韩丹璐, 赖建彬, 阳成伟*, 华南师范大学生命科学学院, 广东省植物发育生物工程重点实验室, 广州 510631 HanDanlu, LaiJianbin, YangChengwei*, Guangdong Provincial Key Laboratory of Biotechnology for Plant Development, College of Life Sciences, South China Normal University, Guangzhou 510631, China 引用本文 韩丹璐, 赖建彬, 阳成伟. SUMO E3连接酶在植物生长发育中的功能研究进展. 植物学报, 2018, 53(2): 175-184
Abstract: SUMOylation is an important type of protein posttranslational modification. SUMO E3 ligases are critical in this process via substrate recognition and reaction enhancement. Several SUMO E3 ligases, involved in development regulation, have been identified in plants. Here, we summarize the research advances in the functions of SUMO E3 ligases in root development, flowering, gametogenesis and photomorphogenesis and prospects for further study.
Key words:MMS21 ; plant growth and development ; SIZ1 ; SUMO E3 ligases ; SUMOylation
1 植物SUMO E3连接酶的结构与功能SP-RING (Siz/Pias)家族是第一个被发现的SUMO E3连接酶家族, SIZ1和MMS21都属于这个家族, 它们的C端都具有1个高度保守的SP-RING结构域。这个结构域与泛素RING结构域相似, 但SP-RING结构域只有1个与锌离子成环的位点, 另1个位点则被氢键和范德华力所代替, 从而维持整个建构的稳定性(图1) (Johnson and Gupta, 2001; Yunus and Lima, 2009; Ishida et al., 2012)。植物中有2个SUMO E3连接酶, 其中MMS21较小, 只含有1个SP-RING结 构域; 而SIZ1除了包括SP-RING结构域之外, 还包含 4个保守的结构域: N端的SAP (scaffold attachment factor A/B/acinus/PIAS)结构域、PINIT (Pro-Ile-Asn-Ile- Thr)基序、SXS (Ser-X-Ser; X代表任意氨基酸)结构域以及一段植物特有的锌指结构域PHD (plant homeodomain) (Miura et al., 2007a; Cheong et al., 2009)。Cheong等(2009)分别突变AtSIZ1的结构域, 发现5个结构域行使不同的功能, 其中SP-RING结构域赋予AtSIZ1连接酶催化的功能, 决定AtSIZ1的亚细胞定位和控制水杨酸(SA)水平, 同时也调节AtSIZ1独立于SA信号途径的热胁迫响应; PHD结构域也参与连接酶催化底物连接的过程, 但并不是SUMO化所必需的结构, 其可能与PINIT结构域共同调控SUMO与糖和光响应中下胚轴伸长的信号途径; SXS基序可能参与SIZ1介导的ABA信号途径; SAP结构域在酵母中与DNA结合相关, 但在植物中的功能尚未明确。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-175/img_1.png<b>图1</b> 植物SUMO E3连接酶的保守结构域<br/>(A) 植物SUMO E3连接酶MMS21和SIZ1的特征结构域。SIZ1具有SAP、PHD、PINIT、SP-RING和SXS结构域, MMS21只具有SP-RING结构域(<xref ref-type="bibr" rid="b19-1674-3466-53-2-175">Ishida et al., 2012</xref>); (B) SUMO E3连接酶的SP-RING结构域和泛素E3连接酶的RING结构域构型。RING结构域有2个锌离子结合环, SP-RING只含有1个, 第2个环被范德华力和氢键所替代(<xref ref-type="bibr" rid="b14-1674-3466-53-2-175">Duan et al., 2009</xref>; <xref ref-type="bibr" rid="b49-1674-3466-53-2-175">Yunus and Lima, 2009</xref>; <xref ref-type="bibr" rid="b19-1674-3466-53-2-175">Ishida et al., 2012</xref>; <xref ref-type="bibr" rid="b3-1674-3466-53-2-175">石田喬志和杉本慶子, 2012</xref>); (C), (D) 分别为植物SIZ1和MMS21的同源序列比对。数据来源于NCBI。由DNAMAN定义的氨基酸序列、保守序列(黑色)和相似序列(灰色)。SIZ1的蛋白质氨基酸序列为拟南芥(<i>Arabidopsis thaliana</i>) AtSIZ1 (AAU00414.1)、大豆(<i>Glycine max</i>) GmSIZ1a (KRH24918.1)和GmSIZ1b (KRG89029.1)、水稻(<i>Oryza sativa</i>) OsSIZ1 (BAG97182.1)和OsSIZ2 (BAG89374.1)以及玉米(<i>Zea mays</i>) ZmSIZ1a (AQK95338.1)、ZmSIZ1b (AQK79048.1)和ZmSIZc (ONM41936.1)。MMS21的蛋白质氨基酸序列为拟南芥AtMMS21 (NP_188133.2)、大豆GmMMS21 (XP_003541835.1)、水稻OsMMS21 (XP_015640264.1)和玉米ZmMMS21 (AQK88509.1)。<br/><b>Figure 1</b> The domains are conserved in plant SUMO E3 ligases<br/>(A) Characteristic domains of plant SUMO E3 ligases. MMS21 and SIZ1 are shown by boxes, SIZ1 possesses SAP, PHD, PINIT, SP-RING and SXS domains, while MMS21 possesses only SP-RING domain (<xref ref-type="bibr" rid="b19-1674-3466-53-2-175">Ishida et al., 2012</xref>); (B) A schematic model of the SP-RING domain of SUMO E3 ligases and the RING finger of ubiquitin E3 ligases. The RING domain sports two zinc-coordinating loops, the SP-RING domain contains only one, the second loop is instead held together by hydrogen bonds and Van der Waals forces (<xref ref-type="bibr" rid="b14-1674-3466-53-2-175">Duan et al., 2009</xref>; <xref ref-type="bibr" rid="b49-1674-3466-53-2-175">Yunus and Lima, 2009</xref>; <xref ref-type="bibr" rid="b19-1674-3466-53-2-175">Ishida et al., 2012</xref>); (C), (D) The sequence data for the plant SIZ1 homologues were obtained from the NCBI protein database. Sequence identities (black boxes) and similarities (gray boxes) of amino acids were identified by DNAMAN. Amino acid sequences of SIZ1 proteins are from <i>Arabidopsis thaliana </i>AtSIZ1 (AAU00414.1), <i>Glycine max </i>GmSIZ1a (KRH24918.1), GmSIZ1b (KRG89029.1), <i>Oryza sativa </i>OsSIZ1 (BAG97182.1), OsSIZ2 (BAG89374.1), and <i>Zea mays</i> ZmSIZ1a (AQK95338.1), ZmSIZ1b (AQK79048.1), ZmSIZc (ONM41936.1). Amino acid sequences of MMS21 proteins are from <i>Arabidopsis thaliana </i>AtMMS21 (NP_188133.2),<i> Glycine max</i> GmMMS21 (XP_003541835.1), <i>Oryza sativa </i>OsMMS21 (XP_015640264.1), and<i> Zea mays</i> ZmMMS21 (AQK88509.1). Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-175/img_1.png<b>图1</b> 植物SUMO E3连接酶的保守结构域<br/>(A) 植物SUMO E3连接酶MMS21和SIZ1的特征结构域。SIZ1具有SAP、PHD、PINIT、SP-RING和SXS结构域, MMS21只具有SP-RING结构域(<xref ref-type="bibr" rid="b19-1674-3466-53-2-175">Ishida et al., 2012</xref>); (B) SUMO E3连接酶的SP-RING结构域和泛素E3连接酶的RING结构域构型。RING结构域有2个锌离子结合环, SP-RING只含有1个, 第2个环被范德华力和氢键所替代(<xref ref-type="bibr" rid="b14-1674-3466-53-2-175">Duan et al., 2009</xref>; <xref ref-type="bibr" rid="b49-1674-3466-53-2-175">Yunus and Lima, 2009</xref>; <xref ref-type="bibr" rid="b19-1674-3466-53-2-175">Ishida et al., 2012</xref>; <xref ref-type="bibr" rid="b3-1674-3466-53-2-175">石田喬志和杉本慶子, 2012</xref>); (C), (D) 分别为植物SIZ1和MMS21的同源序列比对。数据来源于NCBI。由DNAMAN定义的氨基酸序列、保守序列(黑色)和相似序列(灰色)。SIZ1的蛋白质氨基酸序列为拟南芥(<i>Arabidopsis thaliana</i>) AtSIZ1 (AAU00414.1)、大豆(<i>Glycine max</i>) GmSIZ1a (KRH24918.1)和GmSIZ1b (KRG89029.1)、水稻(<i>Oryza sativa</i>) OsSIZ1 (BAG97182.1)和OsSIZ2 (BAG89374.1)以及玉米(<i>Zea mays</i>) ZmSIZ1a (AQK95338.1)、ZmSIZ1b (AQK79048.1)和ZmSIZc (ONM41936.1)。MMS21的蛋白质氨基酸序列为拟南芥AtMMS21 (NP_188133.2)、大豆GmMMS21 (XP_003541835.1)、水稻OsMMS21 (XP_015640264.1)和玉米ZmMMS21 (AQK88509.1)。<br/><b>Figure 1</b> The domains are conserved in plant SUMO E3 ligases<br/>(A) Characteristic domains of plant SUMO E3 ligases. MMS21 and SIZ1 are shown by boxes, SIZ1 possesses SAP, PHD, PINIT, SP-RING and SXS domains, while MMS21 possesses only SP-RING domain (<xref ref-type="bibr" rid="b19-1674-3466-53-2-175">Ishida et al., 2012</xref>); (B) A schematic model of the SP-RING domain of SUMO E3 ligases and the RING finger of ubiquitin E3 ligases. The RING domain sports two zinc-coordinating loops, the SP-RING domain contains only one, the second loop is instead held together by hydrogen bonds and Van der Waals forces (<xref ref-type="bibr" rid="b14-1674-3466-53-2-175">Duan et al., 2009</xref>; <xref ref-type="bibr" rid="b49-1674-3466-53-2-175">Yunus and Lima, 2009</xref>; <xref ref-type="bibr" rid="b19-1674-3466-53-2-175">Ishida et al., 2012</xref>); (C), (D) The sequence data for the plant SIZ1 homologues were obtained from the NCBI protein database. Sequence identities (black boxes) and similarities (gray boxes) of amino acids were identified by DNAMAN. Amino acid sequences of SIZ1 proteins are from <i>Arabidopsis thaliana </i>AtSIZ1 (AAU00414.1), <i>Glycine max </i>GmSIZ1a (KRH24918.1), GmSIZ1b (KRG89029.1), <i>Oryza sativa </i>OsSIZ1 (BAG97182.1), OsSIZ2 (BAG89374.1), and <i>Zea mays</i> ZmSIZ1a (AQK95338.1), ZmSIZ1b (AQK79048.1), ZmSIZc (ONM41936.1). Amino acid sequences of MMS21 proteins are from <i>Arabidopsis thaliana </i>AtMMS21 (NP_188133.2),<i> Glycine max</i> GmMMS21 (XP_003541835.1), <i>Oryza sativa </i>OsMMS21 (XP_015640264.1), and<i> Zea mays</i> ZmMMS21 (AQK88509.1).
图1 植物SUMO E3连接酶的保守结构域 (A) 植物SUMO E3连接酶MMS21和SIZ1的特征结构域。SIZ1具有SAP、PHD、PINIT、SP-RING和SXS结构域, MMS21只具有SP-RING结构域(Ishida et al., 2012); (B) SUMO E3连接酶的SP-RING结构域和泛素E3连接酶的RING结构域构型。RING结构域有2个锌离子结合环, SP-RING只含有1个, 第2个环被范德华力和氢键所替代(Duan et al., 2009; Yunus and Lima, 2009; Ishida et al., 2012; 石田喬志和杉本慶子, 2012); (C), (D) 分别为植物SIZ1和MMS21的同源序列比对。数据来源于NCBI。由DNAMAN定义的氨基酸序列、保守序列(黑色)和相似序列(灰色)。SIZ1的蛋白质氨基酸序列为拟南芥(Arabidopsis thaliana) AtSIZ1 (AAU00414.1)、大豆(Glycine max) GmSIZ1a (KRH24918.1)和GmSIZ1b (KRG89029.1)、水稻(Oryza sativa) OsSIZ1 (BAG97182.1)和OsSIZ2 (BAG89374.1)以及玉米(Zea mays) ZmSIZ1a (AQK95338.1)、ZmSIZ1b (AQK79048.1)和ZmSIZc (ONM41936.1)。MMS21的蛋白质氨基酸序列为拟南芥AtMMS21 (NP_188133.2)、大豆GmMMS21 (XP_003541835.1)、水稻OsMMS21 (XP_015640264.1)和玉米ZmMMS21 (AQK88509.1)。 Figure 1 The domains are conserved in plant SUMO E3 ligases (A) Characteristic domains of plant SUMO E3 ligases. MMS21 and SIZ1 are shown by boxes, SIZ1 possesses SAP, PHD, PINIT, SP-RING and SXS domains, while MMS21 possesses only SP-RING domain (Ishida et al., 2012); (B) A schematic model of the SP-RING domain of SUMO E3 ligases and the RING finger of ubiquitin E3 ligases. The RING domain sports two zinc-coordinating loops, the SP-RING domain contains only one, the second loop is instead held together by hydrogen bonds and Van der Waals forces (Duan et al., 2009; Yunus and Lima, 2009; Ishida et al., 2012); (C), (D) The sequence data for the plant SIZ1 homologues were obtained from the NCBI protein database. Sequence identities (black boxes) and similarities (gray boxes) of amino acids were identified by DNAMAN. Amino acid sequences of SIZ1 proteins are from Arabidopsis thaliana AtSIZ1 (AAU00414.1), Glycine max GmSIZ1a (KRH24918.1), GmSIZ1b (KRG89029.1), Oryza sativa OsSIZ1 (BAG97182.1), OsSIZ2 (BAG89374.1), and Zea mays ZmSIZ1a (AQK95338.1), ZmSIZ1b (AQK79048.1), ZmSIZc (ONM41936.1). Amino acid sequences of MMS21 proteins are from Arabidopsis thaliana AtMMS21 (NP_188133.2), Glycine max GmMMS21 (XP_003541835.1), Oryza sativa OsMMS21 (XP_015640264.1), and Zea mays ZmMMS21 (AQK88509.1).
