Abstract: We chose Arabidopsis thaliana and Setaria italic as material to study the character of HL1 that can specifically combine with zinc, and the distribution of extracellular Zn2+ by using fluorescence spectrophotometry, isothermal titration calorimetry (ITC200) and inverted fluorescence microscopy. Fluorescence intensity for HL1 was greatly enhanced with the addition of Zn2+ but not other ions. The dissociation constant (KD=7.02×10-4 mol·L-1) exhibited product stability with the combined reaction of HL1 and Zn2+. In A. thaliana, the distribution of extracellular free Zn2+ was mainly located in leaf intercellular space and surface of the trichome where fluorescence intensity was corresponding to the concentration of Zn2+. The distribution of Zn2+ were located in intercellular space and fibrovascular tissue in the leaf of S. italic. The root of elongation zone was existing the blue fluorescence corresponded to the presence and concentration of Zn2+. The root elongation zone relates to Zn2+ transportation, and the leaf intercellular space and trichome surface are related to Zn2+ storage. In conclusion, Investigation of extracellular free Zn2+ by using HL1 is efficient.
Key words:zinc ; blue fluorescence ; leaf trichome ; vascular tissue ; intercellular space
锌(Zn)是所有生物体生长繁殖所需要的基本元素。目前认为, 锌在生物体内具有催化和辅因子的作用(Maret, 2009)。当植物缺锌时, 普遍会出现生物产量降低、生育力下降、叶片萎黄和过早衰老的症状。长期锌摄入量过低会对成人的健康以及婴儿的行动能力造成较大影响, 引起生长、认知和免疫功能的损伤(Hambidge et al., 2010)。普遍的观点认为, 锌至少存在6种转运体家族(Hantke, 2001), 其中ABC转运体家族、RND转运体家族和CorA蛋白家族在细菌中具有转运锌的功能, 然而并未在真核生物中发现类似的功能(Liuzzi and Cousins, 2004)。在拟南芥(Ara- bidopsis thaliana)中, 锌转运蛋白(ZIP)家族负责对Zn2+装载, 阳离子扩散促进因子(CDF)负责Zn2+分配, P1B型ATP酶泵蛋白家族负责Zn2+从根到茎的运输(Hussain et al., 2004)。在酵母中, ZIP家族中的ZRT1和ZRT2蛋白负责将外界Zn2+转运到细胞内(Zhao and Eide, 1996)。在人体内, ZIP4蛋白负责转运肠道内的Zn2+, ZIP10蛋白负责肾脏中Zn2+的重新吸收(Kelleher and Lonnerdal, 2005)。植物需要吸收并转运体外的锌来满足生存的需要。首先植物的根部向根周围分泌小分子有机螯合物并使周围的环境酸化, 来增加锌的溶解, 随后Zn2+以游离态穿过细胞膜进入根表皮细胞。进入细胞内的Zn2+以小分子配体螯合的状态存在, 它可被转运至液泡中贮存, 也可通过胞间连丝或共质体途径进入维管束的木质部。进入木质部的Zn2+仍以小分子配体螯合的状态存在, 在集流的驱动下进入根上部分(Sinclair and Krämer, 2012)。植物体内小分子配体具有重要作用, 它维持Zn2+的稳定, 防止Zn2+与其它物质结合或发生无意义的反应, 这对Zn2+的贮藏和运输具有重大意义(Klatte et al., 2009)。
