Effects of dissolved organic matter from different plant sources on soil enzyme activities in subtropical forests
Kong-Can MEI1,2, Lei CHENG1,2, Qiu-Fang ZHANG1,2, Kai-Miao LIN2,3, Jia-Cong ZHOU1,2, Quan-Xin ZENG1,2, Yue WU1,2, Jian-Guo XU3, Jin-Rong ZHOU1,2, Yue-Min CHEN,1,2,*1School of Geographical Science, Fujian Normal University, Fuzhou 350007, China 2Cultivation Base of State Key Laboratory of Humid Subtropical Mountain Ecology, Fuzhou 350007, China 3Daiyun Mountain National Nature Reserve Administration Bureau, Quanzhou, Fujian 362500, China
National Natural Science Foundation of China (31670620),(31670620) Fujian Provincial Training Program of Innovation and Entrepreneurship for Undergraduates(S201910394028)
Abstract Aims This study aims to explore the effects of dissolved organic matter (DOM) from different plant sources on subtropical forest soil enzyme activities in order to provide a scientific basis for soil carbon cycling under rainfall leaching conditions in different forest ecosystems of subtropical areas. Methods Three kinds of DOM, extracted from the fresh leaves of Cunninghamia lanceolata, Schima superb, and Phoebe zherman, were added to the soils of C. lanceolata plantation. Soil treated with deionized water of the same amount as the DOM solution served as the control. Incubation of treated and control soils was conducted for 25 days in the laboratory. The physical and chemical properties, microbial biomass, and enzyme activities of the soils were determined after incubation. Important findings The results showed that, compared with the control treatment (CT), soil total organic carbon (SOC) content, soil total nitrogen (TN) content, and carbon-nitrogen ratio had no significant changes after adding DOM. However, the TN of treatment with added C. lanceolata leaf DOM (CL) was significantly lower than that of treatments with added S. superb leaf DOM (SL) and P. zherman leaf DOM (PL), and the carbon-nitrogen ratio in CL was significantly higher than that in SL and PL. The three treatments with added DOM increased soil dissolved organic carbon (DOC) content and dissolved organic nitrogen (DON) content overall. There was no significant change in soil microbial biomass carbon (MBC) content after all three DOM inputs, while the microbial biomass nitrogen (MBN) content in CL and SL treatments was reduced by 50.9% and 51.1%, respectively, compared to CT. However, MBN content in PL treatment was increased by 54.0% than CT. DOM input significantly increased the activities of β-glucosidase, cellobiohydrolase, and peroxidase in comparison with CT, but significantly decreased the activity of polyphenol oxidase. In addition, both β-glucosidase and cellobiohydrolase activities showed the following characteristics: CL > SL > PL. The results of correlation analysis showed that the contents of SOC, TN and MBN and the activities of β-glucosidase and cellobiohydrolase of the treatments with DOM were significantly correlated with the DOC content and humification index (HIX) of added DOM. In addition, soil MBN content and polyphenol oxidase activity were positively correlated with the pH value of added DOM. The results of redundancy analysis (RDA) showed that the key factors causing the change in soil enzyme activities after DOM input were DON and DOC content. In general, differences in the properties of DOM obtained from different plant sources affect the activities of soil carbon-acquisition hydrolase. DOM input increases the availability of soil carbon and nitrogen, and elicits different responses from the four carbon-acquisition enzymes. This study provides a theoretical basis for exploring the process of carbon cycling in the rain-rich subtropical forest ecosystem. Keywords:dissolved organic matter;soil enzyme;soil nutrient availability;fresh leaves;subtropical forest
PDF (1168KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 梅孔灿, 程蕾, 张秋芳, 林开淼, 周嘉聪, 曾泉鑫, 吴玥, 徐建国, 周锦容, 陈岳民. 不同植物来源可溶性有机质对亚热带森林土壤酶活性的影响. 植物生态学报, 2020, 44(12): 1273-1284. DOI: 10.17521/cjpe.2020.0097 MEI Kong-Can, CHENG Lei, ZHANG Qiu-Fang, LIN Kai-Miao, ZHOU Jia-Cong, ZENG Quan-Xin, WU Yue, XU Jian-Guo, ZHOU Jin-Rong, CHEN Yue-Min. Effects of dissolved organic matter from different plant sources on soil enzyme activities in subtropical forests. Chinese Journal of Plant Ecology, 2020, 44(12): 1273-1284. DOI: 10.17521/cjpe.2020.0097
Table 1 表1 表1不同植物叶片及其可溶性有机质(DOM)的性质(平均值±标准偏差) Table 1Characterizations of leaves and dissolved organic matter (DOM) from different plant sources (mean ± SD)
同一列不同小写字母表示不同叶片DOM的性质差异显著(p < 0.05)。 Different lowercase letters in the same column indicate significant differences among different properties of DOM extracted from different leaves (p < 0.05).
Table 2 表2 表2与碳循环相关的4种土壤酶的缩写、底物和功能 Table 2Substrates and functions of four soil enzymes related to carbon cycling
土壤酶 Soil enzyme
缩写 Abbreviation
底物 Substrate
功能 Function
β-葡萄糖苷酶 β-glucosidase
βG
4-MUB-β-D-glucoside
水解纤维素 Hydrolyze cellulose
纤维素水解酶 Cellobiohydrolase
CBH
4-MUB-β-D-cellobioside
水解纤维素 Hydrolyze cellulose
过氧化物酶 Peroxidase
PEO
L-dihydroxyphenylalanine
降解木质素 Degrade lignin
多酚氧化酶 Polyphenol oxidase
PPO
L-dihydroxyphenylalanine
降解木质素 Degrade lignin
两种氧化酶都是以L-DOPA为底物, 但测定过氧化物酶的微孔板中会额外加入10 μL 0.3%的H2O2。 The activities of two oxidative enzymes were determined with L-dihydroxyphenylalanine as the substrate, but 10 μL of 0.3% H2O2 were added to each well of the microplates prepared for Peroxidase measurement.
同一列不同小写字母表示不同处理间差异显著(p < 0.05)。CL, 输入杉木叶片DOM处理; CT, 对照处理; PL, 输入楠木叶片DOM处理; SL, 输入木荷叶片DOM处理。 Different lowercase letters in the same column indicate significant difference among different treatments (p < 0.05). CL, Cunninghamia lanceolata leaf DOM addition treatment; CT, control treatment; PL, Phoebe zherman leaf DOM addition treatment; SL, Schima superba leaf DOM addition treatment.
