Abstract: The SiSAD gene in Saussurea involucrata and its homologous gene AtFAB2 in Arabidopsis thaliana have been reported to encode homeologous Δ9 stearoyl-acp desaturases. To investigate the function of these genes in plants’ response to cold stress, we constructed two expression vectors PSiSAD:AtFAB2 and PSiSAD:SiSAD, and were Agrobacterium-infiltrated in tobacco. These two kinds of transgenic plants and wild-type tobacco were treated at 20°C, 10°C, 5°C, 0°C, and -2°C for 2 h and then to determine the relative conductivity, malondialdehyde (MDA) and fatty acid content and chlorophyll fluorescence parameters (Fv/Fm). Furthermore, after -2°C treatment for 2 h, and recovery at 25°C for 1 week, we examined recovery rate of these tobacco plants. The recovery rate of SiSAD transgenic tobacco was much better than AtFAB2 transgenic tobacco and wild type. After treatment at 0°C and -2°C for 2 h, the relative conductivity and MDA content of the SiSAD and AtFAB2 transgenic tobacco and wild-type tobacco showed a significant increasing trend. The Fv/Fm of SiSAD and AtFAB2 transgenic tobacco were significantly higher than wild-type tobacco and the Fv/Fm of SiSAD transgenic tobacco was significantly higher than AtFAB2 transgenic tobacco. The content of oleic acid (C18:1) in AtFAB2 transgenic tobacco and wild type were decreased gradually with decreasing temperature and reached the lowest level at 0°C, whereas the content of C18:1 in SiSAD transgenic tobacco increased and peaked at -2°C; The C18:1 contents in SiSAD transgenic tobacco were at least 1.58 and 1.7 folds when compared to AtFAB2 transgenic tobacco and wild type. These results indicate that SiSAD and AtFAB2 genes can significantly enhance the cold tolerance of a non-cold acclimated tobacco. Moreover, the SiSAD gene plays more important role than AtFAB2 gene in cold tolerance.
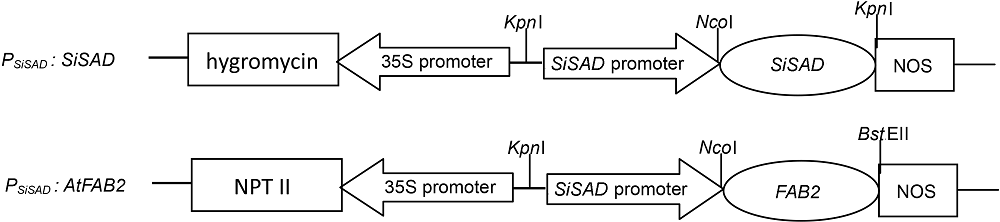
1.2 方法1.2.1 PSiSAD启动子、SiSAD基因和AtFAB2基因的克隆及表达载体的构建 本研究所用的PSiSAD启动子、SiSAD基因、AtFAB2基因的克隆及表达载体的构建均由本实验室完成。PSiSAD启动子为本实验室首次克隆, 为冷诱导启动子(未发表数据)。PSiSAD:SiSAD和PSiSAD:AtFA- B2表达载体结构如图1所示。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-603/img_1.png<b>图1</b> 2种植物表达载体结构<br/><b>Figure 1</b> The structure of two plant expression vectors Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-603/img_1.png<b>图1</b> 2种植物表达载体结构<br/><b>Figure 1</b> The structure of two plant expression vectors