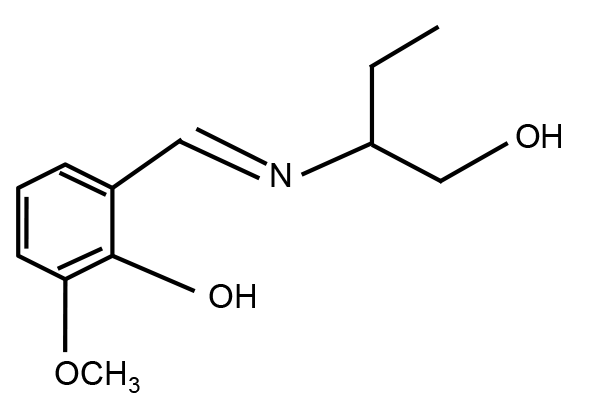
图1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-608/img_1.png<b>图1</b> HL<sup>1</sup>的结构<br/><b>Figure 1</b> The structure of HL<sup>1</sup> Figure 1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-608/img_1.png<b>图1</b> HL<sup>1</sup>的结构<br/><b>Figure 1</b> The structure of HL<sup>1</sup>
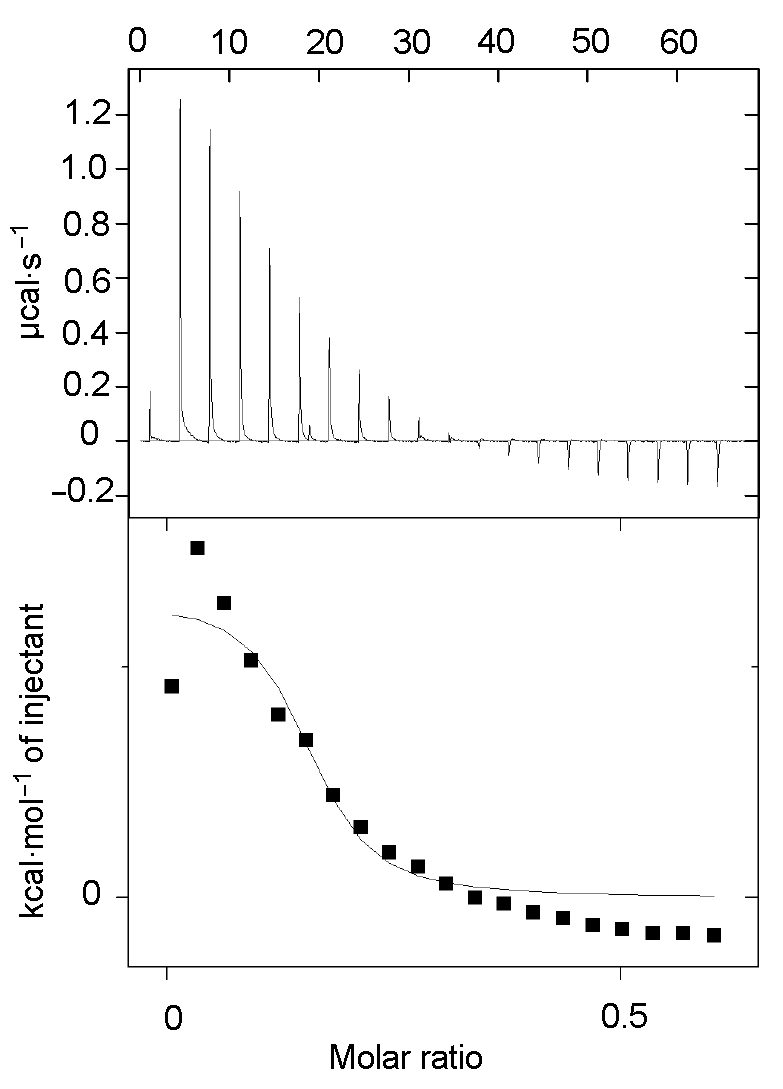
图3 利用ITC200检测HL1与Zn2+的反应 Figure 3 ITC200 detection plot of combination of HL1 and Zn2+
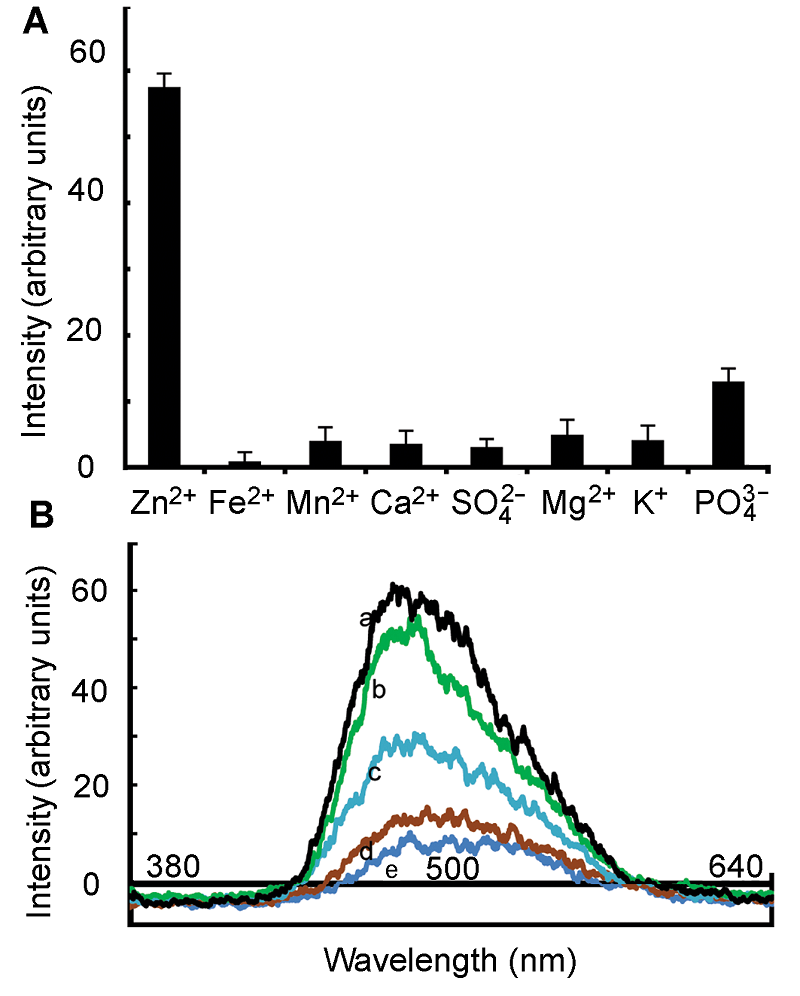
2.3 拟南芥叶中的Zn2+使用含有不同浓度Zn2+的溶液处理叶片, 结果(图4A)表明, 叶片上的叶表皮毛和细胞间隙具有明显的蓝色荧光。在具有蓝色荧光的叶表皮毛上均匀取12个点, 根据这些点的荧光强度值, 计算出荧光强度的平均值和标准差并作图。通过观察叶片上的细胞, 发现蓝色荧光主要位于细胞间隙, 我们并未在细胞内观察到明显的蓝色荧光(图5)。这可能是由于植物细胞内存在某些物质或条件影响HL1与Zn2+的结合, 也有可能因为自身存在N和O原子, 使其具有一定的极性, 从而导致进入的HL1很少, 无法发出明显的蓝色荧光。 图4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-608/img_4.png<b>图4</b> 拟南芥叶细胞外Zn<sup>2+</sup>分布荧光图像<br/>(A) 用不同浓度ZnSO<sub>4</sub>溶液及HL<sup>1</sup>溶液处理成熟的拟南芥叶片, a1和a2为对照, b1和b2处理ZnSO<sub>4</sub>溶液浓度为0.3 μmol∙L<sup>-1</sup>, c1和c2处理ZnSO<sub>4</sub>溶液浓度为3 μmol∙L<sup>-1</sup>, d1和d2处理ZnSO<sub>4</sub>溶液浓度为30 μmol∙L<sup>-1</sup>, e1和e2处理ZnSO<sub>4</sub>溶液浓度为300 μmol∙L<sup>-1</sup>; (B) 不同浓度ZnSO<sub>4</sub>溶液及HL<sup>1</sup>溶液处理下的拟南芥叶表皮毛的荧光强度, 其中a为对照, b、c、d和e分别为叶表皮毛在Zn<sup>2+</sup>浓度为0.3、3、30和300 μmol∙L<sup>-1</sup>时的荧光强度。蓝色荧光代表Zn<sup>2+</sup>, 红色荧光代表叶绿素。