伍国强*,, 水清照, 冯瑞军 兰州理工大学生命科学与工程学院, 兰州 730050 WuGuoqiang*, ShuiQingzhao, FengRuijun School of Life Science and Engineering, Lanzhou University of Technology, Lanzhou 730050, China 引用本文 伍国强, 水清照, 冯瑞军. 植物K通道AKT1的研究进展. , 2017, 52(2): 225-234
Abstract: Potassium (K) is an essential macronutrient for plant growth and development. K+ uptake was mainly mediated by K+ channels and transporters in root cells. Arabidopsis K+ transporter 1 (AKT1) is one of the important members of the Shaker type K+ channel families and plays crucial roles in root K+ uptake and transmembrane transport of plants. Here, we summarize the latest research advances for the K+ channel AKT1, mainly its molecular structure, tissue-specific expression, regulation mechanisms, and biological function. We propose research hotspots and directions for research into the AKT1 channel.
2 AKT1的分子调控机制尽管自20世纪90年代从拟南芥中克隆到第1个AKT1基因以来, 人们相继在其它植物中克隆得到同源基因。然而, AKT1基因所编码蛋白的调控机制仍不清楚。Xu等(2006)首次在植物中报道了低K+条件下蛋白磷酸化对K+吸收的调控机制。在爪蟾卵母细胞(Xeno- pus oocytes)中, CBL1 (calcineurin B-like protein1)/ CBL9与CIPK23 (CBL-interacting protein kinases 23)互作, 通过磷酸化作用激活AKT1介导的K+吸收(Li et al., 2006; Xu et al., 2006; Cuéllar et al., 2010, 2013)。低K+处理下, CIPK23表达上调, 而CBL1和CBL9似乎是组成型表达(Xu et al., 2006; Cheong et al., 2007); 过量表达CIPK23可显著增强拟南芥K+的吸收能力和低K+耐受性(Xu et al., 2006)。CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009)。由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009)。AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011)。Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应。此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013)。CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015)。 AtKC1是Reintanz等(2002)从拟南芥中分离克隆的α-亚基Shaker通道基因, 主要在根毛、根表皮及皮层细胞中表达。AtKC1作为四聚体的组成亚基参与K+通道四聚体的形成, 其存在对四聚体的K+通透能力产生抑制作用(Reintanz et al., 2002; Geiger et al., 2009)。可见, AtKC1是负调控因子, 也是1个调控亚基(Wang et al., 2010)。另外, AtKC1的存在使四聚体的K+通道活性激活电压向负电压方向偏移, 离子通道更难以被激活, 即改变了其对电压的敏感性(Duby et al., 2008)。因此, 这种负向调控阻断了由AKT1介导的吸收。短时间缺K+处理则上调AtKC1的转录丰度, 随后其转录丰度下调(Shin and Schachtman, 2004), 这种现象可通过缺K+处理几天后的AtKC1转录物消失而得到解释(Pilot et al., 2003)。在异源表达系统中, AtKC1不能独立形成有功能的K+通道(Duby et al., 2008)。然而, 当AtKC1不能正常发挥功能时, 根毛中AKT1介导的内向K+电流的生物物理特征就会发生改变(Jeanguenin et al., 2011)。Wang等(2016)进一步研究证实, 在低K+条件下, CIPK23促进AKT1介导的K+吸收, 而AtKC1抑制AKT1介导的K+内流。他们认为, CIPK23和AtKC1协同调节AKT1的功能, 二者具有一定的互补性。另有研究表明, Atkc1与Atakt1突变体植株的表型相似或相反, 这主要取决于外界环境条件(Geiger et al., 2009; Wang et al., 2010)。AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009)。低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似。在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014)。最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014)。PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性。可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014)。
3 AKT1在植物逆境胁迫响应中的功能3.1 AKT1与K+饥饿耐受性低K+环境是影响植物正常生长发育及作物品质的主要非生物因子之一。K+是种子萌发所需的重要渗透调节物质。在低K+ (50-100 μmol·L-1)条件下, 拟南芥Atakt1突变体种子的胚根不能正常生长; 然而, 当在Atakt1突变体中超表达AtAKT1后, 其种子萌发活力得到恢复, 说明AKT1参与种子萌发期间K+的吸收(Pyo et al., 2010; Xu et al., 2014)。AKT1功能缺失使植物根细胞膜表现出超级化, 对外界环境K+浓度变化异常敏感(Hirsch et al., 1998)。在低K+环境中, 拟南芥Atakt1突变体植株细胞内的K+含量明显降低(Hirsch et al., 1998; Spalding et al., 1999)。电生理学实验表明, 水稻OsAKT1介导内向K+电流, 即使在低K+ (5 μmol·L-1-1 mmol·L-1)条件下也是如此(Li et al., 2014)。有研究表明, 拟南芥Atakt1和水稻Osakt1突变体植株的内向K+电流减弱, K+吸收功能紊乱, 从而影响其生长发育(Reintanz et al., 2002; Xu et al., 2006; Li et al., 2014)。当在水稻Osakt1突变体中超表达OsAKT1后, 低K+胁迫下转基因植株的内向K+电流增强, K+含量显著增加(Li et al., 2014)。这进一步证实了AKT1介导植物根吸收K+。Xu等(2014)采用非损伤微测技术(no-invasive micro-test technology, NMT), 在棉花GhAKT1功能研究中也得到类似的结果。可见, 在低K+环境中, AKT1介导的K+吸收系统对维持植物正常生长发育具有重要作用。
3.2 AKT1与植物耐盐性盐胁迫下, 植物体内积累大量的Na+, 一方面抑制K+的吸收, 另一方面与K+竞争一些酶活性结合位点, 影响蛋白质合成及核糖体功能, 从而产生Na+毒害(Horie et al., 2009; Zhang et al., 2010)。一般认为, 维持细胞质内较高的K+/Na+比是植物适应盐胁迫的重要策略之一(Horie et al., 2009; Shabala et al., 2010; Chérel et al., 2014)。AKT1是参与K+吸收的重要通道蛋白, 在植物响应盐胁迫中起着重要作用。异源表达系统研究表明, 尽管AKT1对K+具有更强的选择性, 但并不排除对Na+的吸收(Maathuis et al., 1997)。低Na+条件下, 内向整流K+通道未表现出明显的Na+进出现象; 但在高盐条件下, 其对K+表现出明显的选择性(Amtmann and Sanders, 1999)。Goll- dack等(2003)研究表明, 盐处理下水稻根吸收Na+与OsAKT1的表达水平有直接关系。由此证明OsAKT1参与盐胁迫下Na+/K+比调控, 从而增强水稻的耐盐性。Wang等(2015)研究发现, 无论是低盐(25 mmol·L-1 NaCl)还是高盐(150 mmol·L-1 NaCl)条件下, 小花碱茅PtAKT1选择性吸收K+的能力显著强于Na+ (selective absorption capacity for K+ over Na+, SA)。进一步研究发现, PtAKT1的转录水平与SA值之间呈显著正相关, 可见盐胁迫下PtAKT1是调控小花碱茅K+选择性吸收的关键因子(Wang et al., 2015)。与野生型相比, 盐胁迫下超表达PtAKT1的拟南芥转基因植株K+含量增加, 而Na+含量降低, 表明PtAKT1增强了转基因植株对K+的吸收能力, 从而提高其耐盐性(Ardie et al., 2010)。Duan等(2015)研究发现, 盐胁迫下盐地碱蓬SsAKT1能促进植株体内K+积累, 进而增强植物耐盐性。在盐(100 mmol·L-1 NaCl)处理下, 大麦HvAKT1在叶伸长区的表达水平明显增加, 说明HvAKT1参与维持叶肉细胞K+的稳态平衡(Boscari et al., 2009)。我们的研究发现, 在5-50 mmol·L-1 NaCl处理下, 分别添加3和6 mmol·L-1 Cs+ (K+通道AKT1特异性抑制剂)均可显著降低甜菜(Beta vulgaris)根的K+净吸收速率, 可见AKT1可介导甜菜根K+的吸收(Wu et al., 2015b)。这些研究结果表明, AKT1在K+和Na+选择性吸收、维持植物体内K+含量以及增强植物耐盐性等方面具有重要作用。
3.3 AKT1与植物抗旱性干旱是制约全球作物生长和产量的主要非生物因素之一(Chaves and Oliveira, 2004; 山仑, 2011)。植物响应干旱胁迫的重要策略之一是吸收并在体内积累大量的溶质(如K+、Na+和脯氨酸等) (Wang et al., 2004; Mahouachi et al., 2006; Wu et al., 2015a), 降低细胞渗透势, 从而增强植物细胞吸水能力。AKT1在植物响应干旱胁迫中的作用主要有2方面。(1) 干旱胁迫下AKT1增强植物根的K+吸收能力。在水稻中超表达OsAKT1基因后, 转基因植株在干旱条件下生长良好, 且根中积累较多的K+; 相反, Osakt1突变体植物表现为生长不良, 且根中K+含量明显减少(Ahmad et al., 2016)。由此表明, 超表达AKT1转基因植株通过增加根部K+的积累来改善细胞渗透调节能力, 从而增强植物的抗旱性。(2) AKT1通过调节气孔的运动影响植物体内的水分平衡。研究表明, 拟南芥AtAKT1在保卫细胞质膜上呈低丰度表达(Lagarde et al., 1996)。在渗透胁迫下, 拟南芥Atakt1突变体植株的蒸腾速率和水分消耗量明显低于野生型植株(Nieves- Cordones et al., 2012)。由此可见, AKT1功能缺失致使保卫细胞K+跨膜转运体系受损, 气孔导度下降。Ahmad等(2016)进一步研究发现, 与野生型相比, 在干旱胁迫下水稻Osakt1突变体植株的气孔导度明显降低; 而超表达OsAKT1显著增强了转基因植株的气孔导度。这些研究结果表明, AKT1在介导保卫细胞K+转运、调控气孔运动及维持细胞水分平衡中发挥重要作用。
CuéllarT, AzeemF, AndrianteranagnaM, PascaudF, VerdeilJL, SentenacH, ZimmermannS, GaillardI (2013). Potassium transport in developing fleshy fruits: the grapevine inward K+ channel VvK1.2 is activated by CIPK-CBL complexes and induced in ripening berry flesh cells.Plant J 73, 1006-1018. [本文引用: 3]
[15]
CuéllarT, PascaudF, VerdeilJL, TorregrosaL, Adam- BlondonAF, ThibaudJB, SentenacH, GaillardI (2010). A grapevine Shaker inward K+ channel activated by the calcineurin B-like calcium sensor 1-protein kinase CIPK23 network is expressed in grape berries under dr- ought stress conditions. Plant J 61, 58-69. [本文引用: 3]
[16]
DemidchikV (2014). Mechanisms and physiological roles of K+ efflux from root cells.J Plant Physiol 171, 696-707. [本文引用: 2]
[17]
DemidchikV, MaathuisFJM (2007). Physiological roles of nonselective cation channels in plants: from salt stress to signaling and development.New Phytol 175, 387-404. [本文引用: 1]
[18]
DietrichP, AnschützU, KuglerA, BeckerD (2010). Physiology and biophysics of plant ligand gated ion chan- nels.Plant Biol 12, 80-93. [本文引用: 1]
[19]
DreyerI, BlattMR (2009). What makes a gate? The ins and outs of Kv-like K+ channels in plants.Trends Plant Sci 14, 383-390. [本文引用: 1]
DuanRH, MaQ, ZhangJL, HuJ, BaoAK, WeiL, WangQ, LuanS, WangSM (2015). The inward-rectifying K+ channel SsAKT1 is a candidate involved in K+ uptake in the halophyteSuaeda salsa under saline condition. Plant Soil 395, 173-187. [本文引用: 2]
[22]
DubyG, HosyE, FizamesC, AlconC, CostaA, SentenacH, ThibaudJB (2008). AtKC1, a conditionally targeted Shaker-type subunit, regulates the activity of plant K+ ch- annels.Plant J 53, 115-123. [本文引用: 2]
[23]
FormentinE, VarottoS, CostaA, DowneyP, BreganteM, NasoA, PiccoC, GambaleF, SchiavoFL (2004). DKT1, a novel K+ channel from carrot, forms functional heteromeric channels with KDC1.FEBS Lett 573, 61-67. [本文引用: 1]
[24]
FuchsI, StolzleS, IvashikinaN, HedrichR (2005). Rice K+ uptake channel OsAKT1 is sensitive to salt stress.Planta 221, 212-221. [本文引用: 1]
[25]
GambaleF, UozumiN (2006). Properties of Shaker-type potassium channels in higher plants.J Membrane Biol 210, 1-19. [本文引用: 1]
GierthM, MäserP (2007). Potassium transporters in plants-involvement in K+ acquisition, redistribution and homeostasis.FEBS Lett 581, 2348-2356. [本文引用: 1]
GrefenC, BlattMR (2012). Do calcineurin B-like proteins interact independently of the serine threonine kinase CIPK23 with the K+ channel AKT1?Plant Physiol 159, 915-919. [本文引用: 1]
[30]
HanM, WuW, WuWH, WangY (2016). Potassium transporter KUP7 is involved in K+ acquisition and translocation in Arabidopsis root under K+-limited conditions.Mol Plant 9, 437-446. [本文引用: 1]
[31]
HartjeS, ZimmermannS, KlonusD, Mueller-RoeberB (2000). Functional characterization of LKT1, a K+ uptake channel from tomato root hairs, and comparison with the closely related potato inwardly rectifying K+ channel SKT1 after expression inXenopus oocytes. Planta 210, 723-731. [本文引用: 1]
[32]
HedrichP (2012). Ion channels in plants.Physiol Rev 92, 1777-1811. [本文引用: 2]
[33]
HirschRE, LewisBD, SpaldingEP, SussmanMR (1998). A role for the AKT1 potassium channel in plant nutrition.Science 280, 918-921. [本文引用: 2]
[34]
HonsbeinA, SokolovskiS, GrefenC, CampanoniP, PratelliR, PanequeM, ChenZ, JohanssonI, BlattMR (2009). A tripartite SNARE-K+ channel complex mediates in channel-dependent K+ nutrition in Arabidopsis.Plant Cell 21, 2859-2877. [本文引用: 1]
JeangueninL, AlconC, DubyG, BoeglinM, ChérelI, GaillardI, ZimmermannS, SentenacH, VéryAA (2011). AtKC1 is a general modulator of Arabidopsis inward Shaker channel activity.Plant J 67, 570-582. [本文引用: 1]
[37]
KronzuckerHJ, BrittoDT (2011). Sodium transport in plants: a critical review.New Phytol 189, 54-81. [本文引用: 1]
[38]
LagardeD, BassetM, LepetitM, ConejeroG, GaymardF, AstrucS, GrignonC (1996). Tissue-specific expression of ArabidopsisAKT1 gene is consistent with a role in K+ nutrition. Plant J 9, 195-203. [本文引用: 2]
[39]
LanWZ, LeeSC, CheYF, JiangYQ, LuanS (2011). Mechanistic analysis of AKT1 regulation by the CBL- CIPK-PP2CA interactions.Mol Plant 4, 527-536. [本文引用: 1]
[40]
LeeSC, LanWZ, KimBG, LiL, CheongYH, PandeyGK, LuG, BuchananBB, LuanS (2007). A protein phosphorylation/dephosphorylation network regulates a plant potassium channel.Proc Natl Acad Sci USA 104, 15959-15964. [本文引用: 1]
[41]
LiJ, LongY, QiGN, XuZJ, WuWH, WangY (2014). The Os-AKT1 channel is critical for K+ uptake in rice roots and is modulated by the rice CBL1-CIPK23 complex.Plant Cell 26, 3387-3402. [本文引用: 3]
[42]
LiL, KimBG, CheongYH, PandeyGK, LuanS (2006). A Ca2+ signaling pathway regulates a K+ channel for low-K response in Arabidopsis.Proc Natl Acad Sci USA 103, 12625-12630. [本文引用: 2]
[43]
LiR, ZhangJ, WeiJ, WangH, WangY, MaR (2009). Functions and mechanisms of the CBL-CIPK signaling system in plant response to abiotic stress.Proc Natl Acad Sci USA 19, 667-676. [本文引用: 2]
[44]
MaathuisFJM (2009). Physiological functions of mineral macronutrients.Curr Opin Plant Biol 12, 250-258. [本文引用: 1]
[45]
MaathuisFJM, FilatovV, HerzykP, KrijgerGC, AxelsenKB, ChenSX (2003). Transcriptome analysis of root trans- porters reveals participation of multiple gene families in the response to cation stress.Plant J 35, 675-692. [本文引用: 1]
MaathuisFJM, SandersD (1993). Energization of potassium uptake in Arabidopsis thaliana. Planta 191, 302-307. [本文引用: 1]
[48]
MahouachiJ, SocorroAR, TalonM (2006). Responses of papaya seedlings (Carica papaya L.) to water stress and re-hydration: growth, photosynthesis and mineral nutrient imbalance. Plant Soil 281, 137-146. [本文引用: 1]
Nieves-CordonesM, AlemánF, MartínezV, RubioF (2014a). K+ uptake in plant roots. The systems involved, their regulation and parallels in other organisms.J Plant Physiol 171, 688-695.
[51]
Nieves-CordonesM, CaballeroF, MartínezV, RubioF (2012). Disruption of theArabidopsis thaliana inward- rectifier K+ channel AKT1 improves plant responses to water stress. Plant Cell Physiol 53, 423-432. [本文引用: 1]
[52]
Nieves-CordonesM, ChavanieuA, JeangueninL, AlconC, SzponarskiW, EstaranS, CherelI, ZimmermannS, SentenacH, GaillardI (2014b). Distinct amino acids in the C-linker domain of the Arabidopsis K+ channel KAT2 determine its subcellular localization and activity at the plasma membrane. Plant Physiol 164, 1415-1429. [本文引用: 2]
[53]
Nieves-CordonesM, GaillardI (2014). Involvement of the S4-S5 linker and the C-linker domain regions to voltage-gating in plant Shaker channels: comparison with animal HCN and Kv channels.Plant Signal Behav 9, 10. [本文引用: 7]
[54]
PhilipparK, FuchsI, LuthenH, HothS, BauerCS, HagaK, ThielG, LjungK, SandbergG, BottgerM, BeckerD, HedrlchR (1999). Auxin-induced K+ channel expression represents an essential step in coleoptile growth and gravitropism.Proc Natl Acad Sci USA 96, 12186-12191. [本文引用: 1]
[55]
PilotG, GaymardF, MoulineK, ChérelI, SentenacH (2003). Regulated expression of Arabidopsis Shaker K+ channel genes involved in K+ uptake and distribution in the plant.Plant Mol Biol 51, 773-787. [本文引用: 3]
[56]
PyoYJ, GierthM, SchroederJI, ChoMH (2010). High- affinity K+ transport in Arabidopsis: AtHAK5 and AKT1 are vital for seedling establishment and post germination growth under low-potassium conditions.Plant Physiol 153, 863-875. [本文引用: 1]
[57]
RagelP, RódenasR, García-MartínE, AndrésZ, VillaltaI, Nieves-CordonesM, RiveroRM, MartínezV, PardoJM, QuinteroFJ, RubioF (2015). The CBL-interacting protein kinase CIPK23 regulates HAK5-mediated high-affinity K+ uptake in Arabidopsis roots.Plant Physiol 169, 2863-2873. [本文引用: 1]
RenXL, QiGN, FengHQ, ZhaoS, ZhaoSS, WangY, WuWH (2013). Calcineurin B-like protein CBL10 directly interacts with AKT1 and modulates K+ homeostasis in Ara- bidopsis. Plant J 74, 258-266. [本文引用: 1]
[60]
RiedelsbergerJ, SharmaT, GonzalezW, GajdanowiczP, Morales-NavarroSE, Garcia-MataC, Mueller-RoeberB, Gonzalez-NiloFD, BlattMR, DreyerI (2010). Distri- buted structures underlie gating differences between the kin channel KAT1 and the K out channel SKOR.Mol Plant 3, 236-245. [本文引用: 1]
[61]
SanoT, BeckerD, IvashikinaN, WegnerLH, Zimmer- mannU, RoelfsemaMRG, NagataT, HedrichR (2007). Plant cells must pass a K+ threshold to re-enter the cell cycle.Plant J 50, 401-413.
[62]
SentenacH, BonneaudN, MinetM, LacrouteF, SalmonJM, GaymardF, GrignonC (1992). Cloning and expression in yeast of a plant potassium ion transport system. Science 256, 663-665.
[63]
ShabalaS (2003). Regulation of potassium transport in leaves: from molecular to tissue level.Ann Bot 92, 627-634. [本文引用: 1]
[64]
ShabalaS, CuinTA, PottosinI (2010). Polyamines prevent NaCl-induced K+ efflux from pea mesophyll by blocking non-selective cation channels. FEBS Lett 15, 1993-1999. [本文引用: 1]
SpaldingEP, HirschRE, LewisDR, QiZ, SussmanMR, LewisBD (1999). Potassium uptake supporting plant growth in the absence of AKT1 channel activity: inhibition by ammonium and stimulation by sodium. J Gen Physiol 113, 909-918. [本文引用: 1]
[67]
SuH, GolldackD, KatsuharaM, ZhaoC, BohnertHJ (2001). Expression and stress-dependent induction of potassium channel transcripts in the common ice plant. Plant Physiol 125, 604-614.
