Corresponding authors:* E-mail: wujianming2004@126.com* E-mail: 389001826@qq.com First author contact:? These authors contributed equally to this paper Received:2018-06-19Accepted:2018-12-10Online:2019-07-10
Abstract In this study, Saccharum officinarum GT42 was used as the research material. Second internode tissue of sugarcane at the non-elongation stage (Ls1), early elongation stage (Ls2) and elongation stage (Ls3) was collected to measure plant hormones such as gibberellic acid (GA), indole acetic acid (IAA), brassinosteroids (BR), cytokinin (CTK), ethylene (ETH) and abscisic acid (ABA). The differential expression of the key genes GA20-Oxidase1, GID1 and GAI was analyzed by qRT-PCR. During the elongation stage, the contents of GA and IAA showed an upward trend, but CTK and ABA contents showed a downward trend. ETH content was increased at first and then decreased, whereas BR content did not change. The expression of GA20-Oxidase 1 and GID1 increased, and that of GAI decreased, which was closely related to the changes in GA content. Therefore, we considered that the internode elongation of sugarcane is mainly related to GA and IAA, then are CTK and ABA, and ETH is regulated by IAA to affect internode elongation. The interaction between these phytohormones regulates the expression of GA20-Oxidase 1, GID1 and GAI, which affects the content and signal transduction of GA, thereby affecting the internode elongation of sugarcane. The study clarifies the differential expression of key genes in the gibberellin biosynthesis pathway and signal transduction process, along with the dynamic changes of plant hormone content during internode elongation of sugarcane. Keywords:phytohormone;gibberellin;gene expression analysis;internode elongation;sugarcane
PDF (1342KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 范业赓, 丘立杭, 黄杏, 周慧文, 甘崇琨, 李杨瑞, 杨荣仲, 吴建明, 陈荣发. 甘蔗节间伸长过程赤霉素生物合成关键基因的表达及相关植物激素动态变化. 植物学报, 2019, 54(4): 486-496 doi:10.11983/CBB18139 Fan Yegeng, Qiu Lihang, Huang Xing, Zhou Huiwen, Gan Chongkun, Li Yangrui, Yang Rongzhong, Wu Jianming, Chen Rongfa. Expression Analysis of Key Genes in Gibberellin Biosynthesis and Related Phytohormonal Dynamics During Sugarcane Internode Elongation. Chinese Bulletin of Botany, 2019, 54(4): 486-496 doi:10.11983/CBB18139
Table 2 表2 表2不同时期甘蔗的伸长情况(平均值±标准差) Table 2The elongation of sugarcane at different stages (means±SD)
Period
Plant height (cm)
Elongation rate (cm·d-1)
Internode number
Average internode length (cm)
Ls1
39.38±1.10
0.44±0.01
4±1
6.09±0.28
Ls2
68.37±1.25**
0.97±0.07**
7±1**
8.05±0.18**
Ls3
147.13±0.85**
2.63±0.06**
12±1**
11.43±0.07**
** indicate extremely significant differences (P<0.01). Ls1, Ls2 and Ls3 represents the non-elongation period, early elongation period and elongation period of sugarcane, respectively. ** 表示极显著性差异(P<0.01)。Ls1、Ls2和Ls3分别表示甘蔗未伸长期、伸长初期和伸长盛期。
(A) 未伸长期(Ls1)甘蔗株高; (B) 伸长初期(Ls2)甘蔗株高; (C) 伸长盛期(Ls3)甘蔗株高。Bars=10 cm Figure 1Changes of sugarcane elongation at different stages
(A) The sugarcane plant height of non-elongation stage (Ls1); (B) The sugarcane plant height of early elongation stage (Ls2); (C) The sugarcane plant height of elongation stage (Ls3). Bars=10 cm
(A) 赤霉素(GA)含量; (B) GA20-Oxidase1的表达量; (C) GID1的表达量; (D) GAI的表达量。不同小写字母表示差异显著(P<0.05), 不同大写字母表示差异极显著(P<0.01)。Ls1-Ls3同表2。 Figure 3The changes of gibberellin content and genes differential expression of synthetic pathways during sugarcane internode elongation
(A) The content of gibberellic acid (GA); (B) The expression level of GA20-Oxidase1; (C) The expression level of GID1; (D) The expression level of GAI. The different lowercase letters indicate significant differences (P<0.05), and the different capital letters indicate extremely significant differences (P<0.01). Ls1-Ls3 see Table 2.
AbelesFB, MorganPW, SaltveitME ( 1992). Ethylene in Plant Biology. San Diego: Academic Press. pp. 123-135. [本文引用: 1]
AyanoM, KaniT, KojimaM, SakakibaraH, KitaokaT, KurohaT, Angeles-ShimRB, KitanoH, NagaiK, AshikariM ( 2014). Gibberellin biosynthesis and signal transduction is essential for internode elongation in deepwater rice Plant Cell Environ 37, 2313-2324. [本文引用: 1]
ChoiYH, YoshizawaK, KobayashiM, SakuraiA ( 1995). Distribution of endogenous gibberellins in vegetative shoots of rice Plant Cell Physiol 36, 997-1001. [本文引用: 1]
ColesJP, PhillipsAL, CrokerSJ, García-LepeR, LewisMJ, HeddenP ( 1999). Modification of gibberellin production and plant development in Arabidopsis by sense and antisense expression of gibberellin 20-oxidase genes Plant J 17, 547-556. [本文引用: 2]
CuiK, HeCY, ZhangJG, DuanAG, ZengYF ( 2012). Temporal and spatial profiling of internode elongation-associated protein expression in rapidly growing culms of bamboo J Proteome Res 11, 2492-2507. [本文引用: 2]
CutlerAJ, KrochkoJE ( 1999). Formation and breakdown of ABA Trends Plant Sci 4, 472-478. [本文引用: 1]
De LucasM, DavièreJM, Rodríguez-FalcónM, PontinM, Iglesias-PedrazJM, LorrainS, FankhauserC, BlázquezMA, TitarenkoE, PratS ( 2008). A molecular framework for light and gibberellin control of cell elongation Nature 451, 480-484. [本文引用: 1]
