Abstract: The coordination between the maintenance of stem-cell niches and differentiation of their progeny sets the developmental basis of higher multiple-cellular organisms to ensure coherent organ growth. The plant hormone auxin plays important roles in the morphological establishment of shoot and root apical meristem mediated by the maintenance of shoot and root stem-cell niches. Here we review multiple signaling modules discovered in recent years, including the PLETHORA pathway, SCARECROW-SHORT ROOT pathway and environmental factors, for their mechanisms in maintaining root stem cell niches. Auxin participates in these mechanisms by regulating local synthesis, polar transport and signal transduction. By these means, auxin concentration gradient can be maintained accurately in stem-cell niches around the root apical quiescent center, for balancing the proliferation and differentiation of stem-cell niches.
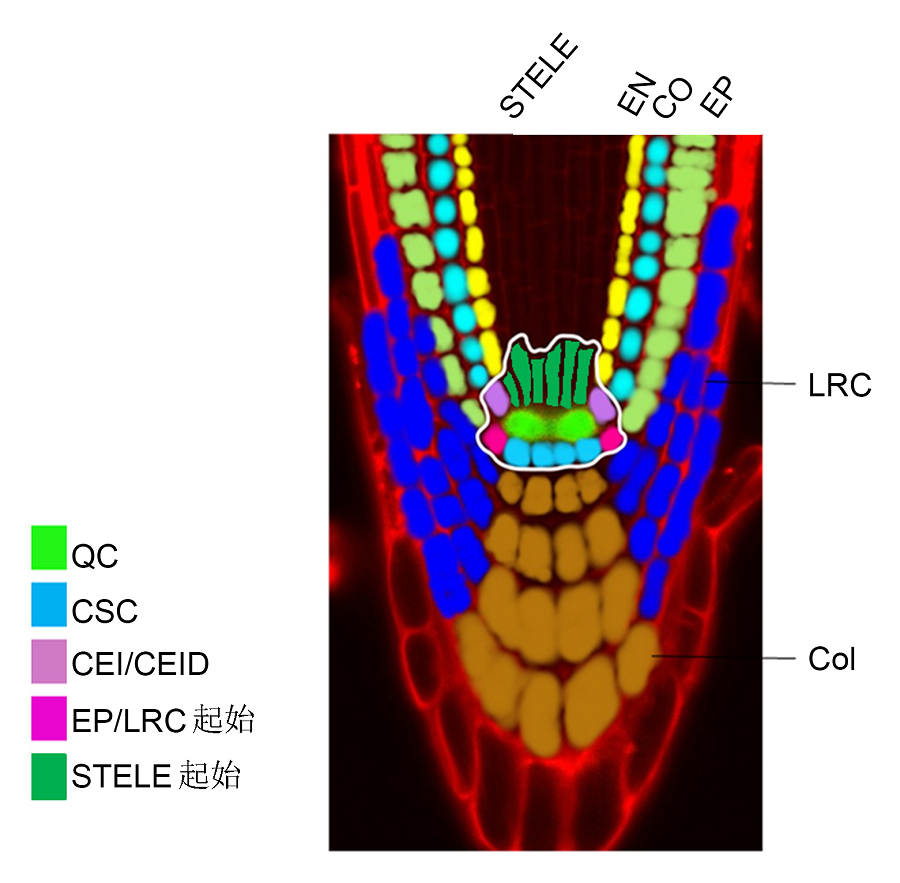
5.3 干细胞维持的空间协调性植物以及一些动物有从成熟的器官中再生新器官的能力。关于植物干细胞的再生能力, 前人也做了一些探索与研究(Xu et al., 2006; Sena et al., 2009)。Ben Scheres实验室利用激光敲除技术将QC切除, 发现切除QC会导致干细胞巢区从原维管组织中重新获得特异性(Xu et al., 2006)。QC切除后, 生长素积累转移, 接着是PLT1的重定位, 且下调PIN的表达。PLTs基因促进SHR核定位, 进而促进SCR以及PLTs基因的表达, 最终新的QC得以建立。当新的细胞特异性产生后, PIN蛋白重新出现并进行正确的极性定位。QC被切除后, plt1/plt2突变体由于根尖生长素最大浓度建立出现缺陷而不能产生新的QC和根冠细胞。在突变体shr和scr中, 一个新的生长素最大浓度在一定区域产生, 紧接着根冠重新形成, 但是QC并没有形成, 且WOX5的表达丢失。这些初步结果都表明PLTs、SHR和SCR对于根尖干细胞巢的命运以及再生能力可能发挥重要作用。 植物依赖顶端干细胞巢的维持来进行根尖以及茎尖的持续生长。虽然植物细胞发育的可塑性已经得到证明, 但是植物干细胞巢区对于器官再生的影响还不清楚, 生长素被认为是其中的一个重要因子。研究者利用切除部分分生区(包括干细胞巢)的根尖再生系统来追踪细胞命运恢复的分子功能, 发现丢失的细胞识别特异性在几小时内就能恢复, 而功能特异性细胞在1天之内也能恢复(Sena et al., 2009)。根尖分生区切除5小时之内, 切口处细胞特异性开始重新建立, 超过50%的根冠小柱细胞的特异表达基因开始重新表达。切除1天后, 尽管只有四分之一QC富集的基因恢复表达, 然而通过淀粉染色及重力响应实验, 已经能观察到分化的小柱细胞, 与此同时WOX5仍然异常表达, 且干细胞巢的形态仍未恢复。意外的是, 无论是plt1/plt2双突变体或者scr突变体, 均能修复根尖分生区(包括完整根尖干细胞巢)的分离损伤。这些实验表明干细胞巢活性对于早期根尖重建不是必需的。 在根尖分生组织中, 干细胞活性和细胞分化间存在空间协调性。已有研究表明, 生长素与细胞分裂素相互作用来协调根尖分生区和伸长区的转变, 生长素促进细胞分裂而细胞分裂素促进细胞分化(Blilou et al., 2005; Dello Ioio et al., 2007)。在拟南芥根尖转换区(TZ), 细胞分裂素可以通过细胞分裂素受体(Ara- bidopsis histidine kinase, AHK3)来激活2个细胞分裂素主要响应转录因子ARR1 (Arabidopsis response regulators 1)和ARR12 (Hwang et al., 2012)。这些基因又能激活SHY2 (SHORT HYPOCOTYL2)基因的转录, SHY2是生长素信号的抑制子(Dello Ioio et al., 2008)。反之SHY2负调节PIN基因的表达, 从而限制生长素的运输和分布, 进而导致细胞分化(Dello Ioio et al., 2008; Moubayidin et al., 2010)。 最新研究揭示了SCR起空间协调作用的分子机理。在静止中心, SCR直接抑制ARR1的表达, ARR1可诱导ASB1 (ANTHRANILATE SYNTHASE BETA SUBUNIT 1 )基因的转录, 所以SCR可直接在干细胞区抑制生长素的合成; 而在TZ区SCR抑制ARR1的作用得到解除, ARR1蛋白的有效表达促进SHY2的表达, 进而促进TZ区细胞的分化(Moubayidin et al., 2013)。通过这种方法, SCR同时控制干细胞的分裂和分化, 确保根的生长。在scr突变体中, ARR1异常积累, 导致静止中心生长素水平及响应能力升高(Mou- bayidin et al., 2013)。这一发现与高浓度生长素能促进干细胞的分化一致。
