 ,1,*
,1,*Research Progress into the Function of Purple Acid Phosphatase Gene Family in Plants
Ming Wei1,2, Xinwei Wang1, Bo Chen1, Chengwei Song1, Liang Du1, Jianwei Xiao1, Jinxing Lin,1,*通讯作者:
收稿日期:2018-02-9接受日期:2018-05-23网络出版日期:2019-01-30
| 基金资助: |
Corresponding authors:
Received:2018-02-9Accepted:2018-05-23Online:2019-01-30

摘要
关键词:
Abstract
Keywords:
PDF (6454KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
魏铭, 王鑫伟, 陈博, 宋程威, 杜亮, 肖建伟, 林金星. 植物紫色酸性磷酸酶基因家族功能研究进展. 植物学报, 2019, 54(1): 93-101 doi:10.11983/CBB18044
Wei Ming, Wang Xinwei, Chen Bo, Song Chengwei, Du Liang, Xiao Jianwei, Lin Jinxing.
磷是植物生长发育过程中必需的六大营养元素之一, 对植物新陈代谢和正常生长发育有着极其重要的作用。磷元素不仅参与植物细胞内许多化合物的生物合成, 蛋白磷酸化和去磷酸化等生理生化过程, 而且对植物生长发育及作物产量形成等也有一定的影响(James and David, 1992; Hammond et al., 2004)。尽管土壤中含有大量的磷, 但主要以磷酸酯和磷酸酐等有机形式存在, 不能直接被植物吸收和利用, 因此土壤中有效磷含量不足, 限制着植物的生长发育(Vance et al., 2003; 刘涛等, 2016)。植物在缺磷状态下, 会表现出茎短而细、叶片基部变黄、植株矮小、生长延缓、分枝或分蘖减少、种子小且不饱满等缺素症状。农业生产上普遍采用外施磷肥来提高植物对磷素的吸收利用, 这不仅增加了农业生产成本, 而且施肥过量还会污染水环境, 导致水体富营养化, 同时加剧磷资源的消耗(Vance, 2001; Cakmak, 2002; Smith and Schindler, 2009)。因此, 通过基因工程手段对植物进行改良, 研究植物磷营养代谢的生理学和分子生物学机制, 深入挖掘植物自身磷吸收利用潜力, 对节约磷矿资源、提高作物产量和品质以及保护生态环境具有重要意义(周志高等, 2005)。
紫色酸性磷酸酶(purple acid phosphatases, PAPs)属于双核金属脱氢酶(binuclear metallohydro- lase)家族, 是一种广泛存在于动物和植物体内的酸性磷酸酶类。在植物体的弱酸性(pH4-7)条件下, PAPs能够催化水解磷酸单酯和酸酐类有机物(如ATP、ADP和糖脂)并释放出无机磷, 供植物吸收利用, 从而提高植物对磷的利用率(Olczak et al., 2003; 卢坤等, 2010)。Zimmermann等(2004)研究表明, 在低磷胁迫下, 植物体内和根际分泌的PAPs活性均显著提高, 因此该酶能够分泌到植物细胞外活化植物根际周围的有机磷, 从而促进植株体内磷素的再循环利用。植物PAPs的成功分离对研究植物磷营养代谢机制具有重要意义。近年来, 在植物中已鉴定出许多PAPs编码基因。本文将系统论述植物中紫色酸性磷酸酶的结构及其家族成员的分类, 并着重介绍紫色酸性磷酸酶生物学功能的研究进展。
1 植物PAPs的发现及其结构
1998年, Hiroshi等用同位素标记法, 发现低磷会诱导浮萍(Spirodela oligorrhiza)产生57 kDa的磷脂酰肌醇锚定磷酸酶, 该酶含有Fe3+-Mn2+双核金属中心, 活性受到酒石酸抑制。这是首次在高等植物中发现紫色酸性磷酸酶(Nakazato et al., 1998), 由此揭开了植物PAPs研究的序幕。之后, 在单子叶和双子叶植物中均已鉴定出PAPs (Li et al., 2002; Zhang et al., 2011)。紫色酸性磷酸酶因其在溶液中表现出特殊的紫色或粉色而命名, 而呈现粉紫色是由发色基团铁离子和酪氨酸之间的电子转移所致。PAPs的催化位点和结构域高度保守, 其结构的基本特征在于含有5个保守基序(β/α/β/α/β)、7个结合金属离子的氨基酸残基(DXG/GDXXY/GNH(D/E)/VXXH/GHXH)和1个金属离子的双核中心(Schenk et al., 2000b)。菜豆(Pha- seolus vulgaris)紫色酸性磷酸酶(KBPAP)的三级结构见图1。其三维大小为40×60×75A, 二聚体呈现出心形结构, 每个亚基具有1个N端结构域(120个氨基酸残基), 1个C端结构域(210个氨基酸残基)和Fe-Zn金属离子中心(Strater et al., 1995)。植物、哺乳动物和细菌PAPs的催化结构域都含有“三明治”形状的β/α/β/α/β基序。植物PAPs的双核金属中心一般由Fe3+-Zn2+或Fe3+-Mn2+构成, 而动物PAPs的双核金属中心一般由Fe3+-Fe2+构成(Schenk et al., 2008; Jarenmark et al., 2011)。
图1
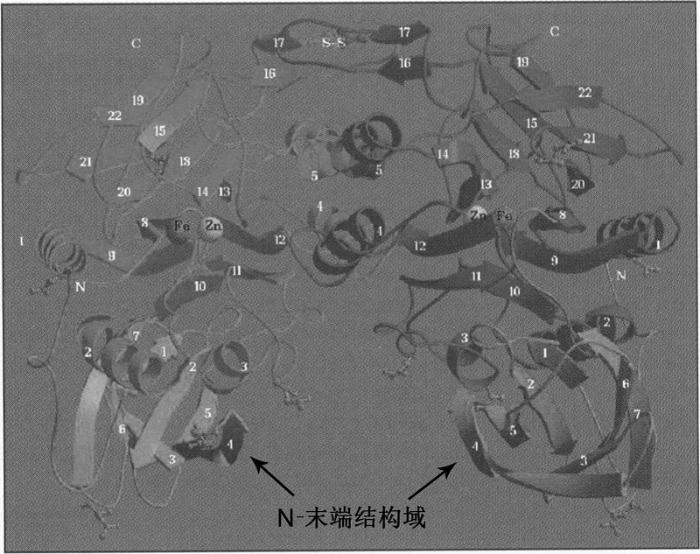 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1菜豆紫色酸性磷酸酶(KBPAP)的三维结构(改自Strater et al., 1995)
Figure 13D-structure of KBPAP of Phaseolus vulgaris (modified from Strater et al., 1995)
2 PAPs的分类及植物PAPs家族分析
根据紫色酸性磷酸酶基因编码蛋白的结构和分子量大小, 可将PAPs蛋白成员分为2类: 一类为大分子量PAPs, 其单体亚基分子量约为55 kDa; 另一类为小分子量PAPs, 其单体亚基约为35 kDa。植物大分子量PAPs与真菌的同源蛋白进化关系较近; 而植物小分子量PAPs与动物的同源蛋白进化关系较近, 从而反映PAPs家族成员在结构和功能上进化出的多样性。2.1 植物大分子量PAPs
大分子量的紫色酸性磷酸酶(high molecular weight PAPs, HMW PAPs)通常以同源二聚体的形式存在, 其单体分子量约为55 kDa, 此类PAPs在植物中所占比例较大, 并且与真菌和分枝杆菌的此类蛋白之间同源性较高(Schenk et al., 2000b)。菜豆中克隆出的KBPAP (检索号P80366)分子量为111 kDa, 属于大分子量PAP。KBPAP是1个二聚体酶, 包含2个糖基化的亚基, 2个亚基通过二硫键共价相连。每个亚基包含432个氨基酸残基和5个N-糖基化的天冬酰胺残基, 并且同时具有N-端和C-端2个结构域。C-端结构域分子量较大, 具有催化功能; N-端结构域较小, 由2个β-折叠组成。每个β-折叠由3个反向平行的β-链构成, 且不具有金属磷酸酶催化活性(Klabunde et al., 1994, 1995)。
2.2 植物小分子量PAPs
小分子量的紫色酸性磷酸酶(low molecular weight PAPs, LMW PAPs)以单聚体形式存在, 与哺乳动物和蓝藻体内的小分子量PAPs极为相似, 单体分子量一般约为35 kDa, 在结构上仅保留了典型的C-端结构域(Schenk et al., 2000a, 2000b)。菜豆根中提取的PvPAP3, 分子量为34 kDa, 属小分子量PAPs, 具有较高的热稳定性, 作用底物为ATP。PvPAP3定位于细胞质膜。低磷条件下, PvPAP3可在大豆(Glycine max)叶片和根中诱导表达, 并在磷高效基因型中表达量迅速增加(Liang et al., 2010)。马铃薯(Solanum tuberosu) StPAP1基因编码LMW PAPs, 类似于哺乳动物的PAPs, 在茎和根部高表达并对低磷环境敏感; 而StPAP2和StPAP3编码2种典型的植物HMW PAPs, 分别在茎和根部受低磷诱导表达(Zimmer- mann et al., 2004)。2.3 PAPs家族及其进化关系
目前, PAPs已在多种植物中被鉴定和分离, 通过对拟南芥(Arabidopsis thaliana)和大豆等植物中的PAPs进行系统研究, 发现拟南芥中存在29个PAPs基因(Li et al., 2002), 大豆中存在35个PAPs基因(Li et al., 2012), 水稻(Oryza sativa)中存在26个PAPs基因(Zhang et al., 2011)。将这些植物PAPs与已报道的烟草(Nicotiana tabacum)、玉米(Zea mays)、小麦(Triticum aestivum)和菜豆等物种PAPs的氨基酸序列比对, 构建进化树进行聚类分析(Tian and Liao, 2015), 可将其分成4个家族(图2)。图2
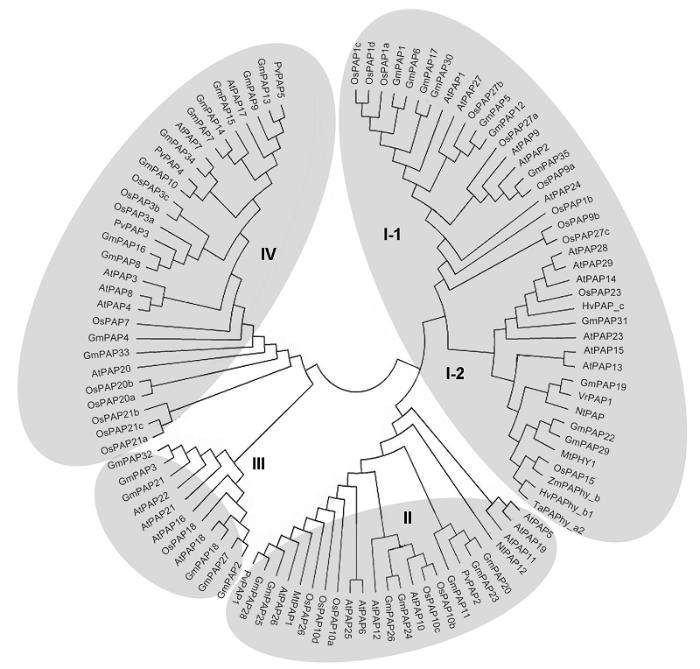 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2植物PAPs物种间的进化关系(改自Tian and Liao, 2015)
At: 拟南芥; Os: 水稻; Gm: 大豆; Mt: 蒺藜苜蓿; Nt: 烟草; Zm: 玉米; Hv: 大麦; Ta: 小麦; Vr: 绿豆; Pv: 菜豆
Figure 2Phylogenetic relationships of plant PAPs (modified from Tian and Liao, 2015)
At: Arabidopsis thaliana; Os: Oryza sativa; Gm: Glycine max; Mt: Medicago truncatula; Nt: Nicotiana tabacum; Zm: Zea mays; Hv: Hordeum vulgare; Ta: Triticum aestivum; Vr: Vigna radiata; Pv: Phaseolus vulgaris
PAPs家族I所包含的种类最多, 该家族可进一步分为I-1和I-2两个亚家族。I-1亚家族成员已被证实参与植物磷和碳代谢途径(Hur et al., 2010; Sun et al., 2012a)。目前, 已报道的I-2亚家族蛋白均具有植酸酶活性(Zhang et al., 2008; Kuang et al., 2009)。PAPs家族II是一个多功能家族, 参与植物抗病、磷饥饿诱导和细胞壁合成等多种生物学过程(Kaida et al., 2010; Robinson et al., 2012a; Ravichandran et al., 2013)。PAPs家族III和IV成员相关研究报道较少, 已鉴定的GmPAP3和PvPAP3与植物响应非生物逆境胁迫和磷利用效率有关(Li et al., 2008; Liang et al., 2010)。
3 植物PAPs的生物学功能
紫色酸性磷酸酶在植物各个组织均有分布, 参与各种生命活动(Zhu et al., 2005)。多数PAPs蛋白能够非特异性水解很多含磷脂键的化合物(包括ATP、PEP和植酸等), 释放磷酸基团(Olczak et al., 2003)。PAPs在植物不同生长发育过程中, 具有多种生物学功能, 具体功能由其在植物体内作用的对象决定。近几年, 越来越多的植物紫色酸性磷酸酶被鉴定和克隆出来。我们对植物PAPs的生物学功能进行了总结, 详见表1。Table 1
表1
表1植物PAP的生物学特性和功能(改自Tian and Liao, 2015)
Table 1
| 蛋白 | 物种 | 功能 | 参考文献 |
|---|---|---|---|
| AtPAP2 | 拟南芥 | 碳代谢 | Sun et al., 2012a |
| AtPAP10 | 拟南芥 | 参与初生壁合成 | Kaida et al., 2003 |
| AtPAP12 | 拟南芥 | 磷代谢 | Wang et al., 2014 |
| AtPAP15 | 拟南芥 | 响应非生物逆境胁迫 | Zhang et al., 2008 |
| AtPAP25 | 拟南芥 | 磷代谢 | Del Vecchio et al., 2014 |
| AtPAP26 | 拟南芥 | 磷代谢 | Wang et al., 2014 |
| GmPAP3 | 大豆 | 响应非生物逆境胁迫 | Li et al., 2008 |
| GmPAP4 | 大豆 | 响应非生物逆境胁迫 | Kong et al., 2014 |
| NtPAP12 | 烟草 | 参与初生壁合成 | Kaida et al., 2010 |
| PvPAP3 | 菜豆 | 磷代谢 | Liang et al., 2010 |
| StPAP1 | 马铃薯 | 磷代谢 | Zimmermann et al., 2004 |
新窗口打开|下载CSV
3.1 调节植物的磷代谢
植物PAPs与土壤有机磷的分解吸收及植株体内磷素的再利用有密切关系, 很多植物来源的PAPs都具有酸性磷酸酶活性, 能够将植物体内外的有机磷水解成无机磷, 供植物吸收利用。根据PAPs是否具有分泌到细胞外的性质, 可将其分为细胞内酸性磷酸酶(intracellular acid phosphatase, IAP)和分泌型酸性磷酸酶(secreted acid phosphatase, SAP)。细胞内酸性磷酸酶主要水解植物体内磷库的磷, 进行细胞内磷素的再利用; 分泌型酸性磷酸酶则被分泌出细胞外, 活化土壤中的有机磷组分, 释放出供植物利用的无机磷(Wasaki et al., 2003)。细胞内的无机磷含量与IAP和SAP活性密切相关。在磷饥饿诱导下, IAP会迅速催化植物体内储存的有机磷, 释放无机磷, 保证磷源的供应(Veneklaas et al., 2012)。随后, 植物会诱导合成SAP并且分泌到细胞外, 水解植物体外的磷源, 将释放的无机磷吸收到植物体内(Wasaki et al., 2003) (图3)。拟南芥AtPAP12和AtPAP26分别是2个主要的细胞内和分泌型酸性磷酸酶。缺磷条件下, 它们被检测到超量表达(Wang et al., 2014)。在磷胁迫下, 可检测到番茄(Solanum lycopersicum)悬浮细胞中IAP和SAP先后被诱导表达(Kusudo et al., 2003)。图3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3缺磷情况下分泌型酸性磷酸酶的诱导表达及其根际分泌与排根形成示意图(改自Wasaki et al., 2003)
(A) 磷充足条件下, SAP聚集在根表皮细胞周围, 只有少量SAP分泌; (B) 磷胁迫条件下, SAP在12小时内从根部迅速分泌, 但在长时间保持较低水平; (C) 组织中磷含量明显下降后, 根系成簇排列, 形成排根, SAP分泌量显著增加。
Figure 3Schematic model of SAP expression and formation of cluster root under phosphorus deficiency (modified from Wasaki et al., 2003)
(A) Under sufficient P conditions, SAP localizes around the epidermal cells of roots, and only a small amount of SAP is secreted; (B) After P stress treatment, SAP is released from roots rapidly within 12 h, however, SAP secretion remains at a low level for a long time; (C) After the decrease of P content in tissues, cluster roots form and SAP secretion significantly increases.