2 植物SUMO E3连接酶调节植物根系的发育根作为植物的关键器官, 对整个植物的生长发育至关重要。拟南芥突变体atsiz1和atmms21都出现根发育缺陷表型, 表明这2个SUMO E3连接酶在根系发育调控中具有重要功能。 2.1 SIZ1对根发育的调节外界环境中营养元素水平的波动对根的形态建成有极大的影响。磷酸盐是根吸收和同化磷的主要形式。缺磷条件下, 植物会通过减少主根的生长而增加侧根的伸长来增加根系与土壤的接触面积, 从而获取更多磷(Dong et al., 2017)。突变体atsiz1出现缺磷超敏感表型: 主根变短, 侧根增加和伸长, 根毛发达。一方面AtSIZ1可以介导调控低磷反应关键因子PHR1的SUMO化修饰, 进而正调控磷酸饥饿响应基因AtPS1和AtRNS1的表达以增加磷的吸收(Miura et al., 2005; 杨辉霞等, 2007)。另一方面, Miura等(2011)通过生长素响应因子DR5启动的GUS系统来检测根系中生长素的分布, 发现缺磷后根系的重建与生长素在根系中分布变化相关, 进一步揭示AtSIZ1也可以通过调控生长素积累负调控缺磷响应中的根形态建成。Wang等(2015)在水稻中也发现, 磷胁迫减弱了OsSIZ1缺失增强的根系生长素输出能力。说明SIZ1在低磷条件下对根构型的重建具有重要作用。 此外, SIZ1还通过调节脱落酸(ABA)和糖代谢途径参与根系调节(Castro et al., 2015)。拟南芥突变体atsiz1对ABA超敏感且主根生长受抑制。研究发现这是由于AtSIZ1介导了ABA信号相关bZIP转录因子ABI5和MYB30的SUMO化, 从而影响ABA对幼苗初生根的调节与种子的萌发(Miura et al., 2009; Zheng et al., 2012)。外源糖会促进体内SUMO分子的积累, 同时AtSIZ1负调节淀粉和糖代谢相关基因的转录水平而影响初生根的发育(Castro et al., 2015)。
2.2 MMS21对根发育的调节根尖干细胞的命运是由多能转录因子调节的, 如维持干细胞活性的静止中心(quiescent center, QC)特异转录因子WOX5、决定干细胞微环境横向模式建立的SHR和SCR以及调控干细胞微环境纵向模式建立的PLTs。拟南芥突变体atmms21根尖分生组织的细胞分裂和细胞分化模式发生改变(徐庞连等, 2008; Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010), 说明AtMMS21是根尖分生组织的结构与功能维持所必需的。Xu等(2013)通过检测QC特异性标记的表达和根尖干细胞的命运, 发现突变体atmms21根尖中WOX5出现异位表达; SHR和SCR的正常空间表达模式出现部分缺失, 进而呈现不连续表达; PLT1和PLT2在蛋白水平发生严重下调, 进而呈现表达/积累减少的模式。此外, 拟南芥突变体atmms21胚胎中胚根原基的后代细胞出现异常分裂, 导致根尖干细胞微环境起始的位置——胚胎基极发生结构错乱。这些现象表明, AtMMS21在胚胎和胚后发育阶段是根尖QC组织维持及调节多能性转录因子在根尖干细胞微环境中稳定表达所必需的。另一方面, Ishida等(2009)发现PLT1和PLT2反过来对AtMMS21的表达或积累起着正调控作用; Zhang等(2010)发现突变体atmms- 21对外源细胞分裂素的敏感性降低, ARRs基因表达下调。因此可以推断AtMMS21可能是细胞分裂素与生长素相互作用的一个节点, 通过调控植物激素来维持拟南芥根尖干细胞的稳态。 突变体atmms21对DNA损伤剂超敏感, 根尖干细胞及其子代细胞出现明显的细胞死亡。对酵母(Sac- charomyces cerevisiae)和人类(Homo sapiens)的研究表明, MMS21是染色体结构维持复合体(Structure Maintenance of Chromosomes5/6, SMC5/6)的关键组分, 是促进DNA双链断裂(DNA double-strand break, DSB)修复所必需的(De Piccoli et al., 2006; Watanabe et al., 2009; Almedawar et al., 2012)。在拟南芥中, atmms21突变体中DSB的标记基因和DSB修复标记基因都有较强的表达(Xu et al., 2013; Yuan et al., 2014), 说明MMS21在物种间有很强的保守性。因此推测AtMMS21可能通过与SMC5相互作用在DNA双链断裂修复中发挥功能, 从而维持根尖干细胞微环境的稳定以调控根系的正常发育。 除了启动DNA修复机制, 细胞还可以通过严格控制细胞周期进程对DSB进行响应。突变体atmms21根尖分生组织中的细胞过早地从有丝分裂周期进入内复制周期, 细胞周期G2/M期转换相关基因CYCB;1、CDKB1和CDKB2等的表达也显著降低(Huang et al., 2009; Ishida et al., 2009)。Liu等(2016)发现AtMMS21与细胞周期调控因子DPa相互作用并介导其SUMO化修饰, 同时AtMMS21可以恢复E2Fa/DPa双过表达植物的发育缺陷, 进一步研究表明AtMMS21通过SUMO化及物理竞争作用导致DPa与E2Fa的结合能力下降, 影响E2Fa/DPa复合物的核定位, 负调控G1/S期转换, 进而调节根分生组织中细胞分裂和分化的平衡。 染色质重塑复合物在植物根系发育相关基因的表达调控方面具有重要作用。在有关酵母和人类的研究报道中, SUMO E3连接酶可以作为转录调节复合物调节基因的表达和染色质重塑(Wotton et al., 2017)。Zhang等(2017a)发现AtMMS21可以与染色质重塑复合物BRM相互作用并促进其SUMO化修饰, 维持BRM蛋白的稳定性, 进而调控根发育相关基因的正常表达, 以维持根尖微环境的稳态。
3 植物SUMO E3连接酶调节植物开花途径开花是植物由营养生长过渡到生殖生长的重要转折点。突变体atsiz1和atmms21在短日照下都具有明显的早花表型(Jin et al., 2008; Kwak et al., 2016)。Jin等(2008)发现拟南芥突变体atsiz1中的SA水平升高, FLC的mRNA水平降低, 表达nahG基因(降低SA水平)可以抑制atsiz1的早花表型, 所以AtSIZ1可以通过调节SA水平调节开花途径。同时FLC也是一个SUMO化修饰的底物, AtSIZ1和AtMMS21都与其相互作用, 共同调节FLC的稳定性: 即AtMMS21介导FLC的SUMO化修饰, 正调控FLC的功能; AtSIZ1抑制FLC的SUMO化, 从而负调控FLC (Kwak et al., 2016)。此外, AtSIZ1介导FLD的SUMO化, 进而抑制FLC染色质区域组蛋白H4的去乙酰化, 从而抑制了FLC的表达(Jin et al., 2008)。在石斛中也发现DnSIZ1可以通过诱导春化途径调节开花(Liu et al., 2015)。以上实验证据表明, SUMO E3连接酶介导的SUMO化修饰从多个层次对FLC依赖的开花途径进行调节。
4 植物SUMO E3连接酶调节配子发育SUMO E3连接酶也参与植物的生殖发育进程。拟南芥突变体atmms21有严重的生育缺陷, 虽然可以完成其生殖阶段, 但结实率低, 种子败育率高。Liu等(2014)用突变体atmms21与野生型异杂交, 发现大部分突变体结实率下降可能是由于花粉管的缺陷导致的。虽然一些雌配子的缺陷也可能导致不育, 但是相对来说影响较小。突变体atmms21在生殖发育的第7阶段就出现花药发育缺陷, 而不是在花药的成熟时期。随后的阶段中, 突变体atmms21的缺陷越来越明显, 第8阶段中出现小孢子形状和大小异常, 到第11阶段甚至发生退化。大孢子母细胞减数分裂异常, 染色体在前期I就出现缺陷, 后期II中姐妹染色单体分离也观察到缺陷, 这导致减数分裂后DNA分布不均。因此, AtMMS21是拟南芥减数分裂与配子体发育所必需的(Liu et al., 2014)。 与AtMMS21不同, AtSIZ1在花、萼片和雌蕊中的表达量最高。Ling等(2012)发现, 拟南芥突变体atsiz1雌蕊中有23%的胚珠是干瘪的, 花粉管萌发及花粉管进入珠孔的过程都正常, 但是由于雌配子发育异常导致部分配子在最后阶段无法完成受精(Ling et al., 2012), 进而结实率降低(Kim et al., 2015, 2016b)。由此表明, AtSIZ1是拟南芥雌配子正常发育和种子萌发所必需。而水稻ossiz1突变体则出现花药不开裂和结实率下降的表型(Wang et al., 2015)。Thangasamy等(2001)推断OsSIZ1可能通过调节药室内壁发育而影响花药的开裂。这表明SUMO E3连接酶在不同植物中的功能有所差异。
5 植物SUMO E3连接酶调节光形态建成泛素E3连接酶COP1是植物光形态建成的关键调控因子, 拟南芥突变体cop1出现光生长抑制, 即使在黑暗条件下也会出现下胚轴短的表型。与突变体cop1相似, 突变体atsiz1幼苗也会在黑暗或白光、红光、蓝光和远红光下出现下胚轴短的表型, 该表型不能通过表达nahG (降低SA水平)恢复, 说明AtSIZ1对光形态建成的调节独立于SA信号途径。进一步研究发现, AtSIZ1可以介导COP1 (K193)的SUMO化修饰。突变体atsiz1中过表达COP1或是在野生型中过表达COP1 K193 (突变其SUMO化修饰位点)与过表达完整的COP表型不一致。这是由于AtSIZ1介导COP1的SUMO化增强了COP1的泛素E3连接酶活性, 导致COP1对HY5的泛素化修饰水平增加, 从而促进HY5降解。另一方面, COP1促进AtSIZ1的泛素化修饰, 导致AtSIZ1通过26S蛋白酶体降解。光照条件下, COP1的SUMO化水平下降, 其活性受到抑制。因此, COP1和AtSIZ1之间形成了一个光形态建成的反馈调节机制(Kim et al., 2016a; Lin et al., 2016)。
6 总结与展望从SUMO分子被发现至今, 科学家对植物生物学中SUMO化修饰系统的研究已经延伸到各个方面。SUMO E3连接酶参与植物根、花、种子以及叶片等重要器官的发育和环境胁迫响应(图2)。植物细胞中少量的连接酶E3对大量的SUMO修饰靶蛋白进行调控, 为后续研究提供了广大的空间。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-175/img_2.png<b>图2</b> SUMO E3连接酶在拟南芥生长发育中的功能<br/>(A) AtMMS21对细胞周期的调节功能(<xref ref-type="bibr" rid="b31-1674-3466-53-2-175">Liu et al., 2016</xref>); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(<xref ref-type="bibr" rid="b26-1674-3466-53-2-175">Kwak et al., 2016</xref>); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(<xref ref-type="bibr" rid="b46-1674-3466-53-2-175">Xu et al., 2013</xref>; <xref ref-type="bibr" rid="b48-1674-3466-53-2-175">Yuan et al., 2014</xref>); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(<xref ref-type="bibr" rid="b18-1674-3466-53-2-175">Ishida et al., 2009</xref>; <xref ref-type="bibr" rid="b17-1674-3466-53-2-175">Huang et al., 2009</xref>; <xref ref-type="bibr" rid="b53-1674-3466-53-2-175">Zhang et al., 2010</xref>); (E) MMS21通过染色质重塑复合物调节根分生组织发育(<xref ref-type="bibr" rid="b50-1674-3466-53-2-175">Zhang et al., 2017a</xref>); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(<xref ref-type="bibr" rid="b35-1674-3466-53-2-175">Miura et al., 2009</xref>); (G) SIZ1对开花途径的调节(<xref ref-type="bibr" rid="b20-1674-3466-53-2-175">Jin et al., 2008</xref>); (H) SIZ1对植物光形态建成的调控(<xref ref-type="bibr" rid="b23-1674-3466-53-2-175">Kim et al., 2016a</xref>; <xref ref-type="bibr" rid="b27-1674-3466-53-2-175">Lin et al., 2016</xref>); (I) SIZ1对磷酸饥饿响应的调控(<xref ref-type="bibr" rid="b36-1674-3466-53-2-175">Miura et al., 2005</xref>; <xref ref-type="bibr" rid="b34-1674-3466-53-2-175">2011</xref>); (J) SIZ1通过糖代谢途径调节根的发育(<xref ref-type="bibr" rid="b7-1674-3466-53-2-175">Castro et al., 2015</xref>)<br/><b>Figure 2</b> The functions of SUMO E3 ligases in Arabidopsis<br/>(A) The function of MMS21 regulates in cell cycle (<xref ref-type="bibr" rid="b31-1674-3466-53-2-175">Liu et al., 2016</xref>); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (<xref ref-type="bibr" rid="b26-1674-3466-53-2-175">Kwak et al., 2016</xref>); (C) MMS21 responses to the DNA damage to regulate plant growth and development (<xref ref-type="bibr" rid="b46-1674-3466-53-2-175">Xu et al., 2013</xref>; <xref ref-type="bibr" rid="b48-1674-3466-53-2-175">Yuan et al., 2014</xref>); (D) MMS21 is involved in Arabidopsis root development by auxin (<xref ref-type="bibr" rid="b18-1674-3466-53-2-175">Ishida et al., 2009</xref>; <xref ref-type="bibr" rid="b17-1674-3466-53-2-175">Huang et al., 2009</xref>; <xref ref-type="bibr" rid="b53-1674-3466-53-2-175">Zhang et al., 2010</xref>); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (<xref ref-type="bibr" rid="b50-1674-3466-53-2-175">Zhang et al., 2017a</xref>); (F) SIZ1 regulate glucose metabolism germinated/seedling development (<xref ref-type="bibr" rid="b35-1674-3466-53-2-175">Miura et al., 2009</xref>); (G) SIZ1 is involved in flowering pathway (<xref ref-type="bibr" rid="b20-1674-3466-53-2-175">Jin et al., 2008</xref>); (H) SIZ1 regulates the photomorphogenesis (<xref ref-type="bibr" rid="b23-1674-3466-53-2-175">Kim et al., 2016a</xref>; <xref ref-type="bibr" rid="b27-1674-3466-53-2-175">Lin et al., 2016</xref>); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (<xref ref-type="bibr" rid="b36-1674-3466-53-2-175">Miura et al., 2005</xref>, <xref ref-type="bibr" rid="b34-1674-3466-53-2-175">2011</xref>); (J) SIZ1 regulates development of root by sugar pathway (<xref ref-type="bibr" rid="b7-1674-3466-53-2-175">Castro et al., 2015</xref>) Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-2-175/img_2.png<b>图2</b> SUMO E3连接酶在拟南芥生长发育中的功能<br/>(A) AtMMS21对细胞周期的调节功能(<xref ref-type="bibr" rid="b31-1674-3466-53-2-175">Liu et al., 2016</xref>); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(<xref ref-type="bibr" rid="b26-1674-3466-53-2-175">Kwak et al., 2016</xref>); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(<xref ref-type="bibr" rid="b46-1674-3466-53-2-175">Xu et al., 2013</xref>; <xref ref-type="bibr" rid="b48-1674-3466-53-2-175">Yuan et al., 2014</xref>); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(<xref ref-type="bibr" rid="b18-1674-3466-53-2-175">Ishida et al., 