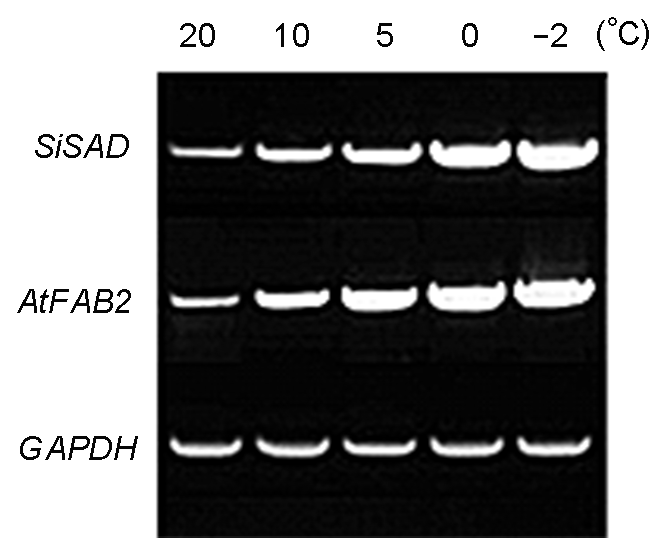
图3 转PSiSAD:SiSAD和PSiSAD:AtFAB2重组质粒烟草的RT- PCR鉴定 Figure 3 RT-PCR identification of tobacco transferred with PSiSAD:SiSAD and PSiSAD:AtFAB2 recombinant plasmid, respectively
2.2 转基因烟草的抗寒性鉴定通过生长恢复实验(图4)可以看出, 在20°C、10°C、5°C和0°C各处理2小时后转基因烟草表型无显著变化; 在-2°C处理2小时后转基因烟草均有不同程度的萎蔫, 其中以野生型最严重, 转SiSAD基因烟草最轻。恢复生长1周后, 野生型烟草未能恢复至处理前的生长状态, 转AtFAB2基因烟草基本恢复到处理前的生长状态, 转SiSAD基因烟草可完全恢复到处理前的生长状态。结果表明AtFAB2基因和SiSAD基因均可提高烟草的抗寒能力, 但SiSAD基因的抗寒效果显著优于AtFAB2基因。 图4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-603/img_4.png<b>图4</b> 不同温度处理下野生型和转基因烟草表型<br/>(A)-(E) 野生型和转基因烟草在20°C、10°C、5°C、0°C及-2°C各处理2小时; (F) -2°C处理后, 25°C恢复培养1周。s-f: <i>P<sub>SiSAD</sub></i>: <i>AtFAB2</i>; s-s: <i>P<sub>SiSAD</sub></i>:<i>SiSAD</i>; WT: 野生型<br/><b>Figure 4</b> Phenotype of wild-type and transgenic tobacco under different temperatures<br/>(A)-(E) Wild-type and transgenic tobacco plants grown at 20°C, 10°C, 5°C, 0°C, and -2°C for 2 hours, respectively; (F) After -2°C treatment recovering in 25°C for one week.s-f: <i>P<sub>SiSAD</sub></i>:<i>AtFAB2</i>; s-s: <i>P<sub>SiSAD</sub></i>:<i>SiSAD</i>; WT: Wild-type Figure 4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-603/img_4.png<b>图4</b> 不同温度处理下野生型和转基因烟草表型<br/>(A)-(E) 野生型和转基因烟草在20°C、10°C、5°C、0°C及-2°C各处理2小时; (F) -2°C处理后, 25°C恢复培养1周。s-f: <i>P<sub>SiSAD</sub></i>: <i>AtFAB2</i>; s-s: <i>P<sub>SiSAD</sub></i>:<i>SiSAD</i>; WT: 野生型<br/><b>Figure 4</b> Phenotype of wild-type and transgenic tobacco under different temperatures<br/>(A)-(E) Wild-type and transgenic tobacco plants grown at 20°C, 10°C, 5°C, 0°C, and -2°C for 2 hours, respectively; (F) After -2°C treatment recovering in 25°C for one week.s-f: <i>P<sub>SiSAD</sub></i>:<i>AtFAB2</i>; s-s: <i>P<sub>SiSAD</sub></i>:<i>SiSAD</i>; WT: Wild-type
图4 不同温度处理下野生型和转基因烟草表型 (A)-(E) 野生型和转基因烟草在20°C、10°C、5°C、0°C及-2°C各处理2小时; (F) -2°C处理后, 25°C恢复培养1周。s-f: PSiSAD: AtFAB2; s-s: PSiSAD:SiSAD; WT: 野生型 Figure 4 Phenotype of wild-type and transgenic tobacco under different temperatures (A)-(E) Wild-type and transgenic tobacco plants grown at 20°C, 10°C, 5°C, 0°C, and -2°C for 2 hours, respectively; (F) After -2°C treatment recovering in 25°C for one week.s-f: PSiSAD:AtFAB2; s-s: PSiSAD:SiSAD; WT: Wild-type