ErikssonME, HoffmanD, KadukM, MauriatM, MoritzT ( 2015). Transgenic hybrid aspen trees with increased gibberellin (GA) concentrations suggest that GA acts in parallel with FLOWERING LOCUS T2 to control shoot elongation New Phytol 205, 1288-1295. [本文引用: 2]
FengSH, MartinezC, GusmaroliG, WangY, ZhouJL, WangF, ChenLY, YuL, Iglesias-PedrazJM, KircherS, Sch?ferE, FuXD, FanLM, DengXW ( 2008). Coordinated regulation of Arabidopsis thaliana development by light and gibberellins Nature 451, 475-479. [本文引用: 1]
GaoJ, ZhangY, ZhangCL, QiFY, LiXP, MuSH, PengZH ( 2014). Characterization of the floral transcriptome of Moso bamboo (Phyllostachys edulis) at different flowering developmental stages by transcriptome sequencing and RNA-seq analysis PLoS One 9, e98910. [本文引用: 1]
HattoriY, NagaiK, FurukawaS, SongXJ, KawanoR, SakakibaraH, WuJZ, MatsumotoT, YoshimuraA, KitanoH, MatsuokaM, MoriH, AshikariM ( 2009). The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water Nature 460, 1026-1030. [本文引用: 3]
HeCY, CuiK, ZhangJG, DuanAG, ZengYF ( 2013). Next-generation sequencing-based mRNA and microRNA expression profiling analysis revealed pathways involved in the rapid growth of developing culms in Moso bamboo BMC Plant Biol 13, 119. [本文引用: 1]
HeddenP, PhillipsAL ( 2000). Gibberellin metabolism: new insights revealed by the genes Trends Plant Sci 5, 523-530. [本文引用: 1]
KendeH, van der KnaapE, ChoHT ( 1998). Deepwater rice: a model plant to study stem elongation Plant Physiol 118, 1105-1110. [本文引用: 2]
LawitSJ, WychHM, XuDP, KunduS, TomesDT ( 2010). Maize DELLA proteins dwarf plant8 and dwarf plant9 as modulators of plant development Plant Cell Physiol 51, 1854-1868. [本文引用: 1]
LeeS, ChengH, KingKE, WangW, HeYW, HussainA, LoJ, HarberdNP, PengJR ( 2002). Gibberellin regulates Arabidopsis seed germination via RGL2, a GAI/RGA-like gene whose expression is up-regulated following imbibition Genes Dev 16, 646-658. [本文引用: 1]
LiLF, ZhangWB, ZhangLL, LiN, PengJZ, WangYQ, ZhongCM, YangYP, SunSL, LiangS, WangXJ ( 2015). Transcriptomic insights into antagonistic effects of gibberellin and abscisic acid on petal growth in Gerbera hybrida Front Plant Sci 6, 168. [本文引用: 2]
MagomeH, NomuraT, HanadaA, Takeda-KamiyaN, OhnishiT, ShinmaY, KatsumataT, KawaideH, KamiyaY, YamaguchiS ( 2013). CYP714B1 and CYP 714B2 encode gibberellin 13-oxidases that reduce gibberellin activity in rice Proc Natl Acad Sci USA 110, 1947-1952. [本文引用: 1]
MauriatM, MoritzT ( 2009). Analyses of GA20ox- and GID1-over-expressing aspen suggest that gibberellins play two distinct roles in wood formation Plant J 58, 989-1003. [本文引用: 1]
PengJR, CarolPO, RichardsDE, KingKE, CowlingRJ, MurphyGP, HarberdNP ( 1997). The Arabidopsis GAI gene defines a signaling pathway that negatively regulates gibberellin responses Genes Dev 11, 3194-3205. [本文引用: 1]
PengZH, ZhangCL, ZhangY, HuT, MuSH, LiXP, GaoJ ( 2013). Transcriptome sequencing and analysis of the fast growing shoots of Moso bamboo (Phyllostachys edulis) PLoS One 8, e78944. [本文引用: 1]
PierikR, CuppensMLC, VoesenekLACJ, VisserEJW ( 2004). Interactions between ethylene and gibberellins in phytochrome-mediated shade avoidance responses in tobacco Plant Physiol 136, 2928-2936. [本文引用: 1]
PierikR, VisserEJW, De KroonH, VoesenekLACJ ( 2003). Ethylene is required in tobacco to successfully compete with proximate neighbours Plant Cell Environ 26, 1229-1234. [本文引用: 1]
QinXQ, ZeevaartJAD ( 2002). Overexpression of a 9-cis-epoxycarotenoid dioxygenase gene in Nicotiana plumbaginifolia increases abscisic acid and phaseic acid levels and enhances drought tolerance Plant Physiol 128, 544-551. [本文引用: 1]
RieuI, Ruiz-RiveroO, Fernandez-GarciaN, GriffithsJ, PowersSJ, GongF, LinhartovaT, ErikssonS, NilssonO, ThomasSG, PhillipsAL, HeddenP ( 2008). The gibberellin biosynthetic genes AtGA20ox1 and AtGA20- ox2 act, partially redundantly, to promote growth and development throughout the Arabidopsis life cycle Plant J 53, 488-504. [本文引用: 2]
RossJJ, O’NeillDP, SmithJJ, KerckhoffsLHJ, ElliotRC ( 2000). Evidence that auxin promotes gibberellin A1 biosynthesis in pea Plant J 21, 547-552. [本文引用: 2]
TakatsukaH, UmedaM ( 2014). Hormonal control of cell division and elongation along differentiation trajectories in roots J Exp Bot 65, 2633-2643. [本文引用: 2]
ThomasGS, HeddenP ( 2018). Gibberellin metabolism and signal transduction. In: Hedden P, Thomas SG, eds. Annual Plant Reviews Washington: John Wiley & Sons. pp. 147-184. [本文引用: 1]
... 关基因与植物节间伸长的关系进行探究(Rieu et al., 2008; Mauriat and Moritz, 2009; Lawit et al., 2010; 李兴亮等, 2011; Eriksson et al., 2015; 魏强和丁雨龙, 2017), 通过对植物节间伸长动态发生过程中的差异表达基因进行分析, 获得了一些与节间伸长相关的差异基因或蛋白(Cui et al., 2012; He et al., 2013; Peng et al., 2013; Gao et al., 2014; 张珂, 2014; 张春玲, 2014; 张莹等, 2015).但上述研究主要集中于拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、毛竹(Phyllostachys heterocycla)和玉米(Zea mays)等植物, 而在甘蔗中的研究深度和广度亟待拓宽.本研究以甘蔗优良品种桂糖42号(GT42)为材料, 分别于未伸长期(9-10叶龄) (Ls1)、伸长初期(12-13叶龄) (Ls2)和伸长盛期(15-16叶龄) (Ls3) 3个时期取甘蔗第2片真叶(自顶部起)对应的节间组织, 测定其赤霉素(GA)、生长素(IAA)、油菜素甾醇(BR)、细胞分裂素(CTK)、乙烯(ETH)和脱落酸(ABA)的含量, 并分析赤霉素生物合成和信号转导关键基因GA20- Oxidase1、GID1和GAI的差异表达, 旨在为解析赤霉素等植物激素对甘蔗节间伸长过程的调控作用奠定基础. ...
