 ,
, ,*, 刘洪妤
,*, 刘洪妤Using Strauss-Hardcore model to detect vessel spatial distribution in angiosperms with various vessel configurations
Jing-Ming ZHENG,,*, Hong-Yu LIU通讯作者: * 郑景明: ORCID:0000-0002-8517-9999,E-mail:zhengjm@bjfu.edu.cn
编委: 梁尔源
责任编辑: 李敏
收稿日期:2021-03-10接受日期:2021-05-19
| 基金资助: |
Corresponding authors: * E-mail:zhengjm@bjfu.edu.cn
Received:2021-03-10Accepted:2021-05-19
| Fund supported: |

摘要
被子植物木质部导管的分布格局非常多样, 并且与木质部的输水功能有密切的联系, 然而在木材解剖学中对导管分布格局往往采用定性描述, 不利于分析该特征与物种的水力功能、生态地理分布的关系。该文采用点格局分析方法, 依据木材孔型、导管空间排列和导管群集度三类木材宏观结构特征组合, 选取不同导管分布类型的17种代表性阔叶树种, 利用Strauss-Hardcore模型对其木质部横切面解剖影像进行定量分析。Strauss-Hardcore模型能够很好地拟合木质部中导管二维空间位点的分布特征, 该模型的3个参数: 硬核距离、局部聚集距离、点对交互作用强度(局部聚集指数)都有着明确的生物学意义。传统解剖学对导管构型的定性分类同模型相比不能准确表现被子植物的木质部导管空间分布特征, Strauss-Hardcore模型的局部聚集度指数主要受导管群集度影响, 尤其是复导管和导管团的存在都会增大导管小尺度聚集程度。对散孔材、半环孔材的生长轮及环孔材的晚材部分解剖图像分析表明, 导管以单导管为主且没有明显分布方向的散孔材树种, 其木质部导管点对交互作用强度为负值, 局部聚集指数一般小于0.4, 导管空间分布依次在3个局部尺度表现出排斥-排斥-随机格局; 而导管具有径向、弦向、锯齿形等明显目视识别特征的物种, 无论孔型和是否以单复导管为主, 其导管点对交互作用强度为正值, 局部聚集指数均大于0.4, 导管依次在3个局部尺度上表现出排斥-聚集-随机的分布格局。采用点过程模型有利于准确描述导管二维空间分布规律, 增强对导管空间格局形成机理的理解, 可有力地支撑木质部三维导管系统的理论研究和木质部结构-功能的实验研究。
关键词:
Abstract
Aims Spatial patterns of vessel in xylem are diverse and closely related with water transportation functions in angiosperms. However, the pattern was generally described qualitatively in anatomy, which were unable to reveal their links to xylem functions and to species distribution. We used point pattern analysis to study vessel spatial pattern in xylem cross-sectional images to quantify their features. Methods Images of 17 types of vessel configurations were selected in terms of wood porosity, vessel arrangement, and vessel grouping. Optimum Strauss-Hardcore models for coordinates in the images were fitted. Correlations among vessel variables and model coefficients were tested. Important findings We found that (1) Strauss-Hardcore model fitted all the data well and its three parameters, i.e., hardcore distance, local aggregation distance, and point-pair interaction or point aggregation index, and had apparent biological significance. (2) Classifications of wood xylem by traditional anatomical indices could not precisely present the spatial pattern of vessels compared to spatial point analysis, and local aggregation index from Strauss-Hardcore model was mainly influenced by vessel grouping, especially frequency of radial multiples and vessel clusters. (3) Among the 17 vessel patterns analyzed, diffusive or semi-ring species with xylem consisting of solidary vessels showed negative point-pair interaction and aggregation index was less than 0.4, whereas species with obvious vessel arrangement and multiple or clusters of vessel grouping in xylem owned positive point-pair interaction and bigger aggregation index. (4) The former group of species demonstrated inhibition- inhibition-random pattern at three local scales while the latter species showed inhibition-aggregation-random pattern according to the fitted Strauss-Hardcore models. The findings showed that point process modeling could precisely describe vessel distribution features in 2-D xylem sections and provide insights on vessel development. Therefore, this method may support 3-D vessel system simulation and experimental studies on structure-function of angiosperm xylem.
Keywords:
PDF (1240KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
郑景明, 刘洪妤. 采用Strauss-Hardcore模型研究不同导管构型被子植物的导管空间分布特征. 植物生态学报, 2021, 45(9): 1024-1032. DOI: 10.17521/cjpe.2021.0083
ZHENG Jing-Ming, LIU Hong-Yu.

树木木质部在植物水分运输效率以及输水安全方面发挥着重要作用, 理解其结构和功能之间的关系对于理解森林响应全球变化的机理有重要的意义(Choat et al., 2012)。裸子植物的木质部主要由管胞和射线薄壁细胞构成, 结构相对均质; 而被子植物的木质部是由导管、轴向和径向薄壁细胞、纤维等细胞共同构成的一个复杂组织, 并同时完成水分运输、机械支撑、物质储藏以及防御等多项功能(Pfautsch, 2016)。被子植物的导管进化及不同组织分工而导致的输水功能提升被认为是白垩纪后期裸子植物衰落而被子植物繁盛的主要原因(Carlquist, 2012)。全球气候变化下, 欧洲及北美一些地区的森林树木由于频繁、持续的干旱而死亡, 木质部水力失效是其中一个最重要的机理假说(Adams et al., 2017), 被子植物木质部的结构和功能关系受到广泛关注。
被子植物的木质部具有极高的导管构型多样性, 包括早晚材导管差异、导管大小和数量、导管的空间分布格局、复导管(木质部横切面上具有共同细胞壁的相邻导管, 多为径向排列)和管孔团(多个导管共用相邻细胞壁, 或者局部单导管明显聚集)出现频率等。传统木材解剖研究对木质部导管空间排列方式的研究主要是通过观察木材横切面上的导管分布特征并做定性描述, 例如, 根据导管空间排列模式分为散生、径列、斜列、弦列等(Wheeler & Baas, 1998)。这种途径虽然有利于木材识别, 但无法将物种的木质部结构特征与功能联系起来, 因而缺乏其与木质部功能的相关性探讨(Martínez-Vilalta et al., 2012)。树木水分生理、生态学研究中, 一般采用导管群聚度(vessel grouping)的指标来刻画导管聚集程度, 它主要指示的是单复导管比例(Scholz et al., 2013)。这种方法强调了复导管在导水效率和安全方面的作用, 在一定程度上把木质部的结构和功能联系起来, 但忽略了比导管细胞更大尺度上的导管分布特征, 例如一些复杂的导管排列模式, 如某些物种的导管分布在径向或斜向伸展看起来越过了生长轮边界(称为溪流状、火焰状排列), 或者在弦向上构成形状繁复的构型(称为花彩状排列)等, 能够跨越数个年轮存在, 并可能对整个木质部的导水功能有重要的影响(Carlquist, 2001)。随着现代立体成像技术引入木质部解剖、导水生理研究中, 近期对二维导管空间分布格局、三维导管网络特征及其输水功能的关系研究有了进一步发展。现有实验和模型研究表明, 聚集分布导管的物种具有较高的相邻导管连通性, 因此能获得较高的水力传导率; 而木质部导管系统的空间三维结构对木质部的导水效率和抗栓塞能力都有重要的影响(Loepfe et al., 2007; Martínez-Vilalta et al., 2012; Mrad et al., 2018)。因此定量分析导管在木质部中的二维空间分布特征可为木质部的结构-功能关系研究提供更多的视角, 并可以对精确模拟三维导管网络系统的结构提供理论支持。