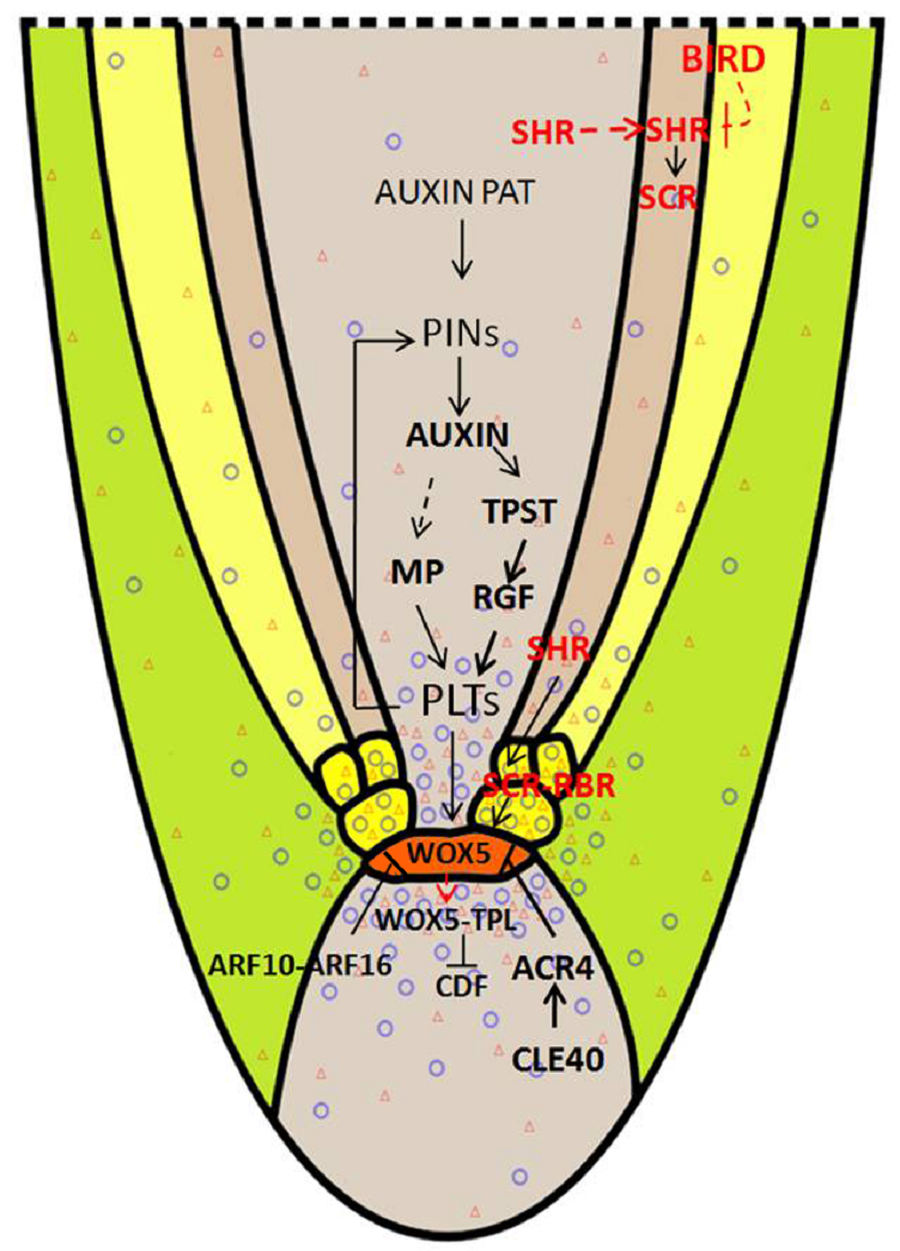
6 结论和展望目前, 研究表明多种信号途径参与拟南芥根尖及茎尖干细胞巢区的定义及维持。根尖干细胞巢维持的途径主要有2条, 一条是PLT途径, 另一条是SHR-SCR途径。其中, 生长素浓度梯度的积累在这2条途径中起着非常重要的作用。在植物界, 尽管根尖的进化晚于茎尖, 但在根尖及茎尖干细胞区维持上却有相似的保守机制。研究表明, WUS定义了组织中心并且自身也作为一个信号分子, 通过在组织中心移动来控制上层干细胞, 且同一研究也揭示出WUS-CLV3/CLV1对于茎尖干细胞巢维持的反馈调节机制(Brand et al., 2000)。另外, 在根尖干细胞巢区, WOX5在静止中心表达, 产生信号来抑制周围干细胞的分化, CLE40与CLV3类似, 通过与ACR4结合, 限制WOX5的表达, 从而在根尖形成了WOX5-CLE40/ACR40信号模块(Stahl et al., 2009)。同时WOX5也可以移动到根冠小柱起始细胞区(CSC), 与TPL蛋白相互作用, 抑制根冠分化蛋白CDF编码基因的转录, 从而维持CSC的干细胞功能(Pi et al., 2015)。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-126/img_2.png<b>图2</b> 根尖干细胞巢维持的调控网络模式图<br/>根尖干细胞巢维持途径主要有2条: PLT途径和SHR-SCR途径。PLTs (红色三角的密度)蛋白梯度和生长素的浓度梯度(紫色圆圈的密度)维持干细胞巢区的活性; BIRDs蛋白参与限制SHR转移到皮层, 从而与SCR蛋白一起作用界定内皮层与皮层细胞的界限。WOX5既可以维持QC的稳定性, 也可在CSC细胞抑制CDF4活性, 进而维持干细胞活性。CLE40通过与ACR4结合, 以及ARF10和ARF16的表达, 也可将WOX5的表达限制在QC内。橘黄色代表静止中心细胞, 浅黄色代表皮层细胞, 淡红色代表内皮层细胞; 黑色箭头表示促进作用; ⊥表示抑制作用;红色虚线箭头表示蛋白在细胞间移动。<br/><b>Figure 2</b> The regulation network for maintenance of the root stem cell niche<br/>The two major pathways for root stem cell niche maintenance: PLT pathway and SHR-SCR pathway. PLTs protein gradient (density of red triangles) and auxin concentration gradient (density of purple circles) maintain the activity of stem cell niche; BIRDs proteins are involved in repressing the transfer of SHR from endodermis to cortex cells, and work with SCR protein to define the boundary between endodermis and cortex. WOX5 not only maintains the stability of QC, but also repress the activity of CDF4 in CSC cells to maintain the acti- vity of stem cells. CLE40 restricts the expression of WOX5 in QC through binding to ACR4.And the expression of ARF10 and ARF16 has the same effect on WOX5. QC cells are co- lored in orange, cortex cells are colored in light yellow, and endodermis cells are colored in light red; black arrow represents promoted function; ⊥ represents inhibited function; red dotted arrow represents protein movement between cells. Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-126/img_2.png<b>图2</b> 根尖干细胞巢维持的调控网络模式图<br/>根尖干细胞巢维持途径主要有2条: PLT途径和SHR-SCR途径。PLTs (红色三角的密度)蛋白梯度和生长素的浓度梯度(紫色圆圈的密度)维持干细胞巢区的活性; BIRDs蛋白参与限制SHR转移到皮层, 从而与SCR蛋白一起作用界定内皮层与皮层细胞的界限。WOX5既可以维持QC的稳定性, 也可在CSC细胞抑制CDF4活性, 进而维持干细胞活性。CLE40通过与ACR4结合, 以及ARF10和ARF16的表达, 也可将WOX5的表达限制在QC内。