3.2 参与植物应对逆境
大豆酸性磷酸酶3基因(GmPAP3)定位在线粒体(活性氧产生的主要部位), 其表达受盐和氧胁迫诱导, 可能在非生物逆境响应过程中发挥作用。将GmPAP3过表达植株分别进行盐、PEG以及农药胁迫处理, 对比野生型发现过表达植株的根长增加。此外, Gm- PAP3能够通过抑制活性氧(ROS)的积累, 减轻盐和过氧化胁迫造成的损伤(Liao et al., 2003; Li et al., 2008)。酸性磷酸酶15基因(AtPAP15)在拟南芥中组成型表达, 具有植酸酶活性。在atpap15突变体材料中, 抗坏血酸盐(ascorbic acid sodium salt, AsA)的含量下降, 植酸的含量升高, 植物对氧胁迫逆境的抗性降低。反之, 在AtPAP15过量表达的转基因拟南芥中, AsA含量升高, 抗性增强。AtPAP15通过水解植 酸盐释放出肌醇和无机磷, 肌醇参与抗坏血酸合成 途径, 进而调节植物的逆境适应性(Zhang et al., 2008)。3.3 调节植物的碳代谢
Sun等(2012b)研究表明, AtPAP2是一种尾部锚定蛋白, 含有N-端信号肽和C-端膜锚定信号肽。AtPAP2在拟南芥角果、茎、花、根和开始衰老的叶中高度表达, 但在叶和成熟的种子中表达量相对较低(Sun et al., 2012a)。AtPAP2在C-末端具有保守的疏水结构域, 这决定了其亚细胞定位及对叶绿体和线粒体的双向靶定; 同时, C-端锚定信号肽的存在直接影响AtPAP2的基因功能, 将C-端信号肽去掉, AtPAP2将无法继续行使功能(Sun et al., 2012a)。AtPAP2过量表达的拟南芥、亚麻芥(Camelina sativa)和马铃薯转基因植株抽苔提前, 生长速度加快, 叶片光合速率提高10%-20%, 种子体积及生物量增加; 同时, 蔗糖磷酸合酶的活性显著升高, 嫩枝含有更高水平的糖类(蔗糖和己糖)、丙氨酸和脯氨酸等三羧酸循环(TCA)代谢产物(Sun et al., 2012a; Zhang et al., 2012, 2014)。此外, AtPAP2转基因植株的ATP含量升高, 细胞壁合成相关酶(果胶酯酶和纤维素合酶)基因表达量上调(Sun et al., 2013; Liang et al., 2014, 2015)。AtPAP2是第1个被证实参与碳代谢的紫色酸性磷酸酶。最新研究表明, AtPAP2能与线粒体和叶绿体中参与RNA编辑的MORFs因子互作, 控制相应蛋白进出线粒体和叶绿体的效率, 最终调节质体基因的表达(Law et al., 2015; Zhang et al., 2016; Sun et al., 2017)。3.4 参与植物细胞壁合成
烟草中, NtPAP12可催化重组的木糖苷酶磷酸化和去磷酸化以及葡糖苷酶去磷酸化, 抑制木葡聚糖降解, 增加木聚糖的合成; 还可通过葡糖苷酶去磷酸化, 抑制纤维寡糖的降解, 进而增加纤维素和胼胝质的合成。NtPAP12过表达株系糖苷酶活性下降, 质外体中木葡聚糖和纤维寡糖含量上升, 细胞壁主要成分(木聚糖、胼胝质和纤维素)的含量增加, 证实NtPAP12能够调节木糖苷酶以及葡糖苷酶的活性, 从而促进细胞壁的合成(Kaida et al., 2008, 2009, 2010)。拟南芥AtPAP10与烟草NtPAP12的功能相似性较高, 能够活化β-葡聚糖合成酶, 推测其可能参与初生壁的合成(Kaida et al., 2003)。4 植物PAPs基因的表达调控
不同PAPs在植物体内的表达模式不尽相同, 它们分工合作, 在转录水平、翻译水平和翻译后水平受到调控。绝大部分植物PAPs的表达水平不仅与植物体内的无机磷含量相关, 而且与植物所处的逆境(氧和盐胁迫等)环境有关。对26个水稻PAPs编码基因进行启动子元件分析, 发现其中12个基因的启动子含有顺式作用元件P1BS (PHR1-binding site), P1BS是水稻磷信号代谢途径转录因子OsPHR2的结合元件(Rubio et al., 2001)。PHR1在水稻中的同源基因为OsPHR2 (Zhou et al., 2008)。通过对OsPHR2过表达材料进行定量检测表明, 其中10个PAPs受到Os- PHR2诱导表达(Zhang et al., 2011)。与水稻PAPs类似, 拟南芥AtPAP10、AtPAP12和AtPAP15等在转录水平受到PHR1正调控(Wang et al., 2011; Robinson et al., 2012b)。大部分PAP都具有N-端信号肽, 这个信号肽对PAP蛋白的锚定和生物学功能具有重要作用。信号肽定位预测表明, 少数PAPs定位于线粒体(Zhang et al., 2011)。虽然很多细胞内和分泌型酸性磷酸酶编码基因已被克隆, 但是其表达调控机制和分子生物学功能还不是很清楚。低磷胁迫下, AtPAP10和AtPAP12的表达受到诱导, 由此可知这2个基因存在转录水平的调控; AtPAP26在转录水平没有变化, 但是细胞内外AtPAP26蛋白含量显著增加, 表明该基因受到转录后水平的调控(Tran et al., 2010b)。另外, AtPAP1也被证实可能受到转录后水平的调控(Wang et al., 2011)。由于AtPAP10、AtPAP12和AtPAP26蛋白的N-端都存在至少1个糖基化修饰位点, 故这些蛋白的糖基化修饰可能直接影响它们的细胞定位(Tran et al., 2010b), 但具体的糖基化调控方式尚有待研究。
5 展望
植物PAPs是植物体对低磷应答反应及其信号转导途径的重要调控因子。克隆与解析PAPs基因功能, 对认识植物对磷高效利用的分子机制、解决当前土壤有效磷不足以及植物磷缺乏等问题具有重要意义。通过对植物各物种PAPs家族的不同成员进行分析, 发现除与金属离子活性中心结合的5段氨基酸序列高度保守外, PAPs蛋白分子大小和氨基酸序列均存在较大差异, 暗示PAPs成员间存在功能多样性。近年来, 已从高等植物中分离鉴定出许多PAPs的编码基因, 它们参与了次生壁合成、活性氧代谢和抵御病原菌侵染等各种生命过程, 但大多数植物PAPs基因的功能未知, 值得进一步挖掘和探讨。此外, 虽然许多植物细胞内和分泌型酸性磷酸酶编码基因已被克隆, 但其表达调控机制至今未有详尽的解释。磷胁迫条件下, AtPAP26在转录水平并未发生变化。免疫印迹检测显示, AtPAP26在蛋白水平受到缺磷诱导, 但是其在蛋白水平具体的调控机制还不清楚(Tran et al., 2010a; Robinson et al., 2012b)。序列分析和生化实验证明, 大多数植物紫色酸性磷酸酶都具有翻译后糖基化修饰的功能。糖基化修饰可调节酶的定位、溶解性、稳定性和动力学特征。现已发现不同物种PAPs家族的很多成员也存在潜在的N-端糖基化修饰位点, 但糖基化如何改变PAPs定位及动力学活性还有待探讨。未来对PAPs的研究如果能够解释上述问题, 将对阐明植物PAPs的功能以及调控大有助益。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.11983/CBB16028URL [本文引用: 1]

以大麦(Hordeum vulgare)磷高效基因型(DH110和DH147)和低效基因型(DH49)为材料,采用盆栽实验研究大麦在极低磷(25 mg·kg–1土)、低磷(50 mg·kg–1土)和正常磷(75 mg·kg–1土)处理下叶片的磷组分和酸性磷酸酶活性特征。结果表明,低磷胁迫显著降低大麦叶片的无机磷含量,但对难溶态磷含量影响较小。高效基因型上部叶核酸态磷含量显著高于低效基因型,而下部叶则显著低于低效基因型,是低效基因型的18.4%–91.4%。大麦下部叶酯磷含量和分配比例表现为高效基因型低于低效基因型,而上部叶仅在低效基因型中显著低于高效基因型。核酸态磷和酯磷在高效基因型叶片中的含量分配表明其上部叶的磷素营养状况较优,而下部叶易溶性有机磷的分解转化作用更强。低磷和极低磷胁迫下,下部叶酸性磷酸酶的活性显著增加,且高效基因型显著高于低效基因型,分别为低效基因型的1.29–1.41倍。磷高效基因型大麦通过提高下部叶酸性磷酸酶活性加强酯磷和核酸态磷的分解,转化为无机磷,增加可移动性磷源的含量和比例,以提高生育后期大麦的磷素再利用能力。
DOI:10.3724/SP.J.1006.2010.00517URL [本文引用: 1]
PAP17 BrPAP17-1) and type II (BoPAP17-2 and BrPAP17-2). Southern hybridization resulted in two bands both in B. oleracea and B. rapa, this is accordance with former cloning results. Phytogenetic and molecular evolution analysis indicated that PAP17 genes in Brassica species underwent purifying selection, and their deduced proteins are typical low molecular weight PAP proteins. Expression patterns of BoPAP17 and BrPAP17 genes were assayed by fluorescent quantitative PCR. The results revealed that PAP17 genes expressed in all nine tested tissues and organs, with the extremely high expression in flower and bud, and certain expression in seeds at different stages, implying these PAP17 genes most likely mobilize phosphorus reserves in plants, particularly during flower and bud development stages. Under phosphate starvation conditions, expression of BrPAP17-2 in seedling leaf was restrained, while that of BrPAP17 and BoPAP17 in seedling root and leaf was induced, the expression levels declined in the first 24 hours, and then continuously increased with the maximal levels between four days and eight days after treatment. After four days of Pi-resupply, their expression declined below un-induced basal levels. In comparison with seedling leaf, it seems that BrPAP17 and BoPAP17 showed stronger phosphate starvation induced expression in seedling root. These results thus suggested that PAP17 genes in B. oleracea and B. rapa may be involved in external phosphorus assimilation and transferring inorganic phosphate from root to other tissues or organs.
DOI:10.3969/j.issn.1674-3466.2005.01.014URL [本文引用: 1]
Exploration and utilization of the genetic resources of high efficient phosphorus (P) nutrition in plants are the key to sustainable agricultural development. High efficiency of P nutritionin plants is closely related to root morphology, root exudates, P transport across plasmalemma, phosphorus translocation and symbiosis with microorganisms. These nutritional properties are geneticallycontrolled by multi-genes and characterized by quantitative trait loci. In past years, many genes encoding high-affinity P transporters have been cloned; the genes regulating root-to-shoot P translocation and down-regulating P absorption have been identified. The genetic regulation of root exudation of organic acids and acid phosphatases has been moderately understood. Bacterial citric synthase has been successfully introduced into and overexpressed in plant cells. However, few advances have been made in the molecular biology of root hair, cluster roots, root architecture and mycorrhiza in relation to high efficient P nutrition.
DOI:10.1023/A:1021194511492URL [本文引用: 1]
The world population is expanding rapidly and will likely be 10 billion by the year 2050. Limited availability of additional arable land and water resources, and the declining trend in crop yields globally make food security a major challenge in the 21st century. According to the projections, food production on presently used land must be doubled in the next two decades to meet food demand of the growing world population. To achieve the required massive increase in food production, large enhancements in application of fertilizers and improvements of soil fertility are indispensable approaches. Presently, in many developing countries, poor soil fertility, low levels of available mineral nutrients in soil, improper nutrient management, along with the lack of plant genotypes having high tolerance to nutrient deficiencies or toxicities are major constraints contributing to food insecurity, malnutrition (i.e., micronutrient deficiencies) and ecosystem degradation. Plant nutrition research provides invaluable information highly useful in elimination of these constraints, and thus, sustaining food security and well-being of humans without harming the environment. The fact that at least 60% of cultivated soils have growth-limiting problems with mineral-nutrient deficiencies and toxicities, and about 50% of the world population suffers from micronutrient deficiencies make plant nutrition research a major promising area in meeting the global demand for sufficient food production with enhanced nutritional value in this millennium. Integration of plant nutrition research with plant genetics and molecular biology is indispensable in developing plant genotypes with high genetic ability to adapt to nutrient deficient and toxic soil conditions and to allocate more micronutrients into edible plant products such as cereal grains.
.
