Detection and Analysis of Protein S-nitrosylation in Plants
Lichao Chen?, Ni Zhan?, Yansha Li, Jian Feng, Jianru Zuo,*Institute of Genetics and Developmental Biology, Chinese Academy of Sciences, Beijing 100101, China
Corresponding authors:* E-mail: jrzuo@genetics.ac.cn First author contact:? These authors contributed equally to this paper Received:2019-06-13Accepted:2019-06-20Online:2019-07-10
Abstract S-nitrosylation is an important protein posttranslational modification, involved in covalently linking a nitric oxide (NO) molecule to the thiol group of a cysteine residue to generate S-nitrosothiols. S-nitrosylation regulates multiple biological processes by modulating protein activity, stability, subcellular localization and protein-protein interactions. The biotin-switch assay is one of the most-often used methods to detect and analyze protein S-nitrosylation. In principle, the free thiols in a target protein are first blocked, followed by reducing the S-nitrosothiols of the target protein to free thiols by ascorbate, which are subsequently labelled by biotin to form biotinylated proteins. The biotin-labelled sample was assayed by immunoblotting and mass spectrometry. Here, we present detailed experimental procedures for the in vitro and in vivo biotin-switch methods and give advice on key troubleshooting solutions. Keywords:nitric oxide;S-nitrosylation;biotin-switch assay;plants
PDF (773KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 陈立超, 詹妮, 李彦莎, 冯健, 左建儒. 植物蛋白质S-亚硝基化修饰的检测与分析. 植物学报, 2019, 54(4): 497-502 doi:10.11983/CBB19108 Chen Lichao, Zhan Ni, Li Yansha, Feng Jian, Zuo Jianru. Detection and Analysis of Protein S-nitrosylation in Plants. Chinese Bulletin of Botany, 2019, 54(4): 497-502 doi:10.11983/CBB19108
S-亚硝基化修饰发生在蛋白质的半胱氨酸残基上。一般而言, 蛋白质能否发生亚硝基化修饰取决于半胱氨酸残基周围的氨基酸组成及半胱氨酸残基在蛋白质三维结构上的相对空间位置。通过对不同靶蛋白的修饰位点进行系统分析, 人们发现发生S-亚硝基化修饰的半胱氨酸残基通常位于一个酸-碱-疏水结构域中([HKR]-C-[hydrophobic]X[D-E])或偏酸性环境中(Hess et al., 2005; Hu et al., 2015)。基于上述发现开发的亚硝基化修饰位点预测软件工具(如GPS-SNO)能够预测蛋白质中亚硝基化修饰的候选位点(Xue et al., 2010)。此外, 利用植物PTM Viewer数据库也可以预测植物中S-亚硝基化修饰的潜在位点, 其中包括多个已被实验验证的修饰位点(Willems et al., 2019)。与其它蛋白质翻译后修饰类似, 预测的S-亚硝基化修饰位点及其功能需要实验生物学验证。
在S-亚硝基化修饰的研究中, 通过生物素转化法(biotin switch assay)鉴定蛋白质中S-亚硝基化的修饰位点是目前最常用的方法(Jaffrey and Snyder, 2001)。其原理是将未发生S-亚硝基化修饰的自由巯基用烷化剂封闭, 然后用还原剂(通常用抗坏血酸盐)将被S-亚硝基化的巯基还原为自由巯基, 继而用生物素标记新形成的自由巯基。生物素化的半胱氨酸残基(即被S-亚硝基化修饰的半胱氨酸残基)可以利用生物素抗体进行蛋白质免疫印迹(western blotting)或通过液相色谱-串联质谱(liquid chromatography coupled to tandem mass spectrometry; LC-MS/MS)技术进行检测分析。后者也被广泛用于高通量鉴定S-亚硝基化靶蛋白及修饰靶位点(Hu et al., 2015)。本文对重组蛋白质的体外生物素转化以及拟南芥(Arabidopsis thaliana)体内蛋白质的生物素转化实验流程进行详细介绍, 并对实验中的注意事项做出具体说明, 旨在为本领域的研究提供借鉴及帮助。
1 实验材料
(1) 纯化后的重组蛋白
(2) 拟南芥(Arabidopsis thaliana L.)或其它植物材料
(3) 植物蛋白质抽提物(plant protein extracts)
2 试剂
•棕色离心管: Axygen (Cat No.MCT-150-X)
•丙酮(分析纯)
•Antibodies for target proteins (自制特异抗体或商用抗体; 后者需具有携带特异标签(GFP、FLAG或MYC等)的转基因植物)
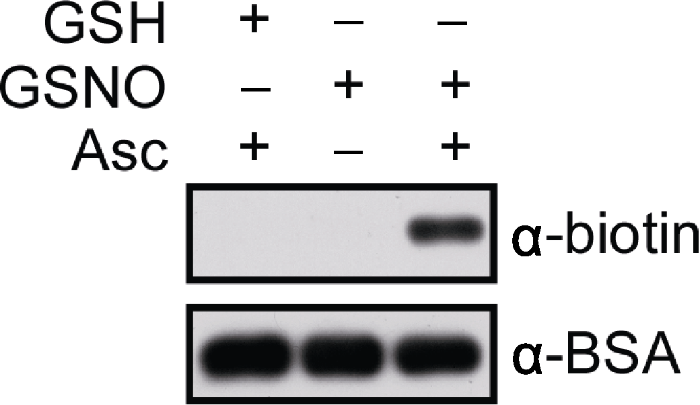
使用biotin-maleimide标记BSA样品, 经SDS-PAGE胶分离后进行免疫印迹分析。分别采用anti-biotin和anti-BSA抗体作为第一抗体(primary antibodies; 1:20 000稀释), anti-mouse IgG为第二抗体(secondary antibodies)。样品信号用SuperSignal Western Femto Maximun Sensitivity Substrate Kit检测。曝光时间分别为20秒(上)和30秒(下)。GSH: 谷胱甘肽; GSNO: S-亚硝基谷胱甘肽; Asc: 抗坏血酸盐 Figure 1In vitro analysis of S-nitrosylated bovine serum albumin (BSA)
BSA is labelled with biotin-maleimide, subjected to SDS- PAGE and western blotting. Anti-biotin and anti-BSA antibodies are used as primary antibodies, respectively (dilution in 1:20 000) and anti-mouse IgG as a secondary antibody. Signals are detected by using the SuperSignal Western Femto Maximun Sensitivity Substrate Kit. The blots are exposed for 20 sec (top) and 30 sec (bottom), respectively. GSH: Glutathione; GSNO: S-nitrosoglutathione; Asc: Sodium ascorbate
