 , ,
, , 引用本文
贡献者
基金资助
接受日期:2016-12-6接受日期:2017-05-4网络出版日期:2017-11-1
-->Copyright
2017《植物学报》编辑部
Contributors
History
Received:Accepted:Online:
摘要:
Abstract:
Key words:
负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(Krueger, 1962, 1972, 1985; Krueger and Reed, 1976)。其作为衡量空气质量的一个重要指标, 具有预防疾病和增强人体免疫力的功效, 呼吸富含负离子的空气, 能提高人体的舒适度(Griffin and Kornblueh, 1962; Krueger and Reed, 1976; Kosenko et al., 1997; Kondrashove et al., 2000)。许多****对负离子的生物学效应进行过研究, 如负离子可调节大脑皮层的功能, 促进氧的吸收和利用(孙继良等, 2010), 降低直肠的温度(Wakamura et al., 2004); 缓解高强度室内工作的压力, 消除疲劳(Nakane et al., 2002)等。反之, 在缺乏负离子的空气中工作或生活, 人体的功能可能会出现紊乱(Krue- ger, 1985; Wu et al., 2011)。故负离子常被誉为空气中的“维生素”(李安伯, 1983, 2001; 李安伯和张振军, 1996)。负离子主要以O2-(H2O)n和(OH)-(H2O)n两种形式存在(Griffin and Kornblueh, 1962; Wang and Li, 2009; 吴仁烨等, 2015)。自然界中的负离子大部分来自宇宙射线和雷电等对大气分子的电离作用、水的喷筒电效应、树冠与枝叶的尖端放电, 及植物光合作用形成的光电效应促使空气电离。自然状态下, 空气通过空气电离与离子复合两个过程使空气中的离子保持动态平衡(Wang and Li, 2009; Wu et al., 2011)。
负离子浓度随着环境的变化而呈现差异。例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3)。而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(李安伯, 2001; 吴楚材等, 2001; 吴仁烨等, 2014b)。原因在于植物的光合作用促使空气发生电离, 进而增加了负离子的浓度(吴楚材和钟林生, 1998; 曾曙才等, 2007; 刘新等, 2011)。同时, 负离子浓度受环境气象因素的影响较大, 温度、湿度、风、空气中的颗粒物和辐射等都影响负离子浓度的时空差异(秦俊等, 2008; 黄向华等, 2013; 王成等, 2014; 王薇, 2014; 王晓磊和王成, 2014)。总之, 室外环境中负离子的浓度远高于室内环境, 然而大部分人群在室内工作和活动的时间较长, 几乎占全天24小时的80%-90%, 这使得他们无法享受到大自然环境中富含植物源负离子的清新空气(王艳英等, 2014; 吴仁烨等, 2014a), 显然, 提高室内环境的负离子浓度十分重要。
目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(邵海荣和贺庆棠, 2000; 秦俊等, 2008; 鲍风宇等, 2013; 陈雷等, 2015), 环境中负离子浓度与气象因子的关系(穆丹和梁英辉, 2009; 石彦军等, 2010; 黄向华等, 2013), 城市绿地及景区不同植被的负离子浓度与空气质量评价(李少宁等, 2010; Wu et al., 2011; 张凯旋和张建华, 2013; Liang et al., 2014; 任洪昌等, 2014; 关蓓蓓等, 2016), 及负离子的生态效益评价(钟林生等, 1998; 范亚民等, 2005; 董莎莎等, 2013)等方面。对植物释放负离子的机理则未见报道。吴仁烨等(2011b)研究了植物在自然状态下产生负离子的能力, 指出自然状态下植物释放负离子的能力均很弱, 单株植物释放负离子的浓度均值最大不超过300 ion?cm-3, 能够引起的生物学和生态效应极为有限。另有研究表明, 高压脉冲(Tikhonov et al., 2002, 2004; 吴仁烨等, 2014b, 2015; 张万超等, 2015, 2016)和强光照度(Wang and Li, 2009)可显著提高植物释放负离子的能力, 推测植物释放负离子源自植物的生理变化过程而非物理变化过程。
为了研究脉冲电场和光照刺激对植物释放负离子的影响, 以及植物叶片气孔数量特征的变化与植物释放负离子能力之间的关系, 我们选取合果芋(Syngo- nium podophyllum)、百合(Lilium brownii var. viridu- lum)和剑麻(Agave sisalana)等10种常见、易于种植且具有观赏价值的植物为研究对象, 探索施加不同参数的脉冲电场和不同强度的光照刺激时植物释放负离子的能力, 及脉冲电场作用下, 植物释放负离子的能力与植物叶片气孔特征变化的关系, 以期寻找到提高植物释放负离子能力的最佳方法。
1 材料与方法1.1 植物材料为研究叶片形状以及植株形态对植物释放负离子的影响, 我们选取合果芋(Syngonium podophyllum Schott)、百合(Lilium brownii var. viridulum Baker)、剑麻(Agave sisalana Perr. ex Engelm.)、紫背竹芋(Stromanthe sanguinea Sond.)、朱蕉(Cordyline fruticosa (L.) A.Cheval.)、绒叶肖竹芋(Calathea zeb- rina (Sims) Lindl.)、朱顶红(Hippeastrum rutilum (Ker-Gawl.) Herb.)、大鳞巢蕨(Neottopteris antiqua (Makino) Masamune)、虎耳草(Saxifraga stolonifera Curt.)和八角金盘(Fatsia japonica (Thunb.) Decne. et Planch) 10种植物(表1)为研究对象。供试植物均购于福建漳州百花村花卉交易市场, 每种植物选取外形和长势一致的植株3棵, 作为实验的3次重复。所有植物均种植在塑料花盆(外口径23 cm, 高19 cm, 底15 cm)中, 采取统一的水肥管理和养护方法。
表1
Table 1
表1
| Code | Plants | Age (mon- th) | Plant height× Grown bread- th (cm) |
|---|---|---|---|
| P1 | Syngonium podophyllum | 14 | 20×20 |
| P2 | Lilium brownii var. viridulum | 13 | 70×20 |
| P3 | Agave sisalana | 24 | 45×30 |
| P4 | Stromanthe sanguinea | 12 | 30×30 |
| P5 | Cordyline fruticosa | 14 | 60×50 |
| P6 | Calathea zebrina | 12 | 40×45 |
| P7 | Hippeastrum rutilum | 13 | 60×40 |
| P8 | Neottopteris antiqua | 14 | 40×40 |
| P9 | Saxifraga stolonifera | 12 | 30×30 |
| P10 | Fatsia japonica | 12 | 50×40 |
表1
供试植物名录
Table 1
Tested plants species
1.2 负离子浓度的测定实验于2016年6-8月进行, 选择该段时间的晴天, 采用厚度为4 mm玻璃制成的规格为80 cm×80 cm×80 cm的可密封式玻璃室, 在其一个侧面截取104 mm×104 mm的小窗口, 窗口大小与DLY-4G-232型空气离子测量仪的进风口相匹配, 未测量时用玻璃挡板封闭窗口(图1)。调节盆栽植物中心, 使其与玻璃箱的中心重叠, 以保持空气离子测量仪进气口的中心与株顶处于同一水平。负离子浓度使用DLY-4G-232 型空气离子测量仪测定, 每秒读取1个数据, 并通过RS-232接口传输至电脑, 连续读取有效时间150秒, 取150个数据的平均值作为该植株释放负离子的浓度值, 将3组重复的负离子浓度平均值作为该种植物在该状态下释放的负离子浓度值。
图1
Figure 1

下载原图ZIP
生成PPT
图1
密闭玻璃室内测量植物释放负离子浓度示意图
a: 高压脉冲刺激仪; b: 脉冲探针; c: 空气离子测量仪; d: 电脑; e: 密闭玻璃室; f: 植物; g: 可调式绝缘平台
Figure 1
Schematic of detecting negative air ions concentration by plant in a sealed chamber
a: High-voltage pulsed generator; b: Pulse probe; c: Air ions detector; d: Computer; e: Glass chamber; f: Plant; g: Adjustable insulation platform
自然状态下植物释放负离子浓度的测量实验设置2个对照组, 即盆土(无植物)和空白组(无土壤且无植物)。测量前先将植物放入玻璃室中静置15分钟, 以消除玻璃室中原有负离子浓度的影响。每小时测量1次, 全天共测定24次, 将3次重复的负离子浓度均值作为自然状态下该植物在该时段释放的负离子浓度值, 24个时段的负离子浓度均值作为该植物在自然状态下释放负离子的浓度值。
脉冲电场作用下植物释放负离子浓度的测量实验设置1个对照组, 即土壤(无植物)。采用福建农林大学研制的电脉冲刺激仪进行测定, 该仪器平均脉冲输出电压U=0.8×104-2.0×104 V, 脉冲间隔T=0.5-2.0 s, 脉冲宽度τ=5-90 ms。为研究施加不同强度脉冲电场对植物释放负离子能力的影响, 采用3因素(脉冲电压A、脉冲间隔B和脉冲宽度C) 4水平(表2)的正交设计实验, 共设置16个处理, 每个处理3个重复。测量前将脉冲刺激仪的探针在离株茎外围5 cm处埋入土壤(深5 cm), 之后施加脉冲电场刺激。3次重复的负离子浓度均值作为该脉冲电场作用下该植物释放负离子的浓度值。
表2
Table 2
表2
| Level | Factors | ||
|---|---|---|---|
| A (103 V) | B (s) | C (ms) | |
| 1 | 8 | 0.5 | 5 |
| 2 | 10 | 1.0 | 35 |
| 3 | 15 | 1.5 | 65 |
| 4 | 20 | 2.0 | 90 |
表2
正交实验设计表
Table 2
Factors and levels for orthogonal test
1.3 脉冲电场作用时植物体电压分析为探究植物体电压与释放负离子能力之间的关系, 我们设置了3个实验组, 即对照组(CK)、实验A组和B组, 各设3次重复。对照组(CK): 对植物根际土壤施加最佳脉冲电场刺激, 用PINTEK HVP-40高压测量笔连接TDO1000/2000数字储存示波器, 读取植物体上的电压数值, 同时检测负离子浓度。实验A组: 最佳脉冲电场作用时, 导线连接植物体与物理地(大地); 实验B组: 最佳脉冲电场作用时, 导线连接植物体与地线, 同时均检测植物体电压和负离子的浓度。
1.4 脉冲电场和光照度作用下植物释放负离子 分析实验在Hipoint 740FHC光照培养箱中进行, 光照度的测量和校准采用Hipoint HR-350光谱分析仪(测量范围为0-100 000 lx, 测量波长为360-760 nm)。实验设置对照组(不施加脉冲电场作用)和刺激组(对每个供试植物施加最佳脉冲电场刺激)。每实验组设置6个处理(光照度水平), 即0、500、1 500、3 000、6 000和12 000 lx, 每个处理3次重复。研究常态和最佳脉冲电场刺激下, 光照度变化对植物释放负离子能力的影响。
1.5 植物叶表皮气孔分析为研究植物释放负离子与植物叶片气孔数量特征的关系, 实验设置1个对照组(CK)和2个处理组。对照组: 对自然状态下生长的植物进行采样; 实验组: 对植物施加最佳脉冲电场刺激, 持续600秒后进行采样, 同时检测负离子的浓度值。取样部位为叶片靠近中脉两侧约0.3-0.5 cm处, 将供试材料切成3-5 cm2正方形小块, 于FAA溶液中固定24小时, 60.0°C水浴40分钟后, 用30%-40%次氯酸钠浸泡24小时, 之后换用蒸馏水, 撕取叶片的上下表皮, 用0.5%固绿染色, 制成临时切片(张亚冰等, 2008), 在OPTEC (DV320)光学显微镜下观察并拍照。叶表皮的观察在叶脉间区域进行, 数量特征统计取10个视野进行分析。光学显微镜20x镜下观察气孔数量, 40x镜下测量植物叶片表皮气孔的长、宽和面积。在观察气孔数量特征时, 测定200个数值后取平均值, 气孔长宽比按对应值计算后求200个数值的平均值。
2 结果与讨论2.1 常态下植物释放负离子浓度的日变化由图2可知, 自然状态下10种植物在全天各时段释放负离子浓度的均值都很低。空白对照(CK)和盆土(P0)的负离子浓度在全天各时段的变化较为平稳。其中,空白对照全天的负离子浓度最小值为29 ion·cm-3, 最大值为38 ion·cm-3。从全天释放负离子浓度的最大值来看, 百合在14:00释放的负离子浓度值最大, 为94 ion·cm-3, 是空白对照负离子浓度最大值的2.5倍; 剑麻在23:00释放能力最小, 为35 ion·cm-3。以全天释放负离子浓度的均值分析, 绒叶肖竹芋释放负离子的浓度均值最大, 为81 ion·cm-3; 紫背竹芋位居第2, 数值为76 ion·cm-3; 朱顶红为75 ion·cm-3, 位列第3; 合果芋最小, 为46 ion·cm-3。尽管10种植物释放负离子浓度的均值间存在显著差异(P<0.5), 但各植物释放负离子浓度值的绝对差异非常小。从白天均值和夜间均值的分析结果可以看出, 除绒叶肖竹芋和虎耳草外, 其余植物释放负离子的浓度均值白天时段(7:00AM- 7:00PM)均高于夜间时段(7:00PM-7:00AM)(表3), 其中以大鳞巢蕨最为明显, 相对增幅为25%。