DOI:10.1111/tpj.12663URLPMID:25270985
Summary Plant purple acid phosphatases (PAPs) belong to a relatively large gene family whose individual functions are poorly understood. Three PAP isozymes that are up-regulated in the cell walls of phosphate (Pi)-starved (61Pi) Arabidopsis thaliana suspension cells were purified and identified by MS as AtPAP12 (At2g27190), AtPAP25 (At4g36350) and AtPAP26 (At5g34850). AtPAP12 and AtPAP26 were previously isolated from the culture medium of 61Pi cell cultures, and shown to be secreted by roots of Arabidopsis seedlings to facilitate Pi scavenging from soil-localized organophosphates. AtPAP25 exists as a 5502kDa monomer containing complex NX(S/T) glycosylation motifs at Asn172, Asn367 and Asn424. Transcript profiling and immunoblotting with anti-AtPAP25 immune serum indicated that AtPAP25 is exclusively synthesized under 61Pi conditions. Coupled with potent mixed-type inhibition of AtPAP25 by Pi ( I 5002=025002μ m ), this indicates a tight feedback control by Pi that prevents AtPAP25 from being synthesized or functioning as a phosphatase except when Pi levels are quite low. Promoter–GUS reporter assays revealed AtPAP25 expression in shoot vascular tissue of 61Pi plants. Development of an atpap25 T-DNA insertion mutant was arrested during cultivation on soil lacking soluble Pi, but rescued upon Pi fertilization or complementation with AtPAP25 . Transcript profiling by quantitative RT-PCR indicated that Pi starvation signaling was attenuated in the atpap25 mutant. AtPAP25 exhibited near-optimal phosphatase activity with several phosphoproteins and phosphoamino acids as substrates. We hypothesize that AtPAP25 plays a key signaling role during Pi deprivation by functioning as a phosphoprotein phosphatase rather than as a non-specific scavenger of Pi from extracellular P-monoesters.
DOI:10.1093/aob/mch156URLPMID:4242181 [本文引用: 1]
Background Phosphorus (P) is an essential macronutrient for plants. Plants take up P as phosphate (Pi) from the soil solution. Since little Pi is available in most soils, P fertilizers are applid to crops. However, the use of P fertilizers is unsustainable and may cause pollution. Consequently, there is a need to develop more P-use-efficient (PUE) crops and precise methods to monitor crop P-status. Scope Manipulating the expression of genes to improve the PUE of crops could reduce their P fertilizer requirement. This has stimulated research towards the identification of genes and signalling cascades involved in plant responses to P deficiency. Genes that respond to P deficiency can be grouped into 'early' genes that respond rapidly and often non-specifically to P deficiency, or 'late' genes that impact on the morphology, physiology or metabolism of plants upon prolonged P deficiency. Summary The use of micro-array technology has allowed researchers to catalogue the genetic responses of plants to P deficiency. Genes whose expression is altered by P deficiency include various transcription factors, which are thought to coordinate plant responses to P deficiency, and other genes involved in P acquisition and tissue P economy. Several common cw-regulatory elements have been identified in the promoters of these genes, suggesting that their expression might be coordinated. It is suggested that knowledge of the genes whose expression changes in response to P deficiency might allow the development of crops with improved PUE, and could be used in diagnostic techniques to monitor P deficiency in crops either directly using 'smart' indicator plants or indirectly through transcript profiling. The development of crops with improved PUE and the adoption of diagnostic technology could reduce production costs, minimize the use of a non-renewable resource, reduce pollution and enhance biodiversity.
.
DOI:10.1007/s10529-009-0131-1URLPMID:19838636 [本文引用: 1]
A phosphate starvation-induced, purple, acid phosphatase cDNA was cloned from rice, Oryza 02 sativa . The cDNA encoding the phosphatase ( OsPAP2 ) has 1,89302bp with an open reading frame of 630 amino acid residues. The deduced amino acid sequence of OsPAP2 shows identities of 60–63% with other plant purple acid phosphatases and appears to have five conserved motifs containing the residues involved in metal binding. OsPAP2 expression is up-regulated in the rice plant and in cell cultures in the absence of phosphate (P i ). The induced expression of OsPAP2 is a specific response to P i starvation, and is not affected by the deprivation of other nutrients. OsPAP2 expression was responsive to the level of P i -supply, and transcripts of OsPAP2 were abundant in P i -deprived roots. The OsPAP2 cDNA was expressed as a 6902kDa polypeptide in baculovirus-infected insect Sf9 cells. In addition, the OsPAP2 gene was introduced into Arabidopsis via an Agrobacterium -mediated transformation. Functional expression of the OsPAP2 gene in the transgenic Arabidopsis line was confirmed by northern and western blot analyses, as well as by phosphatase activity assays. These results suggest that the OsPAP2 gene can be used to develop new transgenic dicotyledonous plants that are able to adapt to P i -deficient conditions.
DOI:10.1104/pp.98.3.801URLPMID:16668751 [本文引用: 1]
Sunflower (Helianthus annuus L. cv Asmer) and maize (Zea mays L. cv Eta) plants were grown under controlled environmental conditions with a nutrient solution containing 0, 0.5, or 10 millimolar inorganic phosphate. Phosphate-deficient leaves had lower photosynthetic rates at ambient and saturating CO(2) and much smaller carboxylation efficiencies than those of plants grown with ample phosphate. In addition, phosphate-deficient leaves contained smaller quantities of total soluble proteins and ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) per unit area, although the relative proportions of these components remained unchanged. The specific activity of Rubisco (estimated in the crude extracts of leaves) was significantly reduced by phosphate deficiency in sunflower but not in maize. Thus, there was a strong dependence of carboxylation efficiency and CO(2)-saturated photosynthetic rate on Rubisco activity only in sunflower. Phosphate deficiency decreased the 3-phosphoglycerate and ribulose-1,5-bisphosphate (RuBP) contents of the leaf in both species. The ratio of 3-phosphoglycerate to RuBP decreased in sunflower but increased in maize with phosphate deficiency. The calculated concentrations of RuBP and RuBP-binding sites in the chloroplast stroma decreased markedly with phosphate deficiency. The ratio of the stromal concentration of RuBP to that of RuBP-binding sites decreased in sunflower but was not affected in maize with phosphate deficiency. We suggest that a decrease in this ratio made the RuBP-binding sites more vulnerable to blockage or inactivation by tight-binding metabolites/inhibitors, causing a decrease in the initial specific activity of Rubisco in the crude extract from phosphate-deficient sunflower leaves. However, the decrease in Rubisco specific activity was much less than the decrease in the RuBP content in the leaf and its concentration in the stroma. A large ratio of RuBP to RuBP-binding sites may have maintained the Rubisco-specific activity in phosphate-deficient maize leaves. We conclude that the effect of phosphate deficiency is more on RuBP regeneration than on Rubisco activity in both sunflower and maize.
DOI:10.1021/ic1020324URLPMID:21452875 [本文引用: 1]
To model the heterodinuclear active sites in plant purple acid phosphatases, a mononuclear synthon, [Fe(III)(H(2)IPCPMP)(Cl(2))][PF(6)] (1), has been generated in situ from the ligand 2-(N-isopropyl-N-((2-pyridyl)methyl)aminomethyl)-6-(N-(carboxylmethyl)-N-((2-pyridyl)methyl)amino methyl)-4-methylphenol (IPCPMP) and used to synthesize heterodinuclear complexes of the formulas [Fe(III)M(II)(IPCPMP)(OAc)(2)(CH(3)OH)][PF(6)] (M = Zn (2), Co (3), Ni (4), Mn (5)), [Fe(III)Zn(II)(IPCPMP)(mpdp)][PF(6)] (6) (mpdp = meta-phenylene-dipropionate), and [Fe(III)Cu(II)(IPCPMP) (OAc)}(2)(O)][PF(6)] (7). Complexes 2-4, 6, and 7 have been crystallographically characterized. The structure of 6 is a solid state coordination polymer with heterodinuclear monomeric units, and 7 is a tetranuclear complex consisting of two heterodinuclear phenolate-bridged Fe(III)Cu(II) units bridged through a -oxido group between the two Fe(III) ions. Mo ssbauer spectra confirm the presence of high spin Fe(III) in an octahedral environment for 1, 3, and 5 while 2 and 4 display relaxation effects. Magnetic susceptibility measurements indicate weak antiferromagnetic coupling for 3, 4, and 5 and confirm the assignment of the metal centers in 2-5 as high spin Fe(III)-M(II) (M = Zn, Co (high spin), Ni (high spin), Mn (high spin)). Complexes 2-5 are intact in acetonitrile solution as indicated by IR spectroscopy (for 2-4) and electrospray ionization mass spectrometry (ESI-MS) but partly dissociate to hydroxide species and a mononuclear complex in water/acetonitrile solutions. UV-vis spectroscopy reveal pH-dependent behavior, and species that form upon increasing the pH have been assigned to -hydroxido-bridged Fe(III)M(II) complexes for 2-5 although 2 and 3 is further transformed into what is propsed to be a -oxido-bridged tetranuclear complex similar to 7. Complexes 2-5 enhance phosphodiester cleavage of 2-hydroxy-propyl-p-nitrophenyl phosphate (HPNP) and bis(2,4-dinitrophenyl)phosphate (BDNPP), but the reactivities are different for different complexes and generally show strong pH dependence.
DOI:10.1016/j.phytochem.2008.07.008URLPMID:18762304 [本文引用: 1]
Purple acid phosphatase isolated from the walls of tobacco cells appears to be a 220 kDa homotetramer composed of 60 kDa subunits, which is purple in color and which contains iron as its only metal ion. Although the phosphatase did not require dithiothreitol for activity and was not inhibited by phenylarsine oxide, the enzyme showed a higher catalytic efficiency ( k cat/ K m) for phosphotyrosine-containing peptides than for other substrates including p-nitrophenyl-phosphate and ATP. The phosphatase formed as a 120 kDa dimer in the cytoplasm and as a 220 kDa tetramer in the walls, where Brefeldin A blocked its secretion during wall regeneration. According to our double-immunofluorescence labeling results, the enzyme might be translocated through the Golgi apparatus to the walls at the interphase and to the cell plate during cytokinesis.
DOI:10.1016/S0167-4781(02)00599-7URLPMID:12531472 [本文引用: 1]
Four full-length cDNAs were isolated from a cDNA library prepared from tobacco cultured cells and designated NtPAP4, NtPAP12, NtPAP19 and NtPAP21, which could correspond to purple acid phosphatase (PAP). Levels of both NtPAP12 and NtPAP21 mRNA in the protoplasts immediately increased after the protoplasts were transferred to a medium for cell wall regeneration, and the accumulation of the mRNA was correlated with cell wall regeneration for 3 h. It is likely that the NtPAP12 and NtPAP21 gene products are wall-bound PAPs at the early stage of regenerating walls in tobacco protoplasts.
DOI:10.1104/pp.109.139287URLPMID:19493971 [本文引用: 1]
Wall-bound purple acid phosphatases have been shown to be potentially involved in the regulation of plant cell growth. The aim of this work was to further investigate the function of one of these phosphatases in tobacco (Nicotiana tabacum), NtPAP12, using transgenic cells overexpressing the enzyme. The transgenic cells exhibited a higher level of phosphatase activity in their walls. The corresponding protoplasts regenerating a cell wall exhibited a higher rate of /3-glucan synthesis and cellulose deposition was increased in the walls of the transgenic cells. A higher level of plasma membrane glucan synthase activities was also measured in detergent extracts of membrane fractions from the transgenic line, while no activation of Golgi-bound glycan synthases was detected. Enzymatic hydrolysis and methylation analysis were performed on the products synthesized in vitro by the plasma membrane enzymes from the wild-type and transgenic lines extracted with digitonin and incubated with radioactive UDP-glucose. The data showed that the glucans consisted of callose and cellulose and that the amount of each glucan synthesized by the enzyme preparation from the transgenic cells was significantly higher than in the case of the wildtype cells. The demonstration that callose and cellulose synthases are activated in cells overexpressing the wall-bound phosphatase NtPAP12 suggests a regulation of these carbohydrate synthases by a phosphorylation/dephosphorylation process, as well as a role of wall-bound phosphatases in the regulation of cell wall biosynthesis.
DOI:10.1104/pp.110.154138URLPMID:20357138 [本文引用: 1]
It is not yet known whether dephosphorylation of proteins catalyzed by phosphatases occurs in the apoplastic space. In this study, we found that tobacco (Nicotiana tabacum) purple acid phosphatase could dephosphorylate the phosphoryl residues of three apoplastic proteins, two of which were identified as α-xylosidase and β-glucosidase. The dephosphorylation and phosphorylation of recombinant α-xylosidase resulted in a decrease and an increase in its activity, respectively, when xyloglucan heptasaccharide was used as a substrate. Attempted overexpression of the tobacco purple acid phosphatase NtPAP12 in tobacco cells not only decreased the activity levels of the glycosidases but also increased levels of xyloglucan oligosaccharides and cello-oligosaccharides in the apoplast during the exponential phase. We suggest that purple acid phosphatase controls the activity of α-xylosidase and β-glucosidase, which are responsible for the degradation of xyloglucan oligosaccharides and cello-oligosaccharides in the cell walls.
DOI:10.1111/j.1432-1033.1994.tb20061.xURLPMID:8001554 [本文引用: 1]
Purple acid phosphatase of the common bean Phaseolus vulgaris is a homodimeric 110090009kDa glycoprotein with a Fe(III)-Zn(II) center in the active site of each monomer. After exchange of Zn(II) for Fe(II), the enzyme spectroscopically and kinetically resembles the mammalian purple acid phosphatases with Fe(III)-Fe(II) centers in monomeric 35090009kDa proteins. The kidney bean enzyme consists of 432 amino acids/monomer with five N-glycosylated asparagine residues. The complete amino acid sequence was determined by a combination of matrix-assisted laser desorption/ionization mass spectrometry (MALDI-MS) and classical sequencing methods. Our strategy involved mass determination and sequence analysis of all cyanogen-bromide-generated fragments by automated Edman degradation. Limited cleavages with cyanogen bromide were performed to obtain fragments containing still uncleaved Met-Xaa linkages. MALDI mass spectra of these products allowed the characterization of each fragment and the determination of the order of the cyanogen bromide fragments in the intact protein without producing overlapping peptides. For one large 30090009kDa methionine-free fragment, the alignment of the Edman-degraded tryptic peptides was obtained by MALDI-MS analysis and enzymic microscale peptide laddering of overlapping Glu-C-generated fragments. The employed strategy shows that the classical method, in combination with modern mass spectrometry, is an attractive approach for primary structure determination in addition to the DNA sequencing method.
DOI:10.1016/0014-5793(95)00536-IURLPMID:7601285 [本文引用: 1]
Abstract The primary structure of uteroferrin (Uf), a 35 kDa monomeric mammalian purple acid phosphatase (PAP) containing a Fe(III)-Fe(II) center, has been compared with the sequence of the homodimeric 111 kDa Fe(III)-Zn(II) kidney bean purple acid phosphatase (KBPAP). The alignment suggests that the amino acid residues ligating the dimetal center are identical in Uf and KBPAP, although the geometry of the coordination sphere might slightly differ. Secondary structure predictions indicate that Uf contains two beta alpha beta alpha beta motifs thus resembling the folding topology of the plant enzyme. Guided by the recently determined X-ray structure of KBPAP a tentative model for the mammalian PAP can be constructed.
