 ,*, 张文忠,*沈阳农业大学水稻研究所, 农业部东北水稻生物学与遗传育种重点实验室, 北方超级粳稻育种教育部重点实验室, 沈阳 110866
,*, 张文忠,*沈阳农业大学水稻研究所, 农业部东北水稻生物学与遗传育种重点实验室, 北方超级粳稻育种教育部重点实验室, 沈阳 110866Heterologous Overexpression of Autophagy-related Gene OsATG8b from Rice Confers Tolerance to Nitrogen/Carbon Starvation and Increases Yield in Arabidopsis
Xiaoxi Zhen, Haoran Liu, Xin Li, Fan Xu,*, Wenzhong Zhang,*Key Laboratory of Northern Japonica Rice Genetics and Breedings, Ministry of Education, Key Laboratory of Northeast Rice Biology and Breeding, Ministry of Agriculture, Rice Research Institute, Shenyang Agriculture University, Shenyang 110866, China通讯作者:
收稿日期:2018-03-12接受日期:2018-07-16网络出版日期:2019-01-30
| 基金资助: |
Corresponding authors:
Received:2018-03-12Accepted:2018-07-16Online:2019-01-30

摘要
关键词:
Abstract
Keywords:
PDF (19481KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
甄晓溪, 刘浩然, 李鑫, 徐凡, 张文忠. 异源过表达OsATG8b基因提高转基因拟南芥的 氮/碳胁迫耐受性和产量. 植物学报, 2019, 54(1): 23-36 doi:10.11983/CBB18064
Zhen Xiaoxi, Liu Haoran, Li Xin, Xu Fan, Zhang Wenzhong.
氮素是植物体内最重要的营养元素, 是植物生长发育过程中必需的大量元素之一, 是氨基酸、蛋白质、核酸、叶绿素和激素等的重要组分(Kraiser et al., 2011)。氮素对植物(尤其是谷类作物)的产量和品质具有重要作用, 籽粒产量不仅取决于植株在开花前对氮素的吸收, 还取决于种子成熟期间氮素的动员再利用(Kichey et al., 2007), 因此对氮素循环再利用进行研究具有重要意义(Chardon et al., 2012)。
植物无法选择自身所需的营养环境, 只能依靠根际吸收有限的矿质营养维持生长发育, 因此植物必须有效代谢并循环利用体内的营养物质(Avila-Ospina et al., 2014; Krapp, 2015)。植物中大部分营养物质被分配到叶绿体中, C3植物中叶绿体蛋白质占叶片总氮的75%-80%, 因此叶绿体是植物中碳、氮循环再利用的主要来源(Makino and Osmond, 1991)。光合作用CO2固定酶Rubisco占叶片中可溶性蛋白质的50%以上, 是氮素动员再利用的主要来源(Masclaux- Daubresse et al., 2010)。
在植物发育的后期阶段, 叶片蛋白(尤其是叶绿体蛋白)被迅速降解, 从而输出大量可再利用的氮, 其释放的游离氨基酸及其它化合物被转运至新生的生殖器官和储存器官中(如新生的叶片或种子) (Masclaux-Daubresse et al., 2008), 这部分被重新再动员的氮也是种子中氮素的重要来源(Patrick and Offler, 2001)。植物中有机氮从衰老器官组织向籽粒中转移的循环过程(尤其是在氮胁迫条件下)是植物生产力和产量的重要决定因素(Masclaux-Daubresse et al., 2010)。虽然衰老叶片的蛋白质降解途径还没有被完全解析清楚, 但目前公认的代谢机制主要包括3种: 细胞质蛋白酶、衰老相关小泡(SAVs)和巨自噬(autophagy, 以下简称自噬) (Otegui et al., 2005; Ishida et al., 2008; Wada et al., 2009; Roberts et al., 2012)。
自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态。当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(Liu and Bassham, 2012; Yoshimoto, 2012; Avila-Ospinaet al., 2014; Yang and Bassham, 2015)。在植物细胞受到胁迫时, 自噬通过大规模的胞质降解路径降解单个长寿蛋白、蛋白质复合体及细胞器等来响应和适应胁迫条件(景红娟等, 2012)。Tsukada和Ohsumi (1993)首先在酿酒酵母(Saccharomyces cerevisiae)中发现了自噬相关基因。目前已发现40多个自噬相关基因(ATGs), 它们大多编码参与自噬发生与调控的核心蛋白(Ohsumi, 2001; Feng et al., 2014; Yao et al., 2015), 且大多数高等植物中都存在自噬相关同源基因。已在拟南芥(Arabidopsis thaliana)、水稻(Oryza sativa)和玉米(Zea mays)等植物中证实了自噬对氮素再利用的重要作用。Guiboileau等(2012)利用15N同位素标记实验首次在拟南芥中证明了氮素从叶片到种子的再动员受自噬控制, 拟南芥自噬缺失导致突变体种子中氮素再利用效率显著降低至50%左右。水稻自噬缺失突变体Osatg7-1在营养生长阶段的生物量和氮素利用效率明显低于对照, 叶片早衰且衰老的叶片中氮含量较高, 说明Osatg7-1无法动员再利用衰老叶片中的氮素(Wada et al., 2015)。通过15N同位素标记发现, 玉米自噬缺失突变体atg12籽粒中再利用的氮素减少50%, 新生上部叶中的15N主要位于玉米棒上方, 说明玉米自噬突变体atg12中氮素从衰老叶向籽粒中再动员受到抑制(Li et al., 2015a)。在众多ATG蛋白中, ATG8蛋白在细胞水平的丰度可以调节自噬体的大小。在多种植物中过表达ATG8基因都表现出有利于整体植株生长发育的表型。例如, 过表达AtATG8f转基因拟南芥莲座叶显著增大, 生长发育速度加快, 且能有效抵抗碳氮胁迫(Slavikova et al., 2008); 过表达谷子(Setaria italica)自噬相关基因SiATG8a能够增强转基因拟南芥对氮胁迫和干旱胁迫的耐受性, 使氮胁迫下转基因拟南芥生物量与氮素含量增加, 而干旱胁迫下转基因拟南芥存活率较高(Li et al., 2015b); 异源表达谷子自噬相关基因SiATG8a还能够提高水稻对氮胁迫的耐受性(Li et al., 2016)。此外, 异源表达苹果(Malus domestica)自噬相关基因MdATG8i能够促进转基因拟南芥的营养生长, 增强转基因拟南芥和苹果愈伤组织对氮胁迫的耐受性(Wang et al., 2016); 过表达大豆(Glycine max)自噬相关基因Gm- ATG8c能够促进转基因大豆愈伤组织与转基因拟南芥抵抗氮胁迫及拟南芥的生长发育和产量提高(Xia et al., 2012)。Xia等(2011)通过对水稻全基因组数据分析, 鉴定了7个OsATG8的编码基因(OsATG8a、OsATG8b、OsATG8c、OsATG8d、OsATG8e、OsATG8f和OsATG8i)。Izumi等(2015)对OsATG8s的氨基酸序列进行比对分析, 发现OsATG8a-c氨基酸序列高度一致, 而OsATG8d与AtATG8i较为相似, 他们通过将荧光蛋白(FP)与OsATG8a和OsATG8d融合, 成功建立了FPs-OsATG8水稻自噬监测系统, 并用该系统证明能量缺乏能够诱导水稻叶绿体和根质体发生自噬, 使叶绿体蛋白得到循环再利用, 同时也表明OsATG8s在作物自噬研究中具有至关重要的作用。
为了筛选水稻的可能氮高效利用候选基因, 我们前期克隆到1个新的水稻自噬核心基因(LOC_Os04g 53240; Os04g0624000) OsATG8b (Oryza sativa Autophagy Related 8 gene)。该基因位于第4号染色体, 全长2 754 bp, CDS全长360 bp, 编码120个氨基酸残基。我们克隆了OsATG8b基因ATG上游1 988 bp的启动子序列, 并使用PlantCare对该启动子序列进行顺式作用元件分析, 结果显示该序列包含启动子基本转录元件TATA-box和CAAT-box, 同时还含有光响应元件(Box I、CATT-motif和G-Box)、与水杨酸的响应相关元件(TCA-element)、生长发育相关元件(Skn-1 motif)和参与逆境胁迫相关的作用元件(TCT-motif、CCAAT-box和MBS)等顺式调控元件。
在本研究中, 我们发现OsATG8b基因响应低氮胁迫诱导。与野生型相比, 过表达OsATG8b转基因拟南芥表现出明显的生长优势, 莲座叶大小和株高显著增加、抽薹开花提前、植株生物量和产量也显著增加。同时, 缺氮胁迫下过表达OsATG8b转基因拟南芥叶片中的自噬活性显著增强, 且能够有效缓解碳、氮胁迫造成的生长抑制。因此, 我们认为OsATG8b基因可能是1个协同提高氮素利用效率和生产潜力的关键基因。
1 材料与方法
1.1 植物材料及培养条件
实验所用水稻品种为粳稻(Oryza sativa L. subsp. japonica)沈农9816。培养条件: 光周期为14小时光照/ 10小时黑暗, 温度28°C/25°C, 光照强度为90 μmol·m-2·s-1。拟南芥生态型为Columbia-0。培养条件: 光周期为16小时光照/8小时黑暗, 温度(22±1)°C, 光照强度为80 μmol·m-2·s-1。
1.2 水稻幼苗中OsATG8b基因对氮胁迫的响应
挑选籽粒饱满且大小一致的水稻种子, 经表面消毒后于28°C催芽2天。选取发芽状态一致的水稻种子置于1/2Hoagland营养液中培养14天, 然后将生长状态一致的水稻幼苗以全氮(NS)、低氮(NL)和缺氮(ND) 1/2 Hoagland营养液继续培养。全氮1/2Hoagland营养液中氮的浓度为3 mmol·L-1, 用KNO3和(NH4)2SO4来改变营养液中氮的浓度; 低氮条件下二者的浓度分别为0.6和0.1 mmol·L-1; 缺氮条件下二者浓度均为0 mmol·L-1, 缺少的K+用KCl来补足。培养1和3天时分别取水稻幼苗的叶片和根, 液氮速冻后保存于-80°C冰箱, 用于总RNA提取和基因表达分析。1.3 植物表达载体构建及拟南芥遗传转化
以水稻cDNA为模板, 利用常规PCR方法扩增Os- ATG8b基因的CDS序列(360 bp), 分别在上、下游引物中引入Nco I和BstE II的酶切位点, 通过Nco I/BstE II双酶切插入到双元表达载体pCAMBIA1301, 完成携带35S-OsATG8b的植物表达载体构建。以水稻基因组DNA为模板, 扩增OsATG8b基因启动子序列(1 988 bp), 分别在上、下游引物中引入EcoR I和Nco I的酶切位点, 通过EcoR I/Nco I双酶切插入到双元表达载体pCAMBIA1301, 完成ProOsATG8b-GUS双元表达载体的构建。使用农杆菌(Agrobacterium tumefaciens) GV3101介导的花序浸泡法(Clough and Bent, 1998)转化野生型拟南芥。将35S-OsATG8b转基因拟南芥T0代种子在含有30 mg·L-1潮霉素的筛选培养基上经过3代筛选获得5个纯合株系, 选取2个代表株系用于后续实验。通过GUS组织化学染色鉴定选取的启动子区域具有活性的ProOsATG8b-GUS转基因拟南芥。所有构建均经测序验证正确, 构建引物信息见表1。Table 1
表1
表1引物信息
Table 1