Bar=100 μm<br/><b>Figure 4</b> Fluorescence imaging showing the distribution of extracellular Zn<sup>2+</sup> in Arabidopsis leaves<br/>(A) Mature Arabidopsis leaves pre-treated with different concentrations of ZnSO<sub>4</sub> and HL<sup>1</sup> (1 mmol∙L<sup>-1</sup>) for 4 h, a1 and a2 for control, b1 and b2 for 0.3 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, c1 and c2 for 3 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, d1 and d2 for 30 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, e1 and e2 for 300 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>; (B) The fluorescence intensity of trichome in Arabidopsis leaf with different concentration of ZnSO<sub>4</sub> and HL<sup>1 </sup>(1 mmol∙L<sup>-1</sup>) for 4 h, a for control, b for 0.3 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, c for 3 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, d for 30 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, and e for 300 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>. Blue fluorescence represents Zn<sup>2+</sup> and red fluorescence for chlorophyll. Bar=100 μm Figure 4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-608/img_4.png<b>图4</b> 拟南芥叶细胞外Zn<sup>2+</sup>分布荧光图像<br/>(A) 用不同浓度ZnSO<sub>4</sub>溶液及HL<sup>1</sup>溶液处理成熟的拟南芥叶片, a1和a2为对照, b1和b2处理ZnSO<sub>4</sub>溶液浓度为0.3 μmol∙L<sup>-1</sup>, c1和c2处理ZnSO<sub>4</sub>溶液浓度为3 μmol∙L<sup>-1</sup>, d1和d2处理ZnSO<sub>4</sub>溶液浓度为30 μmol∙L<sup>-1</sup>, e1和e2处理ZnSO<sub>4</sub>溶液浓度为300 μmol∙L<sup>-1</sup>; (B) 不同浓度ZnSO<sub>4</sub>溶液及HL<sup>1</sup>溶液处理下的拟南芥叶表皮毛的荧光强度, 其中a为对照, b、c、d和e分别为叶表皮毛在Zn<sup>2+</sup>浓度为0.3、3、30和300 μmol∙L<sup>-1</sup>时的荧光强度。蓝色荧光代表Zn<sup>2+</sup>, 红色荧光代表叶绿素。Bar=100 μm<br/><b>Figure 4</b> Fluorescence imaging showing the distribution of extracellular Zn<sup>2+</sup> in Arabidopsis leaves<br/>(A) Mature Arabidopsis leaves pre-treated with different concentrations of ZnSO<sub>4</sub> and HL<sup>1</sup> (1 mmol∙L<sup>-1</sup>) for 4 h, a1 and a2 for control, b1 and b2 for 0.3 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, c1 and c2 for 3 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, d1 