图2
Figure 2

下载原图ZIP
生成PPT
图2
自然状态下植物24小时释放负离子浓度的变化趋势
CK: 空白对照; P0: 盆土; P1-P10同
Figure 2
The changes in negative air ions concentration over 24 h for plant varieties under normal condition
CK: P0 soil without plant; P0: Soil; P1-P10 see
表3
Table 3
表3
| Code | 24 h mini- mum | 24 h maxi- mum | 24 h mean | Daytime mean | Nighttime mean | Max/ Min | (Day mean-night mean)/ Night mean | Day mean/Night mean |
|---|---|---|---|---|---|---|---|---|
| CK | 29 | 38 | 32 k | 34 | 31 | 1.31 | 0.10 | 1.1 |
| P0 | 30 | 36 | 33 j | 34 | 32 | 1.2 | 0.06 | 1.06 |
| P1 | 36 | 56 | 46 i | 48 | 44 | 1.56 | 0.09 | 1.09 |
| P2 | 48 | 94 | 63 d | 68 | 57 | 1.96 | 0.19 | 1.19 |
| P3 | 35 | 81 | 58 f | 63 | 53 | 2.31 | 0.19 | 1.19 |
| P4 | 64 | 91 | 76 b | 79 | 74 | 1.42 | 0.07 | 1.07 |
| P5 | 48 | 71 | 57 g | 58 | 57 | 1.48 | 0.02 | 1.02 |
| P6 | 71 | 90 | 81 a | 80 | 83 | 1.27 | -0.04 | 0.96 |
| P7 | 56 | 90 | 75 c | 76 | 74 | 1.61 | 0.03 | 1.03 |
| P8 | 39 | 82 | 57 g | 64 | 51 | 2.1 | 0.25 | 1.25 |
| P9 | 44 | 80 | 62 e | 58 | 66 | 1.82 | -0.12 | 0.88 |
| P10 | 40 | 58 | 48 h | 48 | 47 | 1.45 | 0.02 | 1.02 |
表3
自然状态下植物24小时释放负离子浓度的分析(ion·cm-3)
Table 3
Analysis of negative air ions concentration generated by plants among 24 h in natural conditions (ion·cm-3)
2.2 植物释放负离子最佳参数组合的脉冲电场不同强度的脉冲电场对盆土释放负离子的作用很小(36-140 ion·cm-3), 处理间均无显著差异(表4)。紫背竹芋在不同强度脉冲电场作用下, 释放负离子的能力存在差异, 浓度均值为452 644-1 730 800 ion·cm-3 (表4)。A3B3C3处理下的负离子浓度均值最高, 是常态下释放能力(未刺激)的22 773.7倍。负离子浓度最低的处理为A1B1C1, 浓度均值为452 644 ion·cm-3, 是其常态下释放能力的5 955.8倍。方差分析表明, 紫背竹芋高效释放负离子的最优处理为A3B3C3。在该强度脉冲电场作用下, 其释放负离子能力的倍增效应最为显著。
表4
Table 4
表4
| Treatment | Factors | Average of negative air ions concentration (ion·cm-3) | ||||||
|---|---|---|---|---|---|---|---|---|
| A | B | C | P0 | P4 | P6 | P7 | ||
| A1B1C1 | 8 | 0.5 | 5 | 36±3 a | 452644±21866 n | 91±6 a | 8605±983 f | |
| A1B2C3 | 8 | 1.0 | 65 | 38±2 a | 471667±21881 n | 92±8 a | 9581±948 f | |
| A1B3C4 | 8 | 1.5 | 90 | 39±2 a | 535311±25007 l | 94±5 a | 8559±874 f | |
| A1B4C2 | 8 | 2.0 | 35 | 44±2 a | 494667±27146 m | 98±8 a | 8832±543 f | |
| A2B1C4 | 10 | 0.5 | 90 | 39±2 a | 795822±53569 k | 95±9 a | 20200±1478 f | |
| A2B2C2 | 10 | 1.0 | 35 | 38±2 a | 822267±54244 j | 88±8 a | 22473±1381 f | |
| A2B3C1 | 10 | 1.5 | 5 | 37±3 a | 813022±50909 jk | 85±7 a | 23236±2467 f | |
| A2B4C3 | 10 | 2.0 | 65 | 41±3 a | 872734±55664 i | 91±4 a | 21801±1643 f | |
| A3B1C2 | 15 | 0.5 | 35 | 105±2 a | 1564444±119680 d | 226±20 a | 181311±20261 e | |
| A3B2C4 | 15 | 1.0 | 90 | 107±2 a | 1628244±191548 c | 250±40 a | 218444±33270 c | |
| A3B3C3 | 15 | 1.5 | 65 | 109±3 a | 1730800±195344 a | 233±16 a | 191867±32167 de | |
| A3B4C1 | 15 | 2.0 | 5 | 107±2 a | 1670933±187634 b | 262±28 a | 208067±34590 cd | |
| A4B1C3 | 20 | 0.5 | 65 | 130±2 a | 1186667±103135 f | 170±39 a | 301933±30237 b | |
| A4B2C1 | 20 | 1.0 | 5 | 129±5 a | 1264000±117184 e | 179±18 a | 330356±30322 a | |
| A4B3C2 | 20 | 1.5 | 35 | 134±3 a | 1136800±83461 g | 215±33 a | 322644±29865 ab | |
| A4B4C4 | 20 | 2.0 | 90 | 140±4 a | 1038133±83109 h | 164±51 a | 318156±36925 ab | |
表4
不同参数的脉冲电场刺激下植物释放负离子浓度的分析(平均值±标准差)
Table 4
Analysis of negative air ions concentration generated by plants upon different combinational parameters of pulsed electrical stimulation (means±SD)
绒叶肖竹芋释放负离子的浓度均值为85-262 ion·cm-3 (表4), 以A3B4C1处理下负离子的释放能力最强, 是其常态下释放能力的3.2倍。而以A2B3C1作用下的释放能力最弱, 仅为85 ion·cm-3, 与自然状态下的释放能力(81 ion·cm-3)相近。尽管各处理绒叶肖竹芋释放负离子的能力不同, 但各处理间均无显著差异。方差分析表明, 仅脉冲电压对其释放负离子的能力存在显著影响(P<0.05)。验证性实验表明, A3B4C1为绒叶肖竹芋高效释放负离子最佳脉冲电场的组合参数。
朱顶红释放负离子的浓度均值为8 559-330 356 ion·cm-3 (表4), 以A4B2C1作用下的释放能力最强, 是其常态下的4 404.7倍, 与其它各处理均呈显著差异(P<0.05)。释放负离子能力最小的是A1B3C4, 为8 559 ion·cm-3, 是其常态下的114.1倍。方差分析表明, 脉冲电压、脉冲间隔和脉冲宽度均对其释放负离子的浓度具有显著的影响(P<0.05)。验证性实验表明, A4B4- C4是其高效释放负离子的最佳脉冲电场组合参数, 浓度均值为330 356 ion·cm-3。
与自然状态下各植物释放负离子的能力相比, 施加不同强度的脉冲电场, 植物释放负离子的浓度有不同程度的提高。除绒叶肖竹芋外, 紫背竹芋和朱顶红释放负离子的能力在不同强度脉冲电场作用下均存在显著差异(P<0.05) (表4)。
2.3 释放负离子能力与植物体电压的关系表5显示, 同一植物体上储存的电压与其释放负离子的能力呈正相关。紫背竹芋、绒叶肖竹芋和朱顶红对照组的负离子浓度均值都高于处理A和B组。除绒叶肖竹芋的负离子浓度在3个处理间无显著差异外(P> 0.05), 紫背竹芋和朱顶红在处理A、B两组的浓度值与对照组相比均差异显著(P<0.05)。
表5
Table 5
表5
| Treatment | Calathea insignis | Calathea zebrina | Hippeastrum rutilum | |||||
|---|---|---|---|---|---|---|---|---|
| Voltage (103 V) | NAIC (ion·cm-3) | Voltage (103 V) | NAIC (ion·cm-3) | Voltage (103 V) | NAIC (ion·cm-3) | |||
| CK | 5.13±0.33 a | 1757467±218808 a | 5.50±0.14 a | 260±33 a | 4.32±0.11 a | 362000±35957 a | ||
| A | 1.78±0.10 b | 2119±88 b | 1.44±0.26 b | 152±11 a | 0.91±0.06 b | 706±29 b | ||
| B | 0.51±0.03 c | 89±6 b | 0.66±0.04 c | 85±6 a | 0.50±0.06 c | 77±5 b | ||
表5
植物体电压与释放负离子浓度的关系(平均值±标准差)
Table 5
Analysis between voltage of plants and negative air ions concentration (means±SD)
3种植物对照组的电压与输入电压(最佳参数组合的脉冲电压)相比都有不同程度的衰减, 其中以朱顶红的电压衰减量最大, 减少了78.4%, 负离子浓度为362 000 ion·cm-3。绒叶肖竹芋和朱顶红分别减少65.8%和63.3%, 负离子浓度分别为1 757 467和260 ion·cm-3。尽管绒叶肖竹芋植物体上的电压为三者中最高, 但其释放负离子的能力却最小。从处理组A的结果可以看出, 3种植物体上的电压衰减量与CK相比存在显著差异(P<0.05); 紫背竹芋植物体电压与输入电压相比减少了88.1%, 负离子浓度比CK减少了99.9%; 绒叶肖竹芋和朱顶红的植物体电压分别减少了90.4%和95.5%, 负离子浓度分别减少41.5%和99.8%。处理B组中, 3种植物的植物体电压进一步衰减, 释放负离子的能力接近自然状态下的水平(表5)。
2.4 释放负离子能力与光照度的关系由表6可知, 不施加脉冲电场作用时, 土壤释放负离子的能力在各光照度下变幅很小, 其释放负离子的浓度均值为35-43 ion·cm-3。3种植物释放负离子的能力随光照度的增加呈上升趋势, 尽管绝对变幅很小, 但各处理间均存在显著差异(P<0.05)(表6)。在脉冲电场作用下, 土壤和3种植物释放负离子的能力与常态下相比均有显著提高(P<0.05), 且随着光照度的增加, 植物释放负离子的能力呈升高趋势。其中以朱顶红的增幅最小, 在12 000 lx光照度下其释放负离子的浓度为黑暗状态下的1.5倍; 以绒叶肖竹芋的增幅最大, 其释放负离子的能力是0 lx下的2.0倍, 但释放负离子的浓度均值很小, 最大值仅为310 ion·cm-3。除土壤和绒叶肖竹芋外, 紫背竹芋和朱顶红在不同的光照度刺激下释放负离子的浓度均存在显著差异(P< 0.05)。在光照度为3 000-6 000 lx时, 3种植物释放负离子的能力均差异不显著。
表6
Table 6
表6
| Light intensity (lx) | Negative air ions concentration (ion·cm-3) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Soil | Calathea insignis | Calathea zebrina | Hippeastrum rutilum | ||||||||
| CK | S | CK | S | CK | S | CK | S | ||||
| 0 | 35±2 c | 127±3 a | 53±2 e | 824400±88904 e | 76±2 e | 156±16 a | 66±2 e | 202067±24109 c | |||
| 500 | 36±2 c | 136±2 a | 69±3 d | 1726200±186594 d | 80±4 d | 275±28 a | 69±3 d | 298289±36482 b | |||
| 1500 | 37±3 bc | 138±2 a | 79±2 c | 1769911±191872 c | 83±3 c | 284±28 a | 77±2 c | 308156±36967 b | |||
