Abstract: Cadmium (Cd) is a highly toxic heavy metal that threatens human health. Rice is one of food crops that can accumulate Cd in the grain to levels that are unsafe for human consumption. With increasing contamination of heavy metals in paddy soils in China, considerable proportions of rice grain produced in some areas of southern China exceed the 0.2 mg·kg-1 Cd limit of the Chinese food standard, which causes widespread public concern. Molecular breeding of rice varieties that accumulate Cd in straw for removing Cd from paddy soil while producing safe grain is one of the strategies for phytoremediation of contaminated soils. Recently, Luo et al. identified a quantitative trait locus CAL1 in rice that specifically regulates the accumulation of Cd in leaves. CAL1 encodes a defensin-like protein that can chelate Cd in the cytosol and facilitates Cd secretion from xylem parenchyma cells into xylem vessels for long-distance transport. The chelation of Cd to CAL1 appears to prevent Cd from being loaded into the phloem for transport to rice grain. Thus, CAL1 does not affect the accumulation of Cd in rice grain. These findings shed light on understanding the molecular mechanism of Cd translocation and allocation in rice and provide a molecular tool to breed rice varieties that may be used to remove Cd from the soil without affecting grain Cd concentration.
Key words:rice ; cadmium ; phytoremediation ; defensin protein
重金属镉是我国农田和农产品的重要污染物之一。随着我国工业化和城市化的快速发展, 土壤重金属污染问题日益突出。环保部和国土资源部2014年发布的全国土壤污染调查报告显示, 我国耕地土壤19.4%点位污染物含量超标, 其中重金属镉的超标率为7%, 污染程度以中、低污染为主(环境保护部和国土资源部, 2014)。农田重金属污染造成农产品重金属含量超标时常发生, 我国南方稻米重金属含量超标现象尤为严重(Zhao et al., 2015)。调查结果显示, 我国湖南益阳和长株潭地区分别有60%和76%的稻米镉含量超过我国食品卫生标准稻米镉限量值——0.2 mg∙kg-1, 样品中最高镉含量分别达2.77和4.80 mg∙kg-1, 超过限量值13和24倍(Du et al., 2013; Zhu et al., 2016)。土壤重金属污染已经导致我国居民的镉摄入量逐年增加。据国家卫生和计划生育委员会长达25年的调查统计结果显示, 我国居民人均镉的摄入量已从1990年的13.8 μg∙d-1上升到2015年的30.6 μg∙d-1, 25年间升高了122%, 达到世界卫生组织和世界粮农组织规定的日均镉允许摄入量(FAO/WHO Cd Tole- rable Daily Intake)的61%。其中, 稻米是膳食中镉的主要来源, 占我国人均每天镉摄入量的55%, 南方人群则占到了65% (Song et al., 2017)。因此, 农田重金属污染造成的农产品重金属含量超标已经严重影响农产品的质量安全, 对我国居民的身体健康构成威胁, 已成为限制我国农产品质量安全和农业生产可持续发展的重要因素。因此, 重金属污染治理迫在眉睫。
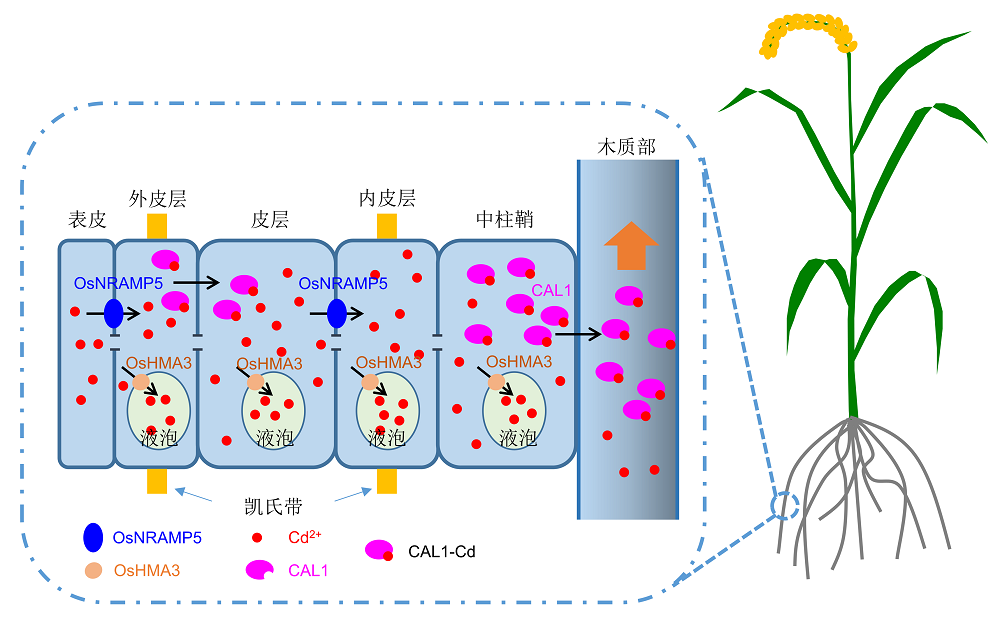
图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-451/img_1.png<b>图1</b> 水稻根部吸收和转运镉的示意图<br/>镉离子在水稻根部通过OsNRAMP5吸收进入外皮层, 随后部分镉被OsHMA3转运到液泡中进行区隔化。OsNRAMP5同时负责将镉离子从皮层细胞转运到内皮层细胞。位于木质部薄壁细胞的CAL1蛋白与镉进行螯合后被分泌到胞外, 进入木质部导管向地上部进行长距离运输。CAL1蛋白也在外皮层细胞中表达, 可能也与镉进行螯合之后分泌到皮层细胞中。然而, 介导CAL1-Cd螯合物跨膜转运是通过转运蛋白还是囊泡运输尚不清楚。