and d2 for 30 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, e1 and e2 for 300 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>; (B) The fluorescence intensity of trichome in Arabidopsis leaf with different concentration of ZnSO<sub>4</sub> and HL<sup>1 </sup>(1 mmol∙L<sup>-1</sup>) for 4 h, a for control, b for 0.3 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, c for 3 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, d for 30 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, and e for 300 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>. Blue fluorescence represents Zn<sup>2+</sup> and red fluorescence for chlorophyll. Bar=100 μm
图4 拟南芥叶细胞外Zn2+分布荧光图像 (A) 用不同浓度ZnSO4溶液及HL1溶液处理成熟的拟南芥叶片, a1和a2为对照, b1和b2处理ZnSO4溶液浓度为0.3 μmol∙L-1, c1和c2处理ZnSO4溶液浓度为3 μmol∙L-1, d1和d2处理ZnSO4溶液浓度为30 μmol∙L-1, e1和e2处理ZnSO4溶液浓度为300 μmol∙L-1; (B) 不同浓度ZnSO4溶液及HL1溶液处理下的拟南芥叶表皮毛的荧光强度, 其中a为对照, b、c、d和e分别为叶表皮毛在Zn2+浓度为0.3、3、30和300 μmol∙L-1时的荧光强度。蓝色荧光代表Zn2+, 红色荧光代表叶绿素。Bar=100 μm Figure 4 Fluorescence imaging showing the distribution of extracellular Zn2+ in Arabidopsis leaves (A) Mature Arabidopsis leaves pre-treated with different concentrations of ZnSO4 and HL1 (1 mmol∙L-1) for 4 h, a1 and a2 for control, b1 and b2 for 0.3 μmol∙L-1 ZnSO4, c1 and c2 for 3 μmol∙L-1 ZnSO4, d1 and d2 for 30 μmol∙L-1 ZnSO4, e1 and e2 for 300 μmol∙L-1 ZnSO4; (B) The fluorescence intensity of trichome in Arabidopsis leaf with different concentration of ZnSO4 and HL1 (1 mmol∙L-1) for 4 h, a for control, b for 0.3 μmol∙L-1 ZnSO4, c for 3 μmol∙L-1 ZnSO4, d for 30 μmol∙L-1 ZnSO4, and e for 300 μmol∙L-1 ZnSO4. Blue fluorescence represents Zn2+ and red fluorescence for chlorophyll. Bar=100 μm