| 3000 | 37±3 bc | 135±2 a | 81±3 c | 1831378±198645 b | 80±3 d | 297±30 a | 79±5 c | 328644±39232 a | |||
| 6000 | 43±2 a | 140±2 a | 90±3 b | 1813622±197187 b | 106±3 b | 296±30 a | 93±3 b | 328200±38992 a | |||
| 12000 | 39±3 b | 139±3 a | 138±4 a | 1895200±205601 a | 128±3 a | 310±31 a | 134±3 a | 308156±36967 b | |||
表6
脉冲电场作用对植物不同光照度下释放负离子浓度的分析(平均值±标准差)
Table 6
The negative air ion concentration of plants under pulsed electrical field stimulation in different light intensity (means± SD)
2.5 释放负离子能力与气孔数量特征的关系紫背竹芋在最佳脉冲电场作用下, 其气孔形态由常态下的紧闭变为开启状态(图3A, B)。与对照相比, 电场作用下的叶片气孔长度增加了65.9%, 宽度增加了67.1%, 均差异显著(P<0.05); 气孔周长比对照组增加了66.0%, 面积增大了175.5%, 且差异显著(P<0.05); 而气孔密度比对照组降低了11.0%, 释放负离子的能力为对照组的21 760.0倍, 具显著性差异(P<0.05) (表7)。
图3
Figure 3
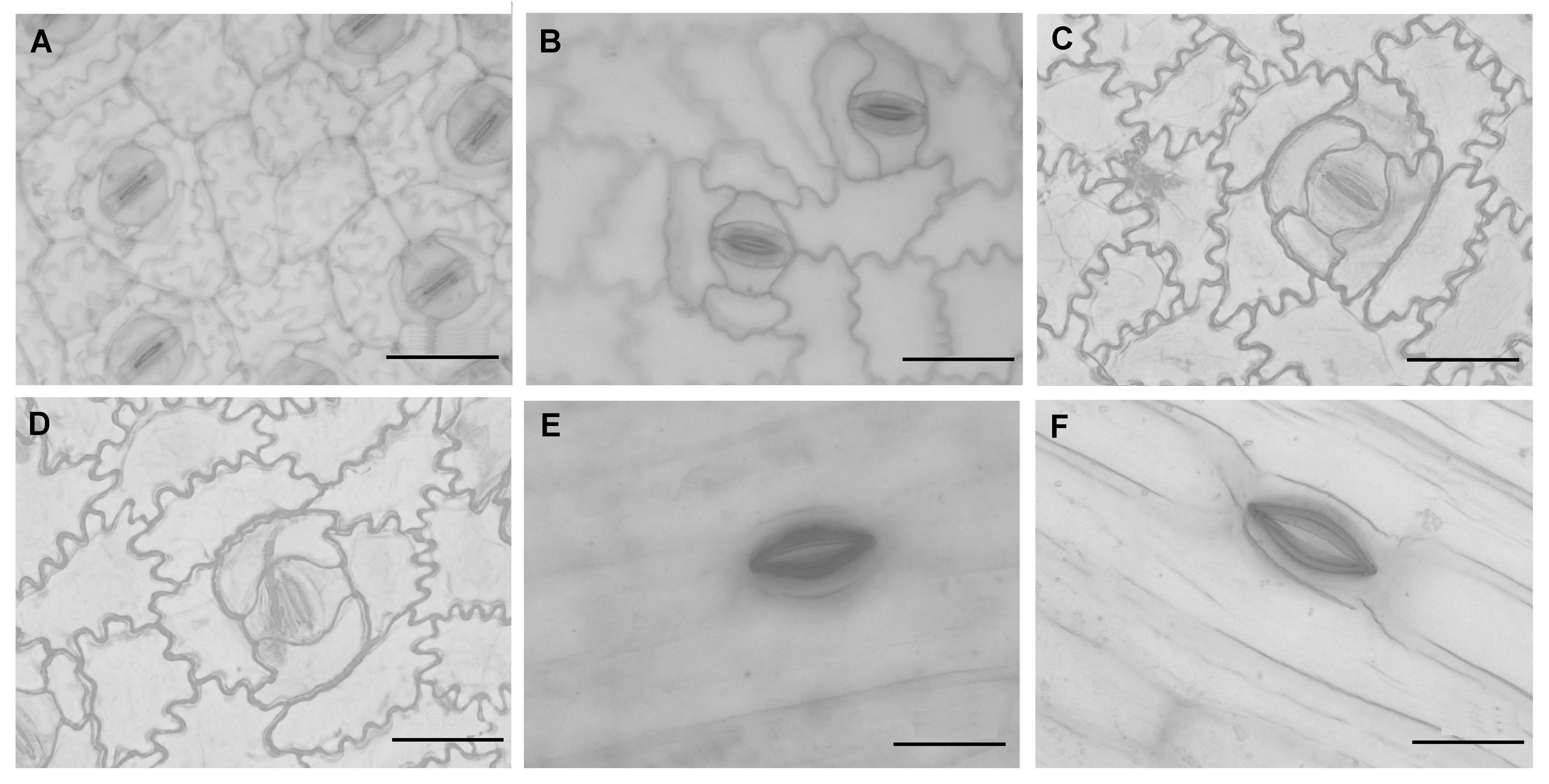
下载原图ZIP
生成PPT
图3
最佳脉冲电场作用下3种植物叶表皮气孔形态特征的显微观察(40倍物镜下3种植物的气孔特征)
(A), (B) 紫背竹芋在常态和施加脉冲电场刺激下的气孔特征; (C), (D) 绒叶肖竹芋在常态和施加脉冲电场刺激下的气孔特征; (E), (F) 朱顶红在常态和施加脉冲电场刺激下的气孔特征。Bars=50 μm
Figure 3
The stomatal shape feature of three plant species under the stimulation of high voltage pulsed electrical field with optimal combinational parameters (The shape feature of three plant species were observed under 40× objectives)
(A), (B) The stomatal feature of Stromanthe sanguinea under normal and electrostimulation conditions, separately; (C), (D) The stomatal feature of Calathea zebrina under normal and electrostimulation conditions, separately; (E), (F) The stomatal feature of Hippeastrum rutilum under normal and electrostimulation conditions, separately. Bars=50 μm
最佳脉冲电场作用下, 绒叶肖竹芋的气孔形态与对照的变化较为一致, 开合度呈增大趋势(图3C, D)。叶表皮气孔的长度与对照相比增加了55.4%, 宽度增加了209.4%, 且两者均差异显著(P<0.05); 气孔面积为对照组的3.8倍(表7; 图3C, D), 气孔密度在电场刺激前后无显著差异(P>0.05), 释放负离子的浓度均值为对照组的3.4倍, 两者差异不显著(P>0.05)。朱顶红的气孔形态与对照组相比开张状态明显(图3E, F),叶表皮气孔的长度增加了20.2%, 差异显著(P<0.05); 叶片气孔宽度比对照组缩小了10.2%; 气孔面积增加了8.4%, 且差异显著(P<0.05); 释放负离子的能力是对照组的4 696.6倍, 两者呈显著性差异(P<0.05)。
表7
Table 7
表7
| Plants | Treat- ment | Length (μm) | Width (μm) | Length/ width | Perimeter | Area (μm2) | Stomatal density (·mm-2) | Negative air ions concentration (ion·cm-3) |
|---|---|---|---|---|---|---|---|---|
| P4 | CK | 9.7±2.47 b | 1.58±0.49 b | 6.84±3.26 a | 21.2±4.92 b | 11.98±4.64 b | 250.88±31.25 a | 80±2 b |
| S | 16.09±2.81a | 2.64±0.84 ab | 6.89±3.11 a | 35.19±5.52 a | 33.01±11.27 a | 223.16±45.6 b | 1740800±195562 a | |
| P6 | CK | 12.79±2.96 b | 1.38±0.4 b | 9.93±3.42 a | 27.16±6 b | 14±5.47 b | 84.28±21.45 a | 83±3 a |
| S | 19.87±2.84 a | 4.27±1 a | 4.94±1.49 b | 44.62±5.85 a | 66.77±18.54 a | 87.48±19.28 a | 284±29 a | |
| P7 | CK | 28.07±4.77 b | 3.84±0.41 a | 7.38±1.43 ab | 60.52±9.58 b | 84.77±17.88 b | 51.14±4.66 a | 72±2 b |
| S | 33.75±2.6 a | 3.45±0.78 a | 10.46±3.62 a | 71.43±5.56 a | 91.93±23.81 a | 54.22±4.06 a | 338156±36967 a |
表7
最佳参数脉冲电场刺激下植物气孔数量特征和释放负离子的浓度(平均值±标准差)
Table 7
The negative air ions concentration and stomata quantitative feature of plants under high voltage pulsed electrical field stimulation (means±SD)
2.6 讨论负离子因具有改善空气质量和有利于人体健康等作用, 常被誉为空气中的“维生素”和“生长素”, 备受人们的关注。目前, 对植物释放负离子的研究主要集中在植物群落和室内环境单株植物两个领域。自然状态下, 植物释放负离子的能力均很弱。吴仁烨等(2011b)对自然状态下能够释放负离子的种质资源进行了调查, 发现所研究的68种植物, 释放负离子浓度均值最高的仅为73 ion·cm-3。本研究也得出了相似的结果, 即常态下植物释放负离子浓度值远低于宜居室内的标准值(1 500 ion·cm-3) (李安伯, 2001), 因此无法满足人们对健康的需求。但对植物施加脉冲电场刺激时, 其释放负离子的能力可大幅提高。吴仁烨等(2015)对蟹爪兰(Zygocactus truncatus), Tikhonov等(2004)对中华芦荟(Aloe vera var. chinensis)施加脉冲电场刺激, 发现其释放负离子的能力与常态相比均显著提高。习岗和杨运经(2008)以及杨运经等(2009, 2011)也进一步验证了该现象。本研究发现, 对植物施加不同强度的脉冲电场刺激, 其释放负离子的能力存在较大差异, 其中紫背竹芋在最佳脉冲电场A3B3C3作用下释放负离子的能力是最低处理A1B1C1的3.8倍。在不同参数脉冲电场作用下植物释放负离子的能力不同, 要使植物释放负离子的能力显著提高, 需给植物施加足够强的脉冲电场。但另一方面, 如果电场强度过大, 会造成植物细胞的不可修复性损伤(习岗和杨运经, 2008; 杨运经等, 2011)。在最佳脉冲电场作用下, 植物释放负离子的能力最强, 施加低于或高于此最佳脉冲电场, 其释放负离子的能力均有不同程度的降低, 原因可能在于低频电场生物学效应具有“阈值”与“功率窗”的特性(习岗和杨运经, 2008)。每种植物都有适合其本身高效释放负离子的最佳脉冲电场, 在这种强度和频率的电场刺激下, 植物生长状态良好, 释放负离子的能力显著提高且其本身也未受到损伤。
同时, 本研究发现, 绒叶肖竹芋在各强度脉冲电场作用下, 释放负离子的浓度均值变化幅度很小。有研究表明, 植物本身具有特定的电位特征频率, 如果施加的脉冲电场适宜, 则会与植物电场共振进而提高释放负离子的能力(习岗和杨运经, 2008; 杨运经等, 2009, 2011)。因此, 我们推测可能施加的脉冲电场频率与绒叶肖竹芋固有的电位频率不符, 未使其发生电场共振, 从而使释放负离子的倍增效应不明显。
对3种植物施加最佳脉冲电场作用的研究结果表明, 植物体上的电压值与释放负离子的能力呈正相关, 即植物体所储存的电压越高, 其释放负离子的能力越强。以紫背竹芋为例, 当植物体电压由5.13×103 V降为0.51×103 V时, 其释放负离子的能力接近自然状态下的水平(89 ion·cm-3)。我们认为储存在植物体上的电压相当于施加给植物体的“能量”, 这个能量需达到激发该植物生物学效应具有的“阈值”或“功率窗”, 即该脉冲电压正是打开植物释放负离子通道所必需的“能量”。
与脉冲电场作用对植物释放负离子的影响相似, 光照在一定程度上也可提高植物释放负离子的能力, 光照度是影响植物群落释放负离子能力的主要因素之一(刘新等, 2011)。Wang和Li (2009)对芦荟等植物进行光照刺激, 发现芦荟对光照的响应最敏感, 随着光照度的增加, 其释放负离子的浓度均值显著升高。本研究中的3种植物在不施加脉冲电场刺激时对光照响应不太敏感, 随着光照度的增强其释放负离子的浓度均值均呈上升趋势, 但绝对增幅很小, 原因在于不同植物在自然状态下释放负离子的浓度存在差异, 且对光照的反应也不同。当施加最佳脉冲电场时, 3种植物在不同光照度下释放负离子的能力均出现激增效应, 原因可能是光照激发了植物叶片表层的光电效应, 使大量自由离子从植物体逃逸而产生负离子。另外, 本研究还发现, 3种植物在最佳脉冲电场作用下, 光照度为3 000和6 000 lx时释放的负离子浓度无 显著差异(P>0.05); 而当光照度小于3 000 lx或大于 6 000 lx时, 释放的负离子浓度均值均差异显著(P< 0.05)。我们认为这两个强度的光照所提供给植物体的“能量”可能位于同一级别, 与3 000 lx的光照度相比, 6 000 lx的光照度还未能激活植物体更高一阶的负离子释放能力。
脉冲电场作用后, 植物叶片气孔的数量特征与自然状态下相比均有不同程度的差异, 3种植物的叶片气孔面积在脉冲电场刺激后均呈增加趋势。以紫背竹芋为例, 在施加脉冲电场前后, 其气孔面积分别为11.98和33.01 μm2, 释放负离子的能力提高了21 759.0倍。此外, 气孔密度也是影响植物释放负离子的一个重要因素, 3种植物中以紫背竹芋的气孔密度最大, 脉冲电场作用下释放负离子的能力最强, 原因可能在于叶片气孔是植物释放负离子的一个主要通道。张万超等(2016)对银冠(Cotyledon orbiculata var. oblonga)等3种仙人掌科植物的研究表明, 刺尖的数量和气孔均是植物释放负离子的通道。习岗和杨运经(2008)提出了植物通过气孔蒸腾放电释放负离子的假说。我们的实验结果在一定程度上证实了气孔作为植物释放负离子重要通道的假说, 气孔可能通过蒸腾放电, 聚集在植物叶片上的阴离子通过植物叶片的气孔蒸腾释放出来而形成负离子。同时本研究发现, 绒叶肖竹芋的气孔面积比对照提高了3.8倍, 而负离子的释放量却很低, 仅为284 ion·cm-3。这可能与不同植物对脉冲电场刺激的承受能力不同和气孔类型有关, 实验中所施加的脉冲电场强度可能未达到绒叶肖竹芋释放负离子所需“功率窗”的阈值, 尽管气孔面积有所增加, 但所施加的“能量”未能使释放负离子的通道完全打开。因此, 如何提高植物气孔的开张度, 并保证所施加的脉冲电场强度在提高植物释放负离子能力的同时又不对植物造成损伤, 是植物释放负离子研究工作的首要问题。
本文从植物释放负离子的最佳脉冲电场, 植物体上的电压、光照度及植物叶片气孔特征等方面, 初步分析了其与植物释放负离子能力间的关系, 其中以脉冲电场的影响最为显著。在自然状态下植物释放负离子的能力很弱, 施加脉冲电场后其能力显著增强, 且植物具有高效释放负离子的最佳脉冲电场。在该最佳脉冲电场作用下, 随着光照度的增加植物释放负离子的能力呈上升趋势。此外, 植物释放负离子的能力与其携带的电压呈正相关, 电压越大, 植物释放负离子的能力越强; 另外, 植物释放负离子的能力也受叶片气孔面积和气孔密度等特征, 及其它外界因素(温度、湿度和气压等)和植物本身特性的影响。因此, 在后续工作中, 我们希望能将以上几个因素综合起来进行更为系统的研究。
参考文献
文献选项
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [9] | URL室内空气污染物是多种多样的,进行空气采样和分析,藉以评价空气质量是必要的,分析过程是比较繁复费时的,采用空气离子测量评价法,可在短时间内获得空气洁净与否的初步印象。 [本文引用: 1] |
| [10] | URL对北京典型园林植物区香山、颐和园、植物园等地开展了不同季节空气负离子浓度的测定,并结合空旷地和市区的观测结果进行了对比分析。结果表明:园林植物的存在可以明显提高空气负离子的浓度,有植被地区空气负离子数值明显高于无植被区。空气负离子浓度随海拔高度的增加呈单峰曲线形式,与各海拔高度的植被状况关系密切,并随着空间梯度的增加而减小。空气负离子浓度在园林植物区日动态变化明显,呈双峰曲线形式。不同生境空气负离子浓度随季节不同变化明显,夏秋季节浓度较高。空气负离子浓度与温度和相对湿度成正相关关系。 [本文引用: 1] |
