Effects of nitrogen addition amount and frequency on soil respiration and its components in a temperate semiarid grassland
Ze YANG1,2, Xing-Ru TAN1,2, Cui-Hai YOU1,2, Yan-Bing WANG1,2, Jun-Jie YANG1, Xing-Guo HAN1,2, Shi-Ping CHEN,,1,2,*1State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2University of Chinese Academy of Sciences, Beijing 100049, China
Abstract Aims Increasing global nitrogen (N) deposition has exerted significant influences on productivity and carbon cycle of terrestrial ecosystems. More than 90% of the carbon in grasslands is stored in the soil, therefore any changes in soil total respiration (Rs) might have a vital impact on the carbon balance and the stability of soil carbon pool of grassland ecosystems. Most of our understanding about the responses of Rs to N deposition was based on N deposition manipulative experiments with short-term (<5 years) and low frequency (1-2 times per year) N addition treatments. It is still unclear how the long term N addition and different N addition frequency will affect Rs and its components in semiarid grasslands. Methods Our study is based on a long term N addition manipulative experiment platform conducted in a typical temperate semiarid steppe, Nei Mongol. The experimental treatment consisted of six N addition amounts and two N addition frequencies. N addition treatments began at 2008. Soil respiration and its components were measured every two weeks during the growing season in 2018 and 2019. Important findings 1) Rs significantly decreased with increasing N addition amount. The negative impact of N addition on Rs was mainly resulted from the inhibition of heterotrophic respiration (Rh). 2) No significant differences were observed in responses of Rs and its components to low and high frequency N addition treatments. 3) Soil acidification caused by long term N addition inhibited soil microbial activity and changed soil microbial community composition, consequently decreased Rs and Rh. Our results suggested that the negative effect of N addition on soil carbon release still lasted after a decade of N addition treatment. In particular, the decrease of Rh would enhance the stability of soil carbon pool. No significant differences in the two N addition frequency treatments indicated that the potential impacts caused by simulated N addition with different frequencies would be diminished with prolonged treatment period. Therefore, the results of long-term (>10 years) simulated N addition experiments can provide reliable references for evaluating the responses of natural ecosystems to atmospheric N deposition. Keywords:long-term nitrogen addition;nitrogen addition frequency;soil respiration;autotrophic respiration;heterotrophic respiration
PDF (2225KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 杨泽, 嘎玛达尔基, 谭星儒, 游翠海, 王彦兵, 杨俊杰, 韩兴国, 陈世苹. 氮添加量和施氮频率对温带半干旱草原土壤呼吸及组分的影响. 植物生态学报, 2020, 44(10): 1059-1072. DOI: 10.17521/cjpe.2020.0171 YANG Ze, TAN Xing-Ru, YOU Cui-Hai, WANG Yan-Bing, YANG Jun-Jie, HAN Xing-Guo, CHEN Shi-Ping. Effects of nitrogen addition amount and frequency on soil respiration and its components in a temperate semiarid grassland. Chinese Journal of Plant Ecology, 2020, 44(10): 1059-1072. DOI: 10.17521/cjpe.2020.0171
土壤微生物生物量(MB)和土壤微生物功能群组成采用磷脂脂肪酸(PLFA)方法测定(Frosteg?rd et al., 2011)。根据细菌的特异性脂肪酸(i15:0、i16:0、i17:0、18:1ω7、cy17:0和cy19:0)和真菌的特异性脂肪酸(18:2ω6, 9)的含量代表土壤中细菌和真菌的生物量(Wang et al., 2019c), 并计算真菌/细菌比(F/B)。
Table 1 表1 表1测量年份(Y)、氮添加量(N)和施氮频率(NF)及其交互作用对土壤温度、土壤水分含量、土壤总呼吸速率及组分(异养和自养呼吸)和异养呼吸比值的影响 Table 1Results of the effects of year (Y), nitrogen addition amount (N), nitrogen addition frequency (NF) and their interactions on soil temperature (ST), soil water content (SWC), soil total respiration rate (Rs) and its components (Rh and Ra) and Rh/Rs ratio
新窗口打开|下载原图ZIP|生成PPT 图12018和2019年氮添加量和施氮频率对表层(0-10 cm)土壤温度(ST)(A、C)和土壤水分含量(SWC)(B、D)的影响。图中数据为平均值±标准误差。图中不同颜色的图标分别代表0、2、5、10、20、50 g·m-2·a-1氮添加处理。图中给出了氮添加量(N)、施氮频率(NF)及其交互作用(N × NF)对土壤温度和含水量的差异显著性检验结果(***, p < 0.001; *, p < 0.05; ns, p > 0.05)。图中的小写字母代表不同氮添加量处理间的差异显著性, 相同字母表示无显著差异(p > 0.05), 不同字母表示具有显著差异(p < 0.05)。
Fig. 1Changes in soil temperature (ST)(A, C) and soil water content (SWC)(B, D) in 0-10 cm surface soil layer under different N addition amount and frequency treatments in 2018 and 2019. Data are mean ± SE. The different color icons in the figure represent N addition treatments with 0, 2, 5, 10, 20, 50 g·m-2·a-1, respectively. The ANOVA results were shown in the figure to address the significance of effects of N addition amount (N), frequency (NF) and their interactions (N × NF) on the parameters (***, p < 0.001; *, p < 0.05; ns, p > 0.05). The lowercase letters in the figure represent the significance among different nitrogen addition treatments. The same letter means no significance (p > 0.05), while different letters mean significant differences (p < 0.05). HF, N addition with high frequency; LF, N addition with low frequency.
新窗口打开|下载原图ZIP|生成PPT 图22018和2019年不同氮添加量和施氮频率处理下土壤呼吸速率(Rs)及其异养(Rh)和自养(Ra)组分的季节和年际动态(平均值±标准误差)。图中不同颜色的图标分别代表0、2、5、10、20、50 g·m-2·a-1氮添加处理。图中给出了氮添加量(N)、施氮频率(NF)及其交互作用(N × NF)对土壤呼吸及其组分的差异显著性检验结果(***, p < 0.001; ns, p > 0.05)。
Fig. 2Inter- and intra-annual variations in soil respiration rate (Rs) and its heterotrophic (Rh) and autotrophic (Ra) components under different N addition amount and frequency treatments in 2018 and 2019 (mean ± SE). The different color icons in the figure represent N addition treatments with 0, 2, 5, 10, 20, 50 g·m-2·a-1, respectively. The ANOVA results were shown in the figure to address the significance of effects of N addition amount (N), frequency (NF) and their interactions (N × NF) on the parameters (***, p < 0.001; ns, p > 0.05). HF, N addition with high frequency; LF, N addition with low frequency.