[68]
WangP, GuoQ, WangQ, ZhouXR, WangSM (2015). PtAKT1 maintains selective absorption capacity for K+ over Na+ in halophyte Puccinellia tenuiflora under salt stress. Acta Physiol Plant 37, 100. [本文引用: 3]
[69]
WangSM, WanCG, WangYR, ChenH, ZhouZY, FuH, SosebeebRE (2004). The characteristics of Na+, K+ and free proline distribution in several drought-resistant plants of the Alxa Desert, China.J Arid Environ 56, 525-539. [本文引用: 1]
XuJ, LiHD, ChenLQ, WangY, LiuLL, HeL, WuWH (2006). A protein kinase, interacting with two calcineurin B-like proteins, regulates K+ transporter AKT1 in Arabidopsis.Cell 125, 1347-1360. [本文引用: 4]
[78]
XuJ, TianXL, EnejiAE, LiZH (2014). Functional characterization of GhAKT1, a novel Shaker-like K+ channel gene involved in K+ uptake from cotton(Gossypium hirsutum). Gene 545, 61-71. [本文引用: 2]
[79]
ZhangJL, FlowersTJ, WangSM (2010). Mechanism of sodium uptake by roots of higher plants.Plant Soil 326, 45-60. [本文引用: 1]
[80]
ZimmermannS, TalkeI, EhrhardtT, NastG, MüllerröberB (1998). Characterization of SKT1, an inwardly rectifying potassium channel from potato, by heterologous expression in insect cells.Plant Physiol 116, 879-890. [本文引用: 1]
科学应对农业干旱 1 2011
... 干旱是制约全球作物生长和产量的主要非生物因素之一(Chaves and Oliveira, 2004; 山仑, 2011).植物响应干旱胁迫的重要策略之一是吸收并在体内积累大量的溶质(如K+、Na+和脯氨酸等) (Wang et al., 2004; Mahouachi et al., 2006; Wu et al., 2015a), 降低细胞渗透势, 从而增强植物细胞吸水能力.AKT1在植物响应干旱胁迫中的作用主要有2方面.(1) 干旱胁迫下AKT1增强植物根的K+吸收能力.在水稻中超表达OsAKT1基因后, 转基因植株在干旱条件下生长良好, 且根中积累较多的K+; 相反, Osakt1突变体植物表现为生长不良, 且根中K+含量明显减少(Ahmad et al., 2016).由此表明, 超表达AKT1转基因植株通过增加根部K+的积累来改善细胞渗透调节能力, 从而增强植物的抗旱性.(2) AKT1通过调节气孔的运动影响植物体内的水分平衡.研究表明, 拟南芥AtAKT1在保卫细胞质膜上呈低丰度表达(Lagarde et al., 1996).在渗透胁迫下, 拟南芥Atakt1突变体植株的蒸腾速率和水分消耗量明显低于野生型植株(Nieves- Cordones et al., 2012).由此可见, AKT1功能缺失致使保卫细胞K+跨膜转运体系受损, 气孔导度下降.Ahmad等(2016)进一步研究发现, 与野生型相比, 在干旱胁迫下水稻Osakt1突变体植株的气孔导度明显降低; 而超表达OsAKT1显著增强了转基因植株的气孔导度.这些研究结果表明, AKT1在介导保卫细胞K+转运、调控气孔运动及维持细胞水分平衡中发挥重要作用. ...
植物钾营养高效分子遗传机制 1 2009
... K+是植物细胞中含量最丰富的无机阳离子之一, 主要通过根表皮细胞和皮层细胞的质膜K+转运系统吸收并运输到植物体内(王毅和武维华, 2009; Alemán et al., 2011; Dreyer and Uozumi, 2011; Wang and Wu, 2013; Nieves-Cordones et al., 2014a).K+在植物生长发育、逆境胁迫响应及生理生化反应等方面起着重要作用(Alemán et al., 2011; Nieves-Cordones et al., 2014a).一般而言, 植物细胞维持正常新陈代谢所需K+的最佳浓度为100 mmol·L-1, 但土壤中能被植物利用的K+浓度仅为0.1-1 mmol·L-1, 在一些盐碱地中甚至更低(Maathuis and Sanders, 1993; Maathuis, 2009).因此, 土壤中的K+远远不能满足植物正常生长需要, 使植物经常处于K+饥饿状态, 严重影响其生长发育.鉴于复杂多变的环境及K+的重要作用, 众多研究者对植物K+吸收和转运机制进行了深入研究. ...
2001
2 2016
... 在K+饥饿状态下, 小麦根TaAKT1 (Buschmann et al., 2000)和小花碱茅(Puccinellia tenuiflora) PtAKT1 (Ardie et al., 2010)的转录水平显著提高.在拟南芥中, 外部K+浓度在5 μmol·L-1至5 mmol·L-1范围内未影响AtAKT1的转录丰度(Lagarde et al., 1996).在棉花(Xu et al., 2014)和小花碱茅(Wang et al., 2015)中也有类似报道, 由此推测AKT1可能受转录后调控.然而, 25-250 mmol·L-1 NaCl短期(6小时)处理能够诱导盐地碱蓬根SsAKT1的表达(Duan et al., 2015).研究表明, 水稻OsAKT1主要在根的外皮层和内皮层细胞以及叶的木质部薄壁组织、韧皮部和叶肉细胞中表达(Golldack et al., 2003).盐胁迫(150 mmol·L-1 NaCl处理48小时)下, 盐敏感型水稻品种Pokkali和BK根外皮层细胞中的OsAKT1转录物消失, 而耐盐型品种IR29中的则未受到影响(Golldack et al., 2003).Su等(2001)的研究表明, 盐胁迫下盐生植物冰叶日中花(Mesembryanthemum crystallinum)根中与AtAKT1同源的K+通道基因McAKT1表达水平下调.在小花碱茅中, 尽管盐处理会下调PtAKT1的表达, 但150 mmol·L-1 NaCl处理下PtAKT1的转录丰度显著高于25 mmol·L-1 (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ... ... 干旱是制约全球作物生长和产量的主要非生物因素之一(Chaves and Oliveira, 2004; 山仑, 2011).植物响应干旱胁迫的重要策略之一是吸收并在体内积累大量的溶质(如K+、Na+和脯氨酸等) (Wang et al., 2004; Mahouachi et al., 2006; Wu et al., 2015a), 降低细胞渗透势, 从而增强植物细胞吸水能力.AKT1在植物响应干旱胁迫中的作用主要有2方面.(1) 干旱胁迫下AKT1增强植物根的K+吸收能力.在水稻中超表达OsAKT1基因后, 转基因植株在干旱条件下生长良好, 且根中积累较多的K+; 相反, Osakt1突变体植物表现为生长不良, 且根中K+含量明显减少(Ahmad et al., 2016).由此表明, 超表达AKT1转基因植株通过增加根部K+的积累来改善细胞渗透调节能力, 从而增强植物的抗旱性.(2) AKT1通过调节气孔的运动影响植物体内的水分平衡.研究表明, 拟南芥AtAKT1在保卫细胞质膜上呈低丰度表达(Lagarde et al., 1996).在渗透胁迫下, 拟南芥Atakt1突变体植株的蒸腾速率和水分消耗量明显低于野生型植株(Nieves- Cordones et al., 2012).由此可见, AKT1功能缺失致使保卫细胞K+跨膜转运体系受损, 气孔导度下降.Ahmad等(2016)进一步研究发现, 与野生型相比, 在干旱胁迫下水稻Osakt1突变体植株的气孔导度明显降低; 而超表达OsAKT1显著增强了转基因植株的气孔导度.这些研究结果表明, AKT1在介导保卫细胞K+转运、调控气孔运动及维持细胞水分平衡中发挥重要作用. ...
3 2011
... K+是植物细胞中含量最丰富的无机阳离子之一, 主要通过根表皮细胞和皮层细胞的质膜K+转运系统吸收并运输到植物体内(王毅和武维华, 2009; Alemán et al., 2011; Dreyer and Uozumi, 2011; Wang and Wu, 2013; Nieves-Cordones et al., 2014a).K+在植物生长发育、逆境胁迫响应及生理生化反应等方面起着重要作用(Alemán et al., 2011; Nieves-Cordones et al., 2014a).一般而言, 植物细胞维持正常新陈代谢所需K+的最佳浓度为100 mmol·L-1, 但土壤中能被植物利用的K+浓度仅为0.1-1 mmol·L-1, 在一些盐碱地中甚至更低(Maathuis and Sanders, 1993; Maathuis, 2009).因此, 土壤中的K+远远不能满足植物正常生长需要, 使植物经常处于K+饥饿状态, 严重影响其生长发育.鉴于复杂多变的环境及K+的重要作用, 众多研究者对植物K+吸收和转运机制进行了深入研究. ... ... 在植物生长发育、逆境胁迫响应及生理生化反应等方面起着重要作用(Alemán et al., 2011; Nieves-Cordones et al., 2014a).一般而言, 植物细胞维持正常新陈代谢所需K+的最佳浓度为100 mmol·L-1, 但土壤中能被植物利用的K+浓度仅为0.1-1 mmol·L-1, 在一些盐碱地中甚至更低(Maathuis and Sanders, 1993; Maathuis, 2009).因此, 土壤中的K+远远不能满足植物正常生长需要, 使植物经常处于K+饥饿状态, 严重影响其生长发育.鉴于复杂多变的环境及K+的重要作用, 众多研究者对植物K+吸收和转运机制进行了深入研究. ... ... AKT1 (Arabidopsis K+ transporter 1)是植物中广泛存在的一类内向整流K+通道, 与动物中的Shaker型K+通道具有高度的同源性(Pilot et al., 2003).Sentenac等(1992)利用酵母双突变体互补法从拟南芥中克隆到第1个高等植物的Shaker型K+通道基因AtAKT1.该基因位于拟南芥第2号染色体上, 含有11个外显子和10个内含子, 编码的多肽由857个氨基酸残基组成.此后, 科研人员相继在马铃薯(Sola- num tuberosum) (Zimmermann et al., 1998)、玉米(Zea mays) (Philippar et al., 1999)、小麦(Triticum aestivum) (Buschmann et al., 2000)、番茄(Lycoper- sicon esculentum) (Hartje et al., 2000)、水稻(Oryza sativa) (Fuchs et al., 2005)、大麦(Hordeum vulgare) (Boscari et al., 2009)、葡萄(Vitis vinifera) (Cuéllar et al., 2010, 2013)和盐地碱蓬(Suaeda salsa) (Duan et al., 2015)等植物中分离得到AKT1基因.AKT1在维持植物离子稳态平衡及增强植物逆境胁迫耐受性等方面起着重要作用(Alemán et al., 2011; Wang and Wu, 2013).本文从植物AKT1的分子结构特点、调控机制及生物学功能等方面对其主要研究进展进行了归纳总结, 旨在为植物AKT1的进一步研究奠定基础. ...
1 1999
... 盐胁迫下, 植物体内积累大量的Na+, 一方面抑制K+的吸收, 另一方面与K+竞争一些酶活性结合位点, 影响蛋白质合成及核糖体功能, 从而产生Na+毒害(Horie et al., 2009; Zhang et al., 2010).一般认为, 维持细胞质内较高的K+/Na+比是植物适应盐胁迫的重要策略之一(Horie et al., 2009; Shabala et al., 2010; Chérel et al., 2014).AKT1是参与K+吸收的重要通道蛋白, 在植物响应盐胁迫中起着重要作用.异源表达系统研究表明, 尽管AKT1对K+具有更强的选择性, 但并不排除对Na+的吸收(Maathuis et al., 1997).低Na+条件下, 内向整流K+通道未表现出明显的Na+进出现象; 但在高盐条件下, 其对K+表现出明显的选择性(Amtmann and Sanders, 1999).Goll- dack等(2003)研究表明, 盐处理下水稻根吸收Na+与OsAKT1的表达水平有直接关系.由此证明OsAKT1参与盐胁迫下Na+/K+比调控, 从而增强水稻的耐盐性.Wang等(2015)研究发现, 无论是低盐(25 mmol·L-1 NaCl)还是高盐(150 mmol·L-1 NaCl)条件下, 小花碱茅PtAKT1选择性吸收K+的能力显著强于Na+ (selective absorption capacity for K+ over Na+, SA).进一步研究发现, PtAKT1的转录水平与SA值之间呈显著正相关, 可见盐胁迫下PtAKT1是调控小花碱茅K+选择性吸收的关键因子(Wang et al., 2015).与野生型相比, 盐胁迫下超表达PtAKT1的拟南芥转基因植株K+含量增加, 而Na+含量降低, 表明PtAKT1增强了转基因植株对K+的吸收能力, 从而提高其耐盐性(Ardie et al., 2010).Duan等(2015)研究发现, 盐胁迫下盐地碱蓬SsAKT1能促进植株体内K+积累, 进而增强植物耐盐性.在盐(100 mmol·L-1 NaCl)处理下, 大麦HvAKT1在叶伸长区的表达水平明显增加, 说明HvAKT1参与维持叶肉细胞K+的稳态平衡(Boscari et al., 2009).我们的研究发现, 在5-50 mmol·L-1 NaCl处理下, 分别添加3和6 mmol·L-1 Cs+ (K+通道AKT1特异性抑制剂)均可显著降低甜菜(Beta vulgaris)根的K+净吸收速率, 可见AKT1可介导甜菜根K+的吸收(Wu et al., 2015b).这些研究结果表明, AKT1在K+和Na+选择性吸收、维持植物体内K+含量以及增强植物耐盐性等方面具有重要作用. ...
2 2010
... 在K+饥饿状态下, 小麦根TaAKT1 (Buschmann et al., 2000)和小花碱茅(Puccinellia tenuiflora) PtAKT1 (Ardie et al., 2010)的转录水平显著提高.在拟南芥中, 外部K+浓度在5 μmol·L-1至5 mmol·L-1范围内未影响AtAKT1的转录丰度(Lagarde et al., 1996).在棉花(Xu et al., 2014)和小花碱茅(Wang et al., 2015)中也有类似报道, 由此推测AKT1可能受转录后调控.然而, 25-250 mmol·L-1 NaCl短期(6小时)处理能够诱导盐地碱蓬根SsAKT1的表达(Duan et al., 2015).研究表明, 水稻OsAKT1主要在根的外皮层和内皮层细胞以及叶的木质部薄壁组织、韧皮部和叶肉细胞中表达(Golldack et al., 2003).盐胁迫(150 mmol·L-1 NaCl处理48小时)下, 盐敏感型水稻品种Pokkali和BK根外皮层细胞中的OsAKT1转录物消失, 而耐盐型品种IR29中的则未受到影响(Golldack et al., 2003).Su等(2001)的研究表明, 盐胁迫下盐生植物冰叶日中花(Mesembryanthemum crystallinum)根中与AtAKT1同源的K+通道基因McAKT1表达水平下调.在小花碱茅中, 尽管盐处理会下调PtAKT1的表达, 但150 mmol·L-1 NaCl处理下PtAKT1的转录丰度显著高于25 mmol·L-1 (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ... ... 盐胁迫下, 植物体内积累大量的Na+, 一方面抑制K+的吸收, 另一方面与K+竞争一些酶活性结合位点, 影响蛋白质合成及核糖体功能, 从而产生Na+毒害(Horie et al., 2009; Zhang et al., 2010).一般认为, 维持细胞质内较高的K+/Na+比是植物适应盐胁迫的重要策略之一(Horie et al., 2009; Shabala et al., 2010; Chérel et al., 2014).AKT1是参与K+吸收的重要通道蛋白, 在植物响应盐胁迫中起着重要作用.异源表达系统研究表明, 尽管AKT1对K+具有更强的选择性, 但并不排除对Na+的吸收(Maathuis et al., 1997).低Na+条件下, 内向整流K+通道未表现出明显的Na+进出现象; 但在高盐条件下, 其对K+表现出明显的选择性(Amtmann and Sanders, 1999).Goll- dack等(2003)研究表明, 盐处理下水稻根吸收Na+与OsAKT1的表达水平有直接关系.由此证明OsAKT1参与盐胁迫下Na+/K+比调控, 从而增强水稻的耐盐性.Wang等(2015)研究发现, 无论是低盐(25 mmol·L-1 NaCl)还是高盐(150 mmol·L-1 NaCl)条件下, 小花碱茅PtAKT1选择性吸收K+的能力显著强于Na+ (selective absorption capacity for K+ over Na+, SA).进一步研究发现, PtAKT1的转录水平与SA值之间呈显著正相关, 可见盐胁迫下PtAKT1是调控小花碱茅K+选择性吸收的关键因子(Wang et al., 2015).与野生型相比, 盐胁迫下超表达PtAKT1的拟南芥转基因植株K+含量增加, 而Na+含量降低, 表明PtAKT1增强了转基因植株对K+的吸收能力, 从而提高其耐盐性(Ardie et al., 2010).Duan等(2015)研究发现, 盐胁迫下盐地碱蓬SsAKT1能促进植株体内K+积累, 进而增强植物耐盐性.在盐(100 mmol·L-1 NaCl)处理下, 大麦HvAKT1在叶伸长区的表达水平明显增加, 说明HvAKT1参与维持叶肉细胞K+的稳态平衡(Boscari et al., 2009).我们的研究发现, 在5-50 mmol·L-1 NaCl处理下, 分别添加3和6 mmol·L-1 Cs+ (K+通道AKT1特异性抑制剂)均可显著降低甜菜(Beta vulgaris)根的K+净吸收速率, 可见AKT1可介导甜菜根K+的吸收(Wu et al., 2015b).这些研究结果表明, AKT1在K+和Na+选择性吸收、维持植物体内K+含量以及增强植物耐盐性等方面具有重要作用. ...
1 2014
... 植物AKT1与动物Shaker型K+通道结构高度相似(图1), 含有6个跨膜片段(S1-S6), 其中第4个跨膜片段S4含有大量带正电荷的氨基酸残基, 是电压感受器, 其主要功能为响应膜电势的变化.该片段可在膜上移动, 使得膜通道构象改变, 从而控制通道孔的开放与关闭(Benito et al., 2014; Nieves-Cordones and Gaillard, 2014).S5和S6之间含有1个高度保守的P环结构域, 该结构域是陷入细胞膜内的一段多肽片段, 构成通道孔.AKT1含有典型的TxGYG (Thr-X- Gly-Tyr-Gly)序列, 为K+选择器, 这是Shaker型K+通道的共同特点.胞质C末端从第6个跨膜片段末尾起, 含有1个C-接头(约有80个氨基酸残基), 1个环核苷酸结合域(cyclic-nucleotide binding domain, CNBD), 1个锚蛋白域和1个富含疏水性、酸性残基的KHA域(Nieves-Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).对大多数Shaker型K+通道家族成员来说, 锚蛋白域有助于通道与细胞骨架的连接、蛋白质的相互作用和细胞溶质的调节(Dreyer and Blatt, 2009; Riedelsberger et al., 2010; Nieves- Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).Shaker型K+通道的重要特点之一是能够形成异源四聚体结构, 可使植物调控不同细胞中的K+转运活性.这种调控在每个器官或者组织中相对独立, 并且受到环境的影响(Gambale and Uozumi, 2006). ...
2 2009
... AKT1 (Arabidopsis K+ transporter 1)是植物中广泛存在的一类内向整流K+通道, 与动物中的Shaker型K+通道具有高度的同源性(Pilot et al., 2003).Sentenac等(1992)利用酵母双突变体互补法从拟南芥中克隆到第1个高等植物的Shaker型K+通道基因AtAKT1.该基因位于拟南芥第2号染色体上, 含有11个外显子和10个内含子, 编码的多肽由857个氨基酸残基组成.此后, 科研人员相继在马铃薯(Sola- num tuberosum) (Zimmermann et al., 1998)、玉米(Zea mays) (Philippar et al., 1999)、小麦(Triticum aestivum) (Buschmann et al., 2000)、番茄(Lycoper- sicon esculentum) (Hartje et al., 2000)、水稻(Oryza sativa) (Fuchs et al., 2005)、大麦(Hordeum vulgare) (Boscari et al., 2009)、葡萄(Vitis vinifera) (Cuéllar et al., 2010, 2013)和盐地碱蓬(Suaeda salsa) (Duan et al., 2015)等植物中分离得到AKT1基因.AKT1在维持植物离子稳态平衡及增强植物逆境胁迫耐受性等方面起着重要作用(Alemán et al., 2011; Wang and Wu, 2013).本文从植物AKT1的分子结构特点、调控机制及生物学功能等方面对其主要研究进展进行了归纳总结, 旨在为植物AKT1的进一步研究奠定基础. ... ... 盐胁迫下, 植物体内积累大量的Na+, 一方面抑制K+的吸收, 另一方面与K+竞争一些酶活性结合位点, 影响蛋白质合成及核糖体功能, 从而产生Na+毒害(Horie et al., 2009; Zhang et al., 2010).一般认为, 维持细胞质内较高的K+/Na+比是植物适应盐胁迫的重要策略之一(Horie et al., 2009; Shabala et al., 2010; Chérel et al., 2014).AKT1是参与K+吸收的重要通道蛋白, 在植物响应盐胁迫中起着重要作用.异源表达系统研究表明, 尽管AKT1对K+具有更强的选择性, 但并不排除对Na+的吸收(Maathuis et al., 1997).低Na+条件下, 内向整流K+通道未表现出明显的Na+进出现象; 但在高盐条件下, 其对K+表现出明显的选择性(Amtmann and Sanders, 1999).Goll- dack等(2003)研究表明, 盐处理下水稻根吸收Na+与OsAKT1的表达水平有直接关系.由此证明OsAKT1参与盐胁迫下Na+/K+比调控, 从而增强水稻的耐盐性.Wang等(2015)研究发现, 无论是低盐(25 mmol·L-1 NaCl)还是高盐(150 mmol·L-1 NaCl)条件下, 小花碱茅PtAKT1选择性吸收K+的能力显著强于Na+ (selective absorption capacity for K+ over Na+, SA).进一步研究发现, PtAKT1的转录水平与SA值之间呈显著正相关, 可见盐胁迫下PtAKT1是调控小花碱茅K+选择性吸收的关键因子(Wang et al., 2015).与野生型相比, 盐胁迫下超表达PtAKT1的拟南芥转基因植株K+含量增加, 而Na+含量降低, 表明PtAKT1增强了转基因植株对K+的吸收能力, 从而提高其耐盐性(Ardie et al., 2010).Duan等(2015)研究发现, 盐胁迫下盐地碱蓬SsAKT1能促进植株体内K+积累, 进而增强植物耐盐性.在盐(100 mmol·L-1 NaCl)处理下, 大麦HvAKT1在叶伸长区的表达水平明显增加, 说明HvAKT1参与维持叶肉细胞K+的稳态平衡(Boscari et al., 2009).我们的研究发现, 在5-50 mmol·L-1 NaCl处理下, 分别添加3和6 mmol·L-1 Cs+ (K+通道AKT1特异性抑制剂)均可显著降低甜菜(Beta vulgaris)根的K+净吸收速率, 可见AKT1可介导甜菜根K+的吸收(Wu et al., 2015b).这些研究结果表明, AKT1在K+和Na+选择性吸收、维持植物体内K+含量以及增强植物耐盐性等方面具有重要作用. ...