橘黄色代表静止中心细胞, 浅黄色代表皮层细胞, 淡红色代表内皮层细胞; 黑色箭头表示促进作用; ⊥表示抑制作用;红色虚线箭头表示蛋白在细胞间移动。<br/><b>Figure 2</b> The regulation network for maintenance of the root stem cell niche<br/>The two major pathways for root stem cell niche maintenance: PLT pathway and SHR-SCR pathway. PLTs protein gradient (density of red triangles) and auxin concentration gradient (density of purple circles) maintain the activity of stem cell niche; BIRDs proteins are involved in repressing the transfer of SHR from endodermis to cortex cells, and work with SCR protein to define the boundary between endodermis and cortex. WOX5 not only maintains the stability of QC, but also repress the activity of CDF4 in CSC cells to maintain the acti- vity of stem cells. CLE40 restricts the expression of WOX5 in QC through binding to ACR4.And the expression of ARF10 and ARF16 has the same effect on WOX5. QC cells are co- lored in orange, cortex cells are colored in light yellow, and endodermis cells are colored in light red; black arrow represents promoted function; ⊥ represents inhibited function; red dotted arrow represents protein movement between cells.
图2 根尖干细胞巢维持的调控网络模式图 根尖干细胞巢维持途径主要有2条: PLT途径和SHR-SCR途径。PLTs (红色三角的密度)蛋白梯度和生长素的浓度梯度(紫色圆圈的密度)维持干细胞巢区的活性; BIRDs蛋白参与限制SHR转移到皮层, 从而与SCR蛋白一起作用界定内皮层与皮层细胞的界限。WOX5既可以维持QC的稳定性, 也可在CSC细胞抑制CDF4活性, 进而维持干细胞活性。CLE40通过与ACR4结合, 以及ARF10和ARF16的表达, 也可将WOX5的表达限制在QC内。橘黄色代表静止中心细胞, 浅黄色代表皮层细胞, 淡红色代表内皮层细胞; 黑色箭头表示促进作用; ⊥表示抑制作用;红色虚线箭头表示蛋白在细胞间移动。 Figure 2 The regulation network for maintenance of the root stem cell niche The two major pathways for root stem cell niche maintenance: PLT pathway and SHR-SCR pathway. PLTs protein gradient (density of red triangles) and auxin concentration gradient (density of purple circles) maintain the activity of stem cell niche; BIRDs proteins are involved in repressing the transfer of SHR from endodermis to cortex cells, and work with SCR protein to define the boundary between endodermis and cortex. WOX5 not only maintains the stability of QC, but also repress the activity of CDF4 in CSC cells to maintain the acti- vity of stem cells. CLE40 restricts the expression of WOX5 in QC through binding to ACR4.And the expression of ARF10 and ARF16 has the same effect on WOX5. QC cells are co- lored in orange, cortex cells are colored in light yellow, and endodermis cells are colored in light red; black arrow represents promoted function; ⊥ represents inhibited function; red dotted arrow represents protein movement between cells.