.
DOI:10.1104/pp.109.143180URLPMID:19633233 [本文引用: 1]
Purple acid phosphatase (PAP) catalyzes the hydrolysis of phosphate monoesters and anhydrides to release phosphate within an acidic pH range. Among the 29 PAP-like proteins in Arabidopsis (Arabidopsis thaliana), AtPAP15 (At3g07130) displays a greater degree of amino acid identity with soybean (Glycine max; GmPHY) and tobacco (Nicotiana tabacum) PAP (NtPAP) with phytase activity than the other AtPAPs. In this study, transgenic Arabidopsis that expressed an AtPAP15 promoter:-glucuronidase (GUS) fusion protein showed that AtPAP15 expression was developmentally and temporally regulated, with strong GUS staining at the early stages of seedling growth and pollen germination. The expression was also organ/tissue specific, with strongest GUS staining in the vasculature, pollen grains, and roots. The recombinant AtPAP purified from transgenic tobacco exhibited broad substrate specificity with moderate phytase activity. AtPAP15 T-DNA insertion lines exhibited a lower phytase and phosphatase activity in seedling and germinating pollen and lower pollen germination rate compared with the wild type and their complementation lines. Therefore, AtPAP15 likely mobilizes phosphorus reserves in plants, particularly during seed and pollen germination. Since AtPAP15 is not expressed in the root hair or in the epidermal cells, it is unlikely to play any role in external phosphorus assimilation.
DOI:10.1271/bbb.67.1609URL [本文引用: 1]
Purple acid phosphatase (PAP) was purified from sweet potato dry powder, which is used as a food additive. Spectrometric and enzymatic analyses, and analysis of the amino-terminal sequence indicated that the purified purple acid phosphatase was PAP1. High activity in neutral and acidic conditions, broad substrate specificity, and good thermal stability of PAP1 suggest the possibility of practical applications of PAP1.
DOI:10.1104/pp.15.01115URLPMID:26304849 [本文引用: 1]
Abstract The nuclear-encoded mitochondrial-targeted proteins, multiple organellar RNA editing factors (MORF3, MORF5, MORF6) interact with AtPAP2 (Purple acid phosphatase 2) located on the chloroplast and mitochondrial outer membranes in a presequence dependent manner. Phosphorylation of the presequence of the precursor MORF3 (pMORF3) by endogenous kinases in wheat germ translation lysate, leaf extracts, or STY kinases, but not in rabbit reticulocyte translation lysate, resulted in the inhibition of protein import into mitochondria. This inhibition of import could be overcome by altering threonine/serine residues to alanine on the presequence, thus preventing phosphorylation. Phosphorylated pMORF3, but not the phosphorylation deficient pMORF3, can form a complex with 14-3-3 proteins and HSP70. The phosphorylation deficient mutant of pMORF3 also displayed faster rates of import when translated in wheat germ lysates. Mitochondria isolated from plants with altered amounts of AtPAP2 displayed altered protein import kinetics. The import rate of pMORF3 synthesized in wheat germ translation lysate into pap2 mitochondria was slower than that into wild-type mitochondria, and this rate disparity was not seen for pMORF3 synthesized in rabbit reticulocyte translation lysate, the latter translation lysate largely deficient in kinase activity. Taken together, these results support a role for the phosphorylation and dephosphorylation of pMORF3 during the import into plant mitochondria. These results suggest that kinases, possibly STY kinases, and AtPAP2 are involved in the import of protein into both mitochondria and chloroplasts, and provides a mechanism by which the import of proteins into both organelles may be coordinated. Copyright 2015, Plant Physiology.
DOI:10.1093/aob/mcr246URLPMID:3241574 [本文引用: 1]
Purple acid phosphatases (PAPs) are members of the metallo-phosphoesterase family and have been known to play important roles in phosphorus (P) acquisition and recycling in plants. Low P availability is a major constraint to growth and production of soybean, Glycine max. Comparative studies on structure, transcription regulation and responses to phosphate (Pi) deprivation of the soybean PAP gene family should facilitate further insights into the potential physiological roles of GmPAPs. BLAST searches were performed to identify soybean PAP genes at the phytozome website. Bioinformatic analyses were carried out to investigate their gene structure, conserve motifs and phylogenetic relationships. Hydroponics and sand-culture experiments were carried out to obtain the plant materials. Quantitative real-time PCR was employed to analyse the expression patterns of PAP genes in response to P deficiency and symbiosis. In total, 35 PAP genes were identified from soybean genomes, which can be classified into three distinct groups including six subgroups in the phylogenetic tree. The expression pattern analysis showed flowers possessed the largest number of tissue-specific GmPAP genes under normal P conditions. The expression of 23 GmPAPs was induced or enhanced by Pi starvation in different tissues. Among them, nine GmPAP genes were highly expressed in the Pi-deprived nodules, whereas only two GmPAP genes showed significantly increased expression in the arbuscular mycorrhizal roots under low-P conditions. Most GmPAP genes are probably involved in P acquisition and recycling in plants. Also we provide the first evidence that some members of the GmPAP gene family are possibly involved in the response of plants to symbiosis with rhizobia or arbuscular mycorrhizal fungi under P-limited conditions.
DOI:10.1074/jbc.M204183200URLPMID:12021284 [本文引用: 2]
Purple acid phosphatases (PAPs) are members of the metallo-phosphoesterase family. They are characterized by the presence of seven conserved amino acid residues involved in coordinating the dimetal nuclear center in theirreactive site. We compared the 29 PAPs predicted for Arabidopsis thaliana in their varieties of potential metal-ligating residues. Although 24 members possessed sets of metal-ligating residues typical of known PAPs, 1 member lacked four of the seven residues. For the remaining four members, potential metal-ligating residues were generally more similar to those in metal-dependent exonucleases and related proteins. Evidence was obtained for the expression of the majority of the 29 PAPs. To facilitate future investigations, a scheme for naming Arabidopsis PAPs and a system for classifying the 29 PAPs are proposed. The cDNA sequences and the responses to phosphate deprivation of seven Arabidopsis PAPs (AtPAP7-AtPAP13) were characterized. For some AtPAPs analyzed, there were fully processed transcripts as well as splice variants. The splice variants of AtPAP10 were found to associate with polyribosomes and may be translated into a NH
.
DOI:10.1111/j.1469-8137.2007.02356.xURLPMID:18208471 [本文引用: 2]
090004 The primary biochemical reaction of purple acid phosphatases (PAP) is to catalyze the hydrolysis of phosphate esters and anhydrides. However, the soybean GmPAP3 gene expression is induced by NaCl, osmotic, and oxidative treatments, indicating a possible role of PAP in abiotic stress responses. 090004 Confocal and electron microscopic studies demonstrated that GmPAP3 protein is mainly localized in mitochondria, a primary site for reactive oxygen species (ROS) production. 090004 When subjected to NaCl and polyethylene glycol (PEG) treatments, ectopic expression of GmPAP3 in transgenic tobacco BY-2 cells mimicked the protective effects exhibited by the antioxidant ascorbic acid: increase in the percentage of cells with active mitochondria; reduction in the percentage of dead cells; and reduced accumulation of ROS. In addition, when GmPAP3 transgenic Arabidopsis thaliana seedlings were subjected to NaCl, PEG, and paraquat (PQ) treatments, the percentage of root elongation was significantly higher than the wild type. Furthermore, PQ-induced lipid peroxidation in these transgenic seedlings was also reduced. 090004 In summary, the mitochondrial localized GmPAP3 may play a role in stress tolerance by enhancing ROS scavenging.
.
DOI:10.1186/1471-2164-15-116URLPMID:24507710 [本文引用: 1]
In higher eukaryotes, small RNAs play a role in regulating gene expression. Overexpression (OE) lines ofArabidopsis thalianapurple acid phosphatase 2 (AtPAP2) were shown to grow faster and exhibit higher ATP and sugar contents. Leaf microarray studies showed that many genes involved in microRNAs (miRNAs) and trans-acting siRNAs (tasiRNAs) biogenesis were significantly changed in the fast-growing lines. In this study, the sRNA profiles of the leaf and the root of 20-day-old plants were sequenced and the impacts of high energy status on sRNA expression were analyzed. 9-13 million reads from each library were mapped to genome. miRNAs, tasiRNAs and natural antisense transcripts-generated small interfering RNAs (natsiRNAs) were identified and compared between libraries. In the leaf of OE lines, 15 known miRNAs increased in abundance and 9 miRNAs decreased in abundance, whereas in the root of OE lines, 2 known miRNAs increased in abundance and 9 miRNAs decreased in abundance. miRNAs with increased abundance in the leaf and root samples of both OE lines (miR158b and miR172a/b) were predicted to target mRNAs coding for Dof zinc finger protein and Apetala 2 (AP2) proteins, respectively. Furthermore, a significant change in the miR173-tasiRNAs-PPR/TPRnetwork was observed in the leaves of both OE lines. In this study, the impact of high energy content on the sRNA profiles of Arabidopsis is reported. While the abundance of many stress-induced miRNAs is unaltered, the abundance of some miRNAs related to plant growth and development (miR172 and miR319) is elevated in the fast-growing lines. An induction of miR173-tasiRNAs-PPR/TPRnetwork was also observed in the OE lines. In contrast, only few cis- and trans-natsiRNAs are altered in the fast-growing lines.
.
DOI:10.3389/fpls.2015.00922URLPMID:4623399 [本文引用: 1]
Chloroplasts and mitochondria are the major ATP producing organelles in plant leaves.Arabidopsis thalianapurple acid phosphatase 2 (AtPAP2) is a phosphatase dually targeted to the outer membranes of both organelles and it plays a role in the import of selected nuclear-encoded proteins into these two organelles. Overexpression (OE) of AtPAP2 inA. thalianaaccelerates plant growth and promotes flowering, seed yield, and biomass at maturity. Measurement of ADP/ATP/NADP+/NADPH contents in the leaves of 20-day-old OE and wild-type (WT) lines at the end of night and at 1 and 8 h following illumination in a 16/8 h photoperiod revealed that the ATP levels and ATP/NADPH ratios were significantly increased in the OE line at all three time points. The AtPAP2 OE line is therefore a good model to investigate the impact of high energy on the global molecular status ofArabidopsis. In this study, transcriptome, proteome, and metabolome profiles of the high ATP transgenic line were examined and compared with those of WT plants. A comparison of OE and WT at the end of the night provide valuable information on the impact of higher ATP output from mitochondria on plant physiology, as mitochondrial respiration is the major source of ATP in the dark in leaves. Similarly, comparison of OE and WT following illumination will provide information on the impact of higher energy output from chloroplasts on plant physiology. OE of AtPAP2 was found to significantly affect the transcript and protein abundances of genes encoded by the two organellar genomes. For example, the protein abundances of many ribosomal proteins encoded by the chloroplast genome were higher in the AtPAP2 OE line under both light and dark conditions, while the protein abundances of multiple components of the photosynthetic complexes were lower. RNA-seq data also showed that the transcription of the mitochondrial genome is greatly affected by the availability of energy. These data reflect that the transcription and translation of organellar genomes are tightly coupled with the energy status. This study thus provides comprehensive information on the impact of high ATP level on plant physiology, from organellar biology to primary and secondary metabolism.
DOI:10.2307/25680699URL [本文引用: 2]
http://www.jstor.org/stable/25680699
DOI:10.1016/S0378-1119(03)00764-9URLPMID:14585503 [本文引用: 1]
Purple acid phosphatases (PAPs) are commonly found in plants, but the physiological functions of different classes of PAPs are not thoroughly understood. In the present study, we identified a novel gene, GmPAP3, from salt-stressed soybean using suppression subtractive hybridization (SSH) techniques. Protein sequence alignment studies and phylogenetic analysis strongly suggested that GmPAP3 belongs to the group of plant PAPs and PAP-like proteins that are distinct from those of fungi and animals. In addition, the invariable consensus metal binding residues of PAPs were all conserved in GmPAP3. Surprisingly, analysis of protein sorting signals showed that a putative mitochondrion targeting transit peptide is present on GmPAP3. Northern blot analysis revealed that NaCl stress causes a general induction of GmPAP3 expression in both roots and leaves of various cultivated ( Glycine max) and wild ( Glycine soja) soybean varieties. Further test using two genetically unrelated cultivated soybean varieties showed that the expression pattern of GmPAP3 is distinct from other PAP genes in soybeans. NaCl stress and oxidative stress but not phosphorus (P) starvation induces the expression of GmPAP3. These results suggest that the physiological role of GmPAP3 might be related to the adaptation of soybean to NaCl stress, possibly through its involvement in reactive oxygen species (ROS) forming and/or scavenging or stress-responding signal transduction pathways.
.
URL [本文引用: 1]
DOI:10.1016/S1381-1177(02)00143-1URLPMID:14740011 [本文引用: 2]
Abstract The properties of plant purple acid phosphatases (PAPs), metallophosphoesterases present in some bacteria, plants and animals are reviewed. All members of this group contain a characteristic set of seven amino-acid residues involved in metal ligation. Animal PAPs contain a binuclear metallic center composed of two irons, whereas in plant PAPs one iron ion is joined by zinc or manganese ion. Among plant PAPs two groups can be distinguished: small PAPs, monomeric proteins with molecular mass around 35 kDa, structurally close to mammalian PAPs, and large PAPs, homodimeric proteins with a single polypeptide of about 55 kDa. Large plant PAPs exhibit two types of structural organization. One type comprises enzymes with subunits bound by a disulfide bridge formed by cysteines located in the C-terminal region around position 350. In the second type no cysteines are located in this position and no disulfide bridges are formed between subunits. Differences in structural organisation are reflected in substrate preferences. Recent data reveal in plants the occurrence of metallophosphoesterases structurally different from small or large PAPs but with metal-ligating sequences characteristic for PAPs and expressing pronounced specificity towards phytate or diphosphate nucleosides and inorganic pyrophosphate.
.
[本文引用: 1]
.
DOI:10.1111/nph.12006URLPMID:23072540 [本文引用: 1]
Abstract Limitation of crop productivity by suboptimal phosphorus (P) nutrition is a widespread concern. Enhanced crop P-use efficiency could be achieved by improving P remobilization from senescing leaves to developing tissues and seeds. Transcriptomic studies indicate that hundreds of Arabidopsis thaliana genes are up-regulated during leaf senescence, including that encoding the purple acid phosphatase (PAP) AtPAP26 (At5g34850). In this study, biochemical and functional genomic tools were integrated to test the hypothesis that AtPAP26 participates in P remobilization during leaf senescence. An eightfold increase in acid phosphatase activity of senescing leaves was correlated with the accumulation of AtPAP26 transcripts and immunoreactive AtPAP26 polypeptides. Senescing leaves of an atpap26 T-DNA insertion mutant displayed a> 90% decrease in acid phosphatase activity, markedly impaired P remobilization efficiency and delayed senescence. This was paralleled by reduced seed total P concentrations and germination rates. These results demonstrate that AtPAP26 loss of function causes dramatic effects that cannot be compensated for by any other PAP isozyme, even though Arabidopsis contains 29 different PAP genes. Our current and earlier studies establish that AtPAP26 not only helps to scavenge P from organic P sources when Arabidopsis is cultivated in inorganic orthophosphate (Pi)-deficient soils, but also has an important P remobilization function during leaf senescence.2012 The Authors. New Phytologist 2012 New Phytologist Trust.