<br/><b>Figure 1</b> Schematic diagram of the uptake and transport of Cd in rice roots<br/>The uptake of Cd into rice roots is mediated by OsNRAMP5, which is also responsible for the transport of Cd from cortex cells into endodermis. Part of Cd is then sequestered into vacuoles by OsHMA3. In xylem parenchyma cells, Cd is chelated with CAL1 in the cytosol and then is secreted into the xylem vessels for long-distance transport to shoots. CAL1 is also expressed in exodermis, where CAL1 is also able to chelate Cd and potentially facilitates the Cd secretion from exodermis into cortex cells. However, it is still not clear whether transporters or vesicular trafficking pathways are responsible for translocating the CAL1-Cd complex across the plasma membrane. Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-4-451/img_1.png<b>图1</b> 水稻根部吸收和转运镉的示意图<br/>镉离子在水稻根部通过OsNRAMP5吸收进入外皮层, 随后部分镉被OsHMA3转运到液泡中进行区隔化。OsNRAMP5同时负责将镉离子从皮层细胞转运到内皮层细胞。位于木质部薄壁细胞的CAL1蛋白与镉进行螯合后被分泌到胞外, 进入木质部导管向地上部进行长距离运输。CAL1蛋白也在外皮层细胞中表达, 可能也与镉进行螯合之后分泌到皮层细胞中。然而, 介导CAL1-Cd螯合物跨膜转运是通过转运蛋白还是囊泡运输尚不清楚。<br/><b>Figure 1</b> Schematic diagram of the uptake and transport of Cd in rice roots<br/>The uptake of Cd into rice roots is mediated by OsNRAMP5, which is also responsible for the transport of Cd from cortex cells into endodermis. Part of Cd is then sequestered into vacuoles by OsHMA3. In xylem parenchyma cells, Cd is chelated with CAL1 in the cytosol and then is secreted into the xylem vessels for long-distance transport to shoots. CAL1 is also expressed in exodermis, where CAL1 is also able to chelate Cd and potentially facilitates the Cd secretion from exodermis into cortex cells. However, it is still not clear whether transporters or vesicular trafficking pathways are responsible for translocating the CAL1-Cd complex across the plasma membrane.
图1 水稻根部吸收和转运镉的示意图 镉离子在水稻根部通过OsNRAMP5吸收进入外皮层, 随后部分镉被OsHMA3转运到液泡中进行区隔化。OsNRAMP5同时负责将镉离子从皮层细胞转运到内皮层细胞。位于木质部薄壁细胞的CAL1蛋白与镉进行螯合后被分泌到胞外, 进入木质部导管向地上部进行长距离运输。CAL1蛋白也在外皮层细胞中表达, 可能也与镉进行螯合之后分泌到皮层细胞中。然而, 介导CAL1-Cd螯合物跨膜转运是通过转运蛋白还是囊泡运输尚不清楚。 Figure 1 Schematic diagram of the uptake and transport of Cd in rice roots The uptake of Cd into rice roots is mediated by OsNRAMP5, which is also responsible for the transport of Cd from cortex cells into endodermis. Part of Cd is then sequestered into vacuoles by OsHMA3. In xylem parenchyma cells, Cd is chelated with CAL1 in the cytosol and then is secreted into the xylem vessels for long-distance transport to shoots. CAL1 is also expressed in exodermis, where CAL1 is also able to chelate Cd and potentially facilitates the Cd secretion from exodermis into cortex cells. However, it is still not clear whether transporters or vesicular trafficking pathways are responsible for translocating the CAL1-Cd complex across the plasma membrane.