Table 4 表4 表4土壤理化性质、微生物生物量和酶活性与叶片可溶性有机质(DOM)性质的相关性分析 Table 4Correlation analysis of soil physical and chemical properties, microbial biomass and enzyme activities with properties of dissolved organic matter (DOM) of fresh leaves
DOM性质 DOM property
土壤理化性质 Soil physical and chemical properties
微生物生物量Microbial biomass
土壤酶活性 Soil enzyme activities
SOC
TN
C:N
pH
DOC
DON
MBC
MBN
βG
CBH
PEO
PPO
L-DOC
-0.79*
-0.73*
0.69*
0.59
0.12
0.83**
0.14
-0.78*
0.87**
0.94**
-0.10
-0.25
L-DON
-0.15
0.16
-0.52
0.14
0.16
-0.46
-0.25
-0.27
-0.51
-0.30
-0.20
-0.84**
L-HIX
0.81**
0.79*
-0.69*
-0.59
-0.52
-0.76*
0.04
0.90**
-0.86*
-0.93*
-0.08
0.44
L-pH
0.52
0.65
-0.14
-0.47
-0.39
-0.19
0.14
0.86**
-0.34
-0.54
0.09
0.88**
DOC, 土壤溶解有机碳含量; DON, 土壤溶解有机氮含量; MBC, 微生物生物量碳含量; MBN, 微生物生物量氮含量; SOC, 土壤总有机碳含量; TN, 土壤总氮含量。ΒG, β-葡萄糖苷酶; CBH, 纤维素水解酶; PEO, 过氧化物酶; PPO, 多酚氧化酶。L-DOC, 叶片DOM的可溶性有机碳含量; L-DON, 叶片DOM的可溶性有机氮含量; L-HIX, 叶片DOM的腐殖化指数; L-pH, 叶片DOM的pH。*, p < 0.05; **, p < 0.01。 DOC, soil dissolved organic carbon content; DON, soil dissolved organic nitrogen content; MBC, microbial biomass carbon content; MBN, microbial biomass nitrogen content; SOC, total soil organic carbon content; TN, total soil nitrogen content. βG, β-glucosidase; CBH, cellobiohydrolase; PEO, peroxidase; PPO, polyphenol oxidase. L-DOC, dissolved organic carbon content of the added leaf DOM; L-DON, dissolved organic nitrogen content of the added leaf DOM; L-HIX, humification index of the added leaf DOM; L-pH, pH values of the added leaf DOM. *, p < 0.05; **, p < 0.01.
CL, 输入杉木叶片DOM处理; CT, 对照处理; PL, 输入楠木叶片DOM处理; SL, 输入木荷叶片DOM处理。βG, β-葡萄糖苷酶; CBH, 纤维素水解酶; PEO, 过氧化物酶; PPO, 多酚氧化酶。DOC, 土壤溶解有机碳含量; DON, 土壤溶解有机氮含量; MBN, 微生物生物量氮含量; SOC, 土壤总有机碳含量。图中实线箭头表示物种因子, 虚线箭头表示环境因子。图右下角变量注释表示对土壤酶活性变化有显著影响的环境因子的相应解释比例。 Fig. 3Redundant analysis of the effect of dissolved organic matter (DOM) input on soil enzyme activities.
CL, Cunninghamia lanceolata leaf DOM addition treatment; CT, control treatment; PL, Phoebe zherman leaf DOM addition treatment; SL, Schima superba leaf DOM addition treatment. βG, β-glucosidase; CBH, cellulolytic enzymes; PEO, peroxidase; PPO, polyphenol oxidase. DOC, soil dissolved organic carbon content; DON, soil dissolved organic nitrogen content; MBN, microbial biomass nitrogen content; SOC, total soil organic carbon content. The solid arrows represent species factors and the dotted arrows represent environmental factors. The notes of variables in lower right corner of the picture represent the corresponding proportion of environmental factors that have a significant effect on changes in soil enzyme activities.
3 讨论
3.1 土壤理化性质对不同植物来源DOM输入的响应
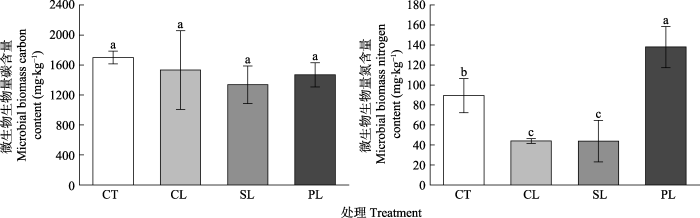
土壤SOC和TN含量的变化对森林生态系统主要功能、组成和发展方向具有重要影响(Zhou et al., 2018)。本研究发现, 杉木和木荷叶片DOM输入后SOC和TN含量降低(表3), 但是土壤DOC和DON浓度升高, 这可能是因为外源DOM输入刺激了土壤有机质的分解(Fontaine et al., 2007), 产生了激发效应, 使SOC和TN转化为DOC和DON或以气体的形式释放到外界。相反, 楠木叶片DOM输入后土壤TN含量略有上升, 这可能与所添加的楠木叶片DOM中的DOC含量较低, HIX较高(表1)有关。输入的叶片DOC含量较低, 微生物可利用的碳源相对较少; HIX越高表示DOM成分越复杂(Miano & Senesi, 1992; Du et al., 2016), 因此楠木叶片DOM较难被微生物利用与分解。相关性分析中DOM添加处理下SOC和TN含量与输入的叶片DOM中DOC含量和HIX显著相关(表4)也进一步支持了这一说法。
?grenGI, BosattaE, MagillAH (2001). Combining theory and experiment to understand effects of inorganic nitrogen on litter decomposition Oecologia, 128, 94-98. DOI:10.1007/s004420100646URLPMID:28547095 [本文引用: 1] It has been long recognised that mineral elements, and nitrogen in particular, play an important role in determining the rate at which organic matter is decomposed. The magnitude and even the sign of the effects are, however, not universal and the underlying mechanisms are not well understood. In this paper, an explanation for the observed decreases in decomposition/CO2 evolution rates when inorganic nitrogen increases is proposed by combining a theoretical approach with the results of a 6-year litter decomposition-forest nitrogen fertilisation experiment. Our results show that the major causes of observed changes in decomposition rate after nitrogen fertilisation are increases in decomposer efficiency, more rapid formation of recalcitrant material, and, although less pronounced, decreased growth rate of decomposers. This gives a more precise description of how inorganic nitrogen modifies decomposition rates than the previously loosely used
AllisonSD, VitousekPM (2005). Responses of extracellular enzymes to simple and complex nutrient inputs Soil Biology & Biochemistry, 37, 937-944. [本文引用: 1]
AllisonSD, WeintraubMN, GartnerTB, WaldropMP (2010). Evolutionary-economic principles as regulators of soil enzyme production and ecosystem function//Shukla G, Varma A Soil Enzymology. Springer, Berlin, Germany. 229-243. [本文引用: 1]
BaoY, GaoY, ZengXM, YuanP, SiYT, ChenYM, ChenYY (2018). Relationships between carbon and nitrogen contents and enzyme activities in soil of three typical subtropical forests in China Chinese Journal of Plant Ecology, 42, 508-516. [本文引用: 1]
BoddyE, HillPW, FarrarJ, JonesDL (2007). Fast turnover of low molecular weight components of the dissolved organic carbon pool of temperate grassland field soils Soil Biology & Biochemistry, 39, 827-835. [本文引用: 1]
ChenGS, YangZJ, GaoR, XieJS, GuoJF, HuangZQ, YangYS (2013). Carbon storage in a chronosequence of Chinese fir plantations in southern China Forest Ecology and Management, 300, 68-76. [本文引用: 1]
