Research Progress in Protein Phosphorylation in Plant-pathogen Interactions
Yaqiong Liu, Suiwen Hou,*Ministry of Education Key Laboratory of Cell Activities and Stress Adaptations, School of Life Sciences, Lanzhou University, Lanzhou 730000, China
Abstract Reversible protein phosphorylation is a common mechanism regulating plant signaling pathways. Phosphorylation of key components in plant-pathogen interactions affects the activation of defense signaling. Many pathogens attack the plant immune system and enhance pathogenic toxicity by disturbing the phosphorylation status of defense regulators. In this review, we summarize the phosphorylation of regulators in plant defense responses and its regulating effect in plant immunity. Understanding the phosphorylation of key regulators in the plant-pathogen interaction may help to explore new mechanism of plant immune regulation. This review may provide support and a basis for studying new approaches of broad-spectrum disease resistance. Keywords:protein phosphorylation;protein kinase;protein phosphatase;plant-pathogen interaction
PDF (4098KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 刘雅琼, 侯岁稳. 蛋白磷酸化修饰在植物-病原微生物互作中的作用研究进展. 植物学报, 2019, 54(2): 168-184 doi:10.11983/CBB18219 Liu Yaqiong, Hou Suiwen. Research Progress in Protein Phosphorylation in Plant-pathogen Interactions. Chinese Bulletin of Botany, 2019, 54(2): 168-184 doi:10.11983/CBB18219
植物在漫长的进化过程中形成至少2种免疫途径来抵抗自然界中各种病原微生物的入侵。一种是由定位于细胞质膜的受体识别病原菌关联的分子模式(pathogen-associated molecular patterns, PAMPs)激活的防御反应, 称为PAMP触发的免疫(PAMP- triggered immunity, PTI)。病原微生物分泌效应蛋白进入植物细胞, 干扰宿主免疫激活并增强致病性。另一种免疫途径是通过植物抗病蛋白直接或间接识别效应蛋白后激活, 即效应子触发的免疫(effector- triggered immunity, ETI) (Dangl and Jones, 2001; Jones and Dangl, 2006; Dodds and Rathjen, 2010)。蛋白磷酸化修饰作用是调控细胞信号转导的主要机制, 这一过程依赖于蛋白激酶和蛋白磷酸酶。在植物-病原微生物互作的复杂过程中, 蛋白激酶和蛋白磷酸酶通过调节多种蛋白的磷酸化状态调控植物免疫途径。
1 蛋白激酶和蛋白磷酸酶
模式植物拟南芥(Arabidopsis thaliana)的基因组编码大约1 100种蛋白激酶以及约150种蛋白磷酸酶(Schweighofer and Meskiene, 2015)。根据磷酸化修饰的氨基酸残基不同, 蛋白激酶主要分为两大类, 即丝氨酸/苏氨酸蛋白激酶和酪氨酸蛋白激酶。此外, 有些激酶同时具有丝/苏氨酸蛋白激酶活性和酪氨酸蛋白激酶活性。目前, 在植物免疫途径中研究相对较多的主要有4种激酶, 即跨膜的受体激酶(receptor like kinase, RLK)、胞质内受体激酶(receptor-like cytoplasmic kinase, RLCK)、丝裂原活化激酶(mitogen- activated protein kinase, MAPK)以及钙依赖蛋白激酶(calcium-dependent protein kinase, CPK)。根据结构、催化方式以及底物特异性的不同蛋白磷酸酶分为3种类型, 即蛋白丝氨酸/苏氨酸磷酸酶、蛋白酪氨酸磷酸酶以及双特异性磷酸酶(Luan, 2003)。在拟南芥中, 约96%的蛋白磷酸化作用发生在丝氨酸/苏氨酸残基上。催化丝氨酸/苏氨酸残基去磷酸化的磷酸酶包括2个家族, 即丝氨酸/苏氨酸蛋白磷酸酶家族(pho- sphoprotein phosphatase, PPP)和金属离子依赖的蛋白磷酸酶(metal-dependent protein phosphatases, PPM)。PPP类蛋白磷酸酶的催化亚基结合不同的调节亚基组成的全酶具有不同的底物特异性。PPM家族蛋白磷酸酶主要由PP2C组成, 其本身同时具有催化活性和调节功能, 不需要单独的调节亚基进行调控(Schweighofer and Meskiene, 2015)。
(1) 拟南芥中, LRR类受体激酶和受体蛋白识别病原微生物蛋白或多肽序列; (2) 拟南芥EGF类受体激酶WAK1识别寡聚半乳糖醛酸; (3) Lectin类受体激酶参与免疫信号识别; (4) 拟南芥和水稻LysM类受体激酶或受体蛋白识别几丁质和肽聚糖。 Figure 1Recognition of the immune signals by receptor kinases and receptor proteins
(1) In Arabidopsis, LRR-type receptor kinase and receptor protein recognize pathogenic protein or polypeptide; (2) EGF-type receptor WAK1 recognize oligogalacturonides (OGs); (3) Lectin-type receptor kinase participate in immune signaling recognition; (4) LysM-type receptor kinase or receptor protein recognize chitin and peptidoglycan (PGN) signal in Arabidopsis and rice.
(1) In Arabidopsis, PP2A negatively regulates the activation of BAK1, meanwhile PP2C38 and CPK28 negatively regulate the phosphorylation of BIK1 without pathogen infection. (2), (3) LRR-type PRRs combine with BAK1 and LysM-type RLK CERK1 combine with LYKs to activate immune signaling after perception of pathogen attack. (4) Activated PRRs phosphorylate BIK1 or PBL27. (5) BIK1 and CPKs phosphorylate RBOHD to promote ROS generation. (6), (7) MAPK cascades are activated and then phosphorylate different substrates to regulate different defense responses. (8) MKP1, MKP2, PTP1 and AP2C1 negatively regulate the activity of MAPKs. (9) Bacterial pathogens inject effector proteins HopF2 and HopAI1 to inhibit the activation of MAPK cascades. (10) Resistance protein SUMM2 activate ETI pathway in time by monitoring the phosphorylation statue of MEKK2, CRCK3 and PAT1, the substrates of MPK4.
2.2 LysM类和EGF类受体激酶对免疫信号的识别和激活
LysM类和EGF类PRRs主要识别包含碳水化合物的分子, 如真菌几丁质(chitin)、细菌肽聚糖(peptidog- lycan, PGN)和丛枝菌根菌分泌的脂质几丁寡糖(lipo- chitooligosaccharides, LCOs)或者植物细胞壁衍生的寡聚半乳糖醛酸(oligogalacturonides, OGs)等(图1)。拟南芥中的3个LysM类受体激酶CERK1 (CHITIN ELICITOR RECEPTOR KINASE 1)、LYK4 (LysM- CONTAINING RECEPTOR-LIKE KINASE 4)和LYK5共同调控几丁质的识别及免疫信号的激活(Miya et al., 2007; Wan et al., 2012; Cao et al., 2014); LYK1和LYK3参与对不同结构的PGN信号的识别, 在AtCERK1存在的条件下激活PGN诱导的防御信号(Willmann et al., 2011)。水稻LysM类RLK OsCERK1胞外的LysM结构域不能直接结合几丁质, LysM类受体蛋白OsCEBiP (CHITIN OLIFOSACCHARIDE ELICITOR BINDING PROTEIN)作为受体结合几丁质, 随后结合OsCERK1激活几丁质诱导的防御反应(Kaku et al., 2006; Shinya et al., 2015)。拟南芥EGF类受体激酶WAK1 (WALL-ASSOCIATED KINASE 1)接受OGs信号并激活防御反应(Brutus et al., 2010)。水稻免疫信号激活过程中, OsWAK1发生自磷酸化并磷酸化下游转录因子OsRFP1。此外, 水稻稻瘟菌侵染、机械损伤及水杨酸和茉莉酸甲酯处理均可以激活OsWAK1转录水平的表达(Li et al., 2009)。
2.3 Lectin类受体激酶参与植物防御反应
根据胞外凝集素结构域的不同, 凝集素样受体激酶(lectin receptor kinase, LecRK)又分为3种类型: G (GNA-related or S-locus)-、C (calcium-dependent)-以及L (legume)-类(Bouwmeester and Govers, 2009)。目前已经鉴定到的L-类LecRK有45个, 它们的功能大部分尚属未知。越来越多的研究表明, L-类LecRK广泛参与植物免疫调控。病原菌或PAMPs诱导LecRK-I.9、LecRK-V.5、LecRK-VI.2以及LecRK- IX.2激活表达; 这些LecRKs功能缺失表现出对不同病原菌的敏感性, 过表达则对病原菌抗性增强(Bou- wmeester et al., 2011; Singh et al., 2012; Wang et al., 2014b; Luo et al., 2017)。LecRK-V.5抑制病原菌侵染后的气孔关闭过程; lecrk-V.5缺失突变体对疫霉菌属的抗病性减弱, 对丁香假单胞菌(Pseudomonas syringae pv. tomato (Pst) DC3000)的抗病性增强(Desclos-Theveniau et al., 2012; Wang et al., 2014)。L-类LecRK-V.2和LecRK-VII.1结合FLS2, 正调控气孔免疫途径(Yekondi et al., 2018)。LecRK- VI.2结合FLS2, 正调控PTI的响应, 参与对丁香假单胞菌和果胶杆菌(Pectobacterium carotovorum)的抗性(Singh et al., 2012; Huang et al., 2014)。LecRK- IX.2招募CPKs并磷酸化修饰RBOHD (respiratory burst oxidase homolog protein D), 正调控依赖于RBOHD的水杨酸(salicylic acid, SA)积累以及PTI信号途径(Luo et al., 2017)。此外, 有研究表明, 拟南芥Lectin类受体激酶LORE识别细菌脂多糖(lipopoly- saccharides, LPSs)并激活下游免疫信号(Ranf et al., 2015)。
2.4 细胞内免疫信号的传递依赖于蛋白磷酸化 修饰
2.4.1 RLCK参与免疫信号的传递
胞质内的受体激酶结合RLK/RLP, 共同调控植物的生长发育、激素信号以及生物和非生物胁迫等过程。研究表明, 拟南芥RLCK-VII家族的PBS1 (AvrPphB SUSCEPTIBLE 1)、BIK1 (BOTRYTIS-INDUCED KINASE 1)、PBL1 (PBS1- LIKE 1)、PBL2、PBL27、PCRKs (PTI COMPROMISED RECEPTOR-LIKE CYTOPLASMIC KINASE)以及RLCK-XII家族成员BSK1 (BRASSINOSTEROID-SIGNALING KINASE 1)参与植物免疫信号传递(图1)。例如, FLS2、EFR、PEPRs以及CERK1等多种PRR被激活后磷酸化修饰BIK1, 导致BIK1与PRRs复合体分离并进入细胞质激活下游免疫信号(Lu et al., 2010; Yang et al., 2010; Liu et al., 2013)。PCRK1和PCRK2结合FLS2并在PAMPs诱导后被磷酸化激活, 冗余地诱导转录因子SARD1 (SYSTEMIC ACQUIRED RESISTANCE DEFICIENT 1)和CBP60g (CALMODULIN-BINFING PROTEIN 60g)的表达, 促进SA的合成(Kong et al., 2016)。水稻RLCK-VII家族的OsRLCK185和OsRL- CK176结合CERK1, 传递PGN和几丁质激活的免疫信号(Yamaguchi et al., 2013; Ao et al., 2014)。Os- RLCK185在拟南芥中的同源蛋白PBL27被CERK1磷酸化激活后, 正调控几丁质诱导的防御反应(Shinya et al., 2014)。
2.4.2 RLCK对底物的磷酸化修饰
RLCK经PRRs激活后, 一方面招募钙离子依赖的蛋白激酶(CPKs)促进产生活性氧(ROS); 另一方面激活下游MAPK信号级联反应(图2)。植物中ROS主要由NADPH氧化酶RBOHD催化生成。目前已报道多种RLCKs调控RBOHD的活性。例如, 被flg22诱导激活的BIK1一方面通过磷酸化修饰RBOHD的多个氨基酸位点激活RBOHD的活性; 另一方面通过磷酸化植物Gα蛋白XLG2 (EXTRA LARGE G-PROTEIN 2)促进ROS的产生(Kadota et al., 2014; Li et al., 2014b; Liang et al., 2016)。GTP酶促进蛋白(RGS1)维持FLS2结合的G蛋白不活跃状态。被?g22诱导激活的BIK1/PBL1磷酸化RGS1的S428和S431位点, 促进RGS1与FLS2-G蛋白复合体分离, 从而正调控免疫信号的激活(Liang et al., 2018)。BSK1结合FLS2调控ROS的生成。bsk1缺失突变体中, flg22诱导的ROS爆发减弱, 病原菌诱导的SA积累受到阻碍并且对白粉菌、丁香假单胞菌和卵菌的敏感性增强(Shi et al., 2013)。在无病原菌侵染时, PBL13通过结合RBOHD负调控植物对致病菌的先天免疫途径(Lin et al., 2015)。
MAPKs磷酸化修饰某些WRKY类转录因子, 调控转录重编程。烟草SIPK和WIPK识别并直接磷酸化I类WRKY转录因子WRKY8, 增强WRKY8与W-box的结合和反式激活活性, 进而激活HMGR2 (3-HYD- ROXY-3-METHYLGLUTARYL CoA REDUCTASE 2)的表达, 产生类异戊二烯植物抗毒素, 促进植物的免疫反应(Ishihama et al., 2011)。拟南芥中, 与烟草WRKY8同源的转录因子WRKY33与MPK4以及MKS1在细胞核内形成复合体; 受到flg22或者病原菌刺激时, MPK4磷酸化修饰MKS1并促使WRKY33- MKS1与MPK4分离, 磷酸化的WRKY33随后结合植物抗毒素(camalexin)合成相关基因PAD3启动子区的W-box, 激活PAD3的表达并促进camalexin的合成(Andreasson et al., 2005; Qiu et al., 2008; Len- zoni et al., 2018)。WRKY33还参与MPK3/6调控的camalexin的合成过程。灰霉菌诱导时, MPK3/6磷酸化修饰WRKY33, 促进相关基因转录重编程(Mao et al., 2011; Li et al., 2012)。MPK6结合并磷酸化ERF104调控防御相关基因的表达(Bethke et al., 2009)。拟南芥中ASR3作为转录抑制因子负调控PAMP诱导的免疫响应。PAMP处理后MPK4磷酸化ASR3第189位苏氨酸, 磷酸化的ASR3与DNA结合能力增强, 从而抑制flg22诱导的防御基因的表达(Li et al., 2015)。
MAPKs对底物蛋白的磷酸化修饰影响底物蛋白的稳定性。拟南芥中, MPK3/MPK6通过磷酸化修饰ACS2/ACS6调控乙烯产生。泛素蛋白酶降解复合体结合未被磷酸化修饰的ACS6的C末端非催化区域, 导致ACS6经泛素化途径降解。MPK3/MPK6对ACS6的磷酸化修饰在其C末端引入了负电荷, 抑制ACS6的泛素化降解, 增强其蛋白的稳定性, 进而促进乙烯生成(Joo et al., 2008; Han et al., 2010)。MPK3/ MPK6磷酸化修饰ERF家族成员ERF6, 增强其蛋白稳定性, 激活PDF1.1 (plant defensin 1.1)、PDF- 1.2a、PDF1.2b和ChiB等多种防御相关基因的表达(Meng et al., 2013)。受到PAMP诱导后, MPK3磷酸化修饰WRKY46的S168和S250位点, 促进WRKY46的降解(Sheikh et al., 2016)。LIP5参与病原菌诱导的内吞作用和囊泡运输, MPK3和MPK6磷酸化修饰LIP5, 增强LIP5的蛋白稳定性, 从而正调控多泡体(multivesicular bodies, MVB)通路, 影响免疫应答过程中相关因子的重定位(Wang et al., 2014a)。E3泛素连接酶PUB22是拟南芥中PTI响应的抑制因子。MPK3对PUB22的磷酸化修饰抑制其自泛素化, 从而导致PUB22积累, 抑制免疫信号激活(Furlan et al., 2017)。
MAPKs通过磷酸化修饰底物蛋白改变底物蛋白的亚细胞定位。激活的MPK3对bZIP类转录因子VIP1的磷酸化修饰促进其从细胞质进入细胞核, 激活病程相关基因(pathogenesis-related gene, PR gene)的表达(Djamei et al., 2007)。
R蛋白监控MAPKs对底物蛋白的磷酸化修饰(图2)。多种病原菌分泌效应蛋白攻击宿主细胞内MAPKs信号级联组成元件, 抑制MAPKs调控的防御反应(Zhang et al., 2007, 2012; Wang et al., 2010)。拟南芥抗病蛋白SUMM2 (SUPPRESSOR OF MKK1 MKK2 2)在MAPKs信号的下游, 监控MPK4底物PAT1 (PROTEIN ASSOCIATED WITH TOPOISOMERASE II)、CRCK3 (CALMODULIN-BINDGIN RECEPTOR LIKE CYTOPLASMIC KINASE 3)以及MEKK2的磷酸化状态, 从而调控免疫应答。拟南芥PAT1编码mRNA脱帽装置的组成元件, 与MPK4结合形成复合体。PAMP诱导后, PAT1被MPK4磷酸化并在细胞溶质中聚集成离散的点状物。pat1缺失突变体表现出依赖于SUMM2的免疫自激活(Roux et al., 2015)。CRCK3编码钙离子结合的胞质内受体激酶,结合MPK4并被MPK4磷酸化修饰。CRCK3缺失可恢复mekk1、mkk1/mkk2双突变体以及mpk4中SUMM2介导的免疫自激活。此外, CRCK3和PAT1均可以与SUMM2相互作用, 表明SUMM2可能通过监控CR- CK3和PAT1的磷酸化状态, 从而确定MEKK1- MKK1/MKK2-MPK4信号通路的完整性, 并适时启动免疫应答(Bi and Zhou, 2017; Zhang et al., 2017)。
3.3 磷酸酶对MAPK信号级联的调控
MAPK的持续磷酸化会导致免疫反应被持续激活, 因此MAPK的活性需要受到严格控制。目前已报道多种磷酸酶在植物免疫过程中负调控MAPK的激活。这些磷酸酶主要包括蛋白酪氨酸磷酸酶(PTPs)、双特异性磷酸酶(DSPs)以及PP2C类蛋白磷酸酶。不同的磷酸酶对MAPK的去磷酸化修饰导致不同的下游信号事件。拟南芥蛋白酪氨酸磷酸酶PTP1 (PROTEIN TYROSINE PHOSPHATASE 1)以及双特异性磷酸酶MKP1 (MAP KINASE PHOSPHATASE 1)和MKP2参与对MPK3/MPK6活性的调控。MKP1通过负调控MPK6途径抑制PAMP激活的免疫应答(Anderson et al., 2011); PTP1结合MPK6与MKP1, 协同抑制MPK6调控的防御反应激活。MKP2与MPK3/MPK6结合并且可在体外去磷酸化修饰MAK3和MPK6 (Lee and Ellis, 2007)。真菌激发子诱导的过程中, MKP2抑制MPK6介导的超敏反应(hypersen- sitive response, HR), 从而调控氧化胁迫以及对青枯病病原菌(Ralstonia solanacearum)的抗性(Lumb- reras et al., 2010)。此外, MKP1的活性也受到MPK6的调控。PAMP诱导后MKP1蛋白迅速积累, 而这一过程依赖于MPK6对MKP1的磷酸化修饰(Jiang et al., 2017)。拟南芥PP2C类蛋白磷酸酶(AP2Cs蛋白磷酸酶)通过激酶结合基序(kinase interaction motif, KIM)与MAPK相互作用(Fuchs et al., 2013)。AP2C1负调控植物免疫应答过程中MAPK信号的激活, 过表达AP2C1抑制损伤诱导的乙烯产生以及对死体营养型真菌灰霉菌的抗性。AP2C1功能缺失增强PAMP诱导的MAPK激活、胼胝质积累以及对细菌的抗病性(Schweighofer et al., 2007; Galletti et al., 2011; Sidonskaya et al., 2016; Shubchynskyy et al., 2017)。
AndersonJC, BartelsS, González BesteiroMA, Sha- hollariB, UlmR, PeckSC ( 2011). Arabidopsis MAP Kinase Phosphatase 1 (AtMKP1) negatively regulates MPK6-mediated PAMP responses and resistance against bacteria Plant J 67, 258-268. DOI:10.1111/j.1365-313X.2011.04588.xURLPMID:21447069 [本文引用: 1] A primary component of plant defense is the detection of pathogen-associated molecular patterns (PAMPs) by plasma membrane-localized pathogen recognition receptors. PAMP perception results in rapid and transient activation of phosphorylation-dependent signaling pathways that lead to a wide array of defense-related responses, including extensive changes in gene expression. In Arabidopsis, several kinases, including the mitogen-activated protein kinases (MAPKs) MPK6 and MPK3, are rapidly activated after PAMP treatment, and are thought to positively regulate a wide array of defense-related responses. In contrast, negative regulation of PAMP responses by downstream phosphatases remains poorly understood. Here we report the identification of Arabidopsis MAP Kinase Phosphatase 1 (MKP1) as a negative regulator of diverse PAMP responses, including activation of MPK6 and MPK3, transient production of extracellular reactive oxygen species, accumulation of a subset of PAMP-regulated transcripts, and inhibition of seedling growth. In agreement with the enhanced PAMP response phenotypes observed in the mkp1 mutant, we found that mkp1 seedlings and adult plants are more resistant to the virulent bacterial pathogen Pseudomonas syringae pv. tomato (Pto) DC3000. Further genetic analysis revealed that MPK6, but not MPK3, is required for the mkp1-dependent increase in resistance to Pto and enhanced PAMP-induced growth inhibition observed in mkp1 seedlings. Together, our data support a role for MKP1 as a negative regulator of MPK6-mediated PAMP responses.
AndreassonE, JenkinsT, BrodersenP, ThorgrimsenS, PetersenNHT, ZhuSJ, QiuJL, MicheelsenP, RocherA, PetersenM, NewmanMA, Bjørn NielsenH, HirtH, SomssichI, MattssonO, MundyJ ( 2005). The MAP kinase substrate MKS1 is a regulator of plant defense responses EMBO J 24, 2579-2589. DOI:10.1038/sj.emboj.7600737URLPMID:15990873 [本文引用: 1] Arabidopsis MAP kinase 4 (MPK4) functions as a regulator of pathogen defense responses, because it is required for both repression of salicylic acid (SA)-dependent resistance and for activation of jasmonate (JA)-dependent defense gene expression. To understand MPK4 signaling mechanisms, we used yeast two-hybrid screening to identify the MPK4 substrate MKS1. Analyses of transgenic plants and genome-wide transcript profiling indicated that MKS1 is required for full SA-dependent resistance in mpk4 mutants, and that overexpression of MKS1 in wild-type plants is sufficient to activate SA-dependent resistance, but does not interfere with induction of a defense gene by JA. Further yeast two-hybrid screening revealed that MKS1 interacts with the WRKY transcription factors WRKY25 and WRKY33. WRKY25 and WRKY33 were shown to be in vitro substrates of MPK4, and a wrky33 knockout mutant was found to exhibit increased expression of the SA-related defense gene PR1. MKS1 may therefore contribute to MPK4-regulated defense activation by coupling the kinase to specific WRKY transcription factors.
AoY, LiZQ, FengDR, XiongF, LiuJ, LiJF, WangML, WangJF, LiuB, WangHB ( 2014). OsCERK1 and OsRLCK176 play important roles in peptidoglycan and chitin signaling in rice innate immunity Plant J 80, 1072-1084. DOI:10.1111/tpj.12710URLPMID:25335639 [本文引用: 1] Summary Microbe-associated molecular pattern (MAMP)-triggered immunity plays critical roles in the basal resistance defense response in plants. Chitin and peptidoglycan (PGN) are major molecular patterns for fungi and bacteria, respectively. Two rice ( Oryza sativa ) lysin motif-containing proteins, OsLYP4 and OsLYP6, function as receptors that sense bacterial PGN and fungal chitin. These membrane receptors, which lack intracellular kinase domains, likely contain another component for transmembrane immune signal transduction. Here, we demonstrate that the rice LysM receptor-like kinase OsCERK1, a key component of the chitin elicitor signaling pathway, also plays an important role in PGN-triggered immunity in rice. Silencing of OsCERK1 suppressed PGN-induced (and chitin-induced) immunity responses, including reactive oxygen species generation, defense gene expression, and callose deposition, indicating that OsCERK1 is essential for both PGN and chitin signaling initiated by OsLYP4 and OsLYP6. OsLYP4 associated with OsLYP6 and the rice chitin receptor chitin oligosaccharide elicitor-binding protein (CEBiP) in the absence of PGN or chitin, and treatment with PGN or chitin led to their disassociation in vivo . OsCERK1 associated with OsLYP4 or OsLYP6 when induced by PGN but it associated with OsLYP4, OsLYP6, or CEBiP under chitin treatment, suggesting the presence of different patterns of ligand-induced heterooligomeric receptor complexes. Furthermore, the receptor-like cytoplasmic kinase OsRLCK176 functions downstream of OsCERK1 in the PGN and chitin signaling pathways, suggesting that these MAMPs share overlapping intracellular signaling components. Therefore, OsCERK1 plays dual roles in PGN and chitin signaling in rice innate immunity and as an adaptor involved in signal transduction at the plasma membrane in conjunction with OsLYP4 and OsLYP6.
AxtellMJ, StaskawiczBJ ( 2003). Initiation of RPS2- specified disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4 Cell 112, 369-377. DOI:10.1016/S0092-8674(03)00036-9URLPMID:12581526 [本文引用: 1] Plants have evolved a sophisticated innate immune system to recognize invading pathogens and to induce a set of host defense mechanisms resulting in disease resistance. Pathogen recognition is often mediated by plant disease resistance (R) proteins that respond specifically to one or a few pathogen-derived molecules. This specificity has led to suggestions of a receptor-ligand mode of R protein function. Delivery of the bacterial effector protein AvrRpt2 by Pseudomonas syringae specifically induces disease resistance in Arabidopsis plants expressing the RPS2 R protein. We demonstrate that RPS2 physically interacts with Arabidopsis RIN4 and that AvrRpt2 causes the elimination of RIN4 during activation of the RPS2 pathway. AvrRpt2-mediated RIN4 elimination also occurs in the rps2, ndr1, and Atrar1 mutant backgrounds, demonstrating that this activity can be achieved independent of an RPS2-mediated signaling pathway. Therefore, we suggest that RPS2 initiates signaling based upon perception of RIN4 disappearance rather than direct recognition of AvrRpt2.
BethkeG, UnthanT, UhrigJF, PoschlY, GustAA, ScheelD, LeeJ ( 2009). Flg22 regulates the release of an ethy- lene response factor substrate from MAP kinase 6 in Arabidopsis thaliana via ethylene signaling Proc Natl Acad Sci USA 106, 8067-8072. [本文引用: 1]
BiGZ, ZhouJM ( 2017). MAP kinase signaling pathways: a hub of plant-microbe interactions Cell Host Microbe 21, 270-273. DOI:10.1016/j.chom.2017.02.004URLPMID:28279328 [本文引用: 1] In 2007, we reported that a phytopathogen effector directly inhibits a MAP kinase cascade. In the decade since, many more effectors have been found to inhibit MAP kinase cascades, providing not only a mechanistic understanding of pathogenesis and immunity in plants, but also the identification of previously unknown enzymes.
BiGZ, ZhouZY, WangWB, LiL, RaoSF, WuY, ZhangXJ, MenkeFLH, ChenS, ZhouJM ( 2018). Receptor-like cytoplasmic kinases directly link diverse pattern recog- nition receptors to the activation of mitogen-activated protein kinase cascades in Arabidopsis Plant Cell 30, 1543-1561. [本文引用: 2]
BigeardJ, ColcombetJ, HirtH ( 2015). Signaling mech- anisms in pattern-triggered immunity (PTI) Mol Plant 8, 521-539. DOI:10.1016/j.molp.2014.12.022URLPMID:25744358 [本文引用: 1] The pathogens that can go through plant preformed defenses are perceived by two different recognition systems that initiate the pattern-triggered immunity (PTI) or effector-triggered immunity (ETI), both of which are accompanied by a set of induced defenses that usually repel pathogen attacks. We discuss the complex network of signaling pathways occurring during PTI, focusing on the involvement of mitogen-activated protein kinases.
BigeardJ, RayapuramN, PfliegerD, HirtH ( 2014). Phosphorylation-dependent regulation of plant chromatin and chromatin-associated proteins Proteomics 14, 2127-2140. DOI:10.1002/pmic.201400073URLPMID:24889195 [本文引用: 1] In eukaryotes, most of the DNA is located in the nucleus where it is organized with histone proteins in a higher order structure as chromatin. Chromatin and chromatin-associated proteins contribute to DNA-related processes such as replication and transcription as well as epigenetic regulation. Protein functions are often regulated by PTMs among which phosphorylation is one of the most abundant PTM. Phosphorylation of proteins affects important properties, such as enzyme activity, protein stability, or subcellular localization. We here describe the main specificities of protein phosphorylation in plants and review the current knowledge on phosphorylation-dependent regulation of plant chromatin and chromatin-associated proteins. We also outline some future challenges to further elucidate protein phosphorylation and chromatin regulation.