2009</xref>; <xref ref-type="bibr" rid="b17-1674-3466-53-2-175">Huang et al., 2009</xref>; <xref ref-type="bibr" rid="b53-1674-3466-53-2-175">Zhang et al., 2010</xref>); (E) MMS21通过染色质重塑复合物调节根分生组织发育(<xref ref-type="bibr" rid="b50-1674-3466-53-2-175">Zhang et al., 2017a</xref>); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(<xref ref-type="bibr" rid="b35-1674-3466-53-2-175">Miura et al., 2009</xref>); (G) SIZ1对开花途径的调节(<xref ref-type="bibr" rid="b20-1674-3466-53-2-175">Jin et al., 2008</xref>); (H) SIZ1对植物光形态建成的调控(<xref ref-type="bibr" rid="b23-1674-3466-53-2-175">Kim et al., 2016a</xref>; <xref ref-type="bibr" rid="b27-1674-3466-53-2-175">Lin et al., 2016</xref>); (I) SIZ1对磷酸饥饿响应的调控(<xref ref-type="bibr" rid="b36-1674-3466-53-2-175">Miura et al., 2005</xref>; <xref ref-type="bibr" rid="b34-1674-3466-53-2-175">2011</xref>); (J) SIZ1通过糖代谢途径调节根的发育(<xref ref-type="bibr" rid="b7-1674-3466-53-2-175">Castro et al., 2015</xref>)<br/><b>Figure 2</b> The functions of SUMO E3 ligases in Arabidopsis<br/>(A) The function of MMS21 regulates in cell cycle (<xref ref-type="bibr" rid="b31-1674-3466-53-2-175">Liu et al., 2016</xref>); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (<xref ref-type="bibr" rid="b26-1674-3466-53-2-175">Kwak et al., 2016</xref>); (C) MMS21 responses to the DNA damage to regulate plant growth and development (<xref ref-type="bibr" rid="b46-1674-3466-53-2-175">Xu et al., 2013</xref>; <xref ref-type="bibr" rid="b48-1674-3466-53-2-175">Yuan et al., 2014</xref>); (D) MMS21 is involved in Arabidopsis root development by auxin (<xref ref-type="bibr" rid="b18-1674-3466-53-2-175">Ishida et al., 2009</xref>; <xref ref-type="bibr" rid="b17-1674-3466-53-2-175">Huang et al., 2009</xref>; <xref ref-type="bibr" rid="b53-1674-3466-53-2-175">Zhang et al., 2010</xref>); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (<xref ref-type="bibr" rid="b50-1674-3466-53-2-175">Zhang et al., 2017a</xref>); (F) SIZ1 regulate glucose metabolism germinated/seedling development (<xref ref-type="bibr" rid="b35-1674-3466-53-2-175">Miura et al., 2009</xref>); (G) SIZ1 is involved in flowering pathway (<xref ref-type="bibr" rid="b20-1674-3466-53-2-175">Jin et al., 2008</xref>); (H) SIZ1 regulates the photomorphogenesis (<xref ref-type="bibr" rid="b23-1674-3466-53-2-175">Kim et al., 2016a</xref>; <xref ref-type="bibr" rid="b27-1674-3466-53-2-175">Lin et al., 2016</xref>); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (<xref ref-type="bibr" rid="b36-1674-3466-53-2-175">Miura et al., 2005</xref>, <xref ref-type="bibr" rid="b34-1674-3466-53-2-175">2011</xref>); (J) SIZ1 regulates development of root by sugar pathway (<xref ref-type="bibr" rid="b7-1674-3466-53-2-175">Castro et al., 2015</xref>)
DesterroJMP, RodriguezMS, KempGD, HayRT (1999). Identification of the enzyme required for activation of the small ubiquitin-like protein SUMO-1.J Biol Chem 274, 10618-10624. [本文引用: 1]
[12]
DesterroJMP, ThomsonJ, HayRT (1997). Ubch9 conjugates SUMO but not ubiquitin.FEBS Lett 417, 297-300. [本文引用: 1]
[13]
DongJS, PiñerosMA, LiXX, YangHB, LiuY, MurphyAS, KochianLV, LiuD (2017). An Arabidopsis ABC transporter mediates phosphate deficiency-induced remodeling of root architecture by modulating iron homeostasis in roots.Mol Plant 10, 244-259. [本文引用: 1]
[14]
DuanX, SarangiP, LiuX, RangiGK, ZhaoX, YeH (2009). Structural and functional insights into the roles of the Mms21 subunit of the Smc5/6 complex.Mol Cell 35, 657-668. [本文引用: 2]
[15]
EisenhardtN, ChauguleVK, KoidlS, DroescherM, DoganE, RettichJ, SutinenP, ImanishiSY, HofmannK, PalvimoJJ, PichlerA (2015). A new vertebrate SUMO enzyme family reveals insights into SUMO-chain assembly.Nat Struct Mol Biol 22, 959-967. [本文引用: 1]
[16]
GongLM, LiB, MillasS, YehETH (1999). Molecular cloning and characterization of human AOS1 and UBA2, components of the sentrin-activating enzyme complex.FEBS Lett 448, 185-189. [本文引用: 1]
[17]
HuangLX, YangSG, ZhangSC, LiuM, LaiJB, QiYL, ShiSF, WangJX, WangYQ, XieQ, YangCW (2009). The Arabidopsis SUMO E3 ligase AtMMS21, a homologue of NSE2/MMS21, regulates cell proliferation in the root.Plant J 60, 666-678. [本文引用: 7]
[18]
IshidaT, FujiwaraS, MiuraK, StaceyN, YoshimuraM, SchneiderK, AdachiS, MinamisawaK, UmedaM, SugimotoK (2009). SUMO E3 ligase HIGH PLOIDY2 regul- ates endocycle onset and meristem maintenance in Arabidopsis.Plant Cell 21, 2284-2297. [本文引用: 7]
[19]
IshidaT, YoshimuraM, MiuraK, SugimotoK (2012). MMS21/HPY2 and SIZ1, two Arabidopsis SUMO E3 ligases, have distinct functions in development.PLoS One 7, e46897. [本文引用: 7]
[20]
JinJB, JinYH, LeeJ, MiuraK, YooCY, KimWY, Van OostenM, HyunY, SomersDE, LeeI, YunDJ, BressanRA, HasegawaPM (2008). The SUMO E3 ligase, AtSIZ1, regulates flowering by controlling a salicylic acid-mediated floral promotion pathway and through affects onFLC chromatin structure. Plant J 53, 530-540. [本文引用: 5]
[21]
JohnsonES, BlobelG (1997). Ubc9p is the conjugating enzyme for the ubiquitin-like protein Smt3p.J Biol Chem 272, 26799-26802. [本文引用: 1]
[22]
JohnsonES, GuptaAA (2001). An E3-like factor that promotes SUMO conjugation to the yeast septins.Cell 106, 735-744. [本文引用: 1]
[23]
KimJY, JangIC, SeoHS (2016a). COP1 controls abiotic stress responses by modulating AtSIZ1 function through its E3 ubiquitin ligase activity.Front Plant Sci 7, 1182. [本文引用: 3]
[24]
KimSI, KwakJS, SongJT, SeoHS (2016b). The E3 SUMO ligase AtSIZ1 functions in seed germination in Arabidopsis.Physiol Plant 158, 256-271. [本文引用: 1]
KwakJS, SonGH, KimSI, SongJT, SeoHS (2016). Arabidopsis HIGH PLOIDY2 sumoylates and stabilizes Flowering Locus C through its E3 ligase activity.Front Plant Sci 7, 530. [本文引用: 4]
ReverterD, LimaCD (2005). Insights into E3 ligase activity revealed by a SUMO-RanGAP1-Ubc9-Nup358 complex.Nature 435, 687-692. [本文引用: 1]
[41]
Streich JrFC, LimaCD (2016). Capturing a substrate in an activated RING E3/E2-SUMO complex.Nature 536, 304-308. [本文引用: 1]
[42]
ThangasamyS, GuoCL, ChuangMH, LaiMH, ChenJ, JauhGY (2011). Rice SIZ1, a SUMO E3 ligase, controls spikelet fertility through regulation of anther dehiscence.New Phytol 189, 869-882. [本文引用: 1]
[43]
WangHD, SunR, CaoY, PeiWX, SunYF, ZhouHM, WuXN, ZhangF, LuoL, ShenQR, XuGH (2015). OsSIZ1, a SUMO E3 ligase gene, is involved in the regulation of the responses to phosphate and nitrogen in rice. Plant Cell Physiol 56, 2381-2395 [本文引用: 2]
[44]
WatanabeK, PacherM, DukowicS, SchubertV, PuchtaH, SchubertI (2009). The STRUCTURAL MAINTENANCE OF CHROMOSOMES 5/6 complex promotes sister chromatid alignment and homologous recombination after DNA damage in Arabidopsis thaliana.Plant Cell 21, 2688-2699. [本文引用: 1]
[45]
WottonD, PembertonLF, Merrill-SchoolsJ (2017). SUMO and Chromatin Remodeling. In: Wilson VG, ed. SUMO Regulation of Cellular Processes. Cham: Springer. pp. 35-50. [本文引用: 1]
[46]
XuPL, YuanDK, LiuM, LiCX, LiuYY, ZhangSC, YaoN, YangCW (2013). AtMMS21, an SMC5/6 complex subunit, is involved in stem cell niche maintenance and DNA damage responses in Arabidopsis roots.Plant Physiol 161, 1755-1768. [本文引用: 4]
[47]
YooCY, MiuraK, JinJB, LeeJ, ParkHC, SaltDE, YunDJ, BressanRA, HasegawaPM (2006). SIZ1 small ubiquitin-like modifier E3 ligase facilitates basal thermotolerance in Arabidopsis independent of salicylic acid.Plant Physiol 142, 1548-1558. [本文引用: 1]
[48]
YuanDK, LaiJB, XuPL, ZhangSC, ZhangJJ, LiCX, WangYQ, DuJJ, LiuYY, YangCW (2014). AtMMS21 regulates DNA damage response and homologous recombination repair in Arabidopsis.DNA Repair 21, 140-147. [本文引用: 3]
[49]
YunusAA, LimaCD (2009). Structure of the Siz/PIAS SUMO E3 ligase Siz1 and determinants required for SUMO modification of PCNA.Mol Cell 35, 669-682. [本文引用: 3]
[50]
ZhangJJ, LaiJB, WangFG, YangSG, HeZP, JiangJM, LiQL, WuQ, LiuYY, YuMY, DuMY, DuJJ, XieQ, WuKQ (2017).a A SUMO ligase AtMMS21 regulates the stability of the chromatin remodeler BRAHMA in root deve- lopment.Plant Physiol 173 1574-1582. [本文引用: 3]
[51]
ZhangRF, GuoY, LiYY, ZhouLJ, HaoYJ, YouCX (2016). Functional identification of MdSIZ1 as a SUMO E3 ligase in apple.J Plant Physiol 198, 69-80. [本文引用: 1]
... 突变体atmms21对DNA损伤剂超敏感, 根尖干细胞及其子代细胞出现明显的细胞死亡.对酵母(Sac- charomyces cerevisiae)和人类(Homo sapiens)的研究表明, MMS21是染色体结构维持复合体(Structure Maintenance of Chromosomes5/6, SMC5/6)的关键组分, 是促进DNA双链断裂(DNA double-strand break, DSB)修复所必需的(De Piccoli et al., 2006; Watanabe et al., 2009; Almedawar et al., 2012).在拟南芥中, atmms21突变体中DSB的标记基因和DSB修复标记基因都有较强的表达(Xu et al., 2013; Yuan et al., 2014), 说明MMS21在物种间有很强的保守性.因此推测AtMMS21可能通过与SMC5相互作用在DNA双链断裂修复中发挥功能, 从而维持根尖干细胞微环境的稳定以调控根系的正常发育. ...
