Abstract: In recent years, Physalis plants have attracted increasing attention worldwide due to their high nutritional value, edible fruit, and potential medicinal value. In this study, simple sequence repeat (SSR) markers were used to assess genetic relationships with 22 samples of four Physalis species mainly distributed in China. Twenty SSR primer pairs produced 118 loci, 90.7% (107) of which showed polymorphism. The average interspecies similarity coefficient was 0.501, which indicates a degree of genetic relationship among Physalis species. The results of UPGMA dendrography and PCoA plotting were similar, and all Physalis samples were grouped into two clusters. All P. alkekengi var. francheti samples, distant from any other Physalis species, constituted a separate cluster, which confirmed findings of previous studies. This study also indicated that SSR markers are rich in genetic information and could be used to assess the genetic diversity of Physalis species which provides rich useful information for protecting the Physalis germplasm resource and an important foundation for molecular assisted-breeding programs with Physalis.
Key words:Physalis ; genetic relationship ; SSR markers
1.5 数据统计分析利用Quantity one软件(Version 4.6.2)及人工辅助判读的方法进行电泳图拷带。为了保证结果的准确性, 每对引物进行2次重复PCR扩增和电泳检测, 并只统计清晰度高且稳定性好的电泳位点。同一位点有条带记作1, 无条带记作0。将整理好的0/1原始矩阵数据输入NTSYS-pc 2.10e软件(Rohlf, 2000), 计算样品间的遗传相似系数(simple matching coefficie, SM) (Nei and Li, 1979), 并基于非加权组平均法(unweighted pair group method with an arithmetic mean, UPGMA)进行聚类分析。另外, 运用NTSYSpc 2.10e软件中的Dcenter及Eigen程序进行主坐标分析(principal coordinate analysis, PCoA) (Gower, 1966)。
2 结果与讨论2.1 SSR多态性分析20对SSR引物共扩增出118个可靠的电泳位点, 每对引物扩增出的电泳条带数分布于2-11之间, 每对引物平均扩增出5.9个位点(表2)。在所有电泳条带中, 107个条带是多态性位点, 每条引物扩增出的多态性位点数分布于2-11之间, 平均每对引物扩增出5.4个多态性位点。每对引物的多态性比例分布于75.0%-100.0%之间, 平均多态性百分比为90.7%。引物SSR55和SSR112的电泳检测结果见图1。 表2 Table 2 表2 表2 20对简单重复序列(SSR)引物的扩增结果及多态性信息 Table 2 Amplification results and polymorphism information of 20 simple sequence repeats (SSR) primer pairs
Primer name
Primer sequence (5'-3')
Repeat type
Tm
No. of loci
Polymorphic loci
Polymorphism rate (%)
SSR2
F: CATTGGGTTTCGCATCCAT
AG
60
6
6
100.0
R: AGACAAGCCTAGGGGAAAGG
SSR9
F: TGCTCCGAGTTTTAGGGTTC
AG
60
8
7
87.5
R: GCAGTTGGTAAAGTTGAGAGACG
SSR10
F: GCTTCCTATTGTGTTGCCTGA
AT
58
5
4
80.0
R: ACTTTGGGTTTCGGGAATTG
SSR11
F: CAGCTGAAATAAGAGAGTGATTGG
AG
57
4
3
75.0
R: CCCTCTTTTTCTCCTCCGAGT
SSR13
F: GCGGAATCCATTGTTTTTCA
AC
58
9
8
88.9
R: CCGATGGAGTATAGTCACGCAAA
SSR15
F: GCTTGTTGATCAGCTTTCTTTG
AT
57
7
6
85.7
R: TGGATCATAACCTTGCTAATGC
SSR36
F: ATGAACCACATGTCGGAGGA
AG
58
7
6
85.7
R: GGGGATCCAAACGAAGTGTA
SSR54
F: CGGCTGGTATGCTTACAAAGAT
AC
58
4
4
100.0
R: GCACTTCCACTGTTTTTAACTTCC
SSR55
F: CACCTACATAGGCAGCCAAAA
AG
58
7
6
85.7
R: ATTTGTGGGCGGAGGAAG
SSR57
F: AGTGAAAAGCAGCCCATTCT
AT
56
9
8
88.9
R: GGCGAAGCTGAATTGAAAAA
SSR67
F: GCTTCTGTTCCATTATTCACCA
AG
56
5
5
100.0
R: GCAGTGTGGGATCAATCAAT
SSR68
F: GAAGCAAACAACTACACCCAAA
AG
56
8
8
100.0
R: AAGCCTCGGATTTCATAGCA
SSR77
F: CATACCATAACTCCCCATCTCTC
AG
57
4
3
75.0
R: TGCCGATTCTGATTTCTTCC
SSR92
F: TGGTTTGAGGATCAAGAAAGAA
AAG
56
5
4
80.0
R: GTGGTATCAACGCAGAGTGG
SSR107
F: CATCCAACACCAGAAATACGC
AAG
58
4
4
100.0
R: TCCAACTTTATCATTTCTTCCAC
SSR110
F: CACCCATATCCCAATCTTCTTC
CTT
60
4
4
100.0
R: GGGTAATTTTCACGGGGAAT
SSR112
F: CTACGCCTACCACTTGCACA
TCT
60
11
11
100.0
R: CAGTGGAAGCCTCAAGATCC
SSR118
F: AATCAAGGGTCAGAAGAAATGG
ATC
58
2
2
100.0
R: GCAAGAATGGATGTGGGTGT
SSR123
F: TCAGTGGAGCGCGTATATCT
ATC
60
5
5
100.0
R: GCGATCTCACCAAACCTCTC
SSR127
F: TTGGTTTGGCATAACTGCAA
AAT
58
4
3
75.0
R: GGTTTGCAACTCTCATGCTG
Average
-
-
-
5.9
5.4
90.4
Total
-
-
-
118
107
-
表2 20对简单重复序列(SSR)引物的扩增结果及多态性信息 Table 2 Amplification results and polymorphism information of 20 simple sequence repeats (SSR) primer pairs
图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-305/img_1.png<b>图1</b> 引物SSR55 (A)和SSR112 (B)在酸浆属植物中的电泳检测结果<br/>M: DNA分子标准。泳道1-22: 22个酸浆属样品(编号同<xref ref-type="table" rid="T1-1674-3466-53-3-305">表1</xref>)<br/><b>Figure 1</b> Amplification profile of primer SSR55 (A) and SSR112 (B) in <i>Physalis</i> samples<br/>M: DNA molecular standards. Lane 1-22: The 22 <i>Physalis</i> samples (sample number is the same as in <xref ref-type="table" rid="T1-1674-3466-53-3-305">Table 1</xref>). Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-305/img_1.png<b>图1</b> 引物SSR55 (A)和SSR112 (B)在酸浆属植物中的电泳检测结果<br/>M: DNA分子标准。泳道1-22: 22个酸浆属样品(编号同<xref ref-type="table" rid="T1-1674-3466-53-3-305">表1</xref>)<br/><b>Figure 1</b> Amplification profile of primer SSR55 (A) and SSR112 (B) in <i>Physalis</i> samples<br/>M: DNA molecular standards. Lane 1-22: The 22 <i>Physalis</i> samples (sample number is the same as in <xref ref-type="table" rid="T1-1674-3466-53-3-305">Table 1</xref>).
图1 引物SSR55 (A)和SSR112 (B)在酸浆属植物中的电泳检测结果 M: DNA分子标准。泳道1-22: 22个酸浆属样品(编号同表1) Figure 1 Amplification profile of primer SSR55 (A) and SSR112 (B) in Physalis samples M: DNA molecular standards. Lane 1-22: The 22 Physalis samples (sample number is the same as in Table 1).