Formation and breakdown of ABA 1 1999
... ABA是植物生长发育过程中的重要信号分子, 与GA协同调控组织细胞的伸长(Li et al., 2015).本研究表明, 在不同伸长时期, 节间组织中ABA的含量随着节间伸长而不断减少, 可能是与GA拮抗作用的结果(Li et al., 2015), 也可能是在非胁迫条件下ABA通过激活其分解代谢途径进而限制自身的累积, 以响应自身生长发育的需要(Cutler and Krochko, 1999; Qin and Zeevaart, 2002).本研究结果与在毛竹和小麦(Triticum aestivum)的不同发育期, 节间发育过程中不同器官GA和ABA含量的动态变化相一致(丁兴萃, 1997; 刁家连和何钟佩, 1998; 吴良如和萧江华, 1998).由此表明, ABA参与正常生理条件下甘蔗节间伸长调控, 其在茎节中的过量积累可能对茎秆伸长不利. ...
A molecular framework for light and gibberellin control of cell elongation 1 2008
... 赤霉素合成和代谢受到许多内外因素的影响(De Lucas et al., 2008), 适当浓度的GAs能够促进植物的伸长生长.GA20-Oxidase是植物体GAs生物合成的关键限速酶, 它的表达量决定着活性GAs的含量; 内源GAs及外界环境因子对植物生长发育的调节与GA20-Oxidase1转录水平有关(刘芳等, 2013).已有大量研究证明, 过表达GA20-Oxidase1会引起植株体内赤霉素含量增加, 进而促进植物节间的伸长(Coles et al., 1999; Rieu et al., 2008; Eriksson et al., 2015; Voorend et al., 2016).本研究表明, 在甘蔗节间伸长过程中, GA20-Oxidase1的表达量总体呈上升趋势, 虽然伸长初期(Ls2)其表达下降, 但与未伸长期(Ls1)的差异不显著, 而伸长盛期(Ls3)其表达量大幅增加(图3B); 此时的GA含量呈逐渐上升趋势(图3A).这表明GA20-Oxidase1的总体表达与GA含量变化一致, 暗示GA20-Oxidase1的表达量与GA含量呈正相关, 也表明GA20-Oxidase1的表达对节间伸长有利.这与前人的研究结果基本相符.而伸长初期(Ls2) GA20-Oxidase1表达量下降可能是GA反馈调节引起(Hedden and Phillips, 2000). ...
Transgenic hybrid aspen trees with increased gibberellin (GA) concentrations suggest that GA acts in parallel with FLOWERING LOCUS T2 to control shoot elongation 2 2015
... 关基因与植物节间伸长的关系进行探究(Rieu et al., 2008; Mauriat and Moritz, 2009; Lawit et al., 2010; 李兴亮等, 2011; Eriksson et al., 2015; 魏强和丁雨龙, 2017), 通过对植物节间伸长动态发生过程中的差异表达基因进行分析, 获得了一些与节间伸长相关的差异基因或蛋白(Cui et al., 2012; He et al., 2013; Peng et al., 2013; Gao et al., 2014; 张珂, 2014; 张春玲, 2014; 张莹等, 2015).但上述研究主要集中于拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、毛竹(Phyllostachys heterocycla)和玉米(Zea mays)等植物, 而在甘蔗中的研究深度和广度亟待拓宽.本研究以甘蔗优良品种桂糖42号(GT42)为材料, 分别于未伸长期(9-10叶龄) (Ls1)、伸长初期(12-13叶龄) (Ls2)和伸长盛期(15-16叶龄) (Ls3) 3个时期取甘蔗第2片真叶(自顶部起)对应的节间组织, 测定其赤霉素(GA)、生长素(IAA)、油菜素甾醇(BR)、细胞分裂素(CTK)、乙烯(ETH)和脱落酸(ABA)的含量, 并分析赤霉素生物合成和信号转导关键基因GA20- Oxidase1、GID1和GAI的差异表达, 旨在为解析赤霉素等植物激素对甘蔗节间伸长过程的调控作用奠定基础. ...
... 赤霉素合成和代谢受到许多内外因素的影响(De Lucas et al., 2008), 适当浓度的GAs能够促进植物的伸长生长.GA20-Oxidase是植物体GAs生物合成的关键限速酶, 它的表达量决定着活性GAs的含量; 内源GAs及外界环境因子对植物生长发育的调节与GA20-Oxidase1转录水平有关(刘芳等, 2013).已有大量研究证明, 过表达GA20-Oxidase1会引起植株体内赤霉素含量增加, 进而促进植物节间的伸长(Coles et al., 1999; Rieu et al., 2008; Eriksson et al., 2015; Voorend et al., 2016).本研究表明, 在甘蔗节间伸长过程中, GA20-Oxidase1的表达量总体呈上升趋势, 虽然伸长初期(Ls2)其表达下降, 但与未伸长期(Ls1)的差异不显著, 而伸长盛期(Ls3)其表达量大幅增加(图3B); 此时的GA含量呈逐渐上升趋势(图3A).这表明GA20-Oxidase1的总体表达与GA含量变化一致, 暗示GA20-Oxidase1的表达量与GA含量呈正相关, 也表明GA20-Oxidase1的表达对节间伸长有利.这与前人的研究结果基本相符.而伸长初期(Ls2) GA20-Oxidase1表达量下降可能是GA反馈调节引起(Hedden and Phillips, 2000). ...
Coordinated regulation of Arabidopsis thaliana development by light and gibberellins 1 2008
... DELLA蛋白负向调节GA信号转导, 抑制赤霉素的合成, 从而阻遏植物生长发育.敲除GAI后, 植株表现出PAC (paclobutrazol, GAs生物合成抑制剂)抗性, 但不能恢复其矮化表型(Peng et al., 1997).进一步分析显示, GAs促进植物生长发育是通过GID1介导活性GAs降解DELLA蛋白来完成(Feng et al., 2008; Gao et al., 2008; Wang and Deng, 2014).因此, GID1基因过量表达能够使植物矮化突变体株高得到较大程度的恢复(Xiao et al., 2006; 董静等, 2009; 李兴亮等, 2011); 相反, GAI基因的表达促使DELLA蛋白积累进而抑制生长(陈宇杰等, 2017).在甘蔗节间伸长不同时期, GID1基因的表达量呈单边显著上升趋势, 在伸长盛期(Ls3)表达最活跃, 与此同时GA含量积累最大, 节间也以2.63 cm·d-1的速度快速生长; 相反, GAI基因的表达量呈单边明显下降趋势.结合前人的研究结果, 本研究表明甘蔗节间GID1和GAI的差异表达与GA含量动态密切相关, 且GID1的高表达和GAI的低表达是赤霉素响应甘蔗节间伸长的遗传基础. ...