| [11] | [本文引用: 2] |
| [12] | URL2006年3月至2007年2月,对佳木斯绿地的空气负离子浓度和相关气象因子进行同步观测,并分析了二者的关系.结果表明:研究区空气负离子浓度的日变化和月变化较明显;清晨和午夜前后,该区空气负离子浓度较高,中午和傍晚前后,空气负离子浓度较低,最低时只有217个.cm-3;7月的空气负离子浓度(782个.cm-3)最高,12月最低(283个.cm-3);该区空气负离子浓度以夏季最高,春、秋两季相当,冬季最低;空气负离子浓度与温度呈极显著负相关,与相对湿度、气压呈极显著正相关. [本文引用: 1] |
| [13] | DOI:10.3321/j.issn:1000-2421.2008.02.029URL对上海地区常见的21个群落距地面1.5m的空气正、负离子浓度进行了测试分析,结果表明:群落增加空气负离子浓度极显著高于水泥地,经多重比较发现草坪提高空气负离子浓度的能力最弱,竹类群落提高空气负离子浓度的能力最强;空气质量指数CI值呈正态分布,CI值在0.5~0.7的群落最多;竹类群落的CI最高,均值为0.83,比草坪CI值提高了117%;针叶群落与阔叶群落的CI相近,比草坪CI值提高了70.7%;CI与空气负离子浓度呈极显著正相关,与群落叶面积指数呈显著正相关,与群落优势种的胸径、树高无关;聚类分析结果表明,植物群落可明显改善微环境的空气质量,其中改善空气质量能力较高的群落为群落8、15、16、17等,改善空气质量的能力较差的是草坪、群落12、群落9、群落7。 [本文引用: 2] |
| [14] | URL为研究福州市鼓山茶园不同生境空气负离子浓度空间分布规律以及空气负离子与其环境因子的关系,以茶园内7种生境作为研究对象,对各生境空气负离子浓度及环境因子进行测定。结果表明,茶园内7种生境空气负离子浓度由高到低为乔木群落高海拔茶园池塘有机茶园草地空地常规茶园,其平均负离子浓度分别为924、892、770、761、754、617、601个/cm3;在观测高度范围内(50~150cm),空气负离子浓度大致存在随海拔高度升高而增加的趋势,且150cm高度处空气负离子浓度显著大于50cm高度;各生境空气负离子与相对湿度呈极显著正相关关系,与温度、风速、气压及海拔相关性不显著;鼓山茶园空气质量总体较为清洁,各生境空气质量评价系数CI均在0.50以上,且空气质量评价系数CI值大致存在随海拔高度而增加的趋势。 [本文引用: 1] |
| [15] | DOI:10.3969/j.issn.1001-4241.2000.05.004URL近年来,空气离子特别是空气负离子的保健作用受到人们的普遍关注,空气离子浓度也是衡量空气清洁度的重要指标之一,森林地区空气负离子浓度明显高于其它地区,成为重要的森林旅游资源。文中就空气负离子的来源,时空变化、森林与空气负离子的关系,空气负离子的保健及治疗功能,以及以空气负离子浓度为指标的空气清洁度的评价标准等一些问题进行了综述。 [本文引用: 1] |
| [16] | DOI:10.3969/j.issn.2095-0756.2010.02.004URLA comparison of 1.5 m above ground aero-anion and positive air ion concentration for six plant communities,Sapindus mukorossiMagnolia grandifloraPhyllostachys praecoxCunninghamia lanceolataLagerstroemia indicaS. mukorossiP. praecoxC. lanceolataL. indica<0.01) positive correlation (= 0.702) between aero-anion concentration and relative humidity,a significant(<0.05) negative correlation( = - 0.637) between aero-anion concentration and temperature,a negative correlation( = - 0.191) between aero-anion concentration and solar radiation intensity,and a significant(<0.05) positive correlation( = 0.580) between aero-anion concentration and ultraviolet radiation intensity(UVB).[Ch,2 fig. 3 tab. 15 ref.] [本文引用: 1] |
| [17] | DOI:10.3969/j.issn.1000-8535.2010.02.009URL目的探讨低强度运动匹配高浓度 负离子对老年高血压患者血压及血脂的影响。方法122例老年轻、中度高血压患者随机分为治疗组58例和对照组64例,两组均给予非洛地平缓释片5 mg/d+贝那普利片10 mg/d口服,治疗组在服药同时予每周4次以上的低强度运动匹配30~60 min在高浓度负离子区的空气浴,4周后依血压情况调整服药剂量,随访6个月,观察两组血压、血脂及服药量的差异。结果4周后治疗组降压效果优于对照组 (P0.05),血压控制达标率67.2%vs 48.4%,高于对照组(P0.05);6个月后在血压均控制在目标值时,治疗组服药剂量明显低于对照组(P0.01),药物不良反应发生率明显低于对照 组(P0.01),治疗组血脂明显下降(P0.05),对照组血脂无明显变化(P0.05)。结论低强度运动匹配高浓度负离子对老年高血压患者具有降压及 降血脂作用。 [本文引用: 1] |
| [18] | DOI:10.5846/stxb201301180115URL城市森林内PM2.5浓度的状况可以直接反映城市森林对PM2.5的净化效果,也是居民休闲游憩关心的森林环境问题。选择北京西山3种典型的游憩型城市森林,通过对林内PM2.5浓度一年四季昼夜24h内变化的同步观测,分析了不同类型城市森林内PM2.5浓度的季节变化、日变化以及影响因素,结果表明:(1)北京西山3种游憩林内PM2.5浓度多数时候远低于城区对照值,在春、夏、秋三季都达到了国家城市化地区的标准,甚至在春季、秋季还达到了国家一类地区的标准。(2)城市森林在不同季节对PM2.5的净化效果存在差异,林内PM2.5浓度总体上呈现冬季夏季秋季春季的规律。(3)林内PM2.5浓度在一天24h内有很大变化波动,夜间浓度总体上高于白天,日变化曲线近似呈"双峰双谷"型,两个高峰出现在夜晚和早上,两个低谷出现在凌晨和中午前后。一年四季白天低谷出现时间有所不同,春季15:00左右、夏季13:00—17:00、秋季13:00—15:00、冬季9:00—11:00。(4)PM2.5在不同类型游憩林内的变化趋势和浓度值存在一定差异。郁闭度较大的侧柏林夜间PM2.5浓度总体上高于其它两种林型,其高峰和低谷出现时间延迟,高峰值大,高峰期持续时间长,且这种规律在秋季表现得更明显。(5)基于上述研究认为,北京西山城市森林为居民在PM2.5污染比较突出的都市背景下提供了一个相对清洁、健康的森林游憩环境,春季、夏季、秋季全天以及冬季9:00—11:00均是森林中PM2.5状况健康而适宜外出游憩的时段。 [本文引用: 1] |
| [19] | DOI:10.3969/j.issn.1674-5906.2014.06.011URL大气离子是由许多自然和人为的原因产生,并且它们的浓度在不同环境场所下差别很大。选取我国南部沿海某省份作为研究区域,挑选近10处具有代表性的环境场所进行实测研究,收集了空气正、负离子浓度、风速、空气温度、相对湿度、材料和植物负离子浓度等数据。实测数据表明:地面上的空气负离子浓度随着地理环境因素(瀑布、海边、峡谷、乡村田野、郊区旷野、县城等)不同差别很大,其中瀑布海边峡谷溪流县城;同时空气负离子浓度与风速、水、植物、相对湿度等有较为密切的关系。通过对实测数据进行分析得出以下结论,1)空气的摩擦可以有效显著地增加空气中负离子的浓度,两者呈现出一定的正相关关系。2)水体在撞击和喷射过程能加快正负电荷的分离,水速流动地越快,相应摩擦产生的电离能越大,周边环境的空气负离子浓度就越高;同时随着距离的衰减,环境中的空气负离子浓度也在不断地降低。3)空气负离子浓度与植物群落的种类结构和配置相关,其中高处复层结构植物低处复层结构植物低处单层结构植物。4)相对湿度与空气负离子具有良好的相关性。5)温度与空气负离子的关系有待进一步明确。6)适宜的温度、湿度以及风速能使人感到舒服,有益于人体的健康。7)自然生态环境整体而言,空气负离子浓度大,空气清洁度均为A级最清洁。通过结果和分析给城市生态环境建设方面以重要的启示,因此作者从城市规划层面进一步提出以下建议:1)以自然环境的空气负离子浓度值为参考标准研究城市环境的空气负离子浓度和空气清洁度,进行符合气候和生态资源运行状态的设计,对于优化城市生态环境和城乡规划建设的水平有着重要的意义。2)加强自然通风、增加水体景观设计以及扩大绿地并减少不必要的硬质广场或铺装,从而不断激发并保持周边环境的空气负离子浓度,减少空气污染,有效地改善城市生态环境。 [本文引用: 1] |
| [20] | DOI:10.5846/stxb201305301239URL受城市扩张、工业发展、汽车保有量增加的影响,空气颗粒物目前已成为诸多城市空气的首要污染物。而城市森林作为城市生态建设中最大的唯一具有自净功能的生态系统,不仅为城市高污染环境下的居民提供了相对洁净的休闲游憩空间,还对净化空气颗粒物起重要作用。从城市森林调控空气颗粒物的机理、分析方法、植物个体、群落及不同类型城市森林调控空气颗粒物的功能差异及其时空变化规律等方面进行阐述。结果表明:目前有关物理降尘的研究较多,涉及化学除尘的研究尚缺;应用重量法测定植物滞尘量的研究较多,质量浓度法测定城市森林净化空气颗粒物功能的研究较少。在未来一段时间,森林植被化学除尘的过程与机理,城市森林调控空气颗粒物的多方面、系统性研究及相关研究成果的转化与实际应用将会是重要的研究方向。 [本文引用: 1] |
| [21] | [本文引用: 1] |
| [22] | DOI:10.11707/j.1001-7488.20010513Magsci<p>选择南方有代表性的森林游憩区进行森林环境中空气负离子水平研究,结果表明:(1)不同林分类型的森林环境中空气负离子水平差异较大。(2)森林环境中空气负离子浓度与温度成显著负相关,与空气相对湿度成正相关。(3)森林游憩区中水体对负离子水平的影响较大。其中动态水大于静态水,瀑布大于溪流。(4)森林中空气负离子浓度还呈现出一定的日变化和年变化。(5 )建筑材料对空气负离子水平也有一定的影响。</p> [本文引用: 1] |
| [23] | DOI:10.1017/S0266078400010713URL在南岳衡山马尾松纯林内,设立了立地条件相类似的15个测点,测定空气负离于浓度及相应的林分因子,采用数量化理论Ⅰ模型的分析表明,树龄、郁闭度、树高为影响空气负离子浓度的主导因子,且相关系数显著。 [本文引用: 1] |
| [24] | |
| [25] | DOI:10.3969/j.issn.0517-6611.2014.27.084URL[目的]研究改善室内空气质量的新途径.[方法]在自然状态和放置植物源负离子发生器状态下,对玻璃室内的负离子、正离子和安倍指数等指标进行研究.[结果]在自然状态下,玻璃室内负离子浓度均值较低,为1 084 ion/cm3;在放置植物源负离子发生器状态下,负离子浓度均值为684 190 ion/cm3,是自然状态下的631倍.正离子在自然状态下的浓度均值为946 ion/cm3,在植物源负离子发生器开启状态下值为976 ion/cm3,两者差异不显著(P>0.05).从安倍指数看,植物源负离子发生器开启状态下值为685 457.62,是自然状态下的539 730倍.[结论]在玻璃室内放置植物源负离子发生器可显著提高室内空气质量. [本文引用: 1] |
| [26] | [本文引用: 2] |
| [27] | |
| [28] | DOI:10.3969/j.issn.1671-5470.2014.05.012URL脉冲电场作用下对丰两优1号(r1)等6个水稻品种释放负离子的浓度特性进行研究.结果表明:在自然状态下,6个水稻品种的植株释放负离子的能力很弱,负离子浓度最高为特优420(22 ion·cm^-3),最低为冈优527(15 ion·cm^-3).适当的脉冲电场刺激水稻根部,6个水稻品种较常态下释放负离子的能力均大幅度提高,以脉冲电场u2(脉冲电压15 kV,脉冲频率1Hz,脉冲宽度50ms)的效果最大,释放负离子浓度最高为冈优527(r2),达4168100 ion·cm^-3,是对照组刺激无水稻植株的土壤(r7)的24959倍,是自然状态下(未刺激)的208405倍;最低为糯广优201(r3),为2786000ion·cm^-3,是对照组刺激无水稻植株的土壤(r7)的16683倍,是自然状态下(未刺激)的139300倍.以同一强度脉冲电场对不同生长期的水稻进行刺激,以四叶期的释放量最大,抽穗期阶段均出现下降趋势. [本文引用: 2] |
| [29] | DOI:10.3969/j.issn.1000-0712.2008.11.014URL由于电力和通讯的快速发展,人类面临着日益严重的电磁污染,低强度电磁场对生物体系的非热效应引起人们的关注.本文介绍了极低频电磁场和射频电磁场对生物体系非热效应及其机理研究,并提出了一些看法. [本文引用: 3] |
| [30] | URL现代城市中的室内空气污染日益严重,室内空气污染的净化与治理已成为社会发展中迫切需要解决的关键科技问题之一。为此,根据植物电信号的低频特征与电共振原理,利用极低频负高压脉冲连续刺激盆栽吊兰植物的根际土壤,使吊兰叶片产生了显著的空气负离子倍增效应。在此基础上依据评价空气质量的常用指标:单极系数q、安培空气质量评价系数CI、空气负离子系数P和森林空气离子评价指数FCI,对应用负高压脉冲技术提高植物的空气净化能力进行了计算与分析。结果表明,极低频负高压脉冲电场可以有效地提高植物的空气净化能力,提出了极低频负高压脉冲电场介导的植物空气净化新技术。 [本文引用: 2] |