从烟草脂肪酸含量(表2)可以看出, 随着温度的降低, 转SiSAD基因、转AtFAB2基因和野生型烟草的硬脂酸(C18:0)含量呈现显著递减趋势, 并在-2°C时达到最低值, 降幅分别为9.73%、9.69%和9.09%; 转AtFAB2基因和野生型烟草的油酸(C18:1)含量随着温度的降低逐渐升高后降低, 并在0°C时达到最高值, 分别为19.17%和17.90%, 而转SiSAD基因烟草C18:1含量持续升高, 并在-2°C时达到最高值(30.47%), 其含量分别是转AtFAB2基因和野生型烟草的1.58和1.7 倍。以上结果表明, SiSAD基因比AtFAB2基因更能提高烟草的耐低温能力。 表2 Table 2 表2 表2 不同温度下转基因烟草的脂肪酸含量测定 Table 2 The analysis of the content of desaturation products in transgenic tobacco under different temperatures
Temperature (°C)
Plant
Fatty acid (%)
C16:0
C18:0
C18:1
C18:2
C18:3
Total desaturation products
20
WT
43.70±1.10 a
24.77±0.66 a
14.33±0.94 a
3.27±0.43 a
12.97±1.56 a
16.57
s-f
42.27±1.11 a
20.89±1.09 b
15.20±0.38 b
2.77±0.92 a
7.37±1.24 b
17.34
s-s
46.53±0.90 a
20.03±0.32 c
17.23±1.60 c
4.61±1.15 a
7.07±0.93 b
21.91
10
WT
38.99±0.58 a
19.83±0.65 a
15.87±0.92 a
2.53±0.30 a
8.17±1.22 a
17.57
s-f
42.97±0.62 a
15.87±0.86 a
17.43±0.48 a
2.07±0.12 a
8.19±1.12 a
19.69
s-s
40.76±0.79 a
15.70±2.80 a
19.40±1.23 c
1.77±0.38 a
6.00±1.11 a
23.17
5
WT
30.30±0.80 a
16.47±0.66 a
17.50±0.44 a
3.70±0.12 a
17.20±2.02 a
19.4
s-f
25.27±0.66 b
13.83±0.71 b
18.70±1.29 b
1.90±0.17 a
10.20±0.86 b
20.8
s-s
46.43±0.41 c
12.30±0.67 a
20.70±0.57 b
2.67±0.09 a
6.60±0.68 c
31.97
0
WT
40.53±0.44 a
16.23±0.32 a
17.90±0.76 a
2.20±0.17 a
8.47±0.41 a
22.67
s-f
47.63±0.47 b
13.67±0.73 b
19.17±1.45 b
2.33±0.27 a
11.97±1.44 a
28.47
s-s
44.33±0.07 b
12.47±0.45 c
23.50±1.33 a
2.30±0.46 a
11.87±2.07 a
37.67
-2
WT
38.20±1.07 a
15.57±0.76 a
15.63±2.21 a
2.87±0.20 a
10.23±0.72 a
26.73
s-f
40.20±0.23 b
11.20±1.56 b
16.54±0.49 a
3.13±0.59 a
16.30±1.22 b
31.97
s-s
53.60±0.47 c
10.30±0.64 b
30.47±0.89 b
2.33±0.48 a
9.83±2.24 a
49.63
Recover treatment
WT
36.30±1.04 a
14.46±0.68 a
14.77±2.11 a
2.46±0.22 a
10.02±0.68 a
25.94
s-f
41.40±1.03 a
20.05±1.03 b
14.91±0.34 b
2.58±0.85 a
7.19±1.18 b
16.87
s-s
46.33±0.87 a
20.01±0.30 c
17.14±1.53 c
4.57±1.12 a
6.98±0.90 b
21.23
s-f: PSiSAD:AtFAB2; s-s: PSiSAD:SiSAD; WT: Wild-type. Different lowercase letters in the same column indicate significant differences at P<0.05. s-f: PSiSAD:AtFAB2; s-s: PSiSAD:SiSAD; WT: 野生型。不同小写字母表示差异显著(P<0.05)。
表2 不同温度下转基因烟草的脂肪酸含量测定 Table 2 The analysis of the content of desaturation products in transgenic tobacco under different temperatures
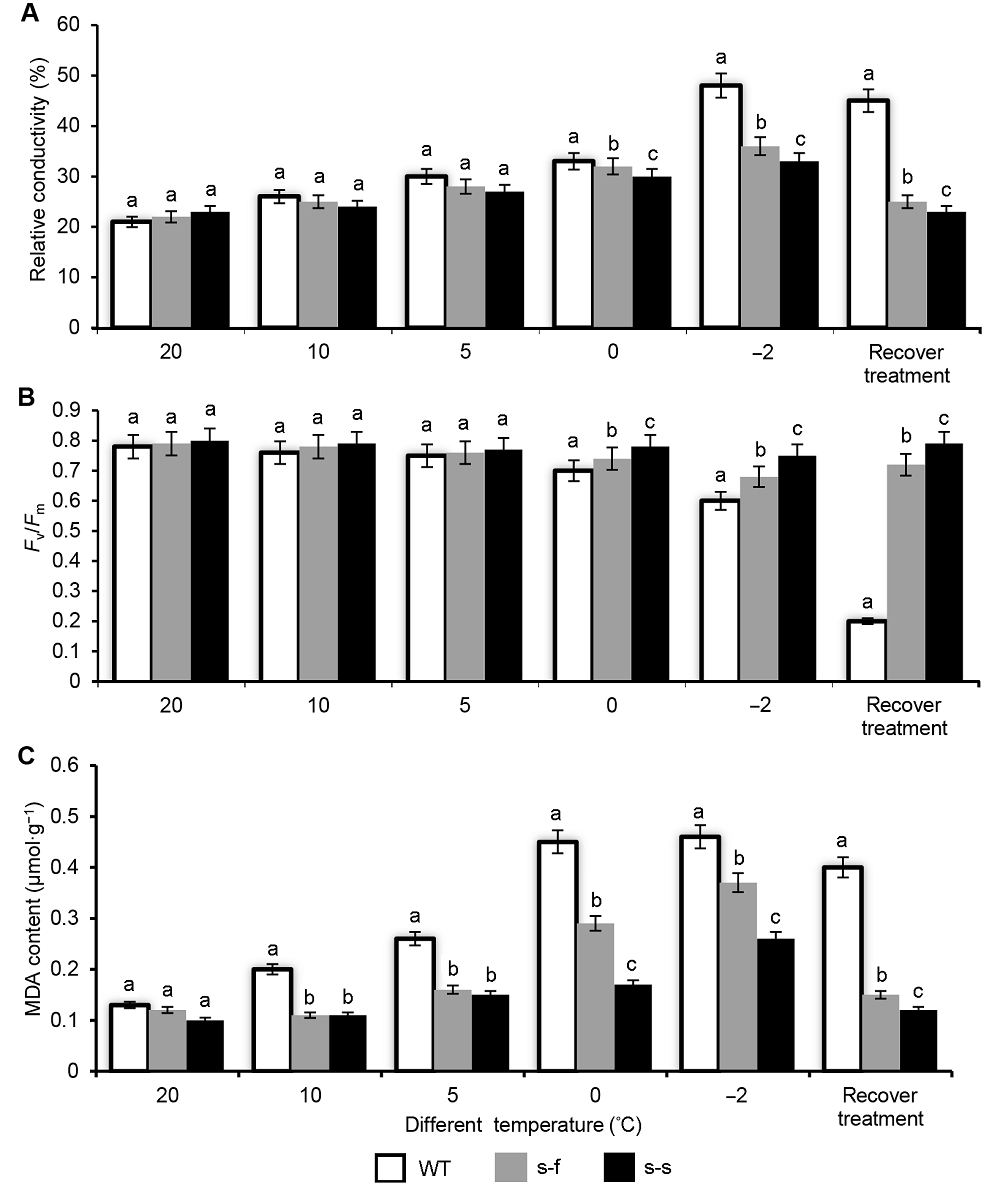
