Differences in anatomical structure and hydraulic function of xylem in branches of angiosperms in field and garden habitats
Jing FANG1,2, Lin-Feng YE1,2, Sen CHEN1,2, Shi-Tong LU1,2, Tian-Tian PAN1,2, Jiang-Bo XIE1,2,3, Yan LI1,2,3, Zhong-Yuan WANG,1,*1State Key Laboratory of Subtropical Silviculture, Zhejiang A&F University, Hangzhou 311300, China 2College of Forestry and Biotechnology, Zhejiang A&F University, Hangzhou 311300, China 3State Key Laboratory of Desert and Oasis Ecology, Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences, Ürümqi 830011, China
National Natural Science Foundation of China(31770651) National Natural Science Foundation of China(41730638) National Natural Science Foundation of China(31901280)
Abstract Aims Water is essential for plant survival and growth. Water availability affects the anatomical structure and hydraulic function of xylem, and finally makes trees form specific acclimating characters. Therefore, comparing the differences of hydraulic function and anatomical structure between plants growing in field and garden habitats, can facilitate a better understanding on the acclimation of plants to water conditions. Methods We compared hydraulic safety (water potential at 50% loss of conductivity, P50), hydraulic efficiency (specific hydraulic conductivity, Ks) and xylem anatomy (vessel diameter (D), double thickness of vessel wall (T), vessel density (N), xylem density (WD) and thickness-to-span ratio of vessels ((t/b)2)), between field and garden plants of Acer buergerianum, Cyclobalanopsis glauca and Ligustrum lucidum. We also analyzed the differences of the relationship between xylem hydraulic function and anatomical structure in field and garden habitats. Important findings We found that: 1) The Ks was higher and P50 was lower in field habitat of the three angiosperms, which were related to the larger D and smaller (t/b)2. 2) The intraspecific correlation analysis between Ks and P50 showed that there were no efficiency-safety trade-offs. 3) Intraspecific correlation analysis of anatomical structure and functional traits showed that there was no significant correlation between D and P50; except for L. lucidum in garden habitat, the T and (t/b)2of the other trees was positively correlated with P50. Comparing to garden habitat, plants in field habitat with low water availability or no additional irrigation increased their diameter of vessels to improve water transport efficiency, so as to avoid the decrease of water potential and effectively reduce the potential risk of embolism. Keywords:embolism;cavitation resistance;hydraulic efficiency;xylem anatomy;efficiency-safety trade-off
PDF (3001KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 方菁, 叶琳峰, 陈森, 陆世通, 潘天天, 谢江波, 李彦, 王忠媛. 自然和人工生境被子植物枝木质部结构与功能差异. 植物生态学报, 2021, 45(6): 650-658. DOI: 10.17521/cjpe.2020.0430 FANG Jing, YE Lin-Feng, CHEN Sen, LU Shi-Tong, PAN Tian-Tian, XIE Jiang-Bo, LI Yan, WANG Zhong-Yuan. Differences in anatomical structure and hydraulic function of xylem in branches of angiosperms in field and garden habitats. Chinese Journal of Plant Ecology, 2021, 45(6): 650-658. DOI: 10.17521/cjpe.2020.0430
Table 2 表2 表2自然和人工生境3种被子植物样树的基本特性(平均值±标准误) Table 2Basic characteristics of the sampled trees for the three species in field and garden habitats (mean ± SE)
Fig. 1Hydraulic functional traits of three species in field and garden habitats (mean ± SE). A, The specific hydraulic conductivity (Ks). B, Embolism resistance (water potential at 50% loss of conductivity, P50). Different lowercase letters indicate significant differences within species (p < 0.05).
Fig. 2Examples of light microscopy images of xylem cross sections of three species in field and garden habitats. A, Acer buergerianum in the field. B, Cyclobalanopsis glauca in the field. C, Ligustrum lucidum in the field. D, A. buergerianum in the garden. E, C. glauca in the garden. F, L. lucidum in the garden.
Fig. 3Xylem anatomical structure traits of three species in field and garden habitats (mean ± SE). A, Vessel diameter (D). B, Double thickness of vessel wall (T). C, Vessel density (N). D, Xylem density (WD). E, Thickness-to-span ratio of vessels ((t/b)2). Different lowercase letters indicate significant differences within species (p < 0.05).
Fig. 4Correlation networks between functional traits (Ks and P50) and structural traits of xylem for the three species in field and garden habitat. A, Acer buergerianum in the field. B, Cyclobalanopsis glauca in the field. C, Ligustrum lucidum in the field. D, A. buergerianum in the garden. E, C. glauca in the garden. F, L. lucidum in the garden. Solid lines, positive correlations; dashed lines, negative correlations. Red lines, p < 0.05; grey lines, p > 0.05. Line thickness indicate the correlation coefficient (r) values. D, vessel diameter (μm); Ks, specific hydraulic conductivity (kg·m-1∙MPa-1·s-1); N, vessel density (103∙mm-2); P50, water potential at 50% loss of conductivity (-MPa); T, double thickness of vessel wall (μm); Ttob, thickness-to-span ratio of vessels ((t/b)2); WD, xylem density (g∙cm-3).
AllenCD, MacaladyAK, ChenchouniH, BacheletD, McDowellN, VennetierM, KitzbergerT, RiglingA, BreshearsDD, HoggEH, GonzalezP, FenshamR, ZhangZ, CastroJ, DemidovaN, et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests Forest Ecology and Management, 259, 660-684. DOI:10.1016/j.foreco.2009.09.001URL [本文引用: 1]
AnF, ZhangSX, ZhaoPJ(2002). Progress on study of vulnerability of xylem embolism in woody plants Journal of Northwest Forestry University, 17(3), 30-34. [本文引用: 1]
AndereggWRL, AndereggLDL, BerryJA, FieldCB (2014). Loss of whole-tree hydraulic conductance during severe drought and multi-year forest die-off Oecologia, 175, 11-23. DOI:10.1007/s00442-013-2875-5PMID:24394863 [本文引用: 1] Understanding the pathways through which drought stress kills woody vegetation can improve projections of the impacts of climate change on ecosystems and carbon-cycle feedbacks. Continuous in situ measurements of whole trees during drought and as trees die hold promise to illuminate physiological pathways but are relatively rare. We monitored leaf characteristics, water use efficiency, water potentials, branch hydraulic conductivity, soil moisture, meteorological variables, and sap flux on mature healthy and sudden aspen decline-affected (SAD) trembling aspen (Populus tremuloides) ramets over two growing seasons, including a severe summer drought. We calculated daily estimates of whole-ramet hydraulic conductance and modeled whole-ramet assimilation. Healthy ramets experienced rapid declines of whole-ramet conductance during the severe drought, providing an analog for what likely occurred during the previous drought that induced SAD. Even in wetter periods, SAD-affected ramets exhibited fivefold lower whole-ramet hydraulic conductance and sevenfold lower assimilation than counterpart healthy ramets, mediated by changes in leaf area, water use efficiency, and embolism. Extant differences between healthy and SAD ramets reveal that ongoing multi-year forest die-off is primarily driven by loss of whole-ramet hydraulic capability, which in turn limits assimilation capacity. Branch-level measurements largely captured whole-plant hydraulic limitations during drought and mortality, but whole-plant measurements revealed a potential role of other losses in the hydraulic continuum. Our results highlight the importance of a whole-tree perspective in assessing physiological pathways to tree mortality and indicate that the effects of mortality on these forests' assimilation and productivity are larger than expected based on canopy leaf area differences.
AndereggWRL, KleinT, BartlettM, SackL, PellegriniAFA, ChoatB, JansenS (2016). Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought- induced tree mortality across the globe Proceedings of the National Academy of Sciences of the United States of America, 113, 5024-5029. DOI:10.1073/pnas.1525678113PMID:27091965 [本文引用: 1] Drought-induced tree mortality has been observed globally and is expected to increase under climate change scenarios, with large potential consequences for the terrestrial carbon sink. Predicting mortality across species is crucial for assessing the effects of climate extremes on forest community biodiversity, composition, and carbon sequestration. However, the physiological traits associated with elevated risk of mortality in diverse ecosystems remain unknown, although these traits could greatly improve understanding and prediction of tree mortality in forests. We performed a meta-analysis on species' mortality rates across 475 species from 33 studies around the globe to assess which traits determine a species' mortality risk. We found that species-specific mortality anomalies from community mortality rate in a given drought were associated with plant hydraulic traits. Across all species, mortality was best predicted by a low hydraulic safety margin-the difference between typical minimum xylem water potential and that causing xylem dysfunction-and xylem vulnerability to embolism. Angiosperms and gymnosperms experienced roughly equal mortality risks. Our results provide broad support for the hypothesis that hydraulic traits capture key mechanisms determining tree death and highlight that physiological traits can improve vegetation model prediction of tree mortality during climate extremes.
BeikircherB, MayrS (2009). Intraspecific differences in drought tolerance and acclimation in hydraulics of Ligustrum vulgare and Viburnum lantana Tree Physiology, 29, 765-775. DOI:10.1093/treephys/tpp018PMID:19364707 [本文引用: 5] An adequate general drought tolerance and the ability to acclimate to changing hydraulic conditions are important features for long-lived woody plants. In this study, we compared hydraulic safety (water potential at 50% loss of conductivity, Psi(50)), hydraulic efficiency (specific conductivity, k(s)), xylem anatomy (mean tracheid diameter, d(mean), mean hydraulic diameter, d(h), conduit wall thickness, t, conduit wall reinforcement, (t/b)(h)(2)) and stomatal conductance, g(s), of forest plants as well as irrigated and drought-treated garden plants of Ligustrum vulgare L. and Viburnum lantana L. Forest plants of L. vulgare and V. lantana were significantly less resistant to drought-induced cavitation (Psi(50) at -2.82 +/- 0.13 MPa and -2.79 +/- 0.17 MPa) than drought-treated garden plants (- 4.58 +/- 0.26 MPa and -3.57 +/- 0.15 MPa). When previously irrigated garden plants were subjected to drought, a significant decrease in d(mean) and d(h) and an increase in t and (t/b)(h)(2) were observed in L. vulgare. In contrast, in V. lantana conduit diameters increased significantly but no change in t and (t/b)(h)(2) was found. Stomatal closure occurred at similar water potentials (Psi(sc)) in forest plants and drought-treated garden plants, leading to higher safety margins (Psi(sc) - Psi(50)) of the latter (L. vulgare 1.63 MPa and V. lantana 0.43 MPa). These plants also showed higher g(s) at moderate Psi, more abrupt stomatal closure and lower cuticular conductivity. Data indicate that the development of drought-tolerant xylem as well as stomatal regulation play an important role in drought acclimation, whereby structural and physiological responses to drought are species-specific and depend on the plant's hydraulic strategy.
