 ,1,*, 戴绍军,1,2,*1.
,1,*, 戴绍军,1,2,*1. 2.
3.
Research Advances in DREPP Gene Family in Plants
Xun Zhang1, Juanjuan Yu1,3, Sizhu Wang1, Ying Li,1,*, Shaojun Dai,1,2,*1. 2.
3.
通讯作者:
责任编辑: 白羽红
收稿日期:2019-01-21接受日期:2019-04-23网络出版日期:2019-09-01
| 基金资助: |
Editor:
Received:2019-01-21Accepted:2019-04-23Online:2019-09-01

摘要
关键词:
Abstract
Keywords:
PDF (3747KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
张洵, 喻娟娟, 王思竹, 李莹, 戴绍军. 植物DREPP基因家族研究进展. 植物学报, 2019, 54(5): 582-595 doi:10.11983/CBB19014
Zhang Xun, Yu Juanjuan, Wang Sizhu, Li Ying, Dai Shaojun.
发育调节质膜多肽(developmentally regulated plasma membrane polypeptide, DREPP)蛋白是一类与质膜相关的Ca2+结合蛋白。它可以通过与微管和微丝结合调节细胞骨架结构, 从而调控根和花粉管的发育(Zhu et al., 2013; Qin et al., 2014), 也可以参与脱落酸(abscisic acid, ABA)与水杨酸(salicylic acid, SA)介导的多种信号通路, 从而调控植物应答干旱与低温等非生物胁迫(Wang et al., 2018a, 2018b)。因此, 阐明DREPP的功能对于认识植物生长发育及抵御逆境的分子调控机制具有重要意义。
1 DREPP蛋白家族组成与进化关系
人们最早从烟草(Nicotiana tabacum)中克隆到DREPP编码基因(Gantet et al., 1996)。此后, 在拟南芥(Arabidopsis thaliana)中发现了2个质膜定位的DREPP蛋白, 分别将其命名为质膜相关阳离子结合蛋白(plasma membrane-associated cation binding protein, PCaP) 1和2 (Ide et al., 2007; Kato et al., 2010a)。AtPCaP1和AtPCaP2分别被称为微管去稳定蛋白25 (microtubule-destabilizing protein 25, MDP25)和微管相关蛋白18 (microtubule-associated protein 18, MAP18) (Wang et al., 2007; Li et al.,2011)。近年来, 人们陆续在几十种植物中发现了多个DREPP家族成员。我们用AtPCaP1 (AtMDP25)和AtPCaP2 (AtMAP18)的氨基酸序列在NCBI蛋白质数据库中进行BLAST搜索, 同时参考相关文献报道的DREPP蛋白质序列, 共得到191条DREPP蛋白序列(附表1)。我们对这些DREPP蛋白序列的完整性和功能结构域进行分析, 最终确定了79种植物中120条完整的DREPP蛋白(表1)。我们用MEGA7.0软件中的Clustal W方法对这120种DREPP蛋白的氨基酸序列进行多重比对, 并采用邻接法构建系统发生树(图1)。Table 1
表1
表1植物DREPP蛋白家族成员组成
Table 1
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 1 | 番茄 | Solanum lycopersicum | Sl | 茄科 | 茄属 | XP_004248413.1 | - |
| 2 | 野生潘那利番茄 | S. pennellii | Sp | 茄科 | 茄属 | XP_015055582.1 | - |
| 3 | 马铃薯 | S. tuberosum | St | 茄科 | 茄属 | XP_006352621.1 | - |
| 4 | 风铃辣椒 | Capsicum baccatum | Cb | 茄科 | 辣椒属 | PHT35828.1 | Kim et al., 2017 |
| 5 | 黄灯笼辣椒 | C. chinense | Cch | 茄科 | 辣椒属 | PHU04552.1 | Kim et al., 2017 |
| 6 | 辣椒 | C. annuum | Can | 茄科 | 辣椒属 | PHT70067.1 | Kim et al., 2017 |
| 7 | 本氏烟草 | Nicotiana benthamiana | Nb | 茄科 | 烟草属 | ASW15993.1 | Huang et al., 2017 |
| 8 | 渐狭叶烟草 | N. attenuata | Na | 茄科 | 烟草属 | XP_019226343.1 | - |
| 9 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69901.1 | Logan et al., 1998 |
| 10 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69900.1 | Logan et al., 1998 |
| 11 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016438748.1 | - |
| 12 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | X95957.1 | Gantet et al., 1996 |
| 13 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016451342.1 | - |
| 14 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_018631801.1 | - |
| 15 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_009600368.1 | - |
| 16 | 林烟草 | N. sylvestris | Ns | 茄科 | 烟草属 | XP_009763203.1 | - |
| 17 | 中粒咖啡 | Coffea canephora | Cca | 茜草科 | 咖啡属 | CDP04109.1 | - |
| 18 | 小粒咖啡 | C. arabica | Car | 茜草科 | 咖啡属 | XP_027089181.1 | - |
| 19 | 木犀榄 | Olea europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022847863.1 | - |
| 20 | 木犀榄 | O. europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022870576.1 | - |
| 21 | 芝麻 | Sesamum indicum | Si | 胡麻科 | 胡麻属 | XP_011085144.1 | - |
| 22 | 中华猕猴桃 | Actinidia chinensis var. chinensis | Ac | 猕猴桃科 | 猕猴桃属 | PSS11063.1 | - |
| 23 | 苹果 | Malus domestica | Md | 蔷薇科 | 苹果属 | XP_008383239.1 | - |
| 24 | 月季 | Rosa chinensis | Rch | 蔷薇科 | 蔷薇属 | XP_024156781.1 | - |
| 25 | 桃 | Prunus persica | Pp | 蔷薇科 | 桃属 | XP_007212006.1 | - |
| 26 | 樱花 | P. yedoensis var. nudiflora | Py | 蔷薇科 | 桃属 | PQP97549.1 | - |
| 27 | 甜樱桃 | P. avium | Pa | 蔷薇科 | 桃属 | XP_021822163.1 | - |
| 28 | 刺苞菜蓟 | Cynara cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | XP_024964532.1 | - |
| 29 | 刺苞菜蓟 | C. cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | KVH87676.1 | Scaglione et al., 2016 |
| 30 | 莴苣 | Lactuca sativa | Ls | 菊科 | 莴苣属 | XP_023745485.1 | - |
| 31 | 莴苣 | L. sativa | Ls | 菊科 | 莴苣属 | XP_023768949.1 | - |
| 32 | 向日葵 | Helianthus annuus | Ha | 菊科 | 向日葵属 | XP_021982589.1 | - |
| 33 | 向日葵 | H. annuus | Ha | 菊科 | 向日葵属 | XP_022009496.1 | - |
| 34 | 黄花蒿 | Artemisia annua | Aa | 菊科 | 蒿亚属 | PWA97158.1 | Shen et al., 2018 |
| 35 | 蒺藜苜蓿 | Medicago truncatula | Mt | 豆科 | 苜蓿属 | XP_013463310.1 | - |
| 36 | 地三叶 | Trifolium subterraneum | Ts | 豆科 | 三叶草属 | GAU45950.1 | Kaur et al., 2017 |
| 37 | 鹰嘴豆 | Cicer arietinum | Ca | 豆科 | 鹰嘴豆属 | Q9SMK5 | Subba et al., 2013 |
| 38 | 野生落花生 | Arachis ipaensis | Ai | 豆科 | 落花生属 | XP_016197976.1 | - |
| 39 | 蔓花生 | A. duranensis | Ad | 豆科 | 落花生属 | XP_015959777.1 | - |
| 40 | 百脉根 | Lotus japonicus | Lj | 豆科 | 百脉根属 | AFK34640.1 | - |
| 41 | 菜豆 | Phaseolus vulgaris | Pv | 豆科 | 菜豆属 | AGV54625.1 | - |
新窗口打开|下载CSV
Table 1
表1
表1(续)
Table 1
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 42 | 大豆 | Glycine max | Gm | 豆科 | 大豆属 | XP_003546380.1 | Matthews et al., 2013 |
| 43 | 大豆 | G. max | Gm | 豆科 | 大豆属 | NP_001237575.1 | - |
| 44 | 欧洲栓皮栎 | Quercus suber | Qs | 壳斗科 | 栎属 | XP_023919313.1 | - |
| 45 | 土瓶草 | Cephalotus follicularis | Cf | 土瓶草科 | 土瓶草属 | GAV80098.1 | Fukushima et al., 2017 |
| 46 | 胡桃 | Juglans regia | Jr | 胡桃科 | 胡桃属 | XP_018814574.1 | - |
| 47 | 甜橙 | Citrus sinensis | Cs | 芸香科 | 柑橘属 | XP_006468602.1 | - |
| 48 | 克里曼丁桔 | C. clementina | Ccl | 芸香科 | 柑橘属 | XP_006448565.1 | - |
| 49 | 大桉 | Eucalyptus grandis | Eg | 桃金娘科 | 桉属 | XP_010051796.1 | - |
| 50 | 毛果杨 | Populus trichocarpa | Pt | 杨柳科 | 杨属 | XP_006385859.1 | - |
| 51 | 毛果杨 | P. trichocarpa | Pt | 杨柳科 | 杨属 | XP_006368838.1 | - |
| 52 | 蓖麻 | Ricinus communis | Rc | 大戟科 | 蓖麻属 | XP_015583044.1 | - |
| 53 | 麻疯树 | Jatropha curcas | Jc | 大戟科 | 麻疯树属 | XP_012088609.1 | - |
| 54 | 木薯 | Manihot esculenta | Me | 大戟科 | 木薯属 | XP_021602838.1 | - |
| 55 | 橡胶树 | Hevea brasiliensis | Hb | 大戟科 | 橡胶树属 | XP_021686108.1 | - |
| 56 | 糙叶山黄麻 | Parasponia andersonii | Pan | 榆科 | 南山黄麻属 | PON44636.1 | - |
| 57 | 异色山黄麻 | Trema orientale | To | 椴树科 | 山黄麻属 | PON83811.1 | - |
| 58 | 长蒴黄麻 | Corchorus olitorius | Co | 椴树科 | 黄麻属 | OMO77743.1 | - |
| 59 | 博落回 | Macleaya cordata | Mc | 罂粟科 | 博落回属 | OVA01374.1 | Liu et al., 2017 |
| 60 | 莲 | Nelumbo nucifera | Nn | 睡莲科 | 莲属 | XP_010260837.1 | - |
| 61 | 川桑 | Morus notabilis | Mn | 桑科 | 桑属 | XP_010094633.1 | - |
| 62 | 葡萄 | Vitis vinifera | Vv | 葡萄科 | 葡萄属 | XP_002263090.1 | - |
| 63 | 菠菜 | Spinacia oleracea | So | 藜科 | 菠菜属 | KNA18787.1 | Dohm et al., 2014 |
| 64 | 甜菜 | Beta vulgaris subsp. vulgaris | Bv | 藜科 | 甜菜属 | XP_010684190.2 | - |
| 65 | 甜菜 | B. vulgaris subsp. vulgaris | Bv | 藜科 | 甜菜属 | KMT06194.1 | Dohm et al., 2014 |
| 66 | 无油樟 | Amborella trichopoda | Atr | 无油樟科 | 无油樟属 | XP_006859057.2 | - |
| 67 | 醉蝶花 | Tarenaya hassleriana | Th | 醉蝶花科 | 白花菜属 | XP_010537158.1 | - |
| 68 | 芜菁 | Brassica rapa | Br | 十字花科 | 芸苔属 | AHV84997.1 | - |
| 69 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | XP_009133577.1 | - |
| 70 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | RIA04026.1 | - |
| 71 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | XP_009113907.1 | - |
| 72 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | XP_009101693.1 | - |
| 73 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013660923.1 | - |
| 74 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | CDY66354.1 | - |
| 75 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013672014.1 | - |
| 76 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_022572533.1 | - |
| 77 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013717493.1 | - |
| 78 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013659410.1 | - |
| 79 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013730570.1 | - |
| 80 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013715242.1 | - |
| 81 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013733201.1 | - |
| 82 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | CDY63708.1 | - |
| 83 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013602975.1 | - |
新窗口打开|下载CSV
Table 1
表1
表1(续)
Table 1
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 84 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013610402.1 | - |
| 85 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013596813.1 | - |
| 86 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013619975.1 | - |
| 87 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD40021.1 | - |
| 88 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD37718.1 | - |
| 89 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD25026.1 | - |
| 90 | 萝卜 | Raphanus sativus | Rs | 十字花科 | 萝卜属 | XP_018471954.1 | - |
| 91 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | BAA99394.1 | Yuasa and Mae- shima, 2000 |
| 92 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018437382.1 | - |
| 93 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018455198.1 | - |
| 94 | 山嵛菜 | Eutrema salsugineum | Es | 十字花科 | 山嵛菜属 | XP_006413890.1 | - |
| 95 | 山嵛菜 | E. salsugineum | Es | 十字花科 | 山嵛菜属 | XP_024011088.1 | - |
| 96 | 荠菜 | Capsella rubella | Cr | 十字花科 | 荠属 | XP_006284474.1 | - |
| 97 | 荠菜 | C. rubella | Cr | 十字花科 | 荠属 | XP_006279705.1 | - |
| 98 | 亚麻荠 | Camelina sativa | Csa | 十字花科 | 亚麻荠属 | XP_010434307.1 | - |
| 99 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010441852.1 | - |
| 100 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010445250.1 | - |
| 101 | 天蓝遏蓝菜 | Noccaea caerulescens | Nc | 十字花科 | 菥蓂属 | JAU60665.1 | - |
| 102 | 拟南芥 | Arabidopsis thaliana | At | 十字花科 | 拟南芥属 | Q96262.1 | Ide et al., 2007 |
