Abstract Zinc finger-homeodomain (ZF-HD) genes encode plant-specific transcription factors that participate in the regulation of plant growth, development and abiotic stress response. In this study, a total of 62 ZF-HD genes were identified in Brassica napus. Among them, 83.9% lack introns, 93.5% of deduced proteins have the isoelectric point greater than 7. Most of the BnZF-HD proteins with nuclear localization signal were predicted to be more than 100 amino acid residues. Based on phylogenetic analysis, this gene family can be divided into six subgroups. In each subgroup, the number of ZF-HD genes in B. oleracea and B. rapa was the same or nearly so, while the number of ZF-HD genes in B. napus was close to or equal to the sum of ZF-HD genes in B. oleracea and B. rapa. The number and type of motifs in each subgroup were highly conserved. Collinear analysis indicated that the whole genome triplication allowed the ZF-HD gene family to expand in diploid ancestors, and allopolyploidization made the ZF-HD gene family to reexpand in B. napus. Ka/Ks analysis showed that the ZF-HD gene family has experienced purification selection in the evolution process. All BnZF-HD genes have light response elements, and two-thirds of them have MeJA, ABA and anaerobic inducible cis-acting elements. It is speculated that they may participate in related biological processes. This study laid a foundation for further exploring the molecular mechanism of this family in regulating plant growth and stress responses, and provided more reference for future research on the evolution of plant polyploidization. Keywords:Brassica napus;ZF-HD;evolution;collinear analysis;cis-acting elements
PDF (1433KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 宋敏, 张瑶, 王丽莹, 彭向永. 甘蓝型油菜ZF-HD基因家族的鉴定与系统进化分析. 植物学报, 2019, 54(6): 699-710 doi:10.11983/CBB19055 Song Min, Zhang Yao, Wang Liying, Peng Xiangyong. Genome-wide Identification and Phylogenetic Analysis of Zinc Finger Homeodomain Family Genes in Brassica napus. Chinese Bulletin of Botany, 2019, 54(6): 699-710 doi:10.11983/CBB19055
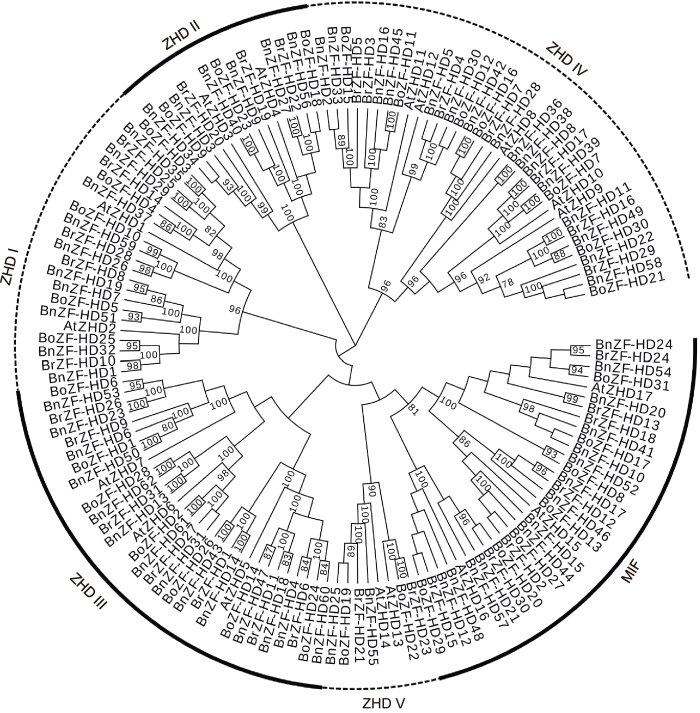
有趣的是, 有一类蛋白MIF (MINI ZINC FINGER)与ZF-HD蛋白的ZF结构域序列相似度高但缺乏HD结构域(Hu and Ma, 2006; Hu et al., 2008), 且是种子植物特有的蛋白。推测MIF蛋白与ZF-HD蛋白的进化关系有2种可能, 一是MIF蛋白由ZF-HD蛋白丢失HD结构域形成; 二是MIF获得HD结构域形成ZF-HD蛋白(Hu et al., 2008; Wang et al., 2016)。按照已发表的模式植物拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)的ZF-HD蛋白的划分方法, 本研究将MIF归入ZF-HD家族。
ZF-HD基因在植物多种发育和生理过程中发挥作用。研究发现ZF-HD基因家族只存在于陆生植物中(Wang et al., 2016), 这类基因可能在陆生植物祖先与单细胞藻类趋异后得到了进化。本研究在甘蓝型油菜中鉴定到62个ZF-HD基因, 参考其它植物中的分类(Hu et al., 2008), 发现MIF与ZF-HD蛋白相似度高, 虽然缺乏HD结构域, 也将14个MIF基因纳入ZF-HD基因家族的范畴。与拟南芥、白菜和甘蓝的ZF-HD蛋白聚类分析后, 分成6个亚群, MIF单独形成1个亚群。同一亚群的motif分布模式基本一致, 如MIF都具有相同的3个基序, 不同亚群之间motif分布差异较大。甘蓝型油菜ZF-HD基因大多缺乏内含子, 这与拟南芥、水稻、葡萄及白菜等植物中的研究结果一致(Hu et al., 2008; Wang et al., 2014, 2016)。
多倍化是物种进化的重要力量。比较基因组学研究显示, 大约2千万年前与拟南芥分离后, 芸薹属物种经历了基因组水平的三倍化(Chalhoub et al., 2014; Liu et al., 2014; Yang et al., 2016)。基于三倍化假说, 每个拟南芥的ZF-HD基因对应3个白菜或甘蓝的同源基因, 而实际上只有AtZHD1/5/17在白菜和甘蓝基因组各有3个同源基因, 其它AtZHD基因在白菜和甘蓝基因组中有1或2个拷贝(附录5), 表明白菜和甘蓝基因在芸薹属三倍化过程中发生了基因丢失。白菜与甘蓝的分化大约发生在400万年以前(Liu et al., 2014), 甘蓝型油菜大约在7 500-12 500年前由白菜和甘蓝天然杂交形成(Chalhoub et al., 2014)。染色体加倍以后, 理论上每个拟南芥ZF-HD基因在甘蓝型油菜中应该有6个同源基因, 且甘蓝型油菜ZF-HD基因等于白菜和甘蓝的同源基因数目之和。实际上除了AtZHD17在甘蓝型油菜有6个同源基因外, 其余AtZHD基因在甘蓝型油菜中的同源基因数目不超过4个, 说明ZF-HD基因在芸薹属三倍化后发生了丢失。推测丢失的原因可能有以下3个: (1) 芸薹属三倍化以后, 在二倍体白菜和甘蓝祖先中发生了基因丢失, 从而导致甘蓝型油菜中共线性同源基因丢失, 如AtZHD3和AtZHD14在白菜和甘蓝中各有1个共线性同源基因, 在油菜中有2个共线性同源基因; (2) 在异源四倍体化以后发生丢失, 如AtZHD1和AtZHD5在白菜和甘蓝中各有3个共线性同源基因, 在油菜中只有3或4个共线性同源基因; (3) 两个过程中都有丢失现象, 如AtZHD13在白菜和甘蓝中各有1个共线性同源基因, 但油菜中没有相应的共线性同源基因, 这可能与三倍化过程中发生大规模的基因丢失和染色体重排有关(Mun et al., 2009; 高虎虎等, 2017)。但总体来看, 62个BnZF-HD基因中有48个与拟南芥、白菜和甘蓝存在共线性关系, 从白菜和甘蓝祖先基因组各继承了24个, 表明甘蓝型油菜ZF-HD基因家族扩张的主要力量是异源四倍体化。
比较基因组学研究表明, 在白菜和甘蓝基因组的不同区域, 基因密度与表达水平存在差异, 可划分为LF (least fractionated)、MF1 (medium fractionated) 和MF2 (most fractionated) 3个亚基因组(subgeno- me) (Wang et al., 2011; Liu et al., 2014)。与拟南芥ZF-HD基因相比, 白菜、甘蓝及甘蓝型油菜中大多数保留下来的ZF-HD基因位于LF亚基因组, 这与整个基因组的保留趋势一致, 其次是MF2亚基因组, MF1亚基因组保留的ZF-HD基因最少, 而基因组整体水平上MF1亚基因组基因丢失的数目最多。
Appendix 1 Name, ID and various features of ZF-HD genes in Brassica napus
附录2 白菜、甘蓝和拟南芥ZF-HD基因
Appendix 2 The members of ZF-HD gene family identified in Brassica rapa, B. oleracea and Arabidopsis thaliana
附录3 基于最大似然(ML)法构建的甘蓝型油菜、拟南芥、白菜和甘蓝的ZF-HD基因系统进化树
Appendix 3 Phylogenetic tree of Brassica napus, Arabidopsis thaliana, B. rapa and B. oleracea ZF-HD genes constructed by the maximum likelihood (ML) method
附录4 甘蓝型油菜ZF-HD蛋白motif保守氨基酸序列
Appendix 4 Motif sequences identified of ZF-HD proteins in Brassica napus
附录5 ZF-HD基因在白菜、甘蓝、甘蓝型油菜和拟南芥共线性区的同源基因
Appendix 5 Homologous ZF-HD genes in collinear regions of Brassica rapa, B. oleracea, B. napus and A. thaliana genomes
附录6 甘蓝型油菜与拟南芥ZF-HD共线性基因的Ka/Ks比值
Appendix 6 The Ka/Ks ratios for orthologous ZF-HD genes between Brassica napus and Arabidopsis thaliana
附录7 甘蓝型油菜与白菜ZF-HD共线性基因的Ka/Ks比值
Appendix 7 The Ka/Ks ratios for orthologous ZF-HD genes between Brassica napus and B. rapa
AbdullahM, ChengX, CaoYP, SuXQ, ManzoorMA, GaoJS, CaiYP, LinY ( 2018). Zinc finger-homeodomain transcriptional factors (ZHDs) in upland cotton ( Gossypium hirsutum ): genome-wide identification and expression analysis in fiber development Front Genet 9, 357. [本文引用: 2]
ArielFD, ManavellaPA, DezarCA, ChanRL ( 2007). The true story of the HD-Zip family Trends Plant Sci 12, 419-426. [本文引用: 1]
BaileyTL, JohnsonJ, GrantCE, NobleWS ( 2015). The MEME suite Nucleic Acids Res 43, W39-W49. [本文引用: 1]
BatemanA, BirneyE, CerrutiL, DurbinR, EtwillerL, EddySR, Griffiths-JonesS, HoweKL, MarshallM, SonnhammerELL ( 2002). The Pfam protein families database Nucleic Acids Res 30, 276-280. [本文引用: 1]