空间点格局分析通常通过建立一定空间内相邻点对间相互作用的点过程模型, 研究在一定区域内对象(如样地中的植物个体、木质部影像中的导管)的空间分布格局特征及格局的形成机理。虽然点格局分析方法在种群生态学研究中已有较多应用, 但在导管分布格局研究方面则很少见。点过程模型种类很多, 本文首次采取斯特劳斯-硬核过程模型(Strauss-Hardcore model, 以下简称SH模型)拟合分析被子植物木质部导管空间格局。SH模型属于吉布斯混合模型(Hybrid Gibbs model), 由一个硬核模型(Hardcore model, 以下简称H模型)和一个斯特劳斯模型(Strauss model, 以下简称S模型)组合构成。H模型和S模型是两种描述相邻点对之间排斥-吸引作用过程的模型, 两者组合则可以描述两个不同尺度上的点对交互过程, 其中H模型描述较小尺度上的点对间排斥作用, S模型描述稍大尺度上的排斥或吸引作用(Baddeley et al., 2015)。
被子植物木质部的生长发育速度通常是有限的, 且因物种而异, 不同物种的年轮宽度、导管大小和密度、空间分布特征都具有发育保守性(Panshin & Zeeuw, 1980)。木质部中导管的主要功能是自下而上输导水分, 单一导管仅能为木质部局部提供水分输导, 多数导管协同才能为植株提供充足导水功能和降低因部分导管气穴化导致的木质部水力功能失效风险。通常各个导管之间因其他细胞类型(纤维、薄壁细胞等)的存在而保持一定距离, 因此可以把木质部所有导管在横切面上的位置作为点格局分析对象, 并可以理解为相邻导管之间存在互相排斥和吸引的关系, 从而采用SH模型来描述木质部导管空间分布格局和导管密度约束的机理。
国际木材解剖学家联合会(IAWA)从3方面指标对被子植物的木质部导管构型进行描述: 木材孔型(包括环孔材、半环孔材、散孔材3类)、导管排列模式(包括弦向带状分布、径向或斜向分布、锯齿形分布3类, 忽略了没有明显分布规律的类型), 导管群聚程度(包括单导管、径向复导管、导管团3类)(IAWA Committee, 1989)。本研究据此选取具有不同导管构型特征的代表树种, 收集其木质部横切面解剖图像, 利用空间点格局方法, 定量分析木质部导管的空间分布特征, 提出导管空间分布定量化描述指标并分析其生物学意义, 为进一步研究导管分布与导水功能关系提供新的角度和基础。
1 方法
1.1 图像的选取与处理
本研究依据IAWA的3类导管构型指标组合, 通过国际木材解剖网站InsideWood (1.2 空间点格局分析方法
SH模型包含了一个H模型, 描述任意相邻两个导管间距离不能超过某一固定值(即最近一对相邻导管半径之和); 同时它还包含一个S模型, 描述相邻导管对间的局部吸引聚集或排斥分散的特征。因此, SH模型允许在3个尺度上的点对间出现不同的相互作用方式, 即它可以表示小尺度排斥、吸引、均匀特征(公式1)。式中, c(u, v)为任意点对(u, v)间相互作用函数, h为硬核距离, r为局部相互作用距离, γ为局部聚集指数。点对间距离为r时的局部聚集指数γ实际上等于点对间相互作用程度s的指数函数, 即γ = exp(s), 因此两者意义相同, 后面相关分析时采用s进行。
SH模型包括两个距离参数, h (硬核距离)和r (局部聚集距离), 可通过设置调参范围根据拟合模型的赤池信息量准则(AIC)值得到最优参数组合的SH模型(AIC值最低模型为最优)。根据拟合成功的最佳SH模型, 可得到描述点格局强度的一个参数, 即点对局部聚集指数γ或点对间交互作用强度s。当γ = 0时, SH模型简化为一般硬核模型(h = r); γ = 1时, SH模型简化为一般硬核模型(h = h); 当0 < γ < 1时, 为硬核模型(h = h), 但带有一定程度的排斥格局(h-r间); 当γ > 1时, 为硬核模型但在0-h间为排斥格局, h-r间为吸引格局(Baddeley et al., 2015)。
对导管点位数据拟合SH模型, 首先根据点对最小邻体距离和最大邻体距离设置参数网格范围, 采用逼近最大似然法(Huang-Ogata approximate maximum likelihood)拟合HS模型系列, 根据拟合模型的AIC值最小的原则, 调整参数(h, c)组合至最佳(Baddeley et al., 2015)。之后, 采用该方法对最佳模型参数执行100次模型拟合以计算参数γ或s的中值和置信区间。模型的拟合优度检验采用L函数的包络检验(envelope test), L函数为点格局分析中常用的Ripley's K函数的变形, 包络检验是采用一定次数的蒙特卡洛模拟方法进行模型检验的常用方式(Perry et al., 2006)。
Ripley's K函数的表达式为:
式中, n为研究区域内事件的个数; A为研究区域的面积; d为空间尺度, 即圆的半径; uij为i个体和j个体之间的距离; Wij为以i为圆心uij为半径的圆落在面积A中的比例。
本研究采用的L函数的表达式:
对最佳(h, r)组合采用拟合好的模型公式随机拟合19次, 可获得的该模型L函数值的95%置信区间, 该检验的显著性为α = 1/(19 + 1) = 0.05。以r为横坐标、上下包迹线的数值为纵坐标作图, 当实际值的线位于上下包络线之间, 说明模型拟合成功, 否则需要调整参数重新进行拟合。通常木质部图像中导管分布的局部单位点密度在空间是不均匀的, 因此L函数包迹线计算采用异质函数形式(Mencuccini et al., 2010)。
本研究同时计算了生态学中常用的空间均质性程度指数(CE)作为空间点格局分析得到的聚集程度的参考指标。CE是基于相邻点对间距离指标计算, 采用实际点格局的平均点对距离除以同样点对密度但空间完全随机的点格局的平均点对距离得到的比值。CE被视为一种简单聚集指数, 可检验偏离完全空间随机分布格局程度, 通常采用蒙特卡洛检验(n = 1 000次), 在植物种群空间分布研究中常用, 但它存在明显的弱点是只能描述一个尺度上的空间点分布特征(Perry et al., 2006)。
本研究木质部解剖影像处理和导管信息提取采用ImageJ软件(Rueden et al., 2017), 点格局分析和相关统计分析采用R 3.6.1软件(R Core Team, 2015)。SH函数空间点格局模型的拟合和评价使用“spatstat”软件包, 其中SH模型拟合采用ppm函数, 包络检验采用envelop函数, 该软件包是专门为各类空间点格局分析方法而研制并不断更新中(Baddeley & Turner, 2005)。木质部特征的导管平均直径及SH模型距离指数h、r之间, 简单聚集指数CE与点间相互作用强度指数s间等指标间的相关显著性检验采用cor.test函数, 由于各变量数据有少数不符合正态分布, 因此相关系数采用斯皮尔曼相关系数。
2 结果
2.1 导管空间点格局分析过程及结果举例
以一个木质部导管空间分布接近随机状态的物种Stewartia pseudocamelliashi为例, 从解剖图像提取导管点位置数据, 采用SH模型对空间点位数据进行最佳参数拟合并采用L函数的蒙特卡洛检验方法检验拟合优度(图1)。图1
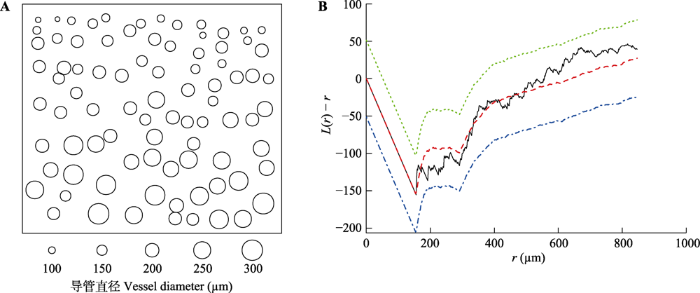 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1Stewartia pseudocamelliashi的导管空间分布及其Strauss-Hardcore (SH)模型的包络检验。A, 木质部横切面的导管分布情况。每个圆圈代表一个导管, 圆圈直径为导管直径(µm)。B, 显著性水平为0.05的L函数包络检验。r, 点对距离; 纵坐标为公式(3)定义的L函数。黑色线代表实际数据拟合得到SH模型的L函数值, 红色线代表采用相同参数的SH理论分布模型的19次拟合平均值, 绿色和蓝色线分别代
Fig. 1Spatial distribution of vessels in xylem of Stewartia pseudocamelliashi and envelope test of fitted Strauss-Hardcore (SH) model. A, Distribution of vessels in xylem cross-section. Each cycle stands for a vessel with cycle diameter as vessel diameter (µm). B, Envelope test for L function at significance of 0.05. r, the distance of paired points; Y-axis is the L function defined in equation (3). Black line represents L value from data fitted SH model, red line for average L value from 19 simulation of theoretical SH model, green and blue lines represent 2.5% and 97.5% quantile of L value from 19 simulation of theoretical SH model respectively.