1 2017
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
1 2015
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
4 2015
... 此外, SIZ1还通过调节脱落酸(ABA)和糖代谢途径参与根系调节(Castro et al., 2015).拟南芥突变体atsiz1对ABA超敏感且主根生长受抑制.研究发现这是由于AtSIZ1介导了ABA信号相关bZIP转录因子ABI5和MYB30的SUMO化, 从而影响ABA对幼苗初生根的调节与种子的萌发(Miura et al., 2009; Zheng et al., 2012).外源糖会促进体内SUMO分子的积累, 同时AtSIZ1负调节淀粉和糖代谢相关基因的转录水平而影响初生根的发育(Castro et al., 2015). ... ... ).外源糖会促进体内SUMO分子的积累, 同时AtSIZ1负调节淀粉和糖代谢相关基因的转录水平而影响初生根的发育(Castro et al., 2015). ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
2 2007
... 2个植物SUMO E3连接酶除了结构域的差异之外, 其功能和分布也大不相同.AtSIZ1主要在叶片、叶柄、根尖、花絮、雄蕊和柱头中表达(Catala et al., 2007), 主要通过调节SA水平参与植物逆境胁迫响应, 如磷酸饥饿响应、耐盐、耐高温及耐冷等性状(Miura et al., 2005, 2007b; Yoo et al., 2006; Catala et al., 2007).AtMMS21则主要在花药、叶脉和下胚轴分生组织的增殖细胞中表达(Ishida et al., 2012), 参与细胞周期进程和分生组织的维持(Ishida et al., 2009; Huang et al., 2009).两者的突变体分别出现多效性表型, 实验表明双突体atsiz1/atmms21胚胎致死, 同时AtSIZ1不能互补突变体atmms21的表型, 而AtMMS21也无法互补突变体atsiz1的表型(Ishida et al., 2012), 说明2个连接酶在功能上相互独立, 共同参与SUMO化对植物生长发育的精细调控. ... ... ; Catala et al., 2007).AtMMS21则主要在花药、叶脉和下胚轴分生组织的增殖细胞中表达(Ishida et al., 2012), 参与细胞周期进程和分生组织的维持(Ishida et al., 2009; Huang et al., 2009).两者的突变体分别出现多效性表型, 实验表明双突体atsiz1/atmms21胚胎致死, 同时AtSIZ1不能互补突变体atmms21的表型, 而AtMMS21也无法互补突变体atsiz1的表型(Ishida et al., 2012), 说明2个连接酶在功能上相互独立, 共同参与SUMO化对植物生长发育的精细调控. ...
... 突变体atmms21对DNA损伤剂超敏感, 根尖干细胞及其子代细胞出现明显的细胞死亡.对酵母(Sac- charomyces cerevisiae)和人类(Homo sapiens)的研究表明, MMS21是染色体结构维持复合体(Structure Maintenance of Chromosomes5/6, SMC5/6)的关键组分, 是促进DNA双链断裂(DNA double-strand break, DSB)修复所必需的(De Piccoli et al., 2006; Watanabe et al., 2009; Almedawar et al., 2012).在拟南芥中, atmms21突变体中DSB的标记基因和DSB修复标记基因都有较强的表达(Xu et al., 2013; Yuan et al., 2014), 说明MMS21在物种间有很强的保守性.因此推测AtMMS21可能通过与SMC5相互作用在DNA双链断裂修复中发挥功能, 从而维持根尖干细胞微环境的稳定以调控根系的正常发育. ...
1 1999
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
1 1997
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
... 植物SUMO E3连接酶的保守结构域 (A) 植物SUMO E3连接酶MMS21和SIZ1的特征结构域.SIZ1具有SAP、PHD、PINIT、SP-RING和SXS结构域, MMS21只具有SP-RING结构域(Ishida et al., 2012); (B) SUMO E3连接酶的SP-RING结构域和泛素E3连接酶的RING结构域构型.RING结构域有2个锌离子结合环, SP-RING只含有1个, 第2个环被范德华力和氢键所替代(Duan et al., 2009; Yunus and Lima, 2009; Ishida et al., 2012; 石田喬志和杉本慶子, 2012); (C), (D) 分别为植物SIZ1和MMS21的同源序列比对.数据来源于NCBI.由DNAMAN定义的氨基酸序列、保守序列(黑色)和相似序列(灰色).SIZ1的蛋白质氨基酸序列为拟南芥(Arabidopsis thaliana) AtSIZ1 (AAU00414.1)、大豆(Glycine max) GmSIZ1a (KRH24918.1)和GmSIZ1b (KRG89029.1)、水稻(Oryza sativa) OsSIZ1 (BAG97182.1)和OsSIZ2 (BAG89374.1)以及玉米(Zea mays) ZmSIZ1a (AQK95338.1)、ZmSIZ1b (AQK79048.1)和ZmSIZc (ONM41936.1).MMS21的蛋白质氨基酸序列为拟南芥AtMMS21 (NP_188133.2)、大豆GmMMS21 (XP_003541835.1)、水稻OsMMS21 (XP_015640264.1)和玉米ZmMMS21 (AQK88509.1). ... ... The domains are conserved in plant SUMO E3 ligases (A) Characteristic domains of plant SUMO E3 ligases. MMS21 and SIZ1 are shown by boxes, SIZ1 possesses SAP, PHD, PINIT, SP-RING and SXS domains, while MMS21 possesses only SP-RING domain (Ishida et al., 2012); (B) A schematic model of the SP-RING domain of SUMO E3 ligases and the RING finger of ubiquitin E3 ligases. The RING domain sports two zinc-coordinating loops, the SP-RING domain contains only one, the second loop is instead held together by hydrogen bonds and Van der Waals forces (Duan et al., 2009; Yunus and Lima, 2009; Ishida et al., 2012); (C), (D) The sequence data for the plant SIZ1 homologues were obtained from the NCBI protein database. Sequence identities (black boxes) and similarities (gray boxes) of amino acids were identified by DNAMAN. Amino acid sequences of SIZ1 proteins are from Arabidopsis thaliana AtSIZ1 (AAU00414.1), Glycine max GmSIZ1a (KRH24918.1), GmSIZ1b (KRG89029.1), Oryza sativa OsSIZ1 (BAG97182.1), OsSIZ2 (BAG89374.1), and Zea mays ZmSIZ1a (AQK95338.1), ZmSIZ1b (AQK79048.1), ZmSIZc (ONM41936.1). Amino acid sequences of MMS21 proteins are from Arabidopsis thaliana AtMMS21 (NP_188133.2), Glycine max GmMMS21 (XP_003541835.1), Oryza sativa OsMMS21 (XP_015640264.1), and Zea mays ZmMMS21 (AQK88509.1). ...
1 2015
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
1 1999
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
7 2009
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ... ... )和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ... ... 2个植物SUMO E3连接酶除了结构域的差异之外, 其功能和分布也大不相同.AtSIZ1主要在叶片、叶柄、根尖、花絮、雄蕊和柱头中表达(Catala et al., 2007), 主要通过调节SA水平参与植物逆境胁迫响应, 如磷酸饥饿响应、耐盐、耐高温及耐冷等性状(Miura et al., 2005, 2007b; Yoo et al., 2006; Catala et al., 2007).AtMMS21则主要在花药、叶脉和下胚轴分生组织的增殖细胞中表达(Ishida et al., 2012), 参与细胞周期进程和分生组织的维持(Ishida et al., 2009; Huang et al., 2009).两者的突变体分别出现多效性表型, 实验表明双突体atsiz1/atmms21胚胎致死, 同时AtSIZ1不能互补突变体atmms21的表型, 而AtMMS21也无法互补突变体atsiz1的表型(Ishida et al., 2012), 说明2个连接酶在功能上相互独立, 共同参与SUMO化对植物生长发育的精细调控. ... ... 根尖干细胞的命运是由多能转录因子调节的, 如维持干细胞活性的静止中心(quiescent center, QC)特异转录因子WOX5、决定干细胞微环境横向模式建立的SHR和SCR以及调控干细胞微环境纵向模式建立的PLTs.拟南芥突变体atmms21根尖分生组织的细胞分裂和细胞分化模式发生改变(徐庞连等, 2008; Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010), 说明AtMMS21是根尖分生组织的结构与功能维持所必需的.Xu等(2013)通过检测QC特异性标记的表达和根尖干细胞的命运, 发现突变体atmms21根尖中WOX5出现异位表达; SHR和SCR的正常空间表达模式出现部分缺失, 进而呈现不连续表达; PLT1和PLT2在蛋白水平发生严重下调, 进而呈现表达/积累减少的模式.此外, 拟南芥突变体atmms21胚胎中胚根原基的后代细胞出现异常分裂, 导致根尖干细胞微环境起始的位置——胚胎基极发生结构错乱.这些现象表明, AtMMS21在胚胎和胚后发育阶段是根尖QC组织维持及调节多能性转录因子在根尖干细胞微环境中稳定表达所必需的.另一方面, Ishida等(2009)发现PLT1和PLT2反过来对AtMMS21的表达或积累起着正调控作用; Zhang等(2010)发现突变体atmms- 21对外源细胞分裂素的敏感性降低, ARRs基因表达下调.因此可以推断AtMMS21可能是细胞分裂素与生长素相互作用的一个节点, 通过调控植物激素来维持拟南芥根尖干细胞的稳态. ... ... 除了启动DNA修复机制, 细胞还可以通过严格控制细胞周期进程对DSB进行响应.突变体atmms21根尖分生组织中的细胞过早地从有丝分裂周期进入内复制周期, 细胞周期G2/M期转换相关基因CYCB;1、CDKB1和CDKB2等的表达也显著降低(Huang et al., 2009; Ishida et al., 2009).Liu等(2016)发现AtMMS21与细胞周期调控因子DPa相互作用并介导其SUMO化修饰, 同时AtMMS21可以恢复E2Fa/DPa双过表达植物的发育缺陷, 进一步研究表明AtMMS21通过SUMO化及物理竞争作用导致DPa与E2Fa的结合能力下降, 影响E2Fa/DPa复合物的核定位, 负调控G1/S期转换, 进而调节根分生组织中细胞分裂和分化的平衡. ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
7 2009
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ... ... ; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ... ... 2个植物SUMO E3连接酶除了结构域的差异之外, 其功能和分布也大不相同.AtSIZ1主要在叶片、叶柄、根尖、花絮、雄蕊和柱头中表达(Catala et al., 2007), 主要通过调节SA水平参与植物逆境胁迫响应, 如磷酸饥饿响应、耐盐、耐高温及耐冷等性状(Miura et al., 2005, 2007b; Yoo et al., 2006; Catala et al., 2007).AtMMS21则主要在花药、叶脉和下胚轴分生组织的增殖细胞中表达(Ishida et al., 2012), 参与细胞周期进程和分生组织的维持(Ishida et al., 2009; Huang et al., 2009).两者的突变体分别出现多效性表型, 实验表明双突体atsiz1/atmms21胚胎致死, 同时AtSIZ1不能互补突变体atmms21的表型, 而AtMMS21也无法互补突变体atsiz1的表型(Ishida et al., 2012), 说明2个连接酶在功能上相互独立, 共同参与SUMO化对植物生长发育的精细调控. ... ... 根尖干细胞的命运是由多能转录因子调节的, 如维持干细胞活性的静止中心(quiescent center, QC)特异转录因子WOX5、决定干细胞微环境横向模式建立的SHR和SCR以及调控干细胞微环境纵向模式建立的PLTs.拟南芥突变体atmms21根尖分生组织的细胞分裂和细胞分化模式发生改变(徐庞连等, 2008; Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010), 说明AtMMS21是根尖分生组织的结构与功能维持所必需的.Xu等(2013)通过检测QC特异性标记的表达和根尖干细胞的命运, 发现突变体atmms21根尖中WOX5出现异位表达; SHR和SCR的正常空间表达模式出现部分缺失, 进而呈现不连续表达; PLT1和PLT2在蛋白水平发生严重下调, 进而呈现表达/积累减少的模式.此外, 拟南芥突变体atmms21胚胎中胚根原基的后代细胞出现异常分裂, 导致根尖干细胞微环境起始的位置——胚胎基极发生结构错乱.这些现象表明, AtMMS21在胚胎和胚后发育阶段是根尖QC组织维持及调节多能性转录因子在根尖干细胞微环境中稳定表达所必需的.另一方面, Ishida等(2009)发现PLT1和PLT2反过来对AtMMS21的表达或积累起着正调控作用; Zhang等(2010)发现突变体atmms- 21对外源细胞分裂素的敏感性降低, ARRs基因表达下调.因此可以推断AtMMS21可能是细胞分裂素与生长素相互作用的一个节点, 通过调控植物激素来维持拟南芥根尖干细胞的稳态. ... ... 除了启动DNA修复机制, 细胞还可以通过严格控制细胞周期进程对DSB进行响应.突变体atmms21根尖分生组织中的细胞过早地从有丝分裂周期进入内复制周期, 细胞周期G2/M期转换相关基因CYCB;1、CDKB1和CDKB2等的表达也显著降低(Huang et al., 2009; Ishida et al., 2009).Liu等(2016)发现AtMMS21与细胞周期调控因子DPa相互作用并介导其SUMO化修饰, 同时AtMMS21可以恢复E2Fa/DPa双过表达植物的发育缺陷, 进一步研究表明AtMMS21通过SUMO化及物理竞争作用导致DPa与E2Fa的结合能力下降, 影响E2Fa/DPa复合物的核定位, 负调控G1/S期转换, 进而调节根分生组织中细胞分裂和分化的平衡. ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
