张金玲 ,
程达 ,
李玉灵 * , ,
陈海鹏 河北农业大学林学院, 保定 071000
Zhang Jinling ,
Cheng Da ,
Li Yuling * ,
Chen Haipeng Forestry College of Agricultural University of Hebei, Baoding 071000, China 引用本文 张金玲 ,
程达 ,
李玉灵 ,
陈海鹏 . 光和水分胁迫对臭柏实生幼苗光化学效率及色素组成的影响. 植物学报, 2017, 52(3): 278-289
贡献者
* 通讯作者。E-mail: liyuling0425@126.com 基金资助
河北省自然科学基金(No.C2011204096)和国家林业公益性行业科研专项(No.201404214) ;
接受日期:2016-10-7接受日期:2017-01-3网络出版日期:2017-05-1
-->Copyright
20172010 《植物学报》编辑部
Contributors
Corresponding * Author for correspondence. E-mail: liyuling0425@126.com History
Received :
Accepted :
Online :
摘要: 为探明毛乌素沙地3年生臭柏(Sabina vulgaris )实生苗在不同光照和水分条件下的光抑制响应机制, 研究了各处理臭柏实生苗的最大光化学效率(F v /F m )及叶绿素(Chla+Chlb)和叶黄素(A+V+Z)含量, 分析了其叶绿素循环和叶黄素循环的变化规律。结果表明, 77%透光区通过减少Chlb含量, 升高Chla/Chlb, 避免光能过剩; 同时, 增加A+V+Z及热散逸色素(A+Z)含量、提高(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)值, 耗散过剩光能, 避免光破坏。25%透光区的叶绿素和叶黄素循环机制随着水分条件的变化迅速发生改变。10%透光区通过增加Chlb含量, 降低Chla/Chlb, 捕捉更多的光能, 几乎不存在光抑制。毛乌素臭柏实生幼苗能够适应不同的光照和水分条件, 在恶劣的沙漠中完成天然更新, 形成独特的群落景观, 与叶绿素循环和叶黄素循环有着密切的关系。 关键词 :
臭柏 ;
光抑制 ;
最大光化学效率 ;
叶绿素循环 ;
叶黄素循环 Abstract :
We aimed to ascertain the photoinhibition response mechanism of 3-year-old Sabina vulgaris seedlings of Mu Us Sandland in different light and water conditions. We examined the photochemical efficiency of photosystem II at predawn (F v /F m ) and the content of chlorophyll (Chla+Chlb) and lutein (A+V+Z) of every treatment and analyzed the change law of chlorophyll cycle and xanthophyll cycle. Sabina vulgaris seedlings avoided excess light energy and photodamage and dissipated excess light energy by decreasing the content of chlorophyllb (Chlb) and increasing the ratio of Chla to Chlb (Chla/Chlb ratio), content of A+V+Z and A+Z, (A+V+Z)/(Chla+Chlb) ratio and (A+V)/(A+V+Z) ratio in 77% light transmittance area. The mechanism of chlorophyll cycle and xanthophyll cycle of S. vulgaris seedlings was changed quickly with the change in water condition in 25% light transmittance area. S. vulgaris seedlings captured more light energy by increasing the content of Chlb and decreasing the Chla/Chlb ratio in 10% light transmittance area, with little photoinhibition in this area. S. vulgaris seedlings in Mu Us Sandland can adapt to different light and water conditions and can complete their natural regeneration as a unique community landscape in the harsh desert, which is closely related to chlorophyll cycle and xanthophyll cycle. Key words :
Sabina vulgaris ;
photoinhibition ;
maximum photochemical efficiency ;
chlorophyll cycle ;
xanthophyll cycle 常绿植物叶片的光合作用和蒸腾作用等生理代谢均受到环境因子变化的影响, 如季节变化引起的强光、干旱和低温胁迫(
张有福等, 2010 )。强光胁迫下, 光合天线色素对太阳光能的过量吸收会造成光抑制, 即植物光合作用受到抑制, 可引起光系统II (PSII)反应中心的不可逆破坏, 即光破坏, 进而影响植物的光合生理代谢, 导致植物生长缓慢(
许大全等, 1992 ;
许大全, 2003 )。虽然强光是植物产生光抑制的直接原因, 但干旱、盐渍和低温等叠加胁迫会加剧植物的光抑制, 所以自然界植物的光抑制通常是多种胁迫共同作用的结果。为了物种的延续, 植物在长期的适应和进化过程中, 形成了减轻和抵御光抑制伤害的多种机制。例如, 叶片和叶绿体的避光运动减少了光能的捕获, 增强了光合作用的光能利用效率; 捕光天线色素的转换增加了非光化学耗散及促进了依赖叶黄素的循环等(
许大全等, 1992 ;
高杰等, 2015 ;
代欣等, 2016 ;
付忠等, 2016 )。其中, 叶绿素和叶黄素循环是植物应对光抑制的主要途径之一。
绿色植物的光合色素主要由叶绿素、类胡萝卜素和藻胆素三大类色素组成。叶绿素主要由叶绿素a (Chla)和叶绿素b (Chlb)组成。少部分的Chla具有转化光能的作用, 大部分Chla和全部Chlb的作用是吸收与传递光能, 是光系统天线复合体的重要组成成分。由叶绿素酸酯a合成Chlb和由Chlb到Chla的还原过程可以看作一个循环, 即叶绿素循环(
周锋利等, 2005 ;
郭春爱等, 2006 ;
郭春爱, 2007 ;
李玉灵等, 2009 )。当光能过剩时, 植物可适时减少Chlb的含量, 进而减少光能的吸收量; 当光照不足时, 植物可增加Chlb的含量, 从而捕捉更多的光能, 满足植物生长的需要。叶绿素两大组分间的比值(Chla/Chlb)与总量(Chla+Chlb)在一定程度上表示了整个光系统对光量子的捕获和利用状况, 也反映了植物处于特定环境下承受潜在光胁迫的程度。因此, 叶绿素循环不仅是植物对不同光环境做出的即时响应, 也是植物光抑制的防御机制之一。叶黄素类则是胡萝卜素的氧化衍生物, 是类胡萝卜素中含有氧的一组多萜类的总称, 它包括双环氧紫黄质(violaxanthin, V)、环氧玉米黄质(antheraxanthin, A)和玉米黄质(zeaxanthin, Z)等。其中, V、A和Z三者间可以相互转化, 称为叶黄素循环,该循环通过热耗散防御光破坏。当出现过剩光能时, V在紫黄质脱环氧化酶(VDE)的作用下, 经A转化成Z, Z在植物叶黄素循环耗散光能的过程中起关键作用, 可以直接猝灭激发态叶绿素(或改变类囊体膜的流动性)及促进PSII的捕光复合物(LHCII)聚集来增加非辐射能量的耗散, 所以A和Z也被称为热散逸色素(
王强等, 2003 )。当光能不再过剩时, 转化方向相反, Z在玉米黄质环氧化酶(ZE)的催化作用下经A形成V, Z减少而V增加。(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)比值越高, 意味着叶黄素含量及其中的热散逸色素含量越多或叶绿素含量越少。所以叶黄素循环也是植物对不同光环境做出的即时响应, 与叶绿素循环一样, 也是植物光抑制的防御机制之一。植物的叶绿素荧光主要由PSII反应中心产生, 与光合作用紧密相关, 能够反映光合生理的信息, 特别是胁迫环境下完整植株光合功能的变化(
师生波等, 2014 )。其中, 荧光参数
F v /
F m 指完整叶片中PSII的原初光能转化效率, 是研究光合生理的重要参数, 它能够反映出PSII利用光能的能力(
查倩等, 2016 ;
孙鲁龙等, 2016 )。在非逆境条件下, 不同物种或生态型的叶片,
F v /
F m 均在0.75- 0.85间, 并且不受生长条件的影响, 但在光抑制下,
F v /
F m 值明显降低(
许大全等, 1992 )。故
F v /
F m 是表征光抑制程度的一个重要指标, 该比值越低说明发生光抑制的程度越高(司继播等, 2007)。综上所述, 叶绿素循环、叶黄素循环和叶绿素荧光参数变化都是植物对光抑制的响应机制, 但不同植物在不同胁迫条件下的响应策略不同。
臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 )。天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区。在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木。它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 )。毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大。故臭柏极易受到强光照、干旱和低温的单一或多重胁迫。臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 )。毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱。 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 )。笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化。其必然要对光和水热胁迫做出响应, 启动光抑制防御机制。有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 )。然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确。尤其对光合器官中各类色素的协同调控机制还知之甚少。为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考。
1 材料与方法1.1 材料和实验设计实验材料为3年生臭柏(
Sabina vulgaris Ant.)实生苗。2014年2月将从内蒙古毛乌素沙地天然臭柏群落内采集的100粒臭柏种子播种在河北农业大学苗圃中。2015年4月将播种实生苗移植到花盆内, 土壤基质为苗圃土。5月选取生长健壮且长势一致的60株苗木, 用透光率不同的黑色尼龙遮阳网进行遮光处理。按照毛乌素沙地乌柳(
Salix cheilophila Schneid.)树下天然臭柏实生苗的透光率, 设置3个透光处理区, 透光率分别为77% (二针遮阳网)、25% (三针遮阳网)和10% (四针遮阳网); 再将每个透光区设置为2个水分条件处理区, 即干旱区(pF4-4.5)和湿润区(pF1.8), 每供试小区10株幼苗。使用土壤pF仪(DIK 3404, 日本), 采用离心法测定土壤pF值。2016年开始荧光参数和色素含量的测定。
1.2 实验方法1.2.1
F v /
F m 的测定
使用便携式荧光仪(Hansatech, 英国), 于2016年2-9月的每月上、中和下旬3天晴朗天气的黎明前, 每小区随机选取3株测定
F 0 和
F m , 由公式
F v /
F m = (
F m -
F 0 )/
F m 计算得最大光化学效率。
1.2.2 Chla和Chlb含量的测定
叶绿素a和叶绿素b含量采用分光光度计法测定。2016年, 选择低温的2月、开始生长的4月和生长旺期的7月三个时期的每月上、中和下旬3天晴朗天气, 每小区随机选取3株, 剪取当年生叶片用锡纸包好后放入装有液氮的冰盒内带回实验室。称取0.1 g装入10 mL离心管中, 加入2 mL无水乙醇, 用数显高数分散均质机(FJ200-SH, 上海)粉碎, 再加入8 mL无水乙醇, 置于离心机(Anke, T.G.L-16-aR)在11 269.44×
g 下4°C离心5分钟, 取3 mL装入玻璃比色皿, 用双光束紫外可见分光光度计(TU-1901, 北京)测定在649和665 nm处的吸光度, 计算各处理的Chla和Chlb含量(
王学奎, 2006 )。
1.2.3 叶黄素各组分含量的测定
叶黄素各组分含量采用高效液相色谱(high performance liquid chromatography, HPLC)法测定, 色谱检测系统为Waters1525型高效液相色谱仪, 2998二级阵列检测器, 2707自动进样器, Empower色谱管理软件和C30 (Devolosil, Nomura Chemical Co. LTD)色谱柱(流速为每分钟1.0 mL), 检测波长为450 nm。称取上述冰盒中的样品0.2 g放入10 mL离心管中, 用液氮粉碎机将其打成粉末状, 加入2 mL色素提取液(正己烷:丙酮:无水乙醇=2:1:1, 含0.01% BHT即2, 6-二叔丁基-4-甲基苯酚), 用超声仪(SCIENTZ, SB-3200- DTDN) 4°C超声10分钟, 置于离心机(Thermo, Fre- sco 17)中, 在13 684.32×
g 下4°C离心5分钟, 重复3次, 色素提取完全后, 加入1 mL 10% KOH-甲醇液, 12小时后加入500 μL 1%的NaCl溶液, 取上清用离心蒸发仪(EYELA, 日本)悬干, 再用甲基叔丁基醚(Methyl tert-Butyl Ether, MtBE)定容至500 μL, 之后用0.22 μm有机微孔滤膜过滤, 并装入高效液相色谱进样瓶(2 mL)内的250 μL的玻璃内衬管中。根据特征吸收峰以及标准品进行信号峰鉴定(
曹洪波, 2012 , 2015)。环氧玉米黄质(A)、双环氧紫黄质(V)和玉米黄质(Z)标准品购自CaroteNature公司(Lupsingen, Swit- zerland)。
1.3 数据处理所测数据采用Excel 2016和SPSS 17.0软件进行统计分析。使用单因素(one-way ANOVA), Duncan SSR test法进行方差分析和差异显著性检验(α=0.05), 及两个样本平均数的统计假设检验(two-sample
t -test, assuming μ
1 2 =μ
2 2 ; α=0.05)。表中数据为平均值±标准误。
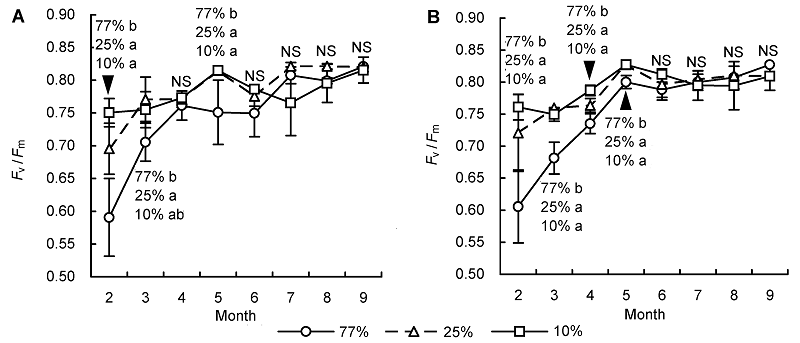
2 结果与讨论2.1 不同光照和水分条件对
F v /
F m 的影响光照、干旱和低温等引起的光抑制现象在自然界植物中普遍存在,
F v /
F m 是表征光抑制程度的良好指标和探针。生长状态良好的植物
F v /
F m 值介于0.75-0.85之间, 低于0.75则表明植物生长不良(
许大全等, 1992 ;
沈亮等, 2015 )。由
图1 可知, 2个水分条件下77%和25%透光区臭柏幼苗的
F v /
F m 均在低温的2月达到全年最低值, 且都在0.75以下; 而10%透光区的
F v /
F m 值在0.75以上。可见光照越强,
F v /
F m 的降低幅度越大, 且77%透光区的
F v /
F m 值均显著低于另外两个透光区。随着气候转暖, 77%和25%透光区的
F v /
F m 值迅速上升, 5月以后趋于稳定, 各透光区间均无显著差异, 且均保持在0.75-0.85之间。说明77%和25%透光区在2月的低温期不同程度地受到了光抑制, 77%透光区受到的光抑制程度最大。随着气温升高, 臭柏幼苗的光能利用能力逐渐增强且趋于稳定, 表明低温胁迫是引起77%和25%透光区出现光抑制现象的主要因素。与77%和25%透光区相比, 10%透光区的
F v /
F m 值随着季节变化的波动幅度较小, 全年维持在0.75以上, 说明其受到光抑制的影响较小。水分条件对3个光照区
F v /
F m 的影响在2-4月间无明显差异(
表1 ); 进入5月, 25%和10%透光区则表现出一定的差异性; 而77%透光区仅在9月出现差异(
表1 )。综上所述, 2个水分条件下低温均导致了77%以及25%透光区 出现光抑制, 且77%透光区的光抑制现象更明显, 10%透光区的光抑制影响较小。水分因子在5月以后作用明显, 特别是对25%和10%透光区的影响更突出。
图1 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-278/img_1.png <b>图1</b> 干旱(A)和湿润区(B)不同光照条件下臭柏幼苗<i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>的季节变化<br/><i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: 最大光化学效率。不同小写字母表示不同光照条件下差异显著(α=0.05)。NS表示不同光照条件下无显著差异。<br/><b>Figure 1</b> Seasonal variation of <i>F</i><sub>v</sub>/<i>F</i><sub>m</sub> of <i>Sabina vulgaris</i> seedlings growing in dry (A) and wet (B) areas under different light conditions <br/><i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: Maximal quantum yield. Different lowercase letters indicate significant difference at 0.05 level under different light conditions. NS indicate no significant difference under different light conditions. Figure 1 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-278/img_1.png <b>图1</b> 干旱(A)和湿润区(B)不同光照条件下臭柏幼苗<i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>的季节变化<br/><i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: 最大光化学效率。不同小写字母表示不同光照条件下差异显著(α=0.05)。NS表示不同光照条件下无显著差异。<br/><b>Figure 1</b> Seasonal variation of <i>F</i><sub>v</sub>/<i>F</i><sub>m</sub> of <i>Sabina vulgaris</i> seedlings growing in dry (A) and wet (B) areas under different light conditions <br/><i>F</i><sub>v</sub>/<i>F</i><sub>m</sub>: Maximal quantum yield. Different lowercase letters indicate significant difference at 0.05 level under different light conditions. NS indicate no significant difference under different light conditions. 在新窗口打开 下载 下载原图ZIP 生成PPT 图1
干旱(A)和湿润区(B)不同光照条件下臭柏幼苗
F v /
F m 的季节变化
F v /
F m : 最大光化学效率。不同小写字母表示不同光照条件下差异显著(α=0.05)。NS表示不同光照条件下无显著差异。
Figure 1
Seasonal variation of
F v /
F m of
Sabina vulgaris seedlings growing in dry (A) and wet (B) areas under different light conditions
F v /
F m : Maximal quantum yield. Different lowercase letters indicate significant difference at 0.05 level under different light conditions. NS indicate no significant difference under different light conditions.