| 103 | 拟南芥 | A. thaliana | At | 十字花科 | 拟南芥属 | NP_568636.1 | Kato et al., 2010a |
| 104 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002869944.1 | - |
| 105 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002863557.1 | - |
| 106 | 番木瓜 | Carica papaya | Cp | 番木瓜科 | 番木瓜属 | XP_021901673.1 | - |
| 107 | 凤梨 | Ananas comosus | Aco | 凤梨科 | 凤梨属 | OAY80473.1 | Redwan et al., 2016 |
| 108 | 深圳拟兰 | Apostasia shenzhenica | As | 兰科 | 拟兰属 | PKA54423.1 | - |
| 109 | - | Sarocalamus faberi | Sf | 兰科 | - | ATE87948.1 | - |
| 110 | 小果野芭蕉 | Musa acuminata subsp. malaccensis | Ma | 芭蕉科 | 芭蕉属 | XP_009409470.1 | - |
| 111 | 小兰屿蝴蝶兰 | Phalaenopsis equestris | Phe | 兰科 | 蝴蝶兰属 | XP_020574860.1 | - |
| 112 | 铁皮石斛 | Dendrobium catenatum | Dc | 兰科 | 石斛属 | XP_020688760.1 | - |
| 113 | 玉米 | Zea mays | Zm | 禾本科 | 玉蜀黍属 | XP_020397542.1 | - |
| 114 | 玉米 | Z. mays | Zm | 禾本科 | 玉蜀黍属 | ACG37450.1 | Alexandrov et al., 2009 |
| 115 | 水稻 | Oryza sativa | Os | 禾本科 | 稻属 | NP_001042508.1 | Yamada et al., 2015 |
| 116 | 水稻 | O. sativa | Os | 禾本科 | 稻属 | NP_001046572.1 | Yamada et al., 2015 |
| 117 | 盐地鼠尾粟 | Sporobolus virginicus | Sv | 禾本科 | 鼠尾粟属 | LC342075.1 | Theerawitaya et al., 2018 |
| 118 | 二穗短柄草 | Brachypodium distachyon | Bd | 禾本科 | 短柄草属 | XP_003572598.1 | - |
| 119 | 高粱 | Sorghum bicolor | Sb | 禾本科 | 高粱属 | XP_002453713.1 | - |
| 120 | - | Dichanthelium oligosanthes | Do | 禾本科 | 二型花属 | OEL25601.1 | Studer et al., 2016 |
新窗口打开|下载CSV
根据进化关系分析结果, DREPP蛋白可分为A、B、C三个分支(图1)。A分支中除了3种DREPP来自单子叶植物兰科外, 其它DREPP均来自双子叶植物。B分支包含了十字花科DREPP蛋白和1种醉蝶花(Tarenaya hassleriana) DREPP蛋白。在C分支中, 除了2种DREPP来自双子叶植物番木瓜(Carica papaya)和长蒴黄麻(Corchorus olitorius)外, 其余11种DREPP蛋白都来自单子叶植物, 且以禾本科植物居多。DREPP蛋白的进化关系与植物系统进化关系基本一致。例如, 莴苣(Lactuca sativa)、黄花蒿(Artemisia annua)和向日葵(Helianthus annuus)都是菊科植物, 它们的DREPP蛋白在系统发生树上处于同一个小分支。欧洲油菜(Brassica napus)与甘蓝(B. oleracea)和芜菁(B. rapa)的亲缘关系较近, 它们的DREPP蛋白也都被分在B类(图1)。但并不是所有的单子叶或双子叶植物的DREPP在一个大分支里, 可能是DREPP在植物进化过程中发生了变异。
图1
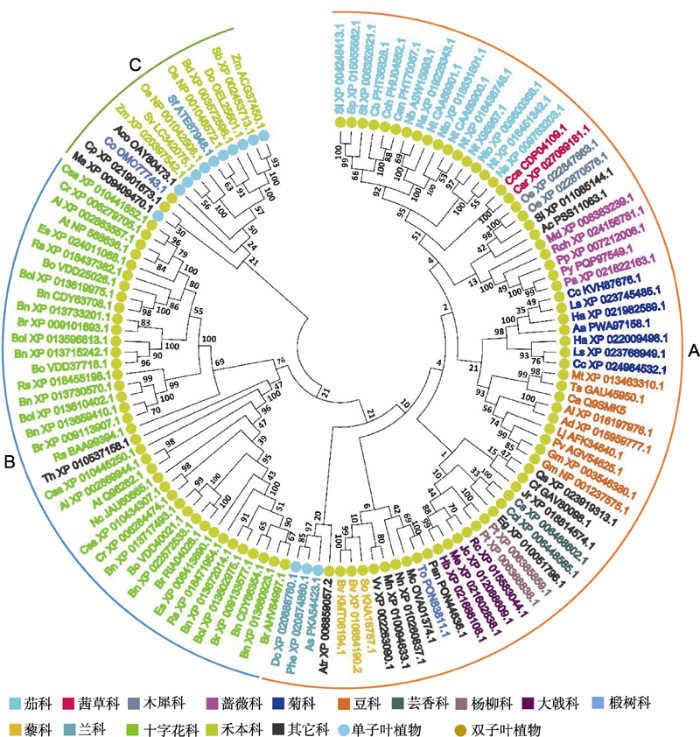 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1DREPP蛋白家族进化关系
Figure 1Evolutionary relationship of DREPP protein family
2 DREPP蛋白序列特征
DREPP蛋白的氨基酸序列一般可分为氨基端区域、中央区域和羧基端区域(图2A), 但AtPCaP2只包含氨基端区域和羧基端区域两部分(Vosolsobě et al., 2017) (图2B)。DREPP蛋白的氨基端区域约含23个高度保守的氨基酸残基, 被称为N23域。AtPCaP1和AtPCaP2的N23域中第2位甘氨酸(Gly2)为豆蔻酰化位点, 第5-23位氨基酸为静电互作区, 豆蔻酰化和静电互作区参与AtDREPPs的质膜定位过程(Vosolsobě et al., 2017)。DREPP蛋白氨基端氨基酸序列比对结果(图2C)表明, Gly2在双子叶植物DREPP中高度保守, 在单子叶植物中不保守。由此可见, 单子叶植物DREPP的定位可能与豆蔻酰化无关, 其可能通过其它氨基酸序列与质膜结合。Gly2也是Ca2+/钙调蛋白(calmodulin, CaM)复合物和磷脂酰肌醇磷酸(phosphatidylinositol phosphates, PtdInsPs)结合DREPP的关键位点(Kato et al., 2010b)。据报道, AtPCaP2的N23域缺失将导致其丧失与PtdInsPs结合的能力(Kato et al., 2013)。N23域中第4位色氨酸(Trp4)和第16位苯丙氨酸(Phe16)在DREPP家族成员中高度保守(图2C)。水稻(Oryza sativa) OsDREPP2的Trp4和Phe16是Ca2+/CaM结合的重要位点, Trp4和Phe16的突变会降低OsDREPP2与Ca2+/CaM结合的能力(Yamada et al., 2015)。AtPCaP1的N23域中第5位天冬氨酸(Asn5)是切割微丝的关键位点, Asn5的突变会导致其切割微丝能力减弱(Qin et al., 2014); 第7位和第18位赖氨酸(Lys)是结合微管的关键位点, 这2个位点突变将导致其丧失结合微管的能力(Li et al., 2011)。因此推测, N23第5、7和18位的氨基酸可能是DREPP与微丝和微管结合的重要位点。DREPP蛋白的中央区域由约110个高度保守的氨基酸组成(图2D), 羧基端区域氨基酸不保守(图2E)。图2
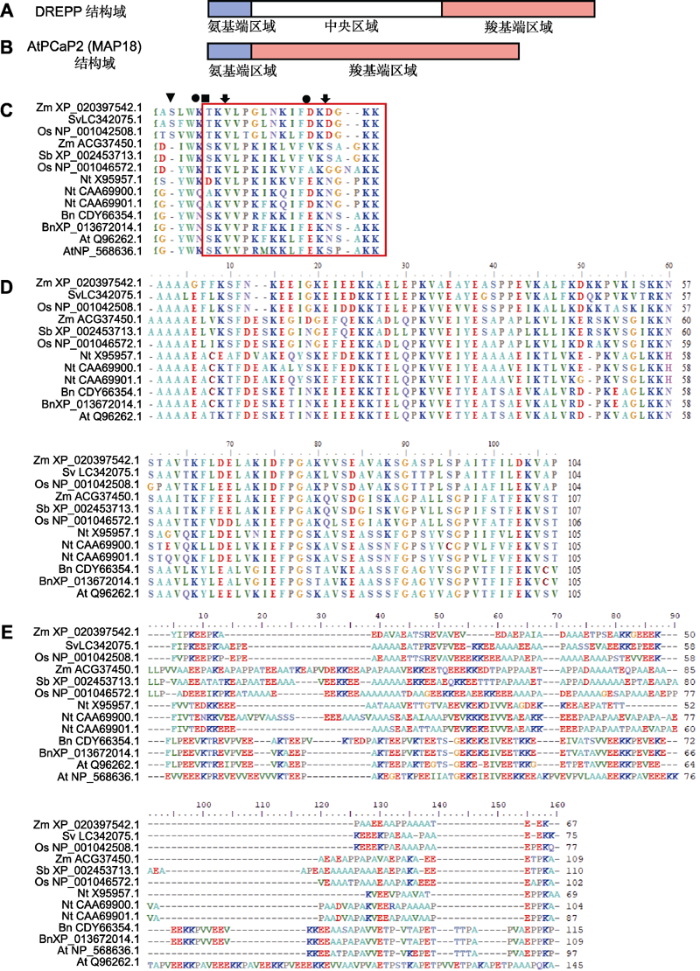 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2植物DREPP蛋白功能结构域示意图
(A) DREPP蛋白功能结构域; (B) AtPCaP2/AtMAP18蛋白功能结构域; (C) DREPP蛋白氨基端区域氨基酸序列(三角形和圆形分别表示推测的豆蔻酰化位点和高度保守位点, 正方形表示推测的微丝切割位点, 箭头示推测的微管结合位点, 红框为质膜静电互作区); (D) DREPP蛋白中央区域氨基酸序列; (E) DREPP蛋白羧基端区域氨基酸序列
Figure 2Schematic diagram of functional domains of plant DREPP proteins
(A) General functional domains of DREPP proteins; (B) Functional domains of AtPCaP2/AtMAP18; (C) Amino acid sequences of N-terminal domain of DREPP proteins (The speculative N-myristoylation site and highly conserved sites are labelled with a triangle and circles respectively, squares represent speculative microfilament cutting sites, arrows represent speculative microtubule binding sites, the region interacting electrostatically with the plasma membrane is highlighted in a red box); (D) Amino acid sequences of the central domain of DREPP proteins; (E) Amino acid sequences of C-terminal domain of DREPP proteins
3 DREPP参与Ca2+介导的PtdInsPs信号转导途径
PtdInsPs是植物生物膜中含量较低的一类磷脂, 对于细胞信号转导、细胞结构维持、膜流动性调控以及物质转运等过程具有重要作用(Kaadige and Ayer, 2006)。正常情况下, AtDREPPs可以与磷脂酰肌醇- 3,4-二磷酸(phosphatidylinositol 3,4-bisphosphate, PtdIns(3,4)P2)、磷脂酰肌醇-3,5-二磷酸(PtdIns (3,5)P2)和磷脂酰肌醇-4,5-二磷酸 (PtdIns(4,5)P2)肌醇环上的磷酸根稳定结合, 形成AtDREPPs/ PtdInsPs复合体, 并结合到细胞膜上(Nagasaki-Takeuchi et al., 2008; Kato et al., 2010a)。当植物细胞感受到外界信号时, 细胞膜上的钙离子通道被激活, 胞外Ca2+内流, 导致细胞内Ca2+浓度升高, Ca2+与胞质中CaM结合, 形成大量Ca2+/CaM复合体。Ca2+/ CaM复合体与PtdInsP2竞争性结合AtDREPPs的N23域, 促使AtDREPPs与PtdInsP2解离(Nagasaki-Takeuchi et al., 2008; Kato et al., 2010a)。游离的PtdInsP2被磷脂酶C (phospholipase C, PLC)水解为肌醇三磷酸(inositol trisphosphates, InsP3)和二酰基甘油(diacyl glycerol, DAG)。InsP3刺激胞外Ca2+进入细胞, 使胞内Ca2+浓度上升并进一步参与信号转导(Hunt et al., 2004; 邱丽丽等, 2017); DAG迅速被磷酸化为磷脂酸(phosphatidic acid, PA), PA作为信号分子, 参与多种信号转导途径(Testerink and Munnik, 2005; Zhang et al., 2014)。游离的AtDREPPs从质膜转至细胞质中, 参与调控植物生长发育及逆境胁迫应答。4 DREPP抑制微管聚合
微管在植物生长发育过程中发挥至关重要的作用, 微管的稳定性、组织结构和动力学都受到微管调控蛋白的调控(Buschmann and Lloyd, 2008)。AtPCaP1可在体内外直接与微管结合并使微管不稳定, 从而促进微管的体外解聚(Li et al., 2011)。与AtPCaP1同属于拟南芥DREPP蛋白家族的AtPCaP2也是微管结合蛋白, 其通过去稳定微管骨架调控拟南芥细胞的各向异性生长(Wang et al., 2007)。对水稻OsDREPP2的研究表明, 微管蛋白和三磷酸鸟苷(guanosine triphosphate, GTP)共孵育时, 微管聚合程度增加。当OsDREPP2加入到微管蛋白中, 微管聚合明显受到抑制, 且OsDREPP2浓度越高抑制作用越明显(Yamada et al., 2015)。盐地鼠尾粟(Sporobdus virginicus)的SvDREPP1与OsDREPP2一样, 也可作为微管去稳定因子抑制微管聚合(Theerawitaya et al., 2018)。5 DREPP调控植物生长发育
AtPCaP2在拟南芥各器官均有不同程度的表达, 但在根/根毛、下胚轴和花粉/花粉管中的表达量较高(Wang et al., 2007; Kato et al., 2010a), 这暗示AtPCaP2可能在根毛和花粉管等顶端生长的细胞中发挥重要作用。5.1 调节根定向生长
植物根部细胞周质微管动态变化调节细胞伸长和形态建成, 对于根部极性生长具有重要作用。AtPCaP2在根部组织中主要在伸长区表达, 而在根尖没有表达(Wang et al., 2007)。野生型拟南芥根表皮细胞微管的排列与细胞长轴方向平行, 而AtPCaP2过表达植株的根部微管排列较乱, 呈斜向或纵向排列(Wang et al., 2007)。对根尖微管列阵数目的分析表明, 过表达AtPCaP2对细胞周期和细胞分裂无影响。在拟南芥中, 过表达AtPCaP2会导致根部呈偏左性生长。用微管解聚试剂黄草消(oryzalin)处理后, 过表达AtPCaP2拟南芥植株与野生型植株根部都发生偏左性生长, 过表达植株偏左性生长更明显, 而atpcap2突变体植株保持正常向地性生长(Wang et al., 2018a)。因此, AtPCaP2可能是作为微管去稳定因子, 可能通过与周质微管结合来抑制微管蛋白聚合, 从而调控周质微管排列, 调节根部定向生长。5.2 调控根毛生长
AtPCaP2在拟南芥根毛中的表达量较高, 这暗示其在根毛中发挥重要作用(Wang et al., 2007; Kato et al., 2010a)。atpcap2突变体与AtPCaP2过表达植株的根毛比野生型短。AtPCaP2过表达植株的根毛分叉数量明显增加, 并且根毛顶端和亚顶端区域生长异常, 且根毛中短微丝片段的数量是野生型根毛的2倍, 而atpcap2突变体植株根毛中短微丝片段的数量仅有野生型的1/2 (Zhu et al., 2013)。由此推测, AtPCaP2通过调节根毛顶端和亚顶端区域的微丝组织动态调控根毛生长。ROPs (Rho-related GTPase from plants)是一类植物特有的调控根毛生长发育的小G蛋白(Kost, 2008; Lee and Yang, 2008)。拟南芥有11个ROPs, 其中ROP2、ROP4和ROP6参与调控根毛的生长(Jones et al., 2002)。ROP4和ROP6在拟南芥根毛的生长调控中与ROP2的功能相反。rop2突变体和ROP2过表达体的根毛生长表型分别与atpcap2突变体和AtPCaP2过表达体一致, 这暗示AtPCaP2可能与ROP2参与相同的信号途径, 从而调控根毛生长。细胞质中的AtRhoGDI1 (RhoGTPase GDP dissociation inhibitor 1)也被称为SCN1 (Super centipede 1), 可与ROPs结合形成复合体, 并抑制ROPs的活性(Carol et al., 2005)。体内与体外实验证实, AtPCaP2可与无活性的ROP2结合, 正调控ROP2的活性。进一步实验证明, AtPCaP2可与AtRhoGDI1 (SCN1)竞争结合无活性的ROP2, 解除AtRhoGDI1 (SCN1)对ROP2的负调控。AtPCaP2的N23域缺失导致其与ROP2结合能力丧失。由此推测, N23域是AtPCaP2与ROP2相互作用的关键结构域(Kang et al., 2017)。然而, AtPCaP2微丝切割活性位点(每个VEEKK基序中的第1个E和第2个K)的突变并未影响其与ROP2的结合, 说明AtPCaP2切割微丝的活性变化不影响其与ROP2的结合。由此推测, AtPCaP2与AtRhoGDI1/ SCN1竞争结合无活性的ROP2, 有助于无活性的ROP2转变为有活性, 激活ROP2下游信号途径, 从而调节根毛的极性生长(Kang et al., 2017)。
5.3 调控花粉管生长
AtPCaP1和AtPCaP2在拟南芥花粉与花粉管中的表达量均很高。atpcap2突变体与AtPCaP2过表达体的花粉管扭曲, 生长方向异常, 甚至环绕胚珠生长, 无法完成受精, 从而造成结实率下降。atpcap2突变体与AtPCaP2过表达体花粉管顶端及亚顶端区内微丝的组织排列方式发生改变。此外, 免疫共沉淀实验结果表明, AtPCaP1可直接与微丝结合并具有微弱的微丝切割活性; 在10 μmol∙L-1 Ca2+的条件下, 微丝切割活性明显增强。与野生型相比, atpcap1突变体植株花粉管亚顶端的微丝束更大, 且密度更高, 这表明AtPCaP1的缺失导致花粉管亚顶端微丝无法被切割。同时, 将AtPCaP1中微丝与Ca2+结合关键位点(分别为Asn5和184-188位VEEKK域中E185和K188)突变后, 回补atpcap1突变体, 无法恢复其正常花粉管表型。由此推测, AtPCaP1和AtPCaP2通过调节花粉管顶端及亚顶端微丝的组织排列和动态变化调控花粉管顶端生长(Zhu et al., 2013; Qin et al., 2014)。5.4 抑制下胚轴生长
AtPCaP2也是微管结合蛋白, 高浓度AtPCaP2抑制微管聚合。AtPCaP2过表达植株的下胚轴细胞内微管排列混乱; 下胚轴表皮细胞肿胀, 细胞伸长被抑制; 铺板细胞突起数明显减少, 细胞边缘光滑(Wang et al., 2007)。此外, 与野生型相比, AtPCaP1过表达植株的下胚轴细胞更短, 细胞内微管排列方向异常。在1 mmol∙L-1 CaCl2的条件下, AtPCaP1过表达植株的下胚轴细胞内微管明显被破坏, 野生型植株的微管受到轻微损伤, 而atpcap1突变体植株的微管无变化。由此推测, AtPCaP1和AtPCaP2都可以依赖Ca2+结合微管, 破坏微管的稳定性, 抑制下胚轴细胞伸长(Li et al., 2011)。6 DREPP参与植物胁迫应答
6.1 干旱胁迫应答
干旱是农业生产中一种常见的非生物胁迫, 严重制约植物生长发育和生产力(Fleta-Soriano and Munné- Bosch, 2016)。植物可以通过细胞膜上特异的受体和信号转导系统感知并转导外界环境中的干旱胁迫信号, 激活或抑制下游相关基因(如RD29A、KIN1和KIN2)的表达, 进而抵御干旱胁迫。随着干旱胁迫时间的延长, AtPCaP2在子叶、下胚轴、主根和侧根的表达逐渐升高。在干旱胁迫下, AtPCaP2过表达植株的种子萌发率高于野生型和atpcap2突变体。这表明, 在干旱胁迫下, AtPCaP2对种子的萌发具有正向调节作用。研究表明, SnRK2s (SNF1-related kinase 2s)和PRs (pathogenesis-related)分别通过ABA和SA介导的信号通路影响植物的耐旱性。ABA能够激活SnRK2s等蛋白激酶, 诱导ABA依赖的转录因子的表达, 如ABIs (abscisic acid insensitive)和ABFs (abscisic acid responsive element binding factor) (Trivedi et al., 2016), 进而促进下游ABA响应基因表达, 抵御逆境。此外, SA响应基因(如PR1、PR2和PR5)也参与植物对干旱胁迫的响应(Ichimura et al., 2000; Liu et al., 2013)。外源施加ABA和SA诱导AtPCaP2大量表达。在AtPCaP2过表达植株中, 受ABA调控的干旱应答基因(SnRK2.2、SnRK2.3、SnRK2.6、ABF1、ABF2、ABF3和ABF4), 受SA调控的干旱应答基因(PR1、PR2和PR5), 以及RD29A、KIN1和KIN2等其它干旱应答基因, 在ABA、SA和干旱处理后表达量都明显上调(Wang et al., 2018b)。研究表明, 水稻含有B-box锌指结构域的OsBBX25蛋白在干旱胁迫时可上调干旱应答基因KIN1、RD22和RD29A的表达, 参与植物逆境响应(刘焱等, 2012)。而DREPP蛋白同样可以调控胁迫相关基因的表达。由此推测, AtPCaP2可能通过DREPP功能结构域参与ABA和SA信号通路调节的干旱胁迫应答。6.2 低温胁迫应答
低温限制植物在地球上的分布, 减少作物产量, 是一种常见的非生物胁迫(Shi et al., 2018)。低温胁迫能快速诱导CBFs (C-repeat binding factors)转录因子的表达, 从而激活下游冷应答(cold-regulated, COR)基因的表达(Zhu, 2016)。随着低温处理时间的延长, 拟南芥根、子叶、叶片和花中的AtPCaP2表达量逐渐上升。在低温胁迫条件下, 过表达AtPCaP2的拟南芥植株耐寒能力明显高于野生型和atpcap2突变体。在低温和ABA处理条件下, AtPCaP2分别正调控低温响应基因(CBF1、CBF3、KIN1、KIN2、RD29A和COR15B)和ABA响应基因(SnRK2.2、SnRK2.3和ABFs)的表达, 负调控低温响应基因CBF2的表达。CBF1、CBF2和CBF3是植物抵抗低温胁迫的主要调控因子。CBF1和CBF3正向调控下游冷胁迫响应基因的表达, 而CBF2是冷信号负调节因子, 负调节CBF1和CBF3在低温胁迫下的表达(Novillo et al., 2004)。当植株受到低温胁迫时, CBFs被激活, 调控其下游CORs基因(COR15A、COR15B、RD29A、RD29B、KIN1和KIN2等)的表达(Novillo et al., 2007), 从而提高植株耐寒性。由此推测, AtPCaP2可能参与调控CBF-COR信号途径和ABA依赖的低温应答途径(Wang et al., 2018a)。6.3 免疫应答