在20°C、10°C和5°C各处理2小时后, 2种转基因型和野生型烟草的相对电导率(图5A)和叶绿素荧光参数Fv/Fm (图5B)无显著差异。而在0°C和-2°C处理2小时后, 2种转基因型烟草的相对电导率显著低于野生型烟草。其中, 转SiSAD基因烟草的相对电导率显著低于转AtFAB2基因的烟草, 表明转SiSAD基因烟草的细胞膜受损程度显著低于转AtFAB2基因烟草和野生型烟草。同时在0°C和-2°C各处理2小时后, 2种转基因烟草的Fv/Fm显著高于野生型烟草, 其中, 转SiSAD基因烟草的Fv/Fm显著高于转AtFAB2基因烟草, 表明SiSAD基因相比AtFAB2基因能够更显著地抑制低温对光系统的伤害, 以维持较高的光能转化效率, 从而增强烟草对低温环境的适应能力。丙二醛含量(图5C)在20°C处理2小时后并无显著差异, 在10°C、5°C、0°C和-2°C处理2小时后, 2种转基因烟草的丙二醛含量显著低于野生型。而在0°C和-2°C时, 转SiSAD基因烟草的丙二醛含量显著低于转AtFAB2基因烟草。结果表明, 在低温胁迫下, SiSAD基因比AtFAB2基因能够更显著地抑制细胞膜脂质氧化的作用。 图5https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-603/img_5.png<b>图5</b> 不同温度处理下野生型和转基因烟草的生理指标<br/>(A) 相对电导率; (B) 叶绿素荧光参数(<i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>); (C) 丙二醛(MDA)含量。处理条件: 野生型和转基因烟草在20°C、10°C、5°C、0°C和-2°C各处理2小时, 恢复处理是在-2°C处理2小时后25°C恢复培养1周。不同小写字母代表差异显著(<i>P<</i>0.05)。s-f: <i>P<sub>SiSAD</sub></i>:<i>AtFAB2</i>; s-s: <i>P<sub>SiSAD</sub></i>:<i>SiSAD</i>; WT: 野生型<br/><b>Figure 5</b> The physiological analysis of wild-type and transgenic tobacco plant after different processing temperature <br/>(A) Relative conductivity; (B) Maximum efficiency of photosystem II photochemistry (<i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>); (C) Malondialdehyde (MDA) content; Wild-type and transgenic tobacco plants grown at 20°C, 10°C, 5°C, 0°C, and -2°C; recovery treatment: after -2°C treatment for 2 hours recovery in 25°C for 1 week. Different lowercase letters indicate significant differences at<i> P</i><0.05. s-f: <i>P<sub>SiSAD</sub></i>:<i>AtFAB2</i>; s-s: <i>P<sub>SiSAD</sub></i>:<i>SiSAD</i>; WT: Wild-type Figure 5https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-603/img_5.png<b>图5</b> 不同温度处理下野生型和转基因烟草的生理指标<br/>(A) 相对电导率; (B) 叶绿素荧光参数(<i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>); (C) 丙二醛(MDA)含量。处理条件: 野生型和转基因烟草在20°C、10°C、5°C、0°C和-2°C各处理2小时, 恢复处理是在-2°C处理2小时后25°C恢复培养1周。不同小写字母代表差异显著(<i>P<</i>0.05)。s-f: <i>P<sub>SiSAD</sub></i>:<i>AtFAB2</i>; s-s: <i>P<sub>SiSAD</sub></i>:<i>SiSAD</i>; WT: 野生型<br/><b>Figure 5</b> The physiological analysis of wild-type and transgenic tobacco plant after different processing temperature <br/>(A) Relative conductivity; (B) Maximum efficiency of photosystem II photochemistry (<i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>); (C) Malondialdehyde (MDA) content; Wild-type and transgenic tobacco plants grown at 20°C, 10°C, 5°C, 0°C, and -2°C; recovery treatment: after -2°C treatment for 2 hours recovery in 25°C for 1 week. Different lowercase letters indicate significant differences at<i> P</i><0.05. s-f: <i>P<sub>SiSAD</sub></i>:<i>AtFAB2</i>; s-s: <i>P<sub>SiSAD</sub></i>:<i>SiSAD</i>; WT: Wild-type