Average interspecies genetic similarity coefficient
0.501±0.074
P. angulata vs P. minima
0.600±0.042
P. alkekengi var. francheti vs P. minima
0.437±0.036
P. pubescens vs P. minima
0.570±0.037
P. angulata vs P. alkekengi var. francheti
0.444±0.044
P. angulata vs P. pubescens
0.514±0.043
P. pubescens vs P. alkekengi var. francheti
0.382±0.040
表3 酸浆属植物平均种间遗传相似系数 Table 3 Average interspecies genetic similarity coefficient in Physalis samples
2.3 聚类分析基于SM遗传相似系数矩阵, 用UPGMA法对所有供试样品进行聚类分析(图2)。从图2可以看出, 在遗传相似系数为0.420处, 可以将22份供试样品归为2大类: I组和II组。其中, I组比较复杂, 包括了毛酸浆、苦蘵和小酸浆的所有样品, I组又可以分为I-1、I-2和I-3共3个亚组, 分别包括了上述3个酸浆属物种的所有供试样本。酸浆的所有供试样品则远离其它酸浆属植物样品, 单独构成II组。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-305/img_2.png<b>图2</b> 4种酸浆属植物22份样品的简单重复序列(SSR)聚类树<br/><b>Figure 2</b> UPGMA dendrogram for 22 samples of 4 <i>Physalis</i> species based on simple sequence repeats (SSR) markers Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-305/img_2.png<b>图2</b> 4种酸浆属植物22份样品的简单重复序列(SSR)聚类树<br/><b>Figure 2</b> UPGMA dendrogram for 22 samples of 4 <i>Physalis</i> species based on simple sequence repeats (SSR) markers
图2 4种酸浆属植物22份样品的简单重复序列(SSR)聚类树 Figure 2 UPGMA dendrogram for 22 samples of 4 Physalis species based on simple sequence repeats (SSR) markers
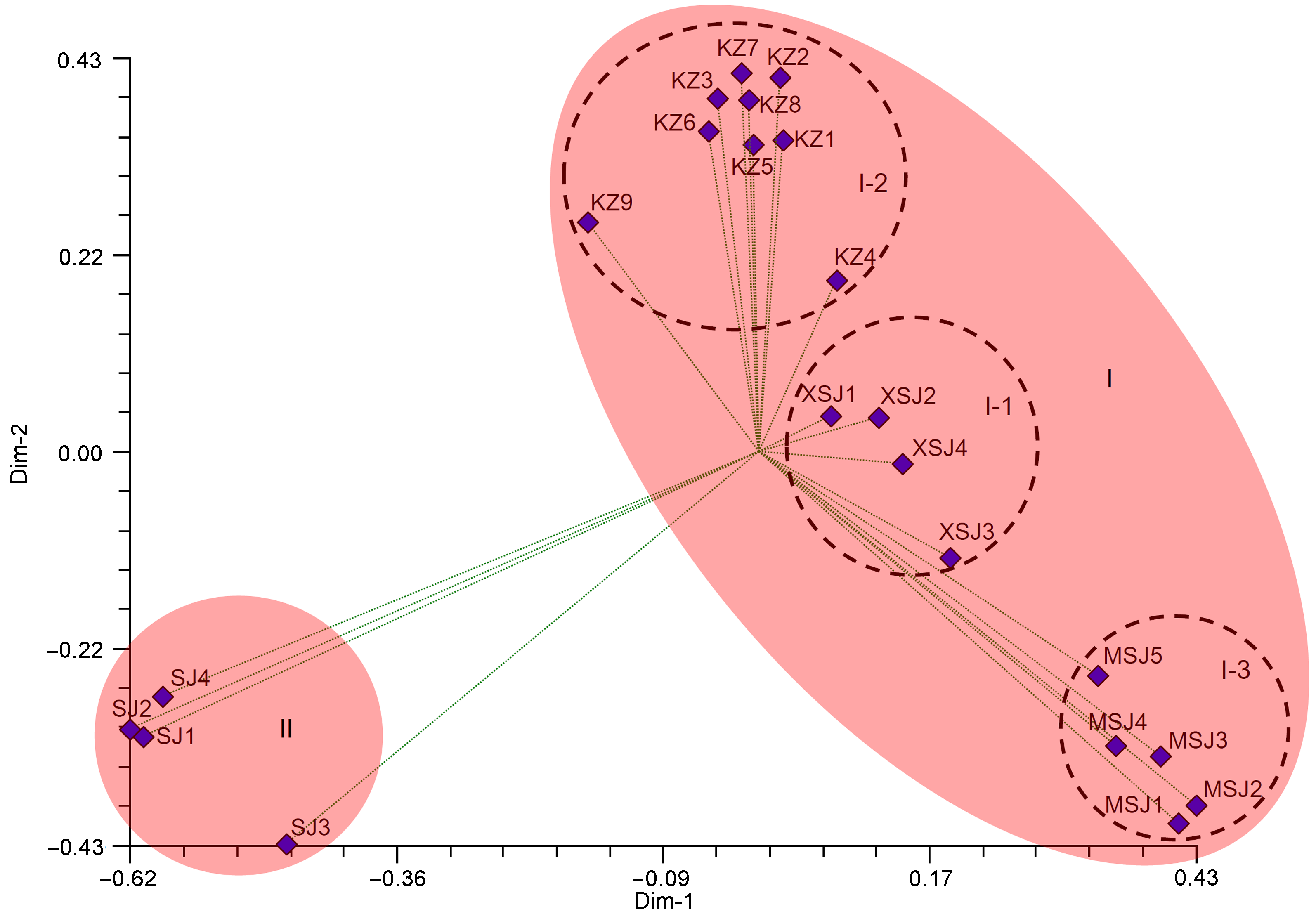
2.4 主坐标分析利用NTSYS-pc 2.10e软件对供试样品进行PCoA分析。22个样品的二维主坐标分析结果如图3所示, 前2个主坐标分别揭示总遗传变异的24.3%和23.9%, 累计可揭示总变异的48.2%。主坐标分析结果与UP- GMA聚类分析结果基本一致, 并且直观地呈现了各个酸浆属物种间的亲缘关系。结果显示, 22个供试样品同样可以分为2大类, 同一个物种的样本倾向于集中在同一个空间区域。 图3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-305/img_3.png<b>图3</b> 基于简单重复序列(SSR)标记的22份酸浆属植物样品二维主坐标分析<br/>编码同<xref ref-type="table" rid="T1-1674-3466-53-3-305">表1</xref>。<br/><b>Figure 3</b> The analysis of PCoA for 22 <i>Physalis</i> samples based on simple sequence repeats (SSR) markers along the first two principal axes<br/>The codes are the same as in <xref ref-type="table" rid="T1-1674-3466-53-3-305">Table 1</xref>. Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-3-305/img_3.png<b>图3</b> 基于简单重复序列(SSR)标记的22份酸浆属植物样品二维主坐标分析<br/>编码同<xref ref-type="table" rid="T1-1674-3466-53-3-305">表1</xref>。<br/><b>Figure 3</b> The analysis of PCoA for 22 <i>Physalis</i> samples based on simple sequence repeats (SSR) markers along the first two principal axes<br/>The codes are the same as in <xref ref-type="table" rid="T1-1674-3466-53-3-305">Table 1</xref>.
图3 基于简单重复序列(SSR)标记的22份酸浆属植物样品二维主坐标分析 编码同表1。 Figure 3 The analysis of PCoA for 22 Physalis samples based on simple sequence repeats (SSR) markers along the first two principal axes The codes are the same as in Table 1.
2.5 讨论与RAPD和ISSR等表现为显性类型的分子标记相比, SSR标记是共显性分子标记, 因此, 更能准确地揭示物种之间的亲缘关系。近年来, SSR分子标记已经成功应用于许多植物的亲缘关系研究(Wei et al., 2012; Teshome et al., 2015; Wang et al., 2015; Feng et al., 2016a)。Simbaqueba等(2011)基于秘鲁苦蘵(P. peruviana)的转录组数据开发出一批可用于秘鲁苦蘵分子鉴定及遗传多样性研究的SSR标记。本研究表明, 66.7% (20/30)的秘鲁苦蘵SSR标记可以有效用于苦蘵、小酸浆、酸浆及毛酸浆的遗传关系研究(表2)。另有结果显示, 90.7% (107/118)的SSR扩增条带是多态性条带, 高于甘草属(Glycyrrhiza)植物中SSR分子标记的多态性(89.44%) (李晓岚等, 2015), 以及芹菜(Apium graveolens)栽培品种中的SSR多态性比例(53.85%) (Fu et al., 2014)。由此可见, 本研究筛选出来的20对SSR分子标记引物在酸浆属内具有很好的适用性, 可以用于该属植物之间的亲缘关系研究。 准确评价不同植物物种之间的亲缘关系是植物分子育种和资源保护的重要基础。由于酸浆属植物种间的形态学性状非常相似, 多年来酸浆属植物的鉴定和分类被认为是茄科植物中非常困难的科学挑战(Maggie and Manos, 2005; Wei et al., 2012; Feng et al., 2016b)。本研究中, 供试的4种酸浆属植物分别属于2个亚属(Maggie and Manos, 2005; Feng et al., 2016b)。其中, 苦蘵和毛酸浆分别属于Rydbergis亚属的Angulatae组和Epeteiorhiza组, 小酸浆同样属于Rydbergis亚属, 但是没有组级别报道; 与其它3种酸浆属植物不同, 酸浆属于Physalis亚属。从UPGMA聚类和PCoA分析结果来看, 来自同一种酸浆属植物的所有样本被聚类在一起。供试酸浆属样品分为2大类(I和II), 其中, 来自Rydbergis亚属的苦蘵、毛酸浆和小酸浆被归为I类; 来自Physalis亚属的酸浆则远离其它酸浆属植物, 单独聚类为II类。我们之前的研究结果表明, 许多酸浆属植物在系统演化过程中的表现并不是单源进化。此外, 我们的研究同样发现酸浆与其它酸浆属植物的遗传关系较远(Feng et al., 2016b, 2018)。Maggie和Manos (2005)研究认为, 酸浆是一种非典型的酸浆属植物, 应该单独列为一个亚属。依据Axelius (1996)关于酸浆属植物的形态学分类方法, 本研究中的苦蘵、毛酸浆和小酸浆属于Rydbergis亚属, 而酸浆属于Physalis亚属。酸浆是目前唯一被《中华人民共和国药典》(2015版)收录的酸浆属药用植物, 其分类归属问题一直以来是分类学家关注的焦点。本研究则在很大程度上与Axelius (1996)以及Maggie和Manos (2005)的研究结论一致, 表明SSR分子标记在酸浆属植物亲缘关系评价中具有较好的应用性。 综上所述, 本研究结果表明, SSR是一种稳定性好且多态性高的分子标记, 可以很好地用于供试酸浆属植物的亲缘关系研究。研究结果为分布在我国的酸浆属植物的合理利用、种质评价和资源保护奠定了重要基础。在后续研究中, 我们将收集更多的酸浆属植物, 以评价SSR分子标记在更多酸浆属植物研究中的适用性。
The authors have declared that no competing interests exist.