7 2012
... SP-RING (Siz/Pias)家族是第一个被发现的SUMO E3连接酶家族, SIZ1和MMS21都属于这个家族, 它们的C端都具有1个高度保守的SP-RING结构域.这个结构域与泛素RING结构域相似, 但SP-RING结构域只有1个与锌离子成环的位点, 另1个位点则被氢键和范德华力所代替, 从而维持整个建构的稳定性(图1) (Johnson and Gupta, 2001; Yunus and Lima, 2009; Ishida et al., 2012).植物中有2个SUMO E3连接酶, 其中MMS21较小, 只含有1个SP-RING结 构域; 而SIZ1除了包括SP-RING结构域之外, 还包含 4个保守的结构域: N端的SAP (scaffold attachment factor A/B/acinus/PIAS)结构域、PINIT (Pro-Ile-Asn-Ile- Thr)基序、SXS (Ser-X-Ser; X代表任意氨基酸)结构域以及一段植物特有的锌指结构域PHD (plant homeodomain) (Miura et al., 2007a; Cheong et al., 2009).Cheong等(2009)分别突变AtSIZ1的结构域, 发现5个结构域行使不同的功能, 其中SP-RING结构域赋予AtSIZ1连接酶催化的功能, 决定AtSIZ1的亚细胞定位和控制水杨酸(SA)水平, 同时也调节AtSIZ1独立于SA信号途径的热胁迫响应; PHD结构域也参与连接酶催化底物连接的过程, 但并不是SUMO化所必需的结构, 其可能与PINIT结构域共同调控SUMO与糖和光响应中下胚轴伸长的信号途径; SXS基序可能参与SIZ1介导的ABA信号途径; SAP结构域在酵母中与DNA结合相关, 但在植物中的功能尚未明确. ... ... 植物SUMO E3连接酶的保守结构域 (A) 植物SUMO E3连接酶MMS21和SIZ1的特征结构域.SIZ1具有SAP、PHD、PINIT、SP-RING和SXS结构域, MMS21只具有SP-RING结构域(Ishida et al., 2012); (B) SUMO E3连接酶的SP-RING结构域和泛素E3连接酶的RING结构域构型.RING结构域有2个锌离子结合环, SP-RING只含有1个, 第2个环被范德华力和氢键所替代(Duan et al., 2009; Yunus and Lima, 2009; Ishida et al., 2012; 石田喬志和杉本慶子, 2012); (C), (D) 分别为植物SIZ1和MMS21的同源序列比对.数据来源于NCBI.由DNAMAN定义的氨基酸序列、保守序列(黑色)和相似序列(灰色).SIZ1的蛋白质氨基酸序列为拟南芥(Arabidopsis thaliana) AtSIZ1 (AAU00414.1)、大豆(Glycine max) GmSIZ1a (KRH24918.1)和GmSIZ1b (KRG89029.1)、水稻(Oryza sativa) OsSIZ1 (BAG97182.1)和OsSIZ2 (BAG89374.1)以及玉米(Zea mays) ZmSIZ1a (AQK95338.1)、ZmSIZ1b (AQK79048.1)和ZmSIZc (ONM41936.1).MMS21的蛋白质氨基酸序列为拟南芥AtMMS21 (NP_188133.2)、大豆GmMMS21 (XP_003541835.1)、水稻OsMMS21 (XP_015640264.1)和玉米ZmMMS21 (AQK88509.1). ... ... ; Ishida et al., 2012; 石田喬志和杉本慶子, 2012); (C), (D) 分别为植物SIZ1和MMS21的同源序列比对.数据来源于NCBI.由DNAMAN定义的氨基酸序列、保守序列(黑色)和相似序列(灰色).SIZ1的蛋白质氨基酸序列为拟南芥(Arabidopsis thaliana) AtSIZ1 (AAU00414.1)、大豆(Glycine max) GmSIZ1a (KRH24918.1)和GmSIZ1b (KRG89029.1)、水稻(Oryza sativa) OsSIZ1 (BAG97182.1)和OsSIZ2 (BAG89374.1)以及玉米(Zea mays) ZmSIZ1a (AQK95338.1)、ZmSIZ1b (AQK79048.1)和ZmSIZc (ONM41936.1).MMS21的蛋白质氨基酸序列为拟南芥AtMMS21 (NP_188133.2)、大豆GmMMS21 (XP_003541835.1)、水稻OsMMS21 (XP_015640264.1)和玉米ZmMMS21 (AQK88509.1). ... ... The domains are conserved in plant SUMO E3 ligases (A) Characteristic domains of plant SUMO E3 ligases. MMS21 and SIZ1 are shown by boxes, SIZ1 possesses SAP, PHD, PINIT, SP-RING and SXS domains, while MMS21 possesses only SP-RING domain (Ishida et al., 2012); (B) A schematic model of the SP-RING domain of SUMO E3 ligases and the RING finger of ubiquitin E3 ligases. The RING domain sports two zinc-coordinating loops, the SP-RING domain contains only one, the second loop is instead held together by hydrogen bonds and Van der Waals forces (Duan et al., 2009; Yunus and Lima, 2009; Ishida et al., 2012); (C), (D) The sequence data for the plant SIZ1 homologues were obtained from the NCBI protein database. Sequence identities (black boxes) and similarities (gray boxes) of amino acids were identified by DNAMAN. Amino acid sequences of SIZ1 proteins are from Arabidopsis thaliana AtSIZ1 (AAU00414.1), Glycine max GmSIZ1a (KRH24918.1), GmSIZ1b (KRG89029.1), Oryza sativa OsSIZ1 (BAG97182.1), OsSIZ2 (BAG89374.1), and Zea mays ZmSIZ1a (AQK95338.1), ZmSIZ1b (AQK79048.1), ZmSIZc (ONM41936.1). Amino acid sequences of MMS21 proteins are from Arabidopsis thaliana AtMMS21 (NP_188133.2), Glycine max GmMMS21 (XP_003541835.1), Oryza sativa OsMMS21 (XP_015640264.1), and Zea mays ZmMMS21 (AQK88509.1). ... ... ; Ishida et al., 2012); (C), (D) The sequence data for the plant SIZ1 homologues were obtained from the NCBI protein database. Sequence identities (black boxes) and similarities (gray boxes) of amino acids were identified by DNAMAN. Amino acid sequences of SIZ1 proteins are from Arabidopsis thaliana AtSIZ1 (AAU00414.1), Glycine max GmSIZ1a (KRH24918.1), GmSIZ1b (KRG89029.1), Oryza sativa OsSIZ1 (BAG97182.1), OsSIZ2 (BAG89374.1), and Zea mays ZmSIZ1a (AQK95338.1), ZmSIZ1b (AQK79048.1), ZmSIZc (ONM41936.1). Amino acid sequences of MMS21 proteins are from Arabidopsis thaliana AtMMS21 (NP_188133.2), Glycine max GmMMS21 (XP_003541835.1), Oryza sativa OsMMS21 (XP_015640264.1), and Zea mays ZmMMS21 (AQK88509.1). ... ... 2个植物SUMO E3连接酶除了结构域的差异之外, 其功能和分布也大不相同.AtSIZ1主要在叶片、叶柄、根尖、花絮、雄蕊和柱头中表达(Catala et al., 2007), 主要通过调节SA水平参与植物逆境胁迫响应, 如磷酸饥饿响应、耐盐、耐高温及耐冷等性状(Miura et al., 2005, 2007b; Yoo et al., 2006; Catala et al., 2007).AtMMS21则主要在花药、叶脉和下胚轴分生组织的增殖细胞中表达(Ishida et al., 2012), 参与细胞周期进程和分生组织的维持(Ishida et al., 2009; Huang et al., 2009).两者的突变体分别出现多效性表型, 实验表明双突体atsiz1/atmms21胚胎致死, 同时AtSIZ1不能互补突变体atmms21的表型, 而AtMMS21也无法互补突变体atsiz1的表型(Ishida et al., 2012), 说明2个连接酶在功能上相互独立, 共同参与SUMO化对植物生长发育的精细调控. ... ... 的表型(Ishida et al., 2012), 说明2个连接酶在功能上相互独立, 共同参与SUMO化对植物生长发育的精细调控. ...
5 2008
... 开花是植物由营养生长过渡到生殖生长的重要转折点.突变体atsiz1和atmms21在短日照下都具有明显的早花表型(Jin et al., 2008; Kwak et al., 2016).Jin等(2008)发现拟南芥突变体atsiz1中的SA水平升高, FLC的mRNA水平降低, 表达nahG基因(降低SA水平)可以抑制atsiz1的早花表型, 所以AtSIZ1可以通过调节SA水平调节开花途径.同时FLC也是一个SUMO化修饰的底物, AtSIZ1和AtMMS21都与其相互作用, 共同调节FLC的稳定性: 即AtMMS21介导FLC的SUMO化修饰, 正调控FLC的功能; AtSIZ1抑制FLC的SUMO化, 从而负调控FLC (Kwak et al., 2016).此外, AtSIZ1介导FLD的SUMO化, 进而抑制FLC染色质区域组蛋白H4的去乙酰化, 从而抑制了FLC的表达(Jin et al., 2008).在石斛中也发现DnSIZ1可以通过诱导春化途径调节开花(Liu et al., 2015).以上实验证据表明, SUMO E3连接酶介导的SUMO化修饰从多个层次对FLC依赖的开花途径进行调节. ... ... ).Jin等(2008)发现拟南芥突变体atsiz1中的SA水平升高, FLC的mRNA水平降低, 表达nahG基因(降低SA水平)可以抑制atsiz1的早花表型, 所以AtSIZ1可以通过调节SA水平调节开花途径.同时FLC也是一个SUMO化修饰的底物, AtSIZ1和AtMMS21都与其相互作用, 共同调节FLC的稳定性: 即AtMMS21介导FLC的SUMO化修饰, 正调控FLC的功能; AtSIZ1抑制FLC的SUMO化, 从而负调控FLC (Kwak et al., 2016).此外, AtSIZ1介导FLD的SUMO化, 进而抑制FLC染色质区域组蛋白H4的去乙酰化, 从而抑制了FLC的表达(Jin et al., 2008).在石斛中也发现DnSIZ1可以通过诱导春化途径调节开花(Liu et al., 2015).以上实验证据表明, SUMO E3连接酶介导的SUMO化修饰从多个层次对FLC依赖的开花途径进行调节. ... ... ).此外, AtSIZ1介导FLD的SUMO化, 进而抑制FLC染色质区域组蛋白H4的去乙酰化, 从而抑制了FLC的表达(Jin et al., 2008).在石斛中也发现DnSIZ1可以通过诱导春化途径调节开花(Liu et al., 2015).以上实验证据表明, SUMO E3连接酶介导的SUMO化修饰从多个层次对FLC依赖的开花途径进行调节. ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
1 1997
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
... 泛素E3连接酶COP1是植物光形态建成的关键调控因子, 拟南芥突变体cop1出现光生长抑制, 即使在黑暗条件下也会出现下胚轴短的表型.与突变体cop1相似, 突变体atsiz1幼苗也会在黑暗或白光、红光、蓝光和远红光下出现下胚轴短的表型, 该表型不能通过表达nahG (降低SA水平)恢复, 说明AtSIZ1对光形态建成的调节独立于SA信号途径.进一步研究发现, AtSIZ1可以介导COP1 (K193)的SUMO化修饰.突变体atsiz1中过表达COP1或是在野生型中过表达COP1 K193 (突变其SUMO化修饰位点)与过表达完整的COP表型不一致.这是由于AtSIZ1介导COP1的SUMO化增强了COP1的泛素E3连接酶活性, 导致COP1对HY5的泛素化修饰水平增加, 从而促进HY5降解.另一方面, COP1促进AtSIZ1的泛素化修饰, 导致AtSIZ1通过26S蛋白酶体降解.光照条件下, COP1的SUMO化水平下降, 其活性受到抑制.因此, COP1和AtSIZ1之间形成了一个光形态建成的反馈调节机制(Kim et al., 2016a; Lin et al., 2016). ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
1 2016
... 与AtMMS21不同, AtSIZ1在花、萼片和雌蕊中的表达量最高.Ling等(2012)发现, 拟南芥突变体atsiz1雌蕊中有23%的胚珠是干瘪的, 花粉管萌发及花粉管进入珠孔的过程都正常, 但是由于雌配子发育异常导致部分配子在最后阶段无法完成受精(Ling et al., 2012), 进而结实率降低(Kim et al., 2015, 2016b).由此表明, AtSIZ1是拟南芥雌配子正常发育和种子萌发所必需.而水稻ossiz1突变体则出现花药不开裂和结实率下降的表型(Wang et al., 2015).Thangasamy等(2001)推断OsSIZ1可能通过调节药室内壁发育而影响花药的开裂.这表明SUMO E3连接酶在不同植物中的功能有所差异. ...
