Leaf and fine root economics spectrum across 49 woody plant species in Wuyi Mountains
Zhao-Ying WANG1, Xiao-Ping CHEN1, Ying CHENG2, Man-Tang WANG3, Quan-Lin ZHONG1,4, Man LI1, Dong-Liang CHENG,1,4,*1Fujian Provincial Key Laboratory of Plant Eco-physiology, Fujian Normal University, Fuzhou 350007, China 2Administrative Bureau of Jiangxi Wuyishan National Nature Reserve, Shangrao, Jiangxi 334500, China 3School of City and Architecture Engineering, Zaozhuang University, Zaozhuang, Shandong 277160, China 4Institute of Geography, Fujian Normal University, Fuzhou 350007, China
National Natural Science Foundation of China(32071555) National Natural Science Foundation of China(31722007) National Natural Science Foundation of China(31971643) National Key R&D Program of China(2017YFC0505400) Fujian Natural Science Funds for Distinguished Young Scholars(2018J07003)
Abstract Aims The plant economics spectrum would explain the trade-off strategies of vascular plants between resource acquisition and resource storage, and provide a scientific basis for understanding the mechanisms of niche differentiation and species coexistence. Methods In this study, we measured leaf/root traits of 49 woody plants species in Wuyi Mountains, including individual leaf area (ILA), specific leaf area (SLA), leaf carbon content (LCC), leaf nitrogen content (LNC) and leaf phosphorus content (LPC), root tissue density (RTD), specific root length (SRL), specific root surface area (SRA), root carbon content (RCC), root nitrogen content (RNC) and root phosphorus content (RPC). Then, we detected if the leaf and fine root economics spectrum of the plants exist, and analyzed the differences of the plant economics spectrum between evergreen and deciduous species. Important findings The results showed that along the PC1 axis, a leaf economics spectrum (LES), a root economics spectrum (RES) and a whole-plant economics spectrum (WPES) can be defined, respectively. Most of the evergreen species were on the conservative side, while deciduous species, on the acquisitive side of the economics spectrum. There were significant positive correlations among the scores of leaf PC1, root PC1 and whole-plant PC1. In the relationships, evergreen and deciduous species shared common scaling exponents, but common scaling constants lack, revealing that the leaf and root strategies of the subtropical species are coordinated toward the integration of WPES. The evergreen and deciduous species distributed at different sides of the economics spectrum are in different ways to construct the WPES. Keywords:trade-off strategy;evergreen species;deciduous species;plant functional traits;plant economics spectrum;Wuyi Mountains
PDF (1307KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 王钊颖, 陈晓萍, 程英, 王满堂, 钟全林, 李曼, 程栋梁. 武夷山49种木本植物叶片与细根经济谱. 植物生态学报, 2021, 45(3): 242-252. DOI: 10.17521/cjpe.2020.0280 WANG Zhao-Ying, CHEN Xiao-Ping, CHENG Ying, WANG Man-Tang, ZHONG Quan-Lin, LI Man, CHENG Dong-Liang. Leaf and fine root economics spectrum across 49 woody plant species in Wuyi Mountains. Chinese Journal of Plant Ecology, 2021, 45(3): 242-252. DOI: 10.17521/cjpe.2020.0280
“植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003)。将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014)。Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005)。谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017)。在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014)。叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012)。细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997)。二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象。
独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018)。有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018)。Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES。但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测。因此木本植物RES是否存在仍值得研究。除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES)。关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点。然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化。有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010)。因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题。通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980)。植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014)。因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1)。
植物经济谱能够反映具有相似策略的物种如何沿着性状变异轴汇集或分散(Díazet al., 2016; Zhao et al., 2017)。Wright等(2004)指出当物种按照叶习性来划分时, 不同物种的叶片投资策略会沿着全球叶经济谱分布, 即在LES上能够区分出常绿和落叶物种。常绿和落叶物种在叶经济性状方面的确存在一定的差异。与常绿物种相比, 落叶物种的叶面积较大(唐青青等, 2016), 单位面积的叶质量较低(即LMA低), 叶片养分的含量相对较高, 光合速率也较高(Reich et al., 1997; Aerts, 1999; Westoby et al., 2002; Wright et al., 2004)。除此之外, 在叶和根形态性状关系上, 落叶进化枝在比叶面积(SLA)和比根长(SRL)间表现出显著的负相关关系(高SLA可以在短生长季节促进更快生长, 而低SRL可以保护根免于冻结(Geng et al., 2014)); 但常绿进化枝显示出较弱的正相关关系或无显著相关性(Medeiros et al., 2017)。落叶物种由于相对频繁地落叶和生长新叶, 对土壤养分的需求量大, 有高的资源获取率, 属于营养获取型; 常绿物种叶片寿命长, 对养分需求较少, 故常绿习性被认为是对贫瘠土壤生境的一种适应性(Aerts, 1995; Givnish, 2002)。上述研究结果表明, 落叶物种倾向于采取获取策略, 而常绿种类通常采取保守策略(Givnish, 2002)。但研究叶性状关系并不能简单地直接套用其他尺度的研究结果, 全球尺度的LES是否适用于局地尺度仍值得探究(何芸雨等, 2019)。本研究力求验证武夷山的常绿和落叶物种是否也能够沿着综合的WPES被区分开来。因此, 本研究假设武夷山的常绿和落叶物种将分布于植物经济谱的两侧: 常绿物种分布在保守的一侧, 落叶物种分布在获取的一侧(假设2)。
Table 1 表1 表1武夷山不同群落的样地概况(平均值±标准误) Table 1Site status of different forest communities in the Wuyi Mountains (mean ± SE)
群落 Community
林分密度 Stand density (trees·hm-2)
平均胸径 Mean DBH (cm)
平均树高 Mean height (m)
土壤总碳含量 Soil total C content (mg·g-1)
土壤总氮含量 Soil total N content (mg·g-1)
土壤总磷含量 Soil total P content (mg·g-1)
EF
3 033.33 ± 200.00 a
13.77 ± 1.46 b
7.87 ± 0.07 b
68.88 ± 0.59 a
4.84 ± 0.04 a
0.46 ± 0.01 b
MF
1 133.33 ± 164.15 b
21.39 ± 0.80 a
10.56 ± 0.21 a
78.71 ± 4.36 a
5.25 ± 0.27 a
0.38 ± 0.02 c
DF
2 725.00 ± 163.94 a
11.47 ± 0.67 b
6.94 ± 0.24 b
75.16 ± 5.23 a
6.05 ± 0.22 a
0.65 ± 0.01 a
DF, 落叶林; EF, 常绿阔叶林; MF, 针阔混交林。同列不同小写字母表示群落间差异显著(p < 0.05)。 DF, deciduous forest; EF, evergreen forest; MF, coniferous and broad-leaved mixed forest. DBH, diameter at breast height. Different small letters in the same column indicate significant differences among communities (p < 0.05).
Fig. 1Principal components analysis (PCA) of the leaf traits, fine root traits and whole-plant traits. A, PCA of leaf traits. B, PCA of fine root traits. C, PCA of whole-plant traits. ILA, individual leaf area; LCC, leaf carbon content; LNC, leaf nitrogen content; LPC, leaf phosphor content; RCC, root carbon content; RNC, root nitrogen content; RPC, root phosphor content; RTD, root tissue density; SLA, specific leaf area; SRA, specific root surface area; SRL, specific root length.
Table 4 表4 表4叶片、细根和整株植物经济谱中各个性状与第一和第二主成分得分之间的相关系数 Table 4Correlation coefficients between individual traits and the scores of the first and second principal components in each of the leaf, fine root and whole-plant economics spectrum
Fig. 2Regression relationships among leaf PC1, fine root PC1 and whole-plant PC1. A, Regression relationships of leaf PC1 and fine root PC1. B, Regression relationships of leaf PC1 and whole-plant PC1. C, Regression relationships of fine root PC1 and whole-plant PC1.