BouwmeesterK, de SainM, WeideR, GougetA, KlamerS, CanutH, GoversF ( 2011). The lectin receptor kinase LecRK-I.9 is a novel Phytophthora resistance component and a potential host target for a RXLR effector PLoS Pathog 7, e1001327. DOI:10.1371/journal.ppat.1001327URLPMID:3068997 [本文引用: 1] In plants, an active defense against biotrophic pathogens is dependent on a functional continuum between the cell wall (CW) and the plasma membrane (PM). It is thus anticipated that proteins maintaining this continuum also function in defense. The legume-like lectin receptor kinase LecRK-I.9 is a putative mediator of CW-PM adhesions in Arabidopsis and is known to bind in vitro to the Phytophthora infestans RXLR-dEER effector IPI-O via a RGD cell attachment motif present in IPI-O. Here we show that LecRK-I.9 is associated with the plasma membrane, and that two T-DNA insertions lines deficient in LecRK-I.9 (lecrk-I.9) have a ‘gain-of-susceptibility’ phenotype specifically towards the oomycete Phytophthora brassicae. Accordingly, overexpression of LecRK-I.9 leads to enhanced resistance to P. brassicae. A similar ‘gain-of-susceptibility’ phenotype was observed in transgenic Arabidopsis lines expressing ipiO (35S-ipiO1). This phenocopy behavior was also observed with respect to other defense-related functions; lecrk-I.9 and 35S-ipiO1 were both disturbed in pathogen- and MAMP-triggered callose deposition. By site-directed mutagenesis, we demonstrated that the RGD cell attachment motif in IPI-O is not only essential for disrupting the CW-PM adhesions, but also for disease suppression. These results suggest that destabilizing the CW-PM continuum is one of the tactics used by Phytophthora to promote infection. As countermeasure the host may want to strengthen CW-PM adhesions and the novel Phytophthora resistance component LecRK-I.9 seems to function in this process.
BouwmeesterK, GoversF ( 2009). Arabidopsis L-type lectin receptor kinases: phylogeny, classification, and expression profiles J Exp Bot 60, 4383-4396. DOI:10.1093/jxb/erp277URLPMID:19773388 [本文引用: 1] Abstract In plants, lectin receptor kinases are considered to play crucial roles during development and in the adaptive response to various stimuli. Arabidopsis lectin receptor kinases can be divided into three type-classes based on sequence similarity of their extracellular lectin motifs. The current study focuses on the legume-like lectin receptor kinases (LecRKs), which are regarded as ideal candidates for monitoring cell wall integrity and are possibly functional in adaptive responses. An inventory of the Arabidopsis LecRK gene family is presented here. It consists of 45 members including three that were recently identified; two encode N-terminal truncated variants one of which has two in tandem kinase domains. Phylogenetic trees derived from full-length amino acid sequence alignments were highly concordant to phylograms that were purely based on lectin motifs or kinase domains. The phylograms allowed reclassification of the LecRK genes and hence a new proposal for gene nomenclature was suggested. In addition, a comprehensive expression analysis was executed by exploring public repositories. This revealed that several LecRK genes are differentially expressed during plant growth and development. Moreover, multiple LecRKs appear to be induced upon treatment with elicitors and pathogen infection. Variation in gene expression was also analysed in seedlings of diverse Arabidopsis accessions. Taken together, this study provides a genome-wide overview of the LecRK gene family and an up-to-date classification using a novel and systematic gene nomenclature.
BrutusA, SiciliaF, MaconeA, CervoneF, De LorenzoG ( 2010). A domain swap approach reveals a role of the plant wall-associated kinase 1 (WAK1) as a receptor of oligogalacturonides Proc Natl Acad Sci USA 107, 9452-9457. DOI:10.1073/pnas.1000675107URLPMID:20439716 [本文引用: 1] Oligogalacturonides (OGs) released from the plant cell wall are active both as damage-associated molecular patterns (DAMPs) for the activation of the plant immune response and regulators of plant growth and development. Members of the Wall-Associated Kinase (WAK) family are candidate receptors of OGs, due to their ability to bind in vitro these oligosaccharides. Because lethality and redundancy have hampered the study of WAKs by reverse genetics, we have adopted a chimeric receptor approach to elucidate the role of Arabidopsis WAK1. In a test-of-concept study, we first defined the appropriate chimera design and demonstrated that the Arabidopsis pattern recognition receptor (PRR) EFR is amenable to the construction of functional and resistance-conferring chimeric receptors carrying the ectodomain of another Arabidopsis PRR, FLS2. After, we analyzed chimeras derived from EFR and WAK1. Our results show that, upon stimulation with OGs, the WAK1 ectodomain is capable of activating the EFR kinase domain. On the other hand, upon stimulation with the cognate ligand elf18, the EFR ectodomain activates the WAK1 kinase, triggering defense responses that mirror those normally activated by OGs and are effective against fungal and bacterial pathogens. Finally, we show that transgenic plants overexpressing WAK1 are more resistant to Botrytis cinerea.
CaoYR, LiangY, TanakaK, NguyenCT, JedrzejczakRP, JoachimiakA, StaceyG ( 2014). The kinase LYK5 is a major chitin receptor in Arabidopsis and forms a chitin- induced complex with related kinase CERK1 eLife 3, e03766. DOI:10.7554/eLife.03766URLPMID:25340959 [本文引用: 1] 10.7554/eLife.03766.001Chitin is a fungal microbe-associated molecular pattern recognized in Arabidopsis by a lysin motif receptor kinase (LYK), AtCERK1. Previous research suggested that AtCERK1 is the major chitin receptor and mediates chitin-induced signaling through homodimerization and phosphorylation. However, the reported chitin binding affinity of AtCERK1 is quite low, suggesting another receptor with high chitin binding affinity might be present. Here, we propose that AtLYK5 is the primary chitin receptor in Arabidopsis. Mutations in AtLYK5 resulted in a significant reduction in chitin response. However, AtLYK5 shares overlapping function with AtLYK4 and, therefore, Atlyk4/Atlyk5-2 double mutants show a complete loss of chitin response. AtLYK5 interacts with AtCERK1 in a chitin-dependent manner. Chitin binding to AtLYK5 is indispensable for chitin-induced AtCERK1 phosphorylation. AtLYK5 binds chitin at a much higher affinity than AtCERK1. The data suggest that AtLYK5 is the primary receptor for chitin, forming a chitin inducible complex with AtCERK1 to induce plant immunity.DOI: http://dx.doi.org/10.7554/eLife.03766.001
ChenXW, ZuoSM, SchwessingerB, ChernM, CanlasPE, RuanDL, ZhouXG, WangJ, DaudiA, PetzoldCJ, HeazlewoodJL, RonaldPC ( 2014). An XA21-associated kinase (OsSERK2) regulates immunity mediated by the XA21 and XA3 immune receptors Mol Plant 7, 874-892. DOI:10.1093/mp/ssu003URLPMID:4064043 [本文引用: 1]
ChengSH, WillmannMR, ChenHC, SheenJ ( 2002). Calcium signaling through protein kinases. The Arabi- dopsis calcium-dependent protein kinase gene family Plant Physiol 129, 469-485. DOI:10.1104/pp.005645URLPMID:12068094 [本文引用: 1] In plants, numerous Ca2+-stimulated protein kinase activities occur through calcium-dependent protein kinases (CDPKs). These novel calcium sensors are likely to be crucial mediators of responses to diverse endogenous and environmental cues. However, the precise biological function(s) of most CDPKs remains elusive. The Arabidopsis genome is predicted to encode 34 different CDPKs. In this Update, we analyze the Arabidopsis CDPK gene family and review the expression, regulation, and possible functions of plant CDPKs. By combining emerging cellular and genomic technologies with genetic and biochemical approaches, the characterization of Arabidopsis CDPKs provides a valuable opportunity to understand the plant calcium-signaling network.
ChinchillaD, BauerZ, RegenassM, BollerT, FelixG ( 2006). The Arabidopsis receptor kinase FLS2 binds flg22 and determines the specificity of flagellin perception Plant Cell 18, 465-476. DOI:10.1105/tpc.105.036574URL [本文引用: 1] Flagellin, the main building block of the bacterial flagellum, acts as a pathogen-associated molecular pattern triggering the innate immune response in animals and plants. In Arabidopsis thaliana, the Leu-rich repeat transmembrane receptor kinase FLAGELLIN SENSITIVE2 (FLS2) is essential for flagellin perception. Here, we demonstrate the specific interaction of the elicitor-active epitope flg22 with the FLS2 protein by chemical cross-linking and immunoprecipitation. The functionality of this receptor was further tested by heterologous expression of the Arabidopsis FLS2 gene in tomato (Lycopersicon esculentum) cells. The perception of flg22 in tomato differs characteristically from that in Arabidopsis. Expression of Arabidopsis FLS2 conferred an additional flg22-perception system on the cells of tomato, which showed all of the properties characteristic of the perception of this elicitor in Arabidopsis. In summary, these results show that FLS2 constitutes the pattern-recognition receptor that determines the specificity of flagellin perception.
ChungEH, El-KasmiF, HeYJ, LoehrA, DanglJL ( 2014). A plant phosphoswitch platform repeatedly targeted by type III effector proteins regulates the output of both tiers of plant immune receptors Cell Host Microbe 16, 484-494. DOI:10.1016/j.chom.2014.09.004URLPMID:25299334 [本文引用: 2] Plant immunity is repressed, in part, by RIN4. Chung et al. show that upon pathogen perception, RIN4 is phosphorylated at Ser141, causing derepression of immune responses. The bacterial type III effector AvrB acts with a host kinase to enhance RIN4 phosphorylation at another site to rerepress immune response and promote virulence.
CoutoD, NiebergallR, LiangXX, BucherlCA, SklenarJ, MachoAP, NtoukakisV, DerbyshireP, AltenbachD, MacleanD, RobatzekS, UhrigJ, MenkeF, ZhouJM, ZipfelC ( 2016). The Arabidopsis protein phosphatase PP2C38 negatively regulates the central immune kinase BIK1 PLoS Pathog 12, e1005811. DOI:10.1371/journal.ppat.1005811URLPMID:4975489 [本文引用: 1] Plants recognize pathogen-associated molecular patterns (PAMPs) via cell surface-localized pattern recognition receptors (PRRs), leading to PRR-triggered immunity (PTI). TheArabidopsiscytoplasmic kinase BIK1 is a downstream substrate of several PRR complexes. How plant PTI is negatively regulated is not fully understood. Here, we identify the protein phosphatase PP2C38 as a negative regulator of BIK1 activity and BIK1-mediated immunity. PP2C38 dynamically associates with BIK1, as well as with the PRRs FLS2 and EFR, but not with the co-receptor BAK1. PP2C38 regulates PAMP-induced BIK1 phosphorylation and impairs the phosphorylation of the NADPH oxidase RBOHD by BIK1, leading to reduced oxidative burst and stomatal immunity. Upon PAMP perception, PP2C38 is phosphorylated on serine 77 and dissociates from the FLS2/EFR-BIK1 complexes, enabling full BIK1 activation. Together with our recent work on the control of BIK1 turnover, this study reveals another important regulatory mechanism of this central immune component. Plants use immune receptors at the cell surface to perceive microbial molecules and initiate a broad-spectrum defence response against pathogens. However, the induction and amplitude of immune signalling must be tightly regulated. Immune responses are triggered by ligand binding to a cognate receptor, which is present in dynamic kinase complexes that heavily rely on trans-phosphorylation to initiate signalling. The cytoplasmic kinase BIK1 associates with different immune receptors and plays a central role in the activation of downstream immune signalling. We show here that theArabidopsis thalianaprotein phosphatase PP2C38 negatively regulates immune responses by controlling the phosphorylation and activation status of BIK1. Furthermore, we propose a mechanism that relieves this negative regulation involving PP2C38 phosphorylation and dissociation from BIK1. These findings extend our knowledge on how plant immunity is appropriately regulated.
CoutoD, ZipfelC ( 2016). Regulation of pattern recognition receptor signaling in plants Nat Rev Immunol 16, 537-552. DOI:10.1038/nri.2016.77 [本文引用: 1]
DanglJL, JonesJDG ( 2001). Plant pathogens and integ- rated defence responses to infection Nature 411, 826-833. DOI:10.1038/35081161 [本文引用: 1]
del PozoO, PedleyKF, MartinGB ( 2004). MAPKKKα is a positive regulator of cell death associated with both plant immunity and disease EMBO J 23, 3072-3082. DOI:10.1038/sj.emboj.7600283URL [本文引用: 1]
FurlanG, NakagamiH, Eschen-LippoldL, JiangXY, MajovskyP, KowarschikK, HoehenwarterW, LeeJ, TrujilloM ( 2017). Changes in PUB22 ubiquitination modes triggered by MITOGEN-ACTIVATED PROTEIN KINASE3 dampen the immune response Plant Cell 29, 726-745. DOI:10.1105/tpc.16.00654URL [本文引用: 1]
GallettiR, FerrariS, De LorenzoG ( 2011). Arabidopsis MPK3 and MPK6 play different roles in basal and oligo- galacturonide- or flagellin-induced resistance against Bot- rytis cinerea Plant Physiol 157, 804-814. [本文引用: 1]
GaoXG, ChenX, LinWW, ChenSX, LuDP, NiuYJ, LiL, ChengC, McCormackM, SheenJ, ShanLB, HeP ( 2013). Bifurcation of Arabidopsis NLR immune signaling via Ca 2+-dependent protein kinases PLoS Pathog 9, e1003127. DOI:10.1371/journal.ppat.1003127URL [本文引用: 2]
Gómez-GómezL, BollerT ( 2000). FLS2: an LRR receptor- like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis Mol Cell 5, 1003-1011. DOI:10.1016/S1097-2765(00)80265-8URL [本文引用: 1]
GravinoM, SavatinDV, MaconeA, De LorenzoG ( 2015). Ethylene production in Botrytis cinerea - and oligogala- cturonide-induced immunity requires calcium-dependent protein kinases Plant J 84, 1073-1086. [本文引用: 1]
GustAA, FelixG ( 2014). Receptor like proteins associate with SOBIR1-type of adaptors to form bimolecular receptor kinases Curr Opin Plant Biol 21, 104-111. DOI:10.1016/j.pbi.2014.07.007URL [本文引用: 1]
HalterT, ImkampeJ, MazzottaS, WierzbaM, PostelS, BücherlC, KieferC, StahlM, ChinchillaD, WangXF, NürnbergerT, ZipfelC, ClouseS, BorstJW, BoerenS, de VriesSC, TaxF, KemmerlingB ( 2014). The leucine- rich repeat receptor kinase BIR2 is a negative regulator of BAK1 in plant immunity Curr Biol 24, 134-143. DOI:10.1016/j.cub.2013.11.047URL [本文引用: 1]
HanL, LiGJ, YangKY, MaoGH, WangRG, LiuYD, ZhangSQ ( 2010). Mitogen-activated protein kinase 3 and 6 regulate Botrytis cinerea -induced ethylene production in Arabidopsis Plant J 64, 114-127. [本文引用: 1]
JooS, LiuYD, LuethA, ZhangSQ ( 2008). MAPK phos- phorylation-induced stabilization of ACS6 protein is me- diated by the non-catalytic C-terminal domain, which also contains the cis -determinant for rapid degradation by the 26S proteasome pathway Plant J 54, 129-140. [本文引用: 1]
KadotaY, SklenarJ, DerbyshireP, StransfeldL, AsaiS, NtoukakisV, JonesJD, ShirasuK, MenkeF, JonesA, ZipfelC ( 2014). Direct regulation of the NADPH oxidase RBOHD by the PRR-associated kinase BIK1 during plant immunity Mol Cell 54, 43-55. DOI:10.1016/j.molcel.2014.02.021URL [本文引用: 1]
KakuH, NishizawaY, Ishii-MinamiN, Akimoto-TomiyamaC, DohmaeN, TakioK, MinamiE, ShibuyaN ( 2006). Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor Proc Natl Acad Sci USA 103, 11086-11091. DOI:10.1073/pnas.0508882103URL [本文引用: 1]
KongQ, SunTJ, QuN, MaJL, LiM, ChengYT, ZhangQ, WuD, ZhangZB, ZhangYL ( 2016). Two redundant receptor-like cytoplasmic kinases function downstream of pattern recognition receptors to regulate activation of SA biosynthesis Plant Physiol 171, 1344-1354. [本文引用: 1]
LalNK, NagalakshmiU, HurlburtNK, FloresR, BakA, SoneP, MaXY, SongGY, WalleyJ, ShanLB, HeP, CasteelC, FisherAJ, Dinesh-KumarSP ( 2018). The receptor-like cytoplasmic kinase BIK1 localizes to the nucleus and regulates defense hormone expression during plant innate immunity Cell Host Microbe 23, 485-497. DOI:10.1016/j.chom.2018.03.010URL [本文引用: 1]
LeeD, BourdaisG, YuG, RobatzekS, CoakerG ( 2015). Phosphorylation of the plant immune regulator RPM1- INTERACTING PROTEIN4 enhances plant plasma mem- brane H +-ATPase activity and inhibits flagellin-triggered immune responses in Arabidopsis Plant Cell 27, 2042-2056. DOI:10.1105/tpc.114.132308URL [本文引用: 1]
LeeJS, EllisBE ( 2007). Arabidopsis MAPK phosphatase 2 (MKP2) positively regulates oxidative stress tolerance and inactivates the MPK3 and MPK6 MAPKs J Biol Chem 282, 25020-25029. DOI:10.1074/jbc.M701888200URL [本文引用: 1]
LenzoniG, LiuJL, KnightMR ( 2018). Predicting plant immunity gene expression by identifying the decoding mechanism of calcium signatures New Phytol 217, 1598-1609. DOI:10.1111/nph.14924URL [本文引用: 1]
LuoY, CaldwellKS, WroblewskiT, WrightME, Mich- elmoreRW ( 2009). Proteolysis of a negative regulator of innate immunity is dependent on resistance genes in tomato and Nicotiana benthamiana and induced by mul- tiple bacterial effectors Plant Cell 21, 2458-2472. [本文引用: 1]
MackeyD, BelkhadirY, AlonsoJM, EckerJR, DanglJL ( 2003). Arabidopsis RIN4 is a target of the type III virulence effector AvrRpt2 and modulates RPS2-mediated resistance Cell 112, 379-389. DOI:10.1016/S0092-8674(03)00040-0URL [本文引用: 1]
MackeyD, Holt BF3rd, WiigA, DanglJL ( 2002). RIN4 interacts with Pseudomonas syringae type III effector molecules and is required for RPM1-mediated resistance in Arabidopsis Cell 108, 743-754. [本文引用: 1]
MaoGH, MengXZ, LiuYD, ZhengZY, ChenZX, ZhangSQ ( 2011). Phosphorylation of a WRKY transcription factor by two pathogen-responsive MAPKs drives phytoalexin bio- synthesis in Arabidopsis Plant Cell 23, 1639-1653. DOI:10.1105/tpc.111.084996URL [本文引用: 1]
MengXZ, XuJ, HeYX, YangKY, MordorskiB, LiuYD, ZhangSQ ( 2013). Phosphorylation of an ERF trans- cription factor by Arabidopsis MPK3/MPK6 regulates plant defense gene induction and fungal resistance Plant Cell 25, 1126-1142. DOI:10.1105/tpc.112.109074URL [本文引用: 1]
NühseTS, StensballeA, JensenON, PeckSC ( 2004). Phosphoproteomics of the Arabidopsis plasma membrane and a new phosphorylation site database Plant Cell 16, 2394-2405. DOI:10.1105/tpc.104.023150URL [本文引用: 1]
SidonskayaE, SchweighoferA, ShubchynskyyV, Kam- merhoferN, HofmannJ, WieczorekK, MeskieneI ( 2016). Plant resistance against the parasitic nematode Heterodera schachtii is mediated by MPK3 and MPK6 kinases, which are controlled by the MAPK phosphatase AP2C1 in Arabidopsis J Exp Bot 67, 107-118. [本文引用: 1]
SinghP, KuoYC, MishraS, TsaiCH, ChienCC, ChenCW, Desclos-TheveniauM, ChuPW, SchulzeB, ChinchillaD, BollerT, ZimmerliL ( 2012). The lectin receptor kinase-VI.2 is required for priming and positively regulates Arabidopsis pattern-triggered immunity Plant Cell 24, 1256-1270. DOI:10.1105/tpc.112.095778URL [本文引用: 2]
SpoelSH, MouZL, TadaY, SpiveyNW, GenschikP, DongXN ( 2009). Proteasome-mediated turnover of the trans- cription coactivator NPR1 plays dual roles in regulating plant immunity Cell 137, 860-872. DOI:10.1016/j.cell.2009.03.038URL [本文引用: 1]
TangDZ, WangGX, ZhouJM ( 2017). Receptor kinases in plant-pathogen interactions: more than pattern recognition Plant Cell 29, 618-637. DOI:10.1105/tpc.16.00891URL [本文引用: 1]
WanJR, TanakaK, ZhangXC, SonGH, BrechenmacherL, TranHNN, StaceyG ( 2012). LYK4, a lysin motif receptor-like kinase, is important for chitin signaling and plant innate immunity in Arabidopsis Plant Physiol 160, 396-406. DOI:10.1104/pp.112.201699URL [本文引用: 1]
WangF, ShangYF, FanBF, YuJQ, ChenZX ( 2014a). Arabidopsis LIP5, a positive regulator of multivesicular body biogenesis, is a critical target of pathogen-responsive MAPK cascade in plant basal defense PLoS Pathog 10, e1004243. DOI:10.1371/journal.ppat.1004243URL [本文引用: 3]
WangJ, ZhouL, ShiH, ChernM, YuH, YiH, HeM, YinJJ, ZhuXB, LiY, LiWT, LiuJL, WangJC, ChenXQ, QingH, WangYP, LiuGF, WangWM, LiP, WuXJ, ZhuLH, ZhouJM, RonaldPC, LiSG, LiJY, ChenXW ( 2018a). A single transcription factor promotes both yield and im- munity in rice Science 361, 1026-1028. DOI:10.1126/science.aat7675URL [本文引用: 2]
WidjajaI, LassowskatI, BethkeG, Eschen-LippoldL, LongHH, NaumannK, DanglJL, ScheelD, LeeJ ( 2010). A protein phosphatase 2C, responsive to the bacterial effector AvrRpm1 but not to the AvrB effector, regulates defense responses in Arabidopsis Plant J 61, 249-258. [本文引用: 1]
WiltonM, SubramaniamR, ElmoreJ, FelsensteinerC, CoakerG, DesveauxD ( 2010). The type III effector HopF2 Pto targets Arabidopsis RIN4 protein to promote Pseudomonas syringae virulence Proc Natl Acad Sci USA 107, 2349-2354. [本文引用: 1]
XieCG, ZhouXN, DengXW, GuoY ( 2010). PKS5, a SNF1- related kinase, interacts with and phosphorylates NPR1, and modulates expression of WRKY38 and WRKY62 J Genet Genomics 37, 359-369. [本文引用: 1]
YamadaK, Yamashita-YamadaM, HiraseT, FujiwaraT, TsudaK, HirumaK, SaijoY ( 2016b). Danger peptide receptor signaling in plants ensures basal immunity upon pathogen-induced depletion of BAK1 EMBO J 35, 46-61. DOI:10.15252/embj.201591807URL [本文引用: 1]
YamaguchiK, YamadaK, IshikawaK, YoshimuraS, HayashiN, UchihashiK, IshihamaN, Kishi-KaboshiM, TakahashiA, TsugeS, OchiaiH, TadaY, ShimamotoK, YoshiokaH, KawasakiT ( 2013). A receptor-like cyto- plasmic kinase targeted by a plant pathogen effector is directly phosphorylated by the chitin receptor and medi- ates rice immunity Cell Host Microbe 13, 347-357. DOI:10.1016/j.chom.2013.02.007URL [本文引用: 1]
YamaguchiY, HuffakerA, BryanAC, TaxFE, RyanCA ( 2010). PEPR2 is a second receptor for the Pep1 and Pep2 peptides and contributes to defense responses in Arabidopsis Plant Cell 22, 508-522. DOI:10.1105/tpc.109.068874URL [本文引用: 1]
YamaguchiY, PearceG, RyanCA ( 2006). The cell surface leucine-rich repeat receptor for At Pep1, an endogenous peptide elicitor in Arabidopsis, is functional in transgenic tobacco cells Proc Natl Acad Sci USA 103, 10104-10109. [本文引用: 1]
YanLM, MaYY, LiuD, WeiXC, SunY, ChenXY, ZhaoHD, ZhouJW, WangZY, ShuiWQ, LouZY ( 2012). Structural basis for the impact of phosphorylation on the activation of plant receptor-like kinase BAK1 Cell Res 22, 1304-1308. DOI:10.1038/cr.2012.74
YangHB, ShiYT, LiuJY, GuoL, ZhangXY, YangSH ( 2010). A mutant CHS3 protein with TIR-NB-LRR-LIM domains modulates growth, cell death and freezing tole- rance in a temperature-dependent manner in Arabi- dopsis Plant J 63, 283-296. DOI:10.1111/j.1365-313X.2010.04241.xURL [本文引用: 1]
YekondiS, LiangFC, OkumaE, RadziejwoskiA, MaiHW, SwainS, SinghP, GauthierM, ChienHC, MurataY, ZimmerliL ( 2018). Nonredundant functions of Arabidopsis LecRK-V.2 and LecRK-VII.1 in controlling stomatal imm- unity and jasmonate-mediated stomatal closure New Phytol 218, 253-268. DOI:10.1111/nph.14953URL [本文引用: 1]
ZhangSQ, KlessigDF ( 1998). Resistance gene N-mediated de novo synthesis and activation of a tobacco mitogen- activated protein kinase by tobacco mosaic virus infection Proc Natl Acad Sci USA 95, 7433-7438. [本文引用: 1]
ZhangWG, FraitureM, KolbD, LöffelhardtB, DesakiY, BoutrotFFG, TörM, ZipfelC, GustAA, BrunnerF ( 2013). Arabidopsis receptor-like protein30 and receptor- like kinase suppressor of BIR1-1/EVERSHED mediate innate immunity to necrotrophic fungi Plant Cell 25, 4227-4241. DOI:10.1105/tpc.113.117010URL [本文引用: 1]
ZhangZB, LiuY, HuangH, GaoMH, WuD, KongQ, ZhangYL ( 2017). The NLR protein SUMM2 senses the disruption of an immune signaling MAP kinase cascade via CRCK3 EMBO Rep 18, 292-302. DOI:10.15252/embr.201642704URL [本文引用: 1]
ZhangZB, WuYL, GaoMH, ZhangJ, KongQ, LiuY, BaHP, ZhouJM, ZhangYL ( 2012). Disruption of PAMP- induced MAP kinase cascade by a Pseudomonas syringae effector activates plant immunity mediated by the NB-LRR protein SUMM2 Cell Host Microbe 11, 253-263. [本文引用: 1]
ZipfelC, KunzeG, ChinchillaD, CaniardA, JonesJDG, BollerT, FelixG ( 2006). Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium -medi- ated transformation Cell 125, 749-760. [本文引用: 1]
An RLP23-SOBIR1-BAK1 complex mediates NLP-triggered immunity 1 2015
... 由于缺少胞内激酶结构域, LRR-RLP识别配体后通过结合RLKs激活胞内免疫信号.LRR类受体激酶 SOBIR1 (SUPPRESSOR OF BIR1-1 1)结合多种LRR-RLP调控防御反应的激活(Gust and Felix, 2014).拟南芥RLP23识别细菌、真菌以及卵菌分泌的坏死/乙烯诱导的多肽蛋白nlp20 (NECROSIS AND ETHYLENE-INDUCING PEPTIDE 1-LIKE PROTEINS 20)后结合SOBIR1, 并招募BAK1形成复合体激活免疫信号(Albert et al., 2015).RLP30结合SOBIR1和BAK1, 识别死体营养型真菌蛋白激发子SCFE1 (SCLEROTINIA CULTURE FILTRATE ELICITOR 1)并激活下游免疫反应(Zhang et al., 2013).番茄中, Cf-4、Ve1和EIX1 (ETHYLENE- INDUCING XYLANASE 1)等RLPs结合SOBIR1激活真菌诱导的免疫途径(Liebrand et al., 2013).番茄受体激酶ELR (ELICITIN RESPONSE)结合免疫共受体BAK1/SERK3, 通过识别多种疫霉菌激肽蛋白(elicitin proteins)激活广谱且持久的抗病性(Du et al., 2015). ...