BrédaN, HucR, GranierA, DreyerE (2006). Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences Annals of Forest Science, 63, 625-644. DOI:10.1051/forest:2006042URL [本文引用: 1]
BrodribbTJ, CochardH (2009). Hydraulic failure defines the recovery and point of death in water-stressed conifers Plant Physiology, 149, 575-584. DOI:10.1104/pp.108.129783PMID:19011001 [本文引用: 1] This study combines existing hydraulic principles with recently developed methods for probing leaf hydraulic function to determine whether xylem physiology can explain the dynamic response of gas exchange both during drought and in the recovery phase after rewatering. Four conifer species from wet and dry forests were exposed to a range of water stresses by withholding water and then rewatering to observe the recovery process. During both phases midday transpiration and leaf water potential (Psileaf) were monitored. Stomatal responses to Psileaf were established for each species and these relationships used to evaluate whether the recovery of gas exchange after drought was limited by postembolism hydraulic repair in leaves. Furthermore, the timing of gas-exchange recovery was used to determine the maximum survivable water stress for each species and this index compared with data for both leaf and stem vulnerability to water-stress-induced dysfunction measured for each species. Recovery of gas exchange after water stress took between 1 and >100 d and during this period all species showed strong 1:1 conformity to a combined hydraulic-stomatal limitation model (r2 = 0.70 across all plants). Gas-exchange recovery time showed two distinct phases, a rapid overnight recovery in plants stressed to <50% loss of leaf hydraulic conductance (Kleaf) and a highly Psileaf-dependent phase in plants stressed to >50% loss of Kleaf. Maximum recoverable water stress (Psimin) corresponded to a 95% loss of Kleaf. Thus, we conclude that xylem hydraulics represents a direct limit to the drought tolerance of these conifer species.
BucciSJ, ScholzFG, CampanelloPI, MonttiL, Jimenez- CastilloM, RockwellFA, MannaLL, GuerraP, BernalPL, TroncosoO, EnricciJ, HolbrookMN, GoldsteinG (2012). Hydraulic differences along the water transport system of South American Nothofagus species: Do leaves protect the stem functionality? Tree Physiology, 32, 880-893. DOI:10.1093/treephys/tps054URL [本文引用: 5]
CarlquistS (1977). Ecological factors in wood evolution: a floristic approach American Journal of Botany, 64, 887-896. DOI:10.1002/ajb2.1977.64.issue-7URL [本文引用: 1]
ChenZC, JiangLN, FengJX, WanXC(2018). Progress and controversy of xylem embolism determination techniques in woody plants Scientia Silvae Sinicae, 54(5), 143-151. [本文引用: 1]
ChoatB, JansenS, BrodribbTJ, CochardH, DelzonS, BhaskarR, BucciSJ, FeildTS, GleasonSM, HackeUG, JacobsenAL, LensF, MaheraliH, Martínez-VilaltaJ, MayrS, et al. (2012). Global convergence in the vulnerability of forests to drought Nature, 491, 752-755. DOI:10.1038/nature11688URL [本文引用: 2]
ChoatB, SackL, HolbrookNM (2007). Diversity of hydraulic traits in nine Cordia species growing in tropical forests with contrasting precipitation New Phytologist, 175, 686-698. DOI:10.1111/nph.2007.175.issue-4URL [本文引用: 1]
CochardH, BarigahST, KleinhentzM, EshelA (2008). Is xylem cavitation resistance a relevant criterion for screening drought resistance among Prunus species? Journal of Plant Physiology, 165, 976-982. PMID:17997190 [本文引用: 2] Fruit trees are likely to suffer from the effects of severe drought in the future; however, sound criteria for evaluating the species' ability to survive these extreme conditions are largely missing. Here, we evaluated the feasibility of using xylem cavitation resistance as a tool for screening Prunus species for drought resistance. Ten different Prunus species were selected to cover a large range of water requirements, from hydrophilic to xerophilic types. Shoot cavitation resistance was evaluated with the new Cavitron technique. At this inter-specific level, cavitation resistance was related to species drought resistance, with xerophilic species being less vulnerable to cavitation. The Cavitron technique enabled species characterization that required a short time and small amounts of plant material. This technique could be used to evaluate the drought resistance of a limited number of fruit tree genotypes. Genotype screening on a larger scale, however, would likely require another approach. Out of a number of anatomical traits tested, a significant correlation was found between cavitation resistance and inter-vessel wall thickness across species. This anatomical trait is, therefore, suggested as a possible alternative to direct cavitation estimates and could be included as a screening criterion in breeding programs for drought resistance of Prunus genotypes.
CochardH, CasellaE, MencucciniM (2007). Xylem vulnerability to cavitation varies among poplar and willow clones and correlates with yield Tree Physiology, 27, 1761-1767. PMID:17938107 [本文引用: 1] Xylem vulnerability to cavitation is a promising criterion for identifying trees with high drought tolerance, but traditional techniques for measuring cavitation resistance are unsuitable for screening large numbers of genotypes. We tested the potential of the new Cavitron technique for high throughput screening of cavitation resistance in five poplar (Populus spp.) and four willow (Salix spp.) clones. The Cavitron technique enabled the screening of three to four clones per day with sufficient accuracy to reveal significant differences between clones. Because intraspecific screening may be better carried out through the identification of correlated and more easily measured traits, we attempted to identify accessible parameters that correlate to cavitation resistance. Variability in vulnerability to cavitation across clones was poorly correlated with anatomical traits such as vessel diameter, vessel wall strength, wood density and fiber wall thickness; however, a striking correlation was established between cavitation resistance and aboveground biomass production, indicating a possible trade-off between xylem safety and growth potential.
CornwellWK, BhaskarR, SackL, CordellS, LunchCK (2007). Adjustment of structure and function of Hawaiian Metrosideros polymorpha at high vs. low precipitation Functional Ecology, 21, 1063-1071. DOI:10.1111/fec.2007.21.issue-6URL [本文引用: 3]
FichotR, BarigahTS, ChamaillardS, Le ThiecD, LauransF, CochardH, BrignolasF (2010). Common trade-offs between xylem resistance to cavitation and other physiological traits do not hold among unrelated Populus deltoides × Populus nigra hybrids Plant, Cell & Environment, 33, 1553-1568. [本文引用: 1]
FrouxF, HucR, DucreyM, DreyerE (2002). Xylem hydraulic efficiency versus vulnerability in seedlings of four contrasting Mediterranean tree species (Cedrus atlantica, Cupressus sempervirens, Pinus halepensis and Pinus nigra) Annals of Forest Science, 59, 409-418. DOI:10.1051/forest:2002015URL [本文引用: 1]
GazolA, CamareroJJ, Vicente-SerranoSM, Sánchez-SalgueroR, GutiérrezE, de LuisM, Sangüesa-BarredaG, NovakK, RozasV, TíscarPA, LinaresJC, Martín-HernándezN, Martínez del CastilloE, RibasM, García-GonzálezI, et al. (2018). Forest resilience to drought varies across biomes Global Change Biology, 24, 2143-2158. DOI:10.1111/gcb.2018.24.issue-5URL [本文引用: 1]
HackeUG, SperryJS, PittermannJ (2000). Drought experience and cavitation resistance in six shrubs from the Great Basin, Utah Basic and Applied Ecology, 1, 31-41. DOI:10.1078/1439-1791-00006URL [本文引用: 1]
HackeUG, SperryJS, PockmanWT, DavisSD, McCullohKA (2001). Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure Oecologia, 126, 457-461. DOI:10.1007/s004420100628PMID:28547229 [本文引用: 5] Wood density (D ), an excellent predictor of mechanical properties, is typically viewed in relation to support against gravity, wind, snow, and other environmental forces. In contrast, we show the surprising extent to which variation in D and wood structure is linked to support against implosion by negative pressure in the xylem pipeline. The more drought-tolerant the plant, the more negative the xylem pressure can become without cavitation, and the greater the internal load on the xylem conduit walls. Accordingly, D was correlated with cavitation resistance. This trend was consistent with the maintenance of a safety factor from implosion by negative pressure: conduit wall span (b) and thickness (t) scaled so that (t/b) was proportional to cavitation resistance as required to avoid wall collapse. Unexpectedly, trends in D may be as much or more related to support of the xylem pipeline as to support of the plant.