3 讨论
植物叶片和细根性状之间的内在联系以及叶片和细根PCA轴的多性状变异共同确定了植物资源分配的权衡策略, 这证实了植物的经济策略不仅仅存在于单一器官的性状中, 而是作用于整株植物(Grime et al., 1997), 表明武夷山49种木本植物中存在着LES、RES和WPES。
3.1 植物叶片性状与细根性状的耦合
叶片和细根性状之间及其性状轴之间的显著相关性表明了LES和RES之间的耦合(Reich, 2014; Zhao et al., 2017), 因此本研究证实了假设1, 即经济谱存在于整株植物水平上。虽然有研究表明叶和根的性状之间存在解耦演化(Fortunel et al., 2012; Isaac et al., 2017), 但叶和根之间性状协调的观点依然得到众多研究的支持(Freschet et al., 2010; Liu et al., 2010; de la Riva et al., 2016; Wang et al., 2017)。有研究表明, 叶、根二者的形态性状(如SLA和SRL)存在显著相关性(Liu et al., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017)。本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL)。除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性。本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变。本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD)。对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资。因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长。有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014)。本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据。
AertsR (1999). Interspecific competition in natural plant communities: mechanisms, trade-offs and plant-soil feedbacks Journal of Experimental Botany, 50, 29-37. DOI:10.1093/jxb/50.330.29URL [本文引用: 2]
BaiKD, MoL, LiuM, ZhangDN, HeCX, WanXC, JiangDB (2015). Nutrient resorption patterns of evergreen and deciduous tree species at different altitudes on Mao’er Mountain, Guangxi Acta Ecologica Sinica, 35, 5776-5787. [本文引用: 1]
ChapinIII FS (1980). The mineral nutrition of wild plants Annual Review of Ecology and Systematics, 11, 233-260. DOI:10.1146/annurev.es.11.110180.001313URL
ChenGS, HobbieSE, ReichPB, YangYS, RobinsonD (2019). Allometry of fine roots in forest ecosystems Ecology Letters, 22, 322-331. DOI:10.1111/ele.13193URL [本文引用: 1]
CornelissenJHC, LavorelS, GarnierE, DíazS, BuchmannN, GurvichDE, ReichPB, terSteege H, MorganHD, van der HeijdenMGA, PausasJG, PoorterH (2003). A handbook of protocols for standardised and easy measurement of plant functional traits worldwide Australian Journal of Botany, 51, 335-380. DOI:10.1071/BT02124URL [本文引用: 1]
CraineJM, LeeWG (2003). Covariation in leaf and root traits for native and non-native grasses along an altitudinal gradient in New Zealand Oecologia, 134, 471-478. PMID:12647118 [本文引用: 1] Across 30 grassland sites in New Zealand that ranged from native alpine grasslands to low elevation improved pastures, there were consistent patterns of leaf and root traits and significant differences between native and non-native grasses. Plants of high altitude sites have low N concentrations in both their leaves and roots, have thick leaves and roots, yet no differences in tissue density or photosynthetic water use efficiency when compared to plants of low altitude sites. Both the leaves and roots of the low altitude plants were enriched in (15)N relative to the plants of higher altitude, indicating that the low-N set of traits is associated with a more closed N cycle at high altitude. A second independent set of correlations shows that plants of wetter habitats have lower photosynthetic water use efficiency (more negative partial differential (13)C) and lower leaf and root tissue density than the plants of drier sites. For both leaves and roots, plants of native species consistently had traits associated with lower resource availability: lower N concentrations, denser tissues, more negative partial differential (15)N, and more positive partial differential (13)C than non-native species. If root %N is correlated with root longevity as has been shown in other systems, root longevity may be able to be predicted from simple measurements of leaf %N, though a hysteresis in the relationship between leaf and root N concentrations may make prediction of high longevity roots difficult.
CraineJM, LeeWG, BondWJ, WilliamsRJ, JohnsonLC (2005). Environmental constraints on a global relationship among leaf and root traits of grasses Ecology, 86, 12-19. DOI:10.1890/04-1075URL [本文引用: 2]
dela Riva EG, TostoA, Pérez-RamosIM, Navarro-FernándezCM, OlmoM, AntenNPR, Mara?ónT, VillarR (2016). A plant economics spectrum in Mediterranean forests along environmental gradients: is there coordination among leaf, stem and root traits Journal of Vegetation Science, 27, 187-199. DOI:10.1111/jvs.12341URL
DíazS, CabidoM, CasanovesF (1998). Plant functional traits and environmental filters at a regional scale Journal of Vegetation Science, 9, 113-122. DOI:10.2307/3237229URL [本文引用: 1]
DíazS, KattgeJ, CornelissenJHC, WrightIJ, LavorelS, DrayS, ReuB, KleyerM, WirthC, ColinPrentice I, GarnierE, B?nischG, WestobyM, PoorterH, ReichPB, et al. (2016). The global spectrum of plant form and function Nature, 529, 167-171. DOI:10.1038/nature16489URL [本文引用: 3]
DingJX, KongDL, ZhangZL, CaiQ, XiaoJ, LiuQ, YinHJ (2020). Climate and soil nutrients differentially drive multidimensional fine root traits in ectomycorrhizal-dominated alpine coniferous forests Journal of Ecology, 108, 2544-2556. DOI:10.1111/jec.v108.6URL [本文引用: 1]
EissenstatDM, YanaiRD (1997). The ecology of root lifespan Advances in Ecological Research, 27, 1-60. [本文引用: 1]
FortunelC, FinePVA, BaralotoC (2012). Leaf, stem and root tissue strategies across 758 Neotropical tree species Functional Ecology, 26, 1153-1161. DOI:10.1111/j.1365-2435.2012.02020.xURL [本文引用: 2]
FreschetGT, CornelissenJHC, van LogtestijnRSP, AertsR (2010). Evidence of the “plant economics spectrum” in a subarctic flora Journal of Ecology, 98, 362-373. DOI:10.1111/jec.2010.98.issue-2URL [本文引用: 6]
FreschetGT, CornwellWK, WardleDA, ElumeevaTG, LiuWD, JacksonBG, OnipchenkoVG, SoudzilovskaiaNA, TaoJP, CornelissenJHC (2013). Linking litter decomposition of above- and below-ground organs to plant-soil feedbacks worldwide Journal of Ecology, 101, 943-952. DOI:10.1111/1365-2745.12092URL [本文引用: 1]
FunkJL, CornwellWK (2013). Leaf traits within communities: context may affect the mapping of traits to function Ecology, 94, 1893-1897. DOI:10.1890/12-1602.1URL [本文引用: 1]
GengY, WangL, JinDM, LiuHY, HeJS (2014). Alpine climate alters the relationships between leaf and root morphological traits but not chemical traits Oecologia, 175, 445-455. DOI:10.1007/s00442-014-2919-5PMID:24633995 [本文引用: 2] Leaves and fine roots are among the most important and dynamic components of terrestrial ecosystems. To what extent plants synchronize their resource capture strategies above- and belowground remains uncertain. Existing results of trait relationships between leaf and root showed great inconsistency, which may be partly due to the differences in abiotic environmental conditions such as climate and soil. Moreover, there is currently little evidence on whether and how the stringent environments of high-altitude alpine ecosystems alter the coordination between above- and belowground. Here we measured six sets of analogous traits for both leaves and fine roots of 139 species collected from Tibetan alpine grassland and Mongolian temperate grassland. N, P and N:P ratio of leaves and fine roots were positively correlated, independent of biogeographic regions, phylogenetic affiliation or climate. In contrast, leaves and fine roots seem to regulate morphological traits more independently. The specific leaf area (SLA)-specific root length (SRL) correlation shifted from negative at sites under low temperature to positive at warmer sites. The cold climate of alpine regions may impose different constraints on shoots and roots, selecting simultaneously for high SLA leaves for rapid C assimilation during the short growing season, but low SRL roots with high physical robustness to withstand soil freezing. In addition, there might be more community heterogeneity in cold soils, resulting in multidirectional strategies of root in resource acquisition. Thus our results demonstrated that alpine climate alters the relationships between leaf and root morphological but not chemical traits.
GivnishTJ (2002). Adaptive significance of evergreen vs. deciduous leaves: solving the triple paradox Silva Fennica, 36, 703-743. [本文引用: 4]
HeYY, GuoSL, WangZ (2019). Research progress of trade-off relationships of plant functional traits Chinese Journal of Plant Ecology, 43, 1021-1035. DOI:10.17521/cjpe.2019.0122URL [本文引用: 1]
IsaacME, MartinAR, deMelo Virginio Filho E, RapidelB, RoupsardO, van den MeerscheK (2017). Intraspecific trait variation and coordination: root and leaf economics spectra in coffee across environmental gradients Frontiers in Plant Science, 8, 1196. DOI: 10.3389/fpls.2017.01196. DOI:10.3389/fpls.2017.01196 [本文引用: 2]
KongDL, WangJJ, WuHF, Valverde-BarrantesOJ, WangRL, ZengH, KardolP, ZhangHY, FengYL (2019). Nonlinearity of root trait relationships and the root economics spectrum Nature Communications, 10, 2203. DOI: 10.1038/s41467-019-10245-6. DOI:10.1038/s41467-019-10245-6 [本文引用: 1]
LiFL, HuH, McCormlackML, FengDF, LiuX, BaoWK (2019). Community-level economics spectrum of fine-roots driven by nutrient limitations in subalpine forests Journal of Ecology, 107, 1238-1249. DOI:10.1111/jec.2019.107.issue-3URL [本文引用: 1]
LiJH, PengGQ, YangDM (2017). Effect of stem length to stem slender ratio of current-year twigs on the leaf display efficiency in evergreen and deciduous broadleaved trees Chinese Journal of Plant Ecology, 41, 650-660. DOI:10.17521/cjpe.2016.0376URL [本文引用: 1]
LiuGF, FreschetGT, PanX, CornelissenJHC, LiY, DongM (2010). Coordinated variation in leaf and root traits across multiple spatial scales in Chinese semi-arid and arid ecosystems New Phytologist, 188, 543-553. DOI:10.1111/nph.2010.188.issue-2URL [本文引用: 5]
LiuL, GeJL, ShuHW, ZhaoCM, XuWT, ShenGZ, XieZQ (2019). C, N and P stoichiometric ratios in mixed evergreen and deciduous broadleaved forests in Shennongjia, China Chinese Journal of Plant Ecology, 43, 482-489. DOI:10.17521/cjpe.2019.0064URL [本文引用: 1]
LuskCH, ReichPB, MontgomeryRA, AckerlyDD, Cavender-Bares J (2008). Why are evergreen leaves so contrary about shade Trends in Ecology & Evolution, 23, 299-303. DOI:10.1016/j.tree.2008.02.006URL [本文引用: 2]
MaZQ, GuoDL, XuXL, LuMZ, BardgettRD, EissenstatDM, McCormackML, HedinLO (2018). Evolutionary history resolves global organization of root functional traits Nature, 555, 94-97. DOI:10.1038/nature25783 [本文引用: 1] Plant roots have greatly diversified in form and function since the emergence of the first land plants(1,2), but the global organization of functional traits in roots remains poorly understood(3,4). Here we analyse a global dataset of 10 functionally important root traits in metabolically active first-order roots, collected from 369 species distributed across the natural plant communities of 7 biomes. Our results identify a high degree of organization of root traits across species and biomes, and reveal a pattern that differs from expectations based on previous studies(5,6) of leaf traits. Root diameter exerts the strongest influence on root trait variation across plant species, growth forms and biomes. Our analysis suggests that plants have evolved thinner roots since they first emerged in land ecosystems, which has enabled them to markedly improve their efficiency of soil exploration per unit of carbon invested and to reduce their dependence on symbiotic mycorrhizal fungi. We also found that diversity in root morphological traits is greatest in the tropics, where plant diversity is highest and many ancestral phylogenetic groups are preserved. Diversity in root morphology declines sharply across the sequence of tropical, temperate and desert biomes, presumably owing to changes in resource supply caused by seasonally inhospitable abiotic conditions. Our results suggest that root traits have evolved along a spectrum bounded by two contrasting strategies of root life: an ancestral 'conservative' strategy in which plants with thick roots depend on symbiosis with mycorrhizal fungi for soil resources and a more-derived 'opportunistic' strategy in which thin roots enable plants to more efficiently leverage photosynthetic carbon for soil exploration. These findings imply that innovations of belowground traits have had an important role in preparing plants to colonize new habitats, and in generating biodiversity within and across biomes.