BhattacharjeeA, GhangalR, GargR, JainM ( 2015). Genome-wide analysis of homeobox gene family in legumes: identification, gene duplication and expression profiling PLoS One 10, e0119198. [本文引用: 1]
BhattacharjeeA, JainM (2013). omeobox genes as potential candidates for crop improvement under abiotic stress In: Tuteja N, Singh Gill S, eds. Plant Acclimation to Environmental Stress. New York: Springer. pp. 163-176. [本文引用: 1]
ChenCJ, XiaR, ChenH, HeYH ( 2018). TBtools, a Toolkit for Biologists integrating various HTS-data handling tools with a user-friendly interface bioRxiv [2018-03-27]. . URL [本文引用: 3]
ChengF, LiuSY, WuJ, FangL, SunSL, LiuB, LiPX, HuaW, WangXW ( 2011). BRAD, the genetics and genomics database for Brassica plants BMC Plant Biol 11, 136. [本文引用: 1]
ChengF, WuJ, FangL, WangXW ( 2012). Syntenic gene analysis between Brassica rapa and other Brassicaceae species Front Plant Sci 3, 198. [本文引用: 1]
DengX, PhillipsJ, MeijerAH, SalaminiF, BartelsD ( 2002). Characterization of five novel dehydration-responsive homeodomain leucine zipper genes from the resurrection plant Craterostigma plantagineum Plant Mol Biol 49, 601-610. [本文引用: 1]
FinnRD, CoggillP, EberhardtRY, EddySR, MistryJ, MitchellAL, PotterSC, PuntaM, QureshiM, Sangrador-VegasA, SalazarGA, TateJ, BatemanA ( 2016). The Pfam protein families database: towards a more sustainable future Nucleic Acids Res 44, D279-D285. [本文引用: 1]
HeZL, ZhangHK, GaoSH, LercherMJ, ChenWH, HuSN ( 2016). Evolview v2: an online visualization and management tool for customized and annotated phylogenetic trees Nucleic Acids Res 44, W236-W241. [本文引用: 1]
HuW, DePamphilisCW, MaH ( 2008). Phylogenetic analysis of the plant-specific Zinc finger-Homeobox and Mini zinc finger gene families J Integr Plant Biol 50, 1031-1045. [本文引用: 8]
HuW, MaH ( 2006). Characterization of a novel putative zinc finger gene MIF1: involvement in multiple hormonal regulation of Arabidopsis development Plant J 45, 399-422. [本文引用: 1]
JainM, TyagiAK, KhuranaJP ( 2008). Genome-wide identification, classification, evolutionary expansion and expression analyses of homeobox genes in rice FEBS J 275, 2845-2861. [本文引用: 1]
JinJP, TianF, YangDC, MengYQ, KongL, LuoJC, GaoG ( 2017a). PlantTFDB 4.0: toward a central hub for transcription factors and regulatory interactions in plants Nucleic Acids Res 45, D1040-D1045.
JinXL, RenJ, NevoE, YinXG, SunDF, PengJH ( 2017b). Divergent evolutionary patterns of NAC transcription factors are associated with diversification and gene duplications in angiosperm Front Plant Sci 8, 1156. [本文引用: 1]
J?rgensenJE, Gr?nlundM, PallisgaardN, LarsenK, MarckerKA, JensenE? ( 1999). A new class of plant homeobox genes is expressed in specific regions of determinate symbiotic root nodules Plant Mol Biol 40, 65-77. [本文引用: 1]
KhadizaK, NathUK, RobinAHK, ParkJI, LeeDJ, KimMB, KimCK, LimLB, NouIS, ChungMY ( 2017). Genome-wide analysis and expression profiling of zinc finger homeodomain ( ZHD ) family genes reveal likely roles in organ development and stress responses in tomato BMC Genomics 18, 695. [本文引用: 2]
KrishnaSS, MajumdarI, GrishinNV ( 2003). SURVEY AND SUMMARY: structural classification of zinc fingers Nucleic Acids Res 31, 532-550. [本文引用: 2]
KrzywinskiM, ScheinJ, Birol?, ConnorsJ, GascoyneR, HorsmanD, JonesSJ, MarraMA ( 2009). Circos: an information aesthetic for comparative genomics Genome Res 19, 1639-1645. [本文引用: 1]
MackayJP, CrossleyM ( 1998). Zinc fingers are sticking together Trends Biochem Sci 23, 1-4. [本文引用: 2]
MunJH, KwonSJ, YangTJ, SeolYJ, JinM, KimJA, LimMH, KimJS, BaekS, ChoiBS, YuHJ, KimDS, KimN, LimKB, LeeSI, HahnJH, LimYP, BancroftI, ParkBS ( 2009). Genome-wide comparative analysis of the Brassica rapa gene space reveals genome shrinkage and differential loss of duplicated genes after whole genome triplication Genome Biol 10, 211. [本文引用: 1]
NamJ, NeiM ( 2005). Evolutionary change of the numbers of homeobox genes in bilateral animals Mol Biol Evol 22, 2386-2394. [本文引用: 1]
NekrutenkoA, MakovaKD, LiWH ( 2002). The KA/KS ratio test for assessing the protein-coding potential of genomic regions: an empirical and simulation study Genome Res 12, 198-202. [本文引用: 1]
ParkHC, KimML, LeeSM, BahkJD, YunDJ, LimCO, HongJC, LeeSY, ChoMJ, ChungWS ( 2007). Pathogen-induced binding of the soybean zinc finger homeodomain proteins GmZF-HD1 and GmZF-HD2 to two repeats of ATTA homeodomain binding site in the calmodulin isoform 4 (GmCaM4) promoter Nucleic Acids Res 35, 3612-3623. [本文引用: 1]
SchultzJ, MilpetzF, BorkP, PontingCP ( 1998). SMART, a simple modular architecture research tool: identification of signaling domains Proc Natl Acad Sci USA 95, 5857-5864. [本文引用: 1]
TanQKG, IrishVF ( 2006). The Arabidopsis zinc finger-homeodomain genes encode proteins with unique biochemical properties that are coordinately expressed during floral development Plant Physiol 140, 1095-1108. [本文引用: 3]
TranLSP, NakashimaK, SakumaY, OsakabeY, QinF, SimpsonSD, MaruyamaK, FujitaY, ShinozakiK, Yamaguchi-ShinozakiK ( 2007). Co-expression of the stress-inducible zinc finger homeodomain ZFHD1 and NAC transcription factors enhances expression of the ERD1 gene in Arabidopsis Plant J 49, 46-63. [本文引用: 2]
WaltherD, BrunnemannR, SelbigJ ( 2007). The regulatory code for transcriptional response diversity and its relation to genome structural properties in A. thaliana PLoS Genet 3, e11. [本文引用: 1]
WangH, YinXJ, LiXQ, WangL, ZhengY, XuXZ, ZhangYC, WangXP ( 2014). Genome-wide identification, evolution and expression analysis of the grape ( Vitis vinifera L.) zinc finger-homeodomain gene family Int J Mol Sci 15, 5730-5748. [本文引用: 4]
WangW, WuP, LiY, HouX ( 2016). Genome-wide analysis and expression patterns of ZF-HD transcription factors under different developmental tissues and abiotic stresses in Chinese cabbage Mol Gen Genomics 291, 1451-1464. [本文引用: 6]