图3
新窗口打开|下载原图ZIP|生成PPT 图32018和2019年氮添加量和施氮频率对土壤呼吸速率(Rs)及其异养(Rh)和自养(Ra)组分季节平均值的影响。图中数据为平均值±标准误差。图中不同颜色的图标分别代表0、2、5、10、20、50 g·m-2·a-1氮添加处理。图中给出了氮添加量(N)、施氮频率(NF)及其交互作用(N × NF)对土壤呼吸及其组分的差异显著性检验结果(***, p < 0.001; **, p < 0.01; *, p < 0.05; ns, p > 0.05)。图中的小写字母代表不同氮添加量处理间的差异显著性, 相同字母表示无显著差异(p > 0.05), 不同字母表示具有显著差异(p < 0.05)。
Fig. 3Seasonal mean values of soil respiration rate (Rs) and its heterotrophic (Rh) and autotrophic (Ra) components under different N addition amount and frequency treatments in 2018 and 2019. Data are mean ± SE. The different color icons in the figure represent N addition treatments with 0, 2, 5, 10, 20, 50 g·m-2·a-1, respectively. The ANOVA results were shown in the figure to address the significance of effects of N addition amount (N), frequency (NF) and their interactions (N × NF) on the parameters (***, p < 0.001; **, p < 0.01; *, p < 0.05; ns, p > 0.05). The lowercase letters in the figure represent the significance among different nitrogen addition treatments. The same letter means no significance (p > 0.05), while different letters mean significant differences (p < 0.05). HF, N addition with high frequency; LF, N addition with low frequency.
Fig. 4Relationships between soil respiration rate (Rs) and its heterotrophic (Rh) and autotrophic (Ra) component with soil temperature (ST), soil water content (SWC) and soil pH value. Different color icons in the figure represent N addition treatments with 0, 2, 5, 10, 20, 50 g·m-2·a-1, respectively. The circle sign and black line represent the low frequency N addition (LF) treatment, and the triangle sign and red line indicate the high frequency N addition (HF) treatment. The solid line represents marginally significant relation (p < 0.1) and the dash line represents insignificant relation (p > 0.1).
新窗口打开|下载原图ZIP|生成PPT 图5不同氮添加量和施氮频率对土壤呼吸速率(Rs)及其异养呼吸组分(Rh)温度敏感性(Q10)(A、B)和水分敏感性(C、D)的影响。图中数据为平均值±标准误差。图中不同颜色的图标分别代表0、2、5、10、20和50 g·m-2·a-1氮添加处理。图中给出了氮添加量(N)、施氮频率(NF)及其交互作用(N × NF)对温度和水分敏感性影响的显著性检验结果(***, p < 0.001; **, p < 0.01; *, p < 0.05; ns, p > 0.05)。图中的小写字母代表不同氮添加量处理间的差异显著性, 相同字母表示无显著差异(p > 0.05), 不同字母表示具显著差异(p < 0.05)。
Fig. 5Changes in temperature (Q10, A, B) and water (C, D) sensitivities of soil respiration (Rs) and its heterotrophic component (Rh) under different nitrogen addition amount and frequency treatments. Data are mean ± SE. The different color icons in the figure represent N addition treatments with 0, 2, 5, 10, 20, 50 g·m-2·a-1, respectively. The ANOVA results were shown in the figure to address the significance of effects of N addition amount (N), frequency (NF) and their interactions (N × NF) on the parameters (***, p < 0.001; **, p < 0.01; *, p < 0.05; ns, p > 0.05). The lowercase letters in the figure represent the significance among different nitrogen addition treatments. The same letter means no significance (p > 0.05), while different letters mean significant differences (p < 0.05). HF, N addition with high frequency; LF, N addition with low frequency.
Fig. 6Relationships between soil total respiration and its components with aboveground net primary productivity (ANPP), soil microbial biomass (MB) and fungal biomass/bacterial biomass ratio (F/B). The different color icons in the figure represent N addition treatments with 0, 2, 5, 10, 20, 50 g·m-2·a-1, respectively. The circle sign and black line represent the low frequency N addition (LF) treatment, and the triangle sign and red line indicate the high frequency N addition (HF) treatment. The solid line represents marginally significant relation (p < 0.1) and the dash line represents insignificant relation (p > 0.1).
Rs对氮添加响应的强度和方向取决于Rh和Ra两个组分的响应, 本研究中氮添加显著降低了Rh, 对Ra并无显著影响。氮添加并没有影响Rh对Rs的贡献比例, Rh平均占Rs的62%, 这与本区域其他研究结果(Zhang et al., 2016a, 2019)一致。大部分陆地生态系统的生产力是受氮限制的, 通常氮添加会通过提高土壤氮可利用性而促进植物生长, 增加植物自养呼吸(Chen et al., 2015b)。但土壤氮限制缓解的同时也会改变植物的地上地下碳分配策略, 减少植物对地下根系的碳分配比例, 该实验平台的研究表明在氮添加处理的第6和第7年, 氮添加使得ANPP增加了45%-53%, 但导致地下净初级生产力(BNPP)平均下降了40%-47% (Wang et al., 2019a)。这可能是本研究中Ra对氮添加处理缺乏响应的原因。一项全球氮添加实验的meta分析表明, 氮添加会显著降低根系生物量和周转速率, 但对根系现存量没有影响(Peng et al., 2017), 由于Ra主要来自于现存根系呼吸, 这可以进一步解释为什么本研究中Ra对氮添加处理没有显著响应。
本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3)。在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现。本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013)。Rh对氮添加的负响应主要有以下几个原因。首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010)。根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源。地下碳输入量的下降会显著降低土壤微生物有机碳降解速率。本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a)。因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5)。第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能。多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017)。这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019)。氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020)。本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%。氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I)。而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5)。第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境。本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1)。但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降。与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高。此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关。降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019)。
3.2 施氮频率对土壤呼吸速率及其组分的影响