| Primer name | Sequence (5′-3′) | Function |
|---|---|---|
| cOsATG8b-F | CCATTCAAGTGGATGGCCAAGAGCTCGTTCAAGC | Gene cloning |
| cOsATG8b-R | GGTGACCTAGAGCAGCCCAAAGGTGTTCTCG | Gene cloning |
| cpOsATG8b-F | AAGCTTAAAATTAAATAAGACGAACAGTCAAACG | Gene cloning |
| cpOsATG8b-R | CCATGGCGCTCCTTCCTGCACACAAT | Gene cloning |
| rtOsATG8b-F | GCTGATCTTACCGTTGGGCA | Real-time RT-PCR |
| rtOsATG8b-R | ATCAGAGCAGCTGTTGGTGG | Real-time RT-PCR |
| rtAtAMT1-F | GCCTCTGCTGACTACTCCAACTT | Real-time RT-PCR |
| rtAtAMT1-R | GACCAGAACCAGTGAGAGACGA | Real-time RT-PCR |
| rtAtNR1-F | AGGATGGGCTAGTAAGCATAAGG | Real-time RT-PCR |
| rtAtNR1-R | GCAAACTGAATCATAGGCGGTG | Real-time RT-PCR |
| rtAtGS2-F | CACCAAACCTTACTCTCTGACA | Real-time RT-PCR |
| rtAtGS2-R | CACTATCTTCACCAGGTGCTTG | Real-time RT-PCR |
| rtAtGDH1-F | GCTTTAGCAGCAACAAACAGAA | Real-time RT-PCR |
| rtAtGDH1-R | TGAGCCAATGCGTTCACTTC | Real-time RT-PCR |
| rtACTIN1-F | ACCATTGGTGCTGAGCGTTT | Real-time RT-PCR |
| rtACTIN1-R | CGCAGCTTCCATTCCTATGAA | Real-time RT-PCR |
| rtTIP41-F | GTATGAAGATGAACTGGCTGACAAT | Real-time RT-PCR |
| rtTIP41-R | ATCAACTCTCAGCCAAAATCGCAAG | Real-time RT-PCR |
新窗口打开|下载CSV
1.4 转基因拟南芥的表型分析
使用10%安替福民对纯合的35S-OsATG8b转基因和野生型(WT)拟南芥种子表面消毒5分钟后, 用无菌蒸馏水冲洗4次, 置于4°C春化处理2天。将种子均匀平铺于1/2MS培养基, 苗龄8天时(萌发后天数)将幼苗转土培养, 用于全氮条件下的基本表型分析。用相机(Nikon D3200)对相应时间点表型进行拍照记录。为了分析转基因拟南芥对氮胁迫的耐受性, 将苗龄7天的转基因拟南芥和野生型(WT)移至1/2MS (NS)及不含氮素的1/2MS (ND)培养基上进行水平和垂直培养, 观察其地上和地下部分的生长变化。使用扫描仪(EPSON Perfection v33)对水平培养和垂直培养9天后的植株进行表型记录, 并利用Image J软件对垂直培养的根长进行比较分析。1.5 叶绿素和可溶性蛋白含量测定
总叶绿素含量测定参照Arnon (1949)的方法。可溶性蛋白含量测定参照Bradford法(Bradford, 1976)。1.6 氮代谢相关基因相对表达量分析
分别取全氮和缺氮处理14天的35SOsATG8b转基因及野生型拟南芥的叶片和根系, 液氮速冻后存于-80°C冰箱, 提取总RNA并反转录得到cDNA。以TIP41为内参基因对所有样品进行归一化处理, 利用荧光定量RT-PCR检测氮代谢相关基因的相对表达水平变化。总RNA提取、cDNA合成和荧光定量RT-PCR分析参照文献所述(Liu et al., 2010)。荧光定量RT-PCR检测所用的基因特异性引物和内参引物见 表1。1.7 外源植物激素/缺氮胁迫处理和GUS组织化学染色
将ProOsATG8b-GUS转基因拟南芥种子消毒后, 均匀平铺在含有30 mg·L-1潮霉素的筛选培养基上, 用于OsATG8b基因启动子的时空表达模式和对植物激素/缺氮胁迫的响应分析。植物激素水杨酸(SA) (SIGMA)浓度为10 μmol·L-1, 培养条件同1.1节所述, 处理时间24小时。GUS组织化学染色分析参照Liu等(2010)的文献。1.8 MDC染色检测自噬体活性
将苗龄7天的转基因和野生型拟南芥幼苗置于含有1 μmol·L-1刀豆素(ConA) (SIGMA)的缺氮培养液中处理12小时, 将叶片取下立即浸没于单丹磺酰戊二胺(monodansylcadaverine, MDC)染液(SIGMA)中进行染色, 通过激光共聚焦显微镜(Zeiss LSM 710)观察自噬体荧光信号, MDC染色方法参照Wang等(2013)的文献。2 结果与讨论
2.1 OsATG8b基因响应氮胁迫
在对沈农9816水稻幼苗进行低氮和缺氮处理1和3天时, 分别利用荧光定量RT-PCR检测了叶片及根系中OsATG8b基因的表达量(图1)。结果显示低氮处理1天, 水稻幼苗叶片中OsATG8b基因的表达量比对照略有上升, 缺氮处理1天则急剧上升(图1A); 水稻幼苗根系在低氮和缺氮处理1天时, OsATG8b基因的表达量随氮胁迫程度加深而上升(图1B); OsATG8b在低氮处理3天的水稻幼苗叶片中表达量增加(图1A), 在缺氮处理3天的叶片和根系中表达量均显著升高(图1)。上述结果表明, 水稻幼苗中OsATG8b基因的表达受氮胁迫诱导。图1
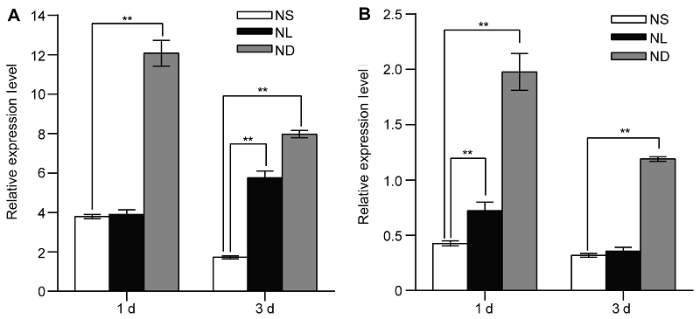 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1氮胁迫处理诱导水稻幼苗叶片及根中OsATG8b基因的表达
(A) 全氮条件下生长14天的水稻幼苗转移至全氮(NS)、低氮(NL)和缺氮(ND)条件下生长1和3天时叶片中OsATG8b基因的表达量; (B) 全氮、低氮和缺氮条件下生长1和3天的水稻根系中OsATG8b基因的表达量。图中数据为平均值±标准差, n=16, 实验经3次生物学重复。**表示全氮条件与低氮和缺氮条件相比差异显著(P<0.01) (Student’s t-test)。
Figure 1Identification of OsATG8b as a nitrogen deficiency inducible/responsive gene in leaves and roots of rice seedlings
(A) The rice seedlings cultured with N-sufficient (NS) solution for 14 days and transferred to the same NS solution, low N (NL) solution and the N-deficient (ND) solution, the expression of OsATG8b gene in leaves after 1 day and 3 days treatment; (B) The expression of OsATG8b gene in roots after 1 day and 3 days treatment. Values are means±SD, n=16, three biological replicates were performed. ** indicate significant differences in NS solution compared with NL and ND solution (P<0.01) (Student’s t-test).
2.2 过表达OsATG8b基因促进转基因拟南芥生长发育并提高产量
为了研究OsATG8b基因的功能, 我们构建了35S- OsATG8b双元表达载体, 获得了多个独立的转基因拟南芥株系。我们选取其中2个纯合株系作为代表株系进行持续的表型观察。首先, 利用荧光定量RT- PCR确定了OsATG8b在转基因拟南芥中的表达水平(图2A); 进一步通过表型观察, 我们发现在整个生长发育过程中, 转基因拟南芥的莲座叶均显著大于野生型(WT) (图2B, D), 且过表达OsATG8b加快了转基因拟南芥的发育进程, 使其抽薹开花明显提前约6天(表2)。此外, 我们统计了转基因拟南芥的株高, 发现转基因拟南芥的株高显著高于野生型(10.42%) (图2C, E); 我们还发现随着苗龄增加, 转基因拟南芥的叶绿素含量始终高于对照(图2F)。8天苗龄的转基因拟南芥莲座叶中的可溶性蛋白含量与对照几乎没有差异; 但从苗龄18天开始一直到植株成熟, 转基因拟南芥莲座叶中的可溶性蛋白含量始终高于对照(图2G)。经统计分析表明, 转基因拟南芥的总果荚数、千粒重和单株总种子重量显著增加(表3)。上述结果表明, 过表达OsATG8b基因能够促进转基因拟南芥的生长发育并提高其产量。Table 2
表2
表2野生型和35S-OsATG8b转基因拟南芥的抽薹及开花 时间
Table 2
| WT | L-13 | L-14 | |
|---|---|---|---|
| Bloting time (d) | 36.56±1.58 | 30.78±2.07** | 31.39±1.91** |
| Flowering time (d) | 42.67±1.75 | 35.94±1.98** | 36.61±1.94** |
新窗口打开|下载CSV
Table 3
表3
表3野生型和35S-OsATG8b转基因拟南芥的产量性状
Table 3
| Total number of siliques | Yield per plant (mg) | Thousand grain weight (mg) | |
|---|---|---|---|
| WT | 35.74±3.86 | 85.34±7.89 | 14.87±0.23 |
| L-13 | 46.26±3.13** | 100.13±6.02** | 16.36±0.21** |
| L-14 | 48.22±3.62** | 99.77±5.76** | 17.54±0.41** |
新窗口打开|下载CSV
2.3 过表达OsATG8b基因可提高转基因拟南芥对缺氮胁迫的耐受性
为研究过表达OsATG8b转基因拟南芥对氮胁迫的响应, 将在1/2MS培养基上生长7天的转基因及其野生型拟南芥幼苗移至1/2MS全氮(NS)和1/2MS缺氮(ND)培养基上处理9天, 采取水平和垂直2种培养方式进行地上部和地下部观察。我们发现无论在全氮还是缺氮条件下, 转基因拟南芥的生长情况都优于对照, 其地上部莲座叶和主根长均显著大于野生型(图3A, B, E, F), 地上部和地下部生物量也显著增加(图3G, H); 说明35SOsATG8b转基因拟南芥与野生型相比能够更好地抵抗缺氮胁迫。此外, 我们还测定了全氮/缺氮条件下转基因拟南芥和野生型的叶绿素和可溶性蛋白含量(图3C, D), 结果表明转基因拟南芥叶绿素和可溶性蛋白含量均高于WT。分别对苗龄为10天的ProOsATG8b-GUS转基因拟南芥进行缺氮处理和10 μmol·L-1水杨酸处理, 24小时后对其进行GUS组织化学染色, 发现OsATG8b启动子活性显著增强(图3I, J)。2.4 过表达OsATG8b促进转基因拟南芥抵抗黑暗诱导的碳胁迫
为进一步探究OsATG8b基因对碳饥饿的响应, 我们将苗龄7天的转基因和野生型拟南芥幼苗同时进行黑暗处理。结果表明, 黑暗处理10天后, 与野生型相比,转基因拟南芥生长速度加快, 莲座叶数目增多, 且叶片黄化的程度较轻, 叶绿素含量显著增加, 说明OsATG8b能够使植物更好地抵抗碳胁迫, 从而促进转基因拟南芥在碳胁迫条件下的生长发育(图4A, B)。2.5 过表达OsATG8b能够增强氮胁迫条件下转基因拟南芥叶片中的自噬活性
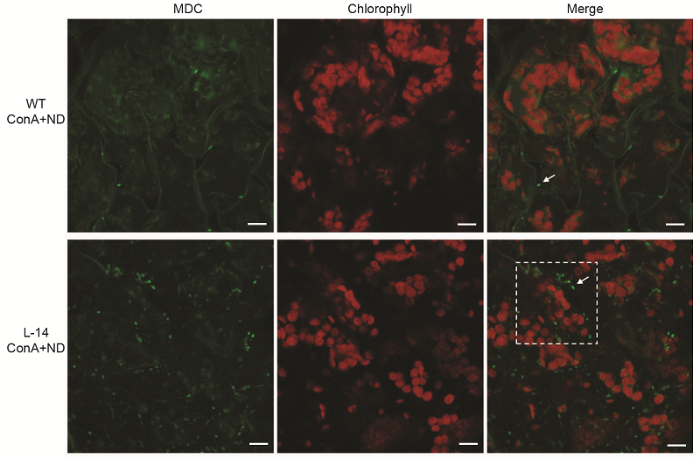