对烟草DREPP家族成员PtBP1的研究表明, 烟草花叶病毒(tobacco mosaic virus, TMV)侵染烟草植株3天后, PeaT1沉默植株叶片的损伤程度约为野生型烟草的3倍, PeaT1的沉默减弱了烟草对TMV的抗性。PeaT1是一种来自真菌病原体细极链格孢菌的蛋白诱导因子, 可以诱导包括烟草在内的各种植物的防御反应并增强抗病性(Mao et al., 2010)。经PeaT1处理后, 与野生型相比, PtBP1沉默植物中PR1a (pathogenesis-related protein 1a)、PR1b (pathogene- sis-related protein 1b)和PAL (phenylalanine am- monia lyase)的表达量显著降低, 推测PtBP1可能通过介导病原相关基因的表达提高植株的抗病性(Meng et al., 2018)。7 展望
DREPPs通过Ca2+依赖的方式调节微丝和微管的组织排列, 从而调控植物的生长发育; 并可通过调控ABA和SA等信号途径参与植物应答干旱和低温等逆境胁迫过程(图3), 但是其详细的调控机理目前并不清楚。近年来, 我们利用蛋白质组学和分子生物学技术(喻娟娟和戴绍军, 2009), 发现星星草(Puccinellia tenuiflora)根中DERPP蛋白质丰度和基因表达水平在应答盐碱和H2O2胁迫时均显著上升(Zhang et al., 2013; Zhao et al., 2016; Yu et al., 2018), 暗示DREPP在应答盐碱和氧化胁迫时也发挥重要作用。然而, DERPP在花粉管和下胚轴中如何调控微管和微丝的动态变化? 与哪些成员相互作用调节ABA信号通路进而参与非生物胁迫应答? 其调节哪些信号通路来影响植物免疫应答? 这些科学问题都有待进一步探讨。因此, 阐明DREPP家族成员在植物发育与胁迫应答过程中的分子调控机理对于解析植物逆境应答机制和提高作物产量具有重要意义。图3
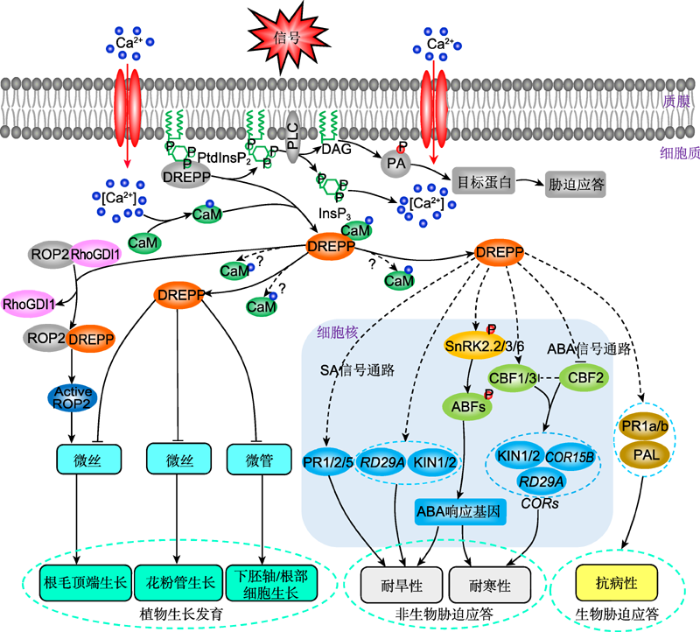 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3DREPP参与调控植物生长与逆境胁迫应答
ABA: 脱落酸; ABF: 脱落酸响应元件结合因子; CaM: 钙调素蛋白; CBF1: C-重复结合因子1; CBF2: C-重复结合因子2; CBF3: C-重复结合因子3; COR: 冷调节基因; COR15B: 冷调节基因15B; DAG: 二酰基甘油; DREPP: 发育调节的质膜多肽; InsP3: 肌醇三磷酸; KIN1: 逆境应答蛋白1; KIN2: 逆境应答蛋白2; PA: 磷脂酸; PLC: 磷脂酶C; PR1: 病程相关因子1; PR2: 病程相关因子2; PR5: 病程相关因子5; PtdInsP2: 磷脂酰肌醇二磷酸; RD29A: 干旱应答基因29A; RhoGDI1: RhoGTP酶GDP解离抑制因子1; ROP2: 植物来源的Rho相关GTP酶; SA: 水杨酸; SnRK2.2: 蔗糖非酵解相关蛋白激酶2.2; SnRK2.3: 蔗糖非酵解相关蛋白激酶2.3; SnRK2.6: 蔗糖非酵解相关蛋白激酶2.6
Figure 3DREPPs are involved in the regulation of plant growth and stress response
ABA: Abscisic acid; ABF: Abscisic acid responsive element binding factor; CaM: Calmodulin protein; CBF1: C-repeat binding factor 1; CBF2: C-repeat binding factor 2; CBF3: C-repeat binding factor 3; COR: Cold-regulated gene; COR15B: Cold-regulated gene 15B; DAG: Diacyl glycerol; DREPP: Developmentally regulated plasma membrane polypeptide; InsP3: Inositol trisphosphates; KIN1: Stress-responsive protein 1; KIN2: Stress-responsive protein 2; PA: Phosphatidic acid; PLC: Phospholipase C; PR1: Pathogenesis-related factor 1; PR2: Pathogenesis-related factor 2; PR5: Pathogenesis-related factor 5; PtdInsP2: Phosphatidylinositol phosphate 2; RD29A: Responsive to dessication gene 29A; RhoGDI1: RhoGTPase GDP dissociation inhibitor 1; ROP2: Rho-related GTPase from plants; SA: Salicylic acid; SnRK2.2: SNF1-related kinase 2.2; SnRK2.3: SNF1-related kinase 2.3; SnRK2.6: SNF1-related kinase 2.6
附表1 植物DREPP蛋白质信息
Appendix table 1 Information of DREPP protein in plantshttp://www.chinbullbotany.com/fileup/1674-3466/PDF/t19014.xlsx
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 5]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 1]
[本文引用: 2]
[本文引用: 7]
[本文引用: 3]
[本文引用: 2]
[本文引用: 4]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 3]
水稻含有B-box锌指结构域的OsBBX25蛋白参与植物对非生物胁迫的响应
1
2012
... 干旱是农业生产中一种常见的非生物胁迫, 严重制约植物生长发育和生产力(
植物质膜蛋白质组的逆境应答研究进展
1
2017
... PtdInsPs是植物生物膜中含量较低的一类磷脂, 对于细胞信号转导、细胞结构维持、膜流动性调控以及物质转运等过程具有重要作用(
植物蛋白质组学研究若干重要进展
1
2009
... DREPPs通过Ca2+依赖的方式调节微丝和微管的组织排列, 从而调控植物的生长发育; 并可通过调控ABA和SA等信号途径参与植物应答干旱和低温等逆境胁迫过程(
Insights into corn genes derived from large-scale cDNA sequencing
1
2009
... (Continued)
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 84 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013610402.1 | - |
| 85 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013596813.1 | - |
| 86 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013619975.1 | - |
| 87 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD40021.1 | - |
| 88 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD37718.1 | - |
| 89 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD25026.1 | - |
| 90 | 萝卜 | Raphanus sativus | Rs | 十字花科 | 萝卜属 | XP_018471954.1 | - |
| 91 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | BAA99394.1 | Yuasa and Mae- shima, 2000 |
| 92 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018437382.1 | - |
| 93 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018455198.1 | - |
| 94 | 山嵛菜 | Eutrema salsugineum | Es | 十字花科 | 山嵛菜属 | XP_006413890.1 | - |
| 95 | 山嵛菜 | E. salsugineum | Es | 十字花科 | 山嵛菜属 | XP_024011088.1 | - |
| 96 | 荠菜 | Capsella rubella | Cr | 十字花科 | 荠属 | XP_006284474.1 | - |
| 97 | 荠菜 | C. rubella | Cr | 十字花科 | 荠属 | XP_006279705.1 | - |
| 98 | 亚麻荠 | Camelina sativa | Csa | 十字花科 | 亚麻荠属 | XP_010434307.1 | - |
| 99 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010441852.1 | - |
| 100 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010445250.1 | - |
| 101 | 天蓝遏蓝菜 | Noccaea caerulescens | Nc | 十字花科 | 菥蓂属 | JAU60665.1 | - |
| 102 | 拟南芥 | Arabidopsis thaliana | At | 十字花科 | 拟南芥属 | Q96262.1 | |
| 103 | 拟南芥 | A. thaliana | At | 十字花科 | 拟南芥属 | NP_568636.1 | |
| 104 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002869944.1 | - |
| 105 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002863557.1 | - |
| 106 | 番木瓜 | Carica papaya | Cp | 番木瓜科 | 番木瓜属 | XP_021901673.1 | - |
| 107 | 凤梨 | Ananas comosus | Aco | 凤梨科 | 凤梨属 | OAY80473.1 | |
| 108 | 深圳拟兰 | Apostasia shenzhenica | As | 兰科 | 拟兰属 | PKA54423.1 | - |
| 109 | - | Sarocalamus faberi | Sf | 兰科 | - | ATE87948.1 | - |
| 110 | 小果野芭蕉 | Musa acuminata subsp. malaccensis | Ma | 芭蕉科 | 芭蕉属 | XP_009409470.1 | - |
| 111 | 小兰屿蝴蝶兰 | Phalaenopsis equestris | Phe | 兰科 | 蝴蝶兰属 | XP_020574860.1 | - |
| 112 | 铁皮石斛 | Dendrobium catenatum | Dc | 兰科 | 石斛属 | XP_020688760.1 | - |
| 113 | 玉米 | Zea mays | Zm | 禾本科 | 玉蜀黍属 | XP_020397542.1 | - |
| 114 | 玉米 | Z. mays | Zm | 禾本科 | 玉蜀黍属 | ACG37450.1 | |
| 115 | 水稻 | Oryza sativa | Os | 禾本科 | 稻属 | NP_001042508.1 | |
| 116 | 水稻 | O. sativa | Os | 禾本科 | 稻属 | NP_001046572.1 | |
| 117 | 盐地鼠尾粟 | Sporobolus virginicus | Sv | 禾本科 | 鼠尾粟属 | LC342075.1 | |
| 118 | 二穗短柄草 | Brachypodium distachyon | Bd | 禾本科 | 短柄草属 | XP_003572598.1 | - |
| 119 | 高粱 | Sorghum bicolor | Sb | 禾本科 | 高粱属 | XP_002453713.1 | - |
| 120 | - | Dichanthelium oligosanthes | Do | 禾本科 | 二型花属 | OEL25601.1 |
Arabidopsis mutants and the network of microtubule-associated functions
1
2008
... 微管在植物生长发育过程中发挥至关重要的作用, 微管的稳定性、组织结构和动力学都受到微管调控蛋白的调控(
A RhoGDP dissociation inhibitor spatially regulates growth in root hair cells
1
2005
... ROPs (Rho-related GTPase from plants)是一类植物特有的调控根毛生长发育的小G蛋白(
The genome of the recently domesticated crop plant sugar beet (Beta vulgaris)
2
2014
... (Continued)
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 42 | 大豆 | Glycine max | Gm | 豆科 | 大豆属 | XP_003546380.1 | Matthews et al., 2013 |
| 43 | 大豆 | G. max | Gm | 豆科 | 大豆属 | NP_001237575.1 | - |
| 44 | 欧洲栓皮栎 | Quercus suber | Qs | 壳斗科 | 栎属 | XP_023919313.1 | - |
| 45 | 土瓶草 | Cephalotus follicularis | Cf | 土瓶草科 | 土瓶草属 | GAV80098.1 | |
| 46 | 胡桃 | Juglans regia | Jr | 胡桃科 | 胡桃属 | XP_018814574.1 | - |
| 47 | 甜橙 | Citrus sinensis | Cs | 芸香科 | 柑橘属 | XP_006468602.1 | - |
| 48 | 克里曼丁桔 | C. clementina | Ccl | 芸香科 | 柑橘属 | XP_006448565.1 | - |
| 49 | 大桉 | Eucalyptus grandis | Eg | 桃金娘科 | 桉属 | XP_010051796.1 | - |
| 50 | 毛果杨 | Populus trichocarpa | Pt | 杨柳科 | 杨属 | XP_006385859.1 | - |
| 51 | 毛果杨 | P. trichocarpa | Pt | 杨柳科 | 杨属 | XP_006368838.1 | - |
| 52 | 蓖麻 | Ricinus communis | Rc | 大戟科 | 蓖麻属 | XP_015583044.1 | - |
| 53 | 麻疯树 | Jatropha curcas | Jc | 大戟科 | 麻疯树属 | XP_012088609.1 | - |
| 54 | 木薯 | Manihot esculenta | Me | 大戟科 | 木薯属 | XP_021602838.1 | - |
| 55 | 橡胶树 | Hevea brasiliensis | Hb | 大戟科 | 橡胶树属 | XP_021686108.1 | - |
| 56 | 糙叶山黄麻 | Parasponia andersonii | Pan | 榆科 | 南山黄麻属 | PON44636.1 | - |
| 57 | 异色山黄麻 | Trema orientale | To | 椴树科 | 山黄麻属 | PON83811.1 | - |
| 58 | 长蒴黄麻 | Corchorus olitorius | Co | 椴树科 | 黄麻属 | OMO77743.1 | - |
| 59 | 博落回 | Macleaya cordata | Mc | 罂粟科 | 博落回属 | OVA01374.1 | |
| 60 | 莲 | Nelumbo nucifera | Nn | 睡莲科 | 莲属 | XP_010260837.1 | - |
| 61 | 川桑 | Morus notabilis | Mn | 桑科 | 桑属 | XP_010094633.1 | - |
| 62 | 葡萄 | Vitis vinifera | Vv | 葡萄科 | 葡萄属 | XP_002263090.1 | - |
| 63 | 菠菜 | Spinacia oleracea | So | 藜科 | 菠菜属 | KNA18787.1 | |
| 64 | 甜菜 | Beta vulgaris subsp. vulgaris | Bv | 藜科 | 甜菜属 | XP_010684190.2 | - |
| 65 | 甜菜 | B. vulgaris subsp. vulgaris | Bv | 藜科 | 甜菜属 | KMT06194.1 | |
| 66 | 无油樟 | Amborella trichopoda | Atr | 无油樟科 | 无油樟属 | XP_006859057.2 | - |
| 67 | 醉蝶花 | Tarenaya hassleriana | Th | 醉蝶花科 | 白花菜属 | XP_010537158.1 | - |
| 68 | 芜菁 | Brassica rapa | Br | 十字花科 | 芸苔属 | AHV84997.1 | - |
| 69 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | XP_009133577.1 | - |
| 70 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | RIA04026.1 | - |
| 71 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | XP_009113907.1 | - |
| 72 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | XP_009101693.1 | - |
| 73 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013660923.1 | - |
| 74 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | CDY66354.1 | - |
| 75 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013672014.1 | - |
| 76 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_022572533.1 | - |
| 77 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013717493.1 | - |
| 78 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013659410.1 | - |
| 79 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013730570.1 | - |
| 80 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013715242.1 | - |
| 81 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013733201.1 | - |
| 82 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | CDY63708.1 | - |
| 83 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013602975.1 | - |
...