DasN, BhattacharyaS, BhattacharyyaS, MaitiMK (2017). Identification of alternatively spliced transcripts of rice phytochelatin synthase 2 gene OsPCS2 involved in miti- gation of cadmium and arsenic stresses. Plant Mol Biol 94, 167-183. [本文引用: 1]
[3]
DuY, HuXF, WuXH, ShuY, JiangY, YanXJ (2013). Affects of mining activities on Cd pollution to the paddy soils and rice grain in Hunan province, Central South China.Environ Monit Assess 185, 9843-9856. [本文引用: 1]
[4]
IshikawaS, IshimaruY, IguraM, KuramataM, AbeT, SenouraT, HaseY, AraoT, NishizawaNK, NakanishiH (2012). Ion-beam irradiation, gene identification, and marker- assisted breeding in the development of low-cadmium rice.Proc Natl Acad Sci USA 109, 19166-19171. [本文引用: 1]
[5]
IshimaruY, TakahashiR, BashirK, ShimoH, SenouraT, SugimotoK, OnoK, YanoM, IshikawaS, AraoT, NakanishiH, NishizawaNK (2012). Characterizing the role of rice NRAMP5 in manganese, iron and cadmium transport.Sci Rep 2, 286. [本文引用: 2]
[6]
LuoJS, HuangJ, ZengDL, PengJS, ZhangGB, MaHL, GuanY, YiHY, FuYL, HanB, LinHX, QianQ, GongJM (2018). A defensin-like protein drives cadmium efflux and allocation in rice.Nat Commun 9, 645. [本文引用: 2]
[7]
MiyadateH, AdachiS, HiraizumiA, TezukaK, NakazawaN, KawamotoT, KatouK, KodamaI, SakuraiK, Tak- ahashiH, Satoh-NagasawaN, WatanabeA, FujimuraT, AkagiH (2011). OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles.New Phytol 189, 190-199. [本文引用: 3]
[8]
SasakiA, YamajiN, YokoshoK, MaJF (2012). Nramp5 is a major transporter responsible for manganese and cad- mium uptake in rice.Plant Cell 24, 2155-2167. [本文引用: 1]
[9]
SongY, WangY, MaoWF, SuiHX, YongL, YangDJ, JiangDG, ZhangL, GongYY (2017). Dietary cadmium expo- sure assessment among the Chinese population.PLoS One 12, e0177978. [本文引用: 1]
[10]
TangL, MaoBG, LiYK, LvQM, ZhangLP, ChenCY, HeHJ, WangWP, ZengXF, ShaoY, PanYL, HuYY, PengY, FuXQ, LiHQ, XiaST, ZhaoBR (2017). Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield.Sci Rep 7, 14438. [本文引用: 3]
YanJL, WangPT, WangP, YangM, LianXM, TangZ, HuangCF, SaltDE, ZhaoFJ (2016). A loss-of-function allele of OsHMA3 associated with high cadmium accu- mulation in shoots and grain of Japonica rice cultivars. Plant Cell Environ 39, 1941-1954. [本文引用: 3]
[13]
YangM, ZhangYY, ZhangLJ, HuJT, ZhangX, LuK, DongHX, WangDJ, ZhaoFJ, HuangCF, LianXM (2014). OsNRAMP5 contributes to manganese translocation and distribution in rice shoots.J Exp Bot 65, 4849-4861. [本文引用: 1]
[14]
ZhaoFJ, MaYB, ZhuYG, TangZ, McGrathSP (2015). Soil contamination in China: current status and mitigation stra- tegies.Environ Sci Technol 49, 750-759. [本文引用: 1]
[15]
ZhuHH, ChenC, XuC, ZhuQH, HuangDY (2016). Effects of soil acidification and liming on the phytoavailability of cadmium in paddy soils of central subtropical China.En- viron Pollut 219, 99-106.
... 重金属镉是我国农田和农产品的重要污染物之一.随着我国工业化和城市化的快速发展, 土壤重金属污染问题日益突出.环保部和国土资源部2014年发布的全国土壤污染调查报告显示, 我国耕地土壤19.4%点位污染物含量超标, 其中重金属镉的超标率为7%, 污染程度以中、低污染为主(环境保护部和国土资源部, 2014).农田重金属污染造成农产品重金属含量超标时常发生, 我国南方稻米重金属含量超标现象尤为严重(Zhao et al., 2015).调查结果显示, 我国湖南益阳和长株潭地区分别有60%和76%的稻米镉含量超过我国食品卫生标准稻米镉限量值——0.2 mg∙kg-1, 样品中最高镉含量分别达2.77和4.80 mg∙kg-1, 超过限量值13和24倍(Du et al., 2013; Zhu et al., 2016).土壤重金属污染已经导致我国居民的镉摄入量逐年增加.据国家卫生和计划生育委员会长达25年的调查统计结果显示, 我国居民人均镉的摄入量已从1990年的13.8 μg∙d-1上升到2015年的30.6 μg∙d-1, 25年间升高了122%, 达到世界卫生组织和世界粮农组织规定的日均镉允许摄入量(FAO/WHO Cd Tole- rable Daily Intake)的61%.其中, 稻米是膳食中镉的主要来源, 占我国人均每天镉摄入量的55%, 南方人群则占到了65% (Song et al., 2017).因此, 农田重金属污染造成的农产品重金属含量超标已经严重影响农产品的质量安全, 对我国居民的身体健康构成威胁, 已成为限制我国农产品质量安全和农业生产可持续发展的重要因素.因此, 重金属污染治理迫在眉睫. ...