.
DOI:10.1093/jxb/ers309URLPMID:3504502 [本文引用: 2]
Orthophosphate (Pi) is an essential but limiting macronutrient for plant growth. Extensive soil P reserves exist in the form of organic P (Po), which is unavailable for root uptake until hydrolysed by secretory acid phosphatases (APases). The predominant purple APase (PAP) isozymes secreted by roots of Pi-deficient (–Pi)Arabidopsis thalianawere recently identified as AtPAP12 (At2g27190) and AtPAP26 (At5g34850). The present study demonstrated that exogenous Pocompounds such as glycerol-3-phosphate or herring sperm DNA: (i) effectively substituted for Piin supporting the P nutrition ofArabidopsisseedlings, and (ii) caused upregulation and secretion of AtPAP12 and AtPAP26 into the growth medium. When cultivated under –Piconditions or supplied with Poas its sole source of P nutrition, anatpap26/atpap12T-DNA double insertion mutant exhibited impaired growth coupled with >60 and >30% decreases in root secretory APase activity and rosette total Piconcentration, respectively. Development of theatpap12/atpap26mutant was unaffected during growth on Pi-replete medium but was completely arrested when 7-day-old Pi-sufficient seedlings were transplanted into a –Pi, Po-containing soil mix. Both PAPs were also strongly upregulated on root surfaces and in shoot cell-wall extracts of –Piseedlings. It is hypothesized that secreted AtPAP12 and AtPAP26 facilitate the acclimation ofArabidopsisto nutritional Pi deficiency by: (i) functioning in the rhizosphere to scavenge Pifrom the soil’s accessible Popool, while (ii) recycling Pifrom endogenous phosphomonoesters that have been leaked into cell walls from the cytoplasm. Thus, AtPAP12 and AtPAP26 are promising targets for improving crop P-use efficiency.
DOI:10.1101/gad.204401URL [本文引用: 1]
Plants have evolved a number of adaptive responses to cope with growth in conditions of limited phosphate (Pi) supply involving biochemical, metabolic, and developmental changes. We prepared an EMS-mutagenized M(2) population of an Arabidopsis thaliana transgenic line harboring a reporter gene specifically responsive to Pi starvation (AtIPS1::GUS), and screened for mutants altered in Pi starvation regulation. One of the mutants, phr1 (phosphate starvation response 1), displayed reduced response of AtIPS1::GUS to Pi starvation, and also had a broad range of Pi starvation responses impaired, including the responsiveness of various other Pi starvation-induced genes and metabolic responses, such as the increase in anthocyanin accumulation. PHR1 was positionally cloned and shown be related to the PHOSPHORUS STARVATION RESPONSE 1 (PSR1) gene from Chlamydomonas reinhardtii. A GFP::PHR1 protein fusion was localized in the nucleus independently of Pi status, as is the case for PSR1. PHR1 is expressed in Pi sufficient conditions and, in contrast to PSR1, is only weakly responsive to Pi starvation. PHR1, PSR1, and other members of the protein family share a MYB domain and a predicted coiled-coil (CC) domain, defining a subtype within the MYB superfamily, the MYB-CC family. Therefore, PHR1 was found to bind as a dimer to an imperfect palindromic sequence. PHR1-binding sequences are present in the promoter of Pi starvation-responsive structural genes, indicating that this protein acts downstream in the Pi starvation signaling pathway.
[本文引用: 1]
DOI:10.1016/S0378-1119(00)00186-4URLPMID:10854785 [本文引用: 1]
Purple acid phosphatases (PAPs) comprise a family of binuclear metal-containing hydrolases, members of which have been isolated from plants, mammals and fungi. Polypeptide chains differ in size (animal 35 kDa, plant 55 kDa) and exhibit low sequence homology between kingdoms but all residues involved in co-ordination of the metal ions are invariant. A search of genomic databases was undertaken using a sequence pattern which includes the conserved residues. Several novel potential PAP sequences were detected, including the first known examples from bacterial sources. Ten plant ESTs were also identified which, although possessing the conserved sequence pattern, were not homologous throughout their sequences to previously known plant PAPs. Based on these EST sequences, novel cDNAs from sweet potato, soybean, red kidney bean and Arabidopsis thaliana were cloned and sequenced. These sequences are more closely related to mammalian PAP than to previously characterized plant enzymes. Their predicted secondary structure is similar to that of the mammalian enzyme. A model of the sweet potato enzyme was generated based on the coordinates of pig PAP. These observations strongly suggest that the cloned cDNA sequences represent a second group of plant PAPs with properties more similar to the mammalian enzymes than to the high molecular weight plant enzymes.
DOI:10.1016/S0378-1119(00)00305-XURLPMID:11024303 [本文引用: 3]
Mammalian and plant purple acid phosphatases have similar active site structures despite low sequence identity (<20%). Although no bacterial enzyme has been purified, a sequence database search revealed that genes that could encode potential purple acid phosphatases may be restricted to a small number of organisms (i.e. myco- and cyanobacteria). Analysis of their deduced amino acid sequences and predicted secondary structures indicates that the cyanobacterial enzyme is similar to both the mammalian and the recently discovered low-molecular-weight plant purple acid phosphatases, while the mycobacterial enzyme is homologous to the fungal and high-molecular-weight plant purple acid phosphatases. Homology models indicate that both bacterial proteins appear to be similar to mammalian purple acid phosphatases in the immediate vicinity of the active site. It is likely that these enzymes act as Fenton-type catalysts in order to prevent damage caused by reactive oxygen species generated by invaded host cells ( M. tuberculosis) or by the light-harvesting complex ( Synechocystis sp.).
DOI:10.1016/j.tree.2008.11.009URLPMID:19246117 [本文引用: 1]
Cultural eutrophication has become the primary water quality issue for most of the freshwater and coastal marine ecosystems in the world. However, despite extensive research during the past four to five decades, many key questions in eutrophication science remain unanswered. Much is yet to be understood concerning the interactions that can occur between nutrients and ecosystem stability: whether they are stable or not, alternate states pose important complexities for the management of aquatic resources. Evidence is also mounting rapidly that nutrients strongly influence the fate and effects of other non-nutrient contaminants, including pathogens. In addition, it will be important to resolve ongoing debates about the optimal design of nutrient loading controls as a water quality management strategy for estuarine and coastal marine ecosystems.
[本文引用: 3]
DOI:10.4161/psb.20769URLPMID:3474687 [本文引用: 3]
To date, Arabidopsis purple acid phosphatase 2 (AtPAP2) is the only known plant protein that is dual-targeted to chloroplasts and mitochondria by a C-terminal targeting signal. Using in vitro organelle import and green fluorescence protein (GFP) localization assays, we showed that AtPAP2 is located on, but not imported across the outer membrane (OM) of chloroplasts and mitochondria and exposed its N-terminal enzymatic domain to the cytosol. It was also found that a short stretch of 30 amino acids (a.a.) at the C-terminal region (a.a. 615-644) that contains a stretch of 18 hydrophobic residues, a WYAK motif and 8 hydrophilic residues is sufficient for dual-targeting. Mutation of WYAK to WYAE had no effect on dual-targeting ability suggesting that the charge within this flanking region alone is not an important determinant for dual-targeting.
.
[本文引用: 1]
.
DOI:10.1111/j.1469-8137.2011.04026.xURLPMID:22269069 [本文引用: 1]
090004Overexpression of AtPAP2, a purple acid phosphatase (PAP) with a unique C-terminal hydrophobic motif in Arabidopsis, resulted in earlier bolting and a higher seed yield. Metabolite analysis showed that the shoots of AtPAP2 overexpression lines contained higher levels of sugars and tricarboxylic acid (TCA) metabolites. Enzyme assays showed that sucrose phosphate synthase (SPS) activity was significantly upregulated in the overexpression lines. The higher SPS activity arose from a higher level of SPS protein, and was independent of SnRK1.090004AtPAP2 was found to be targeted to both plastids and mitochondria via its C-terminal hydrophobic motif. Ectopic expression of a truncated AtPAP2 without this C-terminal motif in Arabidopsis indicated that the subcellular localization of AtPAP2 is essential for its biological actions.090004Plant PAPs are generally considered to mediate phosphorus acquisition and redistribution. AtPAP2 is the first PAP shown to modulate carbon metabolism and the first shown to be dual-targeted to both plastids and mitochondria by a C-terminal targeting signal.090004One PAP-like sequence carrying a hydrophobic C-terminal motif could be identified in the genome of the smallest free-living photosynthetic eukaryote, Ostreococcus tauri. This might reflect a common ancestral function of AtPAP2-like sequences in the regulation of carbon metabolism.
.
DOI:10.1016/j.mito.2017.04.006URLPMID:28478183 [本文引用: 1]
Overexpression of AtPAP2, a phosphatase located on the outer membranes of chloroplasts and mitochondria, leads to higher energy outputs from these organelles. AtPAP2 interacts with seven MORF proteins of the editosome complex. RNA-sequencing analysis showed that the editing degrees of most sites did not differ significantly between OE and WT, except some sites on the transcripts of several cytochrome c maturation (Ccm) genes. Western blotting of 2D BN-PAGE showed that the patterns of CcmF N1 polypeptides were different between the lines. We proposed that AtPAP2 may influence cytochrome c biogenesis by modulating RNA editing through its interaction with MORF proteins.
DOI:10.1002/9781118958841.ch10URL [本文引用: 5]
http://doi.wiley.com/10.1002/9781118958841.ch10
DOI:10.1016/j.plantsci.2010.04.005URL [本文引用: 1]
Phosphate (Pi) is an essential, but limiting macronutrient that plays critical roles in plant metabolism and development. Plants have evolved an intricate array of adaptations to enhance Pi acquisition and utilization from their environment. The availability of the complete genome sequence of the model plant Arabidopsis thaliana, together with a wide assortment of related genomic resources, has significantly advanced our understanding of the adaptations of Pi-starved plants. Information on the genetic identity, subcellular location, biochemical properties, and probable functions of acid phosphatases involved in the Pi metabolism of Pi-starved Arabidopsis is beginning to emerge. Acid phosphatases catalyze the hydrolysis of Pi from a broad range of phosphomonoesters with an acidic pH optimum. The Arabidopsis genome encodes 29 different purple acid phosphatases whose expression is influenced by various developmental and environmental factors. Pi starvation induces de novo synthesis of several extra- and intracellular Arabidopsis purple acid phosphatase isozymes; AtPAP12 and AtPAP26 appear to be the principal root-secreted acid phosphatases that scavenge Pi from extracellular Pi-esters, whereas the dual-targeted AtPAP26 is the predominant intracellular acid phosphatase that functions in vacuolar Pi recycling by Pi-starved Arabidopsis. The identification and functional characterization of intracellular and secreted purple acid phosphatase isozymes upregulated by Pi-deprived plants may help develop strategies for engineering Pi-efficient crops, thereby minimizing the use of unsustainable Pi fertilizers in agriculture.
.
[本文引用: 2]
DOI:10.1104/pp.127.2.390URLPMID:11598215 [本文引用: 1]
Since 1960 the world's population has doubled to 6 billion people. During the next 40 years the population is projected to stabilize at 8-9 billion people. The expanded use of fertilizers, water, and improved germplasm (Green Revolution) has allowed food production to outpace population growth to the present day. Increases in food production to feed a billion people will strain the limits of agriculture. High use of nitrogen and phosphorus in intensive agriculture have compromised air and water quality. Without sustainable practices these problems will be exacerbated in the future. Moreover, the fact that nitrogen and phosphorus fertilizer production is nonrenewable will result in high prices thereby making fertilizer application untenable for subsistence farmers. The crucial question facing agriculture is, "How can plant and crop growth needs be met through sustainable, environmentally friendly approaches?" This essay addresses those concerns.
DOI:10.1046/j.1469-8137.2003.00695.xURL [本文引用: 1]
Phosphorus (P) is limiting for crop yield on > 30% of the world's arable land and, by some estimates, world resources of inexpensive P may be depleted by 2050. Improvement of P acquisition and use by plants is critical for economic, humanitarian and environmental reasons. Plants have evolved a diverse array of strategies to obtain adequate P under limiting conditions, including modifications to root architecture, carbon metabolism and membrane structure, exudation of low molecular weight organic acids, protons and enzymes, and enhanced expression of the numerous genes involved in low-P adaptation. These adaptations may be less pronounced in mycorrhizal-associated plants. The formation of cluster roots under P-stress by the nonmycorrhizal species white lupin (Lupinus albus), and the accompanying biochemical changes exemplify many of the plant adaptations that enhance P acquisition and use. Physiological, biochemical, and molecular studies of white lupin and other species response to P-deficiency have identified targets that may be useful for plant improvement. Genomic approaches involving identification of expressed sequence tags (ESTs) found under low-P stress may also yield target sites for plant improvement. Interdisciplinary studies uniting plant breeding, biochemistry, soil science, and genetics under the large umbrella of genomics are prerequisite for rapid progress in improving nutrient acquisition and use in plants.
DOI:10.1111/j.1469-8137.2012.04190.xURLPMID:22691045 [本文引用: 1]
Summary306I.The need to use phosphorus efficiently307II.P-use efficiency and P dynamics in a growing crop307III.P pools in plants307IV.Phosphorus pools and growth rates310V.Are crops different from other plants in their P concentration?310VI.Phosphorus use and photosynthesis311VII.Crop development and canopy P distribution312VIII.Internal redistribution of P in a growing vegetative plant313IX.Allocation of P to reproductive structures314X.Constraints to P remobilisation315XI.Do physiological or phylogenetic trade-offs constrain traits that could improve PUE?316XII.Identifying genetic loci associated with PUE316XIII.Conclusions317 Acknowledgements317 References317SummaryLimitation of grain crop productivity by phosphorus (P) is widespread and will probably increase in the future. Enhanced P efficiency can be achieved by improved uptake of phosphate from soil (P-acquisition efficiency) and by improved productivity per unit P taken up (P-use efficiency). This review focuses on improved P-use efficiency, which can be achieved by plants that have overall lower P concentrations, and by optimal distribution and redistribution of P in the plant allowing maximum growth and biomass allocation to harvestable plant parts. Significant decreases in plant P pools may be possible, for example, through reductions of superfluous ribosomal RNA and replacement of phospholipids by sulfolipids and galactolipids. Improvements in P distribution within the plant may be possible by increased remobilization from tissues that no longer need it (e.g. senescing leaves) and reduced partitioning of P to developing grains. Such changes would prolong and enhance the productive use of P in photosynthesis and have nutritional and environmental benefits. Research considering physiological, metabolic, molecular biological, genetic and phylogenetic aspects of P-use efficiency is urgently needed to allow significant progress to be made in our understanding of this complex trait.
