吕秀立1,2,3†, 张群1†, 陈香波1, 李圃锦1, 吴伟4, 关媛5,6,* 1上海市园林科学规划研究院, 上海 2002322上海城市困难立地绿化工程技术研究中心, 上海 2002323国家林业种质资源平台上海子平台, 上海 2002324上海植物园, 上海 2002325上海市农业科学研究院林木果树研究所, 上海 2014036上海市农业科学研究院, 上海市设施园艺技术重点实验室, 上海 201403 LüXiuli1,2,3†, ZhangQun1†, ChenXiangbo1, LiPujin1, WuWei4, GuanYuan5,6,* 1Shanghai Academy of Landscape Architecture Science and Planning, Shanghai 200232, China2Shanghai Engineering Research Center of Landscaping on Challenging Urban Sites, Shanghai 200232, China3National Forest Genetic Resources Platform-Shanghai Sub-platform, Shanghai 200232, China4Shanghai Botanical Garden, Shanghai 200232, China5Forestry and Fruit Tree Research Institute, Shanghai Academy of Agricultural Sciences, Shanghai 201403, China6Shanghai Key Laboratory of Protected Horticultural Technology, Shanghai Academy of Agricultural Sciences, Shanghai 201403, China 引用本文 吕秀立, 张群, 陈香波, 李圃锦, 吴伟, 关媛. 岩白菜属植物规模化繁殖及遗传稳定性. 植物学报, 2018, 53(5): 643-652
贡献者 * 通讯作者。E-mail: guanyuan@sibs.ac.cn † 共同第一作者。 基金资助 上海市种业发展项目(沪农科种字(2015)第8号); 接受日期:2017-09-10接受日期:2018-03-13网络出版日期:2018-09-10 -->Copyright 2018《植物学报》编辑部 Contributors * Author for correspondence. E-mail: guanyuan@sibs.ac.cn † These authors contributed equally to this paper History Received:Accepted:Online:
Abstract: According to commercial use, market demand and survival of wild resources, Bergenia crassifolia, B. scopulosa, B. purpurascens were selected for tissue culture in vitro and standardized propagation. ISSR markers were used to analyze the genetic stability of tissue culture plantlets. The optimal multiplication medium was MS medium supplemented with 0.01 mg·L-1 NAA, 0.5 mg·L-1 6-BA, and 2.0 mg·L-1 Vc with shoot tips used as explants. The multiplication coefficients were 3.10, 2.50 and 2.10 for the three species, respectively. The suitable rooting culture medium was 1/2MS medium with 1.0 mg·L-1 IBA and 2.0 mg·L-1 Vc, and the rooting rates for the three Bergenia species were 85%, 80%, and 75%, respectively. The rooted plants were transplanted in a mixed medium of humus, yellow sand, and perlite with volume ratio 2:1:1; the survival rates of transplanted plantlets were 90%, 85% and 80%, respectively. Using this method of rapid propagation, three Bergenia species propagated 200 000, 20 000, and 10 000 plantlets, respectively. ISSR molecular marker detection results showed that genetic variation was greater for regenerated plantlets of B. purpurascens than the mother plant and was lower for regenerated plantlets of B. scopulosa than the mother plant. After 20 generations of regeneration, the plantlets of the 3 Bergenia species showed genetic variation. The average genetic variation rate of B. scopulosa and B. purpurascens increased with increasing subculture times, but the average genetic variation rate of B. crassifolia after the increase in number of generations changed irregularly.