表1 Table 1 表1 表1 不同水分条件下臭柏幼苗
F v /
F m 的比较(平均值±标准误,
n =9)
Table 1 Comparison of
F v /
F m of
Sabina vulgaris seedlings under different water conditions (means±SE,
n =9)
Month Water condition 77% light transmittance area 25% light transmittance area 10% light transmittance area February Dry area 0.5906±0.0592 a 0.6953±0.0389 a 0.7504±0.0217 a Wet area 0.6057±0.0568 a 0.7206±0.0601 a 0.7609±0.0201 a March Dry area 0.7053±0.0290 a 0.7708±0.0339 a 0.7550±0.0274 a Wet area 0.6812±0.0250 a 0.7597±0.0004 a 0.7494±0.0106 a April Dry area 0.7612±0.0223 a 0.7710±0.0126 a 0.7731±0.0105 a Wet area 0.7349±0.0155 a 0.7636±0.0107 a 0.7870±0.0081 a May Dry area 0.7509±0.0491 a 0.8150±0.0053 b 0.8151±0.0068 b Wet area 0.8000±0.0102 a 0.8271±0.0045 a 0.8273±0.0025 a June Dry area 0.7496±0.0358 a 0.7754±0.0153 a 0.7862±0.0047 b Wet area 0.7881±0.0119 a 0.7963±0.0244 a 0.8120±0.0028 a July Dry area 0.8075±0.0128 a 0.8214±0.0058 a 0.7653±0.0501 a Wet area 0.7998±0.0125 a 0.8041±0.0134 a 0.7947±0.0230 a August Dry area 0.7991±0.0043 a 0.8213±0.0042 a 0.7952±0.0291 a Wet area 0.8073±0.0175 a 0.8102±0.0219 a 0.7944±0.0375 a September Dry area 0.8209±0.0009 b 0.8200±0.0007 a 0.8154±0.0197 a Wet area 0.8271±0.0031 a 0.8097±0.0010 b 0.8091±0.0220 a
Different lowercase letters indicate significant difference at 0.05 level under different water conditions. 不同小写字母表示在不同水分条件下差异显著(α=0.05)。 表1
不同水分条件下臭柏幼苗
F v /
F m 的比较(平均值±标准误,
n =9)
Table 1
Comparison of
F v /
F m of
Sabina vulgaris seedlings under different water conditions (means±SE,
n =9)
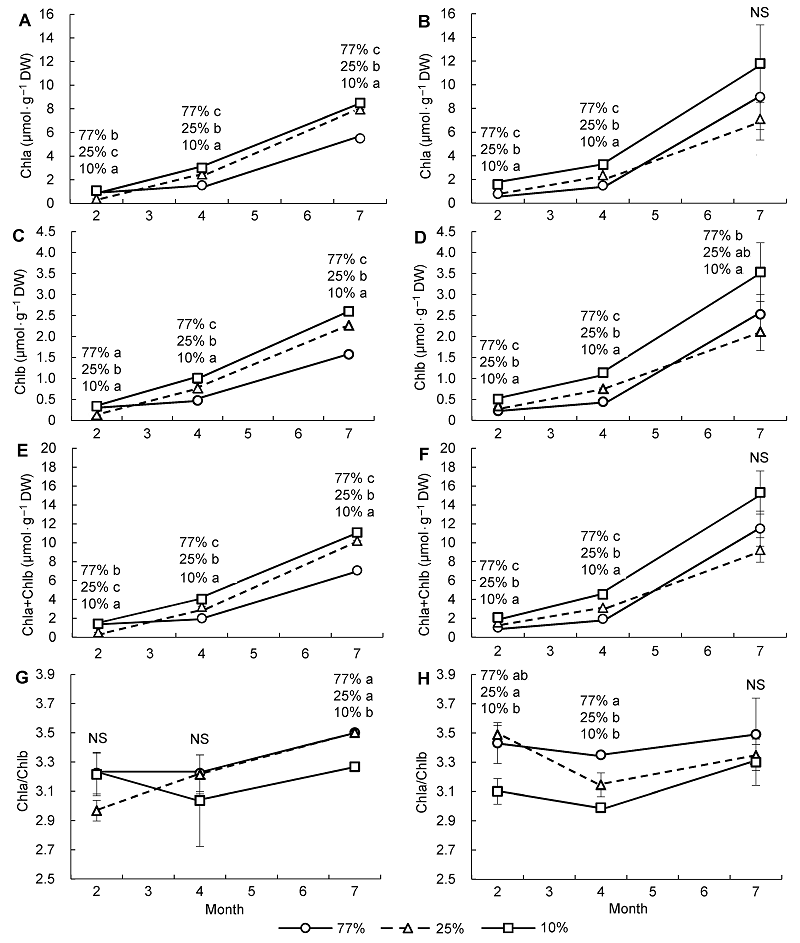
2.2 不同光照和水分条件对叶绿素含量的影响
图2 表明, 3个光照处理区的Chla、Chlb和Chla+Chlb含量在2个水分条件下均随着气候转暖呈上升趋势。在2月低温期所有光照处理区的臭柏幼苗均受到干旱胁迫的影响(
表2 )。4月以后所有光照处理区臭柏的叶绿素含量急剧增加, 说明由冬末经春季到夏季, 随着温度的升高, 臭柏的光合作用越来越活跃, 春季开始进入光合旺盛期。低温和干旱双重胁迫制约了臭柏的叶绿素合成, 但各光照处理区变化不同。随着水分条件的改善和温度的回升, 77%和10%透光区的叶绿素含量逐渐增加。其中, 10%透光区Chla、Chlb和Chla+ Chlb含量始终高于77%和25%透光区, 而Chla/Chlb几乎均低于77%和25%透光区, 说明10%透光区主要是增加Chlb含量以捕捉更多光能。77%透光区几乎与之相反, 除了干旱条件下2月低温期, 其Chla、Chlb以及Chla+Chlb含量始终低于10%和25%透光区, Chla/Chlb则几乎均高于10%和25%透光区, 且随着温度的升高, Chla/Chlb值增大。表明77%透光区主要通过减少Chlb含量来避免光能过剩, 在高温期Chla和Chlb的转换可能更为活跃。25%透光区的Chla、Chlb和Chla+Chlb含量及Chla/Chlb受低温与干旱胁迫的影响更大。干旱条件下, 25%透光区低温2月的4项指标均低于10%和77%透光区。其中, 叶绿素含量与10%和77%透光区相比差异显著(
图2 ); 湿润条件下, 25%透光区低温2月的4项指标均高于77%透光区, 同样其叶绿素含量与77%透光区相比差异显著(
图2 )。结合
表2 可以看出, 虽然所有光照处理区在低温期均不同程度地受到水分和低温胁迫的影响, 但25%透光区受水分和低温胁迫的影响更为突出。因为25%透光区在低温和干旱双重胁迫下不仅叶绿素含量降低, 而且Chla/Chlb也显著降低。说明低温和干旱胁迫下25%透光区的臭柏叶绿素合成受阻, 故可推测低温和干旱胁迫对25%透光区叶绿素合成的影响要大于10%和77%透光区。
图2 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-278/img_2.png <b>图2</b> 干旱(A、C、E、G)和湿润区(B、D、F、H)不同光照条件下臭柏幼苗叶绿素含量随季节的变化 <br/>Chla: 叶绿素a含量; Chlb: 叶绿素b含量; Chla+Chlb: 叶绿素含量; Chla/Chlb: 叶绿素a含量与叶绿素b含量之比。不同小写字母表示不同光照条件下差异显著(α=0.05)。NS表示不同光照条件下无显著差异。<br/><b>Figure 2</b> Seasonal variation of chlorophyll content of <i>Sabina vulgaris</i> seedlings growing in dry (A, C, E, G) and wet (B, D, F, H) areas under different light conditions<br/>Chla: The content of chlorophyll a; Chlb: The content of chlorophyll b; Chla+Chlb: The content of chlorophyll; Chla/Chlb: The ratio of chlorophyll a to chlorophyll b. Different lowercase letters indicate significant difference at 0.05 level under different light conditions. NS meant no significant difference under different light conditions. Figure 2 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-278/img_2.png <b>图2</b> 干旱(A、C、E、G)和湿润区(B、D、F、H)不同光照条件下臭柏幼苗叶绿素含量随季节的变化 <br/>Chla: 叶绿素a含量; Chlb: 叶绿素b含量; Chla+Chlb: 叶绿素含量; Chla/Chlb: 叶绿素a含量与叶绿素b含量之比。不同小写字母表示不同光照条件下差异显著(α=0.05)。NS表示不同光照条件下无显著差异。<br/><b>Figure 2</b> Seasonal variation of chlorophyll content of <i>Sabina vulgaris</i> seedlings growing in dry (A, C, E, G) and wet (B, D, F, H) areas under different light conditions<br/>Chla: The content of chlorophyll a; Chlb: The content of chlorophyll b; Chla+Chlb: The content of chlorophyll; Chla/Chlb: The ratio of chlorophyll a to chlorophyll b. Different lowercase letters indicate significant difference at 0.05 level under different light conditions. NS meant no significant difference under different light conditions. 在新窗口打开 下载 下载原图ZIP 生成PPT 图2
干旱(A、C、E、G)和湿润区(B、D、F、H)不同光照条件下臭柏幼苗叶绿素含量随季节的变化
Chla: 叶绿素a含量; Chlb: 叶绿素b含量; Chla+Chlb: 叶绿素含量; Chla/Chlb: 叶绿素a含量与叶绿素b含量之比。不同小写字母表示不同光照条件下差异显著(α=0.05)。NS表示不同光照条件下无显著差异。
Figure 2
Seasonal variation of chlorophyll content of
Sabina vulgaris seedlings growing in dry (A, C, E, G) and wet (B, D, F, H) areas under different light conditions
Chla: The content of chlorophyll a; Chlb: The content of chlorophyll b; Chla+Chlb: The content of chlorophyll; Chla/Chlb: The ratio of chlorophyll a to chlorophyll b. Different lowercase letters indicate significant difference at 0.05 level under different light conditions. NS meant no significant difference under different light conditions.
表2 Table 2 表2 表2 不同水分条件下臭柏幼苗叶绿素含量的比较(平均值±标准误,
n =9)
Table 2 Comparison of chlorophyll content of
Sabina vulgaris seedlings under different water conditions (means±SE,
n =9)
Index Light transmittance area (%) Water Month February April July The content of-1 DW) 77 Dry area 0.9955±0.0275 a 1.5270±0.1533 a 5.5013±0.0415 b Wet area 0.7795±0.0201 b 1.4898±0.0647 a 8.9617±2.7541 a 25 Dry area 0.3808±0.0085 b 2.4627±0.0565 a 7.9338±0.0171 a Wet area 1.1987±0.0153 a 2.3646±0.0396 b 7.1213±1.7907 a 10 Dry area 1.0732±0.0275 b 3.0017±0.1674 b 8.4823±0.0127 a Wet area 1.5720±0.0431 a 3.3950±0.0250 a 11.7880±3.2639 a The content of-1 DW) 77 Dry area 0.3095±0.0152 a 0.4729±0.0158 a 1.5713±0.0110 b Wet area 0.2277±0.0102 b 0.4447±0.0184 a 2.5279±0.4804 a 25 Dry area 0.1284±0.0024 b 0.7678±0.0377 a 2.2659±0.0046 a Wet area 0.3436±0.3436 a 0.7524±0.7524 a 2.1158±0.4523 a 10 Dry area 0.3348±0.0179 b 1.0034±0.1256 a 2.5961±0.0037 b Wet area 0.5074±0.0107 a 1.1357±0.0053 a 3.5336±0.6975 a The content of-1 DW) 77 Dry area 1.3050±0.0071 a 2.0000±0.0976 a 7.0726±0.0303 b Wet area 1.0072±0.0057 b 1.9345±0.0479 a 11.4896±1.8669 a 25 Dry area 0.5092±0.0035 b 3.2305±0.0109 a 10.1997±0.0125 a Wet area 1.5423±0.0053 a 3.1170±0.0107 b 9.2371±1.2949 a 10 Dry area 1.4080±0.0055 b 4.0051±0.0242 b 11.0784±0.0094 a Wet area 2.0794±0.0187 a 4.5307±0.0175 a 15.3216±2.2867 a Chla/Chlb 77 Dry area 3.2243±0.1428 a 3.2242±0.1252 a 3.5011±0.0011 a Wet area 3.4312±0.1391 a 3.3505±0.0042 a 3.4908±0.2481 a 25 Dry area 2.9679±0.0706 b 3.2149±0.1334 a 3.5014±0.0003 a Wet area 3.4897±0.0615 a 3.1454±0.0810 a 3.3471±0.0757 a 10 Dry area 3.2147±0.1464 a 3.0365±0.3132 a 3.2674±0.0002 a Wet area 3.1006±0.0869 a 2.9893±0.0047 a 3.3000±0.1581 a
Chla/Chlb: The ratio of chlorophyll a to chlorophyll b. Different lowercase letters indicate significant difference at 0.05 level under different water conditions. Chla/Chlb: 叶绿素a含量与叶绿素b含量之比。不同小写字母表示在不同水分条件下差异显著(α=0.05)。 表2
不同水分条件下臭柏幼苗叶绿素含量的比较(平均值±标准误,
n =9)
Table 2
Comparison of chlorophyll content of
Sabina vulgaris seedlings under different water conditions (means±SE,
n =9)
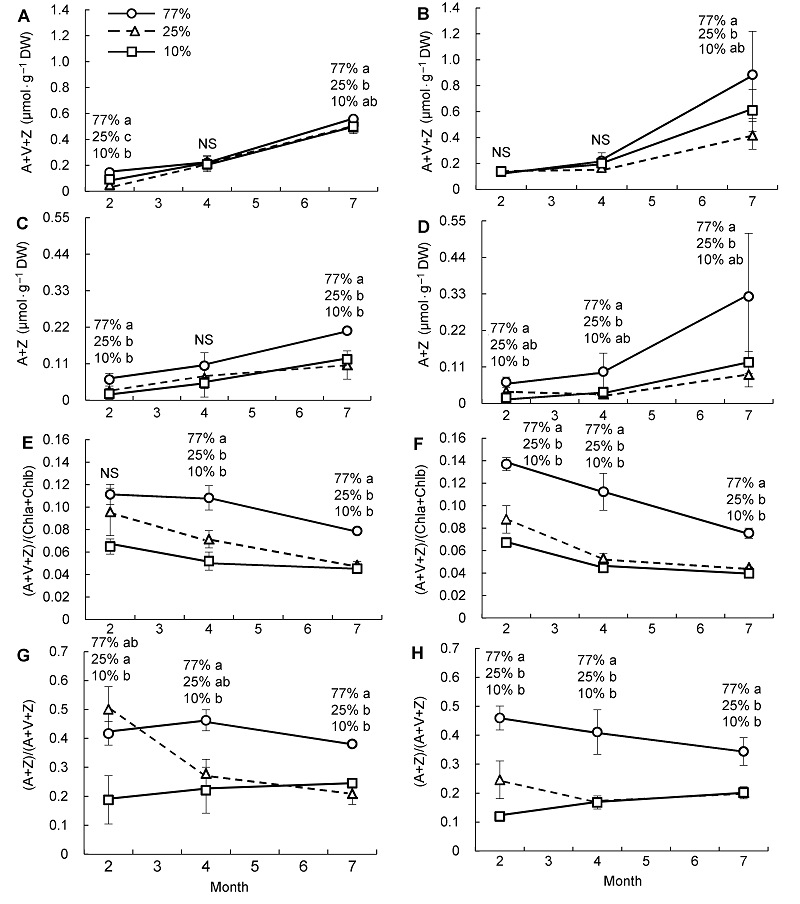
2.3 不同光照和水分条件对叶黄素含量的影响由
图3 可知, 2个水分条件下各透光区的臭柏A+V+Z和A+Z含量均随着季节的变化呈上升趋势, 尤其4月以后增幅变大, 说明由冬末经春季到夏季, 臭柏叶黄素循环越来越活跃, 但(A+V+Z)/(Chla+Chlb)随着季节的变化均呈下降趋势, 这是由于Chla+Chlb含量随着季节的变化呈显著上升所致(
图2 )。进入生长季后, 对植物而言叶绿素的合成较叶黄素合成更重要。虽然生长季植物需要更多的叶绿素, 但不同透光区叶黄素的合成量仍有所不同。77%透光区臭柏的A+V+Z含量、A+Z含量、(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)普遍高于25%和10%透光区(
图3 ;
表3 ), 说明77%透光区光照强, 光能过剩, 为避免光破坏, 臭柏幼苗通过增加叶黄素的含量以提高叶黄素循环的热耗散转化率, 进而耗散过剩的光能。10%透光区与77%透光区的变化则几乎相反, 说明10%透光区光照较弱, 没有过剩光能, 无须合成过多的叶黄素, 而是更多地合成叶绿素捕捉光能。25%透光区叶黄素的含量与10%透光区相近, 而(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V +Z)却大于10%透光区。特别是干旱条件下, 25%透光区低温2月的(A+V)/(A+V+Z)达到3个透光区的最大值, 而湿润条件下介于10%与77%透光区之间(
图3 ;
表3 )。这是由于在干旱和低温双重胁迫下, 25%透光区叶绿素的合成受阻(
图2 ), 所以叶黄素循环的热耗散转化率增加, 从而提高了臭柏幼苗叶黄素循环防御光抑制的能力。
图3 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-278/img_3.png <b>图3</b> 干旱(A、C、E、G)和湿润区(B、D、F、H)不同光照条件下臭柏幼苗叶黄素含量随季节的变化<br/>A+V+Z: 叶黄素含量; A+Z: 热散逸色素含量; (A+V+Z)/(Chla+Chlb): 叶黄素含量与叶绿素含量之比; (A+Z)/(A+V+Z): 叶黄素循环的热耗散转化率。不同小写字母表示不同光照条件下差异显著(α=0.05)。NS表示不同光照条件下无显著差异。<br/><b>Figure 3</b> Seasonal variation of xanthophyll content of <i>Sabina vulgaris</i> seedlings growing in dry (A, C, E, G) and wet (B, D, F, H) areas under different light conditions<br/>A+V+Z: The content of xanthophyll; A+Z: The content of antheraxanthin and zeaxanthin; (A+V+Z)/(Chla+Chlb): The ratio of xanthophyll to chlorophyll; (A+Z)/(A+V+Z): Thermal dissipation transformation rate of xanthophyll cycle. Different lowercase letters indicate significant difference at 0.05 level under different light conditions. NS meant no significant difference under different light conditions. Figure 3 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-278/img_3.png <b>图3</b> 干旱(A、C、E、G)和湿润区(B、D、F、H)不同光照条件下臭柏幼苗叶黄素含量随季节的变化<br/>A+V+Z: 叶黄素含量; A+Z: 热散逸色素含量; (A+V+Z)/(Chla+Chlb): 叶黄素含量与叶绿素含量之比; (A+Z)/(A+V+Z): 叶黄素循环的热耗散转化率。不同小写字母表示不同光照条件下差异显著(α=0.05)。NS表示不同光照条件下无显著差异。<br/><b>Figure 3</b> Seasonal variation of xanthophyll content of <i>Sabina vulgaris</i> seedlings growing in dry (A, C, E, G) and wet (B, D, F, H) areas under different light conditions<br/>A+V+Z: The content of xanthophyll; A+Z: The content of antheraxanthin and zeaxanthin; (A+V+Z)/(Chla+Chlb): The ratio of xanthophyll to chlorophyll; (A+Z)/(A+V+Z): Thermal dissipation transformation rate of xanthophyll cycle. Different lowercase letters indicate significant difference at 0.05 level under different light conditions. NS meant no significant difference under different light conditions. 在新窗口打开 下载 下载原图ZIP 生成PPT 图3
干旱(A、C、E、G)和湿润区(B、D、F、H)不同光照条件下臭柏幼苗叶黄素含量随季节的变化
A+V+Z: 叶黄素含量; A+Z: 热散逸色素含量; (A+V+Z)/(Chla+Chlb): 叶黄素含量与叶绿素含量之比; (A+Z)/(A+V+Z): 叶黄素循环的热耗散转化率。不同小写字母表示不同光照条件下差异显著(α=0.05)。NS表示不同光照条件下无显著差异。
Figure 3
Seasonal variation of xanthophyll content of
Sabina vulgaris seedlings growing in dry (A, C, E, G) and wet (B, D, F, H) areas under different light conditions
A+V+Z: The content of xanthophyll; A+Z: The content of antheraxanthin and zeaxanthin; (A+V+Z)/(Chla+Chlb): The ratio of xanthophyll to chlorophyll; (A+Z)/(A+V+Z): Thermal dissipation transformation rate of xanthophyll cycle. Different lowercase letters indicate significant difference at 0.05 level under different light conditions. NS meant no significant difference under different light conditions.