ChenJ, LuoYQ, LiJW, ZhouXH, CaoJJ, WangRW, WangYQ, SheltonS, JinZ, WalkerLM, FengZZ, NiuSL, FengWT, JianSY, ZhouLY (2017). Costimulation of soil glycosidase activity and soil respiration by nitrogen addition Global Change Biology, 23, 1328-1337. URLPMID:27362835 [本文引用: 1]
ChenX, HaoB, JingX, HeJS, MaW, ZhuB (2019). Minor responses of soil microbial biomass, community structure and enzyme activities to nitrogen and phosphorus addition in three grassland ecosystems Plant and Soil, 444, 21-37. [本文引用: 1]
CurtinD, CampbellCA, JalilA (1998). Effects of acidity on mineralization: pH-dependence of organic matter mineralization in weakly acidic soils Soil Biology & Biochemistry, 30, 57-64. [本文引用: 1]
DuYX, ZhangYY, ChenFZ, ChangYG, LiuZW (2016). Photochemical reactivities of dissolved organic matter (DOM) in a sub-alpine lake revealed by EEM-PARAFAC: an insight into the fate of allochthonous DOM in alpine lakes affected by climate change Science of the Total Environment, 568, 216-225. [本文引用: 1]
FanYX, ZhongXJ, LinTC, LyuMK, WangMH, HuWF, YangZJ, ChenGS, GuoJF, YangYS (2020). Effects of nitrogen addition on DOM-induced soil priming effects in a subtropical plantation forest and a natural forest Biology and Fertility of Soils, 56, 205-216. [本文引用: 2]
FangY, NazariesL, SinghBK, SinghBP (2018). Microbial mechanisms of carbon priming effects revealed during the interaction of crop residue and nutrient inputs in contrasting soils Global Change Biology, 24, 2775-2790. URLPMID:29603502 [本文引用: 1]
FontaineS, BarotS, BarréP, BdiouiN, MaryB, RumpelC (2007). Stability of organic carbon in deep soil layers controlled by fresh carbon supply Nature, 450, 277-280. DOI:10.1038/nature06275URLPMID:17994095 [本文引用: 1] The world's soils store more carbon than is present in biomass and in the atmosphere. Little is known, however, about the factors controlling the stability of soil organic carbon stocks and the response of the soil carbon pool to climate change remains uncertain. We investigated the stability of carbon in deep soil layers in one soil profile by combining physical and chemical characterization of organic carbon, soil incubations and radiocarbon dating. Here we show that the supply of fresh plant-derived carbon to the subsoil (0.6-0.8 m depth) stimulated the microbial mineralization of 2,567 +/- 226-year-old carbon. Our results support the previously suggested idea that in the absence of fresh organic carbon, an essential source of energy for soil microbes, the stability of organic carbon in deep soil layers is maintained. We propose that a lack of supply of fresh carbon may prevent the decomposition of the organic carbon pool in deep soil layers in response to future changes in temperature. Any change in land use and agricultural practice that increases the distribution of fresh carbon along the soil profile could however stimulate the loss of ancient buried carbon.
GaoYQ, DaiXQ, WangJL, FuXL, KouL, WangHM (2019). Characteristics of soil enzymes stoichiometry in rhizosphere of understory vegetation in subtropical forest plantations Chinese Journal of Plant Ecology, 43, 258-272. [本文引用: 1]
GauthierA, Amiotte-SuchetP, NelsonPN, LévêqueJ, ZellerB, HénaultC (2010). Dynamics of the water extractable organic carbon pool during mineralisation in soils from a Douglas fir plantation and an oak-beech forest—An incubation experiment Plant and Soil, 330, 465-479. [本文引用: 1]
GuoJF, YangYS, ChenGS, LinP (2005). Dissolved organic carbon and nitrogen in precipitation, throughfall and stemflow from Schima superba and Cunninghamia lanceolata plantations in subtropical China Journal of Forestry Research, 16, 19-22. [本文引用: 1]
InubushiK, BrookesPC, JenkinsonDS (1991). Soil microbial biomass C, N and ninhydrin-N in aerobic and anaerobic soils measured by the fumigation-extraction method Soil Biology & Biochemistry, 23, 737-741. [本文引用: 1]
JiYH, ZhangQF, ZhouJC, YouZT, XuPC, LinWS, ChenYM, YangYS (2018). Spectral characteristics and quantities of dissolved organic matter released from leaves with competitive and stress-tolerant ecological strategies in a subtropical region Acta Ecologica Sinica, 38, 3998-4007. [本文引用: 2]
JiangY, LeiY, QinW, KorpelainenH, LiC (2019). Revealing microbial processes and nutrient limitation in soil through ecoenzymatic stoichiometry and glomalin-related soil proteins in a retreating glacier forefield Geoderma, 338, 313-324. [本文引用: 1]
JingX, ChenX, TangM, DingZJ, JiangL, LiP, MaSH, TianD, XuLC, ZhuJX, JiCJ, ShenHH, ZhengCY, FangJY, ZhuB (2017). Nitrogen deposition has minor effect on soil extracellular enzyme activities in six Chinese forests Science of the Total Environment, 607-608, 806-815. [本文引用: 1]
JonesDL, WillettVB (2006). Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil Soil Biology & Biochemistry, 38, 991-999. [本文引用: 1]
KalbitzK, GeyerW, GeyerS (1999). Spectroscopic properties of dissolved humic substances—A reflection of land use history in a fen area Biogeochemistry, 47, 219-238. [本文引用: 1]
KalbitzK, SolingerS, ParkJH, MichalzikB, MatznerE (2000). Controls on the dynamics of dissolved organic matter in soils: a review Soil Science, 165, 277-304. [本文引用: 1]
KangGL, GaoR, YangYS, ChenGS, YangZJ, SiYT (2014). Quantities and qualities of leaf-leached DOM of four species in a secondary Castanopsis carlesii forest Journal of Subtropical Resources and Environment, 9, 30-37.