Arabidopsis MAP Kinase Phosphatase 1 (AtMKP1) negatively regulates MPK6-mediated PAMP responses and resistance against bacteria 1 2011
... MAPK的持续磷酸化会导致免疫反应被持续激活, 因此MAPK的活性需要受到严格控制.目前已报道多种磷酸酶在植物免疫过程中负调控MAPK的激活.这些磷酸酶主要包括蛋白酪氨酸磷酸酶(PTPs)、双特异性磷酸酶(DSPs)以及PP2C类蛋白磷酸酶.不同的磷酸酶对MAPK的去磷酸化修饰导致不同的下游信号事件.拟南芥蛋白酪氨酸磷酸酶PTP1 (PROTEIN TYROSINE PHOSPHATASE 1)以及双特异性磷酸酶MKP1 (MAP KINASE PHOSPHATASE 1)和MKP2参与对MPK3/MPK6活性的调控.MKP1通过负调控MPK6途径抑制PAMP激活的免疫应答(Anderson et al., 2011); PTP1结合MPK6与MKP1, 协同抑制MPK6调控的防御反应激活.MKP2与MPK3/MPK6结合并且可在体外去磷酸化修饰MAK3和MPK6 (Lee and Ellis, 2007).真菌激发子诱导的过程中, MKP2抑制MPK6介导的超敏反应(hypersen- sitive response, HR), 从而调控氧化胁迫以及对青枯病病原菌(Ralstonia solanacearum)的抗性(Lumb- reras et al., 2010).此外, MKP1的活性也受到MPK6的调控.PAMP诱导后MKP1蛋白迅速积累, 而这一过程依赖于MPK6对MKP1的磷酸化修饰(Jiang et al., 2017).拟南芥PP2C类蛋白磷酸酶(AP2Cs蛋白磷酸酶)通过激酶结合基序(kinase interaction motif, KIM)与MAPK相互作用(Fuchs et al., 2013).AP2C1负调控植物免疫应答过程中MAPK信号的激活, 过表达AP2C1抑制损伤诱导的乙烯产生以及对死体营养型真菌灰霉菌的抗性.AP2C1功能缺失增强PAMP诱导的MAPK激活、胼胝质积累以及对细菌的抗病性(Schweighofer et al., 2007; Galletti et al., 2011; Sidonskaya et al., 2016; Shubchynskyy et al., 2017). ...
The MAP kinase substrate MKS1 is a regulator of plant defense responses 1 2005
... MAPKs磷酸化修饰某些WRKY类转录因子, 调控转录重编程.烟草SIPK和WIPK识别并直接磷酸化I类WRKY转录因子WRKY8, 增强WRKY8与W-box的结合和反式激活活性, 进而激活HMGR2 (3-HYD- ROXY-3-METHYLGLUTARYL CoA REDUCTASE 2)的表达, 产生类异戊二烯植物抗毒素, 促进植物的免疫反应(Ishihama et al., 2011).拟南芥中, 与烟草WRKY8同源的转录因子WRKY33与MPK4以及MKS1在细胞核内形成复合体; 受到flg22或者病原菌刺激时, MPK4磷酸化修饰MKS1并促使WRKY33- MKS1与MPK4分离, 磷酸化的WRKY33随后结合植物抗毒素(camalexin)合成相关基因PAD3启动子区的W-box, 激活PAD3的表达并促进camalexin的合成(Andreasson et al., 2005; Qiu et al., 2008; Len- zoni et al., 2018).WRKY33还参与MPK3/6调控的camalexin的合成过程.灰霉菌诱导时, MPK3/6磷酸化修饰WRKY33, 促进相关基因转录重编程(Mao et al., 2011; Li et al., 2012).MPK6结合并磷酸化ERF104调控防御相关基因的表达(Bethke et al., 2009).拟南芥中ASR3作为转录抑制因子负调控PAMP诱导的免疫响应.PAMP处理后MPK4磷酸化ASR3第189位苏氨酸, 磷酸化的ASR3与DNA结合能力增强, 从而抑制flg22诱导的防御基因的表达(Li et al., 2015). ...
OsCERK1 and OsRLCK176 play important roles in peptidoglycan and chitin signaling in rice innate immunity 1 2014
... 胞质内的受体激酶结合RLK/RLP, 共同调控植物的生长发育、激素信号以及生物和非生物胁迫等过程.研究表明, 拟南芥RLCK-VII家族的PBS1 (AvrPphB SUSCEPTIBLE 1)、BIK1 (BOTRYTIS-INDUCED KINASE 1)、PBL1 (PBS1- LIKE 1)、PBL2、PBL27、PCRKs (PTI COMPROMISED RECEPTOR-LIKE CYTOPLASMIC KINASE)以及RLCK-XII家族成员BSK1 (BRASSINOSTEROID-SIGNALING KINASE 1)参与植物免疫信号传递(图1).例如, FLS2、EFR、PEPRs以及CERK1等多种PRR被激活后磷酸化修饰BIK1, 导致BIK1与PRRs复合体分离并进入细胞质激活下游免疫信号(Lu et al., 2010; Yang et al., 2010; Liu et al., 2013).PCRK1和PCRK2结合FLS2并在PAMPs诱导后被磷酸化激活, 冗余地诱导转录因子SARD1 (SYSTEMIC ACQUIRED RESISTANCE DEFICIENT 1)和CBP60g (CALMODULIN-BINFING PROTEIN 60g)的表达, 促进SA的合成(Kong et al., 2016).水稻RLCK-VII家族的OsRLCK185和OsRL- CK176结合CERK1, 传递PGN和几丁质激活的免疫信号(Yamaguchi et al., 2013; Ao et al., 2014).Os- RLCK185在拟南芥中的同源蛋白PBL27被CERK1磷酸化激活后, 正调控几丁质诱导的防御反应(Shinya et al., 2014). ...
Initiation of RPS2- specified disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4 1 2003
... 拟南芥包含NOI结构域的蛋白RIN4在陆生植物中非常保守, 其并不具有类似已知的蛋白酶类的特征.二级结构预测发现, RIN4蛋白表现出高度的、固有的无序性, 不能形成二级和三级结构, 是植物中固有的无序蛋白(Lee et al., 2015).无序蛋白在与客户蛋白结合时转变成高度折叠状态, 松散并特异地与多种客户蛋白结合(Dyson and Wright, 2005).研究表明, RIN4是AvrRpt2、AvrRpm1、AvrB、HopF2以及AvrPto等多种病原菌效应子蛋白在宿主内攻击的靶标, 表明RIN4在植物-病原微生物互作中具有重要作用(Mackey et al., 2002, 2003; Axtell and Staskawicz, 2003; Luo et al., 2009; Wilton et al., 2010). ...
Flg22 regulates the release of an ethy- lene response factor substrate from MAP kinase 6 in Arabidopsis thaliana via ethylene signaling 1 2009
... MAPKs磷酸化修饰某些WRKY类转录因子, 调控转录重编程.烟草SIPK和WIPK识别并直接磷酸化I类WRKY转录因子WRKY8, 增强WRKY8与W-box的结合和反式激活活性, 进而激活HMGR2 (3-HYD- ROXY-3-METHYLGLUTARYL CoA REDUCTASE 2)的表达, 产生类异戊二烯植物抗毒素, 促进植物的免疫反应(Ishihama et al., 2011).拟南芥中, 与烟草WRKY8同源的转录因子WRKY33与MPK4以及MKS1在细胞核内形成复合体; 受到flg22或者病原菌刺激时, MPK4磷酸化修饰MKS1并促使WRKY33- MKS1与MPK4分离, 磷酸化的WRKY33随后结合植物抗毒素(camalexin)合成相关基因PAD3启动子区的W-box, 激活PAD3的表达并促进camalexin的合成(Andreasson et al., 2005; Qiu et al., 2008; Len- zoni et al., 2018).WRKY33还参与MPK3/6调控的camalexin的合成过程.灰霉菌诱导时, MPK3/6磷酸化修饰WRKY33, 促进相关基因转录重编程(Mao et al., 2011; Li et al., 2012).MPK6结合并磷酸化ERF104调控防御相关基因的表达(Bethke et al., 2009).拟南芥中ASR3作为转录抑制因子负调控PAMP诱导的免疫响应.PAMP处理后MPK4磷酸化ASR3第189位苏氨酸, 磷酸化的ASR3与DNA结合能力增强, 从而抑制flg22诱导的防御基因的表达(Li et al., 2015). ...
MAP kinase signaling pathways: a hub of plant-microbe interactions 1 2017
... R蛋白监控MAPKs对底物蛋白的磷酸化修饰(图2).多种病原菌分泌效应蛋白攻击宿主细胞内MAPKs信号级联组成元件, 抑制MAPKs调控的防御反应(Zhang et al., 2007, 2012; Wang et al., 2010).拟南芥抗病蛋白SUMM2 (SUPPRESSOR OF MKK1 MKK2 2)在MAPKs信号的下游, 监控MPK4底物PAT1 (PROTEIN ASSOCIATED WITH TOPOISOMERASE II)、CRCK3 (CALMODULIN-BINDGIN RECEPTOR LIKE CYTOPLASMIC KINASE 3)以及MEKK2的磷酸化状态, 从而调控免疫应答.拟南芥PAT1编码mRNA脱帽装置的组成元件, 与MPK4结合形成复合体.PAMP诱导后, PAT1被MPK4磷酸化并在细胞溶质中聚集成离散的点状物.pat1缺失突变体表现出依赖于SUMM2的免疫自激活(Roux et al., 2015).CRCK3编码钙离子结合的胞质内受体激酶,结合MPK4并被MPK4磷酸化修饰.CRCK3缺失可恢复mekk1、mkk1/mkk2双突变体以及mpk4中SUMM2介导的免疫自激活.此外, CRCK3和PAT1均可以与SUMM2相互作用, 表明SUMM2可能通过监控CR- CK3和PAT1的磷酸化状态, 从而确定MEKK1- MKK1/MKK2-MPK4信号通路的完整性, 并适时启动免疫应答(Bi and Zhou, 2017; Zhang et al., 2017). ...
Receptor-like cytoplasmic kinases directly link diverse pattern recog- nition receptors to the activation of mitogen-activated protein kinase cascades in Arabidopsis 2 2018
The lectin receptor kinase LecRK-I.9 is a novel Phytophthora resistance component and a potential host target for a RXLR effector 1 2011
... 根据胞外凝集素结构域的不同, 凝集素样受体激酶(lectin receptor kinase, LecRK)又分为3种类型: G (GNA-related or S-locus)-、C (calcium-dependent)-以及L (legume)-类(Bouwmeester and Govers, 2009).目前已经鉴定到的L-类LecRK有45个, 它们的功能大部分尚属未知.越来越多的研究表明, L-类LecRK广泛参与植物免疫调控.病原菌或PAMPs诱导LecRK-I.9、LecRK-V.5、LecRK-VI.2以及LecRK- IX.2激活表达; 这些LecRKs功能缺失表现出对不同病原菌的敏感性, 过表达则对病原菌抗性增强(Bou- wmeester et al., 2011; Singh et al., 2012; Wang et al., 2014b; Luo et al., 2017).LecRK-V.5抑制病原菌侵染后的气孔关闭过程; lecrk-V.5缺失突变体对疫霉菌属的抗病性减弱, 对丁香假单胞菌(Pseudomonas syringae pv. tomato (Pst) DC3000)的抗病性增强(Desclos-Theveniau et al., 2012; Wang et al., 2014).L-类LecRK-V.2和LecRK-VII.1结合FLS2, 正调控气孔免疫途径(Yekondi et al., 2018).LecRK- VI.2结合FLS2, 正调控PTI的响应, 参与对丁香假单胞菌和果胶杆菌(Pectobacterium carotovorum)的抗性(Singh et al., 2012; Huang et al., 2014).LecRK- IX.2招募CPKs并磷酸化修饰RBOHD (respiratory burst oxidase homolog protein D), 正调控依赖于RBOHD的水杨酸(salicylic acid, SA)积累以及PTI信号途径(Luo et al., 2017).此外, 有研究表明, 拟南芥Lectin类受体激酶LORE识别细菌脂多糖(lipopoly- saccharides, LPSs)并激活下游免疫信号(Ranf et al., 2015). ...
... 根据胞外凝集素结构域的不同, 凝集素样受体激酶(lectin receptor kinase, LecRK)又分为3种类型: G (GNA-related or S-locus)-、C (calcium-dependent)-以及L (legume)-类(Bouwmeester and Govers, 2009).目前已经鉴定到的L-类LecRK有45个, 它们的功能大部分尚属未知.越来越多的研究表明, L-类LecRK广泛参与植物免疫调控.病原菌或PAMPs诱导LecRK-I.9、LecRK-V.5、LecRK-VI.2以及LecRK- IX.2激活表达; 这些LecRKs功能缺失表现出对不同病原菌的敏感性, 过表达则对病原菌抗性增强(Bou- wmeester et al., 2011; Singh et al., 2012; Wang et al., 2014b; Luo et al., 2017).LecRK-V.5抑制病原菌侵染后的气孔关闭过程; lecrk-V.5缺失突变体对疫霉菌属的抗病性减弱, 对丁香假单胞菌(Pseudomonas syringae pv. tomato (Pst) DC3000)的抗病性增强(Desclos-Theveniau et al., 2012; Wang et al., 2014).L-类LecRK-V.2和LecRK-VII.1结合FLS2, 正调控气孔免疫途径(Yekondi et al., 2018).LecRK- VI.2结合FLS2, 正调控PTI的响应, 参与对丁香假单胞菌和果胶杆菌(Pectobacterium carotovorum)的抗性(Singh et al., 2012; Huang et al., 2014).LecRK- IX.2招募CPKs并磷酸化修饰RBOHD (respiratory burst oxidase homolog protein D), 正调控依赖于RBOHD的水杨酸(salicylic acid, SA)积累以及PTI信号途径(Luo et al., 2017).此外, 有研究表明, 拟南芥Lectin类受体激酶LORE识别细菌脂多糖(lipopoly- saccharides, LPSs)并激活下游免疫信号(Ranf et al., 2015). ...
A domain swap approach reveals a role of the plant wall-associated kinase 1 (WAK1) as a receptor of oligogalacturonides 1 2010
... LysM类和EGF类PRRs主要识别包含碳水化合物的分子, 如真菌几丁质(chitin)、细菌肽聚糖(peptidog- lycan, PGN)和丛枝菌根菌分泌的脂质几丁寡糖(lipo- chitooligosaccharides, LCOs)或者植物细胞壁衍生的寡聚半乳糖醛酸(oligogalacturonides, OGs)等(图1).拟南芥中的3个LysM类受体激酶CERK1 (CHITIN ELICITOR RECEPTOR KINASE 1)、LYK4 (LysM- CONTAINING RECEPTOR-LIKE KINASE 4)和LYK5共同调控几丁质的识别及免疫信号的激活(Miya et al., 2007; Wan et al., 2012; Cao et al., 2014); LYK1和LYK3参与对不同结构的PGN信号的识别, 在AtCERK1存在的条件下激活PGN诱导的防御信号(Willmann et al., 2011).水稻LysM类RLK OsCERK1胞外的LysM结构域不能直接结合几丁质, LysM类受体蛋白OsCEBiP (CHITIN OLIFOSACCHARIDE ELICITOR BINDING PROTEIN)作为受体结合几丁质, 随后结合OsCERK1激活几丁质诱导的防御反应(Kaku et al., 2006; Shinya et al., 2015).拟南芥EGF类受体激酶WAK1 (WALL-ASSOCIATED KINASE 1)接受OGs信号并激活防御反应(Brutus et al., 2010).水稻免疫信号激活过程中, OsWAK1发生自磷酸化并磷酸化下游转录因子OsRFP1.此外, 水稻稻瘟菌侵染、机械损伤及水杨酸和茉莉酸甲酯处理均可以激活OsWAK1转录水平的表达(Li et al., 2009). ...
The kinase LYK5 is a major chitin receptor in Arabidopsis and forms a chitin- induced complex with related kinase CERK1 1 2014
... LysM类和EGF类PRRs主要识别包含碳水化合物的分子, 如真菌几丁质(chitin)、细菌肽聚糖(peptidog- lycan, PGN)和丛枝菌根菌分泌的脂质几丁寡糖(lipo- chitooligosaccharides, LCOs)或者植物细胞壁衍生的寡聚半乳糖醛酸(oligogalacturonides, OGs)等(图1).拟南芥中的3个LysM类受体激酶CERK1 (CHITIN ELICITOR RECEPTOR KINASE 1)、LYK4 (LysM- CONTAINING RECEPTOR-LIKE KINASE 4)和LYK5共同调控几丁质的识别及免疫信号的激活(Miya et al., 2007; Wan et al., 2012; Cao et al., 2014); LYK1和LYK3参与对不同结构的PGN信号的识别, 在AtCERK1存在的条件下激活PGN诱导的防御信号(Willmann et al., 2011).水稻LysM类RLK OsCERK1胞外的LysM结构域不能直接结合几丁质, LysM类受体蛋白OsCEBiP (CHITIN OLIFOSACCHARIDE ELICITOR BINDING PROTEIN)作为受体结合几丁质, 随后结合OsCERK1激活几丁质诱导的防御反应(Kaku et al., 2006; Shinya et al., 2015).拟南芥EGF类受体激酶WAK1 (WALL-ASSOCIATED KINASE 1)接受OGs信号并激活防御反应(Brutus et al., 2010).水稻免疫信号激活过程中, OsWAK1发生自磷酸化并磷酸化下游转录因子OsRFP1.此外, 水稻稻瘟菌侵染、机械损伤及水杨酸和茉莉酸甲酯处理均可以激活OsWAK1转录水平的表达(Li et al., 2009). ...
An XA21-associated kinase (OsSERK2) regulates immunity mediated by the XA21 and XA3 immune receptors 1 2014
... 拟南芥LRR类受体激酶SERKs (SOMATIC EMBRYOGENESIS RECEPTOR KINASEs)作为FLS2/EFR/PEPRs等PRRs的共受体参与免疫信号的识别和激活.PRR接受病原信号后与BAK1/SERK3 (BRI1-ASSOCIATED RECEPTOR KINASE 1)结合, 并且胞内激酶结构域发生自磷酸化修饰.与此同时, PRR与BAK1的激酶域相互磷酸化修饰, 迅速激活免疫信号(Gómez-Gómez and Boller, 2000; Yama- guchi et al., 2010; Yamada et al., 2016b).水稻中, OsSERK2正调控XA21、XA3以及OsFLS2介导的免疫响应(Chen et al., 2014). ...
Calcium signaling through protein kinases. The Arabi- dopsis calcium-dependent protein kinase gene family 1 2002
... 钙离子依赖的蛋白激酶(CPK)调控RBOHD及转录因子的激活.拟南芥基因组编码34个CPKs (Cheng et al., 2002; Bigeard et al., 2015), 广泛参与各种胁迫响应的调控.拟南芥CPK5、CPK6和CPK11功能同时缺失可抑制乙烯合成酶ACS2/ACS6的积累及乙烯的合成, 从而影响对死体营养型致病菌灰霉菌的抗病性(Gravino et al., 2015).Gao等(2013)研究表明, 拟南芥抗病蛋白RPS2和RPM1被激活后, CPKs通过磷酸化修饰不同的底物参与调控多种抗病蛋白依赖的免疫激活.CPK1/2/4/11以及CPK5磷酸化质膜定位的NADPH氧化酶促进ROS的生成; 激活的CPK4/ 5/6/11特异地磷酸化修饰WRKY转录因子WRKY8/ 28/48, 协同调控转录重排, 参与抗病蛋白介导的对病原菌的抗性; CPK1/2/4/5则在超敏反应过程中调控程序性细胞死亡的起始(Dubiella et al., 2013; Gao et al., 2013).此外, CPK5参与非典型抗病蛋白TN2激活的抗性.拟南芥胞外复合体亚基EXO70B1缺失突变体中CPK5被过度激活, 参与抗病蛋白TN2介导的免疫自激活过程, 过表达CPK5导致依赖于TN2的免疫自激活以及抗病性增强.TN2结合并且稳定CPK5的活性, 从而调控防御反应(Liu et al., 2017). ...
The Arabidopsis receptor kinase FLS2 binds flg22 and determines the specificity of flagellin perception 1 2006
... LRR类PRRs主要识别蛋白或多肽序列(图1).例如, LRR类受体激酶FLS2 (FLAGELLIN SENSING 2)和EFR (ELONGATION FACTOR-Tu RECEPTOR)分别识别并结合病原菌鞭毛蛋白保守的22个氨基酸残基组成的肽flg22, 以及延伸因子Tu (EF-Tu) N末端的保守氨基酸序列elf18 (Chinchilla et al., 2006; Zipfel et al., 2006); PEPR1/2 (PEP RECEPTOR)识别植物响应病原菌分泌的内源可溶性前体肽pep (pro- peptide) (Yamaguchi et al., 2006); 番茄(Solanum lycopersicum)中FLS3识别细菌flgII-28并增强对细菌的免疫响应(Hind et al., 2016); 水稻(Oryza sativa) LRR-类受体激酶XA21识别水稻白叶枯菌(Xantho- monas oryzae pv. oryzae, Xoo)中保守的蛋白RaxX等(Pruitt et al., 2015). ...
A plant phosphoswitch platform repeatedly targeted by type III effector proteins regulates the output of both tiers of plant immune receptors 2 2014
... RIN4蛋白不同位点的磷酸化修饰具有不同的功能(Nühse et al., 2007; Chung et al., 2014).有研究表明, RIN4第47和141位的丝氨酸是受flg22诱导后被磷酸化修饰的可能位点(Nühse et al., 2004, 2007).Chung等(2014)发现, RIN4第141位的丝氨酸残基的磷酸化修饰影响flg22诱导对病原菌生长的抑制, 导致胼胝质沉积、ROS爆发以及防御基因表达上调.当RIN4第141位丝氨酸突变为不能被磷酸化修饰的丙氨酸时, flg22诱导的PTI响应受到抑制.RIN4蛋白第166位的苏氨酸残基的磷酸化修饰, 发挥与S141位点磷酸化修饰相反的作用.拟南芥的胞质内类受体激酶RIPK磷酸化修饰RIN4蛋白的T21、S160和T166位点.T166位点被磷酸化修饰后, PTI途径的激活受到抑制.与S141位点磷酸化相比, T166位点被磷酸化修饰后发挥的功能具有上位性. ...
The Arabidopsis protein phosphatase PP2C38 negatively regulates the central immune kinase BIK1 1 2016
... RLCK的活性和稳定性受到严格调控.没有PAMP诱导时, 蛋白磷酸酶PP2C38结合并去磷酸化胞质内受体激酶BIK1, 抑制其对RBOHD的磷酸化修饰及下游免疫途径的激活.经PAMP诱导后, PP2C38与FLS2/ EFR-BIK1复合体分离, 解除对BIK1活性的抑制.此外, BIK1对PP2C38第7位丝氨酸残基的磷酸化修饰促进二者分离(Couto et al., 2016).钙离子依赖的蛋白激酶CPK28一方面磷酸化修饰U-box蛋白PUB25/ 26, 增强其泛素蛋白酶活性, 进而促进BIK1的降解; 另一方面CPK28直接磷酸化BIK1, 负调控多种PAMPs诱导后BIK1介导的免疫途径.XLG2/XLG3 (Gα)和AGB1 (Gβ)等G蛋白则通过抑制PUB25/26的活性稳定BIK1 (Monaghan et al., 2014; Liang et al., 2016; Wang et al., 2018a, 2018b). ...
Regulation of pattern recognition receptor signaling in plants 1 2016
... 植物中跨膜的受体类激酶和受体类蛋白(receptor like protein, RLP)识别胞外信号并将其转化为胞内信号.这些RLK或RLP又被称为模式识别受体(pattern recognition receptors, PRRs).典型的RLK由胞外域、单次跨膜的结构域以及胞质内的激酶结构域组成, RLP则不具有胞内激酶结构域.RLK或RLP的胞外域主要有4种类型: 亮氨酸重复序列(leucine-rich repeat, LRR)、赖氨酸基序(lysine motif, LysM)、凝集素样结构域(lectin domain)以及类表皮生长因子(epidermal growth factor, EGF)结构域(Couto and Zipfel, 2016).目前已报道多种受体激酶或受体蛋白参与植物免疫信号的识别和激活(Tang et al., 2017). ...
Plant pathogens and integ- rated defence responses to infection 1 2001
... 植物在漫长的进化过程中形成至少2种免疫途径来抵抗自然界中各种病原微生物的入侵.一种是由定位于细胞质膜的受体识别病原菌关联的分子模式(pathogen-associated molecular patterns, PAMPs)激活的防御反应, 称为PAMP触发的免疫(PAMP- triggered immunity, PTI).病原微生物分泌效应蛋白进入植物细胞, 干扰宿主免疫激活并增强致病性.另一种免疫途径是通过植物抗病蛋白直接或间接识别效应蛋白后激活, 即效应子触发的免疫(effector- triggered immunity, ETI) (Dangl and Jones, 2001; Jones and Dangl, 2006; Dodds and Rathjen, 2010).蛋白磷酸化修饰作用是调控细胞信号转导的主要机制, 这一过程依赖于蛋白激酶和蛋白磷酸酶.在植物-病原微生物互作的复杂过程中, 蛋白激酶和蛋白磷酸酶通过调节多种蛋白的磷酸化状态调控植物免疫途径. ...
MAPKKKα is a positive regulator of cell death associated with both plant immunity and disease 1 2004
... 通过对烟草(Nicotiana benthamiana)和番茄的研究发现, MAPKs除在PTI途径中发挥功能外, 也是ETI途径的组成元件.烟草和番茄中丝/苏氨酸蛋白激酶Pto与丁香假单胞菌分泌的效应子AvrPto/AvrPtoB相互作用, 激活抗病蛋白Prf及ETI响应.此过程中, MAPKKKα和MAPKKKε正调控Pto-介导的ETI途径(del Pozo et al., 2004; Oh and Martin, 2011).在表达抗病蛋白Cf-9的转基因烟草中, 真菌效应子Avr9激活烟草中的MAPK激酶SIPK (SALICYLIC ACID- INDUCED PROTEIN KINASE)和WIPK (WOUND- INDUCED PROTEIN KINASE), 并导致WIPK蛋白积累(Romeis et al., 1999; Pedley and Martin, 2005; Meng and Zhang, 2013).烟草花叶病毒(tobacco mosaic virus, TMV)在烟草抗病蛋白N的作用下磷酸化SIPK和WIPK, 并增加WIPK的mRNA丰度以及蛋白含量(Zhang and Klessig, 1998).此外, AvrRpt2对拟南芥MPK3和MPK6也有激活作用, 与野生型Pst DC3000相比, 携带AvrRpt2效应蛋白的Pst DC3000对MPK3和MPK6的激活能力增强(Eschen-Lippold et al., 2016). ...
The Arabidopsis lectin receptor kinase LecRK-V.5 represses stomatal immunity induced by Pseudomonas syringae pv 1 2012
... 根据胞外凝集素结构域的不同, 凝集素样受体激酶(lectin receptor kinase, LecRK)又分为3种类型: G (GNA-related or S-locus)-、C (calcium-dependent)-以及L (legume)-类(Bouwmeester and Govers, 2009).目前已经鉴定到的L-类LecRK有45个, 它们的功能大部分尚属未知.越来越多的研究表明, L-类LecRK广泛参与植物免疫调控.病原菌或PAMPs诱导LecRK-I.9、LecRK-V.5、LecRK-VI.2以及LecRK- IX.2激活表达; 这些LecRKs功能缺失表现出对不同病原菌的敏感性, 过表达则对病原菌抗性增强(Bou- wmeester et al., 2011; Singh et al., 2012; Wang et al., 2014b; Luo et al., 2017).LecRK-V.5抑制病原菌侵染后的气孔关闭过程; lecrk-V.5缺失突变体对疫霉菌属的抗病性减弱, 对丁香假单胞菌(Pseudomonas syringae pv. tomato (Pst) DC3000)的抗病性增强(Desclos-Theveniau et al., 2012; Wang et al., 2014).L-类LecRK-V.2和LecRK-VII.1结合FLS2, 正调控气孔免疫途径(Yekondi et al., 2018).LecRK- VI.2结合FLS2, 正调控PTI的响应, 参与对丁香假单胞菌和果胶杆菌(Pectobacterium carotovorum)的抗性(Singh et al., 2012; Huang et al., 2014).LecRK- IX.2招募CPKs并磷酸化修饰RBOHD (respiratory burst oxidase homolog protein D), 正调控依赖于RBOHD的水杨酸(salicylic acid, SA)积累以及PTI信号途径(Luo et al., 2017).此外, 有研究表明, 拟南芥Lectin类受体激酶LORE识别细菌脂多糖(lipopoly- saccharides, LPSs)并激活下游免疫信号(Ranf et al., 2015). ...