为了验证过表达OsATG8b转基因拟南芥中的自噬活性是否增强, 我们对苗龄7天的转基因和野生型拟南芥进行缺氮处理, 并通过MDC染色检测其叶片中的自噬活性。由于H+-ATPase抑制剂ConA能够增加植物细胞液泡腔内pH, 通过抑制液泡水解酶活性来促进自噬体在液泡中积累, 使细胞内自噬体便于观察, 我们在缺氮处理液中添加了H+-ATPase抑制剂Con- A。在激光共聚焦显微镜下观察, 转基因拟南芥叶肉细胞中的自噬体荧光信号显著多于野生型, 且信号强度较大(图5)。以上结果表明, 过表达OsATG8b能够显著增加转基因拟南芥自噬体的数量、增强自噬活性, 从而有效促进转基因拟南芥抵抗缺氮胁迫。2.6 过表达OsATG8b转基因拟南芥中氮代谢关键基因的表达变化
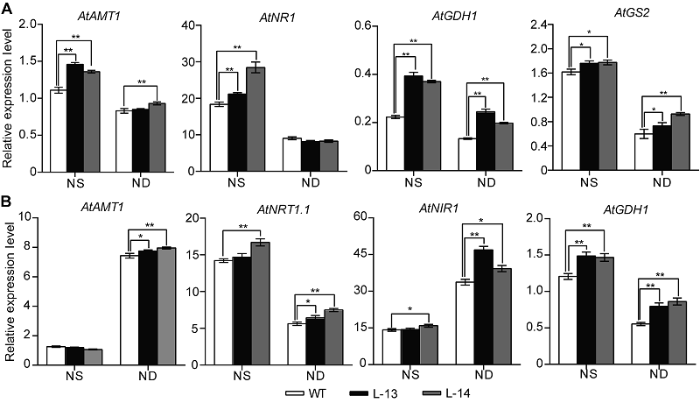
为进一步探究过表达OsATG8b转基因拟南芥对氮素代谢的响应, 我们检测了转基因和野生型拟南芥中一些氮代谢相关基因的相对表达水平。在全氮条件下, 过表达OsATG8b促进了转基因拟南芥叶片中铵转运蛋白基因(AtAMT1)、硝酸还原酶基因(AtNR1)及谷氨酰胺合成酶基因(AtGS2)的表达, 谷氨酸脱氢酶合成基因(AtGDH1)的表达量较低, 但仍显著高于野生型(图6A); 转基因拟南芥地下部硝酸盐转运蛋白基因(AtNRT1.1)和谷氨酸脱氢酶基因(AtGDH1)的表达量显著高于野生型, 亚硝酸还原酶基因(AtNIR1)的表达水平也略有升高(图6B)。在缺氮条件下, 转基因拟南芥地上部AtNR1、AtGDH1和AtGS2的表达水平显著高于野生型, AtAMT1的表达水平也略高于野生型(图6A); 过表达OsATG8b显著促进拟南芥根中AtNRT- 1.1、AtNIR1和AtGDH1的表达, AtAMT1的表达也有所升高(图6B)。上述结果表明, 无论在全氮还是缺氮条件下, 过表达OsATG8b均促进了转基因拟南芥中氮转运蛋白基因和氮代谢关键酶合成基因的表达。2.7 讨论
前人研究表明, 营养饥饿能够诱导植物自噬发生(Moriyasu and Ohsumi, 1996; Thompson, 2005)。碳、氮胁迫下, 许多自噬相关基因的转录水平显著上调(Breeze et al., 2011; Xia et al., 2011)。本研究中, 水稻自噬相关基因OsATG8b在氮胁迫诱导下表达水平显著升高, 同时在缺氮条件下还可以直接观察到过表达OsATG8b基因的转基因拟南芥叶片中自噬体活性增加(图5), 说明OsATG8b可能是参与抵抗氮胁迫的自噬基因(图1)。在其它植物中已有关于ATG8s基因功能的研究。例如, 过表达GmATG8c能够提高转基因拟南芥对氮胁迫的耐受性并使其产量增加(Xia et al., 2012); 异源表达SiATG8a能够促进转基因水稻抵抗氮饥饿(Li et al., 2016)。本研究中, 我们验证了OsATG8b在植株生长发育、产量形成和氮素利用中的作用。在适宜的生长条件下, 35S-OsATG8b转基因拟南芥表现出极强的生长优势, 过表达OsATG8b促进转基因拟南芥的营养生长, 使植株生物量显著增加(图2B, C, 图3A, E); 35S-OsATG8b转基因拟南芥莲座叶中叶绿素和可溶性蛋白含量增加(图2F, G), 反映出转基因拟南芥源器官中氮素含量较高, 表明过表达OsATG8b转基因拟南芥可能通过增加营养生长阶段氮素的吸收和积累, 为生殖生长和产量形成提供了充足的养分。过表达OsATG8b加速了转基因拟南芥营养生长向生殖生长的转变(表2), 延长了转基因拟南芥的生殖生长阶段, 从而使其果荚数量、单株产量和种子千粒重显著增加(图2C; 表3)。
图2
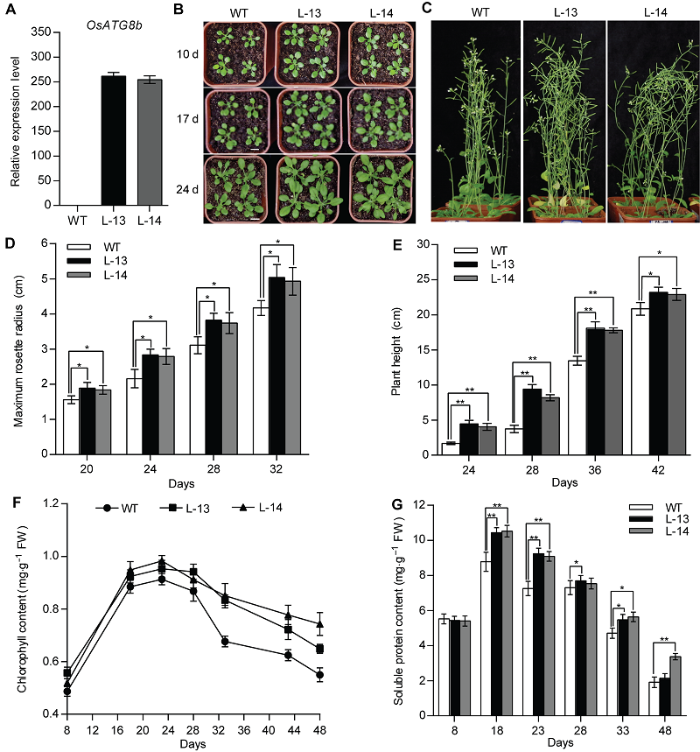 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2过表达OsATG8b促进转基因拟南芥生长发育
将苗龄8天的野生型(WT)和35S-OsATG8b转基因拟南芥幼苗移入蛭石-营养土(1:3, v/v)中。(A) 苗龄14天的35S-OsATG8b转基因拟南芥和WT幼苗中OsATG8b的表达水平; (B) 由上至下依次为移苗后10、17和24天的拟南芥表型; (C) 移苗后42天的表型; (D) 不同苗龄的35S-OsATG8b转基因拟南芥和WT的最大莲座叶半径; (E) 株高; (F) 总叶绿素含量; (G) 可溶性蛋白含量。Days: 萌发后天数。图中数据为平均值±标准差, n=24, * P<0.05, ** P<0.01 (Student’s t-test), 实验经3次生物学重复。Bars=1 cm
Figure 2Over-expression of OsATG8b promotes growth and development of transgenic Arabidopsis
8-day-old seedlings were transferred to vermiculite-nutritional soil (1:3, v/v). (A) Expression level of OsATG8b in 14-day-old seedlings of 35S-OsATG8b transgenic lines and wild type (WT); (B) Panels from top to bottom show phenotypic observations of transgenic lines and WT of Arabidopsis at 10, 17 and 24 days after transfer to soil, respectively; (C) Phenotype of transgenic lines and WT at 42 days after transfer to soil; (D) The maximum rosette radius of 35S-OsATG8b transgenic lines and WT at different seedling age; (E) The plant height; (F) The total chlorophyll content; (G) The soluble protein content. Days: Days after germination. Values are means±SD, n=24, * P<0.05, ** P<0.01 (Student’s t-test), three biological replicates were performed. Bars=1 cm
图3
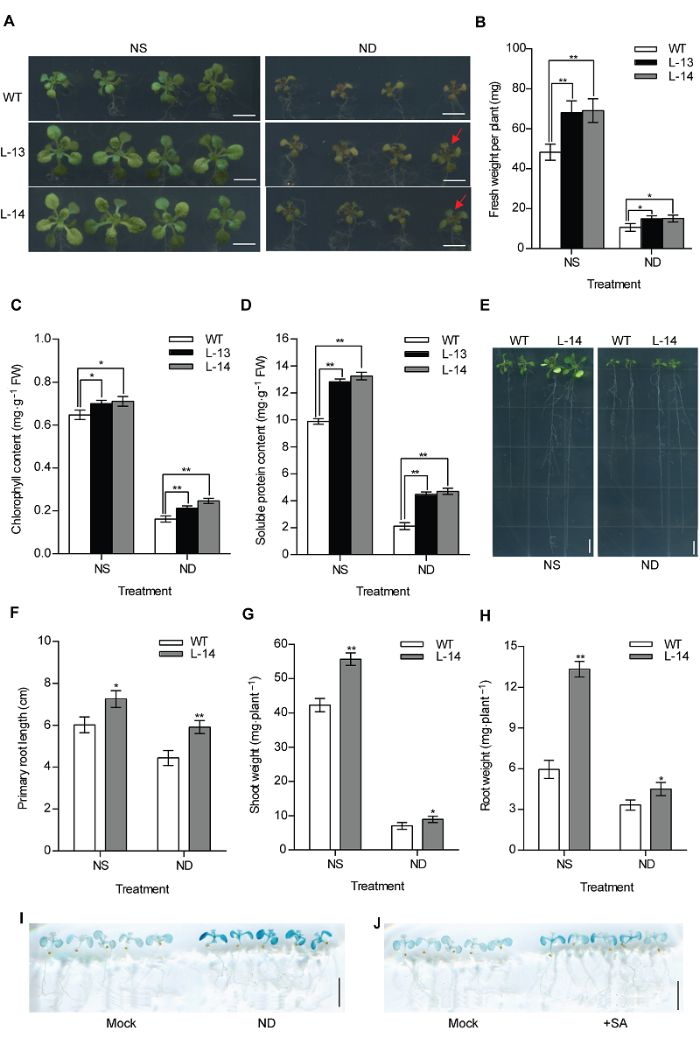 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3过表达OsATG8b促进转基因拟南芥抵抗缺氮胁迫
(A) 将苗龄7天的35S-OsATG8b转基因和野生型(WT)拟南芥幼苗分别移入全氮(NS)和缺氮(ND)1/2MS培养基中培养9天后的表型; (B)-(D) 分别为全氮(NS)和缺氮(ND)处理9天的35S-OsATG8b转基因和WT拟南芥幼苗的鲜重, 叶绿素和可溶性蛋白含量; (E) 苗龄7天的35S-OsATG8b转基因和WT拟南芥幼苗全氮(NS)和缺氮(ND)垂直培养9天后的表型; (F)-(H) 分别为全氮(NS)和缺氮(ND)处理9天的35S-OsATG8b转基因和WT拟南芥幼苗的主根长、地上部和地下部鲜重; (I), (J) 分别为苗龄10天的ProOsATG8b-GUS转基因拟南芥经缺氮处理和10 μmol·L-1水杨酸(SA)处理24小时后的GUS组织化学染色结果。Mock代表未经处理的拟南芥。数据为平均值±标准差, n=16, *和**分别表示各转基因株系与WT之间差异显著(P<0.05)和极显著(P<0.01) (Student’s t-test), 实验经3次生物学重复。Bars=5 mm
Figure 3Overexpression of OsATG8b enhances tolerance to N deficiency in transgenic Arabidopsis
(A) 7-day-old seedlings of 35S-OsATG8b transgenic lines and wild type (WT) were transferred to 1/2MS medium for horizontal culture with sufficient (NS) or deficient (ND) N for 9 days. (B)-(D) The fresh weight, chlorophyll content and soluble protein content in rosette leaves of WT and 35S-OsATG8b transgenic Arabidopsis under NS or ND for 9 days, respectively; (E) The phenotype of 7-day-old seedlings of 35S-OsATG8b transgenic lines and WT were transferred to vertical plates with NS or ND for 9 days; (F)-(H) The primary root length, the shoot weight and the root weight of WT and transgenic Arabidopsis lines under NS or ND for 9 days, respectively; (I), (J) 10-day-old seedlings of ProOsATG8b-GUS transgenic Arabidopsis were transferred to ND and 10 μmol·L-1 SA for 24 h, respectively. Mock represented that the seedlings without treated. Values are means±SD, n=16, * and ** indicate significant (P<0.05) and extremely significant (P<0.01) differences between transgenic lines and WT (Student’s t-test), three biological replicates were performed. Bars=5 mm
细胞自噬是植物中参与营养物质的重新分配、清除受损的蛋白和细胞器及响应生物和非生物胁迫等过程的一种重要的降解途径(黄晓和李发强, 2016)。在所有真核生物的养分循环过程中, 自噬都发挥重要作用, 尤其是对于氮素的再动员并向籽粒中转移至关重要。对拟南芥和玉米自噬相关基因缺失突变体的研究表明, 自噬缺失导致叶片中氮素向籽粒中再动员过程受阻(Guiboileau et al., 2012, 2013; Wada et al., 2015; Li et al., 2015a)。自噬相关基因缺失突变体在营养饥饿时表现出明显的生长发育迟缓、加速衰老及存活率下降等表型(Guiboileau et al., 2012; Wada et al., 2015; Li et al., 2015a; 刘洋等, 2018), 表明植物中自噬相关基因在抵抗营养胁迫过程中发挥重要作用。由于自噬缺陷导致细胞中的含氮资源无法被有效回收和利用, 因此产生的营养物质及废物堆积会促进细胞早衰死亡, 同时影响其它途径的氨基酸产生, 从而抑制植物生长, 并进一步降低氮素利用效率(任晨霞和龚清秋, 2014)。过表达OsATG8b不仅能在全氮条件下显著增加转基因拟南芥的全株生物量, 而且能有效缓解缺氮胁迫对莲座叶和根系造成的生长抑制(图3A, E)。转基因拟南芥莲座叶中叶绿素和可溶性蛋白含量有所增加(图3C, D), 说明过表达OsATG8b能促进转基因拟南芥在全氮条件下积累更多可利用的氮。MDC是一种广泛应用于哺乳动物和植物研究的嗜酸性染料, 它可以作为识别自噬体的探针来检测自噬结构(Biederbick et al., 1995; Contento et al., 2005)。ConA可以通过增大液泡腔内的内部pH值, 使细 胞中的水解酶处于最适环境有利于自噬体的观察(Yoshimoto et al., 2004; Ishida et al., 2008)。在缺氮条件下, 通过MDC染色结合激光共聚焦观察, 我们还发现过表达OsATG8b基因能够显著增加转基因拟南芥叶片中的自噬体活性(图5), 表明转基因拟南芥可以在缺氮条件下通过增加自噬活性更好地利用前期积累的氮素; 缺氮处理24小时的ProOsATG8b-GUS转基因拟南芥中OsATG8b启动子活性增强(图3I), 进一步证明氮胁迫能够诱导OsATG8b表达, 使自噬作用增强, 从而提高转基因拟南芥对氮胁迫的耐受性。根系是植物感知生长环境中氮素供应情况的重要器官, 其形态结构对于氮素的吸收利用非常关键(Walch- Liu et al., 2005), 过表达OsATG8b转基因拟南芥的根系有助于增加其对氮素的吸收。