中国科学院中国植物志编辑委员会 (1978). 中国植物志(第67卷). 北京: 科学出版社. pp. 50.
[12]
AxeliusB (1996). The phylogenetic relationships of the physaloid genera (Solanaceae) based on morphological data.Am J Bot 83, 118-124. [本文引用: 2]
[13]
DingH, HuZJ, YuLY, MaZY, MaXQ, ChenZ, WangD, ZhaoXF (2014). Induction of quinone reductase (QR) by withanolides isolated from Physalis angulata L. var. villosa Bonati (Solanaceae). Steroids 86, 32-38. [本文引用: 2]
[14]
FengSG, HeRF, LuJJ, JiangMY, ShenXY, JiangY, WangZA, WangHZ (2016a). Development of SSR markers and assessment of genetic diversity in medicinal Chrysanthemum morifolium cultivars.Front Genet 7, 113. [本文引用: 2]
[15]
FengSG, JiangMY, ShiYJ, JiaoKL, ShenCJ, LuJJ, YingQC, WangHZ (2016b). Application of the ribosomal DNA ITS2 region of Physalis ( Solanaceae): DNA barcoding and phylogenetic study.Front Plant Sci 7, 1047. [本文引用: 4]
[16]
FengSG, JiaoKL, ZhuYJ, WangHF, JiangMY, WangHZ (2018). Molecular identification of species of Physalis ( Solanaceae) using a candidate DNA barcode: the chloroplast psbA-trnH intergenic region. Genome 61, 15-20. [本文引用: 2]
[17]
FuN, WangPY, LiuXD, ShenHL (2014). Use of EST-SSR markers for evaluating genetic diversity and fingerprinting celery (Apium graveolens L.) cultivars.Molecules 19, 1939-1955. [本文引用: 1]
[18]
Garzon-MartínezGA, Osorio-GuarínJA, Delgádillo-DuranP, MayorgaF, Enciso-RodríguezFE, LandsmanD, Mariño-RamírezL, BarreroLS (2015). Genetic diversity and population structure in Physalis peruviana and related taxa based on InDels and SNPs derived from COSII and IRG markers.Plant Gene 4, 29-37. [本文引用: 1]
[19]
GowerJC (1966). Some distance properties of latent root and vector methods used in multivariate analysis.Biometrika 53, 325-338. [本文引用: 1]
[20]
JiL, YuanYL, MaZJ, ChenZ, GanLS, MaXQ, HuangDS (2013). Induction of quinone reductase (QR) by withanolides isolated from Physalis pubescens L. ( Solanaceae).Steroids 78, 860-865. [本文引用: 1]
[21]
LiX, ZhaoJP, YangM, LiuYL, LiZC, LiRY, LiXR, LiN, XuQM, KhanIA, YangSL (2014). Physalins and withanolides from the fruits ofPhysalis alkekengi L. var. fran- chetii ( Mast.) Makino and the inhibitory activities against human tumor cells.Phytochem Lett 10, 95-100. [本文引用: 1]
[22]
MaggieWPS, ManosPS (2005). Untangling Physalis ( Solanaceae) from the physaloids: a two-gene phylogeny of the Physalinae.Syst Bot 30, 216-230. [本文引用: 5]
[23]
NeiM, LiWH (1979). Mathematical model for studying genetic variation in terms of restriction endonucleases.Proc Natl Acad Sci USA 76, 5269-5273. [本文引用: 1]
[24]
OlmsteadRG, BohsL, MigidHA, Santiago-ValentinE, GarciaVF, CollierSM (2008). A molecular phylogeny of the Solanaceae.Taxon 57, 1159-1181. [本文引用: 1]
[25]
RohlfFJ (2000). NTSYS-PC: numerical taxonomy and multivariate analysis system, version 2.00. Setauket, New York:Exeter Software. [本文引用: 1]
[26]
SimbaquebaJ, SánchezP, SanchezE, ZarantesVMN, ChaconMI, BarreroLS, Mariño-RamírezL (2011). Development and characterization of microsatellite markers for the cape gooseberry Physalis peruviana.PLoS One 6, e26719. [本文引用: 2]
[27]
SunCP, NieXF, KangN, ZhaoF, ChenLX, QiuF (2017). A new phenol glycoside from Physalis angulata.Nat Prod Res 31, 1059-1065. [本文引用: 1]
[28]
TeshomeA, BryngelssonT, DagneK, GeletaM (2015). Assessment of genetic diversity in Ethiopian field pea ( Pisum sativum L.) accessions with newly developed EST- SSR markers.BMC Genet 16, 102. [本文引用: 1]
[29]
WangF, YangT, BurlyaevaM, LiL, JiangJY, FangL, ReddenR, ZongXX (2015). Genetic diversity of grasspea and its relative species revealed by SSR markers.PLoS One 10, e0118542. [本文引用: 1]
[30]
WeiJL, HuXR, YangJJ, YangWC (2012). Identification of single-copy orthologous genes between Physalis and Solanum lycopersicum and analysis of genetic diversity in Physalis using molecular markers.PLoS One 7, e50164. [本文引用: 3]
[31]
XuXM, GuanYZ, ShanSM, LuoJG, KongLY (2016). Withaphysalin-type withanolides from Physalis minima.Phytochem Lett 15, 1-6. [本文引用: 1]
[32]
YangYK, XieSD, XuWX, NianY, LiuXL, PengXR, DingZT, QiuMH (2016). Six new physalins from Physalis alkekengi var. franchetii and their cytotoxicity and antibacterial activity. Fitoterapia 112, 144-152. [本文引用: 1]
[33]
Zamora-TavaresP, Vargas-PonceO, Sanchez-MartínezJ, Cabrera-ToledoD (2015). Diversity and genetic structure of the husk tomato ( Physalis philadelphica Lam.) in Wes- tern Mexico.Genet Res Crop Evol 62, 141-153. [本文引用: 1]
[34]
ZhangWN, TongWY (2016). Chemical constituents and biological activities of plants from the genus Physalis.Chem Biodivers 13, 48-65. [本文引用: 2]
... 酸浆属(Physalis)是茄科(Solanaceae)中一个非常重要的属, 为一年生或多年生草本植物.全球酸浆属植物共约75-120种, 大多分布于美洲热带及温带地区, 少数分布于欧亚大陆及东南亚(中国科学院中国植物志编辑委员会, 1978; Maggie and Manos, 2005; Wei et al., 2012).该属在我国主要分布于华东、华中、华南及西南等地, 常生于海拔500-1 500 m的山谷林下及村边路旁.据《中国植物志》记载, 分布于我国的酸浆属植物主要有5种和2个变种.大多数酸浆属植物具有抗氧化、抗炎及抗癌等多种药理活性(Ding et al., 2014; 杨金颖等, 2014; Zhang and Tong, 2016), 其果、根或全草可入药, 常用来治疗疟疾、风湿、肝炎、哮喘及癌症等疾病(许亮等, 2009; 国家药典委员会, 2015; Xu et al., 2016).其中, 酸浆(P. alkekengi var. francheti), 又名锦灯笼、挂金灯, 被《中华人民共和国药典》(2015年版)收录.另外, 苦蘵(P. angulata)、小酸浆(P. minima)和毛酸浆(P. pubescens)等在很多中药材地方志中也都有记载.除此之外, 大部分酸浆属植物富含多种维生素, 可以用来做罐头、饮料、蜜饯或直接食用的“本草水果”(孙海涛和高玉超, 2013; 王晓英等, 2014; 郭瑜等, 2017; 王赢等, 2017). ...