... MAPKs通过磷酸化修饰底物蛋白改变底物蛋白的亚细胞定位.激活的MPK3对bZIP类转录因子VIP1的磷酸化修饰促进其从细胞质进入细胞核, 激活病程相关基因(pathogenesis-related gene, PR gene)的表达(Djamei et al., 2007). ...
Plant immunity: towards an integrated view of plant-pathogen interactions 1 2010
... 植物在漫长的进化过程中形成至少2种免疫途径来抵抗自然界中各种病原微生物的入侵.一种是由定位于细胞质膜的受体识别病原菌关联的分子模式(pathogen-associated molecular patterns, PAMPs)激活的防御反应, 称为PAMP触发的免疫(PAMP- triggered immunity, PTI).病原微生物分泌效应蛋白进入植物细胞, 干扰宿主免疫激活并增强致病性.另一种免疫途径是通过植物抗病蛋白直接或间接识别效应蛋白后激活, 即效应子触发的免疫(effector- triggered immunity, ETI) (Dangl and Jones, 2001; Jones and Dangl, 2006; Dodds and Rathjen, 2010).蛋白磷酸化修饰作用是调控细胞信号转导的主要机制, 这一过程依赖于蛋白激酶和蛋白磷酸酶.在植物-病原微生物互作的复杂过程中, 蛋白激酶和蛋白磷酸酶通过调节多种蛋白的磷酸化状态调控植物免疫途径. ...
Elicitin recognition confers enhanced resistance to Phytophthora infestans in potato 1 2015
... 由于缺少胞内激酶结构域, LRR-RLP识别配体后通过结合RLKs激活胞内免疫信号.LRR类受体激酶 SOBIR1 (SUPPRESSOR OF BIR1-1 1)结合多种LRR-RLP调控防御反应的激活(Gust and Felix, 2014).拟南芥RLP23识别细菌、真菌以及卵菌分泌的坏死/乙烯诱导的多肽蛋白nlp20 (NECROSIS AND ETHYLENE-INDUCING PEPTIDE 1-LIKE PROTEINS 20)后结合SOBIR1, 并招募BAK1形成复合体激活免疫信号(Albert et al., 2015).RLP30结合SOBIR1和BAK1, 识别死体营养型真菌蛋白激发子SCFE1 (SCLEROTINIA CULTURE FILTRATE ELICITOR 1)并激活下游免疫反应(Zhang et al., 2013).番茄中, Cf-4、Ve1和EIX1 (ETHYLENE- INDUCING XYLANASE 1)等RLPs结合SOBIR1激活真菌诱导的免疫途径(Liebrand et al., 2013).番茄受体激酶ELR (ELICITIN RESPONSE)结合免疫共受体BAK1/SERK3, 通过识别多种疫霉菌激肽蛋白(elicitin proteins)激活广谱且持久的抗病性(Du et al., 2015). ...
Calcium- dependent protein kinase/NADPH oxidase activation cir- cuit is required for rapid defense signal propagation 1 2013
... 钙离子依赖的蛋白激酶(CPK)调控RBOHD及转录因子的激活.拟南芥基因组编码34个CPKs (Cheng et al., 2002; Bigeard et al., 2015), 广泛参与各种胁迫响应的调控.拟南芥CPK5、CPK6和CPK11功能同时缺失可抑制乙烯合成酶ACS2/ACS6的积累及乙烯的合成, 从而影响对死体营养型致病菌灰霉菌的抗病性(Gravino et al., 2015).Gao等(2013)研究表明, 拟南芥抗病蛋白RPS2和RPM1被激活后, CPKs通过磷酸化修饰不同的底物参与调控多种抗病蛋白依赖的免疫激活.CPK1/2/4/11以及CPK5磷酸化质膜定位的NADPH氧化酶促进ROS的生成; 激活的CPK4/ 5/6/11特异地磷酸化修饰WRKY转录因子WRKY8/ 28/48, 协同调控转录重排, 参与抗病蛋白介导的对病原菌的抗性; CPK1/2/4/5则在超敏反应过程中调控程序性细胞死亡的起始(Dubiella et al., 2013; Gao et al., 2013).此外, CPK5参与非典型抗病蛋白TN2激活的抗性.拟南芥胞外复合体亚基EXO70B1缺失突变体中CPK5被过度激活, 参与抗病蛋白TN2介导的免疫自激活过程, 过表达CPK5导致依赖于TN2的免疫自激活以及抗病性增强.TN2结合并且稳定CPK5的活性, 从而调控防御反应(Liu et al., 2017). ...
Intrinsically unstructured proteins and their functions 1 2005
... 拟南芥包含NOI结构域的蛋白RIN4在陆生植物中非常保守, 其并不具有类似已知的蛋白酶类的特征.二级结构预测发现, RIN4蛋白表现出高度的、固有的无序性, 不能形成二级和三级结构, 是植物中固有的无序蛋白(Lee et al., 2015).无序蛋白在与客户蛋白结合时转变成高度折叠状态, 松散并特异地与多种客户蛋白结合(Dyson and Wright, 2005).研究表明, RIN4是AvrRpt2、AvrRpm1、AvrB、HopF2以及AvrPto等多种病原菌效应子蛋白在宿主内攻击的靶标, 表明RIN4在植物-病原微生物互作中具有重要作用(Mackey et al., 2002, 2003; Axtell and Staskawicz, 2003; Luo et al., 2009; Wilton et al., 2010). ...
Bacterial AvrRpt2- like cysteine proteases block activation of the Arabidopsis mitogen-activated protein kinases, MPK4 and MPK11 1 2016
... 通过对烟草(Nicotiana benthamiana)和番茄的研究发现, MAPKs除在PTI途径中发挥功能外, 也是ETI途径的组成元件.烟草和番茄中丝/苏氨酸蛋白激酶Pto与丁香假单胞菌分泌的效应子AvrPto/AvrPtoB相互作用, 激活抗病蛋白Prf及ETI响应.此过程中, MAPKKKα和MAPKKKε正调控Pto-介导的ETI途径(del Pozo et al., 2004; Oh and Martin, 2011).在表达抗病蛋白Cf-9的转基因烟草中, 真菌效应子Avr9激活烟草中的MAPK激酶SIPK (SALICYLIC ACID- INDUCED PROTEIN KINASE)和WIPK (WOUND- INDUCED PROTEIN KINASE), 并导致WIPK蛋白积累(Romeis et al., 1999; Pedley and Martin, 2005; Meng and Zhang, 2013).烟草花叶病毒(tobacco mosaic virus, TMV)在烟草抗病蛋白N的作用下磷酸化SIPK和WIPK, 并增加WIPK的mRNA丰度以及蛋白含量(Zhang and Klessig, 1998).此外, AvrRpt2对拟南芥MPK3和MPK6也有激活作用, 与野生型Pst DC3000相比, 携带AvrRpt2效应蛋白的Pst DC3000对MPK3和MPK6的激活能力增强(Eschen-Lippold et al., 2016). ...
Type 2C protein phosphatases in plants 1 2013
... MAPK的持续磷酸化会导致免疫反应被持续激活, 因此MAPK的活性需要受到严格控制.目前已报道多种磷酸酶在植物免疫过程中负调控MAPK的激活.这些磷酸酶主要包括蛋白酪氨酸磷酸酶(PTPs)、双特异性磷酸酶(DSPs)以及PP2C类蛋白磷酸酶.不同的磷酸酶对MAPK的去磷酸化修饰导致不同的下游信号事件.拟南芥蛋白酪氨酸磷酸酶PTP1 (PROTEIN TYROSINE PHOSPHATASE 1)以及双特异性磷酸酶MKP1 (MAP KINASE PHOSPHATASE 1)和MKP2参与对MPK3/MPK6活性的调控.MKP1通过负调控MPK6途径抑制PAMP激活的免疫应答(Anderson et al., 2011); PTP1结合MPK6与MKP1, 协同抑制MPK6调控的防御反应激活.MKP2与MPK3/MPK6结合并且可在体外去磷酸化修饰MAK3和MPK6 (Lee and Ellis, 2007).真菌激发子诱导的过程中, MKP2抑制MPK6介导的超敏反应(hypersen- sitive response, HR), 从而调控氧化胁迫以及对青枯病病原菌(Ralstonia solanacearum)的抗性(Lumb- reras et al., 2010).此外, MKP1的活性也受到MPK6的调控.PAMP诱导后MKP1蛋白迅速积累, 而这一过程依赖于MPK6对MKP1的磷酸化修饰(Jiang et al., 2017).拟南芥PP2C类蛋白磷酸酶(AP2Cs蛋白磷酸酶)通过激酶结合基序(kinase interaction motif, KIM)与MAPK相互作用(Fuchs et al., 2013).AP2C1负调控植物免疫应答过程中MAPK信号的激活, 过表达AP2C1抑制损伤诱导的乙烯产生以及对死体营养型真菌灰霉菌的抗性.AP2C1功能缺失增强PAMP诱导的MAPK激活、胼胝质积累以及对细菌的抗病性(Schweighofer et al., 2007; Galletti et al., 2011; Sidonskaya et al., 2016; Shubchynskyy et al., 2017). ...
Changes in PUB22 ubiquitination modes triggered by MITOGEN-ACTIVATED PROTEIN KINASE3 dampen the immune response 1 2017
... MAPKs对底物蛋白的磷酸化修饰影响底物蛋白的稳定性.拟南芥中, MPK3/MPK6通过磷酸化修饰ACS2/ACS6调控乙烯产生.泛素蛋白酶降解复合体结合未被磷酸化修饰的ACS6的C末端非催化区域, 导致ACS6经泛素化途径降解.MPK3/MPK6对ACS6的磷酸化修饰在其C末端引入了负电荷, 抑制ACS6的泛素化降解, 增强其蛋白的稳定性, 进而促进乙烯生成(Joo et al., 2008; Han et al., 2010).MPK3/ MPK6磷酸化修饰ERF家族成员ERF6, 增强其蛋白稳定性, 激活PDF1.1 (plant defensin 1.1)、PDF- 1.2a、PDF1.2b和ChiB等多种防御相关基因的表达(Meng et al., 2013).受到PAMP诱导后, MPK3磷酸化修饰WRKY46的S168和S250位点, 促进WRKY46的降解(Sheikh et al., 2016).LIP5参与病原菌诱导的内吞作用和囊泡运输, MPK3和MPK6磷酸化修饰LIP5, 增强LIP5的蛋白稳定性, 从而正调控多泡体(multivesicular bodies, MVB)通路, 影响免疫应答过程中相关因子的重定位(Wang et al., 2014a).E3泛素连接酶PUB22是拟南芥中PTI响应的抑制因子.MPK3对PUB22的磷酸化修饰抑制其自泛素化, 从而导致PUB22积累, 抑制免疫信号激活(Furlan et al., 2017). ...
Arabidopsis MPK3 and MPK6 play different roles in basal and oligo- galacturonide- or flagellin-induced resistance against Bot- rytis cinerea 1 2011
... MAPK的持续磷酸化会导致免疫反应被持续激活, 因此MAPK的活性需要受到严格控制.目前已报道多种磷酸酶在植物免疫过程中负调控MAPK的激活.这些磷酸酶主要包括蛋白酪氨酸磷酸酶(PTPs)、双特异性磷酸酶(DSPs)以及PP2C类蛋白磷酸酶.不同的磷酸酶对MAPK的去磷酸化修饰导致不同的下游信号事件.拟南芥蛋白酪氨酸磷酸酶PTP1 (PROTEIN TYROSINE PHOSPHATASE 1)以及双特异性磷酸酶MKP1 (MAP KINASE PHOSPHATASE 1)和MKP2参与对MPK3/MPK6活性的调控.MKP1通过负调控MPK6途径抑制PAMP激活的免疫应答(Anderson et al., 2011); PTP1结合MPK6与MKP1, 协同抑制MPK6调控的防御反应激活.MKP2与MPK3/MPK6结合并且可在体外去磷酸化修饰MAK3和MPK6 (Lee and Ellis, 2007).真菌激发子诱导的过程中, MKP2抑制MPK6介导的超敏反应(hypersen- sitive response, HR), 从而调控氧化胁迫以及对青枯病病原菌(Ralstonia solanacearum)的抗性(Lumb- reras et al., 2010).此外, MKP1的活性也受到MPK6的调控.PAMP诱导后MKP1蛋白迅速积累, 而这一过程依赖于MPK6对MKP1的磷酸化修饰(Jiang et al., 2017).拟南芥PP2C类蛋白磷酸酶(AP2Cs蛋白磷酸酶)通过激酶结合基序(kinase interaction motif, KIM)与MAPK相互作用(Fuchs et al., 2013).AP2C1负调控植物免疫应答过程中MAPK信号的激活, 过表达AP2C1抑制损伤诱导的乙烯产生以及对死体营养型真菌灰霉菌的抗性.AP2C1功能缺失增强PAMP诱导的MAPK激活、胼胝质积累以及对细菌的抗病性(Schweighofer et al., 2007; Galletti et al., 2011; Sidonskaya et al., 2016; Shubchynskyy et al., 2017). ...
Bifurcation of Arabidopsis NLR immune signaling via Ca 2+-dependent protein kinases 2 2013
... 钙离子依赖的蛋白激酶(CPK)调控RBOHD及转录因子的激活.拟南芥基因组编码34个CPKs (Cheng et al., 2002; Bigeard et al., 2015), 广泛参与各种胁迫响应的调控.拟南芥CPK5、CPK6和CPK11功能同时缺失可抑制乙烯合成酶ACS2/ACS6的积累及乙烯的合成, 从而影响对死体营养型致病菌灰霉菌的抗病性(Gravino et al., 2015).Gao等(2013)研究表明, 拟南芥抗病蛋白RPS2和RPM1被激活后, CPKs通过磷酸化修饰不同的底物参与调控多种抗病蛋白依赖的免疫激活.CPK1/2/4/11以及CPK5磷酸化质膜定位的NADPH氧化酶促进ROS的生成; 激活的CPK4/ 5/6/11特异地磷酸化修饰WRKY转录因子WRKY8/ 28/48, 协同调控转录重排, 参与抗病蛋白介导的对病原菌的抗性; CPK1/2/4/5则在超敏反应过程中调控程序性细胞死亡的起始(Dubiella et al., 2013; Gao et al., 2013).此外, CPK5参与非典型抗病蛋白TN2激活的抗性.拟南芥胞外复合体亚基EXO70B1缺失突变体中CPK5被过度激活, 参与抗病蛋白TN2介导的免疫自激活过程, 过表达CPK5导致依赖于TN2的免疫自激活以及抗病性增强.TN2结合并且稳定CPK5的活性, 从而调控防御反应(Liu et al., 2017). ...
... ; Gao et al., 2013).此外, CPK5参与非典型抗病蛋白TN2激活的抗性.拟南芥胞外复合体亚基EXO70B1缺失突变体中CPK5被过度激活, 参与抗病蛋白TN2介导的免疫自激活过程, 过表达CPK5导致依赖于TN2的免疫自激活以及抗病性增强.TN2结合并且稳定CPK5的活性, 从而调控防御反应(Liu et al., 2017). ...
FLS2: an LRR receptor- like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis 1 2000
... 拟南芥LRR类受体激酶SERKs (SOMATIC EMBRYOGENESIS RECEPTOR KINASEs)作为FLS2/EFR/PEPRs等PRRs的共受体参与免疫信号的识别和激活.PRR接受病原信号后与BAK1/SERK3 (BRI1-ASSOCIATED RECEPTOR KINASE 1)结合, 并且胞内激酶结构域发生自磷酸化修饰.与此同时, PRR与BAK1的激酶域相互磷酸化修饰, 迅速激活免疫信号(Gómez-Gómez and Boller, 2000; Yama- guchi et al., 2010; Yamada et al., 2016b).水稻中, OsSERK2正调控XA21、XA3以及OsFLS2介导的免疫响应(Chen et al., 2014). ...
Ethylene production in Botrytis cinerea - and oligogala- cturonide-induced immunity requires calcium-dependent protein kinases 1 2015
... 钙离子依赖的蛋白激酶(CPK)调控RBOHD及转录因子的激活.拟南芥基因组编码34个CPKs (Cheng et al., 2002; Bigeard et al., 2015), 广泛参与各种胁迫响应的调控.拟南芥CPK5、CPK6和CPK11功能同时缺失可抑制乙烯合成酶ACS2/ACS6的积累及乙烯的合成, 从而影响对死体营养型致病菌灰霉菌的抗病性(Gravino et al., 2015).Gao等(2013)研究表明, 拟南芥抗病蛋白RPS2和RPM1被激活后, CPKs通过磷酸化修饰不同的底物参与调控多种抗病蛋白依赖的免疫激活.CPK1/2/4/11以及CPK5磷酸化质膜定位的NADPH氧化酶促进ROS的生成; 激活的CPK4/ 5/6/11特异地磷酸化修饰WRKY转录因子WRKY8/ 28/48, 协同调控转录重排, 参与抗病蛋白介导的对病原菌的抗性; CPK1/2/4/5则在超敏反应过程中调控程序性细胞死亡的起始(Dubiella et al., 2013; Gao et al., 2013).此外, CPK5参与非典型抗病蛋白TN2激活的抗性.拟南芥胞外复合体亚基EXO70B1缺失突变体中CPK5被过度激活, 参与抗病蛋白TN2介导的免疫自激活过程, 过表达CPK5导致依赖于TN2的免疫自激活以及抗病性增强.TN2结合并且稳定CPK5的活性, 从而调控防御反应(Liu et al., 2017). ...
Receptor like proteins associate with SOBIR1-type of adaptors to form bimolecular receptor kinases 1 2014
... 由于缺少胞内激酶结构域, LRR-RLP识别配体后通过结合RLKs激活胞内免疫信号.LRR类受体激酶 SOBIR1 (SUPPRESSOR OF BIR1-1 1)结合多种LRR-RLP调控防御反应的激活(Gust and Felix, 2014).拟南芥RLP23识别细菌、真菌以及卵菌分泌的坏死/乙烯诱导的多肽蛋白nlp20 (NECROSIS AND ETHYLENE-INDUCING PEPTIDE 1-LIKE PROTEINS 20)后结合SOBIR1, 并招募BAK1形成复合体激活免疫信号(Albert et al., 2015).RLP30结合SOBIR1和BAK1, 识别死体营养型真菌蛋白激发子SCFE1 (SCLEROTINIA CULTURE FILTRATE ELICITOR 1)并激活下游免疫反应(Zhang et al., 2013).番茄中, Cf-4、Ve1和EIX1 (ETHYLENE- INDUCING XYLANASE 1)等RLPs结合SOBIR1激活真菌诱导的免疫途径(Liebrand et al., 2013).番茄受体激酶ELR (ELICITIN RESPONSE)结合免疫共受体BAK1/SERK3, 通过识别多种疫霉菌激肽蛋白(elicitin proteins)激活广谱且持久的抗病性(Du et al., 2015). ...
The leucine- rich repeat receptor kinase BIR2 is a negative regulator of BAK1 in plant immunity 1 2014
... 免疫受体和共受体的活性受到蛋白激酶和蛋白磷酸酶的调控(图2).无病原菌时, 拟南芥LRR-RLK BIR2 (BAK1-INTERACTING RECEPTOR-LIKE KINASE 2)结合BAK1, 防止其与FLS2等PRRs结合而导致PTI途径不必要的激活(Halter et al., 2014).PP2A类蛋白磷酸酶负调控PRR复合体的激活.无PAMP诱导时, 由A1、C4以及B’η/ζ亚基组成的PP2A全酶结合BAK1防止其被磷酸化激活; 配体诱导后, PP2A全酶复合体的活性迅速降低, 导致BAK1被磷酸化激活(Seg-onzac et al., 2014).拟南芥质膜定位的磷蛋白MKKK7结合FLS2, 受鞭毛蛋白诱导时, MKKK7的多个丝氨酸残基被不同程度地磷酸化修饰, 负调控鞭毛蛋白激活的信号以及基础免疫响应.MKKK7通过 直接调控FLS2复合体, 抑制FLS2介导的ROS产生(Mithoe et al., 2016).此外, 水稻PP2C类蛋白磷酸酶XB15 (XA21 BINDING PROTEIN 15)结合并去磷酸化修饰XA21, 负调控XA21介导的免疫激活(Park et al., 2008). ...
Mitogen-activated protein kinase 3 and 6 regulate Botrytis cinerea -induced ethylene production in Arabidopsis 1 2010
... MAPKs对底物蛋白的磷酸化修饰影响底物蛋白的稳定性.拟南芥中, MPK3/MPK6通过磷酸化修饰ACS2/ACS6调控乙烯产生.泛素蛋白酶降解复合体结合未被磷酸化修饰的ACS6的C末端非催化区域, 导致ACS6经泛素化途径降解.MPK3/MPK6对ACS6的磷酸化修饰在其C末端引入了负电荷, 抑制ACS6的泛素化降解, 增强其蛋白的稳定性, 进而促进乙烯生成(Joo et al., 2008; Han et al., 2010).MPK3/ MPK6磷酸化修饰ERF家族成员ERF6, 增强其蛋白稳定性, 激活PDF1.1 (plant defensin 1.1)、PDF- 1.2a、PDF1.2b和ChiB等多种防御相关基因的表达(Meng et al., 2013).受到PAMP诱导后, MPK3磷酸化修饰WRKY46的S168和S250位点, 促进WRKY46的降解(Sheikh et al., 2016).LIP5参与病原菌诱导的内吞作用和囊泡运输, MPK3和MPK6磷酸化修饰LIP5, 增强LIP5的蛋白稳定性, 从而正调控多泡体(multivesicular bodies, MVB)通路, 影响免疫应答过程中相关因子的重定位(Wang et al., 2014a).E3泛素连接酶PUB22是拟南芥中PTI响应的抑制因子.MPK3对PUB22的磷酸化修饰抑制其自泛素化, 从而导致PUB22积累, 抑制免疫信号激活(Furlan et al., 2017). ...
Tomato receptor FLAGELLIN-SENSING 3 binds flgII-28 and acti- vates the plant immune system 1 2016
... LRR类PRRs主要识别蛋白或多肽序列(图1).例如, LRR类受体激酶FLS2 (FLAGELLIN SENSING 2)和EFR (ELONGATION FACTOR-Tu RECEPTOR)分别识别并结合病原菌鞭毛蛋白保守的22个氨基酸残基组成的肽flg22, 以及延伸因子Tu (EF-Tu) N末端的保守氨基酸序列elf18 (Chinchilla et al., 2006; Zipfel et al., 2006); PEPR1/2 (PEP RECEPTOR)识别植物响应病原菌分泌的内源可溶性前体肽pep (pro- peptide) (Yamaguchi et al., 2006); 番茄(Solanum lycopersicum)中FLS3识别细菌flgII-28并增强对细菌的免疫响应(Hind et al., 2016); 水稻(Oryza sativa) LRR-类受体激酶XA21识别水稻白叶枯菌(Xantho- monas oryzae pv. oryzae, Xoo)中保守的蛋白RaxX等(Pruitt et al., 2015). ...
The Arabidopsis LecRK-VI.2 associates with the pattern- recognition receptor FLS2 and primes Nicotiana bent- hamiana pattern-triggered immunity 1 2014
... 根据胞外凝集素结构域的不同, 凝集素样受体激酶(lectin receptor kinase, LecRK)又分为3种类型: G (GNA-related or S-locus)-、C (calcium-dependent)-以及L (legume)-类(Bouwmeester and Govers, 2009).目前已经鉴定到的L-类LecRK有45个, 它们的功能大部分尚属未知.越来越多的研究表明, L-类LecRK广泛参与植物免疫调控.病原菌或PAMPs诱导LecRK-I.9、LecRK-V.5、LecRK-VI.2以及LecRK- IX.2激活表达; 这些LecRKs功能缺失表现出对不同病原菌的敏感性, 过表达则对病原菌抗性增强(Bou- wmeester et al., 2011; Singh et al., 2012; Wang et al., 2014b; Luo et al., 2017).LecRK-V.5抑制病原菌侵染后的气孔关闭过程; lecrk-V.5缺失突变体对疫霉菌属的抗病性减弱, 对丁香假单胞菌(Pseudomonas syringae pv. tomato (Pst) DC3000)的抗病性增强(Desclos-Theveniau et al., 2012; Wang et al., 2014).L-类LecRK-V.2和LecRK-VII.1结合FLS2, 正调控气孔免疫途径(Yekondi et al., 2018).LecRK- VI.2结合FLS2, 正调控PTI的响应, 参与对丁香假单胞菌和果胶杆菌(Pectobacterium carotovorum)的抗性(Singh et al., 2012; Huang et al., 2014).LecRK- IX.2招募CPKs并磷酸化修饰RBOHD (respiratory burst oxidase homolog protein D), 正调控依赖于RBOHD的水杨酸(salicylic acid, SA)积累以及PTI信号途径(Luo et al., 2017).此外, 有研究表明, 拟南芥Lectin类受体激酶LORE识别细菌脂多糖(lipopoly- saccharides, LPSs)并激活下游免疫信号(Ranf et al., 2015). ...
Phosphorylation of the Nicotiana benthamiana WRKY8 transcription factor by MAPK functions in the defense response 1 2011
... MAPKs磷酸化修饰某些WRKY类转录因子, 调控转录重编程.烟草SIPK和WIPK识别并直接磷酸化I类WRKY转录因子WRKY8, 增强WRKY8与W-box的结合和反式激活活性, 进而激活HMGR2 (3-HYD- ROXY-3-METHYLGLUTARYL CoA REDUCTASE 2)的表达, 产生类异戊二烯植物抗毒素, 促进植物的免疫反应(Ishihama et al., 2011).拟南芥中, 与烟草WRKY8同源的转录因子WRKY33与MPK4以及MKS1在细胞核内形成复合体; 受到flg22或者病原菌刺激时, MPK4磷酸化修饰MKS1并促使WRKY33- MKS1与MPK4分离, 磷酸化的WRKY33随后结合植物抗毒素(camalexin)合成相关基因PAD3启动子区的W-box, 激活PAD3的表达并促进camalexin的合成(Andreasson et al., 2005; Qiu et al., 2008; Len- zoni et al., 2018).WRKY33还参与MPK3/6调控的camalexin的合成过程.灰霉菌诱导时, MPK3/6磷酸化修饰WRKY33, 促进相关基因转录重编程(Mao et al., 2011; Li et al., 2012).MPK6结合并磷酸化ERF104调控防御相关基因的表达(Bethke et al., 2009).拟南芥中ASR3作为转录抑制因子负调控PAMP诱导的免疫响应.PAMP处理后MPK4磷酸化ASR3第189位苏氨酸, 磷酸化的ASR3与DNA结合能力增强, 从而抑制flg22诱导的防御基因的表达(Li et al., 2015). ...
Phosphorylation of Arabidopsis MAP Kinase Phosphatase 1 (MKP1) is required for PAMP responses and resistance against Bacteria 1 2017
... MAPK的持续磷酸化会导致免疫反应被持续激活, 因此MAPK的活性需要受到严格控制.目前已报道多种磷酸酶在植物免疫过程中负调控MAPK的激活.这些磷酸酶主要包括蛋白酪氨酸磷酸酶(PTPs)、双特异性磷酸酶(DSPs)以及PP2C类蛋白磷酸酶.不同的磷酸酶对MAPK的去磷酸化修饰导致不同的下游信号事件.拟南芥蛋白酪氨酸磷酸酶PTP1 (PROTEIN TYROSINE PHOSPHATASE 1)以及双特异性磷酸酶MKP1 (MAP KINASE PHOSPHATASE 1)和MKP2参与对MPK3/MPK6活性的调控.MKP1通过负调控MPK6途径抑制PAMP激活的免疫应答(Anderson et al., 2011); PTP1结合MPK6与MKP1, 协同抑制MPK6调控的防御反应激活.MKP2与MPK3/MPK6结合并且可在体外去磷酸化修饰MAK3和MPK6 (Lee and Ellis, 2007).真菌激发子诱导的过程中, MKP2抑制MPK6介导的超敏反应(hypersen- sitive response, HR), 从而调控氧化胁迫以及对青枯病病原菌(Ralstonia solanacearum)的抗性(Lumb- reras et al., 2010).此外, MKP1的活性也受到MPK6的调控.PAMP诱导后MKP1蛋白迅速积累, 而这一过程依赖于MPK6对MKP1的磷酸化修饰(Jiang et al., 2017).拟南芥PP2C类蛋白磷酸酶(AP2Cs蛋白磷酸酶)通过激酶结合基序(kinase interaction motif, KIM)与MAPK相互作用(Fuchs et al., 2013).AP2C1负调控植物免疫应答过程中MAPK信号的激活, 过表达AP2C1抑制损伤诱导的乙烯产生以及对死体营养型真菌灰霉菌的抗性.AP2C1功能缺失增强PAMP诱导的MAPK激活、胼胝质积累以及对细菌的抗病性(Schweighofer et al., 2007; Galletti et al., 2011; Sidonskaya et al., 2016; Shubchynskyy et al., 2017). ...