BlilouI, XuJ, WildwaterM, WillemsenV, PaponovI, FrimlJ, HeidstraR, AidaM, PalmeK, ScheresB (2005). The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots.Nature 433, 39-44. [本文引用: 2]
[6]
BoutilierK, OffringaR, SharmaVK, KieftH, OuelletT, ZhangLM, HattoriJ, LiuCM, van LammerenAAM, MikiBLA, CustersJBM, van Lookeren CampagneMM (2002). Ectopic expression of BABY BOOM triggers a con- version from vegetative to embryonic growth.Plant Cell 14, 1737-1749. [本文引用: 1]
[7]
BrandU, FletcherJC, HobeM, MeyerowitzEM, SimonR (2000). Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity.Science 289, 617-619. [本文引用: 2]
[8]
ChenQG, DaiXH, De-PaoliH, ChengYF, TakebayashiY, KasaharaH, KamiyaY, ZhaoYD (2014a). Auxin over- production in shoots cannot rescue auxin deficiencies in Arabidopsis roots.Plant Cell Physiol 55, 1072-1079. [本文引用: 1]
[9]
ChenX, GrandontL, LiHJ, HauschildR, PaqueS, AbuzeinehA, RakusováH, BenkovaE, Perrot-Rech- enmannC, FrimlJ (2014b). Inhibition of cell expansion by rapid ABP1-mediated auxin effect on microtubules.Nature 516, 90-93. [本文引用: 1]
Dello IoioR, NakamuraK, MoubayidinL, PerilliS, Tani- guchiM, MoritaMT, AoyamaT, CostantinoP, SabatiniS (2008). A genetic framework for the control of cell division and differentiation in the root meristem.Science 322, 1380-1384. [本文引用: 2]
[16]
DharmasiriN, DharmasiriS, EstelleM (2005a). The F-box protein TIR1 is an auxin receptor.Nature 435, 441-445. [本文引用: 2]
[17]
DharmasiriN, DharmasiriS, WeijersD, LechnerE, YamadaM, HobbieL, EhrismannJS, JurgensG, EstelleM (2005b). Plant development is regulated by a family of auxin receptor F box proteins.Dev Cell 9, 109-119. [本文引用: 1]
[18]
Di LaurenzioL, Wysocka-DillerJ, MalamyJE, PyshL, HelariuttaY, FreshourG, HahnMG, FeldmannKA, BenfeyPN (1996). TheSCARECROW gene regulates an asymmetric cell division that is essential for generating the radial organization of the Arabidopsis root. Cell 86, 423-433. [本文引用: 2]
[19]
DingZJ, FrimlJ (2010). Auxin regulates distal stem cell differentiation in Arabidopsis roots.Proc Natl Acad Sci USA 107, 12046-12051. [本文引用: 3]
[20]
DolanL, JanmaatK, WillemsenV, LinsteadP, PoethigS, RobertsK, ScheresB (1993). Cellular organisation of theArabidopsis thaliana root. Development 119, 71-84. [本文引用: 1]
[21]
ForzaniC, AichingerE, SornayE, WillemsenV, LauxT, DewitteW, MurrayJAH (2014). WOX5 suppressesCYC- LIN D activity to establish quiescence at the center of the root stem cell niche. Curr Biol 24, 1939-1944. [本文引用: 1]
GaoYB, ZhangY, ZhangD, DaiXH, EstelleM, ZhaoYD (2015). Auxin binding protein 1 (ABP1) is not required for either auxin signaling or Arabidopsis development.Proc Natl Acad Sci USA 112, 2275-2280. [本文引用: 1]
[24]
GutierrezC (2005). Coupling cell proliferation and develop- ment in plants.Nat Cell Biol 7, 535-541. [本文引用: 1]
[25]
HelariuttaY, FukakiH, Wysocka-DillerJ, NakajimaK, JungJ, SenaG, HauserMT, BenfeyPN (2000). TheSHORT-ROOT gene controls radial patterning of the Arabidopsis root through radial signaling. Cell 101, 555-567. [本文引用: 2]
[26]
HeymanJ, CoolsT, VandenbusscheF, HeyndrickxKS, Van LeeneJ, VercauterenI, VanderauweraS, Van- depoeleK, De JaegerG, Van Der StraetenD, De VeylderL (2013). ERF115 controls root quiescent center cell division and stem cell replenishment.Science 342, 860-863. [本文引用: 1]
KorasickDA, WestfallCS, LeeSG, NanaoMH, DumasR, HagenG, GuilfoyleTJ, JezJM, StraderLC (2014). Molecular basis for AUXIN RESPONSE FACTOR protein interaction and the control of auxin response repression.Proc Natl Acad Sci USA 111, 5427-5432.
SenaG, WangXN, LiuHY, HofhuisH, BirnbaumKD (2009). Organ regeneration does not require a functional stem cell niche in plants.Nature 457, 1150-1153. [本文引用: 2]
[50]
SozzaniR, CuiH, Moreno-RisuenoMA, BuschW, Van NormanJM, VernouxT, BradySM, DewitteW, MurrayJAH, BenfeyPN (2010). Spatiotemporal regulation of cell-cycle genes by SHORTROOT links patterning and growth.Nature 466, 128-132. [本文引用: 1]
[51]
StahlY, WinkRH, IngramGC, SimonR (2009). A signaling module controlling the stem cell niche in Arabidopsis root meristems.Curr Biol 19, 909-914. [本文引用: 2]
YamadaM, GreenhamK, PriggeMJ, JensenPJ, EstelleM (2009). TheTRANSPORT INHIBITOR RESPONSE2 gene is required for auxin synthesis and diverse aspects of plant development. Plant Physiol 151, 168-179. [本文引用: 1]
[62]
ZhangHM, HanW, De SmetI, TalboysP, LoyaR, HassanA, RongHL, JurgensG, Paul KnoxJ, WangMH (2010). ABA promotes quiescence of the quiescent centre and suppresses stem cell differentiation in the Arabidopsis pri- mary root meristem.Plant J 64, 764-774. [本文引用: 3]
[63]
ZhouWK, WeiLR, XuJ, ZhaiQZ, JiangHL, ChenR, ChenQ, SunJQ, ChuJF, ZhuLH, LiuCM, LiCY (2010). Arabidopsis tyrosylprotein sulfotransferase acts in the au- xin/PLETHORA pathway in regulating postembryonic main- tenance of the root stem cell niche.Plant Cell 22, 3692-3709. [本文引用: 2]
中国科学家在生长素信号转导领域取得突破性研究进展 1 2015
... 生长素是一种重要的植物激素, 它调节植物生长发育的各个方面, 包括根尖和茎尖分生组织的形成、胚的发生、下胚轴的伸长、侧根的起始和延伸以及器官极性的建立等.关于生长素对于根尖分生组织维持机理的研究, 虽然仍有需解决的问题, 但目前也取得了很大的进展(Ding and Friml, 2010; Cruz-Ramírez et al., 2012, 2013; Tian et al., 2014; 于倩倩等, 2015; 于燕杰等, 2016).同时, 近年来也有相关文献报道其它一些植物激素和环境因子, 如细胞分裂素、乙烯、脱落酸、油菜素内酯、茉莉酸以及冷适应等对根尖干细胞维持的影响(Ortega-Martinez et al., 2007; Zhang et al., 2010; Moubayidin et al., 2013; Vilarrasa-Blasi et al., 2014; Hong et al., 2017). ...