LiYL, ZhangXB, RenFL, SunN, XuM, XuMG (2020). A meta-analysis of long-term fertilization impact on soil dissolved organic carbon and nitrogen across Chinese cropland Scientia Agricultura Sinica, 53, 1224-1233. [本文引用: 1]
LinKM, ZhangZQ, YeFM, LinY, LiQS (2010). Dynamic analysis of decomposition characteristics and content change of nutrient elements of leaf litter of Cunninghamia lanceolata, Phoebe bournei and Schima superba under C. lanceolata artificial forest Journal of Plant Resources and Environment, 19(2), 34-39. [本文引用: 1]
Pérez-HarguindeguyN, DíazS, GarnierE, LavorelS, PoorterH, JaureguiberryP, Bret-HarteMS, CornwellWK, CraineJM, GurvichDE, UrcelayC, VeneklaasEJ, ReichPB, PoorterL, WrightIJ, et al. (2013). New handbook for standardised measurement of plant functional traits worldwide Australian Journal of Botany, 61, 167-234. [本文引用: 1]
PetersonME, CurtinD, ThomasS, CloughTJ, MeenkenED (2013). Denitrification in vadose zone material amended with dissolved organic matter from topsoil and subsoil Soil Biology & Biochemistry, 61, 96-104. [本文引用: 1]
PiaoS, FangJ, CiaisP, PeylinP, HuangY, SitchS, WangT (2009). The carbon balance of terrestrial ecosystems in China Nature, 458, 1009-1013. URLPMID:19396142 [本文引用: 1]
Saiya-CorkKR, SinsabaughRL, ZakDR (2002). The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil Soil Biology & Biochemistry, 34, 1309-1315. [本文引用: 2]
SchimelJ, BalserTC, WallensteinM (2007). Microbial stress response physiology and its implications for ecosystem function Ecology, 88, 1386-1394. DOI:10.1890/06-0219URLPMID:17601131 [本文引用: 1] Microorganisms have a variety of evolutionary adaptations and physiological acclimation mechanisms that allow them to survive and remain active in the face of environmental stress. Physiological responses to stress have costs at the organismal level that can result in altered ecosystem-level C, energy, and nutrient flows. These large-scale impacts result from direct effects on active microbes' physiology and by controlling the composition of the active microbial community. We first consider some general aspects of how microbes experience environmental stresses and how they respond to them. We then discuss the impacts of two important ecosystem-level stressors, drought and freezing, on microbial physiology and community composition. Even when microbial community response to stress is limited, the physiological costs imposed on soil microbes are large enough that they may cause large shifts in the allocation and fate of C and N. For example, for microbes to synthesize the osmolytes they need to survive a single drought episode they may consume up to 5% of total annual net primary production in grassland ecosystems, while acclimating to freezing conditions switches Arctic tundra soils from immobilizing N during the growing season to mineralizing it during the winter. We suggest that more effectively integrating microbial ecology into ecosystem ecology will require a more complete integration of microbial physiological ecology, population biology, and process ecology.