毛酸浆浆果的化学成分研究 1 2013
... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ...
4种甘草属植物EST-SSR引物开发及其亲缘关系分析 1 2015
... 与RAPD和ISSR等表现为显性类型的分子标记相比, SSR标记是共显性分子标记, 因此, 更能准确地揭示物种之间的亲缘关系.近年来, SSR分子标记已经成功应用于许多植物的亲缘关系研究(Wei et al., 2012; Teshome et al., 2015; Wang et al., 2015; Feng et al., 2016a).Simbaqueba等(2011)基于秘鲁苦蘵(P. peruviana)的转录组数据开发出一批可用于秘鲁苦蘵分子鉴定及遗传多样性研究的SSR标记.本研究表明, 66.7% (20/30)的秘鲁苦蘵SSR标记可以有效用于苦蘵、小酸浆、酸浆及毛酸浆的遗传关系研究(表2).另有结果显示, 90.7% (107/118)的SSR扩增条带是多态性条带, 高于甘草属(Glycyrrhiza)植物中SSR分子标记的多态性(89.44%) (李晓岚等, 2015), 以及芹菜(Apium graveolens)栽培品种中的SSR多态性比例(53.85%) (Fu et al., 2014).由此可见, 本研究筛选出来的20对SSR分子标记引物在酸浆属内具有很好的适用性, 可以用于该属植物之间的亲缘关系研究. ...
毛酸浆的化学成分研究 1 2015
... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ...
毛酸浆番茄复合调味酱的研制 1 2013
... 酸浆属(Physalis)是茄科(Solanaceae)中一个非常重要的属, 为一年生或多年生草本植物.全球酸浆属植物共约75-120种, 大多分布于美洲热带及温带地区, 少数分布于欧亚大陆及东南亚(中国科学院中国植物志编辑委员会, 1978; Maggie and Manos, 2005; Wei et al., 2012).该属在我国主要分布于华东、华中、华南及西南等地, 常生于海拔500-1 500 m的山谷林下及村边路旁.据《中国植物志》记载, 分布于我国的酸浆属植物主要有5种和2个变种.大多数酸浆属植物具有抗氧化、抗炎及抗癌等多种药理活性(Ding et al., 2014; 杨金颖等, 2014; Zhang and Tong, 2016), 其果、根或全草可入药, 常用来治疗疟疾、风湿、肝炎、哮喘及癌症等疾病(许亮等, 2009; 国家药典委员会, 2015; Xu et al., 2016).其中, 酸浆(P. alkekengi var. francheti), 又名锦灯笼、挂金灯, 被《中华人民共和国药典》(2015年版)收录.另外, 苦蘵(P. angulata)、小酸浆(P. minima)和毛酸浆(P. pubescens)等在很多中药材地方志中也都有记载.除此之外, 大部分酸浆属植物富含多种维生素, 可以用来做罐头、饮料、蜜饯或直接食用的“本草水果”(孙海涛和高玉超, 2013; 王晓英等, 2014; 郭瑜等, 2017; 王赢等, 2017). ...
毛酸浆开发利用的研究进展 1 2014
... 酸浆属(Physalis)是茄科(Solanaceae)中一个非常重要的属, 为一年生或多年生草本植物.全球酸浆属植物共约75-120种, 大多分布于美洲热带及温带地区, 少数分布于欧亚大陆及东南亚(中国科学院中国植物志编辑委员会, 1978; Maggie and Manos, 2005; Wei et al., 2012).该属在我国主要分布于华东、华中、华南及西南等地, 常生于海拔500-1 500 m的山谷林下及村边路旁.据《中国植物志》记载, 分布于我国的酸浆属植物主要有5种和2个变种.大多数酸浆属植物具有抗氧化、抗炎及抗癌等多种药理活性(Ding et al., 2014; 杨金颖等, 2014; Zhang and Tong, 2016), 其果、根或全草可入药, 常用来治疗疟疾、风湿、肝炎、哮喘及癌症等疾病(许亮等, 2009; 国家药典委员会, 2015; Xu et al., 2016).其中, 酸浆(P. alkekengi var. francheti), 又名锦灯笼、挂金灯, 被《中华人民共和国药典》(2015年版)收录.另外, 苦蘵(P. angulata)、小酸浆(P. minima)和毛酸浆(P. pubescens)等在很多中药材地方志中也都有记载.除此之外, 大部分酸浆属植物富含多种维生素, 可以用来做罐头、饮料、蜜饯或直接食用的“本草水果”(孙海涛和高玉超, 2013; 王晓英等, 2014; 郭瑜等, 2017; 王赢等, 2017). ...
毛酸浆发酵果脯的研制 1 2017
... 酸浆属(Physalis)是茄科(Solanaceae)中一个非常重要的属, 为一年生或多年生草本植物.全球酸浆属植物共约75-120种, 大多分布于美洲热带及温带地区, 少数分布于欧亚大陆及东南亚(中国科学院中国植物志编辑委员会, 1978; Maggie and Manos, 2005; Wei et al., 2012).该属在我国主要分布于华东、华中、华南及西南等地, 常生于海拔500-1 500 m的山谷林下及村边路旁.据《中国植物志》记载, 分布于我国的酸浆属植物主要有5种和2个变种.大多数酸浆属植物具有抗氧化、抗炎及抗癌等多种药理活性(Ding et al., 2014; 杨金颖等, 2014; Zhang and Tong, 2016), 其果、根或全草可入药, 常用来治疗疟疾、风湿、肝炎、哮喘及癌症等疾病(许亮等, 2009; 国家药典委员会, 2015; Xu et al., 2016).其中, 酸浆(P. alkekengi var. francheti), 又名锦灯笼、挂金灯, 被《中华人民共和国药典》(2015年版)收录.另外, 苦蘵(P. angulata)、小酸浆(P. minima)和毛酸浆(P. pubescens)等在很多中药材地方志中也都有记载.除此之外, 大部分酸浆属植物富含多种维生素, 可以用来做罐头、饮料、蜜饯或直接食用的“本草水果”(孙海涛和高玉超, 2013; 王晓英等, 2014; 郭瑜等, 2017; 王赢等, 2017). ...
中国酸浆属植物药用资源研究 1 2009
... 酸浆属(Physalis)是茄科(Solanaceae)中一个非常重要的属, 为一年生或多年生草本植物.全球酸浆属植物共约75-120种, 大多分布于美洲热带及温带地区, 少数分布于欧亚大陆及东南亚(中国科学院中国植物志编辑委员会, 1978; Maggie and Manos, 2005; Wei et al., 2012).该属在我国主要分布于华东、华中、华南及西南等地, 常生于海拔500-1 500 m的山谷林下及村边路旁.据《中国植物志》记载, 分布于我国的酸浆属植物主要有5种和2个变种.大多数酸浆属植物具有抗氧化、抗炎及抗癌等多种药理活性(Ding et al., 2014; 杨金颖等, 2014; Zhang and Tong, 2016), 其果、根或全草可入药, 常用来治疗疟疾、风湿、肝炎、哮喘及癌症等疾病(许亮等, 2009; 国家药典委员会, 2015; Xu et al., 2016).其中, 酸浆(P. alkekengi var. francheti), 又名锦灯笼、挂金灯, 被《中华人民共和国药典》(2015年版)收录.另外, 苦蘵(P. angulata)、小酸浆(P. minima)和毛酸浆(P. pubescens)等在很多中药材地方志中也都有记载.除此之外, 大部分酸浆属植物富含多种维生素, 可以用来做罐头、饮料、蜜饯或直接食用的“本草水果”(孙海涛和高玉超, 2013; 王晓英等, 2014; 郭瑜等, 2017; 王赢等, 2017). ...
酸浆的药理作用研究进展 1 2014
... 酸浆属(Physalis)是茄科(Solanaceae)中一个非常重要的属, 为一年生或多年生草本植物.全球酸浆属植物共约75-120种, 大多分布于美洲热带及温带地区, 少数分布于欧亚大陆及东南亚(中国科学院中国植物志编辑委员会, 1978; Maggie and Manos, 2005; Wei et al., 2012).该属在我国主要分布于华东、华中、华南及西南等地, 常生于海拔500-1 500 m的山谷林下及村边路旁.据《中国植物志》记载, 分布于我国的酸浆属植物主要有5种和2个变种.大多数酸浆属植物具有抗氧化、抗炎及抗癌等多种药理活性(Ding et al., 2014; 杨金颖等, 2014; Zhang and Tong, 2016), 其果、根或全草可入药, 常用来治疗疟疾、风湿、肝炎、哮喘及癌症等疾病(许亮等, 2009; 国家药典委员会, 2015; Xu et al., 2016).其中, 酸浆(P. alkekengi var. francheti), 又名锦灯笼、挂金灯, 被《中华人民共和国药典》(2015年版)收录.另外, 苦蘵(P. angulata)、小酸浆(P. minima)和毛酸浆(P. pubescens)等在很多中药材地方志中也都有记载.除此之外, 大部分酸浆属植物富含多种维生素, 可以用来做罐头、饮料、蜜饯或直接食用的“本草水果”(孙海涛和高玉超, 2013; 王晓英等, 2014; 郭瑜等, 2017; 王赢等, 2017). ...