| [1] | DOI:10.3969/j.issn.1000-6850.2013.22.006URLHaving made considerable progress in recent years, the urban forestry construction is gradually transforming its concerns from focusing on function and aesthetics to eco-health function. Due to the lack of researches on the eco-health function of most-used plants species, in this study, five typical urban green species (Populus tomentosa, Salix matsudana, Pinus tabulaeformis, Platycladus orientalis, Poa Annua and arbor -shrub-grass green space) in three representative urban green spaces were monitored for the concentration of air anion, the air temperature and relative humidity, the concentration of carbon dioxide (CO2), and the noise level at different locations of green spaces in different seasons. Some conclusions can be seen from the results: (1) compared to the five single-specie arbor groves, arbor -shrub-grass green space has better eco-health function. Using the concentration of CO2 as an example, the highest concentration of air anion was found in a arbor -shrub-grass green space(435 10-6) ;(2) deciduous trees has better eco-health function in foliaceous period, while showed poor eco-health function in foliage-falling period. Using the concentration of air anion as an example, the highest number was found in deciduous tree groves(Salix matsudana) 1500 cm-3; (3) in terms of maintaining the eco-health function, evergreen conifer groves, represented by oriental arborvitae (Platycladus orientalis), showed a more steady result. The difference of the air anion concentrations is 154cm-3 ,which is half of the difference of deciduous specie(Poplus tomentosa) [本文引用: 1] |
| [2] | DOI:10.3969/j.issn.1001-7461.2015.01.39URL为探讨城市绿地植物群落差异对空气负离子特征的影响,研究了5种 不同典型城市绿地植物群落:次生常绿阔叶林群落(SF)、人工改造常绿阔叶林群落(IF)、人工常绿阔叶景观林群落(CF)、人工稀树灌草群落(OF)、 交通绿岛群落(TA)下空气负离子浓度及空气清洁度,并以缺乏绿色植被的火车站广场(RS)作为对照.结果表明,空气负离子浓度和空气清洁度在不同植物群 落和时间段内存在显著差异(p<0.05).其中,城市绿地植物群落中空气负离子浓度日变化普遍呈现双峰曲线;群落结构相对较为复杂的SF群落空气负离子 浓度和空气清洁度最为优秀.总的来说,空气负离子浓度从大到小排序如下:SF>IF>CF>OF>RS>TA;而空气清洁度则为SF>CF>IF>OF> RS> TA;回归分析表明,大气相对湿度显著影响空气负离子浓度(R2=0.478,p<0.05). [本文引用: 1] |
| [31] | DOI:10.3969/j.issn.1000-0712.2009.12.013URL介绍了空气负离子(NAI)的物理概念及其在空气净化中的作用,讨论了正常生态条件下空气中NAI的来源.通过实验结果说明了脉冲电场可以使植物产生NAI发射的倍增效应,分析了其产生的机理. [本文引用: 1] |
| [32] | URL研究了广州城市绿地空气负离子水平以及绿地类型、水体状态、海拔高度、天气阴睛、人类活动等因子对空气负离子浓度的影响。结果表明:广州城市绿地空气负离子浓度在103.3367个·cm^-3,平均为426个·cm^-3,与上海、深圳等地接近。所测定的几种绿地类型空气负离子浓度大小顺序为竹林〉小叶竹柏林〉花卉区〉隆缘桉林〉苗圃和草坪〉住宅区;阔叶林〉针阔混交林〉针叶林。动态水的空气负离子浓度显著高于静态水;在本研究观测范围内,离动态水越远,负离子浓度越低。随着海拔高度增加,空气负离子浓度呈下降趋势。晴天的空气负离子浓度显著高于阴天。人流量和车流量与空气负离子浓度呈显著负相关。 [本文引用: 1] |
| [3] | DOI:10.3969/j.issn.1672-0873.2013.12.031URL以临安市青山湖湿地公园典型植 物群落常绿阔叶林、香樟林、池杉林及人工林为研究对象,研究4种不同植物群落空气负离子效应及其空气清洁度的比较。研究结果表明,不同植物群落空气负离子 浓度大小排序为常绿阔叶林人工林香樟林池杉林,常绿阔叶林空气负离子浓度最高,环境调节功能最强,其对改善湿地公园环境质量发挥着重要的作用。 [本文引用: 1] |
| [4] | URL通过对南宁城区、城郊绿地及农田开发区进行负离子含量的测定,找出不同植被配置类型空气负离子效应的差别。结果表明,就空气负离子而言,城郊大规模绿地的空气质量农田开发区城区;植被配置的复层结构(乔灌草)简单植被配置结构(乔灌、乔草、灌草)单一配置结构(草坪、稀乔、稀灌草)。溪流和瀑布对增加负离子浓度的作用显著。随着海拔和郁闭度的增加,空气负离子含量有上升的趋势。空气负离子含量随季节有一定的波动。 [本文引用: 1] |
| [33] | DOI:10.5846/stxb201209181316URL选取上海环城林带典型的植物群落,分别在夏季和冬季对群落的温湿度调节、负离子提升、抑菌、降噪和大气净化等保健功能进行测定和评价;同时结合群落结构因子,分析群落结构与保健功能之间的关系,并探讨了群落物种组成和结构特征对保健功能的影响,以期揭示群落保健功能发挥的机制。结果表明:夏季,落叶针叶林池杉群落除抑菌功能以外,在其他保健功能上均具有较好的效果,而落叶阔叶林全缘叶栾树群落和悬铃木群落效果较差,其他类型的群落居中;冬季,常绿阔叶林女贞群落在各种保健功能方面均具有较好的效果,其次为常绿落叶阔叶混交林的全缘叶栾树+香樟群落和杂交杨+香樟群落,其他类型的群落效果较差。群落各种保健功能与群落结构特征的相关性分析表明,叶面积指数和郁闭度在夏冬两季均与多种保健功能呈显著正相关,是指示群落保健功能优劣的重要结构因子。研究结果表明人工森林群落的保健功能可通过调整群落结构来提升,为城市森林的群落建构与管理提供了科学依据。 [本文引用: 1] |
| [34] | URL [本文引用: 1] |
| [5] | DOI:10.3969/j.issn.1001-7461.2016.01.49URL为获得崇明岛不同生态用地空气负离子的实时变化趋势,以崇明岛林地、绿地、湿地和对照点为研究对象,利用Epex空气离子测量仪进行实时监测,并采用单级系数和安倍空气质量评价系数进行空气等级评价。结果表明:不同生态用地空气负离子浓度的时间变化趋势有所差异,正离子浓度变化趋势与负离子不完全相同;空气负离子均值浓度表现为夏季:湿地〉绿地〉林地〉对照;秋季:湿地〉林地〉绿地〉对照;夏季和秋季湿地空气负离子浓度显著高于其他3个测点,且均以对照点空气负离子浓度最低;所有观测点空气负离子的单级系数q〈1,空气负离子浓度基本达到1 000个·cm~(-3),空气质量良好。 [本文引用: 1] |
| [6] | URL空气负离子(NAI)是综合反映空气质量的重要指标,对人居环境有重要意义.本文综述了城市空气负离子时空分布特征,并根据不同环境因子对其理化过程的影响及其在城市中的特点探讨了城市NAI时空分布特征的成因:NAI分布的时间动态主要受控于太阳辐射的周期变化;空间分布的城乡梯度差异受城市气溶胶、下垫面性质及城市热岛效应影响;城市绿地的高NAI浓度与植被生命活动和土壤辐射有关;近水环境中NAI浓度较高的原因在于水分子通过多种途径参与NAI生成过程;其余环境因子可在一定程度上影响NAI的生成、寿命、组分、迁移和分布;增加城市绿化面积和大气湿度、保持下垫面土壤自然属性均可有效地提高NAI水平,改善城市空气质量. [本文引用: 2] |
| [7] | [本文引用: 1] |
| [8] | URL一个世纪以来,有关空气离子的研究经历了30,50,70年代三次浪潮。70~80年代大家的愿望和兴趣转到空气离子对细菌、原生动物、昆虫、动物、植物和人类的生物学作用,曾发生过一些学术上的争论。近10年(90年代)继续开展空气离子研究的国家有:中国、俄罗斯、美国、加拿大、日本、德国、丹麦、英国、希腊、波兰、罗马尼亚和南斯拉夫等。研究领域涉及空气离子对生物机体的生物学效应、应激反应、情感精神、听力、免疫和环境空气离子测量等。 1. 空气离子的生物学效应 1.1 空气负离子与组织细胞线粒体赖能(energy dependent)过程 线粒体嵴膜三分子体镶嵌磷脂和结构蛋白,是ATP酶、ADP生成ATP偶联过程和电子传递系统之所在,线粒体的氧化磷酸化依赖于内膜三分子的完整性。Staurouskaia研究大鼠吸入空气负离子后肝脑心匀浆中线粒体赖能过程,发现负离子可改善线粒体的呼吸率,摧毁Ca2+在线粒体内累积,提高ADP/O比值[1]。Temnov将大鼠肝匀浆及媒体冰冻3 h,保持线粒体原有结构,然后向线粒体匀浆和媒体发射空气负离子流。发现线粒体很快形成大的聚合联接体,结构完整,用磁力搅拌器搅拌数分钟也不能打散或消失,同时测定赖能过程加强。空气负离子在加强线粒体氧化磷酸化,调节呼吸率时,主要是保存和维护线粒体三分子体结构的完整性[2]。 [本文引用: 3] |
| [35] | DOI:10.3969/j.issn.1000-2561.2016.07.010URL在室内密闭玻璃箱内测定了3种仙人掌科(Cactaceae)植物,即银冠(Cotyledon orbiculata var.oblonga)、玉翁(Mammillaria hahniana)和金晃(Notocactus leninghausii),在高压脉冲电刺激下不同剪刺阶段负离子释放浓度、刺数量和气孔数量特征,光镜观察了不同剪刺阶段气孔的形态特征,探讨了高压脉冲电场刺激下仙人掌科植物负离子释放量与释放通道的相关性.结果表明:在高压脉冲电刺激下,银冠负离子浓度随气孔长度的增大而升高(p<0.01),玉翁负离子浓度随其刺数量的减少而降低(p<0.0l),随气孔长度的增大而升高(p<0.01),金晃负离子浓度随其刺数量的减少而降低(p<0.01),随气孔宽长比的增大而升高(p<0.01).从结果推断,在高压脉冲电刺激下,仙人掌科植物释放负离子的过程受其生理活动的影响;仙人掌科植物的刺尖和气孔都是其释放空气负离子的通道,当植物生理活动强度大到改变了其气孔数量特征参数时,气孔就可能成为主要的释放通道. [本文引用: 1] |
| [36] | DOI:10.3969/j.issn.0517-6611.2008.29.009URL针对NaOCl法在叶表皮制片上存在的问题,对叶表皮制片方法和工具进行了一些改进,使用改进的方法和工具进行了植物叶表皮制片.结果表明,改进法可提高制片效率,获得较好的制片效果,并有利于初****使用. [本文引用: 1] |
| [37] | DOI:10.1088/0256-307X/15/12/025URL [本文引用: 1] |
| [38] | DOI:10.1007/BF02187010URL [本文引用: 2] |
| [39] | [本文引用: 1] |