The plant immune system 1 2006
... 植物在漫长的进化过程中形成至少2种免疫途径来抵抗自然界中各种病原微生物的入侵.一种是由定位于细胞质膜的受体识别病原菌关联的分子模式(pathogen-associated molecular patterns, PAMPs)激活的防御反应, 称为PAMP触发的免疫(PAMP- triggered immunity, PTI).病原微生物分泌效应蛋白进入植物细胞, 干扰宿主免疫激活并增强致病性.另一种免疫途径是通过植物抗病蛋白直接或间接识别效应蛋白后激活, 即效应子触发的免疫(effector- triggered immunity, ETI) (Dangl and Jones, 2001; Jones and Dangl, 2006; Dodds and Rathjen, 2010).蛋白磷酸化修饰作用是调控细胞信号转导的主要机制, 这一过程依赖于蛋白激酶和蛋白磷酸酶.在植物-病原微生物互作的复杂过程中, 蛋白激酶和蛋白磷酸酶通过调节多种蛋白的磷酸化状态调控植物免疫途径. ...
MAPK phos- phorylation-induced stabilization of ACS6 protein is me- diated by the non-catalytic C-terminal domain, which also contains the cis -determinant for rapid degradation by the 26S proteasome pathway 1 2008
... MAPKs对底物蛋白的磷酸化修饰影响底物蛋白的稳定性.拟南芥中, MPK3/MPK6通过磷酸化修饰ACS2/ACS6调控乙烯产生.泛素蛋白酶降解复合体结合未被磷酸化修饰的ACS6的C末端非催化区域, 导致ACS6经泛素化途径降解.MPK3/MPK6对ACS6的磷酸化修饰在其C末端引入了负电荷, 抑制ACS6的泛素化降解, 增强其蛋白的稳定性, 进而促进乙烯生成(Joo et al., 2008; Han et al., 2010).MPK3/ MPK6磷酸化修饰ERF家族成员ERF6, 增强其蛋白稳定性, 激活PDF1.1 (plant defensin 1.1)、PDF- 1.2a、PDF1.2b和ChiB等多种防御相关基因的表达(Meng et al., 2013).受到PAMP诱导后, MPK3磷酸化修饰WRKY46的S168和S250位点, 促进WRKY46的降解(Sheikh et al., 2016).LIP5参与病原菌诱导的内吞作用和囊泡运输, MPK3和MPK6磷酸化修饰LIP5, 增强LIP5的蛋白稳定性, 从而正调控多泡体(multivesicular bodies, MVB)通路, 影响免疫应答过程中相关因子的重定位(Wang et al., 2014a).E3泛素连接酶PUB22是拟南芥中PTI响应的抑制因子.MPK3对PUB22的磷酸化修饰抑制其自泛素化, 从而导致PUB22积累, 抑制免疫信号激活(Furlan et al., 2017). ...
Direct regulation of the NADPH oxidase RBOHD by the PRR-associated kinase BIK1 during plant immunity 1 2014
... RLCK经PRRs激活后, 一方面招募钙离子依赖的蛋白激酶(CPKs)促进产生活性氧(ROS); 另一方面激活下游MAPK信号级联反应(图2).植物中ROS主要由NADPH氧化酶RBOHD催化生成.目前已报道多种RLCKs调控RBOHD的活性.例如, 被flg22诱导激活的BIK1一方面通过磷酸化修饰RBOHD的多个氨基酸位点激活RBOHD的活性; 另一方面通过磷酸化植物Gα蛋白XLG2 (EXTRA LARGE G-PROTEIN 2)促进ROS的产生(Kadota et al., 2014; Li et al., 2014b; Liang et al., 2016).GTP酶促进蛋白(RGS1)维持FLS2结合的G蛋白不活跃状态.被?g22诱导激活的BIK1/PBL1磷酸化RGS1的S428和S431位点, 促进RGS1与FLS2-G蛋白复合体分离, 从而正调控免疫信号的激活(Liang et al., 2018).BSK1结合FLS2调控ROS的生成.bsk1缺失突变体中, flg22诱导的ROS爆发减弱, 病原菌诱导的SA积累受到阻碍并且对白粉菌、丁香假单胞菌和卵菌的敏感性增强(Shi et al., 2013).在无病原菌侵染时, PBL13通过结合RBOHD负调控植物对致病菌的先天免疫途径(Lin et al., 2015). ...
Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor 1 2006
... LysM类和EGF类PRRs主要识别包含碳水化合物的分子, 如真菌几丁质(chitin)、细菌肽聚糖(peptidog- lycan, PGN)和丛枝菌根菌分泌的脂质几丁寡糖(lipo- chitooligosaccharides, LCOs)或者植物细胞壁衍生的寡聚半乳糖醛酸(oligogalacturonides, OGs)等(图1).拟南芥中的3个LysM类受体激酶CERK1 (CHITIN ELICITOR RECEPTOR KINASE 1)、LYK4 (LysM- CONTAINING RECEPTOR-LIKE KINASE 4)和LYK5共同调控几丁质的识别及免疫信号的激活(Miya et al., 2007; Wan et al., 2012; Cao et al., 2014); LYK1和LYK3参与对不同结构的PGN信号的识别, 在AtCERK1存在的条件下激活PGN诱导的防御信号(Willmann et al., 2011).水稻LysM类RLK OsCERK1胞外的LysM结构域不能直接结合几丁质, LysM类受体蛋白OsCEBiP (CHITIN OLIFOSACCHARIDE ELICITOR BINDING PROTEIN)作为受体结合几丁质, 随后结合OsCERK1激活几丁质诱导的防御反应(Kaku et al., 2006; Shinya et al., 2015).拟南芥EGF类受体激酶WAK1 (WALL-ASSOCIATED KINASE 1)接受OGs信号并激活防御反应(Brutus et al., 2010).水稻免疫信号激活过程中, OsWAK1发生自磷酸化并磷酸化下游转录因子OsRFP1.此外, 水稻稻瘟菌侵染、机械损伤及水杨酸和茉莉酸甲酯处理均可以激活OsWAK1转录水平的表达(Li et al., 2009). ...
Two redundant receptor-like cytoplasmic kinases function downstream of pattern recognition receptors to regulate activation of SA biosynthesis 1 2016
... 胞质内的受体激酶结合RLK/RLP, 共同调控植物的生长发育、激素信号以及生物和非生物胁迫等过程.研究表明, 拟南芥RLCK-VII家族的PBS1 (AvrPphB SUSCEPTIBLE 1)、BIK1 (BOTRYTIS-INDUCED KINASE 1)、PBL1 (PBS1- LIKE 1)、PBL2、PBL27、PCRKs (PTI COMPROMISED RECEPTOR-LIKE CYTOPLASMIC KINASE)以及RLCK-XII家族成员BSK1 (BRASSINOSTEROID-SIGNALING KINASE 1)参与植物免疫信号传递(图1).例如, FLS2、EFR、PEPRs以及CERK1等多种PRR被激活后磷酸化修饰BIK1, 导致BIK1与PRRs复合体分离并进入细胞质激活下游免疫信号(Lu et al., 2010; Yang et al., 2010; Liu et al., 2013).PCRK1和PCRK2结合FLS2并在PAMPs诱导后被磷酸化激活, 冗余地诱导转录因子SARD1 (SYSTEMIC ACQUIRED RESISTANCE DEFICIENT 1)和CBP60g (CALMODULIN-BINFING PROTEIN 60g)的表达, 促进SA的合成(Kong et al., 2016).水稻RLCK-VII家族的OsRLCK185和OsRL- CK176结合CERK1, 传递PGN和几丁质激活的免疫信号(Yamaguchi et al., 2013; Ao et al., 2014).Os- RLCK185在拟南芥中的同源蛋白PBL27被CERK1磷酸化激活后, 正调控几丁质诱导的防御反应(Shinya et al., 2014). ...
The receptor-like cytoplasmic kinase BIK1 localizes to the nucleus and regulates defense hormone expression during plant innate immunity 1 2018
Phosphorylation of the plant immune regulator RPM1- INTERACTING PROTEIN4 enhances plant plasma mem- brane H +-ATPase activity and inhibits flagellin-triggered immune responses in Arabidopsis 1 2015
... 拟南芥包含NOI结构域的蛋白RIN4在陆生植物中非常保守, 其并不具有类似已知的蛋白酶类的特征.二级结构预测发现, RIN4蛋白表现出高度的、固有的无序性, 不能形成二级和三级结构, 是植物中固有的无序蛋白(Lee et al., 2015).无序蛋白在与客户蛋白结合时转变成高度折叠状态, 松散并特异地与多种客户蛋白结合(Dyson and Wright, 2005).研究表明, RIN4是AvrRpt2、AvrRpm1、AvrB、HopF2以及AvrPto等多种病原菌效应子蛋白在宿主内攻击的靶标, 表明RIN4在植物-病原微生物互作中具有重要作用(Mackey et al., 2002, 2003; Axtell and Staskawicz, 2003; Luo et al., 2009; Wilton et al., 2010). ...
Arabidopsis MAPK phosphatase 2 (MKP2) positively regulates oxidative stress tolerance and inactivates the MPK3 and MPK6 MAPKs 1 2007
... MAPK的持续磷酸化会导致免疫反应被持续激活, 因此MAPK的活性需要受到严格控制.目前已报道多种磷酸酶在植物免疫过程中负调控MAPK的激活.这些磷酸酶主要包括蛋白酪氨酸磷酸酶(PTPs)、双特异性磷酸酶(DSPs)以及PP2C类蛋白磷酸酶.不同的磷酸酶对MAPK的去磷酸化修饰导致不同的下游信号事件.拟南芥蛋白酪氨酸磷酸酶PTP1 (PROTEIN TYROSINE PHOSPHATASE 1)以及双特异性磷酸酶MKP1 (MAP KINASE PHOSPHATASE 1)和MKP2参与对MPK3/MPK6活性的调控.MKP1通过负调控MPK6途径抑制PAMP激活的免疫应答(Anderson et al., 2011); PTP1结合MPK6与MKP1, 协同抑制MPK6调控的防御反应激活.MKP2与MPK3/MPK6结合并且可在体外去磷酸化修饰MAK3和MPK6 (Lee and Ellis, 2007).真菌激发子诱导的过程中, MKP2抑制MPK6介导的超敏反应(hypersen- sitive response, HR), 从而调控氧化胁迫以及对青枯病病原菌(Ralstonia solanacearum)的抗性(Lumb- reras et al., 2010).此外, MKP1的活性也受到MPK6的调控.PAMP诱导后MKP1蛋白迅速积累, 而这一过程依赖于MPK6对MKP1的磷酸化修饰(Jiang et al., 2017).拟南芥PP2C类蛋白磷酸酶(AP2Cs蛋白磷酸酶)通过激酶结合基序(kinase interaction motif, KIM)与MAPK相互作用(Fuchs et al., 2013).AP2C1负调控植物免疫应答过程中MAPK信号的激活, 过表达AP2C1抑制损伤诱导的乙烯产生以及对死体营养型真菌灰霉菌的抗性.AP2C1功能缺失增强PAMP诱导的MAPK激活、胼胝质积累以及对细菌的抗病性(Schweighofer et al., 2007; Galletti et al., 2011; Sidonskaya et al., 2016; Shubchynskyy et al., 2017). ...
Predicting plant immunity gene expression by identifying the decoding mechanism of calcium signatures 1 2018
... MAPKs磷酸化修饰某些WRKY类转录因子, 调控转录重编程.烟草SIPK和WIPK识别并直接磷酸化I类WRKY转录因子WRKY8, 增强WRKY8与W-box的结合和反式激活活性, 进而激活HMGR2 (3-HYD- ROXY-3-METHYLGLUTARYL CoA REDUCTASE 2)的表达, 产生类异戊二烯植物抗毒素, 促进植物的免疫反应(Ishihama et al., 2011).拟南芥中, 与烟草WRKY8同源的转录因子WRKY33与MPK4以及MKS1在细胞核内形成复合体; 受到flg22或者病原菌刺激时, MPK4磷酸化修饰MKS1并促使WRKY33- MKS1与MPK4分离, 磷酸化的WRKY33随后结合植物抗毒素(camalexin)合成相关基因PAD3启动子区的W-box, 激活PAD3的表达并促进camalexin的合成(Andreasson et al., 2005; Qiu et al., 2008; Len- zoni et al., 2018).WRKY33还参与MPK3/6调控的camalexin的合成过程.灰霉菌诱导时, MPK3/6磷酸化修饰WRKY33, 促进相关基因转录重编程(Mao et al., 2011; Li et al., 2012).MPK6结合并磷酸化ERF104调控防御相关基因的表达(Bethke et al., 2009).拟南芥中ASR3作为转录抑制因子负调控PAMP诱导的免疫响应.PAMP处理后MPK4磷酸化ASR3第189位苏氨酸, 磷酸化的ASR3与DNA结合能力增强, 从而抑制flg22诱导的防御基因的表达(Li et al., 2015). ...
Phosphorylation of trihelix transcriptional repressor ASR3 by MAP KINASE4 negatively regulates Arabidopsis immunity 1 2015
... MAPKs磷酸化修饰某些WRKY类转录因子, 调控转录重编程.烟草SIPK和WIPK识别并直接磷酸化I类WRKY转录因子WRKY8, 增强WRKY8与W-box的结合和反式激活活性, 进而激活HMGR2 (3-HYD- ROXY-3-METHYLGLUTARYL CoA REDUCTASE 2)的表达, 产生类异戊二烯植物抗毒素, 促进植物的免疫反应(Ishihama et al., 2011).拟南芥中, 与烟草WRKY8同源的转录因子WRKY33与MPK4以及MKS1在细胞核内形成复合体; 受到flg22或者病原菌刺激时, MPK4磷酸化修饰MKS1并促使WRKY33- MKS1与MPK4分离, 磷酸化的WRKY33随后结合植物抗毒素(camalexin)合成相关基因PAD3启动子区的W-box, 激活PAD3的表达并促进camalexin的合成(Andreasson et al., 2005; Qiu et al., 2008; Len- zoni et al., 2018).WRKY33还参与MPK3/6调控的camalexin的合成过程.灰霉菌诱导时, MPK3/6磷酸化修饰WRKY33, 促进相关基因转录重编程(Mao et al., 2011; Li et al., 2012).MPK6结合并磷酸化ERF104调控防御相关基因的表达(Bethke et al., 2009).拟南芥中ASR3作为转录抑制因子负调控PAMP诱导的免疫响应.PAMP处理后MPK4磷酸化ASR3第189位苏氨酸, 磷酸化的ASR3与DNA结合能力增强, 从而抑制flg22诱导的防御基因的表达(Li et al., 2015). ...
Modulation of RNA polymerase II phosphorylation downstream of pathogen perception orchestrates plant immunity 1 2014
Dual-level regulation of ACC synthase activity by MPK3/MPK6 cascade and its downstream WRKY transcription factor during ethylene induction in Arabi- dopsis 1 2012
... MAPKs磷酸化修饰某些WRKY类转录因子, 调控转录重编程.烟草SIPK和WIPK识别并直接磷酸化I类WRKY转录因子WRKY8, 增强WRKY8与W-box的结合和反式激活活性, 进而激活HMGR2 (3-HYD- ROXY-3-METHYLGLUTARYL CoA REDUCTASE 2)的表达, 产生类异戊二烯植物抗毒素, 促进植物的免疫反应(Ishihama et al., 2011).拟南芥中, 与烟草WRKY8同源的转录因子WRKY33与MPK4以及MKS1在细胞核内形成复合体; 受到flg22或者病原菌刺激时, MPK4磷酸化修饰MKS1并促使WRKY33- MKS1与MPK4分离, 磷酸化的WRKY33随后结合植物抗毒素(camalexin)合成相关基因PAD3启动子区的W-box, 激活PAD3的表达并促进camalexin的合成(Andreasson et al., 2005; Qiu et al., 2008; Len- zoni et al., 2018).WRKY33还参与MPK3/6调控的camalexin的合成过程.灰霉菌诱导时, MPK3/6磷酸化修饰WRKY33, 促进相关基因转录重编程(Mao et al., 2011; Li et al., 2012).MPK6结合并磷酸化ERF104调控防御相关基因的表达(Bethke et al., 2009).拟南芥中ASR3作为转录抑制因子负调控PAMP诱导的免疫响应.PAMP处理后MPK4磷酸化ASR3第189位苏氨酸, 磷酸化的ASR3与DNA结合能力增强, 从而抑制flg22诱导的防御基因的表达(Li et al., 2015). ...
A novel wall-associated receptor-like protein kinase gene, Os- WAK1 , plays important roles in rice blast disease resis- tance 1 2009
... LysM类和EGF类PRRs主要识别包含碳水化合物的分子, 如真菌几丁质(chitin)、细菌肽聚糖(peptidog- lycan, PGN)和丛枝菌根菌分泌的脂质几丁寡糖(lipo- chitooligosaccharides, LCOs)或者植物细胞壁衍生的寡聚半乳糖醛酸(oligogalacturonides, OGs)等(图1).拟南芥中的3个LysM类受体激酶CERK1 (CHITIN ELICITOR RECEPTOR KINASE 1)、LYK4 (LysM- CONTAINING RECEPTOR-LIKE KINASE 4)和LYK5共同调控几丁质的识别及免疫信号的激活(Miya et al., 2007; Wan et al., 2012; Cao et al., 2014); LYK1和LYK3参与对不同结构的PGN信号的识别, 在AtCERK1存在的条件下激活PGN诱导的防御信号(Willmann et al., 2011).水稻LysM类RLK OsCERK1胞外的LysM结构域不能直接结合几丁质, LysM类受体蛋白OsCEBiP (CHITIN OLIFOSACCHARIDE ELICITOR BINDING PROTEIN)作为受体结合几丁质, 随后结合OsCERK1激活几丁质诱导的防御反应(Kaku et al., 2006; Shinya et al., 2015).拟南芥EGF类受体激酶WAK1 (WALL-ASSOCIATED KINASE 1)接受OGs信号并激活防御反应(Brutus et al., 2010).水稻免疫信号激活过程中, OsWAK1发生自磷酸化并磷酸化下游转录因子OsRFP1.此外, 水稻稻瘟菌侵染、机械损伤及水杨酸和茉莉酸甲酯处理均可以激活OsWAK1转录水平的表达(Li et al., 2009). ...
BAK1, an Arabidopsis LRR receptor-like protein kinase, interacts with BRI1 and modulates brassinosteroid signaling 0 2002
The FLS2-associated kinase BIK1 directly phosphorylates the NADPH oxidase RbohD to control plant immunity 1 2014
... RLCK经PRRs激活后, 一方面招募钙离子依赖的蛋白激酶(CPKs)促进产生活性氧(ROS); 另一方面激活下游MAPK信号级联反应(图2).植物中ROS主要由NADPH氧化酶RBOHD催化生成.目前已报道多种RLCKs调控RBOHD的活性.例如, 被flg22诱导激活的BIK1一方面通过磷酸化修饰RBOHD的多个氨基酸位点激活RBOHD的活性; 另一方面通过磷酸化植物Gα蛋白XLG2 (EXTRA LARGE G-PROTEIN 2)促进ROS的产生(Kadota et al., 2014; Li et al., 2014b; Liang et al., 2016).GTP酶促进蛋白(RGS1)维持FLS2结合的G蛋白不活跃状态.被?g22诱导激活的BIK1/PBL1磷酸化RGS1的S428和S431位点, 促进RGS1与FLS2-G蛋白复合体分离, 从而正调控免疫信号的激活(Liang et al., 2018).BSK1结合FLS2调控ROS的生成.bsk1缺失突变体中, flg22诱导的ROS爆发减弱, 病原菌诱导的SA积累受到阻碍并且对白粉菌、丁香假单胞菌和卵菌的敏感性增强(Shi et al., 2013).在无病原菌侵染时, PBL13通过结合RBOHD负调控植物对致病菌的先天免疫途径(Lin et al., 2015). ...
Arabidopsis heterotrimeric G proteins regulate immunity by directly coupling to the FLS2 receptor 2 2016
... RLCK经PRRs激活后, 一方面招募钙离子依赖的蛋白激酶(CPKs)促进产生活性氧(ROS); 另一方面激活下游MAPK信号级联反应(图2).植物中ROS主要由NADPH氧化酶RBOHD催化生成.目前已报道多种RLCKs调控RBOHD的活性.例如, 被flg22诱导激活的BIK1一方面通过磷酸化修饰RBOHD的多个氨基酸位点激活RBOHD的活性; 另一方面通过磷酸化植物Gα蛋白XLG2 (EXTRA LARGE G-PROTEIN 2)促进ROS的产生(Kadota et al., 2014; Li et al., 2014b; Liang et al., 2016).GTP酶促进蛋白(RGS1)维持FLS2结合的G蛋白不活跃状态.被?g22诱导激活的BIK1/PBL1磷酸化RGS1的S428和S431位点, 促进RGS1与FLS2-G蛋白复合体分离, 从而正调控免疫信号的激活(Liang et al., 2018).BSK1结合FLS2调控ROS的生成.bsk1缺失突变体中, flg22诱导的ROS爆发减弱, 病原菌诱导的SA积累受到阻碍并且对白粉菌、丁香假单胞菌和卵菌的敏感性增强(Shi et al., 2013).在无病原菌侵染时, PBL13通过结合RBOHD负调控植物对致病菌的先天免疫途径(Lin et al., 2015). ...
... RLCK的活性和稳定性受到严格调控.没有PAMP诱导时, 蛋白磷酸酶PP2C38结合并去磷酸化胞质内受体激酶BIK1, 抑制其对RBOHD的磷酸化修饰及下游免疫途径的激活.经PAMP诱导后, PP2C38与FLS2/ EFR-BIK1复合体分离, 解除对BIK1活性的抑制.此外, BIK1对PP2C38第7位丝氨酸残基的磷酸化修饰促进二者分离(Couto et al., 2016).钙离子依赖的蛋白激酶CPK28一方面磷酸化修饰U-box蛋白PUB25/ 26, 增强其泛素蛋白酶活性, 进而促进BIK1的降解; 另一方面CPK28直接磷酸化BIK1, 负调控多种PAMPs诱导后BIK1介导的免疫途径.XLG2/XLG3 (Gα)和AGB1 (Gβ)等G蛋白则通过抑制PUB25/26的活性稳定BIK1 (Monaghan et al., 2014; Liang et al., 2016; Wang et al., 2018a, 2018b). ...
Ligand-triggered de-repression of Arabidopsis heterotrimeric G proteins coupled to immune receptor kinases 1 2018
... RLCK经PRRs激活后, 一方面招募钙离子依赖的蛋白激酶(CPKs)促进产生活性氧(ROS); 另一方面激活下游MAPK信号级联反应(图2).植物中ROS主要由NADPH氧化酶RBOHD催化生成.目前已报道多种RLCKs调控RBOHD的活性.例如, 被flg22诱导激活的BIK1一方面通过磷酸化修饰RBOHD的多个氨基酸位点激活RBOHD的活性; 另一方面通过磷酸化植物Gα蛋白XLG2 (EXTRA LARGE G-PROTEIN 2)促进ROS的产生(Kadota et al., 2014; Li et al., 2014b; Liang et al., 2016).GTP酶促进蛋白(RGS1)维持FLS2结合的G蛋白不活跃状态.被?g22诱导激活的BIK1/PBL1磷酸化RGS1的S428和S431位点, 促进RGS1与FLS2-G蛋白复合体分离, 从而正调控免疫信号的激活(Liang et al., 2018).BSK1结合FLS2调控ROS的生成.bsk1缺失突变体中, flg22诱导的ROS爆发减弱, 病原菌诱导的SA积累受到阻碍并且对白粉菌、丁香假单胞菌和卵菌的敏感性增强(Shi et al., 2013).在无病原菌侵染时, PBL13通过结合RBOHD负调控植物对致病菌的先天免疫途径(Lin et al., 2015). ...
Receptor-like kinase SOBIR1/EVR interacts with receptor-like proteins in plant immunity against fungal infection 1 2013
... 由于缺少胞内激酶结构域, LRR-RLP识别配体后通过结合RLKs激活胞内免疫信号.LRR类受体激酶 SOBIR1 (SUPPRESSOR OF BIR1-1 1)结合多种LRR-RLP调控防御反应的激活(Gust and Felix, 2014).拟南芥RLP23识别细菌、真菌以及卵菌分泌的坏死/乙烯诱导的多肽蛋白nlp20 (NECROSIS AND ETHYLENE-INDUCING PEPTIDE 1-LIKE PROTEINS 20)后结合SOBIR1, 并招募BAK1形成复合体激活免疫信号(Albert et al., 2015).RLP30结合SOBIR1和BAK1, 识别死体营养型真菌蛋白激发子SCFE1 (SCLEROTINIA CULTURE FILTRATE ELICITOR 1)并激活下游免疫反应(Zhang et al., 2013).番茄中, Cf-4、Ve1和EIX1 (ETHYLENE- INDUCING XYLANASE 1)等RLPs结合SOBIR1激活真菌诱导的免疫途径(Liebrand et al., 2013).番茄受体激酶ELR (ELICITIN RESPONSE)结合免疫共受体BAK1/SERK3, 通过识别多种疫霉菌激肽蛋白(elicitin proteins)激活广谱且持久的抗病性(Du et al., 2015). ...
Tyrosine phosphorylation of protein kinase com- plex BAK1/BIK1 mediates Arabidopsis innate immunity 1 2014
... 目前已报道的参与免疫调控的受体激酶主要是丝氨酸/苏氨酸类蛋白激酶.近年来, 越来越多的研究表明, 这些受体激酶的酪氨酸残基的磷酸化修饰同样在植物免疫调控中发挥重要作用.例如, BAK1第403位酪氨酸残基的磷酸化修饰对于BAK1的功能以及免疫信号的激活非常重要, 该位点在多种植物SERKs家族中非常保守.大多数LRR-RK中存在Y403的类似位点Tyr-Via, 这一位点的磷酸化修饰同样影响多种受体激酶对配体的识别和信号激活(Macho et al., 2014; Perraki et al., 2018).EFR介导的对丁香假单胞菌的免疫途径依赖于其Tyr-Via位点Y836的磷酸化.丁香假单胞菌分泌具有酪氨酸磷酸酶活性的效应子HopAO1, 可减少EFR的磷酸化并抑制随后的免疫响应(Macho et al., 2014).拟南芥CERK1识别几丁质后, 对下游信号的激活依赖于其第428位的酪氨酸残基的自磷酸化修饰.Chitin信号激活后, CERK1招募预测的丝氨酸/苏氨酸蛋白磷酸酶CIPP1 (CERK1- INTERACTING PROTEIN PHOSPHATASE 1), 对Y428去磷酸化修饰, 关闭CERK1介导的免疫信号, 以防止防御反应被持续激活(Liu et al., 2018).此外, BAK1通过磷酸化修饰BIK1的酪氨酸残基调控免疫信号的激活, BIK1第150、243及250位酪氨酸位点的磷酸化修饰影响BIK1在免疫调控中的功能(Lin et al., 2013, 2014). ...
Inverse modulation of plant immune and brassinosteroid signaling pathways by the receptor-like cytoplasmic kinase BIK1 1 2013
... 目前已报道的参与免疫调控的受体激酶主要是丝氨酸/苏氨酸类蛋白激酶.近年来, 越来越多的研究表明, 这些受体激酶的酪氨酸残基的磷酸化修饰同样在植物免疫调控中发挥重要作用.例如, BAK1第403位酪氨酸残基的磷酸化修饰对于BAK1的功能以及免疫信号的激活非常重要, 该位点在多种植物SERKs家族中非常保守.大多数LRR-RK中存在Y403的类似位点Tyr-Via, 这一位点的磷酸化修饰同样影响多种受体激酶对配体的识别和信号激活(Macho et al., 2014; Perraki et al., 2018).EFR介导的对丁香假单胞菌的免疫途径依赖于其Tyr-Via位点Y836的磷酸化.丁香假单胞菌分泌具有酪氨酸磷酸酶活性的效应子HopAO1, 可减少EFR的磷酸化并抑制随后的免疫响应(Macho et al., 2014).拟南芥CERK1识别几丁质后, 对下游信号的激活依赖于其第428位的酪氨酸残基的自磷酸化修饰.Chitin信号激活后, CERK1招募预测的丝氨酸/苏氨酸蛋白磷酸酶CIPP1 (CERK1- INTERACTING PROTEIN PHOSPHATASE 1), 对Y428去磷酸化修饰, 关闭CERK1介导的免疫信号, 以防止防御反应被持续激活(Liu et al., 2018).此外, BAK1通过磷酸化修饰BIK1的酪氨酸残基调控免疫信号的激活, BIK1第150、243及250位酪氨酸位点的磷酸化修饰影响BIK1在免疫调控中的功能(Lin et al., 2013, 2014). ...