Key words:Bergenia crassifolia ; B. scopulosa ; B. purpurascens ; genetic stability ; mass propagation
2 结果与讨论2.1 离体培养2.1.1 无菌苗的获得 本研究表明, 外植体消毒时, 在消毒液中添加0.5- 5.0 mg·L-1 VC有助于获得无菌外植体; 而不添加VC的消毒液, 外植体在初代培养7天内就褐变死亡; 添加浓度超过5.0 mg·L-1 VC的消毒液, 无菌外植体存活率下降甚至死亡。我们最终筛选出2.0 mg·L-1为添加VC的最适浓度, 无菌苗获得率可达75%, 与其它处理相比差异显著(表2)。因此在后续各培养阶段的培养基中添加2.0 mg·L-1 VC。 表2 Table 2 表2 表2 不同浓度VC对无菌苗获得的影响 Table 2 Effects of different concentrations of VC on the ste- rile seedlings
VC (mg·L-1)
Aseptic seedling (individual)
Sterile seedling rate (%)
Browning condition
0.1
0±0 e
0±0 e
Browning
0.2
0±0 e
0±0 e
Browning
0.5
8±2 c
40±10 c
No browning
1.0
13±1 b
65±5 b
No browning
2.0
15±0 a
75±0 a
No browning
4.0
14±1 ab
70±5 ab
No browning
5.0
13±0 b
65±0 b
No browning
6.0
4±0 d
20±0 d
No browning
8.0
0±0 e
0±0 e
Death
The data described as means±SE (n=3). Different lowercase letters behind the data in the same column indicate significant differences at P<0.05. 数据为平均值±标准误(n=3)。同列数字后不同小写字母表示差异显著(P<0.05)。
表2 不同浓度VC对无菌苗获得的影响 Table 2 Effects of different concentrations of VC on the ste- rile seedlings
取样时, 外植体有2种, 一是野生母株顶芽, 二是温室里培养一段时间分株苗的顶芽。外植体修切方式和消毒方法完全相同, 无菌苗获得率却不同(表3)。母株顶芽无菌苗获得率仅为13.33%; 而分株苗顶芽无菌苗获得率高达83.33%, 二者差异显著。其原因是温室中空气较干净, 分株苗感染各种外源微生物、细菌、霉菌和灰尘较少, 植株本身比较洁净。 表3 Table 3 表3 表3 不同外植体对无菌苗获得的影响 Table 3 Effects of different explants on the sterile seedlings
Explant
Inoculation No. (individual)
Septic seedling No. (individual)
Sterile seedling rate (%)
Top bud of mother plant
30
4±1 a
13.33±0.03 a
Shoot apical bud of ramet
30
25±2 b
83.33±0.07 b
The data described as means±SE (n=3). Different lowercase letters behind the data in the same column indicate significant differences at P<0.05. 数据为平均值±标准误(n=3)。同列数字后不同小写字母表示差异显著(P<0.05)。
表3 不同外植体对无菌苗获得的影响 Table 3 Effects of different explants on the sterile seedlings
2.1.2 初代培养 无菌外植体接种到初代培养基上, 1个试管接种1个,以避免交叉污染。初期采用黑暗培养可减少褐变。首次接种7天后将外植体转移至新鲜培养基也可减少褐变。待外植体萌发形成绿色芽点, 再转入光培养; 芽点长至2 cm时, 可转入增殖培养基。 初代培养过程中, 当NAA浓度为0.02 mg·L-1时, 外植体生长发育正常; 当NAA浓度超过0.02 mg·L-1时, 外植体易形成愈伤组织。我们最终筛选0.02 mg·L-1为初代培养NAA最适浓度。当6-BA浓度为0.02 mg·L-1时, 外植体培养30天后, 可以长至2 cm, 但没有出现腋芽分化; 当6-BA浓度超过0.02 mg·L-1时, 外植体在伸长生长的同时, 出现腋芽分化。为了保证下一步增殖实验的准确性, 我们选择0.02 mg·L-1为初代培养6-BA最适浓度。综合外植体愈伤与分化情况, 确定MS+0.02 mg·L-1 6-BA+0.02 mg·L-1 NAA+2.0 mg·L-1 VC为最适初代培养基(表4)。 表4 Table 4 表4 表4 不同激素及不同激素水平对3种岩白菜属植物外植体萌发的影响 Table 4 The effects of different hormone and hormone concentrations on the germination of bud of 3 Bergenia species