| [40] | DOI:10.1109/27.842910URLAbstract The primary chemical mechanism of the beneficial medical/biological action of negative air ions necessary for life was studied. Air ion deficiency is the cause of many illnesses and treatment with air ion inhalation is effective in many cases. However, its application is limited by the absence of knowledge of the primary mechanism of its action. The superoxide anion O2- was detected in the flow of negative air ions generated by an electroeffluvial air ionizer. Earlier, the appearance of hydrogen peroxide in solutions treated with air ions was shown. The presence of these reactive oxygen species in ultralow and low concentrations (10-12-10-6 M) suggested that the primary mechanism for the beneficial medical/biological action of negative air ions is moderate activation of free radical peroxidative oxidation within a physiological range that is lower than in tissues under pathology. It was shown in patients that treatment with inhalation of negative air ions did not induce pathological changes in superoxide dismutase activity and, under simultaneous administration of a food antioxidant, led to its fluid increase. The latter, along with some previous results, supports the proposed mechanism. In addition, taking the proposed mechanism into consideration, air ion doses for treatment can be selected on an individual basis and should depend on the redox state of the patient. This should achieve better results for medical treatment with ionized air |
| [41] | DOI:10.1016/S0014-5793(97)00651-0PMID:9237652URLAbstract The activity of erythrocyte cytosolic superoxide dismutase from rat, bovine, man and duck was considerably increased when measured after preparation or incubation in media pretreated with negative air ions (mostly superoxide) from electroeffluvial ion generator. 0.5-1.0 microM H2O2 was found in incubation medium after treatment with air ions. The stimulatory effect of air ions on superoxide dismutase activity was mimicked by addition of 0.5-6 microM H2O2. The primary physicochemical mechanism of beneficial biological action of negative air ions is suggested to be related to the stimulation of superoxide dismutase activity by micromolar concentrations of H2O2. [本文引用: 1] |
| [42] | DOI:10.1085/jgp.45.4.233PMID:01URLAlthough gaseous ions possess no obvious physical properties which would predict a capability to influence living cells or tissues they can be shown to produce functional changes. The physiological effects so far demonstrated are limited in extent. In the mammalian trachea air ion action appears to depend upon the release of bound 5-hydroxytryptamine by (+) and upon O(-)-induced acceleration of its metabolic destruction. [本文引用: 1] |
| [43] | DOI:10.1007/BF01553616PMID:4571990URLKrueger AP. [本文引用: 1] |
| [44] | DOI:10.1007/BF02189651PMID:4055123URL[Abstract unavailable] [本文引用: 1] |
| [45] | DOI:10.1126/science.959834PMID:959834URLAbstract The thrust of the experimental data presented here is that small air ions are biologically active. There is convincing evidence that both negative and positive ions (i) inhibit growth of bacteria and fungi on solid media; (ii) exert a lethal effect on vegetative forms of bacteria suspended in water when opportunity is provided for contact of cells and ions; and (iii) reduce the viable count of bacterial aerosols. Through physical action, ions of either charge upset the stability of aerolosized bacterial suspensions and, in addition, have a direct lethal effect which is more prominent with negative ions than with positive ions. With regard to the serotonin hypothesis of air ions action, the situation is more complex. The essential fact is that mice and rats display a charge-related metabolic response to air ions and this phenomenon also occurs in humans. Because serotonin is such a potent hormone, the ultimate functional changes incident to air ion action are impressive and account for the signs of symptoms of the sharav syndrome. Alterations in the cumulative mortality rate with three experimental respiratory disease in the mouse also are charge-dependent, positive ions routinely exercising a detrimental effect. Further, in the case of mice infected with influenza virus, ion-deprivation increases the cumulative mortality rate. Since ion depletion is a constant concomitant of modern urban life, one reasonably may speculate about comparable inimical effects on humans. [本文引用: 2] |
| [46] | DOI:10.1007/s11676-014-0475-9URL [本文引用: 1] |
| [47] | DOI:10.1016/S0167-8760(02)00067-3PMID:12374649URLThe effects of negative air ions on computer operation were examined using a biochemical index of the activity of the sympathetic/adrenomedullary system (i.e. salivary chromogranin A-like immunoreactivity (CgA-like IR)) and a self-report questionnaire (State-Trait Anxiety Inventory, Anxiety State TAI-S). Twelve female students carried out a word processing task for 40 min. The salivary CgA-like IR increased more than three times on the task, but the salivary cortisol did not change. The increase in the CgA-like IR level was attenuated by the exposure to negative air ions during the task. The exposure to the ions during the recovery period following the task was effective for rapidly decreasing the CgA-like IR level that had increased after the task. These effects by negative air ions were also observed using STAI-S. Task performance was slightly but significantly improved by the presence of negative air ions. These results suggest that negative air ions are effective for the reduction of and the prompt recovery from stress caused by computer operation. [本文引用: 1] |
| [48] | PMID:11855283URLIt was shown that plantlets of wheat (Triticum vulgare) are capable of generating negative aeroions during the electrization of soil by high-voltage impulses. Soil electrization was carried out either from the moment of planting of seeds or from the appearance of the first seedlings. The concentration of negative ions was measured in the air at a distance of 50 cm from plants. In both variants, similar growth-related changes in the concentration of negative ions were observed. The generation of negative ions began on day 6 after the planting of seeds and reached a concentration of 380 x 10(3) ion/cm3. During the next three days, this level remained unchanged. On day 10, the generation of negative aeroions increased abruptly; on days 10-14, it was twofold as high as on days 7-9. The level of generation of negative aeroions by plants stimulated from the moment of appearance of plantlets was 5-8% higher than by plants stimulated from the moment of planting. The intensity of generation of negative aeroions upon additional illumination and in full darkness remained unchanged. [本文引用: 1] |