PBL13 is a serine/threonine protein kinase that negatively regulates Arabidopsis immune responses 1 2015
... RLCK经PRRs激活后, 一方面招募钙离子依赖的蛋白激酶(CPKs)促进产生活性氧(ROS); 另一方面激活下游MAPK信号级联反应(图2).植物中ROS主要由NADPH氧化酶RBOHD催化生成.目前已报道多种RLCKs调控RBOHD的活性.例如, 被flg22诱导激活的BIK1一方面通过磷酸化修饰RBOHD的多个氨基酸位点激活RBOHD的活性; 另一方面通过磷酸化植物Gα蛋白XLG2 (EXTRA LARGE G-PROTEIN 2)促进ROS的产生(Kadota et al., 2014; Li et al., 2014b; Liang et al., 2016).GTP酶促进蛋白(RGS1)维持FLS2结合的G蛋白不活跃状态.被?g22诱导激活的BIK1/PBL1磷酸化RGS1的S428和S431位点, 促进RGS1与FLS2-G蛋白复合体分离, 从而正调控免疫信号的激活(Liang et al., 2018).BSK1结合FLS2调控ROS的生成.bsk1缺失突变体中, flg22诱导的ROS爆发减弱, 病原菌诱导的SA积累受到阻碍并且对白粉菌、丁香假单胞菌和卵菌的敏感性增强(Shi et al., 2013).在无病原菌侵染时, PBL13通过结合RBOHD负调控植物对致病菌的先天免疫途径(Lin et al., 2015). ...
A tyrosine phosphorylation cycle regulates fungal activation of a plant receptor Ser/Thr kinase 1 2018
... 目前已报道的参与免疫调控的受体激酶主要是丝氨酸/苏氨酸类蛋白激酶.近年来, 越来越多的研究表明, 这些受体激酶的酪氨酸残基的磷酸化修饰同样在植物免疫调控中发挥重要作用.例如, BAK1第403位酪氨酸残基的磷酸化修饰对于BAK1的功能以及免疫信号的激活非常重要, 该位点在多种植物SERKs家族中非常保守.大多数LRR-RK中存在Y403的类似位点Tyr-Via, 这一位点的磷酸化修饰同样影响多种受体激酶对配体的识别和信号激活(Macho et al., 2014; Perraki et al., 2018).EFR介导的对丁香假单胞菌的免疫途径依赖于其Tyr-Via位点Y836的磷酸化.丁香假单胞菌分泌具有酪氨酸磷酸酶活性的效应子HopAO1, 可减少EFR的磷酸化并抑制随后的免疫响应(Macho et al., 2014).拟南芥CERK1识别几丁质后, 对下游信号的激活依赖于其第428位的酪氨酸残基的自磷酸化修饰.Chitin信号激活后, CERK1招募预测的丝氨酸/苏氨酸蛋白磷酸酶CIPP1 (CERK1- INTERACTING PROTEIN PHOSPHATASE 1), 对Y428去磷酸化修饰, 关闭CERK1介导的免疫信号, 以防止防御反应被持续激活(Liu et al., 2018).此外, BAK1通过磷酸化修饰BIK1的酪氨酸残基调控免疫信号的激活, BIK1第150、243及250位酪氨酸位点的磷酸化修饰影响BIK1在免疫调控中的功能(Lin et al., 2013, 2014). ...
IBR5 modulates temperature-dependent, R protein CHS3-mediated defense responses in Arabidopsis 1 2015
... 根据胞外凝集素结构域的不同, 凝集素样受体激酶(lectin receptor kinase, LecRK)又分为3种类型: G (GNA-related or S-locus)-、C (calcium-dependent)-以及L (legume)-类(Bouwmeester and Govers, 2009).目前已经鉴定到的L-类LecRK有45个, 它们的功能大部分尚属未知.越来越多的研究表明, L-类LecRK广泛参与植物免疫调控.病原菌或PAMPs诱导LecRK-I.9、LecRK-V.5、LecRK-VI.2以及LecRK- IX.2激活表达; 这些LecRKs功能缺失表现出对不同病原菌的敏感性, 过表达则对病原菌抗性增强(Bou- wmeester et al., 2011; Singh et al., 2012; Wang et al., 2014b; Luo et al., 2017).LecRK-V.5抑制病原菌侵染后的气孔关闭过程; lecrk-V.5缺失突变体对疫霉菌属的抗病性减弱, 对丁香假单胞菌(Pseudomonas syringae pv. tomato (Pst) DC3000)的抗病性增强(Desclos-Theveniau et al., 2012; Wang et al., 2014).L-类LecRK-V.2和LecRK-VII.1结合FLS2, 正调控气孔免疫途径(Yekondi et al., 2018).LecRK- VI.2结合FLS2, 正调控PTI的响应, 参与对丁香假单胞菌和果胶杆菌(Pectobacterium carotovorum)的抗性(Singh et al., 2012; Huang et al., 2014).LecRK- IX.2招募CPKs并磷酸化修饰RBOHD (respiratory burst oxidase homolog protein D), 正调控依赖于RBOHD的水杨酸(salicylic acid, SA)积累以及PTI信号途径(Luo et al., 2017).此外, 有研究表明, 拟南芥Lectin类受体激酶LORE识别细菌脂多糖(lipopoly- saccharides, LPSs)并激活下游免疫信号(Ranf et al., 2015). ...
... ).LecRK- IX.2招募CPKs并磷酸化修饰RBOHD (respiratory burst oxidase homolog protein D), 正调控依赖于RBOHD的水杨酸(salicylic acid, SA)积累以及PTI信号途径(Luo et al., 2017).此外, 有研究表明, 拟南芥Lectin类受体激酶LORE识别细菌脂多糖(lipopoly- saccharides, LPSs)并激活下游免疫信号(Ranf et al., 2015). ...
Proteolysis of a negative regulator of innate immunity is dependent on resistance genes in tomato and Nicotiana benthamiana and induced by mul- tiple bacterial effectors 1 2009
... 拟南芥包含NOI结构域的蛋白RIN4在陆生植物中非常保守, 其并不具有类似已知的蛋白酶类的特征.二级结构预测发现, RIN4蛋白表现出高度的、固有的无序性, 不能形成二级和三级结构, 是植物中固有的无序蛋白(Lee et al., 2015).无序蛋白在与客户蛋白结合时转变成高度折叠状态, 松散并特异地与多种客户蛋白结合(Dyson and Wright, 2005).研究表明, RIN4是AvrRpt2、AvrRpm1、AvrB、HopF2以及AvrPto等多种病原菌效应子蛋白在宿主内攻击的靶标, 表明RIN4在植物-病原微生物互作中具有重要作用(Mackey et al., 2002, 2003; Axtell and Staskawicz, 2003; Luo et al., 2009; Wilton et al., 2010). ...
... 目前已报道的参与免疫调控的受体激酶主要是丝氨酸/苏氨酸类蛋白激酶.近年来, 越来越多的研究表明, 这些受体激酶的酪氨酸残基的磷酸化修饰同样在植物免疫调控中发挥重要作用.例如, BAK1第403位酪氨酸残基的磷酸化修饰对于BAK1的功能以及免疫信号的激活非常重要, 该位点在多种植物SERKs家族中非常保守.大多数LRR-RK中存在Y403的类似位点Tyr-Via, 这一位点的磷酸化修饰同样影响多种受体激酶对配体的识别和信号激活(Macho et al., 2014; Perraki et al., 2018).EFR介导的对丁香假单胞菌的免疫途径依赖于其Tyr-Via位点Y836的磷酸化.丁香假单胞菌分泌具有酪氨酸磷酸酶活性的效应子HopAO1, 可减少EFR的磷酸化并抑制随后的免疫响应(Macho et al., 2014).拟南芥CERK1识别几丁质后, 对下游信号的激活依赖于其第428位的酪氨酸残基的自磷酸化修饰.Chitin信号激活后, CERK1招募预测的丝氨酸/苏氨酸蛋白磷酸酶CIPP1 (CERK1- INTERACTING PROTEIN PHOSPHATASE 1), 对Y428去磷酸化修饰, 关闭CERK1介导的免疫信号, 以防止防御反应被持续激活(Liu et al., 2018).此外, BAK1通过磷酸化修饰BIK1的酪氨酸残基调控免疫信号的激活, BIK1第150、243及250位酪氨酸位点的磷酸化修饰影响BIK1在免疫调控中的功能(Lin et al., 2013, 2014). ...
... ).EFR介导的对丁香假单胞菌的免疫途径依赖于其Tyr-Via位点Y836的磷酸化.丁香假单胞菌分泌具有酪氨酸磷酸酶活性的效应子HopAO1, 可减少EFR的磷酸化并抑制随后的免疫响应(Macho et al., 2014).拟南芥CERK1识别几丁质后, 对下游信号的激活依赖于其第428位的酪氨酸残基的自磷酸化修饰.Chitin信号激活后, CERK1招募预测的丝氨酸/苏氨酸蛋白磷酸酶CIPP1 (CERK1- INTERACTING PROTEIN PHOSPHATASE 1), 对Y428去磷酸化修饰, 关闭CERK1介导的免疫信号, 以防止防御反应被持续激活(Liu et al., 2018).此外, BAK1通过磷酸化修饰BIK1的酪氨酸残基调控免疫信号的激活, BIK1第150、243及250位酪氨酸位点的磷酸化修饰影响BIK1在免疫调控中的功能(Lin et al., 2013, 2014). ...
Arabidopsis RIN4 is a target of the type III virulence effector AvrRpt2 and modulates RPS2-mediated resistance 1 2003
... 拟南芥包含NOI结构域的蛋白RIN4在陆生植物中非常保守, 其并不具有类似已知的蛋白酶类的特征.二级结构预测发现, RIN4蛋白表现出高度的、固有的无序性, 不能形成二级和三级结构, 是植物中固有的无序蛋白(Lee et al., 2015).无序蛋白在与客户蛋白结合时转变成高度折叠状态, 松散并特异地与多种客户蛋白结合(Dyson and Wright, 2005).研究表明, RIN4是AvrRpt2、AvrRpm1、AvrB、HopF2以及AvrPto等多种病原菌效应子蛋白在宿主内攻击的靶标, 表明RIN4在植物-病原微生物互作中具有重要作用(Mackey et al., 2002, 2003; Axtell and Staskawicz, 2003; Luo et al., 2009; Wilton et al., 2010). ...
RIN4 interacts with Pseudomonas syringae type III effector molecules and is required for RPM1-mediated resistance in Arabidopsis 1 2002
... 拟南芥包含NOI结构域的蛋白RIN4在陆生植物中非常保守, 其并不具有类似已知的蛋白酶类的特征.二级结构预测发现, RIN4蛋白表现出高度的、固有的无序性, 不能形成二级和三级结构, 是植物中固有的无序蛋白(Lee et al., 2015).无序蛋白在与客户蛋白结合时转变成高度折叠状态, 松散并特异地与多种客户蛋白结合(Dyson and Wright, 2005).研究表明, RIN4是AvrRpt2、AvrRpm1、AvrB、HopF2以及AvrPto等多种病原菌效应子蛋白在宿主内攻击的靶标, 表明RIN4在植物-病原微生物互作中具有重要作用(Mackey et al., 2002, 2003; Axtell and Staskawicz, 2003; Luo et al., 2009; Wilton et al., 2010). ...
Phosphorylation of a WRKY transcription factor by two pathogen-responsive MAPKs drives phytoalexin bio- synthesis in Arabidopsis 1 2011
... MAPKs磷酸化修饰某些WRKY类转录因子, 调控转录重编程.烟草SIPK和WIPK识别并直接磷酸化I类WRKY转录因子WRKY8, 增强WRKY8与W-box的结合和反式激活活性, 进而激活HMGR2 (3-HYD- ROXY-3-METHYLGLUTARYL CoA REDUCTASE 2)的表达, 产生类异戊二烯植物抗毒素, 促进植物的免疫反应(Ishihama et al., 2011).拟南芥中, 与烟草WRKY8同源的转录因子WRKY33与MPK4以及MKS1在细胞核内形成复合体; 受到flg22或者病原菌刺激时, MPK4磷酸化修饰MKS1并促使WRKY33- MKS1与MPK4分离, 磷酸化的WRKY33随后结合植物抗毒素(camalexin)合成相关基因PAD3启动子区的W-box, 激活PAD3的表达并促进camalexin的合成(Andreasson et al., 2005; Qiu et al., 2008; Len- zoni et al., 2018).WRKY33还参与MPK3/6调控的camalexin的合成过程.灰霉菌诱导时, MPK3/6磷酸化修饰WRKY33, 促进相关基因转录重编程(Mao et al., 2011; Li et al., 2012).MPK6结合并磷酸化ERF104调控防御相关基因的表达(Bethke et al., 2009).拟南芥中ASR3作为转录抑制因子负调控PAMP诱导的免疫响应.PAMP处理后MPK4磷酸化ASR3第189位苏氨酸, 磷酸化的ASR3与DNA结合能力增强, 从而抑制flg22诱导的防御基因的表达(Li et al., 2015). ...
Phosphorylation of an ERF trans- cription factor by Arabidopsis MPK3/MPK6 regulates plant defense gene induction and fungal resistance 1 2013
... MAPKs对底物蛋白的磷酸化修饰影响底物蛋白的稳定性.拟南芥中, MPK3/MPK6通过磷酸化修饰ACS2/ACS6调控乙烯产生.泛素蛋白酶降解复合体结合未被磷酸化修饰的ACS6的C末端非催化区域, 导致ACS6经泛素化途径降解.MPK3/MPK6对ACS6的磷酸化修饰在其C末端引入了负电荷, 抑制ACS6的泛素化降解, 增强其蛋白的稳定性, 进而促进乙烯生成(Joo et al., 2008; Han et al., 2010).MPK3/ MPK6磷酸化修饰ERF家族成员ERF6, 增强其蛋白稳定性, 激活PDF1.1 (plant defensin 1.1)、PDF- 1.2a、PDF1.2b和ChiB等多种防御相关基因的表达(Meng et al., 2013).受到PAMP诱导后, MPK3磷酸化修饰WRKY46的S168和S250位点, 促进WRKY46的降解(Sheikh et al., 2016).LIP5参与病原菌诱导的内吞作用和囊泡运输, MPK3和MPK6磷酸化修饰LIP5, 增强LIP5的蛋白稳定性, 从而正调控多泡体(multivesicular bodies, MVB)通路, 影响免疫应答过程中相关因子的重定位(Wang et al., 2014a).E3泛素连接酶PUB22是拟南芥中PTI响应的抑制因子.MPK3对PUB22的磷酸化修饰抑制其自泛素化, 从而导致PUB22积累, 抑制免疫信号激活(Furlan et al., 2017). ...
MAPK cascades in plant disease resistance signaling 2 2013
Phosphoproteomics of the Arabidopsis plasma membrane and a new phosphorylation site database 1 2004
... RIN4蛋白不同位点的磷酸化修饰具有不同的功能(Nühse et al., 2007; Chung et al., 2014).有研究表明, RIN4第47和141位的丝氨酸是受flg22诱导后被磷酸化修饰的可能位点(Nühse et al., 2004, 2007).Chung等(2014)发现, RIN4第141位的丝氨酸残基的磷酸化修饰影响flg22诱导对病原菌生长的抑制, 导致胼胝质沉积、ROS爆发以及防御基因表达上调.当RIN4第141位丝氨酸突变为不能被磷酸化修饰的丙氨酸时, flg22诱导的PTI响应受到抑制.RIN4蛋白第166位的苏氨酸残基的磷酸化修饰, 发挥与S141位点磷酸化修饰相反的作用.拟南芥的胞质内类受体激酶RIPK磷酸化修饰RIN4蛋白的T21、S160和T166位点.T166位点被磷酸化修饰后, PTI途径的激活受到抑制.与S141位点磷酸化相比, T166位点被磷酸化修饰后发挥的功能具有上位性. ...
Effector-triggered immunity mediated by the Pto kinase 1 2011
... 通过对烟草(Nicotiana benthamiana)和番茄的研究发现, MAPKs除在PTI途径中发挥功能外, 也是ETI途径的组成元件.烟草和番茄中丝/苏氨酸蛋白激酶Pto与丁香假单胞菌分泌的效应子AvrPto/AvrPtoB相互作用, 激活抗病蛋白Prf及ETI响应.此过程中, MAPKKKα和MAPKKKε正调控Pto-介导的ETI途径(del Pozo et al., 2004; Oh and Martin, 2011).在表达抗病蛋白Cf-9的转基因烟草中, 真菌效应子Avr9激活烟草中的MAPK激酶SIPK (SALICYLIC ACID- INDUCED PROTEIN KINASE)和WIPK (WOUND- INDUCED PROTEIN KINASE), 并导致WIPK蛋白积累(Romeis et al., 1999; Pedley and Martin, 2005; Meng and Zhang, 2013).烟草花叶病毒(tobacco mosaic virus, TMV)在烟草抗病蛋白N的作用下磷酸化SIPK和WIPK, 并增加WIPK的mRNA丰度以及蛋白含量(Zhang and Klessig, 1998).此外, AvrRpt2对拟南芥MPK3和MPK6也有激活作用, 与野生型Pst DC3000相比, 携带AvrRpt2效应蛋白的Pst DC3000对MPK3和MPK6的激活能力增强(Eschen-Lippold et al., 2016). ...
Rice XB15, a protein phosphatase 2C, negatively regulates cell death and XA21-mediated innate immunity 1 2008
... 免疫受体和共受体的活性受到蛋白激酶和蛋白磷酸酶的调控(图2).无病原菌时, 拟南芥LRR-RLK BIR2 (BAK1-INTERACTING RECEPTOR-LIKE KINASE 2)结合BAK1, 防止其与FLS2等PRRs结合而导致PTI途径不必要的激活(Halter et al., 2014).PP2A类蛋白磷酸酶负调控PRR复合体的激活.无PAMP诱导时, 由A1、C4以及B’η/ζ亚基组成的PP2A全酶结合BAK1防止其被磷酸化激活; 配体诱导后, PP2A全酶复合体的活性迅速降低, 导致BAK1被磷酸化激活(Seg-onzac et al., 2014).拟南芥质膜定位的磷蛋白MKKK7结合FLS2, 受鞭毛蛋白诱导时, MKKK7的多个丝氨酸残基被不同程度地磷酸化修饰, 负调控鞭毛蛋白激活的信号以及基础免疫响应.MKKK7通过 直接调控FLS2复合体, 抑制FLS2介导的ROS产生(Mithoe et al., 2016).此外, 水稻PP2C类蛋白磷酸酶XB15 (XA21 BINDING PROTEIN 15)结合并去磷酸化修饰XA21, 负调控XA21介导的免疫激活(Park et al., 2008). ...
Role of mitogen-activated protein kinases in plant immunity 1 2005
... 通过对烟草(Nicotiana benthamiana)和番茄的研究发现, MAPKs除在PTI途径中发挥功能外, 也是ETI途径的组成元件.烟草和番茄中丝/苏氨酸蛋白激酶Pto与丁香假单胞菌分泌的效应子AvrPto/AvrPtoB相互作用, 激活抗病蛋白Prf及ETI响应.此过程中, MAPKKKα和MAPKKKε正调控Pto-介导的ETI途径(del Pozo et al., 2004; Oh and Martin, 2011).在表达抗病蛋白Cf-9的转基因烟草中, 真菌效应子Avr9激活烟草中的MAPK激酶SIPK (SALICYLIC ACID- INDUCED PROTEIN KINASE)和WIPK (WOUND- INDUCED PROTEIN KINASE), 并导致WIPK蛋白积累(Romeis et al., 1999; Pedley and Martin, 2005; Meng and Zhang, 2013).烟草花叶病毒(tobacco mosaic virus, TMV)在烟草抗病蛋白N的作用下磷酸化SIPK和WIPK, 并增加WIPK的mRNA丰度以及蛋白含量(Zhang and Klessig, 1998).此外, AvrRpt2对拟南芥MPK3和MPK6也有激活作用, 与野生型Pst DC3000相比, 携带AvrRpt2效应蛋白的Pst DC3000对MPK3和MPK6的激活能力增强(Eschen-Lippold et al., 2016). ...
Phosphocode-dependent functional dichotomy of a common co-receptor in plant signaling 1 2018
... 目前已报道的参与免疫调控的受体激酶主要是丝氨酸/苏氨酸类蛋白激酶.近年来, 越来越多的研究表明, 这些受体激酶的酪氨酸残基的磷酸化修饰同样在植物免疫调控中发挥重要作用.例如, BAK1第403位酪氨酸残基的磷酸化修饰对于BAK1的功能以及免疫信号的激活非常重要, 该位点在多种植物SERKs家族中非常保守.大多数LRR-RK中存在Y403的类似位点Tyr-Via, 这一位点的磷酸化修饰同样影响多种受体激酶对配体的识别和信号激活(Macho et al., 2014; Perraki et al., 2018).EFR介导的对丁香假单胞菌的免疫途径依赖于其Tyr-Via位点Y836的磷酸化.丁香假单胞菌分泌具有酪氨酸磷酸酶活性的效应子HopAO1, 可减少EFR的磷酸化并抑制随后的免疫响应(Macho et al., 2014).拟南芥CERK1识别几丁质后, 对下游信号的激活依赖于其第428位的酪氨酸残基的自磷酸化修饰.Chitin信号激活后, CERK1招募预测的丝氨酸/苏氨酸蛋白磷酸酶CIPP1 (CERK1- INTERACTING PROTEIN PHOSPHATASE 1), 对Y428去磷酸化修饰, 关闭CERK1介导的免疫信号, 以防止防御反应被持续激活(Liu et al., 2018).此外, BAK1通过磷酸化修饰BIK1的酪氨酸残基调控免疫信号的激活, BIK1第150、243及250位酪氨酸位点的磷酸化修饰影响BIK1在免疫调控中的功能(Lin et al., 2013, 2014). ...
The rice immune receptor XA21 recognizes a tyrosine-sulfated protein from a Gramne- gative bacterium 1 2015
... LRR类PRRs主要识别蛋白或多肽序列(图1).例如, LRR类受体激酶FLS2 (FLAGELLIN SENSING 2)和EFR (ELONGATION FACTOR-Tu RECEPTOR)分别识别并结合病原菌鞭毛蛋白保守的22个氨基酸残基组成的肽flg22, 以及延伸因子Tu (EF-Tu) N末端的保守氨基酸序列elf18 (Chinchilla et al., 2006; Zipfel et al., 2006); PEPR1/2 (PEP RECEPTOR)识别植物响应病原菌分泌的内源可溶性前体肽pep (pro- peptide) (Yamaguchi et al., 2006); 番茄(Solanum lycopersicum)中FLS3识别细菌flgII-28并增强对细菌的免疫响应(Hind et al., 2016); 水稻(Oryza sativa) LRR-类受体激酶XA21识别水稻白叶枯菌(Xantho- monas oryzae pv. oryzae, Xoo)中保守的蛋白RaxX等(Pruitt et al., 2015). ...
Arabidopsis MAP kinase 4 regulates gene ex- pression through transcription factor release in the nu- cleus 1 2008
... MAPKs磷酸化修饰某些WRKY类转录因子, 调控转录重编程.烟草SIPK和WIPK识别并直接磷酸化I类WRKY转录因子WRKY8, 增强WRKY8与W-box的结合和反式激活活性, 进而激活HMGR2 (3-HYD- ROXY-3-METHYLGLUTARYL CoA REDUCTASE 2)的表达, 产生类异戊二烯植物抗毒素, 促进植物的免疫反应(Ishihama et al., 2011).拟南芥中, 与烟草WRKY8同源的转录因子WRKY33与MPK4以及MKS1在细胞核内形成复合体; 受到flg22或者病原菌刺激时, MPK4磷酸化修饰MKS1并促使WRKY33- MKS1与MPK4分离, 磷酸化的WRKY33随后结合植物抗毒素(camalexin)合成相关基因PAD3启动子区的W-box, 激活PAD3的表达并促进camalexin的合成(Andreasson et al., 2005; Qiu et al., 2008; Len- zoni et al., 2018).WRKY33还参与MPK3/6调控的camalexin的合成过程.灰霉菌诱导时, MPK3/6磷酸化修饰WRKY33, 促进相关基因转录重编程(Mao et al., 2011; Li et al., 2012).MPK6结合并磷酸化ERF104调控防御相关基因的表达(Bethke et al., 2009).拟南芥中ASR3作为转录抑制因子负调控PAMP诱导的免疫响应.PAMP处理后MPK4磷酸化ASR3第189位苏氨酸, 磷酸化的ASR3与DNA结合能力增强, 从而抑制flg22诱导的防御基因的表达(Li et al., 2015). ...
A lectin S-domain receptor kinase mediates lipopolysaccharide sensing in Arabidopsis thaliana 1 2015
... 根据胞外凝集素结构域的不同, 凝集素样受体激酶(lectin receptor kinase, LecRK)又分为3种类型: G (GNA-related or S-locus)-、C (calcium-dependent)-以及L (legume)-类(Bouwmeester and Govers, 2009).目前已经鉴定到的L-类LecRK有45个, 它们的功能大部分尚属未知.越来越多的研究表明, L-类LecRK广泛参与植物免疫调控.病原菌或PAMPs诱导LecRK-I.9、LecRK-V.5、LecRK-VI.2以及LecRK- IX.2激活表达; 这些LecRKs功能缺失表现出对不同病原菌的敏感性, 过表达则对病原菌抗性增强(Bou- wmeester et al., 2011; Singh et al., 2012; Wang et al., 2014b; Luo et al., 2017).LecRK-V.5抑制病原菌侵染后的气孔关闭过程; lecrk-V.5缺失突变体对疫霉菌属的抗病性减弱, 对丁香假单胞菌(Pseudomonas syringae pv. tomato (Pst) DC3000)的抗病性增强(Desclos-Theveniau et al., 2012; Wang et al., 2014).L-类LecRK-V.2和LecRK-VII.1结合FLS2, 正调控气孔免疫途径(Yekondi et al., 2018).LecRK- VI.2结合FLS2, 正调控PTI的响应, 参与对丁香假单胞菌和果胶杆菌(Pectobacterium carotovorum)的抗性(Singh et al., 2012; Huang et al., 2014).LecRK- IX.2招募CPKs并磷酸化修饰RBOHD (respiratory burst oxidase homolog protein D), 正调控依赖于RBOHD的水杨酸(salicylic acid, SA)积累以及PTI信号途径(Luo et al., 2017).此外, 有研究表明, 拟南芥Lectin类受体激酶LORE识别细菌脂多糖(lipopoly- saccharides, LPSs)并激活下游免疫信号(Ranf et al., 2015). ...
Rapid Avr9- and Cf-9-dependent activation of MAP kinases in tobacco cell cultures and leaves: con- vergence of resistance gene, elicitor, wound, and salicy- late responses 1 1999
... 通过对烟草(Nicotiana benthamiana)和番茄的研究发现, MAPKs除在PTI途径中发挥功能外, 也是ETI途径的组成元件.烟草和番茄中丝/苏氨酸蛋白激酶Pto与丁香假单胞菌分泌的效应子AvrPto/AvrPtoB相互作用, 激活抗病蛋白Prf及ETI响应.此过程中, MAPKKKα和MAPKKKε正调控Pto-介导的ETI途径(del Pozo et al., 2004; Oh and Martin, 2011).在表达抗病蛋白Cf-9的转基因烟草中, 真菌效应子Avr9激活烟草中的MAPK激酶SIPK (SALICYLIC ACID- INDUCED PROTEIN KINASE)和WIPK (WOUND- INDUCED PROTEIN KINASE), 并导致WIPK蛋白积累(Romeis et al., 1999; Pedley and Martin, 2005; Meng and Zhang, 2013).烟草花叶病毒(tobacco mosaic virus, TMV)在烟草抗病蛋白N的作用下磷酸化SIPK和WIPK, 并增加WIPK的mRNA丰度以及蛋白含量(Zhang and Klessig, 1998).此外, AvrRpt2对拟南芥MPK3和MPK6也有激活作用, 与野生型Pst DC3000相比, 携带AvrRpt2效应蛋白的Pst DC3000对MPK3和MPK6的激活能力增强(Eschen-Lippold et al., 2016). ...
The mRNA decay factor PAT1 functions in a pathway including MAP kinase 4 and immune receptor SUMM2 1 2015
... R蛋白监控MAPKs对底物蛋白的磷酸化修饰(图2).多种病原菌分泌效应蛋白攻击宿主细胞内MAPKs信号级联组成元件, 抑制MAPKs调控的防御反应(Zhang et al., 2007, 2012; Wang et al., 2010).拟南芥抗病蛋白SUMM2 (SUPPRESSOR OF MKK1 MKK2 2)在MAPKs信号的下游, 监控MPK4底物PAT1 (PROTEIN ASSOCIATED WITH TOPOISOMERASE II)、CRCK3 (CALMODULIN-BINDGIN RECEPTOR LIKE CYTOPLASMIC KINASE 3)以及MEKK2的磷酸化状态, 从而调控免疫应答.拟南芥PAT1编码mRNA脱帽装置的组成元件, 与MPK4结合形成复合体.PAMP诱导后, PAT1被MPK4磷酸化并在细胞溶质中聚集成离散的点状物.pat1缺失突变体表现出依赖于SUMM2的免疫自激活(Roux et al., 2015).CRCK3编码钙离子结合的胞质内受体激酶,结合MPK4并被MPK4磷酸化修饰.CRCK3缺失可恢复mekk1、mkk1/mkk2双突变体以及mpk4中SUMM2介导的免疫自激活.此外, CRCK3和PAT1均可以与SUMM2相互作用, 表明SUMM2可能通过监控CR- CK3和PAT1的磷酸化状态, 从而确定MEKK1- MKK1/MKK2-MPK4信号通路的完整性, 并适时启动免疫应答(Bi and Zhou, 2017; Zhang et al., 2017). ...