图5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-608/img_5.png<b>图5</b> 植物细胞在1 mmol∙L<sup>-1 </sup>HL<sup>1</sup>下的荧光图像<br/>蓝色荧光代表Zn<sup>2+</sup>, 红色荧光代表叶绿素。Bar=50 μm<br/><b>Figure 5</b> Fluorescence image of plant cell in presence of 1 mmol∙L<sup>-1</sup> HL<sup>1</sup><br/>Blue represents Zn<sup>2+</sup> and red for chlorophyll. Bar=50 μm Figure 5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-608/img_5.png<b>图5</b> 植物细胞在1 mmol∙L<sup>-1 </sup>HL<sup>1</sup>下的荧光图像<br/>蓝色荧光代表Zn<sup>2+</sup>, 红色荧光代表叶绿素。Bar=50 μm<br/><b>Figure 5</b> Fluorescence image of plant cell in presence of 1 mmol∙L<sup>-1</sup> HL<sup>1</sup><br/>Blue represents Zn<sup>2+</sup> and red for chlorophyll. Bar=50 μm
图5 植物细胞在1 mmol∙L-1 HL1下的荧光图像 蓝色荧光代表Zn2+, 红色荧光代表叶绿素。Bar=50 μm Figure 5 Fluorescence image of plant cell in presence of 1 mmol∙L-1 HL1 Blue represents Zn2+ and red for chlorophyll. Bar=50 μm
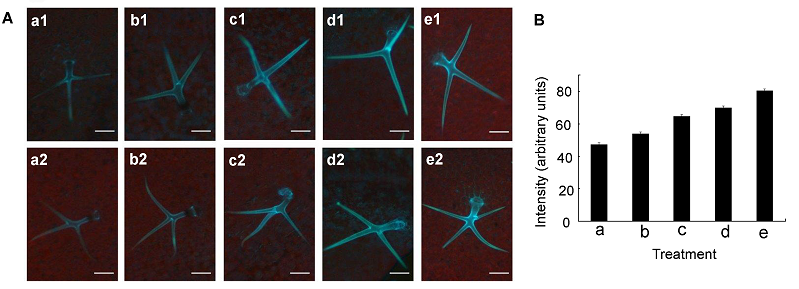
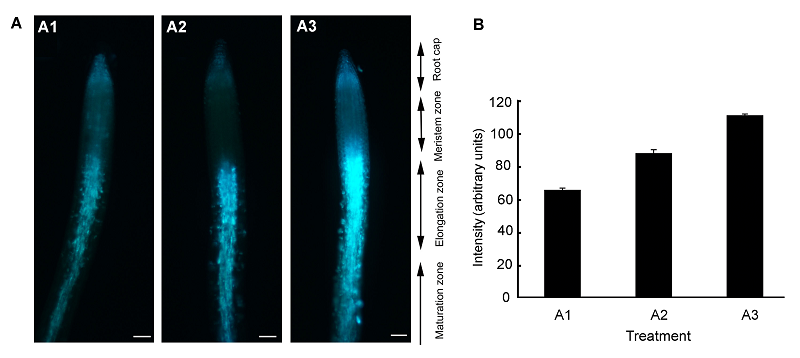
2.4 拟南芥根中的Zn2+用不同浓度Zn2+溶液与HL1溶液共处理根部, 结果(图6)表明, 根的伸长区具有明显的蓝色荧光。在具有蓝色荧光的伸长区取6个点, 根据这些点的荧光强度值, 计算出荧光强度的平均值和标准差并作图。结果表明, 荧光强度从伸长区到成熟区开始逐渐降低, 这说明细胞外游离态Zn2+在根的伸长区浓度最高, 它可能与根上部Zn2+的浓度形成浓度差, 从而促进Zn2+由根尖向根上部的运输。 图6https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-608/img_6.png<b>图6</b> 拟南芥根细胞外Zn<sup>2+</sup>分布荧光图像<br/>(A) 分别用不同浓度的ZnSO<sub>4</sub>溶液与HL<sup>1</sup>溶液共处理2 cm左右的拟南芥根, 处理时间为4小时, 其中A1为对照, A2和A3中Zn<sup>2+</sup>浓度分别为30和300 μmol∙L<sup>-1 </sup>(Bar=100 μm); (B) A1、A2和A3分别对应处理液Zn<sup>2+</sup>浓度0、30和300 μmol∙L<sup>-1</sup>下的荧光强度。蓝色荧光代表Zn<sup>2+</sup>。<br/><b>Figure 6</b> Fluorescence images showing the distribution of extracellular Zn<sup>2+</sup> in Arabidopsis roots<br/>(A) Two-centimeter-long Arabidopsis root pre-treated with different concentrations of ZnSO<sub>4</sub> and HL<sup>1 </sup>(1 mmol∙L<sup>-1</sup>) for 4 h, A1, A2, and A3 for control, 30 μmol∙L<sup>-1</sup> ZnSO<sub>4 </sub>and 300 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, respectively (Bar=100 μm); (B) A1, A2, and A3 corresponding to the fluorescence intensity