJ, GuYN, YanSP, ZavalievR, NomotoM, TadaY, DongXN (2015). Posttranslational modifications of the master transcriptional regulator NPR1 enable dynamic but tight control of plant immune responses Cell Host Microbe 18, 169-182. [本文引用: 1]
SaraccoSA, MillerMJ, KurepaJ, VierstraRD (2007). Genetic analysis of SUMOylation in Arabidopsis: conjugation of SUMO1 and SUMO2 to nuclear proteins is essential Plant Physiol 145, 119-134. [本文引用: 2]
TomanovK, ZeschmannA, HermkesR, EiflerK, ZibaI, GriecoM, NovatchkovaM, HofmannK, HesseH, BachmairA (2014). Arabidopsis PIAL1 and 2 promote SUMO chain formation as E4-type SUMO ligases and are involved in stress responses and sulfur metabolism Plant Cell 26, 4547-4560. [本文引用: 1]
YatesG, SrivastavaAK, SadanandomA (2016). SUMO proteases: uncovering the roles of deSUMOylation in plants J Exp Bot 67, 2541-2548. [本文引用: 1]
YooSD, ChoYH, SheenJ (2007). Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis Nat Protoc 2, 1565-1572. [本文引用: 1]
ZhaoQ, XieYB, ZhengYY, JiangS, LiuWZ, MuWP, LiuZX, ZhaoY, XueY, RenJ (2014). GPS-SUMO: a tool for the prediction of SUMOylation sites and SUMO-interaction motifs Nucleic Acids Res 42, W325-W330. [本文引用: 1]
植物SUMO化修饰及其生物学功能 1 2008
... 拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用.SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007).SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015).拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21).其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018).SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
Structural basis for E2-mediated SUMO conjugation revealed by a complex between ubiquitin-conjugating enzyme Ubc9 and RanGAP1 3 2002
The Arabidopsis E3 SUMO ligase SIZ1 regulates plant growth and drought responses 1 2007
... 拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用.SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007).SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015).拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21).其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018).SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
SUMO-conjugating and SUMO-deconjugating enzymes from Arabidopsis 1 2006
... (1) 体外SUMO反应: pGEX-5X-protein X-Myc (GST- protein X-Myc)、pRQ30-SAE1b-SAE2 (His-SUMO E1) (Colby et al., 2006)、pRSET-A-SCE1 (His- SUMO E2)、pRSET-A-SUMO1GG (His-SUMO1GG)和pRSET-A-SUMO1AA (His-SUMO1AA). ...
Small ubiquitin-like modifier proteases OVERLY TOLERANT TO SALT1 and -2 regulate salt stress responses in Arabidopsis 1 2008
... 拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用.SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007).SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015).拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21).其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018).SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
Proteome-wide screens for small ubiquitin-like modifier (SUMO) substrates identify Arabidopsis proteins implicated in diverse biological processes 1 2010
... SUMO1和SUMO2的N端含有多个赖氨酸, 可形成SUMO链(polySUMOylation).polySUMOylation或多位点SUMO化修饰(mutiSUMOylation or mutiple monoSUMOylation)可以招募含有SIM基序(SUMO interacting motif)的蛋白, 介导蛋白-蛋白的非共价结合.SUMO化修饰除了影响蛋白-蛋白互作外, 还调控底物蛋白的亚细胞定位、活性或稳定性等.截至目前, 通过酵母双杂交(yeast two-hybrid)和质谱分析(mass spectrometry)等方法, 已鉴定出上千个可能的SUMO底物(Elrouby and Coupland, 2010; Miller et al., 2010; Rytz et al., 2018).但由于体内SUMO化修饰水平很低,且处于动态变化中(Sadanandom et al., 2015; Lin et al., 2016), 鉴定特定底物的SUMO化修饰较为困难.本文仅介绍检测植物蛋白SUMO化修饰的常用方法.若想了解更多关于SUMO化修饰的研究方法, 可参阅文献Rodriguez (2016). ...