图5 不同温度处理下野生型和转基因烟草的生理指标 (A) 相对电导率; (B) 叶绿素荧光参数(Fv/Fm); (C) 丙二醛(MDA)含量。处理条件: 野生型和转基因烟草在20°C、10°C、5°C、0°C和-2°C各处理2小时, 恢复处理是在-2°C处理2小时后25°C恢复培养1周。不同小写字母代表差异显著(P<0.05)。s-f: PSiSAD:AtFAB2; s-s: PSiSAD:SiSAD; WT: 野生型 Figure 5 The physiological analysis of wild-type and transgenic tobacco plant after different processing temperature (A) Relative conductivity; (B) Maximum efficiency of photosystem II photochemistry (Fv/Fm); (C) Malondialdehyde (MDA) content; Wild-type and transgenic tobacco plants grown at 20°C, 10°C, 5°C, 0°C, and -2°C; recovery treatment: after -2°C treatment for 2 hours recovery in 25°C for 1 week. Different lowercase letters indicate significant differences at P<0.05. s-f: PSiSAD:AtFAB2; s-s: PSiSAD:SiSAD; WT: Wild-type
2.3 讨论研究表明, 细胞膜中脂肪酸饱和程度与膜的相变温度密切相关, 即不饱和程度越高, 相变温度越低, 抗寒性越强。Δ9硬脂酰-ACP脱饱和酶是植物体中脂肪酸合成途径的第1个脱饱和酶, 催化硬脂酰-ACP脱饱和且在脂肪酸链的C9-C10间引入1个双键形成油酰- ACP的反应(Yukawa et al., 1996; Kachroo et al., 2007)。因此, SAD在很大程度上决定着植物膜脂的饱和脂肪酸与不饱和脂肪酸的比例(Thompson et al., 1991), 而这一比例与生物膜的流动性和选择透性等许多生理功能有密切关系。同时, SAD能响应低温变化, 参与防御和生长调控过程, 维持膜的生理状态并参与膜的形成(Murata and Wada, 1995; Tasseva et al., 2004)。因此, SAD基因被国内外****广泛重视(Uem- ura and Steponkus, 1997; Jung et al., 2003)。本研究表明, 天山雪莲SAD基因启动子调控拟南芥At- FAB2基因和天山雪莲SAD基因在烟草中表达, 均可以抑制细胞膜脂氧化, 减轻细胞膜受损程度, 以维持较高的光能转化效率, 从而提高烟草在低温条件下的捕光能力。不饱和脂肪酸含量增加, 增强了烟草对低温环境的适应能力。在生长恢复实验中, 野生型烟草未能恢复到处理前的状态, 而转AtFAB2基因烟草基本恢复到处理前的生长状态, 转天山雪莲SAD基因烟草可完全恢复到处理前的生长状态, 说明天山雪莲SAD基因和拟南芥AtFAB2基因都具有提高植物抗寒性的功能, 但是天山雪莲SAD显著优于拟南芥At- FAB2。将SiSAD蛋白和GenBank中录入的10种植物SAD蛋白比对, 发现亲缘关系最近的为红花和千年桐。罗通(2006)的研究表明, 在4°C处理过的麻疯树(Jatropha carcas)中, 红花SAD表达量明显增高且其相对电导率上升了18.04%; 而天山雪莲在5°C、0°C和-2°C处理下的相对电导率分别上升了5%、8%和11%, 显著低于麻疯树。陈东亮(2010)和程晨等(2011)的研究表明, 将千年桐SAD基因和天山雪莲SAD基因转入酵母中, 其不饱和脂肪酸含量分别提高了9.39%和13.46%, 说明天山雪莲SAD基因比亲缘性较高的植物SAD基因有更好的抗寒性。这可能是由于天山雪莲在极端低温下通过积极的生命活动来适应严寒, 在雪中能旺盛生长并且开花, 其具有特异性的抗寒机制或富集了耐寒基因, 从而产生了适应极端低温环境的生存机制(陈发菊等, 1999; 郭新勇等, 2012)。 综上所述, 新疆雪莲SiSAD基因在低温胁迫下能改变膜脂比例, 促进植物膜脂中的硬脂酸向油酸转变, 提高细胞膜的流动性, 从而调控下游抗寒基因的表达, 且SiSAD的抗寒功能显著优于AtFAB2。这为今后进一步研究和利用该基因以及新疆雪莲资源的开发利用提供了理论基础和实验依据。该基因是否还具有其它功能, 如抗盐、干旱和氧化性等仍需深入研究。
The authors have declared that no competing interests exist.