HajekP, KurjakD, von WühlischG, DelzonS, SchuldtB (2016). Intraspecific variation in wood anatomical, hydraulic, and foliar traits in ten European beech provenances differing in growth yield Frontiers in Plant Science, 7, 791. DOI: 10.3389/fpls.2016.00791. [本文引用: 2]
JacobsenAL, EwersFW, PrattRB, PaddockWA, DavisSD (2005). Do xylem fibers affect vessel cavitation resistance? Plant Physiology, 139, 546-556. PMID:16100359 [本文引用: 1] Possible mechanical and hydraulic costs to increased cavitation resistance were examined among six co-occurring species of chaparral shrubs in southern California. We measured cavitation resistance (xylem pressure at 50% loss of hydraulic conductivity), seasonal low pressure potential (P(min)), xylem conductive efficiency (specific conductivity), mechanical strength of stems (modulus of elasticity and modulus of rupture), and xylem density. At the cellular level, we measured vessel and fiber wall thickness and lumen diameter, transverse fiber wall and total lumen area, and estimated vessel implosion resistance using (t/b)(h)(2), where t is the thickness of adjoining vessel walls and b is the vessel lumen diameter. Increased cavitation resistance was correlated with increased mechanical strength (r(2) = 0.74 and 0.76 for modulus of elasticity and modulus of rupture, respectively), xylem density (r(2) = 0.88), and P(min) (r(2) = 0.96). In contrast, cavitation resistance and P(min) were not correlated with decreased specific conductivity, suggesting no tradeoff between these traits. At the cellular level, increased cavitation resistance was correlated with increased (t/b)(h)(2) (r(2) = 0.95), increased transverse fiber wall area (r(2) = 0.89), and decreased fiber lumen area (r(2) = 0.76). To our knowledge, the correlation between cavitation resistance and fiber wall area has not been shown previously and suggests a mechanical role for fibers in cavitation resistance. Fiber efficacy in prevention of vessel implosion, defined as inward bending or collapse of vessels, is discussed.
MaheraliH, DeLuciaEH (2000). Xylem conductivity and vulnerability to cavitation of ponderosa pine growing in contrasting climates Tree Physiology, 20, 859-867. PMID:11303576 [本文引用: 1] We examined the effects of increased transpiration demand on xylem hydraulic conductivity and vulnerability to cavitation of mature ponderosa pine (Pinus ponderosa Laws.) by comparing trees growing in contrasting climates. Previous studies determined that trees growing in warm and dry sites (desert) had half the leaf/sapwood area ratio (A(L)/A(S)) and more than twice the transpiration rate of trees growing in cool and moist sites (montane). We predicted that high transpiration rates would be associated with increased specific hydraulic conductivity (K(S)) and increased resistance to xylem cavitation. Desert trees had 19% higher K(S) than montane trees, primarily because of larger tracheid lumen diameters. Predawn water potential and water potential differences between the soil and the shoot were similar for desert and montane trees, suggesting that differences in tracheid anatomy, and therefore K(S), were caused primarily by temperature and evaporative demand, rather than soil drought. Vulnerability to xylem cavitation did not differ between desert and montane populations. A 50% loss in hydraulic conductivity occurred at water potentials between -2.61 and -2.65 MPa, and vulnerability to xylem cavitation did not vary with stem size. Minimum xylem tensions of desert and montane trees did not drop below -2.05 MPa. Foliage turgor loss point did not differ between climate groups and corresponded to mean minimum xylem tensions in the field. In addition to low A(L)/A(S), high K(S) in desert trees may provide a way to increase tree hydraulic conductivity in response to high evaporative demand and prevent xylem tensions from reaching values that cause catastrophic cavitation. In ponderosa pine, the flexible responses of A(L)/A(S) and K(S) to climate may preclude the existence of significant intraspecific variation in the vulnerability of xylem to cavitation.
MaheraliH, PockmanWT, JacksonRB (2004). Adaptive variation in the vulnerability of woody plants to xylem cavitation Ecology, 85, 2184-2199. DOI:10.1890/02-0538URL [本文引用: 1]
Martínez-VilaltaJ, PratE, OliverasI, PiñolJ (2002). Xylem hydraulic properties of roots and stems of nine Mediterranean woody species Oecologia, 133, 19-29. DOI:10.1007/s00442-002-1009-2PMID:24599365 [本文引用: 2] We studied the hydraulic architecture and water relations of nine co-occurring woody species in a Spanish evergreen oak forest over the course of a dry season. Our main objectives were to: (1) test the existence of a trade-off between hydraulic conductivity and security in the xylem, and (2) establish the safety margins at which the species operated in relation to hydraulic failure, and compare these safety margins between species and tissues (roots vs. stems). Our results showed that the relationship between specific hydraulic conductivity (K s) and resistance to cavitation followed a power function with exponent ≈-2, consistent with the existence of a trade-off between conductivity and security in the xylem, and also consistent with a linear relationship between vessel diameter and the size of inter-vessel pores. The diameter of xylem conduits, K s and vulnerability to xylem embolism were always higher in roots than in stems of the same species. Safety margins from hydraulic failure were narrower in roots than in stems. Among species, the water potential (Ψ) at which 50% of conductivity was lost due to embolism ranged between -0.9 and <-8 MPa for roots and between -2.0 and <-8 MPa for stems. Vulnerability to xylem embolism followed a pattern of: Quercus ilex=Acer monspessulanum=Arbutus unedo=Sorbus torminalis=Cistus laurifolius>Cistus albidus=Ilex aquifolium>Phillyrea latifolia>Juniperus oxycedrus. Gas exchange and seasonal Ψ minima were in general correlated with resistance to xylem embolism. Hydraulic safety margins differed markedly among species, with some of them (J. oxycedrus, I. aquifolium, P. latifolia) showing a xylem overly resistant to cavitation. We hypothesize that this overly resistant xylem may be related to the shape of the relationship between K s and security we have found.
MasedaPH, FernándezRJ (2006). Stay wet or else: three ways in which plants can adjust hydraulically to their environment Journal of Experimental Botany, 57, 3963-3977. PMID:17079697 [本文引用: 1] The literature on whole-plant acclimation to drought is reviewed and it is proposed that leaf-level homeostasis in water status is attained during ontogeny largely thanks to whole-plant changes in physical resistance to liquid water flow caused by morphological and anatomical adjustments. It is shown that, in response to water deficits, plant resistance changes at different levels (tissue, organ, individual), levels that are correlated with the time scale of the response. It was found that such adjustments apparently tend to increase resistance to flow in the short term and to reduce it in the long term. A critical view of those findings is provided based on the principle that drought-induced changes cannot be analysed separately from the allometric changes that take place through ontogeny, as for example proposed by the widely cited hydraulic limitation hypothesis. A graphic synthetic model is presented according to which developmental responses to water deficits operate largely through reductions in whole-plant water transport capacity, combined with more or less strong reductions in leaf area (different "hydraulic allometries"), depending on the intrinsic tolerance of leaf tissues to partial desiccation. The model is used to show that, as the result of such adjustments, the water transport capacity per unit leaf area can decrease, remain constant, or increase, and it is argued that the expected leaf-level response would be different in each case, respectively involving a decreased, constant, or increased potential for transpiration. The article ends with a plea to collect the evidence needed to evaluate the occurrence of these three different response types across taxa and their association with different environments, including the reanalysis of existing data.
McDowellN, PockmanWT, AllenCD, BreshearsDD, CobbN, KolbT, PlautJ, SperryJ, WestA, WilliamsDG, YepezEA (2008). Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytologist, 178, 719-739. DOI:10.1111/j.1469-8137.2008.02436.xPMID:18422905 [本文引用: 1] Severe droughts have been associated with regional-scale forest mortality worldwide. Climate change is expected to exacerbate regional mortality events; however, prediction remains difficult because the physiological mechanisms underlying drought survival and mortality are poorly understood. We developed a hydraulically based theory considering carbon balance and insect resistance that allowed development and examination of hypotheses regarding survival and mortality. Multiple mechanisms may cause mortality during drought. A common mechanism for plants with isohydric regulation of water status results from avoidance of drought-induced hydraulic failure via stomatal closure, resulting in carbon starvation and a cascade of downstream effects such as reduced resistance to biotic agents. Mortality by hydraulic failure per se may occur for isohydric seedlings or trees near their maximum height. Although anisohydric plants are relatively drought-tolerant, they are predisposed to hydraulic failure because they operate with narrower hydraulic safety margins during drought. Elevated temperatures should exacerbate carbon starvation and hydraulic failure. Biotic agents may amplify and be amplified by drought-induced plant stress. Wet multidecadal climate oscillations may increase plant susceptibility to drought-induced mortality by stimulating shifts in hydraulic architecture, effectively predisposing plants to water stress. Climate warming and increased frequency of extreme events will probably cause increased regional mortality episodes. Isohydric and anisohydric water potential regulation may partition species between survival and mortality, and, as such, incorporating this hydraulic framework may be effective for modeling plant survival and mortality under future climate conditions.