1 2015
... 与AtMMS21不同, AtSIZ1在花、萼片和雌蕊中的表达量最高.Ling等(2012)发现, 拟南芥突变体atsiz1雌蕊中有23%的胚珠是干瘪的, 花粉管萌发及花粉管进入珠孔的过程都正常, 但是由于雌配子发育异常导致部分配子在最后阶段无法完成受精(Ling et al., 2012), 进而结实率降低(Kim et al., 2015, 2016b).由此表明, AtSIZ1是拟南芥雌配子正常发育和种子萌发所必需.而水稻ossiz1突变体则出现花药不开裂和结实率下降的表型(Wang et al., 2015).Thangasamy等(2001)推断OsSIZ1可能通过调节药室内壁发育而影响花药的开裂.这表明SUMO E3连接酶在不同植物中的功能有所差异. ...
4 2016
... 开花是植物由营养生长过渡到生殖生长的重要转折点.突变体atsiz1和atmms21在短日照下都具有明显的早花表型(Jin et al., 2008; Kwak et al., 2016).Jin等(2008)发现拟南芥突变体atsiz1中的SA水平升高, FLC的mRNA水平降低, 表达nahG基因(降低SA水平)可以抑制atsiz1的早花表型, 所以AtSIZ1可以通过调节SA水平调节开花途径.同时FLC也是一个SUMO化修饰的底物, AtSIZ1和AtMMS21都与其相互作用, 共同调节FLC的稳定性: 即AtMMS21介导FLC的SUMO化修饰, 正调控FLC的功能; AtSIZ1抑制FLC的SUMO化, 从而负调控FLC (Kwak et al., 2016).此外, AtSIZ1介导FLD的SUMO化, 进而抑制FLC染色质区域组蛋白H4的去乙酰化, 从而抑制了FLC的表达(Jin et al., 2008).在石斛中也发现DnSIZ1可以通过诱导春化途径调节开花(Liu et al., 2015).以上实验证据表明, SUMO E3连接酶介导的SUMO化修饰从多个层次对FLC依赖的开花途径进行调节. ... ... 的早花表型, 所以AtSIZ1可以通过调节SA水平调节开花途径.同时FLC也是一个SUMO化修饰的底物, AtSIZ1和AtMMS21都与其相互作用, 共同调节FLC的稳定性: 即AtMMS21介导FLC的SUMO化修饰, 正调控FLC的功能; AtSIZ1抑制FLC的SUMO化, 从而负调控FLC (Kwak et al., 2016).此外, AtSIZ1介导FLD的SUMO化, 进而抑制FLC染色质区域组蛋白H4的去乙酰化, 从而抑制了FLC的表达(Jin et al., 2008).在石斛中也发现DnSIZ1可以通过诱导春化途径调节开花(Liu et al., 2015).以上实验证据表明, SUMO E3连接酶介导的SUMO化修饰从多个层次对FLC依赖的开花途径进行调节. ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
3 2016
... 泛素E3连接酶COP1是植物光形态建成的关键调控因子, 拟南芥突变体cop1出现光生长抑制, 即使在黑暗条件下也会出现下胚轴短的表型.与突变体cop1相似, 突变体atsiz1幼苗也会在黑暗或白光、红光、蓝光和远红光下出现下胚轴短的表型, 该表型不能通过表达nahG (降低SA水平)恢复, 说明AtSIZ1对光形态建成的调节独立于SA信号途径.进一步研究发现, AtSIZ1可以介导COP1 (K193)的SUMO化修饰.突变体atsiz1中过表达COP1或是在野生型中过表达COP1 K193 (突变其SUMO化修饰位点)与过表达完整的COP表型不一致.这是由于AtSIZ1介导COP1的SUMO化增强了COP1的泛素E3连接酶活性, 导致COP1对HY5的泛素化修饰水平增加, 从而促进HY5降解.另一方面, COP1促进AtSIZ1的泛素化修饰, 导致AtSIZ1通过26S蛋白酶体降解.光照条件下, COP1的SUMO化水平下降, 其活性受到抑制.因此, COP1和AtSIZ1之间形成了一个光形态建成的反馈调节机制(Kim et al., 2016a; Lin et al., 2016). ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
2 2012
... 与AtMMS21不同, AtSIZ1在花、萼片和雌蕊中的表达量最高.Ling等(2012)发现, 拟南芥突变体atsiz1雌蕊中有23%的胚珠是干瘪的, 花粉管萌发及花粉管进入珠孔的过程都正常, 但是由于雌配子发育异常导致部分配子在最后阶段无法完成受精(Ling et al., 2012), 进而结实率降低(Kim et al., 2015, 2016b).由此表明, AtSIZ1是拟南芥雌配子正常发育和种子萌发所必需.而水稻ossiz1突变体则出现花药不开裂和结实率下降的表型(Wang et al., 2015).Thangasamy等(2001)推断OsSIZ1可能通过调节药室内壁发育而影响花药的开裂.这表明SUMO E3连接酶在不同植物中的功能有所差异. ... ... 雌蕊中有23%的胚珠是干瘪的, 花粉管萌发及花粉管进入珠孔的过程都正常, 但是由于雌配子发育异常导致部分配子在最后阶段无法完成受精(Ling et al., 2012), 进而结实率降低(Kim et al., 2015, 2016b).由此表明, AtSIZ1是拟南芥雌配子正常发育和种子萌发所必需.而水稻ossiz1突变体则出现花药不开裂和结实率下降的表型(Wang et al., 2015).Thangasamy等(2001)推断OsSIZ1可能通过调节药室内壁发育而影响花药的开裂.这表明SUMO E3连接酶在不同植物中的功能有所差异. ...
2 2015
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ... ... 开花是植物由营养生长过渡到生殖生长的重要转折点.突变体atsiz1和atmms21在短日照下都具有明显的早花表型(Jin et al., 2008; Kwak et al., 2016).Jin等(2008)发现拟南芥突变体atsiz1中的SA水平升高, FLC的mRNA水平降低, 表达nahG基因(降低SA水平)可以抑制atsiz1的早花表型, 所以AtSIZ1可以通过调节SA水平调节开花途径.同时FLC也是一个SUMO化修饰的底物, AtSIZ1和AtMMS21都与其相互作用, 共同调节FLC的稳定性: 即AtMMS21介导FLC的SUMO化修饰, 正调控FLC的功能; AtSIZ1抑制FLC的SUMO化, 从而负调控FLC (Kwak et al., 2016).此外, AtSIZ1介导FLD的SUMO化, 进而抑制FLC染色质区域组蛋白H4的去乙酰化, 从而抑制了FLC的表达(Jin et al., 2008).在石斛中也发现DnSIZ1可以通过诱导春化途径调节开花(Liu et al., 2015).以上实验证据表明, SUMO E3连接酶介导的SUMO化修饰从多个层次对FLC依赖的开花途径进行调节. ...
... 除了启动DNA修复机制, 细胞还可以通过严格控制细胞周期进程对DSB进行响应.突变体atmms21根尖分生组织中的细胞过早地从有丝分裂周期进入内复制周期, 细胞周期G2/M期转换相关基因CYCB;1、CDKB1和CDKB2等的表达也显著降低(Huang et al., 2009; Ishida et al., 2009).Liu等(2016)发现AtMMS21与细胞周期调控因子DPa相互作用并介导其SUMO化修饰, 同时AtMMS21可以恢复E2Fa/DPa双过表达植物的发育缺陷, 进一步研究表明AtMMS21通过SUMO化及物理竞争作用导致DPa与E2Fa的结合能力下降, 影响E2Fa/DPa复合物的核定位, 负调控G1/S期转换, 进而调节根分生组织中细胞分裂和分化的平衡. ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
... 2个植物SUMO E3连接酶除了结构域的差异之外, 其功能和分布也大不相同.AtSIZ1主要在叶片、叶柄、根尖、花絮、雄蕊和柱头中表达(Catala et al., 2007), 主要通过调节SA水平参与植物逆境胁迫响应, 如磷酸饥饿响应、耐盐、耐高温及耐冷等性状(Miura et al., 2005, 2007b; Yoo et al., 2006; Catala et al., 2007).AtMMS21则主要在花药、叶脉和下胚轴分生组织的增殖细胞中表达(Ishida et al., 2012), 参与细胞周期进程和分生组织的维持(Ishida et al., 2009; Huang et al., 2009).两者的突变体分别出现多效性表型, 实验表明双突体atsiz1/atmms21胚胎致死, 同时AtSIZ1不能互补突变体atmms21的表型, 而AtMMS21也无法互补突变体atsiz1的表型(Ishida et al., 2012), 说明2个连接酶在功能上相互独立, 共同参与SUMO化对植物生长发育的精细调控. ...
3 2011
... 外界环境中营养元素水平的波动对根的形态建成有极大的影响.磷酸盐是根吸收和同化磷的主要形式.缺磷条件下, 植物会通过减少主根的生长而增加侧根的伸长来增加根系与土壤的接触面积, 从而获取更多磷(Dong et al., 2017).突变体atsiz1出现缺磷超敏感表型: 主根变短, 侧根增加和伸长, 根毛发达.一方面AtSIZ1可以介导调控低磷反应关键因子PHR1的SUMO化修饰, 进而正调控磷酸饥饿响应基因AtPS1和AtRNS1的表达以增加磷的吸收(Miura et al., 2005; 杨辉霞等, 2007).另一方面, Miura等(2011)通过生长素响应因子DR5启动的GUS系统来检测根系中生长素的分布, 发现缺磷后根系的重建与生长素在根系中分布变化相关, 进一步揭示AtSIZ1也可以通过调控生长素积累负调控缺磷响应中的根形态建成.Wang等(2015)在水稻中也发现, 磷胁迫减弱了OsSIZ1缺失增强的根系生长素输出能力.说明SIZ1在低磷条件下对根构型的重建具有重要作用. ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
3 2009
... 此外, SIZ1还通过调节脱落酸(ABA)和糖代谢途径参与根系调节(Castro et al., 2015).拟南芥突变体atsiz1对ABA超敏感且主根生长受抑制.研究发现这是由于AtSIZ1介导了ABA信号相关bZIP转录因子ABI5和MYB30的SUMO化, 从而影响ABA对幼苗初生根的调节与种子的萌发(Miura et al., 2009; Zheng et al., 2012).外源糖会促进体内SUMO分子的积累, 同时AtSIZ1负调节淀粉和糖代谢相关基因的转录水平而影响初生根的发育(Castro et al., 2015). ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
6 2005
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ... ... ) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ... ... 2个植物SUMO E3连接酶除了结构域的差异之外, 其功能和分布也大不相同.AtSIZ1主要在叶片、叶柄、根尖、花絮、雄蕊和柱头中表达(Catala et al., 2007), 主要通过调节SA水平参与植物逆境胁迫响应, 如磷酸饥饿响应、耐盐、耐高温及耐冷等性状(Miura et al., 2005, 2007b; Yoo et al., 2006; Catala et al., 2007).AtMMS21则主要在花药、叶脉和下胚轴分生组织的增殖细胞中表达(Ishida et al., 2012), 参与细胞周期进程和分生组织的维持(Ishida et al., 2009; Huang et al., 2009).两者的突变体分别出现多效性表型, 实验表明双突体atsiz1/atmms21胚胎致死, 同时AtSIZ1不能互补突变体atmms21的表型, 而AtMMS21也无法互补突变体atsiz1的表型(Ishida et al., 2012), 说明2个连接酶在功能上相互独立, 共同参与SUMO化对植物生长发育的精细调控. ... ... 外界环境中营养元素水平的波动对根的形态建成有极大的影响.磷酸盐是根吸收和同化磷的主要形式.缺磷条件下, 植物会通过减少主根的生长而增加侧根的伸长来增加根系与土壤的接触面积, 从而获取更多磷(Dong et al., 2017).突变体atsiz1出现缺磷超敏感表型: 主根变短, 侧根增加和伸长, 根毛发达.一方面AtSIZ1可以介导调控低磷反应关键因子PHR1的SUMO化修饰, 进而正调控磷酸饥饿响应基因AtPS1和AtRNS1的表达以增加磷的吸收(Miura et al., 2005; 杨辉霞等, 2007).另一方面, Miura等(2011)通过生长素响应因子DR5启动的GUS系统来检测根系中生长素的分布, 发现缺磷后根系的重建与生长素在根系中分布变化相关, 进一步揭示AtSIZ1也可以通过调控生长素积累负调控缺磷响应中的根形态建成.Wang等(2015)在水稻中也发现, 磷胁迫减弱了OsSIZ1缺失增强的根系生长素输出能力.说明SIZ1在低磷条件下对根构型的重建具有重要作用. ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
1 1999
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
1 2010
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
1 2017
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
1 2005
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
1 2016
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
1 2011
... 与AtMMS21不同, AtSIZ1在花、萼片和雌蕊中的表达量最高.Ling等(2012)发现, 拟南芥突变体atsiz1雌蕊中有23%的胚珠是干瘪的, 花粉管萌发及花粉管进入珠孔的过程都正常, 但是由于雌配子发育异常导致部分配子在最后阶段无法完成受精(Ling et al., 2012), 进而结实率降低(Kim et al., 2015, 2016b).由此表明, AtSIZ1是拟南芥雌配子正常发育和种子萌发所必需.而水稻ossiz1突变体则出现花药不开裂和结实率下降的表型(Wang et al., 2015).Thangasamy等(2001)推断OsSIZ1可能通过调节药室内壁发育而影响花药的开裂.这表明SUMO E3连接酶在不同植物中的功能有所差异. ...