ChenR, SunS, WangC, LiY, LiangY, AnF, LiC, DongH, YangX, ZhangJ, ZuoJ ( 2009). The Arabidopsis PARAQUAT RESISTANT2 gene encodes an S-nitrosoglutathione reductase that is a key regulator of cell death Cell Res 19, 1377. [本文引用: 1]
CuiB, PanQ, ClarkeD, VillarrealMO, UmbreenS, YuanB, ShanW, JiangJ, LoakeGJ ( 2018). S-nitrosylation of the zinc finger protein SRG1 regulates plant immunity Nat Commun 9, 4226. [本文引用: 1]
LytvynDI, RaynaudC, YemetsAI, BergouniouxC, BlumeYB ( 2016). Involvement of inositol biosynthesis and nitric oxide in the mediation of UV-B induced oxidative stress Front Plant Sci 7, 430. [本文引用: 1]
PanQN, GengCC, LiDD, XuSW, MaoDD, UmbreenS, LoakeGJ, CuiBM ( 2019). Nitrate reductase-mediated nitric oxide regulates the leaf shape in Arabidopsis by mediating the homeostasis of reactive oxygen species. Int J Mol Sci 20, 2235. [本文引用: 1]
ParísR, VazquezMM, GrazianoM, TerrileMC, MillerND, SpaldingEP, OteguiMS, CasalonguéCA ( 2018). Distribution of endogenous NO regulates early gravitropic response and PIN2 localization in Arabidopsis roots Front Plant Sci 9, 495. [本文引用: 1]
SethD, HessDT, HausladenA, WangL, WangYJ, StamlerJS ( 2018). A multiplex enzymatic machinery for cellular protein S-nitrosylation Mol Cell 69, 451-464. [本文引用: 1]
SethP, HsiehPN, JamalS, WangL, GygiSP, JainMK, CollerJ, StamlerJS ( 2019). Regulation of microRNA machinery and development by interspecies S-nitrosylation Cell 176, 1014-1025. [本文引用: 1]
ShekariesfahlanA, LindermayrC ( 2018). Identification of NO-sensitive cysteine residues using cysteine mutants of recombinant proteins. In: Mengel A, Lindermayr C, eds. Nitric Oxide: Methods and Protocols New York: Springer New York. pp. 183-203. [本文引用: 1]
ShiH, LiuW, WeiY, YeT ( 2017). Integration of auxin/ indole-3-acetic acid 17 and RGA-LIKE3 confers salt stress resistance through stabilization by nitric oxide in Arabidopsis J Exp Bot 68, 1239-1249. [本文引用: 1]
TadaY, SpoelSH, Pajerowska-MukhtarK, MouZ, SongJ, WangC, ZuoJ, DongX ( 2008). Plant immunity requires conformational charges of NPR1 via S-nitrosylation and thioredoxins Science 321, 952-956. [本文引用: 1]
WangP, ZhuJK, LangZ ( 2015). Nitric oxide suppresses the inhibitory effect of abscisic acid on seed germination by S-nitrosylation of SnRK2 proteins Plant Signal Behav 10, e1031939. [本文引用: 1]
WillemsP, HorneA, Van ParysT, GoormachtigS, De SmetI, BotzkiA, Van BreusegemF, GevaertK ( 2019). The Plant PTM Viewer, a central resource for exploring plant protein modifications Plant J 99, 752-762. [本文引用: 1]
XueY, LiuZ, GaoX, JinC, WenL, YaoX, RenJ ( 2010). GPS-SNO: computational prediction of protein S-nitrosylation sites with a modified GPS algorithm PLoS One 5, e11290. [本文引用: 2]
S-nitrosylation triggers ABI5 degradation to promote seed germination and seedling growth 1 2015
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
The Arabidopsis PARAQUAT RESISTANT2 gene encodes an S-nitrosoglutathione reductase that is a key regulator of cell death 1 2009
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
S-nitrosylation of the zinc finger protein SRG1 regulates plant immunity 1 2018
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
Protein S-nitrosylation in plants: current progresses and challenges 4 2019
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
... ; Feng et al., 2019). ...