SinghJS, GuptaVK (2018). Soil microbial biomass: a key soil driver in management of ecosystem functioning Science of the Total Environment, 634, 497-500. [本文引用: 1]
SinsabaughRL (2010). Phenol oxidase, peroxidase and organic matter dynamics of soil Soil Biology & Biochemistry, 42, 391-404. [本文引用: 7]
SinsabaughRL, CarreiroMM, RepertDA (2002). Allocation of extracellular enzymatic activity in relation to litter composition, N deposition, and mass loss Biogeochemistry, 60, 1-24. DOI:10.1023/A:1016541114786URL [本文引用: 1]
?najdrJ, Valá?kováV, MerhautováV, HerinkováJ, CajthamlT, BaldrianP (2008). Spatial variability of enzyme activities and microbial biomass in the upper layers of Quercus petraea forest soil Soil Biology & Biochemistry, 40, 2068-2075. [本文引用: 1]
SongXZ, ChenXF, ZhouGM, JiangH, PengCH (2017). Observed high and persistent carbon uptake by Moso bamboo forests and its response to environmental drivers Agricultural and Forest Meteorology, 247, 467-475. [本文引用: 1]
SoussanaJF, LemaireG (2014). Coupling carbon and nitrogen cycles for environmentally sustainable intensification of grasslands and crop-livestock systems Agriculture, Ecosystems & Environment, 190, 9-17. [本文引用: 1]
VanceED, BrookesPC, JenkinsonDS (1987). An extraction method for measuring soil microbial biomass C Soil Biology & Biochemistry, 19, 703-707. [本文引用: 1]
WanJJ, GuoJF, JiSR, RenWL, YangYS (2016). Effects of dissolved organic matter input on soil CO2 emission and microbial community composition in a subtropical forest Scientia Silvae Sinicae, 52(2), 106-113. [本文引用: 1]
WangJY, SongCC, WangXW, WangLL (2011). Progress in the study of effect of freeze-thaw processes on the organic carbon pool and microorganisms in soils Journal of Glaciology and Geocryology, 33, 442-452. [本文引用: 1]
WangQK, WangSL, HeTX, LiuL, WuJB (2014). Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils Soil Biology & Biochemistry, 71, 13-20. [本文引用: 1]
WangXB, SongDL, LiangGQ, ZhangQ, AiC, ZhouW (2015). Maize biochar addition rate influences soil enzyme activity and microbial community composition in a fluvo-aquic soil Applied Soil Ecology, 96, 265-272. [本文引用: 1]
WeiCC, LiuXF, LinCF, LiXF, LiY, ZhengYX (2018). Response of soil enzyme activities to litter input changes in two secondary Castanopsis carlessii forests in subtropical China Chinese Journal of Plant Ecology, 42, 692-702. [本文引用: 1]
WiederWR, ClevelandCC, TownsendAR (2008). Tropical tree species composition affects the oxidation of dissolved organic matter from litter Biogeochemistry, 88, 127-138. [本文引用: 1]
XiaoW, ChenX, JingX, ZhuB (2018). A meta-analysis of soil extracellular enzyme activities in response to global change Soil Biology & Biochemistry, 123, 21-32. [本文引用: 1]
ZhangCX, NanZB (2010). Research progress of soil microbial biomass in China Pratacultural Science, 27, 50-57. [本文引用: 1]
ZhouJB, ChenZJ, ZhengXF (2005). Soluble organic nitrogen in soil and its roles in the supply and transformation of N Chinese Journal of Soil Science, 36, 244-248. [本文引用: 1]
ZhouY, BouttonTW, WuXB (2018). Soil phosphorus does not keep pace with soil carbon and nitrogen accumulation following woody encroachment Global Change Biology, 24, 1992-2007. DOI:10.1111/gcb.14048URLPMID:29323781 [本文引用: 1] Soil carbon, nitrogen, and phosphorus cycles are strongly interlinked and controlled through biological processes, and the phosphorus cycle is further controlled through geochemical processes. In dryland ecosystems, woody encroachment often modifies soil carbon, nitrogen, and phosphorus stores, although it remains unknown if these three elements change proportionally in response to this vegetation change. We evaluated proportional changes and spatial patterns of soil organic carbon (SOC), total nitrogen (TN), and total phosphorus (TP) concentrations following woody encroachment by taking spatially explicit soil cores to a depth of 1.2 m across a subtropical savanna landscape which has undergone encroachment by Prosopis glandulosa (an N2 fixer) and other woody species during the past century in southern Texas, USA. SOC and TN were coupled with respect to increasing magnitudes and spatial patterns throughout the soil profile following woody encroachment, while TP increased slower than SOC and TN in topmost surface soils (0-5 cm) but faster in subsurface soils (15-120 cm). Spatial patterns of TP strongly resembled those of vegetation cover throughout the soil profile, but differed from those of SOC and TN, especially in subsurface soils. The encroachment of woody species dominated by N2 -fixing trees into this P-limited ecosystem resulted in the accumulation of proportionally less soil P compared to C and N in surface soils; however, proportionally more P accrued in deeper portions of the soil profile beneath woody patches where alkaline soil pH and high carbonate concentrations would favor precipitation of P as relatively insoluble calcium phosphates. This imbalanced relationship highlights that the relative importance of biotic vs. abiotic mechanisms controlling C and N vs. P accumulation following vegetation change may vary with depth. Our findings suggest that efforts to incorporate effects of land cover changes into coupled climate-biogeochemical models should attempt to represent C-N-P imbalances that may arise following vegetation change.
Costimulation of soil glycosidase activity and soil respiration by nitrogen addition 1 2017
... 土壤酶主要由土壤微生物分泌而来, 在土壤有机质(SOM)分解和养分矿化中起着关键的作用(Kotroczó et al., 2014; Jiang et al., 2019).反过来, 土壤碳和养分有效性的变化也会对土壤酶活性产生强烈影响(Chen et al., 2017; 高雨秋等, 2019).已有多项研究表明土壤中溶解有机碳(DOC)的含量与土壤酶活性呈正相关关系(Peterson et al., 2013; 周嘉聪等, 2017).此外, 由于土壤中各元素之间存在耦合关系(Soussana & Lemaire, 2014), 养分获取酶的活性和酶促底物之间的联系还依赖于其他元素含量的高低, 如氮有效性较高会刺激碳获取酶的活性来分解SOM (Sinsabaugh, 2010).根据微生物资源分配理论, 土壤养分含量和酶活性之间也可能呈负相关关系, 即随着养分有效性的增加, 微生物获取该养分的难度降低, 所需的相关获取酶的活性也相应降低(Sinsabaugh, 2010; Fang et al., 2018).由此可见, 土壤酶活性和养分之间关系的研究结果尚未达成共识, 这可能与外源碳和养分输入的差异有关. ...
Minor responses of soil microbial biomass, community structure and enzyme activities to nitrogen and phosphorus addition in three grassland ecosystems 1 2019
Photochemical reactivities of dissolved organic matter (DOM) in a sub-alpine lake revealed by EEM-PARAFAC: an insight into the fate of allochthonous DOM in alpine lakes affected by climate change 1 2016
... 土壤SOC和TN含量的变化对森林生态系统主要功能、组成和发展方向具有重要影响(Zhou et al., 2018).本研究发现, 杉木和木荷叶片DOM输入后SOC和TN含量降低(表3), 但是土壤DOC和DON浓度升高, 这可能是因为外源DOM输入刺激了土壤有机质的分解(Fontaine et al., 2007), 产生了激发效应, 使SOC和TN转化为DOC和DON或以气体的形式释放到外界.相反, 楠木叶片DOM输入后土壤TN含量略有上升, 这可能与所添加的楠木叶片DOM中的DOC含量较低, HIX较高(表1)有关.输入的叶片DOC含量较低, 微生物可利用的碳源相对较少; HIX越高表示DOM成分越复杂(Miano & Senesi, 1992; Du et al., 2016), 因此楠木叶片DOM较难被微生物利用与分解.相关性分析中DOM添加处理下SOC和TN含量与输入的叶片DOM中DOC含量和HIX显著相关(表4)也进一步支持了这一说法. ...