2 2015
... 外界环境中营养元素水平的波动对根的形态建成有极大的影响.磷酸盐是根吸收和同化磷的主要形式.缺磷条件下, 植物会通过减少主根的生长而增加侧根的伸长来增加根系与土壤的接触面积, 从而获取更多磷(Dong et al., 2017).突变体atsiz1出现缺磷超敏感表型: 主根变短, 侧根增加和伸长, 根毛发达.一方面AtSIZ1可以介导调控低磷反应关键因子PHR1的SUMO化修饰, 进而正调控磷酸饥饿响应基因AtPS1和AtRNS1的表达以增加磷的吸收(Miura et al., 2005; 杨辉霞等, 2007).另一方面, Miura等(2011)通过生长素响应因子DR5启动的GUS系统来检测根系中生长素的分布, 发现缺磷后根系的重建与生长素在根系中分布变化相关, 进一步揭示AtSIZ1也可以通过调控生长素积累负调控缺磷响应中的根形态建成.Wang等(2015)在水稻中也发现, 磷胁迫减弱了OsSIZ1缺失增强的根系生长素输出能力.说明SIZ1在低磷条件下对根构型的重建具有重要作用. ... ... 与AtMMS21不同, AtSIZ1在花、萼片和雌蕊中的表达量最高.Ling等(2012)发现, 拟南芥突变体atsiz1雌蕊中有23%的胚珠是干瘪的, 花粉管萌发及花粉管进入珠孔的过程都正常, 但是由于雌配子发育异常导致部分配子在最后阶段无法完成受精(Ling et al., 2012), 进而结实率降低(Kim et al., 2015, 2016b).由此表明, AtSIZ1是拟南芥雌配子正常发育和种子萌发所必需.而水稻ossiz1突变体则出现花药不开裂和结实率下降的表型(Wang et al., 2015).Thangasamy等(2001)推断OsSIZ1可能通过调节药室内壁发育而影响花药的开裂.这表明SUMO E3连接酶在不同植物中的功能有所差异. ...
1 2009
... 突变体atmms21对DNA损伤剂超敏感, 根尖干细胞及其子代细胞出现明显的细胞死亡.对酵母(Sac- charomyces cerevisiae)和人类(Homo sapiens)的研究表明, MMS21是染色体结构维持复合体(Structure Maintenance of Chromosomes5/6, SMC5/6)的关键组分, 是促进DNA双链断裂(DNA double-strand break, DSB)修复所必需的(De Piccoli et al., 2006; Watanabe et al., 2009; Almedawar et al., 2012).在拟南芥中, atmms21突变体中DSB的标记基因和DSB修复标记基因都有较强的表达(Xu et al., 2013; Yuan et al., 2014), 说明MMS21在物种间有很强的保守性.因此推测AtMMS21可能通过与SMC5相互作用在DNA双链断裂修复中发挥功能, 从而维持根尖干细胞微环境的稳定以调控根系的正常发育. ...
... 根尖干细胞的命运是由多能转录因子调节的, 如维持干细胞活性的静止中心(quiescent center, QC)特异转录因子WOX5、决定干细胞微环境横向模式建立的SHR和SCR以及调控干细胞微环境纵向模式建立的PLTs.拟南芥突变体atmms21根尖分生组织的细胞分裂和细胞分化模式发生改变(徐庞连等, 2008; Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010), 说明AtMMS21是根尖分生组织的结构与功能维持所必需的.Xu等(2013)通过检测QC特异性标记的表达和根尖干细胞的命运, 发现突变体atmms21根尖中WOX5出现异位表达; SHR和SCR的正常空间表达模式出现部分缺失, 进而呈现不连续表达; PLT1和PLT2在蛋白水平发生严重下调, 进而呈现表达/积累减少的模式.此外, 拟南芥突变体atmms21胚胎中胚根原基的后代细胞出现异常分裂, 导致根尖干细胞微环境起始的位置——胚胎基极发生结构错乱.这些现象表明, AtMMS21在胚胎和胚后发育阶段是根尖QC组织维持及调节多能性转录因子在根尖干细胞微环境中稳定表达所必需的.另一方面, Ishida等(2009)发现PLT1和PLT2反过来对AtMMS21的表达或积累起着正调控作用; Zhang等(2010)发现突变体atmms- 21对外源细胞分裂素的敏感性降低, ARRs基因表达下调.因此可以推断AtMMS21可能是细胞分裂素与生长素相互作用的一个节点, 通过调控植物激素来维持拟南芥根尖干细胞的稳态. ... ... 突变体atmms21对DNA损伤剂超敏感, 根尖干细胞及其子代细胞出现明显的细胞死亡.对酵母(Sac- charomyces cerevisiae)和人类(Homo sapiens)的研究表明, MMS21是染色体结构维持复合体(Structure Maintenance of Chromosomes5/6, SMC5/6)的关键组分, 是促进DNA双链断裂(DNA double-strand break, DSB)修复所必需的(De Piccoli et al., 2006; Watanabe et al., 2009; Almedawar et al., 2012).在拟南芥中, atmms21突变体中DSB的标记基因和DSB修复标记基因都有较强的表达(Xu et al., 2013; Yuan et al., 2014), 说明MMS21在物种间有很强的保守性.因此推测AtMMS21可能通过与SMC5相互作用在DNA双链断裂修复中发挥功能, 从而维持根尖干细胞微环境的稳定以调控根系的正常发育. ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
1 2006
... 2个植物SUMO E3连接酶除了结构域的差异之外, 其功能和分布也大不相同.AtSIZ1主要在叶片、叶柄、根尖、花絮、雄蕊和柱头中表达(Catala et al., 2007), 主要通过调节SA水平参与植物逆境胁迫响应, 如磷酸饥饿响应、耐盐、耐高温及耐冷等性状(Miura et al., 2005, 2007b; Yoo et al., 2006; Catala et al., 2007).AtMMS21则主要在花药、叶脉和下胚轴分生组织的增殖细胞中表达(Ishida et al., 2012), 参与细胞周期进程和分生组织的维持(Ishida et al., 2009; Huang et al., 2009).两者的突变体分别出现多效性表型, 实验表明双突体atsiz1/atmms21胚胎致死, 同时AtSIZ1不能互补突变体atmms21的表型, 而AtMMS21也无法互补突变体atsiz1的表型(Ishida et al., 2012), 说明2个连接酶在功能上相互独立, 共同参与SUMO化对植物生长发育的精细调控. ...
3 2014
... 突变体atmms21对DNA损伤剂超敏感, 根尖干细胞及其子代细胞出现明显的细胞死亡.对酵母(Sac- charomyces cerevisiae)和人类(Homo sapiens)的研究表明, MMS21是染色体结构维持复合体(Structure Maintenance of Chromosomes5/6, SMC5/6)的关键组分, 是促进DNA双链断裂(DNA double-strand break, DSB)修复所必需的(De Piccoli et al., 2006; Watanabe et al., 2009; Almedawar et al., 2012).在拟南芥中, atmms21突变体中DSB的标记基因和DSB修复标记基因都有较强的表达(Xu et al., 2013; Yuan et al., 2014), 说明MMS21在物种间有很强的保守性.因此推测AtMMS21可能通过与SMC5相互作用在DNA双链断裂修复中发挥功能, 从而维持根尖干细胞微环境的稳定以调控根系的正常发育. ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
3 2009
... SP-RING (Siz/Pias)家族是第一个被发现的SUMO E3连接酶家族, SIZ1和MMS21都属于这个家族, 它们的C端都具有1个高度保守的SP-RING结构域.这个结构域与泛素RING结构域相似, 但SP-RING结构域只有1个与锌离子成环的位点, 另1个位点则被氢键和范德华力所代替, 从而维持整个建构的稳定性(图1) (Johnson and Gupta, 2001; Yunus and Lima, 2009; Ishida et al., 2012).植物中有2个SUMO E3连接酶, 其中MMS21较小, 只含有1个SP-RING结 构域; 而SIZ1除了包括SP-RING结构域之外, 还包含 4个保守的结构域: N端的SAP (scaffold attachment factor A/B/acinus/PIAS)结构域、PINIT (Pro-Ile-Asn-Ile- Thr)基序、SXS (Ser-X-Ser; X代表任意氨基酸)结构域以及一段植物特有的锌指结构域PHD (plant homeodomain) (Miura et al., 2007a; Cheong et al., 2009).Cheong等(2009)分别突变AtSIZ1的结构域, 发现5个结构域行使不同的功能, 其中SP-RING结构域赋予AtSIZ1连接酶催化的功能, 决定AtSIZ1的亚细胞定位和控制水杨酸(SA)水平, 同时也调节AtSIZ1独立于SA信号途径的热胁迫响应; PHD结构域也参与连接酶催化底物连接的过程, 但并不是SUMO化所必需的结构, 其可能与PINIT结构域共同调控SUMO与糖和光响应中下胚轴伸长的信号途径; SXS基序可能参与SIZ1介导的ABA信号途径; SAP结构域在酵母中与DNA结合相关, 但在植物中的功能尚未明确. ... ... 植物SUMO E3连接酶的保守结构域 (A) 植物SUMO E3连接酶MMS21和SIZ1的特征结构域.SIZ1具有SAP、PHD、PINIT、SP-RING和SXS结构域, MMS21只具有SP-RING结构域(Ishida et al., 2012); (B) SUMO E3连接酶的SP-RING结构域和泛素E3连接酶的RING结构域构型.RING结构域有2个锌离子结合环, SP-RING只含有1个, 第2个环被范德华力和氢键所替代(Duan et al., 2009; Yunus and Lima, 2009; Ishida et al., 2012; 石田喬志和杉本慶子, 2012); (C), (D) 分别为植物SIZ1和MMS21的同源序列比对.数据来源于NCBI.由DNAMAN定义的氨基酸序列、保守序列(黑色)和相似序列(灰色).SIZ1的蛋白质氨基酸序列为拟南芥(Arabidopsis thaliana) AtSIZ1 (AAU00414.1)、大豆(Glycine max) GmSIZ1a (KRH24918.1)和GmSIZ1b (KRG89029.1)、水稻(Oryza sativa) OsSIZ1 (BAG97182.1)和OsSIZ2 (BAG89374.1)以及玉米(Zea mays) ZmSIZ1a (AQK95338.1)、ZmSIZ1b (AQK79048.1)和ZmSIZc (ONM41936.1).MMS21的蛋白质氨基酸序列为拟南芥AtMMS21 (NP_188133.2)、大豆GmMMS21 (XP_003541835.1)、水稻OsMMS21 (XP_015640264.1)和玉米ZmMMS21 (AQK88509.1). ... ... The domains are conserved in plant SUMO E3 ligases (A) Characteristic domains of plant SUMO E3 ligases. MMS21 and SIZ1 are shown by boxes, SIZ1 possesses SAP, PHD, PINIT, SP-RING and SXS domains, while MMS21 possesses only SP-RING domain (Ishida et al., 2012); (B) A schematic model of the SP-RING domain of SUMO E3 ligases and the RING finger of ubiquitin E3 ligases. The RING domain sports two zinc-coordinating loops, the SP-RING domain contains only one, the second loop is instead held together by hydrogen bonds and Van der Waals forces (Duan et al., 2009; Yunus and Lima, 2009; Ishida et al., 2012); (C), (D) The sequence data for the plant SIZ1 homologues were obtained from the NCBI protein database. Sequence identities (black boxes) and similarities (gray boxes) of amino acids were identified by DNAMAN. Amino acid sequences of SIZ1 proteins are from Arabidopsis thaliana AtSIZ1 (AAU00414.1), Glycine max GmSIZ1a (KRH24918.1), GmSIZ1b (KRG89029.1), Oryza sativa OsSIZ1 (BAG97182.1), OsSIZ2 (BAG89374.1), and Zea mays ZmSIZ1a (AQK95338.1), ZmSIZ1b (AQK79048.1), ZmSIZc (ONM41936.1). Amino acid sequences of MMS21 proteins are from Arabidopsis thaliana AtMMS21 (NP_188133.2), Glycine max GmMMS21 (XP_003541835.1), Oryza sativa OsMMS21 (XP_015640264.1), and Zea mays ZmMMS21 (AQK88509.1). ...
3 2017
... 染色质重塑复合物在植物根系发育相关基因的表达调控方面具有重要作用.在有关酵母和人类的研究报道中, SUMO E3连接酶可以作为转录调节复合物调节基因的表达和染色质重塑(Wotton et al., 2017).Zhang等(2017a)发现AtMMS21可以与染色质重塑复合物BRM相互作用并促进其SUMO化修饰, 维持BRM蛋白的稳定性, 进而调控根发育相关基因的正常表达, 以维持根尖微环境的稳态. ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
1 2016
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
1 2017
... 蛋白质的SUMO化修饰主要发生在赖氨酸残基侧链, 由SUMO激活酶E1、SUMO结合酶E2和SUMO连接酶E3分级作用完成, 去SUMO化则是由SUMO特异性蛋白酶介导完成.首先, Pre-SUMO (SUMO分子的前体形式)在SUMO特异蛋白酶的作用下, 将C端的2个Gly残基暴露, 成为成熟的SUMO分子; SU- MO分子由E1在ATP供能下通过转酯反应被激活(Desterro et al., 1999; Gong et al., 1999; Okuma et al., 1999); 然后转移到E2上并通过硫酯键与E2的活性位点Cys相连; 最后从E2转移到靶蛋白(Desterro et al., 1997; Johnson and Blobel, 1997).目前通过晶体研究发现, 虽然E2可以直接识别含特异序列(ψKXE\D)的靶蛋白, 但是这种相互作用不足以支持SUMO分子转移, 因此需要E2额外的结构或者是E3维持这一过程的稳定.E3募集特异的靶蛋白和E2, 作为桥梁和支架促进SUMO分子从E2向底物转移(Reverter and Lima 2005; Cappadocia et al., 2015; Eisenhardt et al., 2015; Streich Jr and Lima, 2016; Pichler et al., 2017).因此, SUMO E3连接酶在SUMO修饰调节过程中具有重要作用, 赋予了反应的特异性.至今在植物中发现的SUMO E3连接酶主要包括2类: SIZ1 (SAP AND MIZ1) (Miura et al., 2005)和MMS21/HIGH PLOIDY2 (下文简称MMS21) (Hu- ang et al., 2009; Ishida et al., 2009), 其中已进行深入探究的主要包括拟南芥(Arabidopsis thaliana) AtSIZ1 (Miura et al., 2005)和AtMMS21 (Huang et al., 2009; Ishida et al., 2009)、水稻(Oryza sativa) OsSIZ1和OsSIZ2 (Park et al., 2010)、大豆(Glycine max) GmSIZ1a和GmSIZ1b (Cai et al., 2017)、苹果(Malus domestica) MdSIZ1 (Zhang et al., 2016)、石斛(Dendrobium nobile) DnSIZ1 (Liu et al., 2015)和番茄(Solanum lycopersicum) SISIZ1 (Zhang et al., 2017b).本文对SUMO E3连接酶在植物生长发育中的功能进行综述, 对深入理解翻译后修饰对植物生命活动的精细调节具有重要意义. ...