| [49] | DOI:10.1023/B:RUPP.0000028690.74805.e2URLGeneration of negative air ions (NAI) by pot plants (aloe Aloe arborescens , haworthia Haworthia rasalata , echinopsis Echinopsis tubiflora , mammillaria Mammillaria prolifera , opuntia Opuntia brunnescens , spider plant Chlorophytum comosum , and jade plant Crassula portulacea ) was studied when high-voltage pulses were applied to soil. Plants that generated low amounts of NAI (such as jade plant and echinopsis) elevated NAI level 2–3 times over the mean background level. Plants that produced moderate amounts of NAI (haworthia, opuntia, and mammillaria) increased the NAI level hundredfold, whereas highly active plants (aloe and spider plant) increased the NAI level thousand times. Aloe plants can maintain constant NAI concentration in the ambient air (125 ± 15 × 10 3 ions/cm 3 ) for a long time (7–8 h). Negative air ions were predominantly generated by the leaf tips. The capacity of aloe to produce NAI considerably reduced with plant age and was lost, although reversibly, at temperatures below 8.5°C. The rate of NAI generation considerably depended on weather conditions. It was demonstrated that superoxide anion radical is one of NAI species generated by plants. [本文引用: 1] |
| [50] | DOI:10.1016/j.chemgeo.2012.09.041PMID:15387086URLThis study was conducted to examine whether negative air ions generated from pajamas would influence the rectal temperature and the immune system during night sleep. Nine females (aged 18-23 years) served as participants. They slept during the night in their homes, wearing the pajamas with generation of negative air ions (1260 ions/cm3) and with normal standard (520 ions/cm3). The sequence of wearing the pajamas was: first, standard pajamas; second, pajamas with negative air ions; and third, standard pajamas again, each being worn for three consecutive days. Rectal temperature in the pajamas with negative air ions tended to fall more significantly during the night-time (p = 0.068). Salivary IgA tended to be higher on waking when wearing pajamas with negative air ions (p = 0.094) and its effect continued even after standard pajamas were worn again during last three days. These results suggest that the rectal temperature could possibly be more reduced and the elevation of salivary IgA more marked if the pajamas with negative air ions are worn during nocturnal sleep. [本文引用: 1] |
| [51] | DOI:10.1016/j.jenvman.2009.03.003PMID:19356839URLThe purpose of this research was to determine changes in negative air ion (NAI) concentrations produced by plants grown under different light intensities. NAI concentrations were observed and analyzed in an outdoor green space with five plant species ( Aloe arborescens, Clivia miniata, Chlorophytum comosum, Opuntia brunnescens, Crassula portulacea) and A. arborescens was then grown under controlled-light conditions in an enclosed space for the subsequent light intensity experiment. (1) Two peaks in NAI concentration were observed over the course of 24 h in the outdoor green space: one occurred between 9:00 and 10:00 AM; the other was at approximately 8:00 PM. (2) Among the five plant species, A. arborescens produced the highest levels of NAI and responded most effectively to light with an increase in NAI generation. Accordingly, it was chosen as the model plant for studies on the changes in NAI concentration under different light intensities. (3) In enclosed-space experiments, isolated from sunlight and under controlled-light conditions, the concentrations of NAI varied with changes in light intensities. (4) Using regression analysis, a logistic model was developed showing that changes in NAI concentrations as a function of illumination intensity followed an exponential relationship. [本文引用: 3] |
| [52] | DOI:10.3390/ijerph8062304PMID:3138026URLNegative air ions (NAI) produce biochemical reactions that increase the levels of the mood chemical serotonin in the environment. Moreover, they benefit both the psychological well being and the human body physiological condition. The aim of this research was to estimate and measure the spatial distributions of negative and positive air ions in a residential garden in central Taiwan. Negative and positive air ions were measured at thirty monitoring locations in the study garden from July 2009 to June 2010. Moreover, Kriging was applied to estimate the spatial distribution of negative and positive air ions, as well as the air ion index in the study area. The measurement results showed that the numbers of NAI and PAI differed greatly during the four seasons, the highest and the lowest negative and positive air ion concentrations were found in the summer and winter, respectively. Moreover, temperature was positively affected negative air ions concentration. No matter what temperature is, the ranges of variogram in NAI/PAI were similar during four seasons. It indicated that spatial patterns of NAI/PAI were independent of the seasons and depended on garden elements and configuration, thus the NAP/PAI was a good estimate of the air quality regarding air ions. Kriging maps depicted that the highest negative and positive air ion concentration was next to the waterfall, whereas the lowest air ions areas were next to the exits of the garden. The results reveal that waterscapes are a source of negative and positive air ions, and that plants and green space are a minor source of negative air ions in the study garden. Moreover, temperature and humidity are positively and negatively affected negative air ions concentration, respectively. The proposed monitoring and mapping approach provides a way to effectively assess the patterns of negative and positive air ions in future landscape design projects. [本文引用: 3] |
室内空气质量洁净与否的宏观评价法
1
1996
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
北京典型园林植物区空气负离子分布特征研究
1
2010
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
城市绿地植物群落空气负离子浓度及影响要素研究
2
2011
... 负离子浓度随着环境的变化而呈现差异.例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3).而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(