[本文引用: 2]
[本文引用: 1]
DOI:10.1023/A:1022332320384URL [本文引用: 4]
The roots of white lupin (Lupinus albus L. cv. Kievskij mutant) secrete acid phosphatase, S-APase, when they grow under conditions of low available phosphorus (P). S-APases hydrolyze organic phosphate compounds in the rhizosphere and supply inorganic phosphate to the plants. Low phosphorus availability also induces vigorous growth of cluster roots. In this study, the function of cluster roots was investigated with reference to S-APase secretion. White lupins were grown in hydroponic culture in a greenhouse under P-deficient and P-sufficient conditions. S-APase in the excised roots after treatment was detected by staining with 4-methylumbelliferone phosphate (MUP). Gene expression of S-APase in cluster and normal roots was also investigated. Activity was greatest in the roots of plants grown under conditions of P -deficiency, particularly in cluster roots. S-APase gene expression was induced by a decrease in internal P concentrations, and was especially high in cluster roots formed under conditions of P -deficiency. It was suggested that decrease of internal P concentration stimulated both of the S-APase expression and cluster root formation.
DOI:10.1111/j.1438-8677.2010.00346.xURLPMID:21143719 [本文引用: 4]
Abstract Purple acid phosphatases (PAPs) are a family of metallo-phosphoesterases involved in a variety of physiological functions, especially phosphate deficiency adaptations in plants. We identified 26 putative PAP genes by a genome-wide analysis of rice ( Oryza sativa ), 24 of which have isolated EST sequences in the dbEST database. Amino acid sequence analysis revealed that 25 of these genes possess sets of metal-ligating residues typical of known PAPs. Phylogenetic analysis classified the 26 rice and 29 Arabidopsis PAPs into three main groups and seven subgroups. We detected transcripts of 21 PAP genes in roots or leaves of rice seedlings. The expression levels of ten PAP genes were up-regulated by both phosphate deprivation and over-expression of the transcription factor OsPHR2 . These PAP genes all contained one or two OsPHR2 binding elements in their promoter regions, implying that they are directly regulated by OsPHR2 . Both acid phosphatase (AP) and surface secretory acid phosphatase (SAP) activity assays showed that the up-regulation of PAPs by Pi starvation, OsPHR2 over-expression, PHO2 knockout or OsSPX1 RNA interference led to an increase in AP and SAP activity in rice roots. This study reveals the potential for developing technologies for crop improvement in phosphorus use efficiency.
DOI:10.1080/15592324.2016.1239687URLPMID:27700374 [本文引用: 1]
Arabidopsis thaliana purple acid phosphatase 2 (AtPAP2) is the only phosphatase that is dual-targeted to both chloroplasts and mitochondria. Like Toc33/34 of the TOC and Tom 20 of the TOM, AtPAP2 is anchored to the outer membranes of chloroplasts and mitochondria via a hydrophobic C-terminal motif. AtPAP2 on the mitochondria was previously shown to recognize the presequences of several nuclear-encoded mitochondrial proteins and modulate the import of pMORF3 into the mitochondria. Here we show that AtPAP2 binds to the small subunit of Rubisco (pSSU) and that chloroplast import experiments demonstrated that pSSU was imported less efficiently into pap2 chloroplasts than into wild-type chloroplasts. We propose that AtPAP2 is an outer membrane-bound phosphatase receptor that facilitates the import of selected proteins into chloroplasts.
DOI:10.1104/pp.107.109934URLPMID:18065557 [本文引用: 2]
Ascorbate (AsA) is the most abundant antioxidant in plant cells and a cofactor for a large number of key enzymes. However, the mechanism of how AsA levels are regulated in plant cells remains unknown. The Arabidopsis (Arabidopsis thaliana) activation-tagged mutant AT23040 showed a pleiotropic phenotype, including ozone resistance, rapid growth, and leaves containing higher AsA than wild-type plants. The phenotype was caused by activation of a purple acid phosphatase (PAP) gene, AtPAP15, which contains a dinuclear metal center in the active site. AtPAP15 was universally expressed in all tested organs in wild-type plants. Overexpression of AtPAP15 with the 35S cauliflower mosaic virus promoter produced mutants with up to 2-fold increased foliar AsA, 20% to 30% decrease in foliar phytate, enhanced salt tolerance, and decreased abscisic acid sensitivity. Two independent SALK T-DNA insertion mutants in AtPAP15 had 30% less foliar AsA and 15% to 20% more phytate than wild-type plants and decreased tolerance to abiotic stresses. Enzyme activity of partially purified AtPAP15 from plant crude extract and recombinant AtPAP15 expressed in bacteria and yeast was highest when phytate was used as substrate, indicating that AtPAP15 is a phytase. Recombinant AtPAP15 also showed enzyme activity on the substrate myoinositol-1-phosphate, indicating that the AtPAP15 is a phytase that hydrolyzes myoinositol hexakisphosphate to yield myoinositol and free phosphate. Myoinositol is a known precursor for AsA biosynthesis in plants. Thus, AtPAP15 may modulate AsA levels by controlling the input of myoinositol into this branch of AsA biosynthesis in Arabidopsis.
.
DOI:10.1016/j.febslet.2014.08.019URLPMID:25173632 [本文引用: 1]
Changes in carbon flow and sink/source activities can affect floral, architectural, and reproductive traits of plants. In potato, overexpression (OE) of the purple acid phosphatase 2 of Arabidopsis (AtPAP2) resulted in earlier flowering, faster growth rate, increased tubers and tuber starch content, and higher photosynthesis rate. There was a significant change in sucrose, glucose and fructose levels in leaves, phloem and sink biomass of the OE lines, consistent with an increased expression of sucrose transporter 1 (StSUT1). Furthermore, the expression levels and enzyme activity of sucrose-phosphate synthase (SPS) were also significantly increased in the OE lines. These findings strongly suggest that higher carbon supply from the source and improved sink strength can improve potato tuber yield.
.
DOI:10.1186/1754-6834-5-19URLPMID:3361479 [本文引用: 1]
Background Lipids extracted from seeds of Camelina sativa have been successfully used as a reliable source of aviation biofuels. This biofuel is environmentally friendly because the drought resistance, frost tolerance and low fertilizer requirement of Camelina sativa allow it to grow on marginal lands. Improving the species growth and seed yield by genetic engineering is therefore a target for the biofuels industry. In Arabidopsis, overexpression of purple acid phosphatase 2 encoded by Arabidopsis (AtPAP2) promotes plant growth by modulating carbon metabolism. Overexpression lines bolt earlier and produce 50% more seeds per plant than wild type. In this study, we explored the effects of overexpressing AtPAP2 in Camelina sativa. Results Under controlled environmental conditions, overexpression of AtPAP2 in Camelina sativa resulted in longer hypocotyls, earlier flowering, faster growth rate, higher photosynthetic rate and stomatal conductance, increased seed yield and seed size in comparison with the wild-type line and null-lines. Similar to transgenic Arabidopsis, activity of sucrose phosphate synthase in leaves of transgenic Camelina was also significantly up-regulated. Sucrose produced in photosynthetic tissues supplies the building blocks for cellulose, starch and lipids for growth and fuel for anabolic metabolism. Changes in carbon flow and sink/source activities in transgenic lines may affect floral, architectural, and reproductive traits of plants. Conclusions Lipids extracted from the seeds of Camelina sativa have been used as a major constituent of aviation biofuels. The improved growth rate and seed yield of transgenic Camelina under controlled environmental conditions have the potential to boost oil yield on an area basis in field conditions and thus make Camelina-based biofuels more environmentally friendly and economically attractive.
DOI:10.1104/pp.107.111443URLPMID:18263782 [本文引用: 1]
Previous research has demonstrated that AtPHR1 plays a central role in phosphate (Pi)-starvation signaling in Arabidopsis thaliana. In this work, two OsPHR genes from rice (Oryza sativa) were isolated and designated as OsPHR1 and OsPHR2 based on amino acid sequence homology to AtPHR1. Their functions in Pi signaling in rice were investigated using transgenic plants. Our results showed that both OsPHR1 and OsPHR2 are involved in Pi-starvation signaling pathway by regulation of the expression of Pi-starvation-induced genes, whereas only OsPHR2 overexpression results in the excessive accumulation of Pi in shoots under Pi-sufficient conditions. Under Pi-sufficient conditions, overexpression of OsPHR2 mimics Pi-starvation stress in rice with enhanced root elongation and proliferated root hair growth, suggesting the involvement of OsPHR2 in Pi-dependent root architecture alteration by both systematic and local pathways. In OsPHR2-overexpression plants, some Pi transporters were up-regulated under Pi-sufficient conditions, which correlates with the strongly increased content of Pi. The mechanism behind the OsPHR2 regulated Pi accumulation will provide useful approaches to develop smart plants with high Pi efficiency.
.
DOI:10.1007/s11103-005-0183-0URLPMID:16244908 [本文引用: 1]
Purple acid phosphatases (PAPs) are metallo-phosphoesterases. Their expression and function have not been systematically investigated in higher plants. In this work, we compared the transcript levels of 28 Arabidopsis PAP ( AtPAP ) genes in five Arabidopsis organs. The 28 members, although differed in their expression patterns in vegetative organs, were all transcribed in flower. Furthermore, the transcription of seven members ( AtPAPs 6 , 11 , 14 , 19 , 23 , 24 and 25 ) occurred predominantly in the flower. To begin dissecting the role of AtPAP genes in flower development, further expression and functional analyses were conducted using AtPAP23 . Histochemical staining of transgenic plants expressing AtPAP23 promoter-beta-glucuronidase (GUS) gene construct revealed that AtPAP23 transcription was strong in flower apical meristems, but became restricted to petals and anther filaments in fully developed flower. A GST (glutathione S-transferase) fusion protein of AtPAP23 (GST:AtPAP23) was expressed in bacterial cells, and was found to contain significant amounts of Fe and Mn (whereas the control GST protein contained none). In biochemical tests, GST:AtPAP23 showed typical acid phosphatase activities. The fusion protein was also highly active on phosphoserine, but not phosphotyrosine. Despite its highly specific expression pattern and the demonstrated biochemical function of its protein product, the RNAi (RNA interference), T-DNA knock-out and overexpression lines of AtPAP23 were indistinguishable from wild type plants in the development of flower (or other organs). Interestingly, the Fe and Mn contents were found significantly increased in AtPAP23 overexpression lines, which may offer a new direction for further functional studies of AtPAPs in Arabidopsis .
DOI:10.1055/s-2004-821091URLPMID:15375722 [本文引用: 1]
Abstract: Three cDNAs encoding purple acid phosphatase (PAP) were cloned from potato ( Solanum tuberosum L. cv. D sir e) and expression of the corresponding genes was characterised. StPAP1 encodes a low-molecular weight PAP clustering with mammalian, cyanobacterial, and other plant PAPs. It was highly expressed in stem and root and its expression did not change in response to phosphorus (P) deprivation. StPAP2 and StPAP3 code for high-molecular weight PAPs typical for plants. Corresponding gene expression was shown to be responsive to the level of P supply, with transcripts of StPAP2 and StPAP3 being most abundant in P-deprived roots or both stem and roots, respectively. Root colonisation by arbuscular mycorrhizal fungi had no effect on the expression of any of the three PAP genes. StPAP1 mRNA is easily detectable along the root axis, including root hairs, but is barely detectable in root tips. In contrast, both StPAP2 and StPAP3 transcripts are abundant along the root axis, but absent in root hairs, and are most abundant in the root tip. All three PAPs described contain a predicted N-terminal secretion signal and could play a role in extracellular P scavenging, P mobilisation from the rhizosphere, or cell wall regeneration.