MencucciniM (2003). The ecological significance of long- distance water transport: short-term regulation, long-term acclimation and the hydraulic costs of stature across plant life forms Plant, Cell & Environment, 26, 163-182. [本文引用: 1]
NolfM, PagitzK, MayrS (2014). Physiological acclimation to drought stress in Solidago canadensis Physiologia Plantarum, 150, 529-539. DOI:10.1111/ppl.12100URL [本文引用: 1]
PrattRB, JacobsenAL, EwersFW, DavisSD (2007a). Relationships among xylem transport, biomechanics and storage in stems and roots of nine Rhamnaceae species of the California chaparral New Phytologist, 174, 787-798. DOI:10.1111/nph.2007.174.issue-4URL [本文引用: 1]
PrattRB, JacobsenAL, GolgotiuKA, SperryJS, EwersFW, DavisSD (2007b). Life history type and water stress tolerance in nine California chaparral species (Rhamnaceae) Ecological Monographs, 77, 239-253. DOI:10.1890/06-0780URL [本文引用: 1]
SchuldtB, KnutzenF, DelzonS, JansenS, Müller-HauboldH, BurlettR, CloughY, LeuschnerC (2016). How adaptable is the hydraulic system of European beech in the face of climate change-related precipitation reduction? New Phytologist, 210, 443-458. DOI:10.1111/nph.2016.210.issue-2URL [本文引用: 8]
SchumannK, LeuschnerC, SchuldtB (2019). Xylem hydraulic safety and efficiency in relation to leaf and wood traits in three temperate Acer species differing in habitat preferences Trees, 33, 1475-1490. DOI:10.1007/s00468-019-01874-xURL [本文引用: 4]
SperryJS (2003). Evolution of water transport and xylem structure International Journal of Plant Sciences, 164, S115-S127. DOI:10.1086/368398URL [本文引用: 1]
TissierJ, LambsL, PeltierJP, MarigoG (2004). Relationships between hydraulic traits and habitat preference for six Acer species occurring in the French Alps Annals of Forest Science, 61, 81-86. DOI:10.1051/forest:2003087URL [本文引用: 7]
TyreeMT, DavisSD, CochardH (1994). Biophysical perspectives of xylem evolution: Is there a tradeoff of hydraulic efficiency for vulnerability to dysfunction? IAWA Journal, 15, 335-360. DOI:10.1163/22941932-90001369URL [本文引用: 4]
TyreeMT, SperryJS (1989). Vulnerability of xylem to cavitation and embolism Annual Review of Plant Physiology and Plant Molecular Biology, 40, 19-38. DOI:10.1146/annurev.pp.40.060189.000315URL [本文引用: 2]
TyreeMT, ZimmermannMH (2002). Xylem Structure and the Ascent of Sap Springer, Berlin. 45-56. [本文引用: 2]
WheelerJK, SperryJS, HackeUG, HoangN (2005). Inter- vessel pitting and cavitation in woody Rosaceae and other vesselled plants: a basis for a safety versus efficiency trade-off in xylem transport Plant, Cell & Environment, 28, 800-812. [本文引用: 3]
ZhangJZ, GouXH, ZhaoZQ, LiuWH, ZhangF, CaoZY, ZhouFF(2013). Improved method of obtaining micro- core paraffin sections in dendroecological research Chinese Journal of Plant Ecology, 37, 972-977. DOI:10.3724/SP.J.1258.2013.00100URL [本文引用: 1]
Global convergence in the vulnerability of forests to drought 2 2012
... 干旱是全球性自然灾害, 在过去的一个世纪, 引发了许多森林大规模死亡事件(Allen et al., 2010).干旱引起的森林衰退不仅发生在干旱地区, 也发生在湿润地区(Choat et al., 2012).植物在经历严重干旱后的1至4年里, 普遍存在严重的“遗留效应”, 即生长变缓和恢复不完全(Anderegg et al., 2016).湿润地区的植物因干旱过后的恢复能力较差, 可能难以应对更严重的干旱(Gazol et al., 2018), 且大多数被子植物的水力安全边界(发生气孔关闭与栓塞的水势差)较裸子植物更狭窄(Choat et al., 2012), 因此在湿润地区, 被子植物的生存和分布更容易受到干旱的威胁. ...
... ), 且大多数被子植物的水力安全边界(发生气孔关闭与栓塞的水势差)较裸子植物更狭窄(Choat et al., 2012), 因此在湿润地区, 被子植物的生存和分布更容易受到干旱的威胁. ...
Diversity of hydraulic traits in nine Cordia species growing in tropical forests with contrasting precipitation 1 2007
... 水在植物生存和生长过程中至关重要.木质部解剖结构受水分有效性影响, 因此木质部导管的结构在潮湿和干燥生境中明显不同(Carlquist, 1977).植物水力功能与其木质部导管结构密切相关(Tyree et al., 1994; Hacke et al., 2001; Wheeler et al., 2005), 在不同生境下, 解剖结构的变化势必会引起水力功能方面的变化.植物对不同水分条件生境的适应, 使其形成相应的适应机制(安锋等, 2002).Tissier等(2004)对位于校园和校外低山丘的槭属(Acer)植物进行研究, 发现水分条件较好的校园植物输水效率更高, 栓塞抗性更弱; 而Beikircher和Mayr (2009)的研究表明, Ligustrum vulgare在人工生境下栓塞抗性、输水效率更高, Viburnum lantana则是位于自然生境的栓塞抗性、输水效率更高.Maherali和Delucia (2000)对生长于干旱沙漠和潮湿山地环境下的西黄松(Pinus ponderosa)进行了研究, 发现它们的栓塞抗性没有变化, 但位于沙漠的个体输水效率更高.湿润生境和干燥生境下植物水力性状的差异反映其输水系统的竞争要求(Choat et al., 2007).一般认为水需要以较高的速度被输送到树木冠层, 以使光合作用达到最大, 这是通过最大限度减少通过导管系统引起的栓塞来达到的(Sperry, 2003). ...
Is xylem cavitation resistance a relevant criterion for screening drought resistance among Prunus species? 2 2008
... 干旱导致植物死亡的一个主要原因是水力失效, 土壤供水不足与高蒸发需求矛盾, 导致木质部导管栓塞, 输水效率降低(McDowell et al., 2008), 条件恶劣时植物的死亡不可避免(Tyree & Sperry, 1989).输水效率是植物运输水的能力, 常用比导率(Ks)衡量(Tissier et al., 2004; Gleason et al., 2016; Schumann et al., 2019); 栓塞抗性与植物从持续干旱时期存活和恢复的能力密切相关(Brodribb & Cochard, 2009), 可评估植物抵御干旱环境的能力(Cochard et al., 2008), 由木质部压力和因栓塞引起的导水率损失之间的关系来描述, 常用导水率损失50%时的水势(P50)作为其衡量指标(Tissier et al., 2004; Beikircher & Mayr, 2009; Schumann et al., 2019).Tyree和Zimmermann (2002)认为, 木质部的“效率” (输水效率)和“安全” (栓塞抗性)之间存在权衡, 即导管(或管胞)输水效率越高, 其木质部栓塞抗性就越弱.许多****对此进行了研究, 但关于效率-安全权衡是否普遍存在所有植物中, 尚无统一定论. ...
... 木质部栓塞抗性与导管解剖结构密切相关(Wheeler et al., 2005).本研究结果表明, 自然生境和人工生境下的3种植物导管直径与P50均没有显著的相关关系(图4).与其他研究(Cochard et al., 2007; Fichot et al., 2010; Hajek et al., 2016; Schuldt et al., 2016)相似, 他们也未发现导管直径与栓塞抗性的显著关系.因此, 正如Tyree和Sperry (1989)所推测的, 导管直径与栓塞的形成机制之间没有直接关系.除导管直径外, 木质部栓塞抗性还与导管内径跨度和导管壁厚有关, 导管内径跨度的减小、导管壁厚的增加或二者兼有, 都可以增加导管抵抗内爆的能力, 从而增加其栓塞抗性(Jacobsen et al., 2005).Beikircher和Mayr (2009)对L. vulgare和V. lantana的研究结果表明, 跨物种分析中T、(t/b)2与P50存在正相关关系.Cochard等(2008)对李属(Prunus)植物的研究得出相同结论, 并指出导管壁为37 μm宽时, P50与(t/b)2相关性最好(R2 = 0.76, p = 0.001).与上述发现相似, 本研究中除自然生境女贞外, T、(t/b)2与P50均呈现正相关关系(图4), 因此T和(t/b)2可以较好地指示种内栓塞抗性的强弱. ...
Xylem vulnerability to cavitation varies among poplar and willow clones and correlates with yield 1 2007
... 木质部栓塞抗性与导管解剖结构密切相关(Wheeler et al., 2005).本研究结果表明, 自然生境和人工生境下的3种植物导管直径与P50均没有显著的相关关系(图4).与其他研究(Cochard et al., 2007; Fichot et al., 2010; Hajek et al., 2016; Schuldt et al., 2016)相似, 他们也未发现导管直径与栓塞抗性的显著关系.因此, 正如Tyree和Sperry (1989)所推测的, 导管直径与栓塞的形成机制之间没有直接关系.除导管直径外, 木质部栓塞抗性还与导管内径跨度和导管壁厚有关, 导管内径跨度的减小、导管壁厚的增加或二者兼有, 都可以增加导管抵抗内爆的能力, 从而增加其栓塞抗性(Jacobsen et al., 2005).Beikircher和Mayr (2009)对L. vulgare和V. lantana的研究结果表明, 跨物种分析中T、(t/b)2与P50存在正相关关系.Cochard等(2008)对李属(Prunus)植物的研究得出相同结论, 并指出导管壁为37 μm宽时, P50与(t/b)2相关性最好(R2 = 0.76, p = 0.001).与上述发现相似, 本研究中除自然生境女贞外, T、(t/b)2与P50均呈现正相关关系(图4), 因此T和(t/b)2可以较好地指示种内栓塞抗性的强弱. ...
Adjustment of structure and function of Hawaiian Metrosideros polymorpha at high vs. low precipitation 3 2007
... 一般来说, 湿润生境中的树木输水效率较高(Tissier et al., 2004; Schuldt et al., 2016); 但也有研究表明位于较干旱生境的植物有更高的输水效率, 如位于较干旱生境的Metrosideros polymorpha (Cornwell et al., 2007)、Viburnum lantana (Beikircher & Mayr, 2009)、Nothofagus pumilio和N. antarctica (Bucci et al., 2012)的输水效率更高, 与上述发现相同, 本研究中, 位于自然生境(水分有效性较低)的树种输水效率显著高于位于人工生境的同一树种(图1A).输水效率与导管直径有关, 根据Hagen-Poiseuille方程, 木质部导水率与导管直径的4次方成正比(Tyree & Zimmermann, 2002).本研究中自然生境的植物导管直径更大(图3A), 输水效率更高(图1A), 进一步证实了上述观点.栓塞抗性取决于植物对生境的适应(Cornwell et al., 2007), 在不同生境(Beikircher & Mayr, 2009)、不同物种间(Tissier et al., 2004)存在差异, 在种内也存在差异(Bucci et al., 2012; Aguilar-Romero et al., 2017).本研究表明, 不同生境下3种植物栓塞抗性也有显著差异(图1B), 与一般认为的较湿润生境下树种栓塞抗性更弱不同(Tissier et al., 2004; Nolf et al., 2014; Schuldt et al., 2016), 本研究发现人工生境的树种栓塞抗性更强(图1B), 与Bucci等(2012)的研究结果相同: 与干燥生境的同种植物相比, 位于湿润生境的N. antarctica、N. pumilio和N. dombeyi有更强的栓塞抗性.栓塞抗性取决于导管壁的加厚(Tyree et al., 1994; Hacke et al., 2001).本研究中, 人工生境三角槭和女贞的T较自然生境更大(图3B), 其栓塞抗性也更强(图1B).除T外, (t/b)2也常用来评价物种的栓塞抗性.在Hacke等(2001)的研究中, (t/b)2与P50在针叶树(R2 = 0.78)和被子植物(R2 = 0.81)中都表现出很强的相关性, 因此(t/b)2大的植物可以避免木质部导管壁在负压下坍塌, 从而减缓栓塞的形成.本研究得出相同的结论, 人工生境下3种植物的(t/b)2显著大于自然生境(图3E), 栓塞抗性也更强(图1).(t/b)2与P50的相关性比单独使用b或T更好(Hacke et al., 2001), 本研究中, 3种植物人工生境的(t/b)2均显著大于自然生境, 而T仅有三角槭和女贞表现出人工生境大于自然生境, 因此与T相比, (t/b)2在本研究中能更好地指示P50在不同生境的强弱. ...