4 2010
... 根尖干细胞的命运是由多能转录因子调节的, 如维持干细胞活性的静止中心(quiescent center, QC)特异转录因子WOX5、决定干细胞微环境横向模式建立的SHR和SCR以及调控干细胞微环境纵向模式建立的PLTs.拟南芥突变体atmms21根尖分生组织的细胞分裂和细胞分化模式发生改变(徐庞连等, 2008; Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010), 说明AtMMS21是根尖分生组织的结构与功能维持所必需的.Xu等(2013)通过检测QC特异性标记的表达和根尖干细胞的命运, 发现突变体atmms21根尖中WOX5出现异位表达; SHR和SCR的正常空间表达模式出现部分缺失, 进而呈现不连续表达; PLT1和PLT2在蛋白水平发生严重下调, 进而呈现表达/积累减少的模式.此外, 拟南芥突变体atmms21胚胎中胚根原基的后代细胞出现异常分裂, 导致根尖干细胞微环境起始的位置——胚胎基极发生结构错乱.这些现象表明, AtMMS21在胚胎和胚后发育阶段是根尖QC组织维持及调节多能性转录因子在根尖干细胞微环境中稳定表达所必需的.另一方面, Ishida等(2009)发现PLT1和PLT2反过来对AtMMS21的表达或积累起着正调控作用; Zhang等(2010)发现突变体atmms- 21对外源细胞分裂素的敏感性降低, ARRs基因表达下调.因此可以推断AtMMS21可能是细胞分裂素与生长素相互作用的一个节点, 通过调控植物激素来维持拟南芥根尖干细胞的稳态. ... ... 的表达或积累起着正调控作用; Zhang等(2010)发现突变体atmms- 21对外源细胞分裂素的敏感性降低, ARRs基因表达下调.因此可以推断AtMMS21可能是细胞分裂素与生长素相互作用的一个节点, 通过调控植物激素来维持拟南芥根尖干细胞的稳态. ... ... SUMO E3连接酶在拟南芥生长发育中的功能 (A) AtMMS21对细胞周期的调节功能(Liu et al., 2016); (B) MMS21和SIZ1对拟南芥开花途径和时间的调控(Kwak et al., 2016); (C) MMS21在拟南芥中响应DNA损伤, 进而影响植物生长发育过程(Xu et al., 2013; Yuan et al., 2014); (D) MMS21通过调节生长素信号途径影响拟南芥根的发育(Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21通过染色质重塑复合物调节根分生组织发育(Zhang et al., 2017a); (F) SIZ1通过调节糖代谢途径和ABA途径影响种子的萌发时间(Miura et al., 2009); (G) SIZ1对开花途径的调节(Jin et al., 2008); (H) SIZ1对植物光形态建成的调控(Kim et al., 2016a; Lin et al., 2016); (I) SIZ1对磷酸饥饿响应的调控(Miura et al., 2005; 2011); (J) SIZ1通过糖代谢途径调节根的发育(Castro et al., 2015) ... ... The functions of SUMO E3 ligases in Arabidopsis (A) The function of MMS21 regulates in cell cycle (Liu et al., 2016); (B) MMS21 and SIZ1 are both involved in flowering pathway and time (Kwak et al., 2016); (C) MMS21 responses to the DNA damage to regulate plant growth and development (Xu et al., 2013; Yuan et al., 2014); (D) MMS21 is involved in Arabidopsis root development by auxin (Ishida et al., 2009; Huang et al., 2009; Zhang et al., 2010); (E) MMS21 is involved in root development by regulating chromatin remodeling complex (Zhang et al., 2017a); (F) SIZ1 regulate glucose metabolism germinated/seedling development (Miura et al., 2009); (G) SIZ1 is involved in flowering pathway (Jin et al., 2008); (H) SIZ1 regulates the photomorphogenesis (Kim et al., 2016a; Lin et al., 2016); (I) SIZ1 regulation of phosphate starvation-induced root architecture remodeling involves the control of auxin accumulation (Miura et al., 2005, 2011); (J) SIZ1 regulates development of root by sugar pathway (Castro et al., 2015) ...
1 2012
... 此外, SIZ1还通过调节脱落酸(ABA)和糖代谢途径参与根系调节(Castro et al., 2015).拟南芥突变体atsiz1对ABA超敏感且主根生长受抑制.研究发现这是由于AtSIZ1介导了ABA信号相关bZIP转录因子ABI5和MYB30的SUMO化, 从而影响ABA对幼苗初生根的调节与种子的萌发(Miura et al., 2009; Zheng et al., 2012).外源糖会促进体内SUMO分子的积累, 同时AtSIZ1负调节淀粉和糖代谢相关基因的转录水平而影响初生根的发育(Castro et al., 2015). ...

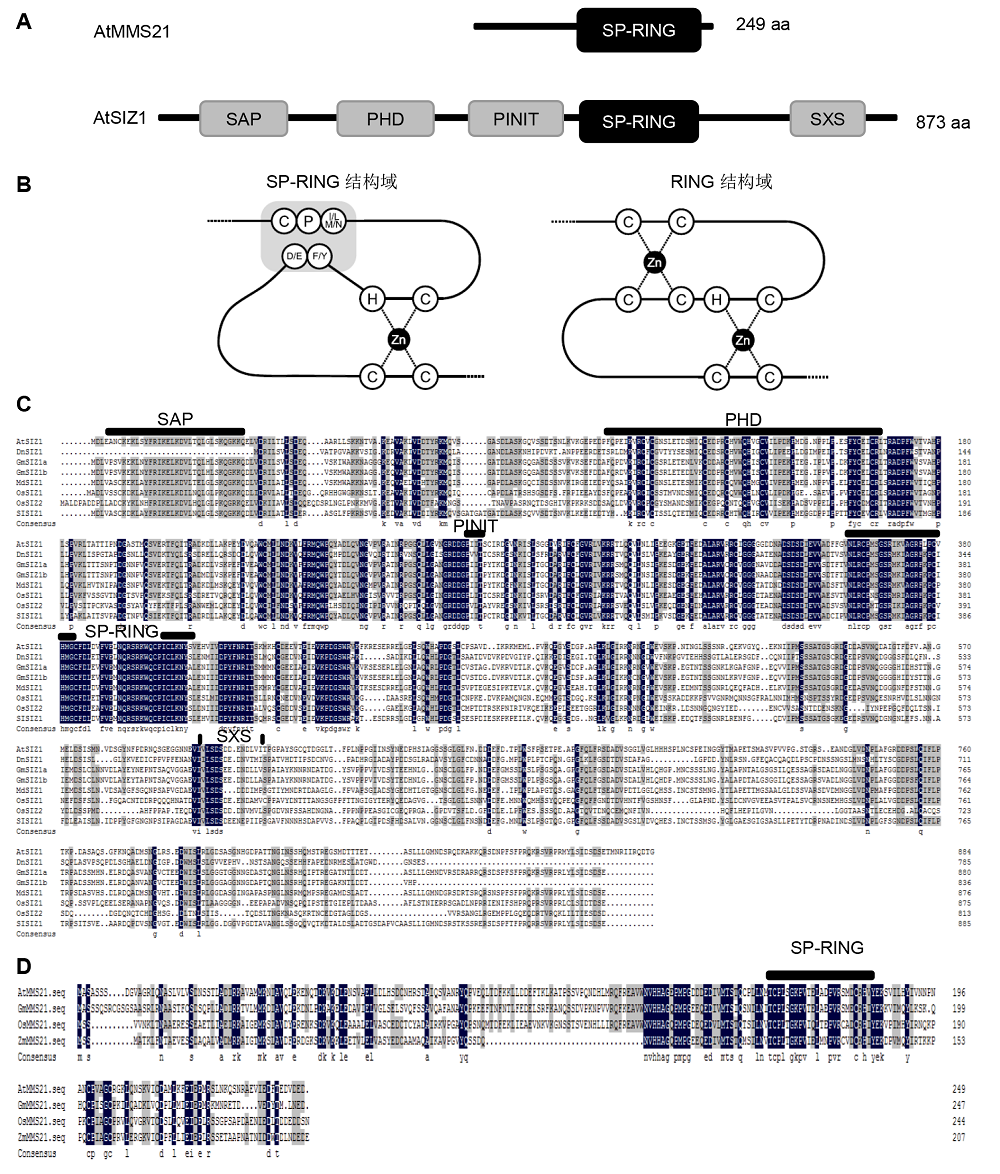
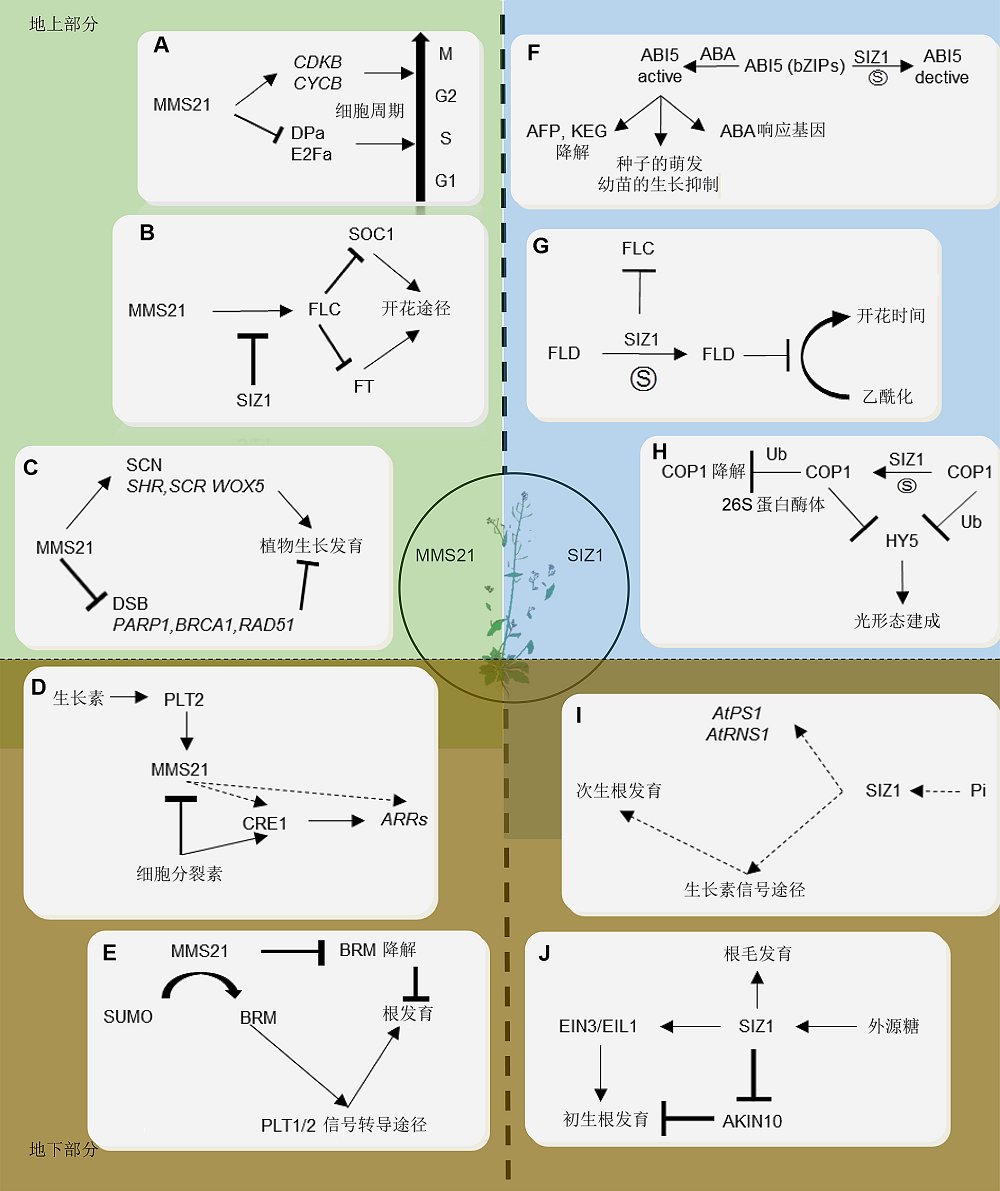

{kind=link}
{kind=link}
{kind=link}
{kind=link}