Effects of nitrogen addition on DOM-induced soil priming effects in a subtropical plantation forest and a natural forest 2 2020
... 可溶性有机质(DOM)被定义为能通过0.45 μm筛孔且能溶解于水、酸或碱溶液的不同大小和结构的有机分子混合体(Kalbitz et al., 2000).我国南方地区森林广布, 碳汇量占中国总量的65%以上(Piao et al., 2009), 受季风气候的影响, 该地区降水丰富, 因此森林长期受到淋溶作用, 源自植物叶片的DOM随降水输入到土壤中(Fan et al., 2020), 如Guo等(2005)的研究表明, 热带亚热带地区森林穿透雨中DOC通量范围一般为97-232 kg·hm-2·a-1, 并有研究指出这是SOM中活性碳库的重要来源(Gauthier et al., 2010).同时, 进入土壤中的DOM也为土壤微生物生长和分解过程提供了底物和能量来源(Boddy et al., 2007), 这可能会进一步影响与土壤底物密切相关的获取酶活性.而不同植物来源的DOM化学组成和性质差异较大(纪宇皝等, 2018), 如阔叶树的DOC含量高于针叶树(Wieder et al., 2008).此外, 根据康根丽等(2014)的研究, 米槠(Castanopsis carlesii)林淋溶产出的DOM比杉木(Cunninghamia lanceolata)林更有利于土壤氮的积累, 纪宇皝等(2018)的研究发现S策略树种叶片DOM可能更有利于土壤维持土壤肥力, 而Fan等(2020)的研究中两种土壤氧化酶的活性对米槠和杉木叶片DOM的输入产生了不同响应.因此我们推测不同植物来源的DOM可能会对林下土壤养分循环及酶活性产生不同影响. ...
Microbial mechanisms of carbon priming effects revealed during the interaction of crop residue and nutrient inputs in contrasting soils 1 2018
... 土壤酶主要由土壤微生物分泌而来, 在土壤有机质(SOM)分解和养分矿化中起着关键的作用(Kotroczó et al., 2014; Jiang et al., 2019).反过来, 土壤碳和养分有效性的变化也会对土壤酶活性产生强烈影响(Chen et al., 2017; 高雨秋等, 2019).已有多项研究表明土壤中溶解有机碳(DOC)的含量与土壤酶活性呈正相关关系(Peterson et al., 2013; 周嘉聪等, 2017).此外, 由于土壤中各元素之间存在耦合关系(Soussana & Lemaire, 2014), 养分获取酶的活性和酶促底物之间的联系还依赖于其他元素含量的高低, 如氮有效性较高会刺激碳获取酶的活性来分解SOM (Sinsabaugh, 2010).根据微生物资源分配理论, 土壤养分含量和酶活性之间也可能呈负相关关系, 即随着养分有效性的增加, 微生物获取该养分的难度降低, 所需的相关获取酶的活性也相应降低(Sinsabaugh, 2010; Fang et al., 2018).由此可见, 土壤酶活性和养分之间关系的研究结果尚未达成共识, 这可能与外源碳和养分输入的差异有关. ...
Stability of organic carbon in deep soil layers controlled by fresh carbon supply 1 2007
... 土壤SOC和TN含量的变化对森林生态系统主要功能、组成和发展方向具有重要影响(Zhou et al., 2018).本研究发现, 杉木和木荷叶片DOM输入后SOC和TN含量降低(表3), 但是土壤DOC和DON浓度升高, 这可能是因为外源DOM输入刺激了土壤有机质的分解(Fontaine et al., 2007), 产生了激发效应, 使SOC和TN转化为DOC和DON或以气体的形式释放到外界.相反, 楠木叶片DOM输入后土壤TN含量略有上升, 这可能与所添加的楠木叶片DOM中的DOC含量较低, HIX较高(表1)有关.输入的叶片DOC含量较低, 微生物可利用的碳源相对较少; HIX越高表示DOM成分越复杂(Miano & Senesi, 1992; Du et al., 2016), 因此楠木叶片DOM较难被微生物利用与分解.相关性分析中DOM添加处理下SOC和TN含量与输入的叶片DOM中DOC含量和HIX显著相关(表4)也进一步支持了这一说法. ...
亚热带人工林下植被根际土壤酶化学计量特征 1 2019
... 土壤酶主要由土壤微生物分泌而来, 在土壤有机质(SOM)分解和养分矿化中起着关键的作用(Kotroczó et al., 2014; Jiang et al., 2019).反过来, 土壤碳和养分有效性的变化也会对土壤酶活性产生强烈影响(Chen et al., 2017; 高雨秋等, 2019).已有多项研究表明土壤中溶解有机碳(DOC)的含量与土壤酶活性呈正相关关系(Peterson et al., 2013; 周嘉聪等, 2017).此外, 由于土壤中各元素之间存在耦合关系(Soussana & Lemaire, 2014), 养分获取酶的活性和酶促底物之间的联系还依赖于其他元素含量的高低, 如氮有效性较高会刺激碳获取酶的活性来分解SOM (Sinsabaugh, 2010).根据微生物资源分配理论, 土壤养分含量和酶活性之间也可能呈负相关关系, 即随着养分有效性的增加, 微生物获取该养分的难度降低, 所需的相关获取酶的活性也相应降低(Sinsabaugh, 2010; Fang et al., 2018).由此可见, 土壤酶活性和养分之间关系的研究结果尚未达成共识, 这可能与外源碳和养分输入的差异有关. ...
亚热带人工林下植被根际土壤酶化学计量特征 1 2019
... 土壤酶主要由土壤微生物分泌而来, 在土壤有机质(SOM)分解和养分矿化中起着关键的作用(Kotroczó et al., 2014; Jiang et al., 2019).反过来, 土壤碳和养分有效性的变化也会对土壤酶活性产生强烈影响(Chen et al., 2017; 高雨秋等, 2019).已有多项研究表明土壤中溶解有机碳(DOC)的含量与土壤酶活性呈正相关关系(Peterson et al., 2013; 周嘉聪等, 2017).此外, 由于土壤中各元素之间存在耦合关系(Soussana & Lemaire, 2014), 养分获取酶的活性和酶促底物之间的联系还依赖于其他元素含量的高低, 如氮有效性较高会刺激碳获取酶的活性来分解SOM (Sinsabaugh, 2010).根据微生物资源分配理论, 土壤养分含量和酶活性之间也可能呈负相关关系, 即随着养分有效性的增加, 微生物获取该养分的难度降低, 所需的相关获取酶的活性也相应降低(Sinsabaugh, 2010; Fang et al., 2018).由此可见, 土壤酶活性和养分之间关系的研究结果尚未达成共识, 这可能与外源碳和养分输入的差异有关. ...