MaoW, LiYL, ZhangTH, ZhaoXY, HuangYX, SongLL (2012). Research advances of plant leaf traits at different ecology scales Journal of Desert Research, 32, 33-41. [本文引用: 1]
MedeirosJS, BurnsJH, NicholsonJ, RogersL, Valverde-Barrantes O (2017). Decoupled leaf and root carbon economics is a key component in the ecological diversity and evolutionary divergence of deciduous and evergreen lineages of genus Rhododendron American Journal of Botany, 104, 803-816. DOI:10.3732/ajb.1700051PMID:28611072 [本文引用: 2] We explored trait-trait and trait-climate relationships for 27 species while accounting for phylogenetic relationships and within-species variation to investigate whether leaf and root traits are coordinated across environments and over evolutionary time, as part of a whole-plant economics spectrum.We examined specific leaf area (SLA) and four root traits: specific root length (SRL), specific root tip abundance (SRTA), first order diameter, and link average length, for plants growing in a cold, seasonal climate (Kirtland, Ohio) and a warmer, less seasonal climate (Federal Way, Washington) in the United States. We estimated a phylogeny and species' climate of origin, determined phylogenetic signal on mean traits and within-species variation, and used phylogenetically informed analysis to compare trait-trait and trait-climate relationships for deciduous and evergreen lineages.Mean SLA and within-species variation in SRL were more similar between close relatives than expected by chance. SLA and root traits differed according to climate of origin and across growth environments, though SLA differed within- and among-species less than roots. A negative SRL-SRTA correlation indicates investment in foraging scale vs. precision as a fundamental trade-off defining the root economic spectrum. Also, the deciduous clade exhibited a strong negative relationship between SLA and SRL, while evergreen clades showed a weaker positive or no relationship.Our work suggests that natural selection has shaped relationships between above- and belowground traits in genus and that leaf and root traits may evolve independently. Morphological decoupling may help explain habitat diversity among species, as well as the changes accompanying the divergence of deciduous and evergreen lineages.? 2017 Botanical Society of America.
Méndez-AlonzoR, PazH, ZuluagaRC, RosellJA, OlsonME (2012). Coordinated evolution of leaf and stem economics in tropical dry forest trees Ecology, 93, 2397-2406. PMID:23236911 [本文引用: 2] With data from 15 species in eight families of tropical dry forest trees, we provide evidence of coordination between the stem and leaf economic spectra. Species with low-density, flexible, breakable, hydraulically efficient but cavitationally vulnerable wood shed their leaves rapidly in response to drought and had low leaf mass per area and dry mass content. In contrast, species with the opposite xylem syndrome shed their costlier but more drought-resistant leaves late in the dry season. Our results explain variation in the timing of leaf shedding in tropical dry forests: selection eliminates combinations such as low-productivity leaves atop highly vulnerable xylem or water-greedy leaves supplied by xylem of low conductive efficiency. Across biomes, rather than a fundamental trade-off underlying a single axis of trait covariation, the relationship between leaf and stem economics is likely to occupy a wide space in which multiple combinations are possible.
MonkCD (1966). An ecological significance of evergreenness Ecology, 47, 504-505. DOI:10.2307/1932995URL [本文引用: 2]
Pérez-RamosIM, RoumetC, CruzP, BlanchardA, AutranP, GarnierE (2012). Evidence for a “plant community economics spectrum” driven by nutrient and water limitations in a Mediterranean rangeland of southern France Journal of Ecology, 100, 1315-1327. DOI:10.1111/jec.2012.100.issue-6URL [本文引用: 2]
PoorterH, LambersH, EvansJR (2014). Trait correlation networks: a whole-plant perspective on the recently criticized leaf economic spectrum New Phytologist, 201, 378-382. DOI:10.1111/nph.2013.201.issue-2URL [本文引用: 1]
PrietoI, RoumetC, CardinaelR, DuprazC, JourdanC, KimJH, MaeghtJL, MaoZ, PierretA, PortilloN, RoupsardO, ThammahacksaC, StokesA (2015). Root functional parameters along a land-use gradient: evidence of a community- level economics spectrum Journal of Ecology, 103, 361-373. DOI:10.1111/1365-2745.12351URL [本文引用: 1]
QuP, XingYJ, WangQG (2018). Research progress of plant economic spectrum Chinese Agricultural Science Bulletin, 34, 88-94. [本文引用: 1]
ReichPB, TjoelkerMG, WaltersMB, VanderkleinDW, BuschenaC (1998). Close association of RGR, leaf and root morphology, seed mass and shade tolerance in seedlings of nine boreal tree species grown in high and low light Functional Ecology, 12, 327-338. DOI:10.1046/j.1365-2435.1998.00208.xURL [本文引用: 1]
ReichPB, WaltersMB, EllsworthDS (1997). From tropics to tundra: global convergence in plant functioning Proceedings of the National Academy of Sciences of the United States of America, 94, 13730-13734. [本文引用: 1]
ReichPB, WrightIJ, Cavender-BaresJ, CraineJM, OleksynJ, WestobyM, WaltersMB (2003). The evolution of plant functional variation: traits, spectra, and strategies International Journal of Plant Sciences, 164, S143-S164. DOI:10.1086/374368URL [本文引用: 1]
RoumetC, BirousteM, Picon-CochardC, GhestemM, OsmanN, Vrignon-BrenasS, CaoKF, StokesA (2016). Root structure-function relationships in 74 species: evidence of a root economics spectrum related to carbon economy New Phytologist, 210, 815-826. DOI:10.1111/nph.2016.210.issue-3URL [本文引用: 1]
TangQQ, HuangYT, DingY, ZangRG (2016). Interspecific and intraspecific variation in functional traits of subtropical evergreen and deciduous broad-leaved mixed forests Biodiversity Science, 24, 262-270. DOI:10.17520/biods.2015200URL [本文引用: 1]
TjoelkerMG, CraineJM, WedinD, ReichPB, TilmanD (2005). Linking leaf and root trait syndromes among 39 grassland and savannah species New Phytologist, 167, 493-508. PMID:15998401 [本文引用: 2] Here, we tested hypothesized relationships among leaf and fine root traits of grass, forb, legume, and woody plant species of a savannah community. CO2 exchange rates, structural traits, chemistry, and longevity were measured in tissues of 39 species grown in long-term monocultures. Across species, respiration rates of leaves and fine roots exhibited a common regression relationship with tissue nitrogen (N) concentration, although legumes had lower rates at comparable N concentrations. Respiration rates and N concentration declined with increasing longevity of leaves and roots. Species rankings of leaf and fine-root N and longevity were correlated, but not specific leaf area and specific root length. The C3 and C4 grasses had lower N concentrations than forbs and legumes, but higher photosynthesis rates across a similar range of leaf N. Despite contrasting photosynthetic pathways and N2-fixing ability among these species, concordance in above- and below-ground traits was evident in comparable rankings in leaf and root longevity, N and respiration rates, which is evidence of a common leaf and root trait syndrome linking traits to effects on plant and ecosystem processes.Copyright New Phytologist (2005).