of Arabidopsis root treated with different concentrations of ZnSO<sub>4</sub> for 0, 30, and 300 μmol∙L<sup>-1</sup>. Blue fluorescence represents Zn<sup>2+</sup>. Figure 6https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-608/img_6.png<b>图6</b> 拟南芥根细胞外Zn<sup>2+</sup>分布荧光图像<br/>(A) 分别用不同浓度的ZnSO<sub>4</sub>溶液与HL<sup>1</sup>溶液共处理2 cm左右的拟南芥根, 处理时间为4小时, 其中A1为对照, A2和A3中Zn<sup>2+</sup>浓度分别为30和300 μmol∙L<sup>-1 </sup>(Bar=100 μm); (B) A1、A2和A3分别对应处理液Zn<sup>2+</sup>浓度0、30和300 μmol∙L<sup>-1</sup>下的荧光强度。蓝色荧光代表Zn<sup>2+</sup>。<br/><b>Figure 6</b> Fluorescence images showing the distribution of extracellular Zn<sup>2+</sup> in Arabidopsis roots<br/>(A) Two-centimeter-long Arabidopsis root pre-treated with different concentrations of ZnSO<sub>4</sub> and HL<sup>1 </sup>(1 mmol∙L<sup>-1</sup>) for 4 h, A1, A2, and A3 for control, 30 μmol∙L<sup>-1</sup> ZnSO<sub>4 </sub>and 300 μmol∙L<sup>-1</sup> ZnSO<sub>4</sub>, respectively (Bar=100 μm); (B) A1, A2, and A3 corresponding to the fluorescence intensity of Arabidopsis root treated with different concentrations of ZnSO<sub>4</sub> for 0, 30, and 300 μmol∙L<sup>-1</sup>. Blue fluorescence represents Zn<sup>2+</sup>.
图6 拟南芥根细胞外Zn2+分布荧光图像 (A) 分别用不同浓度的ZnSO4溶液与HL1溶液共处理2 cm左右的拟南芥根, 处理时间为4小时, 其中A1为对照, A2和A3中Zn2+浓度分别为30和300 μmol∙L-1 (Bar=100 μm); (B) A1、A2和A3分别对应处理液Zn2+浓度0、30和300 μmol∙L-1下的荧光强度。蓝色荧光代表Zn2+。 Figure 6 Fluorescence images showing the distribution of extracellular Zn2+ in Arabidopsis roots (A) Two-centimeter-long Arabidopsis root pre-treated with different concentrations of ZnSO4 and HL1 (1 mmol∙L-1) for 4 h, A1, A2, and A3 for control, 30 μmol∙L-1 ZnSO4 and 300 μmol∙L-1 ZnSO4, respectively (Bar=100 μm); (B) A1, A2, and A3 corresponding to the fluorescence intensity of Arabidopsis root treated with different concentrations of ZnSO4 for 0, 30, and 300 μmol∙L-1. Blue fluorescence represents Zn2+.