Stress memory and the inevitable effects of drought: a physiological perspective
1
2016
... 干旱是农业生产中一种常见的非生物胁迫, 严重制约植物生长发育和生产力(
Genome of the pitcher plant Cephalotus reveals genetic changes associated with carnivory
1
2017
... (Continued)
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 42 | 大豆 | Glycine max | Gm | 豆科 | 大豆属 | XP_003546380.1 | Matthews et al., 2013 |
| 43 | 大豆 | G. max | Gm | 豆科 | 大豆属 | NP_001237575.1 | - |
| 44 | 欧洲栓皮栎 | Quercus suber | Qs | 壳斗科 | 栎属 | XP_023919313.1 | - |
| 45 | 土瓶草 | Cephalotus follicularis | Cf | 土瓶草科 | 土瓶草属 | GAV80098.1 | |
| 46 | 胡桃 | Juglans regia | Jr | 胡桃科 | 胡桃属 | XP_018814574.1 | - |
| 47 | 甜橙 | Citrus sinensis | Cs | 芸香科 | 柑橘属 | XP_006468602.1 | - |
| 48 | 克里曼丁桔 | C. clementina | Ccl | 芸香科 | 柑橘属 | XP_006448565.1 | - |
| 49 | 大桉 | Eucalyptus grandis | Eg | 桃金娘科 | 桉属 | XP_010051796.1 | - |
| 50 | 毛果杨 | Populus trichocarpa | Pt | 杨柳科 | 杨属 | XP_006385859.1 | - |
| 51 | 毛果杨 | P. trichocarpa | Pt | 杨柳科 | 杨属 | XP_006368838.1 | - |
| 52 | 蓖麻 | Ricinus communis | Rc | 大戟科 | 蓖麻属 | XP_015583044.1 | - |
| 53 | 麻疯树 | Jatropha curcas | Jc | 大戟科 | 麻疯树属 | XP_012088609.1 | - |
| 54 | 木薯 | Manihot esculenta | Me | 大戟科 | 木薯属 | XP_021602838.1 | - |
| 55 | 橡胶树 | Hevea brasiliensis | Hb | 大戟科 | 橡胶树属 | XP_021686108.1 | - |
| 56 | 糙叶山黄麻 | Parasponia andersonii | Pan | 榆科 | 南山黄麻属 | PON44636.1 | - |
| 57 | 异色山黄麻 | Trema orientale | To | 椴树科 | 山黄麻属 | PON83811.1 | - |
| 58 | 长蒴黄麻 | Corchorus olitorius | Co | 椴树科 | 黄麻属 | OMO77743.1 | - |
| 59 | 博落回 | Macleaya cordata | Mc | 罂粟科 | 博落回属 | OVA01374.1 | |
| 60 | 莲 | Nelumbo nucifera | Nn | 睡莲科 | 莲属 | XP_010260837.1 | - |
| 61 | 川桑 | Morus notabilis | Mn | 桑科 | 桑属 | XP_010094633.1 | - |
| 62 | 葡萄 | Vitis vinifera | Vv | 葡萄科 | 葡萄属 | XP_002263090.1 | - |
| 63 | 菠菜 | Spinacia oleracea | So | 藜科 | 菠菜属 | KNA18787.1 | |
| 64 | 甜菜 | Beta vulgaris subsp. vulgaris | Bv | 藜科 | 甜菜属 | XP_010684190.2 | - |
| 65 | 甜菜 | B. vulgaris subsp. vulgaris | Bv | 藜科 | 甜菜属 | KMT06194.1 | |
| 66 | 无油樟 | Amborella trichopoda | Atr | 无油樟科 | 无油樟属 | XP_006859057.2 | - |
| 67 | 醉蝶花 | Tarenaya hassleriana | Th | 醉蝶花科 | 白花菜属 | XP_010537158.1 | - |
| 68 | 芜菁 | Brassica rapa | Br | 十字花科 | 芸苔属 | AHV84997.1 | - |
| 69 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | XP_009133577.1 | - |
| 70 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | RIA04026.1 | - |
| 71 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | XP_009113907.1 | - |
| 72 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | XP_009101693.1 | - |
| 73 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013660923.1 | - |
| 74 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | CDY66354.1 | - |
| 75 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013672014.1 | - |
| 76 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_022572533.1 | - |
| 77 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013717493.1 | - |
| 78 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013659410.1 | - |
| 79 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013730570.1 | - |
| 80 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013715242.1 | - |
| 81 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013733201.1 | - |
| 82 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | CDY63708.1 | - |
| 83 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013602975.1 | - |
Cloning of a cDNA encoding a developmentally regulated 22 kDa polypeptide from tobacco leaf plasma membrane
3
1996
... 人们最早从烟草(Nicotiana tabacum)中克隆到DREPP编码基因(
... DREPP protein family members in plants
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 1 | 番茄 | Solanum lycopersicum | Sl | 茄科 | 茄属 | XP_004248413.1 | - |
| 2 | 野生潘那利番茄 | S. pennellii | Sp | 茄科 | 茄属 | XP_015055582.1 | - |
| 3 | 马铃薯 | S. tuberosum | St | 茄科 | 茄属 | XP_006352621.1 | - |
| 4 | 风铃辣椒 | Capsicum baccatum | Cb | 茄科 | 辣椒属 | PHT35828.1 | |
| 5 | 黄灯笼辣椒 | C. chinense | Cch | 茄科 | 辣椒属 | PHU04552.1 | |
| 6 | 辣椒 | C. annuum | Can | 茄科 | 辣椒属 | PHT70067.1 | |
| 7 | 本氏烟草 | Nicotiana benthamiana | Nb | 茄科 | 烟草属 | ASW15993.1 | |
| 8 | 渐狭叶烟草 | N. attenuata | Na | 茄科 | 烟草属 | XP_019226343.1 | - |
| 9 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69901.1 | |
| 10 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69900.1 | |
| 11 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016438748.1 | - |
| 12 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | X95957.1 | |
| 13 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016451342.1 | - |
| 14 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_018631801.1 | - |
| 15 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_009600368.1 | - |
| 16 | 林烟草 | N. sylvestris | Ns | 茄科 | 烟草属 | XP_009763203.1 | - |
| 17 | 中粒咖啡 | Coffea canephora | Cca | 茜草科 | 咖啡属 | CDP04109.1 | - |
| 18 | 小粒咖啡 | C. arabica | Car | 茜草科 | 咖啡属 | XP_027089181.1 | - |
| 19 | 木犀榄 | Olea europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022847863.1 | - |
| 20 | 木犀榄 | O. europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022870576.1 | - |
| 21 | 芝麻 | Sesamum indicum | Si | 胡麻科 | 胡麻属 | XP_011085144.1 | - |
| 22 | 中华猕猴桃 | Actinidia chinensis var. chinensis | Ac | 猕猴桃科 | 猕猴桃属 | PSS11063.1 | - |
| 23 | 苹果 | Malus domestica | Md | 蔷薇科 | 苹果属 | XP_008383239.1 | - |
| 24 | 月季 | Rosa chinensis | Rch | 蔷薇科 | 蔷薇属 | XP_024156781.1 | - |
| 25 | 桃 | Prunus persica | Pp | 蔷薇科 | 桃属 | XP_007212006.1 | - |
| 26 | 樱花 | P. yedoensis var. nudiflora | Py | 蔷薇科 | 桃属 | PQP97549.1 | - |
| 27 | 甜樱桃 | P. avium | Pa | 蔷薇科 | 桃属 | XP_021822163.1 | - |
| 28 | 刺苞菜蓟 | Cynara cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | XP_024964532.1 | - |
| 29 | 刺苞菜蓟 | C. cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | KVH87676.1 | |
| 30 | 莴苣 | Lactuca sativa | Ls | 菊科 | 莴苣属 | XP_023745485.1 | - |
| 31 | 莴苣 | L. sativa | Ls | 菊科 | 莴苣属 | XP_023768949.1 | - |
| 32 | 向日葵 | Helianthus annuus | Ha | 菊科 | 向日葵属 | XP_021982589.1 | - |
| 33 | 向日葵 | H. annuus | Ha | 菊科 | 向日葵属 | XP_022009496.1 | - |
| 34 | 黄花蒿 | Artemisia annua | Aa | 菊科 | 蒿亚属 | PWA97158.1 | |
| 35 | 蒺藜苜蓿 | Medicago truncatula | Mt | 豆科 | 苜蓿属 | XP_013463310.1 | - |
| 36 | 地三叶 | Trifolium subterraneum | Ts | 豆科 | 三叶草属 | GAU45950.1 | |
| 37 | 鹰嘴豆 | Cicer arietinum | Ca | 豆科 | 鹰嘴豆属 | Q9SMK5 | |
| 38 | 野生落花生 | Arachis ipaensis | Ai | 豆科 | 落花生属 | XP_016197976.1 | - |
| 39 | 蔓花生 | A. duranensis | Ad | 豆科 | 落花生属 | XP_015959777.1 | - |
| 40 | 百脉根 | Lotus japonicus | Lj | 豆科 | 百脉根属 | AFK34640.1 | - |
| 41 | 菜豆 | Phaseolus vulgaris | Pv | 豆科 | 菜豆属 | AGV54625.1 | - |
... AtPCaP2在拟南芥各器官均有不同程度的表达, 但在根/根毛、下胚轴和花粉/花粉管中的表达量较高(
Plasma membrane-associated cation- binding protein 1-like protein negatively regulates inter-cellular movement of BaMV
1
2017
... DREPP protein family members in plants
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 1 | 番茄 | Solanum lycopersicum | Sl | 茄科 | 茄属 | XP_004248413.1 | - |
| 2 | 野生潘那利番茄 | S. pennellii | Sp | 茄科 | 茄属 | XP_015055582.1 | - |
| 3 | 马铃薯 | S. tuberosum | St | 茄科 | 茄属 | XP_006352621.1 | - |
| 4 | 风铃辣椒 | Capsicum baccatum | Cb | 茄科 | 辣椒属 | PHT35828.1 | |
| 5 | 黄灯笼辣椒 | C. chinense | Cch | 茄科 | 辣椒属 | PHU04552.1 | |
| 6 | 辣椒 | C. annuum | Can | 茄科 | 辣椒属 | PHT70067.1 | |
| 7 | 本氏烟草 | Nicotiana benthamiana | Nb | 茄科 | 烟草属 | ASW15993.1 | |
| 8 | 渐狭叶烟草 | N. attenuata | Na | 茄科 | 烟草属 | XP_019226343.1 | - |
| 9 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69901.1 | |
| 10 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69900.1 | |
| 11 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016438748.1 | - |
| 12 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | X95957.1 | |
| 13 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016451342.1 | - |
| 14 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_018631801.1 | - |
| 15 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_009600368.1 | - |
| 16 | 林烟草 | N. sylvestris | Ns | 茄科 | 烟草属 | XP_009763203.1 | - |
| 17 | 中粒咖啡 | Coffea canephora | Cca | 茜草科 | 咖啡属 | CDP04109.1 | - |
| 18 | 小粒咖啡 | C. arabica | Car | 茜草科 | 咖啡属 | XP_027089181.1 | - |
| 19 | 木犀榄 | Olea europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022847863.1 | - |
| 20 | 木犀榄 | O. europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022870576.1 | - |
| 21 | 芝麻 | Sesamum indicum | Si | 胡麻科 | 胡麻属 | XP_011085144.1 | - |
| 22 | 中华猕猴桃 | Actinidia chinensis var. chinensis | Ac | 猕猴桃科 | 猕猴桃属 | PSS11063.1 | - |
| 23 | 苹果 | Malus domestica | Md | 蔷薇科 | 苹果属 | XP_008383239.1 | - |
| 24 | 月季 | Rosa chinensis | Rch | 蔷薇科 | 蔷薇属 | XP_024156781.1 | - |
| 25 | 桃 | Prunus persica | Pp | 蔷薇科 | 桃属 | XP_007212006.1 | - |
| 26 | 樱花 | P. yedoensis var. nudiflora | Py | 蔷薇科 | 桃属 | PQP97549.1 | - |
| 27 | 甜樱桃 | P. avium | Pa | 蔷薇科 | 桃属 | XP_021822163.1 | - |
| 28 | 刺苞菜蓟 | Cynara cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | XP_024964532.1 | - |
| 29 | 刺苞菜蓟 | C. cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | KVH87676.1 | |
| 30 | 莴苣 | Lactuca sativa | Ls | 菊科 | 莴苣属 | XP_023745485.1 | - |
| 31 | 莴苣 | L. sativa | Ls | 菊科 | 莴苣属 | XP_023768949.1 | - |
| 32 | 向日葵 | Helianthus annuus | Ha | 菊科 | 向日葵属 | XP_021982589.1 | - |
| 33 | 向日葵 | H. annuus | Ha | 菊科 | 向日葵属 | XP_022009496.1 | - |
| 34 | 黄花蒿 | Artemisia annua | Aa | 菊科 | 蒿亚属 | PWA97158.1 | |
| 35 | 蒺藜苜蓿 | Medicago truncatula | Mt | 豆科 | 苜蓿属 | XP_013463310.1 | - |
| 36 | 地三叶 | Trifolium subterraneum | Ts | 豆科 | 三叶草属 | GAU45950.1 | |
| 37 | 鹰嘴豆 | Cicer arietinum | Ca | 豆科 | 鹰嘴豆属 | Q9SMK5 | |
| 38 | 野生落花生 | Arachis ipaensis | Ai | 豆科 | 落花生属 | XP_016197976.1 | - |
| 39 | 蔓花生 | A. duranensis | Ad | 豆科 | 落花生属 | XP_015959777.1 | - |
| 40 | 百脉根 | Lotus japonicus | Lj | 豆科 | 百脉根属 | AFK34640.1 | - |
| 41 | 菜豆 | Phaseolus vulgaris | Pv | 豆科 | 菜豆属 | AGV54625.1 | - |
Gene-specific expression and calcium activation of Arabidopsis thaliana phospholipase C isoforms
1
2004
... PtdInsPs是植物生物膜中含量较低的一类磷脂, 对于细胞信号转导、细胞结构维持、膜流动性调控以及物质转运等过程具有重要作用(
Various abiotic stresses rapidly activate Arabidopsis MAP kinases ATMPK4 and ATMPK6
1
2000
... 干旱是农业生产中一种常见的非生物胁迫, 严重制约植物生长发育和生产力(
Molecular properties of a novel, hydrophilic cation-binding protein associated with the plasma membrane
2
2007
... 人们最早从烟草(Nicotiana tabacum)中克隆到DREPP编码基因(
... (Continued)
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 84 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013610402.1 | - |
| 85 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013596813.1 | - |
| 86 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013619975.1 | - |
| 87 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD40021.1 | - |
| 88 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD37718.1 | - |
| 89 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD25026.1 | - |
| 90 | 萝卜 | Raphanus sativus | Rs | 十字花科 | 萝卜属 | XP_018471954.1 | - |
| 91 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | BAA99394.1 | Yuasa and Mae- shima, 2000 |
| 92 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018437382.1 | - |
| 93 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018455198.1 | - |
| 94 | 山嵛菜 | Eutrema salsugineum | Es | 十字花科 | 山嵛菜属 | XP_006413890.1 | - |
| 95 | 山嵛菜 | E. salsugineum | Es | 十字花科 | 山嵛菜属 | XP_024011088.1 | - |
| 96 | 荠菜 | Capsella rubella | Cr | 十字花科 | 荠属 | XP_006284474.1 | - |
| 97 | 荠菜 | C. rubella | Cr | 十字花科 | 荠属 | XP_006279705.1 | - |
| 98 | 亚麻荠 | Camelina sativa | Csa | 十字花科 | 亚麻荠属 | XP_010434307.1 | - |
| 99 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010441852.1 | - |
| 100 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010445250.1 | - |
| 101 | 天蓝遏蓝菜 | Noccaea caerulescens | Nc | 十字花科 | 菥蓂属 | JAU60665.1 | - |
| 102 | 拟南芥 | Arabidopsis thaliana | At | 十字花科 | 拟南芥属 | Q96262.1 | |
| 103 | 拟南芥 | A. thaliana | At | 十字花科 | 拟南芥属 | NP_568636.1 | |