大气氮沉降是持续而缓慢的氮添加过程, 而大多数模拟氮沉降实验研究是通过生长季少次脉冲式的氮添加处理进行的(Bai et al., 2015; Fang et al., 2017; Chen et al., 2018; Wang et al., 2019c), 不同的氮添加模式和频次会影响生态系统结构和功能对氮添加的响应(Tu et al., 2013; Zhang et al., 2014)。本平台已有的研究结果表明, 在氮添加处理早期, 不同施氮频率对植物地上生产力并没有显著影响, 而对物种多样性、地下生产力和土壤微生物群落的影响是显著的(Zhang et al., 2016b; Wang et al., 2019a; Cao et al., 2020)。Zhang等(2016b)对氮添加处理前5年的研究发现随着氮添加量的增加, 植物物种多样性显著降低, 而与HF (每年12次)相比, LF (每年2次)处理导致物种多样性丧失的速率更快, 这主要是因为LF处理下较高的氨态氮浓度限制了新物种的定植。Wang等(2019b)在氮添加处理的第5-7年的研究表明, 虽然不同施氮频率并未显著影响植被ANPP, 但LF显著降低了BNPP, 特别是在低氮添加量处理下。Cao等(2020)对施氮后第6和7年的研究表明, HF会显著减弱氮添加处理对植物和土壤细菌多样性的负作用。本研究在氮添加处理第11和12年进行测定, 我们并未发现不同施氮频率对土壤呼吸及其组分影响有显著差别, 而同期两种施氮频率下植物生产力、土壤微生物生物量、土壤细菌和真菌生物量以及F/B均没有显著的差异(附录I, 附录II)。这表明随着氮添加处理时间的延长, 不同施氮频率对植物和微生物群落生产力和结构的影响逐渐减弱。因为随着土壤中可利用氮的不断增加, 造成土壤酸化和有毒离子的积累, 生态系统逐渐呈现出氮饱和的状态(Tian et al., 2016), 进而逐步削弱了不同施氮频率对生态系统影响的差别。本研究的HF处理虽然尽可能地提高了氮添加频次, 但与缓慢持续的大气氮沉降过程仍存在较大差别, 使得我们在研究生态系统结构和功能对大气氮沉降的响应过程中仍存在很大不确定性。随着氮添加手段的改进, 如采用具亲水性包衣的缓释尿素等, 可以更精确地模拟大气氮沉降特别是湿沉降的过程, 为准确模拟大气氮沉降对生态系统碳氮循环过程的影响提供了可能。
Supplement I Effects of different nitrogen addition amount and frequency treatments on aboveground net primary productivity, soil pH, microbial biomass, bacterial biomass, fungal biomass and fungal biomass/bacterial biomass ratio
Supplement II Effects of nitrogen addition amount, nitrogen addition frequency and their interactions on aboveground net primary productivity, soil pH, microbial biomass, bacterial biomass, fungal biomass and fungal biomass/bacterial biomass ratio
AllisonSD, CzimczikCI, TresederKK (2008). Microbial activity and soil respiration under nitrogen addition in Alaskan boreal forest Global Change Biology, 14, 1156-1168. [本文引用: 1]
ArchMillerA, SamuelsonL (2016). Partitioning longleaf pine soil respiration into its heterotrophic and autotrophic components through root exclusion Forests, 7, 1-13. [本文引用: 1]
BaiTS, WangP, HallSJ, WangFW, YeCL, LiZ, LiSJ, ZhouLY, QiuYP, GuoJX, GuoH, WangY, HuSJ (2020). Interactive global change factors mitigate soil aggregation and carbon change in a semi-arid grassland Global Change Biology, 26, 5320-5332. [本文引用: 1]
BaiWM, GuoDL, TianQY, LiuNN, ChengWX, LiLH, ZhangWH (2015). Differential responses of grasses and forbs led to marked reduction in below-ground productivity in temperate steppe following chronic N deposition Journal of Ecology, 103, 1570-1579. [本文引用: 2]
BaiYF, WuJG, ClarkCM, NaeemS, PanQM, HuangJH, ZhangLX, HanXG (2010). Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from Inner Mongolia grasslands Global Change Biology, 16, 358-372. [本文引用: 3]
Bond-LambertyB, ThomsonA (2010). A global database of soil respiration data Biogeosciences, 7, 1915-1926. [本文引用: 1]
Bond-LambertyB, WangC, GowerST (2004). A global relationship between the heterotrophic and autotrophic components of soil respiration? Global Change Biology, 10, 1756-1766. [本文引用: 1]
BowdenRD, DavidsonE, SavageK, ArabiaC, SteudlerP (2004). Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest Forest Ecology and Management, 196, 43-56. [本文引用: 1]
BowdenRD, WurzbacherSJ, WashkoSE, WindL, RiceAM, CobleAE, BaldaufN, JohnsonB, WangJJ, SimpsonM, LajthaK (2019). Long-term nitrogen addition decreases organic matter decomposition and increases forest soil carbon Soil Science Society of America Journal, 83, 82-95. [本文引用: 1]
CaoJR, PangS, WangQB, WilliamsMA, JiaX, DunSS, YangJJ, ZhangYH, WangJ, LüXT, HuYC, LiLH, LiYC, HanXG (2020). Plant-bacteria-soil response to frequency of simulated nitrogen deposition has implications for global ecosystem change Functional Ecology, 34, 723-734. [本文引用: 3]
ChenDM, LanZC, HuSJ, BaiYF (2015a). Effects of nitrogen enrichment on belowground communities in grassland: relative role of soil nitrogen availability vs. soil acidification Soil Biology & Biochemistry, 89, 99-108. [本文引用: 1]
ChenDM, LiJJ, LanZC, HuSJ, BaiYF (2016). Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment Functional Ecology, 30, 658-669. [本文引用: 1]
ChenDM, XingW, LanZC, SaleemM, WuYQQG, HuSJ, BaiYF (2019). Direct and indirect effects of nitrogen enrichment on soil organisms and carbon and nitrogen mineralization in a semi-arid grassland Functional Ecology, 33, 175-187. [本文引用: 1]
ChenH, LiDJ, GurmesaGA, YuGR, LiLH, ZhangW, FangHJ, MoJM (2015b). Effects of nitrogen deposition on carbon cycle in terrestrial ecosystems of China: a meta- analysis Environmental Pollution, 206, 352-360. [本文引用: 1]
ClarkCM, TilmanD (2008). Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands Nature, 451, 712-715. [本文引用: 1]
CoxPM, BettsRA, JonesCD, SpallSA, TotterdellIJ (2000). Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model Nature, 408, 184-187. [本文引用: 1]
CraineJM, WedinDA, ReichPB (2001). The response of soil CO2 flux to changes in atmospheric CO2, nitrogen supply and plant diversity Global Change Biology, 7, 945-953. [本文引用: 1]
FangC, LiFM, PeiJY, RenaJ, GongYH, YuanZG, KeWB, ZhengY, BaiXK, YeJS (2018). Impacts of warming and nitrogen addition on soil autotrophic and heterotrophic respiration in a semi-arid environment Agricultural and Forest Meteorology, 248, 449-457. [本文引用: 1]
FangC, YeJS, GongYH, PeiJY, YuanZQ, XieC, ZhuYS, YuYY (2017). Seasonal responses of soil respiration to warming and nitrogen addition in a semi-arid alfalfa- pasture of the Loess Plateau, China Science of the Total Environment, 590, 729-738. [本文引用: 2]
FornaraDA, TilmanD (2012). Soil carbon sequestration in prairie grasslands increased by chronic nitrogen addition Ecology, 93, 2030-2036. [本文引用: 1]
Frosteg?rd?, TunlidA, B??thE (2011). Use the misuse of PLFA measurements in soils Soil Biology & Biochemistry, 43, 1621-1625. [本文引用: 1]
FuZ, NiuSL, DukesJS (2015). What have we learned from global change manipulative experiments in China? A meta-analysis Scientific reports, 5, 12344. DOI: 10.1038/srep12344. [本文引用: 2]
GallowayJN, TownsendAR, Jan WillemE, MateeteB, ZucongC, FreneyJR, MartinelliLA, SeitzingerSP, SuttonMA (2008). Transformation of the nitrogen cycle: recent trends, questions, and potential solutions Science, 320, 889-892. [本文引用: 2]