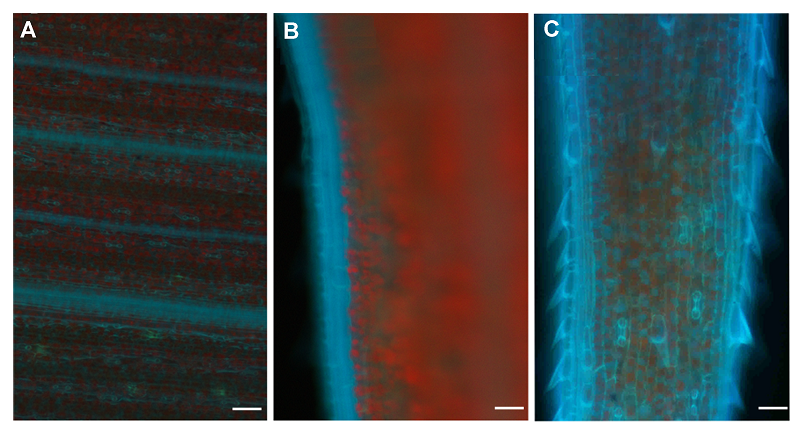
2.5 谷子叶片中的Zn2+用1 mmol∙L-1 HL1溶液处理谷子根部, 利用植物的蒸腾作用使HL1通过根进入叶, 在倒置荧光显微镜下观察, 结果(图7)显示, 蓝色荧光主要分布于叶脉、叶边缘处和细胞间隙。在叶片中央, 蓝色荧光在叶脉处较强, 细胞间隙较弱, 这可能暗示着植物以叶脉作为运输Zn2+的通道, 并以叶脉为中心向两侧的细胞间隙运输Zn2+。在叶边缘处, 蓝色荧光十分明显, 而与其相邻的细胞间隙的蓝色荧光相对较弱, 这表明叶边缘也可能具有运输Zn2+的功能; 相比叶的其它部位, 叶尖处的细胞间隙以及叶边缘蓝色荧光明显, 这可能与位于叶尖处的细胞有关。叶尖处的细胞具有较强的分裂能力, 生理活动及细胞代谢比较旺盛, 因此需要较多的Zn2+来满足代谢的需求, 所以叶尖处具有较强的荧光强度。 图7https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-608/img_7.png<b>图7</b> 谷子叶片细胞外Zn<sup>2+</sup>分布荧光图像<br/>(A) 叶中央; (B) 叶边缘; (C) 叶尖。蓝色荧光代表Zn<sup>2+</sup>, 红色荧光代表叶绿素。Bar=100 μm<br/><b>Figure 7</b> Fluorescence images showing the distribution of extracellular Zn<sup>2+</sup> in <i>Setaria italic </i>leaves<br/>(A) The center of leaf; (B) The margin of leaf; (C) The apex of leaf. Blue fluorescence represents Zn<sup>2+</sup> and red fluorescence for chlorophyll. Bar=100 μm Figure 7https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-608/img_7.png<b>图7</b> 谷子叶片细胞外Zn<sup>2+</sup>分布荧光图像<br/>(A) 叶中央; (B) 叶边缘; (C) 叶尖。蓝色荧光代表Zn<sup>2+</sup>, 红色荧光代表叶绿素。Bar=100 μm<br/><b>Figure 7</b> Fluorescence images showing the distribution of extracellular Zn<sup>2+</sup> in <i>Setaria italic </i>leaves<br/>(A) The center of leaf; (B) The margin of leaf; (C) The apex of leaf. Blue fluorescence represents Zn<sup>2+</sup> and red fluorescence for chlorophyll. Bar=100 μm
图7 谷子叶片细胞外Zn2+分布荧光图像 (A) 叶中央; (B) 叶边缘; (C) 叶尖。蓝色荧光代表Zn2+, 红色荧光代表叶绿素。Bar=100 μm Figure 7 Fluorescence images showing the distribution of extracellular Zn2+ in Setaria italic leaves (A) The center of leaf; (B) The margin of leaf; (C) The apex of leaf. Blue fluorescence represents Zn2+ and red fluorescence for chlorophyll. Bar=100 μm
BermejoC, EwaldJC, LanquarV, JonesAM, FrommerWB (2011). In vivo biochemistry: quantifying ion and metabolite levels in individual cells orcultures of yeast.Biochem J 438, 1-10. [本文引用: 1]
[2]
CakmakI (2000). Tansley review No. 111: possible roles of zinc in protecting plant cells from damage by reactive oxygen species.New Phytol 146, 185-205. [本文引用: 1]
[3]
HacisalihogluG, HartJJ, WangYH, CakmakI, KochianLV (2003). Zinc efficiency is correlated with enhanced expression and activity of zinc-requiring enzymes in whe- at.Plant Physiol 131, 595-602. [本文引用: 1]
HussainD, HaydonMJ, WangYW, WongE, ShersonSM, YoungJ, CamakarisJ, HarperJF, CobbettCS (2004). P-type ATPase heavy metal transporters with roles in es- sential zinc homeostasis in Arabidopsis.Plant Cell 16, 1327-1339. [本文引用: 1]