Concentration (mg·L-1)
No. of germination (individual)
Germination rate (%)
Other condition
6-BA
NAA
0.01
0.01
5±0 d
25±0 d
No callus, undifferentiated
0.01
0.02
12±1 b
60±5 b
No callus, undifferentiated
0.01
0.04
10±0 c
50±0 c
Callus, undifferentiated
0.01
0.08
10±1 c
50±5 c
Callus, undifferentiated
0.01
0.1
10±0 c
50±0 c
Callus, undifferentiated
0.01
0.2
10±1 c
50±5 c
Callus, undifferentiated
0.01
0.5
10±2 c
50±10 c
Callus, undifferentiated
0.02
0.02
15±1 a
75±5 a
No callus, undifferentiated
0.05
0.02
15±2 a
75±10 a
No callus, undifferentiated
0.1
0.02
15±0 a
75±0 a
No callus, undifferentiated
All medium contain 2.0 mg·L-1 VC. The data described as means±SE (n=3). Different lowercase letters behind the data in the same column indicate significant differences at P<0.05. 培养基中添加2.0 mg·L-1 VC。数据为平均值±标准误(n=3)。同列数字后不同小写字母表示差异显著(P<0.05)。
表4 不同激素及不同激素水平对3种岩白菜属植物外植体萌发的影响 Table 4 The effects of different hormone and hormone concentrations on the germination of bud of 3 Bergenia species
2.1.3 增殖培养 挑选大小一致、颜色嫩绿的试管苗, 同批次转入增殖培养基中, 筛选适宜的增殖培养基。当培养基中不添加抗褐化剂VC时, 厚叶岩白菜褐化严重, 增殖率降低; 添加VC后, 仍有少量褐化, 但已减轻在可控范围内。培养基中不添加抗褐化剂VC时, 秦岭岩白菜和岩白菜都没有褐化; 添加VC后, 对试管苗的增殖没有明显影响。当6-BA浓度为0.75 mg·L-1时, 3个种的试管苗都开始出现玻璃化现象, 导致增殖率降低。考虑生产成本和操作便利, 我们选择MS+0.5 mg·L-1 6-BA+ 0.01 mg·L-1 NAA+2.0 mg·L-1 VC为最适增殖培养基, 3个种增殖系数分别为3.10、2.50和2.10 (图1A-C), 与其它处理相比, 差异显著(表5)。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-643/img_1.png<b>图1</b> 3种岩白菜属植物组培快繁过程<br/>(A)-(C) 厚叶岩白菜(A)、秦岭岩白菜(B)和岩白菜(C)的增殖培养; (D)-(F) 厚叶岩白菜(D)、秦岭岩白菜(E)和岩白菜(F)的生根培养; (G)-(I) 厚叶岩白菜(G)、秦岭岩白菜(H)和岩白菜(I)的植株移栽<br/><b>Figure 1</b> Tissue culture and rapid propagation of 3 <i>Bergenia</i> species<br/>(A)-(C) Multiplication culture of <i>B. crassifolia</i> (A), <i>B. scopulosa</i> (B) and <i>B</i>. <i>purpurascens</i> (C); (D)-(F) Rooting culture of <i>B. crassifolia</i> (D), <i>B. scopulosa</i> (E) and <i>B. purpurascens</i> (F); (G)-(I) Transplanting of <i>B. crassifolia</i> (G), <i>B. scopulosa</i> (H) and <i>B. purpurascens</i> (I) Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-643/img_1.png<b>图1</b> 3种岩白菜属植物组培快繁过程<br/>(A)-(C) 厚叶岩白菜(A)、秦岭岩白菜(B)和岩白菜(C)的增殖培养; (D)-(F) 厚叶岩白菜(D)、秦岭岩白菜(E)和岩白菜(F)的生根培养; (G)-(I) 