表3 Table 3 表3 表3 不同水分条件下臭柏幼苗叶黄素含量的比较(平均值±标准误,
n =9)
Table 3 Comparison of xanthophyll content of
Sabina vulgaris seedlings under different water conditions (means±SE,
n =9)
Index Light transmittance area (%) Water Month February April July The content of-1 DW) 77 Dry area 0.1453±0.0215 a 0.2192±0.0557 a 0.5573±0.0103 a Wet area 0.1377±0.0117 a 0.2188±0.0643 a 0.8835±0.3357 a 25 Dry area 0.0489±0.0192 b 0.2308±0.0444 a 0.4958±0.0505 a Wet area 0.1355±0.0334 a 0.1659±0.0221 b 0.4167±0.1074 a 10 Dry area 0.0915±0.0172 b 0.2083±0.0584 a 0.4996±0.0085 a Wet area 0.1395±0.0029 a 0.2016±0.0029 a 0.6092±0.1626 a The content of-1 DW) 77 Dry area 0.1453±0.0215 a 0.2192±0.0557 a 0.5573±0.0103 a Wet area 0.1377±0.0117 a 0.2188±0.0643 a 0.8835±0.3357 a 25 Dry area 0.0489±0.0192 a 0.2308±0.0444 a 0.4958±0.0505 a Wet area 0.1355±0.0334 a 0.1659±0.0221 a 0.4167±0.1074 a 10 Dry area 0.0915±0.0172 a 0.2083±0.0584 a 0.4996±0.0085 a Wet area 0.1395±0.0029 a 0.2016±0.0029 a 0.6092±0.1626 a (A+V+Z)/ 77 Dry area 0.1113±0.0089 b 0.1082±0.0108 a 0.0788±0.0005 a Wet area 0.1369±0.0059 a 0.1123±0.0164 a 0.0754±0.0046 a 25 Dry area 0.0957±0.0211 a 0.0714±0.0077 a 0.0486±0.0028 a Wet area 0.0878±0.0122 a 0.0532±0.0039 a 0.0450±0.0004 a 10 Dry area 0.0649±0.0068 a 0.0519±0.0081 a 0.0451±0.0004 a Wet area 0.0671±0.0002 a 0.0445±0.0002 a 0.0397±0.0002 b (A+V)/ 77 Dry area 0.4170±0.0406 a 0.4624±0.0365 a 0.3805±0.0016 a Wet area 0.4593±0.0416 a 0.4106±0.0770 b 0.3435±0.0479 a 25 Dry area 0.5003±0.0788 a 0.2786±0.0490 a 0.2092±0.0373 a Wet area 0.2461±0.0647 b 0.1687±0.0136 b 0.2012±0.0208 a 10 Dry area 0.1878±0.0834 a 0.2213±0.0790 a 0.2457±0.0003 a Wet area 0.1194±0.0003 a 0.1680±0.0225 a 0.2023±0.0007 b
A+V+Z, A+Z, (A+V+Z)/(Chla+Chlb) and (A+Z)/(A+V+Z) see Figure 3 . Different lowercase letters indicate significant difference at 0.05 level under different water conditions. A+V+Z、A+Z、(A+V+Z)/(Chla+Chlb)和(A+Z)/(A+V+Z)同图3 。不同小写字母表示在不同水分条件下差异显著(α=0.05)。 表3
不同水分条件下臭柏幼苗叶黄素含量的比较(平均值±标准误,
n =9)
Table 3
Comparison of xanthophyll content of
Sabina vulgaris seedlings under different water conditions (means±SE,
n =9)
2.4 讨论如前所述,
F v /
F m 是表征光抑制程度的良好指标和探针。生长状态良好的植物的
F v /
F m 值介于0.75-0.85之间, 低于0.75则生长不良(
许大全等, 1992 ;
沈亮等, 2015 )。本研究结果表明, 冬季低温导致不同透光区臭柏幼苗的
F v /
F m 值差异明显, 77%和25%透光区的
F v /
F m 值降低至0.75以下, 且77%透光区下降幅度最大, 光抑制现象更明显, 10%透光区的
F v /
F m 值全年都介于0.75-0.85之间, 未受到明显的光抑制。但随着季节转换气候变暖, 77%和25%透光区的
F v /
F m 值迅速上升, 5月以后趋于稳定, 且3个透光区间无显著差异, 均保持在0.75-0.85间(
图1 ); 77%和25%透光区的光合作用已从低迷的光抑制状态进入正常的光合旺盛阶段, 那么77%和25%透光区必定启动了光抑制防御机制。朱成刚等(2011)在研究干旱胁迫下胡杨(
Popu- lus euphratica ) PSII光化学效率和激能耗散时也发现, 其
F v /
F m 值在生长季并未降低, 这可能是干旱地区耐旱植物适应干旱强光环境的共性之一。
光抑制防御机制对植物的生存与正常生长发育至关重要。叶绿素循环抵御光抑制的机理是在光能过剩时, 天线色素Chlb转化成特殊激活状态的Chla, 以便将更多的光能转化成化学能, 固定更多的CO
2 , 生产更多的有机物。Chlb是LHCII的重要组成部分, 可调控光合机构天线的大小并维持LHCII的稳定性。强光下LHCII主要通过减少Chlb来降低吸收光能并抵御光胁迫(娄世庆等, 1995; 陈辉等, 2004;
郭春爱等, 2006 )。同时, 叶绿素循环可能是一种不引起光合机构破坏的光抑制防御机制(
许大全, 2003 )。光抑制下, 参与光化学反应的Chla比收集光能的Chlb更重要, 因而Chla/Chlb是植物抵御光抑制的重要指标之一, Chla/Chlb升高有利于充分利用光能。光抑制最直接的表现为光能过剩, 过剩光能的耗散在调节光能吸收和电子传递上起关键作用, 并且在防止光合器官破坏上不可或缺(
王强等, 2003 )。据统计, 在光照和水分充足时, 植物吸收的光有近50%以热耗散的形式消耗掉。例如, 在干旱和低温胁迫下, 热耗散掉的能量可达吸收光能总量的70%-90% (Flexas et al., 2002)。可见被称为“安全阀”的热耗散机制可以高效地保护植物在逆境胁迫下光合机构免受光抑制的破坏, 是光抑制防御的中心和基石, 多年来备受关注(
许大全等, 1992 )。而叶黄素循环是热耗散途径的主要形式之一, 在植物的抗光抑制中非常重要(Niyogi, 1999; ller et al., 2001)。
77%透光区的臭柏实生苗可通过减少Chlb含量, 升高Chla/Chlb值来避免光能过剩, 这也是耐阳植物适应强光环境的正常响应(
陈煜等, 2006 ;
姜武等, 2007 ;
邹长明等, 2015 )。然而, 4月干旱条件下77%透光区的Chla/Chlb与25%和10%透光区相比无显著差异, 但在湿润区显著大于25%和10%透光区(
图2 ), 说明干旱条件下77%透光区的叶绿素循环光抑制防御机制不及湿润区活跃。如果干旱会加重光抑制的程度, 湿润区都已存在叶绿素循环抵御光抑制的现象, 那么为何干旱区却反而未出现此现象?这可能是由于持续的强光和干旱双重胁迫导致了严重的光抑制, 进而产生了光破坏, 使臭柏幼苗无法启动叶绿素循环。光破坏发生在PSII的反应中心, 直接造成了反应中心色素(进行光能传递和转化且具有光化学活性的特殊态Chla)的大量损失, 使得Chla的含量降低, 从而使Chla/Chlb降低, 结果无法完成叶绿素循环的运作(
潘瑞炽, 2004 )。当然, 臭柏幼苗会通过其它途径来应对严重的光抑制, 本研究中, 干旱条件下77%透光区4月的(A+V)/(A+V+Z)显著高于湿润区(
图3 ), 说明臭柏利用较高的叶黄素循环转化率来加强叶黄素循环的运转, 以消耗过剩的光能。已有研究表明, 叶绿素和类胡萝卜素的光能吸收有一定的互补性(司继播等, 2007), 而且植物可能形成一种稳定的叶绿素-叶黄素-蛋白淬灭复合物来耗散过剩的光能(
李玉灵等, 2009 )。由此可见, 臭柏幼苗应对光抑制的色素机制也是协调统一的。77%透光区臭柏幼苗的光抑制响应机制灵敏、快速且高效, 并且光抑制防御能力较强, 叶绿素循环与叶黄素循环机制同时发生作用, 可达到光能的最佳分配。
25%透光区的臭柏幼苗对温度和水分条件较为敏感。在低温的2月, 干旱条件下25%透光区臭柏的叶绿素各项指标均低于77%和10%透光区, (A+V)/ (A+V+Z)达到3个透光区的最大值; 湿润条件下, 25%透光区的叶绿素各项指标均高于77%透光区, (A+V)/ (A+V+Z)介于10%和77%透光区之间(
图2 ,
图3 )。说明25%透光区易受水分和低温胁迫的影响。在干旱和低温双重胁迫下, 25%透光区叶绿素的合成受阻, 叶黄素循环热耗散的转化率增强, 从而提高了臭柏幼苗叶黄素循环防御光抑制的能力。这与简启亮等(2010)对祁连圆柏(
Sabina przewalskii )和青海云杉(
Picea crassifolia )叶片色素的研究结果相一致。25%透光区臭柏幼苗的光抑制响应机制随水分和温度条件的变化而迅速改变, 叶绿素循环和叶黄素循环机制也因水分与温度条件的不同而协同作用, 以达到最佳的生长状态。
10%透光区的臭柏幼苗因光照强度最弱, 叶绿素含量明显高于其它两个遮光处理区, 而不易受到光抑制的影响。在不同水分条件下, 10%透光区的Chla+ Chlb随着季节的变化均显著高于25%透光区, Chla/ Chlb均低于25%透光区, 同时叶黄素组分含量与25%透光区无显著差异(
图2 ,
图3 )。即使在湿润条件下, 光照不足(10%透光区)的影响也很明显, 天线色素的比例与干旱区无明显差异。10%透光区全年都不会发生光能过剩, 自然也没有必要进行热散逸, 因此叶黄素所占比例最小。叶绿素循环和叶黄素循环均不活跃, 很少启动光抑制防御机制。10%透光区主要通过增加Chlb的含量, 降低Chla/Chlb值, 捕捉更多的光能, 这也是耐荫植物对阴暗环境做出的响应(
杨渺等, 2004 ;
史振声等, 2013 )。
臭柏在干旱环境中经过长期的自然选择和协同进化, 从生理到形态上形成了一系列微妙的耐旱、耐低温和耐强光等逆境生理生态适应对策(
李玉灵等, 2009 )。总体而言, 77%透光区无疑最易受到光抑制, 且强光是引起光抑制的主要原因。但77%透光区一方面可通过减少Chlb含量, 升高Chla/Chlb值来避免光能过剩; 另一方面可通过增加A+V+Z和A+Z含量、提高(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)值, 耗散过剩的光能, 避免光破坏。10%透光区的臭柏幼苗则与77%透光区相反, 未受到光抑制, 其主要面临的是光照不足问题。10%透光区主要通过增加Chlb含量, 降低Chla/Chlb值, 捕捉更多的光能, 不启动或很少启动光抑制防御机制。从77%和10%透光区对不同光照强度的响应策略可以看出, 臭柏集减少光能吸收和捕捉光能于一身, 臭柏幼苗叶绿素和叶黄素循环对不同光照环境的响应具有协调互补的灵活性。25%透光区的光资源可能既非过剩也无不足, 在低温和干旱双重胁迫下不仅叶绿素含量降低, 而且Chla/Chlb值也显著降低; 所以对水分和低温胁迫的影响做出了明显的响应, 叶绿素和叶黄素循环机制随着水分条件变化而迅速改变。可见, 臭柏叶绿素循环和叶黄素循环不仅对光照有瞬时响应, 而且对温度和水分也能做出及时响应。这就解释了为何毛乌素沙地的臭柏实生苗能逐渐摆脱乌柳的庇护, 完全暴露在强光下, 完成天然更新; 同时也体现出了物种的适者生存。该研究对臭柏种质资源保护和人工造林具重要意义。
The authors have declared that no competing interests exist. 参考文献 文献选项 原文顺序 文献年度倒序 文中引用次数倒序 被引期刊影响因子 [1] 曹洪波 (2012 ). 转基因调控柑橘类胡萝卜素积累的细胞学和代谢研究. 博士论文. 武汉: 华中农业大学. pp . 35 .URL [2] 陈煜 , 杨志民 , 李志华 (2006 ). 草坪草耐荫性研究进展. 中国草地学报 28, 71 -76 .10.3321/j.issn:1673-5021.2006.03.014 URL [3] 代欣 , 胡举伟 , 张秀丽 , 孙广玉 (2016 ). 植物光合机构对光环境的适应机制. 应用生态学报 27, 1674 -1682 .[4] 董新甜 , 曹洪波 , 张飞燕 , 韩艳 , 贾浩 , 李丹 , 张学英 , 陈海江 (2015 ). 遮光性套袋对黄肉桃类胡萝卜素合成及相关基因表达的影响. 园艺学报 42, 633 -642 .URL [5] 冯玉静 (2015 ). 毛乌素沙地东南缘沉积物光释光测年与环境变迁研究. 硕士论文. 兰州: 兰州大学 . pp. 1 -20 .URL [6] 付忠 , 谢世清 , 徐文果 , 岩所 , 陈军文 (2016 ). 不同光照强度下谢君魔芋的光合作用及能量分配特征. 应用生态学报 27, 1177 -1188 .10.13287/j.1001-9332.201604.035 URL [7] 高杰 , 张仁和 , 王文斌 , 李志伟 , 薛吉全 (2015 ). 干旱胁迫对玉米苗期叶片光系统II性能的影响. 应用生态学报 26, 1391 -1396 .URL [8] 郭春爱 (2007 ). 低叶绿素b对水稻光合机构及其热稳定性的影响. 硕士论文. 南京: 南京农业大学 . pp. 1 -5 .URL [9] 郭春爱 , 刘芳 , 许晓明 (2006 ). 叶绿素b缺失与植物的光合作用. 植物生理学通讯 42, 967 -973 .URL [10] 黄秋娴 , 赵顺 , 刘春梅 , 李玉灵 (2015 ). 遮阴处理对铁尾矿基质臭柏实生苗快速叶绿素荧光特性的影响. 林业科学 51, 17 -26 .10.11707/j.1001-7488.20150603 URL [11] 简启亮 , 文陇英 , 陈拓 , 张满效 , 徐世健 (2010 ). 