... NO发挥其生理效应的主要方式之一是对靶蛋白的特异半胱氨酸位点进行翻译后修饰.NO通过可逆地共价结合到蛋白质的半胱氨酸巯基(Cys-SH)上形成亚硝基硫醇(S-nitrosothiols, SNO), 这一过程被称为S-亚硝基化修饰(Hess et al., 2005).S-亚硝基化修饰通常被认为是一种非酶促反应.然而, 近期的研究表明, 在特定条件下, S-亚硝基化修饰是一种由众多相关蛋白质参与的酶促反应过程(Seth et al., 2018).与其它类型的蛋白质翻译后修饰(如泛素化、磷酸化和乙酰化)类似, S-亚硝基化修饰可以在多个层面改变蛋白质的功能(Hess and Stamler, 2012; Feng et al., 2019), 如蛋白质的亚细胞定位、蛋白质与蛋白质之间的互作、蛋白质的生物学活性以及蛋白质的稳定性.现有的研究表明, S-亚硝基化修饰通过改变蛋白质的功能而调控有机体的生长发育过程以及对胁迫的响应(Tada et al., 2008; Feng et al., 2013; Hu et al., 2017; Ling et al., 2017; Iglesias et al., 2018; Zhan et al., 2018; Feng et al., 2019; Seth et al., 2019).因此, 鉴定S-亚硝基化修饰的靶蛋白以及修饰发生的位点对于研究NO调控某一特定生理过程的机制具有重要意义.在解析修饰位点的基础上, 人们可以通过比较模拟持续S-亚硝基化修饰以及不能发生修饰的突变对蛋白质功能的影响, 进而揭示靶蛋白特异位点的修饰对于蛋白质及其参与的生物学过程的影响(Shekariesfahlan and Lindermayr, 2018). ...
... ; Feng et al., 2019; Seth et al., 2019).因此, 鉴定S-亚硝基化修饰的靶蛋白以及修饰发生的位点对于研究NO调控某一特定生理过程的机制具有重要意义.在解析修饰位点的基础上, 人们可以通过比较模拟持续S-亚硝基化修饰以及不能发生修饰的突变对蛋白质功能的影响, 进而揭示靶蛋白特异位点的修饰对于蛋白质及其参与的生物学过程的影响(Shekariesfahlan and Lindermayr, 2018). ...
S-nitrosylation of phosphotransfer proteins represses cytokinin signaling 2 2013
... NO发挥其生理效应的主要方式之一是对靶蛋白的特异半胱氨酸位点进行翻译后修饰.NO通过可逆地共价结合到蛋白质的半胱氨酸巯基(Cys-SH)上形成亚硝基硫醇(S-nitrosothiols, SNO), 这一过程被称为S-亚硝基化修饰(Hess et al., 2005).S-亚硝基化修饰通常被认为是一种非酶促反应.然而, 近期的研究表明, 在特定条件下, S-亚硝基化修饰是一种由众多相关蛋白质参与的酶促反应过程(Seth et al., 2018).与其它类型的蛋白质翻译后修饰(如泛素化、磷酸化和乙酰化)类似, S-亚硝基化修饰可以在多个层面改变蛋白质的功能(Hess and Stamler, 2012; Feng et al., 2019), 如蛋白质的亚细胞定位、蛋白质与蛋白质之间的互作、蛋白质的生物学活性以及蛋白质的稳定性.现有的研究表明, S-亚硝基化修饰通过改变蛋白质的功能而调控有机体的生长发育过程以及对胁迫的响应(Tada et al., 2008; Feng et al., 2013; Hu et al., 2017; Ling et al., 2017; Iglesias et al., 2018; Zhan et al., 2018; Feng et al., 2019; Seth et al., 2019).因此, 鉴定S-亚硝基化修饰的靶蛋白以及修饰发生的位点对于研究NO调控某一特定生理过程的机制具有重要意义.在解析修饰位点的基础上, 人们可以通过比较模拟持续S-亚硝基化修饰以及不能发生修饰的突变对蛋白质功能的影响, 进而揭示靶蛋白特异位点的修饰对于蛋白质及其参与的生物学过程的影响(Shekariesfahlan and Lindermayr, 2018). ...
Nitric oxide represses the Arabidopsis floral transition 1 2004
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
Protein S-nitrosylation: purview and parameters 2 2005
... NO发挥其生理效应的主要方式之一是对靶蛋白的特异半胱氨酸位点进行翻译后修饰.NO通过可逆地共价结合到蛋白质的半胱氨酸巯基(Cys-SH)上形成亚硝基硫醇(S-nitrosothiols, SNO), 这一过程被称为S-亚硝基化修饰(Hess et al., 2005).S-亚硝基化修饰通常被认为是一种非酶促反应.然而, 近期的研究表明, 在特定条件下, S-亚硝基化修饰是一种由众多相关蛋白质参与的酶促反应过程(Seth et al., 2018).与其它类型的蛋白质翻译后修饰(如泛素化、磷酸化和乙酰化)类似, S-亚硝基化修饰可以在多个层面改变蛋白质的功能(Hess and Stamler, 2012; Feng et al., 2019), 如蛋白质的亚细胞定位、蛋白质与蛋白质之间的互作、蛋白质的生物学活性以及蛋白质的稳定性.现有的研究表明, S-亚硝基化修饰通过改变蛋白质的功能而调控有机体的生长发育过程以及对胁迫的响应(Tada et al., 2008; Feng et al., 2013; Hu et al., 2017; Ling et al., 2017; Iglesias et al., 2018; Zhan et al., 2018; Feng et al., 2019; Seth et al., 2019).因此, 鉴定S-亚硝基化修饰的靶蛋白以及修饰发生的位点对于研究NO调控某一特定生理过程的机制具有重要意义.在解析修饰位点的基础上, 人们可以通过比较模拟持续S-亚硝基化修饰以及不能发生修饰的突变对蛋白质功能的影响, 进而揭示靶蛋白特异位点的修饰对于蛋白质及其参与的生物学过程的影响(Shekariesfahlan and Lindermayr, 2018). ...