S. pseudocamelliashi为散孔材, 导管为单管孔, 木质部横切面上导管空间分布目视较为随机。该幅图像有95个导管, 单位面积导管密度为5 ind.·mm-2, 平均导管直径135 µm, 导管对最近邻体距离为157 µm。通过调参-拟合得到SH模型的最佳参数组合为: h = 155 µm, r = 291 µm, s = -1.028 (-1.456, -0.600), γ = 0.358 (0.233, 0.549), 该模型通过了L函数的蒙特卡洛检验。最优模型结果表明, 该物种的导管分布在0-155 µm范围内相邻导管绝对排斥, 在155-291 µm范围内, 相邻导管之间存在一定强度的排斥作用, 大于291 µm后导管表现为随机分布格局。
2.2 不同导管构型的空间点格局分析结果
按照孔型、导管分布模式、导管群聚类型3类指标组合选出代表性木质部横切面图像数据的SH模型拟合结果见表1。其中孔型指标主要采用了散孔材与环孔材两个水平, 因为半环孔材的特征介于两者之间没有必要详尽罗列; 一些组合, 如散孔材+导管锯齿状空间分布+导管长径列群聚, 不存在典型物种因而不予考虑; 很多图像中导管径向排列和对角线排列难以区分, 则按照IAWA对空间排列的分类规定不予区分。根据上述3类指标的可能组合情况, 通过现有图像的筛选, 得到17个3类指标组合的代表性木质部解剖影像数据。由于被子植物木质部导管构型极其多样, 也可能存在其他组合或导管构型类型本文未纳入考虑, 有待后续进一步研究。木质部影像数据分析结果表明各物种均符合SH分布, 但h与r的分布范围因物种而异; 从γ分布范围看, 单管孔且目视没有明显导管空间分布样式(接近空间随机分布)的物种(如类型1-3), s < 0, γ最大为0.326, 而存在复导管的物种或导管在空间上有明显排布规律(如弦向、径向、斜向排列)的物种, s > 0, 其γ随复导管数量增多而增大; γ最大值出现在具有长径列复导管且整体上呈弦状排列的环孔材物种(类型13)的晚材中, 该物种为五加科树参属植物Dendropanax morbiferus。Table 1
表1
表1不同导管构型代表物种的Straus-Hardcore模型拟合结果
Table 1
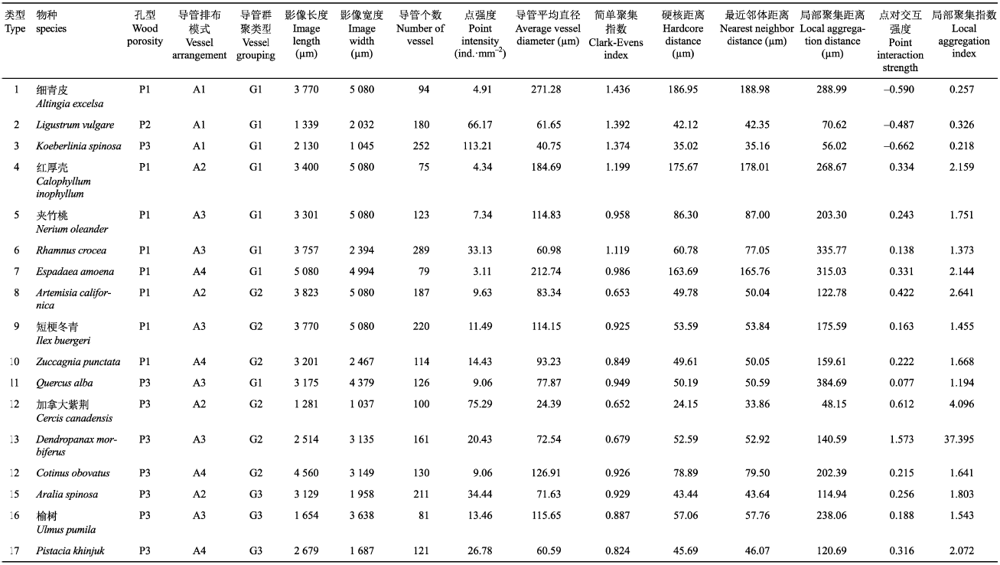 |
新窗口打开|下载CSV
2.3 木质部导管分布特征指标之间的相关性分析
根据SH模型得到的硬核距离与导管平均直径(r = 0.86, p < 0.001)、最近邻体距离间(r = 0.86, p < 0.001)均显著正相关。而根据SH模型得到的局部聚集距离与导管平均直径(r = 0.65, p < 0.01)、最近邻体距离间(r = 0.83, p < 0.001)也显著正相关。表明SH模型的两个距离参数与导管直径和导管间距离有密切联系。根据SH模型得到的点对相互作用强度指数与同样表征聚集程度的简单聚集指数之间(r = -0.68, p < 0.01)为显著负相关关系, 且导管密度与局部聚集指数间相关关系不显著(p > 0.05), 表明局部点对相互作用强度或局部点聚集指数可用于描述木质部导管的空间聚集分布程度, 且不受木质部导管密度影响。
3 讨论
被子植物木质部的导管空间分布格局多样性很高, 不仅可以用于木材种类或树木种类识别, 同时也代表着物种进化过程中的适应环境方式的多样化。木质部导管是植物运输水分的主要通道, 随着全球气候的变化, 植物导管的研究逐渐成为热点(Jacobsen et al., 2018), 木质部导管的空间格局定量化研究也逐渐引起关注(Mencuccini et al., 2010; Sievänen et al., 2014)。本研究采用空间点格局分析方法定量化描述木质部导管的空间分布特点, 初步研究结果表明该方法是可行的, 适用于低倍木质部解剖影像的分析。研究结果表明, 传统解剖学对木质部构型的定性描述不足以准确刻画被子植物木质部导管构型的多样性, 定性分类组合也不能与SH模型的结果构成一致性的对应。SH得到的定量特征值主要受导管群聚程度影响, 导管排列模式的影响主要体现在其中是否存在复导管和导管团, 如果仅由单导管组成则影响很小, 而孔型的作用主要在环孔材的分析木质部部位选择晚材部分而散孔材和半环孔材选用整个年轮, 对局部导管分布聚集程度没有影响。SH模型是基于点过程的机理模型, 可以解释不同点位间相互作用而导致的点格局, 因而不同于基于统计学的传统聚集度指标(如CE), 有助于分析木质部导管空间分布格局多样性背后的生理学、生态学机理。应当说明的是, 本研究中主要采用导管位置信息进行分析, 没有区分单复导管。虽然存在复导管物种确实会增大相应的γ, 然而目前对于单、复导管的输水功能是否存在差异还存在争议(Johnson et al., 2018)。根据导管发生和形成的生长素极性运输假说(canalizations hypothesis), 在木质部形成过程中, 在韧皮部产生的不同浓度生长素刺激下, 形成层原细胞向髓心方向分裂产生不同数量的纺锤状初始细胞和方形初始细胞, 纺锤状细胞进一步发育成为导管, 因而复导管在发育和功能上与单导管很可能没有本质差异(Hacke et al., 2017)。如果需要分别分析单、复导管空间分析格局, 则本方法可以进一步扩展, 采用包括两种点属性的SH模型模拟, 如多类别SH模型(MultiStraussHard), 其中一类点表示单导管而另一类点表示复导管(Baddeley & Turner, 2005)。
本研究通过点格局分析得到的导管分布格局指标具有明确的生物学意义。SH模型有3个量化指标——h、r以及γ, 它们分别指示导管在0-h、h-r、>r 3个局域尺度上导管空间分布的特点, 这意味着导管在木质部中的分布绝对不是完全随机的, 导管分布位置和局部密度受其木质部的多组织结构特点和多功能性约束(Carlquist, 2009)。h与导管平均直径和最近邻体距离显著相关, 意味着在小于h的范围不能存在其他的导管, 可以直接对应于复导管之间存在的最小距离决定于相邻导管半径之和的实际情况; 而在仅有单导管的情况下, h可以大于两个相邻导管半径之和(图1), 可以解释为每个导管仅能为其周围一定面积的木质部组织输水。r表示在h-r的距离范围内, 导管之间存在相互作用(吸引或排斥); 距离超过r的点对不影响点位整体的基线概率(完全随机), 即在大于r的尺度上可以视为随机分布。γ则代表在局部尺度的导管的聚集程度, 当其取值大于1时指示导管间相互吸引作用, 表现为导管的聚集程度高; 当其取值小于1时, 则指示导管间相互排斥作用, 表现为导管趋向于均匀分布。应该指出的是, 任何一种点格局模型都有一定的适用范围, SH模型对于某些植物木质部导管过于复杂的分布格局可能不一定适用, 因而该方法存在一定的局限性。例如, 当某些物种(如榆属(Ulmus)一些种)的一个年轮范围内存在明显的粗射线分隔的密集导管群, 或某些物种(如鼠李属(Rhamnus)一些种)的导管分布形成复杂的多孔网格状等, 这种情况下一个年轮内导管点位分布存在若干个点对交互作用尺度, 而SH模型仅能刻画其中较小尺度上的聚集程度。对此类复杂的导管空间分布特征定量化分析方法尚有待进一步深入研究(表2), 例如可以通过包含多个子模型的另一类混合吉布斯模型——组合式盖耶模型(Piecewise Geyer Model)拟合得到多个尺度上的聚集参数(Mencuccini et al., 2010)。
Table 2
表2
表2用于导管构型定量分析的不同空间点-过程模型的特点
Table 2
| 模型 Model | 点-过程特点 Point-process characteristic | 适用范围 Scenario for model application |
|---|---|---|
| 空间泊松模型 Poisson model | 空间点位置完全随机 Complete spatial randomness of points | 单一尺度, 单一导管属性, 随机分布特征 Single scale, single vessel identity, random distribution |
| 硬核模型 Hardcore model | 相邻点间距低于硬核距离则不能存在 Neighbor point is forbidden at distance smaller than hardcore distance | 单一尺度, 单一导管属性, 均匀分布特征 Single scale, single vessel identity, uniform distribution |
| 施特劳斯模型 Strauss model | 相邻点间距越小则出现概率越低 Neighbor points have lower probability with smaller distance between them | 单一尺度, 单一导管属性, 聚集分布特征 Single scale, single vessel identity, aggregation distribution |
| 盖耶饱和模型 Geyer saturation model | 任一点全部分布概率不超过特定值 Probability of each point is restrained at specific threshold value | 单一尺度, 单一导管属性, 聚集分布特征, 受导管密度影响 Single scale, single vessel identity, aggregation distribution, influenced by total vessel density |