图4
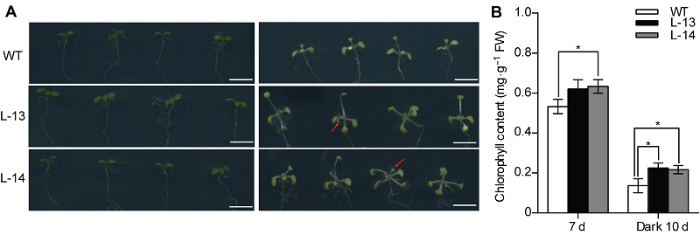 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4过表达OsATG8b促进转基因拟南芥抵抗黑暗诱导的碳胁迫
(A) 对苗龄7天的35S-OsATG8b转基因拟南芥和野生型(WT)黑暗处理10天后记录表型(左为处理前, 右为处理后); (B) 叶绿素含量测定。数据为平均值±标准差, n=10, *表示各转基因株系与WT之间差异显著(P<0.05) (Student’s t-test), 实验经3次生物学重复。Bars=5 mm
Figure 4Overexpression of OsATG8b in Arabidopsis enhanced tolerance to carbon starvation induced by dark treatment
(A) 7-day-old seedlings of 35S-OsATG8b transgenic lines and wild type (WT) were transferred to darkness for 10 days (The left is before treatment, and the right is after treatment); (B) The chlorophyll content determination. Values are means±SD, n=10, * indicate significant difference between transgenic lines and WT (P<0.05) (Student’s t-test), three biological replicates were performed. Bars=5 mm
图5
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5缺氮条件下过表达OsATG8b转基因拟南芥中的自噬活性增加
将苗龄7天的35S-OsATG8b转基因(L-14)和野生型(WT)拟南芥幼苗转移至含有1 μmol·L-1 ConA的缺氮培养液中处理12小时后进行MDC染色, 于激光共聚焦显微镜下观察自噬体荧光。Bars=10 μm
Figure 5Overexpression of OsATG8b in Arabidopsis enhanced the autophagic activity under N deficient condition
7-day-old seedlings of transgenic line (L-14) and wild type (WT) were transferred to in N-deficient (ND) liquid medium with 1 μmol·L-1 ConA for 12 h, MDC-stained autophagosomes in leaves were observed by confocal microscopy. Bars=10 μm
图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6过表达OsATG8b转基因拟南芥中氮代谢关键基因的表达变化
将7天苗龄的35S-OsATG8b转基因和野生型(WT)拟南芥移至全氮(NS)和缺氮(ND) 1/2MS培养基处理14天。(A) 35S-OsATG8b转基因和WT拟南芥中地上部氮代谢相关基因相对表达水平; (B) 地下部氮代谢相关基因相对表达水平。数据为平均值±标准差, n=10, *和**分别表示各转基因株系与WT之间差异显著(P<0.05)和极显著(P<0.01) (Student’s t-test), 实验经3次生物学重复。
Figure 6Overexpression of OsATG8b in Arabidopsis changes the expression of genes in nitrogen metabolic
7-day-old seedlings of transgenic lines and wild type (WT) were transferred to 1/2MS medium with sufficient (NS) or deficient (ND) nitrogen for 14 days. (A) The expression of genes related to nitrogen metabolic in rosette leaves of 35S-OsATG8b transgenic lines and WT; (B) The expression of genes related to nitrogen metabolic in roots of 35S-OsATG8b transgenic lines and WT. Values are means±SD, n=10, * and ** indicate significant (P<0.05) and extremely significant (P<0.01) differences between transgenic lines and WT (Student’s t-test), respectively, three biological replicates were performed.
Yoshimoto等(2009)研究表明, 拟南芥自噬缺失突变体atg5的早衰表型是由水杨酸过量积累造成的, 在拟南芥自噬突变体atg5-2中过表达细菌的水杨酸羟化酶NahG基因, 或者在双突变体atg5-2/sid2 (sid2为水杨酸合成缺陷突变体)中, atg5-2的叶片早衰表型均得到抑制。我们发现OsATG8b启动子响应水杨酸处理, 其活性明显增强(图3J), 表明自噬可能在植物抵抗衰老过程中发挥一定作用。前人研究表明, 自噬可以通过提供游离氨基酸等能源来保证夜间能量供应(Izumi et al., 2013), 且自噬在叶片淀粉降解过程中也起到一定作用(Wang et al., 2013)。本研究表明, 过表达OsATG8b能够有效缓解碳胁迫(黑暗处理10天)造成的转基因拟南芥生长受抑制和叶绿素含量下降(图4); 此外, 将成熟幼苗经黑暗处理后我们发现过表达OsATG8b基因可以显著抑制由于黑暗处理导致的叶片叶绿素含量下降(结果未显示), 表明过表达OsATG8b可能通过增强自噬作用, 为碳饥饿条件下的转基因拟南芥的代谢活动提供能量。
植物通过硝酸盐转运因子(AtNRT)和铵盐转运因子(AtAMTs)转运根系吸收的硝态氮与铵态氮(Good et al., 2004; Rentsch et al., 2007)。硝酸盐在硝酸盐还原酶(NR)和亚硝酸盐还原酶(NiR)的作用下形成铵盐, 铵盐通过谷氨酰胺合成酶-谷氨酸合酶(GS2/ GOGAT)途径生成谷氨酸盐, 谷氨酸盐再通过膜系统转运到木质部并运输到其它库器官中(Meyer and Stitt, 2001)。本研究中, 无论是全氮还是缺氮条件下, 35S-OsATG8b转基因拟南芥莲座叶和根系中氮素吸收与同化相关基因AtAMT1、AtNRT1.1、AtNR1、AtNIR1和AtGS2的表达量均高于野生型(图6), 表明过表达OsATG8b可能促进转基因拟南芥对氮的吸收, 同时也促进硝态氮的还原和氨的同化与利用。谷氨酸脱氢酶(GDH)的某些亚型在氮素再动员过程中被激活(Masclaux-Daubresse et al., 2010); 过表达Os- ATG8b转基因拟南芥中AtGDH1表达量显著增加, 暗示OsATG8b可能促进转基因拟南芥的氮素动员再利用。
综上所述, 水稻自噬相关基因OsATG8b是一个能够促进植物生长发育、增加产量并提高氮素利用效率的重要候选基因。本研究进一步证实了自噬在植物生长发育和氮素利用方面发挥一定作用, 为提高作物氮素利用率和产量指明了一个新的方向。OsATG8b促进氮素的吸收利用并提高产量的具体机制还有待深入研究。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.11983/CBB16011URL [本文引用: 1]

细胞自噬是真核生物中一种由液泡或溶酶体介导的,对细胞内物质进行周转的重要代谢机制.在植物中,细胞自噬作为一种重要的降解手段,参与营养物质的重新分配、受损蛋白和细胞器的清除及生物和非生物胁迫的响应等过程.此外,细胞自噬在各种程序性细胞死亡中也起着重要作用,该文主要综述了近几年来在此方面的研究进展.
DOI:10.3724/SP.J.1259.2012.00534URL [本文引用: 1]
Autophagy is a highly conserved biological progress in degradation of the cell components and has key roles in starvation, senescence and infection by pathogens. Moreover, reactive oxygen species (ROS) is a byproduct of higher plants during normal and stress conditions and plays an important role in regulation of growth and development and programmed cell death during stress response. Recent research has shown that ROS and autophagy both regulate programmed cell death in the hypersensitive reaction during infection by pathogens, growth and development and stress adaptation. We discuss the signal interaction between ROS and autophagy in plants, including species and characteristics of ROS, the molecular basis of plant autophagy, and functions of ROS in plant autophagy.
URL [本文引用: 1]
细胞自噬是一类依赖于溶酶体和液泡的蛋白质降解途径。在动物细胞中,靶物质通过自噬体包裹被运送到溶酶体中,由特定的水解酶降解;而植物和酵母细胞中该过程在液泡内进行。近年来,在模式植物拟南芥(Arabidopsis thaliana)中鉴定到多个关键ATG基因,它们对植物细胞自噬体的形成及自噬调控起到关键作用。该文全面综述了植物细胞自噬的调控及其在植物逆境胁迫中的生理功能。
URL [本文引用: 1]
自噬(autophagy)是真核生物细胞通过形成自噬体,回收利用胞内物质,维持细胞健康的高通量亚细胞降解途径。随着酵母和动物自噬研究的深入,植物自噬也受到越来越多的关注。近期的研究揭示了植物自噬的基本机制及其生理意义,也发现了植物特有的自噬形式与自噬相关基因。该文主要综述了自噬在植物碳、氮营养中的作用。
.