... S-亚硝基化修饰发生在蛋白质的半胱氨酸残基上.一般而言, 蛋白质能否发生亚硝基化修饰取决于半胱氨酸残基周围的氨基酸组成及半胱氨酸残基在蛋白质三维结构上的相对空间位置.通过对不同靶蛋白的修饰位点进行系统分析, 人们发现发生S-亚硝基化修饰的半胱氨酸残基通常位于一个酸-碱-疏水结构域中([HKR]-C-[hydrophobic]X[D-E])或偏酸性环境中(Hess et al., 2005; Hu et al., 2015).基于上述发现开发的亚硝基化修饰位点预测软件工具(如GPS-SNO)能够预测蛋白质中亚硝基化修饰的候选位点(Xue et al., 2010).此外, 利用植物PTM Viewer数据库也可以预测植物中S-亚硝基化修饰的潜在位点, 其中包括多个已被实验验证的修饰位点(Willems et al., 2019).与其它蛋白质翻译后修饰类似, 预测的S-亚硝基化修饰位点及其功能需要实验生物学验证. ...
Regulation by S-nitrosylation of protein post-translational modification 1 2012
... NO发挥其生理效应的主要方式之一是对靶蛋白的特异半胱氨酸位点进行翻译后修饰.NO通过可逆地共价结合到蛋白质的半胱氨酸巯基(Cys-SH)上形成亚硝基硫醇(S-nitrosothiols, SNO), 这一过程被称为S-亚硝基化修饰(Hess et al., 2005).S-亚硝基化修饰通常被认为是一种非酶促反应.然而, 近期的研究表明, 在特定条件下, S-亚硝基化修饰是一种由众多相关蛋白质参与的酶促反应过程(Seth et al., 2018).与其它类型的蛋白质翻译后修饰(如泛素化、磷酸化和乙酰化)类似, S-亚硝基化修饰可以在多个层面改变蛋白质的功能(Hess and Stamler, 2012; Feng et al., 2019), 如蛋白质的亚细胞定位、蛋白质与蛋白质之间的互作、蛋白质的生物学活性以及蛋白质的稳定性.现有的研究表明, S-亚硝基化修饰通过改变蛋白质的功能而调控有机体的生长发育过程以及对胁迫的响应(Tada et al., 2008; Feng et al., 2013; Hu et al., 2017; Ling et al., 2017; Iglesias et al., 2018; Zhan et al., 2018; Feng et al., 2019; Seth et al., 2019).因此, 鉴定S-亚硝基化修饰的靶蛋白以及修饰发生的位点对于研究NO调控某一特定生理过程的机制具有重要意义.在解析修饰位点的基础上, 人们可以通过比较模拟持续S-亚硝基化修饰以及不能发生修饰的突变对蛋白质功能的影响, 进而揭示靶蛋白特异位点的修饰对于蛋白质及其参与的生物学过程的影响(Shekariesfahlan and Lindermayr, 2018). ...
Site-specific nitrosoproteomic identification of endogenously S-nitrosylated proteins in Arabidopsis 2 2015
... S-亚硝基化修饰发生在蛋白质的半胱氨酸残基上.一般而言, 蛋白质能否发生亚硝基化修饰取决于半胱氨酸残基周围的氨基酸组成及半胱氨酸残基在蛋白质三维结构上的相对空间位置.通过对不同靶蛋白的修饰位点进行系统分析, 人们发现发生S-亚硝基化修饰的半胱氨酸残基通常位于一个酸-碱-疏水结构域中([HKR]-C-[hydrophobic]X[D-E])或偏酸性环境中(Hess et al., 2005; Hu et al., 2015).基于上述发现开发的亚硝基化修饰位点预测软件工具(如GPS-SNO)能够预测蛋白质中亚硝基化修饰的候选位点(Xue et al., 2010).此外, 利用植物PTM Viewer数据库也可以预测植物中S-亚硝基化修饰的潜在位点, 其中包括多个已被实验验证的修饰位点(Willems et al., 2019).与其它蛋白质翻译后修饰类似, 预测的S-亚硝基化修饰位点及其功能需要实验生物学验证. ...
... 在S-亚硝基化修饰的研究中, 通过生物素转化法(biotin switch assay)鉴定蛋白质中S-亚硝基化的修饰位点是目前最常用的方法(Jaffrey and Snyder, 2001).其原理是将未发生S-亚硝基化修饰的自由巯基用烷化剂封闭, 然后用还原剂(通常用抗坏血酸盐)将被S-亚硝基化的巯基还原为自由巯基, 继而用生物素标记新形成的自由巯基.生物素化的半胱氨酸残基(即被S-亚硝基化修饰的半胱氨酸残基)可以利用生物素抗体进行蛋白质免疫印迹(western blotting)或通过液相色谱-串联质谱(liquid chromatography coupled to tandem mass spectrometry; LC-MS/MS)技术进行检测分析.后者也被广泛用于高通量鉴定S-亚硝基化靶蛋白及修饰靶位点(Hu et al., 2015).本文对重组蛋白质的体外生物素转化以及拟南芥(Arabidopsis thaliana)体内蛋白质的生物素转化实验流程进行详细介绍, 并对实验中的注意事项做出具体说明, 旨在为本领域的研究提供借鉴及帮助. ...