GieseM, BrueckH, GaoY, LinS, SteffensM, K?gelknabnerI, GlindemannT, SusenbethA, TaubeF, ButterbachbahlK (2013). Data from: N balance and cycling of Inner Mongolia typical steppe—A comprehensive case study of grazing effects Ecological Monographs, 83, 195-219. [本文引用: 2]
HansonPJ, EdwardsNT, GartenCT, AndrewsJA (2000). Separating root and soil microbial contributions to soil respiration—A review of methods and observations Biogeochemistry, 48, 115-146. [本文引用: 1]
HasselquistNJ, MetcalfeDB, H?gbergP (2012). Contrasting effects of low and high nitrogen additions on soil CO2 flux components and ectomycorrhizal fungal sporocarp production in a boreal forest Global Change Biology, 18, 3596-3605. [本文引用: 1]
JanssensIA, DielemanW, LuyssaertS, SubkeJA, ReichsteinM, CeulemansR, CiaisP, DolmanAJ, GraceJ, MatteucciG (2010). Reduction of forest soil respiration in response to nitrogen deposition Nature Geoscience, 3, 315-322. [本文引用: 5]
JiaXX, ShaoMA, WeiXR (2012). Responses of soil respiration to N addition, burning and clipping in temperate semiarid grassland in northern China Agricultural and Forest Meteorology, 166, 32-40. [本文引用: 2]
LeeKH, JoseS (2003). Soil respiration, fine root production, and microbial biomass in cottonwood and loblolly pine plantations along a nitrogen fertilization gradient Forest Ecology and Management, 185, 263-273. [本文引用: 1]
LiYL, HongM, BaiWM, HanGD, WangHM, ZhouM (2015). The responses of soil respiration to water and nitrogen in Stipa breviflora steppe Acta Ecologica Sinica, 35, 1727-1733. [本文引用: 1]
LiangLZ, ChenF, HanHR, ZhangYR, ZhuJ, NiuSK (2019). Pathways regulating decreased soil respiration with nitrogen addition in a subtropical forest in China Water Air and Soil Pollution, 230, 91-100. [本文引用: 1]
LiuW, LuXT, XuWF, ShiHQ, HouLY, LiLH, YuanWP (2018). Effects of water and nitrogen addition on ecosystem respiration across three types of steppe: the role of plant and microbial biomass Science of the Total Environment, 619, 103-111. [本文引用: 1]
LovettGM, ColeJJ, PaceML (2006). Is net ecosystem production equal to ecosystem carbon accumulation? Ecosystems, 9, 152-155. [本文引用: 1]
LucaB, ChrisF, TimothyJ, HaKanR, JuulL, NathalieF, TimE, RenatoG, MichalH, TomásH (2006). Atmospheric nitrogen deposition promotes carbon loss from peat bogs Proceedings of the National Academy of Sciences of the United States of America, 103, 19386-19389. [本文引用: 1]
LuoQP, GongJR, ZhaiZW, PanY, LiuM, XuS, WangYH, YangLL, BaoyinTT (2016). The responses of soil respiration to nitrogen addition in a temperate grassland in northern China Science of the Total Environment, 569, 1466-1477. [本文引用: 2]
MaSY, VerheyenK, PropsR, WasofS, VanhellemontM, BoeckxP, BoonN, de FrenneP (2018). Plant and soil microbe responses to light, warming and nitrogen addition in a temperate forest Functional Ecology, 32, 1293-1303. [本文引用: 1]
MasciandaroG, MacciC, PeruzziE, DoniS (2018). Soil carbon in the world: ecosystem services linked to soil carbon in forest and agricultural soilsGarcia C, Nannipieri P, Hernandez T. Future of Soil Carbon: Its Conservation and Formation. Academic Press, London. 1-38. [本文引用: 1]
MoJM, ZhangW, ZhuWX, GundersenP, FangY, LiD, WangH (2008). Nitrogen addition reduces soil respiration in a mature tropical forest in southern China Global Change Biology, 14, 403-412. [本文引用: 2]
NiuSL, WuMY, HanY, XiaJY, LiLH, WanSQ (2008). Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe New Phytologist, 177, 209-219. [本文引用: 1]
NiuSL, WuMY, HanY, XiaJY, ZhangZ, YangHJ, WanSQ (2010). Nitrogen effects on net ecosystem carbon exchange in a temperate steppe Global Change Biology, 16, 144-155. [本文引用: 1]
PengYF, GuoDL, YangYH (2017). Global patterns of root dynamics under nitrogen enrichment Global Ecology and Biogeography, 26, 102-114. [本文引用: 1]
RaichJW, SchlesingerWH (1992). The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate Tellus B: Chemical and Physical Meteorology, 44, 81-99. [本文引用: 1]
ScharlemannJPW, TannerEVJ, HiedererR, KaposV (2014). Global soil carbon: understanding and managing the largest terrestrial carbon pool Carbon Management, 5, 81-91. [本文引用: 1]
ShanD, HanGD, ZhaoML, WangZ, HanX, GaoFG (2009). The effects of experimental warming and nitrogen addition on soil respiration in desert steppe Journal of Arid Land Resources and Environment, 23, 106-112. [本文引用: 1]
ShiBK, XuWL, ZhuY, WangCL, LoikME, SunW (2019). Heterogeneity of grassland soil respiration: antagonistic effects of grazing and nitrogen addition Agricultural and Forest Meteorology, 268, 215-223. [本文引用: 1]
SmithMD, KnappAK, CollinsSL (2009). A framework for assessing ecosystem dynamics in response to chronic resource alterations induced by global change Ecology, 90, 3279-3289. [本文引用: 1]
SongJ, WanSQ, PiaoSL, KnappAK, ClassenAT, ViccaS, CiaisP, HovendenMJ, LeuzingerS, BeierC, KardolP, XiaJY, LiuQ, RuJY, ZhouZX, et al. (2019). A meta-analysis of 1,119 manipulative experiments on terrestrial carbon-cycling responses to global change Nature Ecology & Evolution, 3, 1309-1320. [本文引用: 3]
TianDS, NiuSL (2015). A global analysis of soil acidification caused by nitrogen addition Environmental Research Letters, 10, 1-10. [本文引用: 1]
TianDS, WangH, SunJ, NiuSL (2016). Global evidence on nitrogen saturation of terrestrial ecosystem net primary productivity Environmental Research Letters, 11, 024012. DOI: 10.1088/1748-9326/11/2/024012. [本文引用: 1]
TresederKK (2008). Nitrogen additions and microbial biomass: a meta-analysis of ecosystem studies Ecology Letters, 11, 1111-1120. [本文引用: 2]
TuLH, HuTX, ZhangJ, LiXW, HuHL, LiuL, XiaoYL (2013). Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem Soil Biology & Biochemistry, 58, 255-264. [本文引用: 1]