Windh?velA, HeinI, DabrowaR, StockhausJ ( 2001). Characterization of a novel class of plant homeodomain proteins that bind to the C4 phosphoenolpyruvate carboxylase gene of Flaveria trinervia Plant Mol Biol 45, 201-214. [本文引用: 4]
Yamaguchi-ShinozakiK, ShinozakiK ( 2005). Organization of cis -acting regulatory elements in osmotic-and cold- stress-responsive promoters Trends Plant Sci 10, 88-94. [本文引用: 1]
... 多倍化是物种进化的重要力量.比较基因组学研究显示, 大约2千万年前与拟南芥分离后, 芸薹属物种经历了基因组水平的三倍化(Chalhoub et al., 2014; Liu et al., 2014; Yang et al., 2016).基于三倍化假说, 每个拟南芥的ZF-HD基因对应3个白菜或甘蓝的同源基因, 而实际上只有AtZHD1/5/17在白菜和甘蓝基因组各有3个同源基因, 其它AtZHD基因在白菜和甘蓝基因组中有1或2个拷贝(附录5), 表明白菜和甘蓝基因在芸薹属三倍化过程中发生了基因丢失.白菜与甘蓝的分化大约发生在400万年以前(Liu et al., 2014), 甘蓝型油菜大约在7 500-12 500年前由白菜和甘蓝天然杂交形成(Chalhoub et al., 2014).染色体加倍以后, 理论上每个拟南芥ZF-HD基因在甘蓝型油菜中应该有6个同源基因, 且甘蓝型油菜ZF-HD基因等于白菜和甘蓝的同源基因数目之和.实际上除了AtZHD17在甘蓝型油菜有6个同源基因外, 其余AtZHD基因在甘蓝型油菜中的同源基因数目不超过4个, 说明ZF-HD基因在芸薹属三倍化后发生了丢失.推测丢失的原因可能有以下3个: (1) 芸薹属三倍化以后, 在二倍体白菜和甘蓝祖先中发生了基因丢失, 从而导致甘蓝型油菜中共线性同源基因丢失, 如AtZHD3和AtZHD14在白菜和甘蓝中各有1个共线性同源基因, 在油菜中有2个共线性同源基因; (2) 在异源四倍体化以后发生丢失, 如AtZHD1和AtZHD5在白菜和甘蓝中各有3个共线性同源基因, 在油菜中只有3或4个共线性同源基因; (3) 两个过程中都有丢失现象, 如AtZHD13在白菜和甘蓝中各有1个共线性同源基因, 但油菜中没有相应的共线性同源基因, 这可能与三倍化过程中发生大规模的基因丢失和染色体重排有关(Mun et al., 2009; 高虎虎等, 2017).但总体来看, 62个BnZF-HD基因中有48个与拟南芥、白菜和甘蓝存在共线性关系, 从白菜和甘蓝祖先基因组各继承了24个, 表明甘蓝型油菜ZF-HD基因家族扩张的主要力量是异源四倍体化. ...
Zinc finger-homeodomain transcriptional factors (ZHDs) in upland cotton ( Gossypium hirsutum ): genome-wide identification and expression analysis in fiber development 2 2018
... 目前, 国内外****已经对水稻、大豆(Glycine max)、拟南芥、小麦(Triticum aestivum)、葡萄(Vitis vinifera)、白菜(Brassica rapa)、番茄(Solanum lycopersicum)和陆地棉(Gossypium hirsutum)等物种中的ZF-HD基因家族进行了鉴定(Jørgensen et al., 1999; Windhövel et al., 2001; Deng et al., 2002; Tan and Irish, 2006; Jain et al., 2008; Bhattacharjee and Jain, 2013; Wang et al., 2014, 2016; Khadiza et al., 2017; Abdullah et al., 2018).已有研究表明, ZF-HD转录因子参与许多生长发育过程和非生物胁迫响应.拟南芥ZF-HD基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
... 基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
The true story of the HD-Zip family 1 2007
... ZF-HD转录因子(zinc finger homeodomain, ZHD)是植物特有的转录因子(Windhovel et al., 2001; Tan and Irish, 2006), 其在C4植物黄顶菊(Flaveria trinervia)中首次被报道, 是磷酸烯醇式丙酮酸羧化酶(PEPCase)编码基因的潜在调控因子(Windhovel et al., 2001).ZF-HD转录因子的命名是因其氨基酸序列中有保守的HD结构域(homeodomain)和C2H2型锌指结构(C2H2-type zinc finger motif).HD结构域是含60个氨基酸的DNA结合域, 大多数具HD结构域的蛋白同时具有其它类型的结构域或基序, 可分为HD- ZIP、ZF-HD、WOX、Bell type HD、PHD和KNOX六个家族(Ariel et al., 2007).含HD的蛋白质广泛存在于动植物体内, 在发育过程中发挥重要作用(Nam and Nei, 2005; Bhattacharjee et al., 2015).锌指结构是由多个保守的氨基酸残基围绕核心的锌离子组成, 形成相对独立的指状四面体结构(Krishna et al., 2003).根据其Cys和His残基的不同组合分成不同类型, 如C3H、C2H2、C2C2和C4C6, 其中以C2H2型最为常见.锌指可以识别和结合DNA、RNA、DNA-RNA双链分子或蛋白质(Mackay and Crossley, 1998; Takatsuji, 1999; Krishna et al., 2003), 在转录和翻译水平调控基因表达, 在植物的应激反应和防御激活中发挥重要作用(Mackay, 1998).在ZF-HD蛋白中, HD结构域能够结合DNA, ZF结构可以增强由HD结构域介导的蛋白质与DNA的相互作用(Windhovel et al., 2001; Hu et al., 2008). ...
The MEME suite 1 2015
... 利用Mapchart软件绘制基因在染色体上的分布图.同时利用在线软件MEME (http://meme-suite.org/ tools/meme) (Bailey et al., 2015)对甘蓝型油菜ZF-HD蛋白的motif进行预测和分析.MEME参数设置: motif数量为15, motif长度为6-60.下载mast.xml结果文件, 用TBtools软件的Redraw motif pattern工具展示motif的分布(Chen et al., 2018). ...
The Pfam protein families database 1 2002
... 通过3种途径搜索甘蓝型油菜和甘蓝的ZF-HD基因.(1) 从植物转录因子网站PlantTFDB (http://planttfdb. cbi.pku.edu.cn/, V4.0) (Jin et al., 2017b)分别下载甘蓝型油菜和甘蓝全部ZF-HD家族成员蛋白序列; (2) 从Pfam数据库(http://pfam.xfam.org/) (Bateman et al., 2002; Finn et al., 2016)下载ZF-HD基因的PF04770结构域种子数据stockholm文件, 使用hmmsearch工具分别搜索甘蓝型油菜和甘蓝的蛋白质数据库, 参数值为1.0; (3) 以拟南芥ZF-HD蛋白序列比对文件, 进行本地Blast P检索, e值设为1e-3; 整理获取的序列, 去掉重复序列, 然后利用在线工具CDD (http://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)和Smart (Schultz et al., 1998) (http://smart. emblheidelberg.de/)检测蛋白序列的结构域, 获得候选基因, 用于后续分析. ...
Genome-wide analysis of homeobox gene family in legumes: identification, gene duplication and expression profiling 1 2015
... ZF-HD转录因子(zinc finger homeodomain, ZHD)是植物特有的转录因子(Windhovel et al., 2001; Tan and Irish, 2006), 其在C4植物黄顶菊(Flaveria trinervia)中首次被报道, 是磷酸烯醇式丙酮酸羧化酶(PEPCase)编码基因的潜在调控因子(Windhovel et al., 2001).ZF-HD转录因子的命名是因其氨基酸序列中有保守的HD结构域(homeodomain)和C2H2型锌指结构(C2H2-type zinc finger motif).HD结构域是含60个氨基酸的DNA结合域, 大多数具HD结构域的蛋白同时具有其它类型的结构域或基序, 可分为HD- ZIP、ZF-HD、WOX、Bell type HD、PHD和KNOX六个家族(Ariel et al., 2007).含HD的蛋白质广泛存在于动植物体内, 在发育过程中发挥重要作用(Nam and Nei, 2005; Bhattacharjee et al., 2015).锌指结构是由多个保守的氨基酸残基围绕核心的锌离子组成, 形成相对独立的指状四面体结构(Krishna et al., 2003).根据其Cys和His残基的不同组合分成不同类型, 如C3H、C2H2、C2C2和C4C6, 其中以C2H2型最为常见.锌指可以识别和结合DNA、RNA、DNA-RNA双链分子或蛋白质(Mackay and Crossley, 1998; Takatsuji, 1999; Krishna et al., 2003), 在转录和翻译水平调控基因表达, 在植物的应激反应和防御激活中发挥重要作用(Mackay, 1998).在ZF-HD蛋白中, HD结构域能够结合DNA, ZF结构可以增强由HD结构域介导的蛋白质与DNA的相互作用(Windhovel et al., 2001; Hu et al., 2008). ...
omeobox genes as potential candidates for crop improvement under abiotic stress 1 2013
... 目前, 国内外****已经对水稻、大豆(Glycine max)、拟南芥、小麦(Triticum aestivum)、葡萄(Vitis vinifera)、白菜(Brassica rapa)、番茄(Solanum lycopersicum)和陆地棉(Gossypium hirsutum)等物种中的ZF-HD基因家族进行了鉴定(Jørgensen et al., 1999; Windhövel et al., 2001; Deng et al., 2002; Tan and Irish, 2006; Jain et al., 2008; Bhattacharjee and Jain, 2013; Wang et al., 2014, 2016; Khadiza et al., 2017; Abdullah et al., 2018).已有研究表明, ZF-HD转录因子参与许多生长发育过程和非生物胁迫响应.拟南芥ZF-HD基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome 5 2014
Syntenic gene analysis between Brassica rapa and other Brassicaceae species 1 2012
... 通过BRAD (http://brassicadb.org/brad/searchSynteny.php) (Cheng et al., 2012)在线检索获取拟南芥ZHD基因在甘蓝型油菜、白菜和甘蓝中的共线性基因及其在染色体上的位置.用Circos (Krzywinski et al., 2009)软件展示这些共线性基因的关系.用TBtools软件计算共线性基因之间的Ka/Ks值(Chen et al., 2018), 估算选择压力. ...