... 与脉冲电场作用对植物释放负离子的影响相似, 光照在一定程度上也可提高植物释放负离子的能力, 光照度是影响植物群落释放负离子能力的主要因素之一(
佳木斯绿地空气负离子浓度及其与气象因子的关系
1
2009
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
植物群落对空气负离子浓度影响的研究
2
2008
... 负离子浓度随着环境的变化而呈现差异.例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3).而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
福州鼓山茶园不同生境空气负离子浓度及其影响因子
1
2014
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
森林与空气负离子
1
2000
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
6种植物群落夏季空气负离子动态及其与气象因子的关系
1
2010
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
低强度运动匹配负离子对老年高血压患者的影响
1
2010
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
北京西山典型城市森林内PM2.5动态变化规律
1
2014
... 负离子浓度随着环境的变化而呈现差异.例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3).而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(
空气负离子浓度分布特征及其与环境因子的关系
1
2014
... 负离子浓度随着环境的变化而呈现差异.例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3).而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(
城市森林调控空气颗粒物功能研究进展
1
2014
... 负离子浓度随着环境的变化而呈现差异.例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3).而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(
植物源负离子发生器室内应用的研究
1
2014
... 负离子浓度随着环境的变化而呈现差异.例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3).而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(
森林游憩区空气负离子水平的研究
1
2001
... 负离子浓度随着环境的变化而呈现差异.例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3).而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(
马尾松纯林林分因子对空气负离子浓度影响的研究
1
1998
... 负离子浓度随着环境的变化而呈现差异.例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3).而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(
a). 具备释放负离子功能室内植物的种质资源研究
2011
a). 植物释放负离子对室内空气质量影响分析
1
2014
... 负离子浓度随着环境的变化而呈现差异.例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3).而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(
脉冲电场作用对植物释放负离子的影响
2
2015
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
具备释放负离子功能室内植物的种质资源研究 II. 常态下室内植物负离子的释放
2011
水稻植株释放负离子研究
2
2014
... 负离子浓度随着环境的变化而呈现差异.例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3).而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
电磁场对生物体系的非热效应及其作用机理
3
2008
... 负离子因具有改善空气质量和有利于人体健康等作用, 常被誉为空气中的“维生素”和“生长素”, 备受人们的关注.目前, 对植物释放负离子的研究主要集中在植物群落和室内环境单株植物两个领域.自然状态下, 植物释放负离子的能力均很弱.吴仁烨等(2011b)对自然状态下能够释放负离子的种质资源进行了调查, 发现所研究的68种植物, 释放负离子浓度均值最高的仅为73 ion·cm-3.本研究也得出了相似的结果, 即常态下植物释放负离子浓度值远低于宜居室内的标准值(1 500 ion·cm-3) (
... ).在最佳脉冲电场作用下, 植物释放负离子的能力最强, 施加低于或高于此最佳脉冲电场, 其释放负离子的能力均有不同程度的降低, 原因可能在于低频电场生物学效应具有“阈值”与“功率窗”的特性(
... 同时, 本研究发现, 绒叶肖竹芋在各强度脉冲电场作用下, 释放负离子的浓度均值变化幅度很小.有研究表明, 植物本身具有特定的电位特征频率, 如果施加的脉冲电场适宜, 则会与植物电场共振进而提高释放负离子的能力(
应用负高压脉冲技术提高植物空气净化能力的探讨
2
2011
... 负离子因具有改善空气质量和有利于人体健康等作用, 常被誉为空气中的“维生素”和“生长素”, 备受人们的关注.目前, 对植物释放负离子的研究主要集中在植物群落和室内环境单株植物两个领域.自然状态下, 植物释放负离子的能力均很弱.吴仁烨等(2011b)对自然状态下能够释放负离子的种质资源进行了调查, 发现所研究的68种植物, 释放负离子浓度均值最高的仅为73 ion·cm-3.本研究也得出了相似的结果, 即常态下植物释放负离子浓度值远低于宜居室内的标准值(1 500 ion·cm-3) (
... 同时, 本研究发现, 绒叶肖竹芋在各强度脉冲电场作用下, 释放负离子的浓度均值变化幅度很小.有研究表明, 植物本身具有特定的电位特征频率, 如果施加的脉冲电场适宜, 则会与植物电场共振进而提高释放负离子的能力(
北京市5种典型城市绿化植物的生态保健功能分析
1
2013
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
广州城市绿地植物群落空气负离子特征研究
1
2015
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
脉冲电场介导的植物空气负离子发射的倍增效应及其意义
1
2009
... 同时, 本研究发现, 绒叶肖竹芋在各强度脉冲电场作用下, 释放负离子的浓度均值变化幅度很小.有研究表明, 植物本身具有特定的电位特征频率, 如果施加的脉冲电场适宜, 则会与植物电场共振进而提高释放负离子的能力(
广州绿地空气负离子水平及其影响因子
1
2007
... 负离子浓度随着环境的变化而呈现差异.例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3).而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(
青山湖不同植物群落空气负离子效应评价
1
2013
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
城市不同植被配置类型空气负离子效应评价
1
2005
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
上海环城林带保健功能评价及其机制
1
2013
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
常态下仙人掌科植物负离子释放量的比较及其与刺数量的关系
1
2015
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
崇明岛不同生态用地空气负离子分布规律研究
1
2016
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
城市空气负离子浓度时空分布及其影响因素综述
2
2013
... 负离子浓度随着环境的变化而呈现差异.例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3).而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
空气离子生物学效应研究的进展
1
1983
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
空气离子实验与临床研究新进展
3
2001
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
... 负离子浓度随着环境的变化而呈现差异.例如, 在窗户紧闭的室内, 负离子的浓度均值可低至500 ion·cm-3; 在污染的空气中, 负离子的浓度甚至可降为0, 远低于清新空气的标准(负离子浓度为1 500 ion·cm-3).而在室外绿地、林区、瀑布和公园等植被丰富区, 负离子浓度均值可达3 000 ion·cm-3以上(
... 负离子因具有改善空气质量和有利于人体健康等作用, 常被誉为空气中的“维生素”和“生长素”, 备受人们的关注.目前, 对植物释放负离子的研究主要集中在植物群落和室内环境单株植物两个领域.自然状态下, 植物释放负离子的能力均很弱.吴仁烨等(2011b)对自然状态下能够释放负离子的种质资源进行了调查, 发现所研究的68种植物, 释放负离子浓度均值最高的仅为73 ion·cm-3.本研究也得出了相似的结果, 即常态下植物释放负离子浓度值远低于宜居室内的标准值(1 500 ion·cm-3) (
3种仙人掌科植物负离子释放量与释放通道的相关性研究
1
2016
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
植物叶表皮制片方法改进
1
2008
... 为研究植物释放负离子与植物叶片气孔数量特征的关系, 实验设置1个对照组(CK)和2个处理组.对照组: 对自然状态下生长的植物进行采样; 实验组: 对植物施加最佳脉冲电场刺激, 持续600秒后进行采样, 同时检测负离子的浓度值.取样部位为叶片靠近中脉两侧约0.3-0.5 cm处, 将供试材料切成3-5 cm2正方形小块, 于FAA溶液中固定24小时, 60.0°C水浴40分钟后, 用30%-40%次氯酸钠浸泡24小时, 之后换用蒸馏水, 撕取叶片的上下表皮, 用0.5%固绿染色, 制成临时切片(
森林旅游资源评价中的空气负离子研究
1
1998
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
2
1962
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
... 两种形式存在(
1
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
2000
1
1997
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
1
1962
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
1
1972
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
1
1985
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
2
1976
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
... ;
1
2014
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
1
2002
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
1
2002
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
1
2004
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
1
2004
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
3
2009
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
... ).自然界中的负离子大部分来自宇宙射线和雷电等对大气分子的电离作用、水的喷筒电效应、树冠与枝叶的尖端放电, 及植物光合作用形成的光电效应促使空气电离.自然状态下, 空气通过空气电离与离子复合两个过程使空气中的离子保持动态平衡(
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(
3
2011
... 负离子的生物学效应及其对环境的生态效应已日益受到人们的关注(
... ;
... 目前, 对负离子的研究主要侧重于植物群落释放负离子及其动态分布特征(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}