| 多类型硬核模型 MultiHardcore model | 点属性2类以上的硬核模型 Hardcore model with more than two point identities | 单一尺度, 两类以上导管属性(如早、晚材导管, 单、复导管等), 同类导管均匀分布特征 Single scale, more than two vessel identities (e.g., vessel for early- and latewood, single vessel and multiple vessel), uniform distribution for each identity |
| 多类型施特劳斯模型 MultiStrauss model | 点属性2类以上的施特劳斯模型 Strauss model with more than two point identities | 单一尺度, 两类以上导管属性, 同类导管聚集分布特征 Single scale, more than two vessel identities, aggregation distribution for each identity |
| 斯特劳斯-硬核模型 Strauss-Hardcore model | 一个硬核模型和一个施特劳斯模型的组合 A combination of a Strauss model and a Hardcore model | 两个尺度, 单一属性的导管, 均匀-聚集分布特征 Two scales, single two vessel identity, uniform-aggregation distribution |
| 多类型施特劳斯-硬核模型 MultiStrauss-Hardcore model | 点属性2类以上的斯特劳斯-硬核模型 Strauss-Hardcore model with more than two point identities | 两个尺度, 两类以上导管属性, 同类导管不同尺度上呈均匀和聚集分布特征 Two scales, more than two vessel identities, uniform-aggregation distribution |
| 组合式盖耶模型 Piecewise Geyer model | 组合模型, 可包括多个盖耶饱和子模型、硬核子模型和施特劳斯子模型 A hybrid model including multiple sub-models such as Strauss model, Hardcore model, and Geyer saturation model | 多个尺度, 单一属性的导管, 均匀和聚集分布特征, 受导管总密度影响 More than two scales, single vessel identity, uniform-aggregation distribution, influenced by total vessel density |
新窗口打开|下载CSV
由于气候变化导致森林衰退树木死亡现象大范围出现, 对于木质部结构和功能的研究成为当前热点领域(Choat et al., 2018)。目前对导管细胞的发育过程的理解还不很清楚, 尚不能解释木质部导管空间格局的形成过程, 如横切面上导管的弦向带状分布、树干轴向上导管的长度分布等(Jacobsen et al., 2012; Hacke et al., 2017)。已有研究表明, 被子植物的导管分布特征对生境水分限制及全球气候变化存在明显的响应, 对导管分布格局的定量化分析有助于揭示全球气候变化下植物导水结构和功能的调整方式。例如, García-Cervigón等(2018)研究发现南美洲两个共存的散孔材树种Embothrium coccineum和Nothofagus antarctica, 都随降水减少呈现相同的木质部导管调整方式, 即导管密度增大、导管水力平均直径降低、但总水力导度不变。有趣的是, 相同气候梯度下这两个物种的导管群聚度指数(VGI)的种内变异趋势却完全不同, E. coccineum的导管群聚度指数在温暖、湿润区域更高, 而N. antarctica则在寒冷、干燥的区域更高(García-Cervigón et al., 2018)。这一相反趋势的出现, 很可能是因为VGI只能说明复导管占导管总数的百分比, 而不能反映木质部整体导管分布格局的变化, 同时还可能存在其他木质部和叶片性状的影响所致。在未来的研究中, 采用本方法对木质部解剖结构的定量分析结合植物水分生理功能的测定将有助于提高我们对木质部结构和功能的认识。
木质部的众多导管构成的是一个三维导管系统, 木质部的输水功能与三维导管系统空间拓扑结构关系密切, 沿导水路径方向上导管长度、垂直导水路径方向上的导管空间分布格局、相邻导管间的连接方式等都是该系统的某一方面的特征。对三维导管系统的模拟和理论研究有助于加深我们对木质部导管系统结构和功能的理解, 而横切面上导管分布特征的机理模型为此提供了一个新思路。本研究通过SH模型获得的木质部中导管空间分布特征指标, 可以用于不同气候条件下种内木质部水力调节方式研究、多物种导水结构和功能比较研究、木质部三维导管系统模拟等方面, 该方法结合年轮学、解剖学、生理学、图像自动识别等最新技术和研究成果, 有利于揭示气候变化及极端干旱条件下树木的导水系统的结构和功能响应机理(von Arx et al., 2013; Beeckman, 2016; Schenk, 2018)。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 2]
[本文引用: 3]
DOI:10.1163/22941932-20160127URL [本文引用: 1]
[本文引用: 2]
[本文引用: 1]
DOI:10.1139/b2012-048URL [本文引用: 1]
DOI:10.1038/s41586-018-0240-xURL [本文引用: 1]
DOI:10.1038/nature11688URL [本文引用: 1]
DOI:10.1093/aob/mcy088PMID:29800073 [本文引用: 2]

Trees adjust the configuration of their conductive system in response to changes in water availability, maximizing efficiency in wet environments and increasing safety in dry habitats. However, evidence of this general trend is not conclusive. Generalist species growing across broad climatic gradients provide an ideal framework to assess intra-specific xylem adjustments under contrasting environmental conditions. Our aims were to compare the response of xylem traits to variations in precipitation of two co-occurring generalist tree species, and to assess climate control on xylem trait variability and co-ordination.We evaluated xylem traits of Embothrium coccineum (Proteaceae, evergreen) and Nothofagus antarctica (Nothofagaceae, deciduous) in three areas across an abrupt precipitation gradient, from 500 to 2500 mm, in southern Chile. We measured wood density, vessel lumen area and density, percentage of conductive area and vessel grouping, and estimated the hydraulic function from anatomical measurements in 60 individuals per species.Both species shared a common pattern of response along the precipitation gradient, with an increase in vessel density with dryness, but without changes in estimated hydraulic conductivity. Xylem traits in E. coccineum were more variable and more responsive to the climate gradient, decreasing vessel lumen area and increasing wood density, whereas vessel grouping showed contrasting patterns between species. Additionally, the analysis of trait co-ordination at the individual level revealed a tighter co-ordination among xylem traits in E. coccineum.Estimated xylem efficiency was maintained in combination with different levels of expected xylem safety within species. Reduction in vessel lumen area was compensated through large increases in vessel density, thus breaking the trade-off between xylem efficiency and safety. Otherwise, the existence of alternative internal adjustments in coexisting species to face similar climatic constraints might increase resilience of temperate forests against unpredictable changes in climatic conditions.