祁连圆柏和圆柏色素含量及其花青苷合成酶活性的季节性变化. 植物学报 45, 698 -704 .10.3969/j.issn.1674-3466.2010.06.007 URL [12] 姜武 , 姜卫兵 , 李志国 (2007 ). 园艺作物光合性状种质差异及遗传表现研究进展. 经济林研究 25, 102 -108 .10.3969/j.issn.1003-8981.2007.04.023 URL [13] 李晨阳 , 高慧 , 陈燕 , 谭为 , 赵军 , 徐芳 (2015 ). HPLC测定不同产地新疆圆柏中的槲皮苷. 华西药学杂志 30, 720 -721 .URL [14] 李玉灵 , 朱帆 , 王俊刚 , 三木直子 , 坂本圭儿 , 吉川贤 (2009 ). 水分胁迫下臭柏(Sabina vulgaris Ant.)光合特性和色素组成的季节变化. 生态学报 29, 4346 -4352 .10.3321/j.issn:1000-0933.2009.08.039 URL [15] 潘瑞炽 (2004 ). 植物生理学(第5版) . 北京: 高等教育出版社. pp. 64 -67 .[16] 沈亮 , 陈君 , 刘赛 , 徐荣 , 徐常青 , 刘同宁 (2015 ). 脱水胁迫和光合日变化对梭梭和白梭梭叶绿素荧光参数的影响. 应用生态学报 26, 2321 -2328 .URL [17] 师生波 , 李和平 , 王学英 , 李惠梅 , 韩发 (2007 ). 高山植物唐古特山莨菪和唐古特大黄对强太阳辐射光能的利用和耗散特性. 植物生态学报 31, 129 -137 .URL [18] 师生波 , 张怀刚 , 师瑞 , 李妙 , 陈文杰 , 孙亚男 (2014 ). 青藏高原春小麦叶片光合作用的光抑制及PSII反应中心光化学效率的恢复分析. 植物生态学报 38, 375 -386 .10.3724/SP.J.1258.2014.00034 URL [19] 史振声 , 钟雪梅 , 黄海皎 , 李凤海 , 王志斌 , 王宏伟 , 吕香玲 , 朱敏 (2013 ). 遮阴胁迫对不同耐阴性玉米叶绿素含量的影响. 玉米科学 21, 55 -58 .10.3969/j.issn.1005-0906.2013.04.012 URL [20] 孙鲁龙 , 耿庆伟 , 宋伟 , 邢浩 , 杜远鹏 , 翟衡 (2016 ). 不同光强对霜冻后葡萄叶片PSII光化学活性恢复的影响. 植物生理学报 8, 1243 -1247 .URL [21] 王林和 , 党宏忠 , 张国盛 , 党国锋 , 冯伟 , 高达布希拉图 , 万俊华 (2014 ). 中国天然臭柏群落的分布与生物量特征. 内蒙古农业大学学报 35, 37 -45 .URL [22] 王林和 , 张国盛 , 温国胜 , 李玉灵 , 吉川贤 (2011 ). 臭柏生理生态学特性及种群恢复与重建(第1版) . 北京: 科学出版社. pp. 13 -20 .[23] 王强 , 温晓刚 , 张其德 (2003 ). 光合作用光抑制的研究进展. 植物学通报 20, 539 -548 .10.3969/j.issn.1674-3466.2003.05.004 URL [24] 王学奎 (2006 ). 植物生理生化实验原理与技术(第2版) . 北京: 高等教育出版社. pp. 134-139, 195 -201 .[25] 许大全 (2003 ). 植物光胁迫研究中的几个问题. 植物生理学通讯 39, 493 -495 .URL [26] 许大全 , 张玉忠 , 张荣铣 (1992 ). 植物光合作用的光抑制. 植物生理学通讯 4, 237 -243 .URL [27] 杨渺 , 毛凯 , 苟文龙 , 赵波 (2004 ). 遮阴胁迫对叶绿素含量的影响. 四川草原 3, 20 -22 .10.3969/j.issn.1673-8403.2004.03.007 URL [28] 查倩 , 奚晓军 , 蒋爱丽 , 田益华 , 黄健 (2016 ). 高温胁迫对葡萄幼树叶绿素荧光特性和抗氧化酶活性的影响. 植物生理学报 52, 525 -532 .URL [29] 张国盛 , 王哲 , 王林和 , 郝云龙 , 温国胜 (2006 ). 毛乌素沙地天然臭柏居群有性更新幼苗动态研究. 林业科学 42, 62 -63 .10.3321/j.issn:1001-7488.2006.05.011 URL [30] 张树斌 , 张教林 , 曹坤芳 (2016 ). 季节性干旱对白皮乌口树(Arenna depauperata Hutchins)水分状况、叶片光谱特征和荧光参数的影响. 植物科学学报 34, 117 -126 .URL [31] 张有福 , 陈春艳 , 陈拓 , 张满效 , 刘光琇 , 安黎哲 (2010 ). 2种圆柏属植物叶绿素荧光对季节变化的响应特征. 植物研究 30, 289 -293 .URL [32] 赵顺 , 黄秋娴 , 李玉灵 , 李晓莹 , 李雯 , 程达 , 尹坤 (2014 ). 遮阴处理对臭柏幼苗光合特性的影响. 生态学报 34, 1994 -2002 .10.5846/stxb201306101606 URL [33] 周锋利 , 宋西德 , 齐高强 , 张永 (2005 ). 臭柏抗旱生理特性研究. 西南林学院学报 25, 1 -4 .10.3969/j.issn.2095-1914.2005.03.001 URL [34] 朱成刚 , 陈亚宁 , 李卫红 , 付爱红 , 杨玉海 (2011 ). 干旱胁迫对胡杨PSII光化学效率和激能耗散的影响. 植物学报 46, 413 -424 .10.3724/SP.J.1259.2011.00413 URL [35] 邹长明 , 王允青 , 曹卫东 , 刘英 , 张晓红 , 唐杉 (2015 ). 不同品种小豆光合作用和生长发育对弱光的响应. 应用生态学报 26, 3687 -3692 .URL [36] Ishii Y Sakamoto K Yamanaka N Wang L Yoshikawa K 2006 ). Light acclimation of needle pigment composition in Sabina vulgaris seedlings under nurse plant canopy. J Arid Environ 403 -415 .10.1016/j.jaridenv.2006.03.004 URL [37] Zhang HF Shi XH Wang LH Zhang GS Zhao LQ 2016 ). Antibacterial effect of waste liquor of essence oil extraction from Sabina vulgaris Ant. in foods. Agric Sci Technol 414 -416 .URL
转基因调控柑橘类胡萝卜素积累的细胞学和代谢研究
1
2012
... 叶黄素各组分含量采用高效液相色谱(high performance liquid chromatography, HPLC)法测定, 色谱检测系统为Waters1525型高效液相色谱仪, 2998二级阵列检测器, 2707自动进样器, Empower色谱管理软件和C30 (Devolosil, Nomura Chemical Co. LTD)色谱柱(流速为每分钟1.0 mL), 检测波长为450 nm.称取上述冰盒中的样品0.2 g放入10 mL离心管中, 用液氮粉碎机将其打成粉末状, 加入2 mL色素提取液(正己烷:丙酮:无水乙醇=2:1:1, 含0.01% BHT即2, 6-二叔丁基-4-甲基苯酚), 用超声仪(SCIENTZ, SB-3200- DTDN) 4°C超声10分钟, 置于离心机(Thermo, Fre- sco 17)中, 在13 684.32×
g 下4°C离心5分钟, 重复3次, 色素提取完全后, 加入1 mL 10% KOH-甲醇液, 12小时后加入500 μL 1%的NaCl溶液, 取上清用离心蒸发仪(EYELA, 日本)悬干, 再用甲基叔丁基醚(Methyl tert-Butyl Ether, MtBE)定容至500 μL, 之后用0.22 μm有机微孔滤膜过滤, 并装入高效液相色谱进样瓶(2 mL)内的250 μL的玻璃内衬管中.根据特征吸收峰以及标准品进行信号峰鉴定(
曹洪波, 2012 , 2015).环氧玉米黄质(A)、双环氧紫黄质(V)和玉米黄质(Z)标准品购自CaroteNature公司(Lupsingen, Swit- zerland). ...
草坪草耐荫性研究进展
1
2006
... 77%透光区的臭柏实生苗可通过减少Chlb含量, 升高Chla/Chlb值来避免光能过剩, 这也是耐阳植物适应强光环境的正常响应(
陈煜等, 2006 ;
姜武等, 2007 ;
邹长明等, 2015 ).然而, 4月干旱条件下77%透光区的Chla/Chlb与25%和10%透光区相比无显著差异, 但在湿润区显著大于25%和10%透光区(
图2 ), 说明干旱条件下77%透光区的叶绿素循环光抑制防御机制不及湿润区活跃.如果干旱会加重光抑制的程度, 湿润区都已存在叶绿素循环抵御光抑制的现象, 那么为何干旱区却反而未出现此现象?这可能是由于持续的强光和干旱双重胁迫导致了严重的光抑制, 进而产生了光破坏, 使臭柏幼苗无法启动叶绿素循环.光破坏发生在PSII的反应中心, 直接造成了反应中心色素(进行光能传递和转化且具有光化学活性的特殊态Chla)的大量损失, 使得Chla的含量降低, 从而使Chla/Chlb降低, 结果无法完成叶绿素循环的运作(
潘瑞炽, 2004 ).当然, 臭柏幼苗会通过其它途径来应对严重的光抑制, 本研究中, 干旱条件下77%透光区4月的(A+V)/(A+V+Z)显著高于湿润区(
图3 ), 说明臭柏利用较高的叶黄素循环转化率来加强叶黄素循环的运转, 以消耗过剩的光能.已有研究表明, 叶绿素和类胡萝卜素的光能吸收有一定的互补性(司继播等, 2007), 而且植物可能形成一种稳定的叶绿素-叶黄素-蛋白淬灭复合物来耗散过剩的光能(
李玉灵等, 2009 ).由此可见, 臭柏幼苗应对光抑制的色素机制也是协调统一的.77%透光区臭柏幼苗的光抑制响应机制灵敏、快速且高效, 并且光抑制防御能力较强, 叶绿素循环与叶黄素循环机制同时发生作用, 可达到光能的最佳分配. ...
植物光合机构对光环境的适应机制
1
2016
... 常绿植物叶片的光合作用和蒸腾作用等生理代谢均受到环境因子变化的影响, 如季节变化引起的强光、干旱和低温胁迫(
张有福等, 2010 ).强光胁迫下, 光合天线色素对太阳光能的过量吸收会造成光抑制, 即植物光合作用受到抑制, 可引起光系统II (PSII)反应中心的不可逆破坏, 即光破坏, 进而影响植物的光合生理代谢, 导致植物生长缓慢(
许大全等, 1992 ;
许大全, 2003 ).虽然强光是植物产生光抑制的直接原因, 但干旱、盐渍和低温等叠加胁迫会加剧植物的光抑制, 所以自然界植物的光抑制通常是多种胁迫共同作用的结果.为了物种的延续, 植物在长期的适应和进化过程中, 形成了减轻和抵御光抑制伤害的多种机制.例如, 叶片和叶绿体的避光运动减少了光能的捕获, 增强了光合作用的光能利用效率; 捕光天线色素的转换增加了非光化学耗散及促进了依赖叶黄素的循环等(
许大全等, 1992 ;
高杰等, 2015 ;
代欣等, 2016 ;
付忠等, 2016 ).其中, 叶绿素和叶黄素循环是植物应对光抑制的主要途径之一. ...
遮光性套袋对黄肉桃类胡萝卜素合成及相关基因表达的影响
2015
毛乌素沙地东南缘沉积物光释光测年与环境变迁研究
1
2015
... 臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 ).天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区.在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木.它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 ).毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大.故臭柏极易受到强光照、干旱和低温的单一或多重胁迫.臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 ).毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱. 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
不同光照强度下谢君魔芋的光合作用及能量分配特征
1
2016
... 常绿植物叶片的光合作用和蒸腾作用等生理代谢均受到环境因子变化的影响, 如季节变化引起的强光、干旱和低温胁迫(
张有福等, 2010 ).强光胁迫下, 光合天线色素对太阳光能的过量吸收会造成光抑制, 即植物光合作用受到抑制, 可引起光系统II (PSII)反应中心的不可逆破坏, 即光破坏, 进而影响植物的光合生理代谢, 导致植物生长缓慢(
许大全等, 1992 ;
许大全, 2003 ).虽然强光是植物产生光抑制的直接原因, 但干旱、盐渍和低温等叠加胁迫会加剧植物的光抑制, 所以自然界植物的光抑制通常是多种胁迫共同作用的结果.为了物种的延续, 植物在长期的适应和进化过程中, 形成了减轻和抵御光抑制伤害的多种机制.例如, 叶片和叶绿体的避光运动减少了光能的捕获, 增强了光合作用的光能利用效率; 捕光天线色素的转换增加了非光化学耗散及促进了依赖叶黄素的循环等(
许大全等, 1992 ;
高杰等, 2015 ;
代欣等, 2016 ;
付忠等, 2016 ).其中, 叶绿素和叶黄素循环是植物应对光抑制的主要途径之一. ...
干旱胁迫对玉米苗期叶片光系统II性能的影响
1
2015
... 常绿植物叶片的光合作用和蒸腾作用等生理代谢均受到环境因子变化的影响, 如季节变化引起的强光、干旱和低温胁迫(
张有福等, 2010 ).强光胁迫下, 光合天线色素对太阳光能的过量吸收会造成光抑制, 即植物光合作用受到抑制, 可引起光系统II (PSII)反应中心的不可逆破坏, 即光破坏, 进而影响植物的光合生理代谢, 导致植物生长缓慢(
许大全等, 1992 ;
许大全, 2003 ).虽然强光是植物产生光抑制的直接原因, 但干旱、盐渍和低温等叠加胁迫会加剧植物的光抑制, 所以自然界植物的光抑制通常是多种胁迫共同作用的结果.为了物种的延续, 植物在长期的适应和进化过程中, 形成了减轻和抵御光抑制伤害的多种机制.例如, 叶片和叶绿体的避光运动减少了光能的捕获, 增强了光合作用的光能利用效率; 捕光天线色素的转换增加了非光化学耗散及促进了依赖叶黄素的循环等(
许大全等, 1992 ;
高杰等, 2015 ;
代欣等, 2016 ;
付忠等, 2016 ).其中, 叶绿素和叶黄素循环是植物应对光抑制的主要途径之一. ...