| 104 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002869944.1 | - |
| 105 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002863557.1 | - |
| 106 | 番木瓜 | Carica papaya | Cp | 番木瓜科 | 番木瓜属 | XP_021901673.1 | - |
| 107 | 凤梨 | Ananas comosus | Aco | 凤梨科 | 凤梨属 | OAY80473.1 | |
| 108 | 深圳拟兰 | Apostasia shenzhenica | As | 兰科 | 拟兰属 | PKA54423.1 | - |
| 109 | - | Sarocalamus faberi | Sf | 兰科 | - | ATE87948.1 | - |
| 110 | 小果野芭蕉 | Musa acuminata subsp. malaccensis | Ma | 芭蕉科 | 芭蕉属 | XP_009409470.1 | - |
| 111 | 小兰屿蝴蝶兰 | Phalaenopsis equestris | Phe | 兰科 | 蝴蝶兰属 | XP_020574860.1 | - |
| 112 | 铁皮石斛 | Dendrobium catenatum | Dc | 兰科 | 石斛属 | XP_020688760.1 | - |
| 113 | 玉米 | Zea mays | Zm | 禾本科 | 玉蜀黍属 | XP_020397542.1 | - |
| 114 | 玉米 | Z. mays | Zm | 禾本科 | 玉蜀黍属 | ACG37450.1 | |
| 115 | 水稻 | Oryza sativa | Os | 禾本科 | 稻属 | NP_001042508.1 | |
| 116 | 水稻 | O. sativa | Os | 禾本科 | 稻属 | NP_001046572.1 | |
| 117 | 盐地鼠尾粟 | Sporobolus virginicus | Sv | 禾本科 | 鼠尾粟属 | LC342075.1 | |
| 118 | 二穗短柄草 | Brachypodium distachyon | Bd | 禾本科 | 短柄草属 | XP_003572598.1 | - |
| 119 | 高粱 | Sorghum bicolor | Sb | 禾本科 | 高粱属 | XP_002453713.1 | - |
| 120 | - | Dichanthelium oligosanthes | Do | 禾本科 | 二型花属 | OEL25601.1 |
The Arabidopsis Rop2 GTPase is a positive regulator of both root hair initiation and tip growth
1
2002
... ROPs (Rho-related GTPase from plants)是一类植物特有的调控根毛生长发育的小G蛋白(
The polybasic region that follows the plant homeodomain zinc finger 1 of Pf1 is necessary and sufficient for specific phosphoinositide binding
1
2006
... PtdInsPs是植物生物膜中含量较低的一类磷脂, 对于细胞信号转导、细胞结构维持、膜流动性调控以及物质转运等过程具有重要作用(
The microtubule-associated protein MAP18 affects ROP2 GTPase activity during root hair growth
2
2017
... ROPs (Rho-related GTPase from plants)是一类植物特有的调控根毛生长发育的小G蛋白(
... ).然而, AtPCaP2微丝切割活性位点(每个VEEKK基序中的第1个E和第2个K)的突变并未影响其与ROP2的结合, 说明AtPCaP2切割微丝的活性变化不影响其与ROP2的结合.由此推测, AtPCaP2与AtRhoGDI1/ SCN1竞争结合无活性的ROP2, 有助于无活性的ROP2转变为有活性, 激活ROP2下游信号途径, 从而调节根毛的极性生长(
The Ca 2+-binding protein PCaP2 located on the plasma membrane is involved in root hair development as a possible signal transducer
1
2013
... DREPP蛋白的氨基酸序列一般可分为氨基端区域、中央区域和羧基端区域(
An Arabidopsis hydrophilic Ca 2+-binding protein with a PEVK-rich domain, PCaP2, is associated with the plasma membrane and interacts with calmodulin and phosphatidylinositol phosphates
5
2010a
... 人们最早从烟草(Nicotiana tabacum)中克隆到DREPP编码基因(
... (Continued)
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 84 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013610402.1 | - |
| 85 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013596813.1 | - |
| 86 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013619975.1 | - |
| 87 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD40021.1 | - |
| 88 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD37718.1 | - |
| 89 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD25026.1 | - |
| 90 | 萝卜 | Raphanus sativus | Rs | 十字花科 | 萝卜属 | XP_018471954.1 | - |
| 91 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | BAA99394.1 | Yuasa and Mae- shima, 2000 |
| 92 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018437382.1 | - |
| 93 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018455198.1 | - |
| 94 | 山嵛菜 | Eutrema salsugineum | Es | 十字花科 | 山嵛菜属 | XP_006413890.1 | - |
| 95 | 山嵛菜 | E. salsugineum | Es | 十字花科 | 山嵛菜属 | XP_024011088.1 | - |
| 96 | 荠菜 | Capsella rubella | Cr | 十字花科 | 荠属 | XP_006284474.1 | - |
| 97 | 荠菜 | C. rubella | Cr | 十字花科 | 荠属 | XP_006279705.1 | - |
| 98 | 亚麻荠 | Camelina sativa | Csa | 十字花科 | 亚麻荠属 | XP_010434307.1 | - |
| 99 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010441852.1 | - |
| 100 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010445250.1 | - |
| 101 | 天蓝遏蓝菜 | Noccaea caerulescens | Nc | 十字花科 | 菥蓂属 | JAU60665.1 | - |
| 102 | 拟南芥 | Arabidopsis thaliana | At | 十字花科 | 拟南芥属 | Q96262.1 | |
| 103 | 拟南芥 | A. thaliana | At | 十字花科 | 拟南芥属 | NP_568636.1 | |
| 104 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002869944.1 | - |
| 105 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002863557.1 | - |
| 106 | 番木瓜 | Carica papaya | Cp | 番木瓜科 | 番木瓜属 | XP_021901673.1 | - |
| 107 | 凤梨 | Ananas comosus | Aco | 凤梨科 | 凤梨属 | OAY80473.1 | |
| 108 | 深圳拟兰 | Apostasia shenzhenica | As | 兰科 | 拟兰属 | PKA54423.1 | - |
| 109 | - | Sarocalamus faberi | Sf | 兰科 | - | ATE87948.1 | - |
| 110 | 小果野芭蕉 | Musa acuminata subsp. malaccensis | Ma | 芭蕉科 | 芭蕉属 | XP_009409470.1 | - |
| 111 | 小兰屿蝴蝶兰 | Phalaenopsis equestris | Phe | 兰科 | 蝴蝶兰属 | XP_020574860.1 | - |
| 112 | 铁皮石斛 | Dendrobium catenatum | Dc | 兰科 | 石斛属 | XP_020688760.1 | - |
| 113 | 玉米 | Zea mays | Zm | 禾本科 | 玉蜀黍属 | XP_020397542.1 | - |
| 114 | 玉米 | Z. mays | Zm | 禾本科 | 玉蜀黍属 | ACG37450.1 | |
| 115 | 水稻 | Oryza sativa | Os | 禾本科 | 稻属 | NP_001042508.1 | |
| 116 | 水稻 | O. sativa | Os | 禾本科 | 稻属 | NP_001046572.1 | |
| 117 | 盐地鼠尾粟 | Sporobolus virginicus | Sv | 禾本科 | 鼠尾粟属 | LC342075.1 | |
| 118 | 二穗短柄草 | Brachypodium distachyon | Bd | 禾本科 | 短柄草属 | XP_003572598.1 | - |
| 119 | 高粱 | Sorghum bicolor | Sb | 禾本科 | 高粱属 | XP_002453713.1 | - |
| 120 | - | Dichanthelium oligosanthes | Do | 禾本科 | 二型花属 | OEL25601.1 |
... PtdInsPs是植物生物膜中含量较低的一类磷脂, 对于细胞信号转导、细胞结构维持、膜流动性调控以及物质转运等过程具有重要作用(
... ;
... AtPCaP2在拟南芥根毛中的表达量较高, 这暗示其在根毛中发挥重要作用(
PCaPs, possible regulators of PtdInsP signals on plasma membrane
1
2010b
... DREPP蛋白的氨基酸序列一般可分为氨基端区域、中央区域和羧基端区域(
Climate clever clovers: new paradigm to reduce the environmental footprint of ruminants by breeding low methanogenic forages utilizing haplotype variation
1
2017
... DREPP protein family members in plants
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 1 | 番茄 | Solanum lycopersicum | Sl | 茄科 | 茄属 | XP_004248413.1 | - |
| 2 | 野生潘那利番茄 | S. pennellii | Sp | 茄科 | 茄属 | XP_015055582.1 | - |
| 3 | 马铃薯 | S. tuberosum | St | 茄科 | 茄属 | XP_006352621.1 | - |
| 4 | 风铃辣椒 | Capsicum baccatum | Cb | 茄科 | 辣椒属 | PHT35828.1 | |
| 5 | 黄灯笼辣椒 | C. chinense | Cch | 茄科 | 辣椒属 | PHU04552.1 | |
| 6 | 辣椒 | C. annuum | Can | 茄科 | 辣椒属 | PHT70067.1 | |
| 7 | 本氏烟草 | Nicotiana benthamiana | Nb | 茄科 | 烟草属 | ASW15993.1 | |
| 8 | 渐狭叶烟草 | N. attenuata | Na | 茄科 | 烟草属 | XP_019226343.1 | - |
| 9 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69901.1 | |
| 10 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69900.1 | |
| 11 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016438748.1 | - |
| 12 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | X95957.1 | |
| 13 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016451342.1 | - |
| 14 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_018631801.1 | - |
| 15 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_009600368.1 | - |
| 16 | 林烟草 | N. sylvestris | Ns | 茄科 | 烟草属 | XP_009763203.1 | - |
| 17 | 中粒咖啡 | Coffea canephora | Cca | 茜草科 | 咖啡属 | CDP04109.1 | - |
| 18 | 小粒咖啡 | C. arabica | Car | 茜草科 | 咖啡属 | XP_027089181.1 | - |
| 19 | 木犀榄 | Olea europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022847863.1 | - |
| 20 | 木犀榄 | O. europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022870576.1 | - |
| 21 | 芝麻 | Sesamum indicum | Si | 胡麻科 | 胡麻属 | XP_011085144.1 | - |
| 22 | 中华猕猴桃 | Actinidia chinensis var. chinensis | Ac | 猕猴桃科 | 猕猴桃属 | PSS11063.1 | - |
| 23 | 苹果 | Malus domestica | Md | 蔷薇科 | 苹果属 | XP_008383239.1 | - |
| 24 | 月季 | Rosa chinensis | Rch | 蔷薇科 | 蔷薇属 | XP_024156781.1 | - |
| 25 | 桃 | Prunus persica | Pp | 蔷薇科 | 桃属 | XP_007212006.1 | - |
| 26 | 樱花 | P. yedoensis var. nudiflora | Py | 蔷薇科 | 桃属 | PQP97549.1 | - |
| 27 | 甜樱桃 | P. avium | Pa | 蔷薇科 | 桃属 | XP_021822163.1 | - |
| 28 | 刺苞菜蓟 | Cynara cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | XP_024964532.1 | - |
| 29 | 刺苞菜蓟 | C. cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | KVH87676.1 | |
| 30 | 莴苣 | Lactuca sativa | Ls | 菊科 | 莴苣属 | XP_023745485.1 | - |
| 31 | 莴苣 | L. sativa | Ls | 菊科 | 莴苣属 | XP_023768949.1 | - |
| 32 | 向日葵 | Helianthus annuus | Ha | 菊科 | 向日葵属 | XP_021982589.1 | - |
| 33 | 向日葵 | H. annuus | Ha | 菊科 | 向日葵属 | XP_022009496.1 | - |
| 34 | 黄花蒿 | Artemisia annua | Aa | 菊科 | 蒿亚属 | PWA97158.1 | |
| 35 | 蒺藜苜蓿 | Medicago truncatula | Mt | 豆科 | 苜蓿属 | XP_013463310.1 | - |
| 36 | 地三叶 | Trifolium subterraneum | Ts | 豆科 | 三叶草属 | GAU45950.1 | |
| 37 | 鹰嘴豆 | Cicer arietinum | Ca | 豆科 | 鹰嘴豆属 | Q9SMK5 | |
| 38 | 野生落花生 | Arachis ipaensis | Ai | 豆科 | 落花生属 | XP_016197976.1 | - |
| 39 | 蔓花生 | A. duranensis | Ad | 豆科 | 落花生属 | XP_015959777.1 | - |
| 40 | 百脉根 | Lotus japonicus | Lj | 豆科 | 百脉根属 | AFK34640.1 | - |
| 41 | 菜豆 | Phaseolus vulgaris | Pv | 豆科 | 菜豆属 | AGV54625.1 | - |
New reference genome sequences of hot pepper reveal the massive evolution of plant disease-resistance genes by retroduplication
3
2017
... DREPP protein family members in plants
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 1 | 番茄 | Solanum lycopersicum | Sl | 茄科 | 茄属 | XP_004248413.1 | - |
| 2 | 野生潘那利番茄 | S. pennellii | Sp | 茄科 | 茄属 | XP_015055582.1 | - |
| 3 | 马铃薯 | S. tuberosum | St | 茄科 | 茄属 | XP_006352621.1 | - |
| 4 | 风铃辣椒 | Capsicum baccatum | Cb | 茄科 | 辣椒属 | PHT35828.1 | |
| 5 | 黄灯笼辣椒 | C. chinense | Cch | 茄科 | 辣椒属 | PHU04552.1 | |
| 6 | 辣椒 | C. annuum | Can | 茄科 | 辣椒属 | PHT70067.1 | |
| 7 | 本氏烟草 | Nicotiana benthamiana | Nb | 茄科 | 烟草属 | ASW15993.1 | |
| 8 | 渐狭叶烟草 | N. attenuata | Na | 茄科 | 烟草属 | XP_019226343.1 | - |
| 9 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69901.1 | |
| 10 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69900.1 | |
| 11 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016438748.1 | - |
| 12 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | X95957.1 | |
| 13 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016451342.1 | - |
| 14 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_018631801.1 | - |
| 15 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_009600368.1 | - |
| 16 | 林烟草 | N. sylvestris | Ns | 茄科 | 烟草属 | XP_009763203.1 | - |
| 17 | 中粒咖啡 | Coffea canephora | Cca | 茜草科 | 咖啡属 | CDP04109.1 | - |
| 18 | 小粒咖啡 | C. arabica | Car | 茜草科 | 咖啡属 | XP_027089181.1 | - |
| 19 | 木犀榄 | Olea europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022847863.1 | - |
| 20 | 木犀榄 | O. europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022870576.1 | - |
| 21 | 芝麻 | Sesamum indicum | Si | 胡麻科 | 胡麻属 | XP_011085144.1 | - |
| 22 | 中华猕猴桃 | Actinidia chinensis var. chinensis | Ac | 猕猴桃科 | 猕猴桃属 | PSS11063.1 | - |
| 23 | 苹果 | Malus domestica | Md | 蔷薇科 | 苹果属 | XP_008383239.1 | - |
| 24 | 月季 | Rosa chinensis | Rch | 蔷薇科 | 蔷薇属 | XP_024156781.1 | - |
| 25 | 桃 | Prunus persica | Pp | 蔷薇科 | 桃属 | XP_007212006.1 | - |
| 26 | 樱花 | P. yedoensis var. nudiflora | Py | 蔷薇科 | 桃属 | PQP97549.1 | - |
| 27 | 甜樱桃 | P. avium | Pa | 蔷薇科 | 桃属 | XP_021822163.1 | - |
| 28 | 刺苞菜蓟 | Cynara cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | XP_024964532.1 | - |
| 29 | 刺苞菜蓟 | C. cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | KVH87676.1 | |
| 30 | 莴苣 | Lactuca sativa | Ls | 菊科 | 莴苣属 | XP_023745485.1 | - |
| 31 | 莴苣 | L. sativa | Ls | 菊科 | 莴苣属 | XP_023768949.1 | - |
| 32 | 向日葵 | Helianthus annuus | Ha | 菊科 | 向日葵属 | XP_021982589.1 | - |
| 33 | 向日葵 | H. annuus | Ha | 菊科 | 向日葵属 | XP_022009496.1 | - |
| 34 | 黄花蒿 | Artemisia annua | Aa | 菊科 | 蒿亚属 | PWA97158.1 | |
| 35 | 蒺藜苜蓿 | Medicago truncatula | Mt | 豆科 | 苜蓿属 | XP_013463310.1 | - |
| 36 | 地三叶 | Trifolium subterraneum | Ts | 豆科 | 三叶草属 | GAU45950.1 | |
| 37 | 鹰嘴豆 | Cicer arietinum | Ca | 豆科 | 鹰嘴豆属 | Q9SMK5 | |
| 38 | 野生落花生 | Arachis ipaensis | Ai | 豆科 | 落花生属 | XP_016197976.1 | - |
| 39 | 蔓花生 | A. duranensis | Ad | 豆科 | 落花生属 | XP_015959777.1 | - |
| 40 | 百脉根 | Lotus japonicus | Lj | 豆科 | 百脉根属 | AFK34640.1 | - |
| 41 | 菜豆 | Phaseolus vulgaris | Pv | 豆科 | 菜豆属 | AGV54625.1 | - |
...