van OmmenKloeke AEE, DoumaJC, Ordo?ezJC, ReichPB, vanBodegom PM (2012). Global quantification of contrasting leaf life span strategies for deciduous and evergreen species in response to environmental conditions Global Ecology and Biogeography, 21, 224-235. DOI:10.1111/j.1466-8238.2011.00667.xURL
VenablesWN, SmithDM, The R Core Team (2019). An introduction to R. Notes on R: a programming environment for data analysis and graphics Version 3.6.0 RC (2019-04- 24). [2019-04-27]. http://ydl.oregonstate.edu/pub/cran/doc/manuals/r-patched/R-intro.pdf. URL [本文引用: 1]
WangM, WanPC, GuoJC, XuJS, ChaiYF, YueM (2017). Relationships among leaf, stem and root traits of the dominant shrubs from four vegetation zones in Shaanxi Province, China Israel Journal of Ecology and Evolution, 63, 25-32. DOI:10.1163/22244662-06301005URL [本文引用: 2]
WarrenCR, AdamsMA (2004). Evergreen trees do not maximize instantaneous photosynthesis Trends in Plant Science, 9, 270-274. DOI:10.1016/j.tplants.2004.04.004URL [本文引用: 2]
WartonDI, WeberNC (2002). Common slope tests for bivariate errors-in-variables models Biometrical Journal, 44, 161-174. DOI:10.1002/(ISSN)1521-4036URL [本文引用: 1]
WartonDI, WrightIJ, FalsterDS, WestobyM (2006). Bivariate line-fitting methods for allometry Biological Reviews, 81, 259-291. PMID:16573844 [本文引用: 1] Fitting a line to a bivariate dataset can be a deceptively complex problem, and there has been much debate on this issue in the literature. In this review, we describe for the practitioner the essential features of line-fitting methods for estimating the relationship between two variables: what methods are commonly used, which method should be used when, and how to make inferences from these lines to answer common research questions. A particularly important point for line-fitting in allometry is that usually, two sources of error are present (which we call measurement and equation error), and these have quite different implications for choice of line-fitting method. As a consequence, the approach in this review and the methods presented have subtle but important differences from previous reviews in the biology literature. Linear regression, major axis and standardised major axis are alternative methods that can be appropriate when there is no measurement error. When there is measurement error, this often needs to be estimated and used to adjust the variance terms in formulae for line-fitting. We also review line-fitting methods for phylogenetic analyses. Methods of inference are described for the line-fitting techniques discussed in this paper. The types of inference considered here are testing if the slope or elevation equals a given value, constructing confidence intervals for the slope or elevation, comparing several slopes or elevations, and testing for shift along the axis amongst several groups. In some cases several methods have been proposed in the literature. These are discussed and compared. In other cases there is little or no previous guidance available in the literature. Simulations were conducted to check whether the methods of inference proposed have the intended coverage probability or Type I error. We identified the methods of inference that perform well and recommend the techniques that should be adopted in future work.
WestobyM, FalsterDS, MolesAT, VeskPA, WrightIJ (2002). Plant ecological strategies: some leading dimensions of variation between species Annual Review of Ecology and Systematics, 33, 125-159. DOI:10.1146/annurev.ecolsys.33.010802.150452URL [本文引用: 3]
WrightIJ, ReichPB, CornelissenJHC, FalsterDS, GroomPK, HikosakaK, LeeW, LuskCH, Niinemetsü, OleksynJ, OsadaN, PoorterH, WartonDI, WestobyM (2005). Modulation of leaf economic traits and trait relationships by climate Global Ecology and Biogeography, 14, 411-421. DOI:10.1111/j.1466-822x.2005.00172.xURL [本文引用: 1]
WrightIJ, ReichPB, WestobyM, AckerlyDD, BaruchZ, BongersF, Cavender-BaresJ, ChapinT, CornelissenJHC, DiemerM, FlexasJ, GarnierE, GroomPK, GuliasJ, HikosakaK, et al. (2004). The worldwide leaf economics spectrum Nature, 428, 821-827. PMID:15103368 [本文引用: 4] Bringing together leaf trait data spanning 2,548 species and 175 sites we describe, for the first time at global scale, a universal spectrum of leaf economics consisting of key chemical, structural and physiological properties. The spectrum runs from quick to slow return on investments of nutrients and dry mass in leaves, and operates largely independently of growth form, plant functional type or biome. Categories along the spectrum would, in general, describe leaf economic variation at the global scale better than plant functional types, because functional types overlap substantially in their leaf traits. Overall, modulation of leaf traits and trait relationships by climate is surprisingly modest, although some striking and significant patterns can be seen. Reliable quantification of the leaf economics spectrum and its interaction with climate will prove valuable for modelling nutrient fluxes and vegetation boundaries under changing land-use and climate.
XiaoD, WangXJ, ZhangK, HeNP, HouJH (2016). Effects of nitrogen addition on leaf traits of common species in natural Pinus tabuliformis forests in Taiyue Mountain, Shanxi Province, China Chinese Journal of Plant Ecology, 40, 686-701. DOI:10.17521/cjpe.2015.1043URL [本文引用: 1]
YuHY, ChenYT, XuZZ, ZhouGS (2014). Analysis of relationships among leaf functional traits and economics spectrum of plant species in the desert steppe of Nei Mongol Chinese Journal of Plant Ecology, 38, 1029-1040. DOI:10.3724/SP.J.1258.2014.00097URL [本文引用: 1]
ZhaoYT, AliA, YanER (2017). The plant economics spectrum is structured by leaf habits and growth forms across subtropical species Tree Physiology, 37, 173-185. [本文引用: 5]
ZhengCY, LiuZL, FangJY (2004). Tree species diversity along altitudinal gradient on southeastern and northwestern slopes of Mt. Huanggang, Wuyi Mountains, Fujian, China Biodiversity Science, 12, 63-74. [本文引用: 1]
Allometry of fine roots in forest ecosystems 1 2019
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
植物叶经济谱的研究进展 1 2014
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
植物叶经济谱的研究进展 1 2014
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
A handbook of protocols for standardised and easy measurement of plant functional traits worldwide 1 2003
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
Covariation in leaf and root traits for native and non-native grasses along an altitudinal gradient in New Zealand 1 2003
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
Environmental constraints on a global relationship among leaf and root traits of grasses 2 2005
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
... 叶片和细根性状之间及其性状轴之间的显著相关性表明了LES和RES之间的耦合(Reich, 2014; Zhao et al., 2017), 因此本研究证实了假设1, 即经济谱存在于整株植物水平上.虽然有研究表明叶和根的性状之间存在解耦演化(Fortunel et al., 2012; Isaac et al., 2017), 但叶和根之间性状协调的观点依然得到众多研究的支持(Freschet et al., 2010; Liu et al., 2010; de la Riva et al., 2016; Wang et al., 2017).有研究表明, 叶、根二者的形态性状(如SLA和SRL)存在显著相关性(Liu et al., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017).本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL).除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性.本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变.本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD).对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资.因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长.有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014).本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据. ...
A plant economics spectrum in Mediterranean forests along environmental gradients: is there coordination among leaf, stem and root traits 2016
Plant functional traits and environmental filters at a regional scale 1 1998
Climate and soil nutrients differentially drive multidimensional fine root traits in ectomycorrhizal-dominated alpine coniferous forests 1 2020
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
The ecology of root lifespan 1 1997
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
Leaf, stem and root tissue strategies across 758 Neotropical tree species 2 2012
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
... 叶片和细根性状之间及其性状轴之间的显著相关性表明了LES和RES之间的耦合(Reich, 2014; Zhao et al., 2017), 因此本研究证实了假设1, 即经济谱存在于整株植物水平上.虽然有研究表明叶和根的性状之间存在解耦演化(Fortunel et al., 2012; Isaac et al., 2017), 但叶和根之间性状协调的观点依然得到众多研究的支持(Freschet et al., 2010; Liu et al., 2010; de la Riva et al., 2016; Wang et al., 2017).有研究表明, 叶、根二者的形态性状(如SLA和SRL)存在显著相关性(Liu et al., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017).本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL).除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性.本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变.本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD).对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资.因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长.有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014).本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据. ...
Evidence of the “plant economics spectrum” in a subarctic flora 6 2010
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
... ., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
... 叶片和细根性状之间及其性状轴之间的显著相关性表明了LES和RES之间的耦合(Reich, 2014; Zhao et al., 2017), 因此本研究证实了假设1, 即经济谱存在于整株植物水平上.虽然有研究表明叶和根的性状之间存在解耦演化(Fortunel et al., 2012; Isaac et al., 2017), 但叶和根之间性状协调的观点依然得到众多研究的支持(Freschet et al., 2010; Liu et al., 2010; de la Riva et al., 2016; Wang et al., 2017).有研究表明, 叶、根二者的形态性状(如SLA和SRL)存在显著相关性(Liu et al., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017).本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL).除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性.本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变.本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD).对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资.因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长.有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014).本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据. ...