Characterization of five novel dehydration-responsive homeodomain leucine zipper genes from the resurrection plant Craterostigma plantagineum 1 2002
... 目前, 国内外****已经对水稻、大豆(Glycine max)、拟南芥、小麦(Triticum aestivum)、葡萄(Vitis vinifera)、白菜(Brassica rapa)、番茄(Solanum lycopersicum)和陆地棉(Gossypium hirsutum)等物种中的ZF-HD基因家族进行了鉴定(Jørgensen et al., 1999; Windhövel et al., 2001; Deng et al., 2002; Tan and Irish, 2006; Jain et al., 2008; Bhattacharjee and Jain, 2013; Wang et al., 2014, 2016; Khadiza et al., 2017; Abdullah et al., 2018).已有研究表明, ZF-HD转录因子参与许多生长发育过程和非生物胁迫响应.拟南芥ZF-HD基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
The Pfam protein families database: towards a more sustainable future 1 2016
... 通过3种途径搜索甘蓝型油菜和甘蓝的ZF-HD基因.(1) 从植物转录因子网站PlantTFDB (http://planttfdb. cbi.pku.edu.cn/, V4.0) (Jin et al., 2017b)分别下载甘蓝型油菜和甘蓝全部ZF-HD家族成员蛋白序列; (2) 从Pfam数据库(http://pfam.xfam.org/) (Bateman et al., 2002; Finn et al., 2016)下载ZF-HD基因的PF04770结构域种子数据stockholm文件, 使用hmmsearch工具分别搜索甘蓝型油菜和甘蓝的蛋白质数据库, 参数值为1.0; (3) 以拟南芥ZF-HD蛋白序列比对文件, 进行本地Blast P检索, e值设为1e-3; 整理获取的序列, 去掉重复序列, 然后利用在线工具CDD (http://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)和Smart (Schultz et al., 1998) (http://smart. emblheidelberg.de/)检测蛋白序列的结构域, 获得候选基因, 用于后续分析. ...
Evolview v2: an online visualization and management tool for customized and annotated phylogenetic trees 1 2016
Phylogenetic analysis of the plant-specific Zinc finger-Homeobox and Mini zinc finger gene families 8 2008
... ZF-HD转录因子(zinc finger homeodomain, ZHD)是植物特有的转录因子(Windhovel et al., 2001; Tan and Irish, 2006), 其在C4植物黄顶菊(Flaveria trinervia)中首次被报道, 是磷酸烯醇式丙酮酸羧化酶(PEPCase)编码基因的潜在调控因子(Windhovel et al., 2001).ZF-HD转录因子的命名是因其氨基酸序列中有保守的HD结构域(homeodomain)和C2H2型锌指结构(C2H2-type zinc finger motif).HD结构域是含60个氨基酸的DNA结合域, 大多数具HD结构域的蛋白同时具有其它类型的结构域或基序, 可分为HD- ZIP、ZF-HD、WOX、Bell type HD、PHD和KNOX六个家族(Ariel et al., 2007).含HD的蛋白质广泛存在于动植物体内, 在发育过程中发挥重要作用(Nam and Nei, 2005; Bhattacharjee et al., 2015).锌指结构是由多个保守的氨基酸残基围绕核心的锌离子组成, 形成相对独立的指状四面体结构(Krishna et al., 2003).根据其Cys和His残基的不同组合分成不同类型, 如C3H、C2H2、C2C2和C4C6, 其中以C2H2型最为常见.锌指可以识别和结合DNA、RNA、DNA-RNA双链分子或蛋白质(Mackay and Crossley, 1998; Takatsuji, 1999; Krishna et al., 2003), 在转录和翻译水平调控基因表达, 在植物的应激反应和防御激活中发挥重要作用(Mackay, 1998).在ZF-HD蛋白中, HD结构域能够结合DNA, ZF结构可以增强由HD结构域介导的蛋白质与DNA的相互作用(Windhovel et al., 2001; Hu et al., 2008). ...
... 有趣的是, 有一类蛋白MIF (MINI ZINC FINGER)与ZF-HD蛋白的ZF结构域序列相似度高但缺乏HD结构域(Hu and Ma, 2006; Hu et al., 2008), 且是种子植物特有的蛋白.推测MIF蛋白与ZF-HD蛋白的进化关系有2种可能, 一是MIF蛋白由ZF-HD蛋白丢失HD结构域形成; 二是MIF获得HD结构域形成ZF-HD蛋白(Hu et al., 2008; Wang et al., 2016).按照已发表的模式植物拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)的ZF-HD蛋白的划分方法, 本研究将MIF归入ZF-HD家族. ...
... ), 且是种子植物特有的蛋白.推测MIF蛋白与ZF-HD蛋白的进化关系有2种可能, 一是MIF蛋白由ZF-HD蛋白丢失HD结构域形成; 二是MIF获得HD结构域形成ZF-HD蛋白(Hu et al., 2008; Wang et al., 2016).按照已发表的模式植物拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)的ZF-HD蛋白的划分方法, 本研究将MIF归入ZF-HD家族. ...
... 目前, 国内外****已经对水稻、大豆(Glycine max)、拟南芥、小麦(Triticum aestivum)、葡萄(Vitis vinifera)、白菜(Brassica rapa)、番茄(Solanum lycopersicum)和陆地棉(Gossypium hirsutum)等物种中的ZF-HD基因家族进行了鉴定(Jørgensen et al., 1999; Windhövel et al., 2001; Deng et al., 2002; Tan and Irish, 2006; Jain et al., 2008; Bhattacharjee and Jain, 2013; Wang et al., 2014, 2016; Khadiza et al., 2017; Abdullah et al., 2018).已有研究表明, ZF-HD转录因子参与许多生长发育过程和非生物胁迫响应.拟南芥ZF-HD基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
... ; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
... ZF-HD基因在植物多种发育和生理过程中发挥作用.研究发现ZF-HD基因家族只存在于陆生植物中(Wang et al., 2016), 这类基因可能在陆生植物祖先与单细胞藻类趋异后得到了进化.本研究在甘蓝型油菜中鉴定到62个ZF-HD基因, 参考其它植物中的分类(Hu et al., 2008), 发现MIF与ZF-HD蛋白相似度高, 虽然缺乏HD结构域, 也将14个MIF基因纳入ZF-HD基因家族的范畴.与拟南芥、白菜和甘蓝的ZF-HD蛋白聚类分析后, 分成6个亚群, MIF单独形成1个亚群.同一亚群的motif分布模式基本一致, 如MIF都具有相同的3个基序, 不同亚群之间motif分布差异较大.甘蓝型油菜ZF-HD基因大多缺乏内含子, 这与拟南芥、水稻、葡萄及白菜等植物中的研究结果一致(Hu et al., 2008; Wang et al., 2014, 2016). ...
... 基因大多缺乏内含子, 这与拟南芥、水稻、葡萄及白菜等植物中的研究结果一致(Hu et al., 2008; Wang et al., 2014, 2016). ...
Characterization of a novel putative zinc finger gene MIF1: involvement in multiple hormonal regulation of Arabidopsis development 1 2006
... 有趣的是, 有一类蛋白MIF (MINI ZINC FINGER)与ZF-HD蛋白的ZF结构域序列相似度高但缺乏HD结构域(Hu and Ma, 2006; Hu et al., 2008), 且是种子植物特有的蛋白.推测MIF蛋白与ZF-HD蛋白的进化关系有2种可能, 一是MIF蛋白由ZF-HD蛋白丢失HD结构域形成; 二是MIF获得HD结构域形成ZF-HD蛋白(Hu et al., 2008; Wang et al., 2016).按照已发表的模式植物拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)的ZF-HD蛋白的划分方法, 本研究将MIF归入ZF-HD家族. ...