低叶绿素b对水稻光合机构及其热稳定性的影响
1
2007
... 绿色植物的光合色素主要由叶绿素、类胡萝卜素和藻胆素三大类色素组成.叶绿素主要由叶绿素a (Chla)和叶绿素b (Chlb)组成.少部分的Chla具有转化光能的作用, 大部分Chla和全部Chlb的作用是吸收与传递光能, 是光系统天线复合体的重要组成成分.由叶绿素酸酯a合成Chlb和由Chlb到Chla的还原过程可以看作一个循环, 即叶绿素循环(
周锋利等, 2005 ;
郭春爱等, 2006 ;
郭春爱, 2007 ;
李玉灵等, 2009 ).当光能过剩时, 植物可适时减少Chlb的含量, 进而减少光能的吸收量; 当光照不足时, 植物可增加Chlb的含量, 从而捕捉更多的光能, 满足植物生长的需要.叶绿素两大组分间的比值(Chla/Chlb)与总量(Chla+Chlb)在一定程度上表示了整个光系统对光量子的捕获和利用状况, 也反映了植物处于特定环境下承受潜在光胁迫的程度.因此, 叶绿素循环不仅是植物对不同光环境做出的即时响应, 也是植物光抑制的防御机制之一.叶黄素类则是胡萝卜素的氧化衍生物, 是类胡萝卜素中含有氧的一组多萜类的总称, 它包括双环氧紫黄质(violaxanthin, V)、环氧玉米黄质(antheraxanthin, A)和玉米黄质(zeaxanthin, Z)等.其中, V、A和Z三者间可以相互转化, 称为叶黄素循环,该循环通过热耗散防御光破坏.当出现过剩光能时, V在紫黄质脱环氧化酶(VDE)的作用下, 经A转化成Z, Z在植物叶黄素循环耗散光能的过程中起关键作用, 可以直接猝灭激发态叶绿素(或改变类囊体膜的流动性)及促进PSII的捕光复合物(LHCII)聚集来增加非辐射能量的耗散, 所以A和Z也被称为热散逸色素(
王强等, 2003 ).当光能不再过剩时, 转化方向相反, Z在玉米黄质环氧化酶(ZE)的催化作用下经A形成V, Z减少而V增加.(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)比值越高, 意味着叶黄素含量及其中的热散逸色素含量越多或叶绿素含量越少.所以叶黄素循环也是植物对不同光环境做出的即时响应, 与叶绿素循环一样, 也是植物光抑制的防御机制之一.植物的叶绿素荧光主要由PSII反应中心产生, 与光合作用紧密相关, 能够反映光合生理的信息, 特别是胁迫环境下完整植株光合功能的变化(
师生波等, 2014 ).其中, 荧光参数
F v /
F m 指完整叶片中PSII的原初光能转化效率, 是研究光合生理的重要参数, 它能够反映出PSII利用光能的能力(
查倩等, 2016 ;
孙鲁龙等, 2016 ).在非逆境条件下, 不同物种或生态型的叶片,
F v /
F m 均在0.75- 0.85间, 并且不受生长条件的影响, 但在光抑制下,
F v /
F m 值明显降低(
许大全等, 1992 ).故
F v /
F m 是表征光抑制程度的一个重要指标, 该比值越低说明发生光抑制的程度越高(司继播等, 2007).综上所述, 叶绿素循环、叶黄素循环和叶绿素荧光参数变化都是植物对光抑制的响应机制, 但不同植物在不同胁迫条件下的响应策略不同. ...
叶绿素b缺失与植物的光合作用
2
2006
... 绿色植物的光合色素主要由叶绿素、类胡萝卜素和藻胆素三大类色素组成.叶绿素主要由叶绿素a (Chla)和叶绿素b (Chlb)组成.少部分的Chla具有转化光能的作用, 大部分Chla和全部Chlb的作用是吸收与传递光能, 是光系统天线复合体的重要组成成分.由叶绿素酸酯a合成Chlb和由Chlb到Chla的还原过程可以看作一个循环, 即叶绿素循环(
周锋利等, 2005 ;
郭春爱等, 2006 ;
郭春爱, 2007 ;
李玉灵等, 2009 ).当光能过剩时, 植物可适时减少Chlb的含量, 进而减少光能的吸收量; 当光照不足时, 植物可增加Chlb的含量, 从而捕捉更多的光能, 满足植物生长的需要.叶绿素两大组分间的比值(Chla/Chlb)与总量(Chla+Chlb)在一定程度上表示了整个光系统对光量子的捕获和利用状况, 也反映了植物处于特定环境下承受潜在光胁迫的程度.因此, 叶绿素循环不仅是植物对不同光环境做出的即时响应, 也是植物光抑制的防御机制之一.叶黄素类则是胡萝卜素的氧化衍生物, 是类胡萝卜素中含有氧的一组多萜类的总称, 它包括双环氧紫黄质(violaxanthin, V)、环氧玉米黄质(antheraxanthin, A)和玉米黄质(zeaxanthin, Z)等.其中, V、A和Z三者间可以相互转化, 称为叶黄素循环,该循环通过热耗散防御光破坏.当出现过剩光能时, V在紫黄质脱环氧化酶(VDE)的作用下, 经A转化成Z, Z在植物叶黄素循环耗散光能的过程中起关键作用, 可以直接猝灭激发态叶绿素(或改变类囊体膜的流动性)及促进PSII的捕光复合物(LHCII)聚集来增加非辐射能量的耗散, 所以A和Z也被称为热散逸色素(
王强等, 2003 ).当光能不再过剩时, 转化方向相反, Z在玉米黄质环氧化酶(ZE)的催化作用下经A形成V, Z减少而V增加.(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)比值越高, 意味着叶黄素含量及其中的热散逸色素含量越多或叶绿素含量越少.所以叶黄素循环也是植物对不同光环境做出的即时响应, 与叶绿素循环一样, 也是植物光抑制的防御机制之一.植物的叶绿素荧光主要由PSII反应中心产生, 与光合作用紧密相关, 能够反映光合生理的信息, 特别是胁迫环境下完整植株光合功能的变化(
师生波等, 2014 ).其中, 荧光参数
F v /
F m 指完整叶片中PSII的原初光能转化效率, 是研究光合生理的重要参数, 它能够反映出PSII利用光能的能力(
查倩等, 2016 ;
孙鲁龙等, 2016 ).在非逆境条件下, 不同物种或生态型的叶片,
F v /
F m 均在0.75- 0.85间, 并且不受生长条件的影响, 但在光抑制下,
F v /
F m 值明显降低(
许大全等, 1992 ).故
F v /
F m 是表征光抑制程度的一个重要指标, 该比值越低说明发生光抑制的程度越高(司继播等, 2007).综上所述, 叶绿素循环、叶黄素循环和叶绿素荧光参数变化都是植物对光抑制的响应机制, 但不同植物在不同胁迫条件下的响应策略不同. ...
... 光抑制防御机制对植物的生存与正常生长发育至关重要.叶绿素循环抵御光抑制的机理是在光能过剩时, 天线色素Chlb转化成特殊激活状态的Chla, 以便将更多的光能转化成化学能, 固定更多的CO
2 , 生产更多的有机物.Chlb是LHCII的重要组成部分, 可调控光合机构天线的大小并维持LHCII的稳定性.强光下LHCII主要通过减少Chlb来降低吸收光能并抵御光胁迫(娄世庆等, 1995; 陈辉等, 2004;
郭春爱等, 2006 ).同时, 叶绿素循环可能是一种不引起光合机构破坏的光抑制防御机制(
许大全, 2003 ).光抑制下, 参与光化学反应的Chla比收集光能的Chlb更重要, 因而Chla/Chlb是植物抵御光抑制的重要指标之一, Chla/Chlb升高有利于充分利用光能.光抑制最直接的表现为光能过剩, 过剩光能的耗散在调节光能吸收和电子传递上起关键作用, 并且在防止光合器官破坏上不可或缺(
王强等, 2003 ).据统计, 在光照和水分充足时, 植物吸收的光有近50%以热耗散的形式消耗掉.例如, 在干旱和低温胁迫下, 热耗散掉的能量可达吸收光能总量的70%-90% (Flexas et al., 2002).可见被称为“安全阀”的热耗散机制可以高效地保护植物在逆境胁迫下光合机构免受光抑制的破坏, 是光抑制防御的中心和基石, 多年来备受关注(
许大全等, 1992 ).而叶黄素循环是热耗散途径的主要形式之一, 在植物的抗光抑制中非常重要(Niyogi, 1999; ller et al., 2001). ...
遮阴处理对铁尾矿基质臭柏实生苗快速叶绿素荧光特性的影响
1
2015
... 臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 ).天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区.在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木.它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 ).毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大.故臭柏极易受到强光照、干旱和低温的单一或多重胁迫.臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 ).毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱. 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
祁连圆柏和圆柏色素含量及其花青苷合成酶活性的季节性变化
2010
园艺作物光合性状种质差异及遗传表现研究进展
1
2007
... 77%透光区的臭柏实生苗可通过减少Chlb含量, 升高Chla/Chlb值来避免光能过剩, 这也是耐阳植物适应强光环境的正常响应(
陈煜等, 2006 ;
姜武等, 2007 ;
邹长明等, 2015 ).然而, 4月干旱条件下77%透光区的Chla/Chlb与25%和10%透光区相比无显著差异, 但在湿润区显著大于25%和10%透光区(
图2 ), 说明干旱条件下77%透光区的叶绿素循环光抑制防御机制不及湿润区活跃.如果干旱会加重光抑制的程度, 湿润区都已存在叶绿素循环抵御光抑制的现象, 那么为何干旱区却反而未出现此现象?这可能是由于持续的强光和干旱双重胁迫导致了严重的光抑制, 进而产生了光破坏, 使臭柏幼苗无法启动叶绿素循环.光破坏发生在PSII的反应中心, 直接造成了反应中心色素(进行光能传递和转化且具有光化学活性的特殊态Chla)的大量损失, 使得Chla的含量降低, 从而使Chla/Chlb降低, 结果无法完成叶绿素循环的运作(
潘瑞炽, 2004 ).当然, 臭柏幼苗会通过其它途径来应对严重的光抑制, 本研究中, 干旱条件下77%透光区4月的(A+V)/(A+V+Z)显著高于湿润区(
图3 ), 说明臭柏利用较高的叶黄素循环转化率来加强叶黄素循环的运转, 以消耗过剩的光能.已有研究表明, 叶绿素和类胡萝卜素的光能吸收有一定的互补性(司继播等, 2007), 而且植物可能形成一种稳定的叶绿素-叶黄素-蛋白淬灭复合物来耗散过剩的光能(
李玉灵等, 2009 ).由此可见, 臭柏幼苗应对光抑制的色素机制也是协调统一的.77%透光区臭柏幼苗的光抑制响应机制灵敏、快速且高效, 并且光抑制防御能力较强, 叶绿素循环与叶黄素循环机制同时发生作用, 可达到光能的最佳分配. ...
HPLC测定不同产地新疆圆柏中的槲皮苷
1
2015
... 臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 ).天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区.在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木.它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 ).毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大.故臭柏极易受到强光照、干旱和低温的单一或多重胁迫.臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 ).毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱. 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
水分胁迫下臭柏(
Sabina vulgaris Ant.)光合特性和色素组成的季节变化
4
2009
... 绿色植物的光合色素主要由叶绿素、类胡萝卜素和藻胆素三大类色素组成.叶绿素主要由叶绿素a (Chla)和叶绿素b (Chlb)组成.少部分的Chla具有转化光能的作用, 大部分Chla和全部Chlb的作用是吸收与传递光能, 是光系统天线复合体的重要组成成分.由叶绿素酸酯a合成Chlb和由Chlb到Chla的还原过程可以看作一个循环, 即叶绿素循环(
周锋利等, 2005 ;
郭春爱等, 2006 ;
郭春爱, 2007 ;
李玉灵等, 2009 ).当光能过剩时, 植物可适时减少Chlb的含量, 进而减少光能的吸收量; 当光照不足时, 植物可增加Chlb的含量, 从而捕捉更多的光能, 满足植物生长的需要.叶绿素两大组分间的比值(Chla/Chlb)与总量(Chla+Chlb)在一定程度上表示了整个光系统对光量子的捕获和利用状况, 也反映了植物处于特定环境下承受潜在光胁迫的程度.因此, 叶绿素循环不仅是植物对不同光环境做出的即时响应, 也是植物光抑制的防御机制之一.叶黄素类则是胡萝卜素的氧化衍生物, 是类胡萝卜素中含有氧的一组多萜类的总称, 它包括双环氧紫黄质(violaxanthin, V)、环氧玉米黄质(antheraxanthin, A)和玉米黄质(zeaxanthin, Z)等.其中, V、A和Z三者间可以相互转化, 称为叶黄素循环,该循环通过热耗散防御光破坏.当出现过剩光能时, V在紫黄质脱环氧化酶(VDE)的作用下, 经A转化成Z, Z在植物叶黄素循环耗散光能的过程中起关键作用, 可以直接猝灭激发态叶绿素(或改变类囊体膜的流动性)及促进PSII的捕光复合物(LHCII)聚集来增加非辐射能量的耗散, 所以A和Z也被称为热散逸色素(
王强等, 2003 ).当光能不再过剩时, 转化方向相反, Z在玉米黄质环氧化酶(ZE)的催化作用下经A形成V, Z减少而V增加.(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)比值越高, 意味着叶黄素含量及其中的热散逸色素含量越多或叶绿素含量越少.所以叶黄素循环也是植物对不同光环境做出的即时响应, 与叶绿素循环一样, 也是植物光抑制的防御机制之一.植物的叶绿素荧光主要由PSII反应中心产生, 与光合作用紧密相关, 能够反映光合生理的信息, 特别是胁迫环境下完整植株光合功能的变化(
师生波等, 2014 ).其中, 荧光参数
F v /
F m 指完整叶片中PSII的原初光能转化效率, 是研究光合生理的重要参数, 它能够反映出PSII利用光能的能力(
查倩等, 2016 ;
孙鲁龙等, 2016 ).在非逆境条件下, 不同物种或生态型的叶片,
F v /
F m 均在0.75- 0.85间, 并且不受生长条件的影响, 但在光抑制下,
F v /
F m 值明显降低(
许大全等, 1992 ).故
F v /
F m 是表征光抑制程度的一个重要指标, 该比值越低说明发生光抑制的程度越高(司继播等, 2007).综上所述, 叶绿素循环、叶黄素循环和叶绿素荧光参数变化都是植物对光抑制的响应机制, 但不同植物在不同胁迫条件下的响应策略不同. ...
... 臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 ).天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区.在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木.它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 ).毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大.故臭柏极易受到强光照、干旱和低温的单一或多重胁迫.臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 ).毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱. 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
... 77%透光区的臭柏实生苗可通过减少Chlb含量, 升高Chla/Chlb值来避免光能过剩, 这也是耐阳植物适应强光环境的正常响应(
陈煜等, 2006 ;
姜武等, 2007 ;
邹长明等, 2015 ).然而, 4月干旱条件下77%透光区的Chla/Chlb与25%和10%透光区相比无显著差异, 但在湿润区显著大于25%和10%透光区(
图2 ), 说明干旱条件下77%透光区的叶绿素循环光抑制防御机制不及湿润区活跃.如果干旱会加重光抑制的程度, 湿润区都已存在叶绿素循环抵御光抑制的现象, 那么为何干旱区却反而未出现此现象?这可能是由于持续的强光和干旱双重胁迫导致了严重的光抑制, 进而产生了光破坏, 使臭柏幼苗无法启动叶绿素循环.光破坏发生在PSII的反应中心, 直接造成了反应中心色素(进行光能传递和转化且具有光化学活性的特殊态Chla)的大量损失, 使得Chla的含量降低, 从而使Chla/Chlb降低, 结果无法完成叶绿素循环的运作(
潘瑞炽, 2004 ).当然, 臭柏幼苗会通过其它途径来应对严重的光抑制, 本研究中, 干旱条件下77%透光区4月的(A+V)/(A+V+Z)显著高于湿润区(
图3 ), 说明臭柏利用较高的叶黄素循环转化率来加强叶黄素循环的运转, 以消耗过剩的光能.已有研究表明, 叶绿素和类胡萝卜素的光能吸收有一定的互补性(司继播等, 2007), 而且植物可能形成一种稳定的叶绿素-叶黄素-蛋白淬灭复合物来耗散过剩的光能(
李玉灵等, 2009 ).由此可见, 臭柏幼苗应对光抑制的色素机制也是协调统一的.77%透光区臭柏幼苗的光抑制响应机制灵敏、快速且高效, 并且光抑制防御能力较强, 叶绿素循环与叶黄素循环机制同时发生作用, 可达到光能的最佳分配. ...