厚叶岩白菜(G)、秦岭岩白菜(H)和岩白菜(I)的植株移栽<br/><b>Figure 1</b> Tissue culture and rapid propagation of 3 <i>Bergenia</i> species<br/>(A)-(C) Multiplication culture of <i>B. crassifolia</i> (A), <i>B. scopulosa</i> (B) and <i>B</i>. <i>purpurascens</i> (C); (D)-(F) Rooting culture of <i>B. crassifolia</i> (D), <i>B. scopulosa</i> (E) and <i>B. purpurascens</i> (F); (G)-(I) Transplanting of <i>B. crassifolia</i> (G), <i>B. scopulosa</i> (H) and <i>B. purpurascens</i> (I)
图1 3种岩白菜属植物组培快繁过程 (A)-(C) 厚叶岩白菜(A)、秦岭岩白菜(B)和岩白菜(C)的增殖培养; (D)-(F) 厚叶岩白菜(D)、秦岭岩白菜(E)和岩白菜(F)的生根培养; (G)-(I) 厚叶岩白菜(G)、秦岭岩白菜(H)和岩白菜(I)的植株移栽 Figure 1 Tissue culture and rapid propagation of 3 Bergenia species (A)-(C) Multiplication culture of B. crassifolia (A), B. scopulosa (B) and B. purpurascens (C); (D)-(F) Rooting culture of B. crassifolia (D), B. scopulosa (E) and B. purpurascens (F); (G)-(I) Transplanting of B. crassifolia (G), B. scopulosa (H) and B. purpurascens (I)
表5 Table 5 表5 表5 不同激素及不同激素水平对3种岩白菜属植物增殖的影响 Table 5 The effects of different hormone and hormone concentrations on shoot multiplication of 3 Bergenia species
Concentration (mg·L-1)
No. of multiplication bud (individual)
Multiplication coefficien
Browning condition
6-BA
NAA
VC
A
B
C
A
B
C
A
B
C
0.1
0.01
24±1 g
29±2 e
23±0 de
1.20±0.03 g
1.45±0.07 e
1.15±0 de
+
-
-
0.1
0.01
2.0
33±2 f
27±1 e
22±2 e
1.65±0.07 f
1.35±0.03 e
1.10±0 e
-
-
-
0.25
0.01
31±1 f
33±2 d
25±1 d
1.55±0.03 f
1.65±0.07 d
1.25±0.03 d
+
-
-
0.25
0.01
2.0
45±2 d
36±1 c
31±2 c
2.25±0.07 d
1.80±0.03 c
1.55±0.07 c
-
-
-
0.5
0.01
48±1 c
51±2 a
41±1 a
2.40±0.03 c
2.55±0.07 a
2.05±0.03 a
+
-
-
0.5
0.01
2.0
62±1 a
50±1 a
42±2 a
3.10±0.03 a
2.50±0.03 a
2.10±0.07 a
-
-
-
0.75
0.01
52±2 b
49±3 a
43±2 a
2.60±0.07 b
2.45±0.1 a
2.15±0.07 a
+
-
-
0.75
0.01
2.0
62±2 a
50±3 a
42±2 a
3.10±0.07 a
2.50±0.1 a
2.10±0.07 a
-
-
-
1.0
0.01
37±2 e
45±4 b
37±2 b
1.85±0.07 e
2.25±0.13 b
1.85±0.07 b
+
-
-
1.0
0.01
2.0
45±2 d
43±3 b
35±2 b
2.25±0.07 d
2.15±0.1 b
1.75±0.07 b
-
-
-
A: B. crassifolia; B: B. scopulosa; C: B. purpurascens. The data described as means±SE (n=3). Different lowercase letters behind the data in the same column indicate significant differences at P<0.05. A: 厚叶岩白菜; B: 秦岭岩白菜; C: 岩白菜。数据为平均值±标准误(n=3)。同列数字后不同小写字母表示差异显著(P<0.05)。