2 2000
... AKT1 (Arabidopsis K+ transporter 1)是植物中广泛存在的一类内向整流K+通道, 与动物中的Shaker型K+通道具有高度的同源性(Pilot et al., 2003).Sentenac等(1992)利用酵母双突变体互补法从拟南芥中克隆到第1个高等植物的Shaker型K+通道基因AtAKT1.该基因位于拟南芥第2号染色体上, 含有11个外显子和10个内含子, 编码的多肽由857个氨基酸残基组成.此后, 科研人员相继在马铃薯(Sola- num tuberosum) (Zimmermann et al., 1998)、玉米(Zea mays) (Philippar et al., 1999)、小麦(Triticum aestivum) (Buschmann et al., 2000)、番茄(Lycoper- sicon esculentum) (Hartje et al., 2000)、水稻(Oryza sativa) (Fuchs et al., 2005)、大麦(Hordeum vulgare) (Boscari et al., 2009)、葡萄(Vitis vinifera) (Cuéllar et al., 2010, 2013)和盐地碱蓬(Suaeda salsa) (Duan et al., 2015)等植物中分离得到AKT1基因.AKT1在维持植物离子稳态平衡及增强植物逆境胁迫耐受性等方面起着重要作用(Alemán et al., 2011; Wang and Wu, 2013).本文从植物AKT1的分子结构特点、调控机制及生物学功能等方面对其主要研究进展进行了归纳总结, 旨在为植物AKT1的进一步研究奠定基础. ... ... 在K+饥饿状态下, 小麦根TaAKT1 (Buschmann et al., 2000)和小花碱茅(Puccinellia tenuiflora) PtAKT1 (Ardie et al., 2010)的转录水平显著提高.在拟南芥中, 外部K+浓度在5 μmol·L-1至5 mmol·L-1范围内未影响AtAKT1的转录丰度(Lagarde et al., 1996).在棉花(Xu et al., 2014)和小花碱茅(Wang et al., 2015)中也有类似报道, 由此推测AKT1可能受转录后调控.然而, 25-250 mmol·L-1 NaCl短期(6小时)处理能够诱导盐地碱蓬根SsAKT1的表达(Duan et al., 2015).研究表明, 水稻OsAKT1主要在根的外皮层和内皮层细胞以及叶的木质部薄壁组织、韧皮部和叶肉细胞中表达(Golldack et al., 2003).盐胁迫(150 mmol·L-1 NaCl处理48小时)下, 盐敏感型水稻品种Pokkali和BK根外皮层细胞中的OsAKT1转录物消失, 而耐盐型品种IR29中的则未受到影响(Golldack et al., 2003).Su等(2001)的研究表明, 盐胁迫下盐生植物冰叶日中花(Mesembryanthemum crystallinum)根中与AtAKT1同源的K+通道基因McAKT1表达水平下调.在小花碱茅中, 尽管盐处理会下调PtAKT1的表达, 但150 mmol·L-1 NaCl处理下PtAKT1的转录丰度显著高于25 mmol·L-1 (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ...
1 2004
... 干旱是制约全球作物生长和产量的主要非生物因素之一(Chaves and Oliveira, 2004; 山仑, 2011).植物响应干旱胁迫的重要策略之一是吸收并在体内积累大量的溶质(如K+、Na+和脯氨酸等) (Wang et al., 2004; Mahouachi et al., 2006; Wu et al., 2015a), 降低细胞渗透势, 从而增强植物细胞吸水能力.AKT1在植物响应干旱胁迫中的作用主要有2方面.(1) 干旱胁迫下AKT1增强植物根的K+吸收能力.在水稻中超表达OsAKT1基因后, 转基因植株在干旱条件下生长良好, 且根中积累较多的K+; 相反, Osakt1突变体植物表现为生长不良, 且根中K+含量明显减少(Ahmad et al., 2016).由此表明, 超表达AKT1转基因植株通过增加根部K+的积累来改善细胞渗透调节能力, 从而增强植物的抗旱性.(2) AKT1通过调节气孔的运动影响植物体内的水分平衡.研究表明, 拟南芥AtAKT1在保卫细胞质膜上呈低丰度表达(Lagarde et al., 1996).在渗透胁迫下, 拟南芥Atakt1突变体植株的蒸腾速率和水分消耗量明显低于野生型植株(Nieves- Cordones et al., 2012).由此可见, AKT1功能缺失致使保卫细胞K+跨膜转运体系受损, 气孔导度下降.Ahmad等(2016)进一步研究发现, 与野生型相比, 在干旱胁迫下水稻Osakt1突变体植株的气孔导度明显降低; 而超表达OsAKT1显著增强了转基因植株的气孔导度.这些研究结果表明, AKT1在介导保卫细胞K+转运、调控气孔运动及维持细胞水分平衡中发挥重要作用. ...
1 2007
... 尽管自20世纪90年代从拟南芥中克隆到第1个AKT1基因以来, 人们相继在其它植物中克隆得到同源基因.然而, AKT1基因所编码蛋白的调控机制仍不清楚.Xu等(2006)首次在植物中报道了低K+条件下蛋白磷酸化对K+吸收的调控机制.在爪蟾卵母细胞(Xeno- pus oocytes)中, CBL1 (calcineurin B-like protein1)/ CBL9与CIPK23 (CBL-interacting protein kinases 23)互作, 通过磷酸化作用激活AKT1介导的K+吸收(Li et al., 2006; Xu et al., 2006; Cuéllar et al., 2010, 2013).低K+处理下, CIPK23表达上调, 而CBL1和CBL9似乎是组成型表达(Xu et al., 2006; Cheong et al., 2007); 过量表达CIPK23可显著增强拟南芥K+的吸收能力和低K+耐受性(Xu et al., 2006).CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ...
2 2014
... AtKC1是Reintanz等(2002)从拟南芥中分离克隆的α-亚基Shaker通道基因, 主要在根毛、根表皮及皮层细胞中表达.AtKC1作为四聚体的组成亚基参与K+通道四聚体的形成, 其存在对四聚体的K+通透能力产生抑制作用(Reintanz et al., 2002; Geiger et al., 2009).可见, AtKC1是负调控因子, 也是1个调控亚基(Wang et al., 2010).另外, AtKC1的存在使四聚体的K+通道活性激活电压向负电压方向偏移, 离子通道更难以被激活, 即改变了其对电压的敏感性(Duby et al., 2008).因此, 这种负向调控阻断了由AKT1介导的吸收.短时间缺K+处理则上调AtKC1的转录丰度, 随后其转录丰度下调(Shin and Schachtman, 2004), 这种现象可通过缺K+处理几天后的AtKC1转录物消失而得到解释(Pilot et al., 2003).在异源表达系统中, AtKC1不能独立形成有功能的K+通道(Duby et al., 2008).然而, 当AtKC1不能正常发挥功能时, 根毛中AKT1介导的内向K+电流的生物物理特征就会发生改变(Jeanguenin et al., 2011).Wang等(2016)进一步研究证实, 在低K+条件下, CIPK23促进AKT1介导的K+吸收, 而AtKC1抑制AKT1介导的K+内流.他们认为, CIPK23和AtKC1协同调节AKT1的功能, 二者具有一定的互补性.另有研究表明, Atkc1与Atakt1突变体植株的表型相似或相反, 这主要取决于外界环境条件(Geiger et al., 2009; Wang et al., 2010).AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009).低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似.在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014).最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014).PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性.可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014). ... ... 盐胁迫下, 植物体内积累大量的Na+, 一方面抑制K+的吸收, 另一方面与K+竞争一些酶活性结合位点, 影响蛋白质合成及核糖体功能, 从而产生Na+毒害(Horie et al., 2009; Zhang et al., 2010).一般认为, 维持细胞质内较高的K+/Na+比是植物适应盐胁迫的重要策略之一(Horie et al., 2009; Shabala et al., 2010; Chérel et al., 2014).AKT1是参与K+吸收的重要通道蛋白, 在植物响应盐胁迫中起着重要作用.异源表达系统研究表明, 尽管AKT1对K+具有更强的选择性, 但并不排除对Na+的吸收(Maathuis et al., 1997).低Na+条件下, 内向整流K+通道未表现出明显的Na+进出现象; 但在高盐条件下, 其对K+表现出明显的选择性(Amtmann and Sanders, 1999).Goll- dack等(2003)研究表明, 盐处理下水稻根吸收Na+与OsAKT1的表达水平有直接关系.由此证明OsAKT1参与盐胁迫下Na+/K+比调控, 从而增强水稻的耐盐性.Wang等(2015)研究发现, 无论是低盐(25 mmol·L-1 NaCl)还是高盐(150 mmol·L-1 NaCl)条件下, 小花碱茅PtAKT1选择性吸收K+的能力显著强于Na+ (selective absorption capacity for K+ over Na+, SA).进一步研究发现, PtAKT1的转录水平与SA值之间呈显著正相关, 可见盐胁迫下PtAKT1是调控小花碱茅K+选择性吸收的关键因子(Wang et al., 2015).与野生型相比, 盐胁迫下超表达PtAKT1的拟南芥转基因植株K+含量增加, 而Na+含量降低, 表明PtAKT1增强了转基因植株对K+的吸收能力, 从而提高其耐盐性(Ardie et al., 2010).Duan等(2015)研究发现, 盐胁迫下盐地碱蓬SsAKT1能促进植株体内K+积累, 进而增强植物耐盐性.在盐(100 mmol·L-1 NaCl)处理下, 大麦HvAKT1在叶伸长区的表达水平明显增加, 说明HvAKT1参与维持叶肉细胞K+的稳态平衡(Boscari et al., 2009).我们的研究发现, 在5-50 mmol·L-1 NaCl处理下, 分别添加3和6 mmol·L-1 Cs+ (K+通道AKT1特异性抑制剂)均可显著降低甜菜(Beta vulgaris)根的K+净吸收速率, 可见AKT1可介导甜菜根K+的吸收(Wu et al., 2015b).这些研究结果表明, AKT1在K+和Na+选择性吸收、维持植物体内K+含量以及增强植物耐盐性等方面具有重要作用. ...
3 2013
... AKT1 (Arabidopsis K+ transporter 1)是植物中广泛存在的一类内向整流K+通道, 与动物中的Shaker型K+通道具有高度的同源性(Pilot et al., 2003).Sentenac等(1992)利用酵母双突变体互补法从拟南芥中克隆到第1个高等植物的Shaker型K+通道基因AtAKT1.该基因位于拟南芥第2号染色体上, 含有11个外显子和10个内含子, 编码的多肽由857个氨基酸残基组成.此后, 科研人员相继在马铃薯(Sola- num tuberosum) (Zimmermann et al., 1998)、玉米(Zea mays) (Philippar et al., 1999)、小麦(Triticum aestivum) (Buschmann et al., 2000)、番茄(Lycoper- sicon esculentum) (Hartje et al., 2000)、水稻(Oryza sativa) (Fuchs et al., 2005)、大麦(Hordeum vulgare) (Boscari et al., 2009)、葡萄(Vitis vinifera) (Cuéllar et al., 2010, 2013)和盐地碱蓬(Suaeda salsa) (Duan et al., 2015)等植物中分离得到AKT1基因.AKT1在维持植物离子稳态平衡及增强植物逆境胁迫耐受性等方面起着重要作用(Alemán et al., 2011; Wang and Wu, 2013).本文从植物AKT1的分子结构特点、调控机制及生物学功能等方面对其主要研究进展进行了归纳总结, 旨在为植物AKT1的进一步研究奠定基础. ... ... 在K+饥饿状态下, 小麦根TaAKT1 (Buschmann et al., 2000)和小花碱茅(Puccinellia tenuiflora) PtAKT1 (Ardie et al., 2010)的转录水平显著提高.在拟南芥中, 外部K+浓度在5 μmol·L-1至5 mmol·L-1范围内未影响AtAKT1的转录丰度(Lagarde et al., 1996).在棉花(Xu et al., 2014)和小花碱茅(Wang et al., 2015)中也有类似报道, 由此推测AKT1可能受转录后调控.然而, 25-250 mmol·L-1 NaCl短期(6小时)处理能够诱导盐地碱蓬根SsAKT1的表达(Duan et al., 2015).研究表明, 水稻OsAKT1主要在根的外皮层和内皮层细胞以及叶的木质部薄壁组织、韧皮部和叶肉细胞中表达(Golldack et al., 2003).盐胁迫(150 mmol·L-1 NaCl处理48小时)下, 盐敏感型水稻品种Pokkali和BK根外皮层细胞中的OsAKT1转录物消失, 而耐盐型品种IR29中的则未受到影响(Golldack et al., 2003).Su等(2001)的研究表明, 盐胁迫下盐生植物冰叶日中花(Mesembryanthemum crystallinum)根中与AtAKT1同源的K+通道基因McAKT1表达水平下调.在小花碱茅中, 尽管盐处理会下调PtAKT1的表达, 但150 mmol·L-1 NaCl处理下PtAKT1的转录丰度显著高于25 mmol·L-1 (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ... ... 尽管自20世纪90年代从拟南芥中克隆到第1个AKT1基因以来, 人们相继在其它植物中克隆得到同源基因.然而, AKT1基因所编码蛋白的调控机制仍不清楚.Xu等(2006)首次在植物中报道了低K+条件下蛋白磷酸化对K+吸收的调控机制.在爪蟾卵母细胞(Xeno- pus oocytes)中, CBL1 (calcineurin B-like protein1)/ CBL9与CIPK23 (CBL-interacting protein kinases 23)互作, 通过磷酸化作用激活AKT1介导的K+吸收(Li et al., 2006; Xu et al., 2006; Cuéllar et al., 2010, 2013).低K+处理下, CIPK23表达上调, 而CBL1和CBL9似乎是组成型表达(Xu et al., 2006; Cheong et al., 2007); 过量表达CIPK23可显著增强拟南芥K+的吸收能力和低K+耐受性(Xu et al., 2006).CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ...
3 2010
... AKT1 (Arabidopsis K+ transporter 1)是植物中广泛存在的一类内向整流K+通道, 与动物中的Shaker型K+通道具有高度的同源性(Pilot et al., 2003).Sentenac等(1992)利用酵母双突变体互补法从拟南芥中克隆到第1个高等植物的Shaker型K+通道基因AtAKT1.该基因位于拟南芥第2号染色体上, 含有11个外显子和10个内含子, 编码的多肽由857个氨基酸残基组成.此后, 科研人员相继在马铃薯(Sola- num tuberosum) (Zimmermann et al., 1998)、玉米(Zea mays) (Philippar et al., 1999)、小麦(Triticum aestivum) (Buschmann et al., 2000)、番茄(Lycoper- sicon esculentum) (Hartje et al., 2000)、水稻(Oryza sativa) (Fuchs et al., 2005)、大麦(Hordeum vulgare) (Boscari et al., 2009)、葡萄(Vitis vinifera) (Cuéllar et al., 2010, 2013)和盐地碱蓬(Suaeda salsa) (Duan et al., 2015)等植物中分离得到AKT1基因.AKT1在维持植物离子稳态平衡及增强植物逆境胁迫耐受性等方面起着重要作用(Alemán et al., 2011; Wang and Wu, 2013).本文从植物AKT1的分子结构特点、调控机制及生物学功能等方面对其主要研究进展进行了归纳总结, 旨在为植物AKT1的进一步研究奠定基础. ... ... 在K+饥饿状态下, 小麦根TaAKT1 (Buschmann et al., 2000)和小花碱茅(Puccinellia tenuiflora) PtAKT1 (Ardie et al., 2010)的转录水平显著提高.在拟南芥中, 外部K+浓度在5 μmol·L-1至5 mmol·L-1范围内未影响AtAKT1的转录丰度(Lagarde et al., 1996).在棉花(Xu et al., 2014)和小花碱茅(Wang et al., 2015)中也有类似报道, 由此推测AKT1可能受转录后调控.然而, 25-250 mmol·L-1 NaCl短期(6小时)处理能够诱导盐地碱蓬根SsAKT1的表达(Duan et al., 2015).研究表明, 水稻OsAKT1主要在根的外皮层和内皮层细胞以及叶的木质部薄壁组织、韧皮部和叶肉细胞中表达(Golldack et al., 2003).盐胁迫(150 mmol·L-1 NaCl处理48小时)下, 盐敏感型水稻品种Pokkali和BK根外皮层细胞中的OsAKT1转录物消失, 而耐盐型品种IR29中的则未受到影响(Golldack et al., 2003).Su等(2001)的研究表明, 盐胁迫下盐生植物冰叶日中花(Mesembryanthemum crystallinum)根中与AtAKT1同源的K+通道基因McAKT1表达水平下调.在小花碱茅中, 尽管盐处理会下调PtAKT1的表达, 但150 mmol·L-1 NaCl处理下PtAKT1的转录丰度显著高于25 mmol·L-1 (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ... ... 尽管自20世纪90年代从拟南芥中克隆到第1个AKT1基因以来, 人们相继在其它植物中克隆得到同源基因.然而, AKT1基因所编码蛋白的调控机制仍不清楚.Xu等(2006)首次在植物中报道了低K+条件下蛋白磷酸化对K+吸收的调控机制.在爪蟾卵母细胞(Xeno- pus oocytes)中, CBL1 (calcineurin B-like protein1)/ CBL9与CIPK23 (CBL-interacting protein kinases 23)互作, 通过磷酸化作用激活AKT1介导的K+吸收(Li et al., 2006; Xu et al., 2006; Cuéllar et al., 2010, 2013).低K+处理下, CIPK23表达上调, 而CBL1和CBL9似乎是组成型表达(Xu et al., 2006; Cheong et al., 2007); 过量表达CIPK23可显著增强拟南芥K+的吸收能力和低K+耐受性(Xu et al., 2006).CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ...
2 2014
... 植物在长期进化过程中形成了一系列复杂的K+吸收和转运系统.已知在模式植物拟南芥(Arabi- dopsis thaliana)中存在15个K+选择性通道, 包括9个Shaker型、5个TPK (two-pore K+ channel)型和1个Kir-like型K+通道(Ward et al., 2009; Hedrich, 2012; Demidchik, 2014).另外, 一些非选择性阳离子通道(non-selective cation channels, NSCC)也可以通透K+, 主要包括20个CNGC (cyclic nucleotide gated channels)通道、20个GLR (glutamate receptor-like)通道和1个TPC (two-pore channel)通道(Shabala, 2003; Demidchik and Maathuis, 2007; Dietrich et al., 2010; Hedrich, 2012; Demidchik, 2014).植物K+转运蛋白主要包括KUP (K+ uptake permease)/HAK (high-affinity K+ transporter)/KT (K+ transporter)、HKT (high-affinity K+ transporter)和CPA (cation proton antiporter) (Gierth and Mäser, 2007; Kron- zucker and Britto, 2011; Wang and Wu, 2013; Han et al., 2016).近年来, 人们相继在植物中克隆到编码这些蛋白的基因, 为深入研究植物K+吸收机制奠定了坚实的基础. ... ... ; Demidchik, 2014).植物K+转运蛋白主要包括KUP (K+ uptake permease)/HAK (high-affinity K+ transporter)/KT (K+ transporter)、HKT (high-affinity K+ transporter)和CPA (cation proton antiporter) (Gierth and Mäser, 2007; Kron- zucker and Britto, 2011; Wang and Wu, 2013; Han et al., 2016).近年来, 人们相继在植物中克隆到编码这些蛋白的基因, 为深入研究植物K+吸收机制奠定了坚实的基础. ...
1 2007
... 植物在长期进化过程中形成了一系列复杂的K+吸收和转运系统.已知在模式植物拟南芥(Arabi- dopsis thaliana)中存在15个K+选择性通道, 包括9个Shaker型、5个TPK (two-pore K+ channel)型和1个Kir-like型K+通道(Ward et al., 2009; Hedrich, 2012; Demidchik, 2014).另外, 一些非选择性阳离子通道(non-selective cation channels, NSCC)也可以通透K+, 主要包括20个CNGC (cyclic nucleotide gated channels)通道、20个GLR (glutamate receptor-like)通道和1个TPC (two-pore channel)通道(Shabala, 2003; Demidchik and Maathuis, 2007; Dietrich et al., 2010; Hedrich, 2012; Demidchik, 2014).植物K+转运蛋白主要包括KUP (K+ uptake permease)/HAK (high-affinity K+ transporter)/KT (K+ transporter)、HKT (high-affinity K+ transporter)和CPA (cation proton antiporter) (Gierth and Mäser, 2007; Kron- zucker and Britto, 2011; Wang and Wu, 2013; Han et al., 2016).近年来, 人们相继在植物中克隆到编码这些蛋白的基因, 为深入研究植物K+吸收机制奠定了坚实的基础. ...
1 2010
... 植物在长期进化过程中形成了一系列复杂的K+吸收和转运系统.已知在模式植物拟南芥(Arabi- dopsis thaliana)中存在15个K+选择性通道, 包括9个Shaker型、5个TPK (two-pore K+ channel)型和1个Kir-like型K+通道(Ward et al., 2009; Hedrich, 2012; Demidchik, 2014).另外, 一些非选择性阳离子通道(non-selective cation channels, NSCC)也可以通透K+, 主要包括20个CNGC (cyclic nucleotide gated channels)通道、20个GLR (glutamate receptor-like)通道和1个TPC (two-pore channel)通道(Shabala, 2003; Demidchik and Maathuis, 2007; Dietrich et al., 2010; Hedrich, 2012; Demidchik, 2014).植物K+转运蛋白主要包括KUP (K+ uptake permease)/HAK (high-affinity K+ transporter)/KT (K+ transporter)、HKT (high-affinity K+ transporter)和CPA (cation proton antiporter) (Gierth and Mäser, 2007; Kron- zucker and Britto, 2011; Wang and Wu, 2013; Han et al., 2016).近年来, 人们相继在植物中克隆到编码这些蛋白的基因, 为深入研究植物K+吸收机制奠定了坚实的基础. ...