Genome-wide identification, classification, evolutionary expansion and expression analyses of homeobox genes in rice 1 2008
... 目前, 国内外****已经对水稻、大豆(Glycine max)、拟南芥、小麦(Triticum aestivum)、葡萄(Vitis vinifera)、白菜(Brassica rapa)、番茄(Solanum lycopersicum)和陆地棉(Gossypium hirsutum)等物种中的ZF-HD基因家族进行了鉴定(Jørgensen et al., 1999; Windhövel et al., 2001; Deng et al., 2002; Tan and Irish, 2006; Jain et al., 2008; Bhattacharjee and Jain, 2013; Wang et al., 2014, 2016; Khadiza et al., 2017; Abdullah et al., 2018).已有研究表明, ZF-HD转录因子参与许多生长发育过程和非生物胁迫响应.拟南芥ZF-HD基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
PlantTFDB 4.0: toward a central hub for transcription factors and regulatory interactions in plants 0 2017
Divergent evolutionary patterns of NAC transcription factors are associated with diversification and gene duplications in angiosperm 1 2017
... 通过3种途径搜索甘蓝型油菜和甘蓝的ZF-HD基因.(1) 从植物转录因子网站PlantTFDB (http://planttfdb. cbi.pku.edu.cn/, V4.0) (Jin et al., 2017b)分别下载甘蓝型油菜和甘蓝全部ZF-HD家族成员蛋白序列; (2) 从Pfam数据库(http://pfam.xfam.org/) (Bateman et al., 2002; Finn et al., 2016)下载ZF-HD基因的PF04770结构域种子数据stockholm文件, 使用hmmsearch工具分别搜索甘蓝型油菜和甘蓝的蛋白质数据库, 参数值为1.0; (3) 以拟南芥ZF-HD蛋白序列比对文件, 进行本地Blast P检索, e值设为1e-3; 整理获取的序列, 去掉重复序列, 然后利用在线工具CDD (http://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)和Smart (Schultz et al., 1998) (http://smart. emblheidelberg.de/)检测蛋白序列的结构域, 获得候选基因, 用于后续分析. ...
A new class of plant homeobox genes is expressed in specific regions of determinate symbiotic root nodules 1 1999
... 目前, 国内外****已经对水稻、大豆(Glycine max)、拟南芥、小麦(Triticum aestivum)、葡萄(Vitis vinifera)、白菜(Brassica rapa)、番茄(Solanum lycopersicum)和陆地棉(Gossypium hirsutum)等物种中的ZF-HD基因家族进行了鉴定(Jørgensen et al., 1999; Windhövel et al., 2001; Deng et al., 2002; Tan and Irish, 2006; Jain et al., 2008; Bhattacharjee and Jain, 2013; Wang et al., 2014, 2016; Khadiza et al., 2017; Abdullah et al., 2018).已有研究表明, ZF-HD转录因子参与许多生长发育过程和非生物胁迫响应.拟南芥ZF-HD基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
Genome-wide analysis and expression profiling of zinc finger homeodomain ( ZHD ) family genes reveal likely roles in organ development and stress responses in tomato 2 2017
... 目前, 国内外****已经对水稻、大豆(Glycine max)、拟南芥、小麦(Triticum aestivum)、葡萄(Vitis vinifera)、白菜(Brassica rapa)、番茄(Solanum lycopersicum)和陆地棉(Gossypium hirsutum)等物种中的ZF-HD基因家族进行了鉴定(Jørgensen et al., 1999; Windhövel et al., 2001; Deng et al., 2002; Tan and Irish, 2006; Jain et al., 2008; Bhattacharjee and Jain, 2013; Wang et al., 2014, 2016; Khadiza et al., 2017; Abdullah et al., 2018).已有研究表明, ZF-HD转录因子参与许多生长发育过程和非生物胁迫响应.拟南芥ZF-HD基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
... 基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
SURVEY AND SUMMARY: structural classification of zinc fingers 2 2003
... ZF-HD转录因子(zinc finger homeodomain, ZHD)是植物特有的转录因子(Windhovel et al., 2001; Tan and Irish, 2006), 其在C4植物黄顶菊(Flaveria trinervia)中首次被报道, 是磷酸烯醇式丙酮酸羧化酶(PEPCase)编码基因的潜在调控因子(Windhovel et al., 2001).ZF-HD转录因子的命名是因其氨基酸序列中有保守的HD结构域(homeodomain)和C2H2型锌指结构(C2H2-type zinc finger motif).HD结构域是含60个氨基酸的DNA结合域, 大多数具HD结构域的蛋白同时具有其它类型的结构域或基序, 可分为HD- ZIP、ZF-HD、WOX、Bell type HD、PHD和KNOX六个家族(Ariel et al., 2007).含HD的蛋白质广泛存在于动植物体内, 在发育过程中发挥重要作用(Nam and Nei, 2005; Bhattacharjee et al., 2015).锌指结构是由多个保守的氨基酸残基围绕核心的锌离子组成, 形成相对独立的指状四面体结构(Krishna et al., 2003).根据其Cys和His残基的不同组合分成不同类型, 如C3H、C2H2、C2C2和C4C6, 其中以C2H2型最为常见.锌指可以识别和结合DNA、RNA、DNA-RNA双链分子或蛋白质(Mackay and Crossley, 1998; Takatsuji, 1999; Krishna et al., 2003), 在转录和翻译水平调控基因表达, 在植物的应激反应和防御激活中发挥重要作用(Mackay, 1998).在ZF-HD蛋白中, HD结构域能够结合DNA, ZF结构可以增强由HD结构域介导的蛋白质与DNA的相互作用(Windhovel et al., 2001; Hu et al., 2008). ...
... ; Krishna et al., 2003), 在转录和翻译水平调控基因表达, 在植物的应激反应和防御激活中发挥重要作用(Mackay, 1998).在ZF-HD蛋白中, HD结构域能够结合DNA, ZF结构可以增强由HD结构域介导的蛋白质与DNA的相互作用(Windhovel et al., 2001; Hu et al., 2008). ...
Circos: an information aesthetic for comparative genomics 1 2009
... 通过BRAD (http://brassicadb.org/brad/searchSynteny.php) (Cheng et al., 2012)在线检索获取拟南芥ZHD基因在甘蓝型油菜、白菜和甘蓝中的共线性基因及其在染色体上的位置.用Circos (Krzywinski et al., 2009)软件展示这些共线性基因的关系.用TBtools软件计算共线性基因之间的Ka/Ks值(Chen et al., 2018), 估算选择压力. ...
MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets 1 2016
... 比较基因组学研究表明, 在白菜和甘蓝基因组的不同区域, 基因密度与表达水平存在差异, 可划分为LF (least fractionated)、MF1 (medium fractionated) 和MF2 (most fractionated) 3个亚基因组(subgeno- me) (Wang et al., 2011; Liu et al., 2014).与拟南芥ZF-HD基因相比, 白菜、甘蓝及甘蓝型油菜中大多数保留下来的ZF-HD基因位于LF亚基因组, 这与整个基因组的保留趋势一致, 其次是MF2亚基因组, MF1亚基因组保留的ZF-HD基因最少, 而基因组整体水平上MF1亚基因组基因丢失的数目最多. ...
Zinc fingers are sticking together 2 1998
... ZF-HD转录因子(zinc finger homeodomain, ZHD)是植物特有的转录因子(Windhovel et al., 2001; Tan and Irish, 2006), 其在C4植物黄顶菊(Flaveria trinervia)中首次被报道, 是磷酸烯醇式丙酮酸羧化酶(PEPCase)编码基因的潜在调控因子(Windhovel et al., 2001).ZF-HD转录因子的命名是因其氨基酸序列中有保守的HD结构域(homeodomain)和C2H2型锌指结构(C2H2-type zinc finger motif).HD结构域是含60个氨基酸的DNA结合域, 大多数具HD结构域的蛋白同时具有其它类型的结构域或基序, 可分为HD- ZIP、ZF-HD、WOX、Bell type HD、PHD和KNOX六个家族(Ariel et al., 2007).含HD的蛋白质广泛存在于动植物体内, 在发育过程中发挥重要作用(Nam and Nei, 2005; Bhattacharjee et al., 2015).锌指结构是由多个保守的氨基酸残基围绕核心的锌离子组成, 形成相对独立的指状四面体结构(Krishna et al., 2003).根据其Cys和His残基的不同组合分成不同类型, 如C3H、C2H2、C2C2和C4C6, 其中以C2H2型最为常见.锌指可以识别和结合DNA、RNA、DNA-RNA双链分子或蛋白质(Mackay and Crossley, 1998; Takatsuji, 1999; Krishna et al., 2003), 在转录和翻译水平调控基因表达, 在植物的应激反应和防御激活中发挥重要作用(Mackay, 1998).在ZF-HD蛋白中, HD结构域能够结合DNA, ZF结构可以增强由HD结构域介导的蛋白质与DNA的相互作用(Windhovel et al., 2001; Hu et al., 2008). ...
... ), 在转录和翻译水平调控基因表达, 在植物的应激反应和防御激活中发挥重要作用(Mackay, 1998).在ZF-HD蛋白中, HD结构域能够结合DNA, ZF结构可以增强由HD结构域介导的蛋白质与DNA的相互作用(Windhovel et al., 2001; Hu et al., 2008). ...