...
Spatial control of Rho (Rac-Rop) signaling in tip-growing plant cells
1
2008
... ROPs (Rho-related GTPase from plants)是一类植物特有的调控根毛生长发育的小G蛋白(
Tip growth: signaling in the apical dome
1
2008
... ROPs (Rho-related GTPase from plants)是一类植物特有的调控根毛生长发育的小G蛋白(
MDP25, a novel calcium regulatory protein, mediates hypocotyl cell elongation by destabilizing cortical microtubules in Arabidopsis
4
2011
... 人们最早从烟草(Nicotiana tabacum)中克隆到DREPP编码基因(
... DREPP蛋白的氨基酸序列一般可分为氨基端区域、中央区域和羧基端区域(
... 微管在植物生长发育过程中发挥至关重要的作用, 微管的稳定性、组织结构和动力学都受到微管调控蛋白的调控(
... AtPCaP2也是微管结合蛋白, 高浓度AtPCaP2抑制微管聚合.AtPCaP2过表达植株的下胚轴细胞内微管排列混乱; 下胚轴表皮细胞肿胀, 细胞伸长被抑制; 铺板细胞突起数明显减少, 细胞边缘光滑(
Arabidopsis Di19 functions as a transcription factor and modulates PR1, PR2, and PR5 expression in response to drought stress
1
2013
... 干旱是农业生产中一种常见的非生物胁迫, 严重制约植物生长发育和生产力(
The genome of medicinal plant Macleaya cordata provides new insights into benzylisoquinoline alkaloids metabolism
1
2017
... (Continued)
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 42 | 大豆 | Glycine max | Gm | 豆科 | 大豆属 | XP_003546380.1 | Matthews et al., 2013 |
| 43 | 大豆 | G. max | Gm | 豆科 | 大豆属 | NP_001237575.1 | - |
| 44 | 欧洲栓皮栎 | Quercus suber | Qs | 壳斗科 | 栎属 | XP_023919313.1 | - |
| 45 | 土瓶草 | Cephalotus follicularis | Cf | 土瓶草科 | 土瓶草属 | GAV80098.1 | |
| 46 | 胡桃 | Juglans regia | Jr | 胡桃科 | 胡桃属 | XP_018814574.1 | - |
| 47 | 甜橙 | Citrus sinensis | Cs | 芸香科 | 柑橘属 | XP_006468602.1 | - |
| 48 | 克里曼丁桔 | C. clementina | Ccl | 芸香科 | 柑橘属 | XP_006448565.1 | - |
| 49 | 大桉 | Eucalyptus grandis | Eg | 桃金娘科 | 桉属 | XP_010051796.1 | - |
| 50 | 毛果杨 | Populus trichocarpa | Pt | 杨柳科 | 杨属 | XP_006385859.1 | - |
| 51 | 毛果杨 | P. trichocarpa | Pt | 杨柳科 | 杨属 | XP_006368838.1 | - |
| 52 | 蓖麻 | Ricinus communis | Rc | 大戟科 | 蓖麻属 | XP_015583044.1 | - |
| 53 | 麻疯树 | Jatropha curcas | Jc | 大戟科 | 麻疯树属 | XP_012088609.1 | - |
| 54 | 木薯 | Manihot esculenta | Me | 大戟科 | 木薯属 | XP_021602838.1 | - |
| 55 | 橡胶树 | Hevea brasiliensis | Hb | 大戟科 | 橡胶树属 | XP_021686108.1 | - |
| 56 | 糙叶山黄麻 | Parasponia andersonii | Pan | 榆科 | 南山黄麻属 | PON44636.1 | - |
| 57 | 异色山黄麻 | Trema orientale | To | 椴树科 | 山黄麻属 | PON83811.1 | - |
| 58 | 长蒴黄麻 | Corchorus olitorius | Co | 椴树科 | 黄麻属 | OMO77743.1 | - |
| 59 | 博落回 | Macleaya cordata | Mc | 罂粟科 | 博落回属 | OVA01374.1 | |
| 60 | 莲 | Nelumbo nucifera | Nn | 睡莲科 | 莲属 | XP_010260837.1 | - |
| 61 | 川桑 | Morus notabilis | Mn | 桑科 | 桑属 | XP_010094633.1 | - |
| 62 | 葡萄 | Vitis vinifera | Vv | 葡萄科 | 葡萄属 | XP_002263090.1 | - |
| 63 | 菠菜 | Spinacia oleracea | So | 藜科 | 菠菜属 | KNA18787.1 | |
| 64 | 甜菜 | Beta vulgaris subsp. vulgaris | Bv | 藜科 | 甜菜属 | XP_010684190.2 | - |
| 65 | 甜菜 | B. vulgaris subsp. vulgaris | Bv | 藜科 | 甜菜属 | KMT06194.1 | |
| 66 | 无油樟 | Amborella trichopoda | Atr | 无油樟科 | 无油樟属 | XP_006859057.2 | - |
| 67 | 醉蝶花 | Tarenaya hassleriana | Th | 醉蝶花科 | 白花菜属 | XP_010537158.1 | - |
| 68 | 芜菁 | Brassica rapa | Br | 十字花科 | 芸苔属 | AHV84997.1 | - |
| 69 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | XP_009133577.1 | - |
| 70 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | RIA04026.1 | - |
| 71 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | XP_009113907.1 | - |
| 72 | 芜菁 | B. rapa | Br | 十字花科 | 芸苔属 | XP_009101693.1 | - |
| 73 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013660923.1 | - |
| 74 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | CDY66354.1 | - |
| 75 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013672014.1 | - |
| 76 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_022572533.1 | - |
| 77 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013717493.1 | - |
| 78 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013659410.1 | - |
| 79 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013730570.1 | - |
| 80 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013715242.1 | - |
| 81 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | XP_013733201.1 | - |
| 82 | 欧洲油菜 | B. napus | Bn | 十字花科 | 芸苔属 | CDY63708.1 | - |
| 83 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013602975.1 | - |
A new family of plasma membrane polypeptides differentially regulated during plant development
2
1998
... DREPP protein family members in plants
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 1 | 番茄 | Solanum lycopersicum | Sl | 茄科 | 茄属 | XP_004248413.1 | - |
| 2 | 野生潘那利番茄 | S. pennellii | Sp | 茄科 | 茄属 | XP_015055582.1 | - |
| 3 | 马铃薯 | S. tuberosum | St | 茄科 | 茄属 | XP_006352621.1 | - |
| 4 | 风铃辣椒 | Capsicum baccatum | Cb | 茄科 | 辣椒属 | PHT35828.1 | |
| 5 | 黄灯笼辣椒 | C. chinense | Cch | 茄科 | 辣椒属 | PHU04552.1 | |
| 6 | 辣椒 | C. annuum | Can | 茄科 | 辣椒属 | PHT70067.1 | |
| 7 | 本氏烟草 | Nicotiana benthamiana | Nb | 茄科 | 烟草属 | ASW15993.1 | |
| 8 | 渐狭叶烟草 | N. attenuata | Na | 茄科 | 烟草属 | XP_019226343.1 | - |
| 9 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69901.1 | |
| 10 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69900.1 | |
| 11 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016438748.1 | - |
| 12 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | X95957.1 | |
| 13 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016451342.1 | - |
| 14 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_018631801.1 | - |
| 15 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_009600368.1 | - |
| 16 | 林烟草 | N. sylvestris | Ns | 茄科 | 烟草属 | XP_009763203.1 | - |
| 17 | 中粒咖啡 | Coffea canephora | Cca | 茜草科 | 咖啡属 | CDP04109.1 | - |
| 18 | 小粒咖啡 | C. arabica | Car | 茜草科 | 咖啡属 | XP_027089181.1 | - |
| 19 | 木犀榄 | Olea europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022847863.1 | - |
| 20 | 木犀榄 | O. europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022870576.1 | - |
| 21 | 芝麻 | Sesamum indicum | Si | 胡麻科 | 胡麻属 | XP_011085144.1 | - |
| 22 | 中华猕猴桃 | Actinidia chinensis var. chinensis | Ac | 猕猴桃科 | 猕猴桃属 | PSS11063.1 | - |
| 23 | 苹果 | Malus domestica | Md | 蔷薇科 | 苹果属 | XP_008383239.1 | - |
| 24 | 月季 | Rosa chinensis | Rch | 蔷薇科 | 蔷薇属 | XP_024156781.1 | - |
| 25 | 桃 | Prunus persica | Pp | 蔷薇科 | 桃属 | XP_007212006.1 | - |
| 26 | 樱花 | P. yedoensis var. nudiflora | Py | 蔷薇科 | 桃属 | PQP97549.1 | - |
| 27 | 甜樱桃 | P. avium | Pa | 蔷薇科 | 桃属 | XP_021822163.1 | - |
| 28 | 刺苞菜蓟 | Cynara cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | XP_024964532.1 | - |
| 29 | 刺苞菜蓟 | C. cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | KVH87676.1 | |
| 30 | 莴苣 | Lactuca sativa | Ls | 菊科 | 莴苣属 | XP_023745485.1 | - |
| 31 | 莴苣 | L. sativa | Ls | 菊科 | 莴苣属 | XP_023768949.1 | - |
| 32 | 向日葵 | Helianthus annuus | Ha | 菊科 | 向日葵属 | XP_021982589.1 | - |
| 33 | 向日葵 | H. annuus | Ha | 菊科 | 向日葵属 | XP_022009496.1 | - |
| 34 | 黄花蒿 | Artemisia annua | Aa | 菊科 | 蒿亚属 | PWA97158.1 | |
| 35 | 蒺藜苜蓿 | Medicago truncatula | Mt | 豆科 | 苜蓿属 | XP_013463310.1 | - |
| 36 | 地三叶 | Trifolium subterraneum | Ts | 豆科 | 三叶草属 | GAU45950.1 | |
| 37 | 鹰嘴豆 | Cicer arietinum | Ca | 豆科 | 鹰嘴豆属 | Q9SMK5 | |
| 38 | 野生落花生 | Arachis ipaensis | Ai | 豆科 | 落花生属 | XP_016197976.1 | - |
| 39 | 蔓花生 | A. duranensis | Ad | 豆科 | 落花生属 | XP_015959777.1 | - |
| 40 | 百脉根 | Lotus japonicus | Lj | 豆科 | 百脉根属 | AFK34640.1 | - |
| 41 | 菜豆 | Phaseolus vulgaris | Pv | 豆科 | 菜豆属 | AGV54625.1 | - |
...
Purification and expression of a protein elicitor from Alternaria tenuissima and elicitor- mediated defence responses in tobacco
1
2010
... 对烟草DREPP家族成员PtBP1的研究表明, 烟草花叶病毒(tobacco mosaic virus, TMV)侵染烟草植株3天后, PeaT1沉默植株叶片的损伤程度约为野生型烟草的3倍, PeaT1的沉默减弱了烟草对TMV的抗性.PeaT1是一种来自真菌病原体细极链格孢菌的蛋白诱导因子, 可以诱导包括烟草在内的各种植物的防御反应并增强抗病性(
Engineered resistance and hypersusceptibility through functional metabolic studies of 100 genes in soybean to its major pathogen, the soybean cyst nematode
0
2013
A DREPP protein interacted with PeaT1 from Alternaria tenuissima and is involved in elicitor-induced disease resistance in Nicotiana plants
1
2018
... 对烟草DREPP家族成员PtBP1的研究表明, 烟草花叶病毒(tobacco mosaic virus, TMV)侵染烟草植株3天后, PeaT1沉默植株叶片的损伤程度约为野生型烟草的3倍, PeaT1的沉默减弱了烟草对TMV的抗性.PeaT1是一种来自真菌病原体细极链格孢菌的蛋白诱导因子, 可以诱导包括烟草在内的各种植物的防御反应并增强抗病性(
A plasma membrane-associated protein of Arabidopsis thaliana AtPCaP1 binds copper ions and changes its higher order structure
2
2008
... PtdInsPs是植物生物膜中含量较低的一类磷脂, 对于细胞信号转导、细胞结构维持、膜流动性调控以及物质转运等过程具有重要作用(
... 解离(
CBF2/ DREB1C is a negative regulator of CBF1/DREB1B and CBF3/DREB1A expression and plays a central role in stress tolerance in Arabidopsis
1
2004
... 低温限制植物在地球上的分布, 减少作物产量, 是一种常见的非生物胁迫(
Arabidopsis CBF1 and CBF3 have a different function than CBF2 in cold acclimation and define different gene classes in the CBF regulon
1
2007
... 低温限制植物在地球上的分布, 减少作物产量, 是一种常见的非生物胁迫(
Arabidopsis microtubule-destabilizing protein 25 functions in pollen tube growth by severing actin filaments
3
2014
... 发育调节质膜多肽(developmentally regulated plasma membrane polypeptide, DREPP)蛋白是一类与质膜相关的Ca2+结合蛋白.它可以通过与微管和微丝结合调节细胞骨架结构, 从而调控根和花粉管的发育(
... DREPP蛋白的氨基酸序列一般可分为氨基端区域、中央区域和羧基端区域(
... AtPCaP1和AtPCaP2在拟南芥花粉与花粉管中的表达量均很高.atpcap2突变体与AtPCaP2过表达体的花粉管扭曲, 生长方向异常, 甚至环绕胚珠生长, 无法完成受精, 从而造成结实率下降.atpcap2突变体与AtPCaP2过表达体花粉管顶端及亚顶端区内微丝的组织排列方式发生改变.此外, 免疫共沉淀实验结果表明, AtPCaP1可直接与微丝结合并具有微弱的微丝切割活性; 在10 μmol∙L-1 Ca2+的条件下, 微丝切割活性明显增强.与野生型相比, atpcap1突变体植株花粉管亚顶端的微丝束更大, 且密度更高, 这表明AtPCaP1的缺失导致花粉管亚顶端微丝无法被切割.同时, 将AtPCaP1中微丝与Ca2+结合关键位点(分别为Asn5和184-188位VEEKK域中E185和K188)突变后, 回补atpcap1突变体, 无法恢复其正常花粉管表型.由此推测, AtPCaP1和AtPCaP2通过调节花粉管顶端及亚顶端微丝的组织排列和动态变化调控花粉管顶端生长(
The draft genome of MD-2 pineapple using hybrid error correction of long reads
1
2016
... (Continued)
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 84 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013610402.1 | - |
| 85 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013596813.1 | - |
| 86 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013619975.1 | - |
| 87 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD40021.1 | - |
| 88 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD37718.1 | - |
| 89 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD25026.1 | - |
| 90 | 萝卜 | Raphanus sativus | Rs | 十字花科 | 萝卜属 | XP_018471954.1 | - |
| 91 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | BAA99394.1 | Yuasa and Mae- shima, 2000 |
| 92 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018437382.1 | - |
| 93 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018455198.1 | - |
| 94 | 山嵛菜 | Eutrema salsugineum | Es | 十字花科 | 山嵛菜属 | XP_006413890.1 | - |
| 95 | 山嵛菜 | E. salsugineum | Es | 十字花科 | 山嵛菜属 | XP_024011088.1 | - |
| 96 | 荠菜 | Capsella rubella | Cr | 十字花科 | 荠属 | XP_006284474.1 | - |
| 97 | 荠菜 | C. rubella | Cr | 十字花科 | 荠属 | XP_006279705.1 | - |
| 98 | 亚麻荠 | Camelina sativa | Csa | 十字花科 | 亚麻荠属 | XP_010434307.1 | - |
| 99 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010441852.1 | - |
| 100 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010445250.1 | - |
| 101 | 天蓝遏蓝菜 | Noccaea caerulescens | Nc | 十字花科 | 菥蓂属 | JAU60665.1 | - |
| 102 | 拟南芥 | Arabidopsis thaliana | At | 十字花科 | 拟南芥属 | Q96262.1 | |
| 103 | 拟南芥 | A. thaliana | At | 十字花科 | 拟南芥属 | NP_568636.1 | |
| 104 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002869944.1 | - |
| 105 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002863557.1 | - |
| 106 | 番木瓜 | Carica papaya | Cp | 番木瓜科 | 番木瓜属 | XP_021901673.1 | - |
| 107 | 凤梨 | Ananas comosus | Aco | 凤梨科 | 凤梨属 | OAY80473.1 | |
| 108 | 深圳拟兰 | Apostasia shenzhenica | As | 兰科 | 拟兰属 | PKA54423.1 | - |
| 109 | - | Sarocalamus faberi | Sf | 兰科 | - | ATE87948.1 | - |