Nitric oxide regulates protein methylation during stress responses in plants 2 2017
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
... NO发挥其生理效应的主要方式之一是对靶蛋白的特异半胱氨酸位点进行翻译后修饰.NO通过可逆地共价结合到蛋白质的半胱氨酸巯基(Cys-SH)上形成亚硝基硫醇(S-nitrosothiols, SNO), 这一过程被称为S-亚硝基化修饰(Hess et al., 2005).S-亚硝基化修饰通常被认为是一种非酶促反应.然而, 近期的研究表明, 在特定条件下, S-亚硝基化修饰是一种由众多相关蛋白质参与的酶促反应过程(Seth et al., 2018).与其它类型的蛋白质翻译后修饰(如泛素化、磷酸化和乙酰化)类似, S-亚硝基化修饰可以在多个层面改变蛋白质的功能(Hess and Stamler, 2012; Feng et al., 2019), 如蛋白质的亚细胞定位、蛋白质与蛋白质之间的互作、蛋白质的生物学活性以及蛋白质的稳定性.现有的研究表明, S-亚硝基化修饰通过改变蛋白质的功能而调控有机体的生长发育过程以及对胁迫的响应(Tada et al., 2008; Feng et al., 2013; Hu et al., 2017; Ling et al., 2017; Iglesias et al., 2018; Zhan et al., 2018; Feng et al., 2019; Seth et al., 2019).因此, 鉴定S-亚硝基化修饰的靶蛋白以及修饰发生的位点对于研究NO调控某一特定生理过程的机制具有重要意义.在解析修饰位点的基础上, 人们可以通过比较模拟持续S-亚硝基化修饰以及不能发生修饰的突变对蛋白质功能的影响, 进而揭示靶蛋白特异位点的修饰对于蛋白质及其参与的生物学过程的影响(Shekariesfahlan and Lindermayr, 2018). ...
Regulation of SCFTIR1/AFBs E3 ligase assembly by S-nitrosylation of Arabidopsis SKP1-like1 impacts on auxin signaling 1 2018
... NO发挥其生理效应的主要方式之一是对靶蛋白的特异半胱氨酸位点进行翻译后修饰.NO通过可逆地共价结合到蛋白质的半胱氨酸巯基(Cys-SH)上形成亚硝基硫醇(S-nitrosothiols, SNO), 这一过程被称为S-亚硝基化修饰(Hess et al., 2005).S-亚硝基化修饰通常被认为是一种非酶促反应.然而, 近期的研究表明, 在特定条件下, S-亚硝基化修饰是一种由众多相关蛋白质参与的酶促反应过程(Seth et al., 2018).与其它类型的蛋白质翻译后修饰(如泛素化、磷酸化和乙酰化)类似, S-亚硝基化修饰可以在多个层面改变蛋白质的功能(Hess and Stamler, 2012; Feng et al., 2019), 如蛋白质的亚细胞定位、蛋白质与蛋白质之间的互作、蛋白质的生物学活性以及蛋白质的稳定性.现有的研究表明, S-亚硝基化修饰通过改变蛋白质的功能而调控有机体的生长发育过程以及对胁迫的响应(Tada et al., 2008; Feng et al., 2013; Hu et al., 2017; Ling et al., 2017; Iglesias et al., 2018; Zhan et al., 2018; Feng et al., 2019; Seth et al., 2019).因此, 鉴定S-亚硝基化修饰的靶蛋白以及修饰发生的位点对于研究NO调控某一特定生理过程的机制具有重要意义.在解析修饰位点的基础上, 人们可以通过比较模拟持续S-亚硝基化修饰以及不能发生修饰的突变对蛋白质功能的影响, 进而揭示靶蛋白特异位点的修饰对于蛋白质及其参与的生物学过程的影响(Shekariesfahlan and Lindermayr, 2018). ...
2001). The biotin switch method for the detection of S-nitrosylated proteins 1 2001
... 在S-亚硝基化修饰的研究中, 通过生物素转化法(biotin switch assay)鉴定蛋白质中S-亚硝基化的修饰位点是目前最常用的方法(Jaffrey and Snyder, 2001).其原理是将未发生S-亚硝基化修饰的自由巯基用烷化剂封闭, 然后用还原剂(通常用抗坏血酸盐)将被S-亚硝基化的巯基还原为自由巯基, 继而用生物素标记新形成的自由巯基.生物素化的半胱氨酸残基(即被S-亚硝基化修饰的半胱氨酸残基)可以利用生物素抗体进行蛋白质免疫印迹(western blotting)或通过液相色谱-串联质谱(liquid chromatography coupled to tandem mass spectrometry; LC-MS/MS)技术进行检测分析.后者也被广泛用于高通量鉴定S-亚硝基化靶蛋白及修饰靶位点(Hu et al., 2015).本文对重组蛋白质的体外生物素转化以及拟南芥(Arabidopsis thaliana)体内蛋白质的生物素转化实验流程进行详细介绍, 并对实验中的注意事项做出具体说明, 旨在为本领域的研究提供借鉴及帮助. ...