1 2013
... 镉是植物非必需的重金属元素之一, 主要通过锰、铁和锌等二价阳离子的吸收途径经根系进入植物体内.在水稻中, 镉主要通过定位于细胞质膜上的OsNRAMP5进行吸收(图1).osnramp5突变体丧失大部分镉的吸收能力, 其秸秆和籽粒中镉的含量都比野生型显著降低(Ishikawa et al., 2012; Sasaki et al., 2012; Yang et al., 2014).通过CRISPR/Cas9基因编辑技术将两系杂交稻双亲中的OsNRAMP5基因同时敲除, 获得的杂交种籽粒镉含量比双亲降低了98%, 在双亲籽粒镉超标10倍的情况下, 杂交种籽粒镉含量仍然低于国家标准, 显示OsNRAMP5基因在培育镉低吸收水稻品种中具有良好的应用前景(Tang et al., 2017).然而, OsNRAMP5同时也是植物必需矿质营养元素锰的转运蛋白(Ishimaru et al., 2012; Sasaki et al., 2012), 参与植物体内锰从根部向地上部的转运以及再分配(Yang et al., 2014).在水培条件下, osnramp5突变体比野生型对缺锰更为敏感(Sasaki et al., 2012; Yang et al., 2014), 这可能限制了osnramp5突变体在控制镉吸收中的实际应用, 尤其是在低锰农田中的应用.镉离子通过根部吸收进入植物体以后, 通常被区隔化到液泡中(图1), 在水稻中, 这一过程由P-1B型重金属ATP酶OsHMA3负责(Ueno et al., 2010).在OsHMA3的T-DNA插入突变体或者OsHMA3功能丧失的水稻品种中, 镉未能被转运到液泡中, 从而导致由根系向地上部和籽粒运输大幅度上升, 引起籽粒中镉含量升高(Ueno et al., 2010; Miyadate et al., 2011; Yan et al., 2016).相反, 过量表达OsHMA3基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ...
1 2012
... 镉是植物非必需的重金属元素之一, 主要通过锰、铁和锌等二价阳离子的吸收途径经根系进入植物体内.在水稻中, 镉主要通过定位于细胞质膜上的OsNRAMP5进行吸收(图1).osnramp5突变体丧失大部分镉的吸收能力, 其秸秆和籽粒中镉的含量都比野生型显著降低(Ishikawa et al., 2012; Sasaki et al., 2012; Yang et al., 2014).通过CRISPR/Cas9基因编辑技术将两系杂交稻双亲中的OsNRAMP5基因同时敲除, 获得的杂交种籽粒镉含量比双亲降低了98%, 在双亲籽粒镉超标10倍的情况下, 杂交种籽粒镉含量仍然低于国家标准, 显示OsNRAMP5基因在培育镉低吸收水稻品种中具有良好的应用前景(Tang et al., 2017).然而, OsNRAMP5同时也是植物必需矿质营养元素锰的转运蛋白(Ishimaru et al., 2012; Sasaki et al., 2012), 参与植物体内锰从根部向地上部的转运以及再分配(Yang et al., 2014).在水培条件下, osnramp5突变体比野生型对缺锰更为敏感(Sasaki et al., 2012; Yang et al., 2014), 这可能限制了osnramp5突变体在控制镉吸收中的实际应用, 尤其是在低锰农田中的应用.镉离子通过根部吸收进入植物体以后, 通常被区隔化到液泡中(图1), 在水稻中, 这一过程由P-1B型重金属ATP酶OsHMA3负责(Ueno et al., 2010).在OsHMA3的T-DNA插入突变体或者OsHMA3功能丧失的水稻品种中, 镉未能被转运到液泡中, 从而导致由根系向地上部和籽粒运输大幅度上升, 引起籽粒中镉含量升高(Ueno et al., 2010; Miyadate et al., 2011; Yan et al., 2016).相反, 过量表达OsHMA3基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ...
... 镉是植物非必需的重金属元素之一, 主要通过锰、铁和锌等二价阳离子的吸收途径经根系进入植物体内.在水稻中, 镉主要通过定位于细胞质膜上的OsNRAMP5进行吸收(图1).osnramp5突变体丧失大部分镉的吸收能力, 其秸秆和籽粒中镉的含量都比野生型显著降低(Ishikawa et al., 2012; Sasaki et al., 2012; Yang et al., 2014).通过CRISPR/Cas9基因编辑技术将两系杂交稻双亲中的OsNRAMP5基因同时敲除, 获得的杂交种籽粒镉含量比双亲降低了98%, 在双亲籽粒镉超标10倍的情况下, 杂交种籽粒镉含量仍然低于国家标准, 显示OsNRAMP5基因在培育镉低吸收水稻品种中具有良好的应用前景(Tang et al., 2017).然而, OsNRAMP5同时也是植物必需矿质营养元素锰的转运蛋白(Ishimaru et al., 2012; Sasaki et al., 2012), 参与植物体内锰从根部向地上部的转运以及再分配(Yang et al., 2014).在水培条件下, osnramp5突变体比野生型对缺锰更为敏感(Sasaki et al., 2012; Yang et al., 2014), 这可能限制了osnramp5突变体在控制镉吸收中的实际应用, 尤其是在低锰农田中的应用.镉离子通过根部吸收进入植物体以后, 通常被区隔化到液泡中(图1), 在水稻中, 这一过程由P-1B型重金属ATP酶OsHMA3负责(Ueno et al., 2010).在OsHMA3的T-DNA插入突变体或者OsHMA3功能丧失的水稻品种中, 镉未能被转运到液泡中, 从而导致由根系向地上部和籽粒运输大幅度上升, 引起籽粒中镉含量升高(Ueno et al., 2010; Miyadate et al., 2011; Yan et al., 2016).