1 2009
... 植物AKT1与动物Shaker型K+通道结构高度相似(图1), 含有6个跨膜片段(S1-S6), 其中第4个跨膜片段S4含有大量带正电荷的氨基酸残基, 是电压感受器, 其主要功能为响应膜电势的变化.该片段可在膜上移动, 使得膜通道构象改变, 从而控制通道孔的开放与关闭(Benito et al., 2014; Nieves-Cordones and Gaillard, 2014).S5和S6之间含有1个高度保守的P环结构域, 该结构域是陷入细胞膜内的一段多肽片段, 构成通道孔.AKT1含有典型的TxGYG (Thr-X- Gly-Tyr-Gly)序列, 为K+选择器, 这是Shaker型K+通道的共同特点.胞质C末端从第6个跨膜片段末尾起, 含有1个C-接头(约有80个氨基酸残基), 1个环核苷酸结合域(cyclic-nucleotide binding domain, CNBD), 1个锚蛋白域和1个富含疏水性、酸性残基的KHA域(Nieves-Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).对大多数Shaker型K+通道家族成员来说, 锚蛋白域有助于通道与细胞骨架的连接、蛋白质的相互作用和细胞溶质的调节(Dreyer and Blatt, 2009; Riedelsberger et al., 2010; Nieves- Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).Shaker型K+通道的重要特点之一是能够形成异源四聚体结构, 可使植物调控不同细胞中的K+转运活性.这种调控在每个器官或者组织中相对独立, 并且受到环境的影响(Gambale and Uozumi, 2006). ...
1 2011
... K+是植物细胞中含量最丰富的无机阳离子之一, 主要通过根表皮细胞和皮层细胞的质膜K+转运系统吸收并运输到植物体内(王毅和武维华, 2009; Alemán et al., 2011; Dreyer and Uozumi, 2011; Wang and Wu, 2013; Nieves-Cordones et al., 2014a).K+在植物生长发育、逆境胁迫响应及生理生化反应等方面起着重要作用(Alemán et al., 2011; Nieves-Cordones et al., 2014a).一般而言, 植物细胞维持正常新陈代谢所需K+的最佳浓度为100 mmol·L-1, 但土壤中能被植物利用的K+浓度仅为0.1-1 mmol·L-1, 在一些盐碱地中甚至更低(Maathuis and Sanders, 1993; Maathuis, 2009).因此, 土壤中的K+远远不能满足植物正常生长需要, 使植物经常处于K+饥饿状态, 严重影响其生长发育.鉴于复杂多变的环境及K+的重要作用, 众多研究者对植物K+吸收和转运机制进行了深入研究. ...
2 2015
... AKT1 (Arabidopsis K+ transporter 1)是植物中广泛存在的一类内向整流K+通道, 与动物中的Shaker型K+通道具有高度的同源性(Pilot et al., 2003).Sentenac等(1992)利用酵母双突变体互补法从拟南芥中克隆到第1个高等植物的Shaker型K+通道基因AtAKT1.该基因位于拟南芥第2号染色体上, 含有11个外显子和10个内含子, 编码的多肽由857个氨基酸残基组成.此后, 科研人员相继在马铃薯(Sola- num tuberosum) (Zimmermann et al., 1998)、玉米(Zea mays) (Philippar et al., 1999)、小麦(Triticum aestivum) (Buschmann et al., 2000)、番茄(Lycoper- sicon esculentum) (Hartje et al., 2000)、水稻(Oryza sativa) (Fuchs et al., 2005)、大麦(Hordeum vulgare) (Boscari et al., 2009)、葡萄(Vitis vinifera) (Cuéllar et al., 2010, 2013)和盐地碱蓬(Suaeda salsa) (Duan et al., 2015)等植物中分离得到AKT1基因.AKT1在维持植物离子稳态平衡及增强植物逆境胁迫耐受性等方面起着重要作用(Alemán et al., 2011; Wang and Wu, 2013).本文从植物AKT1的分子结构特点、调控机制及生物学功能等方面对其主要研究进展进行了归纳总结, 旨在为植物AKT1的进一步研究奠定基础. ... ... 在K+饥饿状态下, 小麦根TaAKT1 (Buschmann et al., 2000)和小花碱茅(Puccinellia tenuiflora) PtAKT1 (Ardie et al., 2010)的转录水平显著提高.在拟南芥中, 外部K+浓度在5 μmol·L-1至5 mmol·L-1范围内未影响AtAKT1的转录丰度(Lagarde et al., 1996).在棉花(Xu et al., 2014)和小花碱茅(Wang et al., 2015)中也有类似报道, 由此推测AKT1可能受转录后调控.然而, 25-250 mmol·L-1 NaCl短期(6小时)处理能够诱导盐地碱蓬根SsAKT1的表达(Duan et al., 2015).研究表明, 水稻OsAKT1主要在根的外皮层和内皮层细胞以及叶的木质部薄壁组织、韧皮部和叶肉细胞中表达(Golldack et al., 2003).盐胁迫(150 mmol·L-1 NaCl处理48小时)下, 盐敏感型水稻品种Pokkali和BK根外皮层细胞中的OsAKT1转录物消失, 而耐盐型品种IR29中的则未受到影响(Golldack et al., 2003).Su等(2001)的研究表明, 盐胁迫下盐生植物冰叶日中花(Mesembryanthemum crystallinum)根中与AtAKT1同源的K+通道基因McAKT1表达水平下调.在小花碱茅中, 尽管盐处理会下调PtAKT1的表达, 但150 mmol·L-1 NaCl处理下PtAKT1的转录丰度显著高于25 mmol·L-1 (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ...
2 2008
... AtKC1是Reintanz等(2002)从拟南芥中分离克隆的α-亚基Shaker通道基因, 主要在根毛、根表皮及皮层细胞中表达.AtKC1作为四聚体的组成亚基参与K+通道四聚体的形成, 其存在对四聚体的K+通透能力产生抑制作用(Reintanz et al., 2002; Geiger et al., 2009).可见, AtKC1是负调控因子, 也是1个调控亚基(Wang et al., 2010).另外, AtKC1的存在使四聚体的K+通道活性激活电压向负电压方向偏移, 离子通道更难以被激活, 即改变了其对电压的敏感性(Duby et al., 2008).因此, 这种负向调控阻断了由AKT1介导的吸收.短时间缺K+处理则上调AtKC1的转录丰度, 随后其转录丰度下调(Shin and Schachtman, 2004), 这种现象可通过缺K+处理几天后的AtKC1转录物消失而得到解释(Pilot et al., 2003).在异源表达系统中, AtKC1不能独立形成有功能的K+通道(Duby et al., 2008).然而, 当AtKC1不能正常发挥功能时, 根毛中AKT1介导的内向K+电流的生物物理特征就会发生改变(Jeanguenin et al., 2011).Wang等(2016)进一步研究证实, 在低K+条件下, CIPK23促进AKT1介导的K+吸收, 而AtKC1抑制AKT1介导的K+内流.他们认为, CIPK23和AtKC1协同调节AKT1的功能, 二者具有一定的互补性.另有研究表明, Atkc1与Atakt1突变体植株的表型相似或相反, 这主要取决于外界环境条件(Geiger et al., 2009; Wang et al., 2010).AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009).低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似.在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014).最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014).PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性.可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014). ... ... 通道(Duby et al., 2008).然而, 当AtKC1不能正常发挥功能时, 根毛中AKT1介导的内向K+电流的生物物理特征就会发生改变(Jeanguenin et al., 2011).Wang等(2016)进一步研究证实, 在低K+条件下, CIPK23促进AKT1介导的K+吸收, 而AtKC1抑制AKT1介导的K+内流.他们认为, CIPK23和AtKC1协同调节AKT1的功能, 二者具有一定的互补性.另有研究表明, Atkc1与Atakt1突变体植株的表型相似或相反, 这主要取决于外界环境条件(Geiger et al., 2009; Wang et al., 2010).AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009).低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似.在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014).最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014).PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性.可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014). ...
... AKT1 (Arabidopsis K+ transporter 1)是植物中广泛存在的一类内向整流K+通道, 与动物中的Shaker型K+通道具有高度的同源性(Pilot et al., 2003).Sentenac等(1992)利用酵母双突变体互补法从拟南芥中克隆到第1个高等植物的Shaker型K+通道基因AtAKT1.该基因位于拟南芥第2号染色体上, 含有11个外显子和10个内含子, 编码的多肽由857个氨基酸残基组成.此后, 科研人员相继在马铃薯(Sola- num tuberosum) (Zimmermann et al., 1998)、玉米(Zea mays) (Philippar et al., 1999)、小麦(Triticum aestivum) (Buschmann et al., 2000)、番茄(Lycoper- sicon esculentum) (Hartje et al., 2000)、水稻(Oryza sativa) (Fuchs et al., 2005)、大麦(Hordeum vulgare) (Boscari et al., 2009)、葡萄(Vitis vinifera) (Cuéllar et al., 2010, 2013)和盐地碱蓬(Suaeda salsa) (Duan et al., 2015)等植物中分离得到AKT1基因.AKT1在维持植物离子稳态平衡及增强植物逆境胁迫耐受性等方面起着重要作用(Alemán et al., 2011; Wang and Wu, 2013).本文从植物AKT1的分子结构特点、调控机制及生物学功能等方面对其主要研究进展进行了归纳总结, 旨在为植物AKT1的进一步研究奠定基础. ...
1 2006
... 植物AKT1与动物Shaker型K+通道结构高度相似(图1), 含有6个跨膜片段(S1-S6), 其中第4个跨膜片段S4含有大量带正电荷的氨基酸残基, 是电压感受器, 其主要功能为响应膜电势的变化.该片段可在膜上移动, 使得膜通道构象改变, 从而控制通道孔的开放与关闭(Benito et al., 2014; Nieves-Cordones and Gaillard, 2014).S5和S6之间含有1个高度保守的P环结构域, 该结构域是陷入细胞膜内的一段多肽片段, 构成通道孔.AKT1含有典型的TxGYG (Thr-X- Gly-Tyr-Gly)序列, 为K+选择器, 这是Shaker型K+通道的共同特点.胞质C末端从第6个跨膜片段末尾起, 含有1个C-接头(约有80个氨基酸残基), 1个环核苷酸结合域(cyclic-nucleotide binding domain, CNBD), 1个锚蛋白域和1个富含疏水性、酸性残基的KHA域(Nieves-Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).对大多数Shaker型K+通道家族成员来说, 锚蛋白域有助于通道与细胞骨架的连接、蛋白质的相互作用和细胞溶质的调节(Dreyer and Blatt, 2009; Riedelsberger et al., 2010; Nieves- Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).Shaker型K+通道的重要特点之一是能够形成异源四聚体结构, 可使植物调控不同细胞中的K+转运活性.这种调控在每个器官或者组织中相对独立, 并且受到环境的影响(Gambale and Uozumi, 2006). ...
2 2009
... AtKC1是Reintanz等(2002)从拟南芥中分离克隆的α-亚基Shaker通道基因, 主要在根毛、根表皮及皮层细胞中表达.AtKC1作为四聚体的组成亚基参与K+通道四聚体的形成, 其存在对四聚体的K+通透能力产生抑制作用(Reintanz et al., 2002; Geiger et al., 2009).可见, AtKC1是负调控因子, 也是1个调控亚基(Wang et al., 2010).另外, AtKC1的存在使四聚体的K+通道活性激活电压向负电压方向偏移, 离子通道更难以被激活, 即改变了其对电压的敏感性(Duby et al., 2008).因此, 这种负向调控阻断了由AKT1介导的吸收.短时间缺K+处理则上调AtKC1的转录丰度, 随后其转录丰度下调(Shin and Schachtman, 2004), 这种现象可通过缺K+处理几天后的AtKC1转录物消失而得到解释(Pilot et al., 2003).在异源表达系统中, AtKC1不能独立形成有功能的K+通道(Duby et al., 2008).然而, 当AtKC1不能正常发挥功能时, 根毛中AKT1介导的内向K+电流的生物物理特征就会发生改变(Jeanguenin et al., 2011).Wang等(2016)进一步研究证实, 在低K+条件下, CIPK23促进AKT1介导的K+吸收, 而AtKC1抑制AKT1介导的K+内流.他们认为, CIPK23和AtKC1协同调节AKT1的功能, 二者具有一定的互补性.另有研究表明, Atkc1与Atakt1突变体植株的表型相似或相反, 这主要取决于外界环境条件(Geiger et al., 2009; Wang et al., 2010).AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009).低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似.在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014).最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014).PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性.可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014). ... ... 突变体植株的表型相似或相反, 这主要取决于外界环境条件(Geiger et al., 2009; Wang et al., 2010).AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009).低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似.在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014).最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014).PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性.可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014). ...
1 2007
... 植物在长期进化过程中形成了一系列复杂的K+吸收和转运系统.已知在模式植物拟南芥(Arabi- dopsis thaliana)中存在15个K+选择性通道, 包括9个Shaker型、5个TPK (two-pore K+ channel)型和1个Kir-like型K+通道(Ward et al., 2009; Hedrich, 2012; Demidchik, 2014).另外, 一些非选择性阳离子通道(non-selective cation channels, NSCC)也可以通透K+, 主要包括20个CNGC (cyclic nucleotide gated channels)通道、20个GLR (glutamate receptor-like)通道和1个TPC (two-pore channel)通道(Shabala, 2003; Demidchik and Maathuis, 2007; Dietrich et al., 2010; Hedrich, 2012; Demidchik, 2014).植物K+转运蛋白主要包括KUP (K+ uptake permease)/HAK (high-affinity K+ transporter)/KT (K+ transporter)、HKT (high-affinity K+ transporter)和CPA (cation proton antiporter) (Gierth and Mäser, 2007; Kron- zucker and Britto, 2011; Wang and Wu, 2013; Han et al., 2016).近年来, 人们相继在植物中克隆到编码这些蛋白的基因, 为深入研究植物K+吸收机制奠定了坚实的基础. ...
2 2003
... 在K+饥饿状态下, 小麦根TaAKT1 (Buschmann et al., 2000)和小花碱茅(Puccinellia tenuiflora) PtAKT1 (Ardie et al., 2010)的转录水平显著提高.在拟南芥中, 外部K+浓度在5 μmol·L-1至5 mmol·L-1范围内未影响AtAKT1的转录丰度(Lagarde et al., 1996).在棉花(Xu et al., 2014)和小花碱茅(Wang et al., 2015)中也有类似报道, 由此推测AKT1可能受转录后调控.然而, 25-250 mmol·L-1 NaCl短期(6小时)处理能够诱导盐地碱蓬根SsAKT1的表达(Duan et al., 2015).研究表明, 水稻OsAKT1主要在根的外皮层和内皮层细胞以及叶的木质部薄壁组织、韧皮部和叶肉细胞中表达(Golldack et al., 2003).盐胁迫(150 mmol·L-1 NaCl处理48小时)下, 盐敏感型水稻品种Pokkali和BK根外皮层细胞中的OsAKT1转录物消失, 而耐盐型品种IR29中的则未受到影响(Golldack et al., 2003).Su等(2001)的研究表明, 盐胁迫下盐生植物冰叶日中花(Mesembryanthemum crystallinum)根中与AtAKT1同源的K+通道基因McAKT1表达水平下调.在小花碱茅中, 尽管盐处理会下调PtAKT1的表达, 但150 mmol·L-1 NaCl处理下PtAKT1的转录丰度显著高于25 mmol·L-1 (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ... ... 转录物消失, 而耐盐型品种IR29中的则未受到影响(Golldack et al., 2003).Su等(2001)的研究表明, 盐胁迫下盐生植物冰叶日中花(Mesembryanthemum crystallinum)根中与AtAKT1同源的K+通道基因McAKT1表达水平下调.在小花碱茅中, 尽管盐处理会下调PtAKT1的表达, 但150 mmol·L-1 NaCl处理下PtAKT1的转录丰度显著高于25 mmol·L-1 (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ...
1 2012
... 尽管自20世纪90年代从拟南芥中克隆到第1个AKT1基因以来, 人们相继在其它植物中克隆得到同源基因.然而, AKT1基因所编码蛋白的调控机制仍不清楚.Xu等(2006)首次在植物中报道了低K+条件下蛋白磷酸化对K+吸收的调控机制.在爪蟾卵母细胞(Xeno- pus oocytes)中, CBL1 (calcineurin B-like protein1)/ CBL9与CIPK23 (CBL-interacting protein kinases 23)互作, 通过磷酸化作用激活AKT1介导的K+吸收(Li et al., 2006; Xu et al., 2006; Cuéllar et al., 2010, 2013).低K+处理下, CIPK23表达上调, 而CBL1和CBL9似乎是组成型表达(Xu et al., 2006; Cheong et al., 2007); 过量表达CIPK23可显著增强拟南芥K+的吸收能力和低K+耐受性(Xu et al., 2006).CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ...
1 2016
... 植物在长期进化过程中形成了一系列复杂的K+吸收和转运系统.已知在模式植物拟南芥(Arabi- dopsis thaliana)中存在15个K+选择性通道, 包括9个Shaker型、5个TPK (two-pore K+ channel)型和1个Kir-like型K+通道(Ward et al., 2009; Hedrich, 2012; Demidchik, 2014).另外, 一些非选择性阳离子通道(non-selective cation channels, NSCC)也可以通透K+, 主要包括20个CNGC (cyclic nucleotide gated channels)通道、20个GLR (glutamate receptor-like)通道和1个TPC (two-pore channel)通道(Shabala, 2003; Demidchik and Maathuis, 2007; Dietrich et al., 2010; Hedrich, 2012; Demidchik, 2014).植物K+转运蛋白主要包括KUP (K+ uptake permease)/HAK (high-affinity K+ transporter)/KT (K+ transporter)、HKT (high-affinity K+ transporter)和CPA (cation proton antiporter) (Gierth and Mäser, 2007; Kron- zucker and Britto, 2011; Wang and Wu, 2013; Han et al., 2016).近年来, 人们相继在植物中克隆到编码这些蛋白的基因, 为深入研究植物K+吸收机制奠定了坚实的基础. ...
1 2000
... AKT1 (Arabidopsis K+ transporter 1)是植物中广泛存在的一类内向整流K+通道, 与动物中的Shaker型K+通道具有高度的同源性(Pilot et al., 2003).Sentenac等(1992)利用酵母双突变体互补法从拟南芥中克隆到第1个高等植物的Shaker型K+通道基因AtAKT1.该基因位于拟南芥第2号染色体上, 含有11个外显子和10个内含子, 编码的多肽由857个氨基酸残基组成.此后, 科研人员相继在马铃薯(Sola- num tuberosum) (Zimmermann et al., 1998)、玉米(Zea mays) (Philippar et al., 1999)、小麦(Triticum aestivum) (Buschmann et al., 2000)、番茄(Lycoper- sicon esculentum) (Hartje et al., 2000)、水稻(Oryza sativa) (Fuchs et al., 2005)、大麦(Hordeum vulgare) (Boscari et al., 2009)、葡萄(Vitis vinifera) (Cuéllar et al., 2010, 2013)和盐地碱蓬(Suaeda salsa) (Duan et al., 2015)等植物中分离得到AKT1基因.AKT1在维持植物离子稳态平衡及增强植物逆境胁迫耐受性等方面起着重要作用(Alemán et al., 2011; Wang and Wu, 2013).本文从植物AKT1的分子结构特点、调控机制及生物学功能等方面对其主要研究进展进行了归纳总结, 旨在为植物AKT1的进一步研究奠定基础. ...
2 2012
... 植物在长期进化过程中形成了一系列复杂的K+吸收和转运系统.已知在模式植物拟南芥(Arabi- dopsis thaliana)中存在15个K+选择性通道, 包括9个Shaker型、5个TPK (two-pore K+ channel)型和1个Kir-like型K+通道(Ward et al., 2009; Hedrich, 2012; Demidchik, 2014).另外, 一些非选择性阳离子通道(non-selective cation channels, NSCC)也可以通透K+, 主要包括20个CNGC (cyclic nucleotide gated channels)通道、20个GLR (glutamate receptor-like)通道和1个TPC (two-pore channel)通道(Shabala, 2003; Demidchik and Maathuis, 2007; Dietrich et al., 2010; Hedrich, 2012; Demidchik, 2014).植物K+转运蛋白主要包括KUP (K+ uptake permease)/HAK (high-affinity K+ transporter)/KT (K+ transporter)、HKT (high-affinity K+ transporter)和CPA (cation proton antiporter) (Gierth and Mäser, 2007; Kron- zucker and Britto, 2011; Wang and Wu, 2013; Han et al., 2016).近年来, 人们相继在植物中克隆到编码这些蛋白的基因, 为深入研究植物K+吸收机制奠定了坚实的基础. ... ... ; Hedrich, 2012; Demidchik, 2014).植物K+转运蛋白主要包括KUP (K+ uptake permease)/HAK (high-affinity K+ transporter)/KT (K+ transporter)、HKT (high-affinity K+ transporter)和CPA (cation proton antiporter) (Gierth and Mäser, 2007; Kron- zucker and Britto, 2011; Wang and Wu, 2013; Han et al., 2016).近年来, 人们相继在植物中克隆到编码这些蛋白的基因, 为深入研究植物K+吸收机制奠定了坚实的基础. ...
2 1998
... 低K+环境是影响植物正常生长发育及作物品质的主要非生物因子之一.K+是种子萌发所需的重要渗透调节物质.在低K+ (50-100 μmol·L-1)条件下, 拟南芥Atakt1突变体种子的胚根不能正常生长; 然而, 当在Atakt1突变体中超表达AtAKT1后, 其种子萌发活力得到恢复, 说明AKT1参与种子萌发期间K+的吸收(Pyo et al., 2010; Xu et al., 2014).AKT1功能缺失使植物根细胞膜表现出超级化, 对外界环境K+浓度变化异常敏感(Hirsch et al., 1998).在低K+环境中, 拟南芥Atakt1突变体植株细胞内的K+含量明显降低(Hirsch et al., 1998; Spalding et al., 1999).电生理学实验表明, 水稻OsAKT1介导内向K+电流, 即使在低K+ (5 μmol·L-1-1 mmol·L-1)条件下也是如此(Li et al., 2014).有研究表明, 拟南芥Atakt1和水稻Osakt1突变体植株的内向K+电流减弱, K+吸收功能紊乱, 从而影响其生长发育(Reintanz et al., 2002; Xu et al., 2006; Li et al., 2014).当在水稻Osakt1突变体中超表达OsAKT1后, 低K+胁迫下转基因植株的内向K+电流增强, K+含量显著增加(Li et al., 2014).这进一步证实了AKT1介导植物根吸收K+.Xu等(2014)采用非损伤微测技术(no-invasive micro-test technology, NMT), 在棉花GhAKT1功能研究中也得到类似的结果.可见, 在低K+环境中, AKT1介导的K+吸收系统对维持植物正常生长发育具有重要作用. ... ... 含量明显降低(Hirsch et al., 1998; Spalding et al., 1999).电生理学实验表明, 水稻OsAKT1介导内向K+电流, 即使在低K+ (5 μmol·L-1-1 mmol·L-1)条件下也是如此(Li et al., 2014).有研究表明, 拟南芥Atakt1和水稻Osakt1突变体植株的内向K+电流减弱, K+吸收功能紊乱, 从而影响其生长发育(Reintanz et al., 2002; Xu et al., 2006; Li et al., 2014).当在水稻Osakt1突变体中超表达OsAKT1后, 低K+胁迫下转基因植株的内向K+电流增强, K+含量显著增加(Li et al., 2014).这进一步证实了AKT1介导植物根吸收K+.Xu等(2014)采用非损伤微测技术(no-invasive micro-test technology, NMT), 在棉花GhAKT1功能研究中也得到类似的结果.可见, 在低K+环境中, AKT1介导的K+吸收系统对维持植物正常生长发育具有重要作用. ...