Characterization of the floral transcriptome of Moso bamboo (Phyllostachys edulis) at different flowering developmental stages by transcriptome sequencing and RNA-seq analysis 1 2014
... 关基因与植物节间伸长的关系进行探究(Rieu et al., 2008; Mauriat and Moritz, 2009; Lawit et al., 2010; 李兴亮等, 2011; Eriksson et al., 2015; 魏强和丁雨龙, 2017), 通过对植物节间伸长动态发生过程中的差异表达基因进行分析, 获得了一些与节间伸长相关的差异基因或蛋白(Cui et al., 2012; He et al., 2013; Peng et al., 2013; Gao et al., 2014; 张珂, 2014; 张春玲, 2014; 张莹等, 2015).但上述研究主要集中于拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、毛竹(Phyllostachys heterocycla)和玉米(Zea mays)等植物, 而在甘蔗中的研究深度和广度亟待拓宽.本研究以甘蔗优良品种桂糖42号(GT42)为材料, 分别于未伸长期(9-10叶龄) (Ls1)、伸长初期(12-13叶龄) (Ls2)和伸长盛期(15-16叶龄) (Ls3) 3个时期取甘蔗第2片真叶(自顶部起)对应的节间组织, 测定其赤霉素(GA)、生长素(IAA)、油菜素甾醇(BR)、细胞分裂素(CTK)、乙烯(ETH)和脱落酸(ABA)的含量, 并分析赤霉素生物合成和信号转导关键基因GA20- Oxidase1、GID1和GAI的差异表达, 旨在为解析赤霉素等植物激素对甘蔗节间伸长过程的调控作用奠定基础. ...
Evolutionarily conserved DELLA-mediated gibberellin signaling in plants 1 2008
... DELLA蛋白负向调节GA信号转导, 抑制赤霉素的合成, 从而阻遏植物生长发育.敲除GAI后, 植株表现出PAC (paclobutrazol, GAs生物合成抑制剂)抗性, 但不能恢复其矮化表型(Peng et al., 1997).进一步分析显示, GAs促进植物生长发育是通过GID1介导活性GAs降解DELLA蛋白来完成(Feng et al., 2008; Gao et al., 2008; Wang and Deng, 2014).因此, GID1基因过量表达能够使植物矮化突变体株高得到较大程度的恢复(Xiao et al., 2006; 董静等, 2009; 李兴亮等, 2011); 相反, GAI基因的表达促使DELLA蛋白积累进而抑制生长(陈宇杰等, 2017).在甘蔗节间伸长不同时期, GID1基因的表达量呈单边显著上升趋势, 在伸长盛期(Ls3)表达最活跃, 与此同时GA含量积累最大, 节间也以2.63 cm·d-1的速度快速生长; 相反, GAI基因的表达量呈单边明显下降趋势.结合前人的研究结果, 本研究表明甘蔗节间GID1和GAI的差异表达与GA含量动态密切相关, 且GID1的高表达和GAI的低表达是赤霉素响应甘蔗节间伸长的遗传基础. ...
The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water 3 2009
... 植物激素参与调控植物生长发育的各个方面, 在植物各生长阶段和环境胁迫响应过程中均发挥重要作用(段娜等, 2015; 杜维等, 2018).前人研究表明, 在禾本科作物中, 赤霉素和生长素促进节间伸长和细胞分裂, 进而显著影响其茎秆伸长, 且赤霉素与茎秆伸长生长的关系最密切(Kende et al., 1998; Ross et al., 2000; 季兰和杨仁崔, 2002; Hattori et al., 2009; Magome et al., 2013; Takatsuka and Umeda, 2014; Ayano et al., 2014).Wang等(2015)对毛竹节间不同发育时期植物激素变化进行探究, 发现IAA和GA含量呈显著正相关(r=0.712, P<0.05), IAA和GA都与ABA含量呈负相关.张春玲(2014)研究表明毛竹笋发育成茎过程中IAA和GA呈总体上升的动态变化.上述研究结果表明, 节间伸长与生长素和赤霉素含量关系密切.本研究也显示, 在甘蔗茎节伸长过程中, Ls1 (未伸长期)伸长最为缓慢, Ls2 (伸长初期)甘蔗节间伸长加快, Ls3 (伸长盛期)的长速进入加速期; 在这一过程中, GA和IAA含量也不断增加, 但GA含量明显高于IAA, 这可能是由于IAA的协同作用或者促进GA转运与积累所致(Ross et al., 2000; Takatsuka and Umeda, 2014); 也可能是顶端优势抑制IAA的含量在茎节间的分布差异引起(吴建明等, 2009).这表明在正常生理条件下, GA和IAA在甘蔗茎节中的积累有利于节间伸长, GA起主要作用, 而高浓度IAA可能发挥相反的作用. ...
... 乙烯以其催熟作用被大家所熟悉, 曾一度被认为抑制植物生长(Abeles et al., 1992).后来发现乙烯的积累是深水水稻节间伸长所必需(Hattori et al., 2009).将外源乙烯作用于深水水稻的茎段, 不仅会诱导茎伸长, 而且诱导GA代谢酶基因的表达(Choi et al., 1995), 而低浓度外源乙烯还能够促进番茄(Lycopersicon esculentum)茎的伸长(Pierik et al., 2003).此外, 乙烯对茎伸长还有间接效应, 即增加幼嫩节间对GA的敏感性(Kende et al., 1998).上述研究表明, 乙烯在不同条件下对茎伸长的效应差异较大.本研究表明, 随着甘蔗节间不断伸长, 乙烯在节间组织中不断积累.从不同时期看, Ls1时含量较低, 到Ls2时剧增, Ls3时有所下降, 这暗示未伸长期(Ls1)细胞横向生长和伸长生长同时进行, 导致乙烯含量较低, 在伸长初期(Ls2), 生长素的浓度可能超过最适浓度, 进而促进乙烯含量增加.Ls2至Ls3时期乙烯的下降可能是由于节间的快速生长需要大量的GA, 而ETH信号的积累可促进活性GA的合成(Pierik et al., 2004). ...