| 110 | 小果野芭蕉 | Musa acuminata subsp. malaccensis | Ma | 芭蕉科 | 芭蕉属 | XP_009409470.1 | - |
| 111 | 小兰屿蝴蝶兰 | Phalaenopsis equestris | Phe | 兰科 | 蝴蝶兰属 | XP_020574860.1 | - |
| 112 | 铁皮石斛 | Dendrobium catenatum | Dc | 兰科 | 石斛属 | XP_020688760.1 | - |
| 113 | 玉米 | Zea mays | Zm | 禾本科 | 玉蜀黍属 | XP_020397542.1 | - |
| 114 | 玉米 | Z. mays | Zm | 禾本科 | 玉蜀黍属 | ACG37450.1 | |
| 115 | 水稻 | Oryza sativa | Os | 禾本科 | 稻属 | NP_001042508.1 | |
| 116 | 水稻 | O. sativa | Os | 禾本科 | 稻属 | NP_001046572.1 | |
| 117 | 盐地鼠尾粟 | Sporobolus virginicus | Sv | 禾本科 | 鼠尾粟属 | LC342075.1 | |
| 118 | 二穗短柄草 | Brachypodium distachyon | Bd | 禾本科 | 短柄草属 | XP_003572598.1 | - |
| 119 | 高粱 | Sorghum bicolor | Sb | 禾本科 | 高粱属 | XP_002453713.1 | - |
| 120 | - | Dichanthelium oligosanthes | Do | 禾本科 | 二型花属 | OEL25601.1 |
The genome sequence of the outbreeding globe artichoke constructed de novo incorporating a phase-aware low-pass sequencing stra-tegy of F1 progeny
1
2016
... DREPP protein family members in plants
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 1 | 番茄 | Solanum lycopersicum | Sl | 茄科 | 茄属 | XP_004248413.1 | - |
| 2 | 野生潘那利番茄 | S. pennellii | Sp | 茄科 | 茄属 | XP_015055582.1 | - |
| 3 | 马铃薯 | S. tuberosum | St | 茄科 | 茄属 | XP_006352621.1 | - |
| 4 | 风铃辣椒 | Capsicum baccatum | Cb | 茄科 | 辣椒属 | PHT35828.1 | |
| 5 | 黄灯笼辣椒 | C. chinense | Cch | 茄科 | 辣椒属 | PHU04552.1 | |
| 6 | 辣椒 | C. annuum | Can | 茄科 | 辣椒属 | PHT70067.1 | |
| 7 | 本氏烟草 | Nicotiana benthamiana | Nb | 茄科 | 烟草属 | ASW15993.1 | |
| 8 | 渐狭叶烟草 | N. attenuata | Na | 茄科 | 烟草属 | XP_019226343.1 | - |
| 9 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69901.1 | |
| 10 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69900.1 | |
| 11 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016438748.1 | - |
| 12 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | X95957.1 | |
| 13 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016451342.1 | - |
| 14 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_018631801.1 | - |
| 15 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_009600368.1 | - |
| 16 | 林烟草 | N. sylvestris | Ns | 茄科 | 烟草属 | XP_009763203.1 | - |
| 17 | 中粒咖啡 | Coffea canephora | Cca | 茜草科 | 咖啡属 | CDP04109.1 | - |
| 18 | 小粒咖啡 | C. arabica | Car | 茜草科 | 咖啡属 | XP_027089181.1 | - |
| 19 | 木犀榄 | Olea europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022847863.1 | - |
| 20 | 木犀榄 | O. europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022870576.1 | - |
| 21 | 芝麻 | Sesamum indicum | Si | 胡麻科 | 胡麻属 | XP_011085144.1 | - |
| 22 | 中华猕猴桃 | Actinidia chinensis var. chinensis | Ac | 猕猴桃科 | 猕猴桃属 | PSS11063.1 | - |
| 23 | 苹果 | Malus domestica | Md | 蔷薇科 | 苹果属 | XP_008383239.1 | - |
| 24 | 月季 | Rosa chinensis | Rch | 蔷薇科 | 蔷薇属 | XP_024156781.1 | - |
| 25 | 桃 | Prunus persica | Pp | 蔷薇科 | 桃属 | XP_007212006.1 | - |
| 26 | 樱花 | P. yedoensis var. nudiflora | Py | 蔷薇科 | 桃属 | PQP97549.1 | - |
| 27 | 甜樱桃 | P. avium | Pa | 蔷薇科 | 桃属 | XP_021822163.1 | - |
| 28 | 刺苞菜蓟 | Cynara cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | XP_024964532.1 | - |
| 29 | 刺苞菜蓟 | C. cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | KVH87676.1 | |
| 30 | 莴苣 | Lactuca sativa | Ls | 菊科 | 莴苣属 | XP_023745485.1 | - |
| 31 | 莴苣 | L. sativa | Ls | 菊科 | 莴苣属 | XP_023768949.1 | - |
| 32 | 向日葵 | Helianthus annuus | Ha | 菊科 | 向日葵属 | XP_021982589.1 | - |
| 33 | 向日葵 | H. annuus | Ha | 菊科 | 向日葵属 | XP_022009496.1 | - |
| 34 | 黄花蒿 | Artemisia annua | Aa | 菊科 | 蒿亚属 | PWA97158.1 | |
| 35 | 蒺藜苜蓿 | Medicago truncatula | Mt | 豆科 | 苜蓿属 | XP_013463310.1 | - |
| 36 | 地三叶 | Trifolium subterraneum | Ts | 豆科 | 三叶草属 | GAU45950.1 | |
| 37 | 鹰嘴豆 | Cicer arietinum | Ca | 豆科 | 鹰嘴豆属 | Q9SMK5 | |
| 38 | 野生落花生 | Arachis ipaensis | Ai | 豆科 | 落花生属 | XP_016197976.1 | - |
| 39 | 蔓花生 | A. duranensis | Ad | 豆科 | 落花生属 | XP_015959777.1 | - |
| 40 | 百脉根 | Lotus japonicus | Lj | 豆科 | 百脉根属 | AFK34640.1 | - |
| 41 | 菜豆 | Phaseolus vulgaris | Pv | 豆科 | 菜豆属 | AGV54625.1 | - |
The genome of Artemisia annua provides insight into the evolution of Asteraceae family and artemisinin biosynthesis
1
2018
... DREPP protein family members in plants
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 1 | 番茄 | Solanum lycopersicum | Sl | 茄科 | 茄属 | XP_004248413.1 | - |
| 2 | 野生潘那利番茄 | S. pennellii | Sp | 茄科 | 茄属 | XP_015055582.1 | - |
| 3 | 马铃薯 | S. tuberosum | St | 茄科 | 茄属 | XP_006352621.1 | - |
| 4 | 风铃辣椒 | Capsicum baccatum | Cb | 茄科 | 辣椒属 | PHT35828.1 | |
| 5 | 黄灯笼辣椒 | C. chinense | Cch | 茄科 | 辣椒属 | PHU04552.1 | |
| 6 | 辣椒 | C. annuum | Can | 茄科 | 辣椒属 | PHT70067.1 | |
| 7 | 本氏烟草 | Nicotiana benthamiana | Nb | 茄科 | 烟草属 | ASW15993.1 | |
| 8 | 渐狭叶烟草 | N. attenuata | Na | 茄科 | 烟草属 | XP_019226343.1 | - |
| 9 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69901.1 | |
| 10 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69900.1 | |
| 11 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016438748.1 | - |
| 12 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | X95957.1 | |
| 13 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016451342.1 | - |
| 14 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_018631801.1 | - |
| 15 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_009600368.1 | - |
| 16 | 林烟草 | N. sylvestris | Ns | 茄科 | 烟草属 | XP_009763203.1 | - |
| 17 | 中粒咖啡 | Coffea canephora | Cca | 茜草科 | 咖啡属 | CDP04109.1 | - |
| 18 | 小粒咖啡 | C. arabica | Car | 茜草科 | 咖啡属 | XP_027089181.1 | - |
| 19 | 木犀榄 | Olea europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022847863.1 | - |
| 20 | 木犀榄 | O. europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022870576.1 | - |
| 21 | 芝麻 | Sesamum indicum | Si | 胡麻科 | 胡麻属 | XP_011085144.1 | - |
| 22 | 中华猕猴桃 | Actinidia chinensis var. chinensis | Ac | 猕猴桃科 | 猕猴桃属 | PSS11063.1 | - |
| 23 | 苹果 | Malus domestica | Md | 蔷薇科 | 苹果属 | XP_008383239.1 | - |
| 24 | 月季 | Rosa chinensis | Rch | 蔷薇科 | 蔷薇属 | XP_024156781.1 | - |
| 25 | 桃 | Prunus persica | Pp | 蔷薇科 | 桃属 | XP_007212006.1 | - |
| 26 | 樱花 | P. yedoensis var. nudiflora | Py | 蔷薇科 | 桃属 | PQP97549.1 | - |
| 27 | 甜樱桃 | P. avium | Pa | 蔷薇科 | 桃属 | XP_021822163.1 | - |
| 28 | 刺苞菜蓟 | Cynara cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | XP_024964532.1 | - |
| 29 | 刺苞菜蓟 | C. cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | KVH87676.1 | |
| 30 | 莴苣 | Lactuca sativa | Ls | 菊科 | 莴苣属 | XP_023745485.1 | - |
| 31 | 莴苣 | L. sativa | Ls | 菊科 | 莴苣属 | XP_023768949.1 | - |
| 32 | 向日葵 | Helianthus annuus | Ha | 菊科 | 向日葵属 | XP_021982589.1 | - |
| 33 | 向日葵 | H. annuus | Ha | 菊科 | 向日葵属 | XP_022009496.1 | - |
| 34 | 黄花蒿 | Artemisia annua | Aa | 菊科 | 蒿亚属 | PWA97158.1 | |
| 35 | 蒺藜苜蓿 | Medicago truncatula | Mt | 豆科 | 苜蓿属 | XP_013463310.1 | - |
| 36 | 地三叶 | Trifolium subterraneum | Ts | 豆科 | 三叶草属 | GAU45950.1 | |
| 37 | 鹰嘴豆 | Cicer arietinum | Ca | 豆科 | 鹰嘴豆属 | Q9SMK5 | |
| 38 | 野生落花生 | Arachis ipaensis | Ai | 豆科 | 落花生属 | XP_016197976.1 | - |
| 39 | 蔓花生 | A. duranensis | Ad | 豆科 | 落花生属 | XP_015959777.1 | - |
| 40 | 百脉根 | Lotus japonicus | Lj | 豆科 | 百脉根属 | AFK34640.1 | - |
| 41 | 菜豆 | Phaseolus vulgaris | Pv | 豆科 | 菜豆属 | AGV54625.1 | - |
Molecular regulation of CBF signaling in cold acclimation
1
2018
... 低温限制植物在地球上的分布, 减少作物产量, 是一种常见的非生物胁迫(
The draft genome of the C3 panicoid grass species Dichanthelium oligosanthes
1
2016
... (Continued)
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 84 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013610402.1 | - |
| 85 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013596813.1 | - |
| 86 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013619975.1 | - |
| 87 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD40021.1 | - |
| 88 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD37718.1 | - |
| 89 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD25026.1 | - |
| 90 | 萝卜 | Raphanus sativus | Rs | 十字花科 | 萝卜属 | XP_018471954.1 | - |
| 91 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | BAA99394.1 | Yuasa and Mae- shima, 2000 |
| 92 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018437382.1 | - |
| 93 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018455198.1 | - |
| 94 | 山嵛菜 | Eutrema salsugineum | Es | 十字花科 | 山嵛菜属 | XP_006413890.1 | - |
| 95 | 山嵛菜 | E. salsugineum | Es | 十字花科 | 山嵛菜属 | XP_024011088.1 | - |
| 96 | 荠菜 | Capsella rubella | Cr | 十字花科 | 荠属 | XP_006284474.1 | - |
| 97 | 荠菜 | C. rubella | Cr | 十字花科 | 荠属 | XP_006279705.1 | - |
| 98 | 亚麻荠 | Camelina sativa | Csa | 十字花科 | 亚麻荠属 | XP_010434307.1 | - |
| 99 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010441852.1 | - |
| 100 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010445250.1 | - |
| 101 | 天蓝遏蓝菜 | Noccaea caerulescens | Nc | 十字花科 | 菥蓂属 | JAU60665.1 | - |
| 102 | 拟南芥 | Arabidopsis thaliana | At | 十字花科 | 拟南芥属 | Q96262.1 | |
| 103 | 拟南芥 | A. thaliana | At | 十字花科 | 拟南芥属 | NP_568636.1 | |
| 104 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002869944.1 | - |
| 105 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002863557.1 | - |
| 106 | 番木瓜 | Carica papaya | Cp | 番木瓜科 | 番木瓜属 | XP_021901673.1 | - |
| 107 | 凤梨 | Ananas comosus | Aco | 凤梨科 | 凤梨属 | OAY80473.1 | |
| 108 | 深圳拟兰 | Apostasia shenzhenica | As | 兰科 | 拟兰属 | PKA54423.1 | - |
| 109 | - | Sarocalamus faberi | Sf | 兰科 | - | ATE87948.1 | - |
| 110 | 小果野芭蕉 | Musa acuminata subsp. malaccensis | Ma | 芭蕉科 | 芭蕉属 | XP_009409470.1 | - |
| 111 | 小兰屿蝴蝶兰 | Phalaenopsis equestris | Phe | 兰科 | 蝴蝶兰属 | XP_020574860.1 | - |
| 112 | 铁皮石斛 | Dendrobium catenatum | Dc | 兰科 | 石斛属 | XP_020688760.1 | - |
| 113 | 玉米 | Zea mays | Zm | 禾本科 | 玉蜀黍属 | XP_020397542.1 | - |
| 114 | 玉米 | Z. mays | Zm | 禾本科 | 玉蜀黍属 | ACG37450.1 | |
| 115 | 水稻 | Oryza sativa | Os | 禾本科 | 稻属 | NP_001042508.1 | |
| 116 | 水稻 | O. sativa | Os | 禾本科 | 稻属 | NP_001046572.1 | |
| 117 | 盐地鼠尾粟 | Sporobolus virginicus | Sv | 禾本科 | 鼠尾粟属 | LC342075.1 | |
| 118 | 二穗短柄草 | Brachypodium distachyon | Bd | 禾本科 | 短柄草属 | XP_003572598.1 | - |
| 119 | 高粱 | Sorghum bicolor | Sb | 禾本科 | 高粱属 | XP_002453713.1 | - |
| 120 | - | Dichanthelium oligosanthes | Do | 禾本科 | 二型花属 | OEL25601.1 |
Phosphoproteomic dynamics of chickpea ( Cicer arietinum L.) reveals shared and distinct components of dehydration response
1
2013
... DREPP protein family members in plants
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 1 | 番茄 | Solanum lycopersicum | Sl | 茄科 | 茄属 | XP_004248413.1 | - |
| 2 | 野生潘那利番茄 | S. pennellii | Sp | 茄科 | 茄属 | XP_015055582.1 | - |
| 3 | 马铃薯 | S. tuberosum | St | 茄科 | 茄属 | XP_006352621.1 | - |
| 4 | 风铃辣椒 | Capsicum baccatum | Cb | 茄科 | 辣椒属 | PHT35828.1 | |
| 5 | 黄灯笼辣椒 | C. chinense | Cch | 茄科 | 辣椒属 | PHU04552.1 | |
| 6 | 辣椒 | C. annuum | Can | 茄科 | 辣椒属 | PHT70067.1 | |
| 7 | 本氏烟草 | Nicotiana benthamiana | Nb | 茄科 | 烟草属 | ASW15993.1 | |
| 8 | 渐狭叶烟草 | N. attenuata | Na | 茄科 | 烟草属 | XP_019226343.1 | - |
| 9 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69901.1 | |
| 10 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | CAA69900.1 | |
| 11 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016438748.1 | - |
| 12 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | X95957.1 | |
| 13 | 烟草 | N. tabacum | Nt | 茄科 | 烟草属 | XP_016451342.1 | - |
| 14 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_018631801.1 | - |
| 15 | 绒毛状烟草 | N. tomentosiformis | Nto | 茄科 | 烟草属 | XP_009600368.1 | - |