WangJ, GaoYZ, ZhangYH, YangJJ, SmithMD, KnappAK, EissenstatDM, HanXG (2019a). Asymmetry in above- and belowground productivity responses to N addition in a semi-arid temperate steppe Global Change Biology, 25, 2958-2969. [本文引用: 6]
WangJB, FuXL, ZhangZ, LiMH, CaoHJ, ZhouXL, NiHW (2019b). Responses of soil respiration to nitrogen addition in the Sanjiang Plain wetland, northeastern China PLOS ONE, 14, e0211456. DOI: 10.1371/journal.pone.0211456. [本文引用: 2]
WangJS, SongB, MaFF, TianDS, LiY, YanT, QuanQ, ZhangFY, LiZL, WangBX, GaoQ, ChenWN, NiuSL (2019c). Nitrogen addition reduces soil respiration but increases the relative contribution of heterotrophic component in an alpine meadow Functional Ecology, 33, 2239-2253. [本文引用: 8]
WeiL, LiuJ, SuJH, JingGH, ZhaoJ, ChengJM, JinJW (2016). Effect of clipping on soil respiration components in temperate grassland of Loess Plateau European Journal of Soil Biology, 75, 157-167. [本文引用: 1]
WeiL, SuJS, JingGH, ZhaoJ, LiuJ, ChengJM, JinJW (2018). Nitrogen addition decreased soil respiration and its components in a longterm fenced grassland on the Loess Plateau Journal of Arid Environments, 152, 37-44. [本文引用: 2]
XiaoH, WangB, LuSB, ChenDM, WuY, ZhuYH, HuSJ, BaiYF (2020). Soil acidification reduces the effects of short-term nutrient enrichment on plant and soil biota and their interactions in grasslands Global Change Biology, 26, 4626-4637. [本文引用: 1]
YanLM, ChenSP, HuangJH, LinGH (2010). Differential responses of auto- and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe Global Change Biology, 16, 2345-2357. [本文引用: 2]
YanWD, ChenXY, PengYY, ZhuF, ZhenW, ZhangXY (2020). Response of soil respiration to nitrogen addition in two subtropical forest types Pedosphere, 30, 478-486. [本文引用: 1]
YangQ, WangW, ZengH (2018). Effects of nitrogen addition on the plant diversity and biomass of degraded grasslands of Nei Mongol, China Chinese Journal of Plant Ecology, 42, 430-441. [本文引用: 1]
ZengWJ, ChenJB, LiuHY, WangW (2018). Soil respiration and its autotrophic and heterotrophic components in response to nitrogen addition among different degraded temperate grasslands Soil Biology & Biochemistry, 124, 255-265. [本文引用: 1]
ZhangBW, LiS, ChenSP, RenTT, YangZQ, ZhaoHL, LiangY, HanXG (2016a). Arbuscular mycorrhizal fungi regulate soil respiration and its response to precipitation change in a semiarid steppe Scientific Reports, 6, 19990. DOI: 10.1038/srep19990. [本文引用: 1]
ZhangBY, LiWJ, ChenSP, TanXR, WangSS, ChenML, RenTT, XiaJY, HuangJH, HanXG (2019). Changing precipitation exerts greater influence on soil heterotrophic than autotrophic respiration in a semiarid steppe Agricultural and Forest Meteorology, 271, 413-421. [本文引用: 7]
ZhangYH, LüXT, IsbellF, StevensC, HanX, HeNP, ZhangGM, YuQ, HuangJH, HanXG (2014). Rapid plant species loss at high rates and at low frequency of N addition in temperate steppe Global Change Biology, 20, 3520-3529. [本文引用: 4]
ZhangYH, StevensCJ, LuXT, HeNP, HuangJH, HanXG (2016b). Fewer new species colonize at low frequency N addition in a temperate grassland Functional Ecology, 30, 1247-1256. [本文引用: 2]
ZhouJD, ShiRJ, ZhaoF, HanSQ, ZhangY (2016). Effects of the frequency and intensity of nitrogen addition on soil pH, the contents of carbon, nitrogen and phosphorus in temperate steppe in Inner Mongolia, China Chinese Journal of Applied Ecology, 27, 2467-2476. [本文引用: 3]
ZhouLY, ZhouXH, ShaoJJ, NieYY, HeYH, JiangLL, WuZT, BaiSH (2016). Interactive effects of global change factors on soil respiration and its components: a meta-analysis Global Change Biology, 22, 3157-3169.
ZhouLY, ZhouXH, ZhangBC, LuM, LuoYQ, LiuLL, BoL (2014). Different responses of soil respiration and its components to nitrogen addition among biomes: a meta-analysis Global Change Biology, 20, 2332-2343. [本文引用: 7]
ZhouXH, WanSQ, LuoYQ (2007). Source components and interannual variability of soil CO2 efflux under experimental warming and clipping in a grassland ecosystem Global Change Biology, 13, 761-775. [本文引用: 1]
ZhouZH, WangCK, LuoYQ (2020). Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality Nature Communications, 11, 3072-3081. DOI:10.1038/s41467-020-16881-7URL [本文引用: 3]
ZhouZH, WangCK, ZhengMH, JiangLF, LuoYQ (2017). Patterns and mechanisms of responses by soil microbial communities to nitrogen addition Soil Biology & Biochemistry, 115, 433-441. DOI:10.1016/j.soilbio.2017.09.015URL [本文引用: 2]
ZhuC, MaYP, WuHH, SunT, La PierreKJ, SunZW, YuQ (2016). Divergent effects of nitrogen addition on soil respiration in a semiarid grassland Scientific Reports, 6, 33541. DOI: 10.1038/srep33541. DOI:10.1038/srep33541URL [本文引用: 1]
Microbial activity and soil respiration under nitrogen addition in Alaskan boreal forest 1 2008
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Partitioning longleaf pine soil respiration into its heterotrophic and autotrophic components through root exclusion 1 2016
... 全球陆地生态系统近3/4的碳(C)储存在土壤中(Masciandaro et al., 2018), 储量高达1 395-2 011 Pg C, 约为大气碳储量的2-4倍(Scharlemann et al., 2014; Bond-Lamberty et al., 2018).作为陆地生态系统第二大碳通量, 土壤呼吸每年释放的CO2为68-98 Pg C, 约为每年化石燃料燃烧释放CO2的10倍(Raich & Schlesinger, 1992; Bond-Lamberty & Thomson, 2010).全球碳模型的预测表明, 由于全球变 化引发的土壤呼吸的微小变动都会显著影响大气 CO2浓度, 进而影响全球碳平衡(Cox et al., 2000; Zhu et al., 2016; Wang et al., 2019c).土壤呼吸根据CO2产生的来源不同主要分为两个组分, 一是自养呼吸, 主要是来自于植物根呼吸以及菌根真菌呼吸产生的CO2; 另一部分是异养呼吸, 主要是来自于凋落物分解和微生物降解土壤有机质产生的CO2 (Hanson et al., 2000; Bond-Lamberty et al., 2004).由于底物来源和对环境因子响应的差异, 土壤呼吸的自养和异养组分对氮沉降等全球变化的响应存在显著差别(Zhou et al., 2014; Wei et al., 2016).因此, 为了准确预测全球变化对土壤呼吸及生态系统碳循环的影响, 就需要区分土壤呼吸的不同组分, 并研究各组分对全球变化因子的响应规律(Wang et al., 2019c), 这对评估整个生态系统的碳收支平衡是非常重要的(Lovett et al., 2006; ArchMiller & Samuelson, 2016). ...