相反, 过量表达OsHMA3基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ... ... 镉离子通过植物根系吸收进入体内以后通常与植物螯合素(phytochelatins, PCs)、谷胱甘肽(GSH)、金属硫蛋白(metallothioneins, MT)以及其它一些寡肽或者小分子化合物等螯合物进行螯合.这些螯合物的含量变化直接影响镉在植物体内的运输和分配.例如, 通过RNAi降低水稻的植物螯合素合成酶基因OsPCS1和OsPCS2的表达, 可以显著降低籽粒镉的含量, 然而其中的机制并不清楚(Das et al., 2017).Luo等(2018)的研究结果表明, 镉可以与植物防御素CAL1蛋白进行螯合, 与镉和植物螯合素的螯合类似, 均通过巯基与镉进行螯合.CAL1蛋白螯合镉后跨细胞膜分泌到胞外, 从而从细胞质中卸载出来, 进入木质部中进行长距离转运.镉与CAL1蛋白螯合之后, 限制了其通过韧皮部向水稻籽粒的再分配, 从而定向调控其在叶片等营养器官的积累而不影响籽粒中镉的积累.这一创新性研究加深了人们对重金属镉在植物体内的转运和再分配机理的认识, 同时也为培育秸秆镉高积累而籽粒镉含量达标的“修复型”水稻品种提供了有用的新基因, 研究成果具有重要的理论意义和应用价值.后续将通过挖掘CAL1强功能型自然变异优异等位基因以及原位过量表达CAL1基因等手段, 进一步提高水稻秸秆中镉的积累量; 同时, 在中低镉污染农田中实际考察镉的移除效率以及籽粒镉的含量, 以进一步提高CAL1在培育“修复型”水稻品种中的应用价值. ...
3 2011
... 镉是植物非必需的重金属元素之一, 主要通过锰、铁和锌等二价阳离子的吸收途径经根系进入植物体内.在水稻中, 镉主要通过定位于细胞质膜上的OsNRAMP5进行吸收(图1).osnramp5突变体丧失大部分镉的吸收能力, 其秸秆和籽粒中镉的含量都比野生型显著降低(Ishikawa et al., 2012; Sasaki et al., 2012; Yang et al., 2014).通过CRISPR/Cas9基因编辑技术将两系杂交稻双亲中的OsNRAMP5基因同时敲除, 获得的杂交种籽粒镉含量比双亲降低了98%, 在双亲籽粒镉超标10倍的情况下, 杂交种籽粒镉含量仍然低于国家标准, 显示OsNRAMP5基因在培育镉低吸收水稻品种中具有良好的应用前景(Tang et al., 2017).然而, OsNRAMP5同时也是植物必需矿质营养元素锰的转运蛋白(Ishimaru et al., 2012; Sasaki et al., 2012), 参与植物体内锰从根部向地上部的转运以及再分配(Yang et al., 2014).在水培条件下, osnramp5突变体比野生型对缺锰更为敏感(Sasaki et al., 2012; Yang et al., 2014), 这可能限制了osnramp5突变体在控制镉吸收中的实际应用, 尤其是在低锰农田中的应用.镉离子通过根部吸收进入植物体以后, 通常被区隔化到液泡中(图1), 在水稻中, 这一过程由P-1B型重金属ATP酶OsHMA3负责(Ueno et al., 2010).在OsHMA3的T-DNA插入突变体或者OsHMA3功能丧失的水稻品种中, 镉未能被转运到液泡中, 从而导致由根系向地上部和籽粒运输大幅度上升, 引起籽粒中镉含量升高(Ueno et al., 2010; Miyadate et al., 2011; Yan et al., 2016).相反, 过量表达OsHMA3基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ... ... ; Sasaki et al., 2012), 参与植物体内锰从根部向地上部的转运以及再分配(Yang et al., 2014).在水培条件下, osnramp5突变体比野生型对缺锰更为敏感(Sasaki et al., 2012; Yang et al., 2014), 这可能限制了osnramp5突变体在控制镉吸收中的实际应用, 尤其是在低锰农田中的应用.镉离子通过根部吸收进入植物体以后, 通常被区隔化到液泡中(图1), 在水稻中, 这一过程由P-1B型重金属ATP酶OsHMA3负责(Ueno et al., 2010).在OsHMA3的T-DNA插入突变体或者OsHMA3功能丧失的水稻品种中, 镉未能被转运到液泡中, 从而导致由根系向地上部和籽粒运输大幅度上升, 引起籽粒中镉含量升高(Ueno et al., 2010; Miyadate et al., 2011; Yan et al., 2016).相反, 过量表达OsHMA3基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ... ... 突变体比野生型对缺锰更为敏感(Sasaki et al., 2012; Yang et al., 2014), 这可能限制了osnramp5突变体在控制镉吸收中的实际应用, 尤其是在低锰农田中的应用.镉离子通过根部吸收进入植物体以后, 通常被区隔化到液泡中(图1), 在水稻中, 这一过程由P-1B型重金属ATP酶OsHMA3负责(Ueno et al., 2010).在OsHMA3的T-DNA插入突变体或者OsHMA3功能丧失的水稻品种中, 镉未能被转运到液泡中, 从而导致由根系向地上部和籽粒运输大幅度上升, 引起籽粒中镉含量升高(Ueno et al., 2010; Miyadate et al., 2011; Yan et al., 2016).相反, 过量表达OsHMA3基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ...