[本文引用: 2]
DOI:10.1163/22941932-90000496URL [本文引用: 1]
DOI:10.3732/ajb.1200140PMID:22965850 [本文引用: 1]
Vessels are the chief conduit for long-distance water transport in the majority of flowering plants. Vessel length is a key trait that determines plant hydraulic efficiency and safety, yet relatively little is known about this xylem feature. •We used previously published studies to generate a new global data set of vessel length in woody plants. These data were used to examine how evolutionary history, plant habit, environment, and growth ring porosity influenced vessel length. We also examined the relationship between mean vessel length and mean vessel diameter and maximum vessel length. •Data on mean vessel length were available for stems of 130 species and on maximum vessel length for stems of 91 species. A phylogenetic analysis indicated that vessel length did not exhibit significant phylogenetic signal. Liana species had longer vessel lengths than in tree or shrub species. Vessel diameter was not predictive of mean vessel length, but maximum vessel length strongly predicted mean vessel length. Vessel length did not vary between species that differed in growth ring porosity. •Many traits often assumed to be linked to vessel length, including growth ring porosity and vessel diameter, are not associated with vessel length when compared interspecifically. Sampling for vessel length has been nonrandom, e.g., there are virtually no data available for roots, and sampling for environment has been confounded with sampling for habit. Increased knowledge of vessel length is key to understanding the structure and function of the plant hydraulic pathway.
DOI:10.1002/ajb2.1029PMID:29570215 [本文引用: 1]
Xylem vessels transition through different stages during their functional lifespan, including expansion and development of vessel elements, transition to vessel hydraulic functionality, and eventual transition to post-functionality. We used information on vessel development and function to develop a model of vessel lifespan for woody plants.We examined vessel functional lifespan using repeated anatomical sampling throughout the growing season, combined with active-xylem staining to evaluate vessel hydraulic transport functionality. These data were combined with a literature review. The transitions between vessel functional lifespans for several species are illustrated, including grapevine (Vitis vinifera L., Vitaceae), English oak (Quercus robur L., Fagaceae), American chestnut [Castanea dentata (Marshall) Borkh.; Fagaceae], and several arid and semi-arid shrub species.In intact woody plants, development and maturation of vessel elements may be gradual. Once hydraulically functional, vessel elements connect to form a vessel network that is responsible for bulk hydraulic flow through the xylem. Vessels become nonfunctional due to the formation of gas emboli. In some species and under some conditions, vessel functionality of embolized conduits may be restored through refilling. Blockages, such as tyloses, gels, or gums, indicate permanent losses in hydraulic functional capacity; however, there may be some interesting exceptions to permanent loss of functionality for gel-based blockages.The gradual development and maturation of vessel elements in woody plants, variation in the onset of functionality between different populations of vessels throughout the growing season, and differences in the timing of vessel transitions to post-functionality are important aspects of plant hydraulic function.© 2018 Botanical Society of America.