... 臭柏在干旱环境中经过长期的自然选择和协同进化, 从生理到形态上形成了一系列微妙的耐旱、耐低温和耐强光等逆境生理生态适应对策(
李玉灵等, 2009 ).总体而言, 77%透光区无疑最易受到光抑制, 且强光是引起光抑制的主要原因.但77%透光区一方面可通过减少Chlb含量, 升高Chla/Chlb值来避免光能过剩; 另一方面可通过增加A+V+Z和A+Z含量、提高(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)值, 耗散过剩的光能, 避免光破坏.10%透光区的臭柏幼苗则与77%透光区相反, 未受到光抑制, 其主要面临的是光照不足问题.10%透光区主要通过增加Chlb含量, 降低Chla/Chlb值, 捕捉更多的光能, 不启动或很少启动光抑制防御机制.从77%和10%透光区对不同光照强度的响应策略可以看出, 臭柏集减少光能吸收和捕捉光能于一身, 臭柏幼苗叶绿素和叶黄素循环对不同光照环境的响应具有协调互补的灵活性.25%透光区的光资源可能既非过剩也无不足, 在低温和干旱双重胁迫下不仅叶绿素含量降低, 而且Chla/Chlb值也显著降低; 所以对水分和低温胁迫的影响做出了明显的响应, 叶绿素和叶黄素循环机制随着水分条件变化而迅速改变.可见, 臭柏叶绿素循环和叶黄素循环不仅对光照有瞬时响应, 而且对温度和水分也能做出及时响应.这就解释了为何毛乌素沙地的臭柏实生苗能逐渐摆脱乌柳的庇护, 完全暴露在强光下, 完成天然更新; 同时也体现出了物种的适者生存.该研究对臭柏种质资源保护和人工造林具重要意义. ...
1
2004
... 77%透光区的臭柏实生苗可通过减少Chlb含量, 升高Chla/Chlb值来避免光能过剩, 这也是耐阳植物适应强光环境的正常响应(
陈煜等, 2006 ;
姜武等, 2007 ;
邹长明等, 2015 ).然而, 4月干旱条件下77%透光区的Chla/Chlb与25%和10%透光区相比无显著差异, 但在湿润区显著大于25%和10%透光区(
图2 ), 说明干旱条件下77%透光区的叶绿素循环光抑制防御机制不及湿润区活跃.如果干旱会加重光抑制的程度, 湿润区都已存在叶绿素循环抵御光抑制的现象, 那么为何干旱区却反而未出现此现象?这可能是由于持续的强光和干旱双重胁迫导致了严重的光抑制, 进而产生了光破坏, 使臭柏幼苗无法启动叶绿素循环.光破坏发生在PSII的反应中心, 直接造成了反应中心色素(进行光能传递和转化且具有光化学活性的特殊态Chla)的大量损失, 使得Chla的含量降低, 从而使Chla/Chlb降低, 结果无法完成叶绿素循环的运作(
潘瑞炽, 2004 ).当然, 臭柏幼苗会通过其它途径来应对严重的光抑制, 本研究中, 干旱条件下77%透光区4月的(A+V)/(A+V+Z)显著高于湿润区(
图3 ), 说明臭柏利用较高的叶黄素循环转化率来加强叶黄素循环的运转, 以消耗过剩的光能.已有研究表明, 叶绿素和类胡萝卜素的光能吸收有一定的互补性(司继播等, 2007), 而且植物可能形成一种稳定的叶绿素-叶黄素-蛋白淬灭复合物来耗散过剩的光能(
李玉灵等, 2009 ).由此可见, 臭柏幼苗应对光抑制的色素机制也是协调统一的.77%透光区臭柏幼苗的光抑制响应机制灵敏、快速且高效, 并且光抑制防御能力较强, 叶绿素循环与叶黄素循环机制同时发生作用, 可达到光能的最佳分配. ...
脱水胁迫和光合日变化对梭梭和白梭梭叶绿素荧光参数的影响
2
2015
... 光照、干旱和低温等引起的光抑制现象在自然界植物中普遍存在,
F v /
F m 是表征光抑制程度的良好指标和探针.生长状态良好的植物
F v /
F m 值介于0.75-0.85之间, 低于0.75则表明植物生长不良(
许大全等, 1992 ;
沈亮等, 2015 ).由
图1 可知, 2个水分条件下77%和25%透光区臭柏幼苗的
F v /
F m 均在低温的2月达到全年最低值, 且都在0.75以下; 而10%透光区的
F v /
F m 值在0.75以上.可见光照越强,
F v /
F m 的降低幅度越大, 且77%透光区的
F v /
F m 值均显著低于另外两个透光区.随着气候转暖, 77%和25%透光区的
F v /
F m 值迅速上升, 5月以后趋于稳定, 各透光区间均无显著差异, 且均保持在0.75-0.85之间.说明77%和25%透光区在2月的低温期不同程度地受到了光抑制, 77%透光区受到的光抑制程度最大.随着气温升高, 臭柏幼苗的光能利用能力逐渐增强且趋于稳定, 表明低温胁迫是引起77%和25%透光区出现光抑制现象的主要因素.与77%和25%透光区相比, 10%透光区的
F v /
F m 值随着季节变化的波动幅度较小, 全年维持在0.75以上, 说明其受到光抑制的影响较小.水分条件对3个光照区
F v /
F m 的影响在2-4月间无明显差异(
表1 ); 进入5月, 25%和10%透光区则表现出一定的差异性; 而77%透光区仅在9月出现差异(
表1 ).综上所述, 2个水分条件下低温均导致了77%以及25%透光区 出现光抑制, 且77%透光区的光抑制现象更明显, 10%透光区的光抑制影响较小.水分因子在5月以后作用明显, 特别是对25%和10%透光区的影响更突出. ...
... 如前所述,
F v /
F m 是表征光抑制程度的良好指标和探针.生长状态良好的植物的
F v /
F m 值介于0.75-0.85之间, 低于0.75则生长不良(
许大全等, 1992 ;
沈亮等, 2015 ).本研究结果表明, 冬季低温导致不同透光区臭柏幼苗的
F v /
F m 值差异明显, 77%和25%透光区的
F v /
F m 值降低至0.75以下, 且77%透光区下降幅度最大, 光抑制现象更明显, 10%透光区的
F v /
F m 值全年都介于0.75-0.85之间, 未受到明显的光抑制.但随着季节转换气候变暖, 77%和25%透光区的
F v /
F m 值迅速上升, 5月以后趋于稳定, 且3个透光区间无显著差异, 均保持在0.75-0.85间(
图1 ); 77%和25%透光区的光合作用已从低迷的光抑制状态进入正常的光合旺盛阶段, 那么77%和25%透光区必定启动了光抑制防御机制.朱成刚等(2011)在研究干旱胁迫下胡杨(
Popu- lus euphratica ) PSII光化学效率和激能耗散时也发现, 其
F v /
F m 值在生长季并未降低, 这可能是干旱地区耐旱植物适应干旱强光环境的共性之一. ...
高山植物唐古特山莨菪和唐古特大黄对强太阳辐射光能的利用和耗散特性
1
2007
... 臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 ).天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区.在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木.它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 ).毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大.故臭柏极易受到强光照、干旱和低温的单一或多重胁迫.臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 ).毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱. 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
青藏高原春小麦叶片光合作用的光抑制及PSII反应中心光化学效率的恢复分析
1
2014
... 绿色植物的光合色素主要由叶绿素、类胡萝卜素和藻胆素三大类色素组成.叶绿素主要由叶绿素a (Chla)和叶绿素b (Chlb)组成.少部分的Chla具有转化光能的作用, 大部分Chla和全部Chlb的作用是吸收与传递光能, 是光系统天线复合体的重要组成成分.由叶绿素酸酯a合成Chlb和由Chlb到Chla的还原过程可以看作一个循环, 即叶绿素循环(
周锋利等, 2005 ;
郭春爱等, 2006 ;
郭春爱, 2007 ;
李玉灵等, 2009 ).当光能过剩时, 植物可适时减少Chlb的含量, 进而减少光能的吸收量; 当光照不足时, 植物可增加Chlb的含量, 从而捕捉更多的光能, 满足植物生长的需要.叶绿素两大组分间的比值(Chla/Chlb)与总量(Chla+Chlb)在一定程度上表示了整个光系统对光量子的捕获和利用状况, 也反映了植物处于特定环境下承受潜在光胁迫的程度.因此, 叶绿素循环不仅是植物对不同光环境做出的即时响应, 也是植物光抑制的防御机制之一.叶黄素类则是胡萝卜素的氧化衍生物, 是类胡萝卜素中含有氧的一组多萜类的总称, 它包括双环氧紫黄质(violaxanthin, V)、环氧玉米黄质(antheraxanthin, A)和玉米黄质(zeaxanthin, Z)等.其中, V、A和Z三者间可以相互转化, 称为叶黄素循环,该循环通过热耗散防御光破坏.当出现过剩光能时, V在紫黄质脱环氧化酶(VDE)的作用下, 经A转化成Z, Z在植物叶黄素循环耗散光能的过程中起关键作用, 可以直接猝灭激发态叶绿素(或改变类囊体膜的流动性)及促进PSII的捕光复合物(LHCII)聚集来增加非辐射能量的耗散, 所以A和Z也被称为热散逸色素(
王强等, 2003 ).当光能不再过剩时, 转化方向相反, Z在玉米黄质环氧化酶(ZE)的催化作用下经A形成V, Z减少而V增加.(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)比值越高, 意味着叶黄素含量及其中的热散逸色素含量越多或叶绿素含量越少.所以叶黄素循环也是植物对不同光环境做出的即时响应, 与叶绿素循环一样, 也是植物光抑制的防御机制之一.植物的叶绿素荧光主要由PSII反应中心产生, 与光合作用紧密相关, 能够反映光合生理的信息, 特别是胁迫环境下完整植株光合功能的变化(
师生波等, 2014 ).其中, 荧光参数
F v /
F m 指完整叶片中PSII的原初光能转化效率, 是研究光合生理的重要参数, 它能够反映出PSII利用光能的能力(
查倩等, 2016 ;
孙鲁龙等, 2016 ).在非逆境条件下, 不同物种或生态型的叶片,
F v /
F m 均在0.75- 0.85间, 并且不受生长条件的影响, 但在光抑制下,
F v /
F m 值明显降低(
许大全等, 1992 ).故
F v /
F m 是表征光抑制程度的一个重要指标, 该比值越低说明发生光抑制的程度越高(司继播等, 2007).综上所述, 叶绿素循环、叶黄素循环和叶绿素荧光参数变化都是植物对光抑制的响应机制, 但不同植物在不同胁迫条件下的响应策略不同. ...
遮阴胁迫对不同耐阴性玉米叶绿素含量的影响
1
2013
... 10%透光区的臭柏幼苗因光照强度最弱, 叶绿素含量明显高于其它两个遮光处理区, 而不易受到光抑制的影响.在不同水分条件下, 10%透光区的Chla+ Chlb随着季节的变化均显著高于25%透光区, Chla/ Chlb均低于25%透光区, 同时叶黄素组分含量与25%透光区无显著差异(
图2 ,
图3 ).即使在湿润条件下, 光照不足(10%透光区)的影响也很明显, 天线色素的比例与干旱区无明显差异.10%透光区全年都不会发生光能过剩, 自然也没有必要进行热散逸, 因此叶黄素所占比例最小.叶绿素循环和叶黄素循环均不活跃, 很少启动光抑制防御机制.10%透光区主要通过增加Chlb的含量, 降低Chla/Chlb值, 捕捉更多的光能, 这也是耐荫植物对阴暗环境做出的响应(
杨渺等, 2004 ;
史振声等, 2013 ). ...
不同光强对霜冻后葡萄叶片PSII光化学活性恢复的影响
1
2016
... 绿色植物的光合色素主要由叶绿素、类胡萝卜素和藻胆素三大类色素组成.叶绿素主要由叶绿素a (Chla)和叶绿素b (Chlb)组成.少部分的Chla具有转化光能的作用, 大部分Chla和全部Chlb的作用是吸收与传递光能, 是光系统天线复合体的重要组成成分.由叶绿素酸酯a合成Chlb和由Chlb到Chla的还原过程可以看作一个循环, 即叶绿素循环(
周锋利等, 2005 ;
郭春爱等, 2006 ;
郭春爱, 2007 ;
李玉灵等, 2009 ).当光能过剩时, 植物可适时减少Chlb的含量, 进而减少光能的吸收量; 当光照不足时, 植物可增加Chlb的含量, 从而捕捉更多的光能, 满足植物生长的需要.叶绿素两大组分间的比值(Chla/Chlb)与总量(Chla+Chlb)在一定程度上表示了整个光系统对光量子的捕获和利用状况, 也反映了植物处于特定环境下承受潜在光胁迫的程度.因此, 叶绿素循环不仅是植物对不同光环境做出的即时响应, 也是植物光抑制的防御机制之一.叶黄素类则是胡萝卜素的氧化衍生物, 是类胡萝卜素中含有氧的一组多萜类的总称, 它包括双环氧紫黄质(violaxanthin, V)、环氧玉米黄质(antheraxanthin, A)和玉米黄质(zeaxanthin, Z)等.其中, V、A和Z三者间可以相互转化, 称为叶黄素循环,该循环通过热耗散防御光破坏.当出现过剩光能时, V在紫黄质脱环氧化酶(VDE)的作用下, 经A转化成Z, Z在植物叶黄素循环耗散光能的过程中起关键作用, 可以直接猝灭激发态叶绿素(或改变类囊体膜的流动性)及促进PSII的捕光复合物(LHCII)聚集来增加非辐射能量的耗散, 所以A和Z也被称为热散逸色素(
王强等, 2003 ).当光能不再过剩时, 转化方向相反, Z在玉米黄质环氧化酶(ZE)的催化作用下经A形成V, Z减少而V增加.(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)比值越高, 意味着叶黄素含量及其中的热散逸色素含量越多或叶绿素含量越少.所以叶黄素循环也是植物对不同光环境做出的即时响应, 与叶绿素循环一样, 也是植物光抑制的防御机制之一.植物的叶绿素荧光主要由PSII反应中心产生, 与光合作用紧密相关, 能够反映光合生理的信息, 特别是胁迫环境下完整植株光合功能的变化(
师生波等, 2014 ).其中, 荧光参数
F v /
F m 指完整叶片中PSII的原初光能转化效率, 是研究光合生理的重要参数, 它能够反映出PSII利用光能的能力(
查倩等, 2016 ;
孙鲁龙等, 2016 ).在非逆境条件下, 不同物种或生态型的叶片,
F v /
F m 均在0.75- 0.85间, 并且不受生长条件的影响, 但在光抑制下,
F v /
F m 值明显降低(
许大全等, 1992 ).故
F v /
F m 是表征光抑制程度的一个重要指标, 该比值越低说明发生光抑制的程度越高(司继播等, 2007).综上所述, 叶绿素循环、叶黄素循环和叶绿素荧光参数变化都是植物对光抑制的响应机制, 但不同植物在不同胁迫条件下的响应策略不同. ...