中国科学院中国植物志编辑委员会 0 1978
2 1996
... 准确评价不同植物物种之间的亲缘关系是植物分子育种和资源保护的重要基础.由于酸浆属植物种间的形态学性状非常相似, 多年来酸浆属植物的鉴定和分类被认为是茄科植物中非常困难的科学挑战(Maggie and Manos, 2005; Wei et al., 2012; Feng et al., 2016b).本研究中, 供试的4种酸浆属植物分别属于2个亚属(Maggie and Manos, 2005; Feng et al., 2016b).其中, 苦蘵和毛酸浆分别属于Rydbergis亚属的Angulatae组和Epeteiorhiza组, 小酸浆同样属于Rydbergis亚属, 但是没有组级别报道; 与其它3种酸浆属植物不同, 酸浆属于Physalis亚属.从UPGMA聚类和PCoA分析结果来看, 来自同一种酸浆属植物的所有样本被聚类在一起.供试酸浆属样品分为2大类(I和II), 其中, 来自Rydbergis亚属的苦蘵、毛酸浆和小酸浆被归为I类; 来自Physalis亚属的酸浆则远离其它酸浆属植物, 单独聚类为II类.我们之前的研究结果表明, 许多酸浆属植物在系统演化过程中的表现并不是单源进化.此外, 我们的研究同样发现酸浆与其它酸浆属植物的遗传关系较远(Feng et al., 2016b, 2018).Maggie和Manos (2005)研究认为, 酸浆是一种非典型的酸浆属植物, 应该单独列为一个亚属.依据Axelius (1996)关于酸浆属植物的形态学分类方法, 本研究中的苦蘵、毛酸浆和小酸浆属于Rydbergis亚属, 而酸浆属于Physalis亚属.酸浆是目前唯一被《中华人民共和国药典》(2015版)收录的酸浆属药用植物, 其分类归属问题一直以来是分类学家关注的焦点.本研究则在很大程度上与Axelius (1996)以及Maggie和Manos (2005)的研究结论一致, 表明SSR分子标记在酸浆属植物亲缘关系评价中具有较好的应用性. ... ... 亚属.酸浆是目前唯一被《中华人民共和国药典》(2015版)收录的酸浆属药用植物, 其分类归属问题一直以来是分类学家关注的焦点.本研究则在很大程度上与Axelius (1996)以及Maggie和Manos (2005)的研究结论一致, 表明SSR分子标记在酸浆属植物亲缘关系评价中具有较好的应用性. ...
2 2014
... 酸浆属(Physalis)是茄科(Solanaceae)中一个非常重要的属, 为一年生或多年生草本植物.全球酸浆属植物共约75-120种, 大多分布于美洲热带及温带地区, 少数分布于欧亚大陆及东南亚(中国科学院中国植物志编辑委员会, 1978; Maggie and Manos, 2005; Wei et al., 2012).该属在我国主要分布于华东、华中、华南及西南等地, 常生于海拔500-1 500 m的山谷林下及村边路旁.据《中国植物志》记载, 分布于我国的酸浆属植物主要有5种和2个变种.大多数酸浆属植物具有抗氧化、抗炎及抗癌等多种药理活性(Ding et al., 2014; 杨金颖等, 2014; Zhang and Tong, 2016), 其果、根或全草可入药, 常用来治疗疟疾、风湿、肝炎、哮喘及癌症等疾病(许亮等, 2009; 国家药典委员会, 2015; Xu et al., 2016).其中, 酸浆(P. alkekengi var. francheti), 又名锦灯笼、挂金灯, 被《中华人民共和国药典》(2015年版)收录.另外, 苦蘵(P. angulata)、小酸浆(P. minima)和毛酸浆(P. pubescens)等在很多中药材地方志中也都有记载.除此之外, 大部分酸浆属植物富含多种维生素, 可以用来做罐头、饮料、蜜饯或直接食用的“本草水果”(孙海涛和高玉超, 2013; 王晓英等, 2014; 郭瑜等, 2017; 王赢等, 2017). ... ... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ...
2 2016
... SSR分子标记技术是基于简单重复序列(simple sequence repeats, SSR)开发的分子标记.根据简单重复序列两侧的保守序列设计引物, 通过检测重复数目的差异反映等位基因间的多态性.由于SSR分子标记在真核生物基因组中分布广泛、数量丰富、类型多样且具有稳定性好、操作简便及共显性等优点, 目前该技术被认为是进行植物遗传研究最理想的分子标记之一(Feng et al., 2016a).本研究采用SSR分子标记对4种酸浆属植物的22份材料进行亲缘关系分析, 为酸浆属植物的有效利用、资源保护及种质创新奠定重要基础. ... ... 与RAPD和ISSR等表现为显性类型的分子标记相比, SSR标记是共显性分子标记, 因此, 更能准确地揭示物种之间的亲缘关系.近年来, SSR分子标记已经成功应用于许多植物的亲缘关系研究(Wei et al., 2012; Teshome et al., 2015; Wang et al., 2015; Feng et al., 2016a).Simbaqueba等(2011)基于秘鲁苦蘵(P. peruviana)的转录组数据开发出一批可用于秘鲁苦蘵分子鉴定及遗传多样性研究的SSR标记.本研究表明, 66.7% (20/30)的秘鲁苦蘵SSR标记可以有效用于苦蘵、小酸浆、酸浆及毛酸浆的遗传关系研究(表2).另有结果显示, 90.7% (107/118)的SSR扩增条带是多态性条带, 高于甘草属(Glycyrrhiza)植物中SSR分子标记的多态性(89.44%) (李晓岚等, 2015), 以及芹菜(Apium graveolens)栽培品种中的SSR多态性比例(53.85%) (Fu et al., 2014).由此可见, 本研究筛选出来的20对SSR分子标记引物在酸浆属内具有很好的适用性, 可以用于该属植物之间的亲缘关系研究. ...