Next-generation sequencing-based mRNA and microRNA expression profiling analysis revealed pathways involved in the rapid growth of developing culms in Moso bamboo 1 2013
... 关基因与植物节间伸长的关系进行探究(Rieu et al., 2008; Mauriat and Moritz, 2009; Lawit et al., 2010; 李兴亮等, 2011; Eriksson et al., 2015; 魏强和丁雨龙, 2017), 通过对植物节间伸长动态发生过程中的差异表达基因进行分析, 获得了一些与节间伸长相关的差异基因或蛋白(Cui et al., 2012; He et al., 2013; Peng et al., 2013; Gao et al., 2014; 张珂, 2014; 张春玲, 2014; 张莹等, 2015).但上述研究主要集中于拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、毛竹(Phyllostachys heterocycla)和玉米(Zea mays)等植物, 而在甘蔗中的研究深度和广度亟待拓宽.本研究以甘蔗优良品种桂糖42号(GT42)为材料, 分别于未伸长期(9-10叶龄) (Ls1)、伸长初期(12-13叶龄) (Ls2)和伸长盛期(15-16叶龄) (Ls3) 3个时期取甘蔗第2片真叶(自顶部起)对应的节间组织, 测定其赤霉素(GA)、生长素(IAA)、油菜素甾醇(BR)、细胞分裂素(CTK)、乙烯(ETH)和脱落酸(ABA)的含量, 并分析赤霉素生物合成和信号转导关键基因GA20- Oxidase1、GID1和GAI的差异表达, 旨在为解析赤霉素等植物激素对甘蔗节间伸长过程的调控作用奠定基础. ...
Gibberellin metabolism: new insights revealed by the genes 1 2000
... 赤霉素合成和代谢受到许多内外因素的影响(De Lucas et al., 2008), 适当浓度的GAs能够促进植物的伸长生长.GA20-Oxidase是植物体GAs生物合成的关键限速酶, 它的表达量决定着活性GAs的含量; 内源GAs及外界环境因子对植物生长发育的调节与GA20-Oxidase1转录水平有关(刘芳等, 2013).已有大量研究证明, 过表达GA20-Oxidase1会引起植株体内赤霉素含量增加, 进而促进植物节间的伸长(Coles et al., 1999; Rieu et al., 2008; Eriksson et al., 2015; Voorend et al., 2016).本研究表明, 在甘蔗节间伸长过程中, GA20-Oxidase1的表达量总体呈上升趋势, 虽然伸长初期(Ls2)其表达下降, 但与未伸长期(Ls1)的差异不显著, 而伸长盛期(Ls3)其表达量大幅增加(图3B); 此时的GA含量呈逐渐上升趋势(图3A).这表明GA20-Oxidase1的总体表达与GA含量变化一致, 暗示GA20-Oxidase1的表达量与GA含量呈正相关, 也表明GA20-Oxidase1的表达对节间伸长有利.这与前人的研究结果基本相符.而伸长初期(Ls2) GA20-Oxidase1表达量下降可能是GA反馈调节引起(Hedden and Phillips, 2000). ...
Deepwater rice: a model plant to study stem elongation 2 1998
... 植物激素参与调控植物生长发育的各个方面, 在植物各生长阶段和环境胁迫响应过程中均发挥重要作用(段娜等, 2015; 杜维等, 2018).前人研究表明, 在禾本科作物中, 赤霉素和生长素促进节间伸长和细胞分裂, 进而显著影响其茎秆伸长, 且赤霉素与茎秆伸长生长的关系最密切(Kende et al., 1998; Ross et al., 2000; 季兰和杨仁崔, 2002; Hattori et al., 2009; Magome et al., 2013; Takatsuka and Umeda, 2014; Ayano et al., 2014).Wang等(2015)对毛竹节间不同发育时期植物激素变化进行探究, 发现IAA和GA含量呈显著正相关(r=0.712, P<0.05), IAA和GA都与ABA含量呈负相关.张春玲(2014)研究表明毛竹笋发育成茎过程中IAA和GA呈总体上升的动态变化.上述研究结果表明, 节间伸长与生长素和赤霉素含量关系密切.本研究也显示, 在甘蔗茎节伸长过程中, Ls1 (未伸长期)伸长最为缓慢, Ls2 (伸长初期)甘蔗节间伸长加快, Ls3 (伸长盛期)的长速进入加速期; 在这一过程中, GA和IAA含量也不断增加, 但GA含量明显高于IAA, 这可能是由于IAA的协同作用或者促进GA转运与积累所致(Ross et al., 2000; Takatsuka and Umeda, 2014); 也可能是顶端优势抑制IAA的含量在茎节间的分布差异引起(吴建明等, 2009).这表明在正常生理条件下, GA和IAA在甘蔗茎节中的积累有利于节间伸长, GA起主要作用, 而高浓度IAA可能发挥相反的作用. ...
... 乙烯以其催熟作用被大家所熟悉, 曾一度被认为抑制植物生长(Abeles et al., 1992).后来发现乙烯的积累是深水水稻节间伸长所必需(Hattori et al., 2009).将外源乙烯作用于深水水稻的茎段, 不仅会诱导茎伸长, 而且诱导GA代谢酶基因的表达(Choi et al., 1995), 而低浓度外源乙烯还能够促进番茄(Lycopersicon esculentum)茎的伸长(Pierik et al., 2003).此外, 乙烯对茎伸长还有间接效应, 即增加幼嫩节间对GA的敏感性(Kende et al., 1998).上述研究表明, 乙烯在不同条件下对茎伸长的效应差异较大.本研究表明, 随着甘蔗节间不断伸长, 乙烯在节间组织中不断积累.从不同时期看, Ls1时含量较低, 到Ls2时剧增, Ls3时有所下降, 这暗示未伸长期(Ls1)细胞横向生长和伸长生长同时进行, 导致乙烯含量较低, 在伸长初期(Ls2), 生长素的浓度可能超过最适浓度, 进而促进乙烯含量增加.Ls2至Ls3时期乙烯的下降可能是由于节间的快速生长需要大量的GA, 而ETH信号的积累可促进活性GA的合成(Pierik et al., 2004). ...
Maize DELLA proteins dwarf plant8 and dwarf plant9 as modulators of plant development 1 2010
... 关基因与植物节间伸长的关系进行探究(Rieu et al., 2008; Mauriat and Moritz, 2009; Lawit et al., 2010; 李兴亮等, 2011; Eriksson et al., 2015; 魏强和丁雨龙, 2017), 通过对植物节间伸长动态发生过程中的差异表达基因进行分析, 获得了一些与节间伸长相关的差异基因或蛋白(Cui et al., 2012; He et al., 2013; Peng et al., 2013; Gao et al., 2014; 张珂, 2014; 张春玲, 2014; 张莹等, 2015).但上述研究主要集中于拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、毛竹(Phyllostachys heterocycla)和玉米(Zea mays)等植物, 而在甘蔗中的研究深度和广度亟待拓宽.本研究以甘蔗优良品种桂糖42号(GT42)为材料, 分别于未伸长期(9-10叶龄) (Ls1)、伸长初期(12-13叶龄) (Ls2)和伸长盛期(15-16叶龄) (Ls3) 3个时期取甘蔗第2片真叶(自顶部起)对应的节间组织, 测定其赤霉素(GA)、生长素(IAA)、油菜素甾醇(BR)、细胞分裂素(CTK)、乙烯(ETH)和脱落酸(ABA)的含量, 并分析赤霉素生物合成和信号转导关键基因GA20- Oxidase1、GID1和GAI的差异表达, 旨在为解析赤霉素等植物激素对甘蔗节间伸长过程的调控作用奠定基础. ...