WOX蛋白家族调控干细胞发育分子机制的研究进展 1 2016
... 生长素是一种重要的植物激素, 它调节植物生长发育的各个方面, 包括根尖和茎尖分生组织的形成、胚的发生、下胚轴的伸长、侧根的起始和延伸以及器官极性的建立等.关于生长素对于根尖分生组织维持机理的研究, 虽然仍有需解决的问题, 但目前也取得了很大的进展(Ding and Friml, 2010; Cruz-Ramírez et al., 2012, 2013; Tian et al., 2014; 于倩倩等, 2015; 于燕杰等, 2016).同时, 近年来也有相关文献报道其它一些植物激素和环境因子, 如细胞分裂素、乙烯、脱落酸、油菜素内酯、茉莉酸以及冷适应等对根尖干细胞维持的影响(Ortega-Martinez et al., 2007; Zhang et al., 2010; Moubayidin et al., 2013; Vilarrasa-Blasi et al., 2014; Hong et al., 2017). ...
1 2015
... 生长素可以通过特定的输入转运蛋白(auxin1, AUX1)和输出转运蛋白(pin-formed, PINs)在植物体及其器官内进行极性运输, 从而维持一个定向的浓度梯度, 为个体发育、器官发育及器官内的细胞形态建成提供信号指导(Adamowski and Friml, 2015).PINs蛋白也可以通过在胚胎基部区域限制PLT的表达来起始胚根原基的形成.反过来, PLT基因维持PIN的转录, 从而稳定末端干细胞巢区的位置.在pin2/pin3/ pin4/pin7中, PLT的表达可以扩大到整个胚; 反过来在plt1/plt2中检测不到PIN4的转录水平, 且在plt1/ plt2/plt3三突变体中只能检测到较少的PIN1、PIN2和PIN3的表达(Aida et al., 2004; Galinha et al., 2007).因此研究者得出结论, 在生长素浓度最大的区域, PLT基因可能维持PINs蛋白编码基因的转录, 从而在植物胚根原基和根尖形成有效的环形生长素极性运输, 植物通过这种方式进一步维持生长素在胚根原基和根尖分生区所形成的浓度梯度, 从而指导根尖的形态发生. ...
... 与生长素相关的许多因子都参与胚根的形成.例如, 参与生长素运输的PIN1,3,4,7 (Blilou et al., 2005)和GNOM (Steinmann et al., 1999); 参与感知生长素信号的受体蛋白TIR1及其家族F-box蛋白(AFB1,2,3) (Dharmasiri et al., 2005a); 参与生长素合成的YUC- CA (Chen et al., 2014a)家族及TAA1 (Yamada et al., 2009), 这些蛋白对应突变体的胚根形成都有缺陷. ...
1 2014
... 很久以来, 研究者一直认为在质膜及胞质中快速的生长素响应并不使用SCFTIR1/AFBs蛋白酶体途径, 而是使用生长素结合蛋白(auxin-binding protein 1, ABP1) 这条非转录依赖的生长素响应途径(Tromas et al., 2013; Paque et al., 2014; Xu et al., 2014).最近的研究发现, ABP1在胞外需要通过跨膜停靠蛋白将生长素信号传送到胞质中, 而跨膜蛋白激酶(transmemb- rane kinase 1, TMK1)则作为停靠蛋白来分泌ABP1, 从而调节生长素信号(Xu et al., 2014).ABP1-TMK生长素感知复合体传递生长素信号, 以激活质膜相关的ROP信号途径, 从而调节非转录的胞质响应及相关生理过程.2014年, Jiri Friml实验室进一步证实了快速生长素响应介导的微管重定向也需要ABP1蛋白(Chen et al., 2014b). ...
2 2012
... 生长素是一种重要的植物激素, 它调节植物生长发育的各个方面, 包括根尖和茎尖分生组织的形成、胚的发生、下胚轴的伸长、侧根的起始和延伸以及器官极性的建立等.关于生长素对于根尖分生组织维持机理的研究, 虽然仍有需解决的问题, 但目前也取得了很大的进展(Ding and Friml, 2010; Cruz-Ramírez et al., 2012, 2013; Tian et al., 2014; 于倩倩等, 2015; 于燕杰等, 2016).同时, 近年来也有相关文献报道其它一些植物激素和环境因子, 如细胞分裂素、乙烯、脱落酸、油菜素内酯、茉莉酸以及冷适应等对根尖干细胞维持的影响(Ortega-Martinez et al., 2007; Zhang et al., 2010; Moubayidin et al., 2013; Vilarrasa-Blasi et al., 2014; Hong et al., 2017). ... ... 另外, 研究表明根尖干细胞巢的ACDs不仅需要SCR和CYCD6;1, 还需要RBR (RETINOBLASTOM- ARELATED)蛋白, 以及生长素在QC附近的最大浓度梯度.RBR与SCR在遗传上存在相互作用, 但是它们与根尖干细胞巢ACDs调控的分子机制尚不清楚(Wildwater et al., 2005).2012年, Ben Scheres实验室阐释了依靠SCR-RBR蛋白相互作用通过磷酸化和脱磷酸化来调控根尖干细胞巢皮层和内皮层母细胞不对称分裂的分子双稳态模型(Cruz-Ramírez et al., 2012).SHR在中柱中表达, 通过蛋白形式移动到内皮层, 与SCR蛋白结合, 而在成熟的内皮层细胞中, RBR蛋白的存在抑制了SHR与SCR的相互作用, 不能诱导ACD的产生.在CEI/CEID细胞中, SHR与SCR结合, 在生长素最大浓度条件下促进了CYCD6;1的转录, 诱导ACD的产生; 产生CYCD6;1后, RBR被CYCD6-CDKB复合体磷酸化, 磷酸化的RBR蛋白通过蛋白降解机制降解, 从而解除了对SCR和SHR的负调节作用, 但因为此时的子细胞没有处在生长素最高浓度, 所以CYCD6;1也不能有效表达, 从而关闭了ACD.如上所述, 植物通过RBR和生长素最高浓度调控SHR和SCR的转录活性, 界定形成了植物2层基本组织——皮层与内皮层. ...