Distinct roles for Arabidopsis SUMO protease ESD4 and its closest homolog ELS1 1 2011
... 拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用.SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007).SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015).拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21).其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018).SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
MMS21/HPY2 and SIZ1, two Arabidopsis SUMO E3 ligases, have distinct functions in development 1 2012
... 拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用.SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007).SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015).拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21).其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018).SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
Arabidopsis SUMO protease ASP1 positively regulates flowering time partially through regulating FLC stability 1 2017
... 拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用.SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007).SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015).拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21).其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018).SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
An Arabidopsis SUMO E3 ligase, SIZ1, negatively regulates photomorphogenesis by promoting COP1 activity 2 2016
... SUMO1和SUMO2的N端含有多个赖氨酸, 可形成SUMO链(polySUMOylation).polySUMOylation或多位点SUMO化修饰(mutiSUMOylation or mutiple monoSUMOylation)可以招募含有SIM基序(SUMO interacting motif)的蛋白, 介导蛋白-蛋白的非共价结合.SUMO化修饰除了影响蛋白-蛋白互作外, 还调控底物蛋白的亚细胞定位、活性或稳定性等.截至目前, 通过酵母双杂交(yeast two-hybrid)和质谱分析(mass spectrometry)等方法, 已鉴定出上千个可能的SUMO底物(Elrouby and Coupland, 2010; Miller et al., 2010; Rytz et al., 2018).但由于体内SUMO化修饰水平很低,且处于动态变化中(Sadanandom et al., 2015; Lin et al., 2016), 鉴定特定底物的SUMO化修饰较为困难.本文仅介绍检测植物蛋白SUMO化修饰的常用方法.若想了解更多关于SUMO化修饰的研究方法, 可参阅文献Rodriguez (2016). ...
... (3) Western blot检测 用30 µL 2×SDS上样缓冲液重悬beads, 100°C煮沸5分钟, 瞬时离心, 混匀.1 000 ×g离心3分钟, 上样.Western blot, anti-SUMO1抗体仅在实验组(转基因材料)检测到目的条带上方间隔×11 kDa (SUMO1大约11 kDa)处有1条或多条条带, 说明目的蛋白在拟南芥中能被SUMO化修饰.如图3所示, Myc-protein X过表达材料, IP (anti-Myc)产物用anti-SUMO1抗体能在处理组(T)特异检测到SUMO化修饰的Myc-protein X, 但在对照组(C)未检测到SUMO化修饰的Myc-protein X, 这说明Myc-protein X仅在特定处理条件下才被SUMO化修饰.因此, 需结合表型, 选择植物特定发育阶段、特定生长条件或特定处理条件进行SUMO化修饰鉴定(Sadanandom et al., 2015; Lin et al., 2016). ...
Two SUMO proteases SUMO PROTEASE RELATED TO FERTILITY1 and 2 are required for fertility in Arabidopsis 1 2017
... 拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用.SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007).SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015).拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21).其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018).SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
Proteomic analyses identify a diverse array of nuclear processes affected by small ubiquitin-like modifier conjugation in Arabidopsis 2 2010
... SUMO1和SUMO2的N端含有多个赖氨酸, 可形成SUMO链(polySUMOylation).polySUMOylation或多位点SUMO化修饰(mutiSUMOylation or mutiple monoSUMOylation)可以招募含有SIM基序(SUMO interacting motif)的蛋白, 介导蛋白-蛋白的非共价结合.SUMO化修饰除了影响蛋白-蛋白互作外, 还调控底物蛋白的亚细胞定位、活性或稳定性等.截至目前, 通过酵母双杂交(yeast two-hybrid)和质谱分析(mass spectrometry)等方法, 已鉴定出上千个可能的SUMO底物(Elrouby and Coupland, 2010; Miller et al., 2010; Rytz et al., 2018).但由于体内SUMO化修饰水平很低,且处于动态变化中(Sadanandom et al., 2015; Lin et al., 2016), 鉴定特定底物的SUMO化修饰较为困难.本文仅介绍检测植物蛋白SUMO化修饰的常用方法.若想了解更多关于SUMO化修饰的研究方法, 可参阅文献Rodriguez (2016). ...