4 2016
... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ... ... 准确评价不同植物物种之间的亲缘关系是植物分子育种和资源保护的重要基础.由于酸浆属植物种间的形态学性状非常相似, 多年来酸浆属植物的鉴定和分类被认为是茄科植物中非常困难的科学挑战(Maggie and Manos, 2005; Wei et al., 2012; Feng et al., 2016b).本研究中, 供试的4种酸浆属植物分别属于2个亚属(Maggie and Manos, 2005; Feng et al., 2016b).其中, 苦蘵和毛酸浆分别属于Rydbergis亚属的Angulatae组和Epeteiorhiza组, 小酸浆同样属于Rydbergis亚属, 但是没有组级别报道; 与其它3种酸浆属植物不同, 酸浆属于Physalis亚属.从UPGMA聚类和PCoA分析结果来看, 来自同一种酸浆属植物的所有样本被聚类在一起.供试酸浆属样品分为2大类(I和II), 其中, 来自Rydbergis亚属的苦蘵、毛酸浆和小酸浆被归为I类; 来自Physalis亚属的酸浆则远离其它酸浆属植物, 单独聚类为II类.我们之前的研究结果表明, 许多酸浆属植物在系统演化过程中的表现并不是单源进化.此外, 我们的研究同样发现酸浆与其它酸浆属植物的遗传关系较远(Feng et al., 2016b, 2018).Maggie和Manos (2005)研究认为, 酸浆是一种非典型的酸浆属植物, 应该单独列为一个亚属.依据Axelius (1996)关于酸浆属植物的形态学分类方法, 本研究中的苦蘵、毛酸浆和小酸浆属于Rydbergis亚属, 而酸浆属于Physalis亚属.酸浆是目前唯一被《中华人民共和国药典》(2015版)收录的酸浆属药用植物, 其分类归属问题一直以来是分类学家关注的焦点.本研究则在很大程度上与Axelius (1996)以及Maggie和Manos (2005)的研究结论一致, 表明SSR分子标记在酸浆属植物亲缘关系评价中具有较好的应用性. ... ... ; Feng et al., 2016b).其中, 苦蘵和毛酸浆分别属于Rydbergis亚属的Angulatae组和Epeteiorhiza组, 小酸浆同样属于Rydbergis亚属, 但是没有组级别报道; 与其它3种酸浆属植物不同, 酸浆属于Physalis亚属.从UPGMA聚类和PCoA分析结果来看, 来自同一种酸浆属植物的所有样本被聚类在一起.供试酸浆属样品分为2大类(I和II), 其中, 来自Rydbergis亚属的苦蘵、毛酸浆和小酸浆被归为I类; 来自Physalis亚属的酸浆则远离其它酸浆属植物, 单独聚类为II类.我们之前的研究结果表明, 许多酸浆属植物在系统演化过程中的表现并不是单源进化.此外, 我们的研究同样发现酸浆与其它酸浆属植物的遗传关系较远(Feng et al., 2016b, 2018).Maggie和Manos (2005)研究认为, 酸浆是一种非典型的酸浆属植物, 应该单独列为一个亚属.依据Axelius (1996)关于酸浆属植物的形态学分类方法, 本研究中的苦蘵、毛酸浆和小酸浆属于Rydbergis亚属, 而酸浆属于Physalis亚属.酸浆是目前唯一被《中华人民共和国药典》(2015版)收录的酸浆属药用植物, 其分类归属问题一直以来是分类学家关注的焦点.本研究则在很大程度上与Axelius (1996)以及Maggie和Manos (2005)的研究结论一致, 表明SSR分子标记在酸浆属植物亲缘关系评价中具有较好的应用性. ... ... 亚属的酸浆则远离其它酸浆属植物, 单独聚类为II类.我们之前的研究结果表明, 许多酸浆属植物在系统演化过程中的表现并不是单源进化.此外, 我们的研究同样发现酸浆与其它酸浆属植物的遗传关系较远(Feng et al., 2016b, 2018).Maggie和Manos (2005)研究认为, 酸浆是一种非典型的酸浆属植物, 应该单独列为一个亚属.依据Axelius (1996)关于酸浆属植物的形态学分类方法, 本研究中的苦蘵、毛酸浆和小酸浆属于Rydbergis亚属, 而酸浆属于Physalis亚属.酸浆是目前唯一被《中华人民共和国药典》(2015版)收录的酸浆属药用植物, 其分类归属问题一直以来是分类学家关注的焦点.本研究则在很大程度上与Axelius (1996)以及Maggie和Manos (2005)的研究结论一致, 表明SSR分子标记在酸浆属植物亲缘关系评价中具有较好的应用性. ...
2 2018
... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ... ... 准确评价不同植物物种之间的亲缘关系是植物分子育种和资源保护的重要基础.由于酸浆属植物种间的形态学性状非常相似, 多年来酸浆属植物的鉴定和分类被认为是茄科植物中非常困难的科学挑战(Maggie and Manos, 2005; Wei et al., 2012; Feng et al., 2016b).本研究中, 供试的4种酸浆属植物分别属于2个亚属(Maggie and Manos, 2005; Feng et al., 2016b).其中, 苦蘵和毛酸浆分别属于Rydbergis亚属的Angulatae组和Epeteiorhiza组, 小酸浆同样属于Rydbergis亚属, 但是没有组级别报道; 与其它3种酸浆属植物不同, 酸浆属于Physalis亚属.从UPGMA聚类和PCoA分析结果来看, 来自同一种酸浆属植物的所有样本被聚类在一起.供试酸浆属样品分为2大类(I和II), 其中, 来自Rydbergis亚属的苦蘵、毛酸浆和小酸浆被归为I类; 来自Physalis亚属的酸浆则远离其它酸浆属植物, 单独聚类为II类.我们之前的研究结果表明, 许多酸浆属植物在系统演化过程中的表现并不是单源进化.此外, 我们的研究同样发现酸浆与其它酸浆属植物的遗传关系较远(Feng et al., 2016b, 2018).Maggie和Manos (2005)研究认为, 酸浆是一种非典型的酸浆属植物, 应该单独列为一个亚属.依据Axelius (1996)关于酸浆属植物的形态学分类方法, 本研究中的苦蘵、毛酸浆和小酸浆属于Rydbergis亚属, 而酸浆属于Physalis亚属.酸浆是目前唯一被《中华人民共和国药典》(2015版)收录的酸浆属药用植物, 其分类归属问题一直以来是分类学家关注的焦点.本研究则在很大程度上与Axelius (1996)以及Maggie和Manos (2005)的研究结论一致, 表明SSR分子标记在酸浆属植物亲缘关系评价中具有较好的应用性. ...
1 2014
... 与RAPD和ISSR等表现为显性类型的分子标记相比, SSR标记是共显性分子标记, 因此, 更能准确地揭示物种之间的亲缘关系.近年来, SSR分子标记已经成功应用于许多植物的亲缘关系研究(Wei et al., 2012; Teshome et al., 2015; Wang et al., 2015; Feng et al., 2016a).Simbaqueba等(2011)基于秘鲁苦蘵(P. peruviana)的转录组数据开发出一批可用于秘鲁苦蘵分子鉴定及遗传多样性研究的SSR标记.本研究表明, 66.7% (20/30)的秘鲁苦蘵SSR标记可以有效用于苦蘵、小酸浆、酸浆及毛酸浆的遗传关系研究(表2).另有结果显示, 90.7% (107/118)的SSR扩增条带是多态性条带, 高于甘草属(Glycyrrhiza)植物中SSR分子标记的多态性(89.44%) (李晓岚等, 2015), 以及芹菜(Apium graveolens)栽培品种中的SSR多态性比例(53.85%) (Fu et al., 2014).由此可见, 本研究筛选出来的20对SSR分子标记引物在酸浆属内具有很好的适用性, 可以用于该属植物之间的亲缘关系研究. ...
1 2015
... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ...
1 1966
... 利用Quantity one软件(Version 4.6.2)及人工辅助判读的方法进行电泳图拷带.为了保证结果的准确性, 每对引物进行2次重复PCR扩增和电泳检测, 并只统计清晰度高且稳定性好的电泳位点.同一位点有条带记作1, 无条带记作0.将整理好的0/1原始矩阵数据输入NTSYS-pc 2.10e软件(Rohlf, 2000), 计算样品间的遗传相似系数(simple matching coefficie, SM) (Nei and Li, 1979), 并基于非加权组平均法(unweighted pair group method with an arithmetic mean, UPGMA)进行聚类分析.另外, 运用NTSYSpc 2.10e软件中的Dcenter及Eigen程序进行主坐标分析(principal coordinate analysis, PCoA) (Gower, 1966). ...
1 2013
... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ...
1 2014
... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ...