[本文引用: 2]
.
DOI:10.1089/dna.1995.14.87URLPMID:7750517 [本文引用: 1]
Eur J Cell Biol. 1995 Jan;66(1):3-14.
DOI:10.1016/0003-2697(76)90527-3URLPMID:942051 [本文引用: 1]
A protein determination method which involves the binding of Coomassie Brilliant Blue G-250 to protein is described. The binding of the dye to protein causes a shift in the absorption maximum of the dye from 465 to 595 nm, and it is the increase in absorption at 595 nm which is monitored. This assay is very reproducible and rapid with the dye binding process virtually complete in approximately 2 min with good color stability for 1 hr. There is little or no interference from cations such as sodium or potassium nor from carbohydrates such as sucrose. A small amount of color is developed in the presence of strongly alkaline buffering agents, but the assay may be run accurately by the use of proper buffer controls. The only components found to give excessive interfering color in the assay are relatively large amounts of detergents such as sodium dodecyl sulfate. Triton X-100, and commercial glassware detergents. Interference by small amounts of detergent may be eliminated by the use of proper controls.
[本文引用: 1]
DOI:10.1093/jxb/err353URLPMID:22231501 [本文引用: 1]
There is evidence that crop yields are showing a trend of stagnation in many countries. This review aims to make an inventory of the last decade's crop productions and the associated economic and environmental challenges. Manipulating nitrogen use efficiency in crops appears to be the best way to conciliate global food security, respecting environmental policies, and the need to produce biofuels. In such a context, the specifications of ideal plants for the future are discussed with regards to human needs and taking into account current physiological and genetic knowledge. The approaches undertaken so far to design an ideal crop and to find suitable new germplasms are discussed. The interest in using model plants in agronomic research is illustrated through the recent data provided by studies exploring natural variation in Arabidopsis thaliana. Efficient Arabidopsis ideotypes are proposed and discussed.
.
[本文引用: 1]
DOI:10.1111/j.1365-313X.2005.02396.xURLPMID:15860017 [本文引用: 1]
Autophagy is a process that is thought to occur in all eukaryotes in which cells recycle cytoplasmic contents when subjected to environmental stress conditions or during certain stages of development. Upon induction of autophagy, double membrane-bound structures called autophagosomes engulf portions of the cytoplasm and transfer them to the vacuole or lysosome for degradation. In this study, we have characterized two potential markers for autophagy in plants, the fluorescent dye monodansylcadaverine (MDC) and a green fluorescent protein (GFP)-AtATG8e fusion protein, and propose that they both label autophagosomes in Arabidopsis. Both markers label the same small, apparently membrane-bound structures found in cells under conditions that are known to induce autophagy such as starvation and senescence. They are usually seen in the cytoplasm, but occasionally can be observed within the vacuole, consistent with a function in the transfer of cytoplasmic material into the vacuole for degradation. MDC-staining and the GFP-AtATG8e fusion protein can now be used as very effective tools to complement biochemical and genetic approaches to the study of autophagy in plant systems.
[本文引用: 1]
DOI:10.1016/j.tplants.2004.10.008URLPMID:15564127 [本文引用: 1]
Plant scientists have long recognized the need to develop crops that absorb and use nutrients more efficiently. Two approaches have been used to increase nutrient use efficiency (NUE) in crop plants. The first involves both traditional breeding and marker-assisted selection in an attempt to identify the genes involved. The second uses novel gene constructs designed to improve specific aspects of NUE. Here, we discuss some recent developments in the genetic manipulation of NUE in crop plants and argue that an improved understanding of the transition between nitrogen assimilation and nitrogen recycling will be important in applying this technology to increasing crop yields. Moreover, we emphasize the need to combine genetic and transgenic approaches to make significant improvements in NUE.
[本文引用: 1]
[本文引用: 2]
.
DOI:10.1104/pp.108.122770URL [本文引用: 2]
During senescence and at times of stress, plants can mobilize needed nitrogen from chloroplasts in leaves to other organs. Much of the total leaf nitrogen is allocated to the most abundant plant protein, Rubisco. While bulk degradation of the cytosol and organelles in plants occurs by autophagy, the role of autophagy in the degradation of chloroplast proteins is still unclear. We have visualized the fate of Rubisco, stroma-targeted green fluorescent protein (GFP) and DsRed, and GFP-labeled Rubisco in order to investigate the involvement of autophagy in the mobilization of stromal proteins to the vacuole. Using immunoelectron microscopy, we previously demonstrated that Rubisco is released from the chloroplast into Rubisco-containing bodies (RCBs) in naturally senescent leaves. When leaves of transgenic Arabidopsis (Arabidopsis thaliana) plants expressing stroma-targeted fluorescent proteins were incubated with concanamycin A to inhibit vacuolar H-ATPase activity, spherical bodies exhibiting GFP or DsRed fluorescence without chlorophyll fluorescence were observed in the vacuolar lumen. Double-labeled immunoelectron microscopy with anti-Rubisco and anti-GFP antibodies confirmed that the fluorescent bodies correspond to RCBs. RCBs could also be visualized using GFP-labeled Rubisco directly. RCBs were not observed in leaves of a T-DNA insertion mutant in ATG5, one of the essential genes for autophagy. Stroma-targeted DsRed and GFP-ATG8 fusion proteins were observed together in autophagic bodies in the vacuole. We conclude that Rubisco and stroma-targeted fluorescent proteins can be mobilized to the vacuole through an ATG gene-dependent autophagic process without prior chloroplast destruction.
DOI:10.2307/41942799URLPMID:23457226 [本文引用: 1]
http://www.jstor.org/stable/41942799
DOI:10.1104/pp.114.254078URLPMID:25717038
Autophagy is an intracellular process leading to vacuolar or lysosomal degradation of cytoplasmic components in eukaryotes. Establishment of proper methods to monitor autophagy was a key step in uncovering its role in organisms, such as yeast (Saccharomyces cerevisiae), mammals, and Arabidopsis (Arabidopsis thaliana), in which chloroplastic proteins were found to be recycled by autophagy. Chloroplast recycling has been predicted to function in nutrient remobilization for growing organs or grain filling in cereal crops. Here, to develop our understanding of autophagy in cereals, we established monitoring methods for chloroplast autophagy in rice (Oryza sativa). We generated transgenic rice-expressing fluorescent protein (FP) OsAuTophaGy8 (OsATG8) fusions as autophagy markers. FP-ATG8 signals were delivered into the vacuolar lumen in living cells of roots and leaves mainly as vesicles corresponding to autophagic bodies. This phenomenon was not observed upon the addition of wortmannin, an inhibitor of autophagy, or in an ATG7 knockout mutant. Markers for the chloroplast stroma, stromal FP, and FP-labeled Rubisco were delivered by a type of autophagic body called the Rubisco-containing body (RCB) in the same manner. RCB production in excised leaves was suppressed by supply of external sucrose or light. The release of free FP caused by autophagy-dependent breakdown of FP-labeled Rubisco was induced during accelerated senescence in individually darkened leaves. In roots, nongreen plastids underwent both RCB-mediated and entire organelle types of autophagy. Therefore, our newly developed methods to monitor autophagy directly showed autophagic degradation of leaf chloroplasts and root plastids in rice plants and its induction during energy limitation.
.
DOI:10.1016/j.fcr.2007.01.002URL [本文引用: 1]
In wheat, nitrogen (N) uptake and remobilisation after flowering contributes largely, in Northern countries, to grain yield and grain protein content. The aim of our study was first to estimate the proportion of N taken up and remobilised to the grain as well as their relative efficiency using 15NO 3 labelling at flowering. The validity of the technique was assessed in comparison to the N budget calculation method on five winter wheat cultivars grown for 2 years at low and high fertilization input. We estimated that on average 71.2% of grain N originates from remobilisation with significant genotypic differences. Among the five genotypes, significant differences were also found for both N remobilisation efficiency (from 69.8 to 88.8%) and N translocation efficiency (from 89.7 to 93.4%). In parallel, during 1 year, we monitored physiological markers representative of N assimilation and recycling at two sampling dates during the grain filling period. We then examined if there was any relationship between these physiological markers, N absorption and remobilisation estimates and agronomic traits related to yield and grain N content. Nitrate reductase (NR) activity was highly correlated to N absorbed post-flowering and to grain protein content. Glutamine synthetase (GS) activity was even more highly correlated than NR activity to the amount of N remobilised and grain yield. The use of physiological traits such as NR and GS activities as markers of the wheat N status is discussed.
[本文引用: 1]
DOI:10.1016/j.pbi.2015.05.010URLPMID:26037390 [本文引用: 1]
Nitrogen (N) is an essential element for plants that is available in agricultural soils mainly as macronutrients in the form of nitrate and ammonium. Interplay between high-affinity and low-affinity transporters ensures efficient uptake from the soil even under highly fluctuating N availability. After uptake, N assimilation comprises the reduction of nitrate to ammonium and its subsequent incorporation into amino acids. Amino acids, but also nitrate, are transported from root to shoot and vice versa. Most steps of N transport and assimilation are tightly controlled by a regulatory network acting both cell-autonomously and systemically. N sensors, transcription factors and further regulatory players have been identified during recent years, elucidating parts of the huge puzzle that represents the efficient use of N by plants.
DOI:10.1105/tpc.15.00158URLPMID:25944100 [本文引用: 3]
Abstract Autophagy is a primary route for nutrient recycling in plants by which superfluous or damaged cytoplasmic material and organelles are encapsulated and delivered to the vacuole for breakdown. Central to autophagy is a conjugation pathway that attaches AUTOPHAGY-RELATED8 (ATG8) to phosphatidylethanolamine, which then coats emerging autophagic membranes and helps with cargo recruitment, vesicle enclosure, and subsequent vesicle docking with the tonoplast. A key component in ATG8 function is ATG12, which promotes lipidation upon its attachment to ATG5. Here, we fully defined the maize (Zea mays) ATG system transcriptionally and characterized it genetically through atg12 mutants that block ATG8 modification. atg12 plants have compromised autophagic transport as determined by localization of a YFP-ATG8 reporter and its vacuolar cleavage during nitrogen or fixed-carbon starvation. Phenotypic analyses showed that atg12 plants are phenotypically normal and fertile when grown under nutrient-rich conditions. However, when nitrogen-starved, seedling growth is severely arrested, and as the plants mature, they show enhanced leaf senescence and stunted ear development. Nitrogen partitioning studies revealed that remobilization is impaired in atg12 plants, which significantly decreases seed yield and nitrogen-harvest index. Together, our studies demonstrate that autophagy, while nonessential, becomes critical during nitrogen stress and severely impacts maize productivity under suboptimal field conditions. 2015 American Society of Plant Biologists. All rights reserved.
.
DOI:10.1186/s12864-016-3113-4URLPMID:5062844 [本文引用: 2]
Autophagy is a cellular degradation process that is highly evolutionarily-conserved in yeast, plants, and animals. In plants, autophagy plays important roles in regulating intracellular degradation and recycling of amino acids in response to nutrient starvation, senescence, and other environmental stresses. Foxtail millet (Setaria italica) has strong resistance to stresses and has been proposed as an ideal material for use in the study of the physiological mechanisms of abiotic stress tolerance in plants. Although the genome sequence of foxtail millet (Setaria italica) is available, the characteristics and functions of abiotic stress-related genes remain largely unknown for this species. A total of 37 putativeATG(autophagy-associated genes) genes in the foxtail millet genome were identified. Gene duplication analysis revealed that both segmental and tandem duplication events have played significant roles in the expansion of theATGgene family in foxtail millet. Comparative synteny mapping between the genomes of foxtail millet and rice suggested that theATGgenes in both species have common ancestors, as theirATGgenes were primarily located in similar syntenic regions. Gene expression analysis revealed the induced expression of 31SiATGgenes by one or more phytohormone treatments, 26SiATGgenes by drought, salt and cold, 24SiATGgenes by darkness and 25SiATGgenes by nitrogen starvation. Results of qRT-PCR showing that among 37SiATGgenes, the expression level ofSiATG8awas the highest after nitrogen starvation treatment 24 h, suggesting its potential role in tolerance to nutrient starvation. Moreover, the heterologous expression ofSiATG8ain rice improved nitrogen starvation tolerance. Compared to wild type rice, the transgenic rice performed better and had higher aboveground total nitrogen content when the plants were grown under nitrogen starvation conditions. Our results deepen understanding about the characteristics and functions ofATGgenes in foxtail millet and also identify promising new genetic resources that should be of use in future efforts to develop varieties of foxtail millet and other crop species that have resistance to nitrogen deficiency stress. The online version of this article (doi:10.1186/s12864-016-3113-4) contains supplementary material, which is available to authorized users.