1 2009
... AtKC1是Reintanz等(2002)从拟南芥中分离克隆的α-亚基Shaker通道基因, 主要在根毛、根表皮及皮层细胞中表达.AtKC1作为四聚体的组成亚基参与K+通道四聚体的形成, 其存在对四聚体的K+通透能力产生抑制作用(Reintanz et al., 2002; Geiger et al., 2009).可见, AtKC1是负调控因子, 也是1个调控亚基(Wang et al., 2010).另外, AtKC1的存在使四聚体的K+通道活性激活电压向负电压方向偏移, 离子通道更难以被激活, 即改变了其对电压的敏感性(Duby et al., 2008).因此, 这种负向调控阻断了由AKT1介导的吸收.短时间缺K+处理则上调AtKC1的转录丰度, 随后其转录丰度下调(Shin and Schachtman, 2004), 这种现象可通过缺K+处理几天后的AtKC1转录物消失而得到解释(Pilot et al., 2003).在异源表达系统中, AtKC1不能独立形成有功能的K+通道(Duby et al., 2008).然而, 当AtKC1不能正常发挥功能时, 根毛中AKT1介导的内向K+电流的生物物理特征就会发生改变(Jeanguenin et al., 2011).Wang等(2016)进一步研究证实, 在低K+条件下, CIPK23促进AKT1介导的K+吸收, 而AtKC1抑制AKT1介导的K+内流.他们认为, CIPK23和AtKC1协同调节AKT1的功能, 二者具有一定的互补性.另有研究表明, Atkc1与Atakt1突变体植株的表型相似或相反, 这主要取决于外界环境条件(Geiger et al., 2009; Wang et al., 2010).AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009).低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似.在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014).最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014).PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性.可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014). ...
2 2009
... 盐胁迫下, 植物体内积累大量的Na+, 一方面抑制K+的吸收, 另一方面与K+竞争一些酶活性结合位点, 影响蛋白质合成及核糖体功能, 从而产生Na+毒害(Horie et al., 2009; Zhang et al., 2010).一般认为, 维持细胞质内较高的K+/Na+比是植物适应盐胁迫的重要策略之一(Horie et al., 2009; Shabala et al., 2010; Chérel et al., 2014).AKT1是参与K+吸收的重要通道蛋白, 在植物响应盐胁迫中起着重要作用.异源表达系统研究表明, 尽管AKT1对K+具有更强的选择性, 但并不排除对Na+的吸收(Maathuis et al., 1997).低Na+条件下, 内向整流K+通道未表现出明显的Na+进出现象; 但在高盐条件下, 其对K+表现出明显的选择性(Amtmann and Sanders, 1999).Goll- dack等(2003)研究表明, 盐处理下水稻根吸收Na+与OsAKT1的表达水平有直接关系.由此证明OsAKT1参与盐胁迫下Na+/K+比调控, 从而增强水稻的耐盐性.Wang等(2015)研究发现, 无论是低盐(25 mmol·L-1 NaCl)还是高盐(150 mmol·L-1 NaCl)条件下, 小花碱茅PtAKT1选择性吸收K+的能力显著强于Na+ (selective absorption capacity for K+ over Na+, SA).进一步研究发现, PtAKT1的转录水平与SA值之间呈显著正相关, 可见盐胁迫下PtAKT1是调控小花碱茅K+选择性吸收的关键因子(Wang et al., 2015).与野生型相比, 盐胁迫下超表达PtAKT1的拟南芥转基因植株K+含量增加, 而Na+含量降低, 表明PtAKT1增强了转基因植株对K+的吸收能力, 从而提高其耐盐性(Ardie et al., 2010).Duan等(2015)研究发现, 盐胁迫下盐地碱蓬SsAKT1能促进植株体内K+积累, 进而增强植物耐盐性.在盐(100 mmol·L-1 NaCl)处理下, 大麦HvAKT1在叶伸长区的表达水平明显增加, 说明HvAKT1参与维持叶肉细胞K+的稳态平衡(Boscari et al., 2009).我们的研究发现, 在5-50 mmol·L-1 NaCl处理下, 分别添加3和6 mmol·L-1 Cs+ (K+通道AKT1特异性抑制剂)均可显著降低甜菜(Beta vulgaris)根的K+净吸收速率, 可见AKT1可介导甜菜根K+的吸收(Wu et al., 2015b).这些研究结果表明, AKT1在K+和Na+选择性吸收、维持植物体内K+含量以及增强植物耐盐性等方面具有重要作用. ... ... 比是植物适应盐胁迫的重要策略之一(Horie et al., 2009; Shabala et al., 2010; Chérel et al., 2014).AKT1是参与K+吸收的重要通道蛋白, 在植物响应盐胁迫中起着重要作用.异源表达系统研究表明, 尽管AKT1对K+具有更强的选择性, 但并不排除对Na+的吸收(Maathuis et al., 1997).低Na+条件下, 内向整流K+通道未表现出明显的Na+进出现象; 但在高盐条件下, 其对K+表现出明显的选择性(Amtmann and Sanders, 1999).Goll- dack等(2003)研究表明, 盐处理下水稻根吸收Na+与OsAKT1的表达水平有直接关系.由此证明OsAKT1参与盐胁迫下Na+/K+比调控, 从而增强水稻的耐盐性.Wang等(2015)研究发现, 无论是低盐(25 mmol·L-1 NaCl)还是高盐(150 mmol·L-1 NaCl)条件下, 小花碱茅PtAKT1选择性吸收K+的能力显著强于Na+ (selective absorption capacity for K+ over Na+, SA).进一步研究发现, PtAKT1的转录水平与SA值之间呈显著正相关, 可见盐胁迫下PtAKT1是调控小花碱茅K+选择性吸收的关键因子(Wang et al., 2015).与野生型相比, 盐胁迫下超表达PtAKT1的拟南芥转基因植株K+含量增加, 而Na+含量降低, 表明PtAKT1增强了转基因植株对K+的吸收能力, 从而提高其耐盐性(Ardie et al., 2010).Duan等(2015)研究发现, 盐胁迫下盐地碱蓬SsAKT1能促进植株体内K+积累, 进而增强植物耐盐性.在盐(100 mmol·L-1 NaCl)处理下, 大麦HvAKT1在叶伸长区的表达水平明显增加, 说明HvAKT1参与维持叶肉细胞K+的稳态平衡(Boscari et al., 2009).我们的研究发现, 在5-50 mmol·L-1 NaCl处理下, 分别添加3和6 mmol·L-1 Cs+ (K+通道AKT1特异性抑制剂)均可显著降低甜菜(Beta vulgaris)根的K+净吸收速率, 可见AKT1可介导甜菜根K+的吸收(Wu et al., 2015b).这些研究结果表明, AKT1在K+和Na+选择性吸收、维持植物体内K+含量以及增强植物耐盐性等方面具有重要作用. ...
1 2011
... AtKC1是Reintanz等(2002)从拟南芥中分离克隆的α-亚基Shaker通道基因, 主要在根毛、根表皮及皮层细胞中表达.AtKC1作为四聚体的组成亚基参与K+通道四聚体的形成, 其存在对四聚体的K+通透能力产生抑制作用(Reintanz et al., 2002; Geiger et al., 2009).可见, AtKC1是负调控因子, 也是1个调控亚基(Wang et al., 2010).另外, AtKC1的存在使四聚体的K+通道活性激活电压向负电压方向偏移, 离子通道更难以被激活, 即改变了其对电压的敏感性(Duby et al., 2008).因此, 这种负向调控阻断了由AKT1介导的吸收.短时间缺K+处理则上调AtKC1的转录丰度, 随后其转录丰度下调(Shin and Schachtman, 2004), 这种现象可通过缺K+处理几天后的AtKC1转录物消失而得到解释(Pilot et al., 2003).在异源表达系统中, AtKC1不能独立形成有功能的K+通道(Duby et al., 2008).然而, 当AtKC1不能正常发挥功能时, 根毛中AKT1介导的内向K+电流的生物物理特征就会发生改变(Jeanguenin et al., 2011).Wang等(2016)进一步研究证实, 在低K+条件下, CIPK23促进AKT1介导的K+吸收, 而AtKC1抑制AKT1介导的K+内流.他们认为, CIPK23和AtKC1协同调节AKT1的功能, 二者具有一定的互补性.另有研究表明, Atkc1与Atakt1突变体植株的表型相似或相反, 这主要取决于外界环境条件(Geiger et al., 2009; Wang et al., 2010).AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009).低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似.在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014).最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014).PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性.可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014). ...
1 2011
... 植物在长期进化过程中形成了一系列复杂的K+吸收和转运系统.已知在模式植物拟南芥(Arabi- dopsis thaliana)中存在15个K+选择性通道, 包括9个Shaker型、5个TPK (two-pore K+ channel)型和1个Kir-like型K+通道(Ward et al., 2009; Hedrich, 2012; Demidchik, 2014).另外, 一些非选择性阳离子通道(non-selective cation channels, NSCC)也可以通透K+, 主要包括20个CNGC (cyclic nucleotide gated channels)通道、20个GLR (glutamate receptor-like)通道和1个TPC (two-pore channel)通道(Shabala, 2003; Demidchik and Maathuis, 2007; Dietrich et al., 2010; Hedrich, 2012; Demidchik, 2014).植物K+转运蛋白主要包括KUP (K+ uptake permease)/HAK (high-affinity K+ transporter)/KT (K+ transporter)、HKT (high-affinity K+ transporter)和CPA (cation proton antiporter) (Gierth and Mäser, 2007; Kron- zucker and Britto, 2011; Wang and Wu, 2013; Han et al., 2016).近年来, 人们相继在植物中克隆到编码这些蛋白的基因, 为深入研究植物K+吸收机制奠定了坚实的基础. ...
2 1996
... 在K+饥饿状态下, 小麦根TaAKT1 (Buschmann et al., 2000)和小花碱茅(Puccinellia tenuiflora) PtAKT1 (Ardie et al., 2010)的转录水平显著提高.在拟南芥中, 外部K+浓度在5 μmol·L-1至5 mmol·L-1范围内未影响AtAKT1的转录丰度(Lagarde et al., 1996).在棉花(Xu et al., 2014)和小花碱茅(Wang et al., 2015)中也有类似报道, 由此推测AKT1可能受转录后调控.然而, 25-250 mmol·L-1 NaCl短期(6小时)处理能够诱导盐地碱蓬根SsAKT1的表达(Duan et al., 2015).研究表明, 水稻OsAKT1主要在根的外皮层和内皮层细胞以及叶的木质部薄壁组织、韧皮部和叶肉细胞中表达(Golldack et al., 2003).盐胁迫(150 mmol·L-1 NaCl处理48小时)下, 盐敏感型水稻品种Pokkali和BK根外皮层细胞中的OsAKT1转录物消失, 而耐盐型品种IR29中的则未受到影响(Golldack et al., 2003).Su等(2001)的研究表明, 盐胁迫下盐生植物冰叶日中花(Mesembryanthemum crystallinum)根中与AtAKT1同源的K+通道基因McAKT1表达水平下调.在小花碱茅中, 尽管盐处理会下调PtAKT1的表达, 但150 mmol·L-1 NaCl处理下PtAKT1的转录丰度显著高于25 mmol·L-1 (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ... ... 干旱是制约全球作物生长和产量的主要非生物因素之一(Chaves and Oliveira, 2004; 山仑, 2011).植物响应干旱胁迫的重要策略之一是吸收并在体内积累大量的溶质(如K+、Na+和脯氨酸等) (Wang et al., 2004; Mahouachi et al., 2006; Wu et al., 2015a), 降低细胞渗透势, 从而增强植物细胞吸水能力.AKT1在植物响应干旱胁迫中的作用主要有2方面.(1) 干旱胁迫下AKT1增强植物根的K+吸收能力.在水稻中超表达OsAKT1基因后, 转基因植株在干旱条件下生长良好, 且根中积累较多的K+; 相反, Osakt1突变体植物表现为生长不良, 且根中K+含量明显减少(Ahmad et al., 2016).由此表明, 超表达AKT1转基因植株通过增加根部K+的积累来改善细胞渗透调节能力, 从而增强植物的抗旱性.(2) AKT1通过调节气孔的运动影响植物体内的水分平衡.研究表明, 拟南芥AtAKT1在保卫细胞质膜上呈低丰度表达(Lagarde et al., 1996).在渗透胁迫下, 拟南芥Atakt1突变体植株的蒸腾速率和水分消耗量明显低于野生型植株(Nieves- Cordones et al., 2012).由此可见, AKT1功能缺失致使保卫细胞K+跨膜转运体系受损, 气孔导度下降.Ahmad等(2016)进一步研究发现, 与野生型相比, 在干旱胁迫下水稻Osakt1突变体植株的气孔导度明显降低; 而超表达OsAKT1显著增强了转基因植株的气孔导度.这些研究结果表明, AKT1在介导保卫细胞K+转运、调控气孔运动及维持细胞水分平衡中发挥重要作用. ...
1 2011
... 尽管自20世纪90年代从拟南芥中克隆到第1个AKT1基因以来, 人们相继在其它植物中克隆得到同源基因.然而, AKT1基因所编码蛋白的调控机制仍不清楚.Xu等(2006)首次在植物中报道了低K+条件下蛋白磷酸化对K+吸收的调控机制.在爪蟾卵母细胞(Xeno- pus oocytes)中, CBL1 (calcineurin B-like protein1)/ CBL9与CIPK23 (CBL-interacting protein kinases 23)互作, 通过磷酸化作用激活AKT1介导的K+吸收(Li et al., 2006; Xu et al., 2006; Cuéllar et al., 2010, 2013).低K+处理下, CIPK23表达上调, 而CBL1和CBL9似乎是组成型表达(Xu et al., 2006; Cheong et al., 2007); 过量表达CIPK23可显著增强拟南芥K+的吸收能力和低K+耐受性(Xu et al., 2006).CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ...
1 2007
... 尽管自20世纪90年代从拟南芥中克隆到第1个AKT1基因以来, 人们相继在其它植物中克隆得到同源基因.然而, AKT1基因所编码蛋白的调控机制仍不清楚.Xu等(2006)首次在植物中报道了低K+条件下蛋白磷酸化对K+吸收的调控机制.在爪蟾卵母细胞(Xeno- pus oocytes)中, CBL1 (calcineurin B-like protein1)/ CBL9与CIPK23 (CBL-interacting protein kinases 23)互作, 通过磷酸化作用激活AKT1介导的K+吸收(Li et al., 2006; Xu et al., 2006; Cuéllar et al., 2010, 2013).低K+处理下, CIPK23表达上调, 而CBL1和CBL9似乎是组成型表达(Xu et al., 2006; Cheong et al., 2007); 过量表达CIPK23可显著增强拟南芥K+的吸收能力和低K+耐受性(Xu et al., 2006).CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ...
3 2014
... 低K+环境是影响植物正常生长发育及作物品质的主要非生物因子之一.K+是种子萌发所需的重要渗透调节物质.在低K+ (50-100 μmol·L-1)条件下, 拟南芥Atakt1突变体种子的胚根不能正常生长; 然而, 当在Atakt1突变体中超表达AtAKT1后, 其种子萌发活力得到恢复, 说明AKT1参与种子萌发期间K+的吸收(Pyo et al., 2010; Xu et al., 2014).AKT1功能缺失使植物根细胞膜表现出超级化, 对外界环境K+浓度变化异常敏感(Hirsch et al., 1998).在低K+环境中, 拟南芥Atakt1突变体植株细胞内的K+含量明显降低(Hirsch et al., 1998; Spalding et al., 1999).电生理学实验表明, 水稻OsAKT1介导内向K+电流, 即使在低K+ (5 μmol·L-1-1 mmol·L-1)条件下也是如此(Li et al., 2014).有研究表明, 拟南芥Atakt1和水稻Osakt1突变体植株的内向K+电流减弱, K+吸收功能紊乱, 从而影响其生长发育(Reintanz et al., 2002; Xu et al., 2006; Li et al., 2014).当在水稻Osakt1突变体中超表达OsAKT1后, 低K+胁迫下转基因植株的内向K+电流增强, K+含量显著增加(Li et al., 2014).这进一步证实了AKT1介导植物根吸收K+.Xu等(2014)采用非损伤微测技术(no-invasive micro-test technology, NMT), 在棉花GhAKT1功能研究中也得到类似的结果.可见, 在低K+环境中, AKT1介导的K+吸收系统对维持植物正常生长发育具有重要作用. ... ... ; Li et al., 2014).当在水稻Osakt1突变体中超表达OsAKT1后, 低K+胁迫下转基因植株的内向K+电流增强, K+含量显著增加(Li et al., 2014).这进一步证实了AKT1介导植物根吸收K+.Xu等(2014)采用非损伤微测技术(no-invasive micro-test technology, NMT), 在棉花GhAKT1功能研究中也得到类似的结果.可见, 在低K+环境中, AKT1介导的K+吸收系统对维持植物正常生长发育具有重要作用. ... ... 含量显著增加(Li et al., 2014).这进一步证实了AKT1介导植物根吸收K+.Xu等(2014)采用非损伤微测技术(no-invasive micro-test technology, NMT), 在棉花GhAKT1功能研究中也得到类似的结果.可见, 在低K+环境中, AKT1介导的K+吸收系统对维持植物正常生长发育具有重要作用. ...
2 2006
... 尽管自20世纪90年代从拟南芥中克隆到第1个AKT1基因以来, 人们相继在其它植物中克隆得到同源基因.然而, AKT1基因所编码蛋白的调控机制仍不清楚.Xu等(2006)首次在植物中报道了低K+条件下蛋白磷酸化对K+吸收的调控机制.在爪蟾卵母细胞(Xeno- pus oocytes)中, CBL1 (calcineurin B-like protein1)/ CBL9与CIPK23 (CBL-interacting protein kinases 23)互作, 通过磷酸化作用激活AKT1介导的K+吸收(Li et al., 2006; Xu et al., 2006; Cuéllar et al., 2010, 2013).低K+处理下, CIPK23表达上调, 而CBL1和CBL9似乎是组成型表达(Xu et al., 2006; Cheong et al., 2007); 过量表达CIPK23可显著增强拟南芥K+的吸收能力和低K+耐受性(Xu et al., 2006).CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ... ... 信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ...
2 2009
... 尽管自20世纪90年代从拟南芥中克隆到第1个AKT1基因以来, 人们相继在其它植物中克隆得到同源基因.然而, AKT1基因所编码蛋白的调控机制仍不清楚.Xu等(2006)首次在植物中报道了低K+条件下蛋白磷酸化对K+吸收的调控机制.在爪蟾卵母细胞(Xeno- pus oocytes)中, CBL1 (calcineurin B-like protein1)/ CBL9与CIPK23 (CBL-interacting protein kinases 23)互作, 通过磷酸化作用激活AKT1介导的K+吸收(Li et al., 2006; Xu et al., 2006; Cuéllar et al., 2010, 2013).低K+处理下, CIPK23表达上调, 而CBL1和CBL9似乎是组成型表达(Xu et al., 2006; Cheong et al., 2007); 过量表达CIPK23可显著增强拟南芥K+的吸收能力和低K+耐受性(Xu et al., 2006).CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ... ... ).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ...
1 2009
... K+是植物细胞中含量最丰富的无机阳离子之一, 主要通过根表皮细胞和皮层细胞的质膜K+转运系统吸收并运输到植物体内(王毅和武维华, 2009; Alemán et al., 2011; Dreyer and Uozumi, 2011; Wang and Wu, 2013; Nieves-Cordones et al., 2014a).K+在植物生长发育、逆境胁迫响应及生理生化反应等方面起着重要作用(Alemán et al., 2011; Nieves-Cordones et al., 2014a).一般而言, 植物细胞维持正常新陈代谢所需K+的最佳浓度为100 mmol·L-1, 但土壤中能被植物利用的K+浓度仅为0.1-1 mmol·L-1, 在一些盐碱地中甚至更低(Maathuis and Sanders, 1993; Maathuis, 2009).因此, 土壤中的K+远远不能满足植物正常生长需要, 使植物经常处于K+饥饿状态, 严重影响其生长发育.鉴于复杂多变的环境及K+的重要作用, 众多研究者对植物K+吸收和转运机制进行了深入研究. ...
1 2003
... 在K+饥饿状态下, 小麦根TaAKT1 (Buschmann et al., 2000)和小花碱茅(Puccinellia tenuiflora) PtAKT1 (Ardie et al., 2010)的转录水平显著提高.在拟南芥中, 外部K+浓度在5 μmol·L-1至5 mmol·L-1范围内未影响AtAKT1的转录丰度(Lagarde et al., 1996).在棉花(Xu et al., 2014)和小花碱茅(Wang et al., 2015)中也有类似报道, 由此推测AKT1可能受转录后调控.然而, 25-250 mmol·L-1 NaCl短期(6小时)处理能够诱导盐地碱蓬根SsAKT1的表达(Duan et al., 2015).研究表明, 水稻OsAKT1主要在根的外皮层和内皮层细胞以及叶的木质部薄壁组织、韧皮部和叶肉细胞中表达(Golldack et al., 2003).盐胁迫(150 mmol·L-1 NaCl处理48小时)下, 盐敏感型水稻品种Pokkali和BK根外皮层细胞中的OsAKT1转录物消失, 而耐盐型品种IR29中的则未受到影响(Golldack et al., 2003).Su等(2001)的研究表明, 盐胁迫下盐生植物冰叶日中花(Mesembryanthemum crystallinum)根中与AtAKT1同源的K+通道基因McAKT1表达水平下调.在小花碱茅中, 尽管盐处理会下调PtAKT1的表达, 但150 mmol·L-1 NaCl处理下PtAKT1的转录丰度显著高于25 mmol·L-1 (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ...