... ), 进一步证实了上述观点.栓塞抗性取决于植物对生境的适应(Cornwell et al., 2007), 在不同生境(Beikircher & Mayr, 2009)、不同物种间(Tissier et al., 2004)存在差异, 在种内也存在差异(Bucci et al., 2012; Aguilar-Romero et al., 2017).本研究表明, 不同生境下3种植物栓塞抗性也有显著差异(图1B), 与一般认为的较湿润生境下树种栓塞抗性更弱不同(Tissier et al., 2004; Nolf et al., 2014; Schuldt et al., 2016), 本研究发现人工生境的树种栓塞抗性更强(图1B), 与Bucci等(2012)的研究结果相同: 与干燥生境的同种植物相比, 位于湿润生境的N. antarctica、N. pumilio和N. dombeyi有更强的栓塞抗性.栓塞抗性取决于导管壁的加厚(Tyree et al., 1994; Hacke et al., 2001).本研究中, 人工生境三角槭和女贞的T较自然生境更大(图3B), 其栓塞抗性也更强(图1B).除T外, (t/b)2也常用来评价物种的栓塞抗性.在Hacke等(2001)的研究中, (t/b)2与P50在针叶树(R2 = 0.78)和被子植物(R2 = 0.81)中都表现出很强的相关性, 因此(t/b)2大的植物可以避免木质部导管壁在负压下坍塌, 从而减缓栓塞的形成.本研究得出相同的结论, 人工生境下3种植物的(t/b)2显著大于自然生境(图3E), 栓塞抗性也更强(图1).(t/b)2与P50的相关性比单独使用b或T更好(Hacke et al., 2001), 本研究中, 3种植物人工生境的(t/b)2均显著大于自然生境, 而T仅有三角槭和女贞表现出人工生境大于自然生境, 因此与T相比, (t/b)2在本研究中能更好地指示P50在不同生境的强弱. ...
... 树种在不同生境的水分利用策略不同, 其水力功能与解剖结构相协调.自然生境3个树种栓塞抗性更弱, 因此存在更高的潜在栓塞风险, 通过形成更大的导管直径来增加Ks可能是有利的: 它可以优化水的运输, 从而在水资源相对充足的时期固定碳, 并可以在较少负水势的情况下促进更高的蒸腾(Bréda et al., 2006; Cornwell et al., 2007).高Ks在避免栓塞方面发挥重要作用(Tyree et al., 1994), 因为更高的输水效率可以为栓塞抗性较弱的树木提供充足的水分, 当土壤水分有效性降低时可以避免叶水势急剧下降而栓塞(Bucci et al., 2012).Maherali等(2004)观察到落叶被子植物的Ks随降水减少而增加, 表明Ks增加可能是对自然生境水分限制的一种重要适应.本研究中除三角槭为落叶被子植物外, 青冈和女贞均为常绿被子植物, 因此Ks增加作为对自然生境水分限制的适应, 不只存在于落叶被子植物中. ...
Common trade-offs between xylem resistance to cavitation and other physiological traits do not hold among unrelated Populus deltoides × Populus nigra hybrids 1 2010
... 木质部栓塞抗性与导管解剖结构密切相关(Wheeler et al., 2005).本研究结果表明, 自然生境和人工生境下的3种植物导管直径与P50均没有显著的相关关系(图4).与其他研究(Cochard et al., 2007; Fichot et al., 2010; Hajek et al., 2016; Schuldt et al., 2016)相似, 他们也未发现导管直径与栓塞抗性的显著关系.因此, 正如Tyree和Sperry (1989)所推测的, 导管直径与栓塞的形成机制之间没有直接关系.除导管直径外, 木质部栓塞抗性还与导管内径跨度和导管壁厚有关, 导管内径跨度的减小、导管壁厚的增加或二者兼有, 都可以增加导管抵抗内爆的能力, 从而增加其栓塞抗性(Jacobsen et al., 2005).Beikircher和Mayr (2009)对L. vulgare和V. lantana的研究结果表明, 跨物种分析中T、(t/b)2与P50存在正相关关系.Cochard等(2008)对李属(Prunus)植物的研究得出相同结论, 并指出导管壁为37 μm宽时, P50与(t/b)2相关性最好(R2 = 0.76, p = 0.001).与上述发现相似, 本研究中除自然生境女贞外, T、(t/b)2与P50均呈现正相关关系(图4), 因此T和(t/b)2可以较好地指示种内栓塞抗性的强弱. ...
Xylem hydraulic efficiency versus vulnerability in seedlings of four contrasting Mediterranean tree species (Cedrus atlantica, Cupressus sempervirens, Pinus halepensis and Pinus nigra) 1 2002
... “安全”的木质部意味着对导管的充分保护, 主要是防止空穴化和栓塞, “效率”意味着一定导管组织投资下, 管道水力阻力较低(Wheeler et al., 2005).许多****认为在“安全”和“效率”之间存在一种结构上的权衡: 高效的木质部易水力失效(Tyree et al., 1994).而近来的研究表明, 树种中Ks和P50之间只存在微弱的权衡或没有权衡(Schuldt et al., 2016)(Bucci et al., 2012; Gleason et al., 2016; Schuldt et al., 2016).本研究中, 不论是自然生境还是人工生境, 种内都不存在效率-安全权衡关系(图4).Schuldt等(2016)的种内研究中并未发现效率-安全权衡, 认为其原因可能是P50变化范围太小, 即效率-安全权衡可能取决于水力功能性状的变化范围.Martínez-Vilalta等(2002)对9种木本植物进行研究, 结果表明跨物种Ks和P50存在权衡, 其拟合的负幂曲线有助于揭示不存在效率-安全权衡的原因: 只有当研究物种的Ks和P50处于负幂曲线的中心部分, 或对变化范围大的Ks和P50进行研究时, 这种权衡才会变得显著(Martínez-Vilalta et al., 2002).本研究中, 树种在同一生境的Ks和P50变化范围较小(图1), 可能因此掩盖了其权衡关系.大的Ks和P50变化范围便于验证效率-安全权衡(Froux et al., 2002), 因此与共生的、系统发育相关的类群研究或种内研究(Hajek et al., 2016; Schuldt et al., 2016)相比, 在不同生境物种的种间研究中(Tissier et al., 2004), 更可能出现效率-安全权衡关系.未来需要对更多物种进行研究, 以便深入理解跨生境跨物种水平的效率-安全权衡. ...
Forest resilience to drought varies across biomes 1 2018
... 干旱是全球性自然灾害, 在过去的一个世纪, 引发了许多森林大规模死亡事件(Allen et al., 2010).干旱引起的森林衰退不仅发生在干旱地区, 也发生在湿润地区(Choat et al., 2012).植物在经历严重干旱后的1至4年里, 普遍存在严重的“遗留效应”, 即生长变缓和恢复不完全(Anderegg et al., 2016).湿润地区的植物因干旱过后的恢复能力较差, 可能难以应对更严重的干旱(Gazol et al., 2018), 且大多数被子植物的水力安全边界(发生气孔关闭与栓塞的水势差)较裸子植物更狭窄(Choat et al., 2012), 因此在湿润地区, 被子植物的生存和分布更容易受到干旱的威胁. ...
Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species 2 2016
... 干旱导致植物死亡的一个主要原因是水力失效, 土壤供水不足与高蒸发需求矛盾, 导致木质部导管栓塞, 输水效率降低(McDowell et al., 2008), 条件恶劣时植物的死亡不可避免(Tyree & Sperry, 1989).输水效率是植物运输水的能力, 常用比导率(Ks)衡量(Tissier et al., 2004; Gleason et al., 2016; Schumann et al., 2019); 栓塞抗性与植物从持续干旱时期存活和恢复的能力密切相关(Brodribb & Cochard, 2009), 可评估植物抵御干旱环境的能力(Cochard et al., 2008), 由木质部压力和因栓塞引起的导水率损失之间的关系来描述, 常用导水率损失50%时的水势(P50)作为其衡量指标(Tissier et al., 2004; Beikircher & Mayr, 2009; Schumann et al., 2019).Tyree和Zimmermann (2002)认为, 木质部的“效率” (输水效率)和“安全” (栓塞抗性)之间存在权衡, 即导管(或管胞)输水效率越高, 其木质部栓塞抗性就越弱.许多****对此进行了研究, 但关于效率-安全权衡是否普遍存在所有植物中, 尚无统一定论. ...