[7]
JiYF, WangR, DingS, DuCF, LiuZL (2012). Synthesis, crystal structures and fluorescence studies of three new Zn(II) complexes with multidentate Schiff base ligands.In- organic Chem Commun 16, 47-50. [本文引用: 1]
[8]
KelleherSL, LönnerdalB (2005). Zip3 plays a major role in zinc uptake into mammary epithelial cells and is regulated by prolactin.Cell Physiol 288, C1042-C1047. [本文引用: 1]
[9]
KlatteM, SchulerM, WirtzM, Fink-StraubeC, HellR, BauerP (2009). The analysis of Arabidopsis nicotiana- mine synthase mutants reveals functions for nicotiana- mine in seed iron loading and iron deficiency responses.Plant Physiol 150, 257-271. [本文引用: 1]
[10]
KrämerU (2010). Metal hyperaccumulation in plants.Annu Rev Plant Biol 61, 517-534. [本文引用: 1]
[11]
LanquarV, LelièvreF, BolteS, HamèsC, AlconC, NeumannD, VansuytG, CurieC, SchroderA, KrämerU, Barbier-BrygooH, ThomineS (2005). Mobilization of vacuolar iron by AtNRAMP3 and AtNRAMP4 is essential for seed germination on low iron.EMBO J 24, 4041-4051. [本文引用: 1]
MaretW (2009). Molecular aspects of human cellular zinc homeostasis: redox control of zinc potentials and zinc signals.Biometals 22, 149-157. [本文引用: 1]
[14]
Seigneurin-BernyD, GravotA, AuroyP, MazardC, KrautA, FinazziG, GrunwaldD, RappaportF, VavasseurA, JoyardJ, RichaudP, RollandN (2006). HMA1, a new Cu-ATPase of the chloroplast envelope, is essential for growth under adverse light conditions.J Biol Chem 281, 2882-2892. [本文引用: 1]
[15]
SinclairSA, KrämerU (2012). The zinc homeostasis network of land plants.Biochim Biophys Acta 1823, 1553-1567. [本文引用: 1]
[16]
SzymanskiDB, LloydAM, MarksMD (2000). Progress in the molecular genetic analysis of trichome initiation and morphogenesis in Arabidopsis.Trends Plant Sci 5, 214-219. [本文引用: 1]
[17]
ZhaoH, EideD (1996). The yeast ZRT1 gene encodes the zinc transporter protein of a high-affinity uptake system induced by zinc limitation.Proc Natl Acad Sci USA 93, 2454-2458. [本文引用: 1]
1 2011
... 目前追踪和检测Zn2+的方法有通过能量色散的X射线分析法(Lanquar et al., 2005)、利用电感耦合等电子体的原子发射光谱法(Seigneurin-Berny et al., 2006) (ICP-AES)、从遗传学角度上编码能量共振转移体法(FRET) (Bermejo et al., 2011)以及化学探针法.此前报道过一种与Zn2+特异性结合的化学探针HL1 (2-((1-羟基-2-亚氨基)甲基)6-甲氧基苯酚) (Ji et al., 2012), 该探针分子式为C12H16NO3, 分子量为222.25 kDa, 结构如图1所示.该化合物通过自身的N、O原子与Zn2+形成配位键, 生成稳定的复合物Zn(HL1)2. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}