1 1997
... 盐胁迫下, 植物体内积累大量的Na+, 一方面抑制K+的吸收, 另一方面与K+竞争一些酶活性结合位点, 影响蛋白质合成及核糖体功能, 从而产生Na+毒害(Horie et al., 2009; Zhang et al., 2010).一般认为, 维持细胞质内较高的K+/Na+比是植物适应盐胁迫的重要策略之一(Horie et al., 2009; Shabala et al., 2010; Chérel et al., 2014).AKT1是参与K+吸收的重要通道蛋白, 在植物响应盐胁迫中起着重要作用.异源表达系统研究表明, 尽管AKT1对K+具有更强的选择性, 但并不排除对Na+的吸收(Maathuis et al., 1997).低Na+条件下, 内向整流K+通道未表现出明显的Na+进出现象; 但在高盐条件下, 其对K+表现出明显的选择性(Amtmann and Sanders, 1999).Goll- dack等(2003)研究表明, 盐处理下水稻根吸收Na+与OsAKT1的表达水平有直接关系.由此证明OsAKT1参与盐胁迫下Na+/K+比调控, 从而增强水稻的耐盐性.Wang等(2015)研究发现, 无论是低盐(25 mmol·L-1 NaCl)还是高盐(150 mmol·L-1 NaCl)条件下, 小花碱茅PtAKT1选择性吸收K+的能力显著强于Na+ (selective absorption capacity for K+ over Na+, SA).进一步研究发现, PtAKT1的转录水平与SA值之间呈显著正相关, 可见盐胁迫下PtAKT1是调控小花碱茅K+选择性吸收的关键因子(Wang et al., 2015).与野生型相比, 盐胁迫下超表达PtAKT1的拟南芥转基因植株K+含量增加, 而Na+含量降低, 表明PtAKT1增强了转基因植株对K+的吸收能力, 从而提高其耐盐性(Ardie et al., 2010).Duan等(2015)研究发现, 盐胁迫下盐地碱蓬SsAKT1能促进植株体内K+积累, 进而增强植物耐盐性.在盐(100 mmol·L-1 NaCl)处理下, 大麦HvAKT1在叶伸长区的表达水平明显增加, 说明HvAKT1参与维持叶肉细胞K+的稳态平衡(Boscari et al., 2009).我们的研究发现, 在5-50 mmol·L-1 NaCl处理下, 分别添加3和6 mmol·L-1 Cs+ (K+通道AKT1特异性抑制剂)均可显著降低甜菜(Beta vulgaris)根的K+净吸收速率, 可见AKT1可介导甜菜根K+的吸收(Wu et al., 2015b).这些研究结果表明, AKT1在K+和Na+选择性吸收、维持植物体内K+含量以及增强植物耐盐性等方面具有重要作用. ...
1 1993
... K+是植物细胞中含量最丰富的无机阳离子之一, 主要通过根表皮细胞和皮层细胞的质膜K+转运系统吸收并运输到植物体内(王毅和武维华, 2009; Alemán et al., 2011; Dreyer and Uozumi, 2011; Wang and Wu, 2013; Nieves-Cordones et al., 2014a).K+在植物生长发育、逆境胁迫响应及生理生化反应等方面起着重要作用(Alemán et al., 2011; Nieves-Cordones et al., 2014a).一般而言, 植物细胞维持正常新陈代谢所需K+的最佳浓度为100 mmol·L-1, 但土壤中能被植物利用的K+浓度仅为0.1-1 mmol·L-1, 在一些盐碱地中甚至更低(Maathuis and Sanders, 1993; Maathuis, 2009).因此, 土壤中的K+远远不能满足植物正常生长需要, 使植物经常处于K+饥饿状态, 严重影响其生长发育.鉴于复杂多变的环境及K+的重要作用, 众多研究者对植物K+吸收和转运机制进行了深入研究. ...
1 2006
... 干旱是制约全球作物生长和产量的主要非生物因素之一(Chaves and Oliveira, 2004; 山仑, 2011).植物响应干旱胁迫的重要策略之一是吸收并在体内积累大量的溶质(如K+、Na+和脯氨酸等) (Wang et al., 2004; Mahouachi et al., 2006; Wu et al., 2015a), 降低细胞渗透势, 从而增强植物细胞吸水能力.AKT1在植物响应干旱胁迫中的作用主要有2方面.(1) 干旱胁迫下AKT1增强植物根的K+吸收能力.在水稻中超表达OsAKT1基因后, 转基因植株在干旱条件下生长良好, 且根中积累较多的K+; 相反, Osakt1突变体植物表现为生长不良, 且根中K+含量明显减少(Ahmad et al., 2016).由此表明, 超表达AKT1转基因植株通过增加根部K+的积累来改善细胞渗透调节能力, 从而增强植物的抗旱性.(2) AKT1通过调节气孔的运动影响植物体内的水分平衡.研究表明, 拟南芥AtAKT1在保卫细胞质膜上呈低丰度表达(Lagarde et al., 1996).在渗透胁迫下, 拟南芥Atakt1突变体植株的蒸腾速率和水分消耗量明显低于野生型植株(Nieves- Cordones et al., 2012).由此可见, AKT1功能缺失致使保卫细胞K+跨膜转运体系受损, 气孔导度下降.Ahmad等(2016)进一步研究发现, 与野生型相比, 在干旱胁迫下水稻Osakt1突变体植株的气孔导度明显降低; 而超表达OsAKT1显著增强了转基因植株的气孔导度.这些研究结果表明, AKT1在介导保卫细胞K+转运、调控气孔运动及维持细胞水分平衡中发挥重要作用. ...
2005
2014
1 2012
... 干旱是制约全球作物生长和产量的主要非生物因素之一(Chaves and Oliveira, 2004; 山仑, 2011).植物响应干旱胁迫的重要策略之一是吸收并在体内积累大量的溶质(如K+、Na+和脯氨酸等) (Wang et al., 2004; Mahouachi et al., 2006; Wu et al., 2015a), 降低细胞渗透势, 从而增强植物细胞吸水能力.AKT1在植物响应干旱胁迫中的作用主要有2方面.(1) 干旱胁迫下AKT1增强植物根的K+吸收能力.在水稻中超表达OsAKT1基因后, 转基因植株在干旱条件下生长良好, 且根中积累较多的K+; 相反, Osakt1突变体植物表现为生长不良, 且根中K+含量明显减少(Ahmad et al., 2016).由此表明, 超表达AKT1转基因植株通过增加根部K+的积累来改善细胞渗透调节能力, 从而增强植物的抗旱性.(2) AKT1通过调节气孔的运动影响植物体内的水分平衡.研究表明, 拟南芥AtAKT1在保卫细胞质膜上呈低丰度表达(Lagarde et al., 1996).在渗透胁迫下, 拟南芥Atakt1突变体植株的蒸腾速率和水分消耗量明显低于野生型植株(Nieves- Cordones et al., 2012).由此可见, AKT1功能缺失致使保卫细胞K+跨膜转运体系受损, 气孔导度下降.Ahmad等(2016)进一步研究发现, 与野生型相比, 在干旱胁迫下水稻Osakt1突变体植株的气孔导度明显降低; 而超表达OsAKT1显著增强了转基因植株的气孔导度.这些研究结果表明, AKT1在介导保卫细胞K+转运、调控气孔运动及维持细胞水分平衡中发挥重要作用. ...
2 2014
... K+是植物细胞中含量最丰富的无机阳离子之一, 主要通过根表皮细胞和皮层细胞的质膜K+转运系统吸收并运输到植物体内(王毅和武维华, 2009; Alemán et al., 2011; Dreyer and Uozumi, 2011; Wang and Wu, 2013; Nieves-Cordones et al., 2014a).K+在植物生长发育、逆境胁迫响应及生理生化反应等方面起着重要作用(Alemán et al., 2011; Nieves-Cordones et al., 2014a).一般而言, 植物细胞维持正常新陈代谢所需K+的最佳浓度为100 mmol·L-1, 但土壤中能被植物利用的K+浓度仅为0.1-1 mmol·L-1, 在一些盐碱地中甚至更低(Maathuis and Sanders, 1993; Maathuis, 2009).因此, 土壤中的K+远远不能满足植物正常生长需要, 使植物经常处于K+饥饿状态, 严重影响其生长发育.鉴于复杂多变的环境及K+的重要作用, 众多研究者对植物K+吸收和转运机制进行了深入研究. ... ... ; Nieves-Cordones et al., 2014a).一般而言, 植物细胞维持正常新陈代谢所需K+的最佳浓度为100 mmol·L-1, 但土壤中能被植物利用的K+浓度仅为0.1-1 mmol·L-1, 在一些盐碱地中甚至更低(Maathuis and Sanders, 1993; Maathuis, 2009).因此, 土壤中的K+远远不能满足植物正常生长需要, 使植物经常处于K+饥饿状态, 严重影响其生长发育.鉴于复杂多变的环境及K+的重要作用, 众多研究者对植物K+吸收和转运机制进行了深入研究. ...
7 2014
... 植物AKT1与动物Shaker型K+通道结构高度相似(图1), 含有6个跨膜片段(S1-S6), 其中第4个跨膜片段S4含有大量带正电荷的氨基酸残基, 是电压感受器, 其主要功能为响应膜电势的变化.该片段可在膜上移动, 使得膜通道构象改变, 从而控制通道孔的开放与关闭(Benito et al., 2014; Nieves-Cordones and Gaillard, 2014).S5和S6之间含有1个高度保守的P环结构域, 该结构域是陷入细胞膜内的一段多肽片段, 构成通道孔.AKT1含有典型的TxGYG (Thr-X- Gly-Tyr-Gly)序列, 为K+选择器, 这是Shaker型K+通道的共同特点.胞质C末端从第6个跨膜片段末尾起, 含有1个C-接头(约有80个氨基酸残基), 1个环核苷酸结合域(cyclic-nucleotide binding domain, CNBD), 1个锚蛋白域和1个富含疏水性、酸性残基的KHA域(Nieves-Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).对大多数Shaker型K+通道家族成员来说, 锚蛋白域有助于通道与细胞骨架的连接、蛋白质的相互作用和细胞溶质的调节(Dreyer and Blatt, 2009; Riedelsberger et al., 2010; Nieves- Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).Shaker型K+通道的重要特点之一是能够形成异源四聚体结构, 可使植物调控不同细胞中的K+转运活性.这种调控在每个器官或者组织中相对独立, 并且受到环境的影响(Gambale and Uozumi, 2006). ... ... 域(Nieves-Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).对大多数Shaker型K+通道家族成员来说, 锚蛋白域有助于通道与细胞骨架的连接、蛋白质的相互作用和细胞溶质的调节(Dreyer and Blatt, 2009; Riedelsberger et al., 2010; Nieves- Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).Shaker型K+通道的重要特点之一是能够形成异源四聚体结构, 可使植物调控不同细胞中的K+转运活性.这种调控在每个器官或者组织中相对独立, 并且受到环境的影响(Gambale and Uozumi, 2006). ... ... ; Nieves-Cordones and Gaillard, 2014).对大多数Shaker型K+通道家族成员来说, 锚蛋白域有助于通道与细胞骨架的连接、蛋白质的相互作用和细胞溶质的调节(Dreyer and Blatt, 2009; Riedelsberger et al., 2010; Nieves- Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).Shaker型K+通道的重要特点之一是能够形成异源四聚体结构, 可使植物调控不同细胞中的K+转运活性.这种调控在每个器官或者组织中相对独立, 并且受到环境的影响(Gambale and Uozumi, 2006). ... ... ; Nieves- Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).Shaker型K+通道的重要特点之一是能够形成异源四聚体结构, 可使植物调控不同细胞中的K+转运活性.这种调控在每个器官或者组织中相对独立, 并且受到环境的影响(Gambale and Uozumi, 2006). ... ... ; Nieves-Cordones and Gaillard, 2014).Shaker型K+通道的重要特点之一是能够形成异源四聚体结构, 可使植物调控不同细胞中的K+转运活性.这种调控在每个器官或者组织中相对独立, 并且受到环境的影响(Gambale and Uozumi, 2006). ... ... 植物Shaker型K+通道拓扑结构示意图(改自Nieves- Cordones and Gaillard, 2014) S1-S6为6个跨膜结构域; S4为电压感受器; CNBD为环核苷酸结合域. ... ... Diagrammatic representation of plant Shaker type K+ channel topological structure (Modified from Nieves- Cordones and Gaillard, 2014) S1-S6 indicate six transmembrane domains; S4 represents voltage sensor; CNBD indicates cyclic-nucleotide binding domain. ...
1 1999
... AKT1 (Arabidopsis K+ transporter 1)是植物中广泛存在的一类内向整流K+通道, 与动物中的Shaker型K+通道具有高度的同源性(Pilot et al., 2003).Sentenac等(1992)利用酵母双突变体互补法从拟南芥中克隆到第1个高等植物的Shaker型K+通道基因AtAKT1.该基因位于拟南芥第2号染色体上, 含有11个外显子和10个内含子, 编码的多肽由857个氨基酸残基组成.此后, 科研人员相继在马铃薯(Sola- num tuberosum) (Zimmermann et al., 1998)、玉米(Zea mays) (Philippar et al., 1999)、小麦(Triticum aestivum) (Buschmann et al., 2000)、番茄(Lycoper- sicon esculentum) (Hartje et al., 2000)、水稻(Oryza sativa) (Fuchs et al., 2005)、大麦(Hordeum vulgare) (Boscari et al., 2009)、葡萄(Vitis vinifera) (Cuéllar et al., 2010, 2013)和盐地碱蓬(Suaeda salsa) (Duan et al., 2015)等植物中分离得到AKT1基因.AKT1在维持植物离子稳态平衡及增强植物逆境胁迫耐受性等方面起着重要作用(Alemán et al., 2011; Wang and Wu, 2013).本文从植物AKT1的分子结构特点、调控机制及生物学功能等方面对其主要研究进展进行了归纳总结, 旨在为植物AKT1的进一步研究奠定基础. ...
3 2003
... AKT1 (Arabidopsis K+ transporter 1)是植物中广泛存在的一类内向整流K+通道, 与动物中的Shaker型K+通道具有高度的同源性(Pilot et al., 2003).Sentenac等(1992)利用酵母双突变体互补法从拟南芥中克隆到第1个高等植物的Shaker型K+通道基因AtAKT1.该基因位于拟南芥第2号染色体上, 含有11个外显子和10个内含子, 编码的多肽由857个氨基酸残基组成.此后, 科研人员相继在马铃薯(Sola- num tuberosum) (Zimmermann et al., 1998)、玉米(Zea mays) (Philippar et al., 1999)、小麦(Triticum aestivum) (Buschmann et al., 2000)、番茄(Lycoper- sicon esculentum) (Hartje et al., 2000)、水稻(Oryza sativa) (Fuchs et al., 2005)、大麦(Hordeum vulgare) (Boscari et al., 2009)、葡萄(Vitis vinifera) (Cuéllar et al., 2010, 2013)和盐地碱蓬(Suaeda salsa) (Duan et al., 2015)等植物中分离得到AKT1基因.AKT1在维持植物离子稳态平衡及增强植物逆境胁迫耐受性等方面起着重要作用(Alemán et al., 2011; Wang and Wu, 2013).本文从植物AKT1的分子结构特点、调控机制及生物学功能等方面对其主要研究进展进行了归纳总结, 旨在为植物AKT1的进一步研究奠定基础. ... ... 在K+饥饿状态下, 小麦根TaAKT1 (Buschmann et al., 2000)和小花碱茅(Puccinellia tenuiflora) PtAKT1 (Ardie et al., 2010)的转录水平显著提高.在拟南芥中, 外部K+浓度在5 μmol·L-1至5 mmol·L-1范围内未影响AtAKT1的转录丰度(Lagarde et al., 1996).在棉花(Xu et al., 2014)和小花碱茅(Wang et al., 2015)中也有类似报道, 由此推测AKT1可能受转录后调控.然而, 25-250 mmol·L-1 NaCl短期(6小时)处理能够诱导盐地碱蓬根SsAKT1的表达(Duan et al., 2015).研究表明, 水稻OsAKT1主要在根的外皮层和内皮层细胞以及叶的木质部薄壁组织、韧皮部和叶肉细胞中表达(Golldack et al., 2003).盐胁迫(150 mmol·L-1 NaCl处理48小时)下, 盐敏感型水稻品种Pokkali和BK根外皮层细胞中的OsAKT1转录物消失, 而耐盐型品种IR29中的则未受到影响(Golldack et al., 2003).Su等(2001)的研究表明, 盐胁迫下盐生植物冰叶日中花(Mesembryanthemum crystallinum)根中与AtAKT1同源的K+通道基因McAKT1表达水平下调.在小花碱茅中, 尽管盐处理会下调PtAKT1的表达, 但150 mmol·L-1 NaCl处理下PtAKT1的转录丰度显著高于25 mmol·L-1 (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ... ... AtKC1是Reintanz等(2002)从拟南芥中分离克隆的α-亚基Shaker通道基因, 主要在根毛、根表皮及皮层细胞中表达.AtKC1作为四聚体的组成亚基参与K+通道四聚体的形成, 其存在对四聚体的K+通透能力产生抑制作用(Reintanz et al., 2002; Geiger et al., 2009).可见, AtKC1是负调控因子, 也是1个调控亚基(Wang et al., 2010).另外, AtKC1的存在使四聚体的K+通道活性激活电压向负电压方向偏移, 离子通道更难以被激活, 即改变了其对电压的敏感性(Duby et al., 2008).因此, 这种负向调控阻断了由AKT1介导的吸收.短时间缺K+处理则上调AtKC1的转录丰度, 随后其转录丰度下调(Shin and Schachtman, 2004), 这种现象可通过缺K+处理几天后的AtKC1转录物消失而得到解释(Pilot et al., 2003).在异源表达系统中, AtKC1不能独立形成有功能的K+通道(Duby et al., 2008).然而, 当AtKC1不能正常发挥功能时, 根毛中AKT1介导的内向K+电流的生物物理特征就会发生改变(Jeanguenin et al., 2011).Wang等(2016)进一步研究证实, 在低K+条件下, CIPK23促进AKT1介导的K+吸收, 而AtKC1抑制AKT1介导的K+内流.他们认为, CIPK23和AtKC1协同调节AKT1的功能, 二者具有一定的互补性.另有研究表明, Atkc1与Atakt1突变体植株的表型相似或相反, 这主要取决于外界环境条件(Geiger et al., 2009; Wang et al., 2010).AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009).低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似.在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014).最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014).PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性.可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014). ...
1 2010
... 低K+环境是影响植物正常生长发育及作物品质的主要非生物因子之一.K+是种子萌发所需的重要渗透调节物质.在低K+ (50-100 μmol·L-1)条件下, 拟南芥Atakt1突变体种子的胚根不能正常生长; 然而, 当在Atakt1突变体中超表达AtAKT1后, 其种子萌发活力得到恢复, 说明AKT1参与种子萌发期间K+的吸收(Pyo et al., 2010; Xu et al., 2014).AKT1功能缺失使植物根细胞膜表现出超级化, 对外界环境K+浓度变化异常敏感(Hirsch et al., 1998).在低K+环境中, 拟南芥Atakt1突变体植株细胞内的K+含量明显降低(Hirsch et al., 1998; Spalding et al., 1999).电生理学实验表明, 水稻OsAKT1介导内向K+电流, 即使在低K+ (5 μmol·L-1-1 mmol·L-1)条件下也是如此(Li et al., 2014).有研究表明, 拟南芥Atakt1和水稻Osakt1突变体植株的内向K+电流减弱, K+吸收功能紊乱, 从而影响其生长发育(Reintanz et al., 2002; Xu et al., 2006; Li et al., 2014).当在水稻Osakt1突变体中超表达OsAKT1后, 低K+胁迫下转基因植株的内向K+电流增强, K+含量显著增加(Li et al., 2014).这进一步证实了AKT1介导植物根吸收K+.Xu等(2014)采用非损伤微测技术(no-invasive micro-test technology, NMT), 在棉花GhAKT1功能研究中也得到类似的结果.可见, 在低K+环境中, AKT1介导的K+吸收系统对维持植物正常生长发育具有重要作用. ...
1 2015
... 尽管自20世纪90年代从拟南芥中克隆到第1个AKT1基因以来, 人们相继在其它植物中克隆得到同源基因.然而, AKT1基因所编码蛋白的调控机制仍不清楚.Xu等(2006)首次在植物中报道了低K+条件下蛋白磷酸化对K+吸收的调控机制.在爪蟾卵母细胞(Xeno- pus oocytes)中, CBL1 (calcineurin B-like protein1)/ CBL9与CIPK23 (CBL-interacting protein kinases 23)互作, 通过磷酸化作用激活AKT1介导的K+吸收(Li et al., 2006; Xu et al., 2006; Cuéllar et al., 2010, 2013).低K+处理下, CIPK23表达上调, 而CBL1和CBL9似乎是组成型表达(Xu et al., 2006; Cheong et al., 2007); 过量表达CIPK23可显著增强拟南芥K+的吸收能力和低K+耐受性(Xu et al., 2006).CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ...