Linking litter decomposition of above- and below-ground organs to plant-soil feedbacks worldwide 1 2013
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
Leaf traits within communities: context may affect the mapping of traits to function 1 2013
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
Alpine climate alters the relationships between leaf and root morphological traits but not chemical traits 2 2014
... 植物经济谱能够反映具有相似策略的物种如何沿着性状变异轴汇集或分散(Díazet al., 2016; Zhao et al., 2017).Wright等(2004)指出当物种按照叶习性来划分时, 不同物种的叶片投资策略会沿着全球叶经济谱分布, 即在LES上能够区分出常绿和落叶物种.常绿和落叶物种在叶经济性状方面的确存在一定的差异.与常绿物种相比, 落叶物种的叶面积较大(唐青青等, 2016), 单位面积的叶质量较低(即LMA低), 叶片养分的含量相对较高, 光合速率也较高(Reich et al., 1997; Aerts, 1999; Westoby et al., 2002; Wright et al., 2004).除此之外, 在叶和根形态性状关系上, 落叶进化枝在比叶面积(SLA)和比根长(SRL)间表现出显著的负相关关系(高SLA可以在短生长季节促进更快生长, 而低SRL可以保护根免于冻结(Geng et al., 2014)); 但常绿进化枝显示出较弱的正相关关系或无显著相关性(Medeiros et al., 2017).落叶物种由于相对频繁地落叶和生长新叶, 对土壤养分的需求量大, 有高的资源获取率, 属于营养获取型; 常绿物种叶片寿命长, 对养分需求较少, 故常绿习性被认为是对贫瘠土壤生境的一种适应性(Aerts, 1995; Givnish, 2002).上述研究结果表明, 落叶物种倾向于采取获取策略, 而常绿种类通常采取保守策略(Givnish, 2002).但研究叶性状关系并不能简单地直接套用其他尺度的研究结果, 全球尺度的LES是否适用于局地尺度仍值得探究(何芸雨等, 2019).本研究力求验证武夷山的常绿和落叶物种是否也能够沿着综合的WPES被区分开来.因此, 本研究假设武夷山的常绿和落叶物种将分布于植物经济谱的两侧: 常绿物种分布在保守的一侧, 落叶物种分布在获取的一侧(假设2). ...
... 叶片和细根性状之间及其性状轴之间的显著相关性表明了LES和RES之间的耦合(Reich, 2014; Zhao et al., 2017), 因此本研究证实了假设1, 即经济谱存在于整株植物水平上.虽然有研究表明叶和根的性状之间存在解耦演化(Fortunel et al., 2012; Isaac et al., 2017), 但叶和根之间性状协调的观点依然得到众多研究的支持(Freschet et al., 2010; Liu et al., 2010; de la Riva et al., 2016; Wang et al., 2017).有研究表明, 叶、根二者的形态性状(如SLA和SRL)存在显著相关性(Liu et al., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017).本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL).除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性.本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变.本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD).对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资.因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长.有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014).本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据. ...
Adaptive significance of evergreen vs. deciduous leaves: solving the triple paradox 4 2002
... 植物经济谱能够反映具有相似策略的物种如何沿着性状变异轴汇集或分散(Díazet al., 2016; Zhao et al., 2017).Wright等(2004)指出当物种按照叶习性来划分时, 不同物种的叶片投资策略会沿着全球叶经济谱分布, 即在LES上能够区分出常绿和落叶物种.常绿和落叶物种在叶经济性状方面的确存在一定的差异.与常绿物种相比, 落叶物种的叶面积较大(唐青青等, 2016), 单位面积的叶质量较低(即LMA低), 叶片养分的含量相对较高, 光合速率也较高(Reich et al., 1997; Aerts, 1999; Westoby et al., 2002; Wright et al., 2004).除此之外, 在叶和根形态性状关系上, 落叶进化枝在比叶面积(SLA)和比根长(SRL)间表现出显著的负相关关系(高SLA可以在短生长季节促进更快生长, 而低SRL可以保护根免于冻结(Geng et al., 2014)); 但常绿进化枝显示出较弱的正相关关系或无显著相关性(Medeiros et al., 2017).落叶物种由于相对频繁地落叶和生长新叶, 对土壤养分的需求量大, 有高的资源获取率, 属于营养获取型; 常绿物种叶片寿命长, 对养分需求较少, 故常绿习性被认为是对贫瘠土壤生境的一种适应性(Aerts, 1995; Givnish, 2002).上述研究结果表明, 落叶物种倾向于采取获取策略, 而常绿种类通常采取保守策略(Givnish, 2002).但研究叶性状关系并不能简单地直接套用其他尺度的研究结果, 全球尺度的LES是否适用于局地尺度仍值得探究(何芸雨等, 2019).本研究力求验证武夷山的常绿和落叶物种是否也能够沿着综合的WPES被区分开来.因此, 本研究假设武夷山的常绿和落叶物种将分布于植物经济谱的两侧: 常绿物种分布在保守的一侧, 落叶物种分布在获取的一侧(假设2). ...
Integrated screening validates primary axes of specialisation in plants 2 1997
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
... 植物叶片和细根性状之间的内在联系以及叶片和细根PCA轴的多性状变异共同确定了植物资源分配的权衡策略, 这证实了植物的经济策略不仅仅存在于单一器官的性状中, 而是作用于整株植物(Grime et al., 1997), 表明武夷山49种木本植物中存在着LES、RES和WPES. ...
植物功能性状权衡关系的研究进展 1 2019
... 植物经济谱能够反映具有相似策略的物种如何沿着性状变异轴汇集或分散(Díazet al., 2016; Zhao et al., 2017).Wright等(2004)指出当物种按照叶习性来划分时, 不同物种的叶片投资策略会沿着全球叶经济谱分布, 即在LES上能够区分出常绿和落叶物种.常绿和落叶物种在叶经济性状方面的确存在一定的差异.与常绿物种相比, 落叶物种的叶面积较大(唐青青等, 2016), 单位面积的叶质量较低(即LMA低), 叶片养分的含量相对较高, 光合速率也较高(Reich et al., 1997; Aerts, 1999; Westoby et al., 2002; Wright et al., 2004).除此之外, 在叶和根形态性状关系上, 落叶进化枝在比叶面积(SLA)和比根长(SRL)间表现出显著的负相关关系(高SLA可以在短生长季节促进更快生长, 而低SRL可以保护根免于冻结(Geng et al., 2014)); 但常绿进化枝显示出较弱的正相关关系或无显著相关性(Medeiros et al., 2017).落叶物种由于相对频繁地落叶和生长新叶, 对土壤养分的需求量大, 有高的资源获取率, 属于营养获取型; 常绿物种叶片寿命长, 对养分需求较少, 故常绿习性被认为是对贫瘠土壤生境的一种适应性(Aerts, 1995; Givnish, 2002).上述研究结果表明, 落叶物种倾向于采取获取策略, 而常绿种类通常采取保守策略(Givnish, 2002).但研究叶性状关系并不能简单地直接套用其他尺度的研究结果, 全球尺度的LES是否适用于局地尺度仍值得探究(何芸雨等, 2019).本研究力求验证武夷山的常绿和落叶物种是否也能够沿着综合的WPES被区分开来.因此, 本研究假设武夷山的常绿和落叶物种将分布于植物经济谱的两侧: 常绿物种分布在保守的一侧, 落叶物种分布在获取的一侧(假设2). ...
植物功能性状权衡关系的研究进展 1 2019
... 植物经济谱能够反映具有相似策略的物种如何沿着性状变异轴汇集或分散(Díazet al., 2016; Zhao et al., 2017).Wright等(2004)指出当物种按照叶习性来划分时, 不同物种的叶片投资策略会沿着全球叶经济谱分布, 即在LES上能够区分出常绿和落叶物种.常绿和落叶物种在叶经济性状方面的确存在一定的差异.与常绿物种相比, 落叶物种的叶面积较大(唐青青等, 2016), 单位面积的叶质量较低(即LMA低), 叶片养分的含量相对较高, 光合速率也较高(Reich et al., 1997; Aerts, 1999; Westoby et al., 2002; Wright et al., 2004).除此之外, 在叶和根形态性状关系上, 落叶进化枝在比叶面积(SLA)和比根长(SRL)间表现出显著的负相关关系(高SLA可以在短生长季节促进更快生长, 而低SRL可以保护根免于冻结(Geng et al., 2014)); 但常绿进化枝显示出较弱的正相关关系或无显著相关性(Medeiros et al., 2017).落叶物种由于相对频繁地落叶和生长新叶, 对土壤养分的需求量大, 有高的资源获取率, 属于营养获取型; 常绿物种叶片寿命长, 对养分需求较少, 故常绿习性被认为是对贫瘠土壤生境的一种适应性(Aerts, 1995; Givnish, 2002).上述研究结果表明, 落叶物种倾向于采取获取策略, 而常绿种类通常采取保守策略(Givnish, 2002).但研究叶性状关系并不能简单地直接套用其他尺度的研究结果, 全球尺度的LES是否适用于局地尺度仍值得探究(何芸雨等, 2019).本研究力求验证武夷山的常绿和落叶物种是否也能够沿着综合的WPES被区分开来.因此, 本研究假设武夷山的常绿和落叶物种将分布于植物经济谱的两侧: 常绿物种分布在保守的一侧, 落叶物种分布在获取的一侧(假设2). ...
Intraspecific trait variation and coordination: root and leaf economics spectra in coffee across environmental gradients 2 2017
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
... 叶片和细根性状之间及其性状轴之间的显著相关性表明了LES和RES之间的耦合(Reich, 2014; Zhao et al., 2017), 因此本研究证实了假设1, 即经济谱存在于整株植物水平上.虽然有研究表明叶和根的性状之间存在解耦演化(Fortunel et al., 2012; Isaac et al., 2017), 但叶和根之间性状协调的观点依然得到众多研究的支持(Freschet et al., 2010; Liu et al., 2010; de la Riva et al., 2016; Wang et al., 2017).有研究表明, 叶、根二者的形态性状(如SLA和SRL)存在显著相关性(Liu et al., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017).本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL).除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性.本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变.本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD).对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资.因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长.有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014).本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据. ...