中国天然臭柏群落的分布与生物量特征
1
2014
... 臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 ).天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区.在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木.它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 ).毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大.故臭柏极易受到强光照、干旱和低温的单一或多重胁迫.臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 ).毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱. 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
1
2011
... 臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 ).天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区.在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木.它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 ).毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大.故臭柏极易受到强光照、干旱和低温的单一或多重胁迫.臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 ).毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱. 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
光合作用光抑制的研究进展
2
2003
... 绿色植物的光合色素主要由叶绿素、类胡萝卜素和藻胆素三大类色素组成.叶绿素主要由叶绿素a (Chla)和叶绿素b (Chlb)组成.少部分的Chla具有转化光能的作用, 大部分Chla和全部Chlb的作用是吸收与传递光能, 是光系统天线复合体的重要组成成分.由叶绿素酸酯a合成Chlb和由Chlb到Chla的还原过程可以看作一个循环, 即叶绿素循环(
周锋利等, 2005 ;
郭春爱等, 2006 ;
郭春爱, 2007 ;
李玉灵等, 2009 ).当光能过剩时, 植物可适时减少Chlb的含量, 进而减少光能的吸收量; 当光照不足时, 植物可增加Chlb的含量, 从而捕捉更多的光能, 满足植物生长的需要.叶绿素两大组分间的比值(Chla/Chlb)与总量(Chla+Chlb)在一定程度上表示了整个光系统对光量子的捕获和利用状况, 也反映了植物处于特定环境下承受潜在光胁迫的程度.因此, 叶绿素循环不仅是植物对不同光环境做出的即时响应, 也是植物光抑制的防御机制之一.叶黄素类则是胡萝卜素的氧化衍生物, 是类胡萝卜素中含有氧的一组多萜类的总称, 它包括双环氧紫黄质(violaxanthin, V)、环氧玉米黄质(antheraxanthin, A)和玉米黄质(zeaxanthin, Z)等.其中, V、A和Z三者间可以相互转化, 称为叶黄素循环,该循环通过热耗散防御光破坏.当出现过剩光能时, V在紫黄质脱环氧化酶(VDE)的作用下, 经A转化成Z, Z在植物叶黄素循环耗散光能的过程中起关键作用, 可以直接猝灭激发态叶绿素(或改变类囊体膜的流动性)及促进PSII的捕光复合物(LHCII)聚集来增加非辐射能量的耗散, 所以A和Z也被称为热散逸色素(
王强等, 2003 ).当光能不再过剩时, 转化方向相反, Z在玉米黄质环氧化酶(ZE)的催化作用下经A形成V, Z减少而V增加.(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)比值越高, 意味着叶黄素含量及其中的热散逸色素含量越多或叶绿素含量越少.所以叶黄素循环也是植物对不同光环境做出的即时响应, 与叶绿素循环一样, 也是植物光抑制的防御机制之一.植物的叶绿素荧光主要由PSII反应中心产生, 与光合作用紧密相关, 能够反映光合生理的信息, 特别是胁迫环境下完整植株光合功能的变化(
师生波等, 2014 ).其中, 荧光参数
F v /
F m 指完整叶片中PSII的原初光能转化效率, 是研究光合生理的重要参数, 它能够反映出PSII利用光能的能力(
查倩等, 2016 ;
孙鲁龙等, 2016 ).在非逆境条件下, 不同物种或生态型的叶片,
F v /
F m 均在0.75- 0.85间, 并且不受生长条件的影响, 但在光抑制下,
F v /
F m 值明显降低(
许大全等, 1992 ).故
F v /
F m 是表征光抑制程度的一个重要指标, 该比值越低说明发生光抑制的程度越高(司继播等, 2007).综上所述, 叶绿素循环、叶黄素循环和叶绿素荧光参数变化都是植物对光抑制的响应机制, 但不同植物在不同胁迫条件下的响应策略不同. ...
... 光抑制防御机制对植物的生存与正常生长发育至关重要.叶绿素循环抵御光抑制的机理是在光能过剩时, 天线色素Chlb转化成特殊激活状态的Chla, 以便将更多的光能转化成化学能, 固定更多的CO
2 , 生产更多的有机物.Chlb是LHCII的重要组成部分, 可调控光合机构天线的大小并维持LHCII的稳定性.强光下LHCII主要通过减少Chlb来降低吸收光能并抵御光胁迫(娄世庆等, 1995; 陈辉等, 2004;
郭春爱等, 2006 ).同时, 叶绿素循环可能是一种不引起光合机构破坏的光抑制防御机制(
许大全, 2003 ).光抑制下, 参与光化学反应的Chla比收集光能的Chlb更重要, 因而Chla/Chlb是植物抵御光抑制的重要指标之一, Chla/Chlb升高有利于充分利用光能.光抑制最直接的表现为光能过剩, 过剩光能的耗散在调节光能吸收和电子传递上起关键作用, 并且在防止光合器官破坏上不可或缺(
王强等, 2003 ).据统计, 在光照和水分充足时, 植物吸收的光有近50%以热耗散的形式消耗掉.例如, 在干旱和低温胁迫下, 热耗散掉的能量可达吸收光能总量的70%-90% (Flexas et al., 2002).可见被称为“安全阀”的热耗散机制可以高效地保护植物在逆境胁迫下光合机构免受光抑制的破坏, 是光抑制防御的中心和基石, 多年来备受关注(
许大全等, 1992 ).而叶黄素循环是热耗散途径的主要形式之一, 在植物的抗光抑制中非常重要(Niyogi, 1999; ller et al., 2001). ...
1
2006
... 叶绿素a和叶绿素b含量采用分光光度计法测定.2016年, 选择低温的2月、开始生长的4月和生长旺期的7月三个时期的每月上、中和下旬3天晴朗天气, 每小区随机选取3株, 剪取当年生叶片用锡纸包好后放入装有液氮的冰盒内带回实验室.称取0.1 g装入10 mL离心管中, 加入2 mL无水乙醇, 用数显高数分散均质机(FJ200-SH, 上海)粉碎, 再加入8 mL无水乙醇, 置于离心机(Anke, T.G.L-16-aR)在11 269.44×
g 下4°C离心5分钟, 取3 mL装入玻璃比色皿, 用双光束紫外可见分光光度计(TU-1901, 北京)测定在649和665 nm处的吸光度, 计算各处理的Chla和Chlb含量(
王学奎, 2006 ). ...
植物光胁迫研究中的几个问题
2
2003
... 常绿植物叶片的光合作用和蒸腾作用等生理代谢均受到环境因子变化的影响, 如季节变化引起的强光、干旱和低温胁迫(
张有福等, 2010 ).强光胁迫下, 光合天线色素对太阳光能的过量吸收会造成光抑制, 即植物光合作用受到抑制, 可引起光系统II (PSII)反应中心的不可逆破坏, 即光破坏, 进而影响植物的光合生理代谢, 导致植物生长缓慢(
许大全等, 1992 ;
许大全, 2003 ).虽然强光是植物产生光抑制的直接原因, 但干旱、盐渍和低温等叠加胁迫会加剧植物的光抑制, 所以自然界植物的光抑制通常是多种胁迫共同作用的结果.为了物种的延续, 植物在长期的适应和进化过程中, 形成了减轻和抵御光抑制伤害的多种机制.例如, 叶片和叶绿体的避光运动减少了光能的捕获, 增强了光合作用的光能利用效率; 捕光天线色素的转换增加了非光化学耗散及促进了依赖叶黄素的循环等(
许大全等, 1992 ;
高杰等, 2015 ;
代欣等, 2016 ;
付忠等, 2016 ).其中, 叶绿素和叶黄素循环是植物应对光抑制的主要途径之一. ...
... 光抑制防御机制对植物的生存与正常生长发育至关重要.叶绿素循环抵御光抑制的机理是在光能过剩时, 天线色素Chlb转化成特殊激活状态的Chla, 以便将更多的光能转化成化学能, 固定更多的CO
2 , 生产更多的有机物.Chlb是LHCII的重要组成部分, 可调控光合机构天线的大小并维持LHCII的稳定性.强光下LHCII主要通过减少Chlb来降低吸收光能并抵御光胁迫(娄世庆等, 1995; 陈辉等, 2004;
郭春爱等, 2006 ).同时, 叶绿素循环可能是一种不引起光合机构破坏的光抑制防御机制(
许大全, 2003 ).光抑制下, 参与光化学反应的Chla比收集光能的Chlb更重要, 因而Chla/Chlb是植物抵御光抑制的重要指标之一, Chla/Chlb升高有利于充分利用光能.光抑制最直接的表现为光能过剩, 过剩光能的耗散在调节光能吸收和电子传递上起关键作用, 并且在防止光合器官破坏上不可或缺(
王强等, 2003 ).据统计, 在光照和水分充足时, 植物吸收的光有近50%以热耗散的形式消耗掉.例如, 在干旱和低温胁迫下, 热耗散掉的能量可达吸收光能总量的70%-90% (Flexas et al., 2002).可见被称为“安全阀”的热耗散机制可以高效地保护植物在逆境胁迫下光合机构免受光抑制的破坏, 是光抑制防御的中心和基石, 多年来备受关注(
许大全等, 1992 ).而叶黄素循环是热耗散途径的主要形式之一, 在植物的抗光抑制中非常重要(Niyogi, 1999; ller et al., 2001). ...
植物光合作用的光抑制
6
1992
... 常绿植物叶片的光合作用和蒸腾作用等生理代谢均受到环境因子变化的影响, 如季节变化引起的强光、干旱和低温胁迫(
张有福等, 2010 ).强光胁迫下, 光合天线色素对太阳光能的过量吸收会造成光抑制, 即植物光合作用受到抑制, 可引起光系统II (PSII)反应中心的不可逆破坏, 即光破坏, 进而影响植物的光合生理代谢, 导致植物生长缓慢(
许大全等, 1992 ;
许大全, 2003 ).虽然强光是植物产生光抑制的直接原因, 但干旱、盐渍和低温等叠加胁迫会加剧植物的光抑制, 所以自然界植物的光抑制通常是多种胁迫共同作用的结果.为了物种的延续, 植物在长期的适应和进化过程中, 形成了减轻和抵御光抑制伤害的多种机制.例如, 叶片和叶绿体的避光运动减少了光能的捕获, 增强了光合作用的光能利用效率; 捕光天线色素的转换增加了非光化学耗散及促进了依赖叶黄素的循环等(
许大全等, 1992 ;
高杰等, 2015 ;
代欣等, 2016 ;
付忠等, 2016 ).其中, 叶绿素和叶黄素循环是植物应对光抑制的主要途径之一. ...
... ).虽然强光是植物产生光抑制的直接原因, 但干旱、盐渍和低温等叠加胁迫会加剧植物的光抑制, 所以自然界植物的光抑制通常是多种胁迫共同作用的结果.为了物种的延续, 植物在长期的适应和进化过程中, 形成了减轻和抵御光抑制伤害的多种机制.例如, 叶片和叶绿体的避光运动减少了光能的捕获, 增强了光合作用的光能利用效率; 捕光天线色素的转换增加了非光化学耗散及促进了依赖叶黄素的循环等(
许大全等, 1992 ;
高杰等, 2015 ;
代欣等, 2016 ;
付忠等, 2016 ).其中, 叶绿素和叶黄素循环是植物应对光抑制的主要途径之一. ...
... 绿色植物的光合色素主要由叶绿素、类胡萝卜素和藻胆素三大类色素组成.叶绿素主要由叶绿素a (Chla)和叶绿素b (Chlb)组成.少部分的Chla具有转化光能的作用, 大部分Chla和全部Chlb的作用是吸收与传递光能, 是光系统天线复合体的重要组成成分.由叶绿素酸酯a合成Chlb和由Chlb到Chla的还原过程可以看作一个循环, 即叶绿素循环(
周锋利等, 2005 ;
郭春爱等, 2006 ;
郭春爱, 2007 ;
李玉灵等, 2009 ).当光能过剩时, 植物可适时减少Chlb的含量, 进而减少光能的吸收量; 当光照不足时, 植物可增加Chlb的含量, 从而捕捉更多的光能, 满足植物生长的需要.叶绿素两大组分间的比值(Chla/Chlb)与总量(Chla+Chlb)在一定程度上表示了整个光系统对光量子的捕获和利用状况, 也反映了植物处于特定环境下承受潜在光胁迫的程度.因此, 叶绿素循环不仅是植物对不同光环境做出的即时响应, 也是植物光抑制的防御机制之一.叶黄素类则是胡萝卜素的氧化衍生物, 是类胡萝卜素中含有氧的一组多萜类的总称, 它包括双环氧紫黄质(violaxanthin, V)、环氧玉米黄质(antheraxanthin, A)和玉米黄质(zeaxanthin, Z)等.其中, V、A和Z三者间可以相互转化, 称为叶黄素循环,该循环通过热耗散防御光破坏.当出现过剩光能时, V在紫黄质脱环氧化酶(VDE)的作用下, 经A转化成Z, Z在植物叶黄素循环耗散光能的过程中起关键作用, 可以直接猝灭激发态叶绿素(或改变类囊体膜的流动性)及促进PSII的捕光复合物(LHCII)聚集来增加非辐射能量的耗散, 所以A和Z也被称为热散逸色素(
王强等, 2003 ).当光能不再过剩时, 转化方向相反, Z在玉米黄质环氧化酶(ZE)的催化作用下经A形成V, Z减少而V增加.(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)比值越高, 意味着叶黄素含量及其中的热散逸色素含量越多或叶绿素含量越少.所以叶黄素循环也是植物对不同光环境做出的即时响应, 与叶绿素循环一样, 也是植物光抑制的防御机制之一.植物的叶绿素荧光主要由PSII反应中心产生, 与光合作用紧密相关, 能够反映光合生理的信息, 特别是胁迫环境下完整植株光合功能的变化(
师生波等, 2014 ).其中, 荧光参数
F v /
F m 指完整叶片中PSII的原初光能转化效率, 是研究光合生理的重要参数, 它能够反映出PSII利用光能的能力(
查倩等, 2016 ;
孙鲁龙等, 2016 ).在非逆境条件下, 不同物种或生态型的叶片,
F v /
F m 均在0.75- 0.85间, 并且不受生长条件的影响, 但在光抑制下,
F v /
F m 值明显降低(
许大全等, 1992 ).故
F v /
F m 是表征光抑制程度的一个重要指标, 该比值越低说明发生光抑制的程度越高(司继播等, 2007).综上所述, 叶绿素循环、叶黄素循环和叶绿素荧光参数变化都是植物对光抑制的响应机制, 但不同植物在不同胁迫条件下的响应策略不同. ...
... 光照、干旱和低温等引起的光抑制现象在自然界植物中普遍存在,
F v /
F m 是表征光抑制程度的良好指标和探针.生长状态良好的植物
F v /
F m 值介于0.75-0.85之间, 低于0.75则表明植物生长不良(
许大全等, 1992 ;
沈亮等, 2015 ).由
图1 可知, 2个水分条件下77%和25%透光区臭柏幼苗的
F v /
F m 均在低温的2月达到全年最低值, 且都在0.75以下; 而10%透光区的
F v /
F m 值在0.75以上.可见光照越强,
F v /
F m 的降低幅度越大, 且77%透光区的
F v /
F m 值均显著低于另外两个透光区.随着气候转暖, 77%和25%透光区的
F v /
F m 值迅速上升, 5月以后趋于稳定, 各透光区间均无显著差异, 且均保持在0.75-0.85之间.说明77%和25%透光区在2月的低温期不同程度地受到了光抑制, 77%透光区受到的光抑制程度最大.随着气温升高, 臭柏幼苗的光能利用能力逐渐增强且趋于稳定, 表明低温胁迫是引起77%和25%透光区出现光抑制现象的主要因素.与77%和25%透光区相比, 10%透光区的
F v /
F m 值随着季节变化的波动幅度较小, 全年维持在0.75以上, 说明其受到光抑制的影响较小.水分条件对3个光照区
F v /
F m 的影响在2-4月间无明显差异(
表1 ); 进入5月, 25%和10%透光区则表现出一定的差异性; 而77%透光区仅在9月出现差异(
表1 ).综上所述, 2个水分条件下低温均导致了77%以及25%透光区 出现光抑制, 且77%透光区的光抑制现象更明显, 10%透光区的光抑制影响较小.水分因子在5月以后作用明显, 特别是对25%和10%透光区的影响更突出. ...
... 如前所述,
F v /
F m 是表征光抑制程度的良好指标和探针.生长状态良好的植物的
F v /
F m 值介于0.75-0.85之间, 低于0.75则生长不良(
许大全等, 1992 ;
沈亮等, 2015 ).本研究结果表明, 冬季低温导致不同透光区臭柏幼苗的
F v /
F m 值差异明显, 77%和25%透光区的
F v /
F m 值降低至0.75以下, 且77%透光区下降幅度最大, 光抑制现象更明显, 10%透光区的
F v /
F m 值全年都介于0.75-0.85之间, 未受到明显的光抑制.但随着季节转换气候变暖, 77%和25%透光区的
F v /
F m 值迅速上升, 5月以后趋于稳定, 且3个透光区间无显著差异, 均保持在0.75-0.85间(
图1 ); 77%和25%透光区的光合作用已从低迷的光抑制状态进入正常的光合旺盛阶段, 那么77%和25%透光区必定启动了光抑制防御机制.朱成刚等(2011)在研究干旱胁迫下胡杨(
Popu- lus euphratica ) PSII光化学效率和激能耗散时也发现, 其
F v /
F m 值在生长季并未降低, 这可能是干旱地区耐旱植物适应干旱强光环境的共性之一. ...