Gibberellin regulates Arabidopsis seed germination via RGL2, a GAI/RGA-like gene whose expression is up-regulated following imbibition 1 2002
... 植物体内GA浓度和分布调控植物的长势.GA20- Oxidase属于可溶性的双加氧酶, 是赤霉素生物合成过程的关键限速酶和决定性因子(Coles et al., 1999; Lee et al., 2002).在甘蔗节间伸长的过程中, 虽然在伸长和未伸长的节间组织中GA20-Oxidase1的表达差异不显著, 但在伸长初期至盛期其表达量差异达极显著水平; 且节间组织中GA20-Oxidase1的表达量呈总体上升趋势(图3B).这与GA和IAA含量的总体变化动态相一致(图2A, 图3A), 可见, GA20-Oxidase1的表达影响GA和IAA的变化. ...
Transcriptomic insights into antagonistic effects of gibberellin and abscisic acid on petal growth in Gerbera hybrida 2 2015
... ABA是植物生长发育过程中的重要信号分子, 与GA协同调控组织细胞的伸长(Li et al., 2015).本研究表明, 在不同伸长时期, 节间组织中ABA的含量随着节间伸长而不断减少, 可能是与GA拮抗作用的结果(Li et al., 2015), 也可能是在非胁迫条件下ABA通过激活其分解代谢途径进而限制自身的累积, 以响应自身生长发育的需要(Cutler and Krochko, 1999; Qin and Zeevaart, 2002).本研究结果与在毛竹和小麦(Triticum aestivum)的不同发育期, 节间发育过程中不同器官GA和ABA含量的动态变化相一致(丁兴萃, 1997; 刁家连和何钟佩, 1998; 吴良如和萧江华, 1998).由此表明, ABA参与正常生理条件下甘蔗节间伸长调控, 其在茎节中的过量积累可能对茎秆伸长不利. ...
... ).本研究表明, 在不同伸长时期, 节间组织中ABA的含量随着节间伸长而不断减少, 可能是与GA拮抗作用的结果(Li et al., 2015), 也可能是在非胁迫条件下ABA通过激活其分解代谢途径进而限制自身的累积, 以响应自身生长发育的需要(Cutler and Krochko, 1999; Qin and Zeevaart, 2002).本研究结果与在毛竹和小麦(Triticum aestivum)的不同发育期, 节间发育过程中不同器官GA和ABA含量的动态变化相一致(丁兴萃, 1997; 刁家连和何钟佩, 1998; 吴良如和萧江华, 1998).由此表明, ABA参与正常生理条件下甘蔗节间伸长调控, 其在茎节中的过量积累可能对茎秆伸长不利. ...
CYP714B1 and CYP 714B2 encode gibberellin 13-oxidases that reduce gibberellin activity in rice 1 2013
... 植物激素参与调控植物生长发育的各个方面, 在植物各生长阶段和环境胁迫响应过程中均发挥重要作用(段娜等, 2015; 杜维等, 2018).前人研究表明, 在禾本科作物中, 赤霉素和生长素促进节间伸长和细胞分裂, 进而显著影响其茎秆伸长, 且赤霉素与茎秆伸长生长的关系最密切(Kende et al., 1998; Ross et al., 2000; 季兰和杨仁崔, 2002; Hattori et al., 2009; Magome et al., 2013; Takatsuka and Umeda, 2014; Ayano et al., 2014).Wang等(2015)对毛竹节间不同发育时期植物激素变化进行探究, 发现IAA和GA含量呈显著正相关(r=0.712, P<0.05), IAA和GA都与ABA含量呈负相关.张春玲(2014)研究表明毛竹笋发育成茎过程中IAA和GA呈总体上升的动态变化.上述研究结果表明, 节间伸长与生长素和赤霉素含量关系密切.本研究也显示, 在甘蔗茎节伸长过程中, Ls1 (未伸长期)伸长最为缓慢, Ls2 (伸长初期)甘蔗节间伸长加快, Ls3 (伸长盛期)的长速进入加速期; 在这一过程中, GA和IAA含量也不断增加, 但GA含量明显高于IAA, 这可能是由于IAA的协同作用或者促进GA转运与积累所致(Ross et al., 2000; Takatsuka and Umeda, 2014); 也可能是顶端优势抑制IAA的含量在茎节间的分布差异引起(吴建明等, 2009).这表明在正常生理条件下, GA和IAA在甘蔗茎节中的积累有利于节间伸长, GA起主要作用, 而高浓度IAA可能发挥相反的作用. ...
Analyses of GA20ox- and GID1-over-expressing aspen suggest that gibberellins play two distinct roles in wood formation 1 2009
... 关基因与植物节间伸长的关系进行探究(Rieu et al., 2008; Mauriat and Moritz, 2009; Lawit et al., 2010; 李兴亮等, 2011; Eriksson et al., 2015; 魏强和丁雨龙, 2017), 通过对植物节间伸长动态发生过程中的差异表达基因进行分析, 获得了一些与节间伸长相关的差异基因或蛋白(Cui et al., 2012; He et al., 2013; Peng et al., 2013; Gao et al., 2014; 张珂, 2014; 张春玲, 2014; 张莹等, 2015).但上述研究主要集中于拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、毛竹(Phyllostachys heterocycla)和玉米(Zea mays)等植物, 而在甘蔗中的研究深度和广度亟待拓宽.本研究以甘蔗优良品种桂糖42号(GT42)为材料, 分别于未伸长期(9-10叶龄) (Ls1)、伸长初期(12-13叶龄) (Ls2)和伸长盛期(15-16叶龄) (Ls3) 3个时期取甘蔗第2片真叶(自顶部起)对应的节间组织, 测定其赤霉素(GA)、生长素(IAA)、油菜素甾醇(BR)、细胞分裂素(CTK)、乙烯(ETH)和脱落酸(ABA)的含量, 并分析赤霉素生物合成和信号转导关键基因GA20- Oxidase1、GID1和GAI的差异表达, 旨在为解析赤霉素等植物激素对甘蔗节间伸长过程的调控作用奠定基础. ...