Posttranslational modifications of the master transcrip- tional regulator NPR1 enable dynamic but tight control of plant immune responses 1 2015
... 拟南芥水杨酸信号通路中, NPR1 (NONEXPRESSOR OF PATHOGENESIS-RELAED GENE 1)作为转录共激活因子调控多种免疫相关基因的表达.其蛋白稳定性受到泛素蛋白酶体途径的调控, 动态地调节免疫相关基因的转录激活或抑制.研究表明, NPR1不同位点的磷酸化修饰对其泛素化产生不同的影响.S11和S15位点的磷酸化修饰促进NPR1与SUMO3的结合, 从而促进NPR1的降解.而NPR1的S55和S59位点的磷酸化修饰则抑制NPR1的SUMO类泛素化, 增强NPR1蛋白的稳定性(Spoel et al., 2009; Saleh et al., 2015).此外, SnRK (SNF1-RELATED PRO- TEIN KINASE)激酶PSK5通过磷酸化修饰NPR1调控转录因子WRKY38和WRKY62的表达(Xie et al., 2010). ...
The PP2C-type phosphatase AP2C1, which negatively regulates MPK4 and MPK6, modulates innate immunity, jasmonic acid, and ethylene levels in Arabidopsis 1 2007
... MAPK的持续磷酸化会导致免疫反应被持续激活, 因此MAPK的活性需要受到严格控制.目前已报道多种磷酸酶在植物免疫过程中负调控MAPK的激活.这些磷酸酶主要包括蛋白酪氨酸磷酸酶(PTPs)、双特异性磷酸酶(DSPs)以及PP2C类蛋白磷酸酶.不同的磷酸酶对MAPK的去磷酸化修饰导致不同的下游信号事件.拟南芥蛋白酪氨酸磷酸酶PTP1 (PROTEIN TYROSINE PHOSPHATASE 1)以及双特异性磷酸酶MKP1 (MAP KINASE PHOSPHATASE 1)和MKP2参与对MPK3/MPK6活性的调控.MKP1通过负调控MPK6途径抑制PAMP激活的免疫应答(Anderson et al., 2011); PTP1结合MPK6与MKP1, 协同抑制MPK6调控的防御反应激活.MKP2与MPK3/MPK6结合并且可在体外去磷酸化修饰MAK3和MPK6 (Lee and Ellis, 2007).真菌激发子诱导的过程中, MKP2抑制MPK6介导的超敏反应(hypersen- sitive response, HR), 从而调控氧化胁迫以及对青枯病病原菌(Ralstonia solanacearum)的抗性(Lumb- reras et al., 2010).此外, MKP1的活性也受到MPK6的调控.PAMP诱导后MKP1蛋白迅速积累, 而这一过程依赖于MPK6对MKP1的磷酸化修饰(Jiang et al., 2017).拟南芥PP2C类蛋白磷酸酶(AP2Cs蛋白磷酸酶)通过激酶结合基序(kinase interaction motif, KIM)与MAPK相互作用(Fuchs et al., 2013).AP2C1负调控植物免疫应答过程中MAPK信号的激活, 过表达AP2C1抑制损伤诱导的乙烯产生以及对死体营养型真菌灰霉菌的抗性.AP2C1功能缺失增强PAMP诱导的MAPK激活、胼胝质积累以及对细菌的抗病性(Schweighofer et al., 2007; Galletti et al., 2011; Sidonskaya et al., 2016; Shubchynskyy et al., 2017). ...
2
... 模式植物拟南芥(Arabidopsis thaliana)的基因组编码大约1 100种蛋白激酶以及约150种蛋白磷酸酶(Schweighofer and Meskiene, 2015).根据磷酸化修饰的氨基酸残基不同, 蛋白激酶主要分为两大类, 即丝氨酸/苏氨酸蛋白激酶和酪氨酸蛋白激酶.此外, 有些激酶同时具有丝/苏氨酸蛋白激酶活性和酪氨酸蛋白激酶活性.目前, 在植物免疫途径中研究相对较多的主要有4种激酶, 即跨膜的受体激酶(receptor like kinase, RLK)、胞质内受体激酶(receptor-like cytoplasmic kinase, RLCK)、丝裂原活化激酶(mitogen- activated protein kinase, MAPK)以及钙依赖蛋白激酶(calcium-dependent protein kinase, CPK).根据结构、催化方式以及底物特异性的不同蛋白磷酸酶分为3种类型, 即蛋白丝氨酸/苏氨酸磷酸酶、蛋白酪氨酸磷酸酶以及双特异性磷酸酶(Luan, 2003).在拟南芥中, 约96%的蛋白磷酸化作用发生在丝氨酸/苏氨酸残基上.催化丝氨酸/苏氨酸残基去磷酸化的磷酸酶包括2个家族, 即丝氨酸/苏氨酸蛋白磷酸酶家族(pho- sphoprotein phosphatase, PPP)和金属离子依赖的蛋白磷酸酶(metal-dependent protein phosphatases, PPM).PPP类蛋白磷酸酶的催化亚基结合不同的调节亚基组成的全酶具有不同的底物特异性.PPM家族蛋白磷酸酶主要由PP2C组成, 其本身同时具有催化活性和调节功能, 不需要单独的调节亚基进行调控(Schweighofer and Meskiene, 2015). ...
... ).在拟南芥中, 约96%的蛋白磷酸化作用发生在丝氨酸/苏氨酸残基上.催化丝氨酸/苏氨酸残基去磷酸化的磷酸酶包括2个家族, 即丝氨酸/苏氨酸蛋白磷酸酶家族(pho- sphoprotein phosphatase, PPP)和金属离子依赖的蛋白磷酸酶(metal-dependent protein phosphatases, PPM).PPP类蛋白磷酸酶的催化亚基结合不同的调节亚基组成的全酶具有不同的底物特异性.PPM家族蛋白磷酸酶主要由PP2C组成, 其本身同时具有催化活性和调节功能, 不需要单独的调节亚基进行调控(Schweighofer and Meskiene, 2015). ...
Negative control of BAK1 by protein phosphatase 2A during plant innate immunity 1 2014
... 免疫受体和共受体的活性受到蛋白激酶和蛋白磷酸酶的调控(图2).无病原菌时, 拟南芥LRR-RLK BIR2 (BAK1-INTERACTING RECEPTOR-LIKE KINASE 2)结合BAK1, 防止其与FLS2等PRRs结合而导致PTI途径不必要的激活(Halter et al., 2014).PP2A类蛋白磷酸酶负调控PRR复合体的激活.无PAMP诱导时, 由A1、C4以及B’η/ζ亚基组成的PP2A全酶结合BAK1防止其被磷酸化激活; 配体诱导后, PP2A全酶复合体的活性迅速降低, 导致BAK1被磷酸化激活(Seg-onzac et al., 2014).拟南芥质膜定位的磷蛋白MKKK7结合FLS2, 受鞭毛蛋白诱导时, MKKK7的多个丝氨酸残基被不同程度地磷酸化修饰, 负调控鞭毛蛋白激活的信号以及基础免疫响应.MKKK7通过 直接调控FLS2复合体, 抑制FLS2介导的ROS产生(Mithoe et al., 2016).此外, 水稻PP2C类蛋白磷酸酶XB15 (XA21 BINDING PROTEIN 15)结合并去磷酸化修饰XA21, 负调控XA21介导的免疫激活(Park et al., 2008). ...
Regulation of WRK- Y46 transcription factor function by mitogen-activated pro- tein kinases in Arabidopsis thaliana 2 2016
... MAPKs对底物蛋白的磷酸化修饰影响底物蛋白的稳定性.拟南芥中, MPK3/MPK6通过磷酸化修饰ACS2/ACS6调控乙烯产生.泛素蛋白酶降解复合体结合未被磷酸化修饰的ACS6的C末端非催化区域, 导致ACS6经泛素化途径降解.MPK3/MPK6对ACS6的磷酸化修饰在其C末端引入了负电荷, 抑制ACS6的泛素化降解, 增强其蛋白的稳定性, 进而促进乙烯生成(Joo et al., 2008; Han et al., 2010).MPK3/ MPK6磷酸化修饰ERF家族成员ERF6, 增强其蛋白稳定性, 激活PDF1.1 (plant defensin 1.1)、PDF- 1.2a、PDF1.2b和ChiB等多种防御相关基因的表达(Meng et al., 2013).受到PAMP诱导后, MPK3磷酸化修饰WRKY46的S168和S250位点, 促进WRKY46的降解(Sheikh et al., 2016).LIP5参与病原菌诱导的内吞作用和囊泡运输, MPK3和MPK6磷酸化修饰LIP5, 增强LIP5的蛋白稳定性, 从而正调控多泡体(multivesicular bodies, MVB)通路, 影响免疫应答过程中相关因子的重定位(Wang et al., 2014a).E3泛素连接酶PUB22是拟南芥中PTI响应的抑制因子.MPK3对PUB22的磷酸化修饰抑制其自泛素化, 从而导致PUB22积累, 抑制免疫信号激活(Furlan et al., 2017). ...
BR-SIGNALING KINASE 1 physically associates with FLAGELLIN SENSING 2 and regulates plant innate immunity in Arabidopsis 1 2013
... RLCK经PRRs激活后, 一方面招募钙离子依赖的蛋白激酶(CPKs)促进产生活性氧(ROS); 另一方面激活下游MAPK信号级联反应(图2).植物中ROS主要由NADPH氧化酶RBOHD催化生成.目前已报道多种RLCKs调控RBOHD的活性.例如, 被flg22诱导激活的BIK1一方面通过磷酸化修饰RBOHD的多个氨基酸位点激活RBOHD的活性; 另一方面通过磷酸化植物Gα蛋白XLG2 (EXTRA LARGE G-PROTEIN 2)促进ROS的产生(Kadota et al., 2014; Li et al., 2014b; Liang et al., 2016).GTP酶促进蛋白(RGS1)维持FLS2结合的G蛋白不活跃状态.被?g22诱导激活的BIK1/PBL1磷酸化RGS1的S428和S431位点, 促进RGS1与FLS2-G蛋白复合体分离, 从而正调控免疫信号的激活(Liang et al., 2018).BSK1结合FLS2调控ROS的生成.bsk1缺失突变体中, flg22诱导的ROS爆发减弱, 病原菌诱导的SA积累受到阻碍并且对白粉菌、丁香假单胞菌和卵菌的敏感性增强(Shi et al., 2013).在无病原菌侵染时, PBL13通过结合RBOHD负调控植物对致病菌的先天免疫途径(Lin et al., 2015). ...
Chitin- mediated plant-fungal interactions: catching, hiding and handshaking 1 2015
... LysM类和EGF类PRRs主要识别包含碳水化合物的分子, 如真菌几丁质(chitin)、细菌肽聚糖(peptidog- lycan, PGN)和丛枝菌根菌分泌的脂质几丁寡糖(lipo- chitooligosaccharides, LCOs)或者植物细胞壁衍生的寡聚半乳糖醛酸(oligogalacturonides, OGs)等(图1).拟南芥中的3个LysM类受体激酶CERK1 (CHITIN ELICITOR RECEPTOR KINASE 1)、LYK4 (LysM- CONTAINING RECEPTOR-LIKE KINASE 4)和LYK5共同调控几丁质的识别及免疫信号的激活(Miya et al., 2007; Wan et al., 2012; Cao et al., 2014); LYK1和LYK3参与对不同结构的PGN信号的识别, 在AtCERK1存在的条件下激活PGN诱导的防御信号(Willmann et al., 2011).水稻LysM类RLK OsCERK1胞外的LysM结构域不能直接结合几丁质, LysM类受体蛋白OsCEBiP (CHITIN OLIFOSACCHARIDE ELICITOR BINDING PROTEIN)作为受体结合几丁质, 随后结合OsCERK1激活几丁质诱导的防御反应(Kaku et al., 2006; Shinya et al., 2015).拟南芥EGF类受体激酶WAK1 (WALL-ASSOCIATED KINASE 1)接受OGs信号并激活防御反应(Brutus et al., 2010).水稻免疫信号激活过程中, OsWAK1发生自磷酸化并磷酸化下游转录因子OsRFP1.此外, 水稻稻瘟菌侵染、机械损伤及水杨酸和茉莉酸甲酯处理均可以激活OsWAK1转录水平的表达(Li et al., 2009). ...
Selective regu- lation of the chitin-induced defense response by the Arabi- dopsis receptor-like cytoplasmic kinase PBL27 1 2014
... 胞质内的受体激酶结合RLK/RLP, 共同调控植物的生长发育、激素信号以及生物和非生物胁迫等过程.研究表明, 拟南芥RLCK-VII家族的PBS1 (AvrPphB SUSCEPTIBLE 1)、BIK1 (BOTRYTIS-INDUCED KINASE 1)、PBL1 (PBS1- LIKE 1)、PBL2、PBL27、PCRKs (PTI COMPROMISED RECEPTOR-LIKE CYTOPLASMIC KINASE)以及RLCK-XII家族成员BSK1 (BRASSINOSTEROID-SIGNALING KINASE 1)参与植物免疫信号传递(图1).例如, FLS2、EFR、PEPRs以及CERK1等多种PRR被激活后磷酸化修饰BIK1, 导致BIK1与PRRs复合体分离并进入细胞质激活下游免疫信号(Lu et al., 2010; Yang et al., 2010; Liu et al., 2013).PCRK1和PCRK2结合FLS2并在PAMPs诱导后被磷酸化激活, 冗余地诱导转录因子SARD1 (SYSTEMIC ACQUIRED RESISTANCE DEFICIENT 1)和CBP60g (CALMODULIN-BINFING PROTEIN 60g)的表达, 促进SA的合成(Kong et al., 2016).水稻RLCK-VII家族的OsRLCK185和OsRL- CK176结合CERK1, 传递PGN和几丁质激活的免疫信号(Yamaguchi et al., 2013; Ao et al., 2014).Os- RLCK185在拟南芥中的同源蛋白PBL27被CERK1磷酸化激活后, 正调控几丁质诱导的防御反应(Shinya et al., 2014). ...
Protein phosphatase AP2C1 negatively regulates basal resistance and defense responses to Pseudomonas syringae 1 2017
... MAPK的持续磷酸化会导致免疫反应被持续激活, 因此MAPK的活性需要受到严格控制.目前已报道多种磷酸酶在植物免疫过程中负调控MAPK的激活.这些磷酸酶主要包括蛋白酪氨酸磷酸酶(PTPs)、双特异性磷酸酶(DSPs)以及PP2C类蛋白磷酸酶.不同的磷酸酶对MAPK的去磷酸化修饰导致不同的下游信号事件.拟南芥蛋白酪氨酸磷酸酶PTP1 (PROTEIN TYROSINE PHOSPHATASE 1)以及双特异性磷酸酶MKP1 (MAP KINASE PHOSPHATASE 1)和MKP2参与对MPK3/MPK6活性的调控.MKP1通过负调控MPK6途径抑制PAMP激活的免疫应答(Anderson et al., 2011); PTP1结合MPK6与MKP1, 协同抑制MPK6调控的防御反应激活.MKP2与MPK3/MPK6结合并且可在体外去磷酸化修饰MAK3和MPK6 (Lee and Ellis, 2007).真菌激发子诱导的过程中, MKP2抑制MPK6介导的超敏反应(hypersen- sitive response, HR), 从而调控氧化胁迫以及对青枯病病原菌(Ralstonia solanacearum)的抗性(Lumb- reras et al., 2010).此外, MKP1的活性也受到MPK6的调控.PAMP诱导后MKP1蛋白迅速积累, 而这一过程依赖于MPK6对MKP1的磷酸化修饰(Jiang et al., 2017).拟南芥PP2C类蛋白磷酸酶(AP2Cs蛋白磷酸酶)通过激酶结合基序(kinase interaction motif, KIM)与MAPK相互作用(Fuchs et al., 2013).AP2C1负调控植物免疫应答过程中MAPK信号的激活, 过表达AP2C1抑制损伤诱导的乙烯产生以及对死体营养型真菌灰霉菌的抗性.AP2C1功能缺失增强PAMP诱导的MAPK激活、胼胝质积累以及对细菌的抗病性(Schweighofer et al., 2007; Galletti et al., 2011; Sidonskaya et al., 2016; Shubchynskyy et al., 2017). ...
Plant resistance against the parasitic nematode Heterodera schachtii is mediated by MPK3 and MPK6 kinases, which are controlled by the MAPK phosphatase AP2C1 in Arabidopsis 1 2016
... MAPK的持续磷酸化会导致免疫反应被持续激活, 因此MAPK的活性需要受到严格控制.目前已报道多种磷酸酶在植物免疫过程中负调控MAPK的激活.这些磷酸酶主要包括蛋白酪氨酸磷酸酶(PTPs)、双特异性磷酸酶(DSPs)以及PP2C类蛋白磷酸酶.不同的磷酸酶对MAPK的去磷酸化修饰导致不同的下游信号事件.拟南芥蛋白酪氨酸磷酸酶PTP1 (PROTEIN TYROSINE PHOSPHATASE 1)以及双特异性磷酸酶MKP1 (MAP KINASE PHOSPHATASE 1)和MKP2参与对MPK3/MPK6活性的调控.MKP1通过负调控MPK6途径抑制PAMP激活的免疫应答(Anderson et al., 2011); PTP1结合MPK6与MKP1, 协同抑制MPK6调控的防御反应激活.MKP2与MPK3/MPK6结合并且可在体外去磷酸化修饰MAK3和MPK6 (Lee and Ellis, 2007).真菌激发子诱导的过程中, MKP2抑制MPK6介导的超敏反应(hypersen- sitive response, HR), 从而调控氧化胁迫以及对青枯病病原菌(Ralstonia solanacearum)的抗性(Lumb- reras et al., 2010).此外, MKP1的活性也受到MPK6的调控.PAMP诱导后MKP1蛋白迅速积累, 而这一过程依赖于MPK6对MKP1的磷酸化修饰(Jiang et al., 2017).拟南芥PP2C类蛋白磷酸酶(AP2Cs蛋白磷酸酶)通过激酶结合基序(kinase interaction motif, KIM)与MAPK相互作用(Fuchs et al., 2013).AP2C1负调控植物免疫应答过程中MAPK信号的激活, 过表达AP2C1抑制损伤诱导的乙烯产生以及对死体营养型真菌灰霉菌的抗性.AP2C1功能缺失增强PAMP诱导的MAPK激活、胼胝质积累以及对细菌的抗病性(Schweighofer et al., 2007; Galletti et al., 2011; Sidonskaya et al., 2016; Shubchynskyy et al., 2017). ...
The lectin receptor kinase-VI.2 is required for priming and positively regulates Arabidopsis pattern-triggered immunity 2 2012
... 根据胞外凝集素结构域的不同, 凝集素样受体激酶(lectin receptor kinase, LecRK)又分为3种类型: G (GNA-related or S-locus)-、C (calcium-dependent)-以及L (legume)-类(Bouwmeester and Govers, 2009).目前已经鉴定到的L-类LecRK有45个, 它们的功能大部分尚属未知.越来越多的研究表明, L-类LecRK广泛参与植物免疫调控.病原菌或PAMPs诱导LecRK-I.9、LecRK-V.5、LecRK-VI.2以及LecRK- IX.2激活表达; 这些LecRKs功能缺失表现出对不同病原菌的敏感性, 过表达则对病原菌抗性增强(Bou- wmeester et al., 2011; Singh et al., 2012; Wang et al., 2014b; Luo et al., 2017).LecRK-V.5抑制病原菌侵染后的气孔关闭过程; lecrk-V.5缺失突变体对疫霉菌属的抗病性减弱, 对丁香假单胞菌(Pseudomonas syringae pv. tomato (Pst) DC3000)的抗病性增强(Desclos-Theveniau et al., 2012; Wang et al., 2014).L-类LecRK-V.2和LecRK-VII.1结合FLS2, 正调控气孔免疫途径(Yekondi et al., 2018).LecRK- VI.2结合FLS2, 正调控PTI的响应, 参与对丁香假单胞菌和果胶杆菌(Pectobacterium carotovorum)的抗性(Singh et al., 2012; Huang et al., 2014).LecRK- IX.2招募CPKs并磷酸化修饰RBOHD (respiratory burst oxidase homolog protein D), 正调控依赖于RBOHD的水杨酸(salicylic acid, SA)积累以及PTI信号途径(Luo et al., 2017).此外, 有研究表明, 拟南芥Lectin类受体激酶LORE识别细菌脂多糖(lipopoly- saccharides, LPSs)并激活下游免疫信号(Ranf et al., 2015). ...
... )的抗性(Singh et al., 2012; Huang et al., 2014).LecRK- IX.2招募CPKs并磷酸化修饰RBOHD (respiratory burst oxidase homolog protein D), 正调控依赖于RBOHD的水杨酸(salicylic acid, SA)积累以及PTI信号途径(Luo et al., 2017).此外, 有研究表明, 拟南芥Lectin类受体激酶LORE识别细菌脂多糖(lipopoly- saccharides, LPSs)并激活下游免疫信号(Ranf et al., 2015). ...
Proteasome-mediated turnover of the trans- cription coactivator NPR1 plays dual roles in regulating plant immunity 1 2009
... 拟南芥水杨酸信号通路中, NPR1 (NONEXPRESSOR OF PATHOGENESIS-RELAED GENE 1)作为转录共激活因子调控多种免疫相关基因的表达.其蛋白稳定性受到泛素蛋白酶体途径的调控, 动态地调节免疫相关基因的转录激活或抑制.研究表明, NPR1不同位点的磷酸化修饰对其泛素化产生不同的影响.S11和S15位点的磷酸化修饰促进NPR1与SUMO3的结合, 从而促进NPR1的降解.而NPR1的S55和S59位点的磷酸化修饰则抑制NPR1的SUMO类泛素化, 增强NPR1蛋白的稳定性(Spoel et al., 2009; Saleh et al., 2015).此外, SnRK (SNF1-RELATED PRO- TEIN KINASE)激酶PSK5通过磷酸化修饰NPR1调控转录因子WRKY38和WRKY62的表达(Xie et al., 2010). ...
Receptor kinases in plant-pathogen interactions: more than pattern recognition 1 2017
... 植物中跨膜的受体类激酶和受体类蛋白(receptor like protein, RLP)识别胞外信号并将其转化为胞内信号.这些RLK或RLP又被称为模式识别受体(pattern recognition receptors, PRRs).典型的RLK由胞外域、单次跨膜的结构域以及胞质内的激酶结构域组成, RLP则不具有胞内激酶结构域.RLK或RLP的胞外域主要有4种类型: 亮氨酸重复序列(leucine-rich repeat, LRR)、赖氨酸基序(lysine motif, LysM)、凝集素样结构域(lectin domain)以及类表皮生长因子(epidermal growth factor, EGF)结构域(Couto and Zipfel, 2016).目前已报道多种受体激酶或受体蛋白参与植物免疫信号的识别和激活(Tang et al., 2017). ...
LYK4, a lysin motif receptor-like kinase, is important for chitin signaling and plant innate immunity in Arabidopsis 1 2012
... LysM类和EGF类PRRs主要识别包含碳水化合物的分子, 如真菌几丁质(chitin)、细菌肽聚糖(peptidog- lycan, PGN)和丛枝菌根菌分泌的脂质几丁寡糖(lipo- chitooligosaccharides, LCOs)或者植物细胞壁衍生的寡聚半乳糖醛酸(oligogalacturonides, OGs)等(图1).拟南芥中的3个LysM类受体激酶CERK1 (CHITIN ELICITOR RECEPTOR KINASE 1)、LYK4 (LysM- CONTAINING RECEPTOR-LIKE KINASE 4)和LYK5共同调控几丁质的识别及免疫信号的激活(Miya et al., 2007; Wan et al., 2012; Cao et al., 2014); LYK1和LYK3参与对不同结构的PGN信号的识别, 在AtCERK1存在的条件下激活PGN诱导的防御信号(Willmann et al., 2011).水稻LysM类RLK OsCERK1胞外的LysM结构域不能直接结合几丁质, LysM类受体蛋白OsCEBiP (CHITIN OLIFOSACCHARIDE ELICITOR BINDING PROTEIN)作为受体结合几丁质, 随后结合OsCERK1激活几丁质诱导的防御反应(Kaku et al., 2006; Shinya et al., 2015).拟南芥EGF类受体激酶WAK1 (WALL-ASSOCIATED KINASE 1)接受OGs信号并激活防御反应(Brutus et al., 2010).水稻免疫信号激活过程中, OsWAK1发生自磷酸化并磷酸化下游转录因子OsRFP1.此外, 水稻稻瘟菌侵染、机械损伤及水杨酸和茉莉酸甲酯处理均可以激活OsWAK1转录水平的表达(Li et al., 2009). ...
Arabidopsis LIP5, a positive regulator of multivesicular body biogenesis, is a critical target of pathogen-responsive MAPK cascade in plant basal defense 3 2014
... 根据胞外凝集素结构域的不同, 凝集素样受体激酶(lectin receptor kinase, LecRK)又分为3种类型: G (GNA-related or S-locus)-、C (calcium-dependent)-以及L (legume)-类(Bouwmeester and Govers, 2009).目前已经鉴定到的L-类LecRK有45个, 它们的功能大部分尚属未知.越来越多的研究表明, L-类LecRK广泛参与植物免疫调控.病原菌或PAMPs诱导LecRK-I.9、LecRK-V.5、LecRK-VI.2以及LecRK- IX.2激活表达; 这些LecRKs功能缺失表现出对不同病原菌的敏感性, 过表达则对病原菌抗性增强(Bou- wmeester et al., 2011; Singh et al., 2012; Wang et al., 2014b; Luo et al., 2017).LecRK-V.5抑制病原菌侵染后的气孔关闭过程; lecrk-V.5缺失突变体对疫霉菌属的抗病性减弱, 对丁香假单胞菌(Pseudomonas syringae pv. tomato (Pst) DC3000)的抗病性增强(Desclos-Theveniau et al., 2012; Wang et al., 2014).L-类LecRK-V.2和LecRK-VII.1结合FLS2, 正调控气孔免疫途径(Yekondi et al., 2018).LecRK- VI.2结合FLS2, 正调控PTI的响应, 参与对丁香假单胞菌和果胶杆菌(Pectobacterium carotovorum)的抗性(Singh et al., 2012; Huang et al., 2014).LecRK- IX.2招募CPKs并磷酸化修饰RBOHD (respiratory burst oxidase homolog protein D), 正调控依赖于RBOHD的水杨酸(salicylic acid, SA)积累以及PTI信号途径(Luo et al., 2017).此外, 有研究表明, 拟南芥Lectin类受体激酶LORE识别细菌脂多糖(lipopoly- saccharides, LPSs)并激活下游免疫信号(Ranf et al., 2015). ...
... ; Wang et al., 2014).L-类LecRK-V.2和LecRK-VII.1结合FLS2, 正调控气孔免疫途径(Yekondi et al., 2018).LecRK- VI.2结合FLS2, 正调控PTI的响应, 参与对丁香假单胞菌和果胶杆菌(Pectobacterium carotovorum)的抗性(Singh et al., 2012; Huang et al., 2014).LecRK- IX.2招募CPKs并磷酸化修饰RBOHD (respiratory burst oxidase homolog protein D), 正调控依赖于RBOHD的水杨酸(salicylic acid, SA)积累以及PTI信号途径(Luo et al., 2017).此外, 有研究表明, 拟南芥Lectin类受体激酶LORE识别细菌脂多糖(lipopoly- saccharides, LPSs)并激活下游免疫信号(Ranf et al., 2015). ...