... “安全”的木质部意味着对导管的充分保护, 主要是防止空穴化和栓塞, “效率”意味着一定导管组织投资下, 管道水力阻力较低(Wheeler et al., 2005).许多****认为在“安全”和“效率”之间存在一种结构上的权衡: 高效的木质部易水力失效(Tyree et al., 1994).而近来的研究表明, 树种中Ks和P50之间只存在微弱的权衡或没有权衡(Schuldt et al., 2016)(Bucci et al., 2012; Gleason et al., 2016; Schuldt et al., 2016).本研究中, 不论是自然生境还是人工生境, 种内都不存在效率-安全权衡关系(图4).Schuldt等(2016)的种内研究中并未发现效率-安全权衡, 认为其原因可能是P50变化范围太小, 即效率-安全权衡可能取决于水力功能性状的变化范围.Martínez-Vilalta等(2002)对9种木本植物进行研究, 结果表明跨物种Ks和P50存在权衡, 其拟合的负幂曲线有助于揭示不存在效率-安全权衡的原因: 只有当研究物种的Ks和P50处于负幂曲线的中心部分, 或对变化范围大的Ks和P50进行研究时, 这种权衡才会变得显著(Martínez-Vilalta et al., 2002).本研究中, 树种在同一生境的Ks和P50变化范围较小(图1), 可能因此掩盖了其权衡关系.大的Ks和P50变化范围便于验证效率-安全权衡(Froux et al., 2002), 因此与共生的、系统发育相关的类群研究或种内研究(Hajek et al., 2016; Schuldt et al., 2016)相比, 在不同生境物种的种间研究中(Tissier et al., 2004), 更可能出现效率-安全权衡关系.未来需要对更多物种进行研究, 以便深入理解跨生境跨物种水平的效率-安全权衡. ...
Drought experience and cavitation resistance in six shrubs from the Great Basin, Utah 1 2000
Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? 1 2008
... 干旱导致植物死亡的一个主要原因是水力失效, 土壤供水不足与高蒸发需求矛盾, 导致木质部导管栓塞, 输水效率降低(McDowell et al., 2008), 条件恶劣时植物的死亡不可避免(Tyree & Sperry, 1989).输水效率是植物运输水的能力, 常用比导率(Ks)衡量(Tissier et al., 2004; Gleason et al., 2016; Schumann et al., 2019); 栓塞抗性与植物从持续干旱时期存活和恢复的能力密切相关(Brodribb & Cochard, 2009), 可评估植物抵御干旱环境的能力(Cochard et al., 2008), 由木质部压力和因栓塞引起的导水率损失之间的关系来描述, 常用导水率损失50%时的水势(P50)作为其衡量指标(Tissier et al., 2004; Beikircher & Mayr, 2009; Schumann et al., 2019).Tyree和Zimmermann (2002)认为, 木质部的“效率” (输水效率)和“安全” (栓塞抗性)之间存在权衡, 即导管(或管胞)输水效率越高, 其木质部栓塞抗性就越弱.许多****对此进行了研究, 但关于效率-安全权衡是否普遍存在所有植物中, 尚无统一定论. ...
The ecological significance of long- distance water transport: short-term regulation, long-term acclimation and the hydraulic costs of stature across plant life forms 1 2003
... 一般来说, 湿润生境中的树木输水效率较高(Tissier et al., 2004; Schuldt et al., 2016); 但也有研究表明位于较干旱生境的植物有更高的输水效率, 如位于较干旱生境的Metrosideros polymorpha (Cornwell et al., 2007)、Viburnum lantana (Beikircher & Mayr, 2009)、Nothofagus pumilio和N. antarctica (Bucci et al., 2012)的输水效率更高, 与上述发现相同, 本研究中, 位于自然生境(水分有效性较低)的树种输水效率显著高于位于人工生境的同一树种(图1A).输水效率与导管直径有关, 根据Hagen-Poiseuille方程, 木质部导水率与导管直径的4次方成正比(Tyree & Zimmermann, 2002).本研究中自然生境的植物导管直径更大(图3A), 输水效率更高(图1A), 进一步证实了上述观点.栓塞抗性取决于植物对生境的适应(Cornwell et al., 2007), 在不同生境(Beikircher & Mayr, 2009)、不同物种间(Tissier et al., 2004)存在差异, 在种内也存在差异(Bucci et al., 2012; Aguilar-Romero et al., 2017).本研究表明, 不同生境下3种植物栓塞抗性也有显著差异(图1B), 与一般认为的较湿润生境下树种栓塞抗性更弱不同(Tissier et al., 2004; Nolf et al., 2014; Schuldt et al., 2016), 本研究发现人工生境的树种栓塞抗性更强(图1B), 与Bucci等(2012)的研究结果相同: 与干燥生境的同种植物相比, 位于湿润生境的N. antarctica、N. pumilio和N. dombeyi有更强的栓塞抗性.栓塞抗性取决于导管壁的加厚(Tyree et al., 1994; Hacke et al., 2001).本研究中, 人工生境三角槭和女贞的T较自然生境更大(图3B), 其栓塞抗性也更强(图1B).除T外, (t/b)2也常用来评价物种的栓塞抗性.在Hacke等(2001)的研究中, (t/b)2与P50在针叶树(R2 = 0.78)和被子植物(R2 = 0.81)中都表现出很强的相关性, 因此(t/b)2大的植物可以避免木质部导管壁在负压下坍塌, 从而减缓栓塞的形成.本研究得出相同的结论, 人工生境下3种植物的(t/b)2显著大于自然生境(图3E), 栓塞抗性也更强(图1).(t/b)2与P50的相关性比单独使用b或T更好(Hacke et al., 2001), 本研究中, 3种植物人工生境的(t/b)2均显著大于自然生境, 而T仅有三角槭和女贞表现出人工生境大于自然生境, 因此与T相比, (t/b)2在本研究中能更好地指示P50在不同生境的强弱. ...
... )、不同物种间(Tissier et al., 2004)存在差异, 在种内也存在差异(Bucci et al., 2012; Aguilar-Romero et al., 2017).本研究表明, 不同生境下3种植物栓塞抗性也有显著差异(图1B), 与一般认为的较湿润生境下树种栓塞抗性更弱不同(Tissier et al., 2004; Nolf et al., 2014; Schuldt et al., 2016), 本研究发现人工生境的树种栓塞抗性更强(图1B), 与Bucci等(2012)的研究结果相同: 与干燥生境的同种植物相比, 位于湿润生境的N. antarctica、N. pumilio和N. dombeyi有更强的栓塞抗性.栓塞抗性取决于导管壁的加厚(Tyree et al., 1994; Hacke et al., 2001).本研究中, 人工生境三角槭和女贞的T较自然生境更大(图3B), 其栓塞抗性也更强(图1B).除T外, (t/b)2也常用来评价物种的栓塞抗性.在Hacke等(2001)的研究中, (t/b)2与P50在针叶树(R2 = 0.78)和被子植物(R2 = 0.81)中都表现出很强的相关性, 因此(t/b)2大的植物可以避免木质部导管壁在负压下坍塌, 从而减缓栓塞的形成.本研究得出相同的结论, 人工生境下3种植物的(t/b)2显著大于自然生境(图3E), 栓塞抗性也更强(图1).(t/b)2与P50的相关性比单独使用b或T更好(Hacke et al., 2001), 本研究中, 3种植物人工生境的(t/b)2均显著大于自然生境, 而T仅有三角槭和女贞表现出人工生境大于自然生境, 因此与T相比, (t/b)2在本研究中能更好地指示P50在不同生境的强弱. ...
... ), 与一般认为的较湿润生境下树种栓塞抗性更弱不同(Tissier et al., 2004; Nolf et al., 2014; Schuldt et al., 2016), 本研究发现人工生境的树种栓塞抗性更强(图1B), 与Bucci等(2012)的研究结果相同: 与干燥生境的同种植物相比, 位于湿润生境的N. antarctica、N. pumilio和N. dombeyi有更强的栓塞抗性.栓塞抗性取决于导管壁的加厚(Tyree et al., 1994; Hacke et al., 2001).本研究中, 人工生境三角槭和女贞的T较自然生境更大(图3B), 其栓塞抗性也更强(图1B).除T外, (t/b)2也常用来评价物种的栓塞抗性.在Hacke等(2001)的研究中, (t/b)2与P50在针叶树(R2 = 0.78)和被子植物(R2 = 0.81)中都表现出很强的相关性, 因此(t/b)2大的植物可以避免木质部导管壁在负压下坍塌, 从而减缓栓塞的形成.本研究得出相同的结论, 人工生境下3种植物的(t/b)2显著大于自然生境(图3E), 栓塞抗性也更强(图1).(t/b)2与P50的相关性比单独使用b或T更好(Hacke et al., 2001), 本研究中, 3种植物人工生境的(t/b)2均显著大于自然生境, 而T仅有三角槭和女贞表现出人工生境大于自然生境, 因此与T相比, (t/b)2在本研究中能更好地指示P50在不同生境的强弱. ...
... “安全”的木质部意味着对导管的充分保护, 主要是防止空穴化和栓塞, “效率”意味着一定导管组织投资下, 管道水力阻力较低(Wheeler et al., 2005).许多****认为在“安全”和“效率”之间存在一种结构上的权衡: 高效的木质部易水力失效(Tyree et al., 1994).而近来的研究表明, 树种中Ks和P50之间只存在微弱的权衡或没有权衡(Schuldt et al., 2016)(Bucci et al., 2012; Gleason et al., 2016; Schuldt et al., 2016).本研究中, 不论是自然生境还是人工生境, 种内都不存在效率-安全权衡关系(图4).Schuldt等(2016)的种内研究中并未发现效率-安全权衡, 认为其原因可能是P50变化范围太小, 即效率-安全权衡可能取决于水力功能性状的变化范围.Martínez-Vilalta等(2002)对9种木本植物进行研究, 结果表明跨物种Ks和P50存在权衡, 其拟合的负幂曲线有助于揭示不存在效率-安全权衡的原因: 只有当研究物种的Ks和P50处于负幂曲线的中心部分, 或对变化范围大的Ks和P50进行研究时, 这种权衡才会变得显著(Martínez-Vilalta et al., 2002).本研究中, 树种在同一生境的Ks和P50变化范围较小(图1), 可能因此掩盖了其权衡关系.大的Ks和P50变化范围便于验证效率-安全权衡(Froux et al., 2002), 因此与共生的、系统发育相关的类群研究或种内研究(Hajek et al., 2016; Schuldt et al., 2016)相比, 在不同生境物种的种间研究中(Tissier et al., 2004), 更可能出现效率-安全权衡关系.未来需要对更多物种进行研究, 以便深入理解跨生境跨物种水平的效率-安全权衡. ...