Dynamics of the water extractable organic carbon pool during mineralisation in soils from a Douglas fir plantation and an oak-beech forest—An incubation experiment 1 2010
... 可溶性有机质(DOM)被定义为能通过0.45 μm筛孔且能溶解于水、酸或碱溶液的不同大小和结构的有机分子混合体(Kalbitz et al., 2000).我国南方地区森林广布, 碳汇量占中国总量的65%以上(Piao et al., 2009), 受季风气候的影响, 该地区降水丰富, 因此森林长期受到淋溶作用, 源自植物叶片的DOM随降水输入到土壤中(Fan et al., 2020), 如Guo等(2005)的研究表明, 热带亚热带地区森林穿透雨中DOC通量范围一般为97-232 kg·hm-2·a-1, 并有研究指出这是SOM中活性碳库的重要来源(Gauthier et al., 2010).同时, 进入土壤中的DOM也为土壤微生物生长和分解过程提供了底物和能量来源(Boddy et al., 2007), 这可能会进一步影响与土壤底物密切相关的获取酶活性.而不同植物来源的DOM化学组成和性质差异较大(纪宇皝等, 2018), 如阔叶树的DOC含量高于针叶树(Wieder et al., 2008).此外, 根据康根丽等(2014)的研究, 米槠(Castanopsis carlesii)林淋溶产出的DOM比杉木(Cunninghamia lanceolata)林更有利于土壤氮的积累, 纪宇皝等(2018)的研究发现S策略树种叶片DOM可能更有利于土壤维持土壤肥力, 而Fan等(2020)的研究中两种土壤氧化酶的活性对米槠和杉木叶片DOM的输入产生了不同响应.因此我们推测不同植物来源的DOM可能会对林下土壤养分循环及酶活性产生不同影响. ...
Dissolved organic carbon and nitrogen in precipitation, throughfall and stemflow from Schima superba and Cunninghamia lanceolata plantations in subtropical China 1 2005
... 可溶性有机质(DOM)被定义为能通过0.45 μm筛孔且能溶解于水、酸或碱溶液的不同大小和结构的有机分子混合体(Kalbitz et al., 2000).我国南方地区森林广布, 碳汇量占中国总量的65%以上(Piao et al., 2009), 受季风气候的影响, 该地区降水丰富, 因此森林长期受到淋溶作用, 源自植物叶片的DOM随降水输入到土壤中(Fan et al., 2020), 如Guo等(2005)的研究表明, 热带亚热带地区森林穿透雨中DOC通量范围一般为97-232 kg·hm-2·a-1, 并有研究指出这是SOM中活性碳库的重要来源(Gauthier et al., 2010).同时, 进入土壤中的DOM也为土壤微生物生长和分解过程提供了底物和能量来源(Boddy et al., 2007), 这可能会进一步影响与土壤底物密切相关的获取酶活性.而不同植物来源的DOM化学组成和性质差异较大(纪宇皝等, 2018), 如阔叶树的DOC含量高于针叶树(Wieder et al., 2008).此外, 根据康根丽等(2014)的研究, 米槠(Castanopsis carlesii)林淋溶产出的DOM比杉木(Cunninghamia lanceolata)林更有利于土壤氮的积累, 纪宇皝等(2018)的研究发现S策略树种叶片DOM可能更有利于土壤维持土壤肥力, 而Fan等(2020)的研究中两种土壤氧化酶的活性对米槠和杉木叶片DOM的输入产生了不同响应.因此我们推测不同植物来源的DOM可能会对林下土壤养分循环及酶活性产生不同影响. ...
Soil microbial biomass C, N and ninhydrin-N in aerobic and anaerobic soils measured by the fumigation-extraction method 1 1991
... 土壤微生物生物量: 微生物生物量碳(MBC)和微生物生物量氮(MBN)采用氯仿熏蒸法浸提(Vance et al., 1987; Inubushi et al., 1991), 分别称取5 g鲜土, 做熏蒸和未熏蒸处理.用20 mL 0.5 mol·L-1 K2SO4溶液浸提, 振荡、离心、过滤, 将浸提液收集到样品杯, 其中, MBC浸提液用总有机碳分析仪测定, MBN浸提液采用连续流动分析仪测定. ...
亚热带地区竞争型和忍耐型树种叶片可溶性有机质数量及光谱学特征 2 2018
... 可溶性有机质(DOM)被定义为能通过0.45 μm筛孔且能溶解于水、酸或碱溶液的不同大小和结构的有机分子混合体(Kalbitz et al., 2000).我国南方地区森林广布, 碳汇量占中国总量的65%以上(Piao et al., 2009), 受季风气候的影响, 该地区降水丰富, 因此森林长期受到淋溶作用, 源自植物叶片的DOM随降水输入到土壤中(Fan et al., 2020), 如Guo等(2005)的研究表明, 热带亚热带地区森林穿透雨中DOC通量范围一般为97-232 kg·hm-2·a-1, 并有研究指出这是SOM中活性碳库的重要来源(Gauthier et al., 2010).同时, 进入土壤中的DOM也为土壤微生物生长和分解过程提供了底物和能量来源(Boddy et al., 2007), 这可能会进一步影响与土壤底物密切相关的获取酶活性.而不同植物来源的DOM化学组成和性质差异较大(纪宇皝等, 2018), 如阔叶树的DOC含量高于针叶树(Wieder et al., 2008).此外, 根据康根丽等(2014)的研究, 米槠(Castanopsis carlesii)林淋溶产出的DOM比杉木(Cunninghamia lanceolata)林更有利于土壤氮的积累, 纪宇皝等(2018)的研究发现S策略树种叶片DOM可能更有利于土壤维持土壤肥力, 而Fan等(2020)的研究中两种土壤氧化酶的活性对米槠和杉木叶片DOM的输入产生了不同响应.因此我们推测不同植物来源的DOM可能会对林下土壤养分循环及酶活性产生不同影响. ...