1 2012
... 重金属镉是我国农田和农产品的重要污染物之一.随着我国工业化和城市化的快速发展, 土壤重金属污染问题日益突出.环保部和国土资源部2014年发布的全国土壤污染调查报告显示, 我国耕地土壤19.4%点位污染物含量超标, 其中重金属镉的超标率为7%, 污染程度以中、低污染为主(环境保护部和国土资源部, 2014).农田重金属污染造成农产品重金属含量超标时常发生, 我国南方稻米重金属含量超标现象尤为严重(Zhao et al., 2015).调查结果显示, 我国湖南益阳和长株潭地区分别有60%和76%的稻米镉含量超过我国食品卫生标准稻米镉限量值——0.2 mg∙kg-1, 样品中最高镉含量分别达2.77和4.80 mg∙kg-1, 超过限量值13和24倍(Du et al., 2013; Zhu et al., 2016).土壤重金属污染已经导致我国居民的镉摄入量逐年增加.据国家卫生和计划生育委员会长达25年的调查统计结果显示, 我国居民人均镉的摄入量已从1990年的13.8 μg∙d-1上升到2015年的30.6 μg∙d-1, 25年间升高了122%, 达到世界卫生组织和世界粮农组织规定的日均镉允许摄入量(FAO/WHO Cd Tole- rable Daily Intake)的61%.其中, 稻米是膳食中镉的主要来源, 占我国人均每天镉摄入量的55%, 南方人群则占到了65% (Song et al., 2017).因此, 农田重金属污染造成的农产品重金属含量超标已经严重影响农产品的质量安全, 对我国居民的身体健康构成威胁, 已成为限制我国农产品质量安全和农业生产可持续发展的重要因素.因此, 重金属污染治理迫在眉睫. ...
1 2017
... 镉是植物非必需的重金属元素之一, 主要通过锰、铁和锌等二价阳离子的吸收途径经根系进入植物体内.在水稻中, 镉主要通过定位于细胞质膜上的OsNRAMP5进行吸收(图1).osnramp5突变体丧失大部分镉的吸收能力, 其秸秆和籽粒中镉的含量都比野生型显著降低(Ishikawa et al., 2012; Sasaki et al., 2012; Yang et al., 2014).通过CRISPR/Cas9基因编辑技术将两系杂交稻双亲中的OsNRAMP5基因同时敲除, 获得的杂交种籽粒镉含量比双亲降低了98%, 在双亲籽粒镉超标10倍的情况下, 杂交种籽粒镉含量仍然低于国家标准, 显示OsNRAMP5基因在培育镉低吸收水稻品种中具有良好的应用前景(Tang et al., 2017).然而, OsNRAMP5同时也是植物必需矿质营养元素锰的转运蛋白(Ishimaru et al., 2012; Sasaki et al., 2012), 参与植物体内锰从根部向地上部的转运以及再分配(Yang et al., 2014).在水培条件下, osnramp5突变体比野生型对缺锰更为敏感(Sasaki et al., 2012; Yang et al., 2014), 这可能限制了osnramp5突变体在控制镉吸收中的实际应用, 尤其是在低锰农田中的应用.镉离子通过根部吸收进入植物体以后, 通常被区隔化到液泡中(图1), 在水稻中, 这一过程由P-1B型重金属ATP酶OsHMA3负责(Ueno et al., 2010).在OsHMA3的T-DNA插入突变体或者OsHMA3功能丧失的水稻品种中, 镉未能被转运到液泡中, 从而导致由根系向地上部和籽粒运输大幅度上升, 引起籽粒中镉含量升高(Ueno et al., 2010; Miyadate et al., 2011; Yan et al., 2016).相反, 过量表达OsHMA3基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ...
3 2017
... 镉是植物非必需的重金属元素之一, 主要通过锰、铁和锌等二价阳离子的吸收途径经根系进入植物体内.在水稻中, 镉主要通过定位于细胞质膜上的OsNRAMP5进行吸收(图1).osnramp5突变体丧失大部分镉的吸收能力, 其秸秆和籽粒中镉的含量都比野生型显著降低(Ishikawa et al., 2012; Sasaki et al., 2012; Yang et al., 2014).通过CRISPR/Cas9基因编辑技术将两系杂交稻双亲中的OsNRAMP5基因同时敲除, 获得的杂交种籽粒镉含量比双亲降低了98%, 在双亲籽粒镉超标10倍的情况下, 杂交种籽粒镉含量仍然低于国家标准, 显示OsNRAMP5基因在培育镉低吸收水稻品种中具有良好的应用前景(Tang et al., 2017).然而, OsNRAMP5同时也是植物必需矿质营养元素锰的转运蛋白(Ishimaru et al., 2012; Sasaki et al., 2012), 参与植物体内锰从根部向地上部的转运以及再分配(Yang et al., 2014).在水培条件下, osnramp5突变体比野生型对缺锰更为敏感(Sasaki et al., 2012; Yang et al., 2014), 这可能限制了osnramp5突变体在控制镉吸收中的实际应用, 尤其是在低锰农田中的应用.镉离子通过根部吸收进入植物体以后, 通常被区隔化到液泡中(图1), 在水稻中, 这一过程由P-1B型重金属ATP酶OsHMA3负责(Ueno et al., 2010).在OsHMA3的T-DNA插入突变体或者OsHMA3功能丧失的水稻品种中, 镉未能被转运到液泡中, 从而导致由根系向地上部和籽粒运输大幅度上升, 引起籽粒中镉含量升高(Ueno et al., 2010; Miyadate et al., 2011; Yan et al., 2016).相反, 过量表达OsHMA3基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ... ... 功能丧失的水稻品种中, 镉未能被转运到液泡中, 从而导致由根系向地上部和籽粒运输大幅度上升, 引起籽粒中镉含量升高(Ueno et al., 2010; Miyadate et al., 2011; Yan et al., 2016).相反, 过量表达OsHMA3基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ... ... 基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ...