[本文引用: 1]
PMID:17509617 [本文引用: 1]
The xylem is one of the two long distance transport tissues in plants, providing a low resistance pathway for water movement from roots to leaves. Its properties determine how much water can be transported and transpired and, at the same time, the plant's vulnerability to transport dysfunctions (the formation and propagation of emboli) associated to important stress factors, such as droughts and frost. Both maximum transport efficiency and safety against embolism have classically been attributed to the properties of individual conduits or of the pit membrane connecting them. But this approach overlooks the fact that the conduits of the xylem constitute a network. The topology of this network is likely to affect its overall transport properties, as well as the propagation of embolism through the xylem, since, according to the air-seeding hypothesis, drought-induced embolism propagates as a contact process (i.e., between neighbouring conduits). Here we present a model of the xylem that takes into account its system-level properties, including the connectivity of the xylem network. With the tools of graph theory and assuming steady state and Darcy's flow we calculated the hydraulic conductivity of idealized wood segments at different water potentials. A Monte Carlo approach was adopted, varying the anatomical and topological properties of the segments within biologically reasonable ranges, based on data available from the literature. Our results showed that maximum hydraulic conductivity and vulnerability to embolism increase with the connectivity of the xylem network. This can be explained by the fact that connectivity determines the fraction of all the potential paths or conduits actually available for water transport and spread of embolism. It is concluded that the xylem can no longer be interpreted as the mere sum of its conduits, because the spatial arrangement of those conduits in the xylem network influences the main functional properties of this tissue. This brings new arguments into the long-standing discussion on the efficiency vs. safety trade-off in the plants' xylem.
DOI:10.3732/ajb.1100384PMID:22733986 [本文引用: 2]
The hydraulic properties of the xylem determine the ability of plants to transport water from the soil to the leaves and to cope with important stress factors such as frost and drought. Hydraulic properties have usually been studied as a function of the anatomy of xylem conduits and their pits, but recent studies have proposed that system-level properties, related to the topology of the xylem network, may also play a role. Here we study how the spatial arrangement of conduits in xylem cross sections affects the relationship between mean conduit lumen area and conduit density (packing function) across species.Point pattern analysis was used to describe the spatial distribution of xylem conduits in 97 woody species. The effect of conduit aggregation on the packing function was tested using phylogenetic generalized least squares. A hydraulic model with an explicit description of the topology of the xylem network was used to interpret the functional significance of our findings.The spatial arrangement of conduits affected the packing function across species, so that species with aggregated distributions tended to have lower conduit densities for a given conduit size and lower conduit lumen fractions. According to our modeling results, the higher conduit-to-conduit connectivity of species with aggregated distributions allows them to achieve higher hydraulic conductivity. Species with aggregated conduits, however, pay a cost in terms of increased vulnerability to embolism.The spatial arrangement of conduits affects the fundamental structural and functional attributes of the xylem.
DOI:10.3732/ajb.0900289PMID:21616877 [本文引用: 3]
Because of their limited length, xylem conduits need to connect to each other to maintain water transport from roots to leaves. Conduit spatial distribution in a cross section plays an important role in aiding this connectivity. While indices of conduit spatial distribution already exist, they are not well defined statistically. •We used point pattern analysis to derive new spatial indices. One hundred and five cross-sectional images from different species were transformed into binary images. The resulting point patterns, based on the locations of the conduit centers-of-area, were analyzed to determine whether they departed from randomness. Conduit distribution was then modeled using a spatially explicit stochastic model. •The presence of conduit randomness, uniformity, or aggregation depended on the spatial scale of the analysis. The large majority of the images showed patterns significantly different from randomness at least at one spatial scale. A strong phylogenetic signal was detected in the spatial variables. •Conduit spatial arrangement has been largely conserved during evolution, especially at small spatial scales. Species in which conduits were aggregated in clusters had a lower conduit density compared to those with uniform distribution. Statistically sound spatial indices must be employed as an aid in the characterization of distributional patterns across species and in models of xylem water transport. Point pattern analysis is a very useful tool in identifying spatial patterns.