Economic strategies of plant absorptive roots vary with root diameter 1 2016
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
Nonlinearity of root trait relationships and the root economics spectrum 1 2019
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
Community-level economics spectrum of fine-roots driven by nutrient limitations in subalpine forests 1 2019
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
Coordinated variation in leaf and root traits across multiple spatial scales in Chinese semi-arid and arid ecosystems 5 2010
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
... ., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
... 叶片和细根性状之间及其性状轴之间的显著相关性表明了LES和RES之间的耦合(Reich, 2014; Zhao et al., 2017), 因此本研究证实了假设1, 即经济谱存在于整株植物水平上.虽然有研究表明叶和根的性状之间存在解耦演化(Fortunel et al., 2012; Isaac et al., 2017), 但叶和根之间性状协调的观点依然得到众多研究的支持(Freschet et al., 2010; Liu et al., 2010; de la Riva et al., 2016; Wang et al., 2017).有研究表明, 叶、根二者的形态性状(如SLA和SRL)存在显著相关性(Liu et al., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017).本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL).除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性.本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变.本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD).对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资.因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长.有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014).本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据. ...
... ., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017).本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL).除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性.本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变.本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD).对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资.因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长.有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014).本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据. ...
Evolutionary history resolves global organization of root functional traits 1 2018
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
不同尺度生态学中植物叶性状研究概述 1 2012
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
不同尺度生态学中植物叶性状研究概述 1 2012
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
Decoupled leaf and root carbon economics is a key component in the ecological diversity and evolutionary divergence of deciduous and evergreen lineages of genus Rhododendron 2 2017
... 植物经济谱能够反映具有相似策略的物种如何沿着性状变异轴汇集或分散(Díazet al., 2016; Zhao et al., 2017).Wright等(2004)指出当物种按照叶习性来划分时, 不同物种的叶片投资策略会沿着全球叶经济谱分布, 即在LES上能够区分出常绿和落叶物种.常绿和落叶物种在叶经济性状方面的确存在一定的差异.与常绿物种相比, 落叶物种的叶面积较大(唐青青等, 2016), 单位面积的叶质量较低(即LMA低), 叶片养分的含量相对较高, 光合速率也较高(Reich et al., 1997; Aerts, 1999; Westoby et al., 2002; Wright et al., 2004).除此之外, 在叶和根形态性状关系上, 落叶进化枝在比叶面积(SLA)和比根长(SRL)间表现出显著的负相关关系(高SLA可以在短生长季节促进更快生长, 而低SRL可以保护根免于冻结(Geng et al., 2014)); 但常绿进化枝显示出较弱的正相关关系或无显著相关性(Medeiros et al., 2017).落叶物种由于相对频繁地落叶和生长新叶, 对土壤养分的需求量大, 有高的资源获取率, 属于营养获取型; 常绿物种叶片寿命长, 对养分需求较少, 故常绿习性被认为是对贫瘠土壤生境的一种适应性(Aerts, 1995; Givnish, 2002).上述研究结果表明, 落叶物种倾向于采取获取策略, 而常绿种类通常采取保守策略(Givnish, 2002).但研究叶性状关系并不能简单地直接套用其他尺度的研究结果, 全球尺度的LES是否适用于局地尺度仍值得探究(何芸雨等, 2019).本研究力求验证武夷山的常绿和落叶物种是否也能够沿着综合的WPES被区分开来.因此, 本研究假设武夷山的常绿和落叶物种将分布于植物经济谱的两侧: 常绿物种分布在保守的一侧, 落叶物种分布在获取的一侧(假设2). ...
... 叶片和细根性状之间及其性状轴之间的显著相关性表明了LES和RES之间的耦合(Reich, 2014; Zhao et al., 2017), 因此本研究证实了假设1, 即经济谱存在于整株植物水平上.虽然有研究表明叶和根的性状之间存在解耦演化(Fortunel et al., 2012; Isaac et al., 2017), 但叶和根之间性状协调的观点依然得到众多研究的支持(Freschet et al., 2010; Liu et al., 2010; de la Riva et al., 2016; Wang et al., 2017).有研究表明, 叶、根二者的形态性状(如SLA和SRL)存在显著相关性(Liu et al., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017).本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL).除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性.本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变.本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD).对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资.因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长.有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014).本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据. ...
Coordinated evolution of leaf and stem economics in tropical dry forest trees 2 2012
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
Evidence for a “plant community economics spectrum” driven by nutrient and water limitations in a Mediterranean rangeland of southern France 2 2012
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
Trait correlation networks: a whole-plant perspective on the recently criticized leaf economic spectrum 1 2014
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
Root functional parameters along a land-use gradient: evidence of a community- level economics spectrum 1 2015
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
植物经济谱研究进展 1 2018
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
植物经济谱研究进展 1 2018
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
The world-wide “fast-slow” plant economics spectrum: a traits manifesto 9 2014
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
... ), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
... 叶片和细根性状之间及其性状轴之间的显著相关性表明了LES和RES之间的耦合(Reich, 2014; Zhao et al., 2017), 因此本研究证实了假设1, 即经济谱存在于整株植物水平上.虽然有研究表明叶和根的性状之间存在解耦演化(Fortunel et al., 2012; Isaac et al., 2017), 但叶和根之间性状协调的观点依然得到众多研究的支持(Freschet et al., 2010; Liu et al., 2010; de la Riva et al., 2016; Wang et al., 2017).有研究表明, 叶、根二者的形态性状(如SLA和SRL)存在显著相关性(Liu et al., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017).本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL).除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性.本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变.本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD).对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资.因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长.有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014).本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据. ...
Generality of leaf trait relationships: a test across six biomes 1 1999
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
Close association of RGR, leaf and root morphology, seed mass and shade tolerance in seedlings of nine boreal tree species grown in high and low light 1 1998
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
From tropics to tundra: global convergence in plant functioning 1 1997
... 植物经济谱能够反映具有相似策略的物种如何沿着性状变异轴汇集或分散(Díazet al., 2016; Zhao et al., 2017).Wright等(2004)指出当物种按照叶习性来划分时, 不同物种的叶片投资策略会沿着全球叶经济谱分布, 即在LES上能够区分出常绿和落叶物种.常绿和落叶物种在叶经济性状方面的确存在一定的差异.与常绿物种相比, 落叶物种的叶面积较大(唐青青等, 2016), 单位面积的叶质量较低(即LMA低), 叶片养分的含量相对较高, 光合速率也较高(Reich et al., 1997; Aerts, 1999; Westoby et al., 2002; Wright et al., 2004).除此之外, 在叶和根形态性状关系上, 落叶进化枝在比叶面积(SLA)和比根长(SRL)间表现出显著的负相关关系(高SLA可以在短生长季节促进更快生长, 而低SRL可以保护根免于冻结(Geng et al., 2014)); 但常绿进化枝显示出较弱的正相关关系或无显著相关性(Medeiros et al., 2017).落叶物种由于相对频繁地落叶和生长新叶, 对土壤养分的需求量大, 有高的资源获取率, 属于营养获取型; 常绿物种叶片寿命长, 对养分需求较少, 故常绿习性被认为是对贫瘠土壤生境的一种适应性(Aerts, 1995; Givnish, 2002).上述研究结果表明, 落叶物种倾向于采取获取策略, 而常绿种类通常采取保守策略(Givnish, 2002).但研究叶性状关系并不能简单地直接套用其他尺度的研究结果, 全球尺度的LES是否适用于局地尺度仍值得探究(何芸雨等, 2019).本研究力求验证武夷山的常绿和落叶物种是否也能够沿着综合的WPES被区分开来.因此, 本研究假设武夷山的常绿和落叶物种将分布于植物经济谱的两侧: 常绿物种分布在保守的一侧, 落叶物种分布在获取的一侧(假设2). ...
The evolution of plant functional variation: traits, spectra, and strategies 1 2003
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
Root structure-function relationships in 74 species: evidence of a root economics spectrum related to carbon economy 1 2016
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
亚热带常绿落叶阔叶混交林植物功能性状的种间和种内变异 1 2016
... 植物经济谱能够反映具有相似策略的物种如何沿着性状变异轴汇集或分散(Díazet al., 2016; Zhao et al., 2017).Wright等(2004)指出当物种按照叶习性来划分时, 不同物种的叶片投资策略会沿着全球叶经济谱分布, 即在LES上能够区分出常绿和落叶物种.常绿和落叶物种在叶经济性状方面的确存在一定的差异.与常绿物种相比, 落叶物种的叶面积较大(唐青青等, 2016), 单位面积的叶质量较低(即LMA低), 叶片养分的含量相对较高, 光合速率也较高(Reich et al., 1997; Aerts, 1999; Westoby et al., 2002; Wright et al., 2004).除此之外, 在叶和根形态性状关系上, 落叶进化枝在比叶面积(SLA)和比根长(SRL)间表现出显著的负相关关系(高SLA可以在短生长季节促进更快生长, 而低SRL可以保护根免于冻结(Geng et al., 2014)); 但常绿进化枝显示出较弱的正相关关系或无显著相关性(Medeiros et al., 2017).落叶物种由于相对频繁地落叶和生长新叶, 对土壤养分的需求量大, 有高的资源获取率, 属于营养获取型; 常绿物种叶片寿命长, 对养分需求较少, 故常绿习性被认为是对贫瘠土壤生境的一种适应性(Aerts, 1995; Givnish, 2002).上述研究结果表明, 落叶物种倾向于采取获取策略, 而常绿种类通常采取保守策略(Givnish, 2002).但研究叶性状关系并不能简单地直接套用其他尺度的研究结果, 全球尺度的LES是否适用于局地尺度仍值得探究(何芸雨等, 2019).本研究力求验证武夷山的常绿和落叶物种是否也能够沿着综合的WPES被区分开来.因此, 本研究假设武夷山的常绿和落叶物种将分布于植物经济谱的两侧: 常绿物种分布在保守的一侧, 落叶物种分布在获取的一侧(假设2). ...