1 2010
... 镉是植物非必需的重金属元素之一, 主要通过锰、铁和锌等二价阳离子的吸收途径经根系进入植物体内.在水稻中, 镉主要通过定位于细胞质膜上的OsNRAMP5进行吸收(图1).osnramp5突变体丧失大部分镉的吸收能力, 其秸秆和籽粒中镉的含量都比野生型显著降低(Ishikawa et al., 2012; Sasaki et al., 2012; Yang et al., 2014).通过CRISPR/Cas9基因编辑技术将两系杂交稻双亲中的OsNRAMP5基因同时敲除, 获得的杂交种籽粒镉含量比双亲降低了98%, 在双亲籽粒镉超标10倍的情况下, 杂交种籽粒镉含量仍然低于国家标准, 显示OsNRAMP5基因在培育镉低吸收水稻品种中具有良好的应用前景(Tang et al., 2017).然而, OsNRAMP5同时也是植物必需矿质营养元素锰的转运蛋白(Ishimaru et al., 2012; Sasaki et al., 2012), 参与植物体内锰从根部向地上部的转运以及再分配(Yang et al., 2014).在水培条件下, osnramp5突变体比野生型对缺锰更为敏感(Sasaki et al., 2012; Yang et al., 2014), 这可能限制了osnramp5突变体在控制镉吸收中的实际应用, 尤其是在低锰农田中的应用.镉离子通过根部吸收进入植物体以后, 通常被区隔化到液泡中(图1), 在水稻中, 这一过程由P-1B型重金属ATP酶OsHMA3负责(Ueno et al., 2010).在OsHMA3的T-DNA插入突变体或者OsHMA3功能丧失的水稻品种中, 镉未能被转运到液泡中, 从而导致由根系向地上部和籽粒运输大幅度上升, 引起籽粒中镉含量升高(Ueno et al., 2010; Miyadate et al., 2011; Yan et al., 2016).相反, 过量表达OsHMA3基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ...
3 2016
... 镉是植物非必需的重金属元素之一, 主要通过锰、铁和锌等二价阳离子的吸收途径经根系进入植物体内.在水稻中, 镉主要通过定位于细胞质膜上的OsNRAMP5进行吸收(图1).osnramp5突变体丧失大部分镉的吸收能力, 其秸秆和籽粒中镉的含量都比野生型显著降低(Ishikawa et al., 2012; Sasaki et al., 2012; Yang et al., 2014).通过CRISPR/Cas9基因编辑技术将两系杂交稻双亲中的OsNRAMP5基因同时敲除, 获得的杂交种籽粒镉含量比双亲降低了98%, 在双亲籽粒镉超标10倍的情况下, 杂交种籽粒镉含量仍然低于国家标准, 显示OsNRAMP5基因在培育镉低吸收水稻品种中具有良好的应用前景(Tang et al., 2017).然而, OsNRAMP5同时也是植物必需矿质营养元素锰的转运蛋白(Ishimaru et al., 2012; Sasaki et al., 2012), 参与植物体内锰从根部向地上部的转运以及再分配(Yang et al., 2014).在水培条件下, osnramp5突变体比野生型对缺锰更为敏感(Sasaki et al., 2012; Yang et al., 2014), 这可能限制了osnramp5突变体在控制镉吸收中的实际应用, 尤其是在低锰农田中的应用.镉离子通过根部吸收进入植物体以后, 通常被区隔化到液泡中(图1), 在水稻中, 这一过程由P-1B型重金属ATP酶OsHMA3负责(Ueno et al., 2010).在OsHMA3的T-DNA插入突变体或者OsHMA3功能丧失的水稻品种中, 镉未能被转运到液泡中, 从而导致由根系向地上部和籽粒运输大幅度上升, 引起籽粒中镉含量升高(Ueno et al., 2010; Miyadate et al., 2011; Yan et al., 2016).相反, 过量表达OsHMA3基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ... ... ), 参与植物体内锰从根部向地上部的转运以及再分配(Yang et al., 2014).在水培条件下, osnramp5突变体比野生型对缺锰更为敏感(Sasaki et al., 2012; Yang et al., 2014), 这可能限制了osnramp5突变体在控制镉吸收中的实际应用, 尤其是在低锰农田中的应用.镉离子通过根部吸收进入植物体以后, 通常被区隔化到液泡中(图1), 在水稻中, 这一过程由P-1B型重金属ATP酶OsHMA3负责(Ueno et al., 2010).在OsHMA3的T-DNA插入突变体或者OsHMA3功能丧失的水稻品种中, 镉未能被转运到液泡中, 从而导致由根系向地上部和籽粒运输大幅度上升, 引起籽粒中镉含量升高(Ueno et al., 2010; Miyadate et al., 2011; Yan et al., 2016).