Genome-wide comparative analysis of the Brassica rapa gene space reveals genome shrinkage and differential loss of duplicated genes after whole genome triplication 1 2009
... 多倍化是物种进化的重要力量.比较基因组学研究显示, 大约2千万年前与拟南芥分离后, 芸薹属物种经历了基因组水平的三倍化(Chalhoub et al., 2014; Liu et al., 2014; Yang et al., 2016).基于三倍化假说, 每个拟南芥的ZF-HD基因对应3个白菜或甘蓝的同源基因, 而实际上只有AtZHD1/5/17在白菜和甘蓝基因组各有3个同源基因, 其它AtZHD基因在白菜和甘蓝基因组中有1或2个拷贝(附录5), 表明白菜和甘蓝基因在芸薹属三倍化过程中发生了基因丢失.白菜与甘蓝的分化大约发生在400万年以前(Liu et al., 2014), 甘蓝型油菜大约在7 500-12 500年前由白菜和甘蓝天然杂交形成(Chalhoub et al., 2014).染色体加倍以后, 理论上每个拟南芥ZF-HD基因在甘蓝型油菜中应该有6个同源基因, 且甘蓝型油菜ZF-HD基因等于白菜和甘蓝的同源基因数目之和.实际上除了AtZHD17在甘蓝型油菜有6个同源基因外, 其余AtZHD基因在甘蓝型油菜中的同源基因数目不超过4个, 说明ZF-HD基因在芸薹属三倍化后发生了丢失.推测丢失的原因可能有以下3个: (1) 芸薹属三倍化以后, 在二倍体白菜和甘蓝祖先中发生了基因丢失, 从而导致甘蓝型油菜中共线性同源基因丢失, 如AtZHD3和AtZHD14在白菜和甘蓝中各有1个共线性同源基因, 在油菜中有2个共线性同源基因; (2) 在异源四倍体化以后发生丢失, 如AtZHD1和AtZHD5在白菜和甘蓝中各有3个共线性同源基因, 在油菜中只有3或4个共线性同源基因; (3) 两个过程中都有丢失现象, 如AtZHD13在白菜和甘蓝中各有1个共线性同源基因, 但油菜中没有相应的共线性同源基因, 这可能与三倍化过程中发生大规模的基因丢失和染色体重排有关(Mun et al., 2009; 高虎虎等, 2017).但总体来看, 62个BnZF-HD基因中有48个与拟南芥、白菜和甘蓝存在共线性关系, 从白菜和甘蓝祖先基因组各继承了24个, 表明甘蓝型油菜ZF-HD基因家族扩张的主要力量是异源四倍体化. ...
Evolutionary change of the numbers of homeobox genes in bilateral animals 1 2005
... ZF-HD转录因子(zinc finger homeodomain, ZHD)是植物特有的转录因子(Windhovel et al., 2001; Tan and Irish, 2006), 其在C4植物黄顶菊(Flaveria trinervia)中首次被报道, 是磷酸烯醇式丙酮酸羧化酶(PEPCase)编码基因的潜在调控因子(Windhovel et al., 2001).ZF-HD转录因子的命名是因其氨基酸序列中有保守的HD结构域(homeodomain)和C2H2型锌指结构(C2H2-type zinc finger motif).HD结构域是含60个氨基酸的DNA结合域, 大多数具HD结构域的蛋白同时具有其它类型的结构域或基序, 可分为HD- ZIP、ZF-HD、WOX、Bell type HD、PHD和KNOX六个家族(Ariel et al., 2007).含HD的蛋白质广泛存在于动植物体内, 在发育过程中发挥重要作用(Nam and Nei, 2005; Bhattacharjee et al., 2015).锌指结构是由多个保守的氨基酸残基围绕核心的锌离子组成, 形成相对独立的指状四面体结构(Krishna et al., 2003).根据其Cys和His残基的不同组合分成不同类型, 如C3H、C2H2、C2C2和C4C6, 其中以C2H2型最为常见.锌指可以识别和结合DNA、RNA、DNA-RNA双链分子或蛋白质(Mackay and Crossley, 1998; Takatsuji, 1999; Krishna et al., 2003), 在转录和翻译水平调控基因表达, 在植物的应激反应和防御激活中发挥重要作用(Mackay, 1998).在ZF-HD蛋白中, HD结构域能够结合DNA, ZF结构可以增强由HD结构域介导的蛋白质与DNA的相互作用(Windhovel et al., 2001; Hu et al., 2008). ...
The KA/KS ratio test for assessing the protein-coding potential of genomic regions: an empirical and simulation study 1 2002
Pathogen-induced binding of the soybean zinc finger homeodomain proteins GmZF-HD1 and GmZF-HD2 to two repeats of ATTA homeodomain binding site in the calmodulin isoform 4 (GmCaM4) promoter 1 2007
... 目前, 国内外****已经对水稻、大豆(Glycine max)、拟南芥、小麦(Triticum aestivum)、葡萄(Vitis vinifera)、白菜(Brassica rapa)、番茄(Solanum lycopersicum)和陆地棉(Gossypium hirsutum)等物种中的ZF-HD基因家族进行了鉴定(Jørgensen et al., 1999; Windhövel et al., 2001; Deng et al., 2002; Tan and Irish, 2006; Jain et al., 2008; Bhattacharjee and Jain, 2013; Wang et al., 2014, 2016; Khadiza et al., 2017; Abdullah et al., 2018).已有研究表明, ZF-HD转录因子参与许多生长发育过程和非生物胁迫响应.拟南芥ZF-HD基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
SMART, a simple modular architecture research tool: identification of signaling domains 1 1998
... 通过3种途径搜索甘蓝型油菜和甘蓝的ZF-HD基因.(1) 从植物转录因子网站PlantTFDB (http://planttfdb. cbi.pku.edu.cn/, V4.0) (Jin et al., 2017b)分别下载甘蓝型油菜和甘蓝全部ZF-HD家族成员蛋白序列; (2) 从Pfam数据库(http://pfam.xfam.org/) (Bateman et al., 2002; Finn et al., 2016)下载ZF-HD基因的PF04770结构域种子数据stockholm文件, 使用hmmsearch工具分别搜索甘蓝型油菜和甘蓝的蛋白质数据库, 参数值为1.0; (3) 以拟南芥ZF-HD蛋白序列比对文件, 进行本地Blast P检索, e值设为1e-3; 整理获取的序列, 去掉重复序列, 然后利用在线工具CDD (http://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)和Smart (Schultz et al., 1998) (http://smart. emblheidelberg.de/)检测蛋白序列的结构域, 获得候选基因, 用于后续分析. ...
Zinc-finger proteins: the classical zinc finger emerges in contemporary plant science 1 1999
... ZF-HD转录因子(zinc finger homeodomain, ZHD)是植物特有的转录因子(Windhovel et al., 2001; Tan and Irish, 2006), 其在C4植物黄顶菊(Flaveria trinervia)中首次被报道, 是磷酸烯醇式丙酮酸羧化酶(PEPCase)编码基因的潜在调控因子(Windhovel et al., 2001).ZF-HD转录因子的命名是因其氨基酸序列中有保守的HD结构域(homeodomain)和C2H2型锌指结构(C2H2-type zinc finger motif).HD结构域是含60个氨基酸的DNA结合域, 大多数具HD结构域的蛋白同时具有其它类型的结构域或基序, 可分为HD- ZIP、ZF-HD、WOX、Bell type HD、PHD和KNOX六个家族(Ariel et al., 2007).含HD的蛋白质广泛存在于动植物体内, 在发育过程中发挥重要作用(Nam and Nei, 2005; Bhattacharjee et al., 2015).锌指结构是由多个保守的氨基酸残基围绕核心的锌离子组成, 形成相对独立的指状四面体结构(Krishna et al., 2003).根据其Cys和His残基的不同组合分成不同类型, 如C3H、C2H2、C2C2和C4C6, 其中以C2H2型最为常见.锌指可以识别和结合DNA、RNA、DNA-RNA双链分子或蛋白质(Mackay and Crossley, 1998; Takatsuji, 1999; Krishna et al., 2003), 在转录和翻译水平调控基因表达, 在植物的应激反应和防御激活中发挥重要作用(Mackay, 1998).在ZF-HD蛋白中, HD结构域能够结合DNA, ZF结构可以增强由HD结构域介导的蛋白质与DNA的相互作用(Windhovel et al., 2001; Hu et al., 2008). ...