1 2013
... 生长素是一种重要的植物激素, 它调节植物生长发育的各个方面, 包括根尖和茎尖分生组织的形成、胚的发生、下胚轴的伸长、侧根的起始和延伸以及器官极性的建立等.关于生长素对于根尖分生组织维持机理的研究, 虽然仍有需解决的问题, 但目前也取得了很大的进展(Ding and Friml, 2010; Cruz-Ramírez et al., 2012, 2013; Tian et al., 2014; 于倩倩等, 2015; 于燕杰等, 2016).同时, 近年来也有相关文献报道其它一些植物激素和环境因子, 如细胞分裂素、乙烯、脱落酸、油菜素内酯、茉莉酸以及冷适应等对根尖干细胞维持的影响(Ortega-Martinez et al., 2007; Zhang et al., 2010; Moubayidin et al., 2013; Vilarrasa-Blasi et al., 2014; Hong et al., 2017). ...
2 2007
... 皮层和内皮层基本组织是通过2次不对称细胞分裂(asymmetric cell division, ACD)形成的.第1次是皮层/内皮层起始(cortex/endodermal initial, CEI)干细胞的横向分裂: 形成1个自我更新的CEI细胞, 同时产生1个皮层/内皮层起始子细胞CEID (cortex/endo- dermis initial daughter).第2次是CEID细胞经过1次纵向的不对称分裂形成2个内皮层和皮层的前体细胞.SHR和SCR编码的GRAS家族转录因子和生长素的最大浓度, 对调节形成这2次连续ACD分裂, 特别是第2次纵向分裂, 以及形成根尖的基本组织起着重要作用(Di Laurenzio et al., 1996; Helariutta et al., 2000; Cui et al., 2007).突变体shr和scr均只有1层基本组织.在shr中, 这个单一的基本组织只有皮层的特征; 而在scr中, 它同时具有皮层和内皮层的特征.由于SHR蛋白在中柱表达, 它可以侧向外移1层细胞, 到达内皮层细胞, 进入细胞核调控SCR蛋白的表达, 从而定义内皮层(Di Laurenzio et al., 1996); 如果通过SCR基因启动子驱动, 在内皮层表达SHR蛋白, 由于SHR蛋白的向外平移特性, 导致邻近的细胞表达SCR蛋白, 最终产生大量的内皮层细胞(Helariutta et al., 2000; Cui et al., 2007).这些研究表明, SHR不仅可以定义特异的内皮层, 而且可以促进外一层细胞发育成皮层. ... ... ; Cui et al., 2007).这些研究表明, SHR不仅可以定义特异的内皮层, 而且可以促进外一层细胞发育成皮层. ...
... 与生长素相关的许多因子都参与胚根的形成.例如, 参与生长素运输的PIN1,3,4,7 (Blilou et al., 2005)和GNOM (Steinmann et al., 1999); 参与感知生长素信号的受体蛋白TIR1及其家族F-box蛋白(AFB1,2,3) (Dharmasiri et al., 2005a); 参与生长素合成的YUC- CA (Chen et al., 2014a)家族及TAA1 (Yamada et al., 2009), 这些蛋白对应突变体的胚根形成都有缺陷. ...
2 2014
... 生长素是一种重要的植物激素, 它调节植物生长发育的各个方面, 包括根尖和茎尖分生组织的形成、胚的发生、下胚轴的伸长、侧根的起始和延伸以及器官极性的建立等.关于生长素对于根尖分生组织维持机理的研究, 虽然仍有需解决的问题, 但目前也取得了很大的进展(Ding and Friml, 2010; Cruz-Ramírez et al., 2012, 2013; Tian et al., 2014; 于倩倩等, 2015; 于燕杰等, 2016).同时, 近年来也有相关文献报道其它一些植物激素和环境因子, 如细胞分裂素、乙烯、脱落酸、油菜素内酯、茉莉酸以及冷适应等对根尖干细胞维持的影响(Ortega-Martinez et al., 2007; Zhang et al., 2010; Moubayidin et al., 2013; Vilarrasa-Blasi et al., 2014; Hong et al., 2017). ... ... Tian等(2014)通过实验数据和计算机模拟的方法进一步得出WOX5-IAA17反馈机制介导的生长素响应对于根尖干细胞的维持非常重要.在根中生长素最大浓度一般出现在QC细胞区域内, 这一过程需要依赖IAA17介导的生长素响应.IAA17介导的生长素响应通过限制WOX5的表达区域来维持干细胞的识别性.WOX5调节QC细胞中游离产生的生长素, 进一步维持在QC细胞中生长素的高含量.进而证明WOX5和IAA17介导的生长素响应会产生反馈机制来调节末端干细胞的分化.在axr3-1突变体中, QC细胞对生长素响应增强, 而在相邻的小柱细胞中对生长素响应减弱, 从而抑制DSC的分化.相反地, 在QC中超表达IAA17强烈抑制生长素响应, 从而增强干细胞的分化.因此, 仅在QC细胞中保持生长素信号最大响应对于维持干细胞的识别性是不够的, 还需要减少相邻小柱细胞对生长素的响应, 从而达到根尖生长素响应梯度以调节根DSC的识别性. ...