| 16 | 林烟草 | N. sylvestris | Ns | 茄科 | 烟草属 | XP_009763203.1 | - |
| 17 | 中粒咖啡 | Coffea canephora | Cca | 茜草科 | 咖啡属 | CDP04109.1 | - |
| 18 | 小粒咖啡 | C. arabica | Car | 茜草科 | 咖啡属 | XP_027089181.1 | - |
| 19 | 木犀榄 | Olea europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022847863.1 | - |
| 20 | 木犀榄 | O. europaea var. sylvestris | Oe | 木犀科 | 木犀榄属 | XP_022870576.1 | - |
| 21 | 芝麻 | Sesamum indicum | Si | 胡麻科 | 胡麻属 | XP_011085144.1 | - |
| 22 | 中华猕猴桃 | Actinidia chinensis var. chinensis | Ac | 猕猴桃科 | 猕猴桃属 | PSS11063.1 | - |
| 23 | 苹果 | Malus domestica | Md | 蔷薇科 | 苹果属 | XP_008383239.1 | - |
| 24 | 月季 | Rosa chinensis | Rch | 蔷薇科 | 蔷薇属 | XP_024156781.1 | - |
| 25 | 桃 | Prunus persica | Pp | 蔷薇科 | 桃属 | XP_007212006.1 | - |
| 26 | 樱花 | P. yedoensis var. nudiflora | Py | 蔷薇科 | 桃属 | PQP97549.1 | - |
| 27 | 甜樱桃 | P. avium | Pa | 蔷薇科 | 桃属 | XP_021822163.1 | - |
| 28 | 刺苞菜蓟 | Cynara cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | XP_024964532.1 | - |
| 29 | 刺苞菜蓟 | C. cardunculus var. scolymus | Cc | 菊科 | 菜蓟属 | KVH87676.1 | |
| 30 | 莴苣 | Lactuca sativa | Ls | 菊科 | 莴苣属 | XP_023745485.1 | - |
| 31 | 莴苣 | L. sativa | Ls | 菊科 | 莴苣属 | XP_023768949.1 | - |
| 32 | 向日葵 | Helianthus annuus | Ha | 菊科 | 向日葵属 | XP_021982589.1 | - |
| 33 | 向日葵 | H. annuus | Ha | 菊科 | 向日葵属 | XP_022009496.1 | - |
| 34 | 黄花蒿 | Artemisia annua | Aa | 菊科 | 蒿亚属 | PWA97158.1 | |
| 35 | 蒺藜苜蓿 | Medicago truncatula | Mt | 豆科 | 苜蓿属 | XP_013463310.1 | - |
| 36 | 地三叶 | Trifolium subterraneum | Ts | 豆科 | 三叶草属 | GAU45950.1 | |
| 37 | 鹰嘴豆 | Cicer arietinum | Ca | 豆科 | 鹰嘴豆属 | Q9SMK5 | |
| 38 | 野生落花生 | Arachis ipaensis | Ai | 豆科 | 落花生属 | XP_016197976.1 | - |
| 39 | 蔓花生 | A. duranensis | Ad | 豆科 | 落花生属 | XP_015959777.1 | - |
| 40 | 百脉根 | Lotus japonicus | Lj | 豆科 | 百脉根属 | AFK34640.1 | - |
| 41 | 菜豆 | Phaseolus vulgaris | Pv | 豆科 | 菜豆属 | AGV54625.1 | - |
Phosphatidic acid: a multifunctional stress signaling lipid in plants
1
2005
... PtdInsPs是植物生物膜中含量较低的一类磷脂, 对于细胞信号转导、细胞结构维持、膜流动性调控以及物质转运等过程具有重要作用(
Isolation, expression, and functional analysis of developmentally regulated plasma membrane polypeptide 1 (DREPP1) in Sporobolus virginicus grown under alkali salt stress
2
2018
... (Continued)
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 84 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013610402.1 | - |
| 85 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013596813.1 | - |
| 86 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013619975.1 | - |
| 87 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD40021.1 | - |
| 88 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD37718.1 | - |
| 89 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD25026.1 | - |
| 90 | 萝卜 | Raphanus sativus | Rs | 十字花科 | 萝卜属 | XP_018471954.1 | - |
| 91 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | BAA99394.1 | Yuasa and Mae- shima, 2000 |
| 92 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018437382.1 | - |
| 93 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018455198.1 | - |
| 94 | 山嵛菜 | Eutrema salsugineum | Es | 十字花科 | 山嵛菜属 | XP_006413890.1 | - |
| 95 | 山嵛菜 | E. salsugineum | Es | 十字花科 | 山嵛菜属 | XP_024011088.1 | - |
| 96 | 荠菜 | Capsella rubella | Cr | 十字花科 | 荠属 | XP_006284474.1 | - |
| 97 | 荠菜 | C. rubella | Cr | 十字花科 | 荠属 | XP_006279705.1 | - |
| 98 | 亚麻荠 | Camelina sativa | Csa | 十字花科 | 亚麻荠属 | XP_010434307.1 | - |
| 99 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010441852.1 | - |
| 100 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010445250.1 | - |
| 101 | 天蓝遏蓝菜 | Noccaea caerulescens | Nc | 十字花科 | 菥蓂属 | JAU60665.1 | - |
| 102 | 拟南芥 | Arabidopsis thaliana | At | 十字花科 | 拟南芥属 | Q96262.1 | |
| 103 | 拟南芥 | A. thaliana | At | 十字花科 | 拟南芥属 | NP_568636.1 | |
| 104 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002869944.1 | - |
| 105 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002863557.1 | - |
| 106 | 番木瓜 | Carica papaya | Cp | 番木瓜科 | 番木瓜属 | XP_021901673.1 | - |
| 107 | 凤梨 | Ananas comosus | Aco | 凤梨科 | 凤梨属 | OAY80473.1 | |
| 108 | 深圳拟兰 | Apostasia shenzhenica | As | 兰科 | 拟兰属 | PKA54423.1 | - |
| 109 | - | Sarocalamus faberi | Sf | 兰科 | - | ATE87948.1 | - |
| 110 | 小果野芭蕉 | Musa acuminata subsp. malaccensis | Ma | 芭蕉科 | 芭蕉属 | XP_009409470.1 | - |
| 111 | 小兰屿蝴蝶兰 | Phalaenopsis equestris | Phe | 兰科 | 蝴蝶兰属 | XP_020574860.1 | - |
| 112 | 铁皮石斛 | Dendrobium catenatum | Dc | 兰科 | 石斛属 | XP_020688760.1 | - |
| 113 | 玉米 | Zea mays | Zm | 禾本科 | 玉蜀黍属 | XP_020397542.1 | - |
| 114 | 玉米 | Z. mays | Zm | 禾本科 | 玉蜀黍属 | ACG37450.1 | |
| 115 | 水稻 | Oryza sativa | Os | 禾本科 | 稻属 | NP_001042508.1 | |
| 116 | 水稻 | O. sativa | Os | 禾本科 | 稻属 | NP_001046572.1 | |
| 117 | 盐地鼠尾粟 | Sporobolus virginicus | Sv | 禾本科 | 鼠尾粟属 | LC342075.1 | |
| 118 | 二穗短柄草 | Brachypodium distachyon | Bd | 禾本科 | 短柄草属 | XP_003572598.1 | - |
| 119 | 高粱 | Sorghum bicolor | Sb | 禾本科 | 高粱属 | XP_002453713.1 | - |
| 120 | - | Dichanthelium oligosanthes | Do | 禾本科 | 二型花属 | OEL25601.1 |
... 微管在植物生长发育过程中发挥至关重要的作用, 微管的稳定性、组织结构和动力学都受到微管调控蛋白的调控(
1
2016
... 干旱是农业生产中一种常见的非生物胁迫, 严重制约植物生长发育和生产力(
Evolutionary plasticity of plasma membrane interaction in DREPP family proteins
2
2017
... DREPP蛋白的氨基酸序列一般可分为氨基端区域、中央区域和羧基端区域(
... ).DREPP蛋白的氨基端区域约含23个高度保守的氨基酸残基, 被称为N23域.AtPCaP1和AtPCaP2的N23域中第2位甘氨酸(Gly2)为豆蔻酰化位点, 第5-23位氨基酸为静电互作区, 豆蔻酰化和静电互作区参与AtDREPPs的质膜定位过程(
Arabidopsis MICROTUBULE-ASSOCIATED PRO- TEIN 18 functions in directional cell growth by destabilizing cortical microtubules
7
2007
... 人们最早从烟草(Nicotiana tabacum)中克隆到DREPP编码基因(
... 微管在植物生长发育过程中发挥至关重要的作用, 微管的稳定性、组织结构和动力学都受到微管调控蛋白的调控(
... AtPCaP2在拟南芥各器官均有不同程度的表达, 但在根/根毛、下胚轴和花粉/花粉管中的表达量较高(
... 植物根部细胞周质微管动态变化调节细胞伸长和形态建成, 对于根部极性生长具有重要作用.AtPCaP2在根部组织中主要在伸长区表达, 而在根尖没有表达(
... 过表达植株的根部微管排列较乱, 呈斜向或纵向排列(
... AtPCaP2在拟南芥根毛中的表达量较高, 这暗示其在根毛中发挥重要作用(
... AtPCaP2也是微管结合蛋白, 高浓度AtPCaP2抑制微管聚合.AtPCaP2过表达植株的下胚轴细胞内微管排列混乱; 下胚轴表皮细胞肿胀, 细胞伸长被抑制; 铺板细胞突起数明显减少, 细胞边缘光滑(
. Arabidopsis PCaP2 plays an important role in chilling tolerance and ABA response by activating CBF- and SnRK2-mediated transcriptional regulatory network
3
2018a
... 发育调节质膜多肽(developmentally regulated plasma membrane polypeptide, DREPP)蛋白是一类与质膜相关的Ca2+结合蛋白.它可以通过与微管和微丝结合调节细胞骨架结构, 从而调控根和花粉管的发育(
... 植物根部细胞周质微管动态变化调节细胞伸长和形态建成, 对于根部极性生长具有重要作用.AtPCaP2在根部组织中主要在伸长区表达, 而在根尖没有表达(
... 低温限制植物在地球上的分布, 减少作物产量, 是一种常见的非生物胁迫(
Arabidopsis PCaP2 functions as a linker between ABA and SA signals in plant water deficit tolerance
2
2018
... 发育调节质膜多肽(developmentally regulated plasma membrane polypeptide, DREPP)蛋白是一类与质膜相关的Ca2+结合蛋白.它可以通过与微管和微丝结合调节细胞骨架结构, 从而调控根和花粉管的发育(
... 干旱是农业生产中一种常见的非生物胁迫, 严重制约植物生长发育和生产力(
Expression of developmentally regulated plasma membrane polypeptide (DREPP2) in rice root tip and interaction with Ca 2+/CaM complex and microtubule
4
2015
... (Continued)
| 序号 | 物种名 | 拉丁名 | 缩写 | 科名 | 属名 | 蛋白质编号 | 参考文献 |
|---|---|---|---|---|---|---|---|
| 84 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013610402.1 | - |
| 85 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013596813.1 | - |
| 86 | 野甘蓝 | B. oleracea var. oleracea | Bol | 十字花科 | 芸苔属 | XP_013619975.1 | - |
| 87 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD40021.1 | - |
| 88 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD37718.1 | - |
| 89 | 甘蓝 | B. oleracea | Bo | 十字花科 | 芸苔属 | VDD25026.1 | - |
| 90 | 萝卜 | Raphanus sativus | Rs | 十字花科 | 萝卜属 | XP_018471954.1 | - |
| 91 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | BAA99394.1 | Yuasa and Mae- shima, 2000 |
| 92 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018437382.1 | - |
| 93 | 萝卜 | R. sativus | Rs | 十字花科 | 萝卜属 | XP_018455198.1 | - |
| 94 | 山嵛菜 | Eutrema salsugineum | Es | 十字花科 | 山嵛菜属 | XP_006413890.1 | - |
| 95 | 山嵛菜 | E. salsugineum | Es | 十字花科 | 山嵛菜属 | XP_024011088.1 | - |
| 96 | 荠菜 | Capsella rubella | Cr | 十字花科 | 荠属 | XP_006284474.1 | - |
| 97 | 荠菜 | C. rubella | Cr | 十字花科 | 荠属 | XP_006279705.1 | - |
| 98 | 亚麻荠 | Camelina sativa | Csa | 十字花科 | 亚麻荠属 | XP_010434307.1 | - |
| 99 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010441852.1 | - |
| 100 | 亚麻荠 | C. sativa | Csa | 十字花科 | 亚麻荠属 | XP_010445250.1 | - |
| 101 | 天蓝遏蓝菜 | Noccaea caerulescens | Nc | 十字花科 | 菥蓂属 | JAU60665.1 | - |
| 102 | 拟南芥 | Arabidopsis thaliana | At | 十字花科 | 拟南芥属 | Q96262.1 | |
| 103 | 拟南芥 | A. thaliana | At | 十字花科 | 拟南芥属 | NP_568636.1 | |
| 104 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002869944.1 | - |
| 105 | 琴叶拟南芥 | A. lyrata subsp. lyrata | Al | 十字花科 | 拟南芥属 | XP_002863557.1 | - |
| 106 | 番木瓜 | Carica papaya | Cp | 番木瓜科 | 番木瓜属 | XP_021901673.1 | - |
| 107 | 凤梨 | Ananas comosus | Aco | 凤梨科 | 凤梨属 | OAY80473.1 | |
| 108 | 深圳拟兰 | Apostasia shenzhenica | As | 兰科 | 拟兰属 | PKA54423.1 | - |
| 109 | - | Sarocalamus faberi | Sf | 兰科 | - | ATE87948.1 | - |
| 110 | 小果野芭蕉 | Musa acuminata subsp. malaccensis | Ma | 芭蕉科 | 芭蕉属 | XP_009409470.1 | - |
| 111 | 小兰屿蝴蝶兰 | Phalaenopsis equestris | Phe | 兰科 | 蝴蝶兰属 | XP_020574860.1 | - |
| 112 | 铁皮石斛 | Dendrobium catenatum | Dc | 兰科 | 石斛属 | XP_020688760.1 | - |
| 113 | 玉米 | Zea mays | Zm | 禾本科 | 玉蜀黍属 | XP_020397542.1 | - |
| 114 | 玉米 | Z. mays | Zm | 禾本科 | 玉蜀黍属 | ACG37450.1 | |
| 115 | 水稻 | Oryza sativa | Os | 禾本科 | 稻属 | NP_001042508.1 | |
| 116 | 水稻 | O. sativa | Os | 禾本科 | 稻属 | NP_001046572.1 | |
| 117 | 盐地鼠尾粟 | Sporobolus virginicus | Sv | 禾本科 | 鼠尾粟属 | LC342075.1 | |
| 118 | 二穗短柄草 | Brachypodium distachyon | Bd | 禾本科 | 短柄草属 | XP_003572598.1 | - |
| 119 | 高粱 | Sorghum bicolor | Sb | 禾本科 | 高粱属 | XP_002453713.1 | - |
| 120 | - | Dichanthelium oligosanthes | Do | 禾本科 | 二型花属 | OEL25601.1 |
...
... DREPP蛋白的氨基酸序列一般可分为氨基端区域、中央区域和羧基端区域(
... 微管在植物生长发育过程中发挥至关重要的作用, 微管的稳定性、组织结构和动力学都受到微管调控蛋白的调控(
Proteomic discovery of H2O2 response in roots and functional characterization of PutGLP gene from alkaligrass
1
2018
... DREPPs通过Ca2+依赖的方式调节微丝和微管的组织排列, 从而调控植物的生长发育; 并可通过调控ABA和SA等信号途径参与植物应答干旱和低温等逆境胁迫过程(
Purification, properties, and molecular cloning of a novel Ca 2+-binding protein in radish vacuoles
0
2000
1
2014
... PtdInsPs是植物生物膜中含量较低的一类磷脂, 对于细胞信号转导、细胞结构维持、膜流动性调控以及物质转运等过程具有重要作用(
Physiological and molecular features of Puccinellia tenuiflora tolerating salt and alkaline-salt stress
1
2013
... DREPPs通过Ca2+依赖的方式调节微丝和微管的组织排列, 从而调控植物的生长发育; 并可通过调控ABA和SA等信号途径参与植物应答干旱和低温等逆境胁迫过程(
Na2CO3-responsive mechanisms in halophyte Puccinellia tenuiflora roots revealed by physiological and proteomic analyses
1
2016
... DREPPs通过Ca2+依赖的方式调节微丝和微管的组织排列, 从而调控植物的生长发育; 并可通过调控ABA和SA等信号途径参与植物应答干旱和低温等逆境胁迫过程(
Abiotic stress signaling and responses in plants
1
2016
... 低温限制植物在地球上的分布, 减少作物产量, 是一种常见的非生物胁迫(
MAP18 regulates the direction of pollen tube growth in Arabidopsis by modulating F-actin organization
3
2013
... 发育调节质膜多肽(developmentally regulated plasma membrane polypeptide, DREPP)蛋白是一类与质膜相关的Ca2+结合蛋白.它可以通过与微管和微丝结合调节细胞骨架结构, 从而调控根和花粉管的发育(
... AtPCaP2在拟南芥根毛中的表达量较高, 这暗示其在根毛中发挥重要作用(
... AtPCaP1和AtPCaP2在拟南芥花粉与花粉管中的表达量均很高.atpcap2突变体与AtPCaP2过表达体的花粉管扭曲, 生长方向异常, 甚至环绕胚珠生长, 无法完成受精, 从而造成结实率下降.atpcap2突变体与AtPCaP2过表达体花粉管顶端及亚顶端区内微丝的组织排列方式发生改变.此外, 免疫共沉淀实验结果表明, AtPCaP1可直接与微丝结合并具有微弱的微丝切割活性; 在10 μmol∙L-1 Ca2+的条件下, 微丝切割活性明显增强.与野生型相比, atpcap1突变体植株花粉管亚顶端的微丝束更大, 且密度更高, 这表明AtPCaP1的缺失导致花粉管亚顶端微丝无法被切割.同时, 将AtPCaP1中微丝与Ca2+结合关键位点(分别为Asn5和184-188位VEEKK域中E185和K188)突变后, 回补atpcap1突变体, 无法恢复其正常花粉管表型.由此推测, AtPCaP1和AtPCaP2通过调节花粉管顶端及亚顶端微丝的组织排列和动态变化调控花粉管顶端生长(
备案号: 京ICP备16067583号-21
版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由北京玛格泰克科技发展有限公司设计开发

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}