Host-mediated S-nitrosylation disarms the bacterial effector HopAI1 to reestablish immunity 1 2017
... NO发挥其生理效应的主要方式之一是对靶蛋白的特异半胱氨酸位点进行翻译后修饰.NO通过可逆地共价结合到蛋白质的半胱氨酸巯基(Cys-SH)上形成亚硝基硫醇(S-nitrosothiols, SNO), 这一过程被称为S-亚硝基化修饰(Hess et al., 2005).S-亚硝基化修饰通常被认为是一种非酶促反应.然而, 近期的研究表明, 在特定条件下, S-亚硝基化修饰是一种由众多相关蛋白质参与的酶促反应过程(Seth et al., 2018).与其它类型的蛋白质翻译后修饰(如泛素化、磷酸化和乙酰化)类似, S-亚硝基化修饰可以在多个层面改变蛋白质的功能(Hess and Stamler, 2012; Feng et al., 2019), 如蛋白质的亚细胞定位、蛋白质与蛋白质之间的互作、蛋白质的生物学活性以及蛋白质的稳定性.现有的研究表明, S-亚硝基化修饰通过改变蛋白质的功能而调控有机体的生长发育过程以及对胁迫的响应(Tada et al., 2008; Feng et al., 2013; Hu et al., 2017; Ling et al., 2017; Iglesias et al., 2018; Zhan et al., 2018; Feng et al., 2019; Seth et al., 2019).因此, 鉴定S-亚硝基化修饰的靶蛋白以及修饰发生的位点对于研究NO调控某一特定生理过程的机制具有重要意义.在解析修饰位点的基础上, 人们可以通过比较模拟持续S-亚硝基化修饰以及不能发生修饰的突变对蛋白质功能的影响, 进而揭示靶蛋白特异位点的修饰对于蛋白质及其参与的生物学过程的影响(Shekariesfahlan and Lindermayr, 2018). ...
Involvement of inositol biosynthesis and nitric oxide in the mediation of UV-B induced oxidative stress 1 2016
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
Nitrate reductase-mediated nitric oxide regulates the leaf shape in 1 2019
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
Distribution of endogenous NO regulates early gravitropic response and PIN2 localization in Arabidopsis roots 1 2018
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
A multiplex enzymatic machinery for cellular protein S-nitrosylation 1 2018
... NO发挥其生理效应的主要方式之一是对靶蛋白的特异半胱氨酸位点进行翻译后修饰.NO通过可逆地共价结合到蛋白质的半胱氨酸巯基(Cys-SH)上形成亚硝基硫醇(S-nitrosothiols, SNO), 这一过程被称为S-亚硝基化修饰(Hess et al., 2005).S-亚硝基化修饰通常被认为是一种非酶促反应.然而, 近期的研究表明, 在特定条件下, S-亚硝基化修饰是一种由众多相关蛋白质参与的酶促反应过程(Seth et al., 2018).与其它类型的蛋白质翻译后修饰(如泛素化、磷酸化和乙酰化)类似, S-亚硝基化修饰可以在多个层面改变蛋白质的功能(Hess and Stamler, 2012; Feng et al., 2019), 如蛋白质的亚细胞定位、蛋白质与蛋白质之间的互作、蛋白质的生物学活性以及蛋白质的稳定性.现有的研究表明, S-亚硝基化修饰通过改变蛋白质的功能而调控有机体的生长发育过程以及对胁迫的响应(Tada et al., 2008; Feng et al., 2013; Hu et al., 2017; Ling et al., 2017; Iglesias et al., 2018; Zhan et al., 2018; Feng et al., 2019; Seth et al., 2019).因此, 鉴定S-亚硝基化修饰的靶蛋白以及修饰发生的位点对于研究NO调控某一特定生理过程的机制具有重要意义.在解析修饰位点的基础上, 人们可以通过比较模拟持续S-亚硝基化修饰以及不能发生修饰的突变对蛋白质功能的影响, 进而揭示靶蛋白特异位点的修饰对于蛋白质及其参与的生物学过程的影响(Shekariesfahlan and Lindermayr, 2018). ...
Regulation of microRNA machinery and development by interspecies S-nitrosylation 1 2019
... NO发挥其生理效应的主要方式之一是对靶蛋白的特异半胱氨酸位点进行翻译后修饰.NO通过可逆地共价结合到蛋白质的半胱氨酸巯基(Cys-SH)上形成亚硝基硫醇(S-nitrosothiols, SNO), 这一过程被称为S-亚硝基化修饰(Hess et al., 2005).S-亚硝基化修饰通常被认为是一种非酶促反应.然而, 近期的研究表明, 在特定条件下, S-亚硝基化修饰是一种由众多相关蛋白质参与的酶促反应过程(Seth et al., 2018).与其它类型的蛋白质翻译后修饰(如泛素化、磷酸化和乙酰化)类似, S-亚硝基化修饰可以在多个层面改变蛋白质的功能(Hess and Stamler, 2012; Feng et al., 2019), 如蛋白质的亚细胞定位、蛋白质与蛋白质之间的互作、蛋白质的生物学活性以及蛋白质的稳定性.现有的研究表明, S-亚硝基化修饰通过改变蛋白质的功能而调控有机体的生长发育过程以及对胁迫的响应(Tada et al., 2008; Feng et al., 2013; Hu et al., 2017; Ling et al., 2017; Iglesias et al., 2018; Zhan et al., 2018; Feng et al., 2019; Seth et al., 2019).因此, 鉴定S-亚硝基化修饰的靶蛋白以及修饰发生的位点对于研究NO调控某一特定生理过程的机制具有重要意义.在解析修饰位点的基础上, 人们可以通过比较模拟持续S-亚硝基化修饰以及不能发生修饰的突变对蛋白质功能的影响, 进而揭示靶蛋白特异位点的修饰对于蛋白质及其参与的生物学过程的影响(Shekariesfahlan and Lindermayr, 2018). ...