[本文引用: 1]
DOI:10.1093/jxb/erq033URLPMID:20194926 [本文引用: 1]
The endosymbiont-derived Sec-dependent protein sorting pathway is essential for protein import into the thylakoid lumen and is important for the proper functioning of the chloroplast. Two loss-of-function mutants of cpSecA, the ATPase subunit of the chloroplast Sec translocation machinery, were analysed in Arabidopsis. The homozygous mutants were albino and seedling lethal under autotrophic conditions and remained dwarf and infertile with an exogenous carbon supply. They were subject to oxidative stress and accumulated superoxide under normal lighting conditions. Electron microscopy revealed that the chloroplast of the mutants had underdeveloped thylakoid structures. Histochemical GUS assay of the AtcpSecA::GUS transgenic plants confirmed that AtcpSecA was expressed in green organs in a light-inducible way. Real-time RT-PCR and microarray analysis revealed repressed transcription of nucleus- and chloroplast- encoded subunits of photosynthetic complexes, and induced transcription of chloroplast protein translocation machinery and mitochondrion-encoded respiratory complexes in the mutants. It is inferred that AtcpSecA plays an essential role in chloroplast biogenesis, the absence of which triggered a retrograde signal, eventually leading to a reprogramming of chloroplast and mitochondrial gene expression.
DOI:10.1146/annurev-arplant-042811-105441URLPMID:22242963 [本文引用: 1]
Abstract Plants have developed sophisticated mechanisms to survive when in unfavorable environments. Autophagy is a macromolecule degradation pathway that recycles damaged or unwanted cell materials upon encountering stress conditions or during specific developmental processes. Over the past decade, our molecular and physiological understanding of plant autophagy has greatly increased. Most of the essential machinery required for autophagy seems to be conserved from yeast to plants. Plant autophagy has been shown to function in various stress responses, pathogen defense, and senescence. Some of its potential upstream regulators have also been identified. Here, we describe recent advances in our understanding of autophagy in plants, discuss areas of controversy, and highlight potential future directions in autophagy research.
DOI:10.1104/pp.96.2.355URLPMID:16668193 [本文引用: 1]
Nitrogen partitioning among proteins in chloroplasts and mitochondria was examined in pea (Pisum sativum L.) and wheat (Triticum aestivum L.) grown hydroponically with different nitrogen concentrations. In pea leaves, chloroplast nitrogen accounted for 75 to 80% of total leaf nitrogen. We routinely found that 8% of total ribulose-1,5-bisphosphate carboxylase/oxygenase adhered to thylakoids during preparation and could be removed with Triton X-100. With this precaution, the ratio of stroma nitrogen increased from 53 to 61% of total leaf nitrogen in response to the nitrogen supply, but thylakoid nitrogen remained almost constant around 20% of total. The changes in the activities of the stromal enzymes and electron transport in response to the nitrogen supply reflected the nitrogen partitioning into stroma and thylakoids. On the other hand, nitrogen partitioning into mitochondria was appreciably smaller than that in chloroplasts, and the ratio of nitrogen allocated to mitochondria decreased with increasing leaf-nitrogen content, ranging from 7 to 4% of total leaf nitrogen. The ratio of mitochondrial respiratory enzyme activities to leaf-nitrogen content also decreased with increasing leaf-nitrogen content. These differences in nitrogen partitioning between chloroplasts and mitochondria were reflected in differences in the rates of photosynthesis and dark respiration in wheat leaves measured with an open gas-exchange system. The response of photosynthesis to nitrogen supply was much greater than that of dark respiration, and the CO(2) compensation point decreased with increasing leaf-nitrogen content.
DOI:10.1093/aob/mcq028URLPMID:2887065 [本文引用: 3]
Background: Productive agriculture needs a large amount of expensive nitrogenous fertilizers. Improving nitrogen use efficiency (NUE) of crop plants is thus of key importance. NUE definitions differ depending on whether plants are cultivated to produce biomass or grain yields. However, for most plant species, NUE mainly depends on how plants extract inorganic nitrogen from the soil, assimilate nitrate and ammonium, and recycle organic nitrogen. Efforts have been made to study the genetic basis as well as the biochemical and enzymatic mechanisms involved in nitrogen uptake, assimilation, and remobilization in crops and model plants. The detection of the limiting factors that could be manipulated to increase NUE is the major goal of such research.
DOI:10.1111/j.1438-8677.2008.00097.xURLPMID:18721309 [本文引用: 1]
A major challenge of modern agriculture is to reduce the excessive input of fertilisers and, at the same time, to improve grain quality without affecting yield. One way to achieve this goal is to improve plant nitrogen economy through manipulating nitrogen recycling, and especially nitrogen remobilisation, from senescing plant organs. In this review, the contribution of nitrogen remobilisation efficiency (NRE) to global nitrogen use efficiency (NUE), and tools dedicated to the determination of NRE are described. An overall examination of the physiological, metabolic and genetic aspects of nitrogen remobilisation is presented.
[本文引用: 1]
DOI:10.1104/pp.111.4.1233URLPMID:12226358 [本文引用: 1]
The response of tobacco (Nicotiana tabacum) suspension-cultured cells (BY-2) to nutrient starvation was investigated. When the cells that were grown in Murashige-Skoog medium containing 3% (w/v) sucrose were transferred to the same medium without sucrose, 30 to 45% of the intracellular proteins were degraded in 2 d. An analysis with sodium dodecyl sulfate-polyacrylamide gel electrophoresis showed that proteins were degraded nonselectively. With the same treatment, protease activity in the cell, which was measured at pH 5.0 using fluorescein thiocarbamoyl-casein as a substrate, increased 3- to 7-fold after 1 d. When the cysteine protease inhibitor (2S,3S)-trans-epoxysuccinyl-L-leucylamido-3-methyl-butane (10 M) was present in the starvation medium, both the protein degradation and the increase in the protease activity were effectively inhibited. Light microscopy analysis showed that many small spherical bodies accumulated in the perinuclear region of the cytosol 8 h after the start of the inhibitor treatment. These bodies were shown to be membrane-bound vesicles of 1 to 6 m in diameter that contained several particles. Quinacrine stained these vesicles and the central vacuole; thus, both organelles are acidic compartments. Cytochemical enzyme analysis using 1-naphthylphosphate and 尾-glycerophosphate as substrates showed that these vesicles contained an acid phosphatase(s). We suggest that these vesicles contribute to cellular protein degradation stimulated under sucrose starvation conditions.
[本文引用: 1]
DOI:10.1111/j.1365-313X.2005.02346.xURLPMID:15743448 [本文引用: 1]
Vacuolar compartments associated with leaf senescence and the subcellular localization of the senescence-specific cysteine-protease SAG12 (senescence-associated gene 12) were studied using specific fluorescent markers, the expression of reporter genes, and the analysis of high-pressure frozen/freeze-substituted samples. Senescence-associated vacuoles (SAVs) with intense proteolytic activity develop in the peripheral cytoplasm of mesophyll and guard cells in Arabidopsis and soybean. The vacuolar identity of these compartments was confirmed by immunolabeling with specific antibody markers. SAVs and the central vacuole differ in their acidity and tonoplast composition: SAVs are more acidic than the central vacuole and, whereas the tonoplast of central vacuoles is highly enriched in -TIP (tonoplast intrinsic protein), the tonoplast of SAVs lacks this aquaporin. The expression of a SAG12-GFP fusion protein in transgenic Arabidopsis plants shows that SAG12 localizes to SAVs. The analysis of Pro SAG 12 : GUS transgenic plants indicates that SAG12 expression in senescing leaves is restricted to SAV-containing cells, for example, mesophyll and guard cells. A homozygous sag12 Arabidopsis mutant develops SAVs and does not show any visually detectable phenotypical alteration during senescence, indicating that SAG12 is not required either for SAV formation or for progression of visual symptoms of senescence. The presence of two types of vacuoles in senescing leaves could provide different lytic compartments for the dismantling of specific cellular components. The possible origin and functions of SAVs during leaf senescence are discussed.
DOI:10.1093/jexbot/52.356.551URLPMID:11373304 [本文引用: 1]
Developing seeds are net importers of organic and inorganic nutrients. Nutrients enter seeds through the maternal vascular system at relatively high concentrations in the phloem. They exit importing sieve elements via interconnecting plasmodesmata and, during subsequent symplasmic passage, are sequestered into labile storage pools (vacuoles; starch). Transporters function to retrieve nutrients leaked to the seed apoplasm during symplasmic passage. Maternal cells responsible for nutrient release to the seed apoplasm are characteristically located at the maternal/filial interface. Their plasma membranes are enriched in transport proteins and, in some species, these cells are modified to a transfer cell morphology. Apoplasmic volumes of seeds are relatively small, but contain high concentrations of sugars, potassium and a range of amino acids. Sucrose and amino acids are taken up from the seed apoplasm by one to two cell layers of filial tissues that juxtapose the maternal tissues. The plasma membranes of the uptake cells are enriched in sucrose and amino acid/H+ transporters which co-localize with H+-ATPases. In some species, these cells are modified to a transfer cell morphology. High densities of plasmodesmata support symplasmic delivery of accumulated nutrients to underlying storage cells where polymer formation (starch, protein) takes place. Hexoses, resulting from sucrose hydrolysis and leakage to the seed apoplasm, are retrieved by hexose/H+ symporters.
DOI:10.1016/j.febslet.2007.04.013URLPMID:17466985 [本文引用: 1]
Nitrogen is an essential macronutrient for plant growth. Following uptake from the soil or assimilation within the plant, organic nitrogen compounds are transported between organelles, from cell to cell and over long distances in support of plant metabolism and development. These translocation processes require the function of integral membrane transporters. The review summarizes our current understanding of the molecular mechanisms of organic nitrogen transport processes, with a focus on amino acid, ureide and peptide transporters.
DOI:10.1111/j.1399-3054.2012.01574.xURLPMID:22242903 [本文引用: 1]
Senescence is the final developmental stage of every plant organ, which leads to cell death. It is a highly regulated process, involving differential gene expression and outstanding increment in the rate of protein degradation. Senescence-associated proteolysis enables the remobilization of nutrients, such as nitrogen (N), from senescent tissues to developing organs or seeds. In addition to the nutrient recycling function, senescence-associated proteases are also involved in the regulation of the senescence process. Nearly, all protease families have been associated with some aspects of plant senescence, and numerous reports addressing the new identification of senescence-associated proteases are published every year. Here, we provide an updated report with the most recent information published in the field, focusing on senescence-associated proteases presumably involved in N remobilization.