Interactive global change factors mitigate soil aggregation and carbon change in a semi-arid grassland 1 2020
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Differential responses of grasses and forbs led to marked reduction in below-ground productivity in temperate steppe following chronic N deposition 2 2015
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
... 大气氮沉降是持续而缓慢的氮添加过程, 而大多数模拟氮沉降实验研究是通过生长季少次脉冲式的氮添加处理进行的(Bai et al., 2015; Fang et al., 2017; Chen et al., 2018; Wang et al., 2019c), 不同的氮添加模式和频次会影响生态系统结构和功能对氮添加的响应(Tu et al., 2013; Zhang et al., 2014).本平台已有的研究结果表明, 在氮添加处理早期, 不同施氮频率对植物地上生产力并没有显著影响, 而对物种多样性、地下生产力和土壤微生物群落的影响是显著的(Zhang et al., 2016b; Wang et al., 2019a; Cao et al., 2020).Zhang等(2016b)对氮添加处理前5年的研究发现随着氮添加量的增加, 植物物种多样性显著降低, 而与HF (每年12次)相比, LF (每年2次)处理导致物种多样性丧失的速率更快, 这主要是因为LF处理下较高的氨态氮浓度限制了新物种的定植.Wang等(2019b)在氮添加处理的第5-7年的研究表明, 虽然不同施氮频率并未显著影响植被ANPP, 但LF显著降低了BNPP, 特别是在低氮添加量处理下.Cao等(2020)对施氮后第6和7年的研究表明, HF会显著减弱氮添加处理对植物和土壤细菌多样性的负作用.本研究在氮添加处理第11和12年进行测定, 我们并未发现不同施氮频率对土壤呼吸及其组分影响有显著差别, 而同期两种施氮频率下植物生产力、土壤微生物生物量、土壤细菌和真菌生物量以及F/B均没有显著的差异(附录I, 附录II).这表明随着氮添加处理时间的延长, 不同施氮频率对植物和微生物群落生产力和结构的影响逐渐减弱.因为随着土壤中可利用氮的不断增加, 造成土壤酸化和有毒离子的积累, 生态系统逐渐呈现出氮饱和的状态(Tian et al., 2016), 进而逐步削弱了不同施氮频率对生态系统影响的差别.本研究的HF处理虽然尽可能地提高了氮添加频次, 但与缓慢持续的大气氮沉降过程仍存在较大差别, 使得我们在研究生态系统结构和功能对大气氮沉降的响应过程中仍存在很大不确定性.随着氮添加手段的改进, 如采用具亲水性包衣的缓释尿素等, 可以更精确地模拟大气氮沉降特别是湿沉降的过程, 为准确模拟大气氮沉降对生态系统碳氮循环过程的影响提供了可能. ...
Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from Inner Mongolia grasslands 3 2010
... 氮是内蒙古半干旱草原生产力和碳交换过程的主要限制因子(Bai et al., 2010).本研究发现随着氮添加量的增加, Rs显著下降(图3), 相同的结果在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草原(Wei et al., 2018)中同样被报道.但是氮添加对Rs无显著影响(Fu et al., 2015)或有促进作用(Zhou et al., 2014, 2016)的结果也多次被报道.通过比较发现, 氮添加对Rs的正作用通常多出现在森林生态系统或短期的氮添加实验中(Zhou et al., 2014; Fu et al., 2015; Luo et al., 2016).而随着氮处理时间的延长,Rs对氮添加的正响应会逐渐减弱, 并转变成负响应(Zhou et al., 2014, 2016).这主要是因为在实验处理初期, 氮添加主要表现出对植物生长的促进作用, 但随着处理时间的延长, 氮添加所导致的土壤pH下降对植物和土壤微生物的负面作用逐渐增强(Janssens et al., 2010; Liang et al., 2019).本研究Rs的测定是在氮添加处理的第11和12年进行的, 历经了10年氮添加处理后, 土壤pH显著降低, 氮添加对Rs的抑制作用十分明显, 特别是当氮添加量大于10 g·m-2·a-1时, Rs显著降低了近22.9 %.Bai等(2010)在本区域的长期氮添加实验研究表明, 10.5 g·m-2·a-1氮添加量是影响内蒙古半干旱草原土壤氮有效性显著增加的关键阈值. ...
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
Plant-bacteria-soil response to frequency of simulated nitrogen deposition has implications for global ecosystem change 3 2020
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
... 大气氮沉降是持续而缓慢的氮添加过程, 而大多数模拟氮沉降实验研究是通过生长季少次脉冲式的氮添加处理进行的(Bai et al., 2015; Fang et al., 2017; Chen et al., 2018; Wang et al., 2019c), 不同的氮添加模式和频次会影响生态系统结构和功能对氮添加的响应(Tu et al., 2013; Zhang et al., 2014).本平台已有的研究结果表明, 在氮添加处理早期, 不同施氮频率对植物地上生产力并没有显著影响, 而对物种多样性、地下生产力和土壤微生物群落的影响是显著的(Zhang et al., 2016b; Wang et al., 2019a; Cao et al., 2020).Zhang等(2016b)对氮添加处理前5年的研究发现随着氮添加量的增加, 植物物种多样性显著降低, 而与HF (每年12次)相比, LF (每年2次)处理导致物种多样性丧失的速率更快, 这主要是因为LF处理下较高的氨态氮浓度限制了新物种的定植.Wang等(2019b)在氮添加处理的第5-7年的研究表明, 虽然不同施氮频率并未显著影响植被ANPP, 但LF显著降低了BNPP, 特别是在低氮添加量处理下.Cao等(2020)对施氮后第6和7年的研究表明, HF会显著减弱氮添加处理对植物和土壤细菌多样性的负作用.本研究在氮添加处理第11和12年进行测定, 我们并未发现不同施氮频率对土壤呼吸及其组分影响有显著差别, 而同期两种施氮频率下植物生产力、土壤微生物生物量、土壤细菌和真菌生物量以及F/B均没有显著的差异(附录I, 附录II).这表明随着氮添加处理时间的延长, 不同施氮频率对植物和微生物群落生产力和结构的影响逐渐减弱.因为随着土壤中可利用氮的不断增加, 造成土壤酸化和有毒离子的积累, 生态系统逐渐呈现出氮饱和的状态(Tian et al., 2016), 进而逐步削弱了不同施氮频率对生态系统影响的差别.本研究的HF处理虽然尽可能地提高了氮添加频次, 但与缓慢持续的大气氮沉降过程仍存在较大差别, 使得我们在研究生态系统结构和功能对大气氮沉降的响应过程中仍存在很大不确定性.随着氮添加手段的改进, 如采用具亲水性包衣的缓释尿素等, 可以更精确地模拟大气氮沉降特别是湿沉降的过程, 为准确模拟大气氮沉降对生态系统碳氮循环过程的影响提供了可能. ...
Effects of nitrogen enrichment on belowground communities in grassland: relative role of soil nitrogen availability vs. soil acidification 1 2015
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
Soil acidification exerts a greater control on soil respiration than soil nitrogen availability in grasslands subjected to long-term nitrogen enrichment 1 2016
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Direct and indirect effects of nitrogen enrichment on soil organisms and carbon and nitrogen mineralization in a semi-arid grassland 1 2019
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
Effects of nitrogen deposition on carbon cycle in terrestrial ecosystems of China: a meta- analysis 1 2015
... Rs对氮添加响应的强度和方向取决于Rh和Ra两个组分的响应, 本研究中氮添加显著降低了Rh, 对Ra并无显著影响.氮添加并没有影响Rh对Rs的贡献比例, Rh平均占Rs的62%, 这与本区域其他研究结果(Zhang et al., 2016a, 2019)一致.大部分陆地生态系统的生产力是受氮限制的, 通常氮添加会通过提高土壤氮可利用性而促进植物生长, 增加植物自养呼吸(Chen et al., 2015b).但土壤氮限制缓解的同时也会改变植物的地上地下碳分配策略, 减少植物对地下根系的碳分配比例, 该实验平台的研究表明在氮添加处理的第6和第7年, 氮添加使得ANPP增加了45%-53%, 但导致地下净初级生产力(BNPP)平均下降了40%-47% (Wang et al., 2019a).这可能是本研究中Ra对氮添加处理缺乏响应的原因.一项全球氮添加实验的meta分析表明, 氮添加会显著降低根系生物量和周转速率, 但对根系现存量没有影响(Peng et al., 2017), 由于Ra主要来自于现存根系呼吸, 这可以进一步解释为什么本研究中Ra对氮添加处理没有显著响应. ...