Biophysical perspectives of xylem evolution: Is there a tradeoff of hydraulic efficiency for vulnerability to dysfunction? 4 1994
... 水在植物生存和生长过程中至关重要.木质部解剖结构受水分有效性影响, 因此木质部导管的结构在潮湿和干燥生境中明显不同(Carlquist, 1977).植物水力功能与其木质部导管结构密切相关(Tyree et al., 1994; Hacke et al., 2001; Wheeler et al., 2005), 在不同生境下, 解剖结构的变化势必会引起水力功能方面的变化.植物对不同水分条件生境的适应, 使其形成相应的适应机制(安锋等, 2002).Tissier等(2004)对位于校园和校外低山丘的槭属(Acer)植物进行研究, 发现水分条件较好的校园植物输水效率更高, 栓塞抗性更弱; 而Beikircher和Mayr (2009)的研究表明, Ligustrum vulgare在人工生境下栓塞抗性、输水效率更高, Viburnum lantana则是位于自然生境的栓塞抗性、输水效率更高.Maherali和Delucia (2000)对生长于干旱沙漠和潮湿山地环境下的西黄松(Pinus ponderosa)进行了研究, 发现它们的栓塞抗性没有变化, 但位于沙漠的个体输水效率更高.湿润生境和干燥生境下植物水力性状的差异反映其输水系统的竞争要求(Choat et al., 2007).一般认为水需要以较高的速度被输送到树木冠层, 以使光合作用达到最大, 这是通过最大限度减少通过导管系统引起的栓塞来达到的(Sperry, 2003). ...
... 一般来说, 湿润生境中的树木输水效率较高(Tissier et al., 2004; Schuldt et al., 2016); 但也有研究表明位于较干旱生境的植物有更高的输水效率, 如位于较干旱生境的Metrosideros polymorpha (Cornwell et al., 2007)、Viburnum lantana (Beikircher & Mayr, 2009)、Nothofagus pumilio和N. antarctica (Bucci et al., 2012)的输水效率更高, 与上述发现相同, 本研究中, 位于自然生境(水分有效性较低)的树种输水效率显著高于位于人工生境的同一树种(图1A).输水效率与导管直径有关, 根据Hagen-Poiseuille方程, 木质部导水率与导管直径的4次方成正比(Tyree & Zimmermann, 2002).本研究中自然生境的植物导管直径更大(图3A), 输水效率更高(图1A), 进一步证实了上述观点.栓塞抗性取决于植物对生境的适应(Cornwell et al., 2007), 在不同生境(Beikircher & Mayr, 2009)、不同物种间(Tissier et al., 2004)存在差异, 在种内也存在差异(Bucci et al., 2012; Aguilar-Romero et al., 2017).本研究表明, 不同生境下3种植物栓塞抗性也有显著差异(图1B), 与一般认为的较湿润生境下树种栓塞抗性更弱不同(Tissier et al., 2004; Nolf et al., 2014; Schuldt et al., 2016), 本研究发现人工生境的树种栓塞抗性更强(图1B), 与Bucci等(2012)的研究结果相同: 与干燥生境的同种植物相比, 位于湿润生境的N. antarctica、N. pumilio和N. dombeyi有更强的栓塞抗性.栓塞抗性取决于导管壁的加厚(Tyree et al., 1994; Hacke et al., 2001).本研究中, 人工生境三角槭和女贞的T较自然生境更大(图3B), 其栓塞抗性也更强(图1B).除T外, (t/b)2也常用来评价物种的栓塞抗性.在Hacke等(2001)的研究中, (t/b)2与P50在针叶树(R2 = 0.78)和被子植物(R2 = 0.81)中都表现出很强的相关性, 因此(t/b)2大的植物可以避免木质部导管壁在负压下坍塌, 从而减缓栓塞的形成.本研究得出相同的结论, 人工生境下3种植物的(t/b)2显著大于自然生境(图3E), 栓塞抗性也更强(图1).(t/b)2与P50的相关性比单独使用b或T更好(Hacke et al., 2001), 本研究中, 3种植物人工生境的(t/b)2均显著大于自然生境, 而T仅有三角槭和女贞表现出人工生境大于自然生境, 因此与T相比, (t/b)2在本研究中能更好地指示P50在不同生境的强弱. ...
... 树种在不同生境的水分利用策略不同, 其水力功能与解剖结构相协调.自然生境3个树种栓塞抗性更弱, 因此存在更高的潜在栓塞风险, 通过形成更大的导管直径来增加Ks可能是有利的: 它可以优化水的运输, 从而在水资源相对充足的时期固定碳, 并可以在较少负水势的情况下促进更高的蒸腾(Bréda et al., 2006; Cornwell et al., 2007).高Ks在避免栓塞方面发挥重要作用(Tyree et al., 1994), 因为更高的输水效率可以为栓塞抗性较弱的树木提供充足的水分, 当土壤水分有效性降低时可以避免叶水势急剧下降而栓塞(Bucci et al., 2012).Maherali等(2004)观察到落叶被子植物的Ks随降水减少而增加, 表明Ks增加可能是对自然生境水分限制的一种重要适应.本研究中除三角槭为落叶被子植物外, 青冈和女贞均为常绿被子植物, 因此Ks增加作为对自然生境水分限制的适应, 不只存在于落叶被子植物中. ...
... “安全”的木质部意味着对导管的充分保护, 主要是防止空穴化和栓塞, “效率”意味着一定导管组织投资下, 管道水力阻力较低(Wheeler et al., 2005).许多****认为在“安全”和“效率”之间存在一种结构上的权衡: 高效的木质部易水力失效(Tyree et al., 1994).而近来的研究表明, 树种中Ks和P50之间只存在微弱的权衡或没有权衡(Schuldt et al., 2016)(Bucci et al., 2012; Gleason et al., 2016; Schuldt et al., 2016).本研究中, 不论是自然生境还是人工生境, 种内都不存在效率-安全权衡关系(图4).Schuldt等(2016)的种内研究中并未发现效率-安全权衡, 认为其原因可能是P50变化范围太小, 即效率-安全权衡可能取决于水力功能性状的变化范围.Martínez-Vilalta等(2002)对9种木本植物进行研究, 结果表明跨物种Ks和P50存在权衡, 其拟合的负幂曲线有助于揭示不存在效率-安全权衡的原因: 只有当研究物种的Ks和P50处于负幂曲线的中心部分, 或对变化范围大的Ks和P50进行研究时, 这种权衡才会变得显著(Martínez-Vilalta et al., 2002).本研究中, 树种在同一生境的Ks和P50变化范围较小(图1), 可能因此掩盖了其权衡关系.大的Ks和P50变化范围便于验证效率-安全权衡(Froux et al., 2002), 因此与共生的、系统发育相关的类群研究或种内研究(Hajek et al., 2016; Schuldt et al., 2016)相比, 在不同生境物种的种间研究中(Tissier et al., 2004), 更可能出现效率-安全权衡关系.未来需要对更多物种进行研究, 以便深入理解跨生境跨物种水平的效率-安全权衡. ...
Vulnerability of xylem to cavitation and embolism 2 1989
... 干旱导致植物死亡的一个主要原因是水力失效, 土壤供水不足与高蒸发需求矛盾, 导致木质部导管栓塞, 输水效率降低(McDowell et al., 2008), 条件恶劣时植物的死亡不可避免(Tyree & Sperry, 1989).输水效率是植物运输水的能力, 常用比导率(Ks)衡量(Tissier et al., 2004; Gleason et al., 2016; Schumann et al., 2019); 栓塞抗性与植物从持续干旱时期存活和恢复的能力密切相关(Brodribb & Cochard, 2009), 可评估植物抵御干旱环境的能力(Cochard et al., 2008), 由木质部压力和因栓塞引起的导水率损失之间的关系来描述, 常用导水率损失50%时的水势(P50)作为其衡量指标(Tissier et al., 2004; Beikircher & Mayr, 2009; Schumann et al., 2019).Tyree和Zimmermann (2002)认为, 木质部的“效率” (输水效率)和“安全” (栓塞抗性)之间存在权衡, 即导管(或管胞)输水效率越高, 其木质部栓塞抗性就越弱.许多****对此进行了研究, 但关于效率-安全权衡是否普遍存在所有植物中, 尚无统一定论. ...
... 木质部栓塞抗性与导管解剖结构密切相关(Wheeler et al., 2005).本研究结果表明, 自然生境和人工生境下的3种植物导管直径与P50均没有显著的相关关系(图4).与其他研究(Cochard et al., 2007; Fichot et al., 2010; Hajek et al., 2016; Schuldt et al., 2016)相似, 他们也未发现导管直径与栓塞抗性的显著关系.因此, 正如Tyree和Sperry (1989)所推测的, 导管直径与栓塞的形成机制之间没有直接关系.除导管直径外, 木质部栓塞抗性还与导管内径跨度和导管壁厚有关, 导管内径跨度的减小、导管壁厚的增加或二者兼有, 都可以增加导管抵抗内爆的能力, 从而增加其栓塞抗性(Jacobsen et al., 2005).Beikircher和Mayr (2009)对L. vulgare和V. lantana的研究结果表明, 跨物种分析中T、(t/b)2与P50存在正相关关系.Cochard等(2008)对李属(Prunus)植物的研究得出相同结论, 并指出导管壁为37 μm宽时, P50与(t/b)2相关性最好(R2 = 0.76, p = 0.001).与上述发现相似, 本研究中除自然生境女贞外, T、(t/b)2与P50均呈现正相关关系(图4), 因此T和(t/b)2可以较好地指示种内栓塞抗性的强弱. ...