5 2005
... 酸浆属(Physalis)是茄科(Solanaceae)中一个非常重要的属, 为一年生或多年生草本植物.全球酸浆属植物共约75-120种, 大多分布于美洲热带及温带地区, 少数分布于欧亚大陆及东南亚(中国科学院中国植物志编辑委员会, 1978; Maggie and Manos, 2005; Wei et al., 2012).该属在我国主要分布于华东、华中、华南及西南等地, 常生于海拔500-1 500 m的山谷林下及村边路旁.据《中国植物志》记载, 分布于我国的酸浆属植物主要有5种和2个变种.大多数酸浆属植物具有抗氧化、抗炎及抗癌等多种药理活性(Ding et al., 2014; 杨金颖等, 2014; Zhang and Tong, 2016), 其果、根或全草可入药, 常用来治疗疟疾、风湿、肝炎、哮喘及癌症等疾病(许亮等, 2009; 国家药典委员会, 2015; Xu et al., 2016).其中, 酸浆(P. alkekengi var. francheti), 又名锦灯笼、挂金灯, 被《中华人民共和国药典》(2015年版)收录.另外, 苦蘵(P. angulata)、小酸浆(P. minima)和毛酸浆(P. pubescens)等在很多中药材地方志中也都有记载.除此之外, 大部分酸浆属植物富含多种维生素, 可以用来做罐头、饮料、蜜饯或直接食用的“本草水果”(孙海涛和高玉超, 2013; 王晓英等, 2014; 郭瑜等, 2017; 王赢等, 2017). ... ... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ... ... 准确评价不同植物物种之间的亲缘关系是植物分子育种和资源保护的重要基础.由于酸浆属植物种间的形态学性状非常相似, 多年来酸浆属植物的鉴定和分类被认为是茄科植物中非常困难的科学挑战(Maggie and Manos, 2005; Wei et al., 2012; Feng et al., 2016b).本研究中, 供试的4种酸浆属植物分别属于2个亚属(Maggie and Manos, 2005; Feng et al., 2016b).其中, 苦蘵和毛酸浆分别属于Rydbergis亚属的Angulatae组和Epeteiorhiza组, 小酸浆同样属于Rydbergis亚属, 但是没有组级别报道; 与其它3种酸浆属植物不同, 酸浆属于Physalis亚属.从UPGMA聚类和PCoA分析结果来看, 来自同一种酸浆属植物的所有样本被聚类在一起.供试酸浆属样品分为2大类(I和II), 其中, 来自Rydbergis亚属的苦蘵、毛酸浆和小酸浆被归为I类; 来自Physalis亚属的酸浆则远离其它酸浆属植物, 单独聚类为II类.我们之前的研究结果表明, 许多酸浆属植物在系统演化过程中的表现并不是单源进化.此外, 我们的研究同样发现酸浆与其它酸浆属植物的遗传关系较远(Feng et al., 2016b, 2018).Maggie和Manos (2005)研究认为, 酸浆是一种非典型的酸浆属植物, 应该单独列为一个亚属.依据Axelius (1996)关于酸浆属植物的形态学分类方法, 本研究中的苦蘵、毛酸浆和小酸浆属于Rydbergis亚属, 而酸浆属于Physalis亚属.酸浆是目前唯一被《中华人民共和国药典》(2015版)收录的酸浆属药用植物, 其分类归属问题一直以来是分类学家关注的焦点.本研究则在很大程度上与Axelius (1996)以及Maggie和Manos (2005)的研究结论一致, 表明SSR分子标记在酸浆属植物亲缘关系评价中具有较好的应用性. ... ... ).本研究中, 供试的4种酸浆属植物分别属于2个亚属(Maggie and Manos, 2005; Feng et al., 2016b).其中, 苦蘵和毛酸浆分别属于Rydbergis亚属的Angulatae组和Epeteiorhiza组, 小酸浆同样属于Rydbergis亚属, 但是没有组级别报道; 与其它3种酸浆属植物不同, 酸浆属于Physalis亚属.从UPGMA聚类和PCoA分析结果来看, 来自同一种酸浆属植物的所有样本被聚类在一起.供试酸浆属样品分为2大类(I和II), 其中, 来自Rydbergis亚属的苦蘵、毛酸浆和小酸浆被归为I类; 来自Physalis亚属的酸浆则远离其它酸浆属植物, 单独聚类为II类.我们之前的研究结果表明, 许多酸浆属植物在系统演化过程中的表现并不是单源进化.此外, 我们的研究同样发现酸浆与其它酸浆属植物的遗传关系较远(Feng et al., 2016b, 2018).Maggie和Manos (2005)研究认为, 酸浆是一种非典型的酸浆属植物, 应该单独列为一个亚属.依据Axelius (1996)关于酸浆属植物的形态学分类方法, 本研究中的苦蘵、毛酸浆和小酸浆属于Rydbergis亚属, 而酸浆属于Physalis亚属.酸浆是目前唯一被《中华人民共和国药典》(2015版)收录的酸浆属药用植物, 其分类归属问题一直以来是分类学家关注的焦点.本研究则在很大程度上与Axelius (1996)以及Maggie和Manos (2005)的研究结论一致, 表明SSR分子标记在酸浆属植物亲缘关系评价中具有较好的应用性. ... ... 以及Maggie和Manos (2005)的研究结论一致, 表明SSR分子标记在酸浆属植物亲缘关系评价中具有较好的应用性. ...
1 1979
... 利用Quantity one软件(Version 4.6.2)及人工辅助判读的方法进行电泳图拷带.为了保证结果的准确性, 每对引物进行2次重复PCR扩增和电泳检测, 并只统计清晰度高且稳定性好的电泳位点.同一位点有条带记作1, 无条带记作0.将整理好的0/1原始矩阵数据输入NTSYS-pc 2.10e软件(Rohlf, 2000), 计算样品间的遗传相似系数(simple matching coefficie, SM) (Nei and Li, 1979), 并基于非加权组平均法(unweighted pair group method with an arithmetic mean, UPGMA)进行聚类分析.另外, 运用NTSYSpc 2.10e软件中的Dcenter及Eigen程序进行主坐标分析(principal coordinate analysis, PCoA) (Gower, 1966). ...
1 2008
... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ...
1 2000
... 利用Quantity one软件(Version 4.6.2)及人工辅助判读的方法进行电泳图拷带.为了保证结果的准确性, 每对引物进行2次重复PCR扩增和电泳检测, 并只统计清晰度高且稳定性好的电泳位点.同一位点有条带记作1, 无条带记作0.将整理好的0/1原始矩阵数据输入NTSYS-pc 2.10e软件(Rohlf, 2000), 计算样品间的遗传相似系数(simple matching coefficie, SM) (Nei and Li, 1979), 并基于非加权组平均法(unweighted pair group method with an arithmetic mean, UPGMA)进行聚类分析.另外, 运用NTSYSpc 2.10e软件中的Dcenter及Eigen程序进行主坐标分析(principal coordinate analysis, PCoA) (Gower, 1966). ...
2 2011
... 参考Simbaqueba等(2011)的研究方法, 共合成SSR引物30对.通过引物筛选, 选出20对多态性高、稳定性好且扩增条带清晰的引物进行后续实验. ... ... 与RAPD和ISSR等表现为显性类型的分子标记相比, SSR标记是共显性分子标记, 因此, 更能准确地揭示物种之间的亲缘关系.近年来, SSR分子标记已经成功应用于许多植物的亲缘关系研究(Wei et al., 2012; Teshome et al., 2015; Wang et al., 2015; Feng et al., 2016a).Simbaqueba等(2011)基于秘鲁苦蘵(P. peruviana)的转录组数据开发出一批可用于秘鲁苦蘵分子鉴定及遗传多样性研究的SSR标记.本研究表明, 66.7% (20/30)的秘鲁苦蘵SSR标记可以有效用于苦蘵、小酸浆、酸浆及毛酸浆的遗传关系研究(表2).另有结果显示, 90.7% (107/118)的SSR扩增条带是多态性条带, 高于甘草属(Glycyrrhiza)植物中SSR分子标记的多态性(89.44%) (李晓岚等, 2015), 以及芹菜(Apium graveolens)栽培品种中的SSR多态性比例(53.85%) (Fu et al., 2014).由此可见, 本研究筛选出来的20对SSR分子标记引物在酸浆属内具有很好的适用性, 可以用于该属植物之间的亲缘关系研究. ...
1 2017
... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ...
1 2015
... 与RAPD和ISSR等表现为显性类型的分子标记相比, SSR标记是共显性分子标记, 因此, 更能准确地揭示物种之间的亲缘关系.近年来, SSR分子标记已经成功应用于许多植物的亲缘关系研究(Wei et al., 2012; Teshome et al., 2015; Wang et al., 2015; Feng et al., 2016a).Simbaqueba等(2011)基于秘鲁苦蘵(P. peruviana)的转录组数据开发出一批可用于秘鲁苦蘵分子鉴定及遗传多样性研究的SSR标记.本研究表明, 66.7% (20/30)的秘鲁苦蘵SSR标记可以有效用于苦蘵、小酸浆、酸浆及毛酸浆的遗传关系研究(表2).另有结果显示, 90.7% (107/118)的SSR扩增条带是多态性条带, 高于甘草属(Glycyrrhiza)植物中SSR分子标记的多态性(89.44%) (李晓岚等, 2015), 以及芹菜(Apium graveolens)栽培品种中的SSR多态性比例(53.85%) (Fu et al., 2014).由此可见, 本研究筛选出来的20对SSR分子标记引物在酸浆属内具有很好的适用性, 可以用于该属植物之间的亲缘关系研究. ...
1 2015
... 与RAPD和ISSR等表现为显性类型的分子标记相比, SSR标记是共显性分子标记, 因此, 更能准确地揭示物种之间的亲缘关系.近年来, SSR分子标记已经成功应用于许多植物的亲缘关系研究(Wei et al., 2012; Teshome et al., 2015; Wang et al., 2015; Feng et al., 2016a).Simbaqueba等(2011)基于秘鲁苦蘵(P. peruviana)的转录组数据开发出一批可用于秘鲁苦蘵分子鉴定及遗传多样性研究的SSR标记.本研究表明, 66.7% (20/30)的秘鲁苦蘵SSR标记可以有效用于苦蘵、小酸浆、酸浆及毛酸浆的遗传关系研究(表2).另有结果显示, 90.7% (107/118)的SSR扩增条带是多态性条带, 高于甘草属(Glycyrrhiza)植物中SSR分子标记的多态性(89.44%) (李晓岚等, 2015), 以及芹菜(Apium graveolens)栽培品种中的SSR多态性比例(53.85%) (Fu et al., 2014).由此可见, 本研究筛选出来的20对SSR分子标记引物在酸浆属内具有很好的适用性, 可以用于该属植物之间的亲缘关系研究. ...