表5 不同激素及不同激素水平对3种岩白菜属植物增殖的影响 Table 5 The effects of different hormone and hormone concentrations on shoot multiplication of 3 Bergenia species
2.1.4 壮苗培养 岩白菜属植物在增殖培养基上分化出的试管苗大小有差异, 需进行壮苗培养。当6-BA浓度达0.05 mg·L-1 时, 试管苗出现分化; 当IBA浓度达0.2 mg·L-1时, 基部有根分化。我们选择MS+0.04 mg·L-1 6-BA+0.1 mg·L-1 IBA+2.0 mg·L-1 VC为最适壮苗培养基(表6)。培养30天, 试管苗叶片面积增大, 长势整齐一致, 高度增加2.28 cm, 有利于后期的生根培养。 表6 Table 6 表6 表6 不同激素及不同激素水平对3种岩白菜属植物壮苗的影响 Table 6 The effects of different hormone and hormone concentrations on making robust seedling of 3 Bergenia species
Concentration (mg·L-1)
Average height before culture (cm)
Average height after culture (cm)
Adding average height (cm)
Other condition
6-BA
IBA
0.01
0.01
1.1
1.9
0.8±0.1 e
Undifferentiated, no root
0.02
0.01
1.2
2.1
0.98±0 e
Undifferentiated, no root
0.03
0.01
1.1
2.0
0.98±0 e
Undifferentiated, no root
0.04
0.01
1.3
2.5
1.28±0.1 d
Undifferentiated, no root
0.05
0.01
1.2
2.1
0.98±0.1 e
Differentiated, no root
0.04
0.02
1.1
2.6
1.58±0.1 c
Undifferentiated, no root
0.04
0.05
1.2
2.3
1.18±0.1 d
Undifferentiated, no root
0.04
0.1
1.1
3.3
2.28±0.1 b
Undifferentiated, no root
0.04
0.2
1.3
4.1
2.88±0.2 a
Undifferentiated, rootage
All media contain 2.0 mg·L-1 VC. The data described as means±SE (n=3). Different lowercase letters behind the data in the same column indicate significant differences at P<0.05. 培养基中添加2.0 mg·L-1 VC。数据为平均值±标准误(n=3)。同列数字后不同小写字母表示差异显著(P<0.05)。
表6 不同激素及不同激素水平对3种岩白菜属植物壮苗的影响 Table 6 The effects of different hormone and hormone concentrations on making robust seedling of 3 Bergenia species
2.1.5 生根培养 生根培养时, 在培养基中添加NAA, 试管苗基部极易形成愈伤组织, 不仅阻碍生根也不利于后期移栽。当IBA浓度低于1.0 mg·L-1时, 生根率随浓度增加而增加; IBA浓度超过1.0 mg·L-1时, 生根率开始下降, 试管苗基部出现愈伤组织。在生根阶段不添加抗褐化剂VC, 厚叶岩白菜试管苗仍会逐渐褐化死亡, 秦岭岩白菜和岩白菜试管苗没有出现明显褐化, 生根率也不受影响。我们选择1/2MS+1.0 mg·L-1 IBA+2.0 mg·L-1 VC为最适生根培养基。在此培养基上培养15天, 试管苗基部分化出根原基, 培养30天根可长至2-4 cm, 根系微红, 叶色浓绿舒展, 形成完整的试管苗(图1D-F), 3种岩白菜生根率分别达85%、80%和75% (表7)。 表7 Table 7 表7 表7 不同激素及不同激素浓度对3种岩白菜属植物生根的影响 Table 7 The effects of different hormone and hormone concentrations on rooting of 3 Bergenia species
Concentration (mg·L-1)
No. of roots (individual)
Rooting rate (%)
Callus status
NAA
IBA
VC
A
B
C
A
B
C
A
B
C
0.1
2.0
4±1 f
5±0 d
3±1 e
20±0.05 f
25±0 d
15±0.05 e
A small amount of browning
A small amount of browning
A small amount of browning
0.5
2.0
9±2 d
9±2 c
7±1 d
45±0.1 d