[本文引用: 1]
[本文引用: 2]
DOI:10.1007/s11258-006-9133-4URL [本文引用: 2]
DOI:10.1007/s40725-016-0046-8URL [本文引用: 1]
[本文引用: 1]
DOI:10.1186/s12859-017-1934-zURL [本文引用: 1]
DOI:10.1002/ajb2.2018.105.issue-2URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
PMID:25469374 [本文引用: 1]
A number of research groups in various areas of plant biology as well as computer science and applied mathematics have addressed modelling the spatiotemporal dynamics of growth and development of plants. This has resulted in development of functional-structural plant models (FSPMs). In FSPMs, the plant structure is always explicitly represented in terms of a network of elementary units. In this respect, FSPMs are different from more abstract models in which a simplified representation of the plant structure is frequently used (e.g. spatial density of leaves, total biomass, etc.). This key feature makes it possible to build modular models and creates avenues for efficient exchange of model components and experimental data. They are being used to deal with the complex 3-D structure of plants and to simulate growth and development occurring at spatial scales from cells to forest areas, and temporal scales from seconds to decades and many plant generations. The plant types studied also cover a broad spectrum, from algae to trees. This special issue of Annals of Botany features selected papers on FSPM topics such as models of morphological development, models of physical and biological processes, integrated models predicting dynamics of plants and plant communities, modelling platforms, methods for acquiring the 3-D structures of plants using automated measurements, and practical applications for agronomic purposes.
DOI:10.1163/22941932-00000035URL [本文引用: 1]
DOI:10.1163/22941932-90001528URL [本文引用: 1]
A multi-species synthesis of physiological mechanisms in drought-induced tree mortality
1
2017
... 树木木质部在植物水分运输效率以及输水安全方面发挥着重要作用, 理解其结构和功能之间的关系对于理解森林响应全球变化的机理有重要的意义(Choat et al.,
Spatstat: an R package for analyzing spatial point patterns
2
2005
... 本研究木质部解剖影像处理和导管信息提取采用ImageJ软件(Rueden et al.,
... SH模型是基于点过程的机理模型, 可以解释不同点位间相互作用而导致的点格局, 因而不同于基于统计学的传统聚集度指标(如CE), 有助于分析木质部导管空间分布格局多样性背后的生理学、生态学机理.应当说明的是, 本研究中主要采用导管位置信息进行分析, 没有区分单复导管.虽然存在复导管物种确实会增大相应的γ, 然而目前对于单、复导管的输水功能是否存在差异还存在争议(Johnson et al.,
3
2015
... 空间点格局分析通常通过建立一定空间内相邻点对间相互作用的点过程模型, 研究在一定区域内对象(如样地中的植物个体、木质部影像中的导管)的空间分布格局特征及格局的形成机理.虽然点格局分析方法在种群生态学研究中已有较多应用, 但在导管分布格局研究方面则很少见.点过程模型种类很多, 本文首次采取斯特劳斯-硬核过程模型(Strauss-Hardcore model, 以下简称SH模型)拟合分析被子植物木质部导管空间格局.SH模型属于吉布斯混合模型(Hybrid Gibbs model), 由一个硬核模型(Hardcore model, 以下简称H模型)和一个斯特劳斯模型(Strauss model, 以下简称S模型)组合构成.H模型和S模型是两种描述相邻点对之间排斥-吸引作用过程的模型, 两者组合则可以描述两个不同尺度上的点对交互过程, 其中H模型描述较小尺度上的点对间排斥作用, S模型描述稍大尺度上的排斥或吸引作用(Baddeley et al.,
... SH模型包括两个距离参数, h (硬核距离)和r (局部聚集距离), 可通过设置调参范围根据拟合模型的赤池信息量准则(AIC)值得到最优参数组合的SH模型(AIC值最低模型为最优).根据拟合成功的最佳SH模型, 可得到描述点格局强度的一个参数, 即点对局部聚集指数γ或点对间交互作用强度s.当γ = 0时, SH模型简化为一般硬核模型(h = r); γ = 1时, SH模型简化为一般硬核模型(h = h); 当0 < γ < 1时, 为硬核模型(h = h), 但带有一定程度的排斥格局(h-r间); 当γ > 1时, 为硬核模型但在0-h间为排斥格局, h-r间为吸引格局(Baddeley et al.,
... 对导管点位数据拟合SH模型, 首先根据点对最小邻体距离和最大邻体距离设置参数网格范围, 采用逼近最大似然法(Huang-Ogata approximate maximum likelihood)拟合HS模型系列, 根据拟合模型的AIC值最小的原则, 调整参数(h, c)组合至最佳(Baddeley et al.,
Wood anatomy and trait-based ecology
1
2016
... 木质部的众多导管构成的是一个三维导管系统, 木质部的输水功能与三维导管系统空间拓扑结构关系密切, 沿导水路径方向上导管长度、垂直导水路径方向上的导管空间分布格局、相邻导管间的连接方式等都是该系统的某一方面的特征.对三维导管系统的模拟和理论研究有助于加深我们对木质部导管系统结构和功能的理解, 而横切面上导管分布特征的机理模型为此提供了一个新思路.本研究通过SH模型获得的木质部中导管空间分布特征指标, 可以用于不同气候条件下种内木质部水力调节方式研究、多物种导水结构和功能比较研究、木质部三维导管系统模拟等方面, 该方法结合年轮学、解剖学、生理学、图像自动识别等最新技术和研究成果, 有利于揭示气候变化及极端干旱条件下树木的导水系统的结构和功能响应机理(von Arx et al.,
2
2001
... 被子植物的木质部具有极高的导管构型多样性, 包括早晚材导管差异、导管大小和数量、导管的空间分布格局、复导管(木质部横切面上具有共同细胞壁的相邻导管, 多为径向排列)和管孔团(多个导管共用相邻细胞壁, 或者局部单导管明显聚集)出现频率等.传统木材解剖研究对木质部导管空间排列方式的研究主要是通过观察木材横切面上的导管分布特征并做定性描述, 例如, 根据导管空间排列模式分为散生、径列、斜列、弦列等(Wheeler & Baas,
... 本研究依据IAWA的3类导管构型指标组合, 通过国际木材解剖网站InsideWood (
Non-random vessel distribution in woods: patterns, modes, diversity, correlations
1
2009
... 本研究通过点格局分析得到的导管分布格局指标具有明确的生物学意义.SH模型有3个量化指标——h、r以及γ, 它们分别指示导管在0-h、h-r、>r 3个局域尺度上导管空间分布的特点, 这意味着导管在木质部中的分布绝对不是完全随机的, 导管分布位置和局部密度受其木质部的多组织结构特点和多功能性约束(Carlquist,
How wood evolves: a new synthesis
1
2012
... 树木木质部在植物水分运输效率以及输水安全方面发挥着重要作用, 理解其结构和功能之间的关系对于理解森林响应全球变化的机理有重要的意义(Choat et al.,
Triggers of tree mortality under drought
1
2018
... 由于气候变化导致森林衰退树木死亡现象大范围出现, 对于木质部结构和功能的研究成为当前热点领域(Choat et al.,
Global convergence in the vulnerability of forests to drought
1
2012
... 树木木质部在植物水分运输效率以及输水安全方面发挥着重要作用, 理解其结构和功能之间的关系对于理解森林响应全球变化的机理有重要的意义(Choat et al.,
Xylem adjusts to maintain efficiency across a steep precipitation gradient in two coexisting generalist species
2
2018
... 由于气候变化导致森林衰退树木死亡现象大范围出现, 对于木质部结构和功能的研究成为当前热点领域(Choat et al.,
... .,
An ecophysiological and developmental perspective on variation in vessel diameter
2
2017
... SH模型是基于点过程的机理模型, 可以解释不同点位间相互作用而导致的点格局, 因而不同于基于统计学的传统聚集度指标(如CE), 有助于分析木质部导管空间分布格局多样性背后的生理学、生态学机理.应当说明的是, 本研究中主要采用导管位置信息进行分析, 没有区分单复导管.虽然存在复导管物种确实会增大相应的γ, 然而目前对于单、复导管的输水功能是否存在差异还存在争议(Johnson et al.,