Revealing microbial processes and nutrient limitation in soil through ecoenzymatic stoichiometry and glomalin-related soil proteins in a retreating glacier forefield 1 2019
... 土壤酶主要由土壤微生物分泌而来, 在土壤有机质(SOM)分解和养分矿化中起着关键的作用(Kotroczó et al., 2014; Jiang et al., 2019).反过来, 土壤碳和养分有效性的变化也会对土壤酶活性产生强烈影响(Chen et al., 2017; 高雨秋等, 2019).已有多项研究表明土壤中溶解有机碳(DOC)的含量与土壤酶活性呈正相关关系(Peterson et al., 2013; 周嘉聪等, 2017).此外, 由于土壤中各元素之间存在耦合关系(Soussana & Lemaire, 2014), 养分获取酶的活性和酶促底物之间的联系还依赖于其他元素含量的高低, 如氮有效性较高会刺激碳获取酶的活性来分解SOM (Sinsabaugh, 2010).根据微生物资源分配理论, 土壤养分含量和酶活性之间也可能呈负相关关系, 即随着养分有效性的增加, 微生物获取该养分的难度降低, 所需的相关获取酶的活性也相应降低(Sinsabaugh, 2010; Fang et al., 2018).由此可见, 土壤酶活性和养分之间关系的研究结果尚未达成共识, 这可能与外源碳和养分输入的差异有关. ...
Nitrogen deposition has minor effect on soil extracellular enzyme activities in six Chinese forests 1 2017
... 由于土壤酶活性控制着土壤中的碳和养分循环, 因此近年来土壤酶活性受到了广泛的关注(Luo et al., 2018; Zheng et al., 2018).一般来说, 酶的生产是能源密集型的; 土壤微生物在缺乏可利用资源的情况下只能以牺牲微生物的生长和代谢为代价生产酶, 因此, 资源的有效性往往与获取酶的活性呈负相关关系(Jing et al., 2017; Zhu et al., 2018).但本研究中, 叶片DOM输入后胞外酶(除PPO外)活性提高(图2), 这主要是因为3种植物来源DOM输入后土壤DOC和DON含量的升高不同程度地为土壤微生物提供了可利用碳源和养分, 进而刺激了微生物的活性, 产生更多的胞外酶(除PPO外).在土壤有机质分解过程中, 含量较高的纤维素被优先分解, 因此纤维素酶活性较高, 在纤维素分解过程中, 葡萄糖二聚体和一些纤维素低聚糖被β-葡萄糖苷酶水解为葡萄糖(?najdr et al., 2008), 因此本研究中叶片DOM添加后CBH与βG这两种酶的活性均显著上升.而不同植物来源DOM添加处理中, βG和CBH活性均表现为CL > SL > PL, 这和叶片DOC含量呈显著正相关关系, 与叶片DOM的HIX呈显著负相关关系(表4), 即随着所添加的叶片DOM中DOC含量的增多与HIX的降低, 两种碳循环相关的水解酶(βG和CBH)活性随之上升.这表明不同植物来源DOM的质量和数量会影响这两种水解酶活性, 可能导致森林土壤碳矿化的差异.杉木叶片DOM中较高的DOC含量为微生物带来更多的能量来源, 较低的HIX表明杉木叶片DOM中易分解成分高于木荷和楠木, 因此CL处理两种碳循环相关的水解酶(CBH和βG)活性高于SL和PL, 说明杉木林土壤碳的周转速率可能大于木荷和楠木. ...
Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil 1 2006
 ,1,2,*
,1,2,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT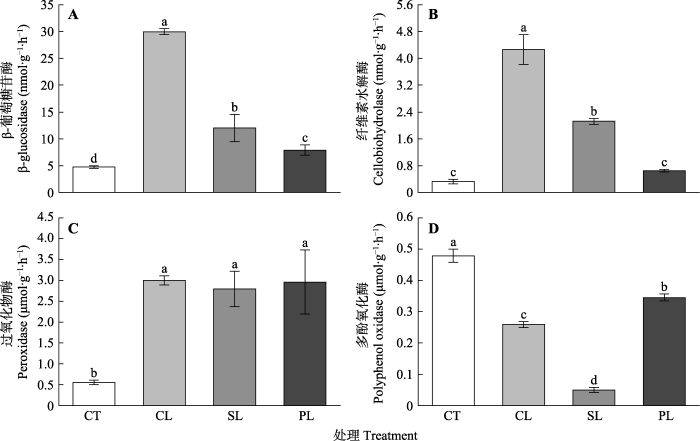 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT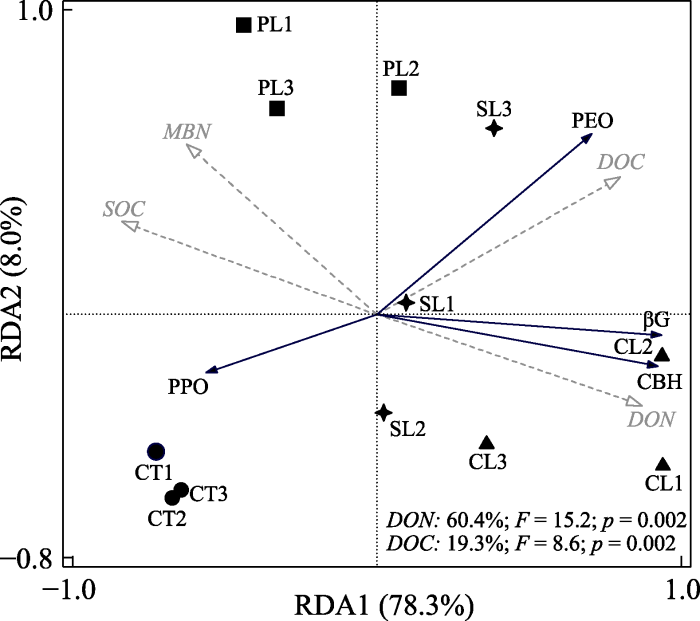 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}