Identification of NO-sensitive cysteine residues using cysteine mutants of recombinant proteins. In: Mengel A, Lindermayr C, eds. Nitric Oxide: Methods and Protocols 1 2018
... NO发挥其生理效应的主要方式之一是对靶蛋白的特异半胱氨酸位点进行翻译后修饰.NO通过可逆地共价结合到蛋白质的半胱氨酸巯基(Cys-SH)上形成亚硝基硫醇(S-nitrosothiols, SNO), 这一过程被称为S-亚硝基化修饰(Hess et al., 2005).S-亚硝基化修饰通常被认为是一种非酶促反应.然而, 近期的研究表明, 在特定条件下, S-亚硝基化修饰是一种由众多相关蛋白质参与的酶促反应过程(Seth et al., 2018).与其它类型的蛋白质翻译后修饰(如泛素化、磷酸化和乙酰化)类似, S-亚硝基化修饰可以在多个层面改变蛋白质的功能(Hess and Stamler, 2012; Feng et al., 2019), 如蛋白质的亚细胞定位、蛋白质与蛋白质之间的互作、蛋白质的生物学活性以及蛋白质的稳定性.现有的研究表明, S-亚硝基化修饰通过改变蛋白质的功能而调控有机体的生长发育过程以及对胁迫的响应(Tada et al., 2008; Feng et al., 2013; Hu et al., 2017; Ling et al., 2017; Iglesias et al., 2018; Zhan et al., 2018; Feng et al., 2019; Seth et al., 2019).因此, 鉴定S-亚硝基化修饰的靶蛋白以及修饰发生的位点对于研究NO调控某一特定生理过程的机制具有重要意义.在解析修饰位点的基础上, 人们可以通过比较模拟持续S-亚硝基化修饰以及不能发生修饰的突变对蛋白质功能的影响, 进而揭示靶蛋白特异位点的修饰对于蛋白质及其参与的生物学过程的影响(Shekariesfahlan and Lindermayr, 2018). ...
Integration of auxin/ indole-3-acetic acid 17 and RGA-LIKE3 confers salt stress resistance through stabilization by nitric oxide in Arabidopsis 1 2017
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
Plant immunity requires conformational charges of NPR1 via S-nitrosylation and thioredoxins 1 2008
... NO发挥其生理效应的主要方式之一是对靶蛋白的特异半胱氨酸位点进行翻译后修饰.NO通过可逆地共价结合到蛋白质的半胱氨酸巯基(Cys-SH)上形成亚硝基硫醇(S-nitrosothiols, SNO), 这一过程被称为S-亚硝基化修饰(Hess et al., 2005).S-亚硝基化修饰通常被认为是一种非酶促反应.然而, 近期的研究表明, 在特定条件下, S-亚硝基化修饰是一种由众多相关蛋白质参与的酶促反应过程(Seth et al., 2018).与其它类型的蛋白质翻译后修饰(如泛素化、磷酸化和乙酰化)类似, S-亚硝基化修饰可以在多个层面改变蛋白质的功能(Hess and Stamler, 2012; Feng et al., 2019), 如蛋白质的亚细胞定位、蛋白质与蛋白质之间的互作、蛋白质的生物学活性以及蛋白质的稳定性.现有的研究表明, S-亚硝基化修饰通过改变蛋白质的功能而调控有机体的生长发育过程以及对胁迫的响应(Tada et al., 2008; Feng et al., 2013; Hu et al., 2017; Ling et al., 2017; Iglesias et al., 2018; Zhan et al., 2018; Feng et al., 2019; Seth et al., 2019).因此, 鉴定S-亚硝基化修饰的靶蛋白以及修饰发生的位点对于研究NO调控某一特定生理过程的机制具有重要意义.在解析修饰位点的基础上, 人们可以通过比较模拟持续S-亚硝基化修饰以及不能发生修饰的突变对蛋白质功能的影响, 进而揭示靶蛋白特异位点的修饰对于蛋白质及其参与的生物学过程的影响(Shekariesfahlan and Lindermayr, 2018). ...
Nitric oxide suppresses the inhibitory effect of abscisic acid on seed germination by S-nitrosylation of SnRK2 proteins 1 2015
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
The Plant PTM Viewer, a central resource for exploring plant protein modifications 1 2019
... S-亚硝基化修饰发生在蛋白质的半胱氨酸残基上.一般而言, 蛋白质能否发生亚硝基化修饰取决于半胱氨酸残基周围的氨基酸组成及半胱氨酸残基在蛋白质三维结构上的相对空间位置.通过对不同靶蛋白的修饰位点进行系统分析, 人们发现发生S-亚硝基化修饰的半胱氨酸残基通常位于一个酸-碱-疏水结构域中([HKR]-C-[hydrophobic]X[D-E])或偏酸性环境中(Hess et al., 2005; Hu et al., 2015).基于上述发现开发的亚硝基化修饰位点预测软件工具(如GPS-SNO)能够预测蛋白质中亚硝基化修饰的候选位点(Xue et al., 2010).此外, 利用植物PTM Viewer数据库也可以预测植物中S-亚硝基化修饰的潜在位点, 其中包括多个已被实验验证的修饰位点(Willems et al., 2019).与其它蛋白质翻译后修饰类似, 预测的S-亚硝基化修饰位点及其功能需要实验生物学验证. ...