2 2002
... AtKC1是Reintanz等(2002)从拟南芥中分离克隆的α-亚基Shaker通道基因, 主要在根毛、根表皮及皮层细胞中表达.AtKC1作为四聚体的组成亚基参与K+通道四聚体的形成, 其存在对四聚体的K+通透能力产生抑制作用(Reintanz et al., 2002; Geiger et al., 2009).可见, AtKC1是负调控因子, 也是1个调控亚基(Wang et al., 2010).另外, AtKC1的存在使四聚体的K+通道活性激活电压向负电压方向偏移, 离子通道更难以被激活, 即改变了其对电压的敏感性(Duby et al., 2008).因此, 这种负向调控阻断了由AKT1介导的吸收.短时间缺K+处理则上调AtKC1的转录丰度, 随后其转录丰度下调(Shin and Schachtman, 2004), 这种现象可通过缺K+处理几天后的AtKC1转录物消失而得到解释(Pilot et al., 2003).在异源表达系统中, AtKC1不能独立形成有功能的K+通道(Duby et al., 2008).然而, 当AtKC1不能正常发挥功能时, 根毛中AKT1介导的内向K+电流的生物物理特征就会发生改变(Jeanguenin et al., 2011).Wang等(2016)进一步研究证实, 在低K+条件下, CIPK23促进AKT1介导的K+吸收, 而AtKC1抑制AKT1介导的K+内流.他们认为, CIPK23和AtKC1协同调节AKT1的功能, 二者具有一定的互补性.另有研究表明, Atkc1与Atakt1突变体植株的表型相似或相反, 这主要取决于外界环境条件(Geiger et al., 2009; Wang et al., 2010).AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009).低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似.在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014).最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014).PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性.可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014). ... ... 低K+环境是影响植物正常生长发育及作物品质的主要非生物因子之一.K+是种子萌发所需的重要渗透调节物质.在低K+ (50-100 μmol·L-1)条件下, 拟南芥Atakt1突变体种子的胚根不能正常生长; 然而, 当在Atakt1突变体中超表达AtAKT1后, 其种子萌发活力得到恢复, 说明AKT1参与种子萌发期间K+的吸收(Pyo et al., 2010; Xu et al., 2014).AKT1功能缺失使植物根细胞膜表现出超级化, 对外界环境K+浓度变化异常敏感(Hirsch et al., 1998).在低K+环境中, 拟南芥Atakt1突变体植株细胞内的K+含量明显降低(Hirsch et al., 1998; Spalding et al., 1999).电生理学实验表明, 水稻OsAKT1介导内向K+电流, 即使在低K+ (5 μmol·L-1-1 mmol·L-1)条件下也是如此(Li et al., 2014).有研究表明, 拟南芥Atakt1和水稻Osakt1突变体植株的内向K+电流减弱, K+吸收功能紊乱, 从而影响其生长发育(Reintanz et al., 2002; Xu et al., 2006; Li et al., 2014).当在水稻Osakt1突变体中超表达OsAKT1后, 低K+胁迫下转基因植株的内向K+电流增强, K+含量显著增加(Li et al., 2014).这进一步证实了AKT1介导植物根吸收K+.Xu等(2014)采用非损伤微测技术(no-invasive micro-test technology, NMT), 在棉花GhAKT1功能研究中也得到类似的结果.可见, 在低K+环境中, AKT1介导的K+吸收系统对维持植物正常生长发育具有重要作用. ...
1 2013
... 尽管自20世纪90年代从拟南芥中克隆到第1个AKT1基因以来, 人们相继在其它植物中克隆得到同源基因.然而, AKT1基因所编码蛋白的调控机制仍不清楚.Xu等(2006)首次在植物中报道了低K+条件下蛋白磷酸化对K+吸收的调控机制.在爪蟾卵母细胞(Xeno- pus oocytes)中, CBL1 (calcineurin B-like protein1)/ CBL9与CIPK23 (CBL-interacting protein kinases 23)互作, 通过磷酸化作用激活AKT1介导的K+吸收(Li et al., 2006; Xu et al., 2006; Cuéllar et al., 2010, 2013).低K+处理下, CIPK23表达上调, 而CBL1和CBL9似乎是组成型表达(Xu et al., 2006; Cheong et al., 2007); 过量表达CIPK23可显著增强拟南芥K+的吸收能力和低K+耐受性(Xu et al., 2006).CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ...
1 2010
... 植物AKT1与动物Shaker型K+通道结构高度相似(图1), 含有6个跨膜片段(S1-S6), 其中第4个跨膜片段S4含有大量带正电荷的氨基酸残基, 是电压感受器, 其主要功能为响应膜电势的变化.该片段可在膜上移动, 使得膜通道构象改变, 从而控制通道孔的开放与关闭(Benito et al., 2014; Nieves-Cordones and Gaillard, 2014).S5和S6之间含有1个高度保守的P环结构域, 该结构域是陷入细胞膜内的一段多肽片段, 构成通道孔.AKT1含有典型的TxGYG (Thr-X- Gly-Tyr-Gly)序列, 为K+选择器, 这是Shaker型K+通道的共同特点.胞质C末端从第6个跨膜片段末尾起, 含有1个C-接头(约有80个氨基酸残基), 1个环核苷酸结合域(cyclic-nucleotide binding domain, CNBD), 1个锚蛋白域和1个富含疏水性、酸性残基的KHA域(Nieves-Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).对大多数Shaker型K+通道家族成员来说, 锚蛋白域有助于通道与细胞骨架的连接、蛋白质的相互作用和细胞溶质的调节(Dreyer and Blatt, 2009; Riedelsberger et al., 2010; Nieves- Cordones et al., 2014b; Nieves-Cordones and Gaillard, 2014).Shaker型K+通道的重要特点之一是能够形成异源四聚体结构, 可使植物调控不同细胞中的K+转运活性.这种调控在每个器官或者组织中相对独立, 并且受到环境的影响(Gambale and Uozumi, 2006). ...
2007
1992
1 2003
... 植物在长期进化过程中形成了一系列复杂的K+吸收和转运系统.已知在模式植物拟南芥(Arabi- dopsis thaliana)中存在15个K+选择性通道, 包括9个Shaker型、5个TPK (two-pore K+ channel)型和1个Kir-like型K+通道(Ward et al., 2009; Hedrich, 2012; Demidchik, 2014).另外, 一些非选择性阳离子通道(non-selective cation channels, NSCC)也可以通透K+, 主要包括20个CNGC (cyclic nucleotide gated channels)通道、20个GLR (glutamate receptor-like)通道和1个TPC (two-pore channel)通道(Shabala, 2003; Demidchik and Maathuis, 2007; Dietrich et al., 2010; Hedrich, 2012; Demidchik, 2014).植物K+转运蛋白主要包括KUP (K+ uptake permease)/HAK (high-affinity K+ transporter)/KT (K+ transporter)、HKT (high-affinity K+ transporter)和CPA (cation proton antiporter) (Gierth and Mäser, 2007; Kron- zucker and Britto, 2011; Wang and Wu, 2013; Han et al., 2016).近年来, 人们相继在植物中克隆到编码这些蛋白的基因, 为深入研究植物K+吸收机制奠定了坚实的基础. ...
1 2010
... 盐胁迫下, 植物体内积累大量的Na+, 一方面抑制K+的吸收, 另一方面与K+竞争一些酶活性结合位点, 影响蛋白质合成及核糖体功能, 从而产生Na+毒害(Horie et al., 2009; Zhang et al., 2010).一般认为, 维持细胞质内较高的K+/Na+比是植物适应盐胁迫的重要策略之一(Horie et al., 2009; Shabala et al., 2010; Chérel et al., 2014).AKT1是参与K+吸收的重要通道蛋白, 在植物响应盐胁迫中起着重要作用.异源表达系统研究表明, 尽管AKT1对K+具有更强的选择性, 但并不排除对Na+的吸收(Maathuis et al., 1997).低Na+条件下, 内向整流K+通道未表现出明显的Na+进出现象; 但在高盐条件下, 其对K+表现出明显的选择性(Amtmann and Sanders, 1999).Goll- dack等(2003)研究表明, 盐处理下水稻根吸收Na+与OsAKT1的表达水平有直接关系.由此证明OsAKT1参与盐胁迫下Na+/K+比调控, 从而增强水稻的耐盐性.Wang等(2015)研究发现, 无论是低盐(25 mmol·L-1 NaCl)还是高盐(150 mmol·L-1 NaCl)条件下, 小花碱茅PtAKT1选择性吸收K+的能力显著强于Na+ (selective absorption capacity for K+ over Na+, SA).进一步研究发现, PtAKT1的转录水平与SA值之间呈显著正相关, 可见盐胁迫下PtAKT1是调控小花碱茅K+选择性吸收的关键因子(Wang et al., 2015).与野生型相比, 盐胁迫下超表达PtAKT1的拟南芥转基因植株K+含量增加, 而Na+含量降低, 表明PtAKT1增强了转基因植株对K+的吸收能力, 从而提高其耐盐性(Ardie et al., 2010).Duan等(2015)研究发现, 盐胁迫下盐地碱蓬SsAKT1能促进植株体内K+积累, 进而增强植物耐盐性.在盐(100 mmol·L-1 NaCl)处理下, 大麦HvAKT1在叶伸长区的表达水平明显增加, 说明HvAKT1参与维持叶肉细胞K+的稳态平衡(Boscari et al., 2009).我们的研究发现, 在5-50 mmol·L-1 NaCl处理下, 分别添加3和6 mmol·L-1 Cs+ (K+通道AKT1特异性抑制剂)均可显著降低甜菜(Beta vulgaris)根的K+净吸收速率, 可见AKT1可介导甜菜根K+的吸收(Wu et al., 2015b).这些研究结果表明, AKT1在K+和Na+选择性吸收、维持植物体内K+含量以及增强植物耐盐性等方面具有重要作用. ...
1 2004
... AtKC1是Reintanz等(2002)从拟南芥中分离克隆的α-亚基Shaker通道基因, 主要在根毛、根表皮及皮层细胞中表达.AtKC1作为四聚体的组成亚基参与K+通道四聚体的形成, 其存在对四聚体的K+通透能力产生抑制作用(Reintanz et al., 2002; Geiger et al., 2009).可见, AtKC1是负调控因子, 也是1个调控亚基(Wang et al., 2010).另外, AtKC1的存在使四聚体的K+通道活性激活电压向负电压方向偏移, 离子通道更难以被激活, 即改变了其对电压的敏感性(Duby et al., 2008).因此, 这种负向调控阻断了由AKT1介导的吸收.短时间缺K+处理则上调AtKC1的转录丰度, 随后其转录丰度下调(Shin and Schachtman, 2004), 这种现象可通过缺K+处理几天后的AtKC1转录物消失而得到解释(Pilot et al., 2003).在异源表达系统中, AtKC1不能独立形成有功能的K+通道(Duby et al., 2008).然而, 当AtKC1不能正常发挥功能时, 根毛中AKT1介导的内向K+电流的生物物理特征就会发生改变(Jeanguenin et al., 2011).Wang等(2016)进一步研究证实, 在低K+条件下, CIPK23促进AKT1介导的K+吸收, 而AtKC1抑制AKT1介导的K+内流.他们认为, CIPK23和AtKC1协同调节AKT1的功能, 二者具有一定的互补性.另有研究表明, Atkc1与Atakt1突变体植株的表型相似或相反, 这主要取决于外界环境条件(Geiger et al., 2009; Wang et al., 2010).AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009).低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似.在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014).最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014).PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性.可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014). ...
1 1999
... 低K+环境是影响植物正常生长发育及作物品质的主要非生物因子之一.K+是种子萌发所需的重要渗透调节物质.在低K+ (50-100 μmol·L-1)条件下, 拟南芥Atakt1突变体种子的胚根不能正常生长; 然而, 当在Atakt1突变体中超表达AtAKT1后, 其种子萌发活力得到恢复, 说明AKT1参与种子萌发期间K+的吸收(Pyo et al., 2010; Xu et al., 2014).AKT1功能缺失使植物根细胞膜表现出超级化, 对外界环境K+浓度变化异常敏感(Hirsch et al., 1998).在低K+环境中, 拟南芥Atakt1突变体植株细胞内的K+含量明显降低(Hirsch et al., 1998; Spalding et al., 1999).电生理学实验表明, 水稻OsAKT1介导内向K+电流, 即使在低K+ (5 μmol·L-1-1 mmol·L-1)条件下也是如此(Li et al., 2014).有研究表明, 拟南芥Atakt1和水稻Osakt1突变体植株的内向K+电流减弱, K+吸收功能紊乱, 从而影响其生长发育(Reintanz et al., 2002; Xu et al., 2006; Li et al., 2014).当在水稻Osakt1突变体中超表达OsAKT1后, 低K+胁迫下转基因植株的内向K+电流增强, K+含量显著增加(Li et al., 2014).这进一步证实了AKT1介导植物根吸收K+.Xu等(2014)采用非损伤微测技术(no-invasive micro-test technology, NMT), 在棉花GhAKT1功能研究中也得到类似的结果.可见, 在低K+环境中, AKT1介导的K+吸收系统对维持植物正常生长发育具有重要作用. ...
2001
3 2015
... 在K+饥饿状态下, 小麦根TaAKT1 (Buschmann et al., 2000)和小花碱茅(Puccinellia tenuiflora) PtAKT1 (Ardie et al., 2010)的转录水平显著提高.在拟南芥中, 外部K+浓度在5 μmol·L-1至5 mmol·L-1范围内未影响AtAKT1的转录丰度(Lagarde et al., 1996).在棉花(Xu et al., 2014)和小花碱茅(Wang et al., 2015)中也有类似报道, 由此推测AKT1可能受转录后调控.然而, 25-250 mmol·L-1 NaCl短期(6小时)处理能够诱导盐地碱蓬根SsAKT1的表达(Duan et al., 2015).研究表明, 水稻OsAKT1主要在根的外皮层和内皮层细胞以及叶的木质部薄壁组织、韧皮部和叶肉细胞中表达(Golldack et al., 2003).盐胁迫(150 mmol·L-1 NaCl处理48小时)下, 盐敏感型水稻品种Pokkali和BK根外皮层细胞中的OsAKT1转录物消失, 而耐盐型品种IR29中的则未受到影响(Golldack et al., 2003).Su等(2001)的研究表明, 盐胁迫下盐生植物冰叶日中花(Mesembryanthemum crystallinum)根中与AtAKT1同源的K+通道基因McAKT1表达水平下调.在小花碱茅中, 尽管盐处理会下调PtAKT1的表达, 但150 mmol·L-1 NaCl处理下PtAKT1的转录丰度显著高于25 mmol·L-1 (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ... ... (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ... ... 盐胁迫下, 植物体内积累大量的Na+, 一方面抑制K+的吸收, 另一方面与K+竞争一些酶活性结合位点, 影响蛋白质合成及核糖体功能, 从而产生Na+毒害(Horie et al., 2009; Zhang et al., 2010).一般认为, 维持细胞质内较高的K+/Na+比是植物适应盐胁迫的重要策略之一(Horie et al., 2009; Shabala et al., 2010; Chérel et al., 2014).AKT1是参与K+吸收的重要通道蛋白, 在植物响应盐胁迫中起着重要作用.异源表达系统研究表明, 尽管AKT1对K+具有更强的选择性, 但并不排除对Na+的吸收(Maathuis et al., 1997).低Na+条件下, 内向整流K+通道未表现出明显的Na+进出现象; 但在高盐条件下, 其对K+表现出明显的选择性(Amtmann and Sanders, 1999).Goll- dack等(2003)研究表明, 盐处理下水稻根吸收Na+与OsAKT1的表达水平有直接关系.由此证明OsAKT1参与盐胁迫下Na+/K+比调控, 从而增强水稻的耐盐性.Wang等(2015)研究发现, 无论是低盐(25 mmol·L-1 NaCl)还是高盐(150 mmol·L-1 NaCl)条件下, 小花碱茅PtAKT1选择性吸收K+的能力显著强于Na+ (selective absorption capacity for K+ over Na+, SA).进一步研究发现, PtAKT1的转录水平与SA值之间呈显著正相关, 可见盐胁迫下PtAKT1是调控小花碱茅K+选择性吸收的关键因子(Wang et al., 2015).与野生型相比, 盐胁迫下超表达PtAKT1的拟南芥转基因植株K+含量增加, 而Na+含量降低, 表明PtAKT1增强了转基因植株对K+的吸收能力, 从而提高其耐盐性(Ardie et al., 2010).Duan等(2015)研究发现, 盐胁迫下盐地碱蓬SsAKT1能促进植株体内K+积累, 进而增强植物耐盐性.在盐(100 mmol·L-1 NaCl)处理下, 大麦HvAKT1在叶伸长区的表达水平明显增加, 说明HvAKT1参与维持叶肉细胞K+的稳态平衡(Boscari et al., 2009).我们的研究发现, 在5-50 mmol·L-1 NaCl处理下, 分别添加3和6 mmol·L-1 Cs+ (K+通道AKT1特异性抑制剂)均可显著降低甜菜(Beta vulgaris)根的K+净吸收速率, 可见AKT1可介导甜菜根K+的吸收(Wu et al., 2015b).这些研究结果表明, AKT1在K+和Na+选择性吸收、维持植物体内K+含量以及增强植物耐盐性等方面具有重要作用. ...
1 2004
... 干旱是制约全球作物生长和产量的主要非生物因素之一(Chaves and Oliveira, 2004; 山仑, 2011).植物响应干旱胁迫的重要策略之一是吸收并在体内积累大量的溶质(如K+、Na+和脯氨酸等) (Wang et al., 2004; Mahouachi et al., 2006; Wu et al., 2015a), 降低细胞渗透势, 从而增强植物细胞吸水能力.AKT1在植物响应干旱胁迫中的作用主要有2方面.(1) 干旱胁迫下AKT1增强植物根的K+吸收能力.在水稻中超表达OsAKT1基因后, 转基因植株在干旱条件下生长良好, 且根中积累较多的K+; 相反, Osakt1突变体植物表现为生长不良, 且根中K+含量明显减少(Ahmad et al., 2016).由此表明, 超表达AKT1转基因植株通过增加根部K+的积累来改善细胞渗透调节能力, 从而增强植物的抗旱性.(2) AKT1通过调节气孔的运动影响植物体内的水分平衡.研究表明, 拟南芥AtAKT1在保卫细胞质膜上呈低丰度表达(Lagarde et al., 1996).在渗透胁迫下, 拟南芥Atakt1突变体植株的蒸腾速率和水分消耗量明显低于野生型植株(Nieves- Cordones et al., 2012).由此可见, AKT1功能缺失致使保卫细胞K+跨膜转运体系受损, 气孔导度下降.Ahmad等(2016)进一步研究发现, 与野生型相比, 在干旱胁迫下水稻Osakt1突变体植株的气孔导度明显降低; 而超表达OsAKT1显著增强了转基因植株的气孔导度.这些研究结果表明, AKT1在介导保卫细胞K+转运、调控气孔运动及维持细胞水分平衡中发挥重要作用. ...
2016
2 2010
... AtKC1是Reintanz等(2002)从拟南芥中分离克隆的α-亚基Shaker通道基因, 主要在根毛、根表皮及皮层细胞中表达.AtKC1作为四聚体的组成亚基参与K+通道四聚体的形成, 其存在对四聚体的K+通透能力产生抑制作用(Reintanz et al., 2002; Geiger et al., 2009).可见, AtKC1是负调控因子, 也是1个调控亚基(Wang et al., 2010).另外, AtKC1的存在使四聚体的K+通道活性激活电压向负电压方向偏移, 离子通道更难以被激活, 即改变了其对电压的敏感性(Duby et al., 2008).因此, 这种负向调控阻断了由AKT1介导的吸收.短时间缺K+处理则上调AtKC1的转录丰度, 随后其转录丰度下调(Shin and Schachtman, 2004), 这种现象可通过缺K+处理几天后的AtKC1转录物消失而得到解释(Pilot et al., 2003).在异源表达系统中, AtKC1不能独立形成有功能的K+通道(Duby et al., 2008).然而, 当AtKC1不能正常发挥功能时, 根毛中AKT1介导的内向K+电流的生物物理特征就会发生改变(Jeanguenin et al., 2011).Wang等(2016)进一步研究证实, 在低K+条件下, CIPK23促进AKT1介导的K+吸收, 而AtKC1抑制AKT1介导的K+内流.他们认为, CIPK23和AtKC1协同调节AKT1的功能, 二者具有一定的互补性.另有研究表明, Atkc1与Atakt1突变体植株的表型相似或相反, 这主要取决于外界环境条件(Geiger et al., 2009; Wang et al., 2010).AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009).低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似.在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014).最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014).PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性.可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014). ... ... ; Wang et al., 2010).AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009).低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似.在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014).最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014).PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性.可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014). ...
3 2013
... K+是植物细胞中含量最丰富的无机阳离子之一, 主要通过根表皮细胞和皮层细胞的质膜K+转运系统吸收并运输到植物体内(王毅和武维华, 2009; Alemán et al., 2011; Dreyer and Uozumi, 2011; Wang and Wu, 2013; Nieves-Cordones et al., 2014a).K+在植物生长发育、逆境胁迫响应及生理生化反应等方面起着重要作用(Alemán et al., 2011; Nieves-Cordones et al., 2014a).一般而言, 植物细胞维持正常新陈代谢所需K+的最佳浓度为100 mmol·L-1, 但土壤中能被植物利用的K+浓度仅为0.1-1 mmol·L-1, 在一些盐碱地中甚至更低(Maathuis and Sanders, 1993; Maathuis, 2009).因此, 土壤中的K+远远不能满足植物正常生长需要, 使植物经常处于K+饥饿状态, 严重影响其生长发育.鉴于复杂多变的环境及K+的重要作用, 众多研究者对植物K+吸收和转运机制进行了深入研究. ... ... 植物在长期进化过程中形成了一系列复杂的K+吸收和转运系统.已知在模式植物拟南芥(Arabi- dopsis thaliana)中存在15个K+选择性通道, 包括9个Shaker型、5个TPK (two-pore K+ channel)型和1个Kir-like型K+通道(Ward et al., 2009; Hedrich, 2012; Demidchik, 2014).另外, 一些非选择性阳离子通道(non-selective cation channels, NSCC)也可以通透K+, 主要包括20个CNGC (cyclic nucleotide gated channels)通道、20个GLR (glutamate receptor-like)通道和1个TPC (two-pore channel)通道(Shabala, 2003; Demidchik and Maathuis, 2007; Dietrich et al., 2010; Hedrich, 2012; Demidchik, 2014).植物K+转运蛋白主要包括KUP (K+ uptake permease)/HAK (high-affinity K+ transporter)/KT (K+ transporter)、HKT (high-affinity K+ transporter)和CPA (cation proton antiporter) (Gierth and Mäser, 2007; Kron- zucker and Britto, 2011; Wang and Wu, 2013; Han et al., 2016).近年来, 人们相继在植物中克隆到编码这些蛋白的基因, 为深入研究植物K+吸收机制奠定了坚实的基础. ... ... AKT1 (Arabidopsis K+ transporter 1)是植物中广泛存在的一类内向整流K+通道, 与动物中的Shaker型K+通道具有高度的同源性(Pilot et al., 2003).Sentenac等(1992)利用酵母双突变体互补法从拟南芥中克隆到第1个高等植物的Shaker型K+通道基因AtAKT1.该基因位于拟南芥第2号染色体上, 含有11个外显子和10个内含子, 编码的多肽由857个氨基酸残基组成.此后, 科研人员相继在马铃薯(Sola- num tuberosum) (Zimmermann et al., 1998)、玉米(Zea mays) (Philippar et al., 1999)、小麦(Triticum aestivum) (Buschmann et al., 2000)、番茄(Lycoper- sicon esculentum) (Hartje et al., 2000)、水稻(Oryza sativa) (Fuchs et al., 2005)、大麦(Hordeum vulgare) (Boscari et al., 2009)、葡萄(Vitis vinifera) (Cuéllar et al., 2010, 2013)和盐地碱蓬(Suaeda salsa) (Duan et al., 2015)等植物中分离得到AKT1基因.AKT1在维持植物离子稳态平衡及增强植物逆境胁迫耐受性等方面起着重要作用(Alemán et al., 2011; Wang and Wu, 2013).本文从植物AKT1的分子结构特点、调控机制及生物学功能等方面对其主要研究进展进行了归纳总结, 旨在为植物AKT1的进一步研究奠定基础. ...