... 由于气候变化导致森林衰退树木死亡现象大范围出现, 对于木质部结构和功能的研究成为当前热点领域(Choat et al.,
IAWA list of microscopic features for hardwood identification
1
1989
... 国际木材解剖学家联合会(IAWA)从3方面指标对被子植物的木质部导管构型进行描述: 木材孔型(包括环孔材、半环孔材、散孔材3类)、导管排列模式(包括弦向带状分布、径向或斜向分布、锯齿形分布3类, 忽略了没有明显分布规律的类型), 导管群聚程度(包括单导管、径向复导管、导管团3类)(IAWA Committee,
A global analysis of xylem vessel length in woody plants
1
2012
... 由于气候变化导致森林衰退树木死亡现象大范围出现, 对于木质部结构和功能的研究成为当前热点领域(Choat et al.,
Functional lifespans of xylem vessels: development, hydraulic function, and post-function of vessels in several species of woody plants
1
2018
... 被子植物木质部的导管空间分布格局多样性很高, 不仅可以用于木材种类或树木种类识别, 同时也代表着物种进化过程中的适应环境方式的多样化.木质部导管是植物运输水分的主要通道, 随着全球气候的变化, 植物导管的研究逐渐成为热点(Jacobsen et al.,
1
2018
... SH模型是基于点过程的机理模型, 可以解释不同点位间相互作用而导致的点格局, 因而不同于基于统计学的传统聚集度指标(如CE), 有助于分析木质部导管空间分布格局多样性背后的生理学、生态学机理.应当说明的是, 本研究中主要采用导管位置信息进行分析, 没有区分单复导管.虽然存在复导管物种确实会增大相应的γ, 然而目前对于单、复导管的输水功能是否存在差异还存在争议(Johnson et al.,
The relevance of xylem network structure for plant hydraulic efficiency and safety
1
2007
... 被子植物的木质部具有极高的导管构型多样性, 包括早晚材导管差异、导管大小和数量、导管的空间分布格局、复导管(木质部横切面上具有共同细胞壁的相邻导管, 多为径向排列)和管孔团(多个导管共用相邻细胞壁, 或者局部单导管明显聚集)出现频率等.传统木材解剖研究对木质部导管空间排列方式的研究主要是通过观察木材横切面上的导管分布特征并做定性描述, 例如, 根据导管空间排列模式分为散生、径列、斜列、弦列等(Wheeler & Baas,
Spatial distribution and packing of xylem conduits
2
2012
... 被子植物的木质部具有极高的导管构型多样性, 包括早晚材导管差异、导管大小和数量、导管的空间分布格局、复导管(木质部横切面上具有共同细胞壁的相邻导管, 多为径向排列)和管孔团(多个导管共用相邻细胞壁, 或者局部单导管明显聚集)出现频率等.传统木材解剖研究对木质部导管空间排列方式的研究主要是通过观察木材横切面上的导管分布特征并做定性描述, 例如, 根据导管空间排列模式分为散生、径列、斜列、弦列等(Wheeler & Baas,
... .,
A quantitative and statistically robust method for the determination of xylem conduit spatial distribution
3
2010
... 对最佳(h, r)组合采用拟合好的模型公式随机拟合19次, 可获得的该模型L函数值的95%置信区间, 该检验的显著性为α = 1/(19 + 1) = 0.05.以r为横坐标、上下包迹线的数值为纵坐标作图, 当实际值的线位于上下包络线之间, 说明模型拟合成功, 否则需要调整参数重新进行拟合.通常木质部图像中导管分布的局部单位点密度在空间是不均匀的, 因此L函数包迹线计算采用异质函数形式(Mencuccini et al.,
... 被子植物木质部的导管空间分布格局多样性很高, 不仅可以用于木材种类或树木种类识别, 同时也代表着物种进化过程中的适应环境方式的多样化.木质部导管是植物运输水分的主要通道, 随着全球气候的变化, 植物导管的研究逐渐成为热点(Jacobsen et al.,
... 本研究通过点格局分析得到的导管分布格局指标具有明确的生物学意义.SH模型有3个量化指标——h、r以及γ, 它们分别指示导管在0-h、h-r、>r 3个局域尺度上导管空间分布的特点, 这意味着导管在木质部中的分布绝对不是完全随机的, 导管分布位置和局部密度受其木质部的多组织结构特点和多功能性约束(Carlquist,
A network model links wood anatomy to xylem tissue hydraulic behaviour and vulnerability to cavitation
1
2018
... 被子植物的木质部具有极高的导管构型多样性, 包括早晚材导管差异、导管大小和数量、导管的空间分布格局、复导管(木质部横切面上具有共同细胞壁的相邻导管, 多为径向排列)和管孔团(多个导管共用相邻细胞壁, 或者局部单导管明显聚集)出现频率等.传统木材解剖研究对木质部导管空间排列方式的研究主要是通过观察木材横切面上的导管分布特征并做定性描述, 例如, 根据导管空间排列模式分为散生、径列、斜列、弦列等(Wheeler & Baas,
2
1980
... 被子植物木质部的生长发育速度通常是有限的, 且因物种而异, 不同物种的年轮宽度、导管大小和密度、空间分布特征都具有发育保守性(Panshin & Zeeuw,
... 本研究依据IAWA的3类导管构型指标组合, 通过国际木材解剖网站InsideWood (
A comparison of methods for the statistical analysis of spatial point patterns in plant ecology
2
2006
... 对导管点位数据拟合SH模型, 首先根据点对最小邻体距离和最大邻体距离设置参数网格范围, 采用逼近最大似然法(Huang-Ogata approximate maximum likelihood)拟合HS模型系列, 根据拟合模型的AIC值最小的原则, 调整参数(h, c)组合至最佳(Baddeley et al.,
... 本研究同时计算了生态学中常用的空间均质性程度指数(CE)作为空间点格局分析得到的聚集程度的参考指标.CE是基于相邻点对间距离指标计算, 采用实际点格局的平均点对距离除以同样点对密度但空间完全随机的点格局的平均点对距离得到的比值.CE被视为一种简单聚集指数, 可检验偏离完全空间随机分布格局程度, 通常采用蒙特卡洛检验(n = 1 000次), 在植物种群空间分布研究中常用, 但它存在明显的弱点是只能描述一个尺度上的空间点分布特征(Perry et al.,
Hydraulic anatomy and function of trees- Basics and critical developments
1
2016
... 树木木质部在植物水分运输效率以及输水安全方面发挥着重要作用, 理解其结构和功能之间的关系对于理解森林响应全球变化的机理有重要的意义(Choat et al.,
1
2015
... 本研究木质部解剖影像处理和导管信息提取采用ImageJ软件(Rueden et al.,
ImageJ2: ImageJ for the next generation of scientific image data
1
2017
... 本研究木质部解剖影像处理和导管信息提取采用ImageJ软件(Rueden et al.,
Wood: biology of a living tissue
1
2018
... 木质部的众多导管构成的是一个三维导管系统, 木质部的输水功能与三维导管系统空间拓扑结构关系密切, 沿导水路径方向上导管长度、垂直导水路径方向上的导管空间分布格局、相邻导管间的连接方式等都是该系统的某一方面的特征.对三维导管系统的模拟和理论研究有助于加深我们对木质部导管系统结构和功能的理解, 而横切面上导管分布特征的机理模型为此提供了一个新思路.本研究通过SH模型获得的木质部中导管空间分布特征指标, 可以用于不同气候条件下种内木质部水力调节方式研究、多物种导水结构和功能比较研究、木质部三维导管系统模拟等方面, 该方法结合年轮学、解剖学、生理学、图像自动识别等最新技术和研究成果, 有利于揭示气候变化及极端干旱条件下树木的导水系统的结构和功能响应机理(von Arx et al.,
How to quantify conduits in wood
1
2013
... 被子植物的木质部具有极高的导管构型多样性, 包括早晚材导管差异、导管大小和数量、导管的空间分布格局、复导管(木质部横切面上具有共同细胞壁的相邻导管, 多为径向排列)和管孔团(多个导管共用相邻细胞壁, 或者局部单导管明显聚集)出现频率等.传统木材解剖研究对木质部导管空间排列方式的研究主要是通过观察木材横切面上的导管分布特征并做定性描述, 例如, 根据导管空间排列模式分为散生、径列、斜列、弦列等(Wheeler & Baas,
1
2008
... 本研究依据IAWA的3类导管构型指标组合, 通过国际木材解剖网站InsideWood (
Functional-structural plant models: a growing paradigm for plant studies
1
2014
... 被子植物木质部的导管空间分布格局多样性很高, 不仅可以用于木材种类或树木种类识别, 同时也代表着物种进化过程中的适应环境方式的多样化.木质部导管是植物运输水分的主要通道, 随着全球气候的变化, 植物导管的研究逐渐成为热点(Jacobsen et al.,
Quantifying plasticity in vessel grouping-added value from the image analysis tool ROXAS
1
2013
... 木质部的众多导管构成的是一个三维导管系统, 木质部的输水功能与三维导管系统空间拓扑结构关系密切, 沿导水路径方向上导管长度、垂直导水路径方向上的导管空间分布格局、相邻导管间的连接方式等都是该系统的某一方面的特征.对三维导管系统的模拟和理论研究有助于加深我们对木质部导管系统结构和功能的理解, 而横切面上导管分布特征的机理模型为此提供了一个新思路.本研究通过SH模型获得的木质部中导管空间分布特征指标, 可以用于不同气候条件下种内木质部水力调节方式研究、多物种导水结构和功能比较研究、木质部三维导管系统模拟等方面, 该方法结合年轮学、解剖学、生理学、图像自动识别等最新技术和研究成果, 有利于揭示气候变化及极端干旱条件下树木的导水系统的结构和功能响应机理(von Arx et al.,
Wood identification-A review
1
1998
... 被子植物的木质部具有极高的导管构型多样性, 包括早晚材导管差异、导管大小和数量、导管的空间分布格局、复导管(木质部横切面上具有共同细胞壁的相邻导管, 多为径向排列)和管孔团(多个导管共用相邻细胞壁, 或者局部单导管明显聚集)出现频率等.传统木材解剖研究对木质部导管空间排列方式的研究主要是通过观察木材横切面上的导管分布特征并做定性描述, 例如, 根据导管空间排列模式分为散生、径列、斜列、弦列等(Wheeler & Baas,

{kind=link}
{kind=link}