The Arabidopsis zinc finger-homeodomain genes encode proteins with unique biochemical properties that are coordinately expressed during floral development 3 2006
... ZF-HD转录因子(zinc finger homeodomain, ZHD)是植物特有的转录因子(Windhovel et al., 2001; Tan and Irish, 2006), 其在C4植物黄顶菊(Flaveria trinervia)中首次被报道, 是磷酸烯醇式丙酮酸羧化酶(PEPCase)编码基因的潜在调控因子(Windhovel et al., 2001).ZF-HD转录因子的命名是因其氨基酸序列中有保守的HD结构域(homeodomain)和C2H2型锌指结构(C2H2-type zinc finger motif).HD结构域是含60个氨基酸的DNA结合域, 大多数具HD结构域的蛋白同时具有其它类型的结构域或基序, 可分为HD- ZIP、ZF-HD、WOX、Bell type HD、PHD和KNOX六个家族(Ariel et al., 2007).含HD的蛋白质广泛存在于动植物体内, 在发育过程中发挥重要作用(Nam and Nei, 2005; Bhattacharjee et al., 2015).锌指结构是由多个保守的氨基酸残基围绕核心的锌离子组成, 形成相对独立的指状四面体结构(Krishna et al., 2003).根据其Cys和His残基的不同组合分成不同类型, 如C3H、C2H2、C2C2和C4C6, 其中以C2H2型最为常见.锌指可以识别和结合DNA、RNA、DNA-RNA双链分子或蛋白质(Mackay and Crossley, 1998; Takatsuji, 1999; Krishna et al., 2003), 在转录和翻译水平调控基因表达, 在植物的应激反应和防御激活中发挥重要作用(Mackay, 1998).在ZF-HD蛋白中, HD结构域能够结合DNA, ZF结构可以增强由HD结构域介导的蛋白质与DNA的相互作用(Windhovel et al., 2001; Hu et al., 2008). ...
... 目前, 国内外****已经对水稻、大豆(Glycine max)、拟南芥、小麦(Triticum aestivum)、葡萄(Vitis vinifera)、白菜(Brassica rapa)、番茄(Solanum lycopersicum)和陆地棉(Gossypium hirsutum)等物种中的ZF-HD基因家族进行了鉴定(Jørgensen et al., 1999; Windhövel et al., 2001; Deng et al., 2002; Tan and Irish, 2006; Jain et al., 2008; Bhattacharjee and Jain, 2013; Wang et al., 2014, 2016; Khadiza et al., 2017; Abdullah et al., 2018).已有研究表明, ZF-HD转录因子参与许多生长发育过程和非生物胁迫响应.拟南芥ZF-HD基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
... 基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
Co-expression of the stress-inducible zinc finger homeodomain ZFHD1 and NAC transcription factors enhances expression of the ERD1 gene in Arabidopsis 2 2007
... 目前, 国内外****已经对水稻、大豆(Glycine max)、拟南芥、小麦(Triticum aestivum)、葡萄(Vitis vinifera)、白菜(Brassica rapa)、番茄(Solanum lycopersicum)和陆地棉(Gossypium hirsutum)等物种中的ZF-HD基因家族进行了鉴定(Jørgensen et al., 1999; Windhövel et al., 2001; Deng et al., 2002; Tan and Irish, 2006; Jain et al., 2008; Bhattacharjee and Jain, 2013; Wang et al., 2014, 2016; Khadiza et al., 2017; Abdullah et al., 2018).已有研究表明, ZF-HD转录因子参与许多生长发育过程和非生物胁迫响应.拟南芥ZF-HD基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
... 基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
The regulatory code for transcriptional response diversity and its relation to genome structural properties in A. thaliana 1 2007
Genome-wide identification, evolution and expression analysis of the grape ( Vitis vinifera L.) zinc finger-homeodomain gene family 4 2014
... 目前, 国内外****已经对水稻、大豆(Glycine max)、拟南芥、小麦(Triticum aestivum)、葡萄(Vitis vinifera)、白菜(Brassica rapa)、番茄(Solanum lycopersicum)和陆地棉(Gossypium hirsutum)等物种中的ZF-HD基因家族进行了鉴定(Jørgensen et al., 1999; Windhövel et al., 2001; Deng et al., 2002; Tan and Irish, 2006; Jain et al., 2008; Bhattacharjee and Jain, 2013; Wang et al., 2014, 2016; Khadiza et al., 2017; Abdullah et al., 2018).已有研究表明, ZF-HD转录因子参与许多生长发育过程和非生物胁迫响应.拟南芥ZF-HD基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
... ; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
... ; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
... ZF-HD基因在植物多种发育和生理过程中发挥作用.研究发现ZF-HD基因家族只存在于陆生植物中(Wang et al., 2016), 这类基因可能在陆生植物祖先与单细胞藻类趋异后得到了进化.本研究在甘蓝型油菜中鉴定到62个ZF-HD基因, 参考其它植物中的分类(Hu et al., 2008), 发现MIF与ZF-HD蛋白相似度高, 虽然缺乏HD结构域, 也将14个MIF基因纳入ZF-HD基因家族的范畴.与拟南芥、白菜和甘蓝的ZF-HD蛋白聚类分析后, 分成6个亚群, MIF单独形成1个亚群.同一亚群的motif分布模式基本一致, 如MIF都具有相同的3个基序, 不同亚群之间motif分布差异较大.甘蓝型油菜ZF-HD基因大多缺乏内含子, 这与拟南芥、水稻、葡萄及白菜等植物中的研究结果一致(Hu et al., 2008; Wang et al., 2014, 2016). ...
Genome-wide analysis and expression patterns of ZF-HD transcription factors under different developmental tissues and abiotic stresses in Chinese cabbage 6 2016
... 有趣的是, 有一类蛋白MIF (MINI ZINC FINGER)与ZF-HD蛋白的ZF结构域序列相似度高但缺乏HD结构域(Hu and Ma, 2006; Hu et al., 2008), 且是种子植物特有的蛋白.推测MIF蛋白与ZF-HD蛋白的进化关系有2种可能, 一是MIF蛋白由ZF-HD蛋白丢失HD结构域形成; 二是MIF获得HD结构域形成ZF-HD蛋白(Hu et al., 2008; Wang et al., 2016).按照已发表的模式植物拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)的ZF-HD蛋白的划分方法, 本研究将MIF归入ZF-HD家族. ...
... 目前, 国内外****已经对水稻、大豆(Glycine max)、拟南芥、小麦(Triticum aestivum)、葡萄(Vitis vinifera)、白菜(Brassica rapa)、番茄(Solanum lycopersicum)和陆地棉(Gossypium hirsutum)等物种中的ZF-HD基因家族进行了鉴定(Jørgensen et al., 1999; Windhövel et al., 2001; Deng et al., 2002; Tan and Irish, 2006; Jain et al., 2008; Bhattacharjee and Jain, 2013; Wang et al., 2014, 2016; Khadiza et al., 2017; Abdullah et al., 2018).已有研究表明, ZF-HD转录因子参与许多生长发育过程和非生物胁迫响应.拟南芥ZF-HD基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
... 基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
... ZF-HD基因在植物多种发育和生理过程中发挥作用.研究发现ZF-HD基因家族只存在于陆生植物中(Wang et al., 2016), 这类基因可能在陆生植物祖先与单细胞藻类趋异后得到了进化.本研究在甘蓝型油菜中鉴定到62个ZF-HD基因, 参考其它植物中的分类(Hu et al., 2008), 发现MIF与ZF-HD蛋白相似度高, 虽然缺乏HD结构域, 也将14个MIF基因纳入ZF-HD基因家族的范畴.与拟南芥、白菜和甘蓝的ZF-HD蛋白聚类分析后, 分成6个亚群, MIF单独形成1个亚群.同一亚群的motif分布模式基本一致, 如MIF都具有相同的3个基序, 不同亚群之间motif分布差异较大.甘蓝型油菜ZF-HD基因大多缺乏内含子, 这与拟南芥、水稻、葡萄及白菜等植物中的研究结果一致(Hu et al., 2008; Wang et al., 2014, 2016). ...
... , 2016). ...
The genome of the mesopolyploid crop species Brassica rapa 1 2011
... 比较基因组学研究表明, 在白菜和甘蓝基因组的不同区域, 基因密度与表达水平存在差异, 可划分为LF (least fractionated)、MF1 (medium fractionated) 和MF2 (most fractionated) 3个亚基因组(subgeno- me) (Wang et al., 2011; Liu et al., 2014).与拟南芥ZF-HD基因相比, 白菜、甘蓝及甘蓝型油菜中大多数保留下来的ZF-HD基因位于LF亚基因组, 这与整个基因组的保留趋势一致, 其次是MF2亚基因组, MF1亚基因组保留的ZF-HD基因最少, 而基因组整体水平上MF1亚基因组基因丢失的数目最多. ...
Characterization of a novel class of plant homeodomain proteins that bind to the C4 phosphoenolpyruvate carboxylase gene of Flaveria trinervia 4 2001
... ZF-HD转录因子(zinc finger homeodomain, ZHD)是植物特有的转录因子(Windhovel et al., 2001; Tan and Irish, 2006), 其在C4植物黄顶菊(Flaveria trinervia)中首次被报道, 是磷酸烯醇式丙酮酸羧化酶(PEPCase)编码基因的潜在调控因子(Windhovel et al., 2001).ZF-HD转录因子的命名是因其氨基酸序列中有保守的HD结构域(homeodomain)和C2H2型锌指结构(C2H2-type zinc finger motif).HD结构域是含60个氨基酸的DNA结合域, 大多数具HD结构域的蛋白同时具有其它类型的结构域或基序, 可分为HD- ZIP、ZF-HD、WOX、Bell type HD、PHD和KNOX六个家族(Ariel et al., 2007).含HD的蛋白质广泛存在于动植物体内, 在发育过程中发挥重要作用(Nam and Nei, 2005; Bhattacharjee et al., 2015).锌指结构是由多个保守的氨基酸残基围绕核心的锌离子组成, 形成相对独立的指状四面体结构(Krishna et al., 2003).根据其Cys和His残基的不同组合分成不同类型, 如C3H、C2H2、C2C2和C4C6, 其中以C2H2型最为常见.锌指可以识别和结合DNA、RNA、DNA-RNA双链分子或蛋白质(Mackay and Crossley, 1998; Takatsuji, 1999; Krishna et al., 2003), 在转录和翻译水平调控基因表达, 在植物的应激反应和防御激活中发挥重要作用(Mackay, 1998).在ZF-HD蛋白中, HD结构域能够结合DNA, ZF结构可以增强由HD结构域介导的蛋白质与DNA的相互作用(Windhovel et al., 2001; Hu et al., 2008). ...