... 光抑制防御机制对植物的生存与正常生长发育至关重要.叶绿素循环抵御光抑制的机理是在光能过剩时, 天线色素Chlb转化成特殊激活状态的Chla, 以便将更多的光能转化成化学能, 固定更多的CO
2 , 生产更多的有机物.Chlb是LHCII的重要组成部分, 可调控光合机构天线的大小并维持LHCII的稳定性.强光下LHCII主要通过减少Chlb来降低吸收光能并抵御光胁迫(娄世庆等, 1995; 陈辉等, 2004;
郭春爱等, 2006 ).同时, 叶绿素循环可能是一种不引起光合机构破坏的光抑制防御机制(
许大全, 2003 ).光抑制下, 参与光化学反应的Chla比收集光能的Chlb更重要, 因而Chla/Chlb是植物抵御光抑制的重要指标之一, Chla/Chlb升高有利于充分利用光能.光抑制最直接的表现为光能过剩, 过剩光能的耗散在调节光能吸收和电子传递上起关键作用, 并且在防止光合器官破坏上不可或缺(
王强等, 2003 ).据统计, 在光照和水分充足时, 植物吸收的光有近50%以热耗散的形式消耗掉.例如, 在干旱和低温胁迫下, 热耗散掉的能量可达吸收光能总量的70%-90% (Flexas et al., 2002).可见被称为“安全阀”的热耗散机制可以高效地保护植物在逆境胁迫下光合机构免受光抑制的破坏, 是光抑制防御的中心和基石, 多年来备受关注(
许大全等, 1992 ).而叶黄素循环是热耗散途径的主要形式之一, 在植物的抗光抑制中非常重要(Niyogi, 1999; ller et al., 2001). ...
遮阴胁迫对叶绿素含量的影响
1
2004
... 10%透光区的臭柏幼苗因光照强度最弱, 叶绿素含量明显高于其它两个遮光处理区, 而不易受到光抑制的影响.在不同水分条件下, 10%透光区的Chla+ Chlb随着季节的变化均显著高于25%透光区, Chla/ Chlb均低于25%透光区, 同时叶黄素组分含量与25%透光区无显著差异(
图2 ,
图3 ).即使在湿润条件下, 光照不足(10%透光区)的影响也很明显, 天线色素的比例与干旱区无明显差异.10%透光区全年都不会发生光能过剩, 自然也没有必要进行热散逸, 因此叶黄素所占比例最小.叶绿素循环和叶黄素循环均不活跃, 很少启动光抑制防御机制.10%透光区主要通过增加Chlb的含量, 降低Chla/Chlb值, 捕捉更多的光能, 这也是耐荫植物对阴暗环境做出的响应(
杨渺等, 2004 ;
史振声等, 2013 ). ...
高温胁迫对葡萄幼树叶绿素荧光特性和抗氧化酶活性的影响
1
2016
... 绿色植物的光合色素主要由叶绿素、类胡萝卜素和藻胆素三大类色素组成.叶绿素主要由叶绿素a (Chla)和叶绿素b (Chlb)组成.少部分的Chla具有转化光能的作用, 大部分Chla和全部Chlb的作用是吸收与传递光能, 是光系统天线复合体的重要组成成分.由叶绿素酸酯a合成Chlb和由Chlb到Chla的还原过程可以看作一个循环, 即叶绿素循环(
周锋利等, 2005 ;
郭春爱等, 2006 ;
郭春爱, 2007 ;
李玉灵等, 2009 ).当光能过剩时, 植物可适时减少Chlb的含量, 进而减少光能的吸收量; 当光照不足时, 植物可增加Chlb的含量, 从而捕捉更多的光能, 满足植物生长的需要.叶绿素两大组分间的比值(Chla/Chlb)与总量(Chla+Chlb)在一定程度上表示了整个光系统对光量子的捕获和利用状况, 也反映了植物处于特定环境下承受潜在光胁迫的程度.因此, 叶绿素循环不仅是植物对不同光环境做出的即时响应, 也是植物光抑制的防御机制之一.叶黄素类则是胡萝卜素的氧化衍生物, 是类胡萝卜素中含有氧的一组多萜类的总称, 它包括双环氧紫黄质(violaxanthin, V)、环氧玉米黄质(antheraxanthin, A)和玉米黄质(zeaxanthin, Z)等.其中, V、A和Z三者间可以相互转化, 称为叶黄素循环,该循环通过热耗散防御光破坏.当出现过剩光能时, V在紫黄质脱环氧化酶(VDE)的作用下, 经A转化成Z, Z在植物叶黄素循环耗散光能的过程中起关键作用, 可以直接猝灭激发态叶绿素(或改变类囊体膜的流动性)及促进PSII的捕光复合物(LHCII)聚集来增加非辐射能量的耗散, 所以A和Z也被称为热散逸色素(
王强等, 2003 ).当光能不再过剩时, 转化方向相反, Z在玉米黄质环氧化酶(ZE)的催化作用下经A形成V, Z减少而V增加.(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)比值越高, 意味着叶黄素含量及其中的热散逸色素含量越多或叶绿素含量越少.所以叶黄素循环也是植物对不同光环境做出的即时响应, 与叶绿素循环一样, 也是植物光抑制的防御机制之一.植物的叶绿素荧光主要由PSII反应中心产生, 与光合作用紧密相关, 能够反映光合生理的信息, 特别是胁迫环境下完整植株光合功能的变化(
师生波等, 2014 ).其中, 荧光参数
F v /
F m 指完整叶片中PSII的原初光能转化效率, 是研究光合生理的重要参数, 它能够反映出PSII利用光能的能力(
查倩等, 2016 ;
孙鲁龙等, 2016 ).在非逆境条件下, 不同物种或生态型的叶片,
F v /
F m 均在0.75- 0.85间, 并且不受生长条件的影响, 但在光抑制下,
F v /
F m 值明显降低(
许大全等, 1992 ).故
F v /
F m 是表征光抑制程度的一个重要指标, 该比值越低说明发生光抑制的程度越高(司继播等, 2007).综上所述, 叶绿素循环、叶黄素循环和叶绿素荧光参数变化都是植物对光抑制的响应机制, 但不同植物在不同胁迫条件下的响应策略不同. ...
毛乌素沙地天然臭柏居群有性更新幼苗动态研究
1
2006
... 臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 ).天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区.在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木.它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 ).毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大.故臭柏极易受到强光照、干旱和低温的单一或多重胁迫.臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 ).毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱. 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
季节性干旱对白皮乌口树(
Arenna depauperata Hutchins)水分状况、叶片光谱特征和荧光参数的影响
1
2016
... 臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 ).天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区.在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木.它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 ).毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大.故臭柏极易受到强光照、干旱和低温的单一或多重胁迫.臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 ).毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱. 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
2种圆柏属植物叶绿素荧光对季节变化的响应特征
2
2010
... 常绿植物叶片的光合作用和蒸腾作用等生理代谢均受到环境因子变化的影响, 如季节变化引起的强光、干旱和低温胁迫(
张有福等, 2010 ).强光胁迫下, 光合天线色素对太阳光能的过量吸收会造成光抑制, 即植物光合作用受到抑制, 可引起光系统II (PSII)反应中心的不可逆破坏, 即光破坏, 进而影响植物的光合生理代谢, 导致植物生长缓慢(
许大全等, 1992 ;
许大全, 2003 ).虽然强光是植物产生光抑制的直接原因, 但干旱、盐渍和低温等叠加胁迫会加剧植物的光抑制, 所以自然界植物的光抑制通常是多种胁迫共同作用的结果.为了物种的延续, 植物在长期的适应和进化过程中, 形成了减轻和抵御光抑制伤害的多种机制.例如, 叶片和叶绿体的避光运动减少了光能的捕获, 增强了光合作用的光能利用效率; 捕光天线色素的转换增加了非光化学耗散及促进了依赖叶黄素的循环等(
许大全等, 1992 ;
高杰等, 2015 ;
代欣等, 2016 ;
付忠等, 2016 ).其中, 叶绿素和叶黄素循环是植物应对光抑制的主要途径之一. ...
... 臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 ).天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区.在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木.它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 ).毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大.故臭柏极易受到强光照、干旱和低温的单一或多重胁迫.臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 ).毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱. 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
遮阴处理对臭柏幼苗光合特性的影响
2
2014
... 臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 ).天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区.在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木.它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 ).毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大.故臭柏极易受到强光照、干旱和低温的单一或多重胁迫.臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 ).毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱. 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
... ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
臭柏抗旱生理特性研究
1
2005
... 绿色植物的光合色素主要由叶绿素、类胡萝卜素和藻胆素三大类色素组成.叶绿素主要由叶绿素a (Chla)和叶绿素b (Chlb)组成.少部分的Chla具有转化光能的作用, 大部分Chla和全部Chlb的作用是吸收与传递光能, 是光系统天线复合体的重要组成成分.由叶绿素酸酯a合成Chlb和由Chlb到Chla的还原过程可以看作一个循环, 即叶绿素循环(
周锋利等, 2005 ;
郭春爱等, 2006 ;
郭春爱, 2007 ;
李玉灵等, 2009 ).当光能过剩时, 植物可适时减少Chlb的含量, 进而减少光能的吸收量; 当光照不足时, 植物可增加Chlb的含量, 从而捕捉更多的光能, 满足植物生长的需要.叶绿素两大组分间的比值(Chla/Chlb)与总量(Chla+Chlb)在一定程度上表示了整个光系统对光量子的捕获和利用状况, 也反映了植物处于特定环境下承受潜在光胁迫的程度.因此, 叶绿素循环不仅是植物对不同光环境做出的即时响应, 也是植物光抑制的防御机制之一.叶黄素类则是胡萝卜素的氧化衍生物, 是类胡萝卜素中含有氧的一组多萜类的总称, 它包括双环氧紫黄质(violaxanthin, V)、环氧玉米黄质(antheraxanthin, A)和玉米黄质(zeaxanthin, Z)等.其中, V、A和Z三者间可以相互转化, 称为叶黄素循环,该循环通过热耗散防御光破坏.当出现过剩光能时, V在紫黄质脱环氧化酶(VDE)的作用下, 经A转化成Z, Z在植物叶黄素循环耗散光能的过程中起关键作用, 可以直接猝灭激发态叶绿素(或改变类囊体膜的流动性)及促进PSII的捕光复合物(LHCII)聚集来增加非辐射能量的耗散, 所以A和Z也被称为热散逸色素(
王强等, 2003 ).当光能不再过剩时, 转化方向相反, Z在玉米黄质环氧化酶(ZE)的催化作用下经A形成V, Z减少而V增加.(A+V+Z)/(Chla+Chlb)和(A+V)/(A+V+Z)比值越高, 意味着叶黄素含量及其中的热散逸色素含量越多或叶绿素含量越少.所以叶黄素循环也是植物对不同光环境做出的即时响应, 与叶绿素循环一样, 也是植物光抑制的防御机制之一.植物的叶绿素荧光主要由PSII反应中心产生, 与光合作用紧密相关, 能够反映光合生理的信息, 特别是胁迫环境下完整植株光合功能的变化(
师生波等, 2014 ).其中, 荧光参数
F v /
F m 指完整叶片中PSII的原初光能转化效率, 是研究光合生理的重要参数, 它能够反映出PSII利用光能的能力(
查倩等, 2016 ;
孙鲁龙等, 2016 ).在非逆境条件下, 不同物种或生态型的叶片,
F v /
F m 均在0.75- 0.85间, 并且不受生长条件的影响, 但在光抑制下,
F v /
F m 值明显降低(
许大全等, 1992 ).故
F v /
F m 是表征光抑制程度的一个重要指标, 该比值越低说明发生光抑制的程度越高(司继播等, 2007).综上所述, 叶绿素循环、叶黄素循环和叶绿素荧光参数变化都是植物对光抑制的响应机制, 但不同植物在不同胁迫条件下的响应策略不同. ...
干旱胁迫对胡杨PSII光化学效率和激能耗散的影响
2011
不同品种小豆光合作用和生长发育对弱光的响应
1
2015
... 77%透光区的臭柏实生苗可通过减少Chlb含量, 升高Chla/Chlb值来避免光能过剩, 这也是耐阳植物适应强光环境的正常响应(
陈煜等, 2006 ;
姜武等, 2007 ;
邹长明等, 2015 ).然而, 4月干旱条件下77%透光区的Chla/Chlb与25%和10%透光区相比无显著差异, 但在湿润区显著大于25%和10%透光区(
图2 ), 说明干旱条件下77%透光区的叶绿素循环光抑制防御机制不及湿润区活跃.如果干旱会加重光抑制的程度, 湿润区都已存在叶绿素循环抵御光抑制的现象, 那么为何干旱区却反而未出现此现象?这可能是由于持续的强光和干旱双重胁迫导致了严重的光抑制, 进而产生了光破坏, 使臭柏幼苗无法启动叶绿素循环.光破坏发生在PSII的反应中心, 直接造成了反应中心色素(进行光能传递和转化且具有光化学活性的特殊态Chla)的大量损失, 使得Chla的含量降低, 从而使Chla/Chlb降低, 结果无法完成叶绿素循环的运作(
潘瑞炽, 2004 ).当然, 臭柏幼苗会通过其它途径来应对严重的光抑制, 本研究中, 干旱条件下77%透光区4月的(A+V)/(A+V+Z)显著高于湿润区(
图3 ), 说明臭柏利用较高的叶黄素循环转化率来加强叶黄素循环的运转, 以消耗过剩的光能.已有研究表明, 叶绿素和类胡萝卜素的光能吸收有一定的互补性(司继播等, 2007), 而且植物可能形成一种稳定的叶绿素-叶黄素-蛋白淬灭复合物来耗散过剩的光能(
李玉灵等, 2009 ).由此可见, 臭柏幼苗应对光抑制的色素机制也是协调统一的.77%透光区臭柏幼苗的光抑制响应机制灵敏、快速且高效, 并且光抑制防御能力较强, 叶绿素循环与叶黄素循环机制同时发生作用, 可达到光能的最佳分配. ...
1
2006
... 臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 ).天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区.在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木.它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 ).毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大.故臭柏极易受到强光照、干旱和低温的单一或多重胁迫.臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 ).毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱. 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
1
2016
... 臭柏(
Sabina vulgaris )又名砂地柏、天山圆柏, 隶属柏科圆柏属, 匍匐灌木, 适应性强、生态功能好且医药价值高(
李晨阳等, 2015 ;
Zhang et al., 2016 ).天然臭柏群落主要分布在我国西北、华北的干旱和半干旱地区.在内蒙古毛乌素沙地植物群落中, 臭柏是唯一的天然常绿针叶灌木.它不但具有防风固沙等生态功能, 还有增加毛乌素沙地绿色景观的作用(
赵顺等, 2014 ).毛乌素沙地地处中纬度北温带, 深居内陆, 远离海洋, 海拔多为1 100-1 300 m, 日照时数为 3 037.80-4 793.01小时, 年降水量为270-395 mm (
王林和等, 2011 ,
2014 ), 集中于7-9月(此3个月的降水占全年的60%-75%) (
冯玉静, 2015 ), 气温的日和年较差大.故臭柏极易受到强光照、干旱和低温的单一或多重胁迫.臭柏对光抑制的防御能力在维持其在该地区生存和生长上具重大意义, 但目前光抑制生理生态机制方面的实验证据很少(
师生波等, 2007 ;
张有福等, 2010 ;
张树斌等, 2016 ).毛乌素沙地中的臭柏实生苗在乌柳(
Salix cheilophila )灌丛下生长, 幼龄时处于遮阴保护状态, 光照较弱. 随着其不断生长, 臭柏逐渐脱离乌柳的荫蔽直至完全暴露于强光之下, 形成纯片灌林, 而当初保护臭柏幼苗的“保姆植物”乌柳衰竭死亡(
Ishii et al., 2006 ;
张国盛等, 2006 ;
赵顺等, 2014 ;
黄秋娴等, 2015 ).笔者在毛乌素沙地也观察到了这一现象, 并由此推测臭柏在完成其天然更新的过程中, 不仅经历了由遮阴到强光的光照变化, 而且经历了由暖湿到干燥的水热条件变化.其必然要对光和水热胁迫做出响应, 启动光抑制防御机制.有研究表明, 成年臭柏在干旱胁迫下通过启动叶绿素循环和叶黄素循环来防止光合机构被破坏(
李玉灵等, 2009 ).然而, 臭柏幼苗如何调控自身的生理机能以逐渐适应强光、干旱和不同的温度环境尚不明确.尤其对光合器官中各类色素的协同调控机制还知之甚少.为此, 本研究结合
F v /
F m , 比较了3年生臭柏实生幼苗在不同光照和水分条件下的叶绿素及叶黄素循环的季节变化, 试图从叶绿素及叶黄素各组分的变化中揭示臭柏实生苗对光及水分胁迫的色素转换响应机制, 为明确臭柏实生苗更新过程中的光响应机制提供理论依据, 并为臭柏种质资源保护和干旱造林技术提供参考. ...
 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}