ArocaR, AmodeoG, Fernández-IllescasS, HermanEM, ChaumontF, ChrispeelsMJ (2005). The role of aqu- aporins and membrane damage in chilling and hydrogen peroxide induced changes in the hydraulic conductance of maize roots.Plant Physiol 137, 341-353. [本文引用: 1]
[19]
BarkanL, VijayanP, CarlssonAS, MekhedovS, BrowseJ (2006). A suppressor of fab1 challenges hypotheses on the role of thylakoid unsaturation in photosynthetic function. Plant Physiol 141, 1012-1020. [本文引用: 1]
[20]
ByfieldGE, XueH, UpchurchRG (2006). Two genes from soybean encoding soluble Δ9 stearoyl-acp desaturas- es.Crop Sci 46, 840-846. [本文引用: 1]
[21]
CraigW, LenziP, ScottiN, De PalmaM, SaggeseP, CarboneV, McGrathCN, MageeAM, MedgyesyP, Kavan- aghTA, DixPJ, GrilloS, CardiT (2008). Transplastomic tobacco plants expressing a fatty acid desaturase gene exhibit altered fatty acid profiles and improved cold tolerance.Transgenic Res 17, 769-782. [本文引用: 1]
James DWJr, DoonerHK (1990). Isolation of EMS-induced mutants in Arabidopsis altered in seed fatty acid composition.Theor Appl Genet 80, 241-245. [本文引用: 1]
[24]
JungS, TatePL, HornR, KochertG, MooreK, AbbottAG (2003). The phylogenetic relationship of possible progenitors of the cultivated peanut. J Hered 94, 334-340. [本文引用: 1]
[25]
KachrooA, ShanklinJ, WhittleE, LapchykL, HildebrandD, KachrooP (2007). The Arabidopsis stearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis.Plant Mol Biol 63, 257-271. [本文引用: 2]
[26]
KrauseGH, WeisE (1991). Chlorophyll fluorescence and photosynthesis: the basics.Ann Rev Plant Physiol Plant Mol Biol 42, 313-349. [本文引用: 1]
[27]
LightnerJ, James DWJr, DoonerHK, BrowseJ (1994a). Altered body morphology is caused by increased stearate levels in a mutant of Arabidopsis.Plant J 6, 401-412. [本文引用: 1]
[28]
LightnerJ, WuJR, BrowseJ (1994b). A mutant of Arabidopsis with increased levels of stearic acid. Plant Physiol 106, 1443-1451.
[29]
LuttsS, KinetJM, BouharmontJ (1996). Nacl-induced senescence in leaves of rice (Oryza sativa L.) cultivars differing in salinity resistance. Ann Bot 78, 389-398. [本文引用: 1]
[30]
MurataN, WadaH (1995). Acyl-lipid desaturases and their importance in the tolerance and acclimatization to cold of cyanobacteria. Biochem J 308, 1-8. [本文引用: 1]
[31]
ShilmanF, BrandY, BrandA, HedvatI, HovavR (2011). Identification and molecular characterization of homeologousΔ9-stearoyl acyl carrier protein desaturase 3 genes from the allotetraploid peanut(Arachis hypogaea).Plant Mol Biol Rep 29, 232-241. [本文引用: 1]
[32]
TassevaG, de VirvilleJD, CantrelC, MoreauF, Zacho- wskiA (2004). Changes in the endoplasmic reticulum lipid properties in response to low temperature in Brassica napus. Plant Physiol Biochem 42, 811-822. [本文引用: 1]
[33]
ThompsonGA, SchererDE, Foxall-Van AkenS, KennyJW, YoungHL, ShintaniDK, KridlJC, KnaufVC (1991). Primary structures of the precursor and mature forms of stearoyl-acyl carrier protein desaturase from safflower embryos and requirement of ferredoxin for enzyme activity.Proc Natl Acad Sci USA 88, 2578-2582. [本文引用: 1]
[34]
UemuraM, SteponkusPL (1997). Effect of cold acclimation on the lipid composition of the inner and outer membrane of the chloroplast envelope isolated from rye leaves.Plant Physiol 114, 1493-1500. [本文引用: 1]
[35]
WhittleE, CahoonEB, SubrahmanyamS, ShanklinJ (2005). A multifunctional acyl-acyl carrier protein desaturase from Hedera helix L.(English ivy) can synthesize 16- and 18-carbon monoene and diene products. J Biol Chem 280, 28169-28176. [本文引用: 1]
[36]
YukawaY, TakaiwaF, ShojiK, MasudaK, YamadaK (1996). Structure and expression of two seed-specific cDNA clones encoding stearoyl-acyl carrier protein desaturase from sesame,Sesamum indicum L. Plant Cell Phy- siol 37, 201-205. [本文引用: 1]
[37]
ZhangP, BurtonJW, UpchurchRG, WhittleE, ShanklinJ, DeweyRE (2008). Mutations in a Δ9-stearoyl-acpdes- aturase gene are associated with enhanced stearic acid levels in soybean seeds.Crop Sci 48, 2305-2313. [本文引用: 1]
植物叶片相对电导率测定方法比较研究 1 2010
... 相对电导率是反映植物膜系统状况的一个重要生理生化指标.植物在受到逆境或其它损伤的情况下细胞膜破裂、膜蛋白受损害, 导致胞质的胞液外渗而使相对电导率增大(陈爱葵, 2010).丙二醛为细胞膜过氧化的产物, 可以与蛋白质和核酸发生反应, 使其分子间结构不稳定, 从而抑制蛋白质的合成, 造成质膜稳定性下降.因此丙二醛含量可以作为衡量植物抗寒性的一个指标(覃鹏等, 2004; 陈思羽等, 2016).诱变处理作为一种逆境因子会对植株的光合性能产生一定的影响, 而通过测定植株叶绿素荧光参数的变化可以很好地鉴定植株对胁迫环境的适应性(Krause and Weis, 1991; 桂仁意等, 2010).当植物受到低温胁迫时, 主要通过增加不饱和脂肪酸的含量和比例来增加膜的流动性, 以维持细胞膜正常的生理功能, 因此不饱和脂肪酸含量/比例也是植物逆境生长中的一项重要生理指标(Aroca et al., 2005; Barkan et al., 2006). ...