低磷胁迫下大麦叶片磷素利用特征
1
2016
... 磷是植物生长发育过程中必需的六大营养元素之一, 对植物新陈代谢和正常生长发育有着极其重要的作用.磷元素不仅参与植物细胞内许多化合物的生物合成, 蛋白磷酸化和去磷酸化等生理生化过程, 而且对植物生长发育及作物产量形成等也有一定的影响(
甘蓝和白菜紫色酸性磷酸酶17基因家族的克隆和比较分析
1
2010
... 紫色酸性磷酸酶(purple acid phosphatases, PAPs)属于双核金属脱氢酶(binuclear metallohydro- lase)家族, 是一种广泛存在于动物和植物体内的酸性磷酸酶类.在植物体的弱酸性(pH4-7)条件下, PAPs能够催化水解磷酸单酯和酸酐类有机物(如ATP、ADP和糖脂)并释放出无机磷, 供植物吸收利用, 从而提高植物对磷的利用率(
植物磷营养高效的分子生物学研究进展
1
2005
... 磷是植物生长发育过程中必需的六大营养元素之一, 对植物新陈代谢和正常生长发育有着极其重要的作用.磷元素不仅参与植物细胞内许多化合物的生物合成, 蛋白磷酸化和去磷酸化等生理生化过程, 而且对植物生长发育及作物产量形成等也有一定的影响(
Plant nutrition research: priorities to meet human needs for food in sustainable ways
1
2002
... 磷是植物生长发育过程中必需的六大营养元素之一, 对植物新陈代谢和正常生长发育有着极其重要的作用.磷元素不仅参与植物细胞内许多化合物的生物合成, 蛋白磷酸化和去磷酸化等生理生化过程, 而且对植物生长发育及作物产量形成等也有一定的影响(
The cell wall- targeted purple acid phosphatase AtPAP25 is critical for acclimation of Arabidopsis thaliana to nutritional phosphorus deprivation
0
2014
Genetic responses to phosphorus deficiency
1
2004
... 磷是植物生长发育过程中必需的六大营养元素之一, 对植物新陈代谢和正常生长发育有着极其重要的作用.磷元素不仅参与植物细胞内许多化合物的生物合成, 蛋白磷酸化和去磷酸化等生理生化过程, 而且对植物生长发育及作物产量形成等也有一定的影响(
Molecular characterization of OsPAP2: transgenic expression of a purple acid phosphatase up-regulated in phosphate-deprived rice sus- pension cells
1
2010
... PAPs家族I所包含的种类最多, 该家族可进一步分为I-1和I-2两个亚家族.I-1亚家族成员已被证实参与植物磷和碳代谢途径(
Dependence of photosynthesis of sunflower and maize leaves on phosphate supply, ribulose-1,5-bisphosphate carboxylase/oxygenase activity, and ribulose-1,5-bisphosphate pool size
1
1992
... 磷是植物生长发育过程中必需的六大营养元素之一, 对植物新陈代谢和正常生长发育有着极其重要的作用.磷元素不仅参与植物细胞内许多化合物的生物合成, 蛋白磷酸化和去磷酸化等生理生化过程, 而且对植物生长发育及作物产量形成等也有一定的影响(
Synthesis, characterization, and reactivity studies of heterodinuclear complexes modeling active sites in purple acid phospha- tases
1
2011
... 紫色酸性磷酸酶因其在溶液中表现出特殊的紫色或粉色而命名, 而呈现粉紫色是由发色基团铁离子和酪氨酸之间的电子转移所致.PAPs的催化位点和结构域高度保守, 其结构的基本特征在于含有5个保守基序(β/α/β/α/β)、7个结合金属离子的氨基酸残基(DXG/GDXXY/GNH(D/E)/VXXH/GHXH)和1个金属离子的双核中心(
Purple acid phosphatase in the walls of tobacco cells
1
2008
... 烟草中, NtPAP12可催化重组的木糖苷酶磷酸化和去磷酸化以及葡糖苷酶去磷酸化, 抑制木葡聚糖降解, 增加木聚糖的合成; 还可通过葡糖苷酶去磷酸化, 抑制纤维寡糖的降解, 进而增加纤维素和胼胝质的合成.NtPAP12过表达株系糖苷酶活性下降, 质外体中木葡聚糖和纤维寡糖含量上升, 细胞壁主要成分(木聚糖、胼胝质和纤维素)的含量增加, 证实NtPAP12能够调节木糖苷酶以及葡糖苷酶的活性, 从而促进细胞壁的合成(
Isolation and characterization of four cell wall purple acid phosphatase genes from tobacco cells
1
2003
... 烟草中, NtPAP12可催化重组的木糖苷酶磷酸化和去磷酸化以及葡糖苷酶去磷酸化, 抑制木葡聚糖降解, 增加木聚糖的合成; 还可通过葡糖苷酶去磷酸化, 抑制纤维寡糖的降解, 进而增加纤维素和胼胝质的合成.NtPAP12过表达株系糖苷酶活性下降, 质外体中木葡聚糖和纤维寡糖含量上升, 细胞壁主要成分(木聚糖、胼胝质和纤维素)的含量增加, 证实NtPAP12能够调节木糖苷酶以及葡糖苷酶的活性, 从而促进细胞壁的合成(
Activation of β-glucan synthases by wall-bound purple acid phosphatase in tobacco cells
1
2009
... 烟草中, NtPAP12可催化重组的木糖苷酶磷酸化和去磷酸化以及葡糖苷酶去磷酸化, 抑制木葡聚糖降解, 增加木聚糖的合成; 还可通过葡糖苷酶去磷酸化, 抑制纤维寡糖的降解, 进而增加纤维素和胼胝质的合成.NtPAP12过表达株系糖苷酶活性下降, 质外体中木葡聚糖和纤维寡糖含量上升, 细胞壁主要成分(木聚糖、胼胝质和纤维素)的含量增加, 证实NtPAP12能够调节木糖苷酶以及葡糖苷酶的活性, 从而促进细胞壁的合成(
Potential role for purple acid phosphatase in the dephosphorylation of wall proteins in tobacco cells
1
2010
... PAPs家族I所包含的种类最多, 该家族可进一步分为I-1和I-2两个亚家族.I-1亚家族成员已被证实参与植物磷和碳代谢途径(
The amino acid sequence of the red kidney bean Fe(III)-Zn(II) purple acid phosphatase. Determination of the amino acid sequence by a combination of matrix-assisted laser desorption/ioni- zation mass spectrometry and automated Edman sequ- encing
1
1994
... 菜豆中克隆出的KBPAP (检索号P80366)分子量为111 kDa, 属于大分子量PAP.KBPAP是1个二聚体酶, 包含2个糖基化的亚基, 2个亚基通过二硫键共价相连.每个亚基包含432个氨基酸残基和5个N-糖基化的天冬酰胺残基, 并且同时具有N-端和C-端2个结构域.C-端结构域分子量较大, 具有催化功能; N-端结构域较小, 由2个β-折叠组成.每个β-折叠由3个反向平行的β-链构成, 且不具有金属磷酸酶催化活性(
Structural relationship between the mammalian Fe(III)-Fe(II) and the Fe(III)-Zn(II) plant purple acid phosphatases
1
1995
... 菜豆中克隆出的KBPAP (检索号P80366)分子量为111 kDa, 属于大分子量PAP.KBPAP是1个二聚体酶, 包含2个糖基化的亚基, 2个亚基通过二硫键共价相连.每个亚基包含432个氨基酸残基和5个N-糖基化的天冬酰胺残基, 并且同时具有N-端和C-端2个结构域.C-端结构域分子量较大, 具有催化功能; N-端结构域较小, 由2个β-折叠组成.每个β-折叠由3个反向平行的β-链构成, 且不具有金属磷酸酶催化活性(
GmPAP4, a novel purple acid phosphatase gene isolated from soybean ( Glycine max), enhanced extracellular phytate utilization in Arabidopsis thaliana
0
2014
Molecular and biochemical characterization of AtPAP15, a purple acid phosphatase with phytase activity, in Arabidopsis
1
2009
... PAPs家族I所包含的种类最多, 该家族可进一步分为I-1和I-2两个亚家族.I-1亚家族成员已被证实参与植物磷和碳代谢途径(
Purification and characterization of purple acid phosphatase PAP1 from dry powder of sweet potato
1
2003
... 植物PAPs与土壤有机磷的分解吸收及植株体内磷素的再利用有密切关系, 很多植物来源的PAPs都具有酸性磷酸酶活性, 能够将植物体内外的有机磷水解成无机磷, 供植物吸收利用.根据PAPs是否具有分泌到细胞外的性质, 可将其分为细胞内酸性磷酸酶(intracellular acid phosphatase, IAP)和分泌型酸性磷酸酶(secreted acid phosphatase, SAP).细胞内酸性磷酸酶主要水解植物体内磷库的磷, 进行细胞内磷素的再利用; 分泌型酸性磷酸酶则被分泌出细胞外, 活化土壤中的有机磷组分, 释放出供植物利用的无机磷(
Phosphorylation and dephosphorylation of the presequence of precursor MULTIPLE ORGANELLAR RNA EDITING FACTOR3 during import into mitochondria from Arabidopsis
1
2015
... Sun等(2012b)研究表明, AtPAP2是一种尾部锚定蛋白, 含有N-端信号肽和C-端膜锚定信号肽.AtPAP2在拟南芥角果、茎、花、根和开始衰老的叶中高度表达, 但在叶和成熟的种子中表达量相对较低(
Identification of soybean purple acid phosphatase genes and their expression responses to phosphorus availability and symbiosis
1
2012
... 目前, PAPs已在多种植物中被鉴定和分离, 通过对拟南芥(Arabidopsis thaliana)和大豆等植物中的PAPs进行系统研究, 发现拟南芥中存在29个PAPs基因(
Purple acid phosphatases of Arabidopsis thaliana.Comparative analysis and differential regulation by phosphate deprivation
2
2002
... 1998年, Hiroshi等用同位素标记法, 发现低磷会诱导浮萍(Spirodela oligorrhiza)产生57 kDa的磷脂酰肌醇锚定磷酸酶, 该酶含有Fe3+-Mn2+双核金属中心, 活性受到酒石酸抑制.这是首次在高等植物中发现紫色酸性磷酸酶(
... 目前, PAPs已在多种植物中被鉴定和分离, 通过对拟南芥(Arabidopsis thaliana)和大豆等植物中的PAPs进行系统研究, 发现拟南芥中存在29个PAPs基因(
Ectopic expression of GmPAP3 alleviates oxidative damage caused by salinity and osmotic stresses
2
2008
... PAPs家族I所包含的种类最多, 该家族可进一步分为I-1和I-2两个亚家族.I-1亚家族成员已被证实参与植物磷和碳代谢途径(
... 大豆酸性磷酸酶3基因(GmPAP3)定位在线粒体(活性氧产生的主要部位), 其表达受盐和氧胁迫诱导, 可能在非生物逆境响应过程中发挥作用.将GmPAP3过表达植株分别进行盐、PEG以及农药胁迫处理, 对比野生型发现过表达植株的根长增加.此外, Gm- PAP3能够通过抑制活性氧(ROS)的积累, 减轻盐和过氧化胁迫造成的损伤(
Global small RNA analysis in fast-growing Arabidopsis thaliana with elevated concentrations of ATP and sugars
1
2014
... Sun等(2012b)研究表明, AtPAP2是一种尾部锚定蛋白, 含有N-端信号肽和C-端膜锚定信号肽.AtPAP2在拟南芥角果、茎、花、根和开始衰老的叶中高度表达, 但在叶和成熟的种子中表达量相对较低(
Impacts of high ATP supply from chloroplasts and mitochondria on the leaf metabolism of Arabidopsis thaliana
1
2015
... Sun等(2012b)研究表明, AtPAP2是一种尾部锚定蛋白, 含有N-端信号肽和C-端膜锚定信号肽.AtPAP2在拟南芥角果、茎、花、根和开始衰老的叶中高度表达, 但在叶和成熟的种子中表达量相对较低(
Biochemical and molecular characterization of PvPAP3, a novel purple acid phosphatase isolated from common bean enhancing extracellular ATP utilization
2
2010
... 小分子量的紫色酸性磷酸酶(low molecular weight PAPs, LMW PAPs)以单聚体形式存在, 与哺乳动物和蓝藻体内的小分子量PAPs极为相似, 单体分子量一般约为35 kDa, 在结构上仅保留了典型的C-端结构域(
... PAPs家族I所包含的种类最多, 该家族可进一步分为I-1和I-2两个亚家族.I-1亚家族成员已被证实参与植物磷和碳代谢途径(
GmPAP3, a novel purple acid phosphatase-like gene in soybean induced by NaCl stress but not phosphorus deficiency
1
2003
... 大豆酸性磷酸酶3基因(GmPAP3)定位在线粒体(活性氧产生的主要部位), 其表达受盐和氧胁迫诱导, 可能在非生物逆境响应过程中发挥作用.将GmPAP3过表达植株分别进行盐、PEG以及农药胁迫处理, 对比野生型发现过表达植株的根长增加.此外, Gm- PAP3能够通过抑制活性氧(ROS)的积累, 减轻盐和过氧化胁迫造成的损伤(
The glycosylphosphatidylinositol-anchored phosphatase from Spirodela oligorrhiza is a purple acid phosphatase
1
1998
... 1998年, Hiroshi等用同位素标记法, 发现低磷会诱导浮萍(Spirodela oligorrhiza)产生57 kDa的磷脂酰肌醇锚定磷酸酶, 该酶含有Fe3+-Mn2+双核金属中心, 活性受到酒石酸抑制.这是首次在高等植物中发现紫色酸性磷酸酶(
Plant purple acid phosphatases-genes, structures and biological function
2
2003
... 紫色酸性磷酸酶(purple acid phosphatases, PAPs)属于双核金属脱氢酶(binuclear metallohydro- lase)家族, 是一种广泛存在于动物和植物体内的酸性磷酸酶类.在植物体的弱酸性(pH4-7)条件下, PAPs能够催化水解磷酸单酯和酸酐类有机物(如ATP、ADP和糖脂)并释放出无机磷, 供植物吸收利用, 从而提高植物对磷的利用率(
... 紫色酸性磷酸酶在植物各个组织均有分布, 参与各种生命活动(
Purple acid phosphatase 5 is required for maintaining basal resistance against Pseudomonas syringae in Arabidopsis
1
2013
... PAPs家族I所包含的种类最多, 该家族可进一步分为I-1和I-2两个亚家族.I-1亚家族成员已被证实参与植物磷和碳代谢途径(
Eliminating the purple acid phosphatase AtPAP26 in Arabidopsis thaliana delays leaf senescence and impairs phosphorus remobilization
1
2012
... PAPs家族I所包含的种类最多, 该家族可进一步分为I-1和I-2两个亚家族.I-1亚家族成员已被证实参与植物磷和碳代谢途径(
b). The secreted purple acid phosphatase isozymes AtPAP12 and AtPAP26 play a pivotal role in extracellular phosphate-scavenging by Arabidopsis thaliana
2
2012
... 不同PAPs在植物体内的表达模式不尽相同, 它们分工合作, 在转录水平、翻译水平和翻译后水平受到调控.绝大部分植物PAPs的表达水平不仅与植物体内的无机磷含量相关, 而且与植物所处的逆境(氧和盐胁迫等)环境有关.对26个水稻PAPs编码基因进行启动子元件分析, 发现其中12个基因的启动子含有顺式作用元件P1BS (PHR1-binding site), P1BS是水稻磷信号代谢途径转录因子OsPHR2的结合元件(
... 此外, 虽然许多植物细胞内和分泌型酸性磷酸酶编码基因已被克隆, 但其表达调控机制至今未有详尽的解释.磷胁迫条件下, AtPAP26在转录水平并未发生变化.免疫印迹检测显示, AtPAP26在蛋白水平受到缺磷诱导, 但是其在蛋白水平具体的调控机制还不清楚(
A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae
1
2001
... 不同PAPs在植物体内的表达模式不尽相同, 它们分工合作, 在转录水平、翻译水平和翻译后水平受到调控.绝大部分植物PAPs的表达水平不仅与植物体内的无机磷含量相关, 而且与植物所处的逆境(氧和盐胁迫等)环境有关.对26个水稻PAPs编码基因进行启动子元件分析, 发现其中12个基因的启动子含有顺式作用元件P1BS (PHR1-binding site), P1BS是水稻磷信号代谢途径转录因子OsPHR2的结合元件(
Crystal structures of a purple acid phosphatase, representing different steps of this enzyme’s catalytic cycle
1
2008
... 紫色酸性磷酸酶因其在溶液中表现出特殊的紫色或粉色而命名, 而呈现粉紫色是由发色基团铁离子和酪氨酸之间的电子转移所致.PAPs的催化位点和结构域高度保守, 其结构的基本特征在于含有5个保守基序(β/α/β/α/β)、7个结合金属离子的氨基酸残基(DXG/GDXXY/GNH(D/E)/VXXH/GHXH)和1个金属离子的双核中心(
a). Identification of mammalian-like purple acid phosphatases in a wide range of plants
1
2000
... 小分子量的紫色酸性磷酸酶(low molecular weight PAPs, LMW PAPs)以单聚体形式存在, 与哺乳动物和蓝藻体内的小分子量PAPs极为相似, 单体分子量一般约为35 kDa, 在结构上仅保留了典型的C-端结构域(
b). Purple acid phosphatases from bacteria: similarities to mammalian and plant enzymes
3
2000
... 紫色酸性磷酸酶因其在溶液中表现出特殊的紫色或粉色而命名, 而呈现粉紫色是由发色基团铁离子和酪氨酸之间的电子转移所致.PAPs的催化位点和结构域高度保守, 其结构的基本特征在于含有5个保守基序(β/α/β/α/β)、7个结合金属离子的氨基酸残基(DXG/GDXXY/GNH(D/E)/VXXH/GHXH)和1个金属离子的双核中心(
... 大分子量的紫色酸性磷酸酶(high molecular weight PAPs, HMW PAPs)通常以同源二聚体的形式存在, 其单体分子量约为55 kDa, 此类PAPs在植物中所占比例较大, 并且与真菌和分枝杆菌的此类蛋白之间同源性较高(
... 小分子量的紫色酸性磷酸酶(low molecular weight PAPs, LMW PAPs)以单聚体形式存在, 与哺乳动物和蓝藻体内的小分子量PAPs极为相似, 单体分子量一般约为35 kDa, 在结构上仅保留了典型的C-端结构域(
Eutrophication science: where do we go from here?