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Nitrogen fertilization stimulated soil heterotrophic but not autotrophic respiration in cropland soils: a greater role of organic over inorganic fertilizer 2 2018
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
... 大气氮沉降是持续而缓慢的氮添加过程, 而大多数模拟氮沉降实验研究是通过生长季少次脉冲式的氮添加处理进行的(Bai et al., 2015; Fang et al., 2017; Chen et al., 2018; Wang et al., 2019c), 不同的氮添加模式和频次会影响生态系统结构和功能对氮添加的响应(Tu et al., 2013; Zhang et al., 2014).本平台已有的研究结果表明, 在氮添加处理早期, 不同施氮频率对植物地上生产力并没有显著影响, 而对物种多样性、地下生产力和土壤微生物群落的影响是显著的(Zhang et al., 2016b; Wang et al., 2019a; Cao et al., 2020).Zhang等(2016b)对氮添加处理前5年的研究发现随着氮添加量的增加, 植物物种多样性显著降低, 而与HF (每年12次)相比, LF (每年2次)处理导致物种多样性丧失的速率更快, 这主要是因为LF处理下较高的氨态氮浓度限制了新物种的定植.Wang等(2019b)在氮添加处理的第5-7年的研究表明, 虽然不同施氮频率并未显著影响植被ANPP, 但LF显著降低了BNPP, 特别是在低氮添加量处理下.Cao等(2020)对施氮后第6和7年的研究表明, HF会显著减弱氮添加处理对植物和土壤细菌多样性的负作用.本研究在氮添加处理第11和12年进行测定, 我们并未发现不同施氮频率对土壤呼吸及其组分影响有显著差别, 而同期两种施氮频率下植物生产力、土壤微生物生物量、土壤细菌和真菌生物量以及F/B均没有显著的差异(附录I, 附录II).这表明随着氮添加处理时间的延长, 不同施氮频率对植物和微生物群落生产力和结构的影响逐渐减弱.因为随着土壤中可利用氮的不断增加, 造成土壤酸化和有毒离子的积累, 生态系统逐渐呈现出氮饱和的状态(Tian et al., 2016), 进而逐步削弱了不同施氮频率对生态系统影响的差别.本研究的HF处理虽然尽可能地提高了氮添加频次, 但与缓慢持续的大气氮沉降过程仍存在较大差别, 使得我们在研究生态系统结构和功能对大气氮沉降的响应过程中仍存在很大不确定性.随着氮添加手段的改进, 如采用具亲水性包衣的缓释尿素等, 可以更精确地模拟大气氮沉降特别是湿沉降的过程, 为准确模拟大气氮沉降对生态系统碳氮循环过程的影响提供了可能. ...
1 2000
... 草原生态系统是我国重要的生态系统类型, 占我国国土面积的近40%, 其中78%的草原位于北方干旱半干旱地区(陈佐忠和汪诗平, 2000), 对全球变化非常敏感(Niu et al., 2008; Liu et al., 2018).本研究基于建立在内蒙古半干旱草原的长期氮添加平台.自2008年平台建立以来, 已经进行了长达10年的氮添加处理.通过连续两年(2018-2019年)土壤呼吸及其组分的测量, 本研究对比了不同氮添加量和氮添加频率对草原土壤碳释放的影响, 阐明调控土壤呼吸及其组分响应的主要环境和生物因子.在长期氮添加背景下, 明确土壤呼吸及其组分对氮添加量和添加频率的响应规律, 对进一步理解氮沉降对草原生态系统土壤碳平衡与固碳潜力的影响有重要的意义. ...
1 2000
... 草原生态系统是我国重要的生态系统类型, 占我国国土面积的近40%, 其中78%的草原位于北方干旱半干旱地区(陈佐忠和汪诗平, 2000), 对全球变化非常敏感(Niu et al., 2008; Liu et al., 2018).本研究基于建立在内蒙古半干旱草原的长期氮添加平台.自2008年平台建立以来, 已经进行了长达10年的氮添加处理.通过连续两年(2018-2019年)土壤呼吸及其组分的测量, 本研究对比了不同氮添加量和氮添加频率对草原土壤碳释放的影响, 阐明调控土壤呼吸及其组分响应的主要环境和生物因子.在长期氮添加背景下, 明确土壤呼吸及其组分对氮添加量和添加频率的响应规律, 对进一步理解氮沉降对草原生态系统土壤碳平衡与固碳潜力的影响有重要的意义. ...
Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands 1 2008
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model 1 2000
... 全球陆地生态系统近3/4的碳(C)储存在土壤中(Masciandaro et al., 2018), 储量高达1 395-2 011 Pg C, 约为大气碳储量的2-4倍(Scharlemann et al., 2014; Bond-Lamberty et al., 2018).作为陆地生态系统第二大碳通量, 土壤呼吸每年释放的CO2为68-98 Pg C, 约为每年化石燃料燃烧释放CO2的10倍(Raich & Schlesinger, 1992; Bond-Lamberty & Thomson, 2010).全球碳模型的预测表明, 由于全球变 化引发的土壤呼吸的微小变动都会显著影响大气 CO2浓度, 进而影响全球碳平衡(Cox et al., 2000; Zhu et al., 2016; Wang et al., 2019c).土壤呼吸根据CO2产生的来源不同主要分为两个组分, 一是自养呼吸, 主要是来自于植物根呼吸以及菌根真菌呼吸产生的CO2; 另一部分是异养呼吸, 主要是来自于凋落物分解和微生物降解土壤有机质产生的CO2 (Hanson et al., 2000; Bond-Lamberty et al., 2004).由于底物来源和对环境因子响应的差异, 土壤呼吸的自养和异养组分对氮沉降等全球变化的响应存在显著差别(Zhou et al., 2014; Wei et al., 2016).因此, 为了准确预测全球变化对土壤呼吸及生态系统碳循环的影响, 就需要区分土壤呼吸的不同组分, 并研究各组分对全球变化因子的响应规律(Wang et al., 2019c), 这对评估整个生态系统的碳收支平衡是非常重要的(Lovett et al., 2006; ArchMiller & Samuelson, 2016). ...