The Arabidopsis GAI gene defines a signaling pathway that negatively regulates gibberellin responses 1 1997
... DELLA蛋白负向调节GA信号转导, 抑制赤霉素的合成, 从而阻遏植物生长发育.敲除GAI后, 植株表现出PAC (paclobutrazol, GAs生物合成抑制剂)抗性, 但不能恢复其矮化表型(Peng et al., 1997).进一步分析显示, GAs促进植物生长发育是通过GID1介导活性GAs降解DELLA蛋白来完成(Feng et al., 2008; Gao et al., 2008; Wang and Deng, 2014).因此, GID1基因过量表达能够使植物矮化突变体株高得到较大程度的恢复(Xiao et al., 2006; 董静等, 2009; 李兴亮等, 2011); 相反, GAI基因的表达促使DELLA蛋白积累进而抑制生长(陈宇杰等, 2017).在甘蔗节间伸长不同时期, GID1基因的表达量呈单边显著上升趋势, 在伸长盛期(Ls3)表达最活跃, 与此同时GA含量积累最大, 节间也以2.63 cm·d-1的速度快速生长; 相反, GAI基因的表达量呈单边明显下降趋势.结合前人的研究结果, 本研究表明甘蔗节间GID1和GAI的差异表达与GA含量动态密切相关, 且GID1的高表达和GAI的低表达是赤霉素响应甘蔗节间伸长的遗传基础. ...
Transcriptome sequencing and analysis of the fast growing shoots of Moso bamboo (Phyllostachys edulis) 1 2013
... 关基因与植物节间伸长的关系进行探究(Rieu et al., 2008; Mauriat and Moritz, 2009; Lawit et al., 2010; 李兴亮等, 2011; Eriksson et al., 2015; 魏强和丁雨龙, 2017), 通过对植物节间伸长动态发生过程中的差异表达基因进行分析, 获得了一些与节间伸长相关的差异基因或蛋白(Cui et al., 2012; He et al., 2013; Peng et al., 2013; Gao et al., 2014; 张珂, 2014; 张春玲, 2014; 张莹等, 2015).但上述研究主要集中于拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、毛竹(Phyllostachys heterocycla)和玉米(Zea mays)等植物, 而在甘蔗中的研究深度和广度亟待拓宽.本研究以甘蔗优良品种桂糖42号(GT42)为材料, 分别于未伸长期(9-10叶龄) (Ls1)、伸长初期(12-13叶龄) (Ls2)和伸长盛期(15-16叶龄) (Ls3) 3个时期取甘蔗第2片真叶(自顶部起)对应的节间组织, 测定其赤霉素(GA)、生长素(IAA)、油菜素甾醇(BR)、细胞分裂素(CTK)、乙烯(ETH)和脱落酸(ABA)的含量, 并分析赤霉素生物合成和信号转导关键基因GA20- Oxidase1、GID1和GAI的差异表达, 旨在为解析赤霉素等植物激素对甘蔗节间伸长过程的调控作用奠定基础. ...
Interactions between ethylene and gibberellins in phytochrome-mediated shade avoidance responses in tobacco 1 2004
... 乙烯以其催熟作用被大家所熟悉, 曾一度被认为抑制植物生长(Abeles et al., 1992).后来发现乙烯的积累是深水水稻节间伸长所必需(Hattori et al., 2009).将外源乙烯作用于深水水稻的茎段, 不仅会诱导茎伸长, 而且诱导GA代谢酶基因的表达(Choi et al., 1995), 而低浓度外源乙烯还能够促进番茄(Lycopersicon esculentum)茎的伸长(Pierik et al., 2003).此外, 乙烯对茎伸长还有间接效应, 即增加幼嫩节间对GA的敏感性(Kende et al., 1998).上述研究表明, 乙烯在不同条件下对茎伸长的效应差异较大.本研究表明, 随着甘蔗节间不断伸长, 乙烯在节间组织中不断积累.从不同时期看, Ls1时含量较低, 到Ls2时剧增, Ls3时有所下降, 这暗示未伸长期(Ls1)细胞横向生长和伸长生长同时进行, 导致乙烯含量较低, 在伸长初期(Ls2), 生长素的浓度可能超过最适浓度, 进而促进乙烯含量增加.Ls2至Ls3时期乙烯的下降可能是由于节间的快速生长需要大量的GA, 而ETH信号的积累可促进活性GA的合成(Pierik et al., 2004). ...
Ethylene is required in tobacco to successfully compete with proximate neighbours 1 2003
... 乙烯以其催熟作用被大家所熟悉, 曾一度被认为抑制植物生长(Abeles et al., 1992).后来发现乙烯的积累是深水水稻节间伸长所必需(Hattori et al., 2009).将外源乙烯作用于深水水稻的茎段, 不仅会诱导茎伸长, 而且诱导GA代谢酶基因的表达(Choi et al., 1995), 而低浓度外源乙烯还能够促进番茄(Lycopersicon esculentum)茎的伸长(Pierik et al., 2003).此外, 乙烯对茎伸长还有间接效应, 即增加幼嫩节间对GA的敏感性(Kende et al., 1998).上述研究表明, 乙烯在不同条件下对茎伸长的效应差异较大.本研究表明, 随着甘蔗节间不断伸长, 乙烯在节间组织中不断积累.从不同时期看, Ls1时含量较低, 到Ls2时剧增, Ls3时有所下降, 这暗示未伸长期(Ls1)细胞横向生长和伸长生长同时进行, 导致乙烯含量较低, 在伸长初期(Ls2), 生长素的浓度可能超过最适浓度, 进而促进乙烯含量增加.Ls2至Ls3时期乙烯的下降可能是由于节间的快速生长需要大量的GA, 而ETH信号的积累可促进活性GA的合成(Pierik et al., 2004). ...
Overexpression of a 9-cis-epoxycarotenoid dioxygenase gene in Nicotiana plumbaginifolia increases abscisic acid and phaseic acid levels and enhances drought tolerance 1 2002
... ABA是植物生长发育过程中的重要信号分子, 与GA协同调控组织细胞的伸长(Li et al., 2015).本研究表明, 在不同伸长时期, 节间组织中ABA的含量随着节间伸长而不断减少, 可能是与GA拮抗作用的结果(Li et al., 2015), 也可能是在非胁迫条件下ABA通过激活其分解代谢途径进而限制自身的累积, 以响应自身生长发育的需要(Cutler and Krochko, 1999; Qin and Zeevaart, 2002).本研究结果与在毛竹和小麦(Triticum aestivum)的不同发育期, 节间发育过程中不同器官GA和ABA含量的动态变化相一致(丁兴萃, 1997; 刁家连和何钟佩, 1998; 吴良如和萧江华, 1998).由此表明, ABA参与正常生理条件下甘蔗节间伸长调控, 其在茎节中的过量积累可能对茎秆伸长不利. ...