亚热带常绿落叶阔叶混交林植物功能性状的种间和种内变异 1 2016
... 植物经济谱能够反映具有相似策略的物种如何沿着性状变异轴汇集或分散(Díazet al., 2016; Zhao et al., 2017).Wright等(2004)指出当物种按照叶习性来划分时, 不同物种的叶片投资策略会沿着全球叶经济谱分布, 即在LES上能够区分出常绿和落叶物种.常绿和落叶物种在叶经济性状方面的确存在一定的差异.与常绿物种相比, 落叶物种的叶面积较大(唐青青等, 2016), 单位面积的叶质量较低(即LMA低), 叶片养分的含量相对较高, 光合速率也较高(Reich et al., 1997; Aerts, 1999; Westoby et al., 2002; Wright et al., 2004).除此之外, 在叶和根形态性状关系上, 落叶进化枝在比叶面积(SLA)和比根长(SRL)间表现出显著的负相关关系(高SLA可以在短生长季节促进更快生长, 而低SRL可以保护根免于冻结(Geng et al., 2014)); 但常绿进化枝显示出较弱的正相关关系或无显著相关性(Medeiros et al., 2017).落叶物种由于相对频繁地落叶和生长新叶, 对土壤养分的需求量大, 有高的资源获取率, 属于营养获取型; 常绿物种叶片寿命长, 对养分需求较少, 故常绿习性被认为是对贫瘠土壤生境的一种适应性(Aerts, 1995; Givnish, 2002).上述研究结果表明, 落叶物种倾向于采取获取策略, 而常绿种类通常采取保守策略(Givnish, 2002).但研究叶性状关系并不能简单地直接套用其他尺度的研究结果, 全球尺度的LES是否适用于局地尺度仍值得探究(何芸雨等, 2019).本研究力求验证武夷山的常绿和落叶物种是否也能够沿着综合的WPES被区分开来.因此, 本研究假设武夷山的常绿和落叶物种将分布于植物经济谱的两侧: 常绿物种分布在保守的一侧, 落叶物种分布在获取的一侧(假设2). ...
Linking leaf and root trait syndromes among 39 grassland and savannah species 2 2005
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
... 叶片和细根性状之间及其性状轴之间的显著相关性表明了LES和RES之间的耦合(Reich, 2014; Zhao et al., 2017), 因此本研究证实了假设1, 即经济谱存在于整株植物水平上.虽然有研究表明叶和根的性状之间存在解耦演化(Fortunel et al., 2012; Isaac et al., 2017), 但叶和根之间性状协调的观点依然得到众多研究的支持(Freschet et al., 2010; Liu et al., 2010; de la Riva et al., 2016; Wang et al., 2017).有研究表明, 叶、根二者的形态性状(如SLA和SRL)存在显著相关性(Liu et al., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017).本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL).除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性.本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变.本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD).对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资.因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长.有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014).本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据. ...
Global quantification of contrasting leaf life span strategies for deciduous and evergreen species in response to environmental conditions 2012
An introduction to R. Notes on R: a programming environment for data analysis and graphics 1 2019
Relationships among leaf, stem and root traits of the dominant shrubs from four vegetation zones in Shaanxi Province, China 2 2017
... 独立于生境的植物经济谱, 能够定量地解释植物总体在生长速率及胁迫环境中的耐受策略差异(曲鹏等, 2018).有研究表明, 植物存在与LES类似的代表根在生产力与持久性之间权衡策略的根经济谱(RES)(Reich, 2014; Prieto et al., 2015), 植物的根性状沿着两种不同的策略边界范围演化: 一种是具有保守性策略的粗根系植物, 而另一种是具有获取性策略的细根系植物(Ma et al., 2018).Ding等(2020)的研究指出, 细根性状的变异分为两个主要维度: 第一维度主要由根直径(RD)和比根长(SRL)代表, 表示根寿命和资源获取效率之间的权衡; 第二维度代表了根养分(即N、P)和根组织密度(RTD)的协调变化, 表示资源吸收的获取-保守权衡, 即RES.但Kong等(2019)研究发现, 木本植物的RD与RTD、根氮含量(RNC)之间存在非线性关系, 这种非线性关系不支持RES的预测.因此木本植物RES是否存在仍值得研究.除此之外, 由于生物普遍存在强烈的物理限制以及性状和生活史的权衡选择(Reich et al., 1999), Reich (2014)认为所有器官中存在统一的资源获取和处理速率(快、中或慢速), 存在一条综合的整株植物经济谱(WPES).关于叶和根之间的关系, Freschet等(2010)、de la Riva等(2016)、Wang等(2017)和Liu等(2010)诸多****的研究均支持二者及其与整株植物之间的功能性状存在高度协调的观点.然而, Fortunel等(2012)研究发现木质根性状与叶性状并无显著相关; Isaac等(2017)还发现种内LES和RES彼此独立, 根性状会随环境条件而变化, 但叶性状及其功能不一定会发生相应的变化.有研究表明, 当设置不同的处理和环境梯度时, 叶、根性状间的相关性并不一致(Reich et al., 1998; Craine & Lee, 2003; Craine et al., 2005; Tjoelker et al., 2005; Withington et al., 2006; Liu et al., 2010).因此, 关于植物不同器官(即叶和根)的性状如何在不同物种之间共变, 以及这种共变与整株植物经济谱的相关程度等均存在争议(Freschet et al., 2010; de la Riva et al., 2016), 表明这仍然是一个值得研究的问题.通常, 良好的叶功能取决于根部吸收的水分和养分, 而根部生长又取决于叶光合作用产生的碳水化合物(Chapin III, 1980).植物叶和根的生态策略应该是建立在WPES上的, 叶和根性状的变异共同决定了物种的表现(Grime et al., 1997; Pérez-Ramoset al., 2012; Poorter et al., 2014; Reich, 2014).因此, 本研究假设武夷山木本植物中存在LES和RES, 它们二者与WPES紧密结合且相互之间高度协调(假设1). ...
... 叶片和细根性状之间及其性状轴之间的显著相关性表明了LES和RES之间的耦合(Reich, 2014; Zhao et al., 2017), 因此本研究证实了假设1, 即经济谱存在于整株植物水平上.虽然有研究表明叶和根的性状之间存在解耦演化(Fortunel et al., 2012; Isaac et al., 2017), 但叶和根之间性状协调的观点依然得到众多研究的支持(Freschet et al., 2010; Liu et al., 2010; de la Riva et al., 2016; Wang et al., 2017).有研究表明, 叶、根二者的形态性状(如SLA和SRL)存在显著相关性(Liu et al., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017).本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL).除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性.本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变.本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD).对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资.因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长.有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014).本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据. ...
Evergreen trees do not maximize instantaneous photosynthesis 2 2004
Plant ecological strategies: some leading dimensions of variation between species 3 2002
... 植物经济谱能够反映具有相似策略的物种如何沿着性状变异轴汇集或分散(Díazet al., 2016; Zhao et al., 2017).Wright等(2004)指出当物种按照叶习性来划分时, 不同物种的叶片投资策略会沿着全球叶经济谱分布, 即在LES上能够区分出常绿和落叶物种.常绿和落叶物种在叶经济性状方面的确存在一定的差异.与常绿物种相比, 落叶物种的叶面积较大(唐青青等, 2016), 单位面积的叶质量较低(即LMA低), 叶片养分的含量相对较高, 光合速率也较高(Reich et al., 1997; Aerts, 1999; Westoby et al., 2002; Wright et al., 2004).除此之外, 在叶和根形态性状关系上, 落叶进化枝在比叶面积(SLA)和比根长(SRL)间表现出显著的负相关关系(高SLA可以在短生长季节促进更快生长, 而低SRL可以保护根免于冻结(Geng et al., 2014)); 但常绿进化枝显示出较弱的正相关关系或无显著相关性(Medeiros et al., 2017).落叶物种由于相对频繁地落叶和生长新叶, 对土壤养分的需求量大, 有高的资源获取率, 属于营养获取型; 常绿物种叶片寿命长, 对养分需求较少, 故常绿习性被认为是对贫瘠土壤生境的一种适应性(Aerts, 1995; Givnish, 2002).上述研究结果表明, 落叶物种倾向于采取获取策略, 而常绿种类通常采取保守策略(Givnish, 2002).但研究叶性状关系并不能简单地直接套用其他尺度的研究结果, 全球尺度的LES是否适用于局地尺度仍值得探究(何芸雨等, 2019).本研究力求验证武夷山的常绿和落叶物种是否也能够沿着综合的WPES被区分开来.因此, 本研究假设武夷山的常绿和落叶物种将分布于植物经济谱的两侧: 常绿物种分布在保守的一侧, 落叶物种分布在获取的一侧(假设2). ...