根癌农杆菌介导的千年桐SAD基因对产油酵母的遗传转化 1 2010
... 研究表明, 细胞膜中脂肪酸饱和程度与膜的相变温度密切相关, 即不饱和程度越高, 相变温度越低, 抗寒性越强.Δ9硬脂酰-ACP脱饱和酶是植物体中脂肪酸合成途径的第1个脱饱和酶, 催化硬脂酰-ACP脱饱和且在脂肪酸链的C9-C10间引入1个双键形成油酰- ACP的反应(Yukawa et al., 1996; Kachroo et al., 2007).因此, SAD在很大程度上决定着植物膜脂的饱和脂肪酸与不饱和脂肪酸的比例(Thompson et al., 1991), 而这一比例与生物膜的流动性和选择透性等许多生理功能有密切关系.同时, SAD能响应低温变化, 参与防御和生长调控过程, 维持膜的生理状态并参与膜的形成(Murata and Wada, 1995; Tasseva et al., 2004).因此, SAD基因被国内外****广泛重视(Uem- ura and Steponkus, 1997; Jung et al., 2003).本研究表明, 天山雪莲SAD基因启动子调控拟南芥At- FAB2基因和天山雪莲SAD基因在烟草中表达, 均可以抑制细胞膜脂氧化, 减轻细胞膜受损程度, 以维持较高的光能转化效率, 从而提高烟草在低温条件下的捕光能力.不饱和脂肪酸含量增加, 增强了烟草对低温环境的适应能力.在生长恢复实验中, 野生型烟草未能恢复到处理前的状态, 而转AtFAB2基因烟草基本恢复到处理前的生长状态, 转天山雪莲SAD基因烟草可完全恢复到处理前的生长状态, 说明天山雪莲SAD基因和拟南芥AtFAB2基因都具有提高植物抗寒性的功能, 但是天山雪莲SAD显著优于拟南芥At- FAB2.将SiSAD蛋白和GenBank中录入的10种植物SAD蛋白比对, 发现亲缘关系最近的为红花和千年桐.罗通(2006)的研究表明, 在4°C处理过的麻疯树(Jatropha carcas)中, 红花SAD表达量明显增高且其相对电导率上升了18.04%; 而天山雪莲在5°C、0°C和-2°C处理下的相对电导率分别上升了5%、8%和11%, 显著低于麻疯树.陈东亮(2010)和程晨等(2011)的研究表明, 将千年桐SAD基因和天山雪莲SAD基因转入酵母中, 其不饱和脂肪酸含量分别提高了9.39%和13.46%, 说明天山雪莲SAD基因比亲缘性较高的植物SAD基因有更好的抗寒性.这可能是由于天山雪莲在极端低温下通过积极的生命活动来适应严寒, 在雪中能旺盛生长并且开花, 其具有特异性的抗寒机制或富集了耐寒基因, 从而产生了适应极端低温环境的生存机制(陈发菊等, 1999; 郭新勇等, 2012). ...
我国雪莲植物的种类、生境分布及化学成分的研究进展 1 1999
... 研究表明, 细胞膜中脂肪酸饱和程度与膜的相变温度密切相关, 即不饱和程度越高, 相变温度越低, 抗寒性越强.Δ9硬脂酰-ACP脱饱和酶是植物体中脂肪酸合成途径的第1个脱饱和酶, 催化硬脂酰-ACP脱饱和且在脂肪酸链的C9-C10间引入1个双键形成油酰- ACP的反应(Yukawa et al., 1996; Kachroo et al., 2007).因此, SAD在很大程度上决定着植物膜脂的饱和脂肪酸与不饱和脂肪酸的比例(Thompson et al., 1991), 而这一比例与生物膜的流动性和选择透性等许多生理功能有密切关系.同时, SAD能响应低温变化, 参与防御和生长调控过程, 维持膜的生理状态并参与膜的形成(Murata and Wada, 1995; Tasseva et al., 2004).因此, SAD基因被国内外****广泛重视(Uem- ura and Steponkus, 1997; Jung et al., 2003).本研究表明, 天山雪莲SAD基因启动子调控拟南芥At- FAB2基因和天山雪莲SAD基因在烟草中表达, 均可以抑制细胞膜脂氧化, 减轻细胞膜受损程度, 以维持较高的光能转化效率, 从而提高烟草在低温条件下的捕光能力.不饱和脂肪酸含量增加, 增强了烟草对低温环境的适应能力.在生长恢复实验中, 野生型烟草未能恢复到处理前的状态, 而转AtFAB2基因烟草基本恢复到处理前的生长状态, 转天山雪莲SAD基因烟草可完全恢复到处理前的生长状态, 说明天山雪莲SAD基因和拟南芥AtFAB2基因都具有提高植物抗寒性的功能, 但是天山雪莲SAD显著优于拟南芥At- FAB2.将SiSAD蛋白和GenBank中录入的10种植物SAD蛋白比对, 发现亲缘关系最近的为红花和千年桐.罗通(2006)的研究表明, 在4°C处理过的麻疯树(Jatropha carcas)中, 红花SAD表达量明显增高且其相对电导率上升了18.04%; 而天山雪莲在5°C、0°C和-2°C处理下的相对电导率分别上升了5%、8%和11%, 显著低于麻疯树.陈东亮(2010)和程晨等(2011)的研究表明, 将千年桐SAD基因和天山雪莲SAD基因转入酵母中, 其不饱和脂肪酸含量分别提高了9.39%和13.46%, 说明天山雪莲SAD基因比亲缘性较高的植物SAD基因有更好的抗寒性.这可能是由于天山雪莲在极端低温下通过积极的生命活动来适应严寒, 在雪中能旺盛生长并且开花, 其具有特异性的抗寒机制或富集了耐寒基因, 从而产生了适应极端低温环境的生存机制(陈发菊等, 1999; 郭新勇等, 2012). ...