The gibberellin biosynthetic genes AtGA20ox1 and AtGA20- ox2 act, partially redundantly, to promote growth and development throughout the Arabidopsis life cycle 2 2008
... 关基因与植物节间伸长的关系进行探究(Rieu et al., 2008; Mauriat and Moritz, 2009; Lawit et al., 2010; 李兴亮等, 2011; Eriksson et al., 2015; 魏强和丁雨龙, 2017), 通过对植物节间伸长动态发生过程中的差异表达基因进行分析, 获得了一些与节间伸长相关的差异基因或蛋白(Cui et al., 2012; He et al., 2013; Peng et al., 2013; Gao et al., 2014; 张珂, 2014; 张春玲, 2014; 张莹等, 2015).但上述研究主要集中于拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)、毛竹(Phyllostachys heterocycla)和玉米(Zea mays)等植物, 而在甘蔗中的研究深度和广度亟待拓宽.本研究以甘蔗优良品种桂糖42号(GT42)为材料, 分别于未伸长期(9-10叶龄) (Ls1)、伸长初期(12-13叶龄) (Ls2)和伸长盛期(15-16叶龄) (Ls3) 3个时期取甘蔗第2片真叶(自顶部起)对应的节间组织, 测定其赤霉素(GA)、生长素(IAA)、油菜素甾醇(BR)、细胞分裂素(CTK)、乙烯(ETH)和脱落酸(ABA)的含量, 并分析赤霉素生物合成和信号转导关键基因GA20- Oxidase1、GID1和GAI的差异表达, 旨在为解析赤霉素等植物激素对甘蔗节间伸长过程的调控作用奠定基础. ...
... 赤霉素合成和代谢受到许多内外因素的影响(De Lucas et al., 2008), 适当浓度的GAs能够促进植物的伸长生长.GA20-Oxidase是植物体GAs生物合成的关键限速酶, 它的表达量决定着活性GAs的含量; 内源GAs及外界环境因子对植物生长发育的调节与GA20-Oxidase1转录水平有关(刘芳等, 2013).已有大量研究证明, 过表达GA20-Oxidase1会引起植株体内赤霉素含量增加, 进而促进植物节间的伸长(Coles et al., 1999; Rieu et al., 2008; Eriksson et al., 2015; Voorend et al., 2016).本研究表明, 在甘蔗节间伸长过程中, GA20-Oxidase1的表达量总体呈上升趋势, 虽然伸长初期(Ls2)其表达下降, 但与未伸长期(Ls1)的差异不显著, 而伸长盛期(Ls3)其表达量大幅增加(图3B); 此时的GA含量呈逐渐上升趋势(图3A).这表明GA20-Oxidase1的总体表达与GA含量变化一致, 暗示GA20-Oxidase1的表达量与GA含量呈正相关, 也表明GA20-Oxidase1的表达对节间伸长有利.这与前人的研究结果基本相符.而伸长初期(Ls2) GA20-Oxidase1表达量下降可能是GA反馈调节引起(Hedden and Phillips, 2000). ...
Evidence that auxin promotes gibberellin A1 biosynthesis in pea 2 2000
... 植物激素参与调控植物生长发育的各个方面, 在植物各生长阶段和环境胁迫响应过程中均发挥重要作用(段娜等, 2015; 杜维等, 2018).前人研究表明, 在禾本科作物中, 赤霉素和生长素促进节间伸长和细胞分裂, 进而显著影响其茎秆伸长, 且赤霉素与茎秆伸长生长的关系最密切(Kende et al., 1998; Ross et al., 2000; 季兰和杨仁崔, 2002; Hattori et al., 2009; Magome et al., 2013; Takatsuka and Umeda, 2014; Ayano et al., 2014).Wang等(2015)对毛竹节间不同发育时期植物激素变化进行探究, 发现IAA和GA含量呈显著正相关(r=0.712, P<0.05), IAA和GA都与ABA含量呈负相关.张春玲(2014)研究表明毛竹笋发育成茎过程中IAA和GA呈总体上升的动态变化.上述研究结果表明, 节间伸长与生长素和赤霉素含量关系密切.本研究也显示, 在甘蔗茎节伸长过程中, Ls1 (未伸长期)伸长最为缓慢, Ls2 (伸长初期)甘蔗节间伸长加快, Ls3 (伸长盛期)的长速进入加速期; 在这一过程中, GA和IAA含量也不断增加, 但GA含量明显高于IAA, 这可能是由于IAA的协同作用或者促进GA转运与积累所致(Ross et al., 2000; Takatsuka and Umeda, 2014); 也可能是顶端优势抑制IAA的含量在茎节间的分布差异引起(吴建明等, 2009).这表明在正常生理条件下, GA和IAA在甘蔗茎节中的积累有利于节间伸长, GA起主要作用, 而高浓度IAA可能发挥相反的作用. ...
 ,1,*, 陈荣发
,1,*, 陈荣发
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT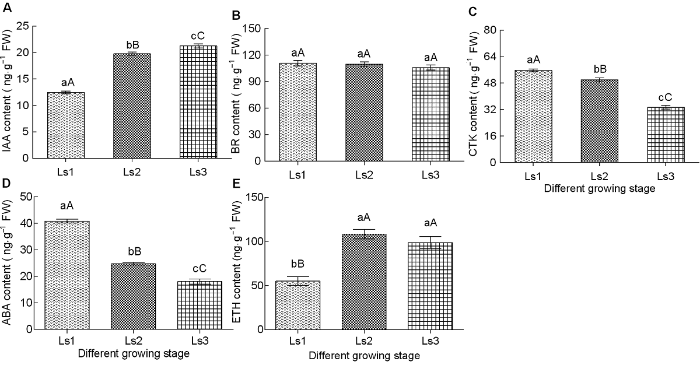 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT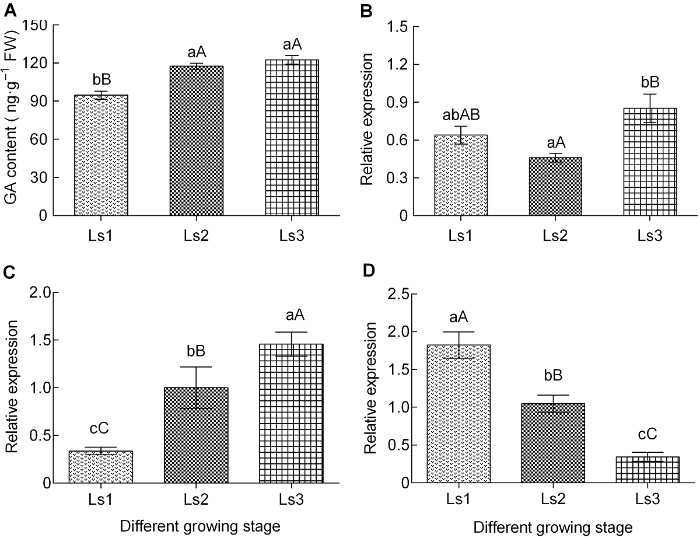 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}