45±0.1 c
35±0.05 d
A small amount of browning
A small amount of browning
A small amount of browning
1.0
2.0
11±2 c
13±2 b
10±1 c
55±0.1 c
65±0.1 b
50±0.05 c
A small amount of browning
A small amount of browning
A small amount of browning
2.0
2.0
12±2 b
11±2 c
12±2 b
55±0.1 c
60±0.1 b
60±0.1 b
Severe browning
Severe browning
Severe browning
0.1
2.0
7±1 e
7±0 cd
7±1 d
35±0.05 e
35±0 cd
35±0.05 d
No browning
No browning
No browning
0.5
2.0
17±2 a
18±1 a
16±2 a
85±0.1 a
90±0.05 a
80±0.1 a
No browning
No browning
No browning
1.0
2.0
17±2 a
16±3 a
15±2 a
85±0.1 a
80±0.15 a
75±0.1 a
No browning
No browning
No browning
2.0
2.0
16±2 b
17±3 a
15±3 a
80±0.1 b
85±0.15 a
75±0.15 a
A small amount of browning
A small amount of browning
A small amount of browning
0.5
0±2 g
17±1 a
15±2 a
0±0.1 g
85±0.05 a
75±0.1 a
Browning death
Normal
Normal
A-C see Table 5. The data described as means±SE (n=3). Different lowercase letters behind the data in the same column indicate significant differences at P<0.05. A-C同表5。数据为平均值±标准误(n=3)。同列数字后不同小写字母表示差异显著(P<0.05)。
表7 不同激素及不同激素浓度对3种岩白菜属植物生根的影响 Table 7 The effects of different hormone and hormone concentrations on rooting of 3 Bergenia species
2.2 遗传稳定性分析2.2.1 组培苗外观表现 3种岩白菜属植物组培苗培养至第20代时, 增殖系数稳定, 叶色翠绿, 外观没有明显变化。至第30代, 种苗生长健壮, 颜色深绿, 外观也没有明显变化。至第40代, 增殖系数略有提高, 芽苗成团状, 色泽深绿。至第50代, 种苗生长速度变慢, 植株纤细, 色泽淡绿, 少量种苗顶端聚合在一起, 呈现畸形(图2A, B)。移栽田间后此类顶端聚合的畸形苗生长慢, 节间短, 不易拔高, 宏观统计所占比例为5/10 000, 此类种苗应及早剔除。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-643/img_2.png<b>图2</b> 3种岩白菜属植物畸形苗和地栽苗<br/>(A), (B) 顶端畸形的种苗; (C) 移栽成活的半年生组培苗; (D) 两年生组培苗开花; (E) 两年生组培苗, 冬季叶色变红<br/><b>Figure 2</b> The phenotype of abnormal seedlings and field grown plants of 3 <i>Bergenia</i> species<br/>(A), (B) The seedlings of abnormal apex; (C) Tissue culture seedlings were transplanted for half a year; (D) Flowering phenotype of two years old tissue culture plants; (E) Phenotypic changes of leaves of two years old tissue culture plants in winter Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-643/img_2.png<b>图2</b> 3种岩白菜属植物畸形苗和地栽苗<br/>(A), (B) 顶端畸形的种苗; (C) 移栽成活的半年生组培苗; (D) 两年生组培苗开花; (E) 两年生组培苗, 冬季叶色变红<br/><b>Figure 2</b> The phenotype of abnormal seedlings and field grown plants of 3 <i>Bergenia</i> species<br/>(A), (B) The seedlings of abnormal apex; (C) Tissue culture seedlings were transplanted for half a year; (D) Flowering phenotype of two years old tissue culture plants; (E) Phenotypic changes of leaves of two years old tissue culture plants in winter