大豆植株不同冠层种子活力及其萌发中抗氧化酶活性 1 2016
... 相对电导率是反映植物膜系统状况的一个重要生理生化指标.植物在受到逆境或其它损伤的情况下细胞膜破裂、膜蛋白受损害, 导致胞质的胞液外渗而使相对电导率增大(陈爱葵, 2010).丙二醛为细胞膜过氧化的产物, 可以与蛋白质和核酸发生反应, 使其分子间结构不稳定, 从而抑制蛋白质的合成, 造成质膜稳定性下降.因此丙二醛含量可以作为衡量植物抗寒性的一个指标(覃鹏等, 2004; 陈思羽等, 2016).诱变处理作为一种逆境因子会对植株的光合性能产生一定的影响, 而通过测定植株叶绿素荧光参数的变化可以很好地鉴定植株对胁迫环境的适应性(Krause and Weis, 1991; 桂仁意等, 2010).当植物受到低温胁迫时, 主要通过增加不饱和脂肪酸的含量和比例来增加膜的流动性, 以维持细胞膜正常的生理功能, 因此不饱和脂肪酸含量/比例也是植物逆境生长中的一项重要生理指标(Aroca et al., 2005; Barkan et al., 2006). ...
转新疆雪莲去饱和酶基因sikSAD重组酵母低温和酒精耐受性分析 1 2011
... 研究表明, 细胞膜中脂肪酸饱和程度与膜的相变温度密切相关, 即不饱和程度越高, 相变温度越低, 抗寒性越强.Δ9硬脂酰-ACP脱饱和酶是植物体中脂肪酸合成途径的第1个脱饱和酶, 催化硬脂酰-ACP脱饱和且在脂肪酸链的C9-C10间引入1个双键形成油酰- ACP的反应(Yukawa et al., 1996; Kachroo et al., 2007).因此, SAD在很大程度上决定着植物膜脂的饱和脂肪酸与不饱和脂肪酸的比例(Thompson et al., 1991), 而这一比例与生物膜的流动性和选择透性等许多生理功能有密切关系.同时, SAD能响应低温变化, 参与防御和生长调控过程, 维持膜的生理状态并参与膜的形成(Murata and Wada, 1995; Tasseva et al., 2004).因此, SAD基因被国内外****广泛重视(Uem- ura and Steponkus, 1997; Jung et al., 2003).本研究表明, 天山雪莲SAD基因启动子调控拟南芥At- FAB2基因和天山雪莲SAD基因在烟草中表达, 均可以抑制细胞膜脂氧化, 减轻细胞膜受损程度, 以维持较高的光能转化效率, 从而提高烟草在低温条件下的捕光能力.不饱和脂肪酸含量增加, 增强了烟草对低温环境的适应能力.在生长恢复实验中, 野生型烟草未能恢复到处理前的状态, 而转AtFAB2基因烟草基本恢复到处理前的生长状态, 转天山雪莲SAD基因烟草可完全恢复到处理前的生长状态, 说明天山雪莲SAD基因和拟南芥AtFAB2基因都具有提高植物抗寒性的功能, 但是天山雪莲SAD显著优于拟南芥At- FAB2.将SiSAD蛋白和GenBank中录入的10种植物SAD蛋白比对, 发现亲缘关系最近的为红花和千年桐.罗通(2006)的研究表明, 在4°C处理过的麻疯树(Jatropha carcas)中, 红花SAD表达量明显增高且其相对电导率上升了18.04%; 而天山雪莲在5°C、0°C和-2°C处理下的相对电导率分别上升了5%、8%和11%, 显著低于麻疯树.陈东亮(2010)和程晨等(2011)的研究表明, 将千年桐SAD基因和天山雪莲SAD基因转入酵母中, 其不饱和脂肪酸含量分别提高了9.39%和13.46%, 说明天山雪莲SAD基因比亲缘性较高的植物SAD基因有更好的抗寒性.这可能是由于天山雪莲在极端低温下通过积极的生命活动来适应严寒, 在雪中能旺盛生长并且开花, 其具有特异性的抗寒机制或富集了耐寒基因, 从而产生了适应极端低温环境的生存机制(陈发菊等, 1999; 郭新勇等, 2012). ...




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}