1 2013
... 很久以来, 研究者一直认为在质膜及胞质中快速的生长素响应并不使用SCFTIR1/AFBs蛋白酶体途径, 而是使用生长素结合蛋白(auxin-binding protein 1, ABP1) 这条非转录依赖的生长素响应途径(Tromas et al., 2013; Paque et al., 2014; Xu et al., 2014).最近的研究发现, ABP1在胞外需要通过跨膜停靠蛋白将生长素信号传送到胞质中, 而跨膜蛋白激酶(transmemb- rane kinase 1, TMK1)则作为停靠蛋白来分泌ABP1, 从而调节生长素信号(Xu et al., 2014).ABP1-TMK生长素感知复合体传递生长素信号, 以激活质膜相关的ROP信号途径, 从而调节非转录的胞质响应及相关生理过程.2014年, Jiri Friml实验室进一步证实了快速生长素响应介导的微管重定向也需要ABP1蛋白(Chen et al., 2014b). ...
1 2014
... 生长素是一种重要的植物激素, 它调节植物生长发育的各个方面, 包括根尖和茎尖分生组织的形成、胚的发生、下胚轴的伸长、侧根的起始和延伸以及器官极性的建立等.关于生长素对于根尖分生组织维持机理的研究, 虽然仍有需解决的问题, 但目前也取得了很大的进展(Ding and Friml, 2010; Cruz-Ramírez et al., 2012, 2013; Tian et al., 2014; 于倩倩等, 2015; 于燕杰等, 2016).同时, 近年来也有相关文献报道其它一些植物激素和环境因子, 如细胞分裂素、乙烯、脱落酸、油菜素内酯、茉莉酸以及冷适应等对根尖干细胞维持的影响(Ortega-Martinez et al., 2007; Zhang et al., 2010; Moubayidin et al., 2013; Vilarrasa-Blasi et al., 2014; Hong et al., 2017). ...
... 植物以及一些动物有从成熟的器官中再生新器官的能力.关于植物干细胞的再生能力, 前人也做了一些探索与研究(Xu et al., 2006; Sena et al., 2009).Ben Scheres实验室利用激光敲除技术将QC切除, 发现切除QC会导致干细胞巢区从原维管组织中重新获得特异性(Xu et al., 2006).QC切除后, 生长素积累转移, 接着是PLT1的重定位, 且下调PIN的表达.PLTs基因促进SHR核定位, 进而促进SCR以及PLTs基因的表达, 最终新的QC得以建立.当新的细胞特异性产生后, PIN蛋白重新出现并进行正确的极性定位.QC被切除后, plt1/plt2突变体由于根尖生长素最大浓度建立出现缺陷而不能产生新的QC和根冠细胞.在突变体shr和scr中, 一个新的生长素最大浓度在一定区域产生, 紧接着根冠重新形成, 但是QC并没有形成, 且WOX5的表达丢失.这些初步结果都表明PLTs、SHR和SCR对于根尖干细胞巢的命运以及再生能力可能发挥重要作用. ... ... ).Ben Scheres实验室利用激光敲除技术将QC切除, 发现切除QC会导致干细胞巢区从原维管组织中重新获得特异性(Xu et al., 2006).QC切除后, 生长素积累转移, 接着是PLT1的重定位, 且下调PIN的表达.PLTs基因促进SHR核定位, 进而促进SCR以及PLTs基因的表达, 最终新的QC得以建立.当新的细胞特异性产生后, PIN蛋白重新出现并进行正确的极性定位.QC被切除后, plt1/plt2突变体由于根尖生长素最大浓度建立出现缺陷而不能产生新的QC和根冠细胞.在突变体shr和scr中, 一个新的生长素最大浓度在一定区域产生, 紧接着根冠重新形成, 但是QC并没有形成, 且WOX5的表达丢失.这些初步结果都表明PLTs、SHR和SCR对于根尖干细胞巢的命运以及再生能力可能发挥重要作用. ...
2 2014
... 很久以来, 研究者一直认为在质膜及胞质中快速的生长素响应并不使用SCFTIR1/AFBs蛋白酶体途径, 而是使用生长素结合蛋白(auxin-binding protein 1, ABP1) 这条非转录依赖的生长素响应途径(Tromas et al., 2013; Paque et al., 2014; Xu et al., 2014).最近的研究发现, ABP1在胞外需要通过跨膜停靠蛋白将生长素信号传送到胞质中, 而跨膜蛋白激酶(transmemb- rane kinase 1, TMK1)则作为停靠蛋白来分泌ABP1, 从而调节生长素信号(Xu et al., 2014).ABP1-TMK生长素感知复合体传递生长素信号, 以激活质膜相关的ROP信号途径, 从而调节非转录的胞质响应及相关生理过程.2014年, Jiri Friml实验室进一步证实了快速生长素响应介导的微管重定向也需要ABP1蛋白(Chen et al., 2014b). ... ... ).最近的研究发现, ABP1在胞外需要通过跨膜停靠蛋白将生长素信号传送到胞质中, 而跨膜蛋白激酶(transmemb- rane kinase 1, TMK1)则作为停靠蛋白来分泌ABP1, 从而调节生长素信号(Xu et al., 2014).ABP1-TMK生长素感知复合体传递生长素信号, 以激活质膜相关的ROP信号途径, 从而调节非转录的胞质响应及相关生理过程.2014年, Jiri Friml实验室进一步证实了快速生长素响应介导的微管重定向也需要ABP1蛋白(Chen et al., 2014b). ...
1 2009
... 与生长素相关的许多因子都参与胚根的形成.例如, 参与生长素运输的PIN1,3,4,7 (Blilou et al., 2005)和GNOM (Steinmann et al., 1999); 参与感知生长素信号的受体蛋白TIR1及其家族F-box蛋白(AFB1,2,3) (Dharmasiri et al., 2005a); 参与生长素合成的YUC- CA (Chen et al., 2014a)家族及TAA1 (Yamada et al., 2009), 这些蛋白对应突变体的胚根形成都有缺陷. ...