... SUMO底物通常含有保守的ΨKXD/E序列, 其中Ψ代表疏水性氨基酸, K是SUMO分子共价结合的赖氨酸, X为任意氨基酸, D/E为酸性氨基酸(天冬氨酸(D)或谷氨酸(E)).SUMO结合酶E2可识别ΨKXD/E基序, 介导底物SUMO化修饰(Bernier-Villamor et al., 2002).常用的SUMO化修饰分析软件有GPS-SUMO (Zhao et al., 2014)、SUMOplot (http://www.abgent.com/umoplot)以及JASSA (http://www.jassa.fr/index.hp?jassa).但由于SUMO化修饰位点并非均是保守的ΨKXD/E基序, 故还需要结合质谱检测进行分析.为提高质谱检测效率, 通常使用SUMO1H89R检测底物的SUMO化修饰位点(Miller et al., 2010). ...
A nuclear protease required for flowering-time regulation in Arabidopsis reduces the abundance of SMALL UBIQUITIN-RELATED MODIFIER conjugates 1 2003
... 拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用.SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007).SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015).拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21).其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018).SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
1 2016
... SUMO1和SUMO2的N端含有多个赖氨酸, 可形成SUMO链(polySUMOylation).polySUMOylation或多位点SUMO化修饰(mutiSUMOylation or mutiple monoSUMOylation)可以招募含有SIM基序(SUMO interacting motif)的蛋白, 介导蛋白-蛋白的非共价结合.SUMO化修饰除了影响蛋白-蛋白互作外, 还调控底物蛋白的亚细胞定位、活性或稳定性等.截至目前, 通过酵母双杂交(yeast two-hybrid)和质谱分析(mass spectrometry)等方法, 已鉴定出上千个可能的SUMO底物(Elrouby and Coupland, 2010; Miller et al., 2010; Rytz et al., 2018).但由于体内SUMO化修饰水平很低,且处于动态变化中(Sadanandom et al., 2015; Lin et al., 2016), 鉴定特定底物的SUMO化修饰较为困难.本文仅介绍检测植物蛋白SUMO化修饰的常用方法.若想了解更多关于SUMO化修饰的研究方法, 可参阅文献Rodriguez (2016). ...
SUMOylome profiling reveals a diverse array of nuclear targets modified by the SUMO ligase SIZ1 during heat stress 3 2018
... 拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用.SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007).SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015).拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21).其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018).SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
... ; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
... SUMO1和SUMO2的N端含有多个赖氨酸, 可形成SUMO链(polySUMOylation).polySUMOylation或多位点SUMO化修饰(mutiSUMOylation or mutiple monoSUMOylation)可以招募含有SIM基序(SUMO interacting motif)的蛋白, 介导蛋白-蛋白的非共价结合.SUMO化修饰除了影响蛋白-蛋白互作外, 还调控底物蛋白的亚细胞定位、活性或稳定性等.截至目前, 通过酵母双杂交(yeast two-hybrid)和质谱分析(mass spectrometry)等方法, 已鉴定出上千个可能的SUMO底物(Elrouby and Coupland, 2010; Miller et al., 2010; Rytz et al., 2018).但由于体内SUMO化修饰水平很低,且处于动态变化中(Sadanandom et al., 2015; Lin et al., 2016), 鉴定特定底物的SUMO化修饰较为困难.本文仅介绍检测植物蛋白SUMO化修饰的常用方法.若想了解更多关于SUMO化修饰的研究方法, 可参阅文献Rodriguez (2016). ...
SUMOylation of phytochrome-B negatively regulates light- induced signaling in Arabidopsis thaliana 2 2015
... SUMO1和SUMO2的N端含有多个赖氨酸, 可形成SUMO链(polySUMOylation).polySUMOylation或多位点SUMO化修饰(mutiSUMOylation or mutiple monoSUMOylation)可以招募含有SIM基序(SUMO interacting motif)的蛋白, 介导蛋白-蛋白的非共价结合.SUMO化修饰除了影响蛋白-蛋白互作外, 还调控底物蛋白的亚细胞定位、活性或稳定性等.截至目前, 通过酵母双杂交(yeast two-hybrid)和质谱分析(mass spectrometry)等方法, 已鉴定出上千个可能的SUMO底物(Elrouby and Coupland, 2010; Miller et al., 2010; Rytz et al., 2018).但由于体内SUMO化修饰水平很低,且处于动态变化中(Sadanandom et al., 2015; Lin et al., 2016), 鉴定特定底物的SUMO化修饰较为困难.本文仅介绍检测植物蛋白SUMO化修饰的常用方法.若想了解更多关于SUMO化修饰的研究方法, 可参阅文献Rodriguez (2016). ...