... )中首次被报道, 是磷酸烯醇式丙酮酸羧化酶(PEPCase)编码基因的潜在调控因子(Windhovel et al., 2001).ZF-HD转录因子的命名是因其氨基酸序列中有保守的HD结构域(homeodomain)和C2H2型锌指结构(C2H2-type zinc finger motif).HD结构域是含60个氨基酸的DNA结合域, 大多数具HD结构域的蛋白同时具有其它类型的结构域或基序, 可分为HD- ZIP、ZF-HD、WOX、Bell type HD、PHD和KNOX六个家族(Ariel et al., 2007).含HD的蛋白质广泛存在于动植物体内, 在发育过程中发挥重要作用(Nam and Nei, 2005; Bhattacharjee et al., 2015).锌指结构是由多个保守的氨基酸残基围绕核心的锌离子组成, 形成相对独立的指状四面体结构(Krishna et al., 2003).根据其Cys和His残基的不同组合分成不同类型, 如C3H、C2H2、C2C2和C4C6, 其中以C2H2型最为常见.锌指可以识别和结合DNA、RNA、DNA-RNA双链分子或蛋白质(Mackay and Crossley, 1998; Takatsuji, 1999; Krishna et al., 2003), 在转录和翻译水平调控基因表达, 在植物的应激反应和防御激活中发挥重要作用(Mackay, 1998).在ZF-HD蛋白中, HD结构域能够结合DNA, ZF结构可以增强由HD结构域介导的蛋白质与DNA的相互作用(Windhovel et al., 2001; Hu et al., 2008). ...
... ).在ZF-HD蛋白中, HD结构域能够结合DNA, ZF结构可以增强由HD结构域介导的蛋白质与DNA的相互作用(Windhovel et al., 2001; Hu et al., 2008). ...
... 目前, 国内外****已经对水稻、大豆(Glycine max)、拟南芥、小麦(Triticum aestivum)、葡萄(Vitis vinifera)、白菜(Brassica rapa)、番茄(Solanum lycopersicum)和陆地棉(Gossypium hirsutum)等物种中的ZF-HD基因家族进行了鉴定(Jørgensen et al., 1999; Windhövel et al., 2001; Deng et al., 2002; Tan and Irish, 2006; Jain et al., 2008; Bhattacharjee and Jain, 2013; Wang et al., 2014, 2016; Khadiza et al., 2017; Abdullah et al., 2018).已有研究表明, ZF-HD转录因子参与许多生长发育过程和非生物胁迫响应.拟南芥ZF-HD基因均在花组织中有表达, 有些表现为花特异性表达(Tan and Irish, 2006).拟南芥AtZHD1蛋白能特异性结合ERD1 (EARLY RESPONSE TO DEHYDRATION STRESS 1)启动子, AtZHD1基因受ABA、盐和干旱诱导(Tran et al., 2007; Wang et al., 2014).过表达AtZHD1和NAC基因的拟南芥植株耐旱性增强(Tran et al., 2007; Hu et al., 2008).4个水稻ZHD基因与基因表达调控相关.大豆GmZHD1和GmZHD2能够结合钙调蛋白编码基因GmCaM4的启动子, 在病原菌侵染后表达量升高(Park et al., 2007; Hu et al., 2008; Wang et al., 2014).15个陆地棉ZHD基因参与纤维的早期发育和纤维色素合成(Abdullah et al., 2018).大多数番茄ZF-HD基因在花芽部位表达, 其中有多个响应非生物胁迫和激素处理(Khadiza et al., 2017).多数白菜ZF-HD基因在花中表达, 且明显受光周期、春化和非生物胁迫诱导(Wang et al., 2016). ...
Organization of cis -acting regulatory elements in osmotic-and cold- stress-responsive promoters 1 2005
 ,*, 张瑶, 王丽莹, 彭向永曲阜师范大学生命科学学院, 曲阜 273165
,*, 张瑶, 王丽莹, 彭向永曲阜师范大学生命科学学院, 曲阜 273165
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT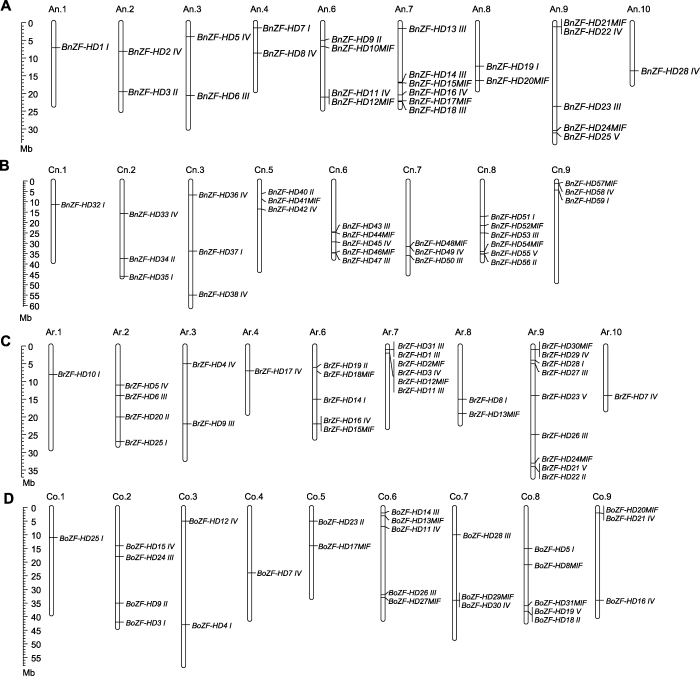 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT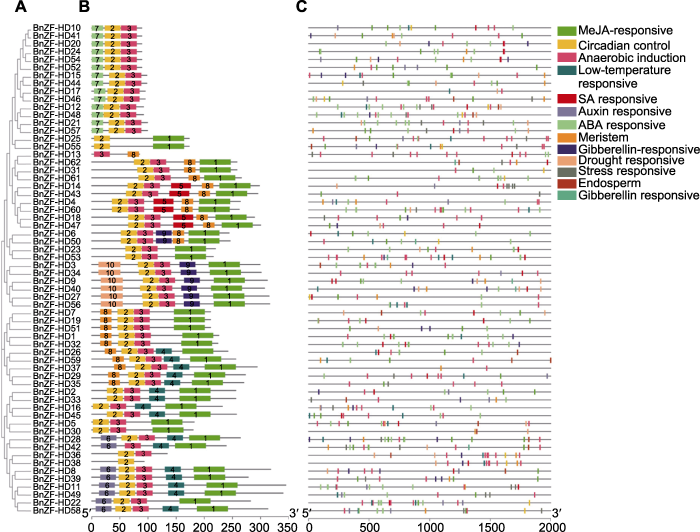 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT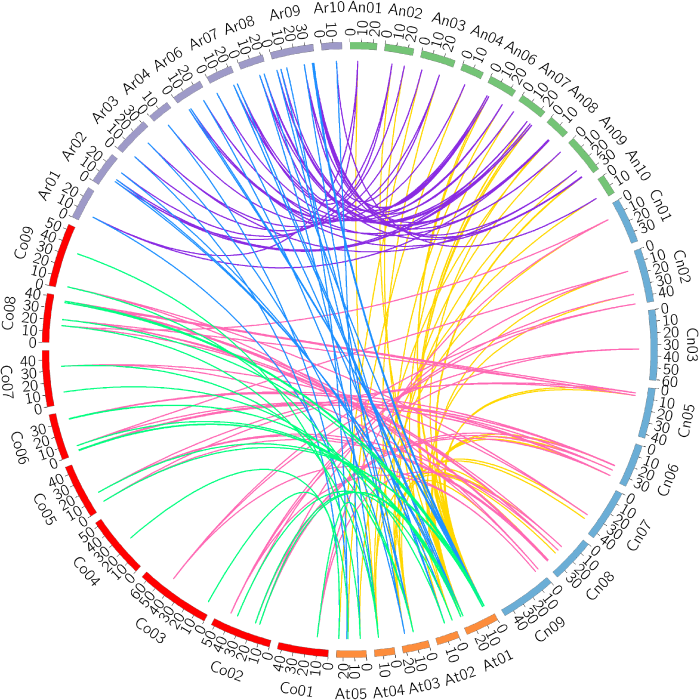 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}