3 2010
... 生长素是一种重要的植物激素, 它调节植物生长发育的各个方面, 包括根尖和茎尖分生组织的形成、胚的发生、下胚轴的伸长、侧根的起始和延伸以及器官极性的建立等.关于生长素对于根尖分生组织维持机理的研究, 虽然仍有需解决的问题, 但目前也取得了很大的进展(Ding and Friml, 2010; Cruz-Ramírez et al., 2012, 2013; Tian et al., 2014; 于倩倩等, 2015; 于燕杰等, 2016).同时, 近年来也有相关文献报道其它一些植物激素和环境因子, 如细胞分裂素、乙烯、脱落酸、油菜素内酯、茉莉酸以及冷适应等对根尖干细胞维持的影响(Ortega-Martinez et al., 2007; Zhang et al., 2010; Moubayidin et al., 2013; Vilarrasa-Blasi et al., 2014; Hong et al., 2017). ... ... QC在根发育中发挥重要作用, 它通过产生一个微环境来维持其周围细胞的干细胞命运.尽管被一些具有高度有丝分裂活性的起始细胞围绕着, 但QC细胞以较低的增殖率进行自我更新.QC周围的起始细胞也可以被QC增殖的细胞所代替, 前者被认为是短期(short-term)干细胞, 而后者被认为是长期(long- term)干细胞.前人研究表明, 乙烯和ABA以及小分子信号肽都能促进QC的分裂(Ortega-Martinez et al., 2007; Matsuzaki et al., 2010; Zhang et al., 2010).功能缺失突变体eto1 (ethylene overproducer 1)会产生大量的乙烯, 而在eto1突变体中可观察到明显的QC分裂现象, 对野生型外源施加ACC (1-aminocyc- lopropane-l-carboxylic acid)也能促进QC的分裂, 从而证明乙烯可以促进QC的分裂.而当外源增高野生型根尖生长素浓度时, 并未观察到额外的QC分裂, 说明生长素自身并不能诱导QC的分裂(Ortega-Mar- tinez et al., 2007).乙烯响应因子ERF115 (ethylene response factor 115)控制根静止中心的分裂以及干细胞重建.ERF115通过PSK信号促进QC的分裂, 它是APC/CCCS52A2的底物且其表达受油菜素内酯的信号调节(Heyman et al., 2013). ... ... 与高等动物不同, 绿色植物固着生长, 因此必须通过自我调节机制来适应外界环境的变化, 如干旱、水涝、病害、高温和低温等.前人研究表明, 通过环境诱导的乙烯和小分子信号肽都能促进QC细胞的分裂, 而ABA可以抑制QC细胞的分裂(Ortega-Martinez et al., 2007; Matsuzaki et al., 2010; Zhang et al., 2010).同时, 温度适应对于植物和动物的发育也起着非常重要的作用, 但温度如何调节植物干细胞活性的维持目前还了解较少.最近须健研究团队发现, 低温可以导致拟南芥根尖干细胞邻近细胞发生DNA损伤, 从而诱导ATM (ataxia telangiectasia mutated)-ATR (ataxia telangiectasia and Rad3-related protein)依赖的程序化死亡, 有利于维持邻近QC细胞中生长素的最高浓度, 从而增强植物根尖对低温的适应性, 保持其有效生长(Hong et al., 2017).这一主动的细胞死亡机制提高了根对环境胁迫的适应能力, 从而维持根的正常发育. ...
2 2010
... 研究人员通过正向遗传学的方法筛选并鉴定了一系列的酪氨酸磺基转移酶(tyrosylprotein sulfot- ransferase, TPST)和根分生生长因子(root meristem growth factor, RGFs)的突变体, 发现单突变体以及多突变体的根尖分生组织存在不同程度的发育缺陷(Matsuzaki et al., 2010; Zhou et al., 2010).李传友课题组发现TPST可以调节根尖干细胞巢的维持(Zhou et al., 2010).TPST突变会导致根尖干细胞维持缺陷, 分生区活性下降, 妨碍根的生长.他们还发现生长素可以正调节TPST的表达, 且TPST突变之后会影响生长素在根尖生长点部位的极性运输、局部合成和局部浓度梯度的形成; TPST突变也导致根尖干细胞转录因子PLTs在转录和蛋白表达水平的降低, 而过表达PLT2可以部分恢复tpst-1根尖分生缺陷.他们最终得出TPST所介导的蛋白质硫基化是生长素和PLT之间的联系纽带.另外, 其它研究组发现, TPST作用产生的小肽RGF信号对于根尖干细胞的维持发挥重要作用(Matsuzaki et al., 2010).他们发现RGFs不仅正向维持根尖分生组织的活性, 而且也正向维持根尖PLTs的表达水平及模式.有意思的是, 研究结果显示, RGFs的功能有可能与生长素途径相互独立, 而且这些肽信号只对根的胚后发育起重要作用. ... ... ).李传友课题组发现TPST可以调节根尖干细胞巢的维持(Zhou et al., 2010).TPST突变会导致根尖干细胞维持缺陷, 分生区活性下降, 妨碍根的生长.他们还发现生长素可以正调节TPST的表达, 且TPST突变之后会影响生长素在根尖生长点部位的极性运输、局部合成和局部浓度梯度的形成; TPST突变也导致根尖干细胞转录因子PLTs在转录和蛋白表达水平的降低, 而过表达PLT2可以部分恢复tpst-1根尖分生缺陷.他们最终得出TPST所介导的蛋白质硫基化是生长素和PLT之间的联系纽带.另外, 其它研究组发现, TPST作用产生的小肽RGF信号对于根尖干细胞的维持发挥重要作用(Matsuzaki et al., 2010).他们发现RGFs不仅正向维持根尖分生组织的活性, 而且也正向维持根尖PLTs的表达水平及模式.有意思的是, 研究结果显示, RGFs的功能有可能与生长素途径相互独立, 而且这些肽信号只对根的胚后发育起重要作用. ...


{kind=link}
{kind=link}
{kind=link}
{kind=link}