相反, 过量表达OsHMA3基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ... ... ; Yang et al., 2014), 这可能限制了osnramp5突变体在控制镉吸收中的实际应用, 尤其是在低锰农田中的应用.镉离子通过根部吸收进入植物体以后, 通常被区隔化到液泡中(图1), 在水稻中, 这一过程由P-1B型重金属ATP酶OsHMA3负责(Ueno et al., 2010).在OsHMA3的T-DNA插入突变体或者OsHMA3功能丧失的水稻品种中, 镉未能被转运到液泡中, 从而导致由根系向地上部和籽粒运输大幅度上升, 引起籽粒中镉含量升高(Ueno et al., 2010; Miyadate et al., 2011; Yan et al., 2016).相反, 过量表达OsHMA3基因可以将镉截留在根部, 从而显著降低籽粒中镉的积累(Ueno et al., 2010).虽然通过敲除OsNRAMP5和过量表达OsHMA3可以有效降低籽粒镉的积累, 但是镉依旧滞留于污染农田中.如何将镉定向分配到水稻地上部秸秆和叶片等非可食部分, 同时保证籽粒镉含量不超标, 在实现水稻安全的同时对镉进行移除修复, 一直停留在概念的阶段. ...
1 2014
... 重金属镉是我国农田和农产品的重要污染物之一.随着我国工业化和城市化的快速发展, 土壤重金属污染问题日益突出.环保部和国土资源部2014年发布的全国土壤污染调查报告显示, 我国耕地土壤19.4%点位污染物含量超标, 其中重金属镉的超标率为7%, 污染程度以中、低污染为主(环境保护部和国土资源部, 2014).农田重金属污染造成农产品重金属含量超标时常发生, 我国南方稻米重金属含量超标现象尤为严重(Zhao et al., 2015).调查结果显示, 我国湖南益阳和长株潭地区分别有60%和76%的稻米镉含量超过我国食品卫生标准稻米镉限量值——0.2 mg∙kg-1, 样品中最高镉含量分别达2.77和4.80 mg∙kg-1, 超过限量值13和24倍(Du et al., 2013; Zhu et al., 2016).土壤重金属污染已经导致我国居民的镉摄入量逐年增加.据国家卫生和计划生育委员会长达25年的调查统计结果显示, 我国居民人均镉的摄入量已从1990年的13.8 μg∙d-1上升到2015年的30.6 μg∙d-1, 25年间升高了122%, 达到世界卫生组织和世界粮农组织规定的日均镉允许摄入量(FAO/WHO Cd Tole- rable Daily Intake)的61%.其中, 稻米是膳食中镉的主要来源, 占我国人均每天镉摄入量的55%, 南方人群则占到了65% (Song et al., 2017).因此, 农田重金属污染造成的农产品重金属含量超标已经严重影响农产品的质量安全, 对我国居民的身体健康构成威胁, 已成为限制我国农产品质量安全和农业生产可持续发展的重要因素.因此, 重金属污染治理迫在眉睫. ...
1 2015
... 重金属镉是我国农田和农产品的重要污染物之一.随着我国工业化和城市化的快速发展, 土壤重金属污染问题日益突出.环保部和国土资源部2014年发布的全国土壤污染调查报告显示, 我国耕地土壤19.4%点位污染物含量超标, 其中重金属镉的超标率为7%, 污染程度以中、低污染为主(环境保护部和国土资源部, 2014).农田重金属污染造成农产品重金属含量超标时常发生, 我国南方稻米重金属含量超标现象尤为严重(Zhao et al., 2015).调查结果显示, 我国湖南益阳和长株潭地区分别有60%和76%的稻米镉含量超过我国食品卫生标准稻米镉限量值——0.2 mg∙kg-1, 样品中最高镉含量分别达2.77和4.80 mg∙kg-1, 超过限量值13和24倍(Du et al., 2013; Zhu et al., 2016).土壤重金属污染已经导致我国居民的镉摄入量逐年增加.据国家卫生和计划生育委员会长达25年的调查统计结果显示, 我国居民人均镉的摄入量已从1990年的13.8 μg∙d-1上升到2015年的30.6 μg∙d-1, 25年间升高了122%, 达到世界卫生组织和世界粮农组织规定的日均镉允许摄入量(FAO/WHO Cd Tole- rable Daily Intake)的61%.其中, 稻米是膳食中镉的主要来源, 占我国人均每天镉摄入量的55%, 南方人群则占到了65% (Song et al., 2017).因此, 农田重金属污染造成的农产品重金属含量超标已经严重影响农产品的质量安全, 对我国居民的身体健康构成威胁, 已成为限制我国农产品质量安全和农业生产可持续发展的重要因素.因此, 重金属污染治理迫在眉睫. ...
 ,
, 

{kind=link}
{kind=link}