DOI:10.1093/jxb/ern244URLPMID:2576633 [本文引用: 1]
Eukaryotes contain a ubiquitous family of autophagy-associated Atg8 proteins. In animal cells, these proteins have multiple functions associated with growth, cancer, and degenerative diseases, but their functions in plants are still largely unknown. To search for novel functions of Atg8 in plants, the present report tested the effect of expression of a recombinant AtAtg8 protein, fused at its N-terminus to green fluorescent protein (GFP) and at its C-terminus to the haemagglutinin epitope tag, on the response of Arabidopsis thaliana plants to the hormones cytokinin and auxin as well as to salt and osmotic stresses. Expression of this AtAtg8 fusion protein modulates the effect of cytokinin on root architecture. Moreover, expression of this fusion protein also reduces shoot anthocyanin accumulation in response to cytokinin feeding to the roots, implying the participation of AtAtg8 in cytokinin-regulated root–shoot communication. External application of cytokinin leads to the formation of novel GFP–AtAtg8-containing structures in cells located in the vicinity of the root vascular system, which are clearly distinct in size and dynamic movement from the GFP–AtAtg8-containing autophagosome-resembling structures that were observed in root epidermis cells. Expression of the AtAtg8 fusion construct also renders the plants more sensitive to a mild salt stress and to a lesser extent to a mild osmotic stress. This sensitivity is also associated with various changes in the root architecture, which are morphologically distinct from those observed in response to cytokinin. The results imply multiple functions for AtAtg8 in different root tissues that may also be regulated by different mechanisms.
.
[本文引用: 1]
.
[本文引用: 3]
DOI:10.1104/pp.108.130013URL [本文引用: 1]
Chloroplasts contain approximately 80% of total leaf nitrogen and represent a major source of recycled nitrogen during leaf senescence. While bulk degradation of the cytosol and organelles in plants is mediated by autophagy, its role in chloroplast catabolism is largely unknown. We investigated the effects of autophagy disruption on the number and size of chloroplasts during senescence. When leaves were individually darkened, senescence was promoted similarly in both wild-type Arabidopsis (Arabidopsis thaliana) and in an autophagy-defective mutant, atg4a4b-l. The number and size of chloroplasts decreased in darkened leaves of wild type, while the number remained constant and the size decrease was suppressed in atg4a4b-2. When leaves of transgenic plants expressing stroma-targeted DsRed were individually darkened, a large accumulation of fluorescence in the vacuolar lumen was observed. Chloroplasts exhibiting chlorophyll fluorescence, as well as Rubisco-containing bodies, were also observed in the vacuole. No accumulation of stroma-targeted DsRed, chloroplasts, or Rubisco-containing bodies was observed in the vacuoles of the autophagy-defective mutant. We have succeeded in demonstrating chloroplast autophagy in living cells and provide direct evidence of chloroplast transportation into the vacuole.
DOI:10.1007/s11120-004-2080-9URLPMID:16143854 [本文引用: 1]
During their life cycle, plants must be able to adapt to wide variations in the supply of soil nitrogen (N). Changes in N availability, and in the relative concentrations of NO 3 61 and NH 4 + , are known to have profound regulatory effects on the N uptake systems in the root, on C and N metabolism throughout the plant, and on root and shoot morphology. Optimising the plant’s responses to fluctuations in the N supply requires co-ordination of the pathways of C and N assimilation, as well as establishment of the appropriate allocation of resources between root and shoot growth. Achieving this integration of responses at the whole plant level implies long-distance signaling mechanisms that can communicate information about the current availability of N from root-to-shoot, and information about the C/N status of the shoot in the reverse direction. In this review we will discuss recent advances which have contributed to our understanding of these long-range signaling pathways.
.
DOI:10.3389/fpls.2016.00720URLPMID:4879346 [本文引用: 1]
Nutrient deficiencies restrict apple (Malussp.) tree growth and productivity in Northwest China. The process of autophagy, a conserved degradation pathway in eukaryotic cells, has important roles in nutrient-recycling and helps improve plant performance during periods of nutrient-starvation. Little is known about the functioning of autophagy-related genes (ATGs) in apple. In this study, one of theATG8gene family membersMdATG8iwas isolated fromMalus domestica. MdATG8i has conserved putative tubulin binding sites and ATG7 interaction domains. A 1865-bp promoter region cloned from apple genome DNA was predicated to havecis-regulatory elements responsive to light, environmental stresses, and hormones. MdATG8i transcriptions were induced in response to leaf senescence, nitrogen depletion, and oxidative stress. At cellular level, MdATG8i protein was expressed in the nucleus and cytoplasm of onion epidermal cells. Yeast two-hybrid tests showed that MdATG8i could interact with MdATG7a and MdATG7b. InArabidopsis, its heterologous expression was associated with enhanced vegetative growth, leaf senescence, and tolerance to nitrogen- and carbon-starvation.MdATG8i-overexpressing rin apple callus lines also displayed improved tolerance to nutrient-limited conditions. Our results demonstrate that MdATG8i protein could function in autophagy in a conserved way, as a positive regulator in the response to nutrient-starvation.
[本文引用: 2]
.
DOI:10.1093/dnares/dsr024URLPMID:21795261 [本文引用: 1]
Autophagy is an intracellular degradation process for recycling macromolecules and organelles. It plays important roles in plant development and in response to nutritional demand, stress, and senescence. Organisms from yeast to plants contain many autophagy-associated genes (ATG). In this study, we found that a total of 33 ATG homologues exist in the rice [Oryza sativa L. (Os)] genome, which were classified into 13 ATG subfamilies. Six of them are alternatively spliced genes. Evolutional analysis showed that expansion of 10 OsATG homologues occurred via segmental duplication events and that the occurrence of these OsATG homologues within each subfamily was asynchronous. The Ka/Ks ratios suggested purifying selection for four duplicated OsATG homologues and positive selection for two. Calculating the dates of the duplication events indicated that all duplication events might have occurred after the origin of the grasses, from 21.43 to 66.77 million years ago. Semi-quantitative RT???PCR analysis and mining the digital expression database of rice showed that all 33 OsATG homologues could be detected in at least one cell type of the various tissues under normal or stress growth conditions, but their expression was tightly regulated. The 10 duplicated genes showed expression divergence. The expression of most OsATG homologues was regulated by at least one treatment, including hormones, abiotic and biotic stresses, and nutrient limitation. The identification of OsATG homologues showing constitutive expression or responses to environmental stimuli provides new insights for in-depth characterization of selected genes of importance in rice.<br>
.
[本文引用: 2]
DOI:10.1016/bs.ircmb.2015.07.005URLPMID:26614870 [本文引用: 1]
Autophagy is a degradation pathway that is conserved throughout eukaryotic organisms and plays important roles in the tolerance of abiotic and biotic stresses. It functions as a housekeeping process to remove unwanted cell components under normal conditions, and is induced during stress and senescence to break down damaged cellular contents and to recycle materials. The target components are engulfed into specialized transport structures termed autophagosomes and are subsequently delivered to the vacuole for degradation. Here, we review milestones in the study of autophagy in plants, discuss recent advances in our understanding of the mechanism and physiological roles of plant autophagy, and highlight potential future directions of research.
DOI:10.1080/15548627.2015.1107692URLPMID:26565778 [本文引用: 1]
Macroautophagy (hereafter autophagy) is one of the major degradation systems in eukaryotic cells, and its dysfunction may result in diseases ranging from neurodegeneration to cancer. Although most of the autophagy-related (Atg) proteins that function in this pathway were first identified in yeast, many were subsequently shown to have homologs in higher eukaryotes including humans, and the overall mechanism of autophagy is highly conserved. The most prominent feature of autophagy is the formation of a double-membrane sequestering compartment, the phagophore; this transient organelle surrounds part of the cytoplasm and matures into an autophagosome, which subsequently fuses with the vacuole or lysosome to allow degradation of the cargo. Much attention has focused on the process involved in phagophore nucleation and expansion, but many questions remain. Here, we identified the yeast protein Icy2, which we now name Atg41, as playing a role in autophagosome formation. Atg41 interacts with the transmembrane protein Atg9, a key component involved in autophagosome biogenesis, and both proteins display a similar localization profile. Under autophagy-inducing conditions the expression level of Atg41 increases dramatically and is regulated by the transcription factor Gcn4. This work provides further insight into the mechanism of Atg9 function and the dynamics of sequestering membrane formation during autophagy.
DOI:10.1093/pcp/pcs099URLPMID:22764279 [本文引用: 1]
Autophagy is an evolutionarily conserved intracellular process for the vacuolar degradation of cytoplasmic components. There is no doubt that autophagy is very important to plant life, especially because plants are immobile and must survive in environmental extremes. Early studies of autophagy provided our first insights into the structural characteristics of the process in plants, but for a long time the molecular mechanisms and the physiological roles of autophagy were not understood. Genetic analyses of autophagy in the yeast Saccharomyces cerevisiae have greatly expanded our knowledge of the molecular aspects of autophagy in plants as well as in animals. Until recently our knowledge of plant autophagy was in its infancy compared with autophagy research in yeast and animals, but recent efforts by plant researchers have made many advances in our understanding of plant autophagy. Here I will introduce an overview of autophagy in plants, present current findings and discuss the physiological roles of self-degradation.
DOI:10.1105/tpc.104.025395URLPMID:15494556 [本文引用: 1]
Autophagy is an intracellular process for vacuolar degradation of cytoplasmic components. Thus far, plant autophagy has been studied primarily using morphological analyses. A recent genome-wide search revealed significant conservation among autophagy genes (ATGs) in yeast and plants. It has not been proved, however, that Arabidopsis thalianaATG genes are required for plant autophagy. To evaluate this requirement, we examined the ubiquitination-like Atg8 lipidation system, whose component genes are all found in the Arabidopsis genome. In Arabidopsis, all nine ATG8 genes and two ATG4 genes were expressed ubiquitously and were induced further by nitrogen starvation. To establish a system monitoring autophagy in whole plants, we generated transgenic Arabidopsis expressing each green fluorescent protein TG8 fusion (GFP-ATG8). In wild-type plants, GFP-ATG8s were observed as ring shapes in the cytoplasm and were delivered to vacuolar lumens under nitrogen-starved conditions. By contrast, in a T-DNA insertion double mutant of the ATG4s (atg4a4b-1), autophagosomes were not observed, and the GFP-ATG8s were not delivered to the vacuole under nitrogen-starved conditions. In addition, we detected autophagic bodies in the vacuoles of wild-type roots but not in those of atg4a4b-1 in the presence of concanamycin A, a V-ATPase inhibitor. Biochemical analyses also provided evidence that autophagy in higher plants requires ATG proteins. The phenotypic analysis of atg4a4b-1 indicated that plant autophagy contributes to the development of a root system under conditions of nutrient limitation.
细胞自噬在植物细胞程序性死亡中的作用
1
2016
... 细胞自噬是植物中参与营养物质的重新分配、清除受损的蛋白和细胞器及响应生物和非生物胁迫等过程的一种重要的降解途径(
活性氧对植物自噬调控的研究进展
1
2012
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
植物细胞自噬研究进展
1
2018
... 细胞自噬是植物中参与营养物质的重新分配、清除受损的蛋白和细胞器及响应生物和非生物胁迫等过程的一种重要的降解途径(
细胞自噬在植物碳氮营养中作用的研究进展
1
2014
... 细胞自噬是植物中参与营养物质的重新分配、清除受损的蛋白和细胞器及响应生物和非生物胁迫等过程的一种重要的降解途径(
Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris
0
1949
Autophagy, plant senescence, and nutrient recycling
2
2014
... 植物无法选择自身所需的营养环境, 只能依靠根际吸收有限的矿质营养维持生长发育, 因此植物必须有效代谢并循环利用体内的营养物质(
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
Monodansylcadaverine (MDC) is a specific in vivo marker for autophagic vacuoles
1
1995
... 细胞自噬是植物中参与营养物质的重新分配、清除受损的蛋白和细胞器及响应生物和非生物胁迫等过程的一种重要的降解途径(
A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding
1
1976
... 总叶绿素含量测定参照Arnon (1949)的方法.可溶性蛋白含量测定参照Bradford法(
High-resolution temporal profiling of transcripts during Arabidopsis leaf senescence reveals a distinct chronology of processes and regulation
1
2011
... 前人研究表明, 营养饥饿能够诱导植物自噬发生(
Exploring NUE in crops and in Arabidopsis ideotypes to improve yield and seed quality
1
2012
... 氮素是植物体内最重要的营养元素, 是植物生长发育过程中必需的大量元素之一, 是氨基酸、蛋白质、核酸、叶绿素和激素等的重要组分(
Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana
1
1998
... 以水稻cDNA为模板, 利用常规PCR方法扩增Os- ATG8b基因的CDS序列(360 bp), 分别在上、下游引物中引入Nco I和BstE II的酶切位点, 通过Nco I/BstE II双酶切插入到双元表达载体pCAMBIA1301, 完成携带35S-OsATG8b的植物表达载体构建.以水稻基因组DNA为模板, 扩增OsATG8b基因启动子序列(1 988 bp), 分别在上、下游引物中引入EcoR I和Nco I的酶切位点, 通过EcoR I/Nco I双酶切插入到双元表达载体pCAMBIA1301, 完成ProOsATG8b-GUS双元表达载体的构建.使用农杆菌(Agrobacterium tumefaciens) GV3101介导的花序浸泡法(
Visualization of autophagy in Arabidopsis using the fluorescent dye monodansylcadaverine and a GFP-AtATG8e fusion protein
1
2005
... 细胞自噬是植物中参与营养物质的重新分配、清除受损的蛋白和细胞器及响应生物和非生物胁迫等过程的一种重要的降解途径(
The machi- nery of macroautophagy
1
2014
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
Can less yield more? Is reducing nutrient input into the environment compatible with maintaining crop production?