图2 3种岩白菜属植物畸形苗和地栽苗 (A), (B) 顶端畸形的种苗; (C) 移栽成活的半年生组培苗; (D) 两年生组培苗开花; (E) 两年生组培苗, 冬季叶色变红 Figure 2 The phenotype of abnormal seedlings and field grown plants of 3 Bergenia species (A), (B) The seedlings of abnormal apex; (C) Tissue culture seedlings were transplanted for half a year; (D) Flowering phenotype of two years old tissue culture plants; (E) Phenotypic changes of leaves of two years old tissue culture plants in winter
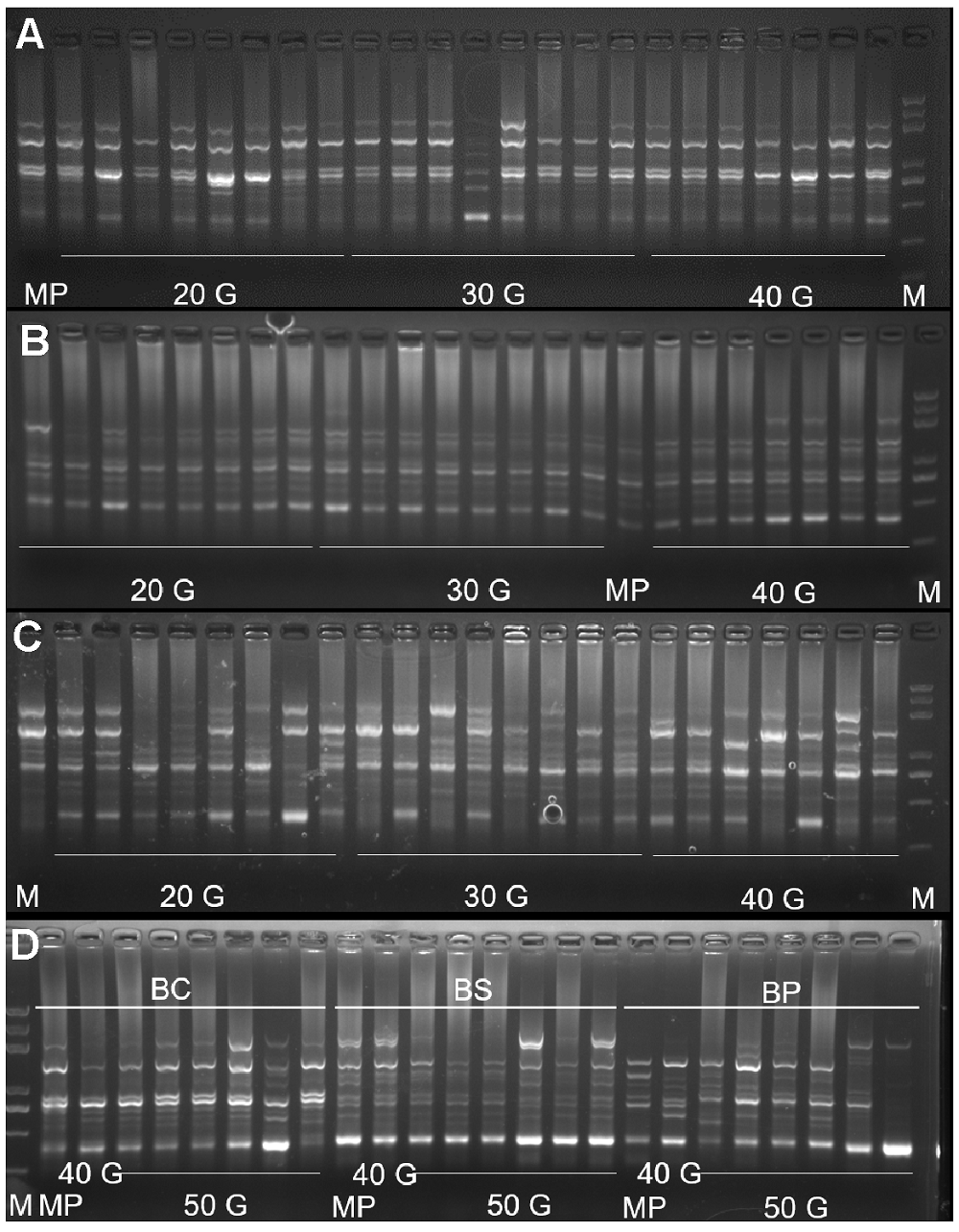
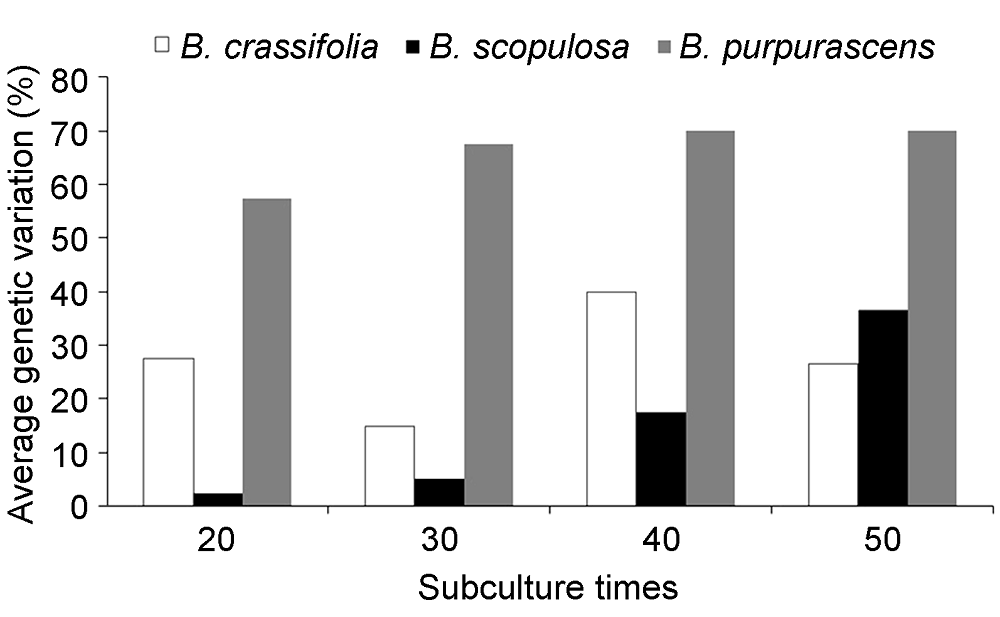
组培苗移栽半年后植株冠幅可达10 cm, 高度可达8 cm (图2C)。于福建省宁德市海拔450 m山地种植2年后, 平均产量达2.48×104 kg·hm-2, 田间长势良好, 75%以上植株早春开花(图2D)。当冬季气温降到0°C以下时, 叶色变红, 变色率达90% (图2E); 在密林、树下栽培, 表现出极耐阴等野生种的特性。 2.2.2 遗传稳定性检测 利用5条扩增效果好的引物, 对3种岩白菜母株和不同继代次数组培苗的基因组DNA进行ISSR分析, 5条引物都扩增出了清晰可辨的谱带。3种岩白菜属植物从第20代开始, 都检测到遗传变异(图3A-D)。厚叶岩白菜随继代次数增加, 遗传变异呈现不规律变化, 平均遗传变异率为15%-40%。而秦岭岩白菜和岩白菜遗传变异率与继代次数呈正相关, 变化规律明显。其中秦岭岩白菜继代至第20代, 组培苗平均遗传变异率为2.5%, 第50代达36.67%; 岩白菜继代至第20代, 组培苗平均遗传变异率为57.5%, 至第50代高达70% (图4)。 图3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-643/img_3.png<b>图3</b> ISSR引物UBC854在3种岩白菜属植物中的扩增结果<br/>(A) 厚叶岩白菜扩增结果; (B) 秦岭岩白菜扩增结果; (C) 岩白菜扩增结果; (D) 3种岩白菜第50代组培苗扩增结果。M: DNA标准样品; MP: 母本; 20 G: 第20代; 30 G: 第30代; 40 G: 第40代; 50 G: 第50代; BC: 厚叶岩白菜; BS: 秦岭岩白菜; BP: 岩白菜<br/><b>Figure 3</b> The amplification results of UBC854 primers in 3 <i>Bergenia</i> species<br/>(A) The amplification results of UBC854 primers in <i>B. crassi- folia</i>; (B) The amplification results of UBC854 primers in <i>B. scopulosa</i>; (C) The amplification results of UBC854 primers in <i>B. purpurascens</i>; (D) The amplification results of UBC854 primers in 50 generation seedlings of 3 <i>Bergenia</i> species. M: DNA marker; MP: Maternal plant; 20 G: 20 generations; 30 G: 30 generations; 40 G: 40 generations; 50 G: 50 generations; BC: <i>B. crassifolia</i>; BS: <i>B. scopulosa</i>; BP: <i>B</i>. <i>purpurascens</i> Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-643/img_3.png<b>图3</b> ISSR引物UBC854在3种岩白菜属植物中的扩增结果<br/>(A) 厚叶岩白菜扩增结果; (B) 秦岭岩白菜扩增结果; (C) 岩白菜扩增结果; (D) 3种岩白菜第50代组培苗扩增结果。