... (3) Western blot检测 用30 µL 2×SDS上样缓冲液重悬beads, 100°C煮沸5分钟, 瞬时离心, 混匀.1 000 ×g离心3分钟, 上样.Western blot, anti-SUMO1抗体仅在实验组(转基因材料)检测到目的条带上方间隔×11 kDa (SUMO1大约11 kDa)处有1条或多条条带, 说明目的蛋白在拟南芥中能被SUMO化修饰.如图3所示, Myc-protein X过表达材料, IP (anti-Myc)产物用anti-SUMO1抗体能在处理组(T)特异检测到SUMO化修饰的Myc-protein X, 但在对照组(C)未检测到SUMO化修饰的Myc-protein X, 这说明Myc-protein X仅在特定处理条件下才被SUMO化修饰.因此, 需结合表型, 选择植物特定发育阶段、特定生长条件或特定处理条件进行SUMO化修饰鉴定(Sadanandom et al., 2015; Lin et al., 2016). ...
Posttranslational modifications of the master transcriptional regulator NPR1 enable dynamic but tight control of plant immune responses 1 2015
... 拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用.SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007).SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015).拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21).其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018).SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
Genetic analysis of SUMOylation in Arabidopsis: conjugation of SUMO1 and SUMO2 to nuclear proteins is essential 2 2007
... 拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用.SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007).SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015).拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21).其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018).SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
... ; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
Arabidopsis PIAL1 and 2 promote SUMO chain formation as E4-type SUMO ligases and are involved in stress responses and sulfur metabolism 1 2014
... 拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用.SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007).SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015).拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21).其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018).SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
SUMO proteases: uncovering the roles of deSUMOylation in plants 1 2016
... 拟南芥(Arabidopsis thaliana)有8个SUMO编码基因, 即SUM1 (SUMO1)-SUM8, 其中SUM4、SUM6、SUM7和SUM8转录水平很低, 可能在特异发育阶段或组织中起作用.SUMO1和SUMO2的氨基酸序列相似性高达93%, sum1-1/sum2-1双突变体胚胎发育畸形, 不能结实(Saracco et al., 2007).SUMO3和SUMO5与SUMO1的氨基酸序列相似性仅为50%, 并且目前SUMO3和SUMO5的功能研究相对较少(Saleh et al., 2015).拟南芥SUMO激活酶E1含有大、小两个亚基, 小亚基有2个编码基因(即SAE1a和SAE1b), 大亚基只有1个编码基因SAE2; SUMO结合酶E2只有1个编码基因SCE1; SUMO连接酶E3有2个编码基因, 即SIZ1 (SAP (scaffold attachment factor, acinus, PIAS), and MIZ1 (Msx2-interacting zinc finger))和MMS21 (methyl methanesulfonate- sensitivity protein 21).其中, 绝大部分底物的SUMO化修饰由SIZ1介导(Rytz et al., 2018).SUMO化修饰过程中, 任何关键酶的缺失均会导致胚胎致死, 或者植株矮小和结实率低等异常发育表型, 说明SUMO化修饰在调控植物正常生长发育过程中起着重要作用(Catala et al., 2007; Saracco et al., 2007; 徐庞连等, 2008; Ishida et al., 2012; Rytz et al., 2018).此外, 还有2个催化SUMO链形成的酶PIAL1 (protein inhibitor of activated stat like 1)和PIAL2, 称为E4连接酶(Tomanov et al., 2014).它们与SIZ1和MMS21都含有保守的SP (SIZ-PIAS)-RING结构域.拟南芥含有多个SUMO蛋白酶, 分别为ESD4 (early in short days 4) (Murtas et al., 2003)、ELS1 (ESD4-like SUMO protease 1) (Hermkes et al., 2011)、OTS1 (overly tolerant to salt 1)、OTS2 (Conti et al., 2008)、ASP1 (Arabidopsis SUMO protease 1)/SPF1 (SUMO protease related to fertility 1)及SPF2 (Kong et al., 2017; Liu et al., 2017).目前, 通常认为SUMO蛋白酶具有底物特异性, 在调控底物SUMO化修饰水平方面发挥重要作用(Yates et al., 2016). ...
Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis 1 2007
 ,*, 金京波中国科学院植物研究所, 植物分子生理学重点实验室, 北京 100093
,*, 金京波中国科学院植物研究所, 植物分子生理学重点实验室, 北京 100093
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT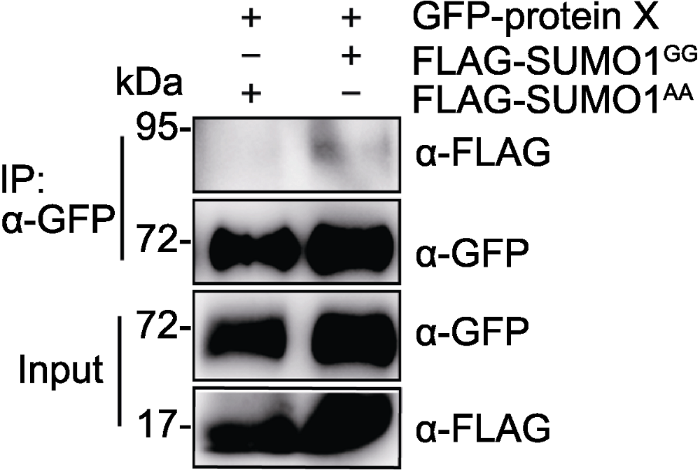 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT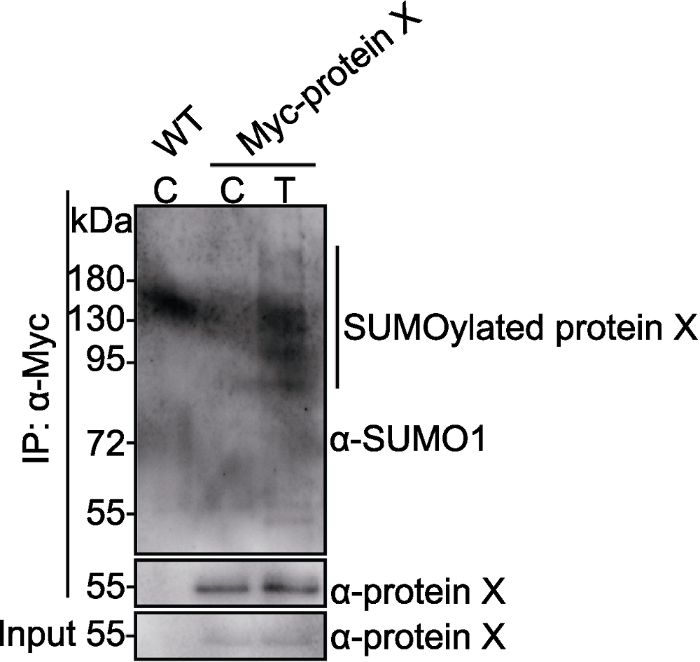 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT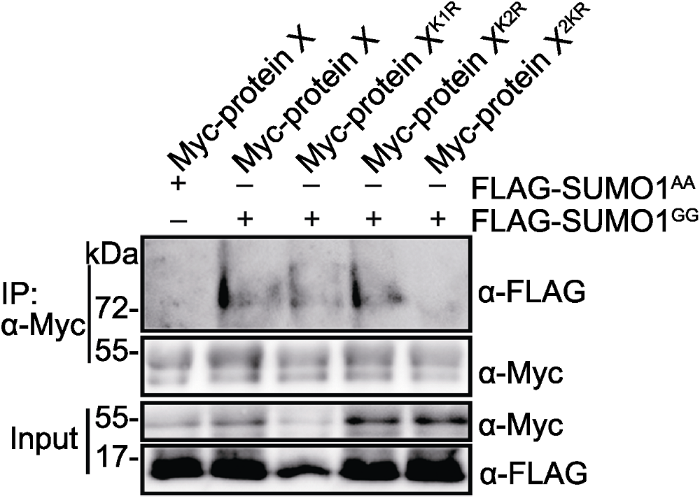 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}