1
2009
... 磷是植物生长发育过程中必需的六大营养元素之一, 对植物新陈代谢和正常生长发育有着极其重要的作用.磷元素不仅参与植物细胞内许多化合物的生物合成, 蛋白磷酸化和去磷酸化等生理生化过程, 而且对植物生长发育及作物产量形成等也有一定的影响(
Crystal structure of a purple acid phosphatase containing a dinuclear Fe(III)-Zn(II) active site
3
1995
... 紫色酸性磷酸酶因其在溶液中表现出特殊的紫色或粉色而命名, 而呈现粉紫色是由发色基团铁离子和酪氨酸之间的电子转移所致.PAPs的催化位点和结构域高度保守, 其结构的基本特征在于含有5个保守基序(β/α/β/α/β)、7个结合金属离子的氨基酸残基(DXG/GDXXY/GNH(D/E)/VXXH/GHXH)和1个金属离子的双核中心(
... 菜豆紫色酸性磷酸酶(KBPAP)的三维结构(
... (
b). AtPAP2 is a tail-anchored protein in the outer membrane of chloroplasts and mitochondria
3
2012
... PAPs家族I所包含的种类最多, 该家族可进一步分为I-1和I-2两个亚家族.I-1亚家族成员已被证实参与植物磷和碳代谢途径(
... Sun等(2012b)研究表明, AtPAP2是一种尾部锚定蛋白, 含有N-端信号肽和C-端膜锚定信号肽.AtPAP2在拟南芥角果、茎、花、根和开始衰老的叶中高度表达, 但在叶和成熟的种子中表达量相对较低(
... )和马铃薯转基因植株抽苔提前, 生长速度加快, 叶片光合速率提高10%-20%, 种子体积及生物量增加; 同时, 蔗糖磷酸合酶的活性显著升高, 嫩枝含有更高水平的糖类(蔗糖和己糖)、丙氨酸和脯氨酸等三羧酸循环(TCA)代谢产物(
Global transcriptome analysis of AtPAP2-overex- pressing Arabidopsis thaliana with elevated ATP
1
2013
... Sun等(2012b)研究表明, AtPAP2是一种尾部锚定蛋白, 含有N-端信号肽和C-端膜锚定信号肽.AtPAP2在拟南芥角果、茎、花、根和开始衰老的叶中高度表达, 但在叶和成熟的种子中表达量相对较低(
A dual-targeted purple acid phosphatase in Arabidopsis thaliana moderates carbon metabolism and its overexpression leads to faster plant growth and higher seed yield
1
2012
... Sun等(2012b)研究表明, AtPAP2是一种尾部锚定蛋白, 含有N-端信号肽和C-端膜锚定信号肽.AtPAP2在拟南芥角果、茎、花、根和开始衰老的叶中高度表达, 但在叶和成熟的种子中表达量相对较低(
RNA editing of cytochrome c maturation transcripts is responsive to the energy status of leaf cells in Arabidopsis thaliana
1
2017
... Sun等(2012b)研究表明, AtPAP2是一种尾部锚定蛋白, 含有N-端信号肽和C-端膜锚定信号肽.AtPAP2在拟南芥角果、茎、花、根和开始衰老的叶中高度表达, 但在叶和成熟的种子中表达量相对较低(
5
... 目前, PAPs已在多种植物中被鉴定和分离, 通过对拟南芥(Arabidopsis thaliana)和大豆等植物中的PAPs进行系统研究, 发现拟南芥中存在29个PAPs基因(
... 植物PAPs物种间的进化关系(
... At: 拟南芥; Os: 水稻; Gm: 大豆; Mt: 蒺藜苜蓿; Nt: 烟草; Zm: 玉米; Hv: 大麦; Ta: 小麦; Vr: 绿豆; Pv: 菜豆
... 植物PAP的生物学特性和功能(
... Biochemical properties functions of plant PAPs (
a). Feeding hungry plants: the role of purple acid phosphatases in phosphate nutrition
1
2010
... 此外, 虽然许多植物细胞内和分泌型酸性磷酸酶编码基因已被克隆, 但其表达调控机制至今未有详尽的解释.磷胁迫条件下, AtPAP26在转录水平并未发生变化.免疫印迹检测显示, AtPAP26在蛋白水平受到缺磷诱导, 但是其在蛋白水平具体的调控机制还不清楚(
b). Biochemical and molecular characterization of AtPAP12 and AtPAP26: the predominant purple acid phosphatase isozymes secreted by phosphate- starved Arabidopsis thaliana
2
2010
... 大部分PAP都具有N-端信号肽, 这个信号肽对PAP蛋白的锚定和生物学功能具有重要作用.信号肽定位预测表明, 少数PAPs定位于线粒体(
... ).由于AtPAP10、AtPAP12和AtPAP26蛋白的N-端都存在至少1个糖基化修饰位点, 故这些蛋白的糖基化修饰可能直接影响它们的细胞定位(
Symbiotic nitrogen fixation and phosphorus acquisition. plant nutrition in a world of declining renewable resources
1
2001
... 磷是植物生长发育过程中必需的六大营养元素之一, 对植物新陈代谢和正常生长发育有着极其重要的作用.磷元素不仅参与植物细胞内许多化合物的生物合成, 蛋白磷酸化和去磷酸化等生理生化过程, 而且对植物生长发育及作物产量形成等也有一定的影响(
Phosphorus acquisition and use: critical adaptations by plants for securing a nonrenewable resource
1
2003
... 磷是植物生长发育过程中必需的六大营养元素之一, 对植物新陈代谢和正常生长发育有着极其重要的作用.磷元素不仅参与植物细胞内许多化合物的生物合成, 蛋白磷酸化和去磷酸化等生理生化过程, 而且对植物生长发育及作物产量形成等也有一定的影响(
Opportunities for improving phosphorus-use efficiency in crop plants
1
2012
... 植物PAPs与土壤有机磷的分解吸收及植株体内磷素的再利用有密切关系, 很多植物来源的PAPs都具有酸性磷酸酶活性, 能够将植物体内外的有机磷水解成无机磷, 供植物吸收利用.根据PAPs是否具有分泌到细胞外的性质, 可将其分为细胞内酸性磷酸酶(intracellular acid phosphatase, IAP)和分泌型酸性磷酸酶(secreted acid phosphatase, SAP).细胞内酸性磷酸酶主要水解植物体内磷库的磷, 进行细胞内磷素的再利用; 分泌型酸性磷酸酶则被分泌出细胞外, 活化土壤中的有机磷组分, 释放出供植物利用的无机磷(
The Arabidopsis purple acid phosphatase AtPAP10 is predominantly associated with the root surface and plays an important role in plant tolerance to phosphate limitation
2
2011
... 不同PAPs在植物体内的表达模式不尽相同, 它们分工合作, 在转录水平、翻译水平和翻译后水平受到调控.绝大部分植物PAPs的表达水平不仅与植物体内的无机磷含量相关, 而且与植物所处的逆境(氧和盐胁迫等)环境有关.对26个水稻PAPs编码基因进行启动子元件分析, 发现其中12个基因的启动子含有顺式作用元件P1BS (PHR1-binding site), P1BS是水稻磷信号代谢途径转录因子OsPHR2的结合元件(
... 大部分PAP都具有N-端信号肽, 这个信号肽对PAP蛋白的锚定和生物学功能具有重要作用.信号肽定位预测表明, 少数PAPs定位于线粒体(
Comparative genetic analysis of Arabidopsis purple acid phosphatases AtPAP10, AtPAP12, and AtPAP26 provides new insights into their roles in plant adaptation to phosphate deprivation
1
2014
... 植物PAPs与土壤有机磷的分解吸收及植株体内磷素的再利用有密切关系, 很多植物来源的PAPs都具有酸性磷酸酶活性, 能够将植物体内外的有机磷水解成无机磷, 供植物吸收利用.根据PAPs是否具有分泌到细胞外的性质, 可将其分为细胞内酸性磷酸酶(intracellular acid phosphatase, IAP)和分泌型酸性磷酸酶(secreted acid phosphatase, SAP).细胞内酸性磷酸酶主要水解植物体内磷库的磷, 进行细胞内磷素的再利用; 分泌型酸性磷酸酶则被分泌出细胞外, 活化土壤中的有机磷组分, 释放出供植物利用的无机磷(
Secreted acid phosphatase is expressed in cluster roots of lupin in response to phosphorus deficiency
4
2003
... 植物PAPs与土壤有机磷的分解吸收及植株体内磷素的再利用有密切关系, 很多植物来源的PAPs都具有酸性磷酸酶活性, 能够将植物体内外的有机磷水解成无机磷, 供植物吸收利用.根据PAPs是否具有分泌到细胞外的性质, 可将其分为细胞内酸性磷酸酶(intracellular acid phosphatase, IAP)和分泌型酸性磷酸酶(secreted acid phosphatase, SAP).细胞内酸性磷酸酶主要水解植物体内磷库的磷, 进行细胞内磷素的再利用; 分泌型酸性磷酸酶则被分泌出细胞外, 活化土壤中的有机磷组分, 释放出供植物利用的无机磷(
... ).随后, 植物会诱导合成SAP并且分泌到细胞外, 水解植物体外的磷源, 将释放的无机磷吸收到植物体内(
... 缺磷情况下分泌型酸性磷酸酶的诱导表达及其根际分泌与排根形成示意图(
... (A) 磷充足条件下, SAP聚集在根表皮细胞周围, 只有少量SAP分泌; (B) 磷胁迫条件下, SAP在12小时内从根部迅速分泌, 但在长时间保持较低水平; (C) 组织中磷含量明显下降后, 根系成簇排列, 形成排根, SAP分泌量显著增加.
Identification of rice purple acid phosphatases related to phosphate starvation signaling
4
2011
... 1998年, Hiroshi等用同位素标记法, 发现低磷会诱导浮萍(Spirodela oligorrhiza)产生57 kDa的磷脂酰肌醇锚定磷酸酶, 该酶含有Fe3+-Mn2+双核金属中心, 活性受到酒石酸抑制.这是首次在高等植物中发现紫色酸性磷酸酶(
... 目前, PAPs已在多种植物中被鉴定和分离, 通过对拟南芥(Arabidopsis thaliana)和大豆等植物中的PAPs进行系统研究, 发现拟南芥中存在29个PAPs基因(
... 不同PAPs在植物体内的表达模式不尽相同, 它们分工合作, 在转录水平、翻译水平和翻译后水平受到调控.绝大部分植物PAPs的表达水平不仅与植物体内的无机磷含量相关, 而且与植物所处的逆境(氧和盐胁迫等)环境有关.对26个水稻PAPs编码基因进行启动子元件分析, 发现其中12个基因的启动子含有顺式作用元件P1BS (PHR1-binding site), P1BS是水稻磷信号代谢途径转录因子OsPHR2的结合元件(
... 大部分PAP都具有N-端信号肽, 这个信号肽对PAP蛋白的锚定和生物学功能具有重要作用.信号肽定位预测表明, 少数PAPs定位于线粒体(
AtPAP2 modulates the import of the small subunit of Rubisco into chloroplasts
1
2016
... Sun等(2012b)研究表明, AtPAP2是一种尾部锚定蛋白, 含有N-端信号肽和C-端膜锚定信号肽.AtPAP2在拟南芥角果、茎、花、根和开始衰老的叶中高度表达, 但在叶和成熟的种子中表达量相对较低(
An Arabidopsis purple acid phosphatase with phytase activity increases foliar ascorbate
2
2008
... PAPs家族I所包含的种类最多, 该家族可进一步分为I-1和I-2两个亚家族.I-1亚家族成员已被证实参与植物磷和碳代谢途径(
... 大豆酸性磷酸酶3基因(GmPAP3)定位在线粒体(活性氧产生的主要部位), 其表达受盐和氧胁迫诱导, 可能在非生物逆境响应过程中发挥作用.将GmPAP3过表达植株分别进行盐、PEG以及农药胁迫处理, 对比野生型发现过表达植株的根长增加.此外, Gm- PAP3能够通过抑制活性氧(ROS)的积累, 减轻盐和过氧化胁迫造成的损伤(
Heterologous expression of AtPAP2 in transgenic potato influences carbon metabolism and tuber development
1
2014
... Sun等(2012b)研究表明, AtPAP2是一种尾部锚定蛋白, 含有N-端信号肽和C-端膜锚定信号肽.AtPAP2在拟南芥角果、茎、花、根和开始衰老的叶中高度表达, 但在叶和成熟的种子中表达量相对较低(
Over-expression of AtPAP2 in Camelina sativa leads to faster plant growth and higher seed yield
1
2012
... Sun等(2012b)研究表明, AtPAP2是一种尾部锚定蛋白, 含有N-端信号肽和C-端膜锚定信号肽.AtPAP2在拟南芥角果、茎、花、根和开始衰老的叶中高度表达, 但在叶和成熟的种子中表达量相对较低(
OsPHR2 is involved in phosphate- starvation signaling and excessive phosphate accumulation in shoots of plants
1
2008
... 不同PAPs在植物体内的表达模式不尽相同, 它们分工合作, 在转录水平、翻译水平和翻译后水平受到调控.绝大部分植物PAPs的表达水平不仅与植物体内的无机磷含量相关, 而且与植物所处的逆境(氧和盐胁迫等)环境有关.对26个水稻PAPs编码基因进行启动子元件分析, 发现其中12个基因的启动子含有顺式作用元件P1BS (PHR1-binding site), P1BS是水稻磷信号代谢途径转录因子OsPHR2的结合元件(
Expression patterns of purple acid phosphatase genes in Arabidopsis organs and functional analysis of AtPAP23 predominantly transcribed in flower
1
2005
... 紫色酸性磷酸酶在植物各个组织均有分布, 参与各种生命活动(
Differential expression of three purple acid phosphatases from potato
1
2004
... 小分子量的紫色酸性磷酸酶(low molecular weight PAPs, LMW PAPs)以单聚体形式存在, 与哺乳动物和蓝藻体内的小分子量PAPs极为相似, 单体分子量一般约为35 kDa, 在结构上仅保留了典型的C-端结构域(
备案号: 京ICP备16067583号-21
版权所有 © 2021 《植物学报》编辑部
地址:北京香山南辛村20号 邮编:100093
电话:010-62836135 010-62836131 E-mail:cbb@ibcas.ac.cn
本系统由北京玛格泰克科技发展有限公司设计开发

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}