3 2012
... 酸浆属(Physalis)是茄科(Solanaceae)中一个非常重要的属, 为一年生或多年生草本植物.全球酸浆属植物共约75-120种, 大多分布于美洲热带及温带地区, 少数分布于欧亚大陆及东南亚(中国科学院中国植物志编辑委员会, 1978; Maggie and Manos, 2005; Wei et al., 2012).该属在我国主要分布于华东、华中、华南及西南等地, 常生于海拔500-1 500 m的山谷林下及村边路旁.据《中国植物志》记载, 分布于我国的酸浆属植物主要有5种和2个变种.大多数酸浆属植物具有抗氧化、抗炎及抗癌等多种药理活性(Ding et al., 2014; 杨金颖等, 2014; Zhang and Tong, 2016), 其果、根或全草可入药, 常用来治疗疟疾、风湿、肝炎、哮喘及癌症等疾病(许亮等, 2009; 国家药典委员会, 2015; Xu et al., 2016).其中, 酸浆(P. alkekengi var. francheti), 又名锦灯笼、挂金灯, 被《中华人民共和国药典》(2015年版)收录.另外, 苦蘵(P. angulata)、小酸浆(P. minima)和毛酸浆(P. pubescens)等在很多中药材地方志中也都有记载.除此之外, 大部分酸浆属植物富含多种维生素, 可以用来做罐头、饮料、蜜饯或直接食用的“本草水果”(孙海涛和高玉超, 2013; 王晓英等, 2014; 郭瑜等, 2017; 王赢等, 2017). ... ... 与RAPD和ISSR等表现为显性类型的分子标记相比, SSR标记是共显性分子标记, 因此, 更能准确地揭示物种之间的亲缘关系.近年来, SSR分子标记已经成功应用于许多植物的亲缘关系研究(Wei et al., 2012; Teshome et al., 2015; Wang et al., 2015; Feng et al., 2016a).Simbaqueba等(2011)基于秘鲁苦蘵(P. peruviana)的转录组数据开发出一批可用于秘鲁苦蘵分子鉴定及遗传多样性研究的SSR标记.本研究表明, 66.7% (20/30)的秘鲁苦蘵SSR标记可以有效用于苦蘵、小酸浆、酸浆及毛酸浆的遗传关系研究(表2).另有结果显示, 90.7% (107/118)的SSR扩增条带是多态性条带, 高于甘草属(Glycyrrhiza)植物中SSR分子标记的多态性(89.44%) (李晓岚等, 2015), 以及芹菜(Apium graveolens)栽培品种中的SSR多态性比例(53.85%) (Fu et al., 2014).由此可见, 本研究筛选出来的20对SSR分子标记引物在酸浆属内具有很好的适用性, 可以用于该属植物之间的亲缘关系研究. ... ... 准确评价不同植物物种之间的亲缘关系是植物分子育种和资源保护的重要基础.由于酸浆属植物种间的形态学性状非常相似, 多年来酸浆属植物的鉴定和分类被认为是茄科植物中非常困难的科学挑战(Maggie and Manos, 2005; Wei et al., 2012; Feng et al., 2016b).本研究中, 供试的4种酸浆属植物分别属于2个亚属(Maggie and Manos, 2005; Feng et al., 2016b).其中, 苦蘵和毛酸浆分别属于Rydbergis亚属的Angulatae组和Epeteiorhiza组, 小酸浆同样属于Rydbergis亚属, 但是没有组级别报道; 与其它3种酸浆属植物不同, 酸浆属于Physalis亚属.从UPGMA聚类和PCoA分析结果来看, 来自同一种酸浆属植物的所有样本被聚类在一起.供试酸浆属样品分为2大类(I和II), 其中, 来自Rydbergis亚属的苦蘵、毛酸浆和小酸浆被归为I类; 来自Physalis亚属的酸浆则远离其它酸浆属植物, 单独聚类为II类.我们之前的研究结果表明, 许多酸浆属植物在系统演化过程中的表现并不是单源进化.此外, 我们的研究同样发现酸浆与其它酸浆属植物的遗传关系较远(Feng et al., 2016b, 2018).Maggie和Manos (2005)研究认为, 酸浆是一种非典型的酸浆属植物, 应该单独列为一个亚属.依据Axelius (1996)关于酸浆属植物的形态学分类方法, 本研究中的苦蘵、毛酸浆和小酸浆属于Rydbergis亚属, 而酸浆属于Physalis亚属.酸浆是目前唯一被《中华人民共和国药典》(2015版)收录的酸浆属药用植物, 其分类归属问题一直以来是分类学家关注的焦点.本研究则在很大程度上与Axelius (1996)以及Maggie和Manos (2005)的研究结论一致, 表明SSR分子标记在酸浆属植物亲缘关系评价中具有较好的应用性. ...
1 2016
... 酸浆属(Physalis)是茄科(Solanaceae)中一个非常重要的属, 为一年生或多年生草本植物.全球酸浆属植物共约75-120种, 大多分布于美洲热带及温带地区, 少数分布于欧亚大陆及东南亚(中国科学院中国植物志编辑委员会, 1978; Maggie and Manos, 2005; Wei et al., 2012).该属在我国主要分布于华东、华中、华南及西南等地, 常生于海拔500-1 500 m的山谷林下及村边路旁.据《中国植物志》记载, 分布于我国的酸浆属植物主要有5种和2个变种.大多数酸浆属植物具有抗氧化、抗炎及抗癌等多种药理活性(Ding et al., 2014; 杨金颖等, 2014; Zhang and Tong, 2016), 其果、根或全草可入药, 常用来治疗疟疾、风湿、肝炎、哮喘及癌症等疾病(许亮等, 2009; 国家药典委员会, 2015; Xu et al., 2016).其中, 酸浆(P. alkekengi var. francheti), 又名锦灯笼、挂金灯, 被《中华人民共和国药典》(2015年版)收录.另外, 苦蘵(P. angulata)、小酸浆(P. minima)和毛酸浆(P. pubescens)等在很多中药材地方志中也都有记载.除此之外, 大部分酸浆属植物富含多种维生素, 可以用来做罐头、饮料、蜜饯或直接食用的“本草水果”(孙海涛和高玉超, 2013; 王晓英等, 2014; 郭瑜等, 2017; 王赢等, 2017). ...
1 2016
... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ...
1 2015
... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ...
2 2016
... 酸浆属(Physalis)是茄科(Solanaceae)中一个非常重要的属, 为一年生或多年生草本植物.全球酸浆属植物共约75-120种, 大多分布于美洲热带及温带地区, 少数分布于欧亚大陆及东南亚(中国科学院中国植物志编辑委员会, 1978; Maggie and Manos, 2005; Wei et al., 2012).该属在我国主要分布于华东、华中、华南及西南等地, 常生于海拔500-1 500 m的山谷林下及村边路旁.据《中国植物志》记载, 分布于我国的酸浆属植物主要有5种和2个变种.大多数酸浆属植物具有抗氧化、抗炎及抗癌等多种药理活性(Ding et al., 2014; 杨金颖等, 2014; Zhang and Tong, 2016), 其果、根或全草可入药, 常用来治疗疟疾、风湿、肝炎、哮喘及癌症等疾病(许亮等, 2009; 国家药典委员会, 2015; Xu et al., 2016).其中, 酸浆(P. alkekengi var. francheti), 又名锦灯笼、挂金灯, 被《中华人民共和国药典》(2015年版)收录.另外, 苦蘵(P. angulata)、小酸浆(P. minima)和毛酸浆(P. pubescens)等在很多中药材地方志中也都有记载.除此之外, 大部分酸浆属植物富含多种维生素, 可以用来做罐头、饮料、蜜饯或直接食用的“本草水果”(孙海涛和高玉超, 2013; 王晓英等, 2014; 郭瑜等, 2017; 王赢等, 2017). ... ... 酸浆属植物的遗传关系评价是开展酸浆属植物品种选育及资源保护的重要基础工作, DNA分子标记技术在其中扮演了重要角色.虽然近年来酸浆属植物的大部分研究主要集中在植物化学和药理特性方面(Ji et al., 2013; 贾远敏等, 2013; Ding et al., 2014; Li et al., 2014; 骆丽萍等, 2015; Yang et al., 2016; Zhang and Tong, 2016; Sun et al., 2017), 但还有一些DNA分子标记技术, 如ISSR、SNPs、ITS、ndhF、trnLF和psbA-trnH等已经应用于酸浆属植物的遗传多样性分析、系统发育评价以及种质资源鉴定等(Maggie and Manos, 2005; Olmstead et al., 2008; Garzon-Martínez et al., 2015; Zamora-Tavares et al., 2015; Feng et al., 2016b, 2018). ...
 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}