... 叶片和细根性状之间及其性状轴之间的显著相关性表明了LES和RES之间的耦合(Reich, 2014; Zhao et al., 2017), 因此本研究证实了假设1, 即经济谱存在于整株植物水平上.虽然有研究表明叶和根的性状之间存在解耦演化(Fortunel et al., 2012; Isaac et al., 2017), 但叶和根之间性状协调的观点依然得到众多研究的支持(Freschet et al., 2010; Liu et al., 2010; de la Riva et al., 2016; Wang et al., 2017).有研究表明, 叶、根二者的形态性状(如SLA和SRL)存在显著相关性(Liu et al., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017).本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL).除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性.本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变.本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD).对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资.因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长.有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014).本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据. ...
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
内蒙古荒漠草原植物叶片功能性状关系及其经济谱分析 1 2014
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
The plant economics spectrum is structured by leaf habits and growth forms across subtropical species 5 2017
... “植物功能性状”是指对植物体定植、存活、适应度等具有潜在显著影响的一系列植物属性(Reichet al., 2003), 是植物在长期进化过程中适应环境的产物(Díazet al., 2016), 并且能对生态系统过程产生强烈影响(Cornelissen et al., 2003).将经济学的“投资-收益权衡”理论应用到植物的资源分配研究中, 可定量分析植物叶片功能性状间的相互关系(陈莹婷和许振柱,2014).Wright等(2004)在全球尺度上定义出一条连续变化的叶功能性状组合谱, 即“叶经济谱(LES)”, 用以阐述维管植物资源获取和储存之间的权衡策略, 其可以通过性状指标的变化范围及其数量关系表现出来(Wrightet al., 2005).谱的一端代表植物的“快速投资-收益型”策略, 拥有该特征的物种具有廉价的组织投资与快速的投资回报(即获取策略); 而另一端则代表植物的“缓慢投资-收益型”策略, 拥有该特征的物种具有昂贵的组织投资和较慢的投资回报(即保守策略)(Wrightet al., 2004; Reich, 2014; Zhao et al., 2017).在后续的研究中, 经济谱研究陆续被相关生态****推广到茎(Baraloto et al., 2010; Méndez-Alonzoet al., 2012), 根(Kong et al., 2016; Roumet et al., 2016), 整株植物(Freschet et al., 2010; de la Riva et al., 2016), 不同层次的群落结构(Pérez-Ramoset al., 2012; Funk & Cornwell, 2013; Li et al., 2019)及生态系统(Freschet et al., 2013; 于鸿莹等, 2014).叶片作为维持陆地生态系统机能的一个最基本要素, 不仅是植物光合作用以及物质生产的主要器官, 而且是植物和大气环境进行水气交换的主要器官(毛伟等, 2012).细根(包括经常定植其中的菌根形成的菌丝)在植物中起着吸收水分和养分的作用(Chen et al., 2019), 细根的生产、周转是森林生态系统进行物质循环以及能量流动的重要组成部分(Eissenstat & Yanai, 1997).二者对植物生长及森林生态系统健康维持具有重要作用, 因此, 本研究将这两个代谢活性较强的植物器官作为研究对象. ...
... 植物经济谱能够反映具有相似策略的物种如何沿着性状变异轴汇集或分散(Díazet al., 2016; Zhao et al., 2017).Wright等(2004)指出当物种按照叶习性来划分时, 不同物种的叶片投资策略会沿着全球叶经济谱分布, 即在LES上能够区分出常绿和落叶物种.常绿和落叶物种在叶经济性状方面的确存在一定的差异.与常绿物种相比, 落叶物种的叶面积较大(唐青青等, 2016), 单位面积的叶质量较低(即LMA低), 叶片养分的含量相对较高, 光合速率也较高(Reich et al., 1997; Aerts, 1999; Westoby et al., 2002; Wright et al., 2004).除此之外, 在叶和根形态性状关系上, 落叶进化枝在比叶面积(SLA)和比根长(SRL)间表现出显著的负相关关系(高SLA可以在短生长季节促进更快生长, 而低SRL可以保护根免于冻结(Geng et al., 2014)); 但常绿进化枝显示出较弱的正相关关系或无显著相关性(Medeiros et al., 2017).落叶物种由于相对频繁地落叶和生长新叶, 对土壤养分的需求量大, 有高的资源获取率, 属于营养获取型; 常绿物种叶片寿命长, 对养分需求较少, 故常绿习性被认为是对贫瘠土壤生境的一种适应性(Aerts, 1995; Givnish, 2002).上述研究结果表明, 落叶物种倾向于采取获取策略, 而常绿种类通常采取保守策略(Givnish, 2002).但研究叶性状关系并不能简单地直接套用其他尺度的研究结果, 全球尺度的LES是否适用于局地尺度仍值得探究(何芸雨等, 2019).本研究力求验证武夷山的常绿和落叶物种是否也能够沿着综合的WPES被区分开来.因此, 本研究假设武夷山的常绿和落叶物种将分布于植物经济谱的两侧: 常绿物种分布在保守的一侧, 落叶物种分布在获取的一侧(假设2). ...
... 叶片和细根性状之间及其性状轴之间的显著相关性表明了LES和RES之间的耦合(Reich, 2014; Zhao et al., 2017), 因此本研究证实了假设1, 即经济谱存在于整株植物水平上.虽然有研究表明叶和根的性状之间存在解耦演化(Fortunel et al., 2012; Isaac et al., 2017), 但叶和根之间性状协调的观点依然得到众多研究的支持(Freschet et al., 2010; Liu et al., 2010; de la Riva et al., 2016; Wang et al., 2017).有研究表明, 叶、根二者的形态性状(如SLA和SRL)存在显著相关性(Liu et al., 2010; Geng et al., 2014; de la Riva et al., 2016; Medeiros et al., 2017).本研究中, ILA和SRL呈显著正相关关系, 表明物种是通过利用较大的叶进行光合作用固碳, 同时增加比根长以充分吸收植物生长所需的营养物质(即高ILA和SRL).除此之外, 叶和根的N含量(Craine et al., 2005; Liu et al., 2010)、C含量、C:N、木质素含量、干物质含量、pH (Freschet et al., 2010)、呼吸速率(Tjoelker et al., 2005)等性状指标之间均被证实存在显著相关性.本研究从群落总体水平上看LCC、LNC和LPC分别与RCC、RNC和RPC呈极显著正相关关系;LNC与RPC呈极显著正相关关系; LPC与RNC也呈显著正相关关系, 这表明武夷山木本植物叶片与细根之间的化学元素存在协同共变.本研究中LNC与RTD的负相关关系表明, 该地区植物群落中, 一方面存在着通过增加LNC以提高其光合能力的快速生长的物种(即高LNC), 另一方面, 还存在具有高根组织支出的物种(即高RTD).对于具有低LNC的物种来说, 建造具有高组织密度的根(即高RTD)用以提高承受物理伤害的能力是一种昂贵的组织投资.因此, 在整株植物水平上看, 具有高根组织密度的物种对应于保守型的叶片策略(Westoby et al., 2002)和缓慢生长(Reich, 2014); 反之, 低RTD的物种因为具有高LNC, 倾向于快速生长.有研究指出, 当植物在任何器官(叶和根)水平上是快速获取或使用碳(营养元素或水)时, 将要求其在同一器官水平对其他资源也是快速获取; 若某一器官水平(如叶水平)上的所有资源均是快速获取, 则将要求其他器官水平(如根水平)对所有资源也是快速获取(Reich, 2014).本研究中, 叶片、细根以及整株植物各性状贡献率相对较大的PC1轴之间均是显著正相关的(p < 0.05), 且常绿和落叶物种在性状轴上具有相同的分异现象, 表明叶片、细根以及整株植物之间的资源策略存在协同变化, 说明亚热带木本植物叶片与细根遵循着WPES策略的协调整合, 研究结果为全球尺度经济谱研究提供了局地性的证据. ...
 ,1,4,*
,1,4,*

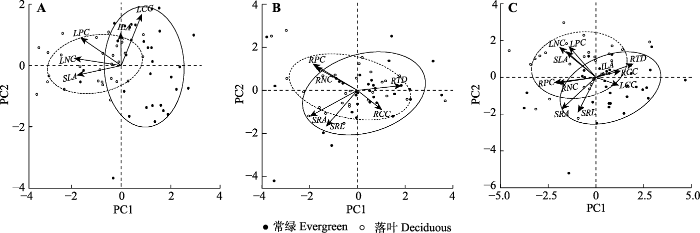 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT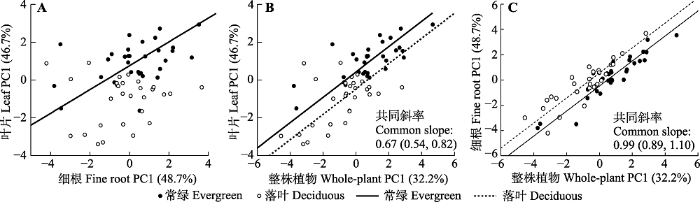 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}