1
2004
... 植物通过硝酸盐转运因子(AtNRT)和铵盐转运因子(AtAMTs)转运根系吸收的硝态氮与铵态氮(
Physiological and metabolic consequences of autophagy deficiency for the management of nitrogen and protein resources in Arabidopsis leaves depending on nitrate availability
1
2013
... 细胞自噬是植物中参与营养物质的重新分配、清除受损的蛋白和细胞器及响应生物和非生物胁迫等过程的一种重要的降解途径(
Autophagy machi- nery controls nitrogen remobilization at the whole-plant level under both limiting and ample nitrate conditions in Arabidopsis
2
2012
... 细胞自噬是植物中参与营养物质的重新分配、清除受损的蛋白和细胞器及响应生物和非生物胁迫等过程的一种重要的降解途径(
... ).自噬相关基因缺失突变体在营养饥饿时表现出明显的生长发育迟缓、加速衰老及存活率下降等表型(
Mobilization of rubisco and stroma-localized fluorescent proteins of chloroplasts to the vacuole by an ATG gene-dependent autophagic process
2
2008
... 在植物发育的后期阶段, 叶片蛋白(尤其是叶绿体蛋白)被迅速降解, 从而输出大量可再利用的氮, 其释放的游离氨基酸及其它化合物被转运至新生的生殖器官和储存器官中(如新生的叶片或种子) (
... 细胞自噬是植物中参与营养物质的重新分配、清除受损的蛋白和细胞器及响应生物和非生物胁迫等过程的一种重要的降解途径(
Autophagy contributes to nighttime energy availability for growth in Arabidopsis
1
2013
... Yoshimoto等(2009)研究表明, 拟南芥自噬缺失突变体atg5的早衰表型是由水杨酸过量积累造成的, 在拟南芥自噬突变体atg5-2中过表达细菌的水杨酸羟化酶NahG基因, 或者在双突变体atg5-2/sid2 (sid2为水杨酸合成缺陷突变体)中, atg5-2的叶片早衰表型均得到抑制.我们发现OsATG8b启动子响应水杨酸处理, 其活性明显增强(
Establishment of monito- ring methods for autophagy in rice reveals autophagic recycling of chloroplasts and root plastids during energy limitation
0
2015
In winter wheat ( Triticum aestivum L.), post-anthesis nitrogen uptake and remobilisation to the grain correlates with agronomic traits and nitrogen physiological markers
1
2007
... 氮素是植物体内最重要的营养元素, 是植物生长发育过程中必需的大量元素之一, 是氨基酸、蛋白质、核酸、叶绿素和激素等的重要组分(
A holistic view of nitrogen acquisition in plants
1
2011
... 氮素是植物体内最重要的营养元素, 是植物生长发育过程中必需的大量元素之一, 是氨基酸、蛋白质、核酸、叶绿素和激素等的重要组分(
Plant nitrogen assimilation and its regulation: a complex puzzle with missing pieces
1
2015
... 植物无法选择自身所需的营养环境, 只能依靠根际吸收有限的矿质营养维持生长发育, 因此植物必须有效代谢并循环利用体内的营养物质(
a). Autophagic recycling plays a central role in maize nitrogen remobilization
3
2015
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
... 细胞自噬是植物中参与营养物质的重新分配、清除受损的蛋白和细胞器及响应生物和非生物胁迫等过程的一种重要的降解途径(
... ;
Genome-wide analysis of autophagy-associated genes in foxtail millet ( Setaria italica L.) and characterization of the function of SiATG8a in conferring tolerance to nitrogen starvation in rice
2
2016
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
... 前人研究表明, 营养饥饿能够诱导植物自噬发生(
b). Overexpression of the autophagy-related gene
1
2015
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
Cpseca, a thylakoid protein translocase subunit, is essential for photosynthetic development in Arabidopsis
1
2010
... 分别取全氮和缺氮处理14天的35SOsATG8b转基因及野生型拟南芥的叶片和根系, 液氮速冻后存于-80°C冰箱, 提取总RNA并反转录得到cDNA.以TIP41为内参基因对所有样品进行归一化处理, 利用荧光定量RT-PCR检测氮代谢相关基因的相对表达水平变化.总RNA提取、cDNA合成和荧光定量RT-PCR分析参照文献所述(
Autophagy: pathways for self-eating in plant cells
1
2012
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
Effects of nitrogen nutrition on nitrogen partitioning between chloroplasts and mitochondria in pea and wheat
1
1991
... 植物无法选择自身所需的营养环境, 只能依靠根际吸收有限的矿质营养维持生长发育, 因此植物必须有效代谢并循环利用体内的营养物质(
Nitrogen uptake, assimilation and remobilization in plants: challenges for sustainable and productive agriculture
3
2010
... 植物无法选择自身所需的营养环境, 只能依靠根际吸收有限的矿质营养维持生长发育, 因此植物必须有效代谢并循环利用体内的营养物质(
... 在植物发育的后期阶段, 叶片蛋白(尤其是叶绿体蛋白)被迅速降解, 从而输出大量可再利用的氮, 其释放的游离氨基酸及其它化合物被转运至新生的生殖器官和储存器官中(如新生的叶片或种子) (
... 植物通过硝酸盐转运因子(AtNRT)和铵盐转运因子(AtAMTs)转运根系吸收的硝态氮与铵态氮(
Leaf nitrogen remobilisation for plant development and gr- ain filling
1
2008
... 在植物发育的后期阶段, 叶片蛋白(尤其是叶绿体蛋白)被迅速降解, 从而输出大量可再利用的氮, 其释放的游离氨基酸及其它化合物被转运至新生的生殖器官和储存器官中(如新生的叶片或种子) (
1
... 植物通过硝酸盐转运因子(AtNRT)和铵盐转运因子(AtAMTs)转运根系吸收的硝态氮与铵态氮(
Autophagy in tobacco suspension-cultured cells in response to sucrose starvation
1
1996
... 前人研究表明, 营养饥饿能够诱导植物自噬发生(
Molecular dissection of autophagy: two ubiquitin-like systems
1
2001
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
Senescence-associated vacuoles with intense proteolytic activity develop in leaves of Arabidopsis and soybean
1
2005
... 在植物发育的后期阶段, 叶片蛋白(尤其是叶绿体蛋白)被迅速降解, 从而输出大量可再利用的氮, 其释放的游离氨基酸及其它化合物被转运至新生的生殖器官和储存器官中(如新生的叶片或种子) (
Compartmentation of transport and transfer events in developing seeds
1
2001
... 在植物发育的后期阶段, 叶片蛋白(尤其是叶绿体蛋白)被迅速降解, 从而输出大量可再利用的氮, 其释放的游离氨基酸及其它化合物被转运至新生的生殖器官和储存器官中(如新生的叶片或种子) (
Transporters for uptake and allocation of organic nitrogen compounds in plants
1
2007
... 植物通过硝酸盐转运因子(AtNRT)和铵盐转运因子(AtAMTs)转运根系吸收的硝态氮与铵态氮(
Senescence-associated proteases in plants
1
2012
... 在植物发育的后期阶段, 叶片蛋白(尤其是叶绿体蛋白)被迅速降解, 从而输出大量可再利用的氮, 其释放的游离氨基酸及其它化合物被转运至新生的生殖器官和储存器官中(如新生的叶片或种子) (
An autophagy-associated Atg8 protein is involved in the responses of Arabidopsis seedlings to hormonal controls and abiotic stresses
1
2008
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
Autophagic nutrient recycling in Arabidopsis directed by the ATG8 and ATG12 conjugation pathways
1
2005
... 前人研究表明, 营养饥饿能够诱导植物自噬发生(
Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae
0
1993
Autophagy supports biomass production and nitrogen use efficiency at the vegetative stage in rice
3
2015
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
... 细胞自噬是植物中参与营养物质的重新分配、清除受损的蛋白和细胞器及响应生物和非生物胁迫等过程的一种重要的降解途径(
... ;
Autophagy plays a role in chloroplast degradation during senescence in individually darkened leaves
1
2009
... 在植物发育的后期阶段, 叶片蛋白(尤其是叶绿体蛋白)被迅速降解, 从而输出大量可再利用的氮, 其释放的游离氨基酸及其它化合物被转运至新生的生殖器官和储存器官中(如新生的叶片或种子) (
Signaling mechanisms integrating root and shoot responses to ch- anges in the nitrogen supply
1
2005
... 细胞自噬是植物中参与营养物质的重新分配、清除受损的蛋白和细胞器及响应生物和非生物胁迫等过程的一种重要的降解途径(
Characterization of an autophagy-related gene MdATG8i from apple
1
2016
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
Autophagy contributes to leaf starch degradation
2
2013
... 将苗龄7天的转基因和野生型拟南芥幼苗置于含有1 μmol·L-1刀豆素(ConA) (SIGMA)的缺氮培养液中处理12小时, 将叶片取下立即浸没于单丹磺酰戊二胺(monodansylcadaverine, MDC)染液(SIGMA)中进行染色, 通过激光共聚焦显微镜(Zeiss LSM 710)观察自噬体荧光信号,
... Yoshimoto等(2009)研究表明, 拟南芥自噬缺失突变体atg5的早衰表型是由水杨酸过量积累造成的, 在拟南芥自噬突变体atg5-2中过表达细菌的水杨酸羟化酶NahG基因, 或者在双突变体atg5-2/sid2 (sid2为水杨酸合成缺陷突变体)中, atg5-2的叶片早衰表型均得到抑制.我们发现OsATG8b启动子响应水杨酸处理, 其活性明显增强(
Genome-wide identification, classification, and expression analysis of autophagy-associated gene homologues in rice ( Oryza sativa L.)
1
2011
... 前人研究表明, 营养饥饿能够诱导植物自噬发生(
Heterologous expression of ATG8c from soybean confers tolerance to nitrogen deficiency and increases yield in Arabidopsis
2
2012
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
... 前人研究表明, 营养饥饿能够诱导植物自噬发生(
New insight into the mechanism and function of autophagy in plant cells
1
2015
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
Atg41/Icy2 regulates autophagosome formation
1
2015
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
Beginning to understand autophagy, an intracellular self-degradation system in plants
1
2012
... 自噬是真核生物中一种保守的基本生物学过程, 正常情况下用于维持细胞稳态.当植物生长发育受到胁迫或营养饥饿时, 细胞通过自噬途径降解一些不需要的大分子物质或受损的细胞器来循环再利用自身养分以维持正常生命活动(
Processing of ATG8s, ubiquitin-like proteins, and their deconjugation by ATG4s are essential for plant autophagy
1
2004
... 细胞自噬是植物中参与营养物质的重新分配、清除受损的蛋白和细胞器及响应生物和非生物胁迫等过程的一种重要的降解途径(
Autophagy negatively regulates cell death by controlling NPR1-dependent salicylic acid signaling during senescence and the innate immune response in Arabidopsis
0
2009
备案号: 京ICP备16067583号-21
版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由北京玛格泰克科技发展有限公司设计开发

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}