Xylem Structure and the Ascent of Sap 2 2002
... 干旱导致植物死亡的一个主要原因是水力失效, 土壤供水不足与高蒸发需求矛盾, 导致木质部导管栓塞, 输水效率降低(McDowell et al., 2008), 条件恶劣时植物的死亡不可避免(Tyree & Sperry, 1989).输水效率是植物运输水的能力, 常用比导率(Ks)衡量(Tissier et al., 2004; Gleason et al., 2016; Schumann et al., 2019); 栓塞抗性与植物从持续干旱时期存活和恢复的能力密切相关(Brodribb & Cochard, 2009), 可评估植物抵御干旱环境的能力(Cochard et al., 2008), 由木质部压力和因栓塞引起的导水率损失之间的关系来描述, 常用导水率损失50%时的水势(P50)作为其衡量指标(Tissier et al., 2004; Beikircher & Mayr, 2009; Schumann et al., 2019).Tyree和Zimmermann (2002)认为, 木质部的“效率” (输水效率)和“安全” (栓塞抗性)之间存在权衡, 即导管(或管胞)输水效率越高, 其木质部栓塞抗性就越弱.许多****对此进行了研究, 但关于效率-安全权衡是否普遍存在所有植物中, 尚无统一定论. ...
... 一般来说, 湿润生境中的树木输水效率较高(Tissier et al., 2004; Schuldt et al., 2016); 但也有研究表明位于较干旱生境的植物有更高的输水效率, 如位于较干旱生境的Metrosideros polymorpha (Cornwell et al., 2007)、Viburnum lantana (Beikircher & Mayr, 2009)、Nothofagus pumilio和N. antarctica (Bucci et al., 2012)的输水效率更高, 与上述发现相同, 本研究中, 位于自然生境(水分有效性较低)的树种输水效率显著高于位于人工生境的同一树种(图1A).输水效率与导管直径有关, 根据Hagen-Poiseuille方程, 木质部导水率与导管直径的4次方成正比(Tyree & Zimmermann, 2002).本研究中自然生境的植物导管直径更大(图3A), 输水效率更高(图1A), 进一步证实了上述观点.栓塞抗性取决于植物对生境的适应(Cornwell et al., 2007), 在不同生境(Beikircher & Mayr, 2009)、不同物种间(Tissier et al., 2004)存在差异, 在种内也存在差异(Bucci et al., 2012; Aguilar-Romero et al., 2017).本研究表明, 不同生境下3种植物栓塞抗性也有显著差异(图1B), 与一般认为的较湿润生境下树种栓塞抗性更弱不同(Tissier et al., 2004; Nolf et al., 2014; Schuldt et al., 2016), 本研究发现人工生境的树种栓塞抗性更强(图1B), 与Bucci等(2012)的研究结果相同: 与干燥生境的同种植物相比, 位于湿润生境的N. antarctica、N. pumilio和N. dombeyi有更强的栓塞抗性.栓塞抗性取决于导管壁的加厚(Tyree et al., 1994; Hacke et al., 2001).本研究中, 人工生境三角槭和女贞的T较自然生境更大(图3B), 其栓塞抗性也更强(图1B).除T外, (t/b)2也常用来评价物种的栓塞抗性.在Hacke等(2001)的研究中, (t/b)2与P50在针叶树(R2 = 0.78)和被子植物(R2 = 0.81)中都表现出很强的相关性, 因此(t/b)2大的植物可以避免木质部导管壁在负压下坍塌, 从而减缓栓塞的形成.本研究得出相同的结论, 人工生境下3种植物的(t/b)2显著大于自然生境(图3E), 栓塞抗性也更强(图1).(t/b)2与P50的相关性比单独使用b或T更好(Hacke et al., 2001), 本研究中, 3种植物人工生境的(t/b)2均显著大于自然生境, 而T仅有三角槭和女贞表现出人工生境大于自然生境, 因此与T相比, (t/b)2在本研究中能更好地指示P50在不同生境的强弱. ...
Inter- vessel pitting and cavitation in woody Rosaceae and other vesselled plants: a basis for a safety versus efficiency trade-off in xylem transport 3 2005
... 水在植物生存和生长过程中至关重要.木质部解剖结构受水分有效性影响, 因此木质部导管的结构在潮湿和干燥生境中明显不同(Carlquist, 1977).植物水力功能与其木质部导管结构密切相关(Tyree et al., 1994; Hacke et al., 2001; Wheeler et al., 2005), 在不同生境下, 解剖结构的变化势必会引起水力功能方面的变化.植物对不同水分条件生境的适应, 使其形成相应的适应机制(安锋等, 2002).Tissier等(2004)对位于校园和校外低山丘的槭属(Acer)植物进行研究, 发现水分条件较好的校园植物输水效率更高, 栓塞抗性更弱; 而Beikircher和Mayr (2009)的研究表明, Ligustrum vulgare在人工生境下栓塞抗性、输水效率更高, Viburnum lantana则是位于自然生境的栓塞抗性、输水效率更高.Maherali和Delucia (2000)对生长于干旱沙漠和潮湿山地环境下的西黄松(Pinus ponderosa)进行了研究, 发现它们的栓塞抗性没有变化, 但位于沙漠的个体输水效率更高.湿润生境和干燥生境下植物水力性状的差异反映其输水系统的竞争要求(Choat et al., 2007).一般认为水需要以较高的速度被输送到树木冠层, 以使光合作用达到最大, 这是通过最大限度减少通过导管系统引起的栓塞来达到的(Sperry, 2003). ...
... “安全”的木质部意味着对导管的充分保护, 主要是防止空穴化和栓塞, “效率”意味着一定导管组织投资下, 管道水力阻力较低(Wheeler et al., 2005).许多****认为在“安全”和“效率”之间存在一种结构上的权衡: 高效的木质部易水力失效(Tyree et al., 1994).而近来的研究表明, 树种中Ks和P50之间只存在微弱的权衡或没有权衡(Schuldt et al., 2016)(Bucci et al., 2012; Gleason et al., 2016; Schuldt et al., 2016).本研究中, 不论是自然生境还是人工生境, 种内都不存在效率-安全权衡关系(图4).Schuldt等(2016)的种内研究中并未发现效率-安全权衡, 认为其原因可能是P50变化范围太小, 即效率-安全权衡可能取决于水力功能性状的变化范围.Martínez-Vilalta等(2002)对9种木本植物进行研究, 结果表明跨物种Ks和P50存在权衡, 其拟合的负幂曲线有助于揭示不存在效率-安全权衡的原因: 只有当研究物种的Ks和P50处于负幂曲线的中心部分, 或对变化范围大的Ks和P50进行研究时, 这种权衡才会变得显著(Martínez-Vilalta et al., 2002).本研究中, 树种在同一生境的Ks和P50变化范围较小(图1), 可能因此掩盖了其权衡关系.大的Ks和P50变化范围便于验证效率-安全权衡(Froux et al., 2002), 因此与共生的、系统发育相关的类群研究或种内研究(Hajek et al., 2016; Schuldt et al., 2016)相比, 在不同生境物种的种间研究中(Tissier et al., 2004), 更可能出现效率-安全权衡关系.未来需要对更多物种进行研究, 以便深入理解跨生境跨物种水平的效率-安全权衡. ...
... 木质部栓塞抗性与导管解剖结构密切相关(Wheeler et al., 2005).本研究结果表明, 自然生境和人工生境下的3种植物导管直径与P50均没有显著的相关关系(图4).与其他研究(Cochard et al., 2007; Fichot et al., 2010; Hajek et al., 2016; Schuldt et al., 2016)相似, 他们也未发现导管直径与栓塞抗性的显著关系.因此, 正如Tyree和Sperry (1989)所推测的, 导管直径与栓塞的形成机制之间没有直接关系.除导管直径外, 木质部栓塞抗性还与导管内径跨度和导管壁厚有关, 导管内径跨度的减小、导管壁厚的增加或二者兼有, 都可以增加导管抵抗内爆的能力, 从而增加其栓塞抗性(Jacobsen et al., 2005).Beikircher和Mayr (2009)对L. vulgare和V. lantana的研究结果表明, 跨物种分析中T、(t/b)2与P50存在正相关关系.Cochard等(2008)对李属(Prunus)植物的研究得出相同结论, 并指出导管壁为37 μm宽时, P50与(t/b)2相关性最好(R2 = 0.76, p = 0.001).与上述发现相似, 本研究中除自然生境女贞外, T、(t/b)2与P50均呈现正相关关系(图4), 因此T和(t/b)2可以较好地指示种内栓塞抗性的强弱. ...
 ,1,*
,1,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT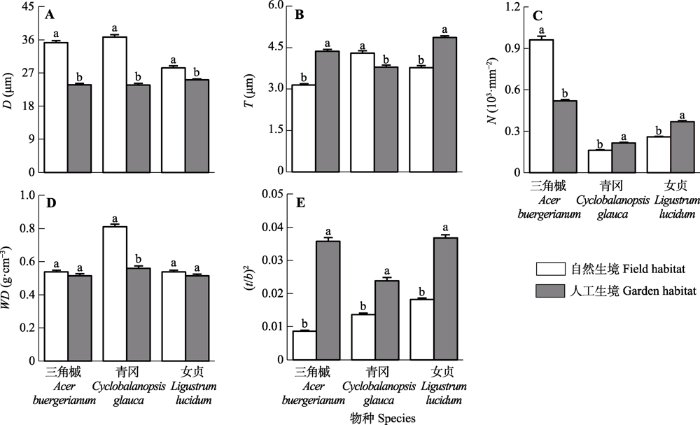 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT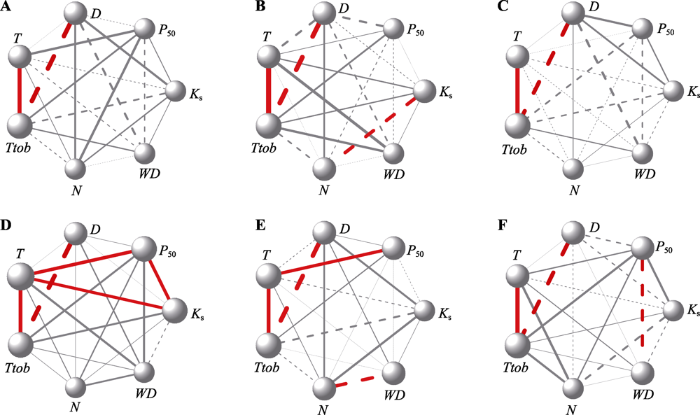 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}