... MAPKs对底物蛋白的磷酸化修饰影响底物蛋白的稳定性.拟南芥中, MPK3/MPK6通过磷酸化修饰ACS2/ACS6调控乙烯产生.泛素蛋白酶降解复合体结合未被磷酸化修饰的ACS6的C末端非催化区域, 导致ACS6经泛素化途径降解.MPK3/MPK6对ACS6的磷酸化修饰在其C末端引入了负电荷, 抑制ACS6的泛素化降解, 增强其蛋白的稳定性, 进而促进乙烯生成(Joo et al., 2008; Han et al., 2010).MPK3/ MPK6磷酸化修饰ERF家族成员ERF6, 增强其蛋白稳定性, 激活PDF1.1 (plant defensin 1.1)、PDF- 1.2a、PDF1.2b和ChiB等多种防御相关基因的表达(Meng et al., 2013).受到PAMP诱导后, MPK3磷酸化修饰WRKY46的S168和S250位点, 促进WRKY46的降解(Sheikh et al., 2016).LIP5参与病原菌诱导的内吞作用和囊泡运输, MPK3和MPK6磷酸化修饰LIP5, 增强LIP5的蛋白稳定性, 从而正调控多泡体(multivesicular bodies, MVB)通路, 影响免疫应答过程中相关因子的重定位(Wang et al., 2014a).E3泛素连接酶PUB22是拟南芥中PTI响应的抑制因子.MPK3对PUB22的磷酸化修饰抑制其自泛素化, 从而导致PUB22积累, 抑制免疫信号激活(Furlan et al., 2017). ...
A single transcription factor promotes both yield and im- munity in rice 2 2018
... RLCK的活性和稳定性受到严格调控.没有PAMP诱导时, 蛋白磷酸酶PP2C38结合并去磷酸化胞质内受体激酶BIK1, 抑制其对RBOHD的磷酸化修饰及下游免疫途径的激活.经PAMP诱导后, PP2C38与FLS2/ EFR-BIK1复合体分离, 解除对BIK1活性的抑制.此外, BIK1对PP2C38第7位丝氨酸残基的磷酸化修饰促进二者分离(Couto et al., 2016).钙离子依赖的蛋白激酶CPK28一方面磷酸化修饰U-box蛋白PUB25/ 26, 增强其泛素蛋白酶活性, 进而促进BIK1的降解; 另一方面CPK28直接磷酸化BIK1, 负调控多种PAMPs诱导后BIK1介导的免疫途径.XLG2/XLG3 (Gα)和AGB1 (Gβ)等G蛋白则通过抑制PUB25/26的活性稳定BIK1 (Monaghan et al., 2014; Liang et al., 2016; Wang et al., 2018a, 2018b). ...
A regulatory module controlling homeostasis of a plant immune kinase 1 2018
... RLCK的活性和稳定性受到严格调控.没有PAMP诱导时, 蛋白磷酸酶PP2C38结合并去磷酸化胞质内受体激酶BIK1, 抑制其对RBOHD的磷酸化修饰及下游免疫途径的激活.经PAMP诱导后, PP2C38与FLS2/ EFR-BIK1复合体分离, 解除对BIK1活性的抑制.此外, BIK1对PP2C38第7位丝氨酸残基的磷酸化修饰促进二者分离(Couto et al., 2016).钙离子依赖的蛋白激酶CPK28一方面磷酸化修饰U-box蛋白PUB25/ 26, 增强其泛素蛋白酶活性, 进而促进BIK1的降解; 另一方面CPK28直接磷酸化BIK1, 负调控多种PAMPs诱导后BIK1介导的免疫途径.XLG2/XLG3 (Gα)和AGB1 (Gβ)等G蛋白则通过抑制PUB25/26的活性稳定BIK1 (Monaghan et al., 2014; Liang et al., 2016; Wang et al., 2018a, 2018b). ...
Sequential transphosph orylation of the BRI1/BAK1 receptor kinase complex im- pacts early events in brassinosteroid signaling 0 2008
Phenotypic analyses of Arabidopsis T-DNA insertion lines and expression profiling reveal that multiple L-type lectin receptor kinases are involved in plant imm- unity 0 2014
A Pseudomonas syringae ADP-ribosyltransferase inhibits Arabidopsis mitogen-acti- vated protein kinase kinases 1 2010
... R蛋白监控MAPKs对底物蛋白的磷酸化修饰(图2).多种病原菌分泌效应蛋白攻击宿主细胞内MAPKs信号级联组成元件, 抑制MAPKs调控的防御反应(Zhang et al., 2007, 2012; Wang et al., 2010).拟南芥抗病蛋白SUMM2 (SUPPRESSOR OF MKK1 MKK2 2)在MAPKs信号的下游, 监控MPK4底物PAT1 (PROTEIN ASSOCIATED WITH TOPOISOMERASE II)、CRCK3 (CALMODULIN-BINDGIN RECEPTOR LIKE CYTOPLASMIC KINASE 3)以及MEKK2的磷酸化状态, 从而调控免疫应答.拟南芥PAT1编码mRNA脱帽装置的组成元件, 与MPK4结合形成复合体.PAMP诱导后, PAT1被MPK4磷酸化并在细胞溶质中聚集成离散的点状物.pat1缺失突变体表现出依赖于SUMM2的免疫自激活(Roux et al., 2015).CRCK3编码钙离子结合的胞质内受体激酶,结合MPK4并被MPK4磷酸化修饰.CRCK3缺失可恢复mekk1、mkk1/mkk2双突变体以及mpk4中SUMM2介导的免疫自激活.此外, CRCK3和PAT1均可以与SUMM2相互作用, 表明SUMM2可能通过监控CR- CK3和PAT1的磷酸化状态, 从而确定MEKK1- MKK1/MKK2-MPK4信号通路的完整性, 并适时启动免疫应答(Bi and Zhou, 2017; Zhang et al., 2017). ...
A protein phosphatase 2C, responsive to the bacterial effector AvrRpm1 but not to the AvrB effector, regulates defense responses in Arabidopsis 1 2010
Arabidopsis lysin-motif proteins LYM1 LYM3 CERK1 mediate bacterial pepti- doglycan sensing and immunity to bacterial infection 1 2011
... LysM类和EGF类PRRs主要识别包含碳水化合物的分子, 如真菌几丁质(chitin)、细菌肽聚糖(peptidog- lycan, PGN)和丛枝菌根菌分泌的脂质几丁寡糖(lipo- chitooligosaccharides, LCOs)或者植物细胞壁衍生的寡聚半乳糖醛酸(oligogalacturonides, OGs)等(图1).拟南芥中的3个LysM类受体激酶CERK1 (CHITIN ELICITOR RECEPTOR KINASE 1)、LYK4 (LysM- CONTAINING RECEPTOR-LIKE KINASE 4)和LYK5共同调控几丁质的识别及免疫信号的激活(Miya et al., 2007; Wan et al., 2012; Cao et al., 2014); LYK1和LYK3参与对不同结构的PGN信号的识别, 在AtCERK1存在的条件下激活PGN诱导的防御信号(Willmann et al., 2011).水稻LysM类RLK OsCERK1胞外的LysM结构域不能直接结合几丁质, LysM类受体蛋白OsCEBiP (CHITIN OLIFOSACCHARIDE ELICITOR BINDING PROTEIN)作为受体结合几丁质, 随后结合OsCERK1激活几丁质诱导的防御反应(Kaku et al., 2006; Shinya et al., 2015).拟南芥EGF类受体激酶WAK1 (WALL-ASSOCIATED KINASE 1)接受OGs信号并激活防御反应(Brutus et al., 2010).水稻免疫信号激活过程中, OsWAK1发生自磷酸化并磷酸化下游转录因子OsRFP1.此外, 水稻稻瘟菌侵染、机械损伤及水杨酸和茉莉酸甲酯处理均可以激活OsWAK1转录水平的表达(Li et al., 2009). ...
The type III effector HopF2 Pto targets Arabidopsis RIN4 protein to promote Pseudomonas syringae virulence 1 2010
... 拟南芥包含NOI结构域的蛋白RIN4在陆生植物中非常保守, 其并不具有类似已知的蛋白酶类的特征.二级结构预测发现, RIN4蛋白表现出高度的、固有的无序性, 不能形成二级和三级结构, 是植物中固有的无序蛋白(Lee et al., 2015).无序蛋白在与客户蛋白结合时转变成高度折叠状态, 松散并特异地与多种客户蛋白结合(Dyson and Wright, 2005).研究表明, RIN4是AvrRpt2、AvrRpm1、AvrB、HopF2以及AvrPto等多种病原菌效应子蛋白在宿主内攻击的靶标, 表明RIN4在植物-病原微生物互作中具有重要作用(Mackey et al., 2002, 2003; Axtell and Staskawicz, 2003; Luo et al., 2009; Wilton et al., 2010). ...
PKS5, a SNF1- related kinase, interacts with and phosphorylates NPR1, and modulates expression of WRKY38 and WRKY62 1 2010
... 拟南芥水杨酸信号通路中, NPR1 (NONEXPRESSOR OF PATHOGENESIS-RELAED GENE 1)作为转录共激活因子调控多种免疫相关基因的表达.其蛋白稳定性受到泛素蛋白酶体途径的调控, 动态地调节免疫相关基因的转录激活或抑制.研究表明, NPR1不同位点的磷酸化修饰对其泛素化产生不同的影响.S11和S15位点的磷酸化修饰促进NPR1与SUMO3的结合, 从而促进NPR1的降解.而NPR1的S55和S59位点的磷酸化修饰则抑制NPR1的SUMO类泛素化, 增强NPR1蛋白的稳定性(Spoel et al., 2009; Saleh et al., 2015).此外, SnRK (SNF1-RELATED PRO- TEIN KINASE)激酶PSK5通过磷酸化修饰NPR1调控转录因子WRKY38和WRKY62的表达(Xie et al., 2010). ...
The Arabidopsis CERK1- associated kinase PBL27 connects chitin perception to MAPK activation 1 2016
Danger peptide receptor signaling in plants ensures basal immunity upon pathogen-induced depletion of BAK1 1 2016
... 拟南芥LRR类受体激酶SERKs (SOMATIC EMBRYOGENESIS RECEPTOR KINASEs)作为FLS2/EFR/PEPRs等PRRs的共受体参与免疫信号的识别和激活.PRR接受病原信号后与BAK1/SERK3 (BRI1-ASSOCIATED RECEPTOR KINASE 1)结合, 并且胞内激酶结构域发生自磷酸化修饰.与此同时, PRR与BAK1的激酶域相互磷酸化修饰, 迅速激活免疫信号(Gómez-Gómez and Boller, 2000; Yama- guchi et al., 2010; Yamada et al., 2016b).水稻中, OsSERK2正调控XA21、XA3以及OsFLS2介导的免疫响应(Chen et al., 2014). ...
A receptor-like cyto- plasmic kinase targeted by a plant pathogen effector is directly phosphorylated by the chitin receptor and medi- ates rice immunity 1 2013
... 胞质内的受体激酶结合RLK/RLP, 共同调控植物的生长发育、激素信号以及生物和非生物胁迫等过程.研究表明, 拟南芥RLCK-VII家族的PBS1 (AvrPphB SUSCEPTIBLE 1)、BIK1 (BOTRYTIS-INDUCED KINASE 1)、PBL1 (PBS1- LIKE 1)、PBL2、PBL27、PCRKs (PTI COMPROMISED RECEPTOR-LIKE CYTOPLASMIC KINASE)以及RLCK-XII家族成员BSK1 (BRASSINOSTEROID-SIGNALING KINASE 1)参与植物免疫信号传递(图1).例如, FLS2、EFR、PEPRs以及CERK1等多种PRR被激活后磷酸化修饰BIK1, 导致BIK1与PRRs复合体分离并进入细胞质激活下游免疫信号(Lu et al., 2010; Yang et al., 2010; Liu et al., 2013).PCRK1和PCRK2结合FLS2并在PAMPs诱导后被磷酸化激活, 冗余地诱导转录因子SARD1 (SYSTEMIC ACQUIRED RESISTANCE DEFICIENT 1)和CBP60g (CALMODULIN-BINFING PROTEIN 60g)的表达, 促进SA的合成(Kong et al., 2016).水稻RLCK-VII家族的OsRLCK185和OsRL- CK176结合CERK1, 传递PGN和几丁质激活的免疫信号(Yamaguchi et al., 2013; Ao et al., 2014).Os- RLCK185在拟南芥中的同源蛋白PBL27被CERK1磷酸化激活后, 正调控几丁质诱导的防御反应(Shinya et al., 2014). ...
PEPR2 is a second receptor for the Pep1 and Pep2 peptides and contributes to defense responses in Arabidopsis 1 2010
... 拟南芥LRR类受体激酶SERKs (SOMATIC EMBRYOGENESIS RECEPTOR KINASEs)作为FLS2/EFR/PEPRs等PRRs的共受体参与免疫信号的识别和激活.PRR接受病原信号后与BAK1/SERK3 (BRI1-ASSOCIATED RECEPTOR KINASE 1)结合, 并且胞内激酶结构域发生自磷酸化修饰.与此同时, PRR与BAK1的激酶域相互磷酸化修饰, 迅速激活免疫信号(Gómez-Gómez and Boller, 2000; Yama- guchi et al., 2010; Yamada et al., 2016b).水稻中, OsSERK2正调控XA21、XA3以及OsFLS2介导的免疫响应(Chen et al., 2014). ...
The cell surface leucine-rich repeat receptor for At Pep1, an endogenous peptide elicitor in Arabidopsis, is functional in transgenic tobacco cells 1 2006
... LRR类PRRs主要识别蛋白或多肽序列(图1).例如, LRR类受体激酶FLS2 (FLAGELLIN SENSING 2)和EFR (ELONGATION FACTOR-Tu RECEPTOR)分别识别并结合病原菌鞭毛蛋白保守的22个氨基酸残基组成的肽flg22, 以及延伸因子Tu (EF-Tu) N末端的保守氨基酸序列elf18 (Chinchilla et al., 2006; Zipfel et al., 2006); PEPR1/2 (PEP RECEPTOR)识别植物响应病原菌分泌的内源可溶性前体肽pep (pro- peptide) (Yamaguchi et al., 2006); 番茄(Solanum lycopersicum)中FLS3识别细菌flgII-28并增强对细菌的免疫响应(Hind et al., 2016); 水稻(Oryza sativa) LRR-类受体激酶XA21识别水稻白叶枯菌(Xantho- monas oryzae pv. oryzae, Xoo)中保守的蛋白RaxX等(Pruitt et al., 2015). ...
BRASSINOSTEROID-SIGNALING KINASE 1 phosphory- lates MAPKKK5 to regulate immunity in Arabidopsis 1 2018
Structural basis for the impact of phosphorylation on the activation of plant receptor-like kinase BAK1 0 2012
A mutant CHS3 protein with TIR-NB-LRR-LIM domains modulates growth, cell death and freezing tole- rance in a temperature-dependent manner in Arabi- dopsis 1 2010
... 胞质内的受体激酶结合RLK/RLP, 共同调控植物的生长发育、激素信号以及生物和非生物胁迫等过程.研究表明, 拟南芥RLCK-VII家族的PBS1 (AvrPphB SUSCEPTIBLE 1)、BIK1 (BOTRYTIS-INDUCED KINASE 1)、PBL1 (PBS1- LIKE 1)、PBL2、PBL27、PCRKs (PTI COMPROMISED RECEPTOR-LIKE CYTOPLASMIC KINASE)以及RLCK-XII家族成员BSK1 (BRASSINOSTEROID-SIGNALING KINASE 1)参与植物免疫信号传递(图1).例如, FLS2、EFR、PEPRs以及CERK1等多种PRR被激活后磷酸化修饰BIK1, 导致BIK1与PRRs复合体分离并进入细胞质激活下游免疫信号(Lu et al., 2010; Yang et al., 2010; Liu et al., 2013).PCRK1和PCRK2结合FLS2并在PAMPs诱导后被磷酸化激活, 冗余地诱导转录因子SARD1 (SYSTEMIC ACQUIRED RESISTANCE DEFICIENT 1)和CBP60g (CALMODULIN-BINFING PROTEIN 60g)的表达, 促进SA的合成(Kong et al., 2016).水稻RLCK-VII家族的OsRLCK185和OsRL- CK176结合CERK1, 传递PGN和几丁质激活的免疫信号(Yamaguchi et al., 2013; Ao et al., 2014).Os- RLCK185在拟南芥中的同源蛋白PBL27被CERK1磷酸化激活后, 正调控几丁质诱导的防御反应(Shinya et al., 2014). ...
Nonredundant functions of Arabidopsis LecRK-V.2 and LecRK-VII.1 in controlling stomatal imm- unity and jasmonate-mediated stomatal closure 1 2018
... 根据胞外凝集素结构域的不同, 凝集素样受体激酶(lectin receptor kinase, LecRK)又分为3种类型: G (GNA-related or S-locus)-、C (calcium-dependent)-以及L (legume)-类(Bouwmeester and Govers, 2009).目前已经鉴定到的L-类LecRK有45个, 它们的功能大部分尚属未知.越来越多的研究表明, L-类LecRK广泛参与植物免疫调控.病原菌或PAMPs诱导LecRK-I.9、LecRK-V.5、LecRK-VI.2以及LecRK- IX.2激活表达; 这些LecRKs功能缺失表现出对不同病原菌的敏感性, 过表达则对病原菌抗性增强(Bou- wmeester et al., 2011; Singh et al., 2012; Wang et al., 2014b; Luo et al., 2017).LecRK-V.5抑制病原菌侵染后的气孔关闭过程; lecrk-V.5缺失突变体对疫霉菌属的抗病性减弱, 对丁香假单胞菌(Pseudomonas syringae pv. tomato (Pst) DC3000)的抗病性增强(Desclos-Theveniau et al., 2012; Wang et al., 2014).L-类LecRK-V.2和LecRK-VII.1结合FLS2, 正调控气孔免疫途径(Yekondi et al., 2018).LecRK- VI.2结合FLS2, 正调控PTI的响应, 参与对丁香假单胞菌和果胶杆菌(Pectobacterium carotovorum)的抗性(Singh et al., 2012; Huang et al., 2014).LecRK- IX.2招募CPKs并磷酸化修饰RBOHD (respiratory burst oxidase homolog protein D), 正调控依赖于RBOHD的水杨酸(salicylic acid, SA)积累以及PTI信号途径(Luo et al., 2017).此外, 有研究表明, 拟南芥Lectin类受体激酶LORE识别细菌脂多糖(lipopoly- saccharides, LPSs)并激活下游免疫信号(Ranf et al., 2015). ...
A Pseudomonas syringae effector inactivates MAPKs to suppress PAMP-induced immunity in plants 1 2007
... R蛋白监控MAPKs对底物蛋白的磷酸化修饰(图2).多种病原菌分泌效应蛋白攻击宿主细胞内MAPKs信号级联组成元件, 抑制MAPKs调控的防御反应(Zhang et al., 2007, 2012; Wang et al., 2010).拟南芥抗病蛋白SUMM2 (SUPPRESSOR OF MKK1 MKK2 2)在MAPKs信号的下游, 监控MPK4底物PAT1 (PROTEIN ASSOCIATED WITH TOPOISOMERASE II)、CRCK3 (CALMODULIN-BINDGIN RECEPTOR LIKE CYTOPLASMIC KINASE 3)以及MEKK2的磷酸化状态, 从而调控免疫应答.拟南芥PAT1编码mRNA脱帽装置的组成元件, 与MPK4结合形成复合体.PAMP诱导后, PAT1被MPK4磷酸化并在细胞溶质中聚集成离散的点状物.pat1缺失突变体表现出依赖于SUMM2的免疫自激活(Roux et al., 2015).CRCK3编码钙离子结合的胞质内受体激酶,结合MPK4并被MPK4磷酸化修饰.CRCK3缺失可恢复mekk1、mkk1/mkk2双突变体以及mpk4中SUMM2介导的免疫自激活.此外, CRCK3和PAT1均可以与SUMM2相互作用, 表明SUMM2可能通过监控CR- CK3和PAT1的磷酸化状态, 从而确定MEKK1- MKK1/MKK2-MPK4信号通路的完整性, 并适时启动免疫应答(Bi and Zhou, 2017; Zhang et al., 2017). ...
Resistance gene N-mediated de novo synthesis and activation of a tobacco mitogen- activated protein kinase by tobacco mosaic virus infection 1 1998
... 通过对烟草(Nicotiana benthamiana)和番茄的研究发现, MAPKs除在PTI途径中发挥功能外, 也是ETI途径的组成元件.烟草和番茄中丝/苏氨酸蛋白激酶Pto与丁香假单胞菌分泌的效应子AvrPto/AvrPtoB相互作用, 激活抗病蛋白Prf及ETI响应.此过程中, MAPKKKα和MAPKKKε正调控Pto-介导的ETI途径(del Pozo et al., 2004; Oh and Martin, 2011).在表达抗病蛋白Cf-9的转基因烟草中, 真菌效应子Avr9激活烟草中的MAPK激酶SIPK (SALICYLIC ACID- INDUCED PROTEIN KINASE)和WIPK (WOUND- INDUCED PROTEIN KINASE), 并导致WIPK蛋白积累(Romeis et al., 1999; Pedley and Martin, 2005; Meng and Zhang, 2013).烟草花叶病毒(tobacco mosaic virus, TMV)在烟草抗病蛋白N的作用下磷酸化SIPK和WIPK, 并增加WIPK的mRNA丰度以及蛋白含量(Zhang and Klessig, 1998).此外, AvrRpt2对拟南芥MPK3和MPK6也有激活作用, 与野生型Pst DC3000相比, 携带AvrRpt2效应蛋白的Pst DC3000对MPK3和MPK6的激活能力增强(Eschen-Lippold et al., 2016). ...
Arabidopsis receptor-like protein30 and receptor- like kinase suppressor of BIR1-1/EVERSHED mediate innate immunity to necrotrophic fungi 1 2013
... 由于缺少胞内激酶结构域, LRR-RLP识别配体后通过结合RLKs激活胞内免疫信号.LRR类受体激酶 SOBIR1 (SUPPRESSOR OF BIR1-1 1)结合多种LRR-RLP调控防御反应的激活(Gust and Felix, 2014).拟南芥RLP23识别细菌、真菌以及卵菌分泌的坏死/乙烯诱导的多肽蛋白nlp20 (NECROSIS AND ETHYLENE-INDUCING PEPTIDE 1-LIKE PROTEINS 20)后结合SOBIR1, 并招募BAK1形成复合体激活免疫信号(Albert et al., 2015).RLP30结合SOBIR1和BAK1, 识别死体营养型真菌蛋白激发子SCFE1 (SCLEROTINIA CULTURE FILTRATE ELICITOR 1)并激活下游免疫反应(Zhang et al., 2013).番茄中, Cf-4、Ve1和EIX1 (ETHYLENE- INDUCING XYLANASE 1)等RLPs结合SOBIR1激活真菌诱导的免疫途径(Liebrand et al., 2013).番茄受体激酶ELR (ELICITIN RESPONSE)结合免疫共受体BAK1/SERK3, 通过识别多种疫霉菌激肽蛋白(elicitin proteins)激活广谱且持久的抗病性(Du et al., 2015). ...
The NLR protein SUMM2 senses the disruption of an immune signaling MAP kinase cascade via CRCK3 1 2017
... R蛋白监控MAPKs对底物蛋白的磷酸化修饰(图2).多种病原菌分泌效应蛋白攻击宿主细胞内MAPKs信号级联组成元件, 抑制MAPKs调控的防御反应(Zhang et al., 2007, 2012; Wang et al., 2010).拟南芥抗病蛋白SUMM2 (SUPPRESSOR OF MKK1 MKK2 2)在MAPKs信号的下游, 监控MPK4底物PAT1 (PROTEIN ASSOCIATED WITH TOPOISOMERASE II)、CRCK3 (CALMODULIN-BINDGIN RECEPTOR LIKE CYTOPLASMIC KINASE 3)以及MEKK2的磷酸化状态, 从而调控免疫应答.拟南芥PAT1编码mRNA脱帽装置的组成元件, 与MPK4结合形成复合体.PAMP诱导后, PAT1被MPK4磷酸化并在细胞溶质中聚集成离散的点状物.pat1缺失突变体表现出依赖于SUMM2的免疫自激活(Roux et al., 2015).CRCK3编码钙离子结合的胞质内受体激酶,结合MPK4并被MPK4磷酸化修饰.CRCK3缺失可恢复mekk1、mkk1/mkk2双突变体以及mpk4中SUMM2介导的免疫自激活.此外, CRCK3和PAT1均可以与SUMM2相互作用, 表明SUMM2可能通过监控CR- CK3和PAT1的磷酸化状态, 从而确定MEKK1- MKK1/MKK2-MPK4信号通路的完整性, 并适时启动免疫应答(Bi and Zhou, 2017; Zhang et al., 2017). ...
Disruption of PAMP- induced MAP kinase cascade by a Pseudomonas syringae effector activates plant immunity mediated by the NB-LRR protein SUMM2 1 2012
... R蛋白监控MAPKs对底物蛋白的磷酸化修饰(图2).多种病原菌分泌效应蛋白攻击宿主细胞内MAPKs信号级联组成元件, 抑制MAPKs调控的防御反应(Zhang et al., 2007, 2012; Wang et al., 2010).拟南芥抗病蛋白SUMM2 (SUPPRESSOR OF MKK1 MKK2 2)在MAPKs信号的下游, 监控MPK4底物PAT1 (PROTEIN ASSOCIATED WITH TOPOISOMERASE II)、CRCK3 (CALMODULIN-BINDGIN RECEPTOR LIKE CYTOPLASMIC KINASE 3)以及MEKK2的磷酸化状态, 从而调控免疫应答.拟南芥PAT1编码mRNA脱帽装置的组成元件, 与MPK4结合形成复合体.PAMP诱导后, PAT1被MPK4磷酸化并在细胞溶质中聚集成离散的点状物.pat1缺失突变体表现出依赖于SUMM2的免疫自激活(Roux et al., 2015).CRCK3编码钙离子结合的胞质内受体激酶,结合MPK4并被MPK4磷酸化修饰.CRCK3缺失可恢复mekk1、mkk1/mkk2双突变体以及mpk4中SUMM2介导的免疫自激活.此外, CRCK3和PAT1均可以与SUMM2相互作用, 表明SUMM2可能通过监控CR- CK3和PAT1的磷酸化状态, 从而确定MEKK1- MKK1/MKK2-MPK4信号通路的完整性, 并适时启动免疫应答(Bi and Zhou, 2017; Zhang et al., 2017). ...
Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium -medi- ated transformation 1 2006
... LRR类PRRs主要识别蛋白或多肽序列(图1).例如, LRR类受体激酶FLS2 (FLAGELLIN SENSING 2)和EFR (ELONGATION FACTOR-Tu RECEPTOR)分别识别并结合病原菌鞭毛蛋白保守的22个氨基酸残基组成的肽flg22, 以及延伸因子Tu (EF-Tu) N末端的保守氨基酸序列elf18 (Chinchilla et al., 2006; Zipfel et al., 2006); PEPR1/2 (PEP RECEPTOR)识别植物响应病原菌分泌的内源可溶性前体肽pep (pro- peptide) (Yamaguchi et al., 2006); 番茄(Solanum lycopersicum)中FLS3识别细菌flgII-28并增强对细菌的免疫响应(Hind et al., 2016); 水稻(Oryza sativa) LRR-类受体激酶XA21识别水稻白叶枯菌(Xantho- monas oryzae pv. oryzae, Xoo)中保守的蛋白RaxX等(Pruitt et al., 2015). ...
 ,*兰州大学生命科学学院, 细胞活动与逆境适应教育部重点实验室, 兰州 730000
,*兰州大学生命科学学院, 细胞活动与逆境适应教育部重点实验室, 兰州 730000
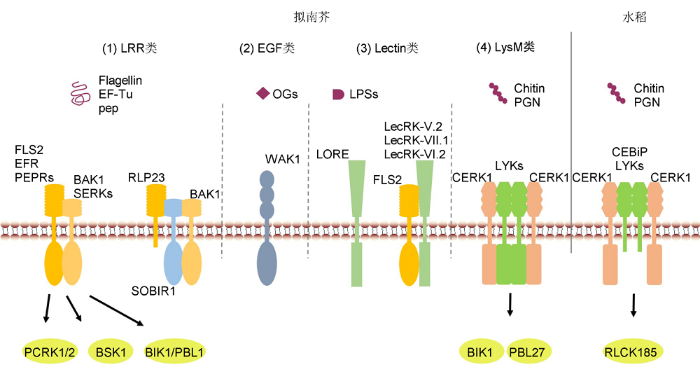 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT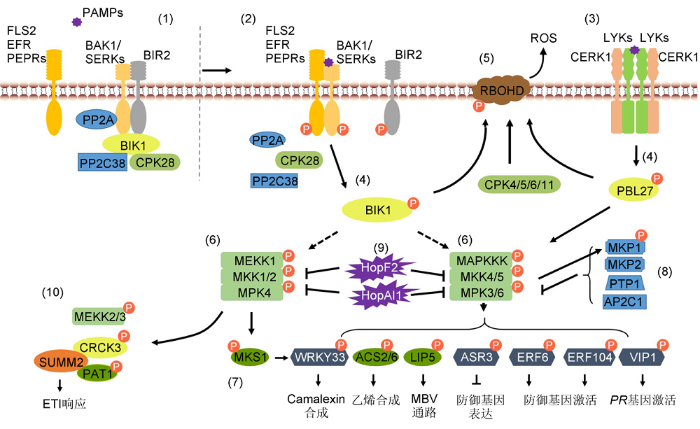 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}