GPS-SNO: computational prediction of protein S-nitrosylation sites with a modified GPS algorithm 2 2010
... S-亚硝基化修饰发生在蛋白质的半胱氨酸残基上.一般而言, 蛋白质能否发生亚硝基化修饰取决于半胱氨酸残基周围的氨基酸组成及半胱氨酸残基在蛋白质三维结构上的相对空间位置.通过对不同靶蛋白的修饰位点进行系统分析, 人们发现发生S-亚硝基化修饰的半胱氨酸残基通常位于一个酸-碱-疏水结构域中([HKR]-C-[hydrophobic]X[D-E])或偏酸性环境中(Hess et al., 2005; Hu et al., 2015).基于上述发现开发的亚硝基化修饰位点预测软件工具(如GPS-SNO)能够预测蛋白质中亚硝基化修饰的候选位点(Xue et al., 2010).此外, 利用植物PTM Viewer数据库也可以预测植物中S-亚硝基化修饰的潜在位点, 其中包括多个已被实验验证的修饰位点(Willems et al., 2019).与其它蛋白质翻译后修饰类似, 预测的S-亚硝基化修饰位点及其功能需要实验生物学验证. ...
... 实验设备包括低温离心机、恒温混匀仪、涡旋仪、蛋白质电泳仪、转膜仪及暗室; 软件用GPS-SNO在线分析软件(Xue et al., 2010) (http://sno.biocuckoo. org/). ...
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
S-nitrosylation targets GSNO reductase for selective autophagy during hypoxia responses in plants 2 2018
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
... NO发挥其生理效应的主要方式之一是对靶蛋白的特异半胱氨酸位点进行翻译后修饰.NO通过可逆地共价结合到蛋白质的半胱氨酸巯基(Cys-SH)上形成亚硝基硫醇(S-nitrosothiols, SNO), 这一过程被称为S-亚硝基化修饰(Hess et al., 2005).S-亚硝基化修饰通常被认为是一种非酶促反应.然而, 近期的研究表明, 在特定条件下, S-亚硝基化修饰是一种由众多相关蛋白质参与的酶促反应过程(Seth et al., 2018).与其它类型的蛋白质翻译后修饰(如泛素化、磷酸化和乙酰化)类似, S-亚硝基化修饰可以在多个层面改变蛋白质的功能(Hess and Stamler, 2012; Feng et al., 2019), 如蛋白质的亚细胞定位、蛋白质与蛋白质之间的互作、蛋白质的生物学活性以及蛋白质的稳定性.现有的研究表明, S-亚硝基化修饰通过改变蛋白质的功能而调控有机体的生长发育过程以及对胁迫的响应(Tada et al., 2008; Feng et al., 2013; Hu et al., 2017; Ling et al., 2017; Iglesias et al., 2018; Zhan et al., 2018; Feng et al., 2019; Seth et al., 2019).因此, 鉴定S-亚硝基化修饰的靶蛋白以及修饰发生的位点对于研究NO调控某一特定生理过程的机制具有重要意义.在解析修饰位点的基础上, 人们可以通过比较模拟持续S-亚硝基化修饰以及不能发生修饰的突变对蛋白质功能的影响, 进而揭示靶蛋白特异位点的修饰对于蛋白质及其参与的生物学过程的影响(Shekariesfahlan and Lindermayr, 2018). ...
CLE9 peptide-induced stomatal closure is mediated by abscisic acid, hydrogen peroxide, and nitric oxide in Arabidopsis thaliana 1 2019
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
Nitrogen and nitric oxide regulate Arabidopsis flowering differently 1 2019
... 一氧化氮(nitric oxide)是一种氮氧化合物, 其化学式为NO, 相对分子量为30.01, 是一种结构简单的气体小分子.NO带有自由基, 其化学性质非常活泼.在常温下, NO为气体, 微溶于水并具有脂溶性, 可以快速透过生物膜扩散.因其上述性质, NO可以作为一种气体信号分子, 将细胞内产生的信息传递至周围的细胞质中, 并参与电子传递反应以及生物体的氧化还原反应.NO在有机体的诸多生物学过程中都发挥重要的调控作用.在高等植物中, NO参与调控众多生长发育过程, 包括开花、种子萌发、叶片形态、气孔开闭及重力响应等(He et al., 2004; Albertos et al., 2015; Wang et al., 2015; París et al., 2018; Feng et al., 2019; Pan et al., 2019; Zhang et al., 2019a, 2019b).此外, NO也参与调控植物对生物与非生物胁迫的响应(Chen et al., 2009; Yang et al., 2015; Lytvyn et al., 2016; Hu et al., 2017; Shi et al., 2017; Cui et al., 2018; Zhan et al., 2018; Feng et al., 2019). ...
 ,*
,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}