M: DNA标准样品; MP: 母本; 20 G: 第20代; 30 G: 第30代; 40 G: 第40代; 50 G: 第50代; BC: 厚叶岩白菜; BS: 秦岭岩白菜; BP: 岩白菜<br/><b>Figure 3</b> The amplification results of UBC854 primers in 3 <i>Bergenia</i> species<br/>(A) The amplification results of UBC854 primers in <i>B. crassi- folia</i>; (B) The amplification results of UBC854 primers in <i>B. scopulosa</i>; (C) The amplification results of UBC854 primers in <i>B. purpurascens</i>; (D) The amplification results of UBC854 primers in 50 generation seedlings of 3 <i>Bergenia</i> species. M: DNA marker; MP: Maternal plant; 20 G: 20 generations; 30 G: 30 generations; 40 G: 40 generations; 50 G: 50 generations; BC: <i>B. crassifolia</i>; BS: <i>B. scopulosa</i>; BP: <i>B</i>. <i>purpurascens</i>
图3 ISSR引物UBC854在3种岩白菜属植物中的扩增结果 (A) 厚叶岩白菜扩增结果; (B) 秦岭岩白菜扩增结果; (C) 岩白菜扩增结果; (D) 3种岩白菜第50代组培苗扩增结果。M: DNA标准样品; MP: 母本; 20 G: 第20代; 30 G: 第30代; 40 G: 第40代; 50 G: 第50代; BC: 厚叶岩白菜; BS: 秦岭岩白菜; BP: 岩白菜 Figure 3 The amplification results of UBC854 primers in 3 Bergenia species (A) The amplification results of UBC854 primers in B. crassi- folia; (B) The amplification results of UBC854 primers in B. scopulosa; (C) The amplification results of UBC854 primers in B. purpurascens; (D) The amplification results of UBC854 primers in 50 generation seedlings of 3 Bergenia species. M: DNA marker; MP: Maternal plant; 20 G: 20 generations; 30 G: 30 generations; 40 G: 40 generations; 50 G: 50 generations; BC: B. crassifolia; BS: B. scopulosa; BP: B. purpurascens
图4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-643/img_4.png<b>图4</b> 3种岩白菜属植物平均遗传变异率比较<br/><b>Figure 4</b> The comparison of average genetic variation in 3 <i>Bergenia </i>species Figure 4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-5-643/img_4.png<b>图4</b> 3种岩白菜属植物平均遗传变异率比较<br/><b>Figure 4</b> The comparison of average genetic variation in 3 <i>Bergenia </i>species
AllenGC, Flores-VergaraMA, KrasynanskiS, KumarS, ThompsonWF (2006). A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide.Nat Protoc 1, 2320-2325. [本文引用: 2]
[26]
ReischBI (1988). Genetic instability in plant cell cultures: utilization in plant breeding and genetic studies. In: Pais MSS, Mavituna F, Novais JM, eds. Plant Cell Biotechnology. Berlin Heidelberg: Springer. pp. 87-95. [本文引用: 1]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}