1 2009
... 植物在长期进化过程中形成了一系列复杂的K+吸收和转运系统.已知在模式植物拟南芥(Arabi- dopsis thaliana)中存在15个K+选择性通道, 包括9个Shaker型、5个TPK (two-pore K+ channel)型和1个Kir-like型K+通道(Ward et al., 2009; Hedrich, 2012; Demidchik, 2014).另外, 一些非选择性阳离子通道(non-selective cation channels, NSCC)也可以通透K+, 主要包括20个CNGC (cyclic nucleotide gated channels)通道、20个GLR (glutamate receptor-like)通道和1个TPC (two-pore channel)通道(Shabala, 2003; Demidchik and Maathuis, 2007; Dietrich et al., 2010; Hedrich, 2012; Demidchik, 2014).植物K+转运蛋白主要包括KUP (K+ uptake permease)/HAK (high-affinity K+ transporter)/KT (K+ transporter)、HKT (high-affinity K+ transporter)和CPA (cation proton antiporter) (Gierth and Mäser, 2007; Kron- zucker and Britto, 2011; Wang and Wu, 2013; Han et al., 2016).近年来, 人们相继在植物中克隆到编码这些蛋白的基因, 为深入研究植物K+吸收机制奠定了坚实的基础. ...
1 2015
... 干旱是制约全球作物生长和产量的主要非生物因素之一(Chaves and Oliveira, 2004; 山仑, 2011).植物响应干旱胁迫的重要策略之一是吸收并在体内积累大量的溶质(如K+、Na+和脯氨酸等) (Wang et al., 2004; Mahouachi et al., 2006; Wu et al., 2015a), 降低细胞渗透势, 从而增强植物细胞吸水能力.AKT1在植物响应干旱胁迫中的作用主要有2方面.(1) 干旱胁迫下AKT1增强植物根的K+吸收能力.在水稻中超表达OsAKT1基因后, 转基因植株在干旱条件下生长良好, 且根中积累较多的K+; 相反, Osakt1突变体植物表现为生长不良, 且根中K+含量明显减少(Ahmad et al., 2016).由此表明, 超表达AKT1转基因植株通过增加根部K+的积累来改善细胞渗透调节能力, 从而增强植物的抗旱性.(2) AKT1通过调节气孔的运动影响植物体内的水分平衡.研究表明, 拟南芥AtAKT1在保卫细胞质膜上呈低丰度表达(Lagarde et al., 1996).在渗透胁迫下, 拟南芥Atakt1突变体植株的蒸腾速率和水分消耗量明显低于野生型植株(Nieves- Cordones et al., 2012).由此可见, AKT1功能缺失致使保卫细胞K+跨膜转运体系受损, 气孔导度下降.Ahmad等(2016)进一步研究发现, 与野生型相比, 在干旱胁迫下水稻Osakt1突变体植株的气孔导度明显降低; 而超表达OsAKT1显著增强了转基因植株的气孔导度.这些研究结果表明, AKT1在介导保卫细胞K+转运、调控气孔运动及维持细胞水分平衡中发挥重要作用. ...
1 2015
... 盐胁迫下, 植物体内积累大量的Na+, 一方面抑制K+的吸收, 另一方面与K+竞争一些酶活性结合位点, 影响蛋白质合成及核糖体功能, 从而产生Na+毒害(Horie et al., 2009; Zhang et al., 2010).一般认为, 维持细胞质内较高的K+/Na+比是植物适应盐胁迫的重要策略之一(Horie et al., 2009; Shabala et al., 2010; Chérel et al., 2014).AKT1是参与K+吸收的重要通道蛋白, 在植物响应盐胁迫中起着重要作用.异源表达系统研究表明, 尽管AKT1对K+具有更强的选择性, 但并不排除对Na+的吸收(Maathuis et al., 1997).低Na+条件下, 内向整流K+通道未表现出明显的Na+进出现象; 但在高盐条件下, 其对K+表现出明显的选择性(Amtmann and Sanders, 1999).Goll- dack等(2003)研究表明, 盐处理下水稻根吸收Na+与OsAKT1的表达水平有直接关系.由此证明OsAKT1参与盐胁迫下Na+/K+比调控, 从而增强水稻的耐盐性.Wang等(2015)研究发现, 无论是低盐(25 mmol·L-1 NaCl)还是高盐(150 mmol·L-1 NaCl)条件下, 小花碱茅PtAKT1选择性吸收K+的能力显著强于Na+ (selective absorption capacity for K+ over Na+, SA).进一步研究发现, PtAKT1的转录水平与SA值之间呈显著正相关, 可见盐胁迫下PtAKT1是调控小花碱茅K+选择性吸收的关键因子(Wang et al., 2015).与野生型相比, 盐胁迫下超表达PtAKT1的拟南芥转基因植株K+含量增加, 而Na+含量降低, 表明PtAKT1增强了转基因植株对K+的吸收能力, 从而提高其耐盐性(Ardie et al., 2010).Duan等(2015)研究发现, 盐胁迫下盐地碱蓬SsAKT1能促进植株体内K+积累, 进而增强植物耐盐性.在盐(100 mmol·L-1 NaCl)处理下, 大麦HvAKT1在叶伸长区的表达水平明显增加, 说明HvAKT1参与维持叶肉细胞K+的稳态平衡(Boscari et al., 2009).我们的研究发现, 在5-50 mmol·L-1 NaCl处理下, 分别添加3和6 mmol·L-1 Cs+ (K+通道AKT1特异性抑制剂)均可显著降低甜菜(Beta vulgaris)根的K+净吸收速率, 可见AKT1可介导甜菜根K+的吸收(Wu et al., 2015b).这些研究结果表明, AKT1在K+和Na+选择性吸收、维持植物体内K+含量以及增强植物耐盐性等方面具有重要作用. ...
2 2014
... AtKC1是Reintanz等(2002)从拟南芥中分离克隆的α-亚基Shaker通道基因, 主要在根毛、根表皮及皮层细胞中表达.AtKC1作为四聚体的组成亚基参与K+通道四聚体的形成, 其存在对四聚体的K+通透能力产生抑制作用(Reintanz et al., 2002; Geiger et al., 2009).可见, AtKC1是负调控因子, 也是1个调控亚基(Wang et al., 2010).另外, AtKC1的存在使四聚体的K+通道活性激活电压向负电压方向偏移, 离子通道更难以被激活, 即改变了其对电压的敏感性(Duby et al., 2008).因此, 这种负向调控阻断了由AKT1介导的吸收.短时间缺K+处理则上调AtKC1的转录丰度, 随后其转录丰度下调(Shin and Schachtman, 2004), 这种现象可通过缺K+处理几天后的AtKC1转录物消失而得到解释(Pilot et al., 2003).在异源表达系统中, AtKC1不能独立形成有功能的K+通道(Duby et al., 2008).然而, 当AtKC1不能正常发挥功能时, 根毛中AKT1介导的内向K+电流的生物物理特征就会发生改变(Jeanguenin et al., 2011).Wang等(2016)进一步研究证实, 在低K+条件下, CIPK23促进AKT1介导的K+吸收, 而AtKC1抑制AKT1介导的K+内流.他们认为, CIPK23和AtKC1协同调节AKT1的功能, 二者具有一定的互补性.另有研究表明, Atkc1与Atakt1突变体植株的表型相似或相反, 这主要取决于外界环境条件(Geiger et al., 2009; Wang et al., 2010).AtKC1也是SNARE (soluble N-ethylmaleimide-sensitive factor protein attachment protein receptor)蛋白SYP121 (syntaxin protein121)的靶蛋白, SYP121参与对AKT1活性的调控(Honsbein et al., 2009).低K+环境中, syp121突变体的根系变短, 其表型与Atkc1和Atakt1突变体相似.在爪蟾卵母细胞中共表达AtAKT1和AtKC1后, SYP121使得K+通道被激活的膜电势负值的绝对值更大, 这种调控方式导致K+流显著增加(Chérel et al., 2014).最近的研究表明, 高盐胁迫下拟南芥植株体内迅速积累一氧化氮(nitric oxi- de, NO), 细胞K+含量降低, 产生过量的吡哆醛5-磷酸(pyridoxal 5-phosphate, PLP) (Xia et al., 2014).PLP作为维生素B6的一种活性形式, 显著抑制爪蟾卵母细胞表达系统和拟南芥根原生质中K+通道AtAKT1的活性.可见, NO通过促进维生素B6 PLP的合成负向调控AKT1介导的K+吸收(Xia et al., 2014). ... ... 吸收(Xia et al., 2014). ...
4 2006
... 尽管自20世纪90年代从拟南芥中克隆到第1个AKT1基因以来, 人们相继在其它植物中克隆得到同源基因.然而, AKT1基因所编码蛋白的调控机制仍不清楚.Xu等(2006)首次在植物中报道了低K+条件下蛋白磷酸化对K+吸收的调控机制.在爪蟾卵母细胞(Xeno- pus oocytes)中, CBL1 (calcineurin B-like protein1)/ CBL9与CIPK23 (CBL-interacting protein kinases 23)互作, 通过磷酸化作用激活AKT1介导的K+吸收(Li et al., 2006; Xu et al., 2006; Cuéllar et al., 2010, 2013).低K+处理下, CIPK23表达上调, 而CBL1和CBL9似乎是组成型表达(Xu et al., 2006; Cheong et al., 2007); 过量表达CIPK23可显著增强拟南芥K+的吸收能力和低K+耐受性(Xu et al., 2006).CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ... ... 似乎是组成型表达(Xu et al., 2006; Cheong et al., 2007); 过量表达CIPK23可显著增强拟南芥K+的吸收能力和低K+耐受性(Xu et al., 2006).CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ... ... 耐受性(Xu et al., 2006).CBL结构中有4个EF手型结构, 能与Ca2+结合; CBL是Ca2+信号感受器(Li et al., 2006, 2009).由于CIPK与CBL互作的特异性, 在植物逆境响应中有多种不同的组合(Li et al., 2009).AKT1-CIPK-CBL复合物能被PP2C (protein phosphatases 2C)负向调控, 参与对ABA信号的响应(Lee et al., 2007; Lan et al., 2011).Nieves- Cordones等(2012)研究发现, AKT1在保卫细胞中受到CIPK23调节, 并参与水分胁迫响应.此外, CBL10能直接与AKT1相互作用, 通过与CIPK23竞争结合AKT1, 从而负向调控K+通道活性(Grefen and Blatt, 2012; Ren et al., 2013).CIPK23也可通过磷酸化作用激活HAK5介导的高亲和性K+吸收(Ragel et al., 2015). ... ... 低K+环境是影响植物正常生长发育及作物品质的主要非生物因子之一.K+是种子萌发所需的重要渗透调节物质.在低K+ (50-100 μmol·L-1)条件下, 拟南芥Atakt1突变体种子的胚根不能正常生长; 然而, 当在Atakt1突变体中超表达AtAKT1后, 其种子萌发活力得到恢复, 说明AKT1参与种子萌发期间K+的吸收(Pyo et al., 2010; Xu et al., 2014).AKT1功能缺失使植物根细胞膜表现出超级化, 对外界环境K+浓度变化异常敏感(Hirsch et al., 1998).在低K+环境中, 拟南芥Atakt1突变体植株细胞内的K+含量明显降低(Hirsch et al., 1998; Spalding et al., 1999).电生理学实验表明, 水稻OsAKT1介导内向K+电流, 即使在低K+ (5 μmol·L-1-1 mmol·L-1)条件下也是如此(Li et al., 2014).有研究表明, 拟南芥Atakt1和水稻Osakt1突变体植株的内向K+电流减弱, K+吸收功能紊乱, 从而影响其生长发育(Reintanz et al., 2002; Xu et al., 2006; Li et al., 2014).当在水稻Osakt1突变体中超表达OsAKT1后, 低K+胁迫下转基因植株的内向K+电流增强, K+含量显著增加(Li et al., 2014).这进一步证实了AKT1介导植物根吸收K+.Xu等(2014)采用非损伤微测技术(no-invasive micro-test technology, NMT), 在棉花GhAKT1功能研究中也得到类似的结果.可见, 在低K+环境中, AKT1介导的K+吸收系统对维持植物正常生长发育具有重要作用. ...
2 2014
... 在K+饥饿状态下, 小麦根TaAKT1 (Buschmann et al., 2000)和小花碱茅(Puccinellia tenuiflora) PtAKT1 (Ardie et al., 2010)的转录水平显著提高.在拟南芥中, 外部K+浓度在5 μmol·L-1至5 mmol·L-1范围内未影响AtAKT1的转录丰度(Lagarde et al., 1996).在棉花(Xu et al., 2014)和小花碱茅(Wang et al., 2015)中也有类似报道, 由此推测AKT1可能受转录后调控.然而, 25-250 mmol·L-1 NaCl短期(6小时)处理能够诱导盐地碱蓬根SsAKT1的表达(Duan et al., 2015).研究表明, 水稻OsAKT1主要在根的外皮层和内皮层细胞以及叶的木质部薄壁组织、韧皮部和叶肉细胞中表达(Golldack et al., 2003).盐胁迫(150 mmol·L-1 NaCl处理48小时)下, 盐敏感型水稻品种Pokkali和BK根外皮层细胞中的OsAKT1转录物消失, 而耐盐型品种IR29中的则未受到影响(Golldack et al., 2003).Su等(2001)的研究表明, 盐胁迫下盐生植物冰叶日中花(Mesembryanthemum crystallinum)根中与AtAKT1同源的K+通道基因McAKT1表达水平下调.在小花碱茅中, 尽管盐处理会下调PtAKT1的表达, 但150 mmol·L-1 NaCl处理下PtAKT1的转录丰度显著高于25 mmol·L-1 (Wang et al., 2015).然而, 也有研究报道认为, NaCl短期处理并未改变AKT1的表达水平(Maathuis et al., 2003; Pilot et al., 2003).在葡萄中, VvAKT1.1主要在根和浆果中表达(Cuéllar et al., 2010); 而VvAKT1.2主要在成熟的浆果中表达, 干旱胁迫可显著提高其表达水平(Cuéllar et al., 2013).此外, 产生活性氧(reactive oxygen species, ROS)的胁迫因子(如砷、缺氧和热胁迫等)均可下调AKT1的转录丰度(Ahmad et al., 2016).由此可见, 植物AKT1的表达不但具有组织特异性, 而且受外界环境的影响. ... ... 低K+环境是影响植物正常生长发育及作物品质的主要非生物因子之一.K+是种子萌发所需的重要渗透调节物质.在低K+ (50-100 μmol·L-1)条件下, 拟南芥Atakt1突变体种子的胚根不能正常生长; 然而, 当在Atakt1突变体中超表达AtAKT1后, 其种子萌发活力得到恢复, 说明AKT1参与种子萌发期间K+的吸收(Pyo et al., 2010; Xu et al., 2014).AKT1功能缺失使植物根细胞膜表现出超级化, 对外界环境K+浓度变化异常敏感(Hirsch et al., 1998).在低K+环境中, 拟南芥Atakt1突变体植株细胞内的K+含量明显降低(Hirsch et al., 1998; Spalding et al., 1999).电生理学实验表明, 水稻OsAKT1介导内向K+电流, 即使在低K+ (5 μmol·L-1-1 mmol·L-1)条件下也是如此(Li et al., 2014).有研究表明, 拟南芥Atakt1和水稻Osakt1突变体植株的内向K+电流减弱, K+吸收功能紊乱, 从而影响其生长发育(Reintanz et al., 2002; Xu et al., 2006; Li et al., 2014).当在水稻Osakt1突变体中超表达OsAKT1后, 低K+胁迫下转基因植株的内向K+电流增强, K+含量显著增加(Li et al., 2014).这进一步证实了AKT1介导植物根吸收K+.Xu等(2014)采用非损伤微测技术(no-invasive micro-test technology, NMT), 在棉花GhAKT1功能研究中也得到类似的结果.可见, 在低K+环境中, AKT1介导的K+吸收系统对维持植物正常生长发育具有重要作用. ...
1 2010
... 盐胁迫下, 植物体内积累大量的Na+, 一方面抑制K+的吸收, 另一方面与K+竞争一些酶活性结合位点, 影响蛋白质合成及核糖体功能, 从而产生Na+毒害(Horie et al., 2009; Zhang et al., 2010).一般认为, 维持细胞质内较高的K+/Na+比是植物适应盐胁迫的重要策略之一(Horie et al., 2009; Shabala et al., 2010; Chérel et al., 2014).AKT1是参与K+吸收的重要通道蛋白, 在植物响应盐胁迫中起着重要作用.异源表达系统研究表明, 尽管AKT1对K+具有更强的选择性, 但并不排除对Na+的吸收(Maathuis et al., 1997).低Na+条件下, 内向整流K+通道未表现出明显的Na+进出现象; 但在高盐条件下, 其对K+表现出明显的选择性(Amtmann and Sanders, 1999).Goll- dack等(2003)研究表明, 盐处理下水稻根吸收Na+与OsAKT1的表达水平有直接关系.由此证明OsAKT1参与盐胁迫下Na+/K+比调控, 从而增强水稻的耐盐性.Wang等(2015)研究发现, 无论是低盐(25 mmol·L-1 NaCl)还是高盐(150 mmol·L-1 NaCl)条件下, 小花碱茅PtAKT1选择性吸收K+的能力显著强于Na+ (selective absorption capacity for K+ over Na+, SA).进一步研究发现, PtAKT1的转录水平与SA值之间呈显著正相关, 可见盐胁迫下PtAKT1是调控小花碱茅K+选择性吸收的关键因子(Wang et al., 2015).与野生型相比, 盐胁迫下超表达PtAKT1的拟南芥转基因植株K+含量增加, 而Na+含量降低, 表明PtAKT1增强了转基因植株对K+的吸收能力, 从而提高其耐盐性(Ardie et al., 2010).Duan等(2015)研究发现, 盐胁迫下盐地碱蓬SsAKT1能促进植株体内K+积累, 进而增强植物耐盐性.在盐(100 mmol·L-1 NaCl)处理下, 大麦HvAKT1在叶伸长区的表达水平明显增加, 说明HvAKT1参与维持叶肉细胞K+的稳态平衡(Boscari et al., 2009).我们的研究发现, 在5-50 mmol·L-1 NaCl处理下, 分别添加3和6 mmol·L-1 Cs+ (K+通道AKT1特异性抑制剂)均可显著降低甜菜(Beta vulgaris)根的K+净吸收速率, 可见AKT1可介导甜菜根K+的吸收(Wu et al., 2015b).这些研究结果表明, AKT1在K+和Na+选择性吸收、维持植物体内K+含量以及增强植物耐盐性等方面具有重要作用. ...
1 1998
... AKT1 (Arabidopsis K+ transporter 1)是植物中广泛存在的一类内向整流K+通道, 与动物中的Shaker型K+通道具有高度的同源性(Pilot et al., 2003).Sentenac等(1992)利用酵母双突变体互补法从拟南芥中克隆到第1个高等植物的Shaker型K+通道基因AtAKT1.该基因位于拟南芥第2号染色体上, 含有11个外显子和10个内含子, 编码的多肽由857个氨基酸残基组成.此后, 科研人员相继在马铃薯(Sola- num tuberosum) (Zimmermann et al., 1998)、玉米(Zea mays) (Philippar et al., 1999)、小麦(Triticum aestivum) (Buschmann et al., 2000)、番茄(Lycoper- sicon esculentum) (Hartje et al., 2000)、水稻(Oryza sativa) (Fuchs et al., 2005)、大麦(Hordeum vulgare) (Boscari et al., 2009)、葡萄(Vitis vinifera) (Cuéllar et al., 2010, 2013)和盐地碱蓬(Suaeda salsa) (Duan et al., 2015)等植物中分离得到AKT1基因.AKT1在维持植物离子稳态平衡及增强植物逆境胁迫耐受性等方面起着重要作用(Alemán et al., 2011; Wang and Wu, 2013).本文从植物AKT1的分子结构特点、调控机制及生物学功能等方面对其主要研究进展进行了归纳总结, 旨在为植物AKT1的进一步研究奠定基础. ...
 ,
, 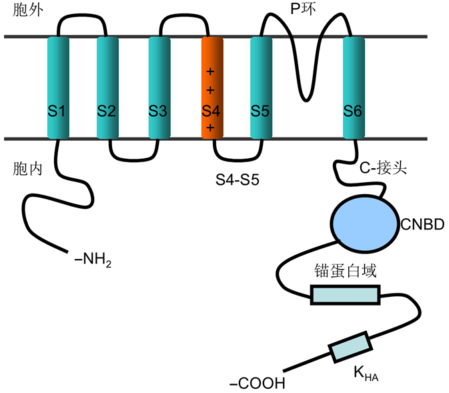


{kind=link}
{kind=link}
{kind=link}
{kind=link}