The response of soil CO2 flux to changes in atmospheric CO2, nitrogen supply and plant diversity 1 2001
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Impacts of warming and nitrogen addition on soil autotrophic and heterotrophic respiration in a semi-arid environment 1 2018
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Seasonal responses of soil respiration to warming and nitrogen addition in a semi-arid alfalfa- pasture of the Loess Plateau, China 2 2017
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
... 大气氮沉降是持续而缓慢的氮添加过程, 而大多数模拟氮沉降实验研究是通过生长季少次脉冲式的氮添加处理进行的(Bai et al., 2015; Fang et al., 2017; Chen et al., 2018; Wang et al., 2019c), 不同的氮添加模式和频次会影响生态系统结构和功能对氮添加的响应(Tu et al., 2013; Zhang et al., 2014).本平台已有的研究结果表明, 在氮添加处理早期, 不同施氮频率对植物地上生产力并没有显著影响, 而对物种多样性、地下生产力和土壤微生物群落的影响是显著的(Zhang et al., 2016b; Wang et al., 2019a; Cao et al., 2020).Zhang等(2016b)对氮添加处理前5年的研究发现随着氮添加量的增加, 植物物种多样性显著降低, 而与HF (每年12次)相比, LF (每年2次)处理导致物种多样性丧失的速率更快, 这主要是因为LF处理下较高的氨态氮浓度限制了新物种的定植.Wang等(2019b)在氮添加处理的第5-7年的研究表明, 虽然不同施氮频率并未显著影响植被ANPP, 但LF显著降低了BNPP, 特别是在低氮添加量处理下.Cao等(2020)对施氮后第6和7年的研究表明, HF会显著减弱氮添加处理对植物和土壤细菌多样性的负作用.本研究在氮添加处理第11和12年进行测定, 我们并未发现不同施氮频率对土壤呼吸及其组分影响有显著差别, 而同期两种施氮频率下植物生产力、土壤微生物生物量、土壤细菌和真菌生物量以及F/B均没有显著的差异(附录I, 附录II).这表明随着氮添加处理时间的延长, 不同施氮频率对植物和微生物群落生产力和结构的影响逐渐减弱.因为随着土壤中可利用氮的不断增加, 造成土壤酸化和有毒离子的积累, 生态系统逐渐呈现出氮饱和的状态(Tian et al., 2016), 进而逐步削弱了不同施氮频率对生态系统影响的差别.本研究的HF处理虽然尽可能地提高了氮添加频次, 但与缓慢持续的大气氮沉降过程仍存在较大差别, 使得我们在研究生态系统结构和功能对大气氮沉降的响应过程中仍存在很大不确定性.随着氮添加手段的改进, 如采用具亲水性包衣的缓释尿素等, 可以更精确地模拟大气氮沉降特别是湿沉降的过程, 为准确模拟大气氮沉降对生态系统碳氮循环过程的影响提供了可能. ...
Soil carbon sequestration in prairie grasslands increased by chronic nitrogen addition 1 2012
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
Use the misuse of PLFA measurements in soils 1 2011
... 土壤微生物生物量(MB)和土壤微生物功能群组成采用磷脂脂肪酸(PLFA)方法测定(Frosteg?rd et al., 2011).根据细菌的特异性脂肪酸(i15:0、i16:0、i17:0、18:1ω7、cy17:0和cy19:0)和真菌的特异性脂肪酸(18:2ω6, 9)的含量代表土壤中细菌和真菌的生物量(Wang et al., 2019c), 并计算真菌/细菌比(F/B). ...
What have we learned from global change manipulative experiments in China? A meta-analysis 2 2015
... 氮是内蒙古半干旱草原生产力和碳交换过程的主要限制因子(Bai et al., 2010).本研究发现随着氮添加量的增加, Rs显著下降(图3), 相同的结果在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草原(Wei et al., 2018)中同样被报道.但是氮添加对Rs无显著影响(Fu et al., 2015)或有促进作用(Zhou et al., 2014, 2016)的结果也多次被报道.通过比较发现, 氮添加对Rs的正作用通常多出现在森林生态系统或短期的氮添加实验中(Zhou et al., 2014; Fu et al., 2015; Luo et al., 2016).而随着氮处理时间的延长,Rs对氮添加的正响应会逐渐减弱, 并转变成负响应(Zhou et al., 2014, 2016).这主要是因为在实验处理初期, 氮添加主要表现出对植物生长的促进作用, 但随着处理时间的延长, 氮添加所导致的土壤pH下降对植物和土壤微生物的负面作用逐渐增强(Janssens et al., 2010; Liang et al., 2019).本研究Rs的测定是在氮添加处理的第11和12年进行的, 历经了10年氮添加处理后, 土壤pH显著降低, 氮添加对Rs的抑制作用十分明显, 特别是当氮添加量大于10 g·m-2·a-1时, Rs显著降低了近22.9 %.Bai等(2010)在本区域的长期氮添加实验研究表明, 10.5 g·m-2·a-1氮添加量是影响内蒙古半干旱草原土壤氮有效性显著增加的关键阈值. ...
... ; Fu et al., 2015; Luo et al., 2016).而随着氮处理时间的延长,Rs对氮添加的正响应会逐渐减弱, 并转变成负响应(Zhou et al., 2014, 2016).这主要是因为在实验处理初期, 氮添加主要表现出对植物生长的促进作用, 但随着处理时间的延长, 氮添加所导致的土壤pH下降对植物和土壤微生物的负面作用逐渐增强(Janssens et al., 2010; Liang et al., 2019).本研究Rs的测定是在氮添加处理的第11和12年进行的, 历经了10年氮添加处理后, 土壤pH显著降低, 氮添加对Rs的抑制作用十分明显, 特别是当氮添加量大于10 g·m-2·a-1时, Rs显著降低了近22.9 %.Bai等(2010)在本区域的长期氮添加实验研究表明, 10.5 g·m-2·a-1氮添加量是影响内蒙古半干旱草原土壤氮有效性显著增加的关键阈值. ...
Nitrogen cycles: past, present, and future 1 2004
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Transformation of the nitrogen cycle: recent trends, questions, and potential solutions 2 2008
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
Data from: N balance and cycling of Inner Mongolia typical steppe—A comprehensive case study of grazing effects 2 2013
... 本研究样地位于内蒙古锡林郭勒盟的内蒙古草原生态系统国家定位研究站(116.70° E, 43.38° N, 海拔1 200 m).该地区属于温带半干旱大陆性气候, 长期(1983-2012年)的气象数据表明, 该区域的年平均气温为0.9 ℃, 月平均气温变化从最冷月(1月) -21.7 ℃到最暖月(7月) 19.7 ℃.年降水量为355.3 mm, 其中60%-80%的降水量集中在生长季(5-8月)(Zhang et al., 2014).该区域主要土壤类型为栗钙土.地带性植被为温带半干旱典型草原, 优势物种为大针茅(Stipa grandis)、羊草(Leymus chinensis)等.该区域大气氮沉降的背景值为1.8 g·m-2·a-1, 其中干、湿氮沉降分别为1.2和0.6 g·m-2·a-1 (Giese et al., 2013). ...
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
Separating root and soil microbial contributions to soil respiration—A review of methods and observations 1 2000
... 全球陆地生态系统近3/4的碳(C)储存在土壤中(Masciandaro et al., 2018), 储量高达1 395-2 011 Pg C, 约为大气碳储量的2-4倍(Scharlemann et al., 2014; Bond-Lamberty et al., 2018).作为陆地生态系统第二大碳通量, 土壤呼吸每年释放的CO2为68-98 Pg C, 约为每年化石燃料燃烧释放CO2的10倍(Raich & Schlesinger, 1992; Bond-Lamberty & Thomson, 2010).全球碳模型的预测表明, 由于全球变 化引发的土壤呼吸的微小变动都会显著影响大气 CO2浓度, 进而影响全球碳平衡(Cox et al., 2000; Zhu et al., 2016; Wang et al., 2019c).土壤呼吸根据CO2产生的来源不同主要分为两个组分, 一是自养呼吸, 主要是来自于植物根呼吸以及菌根真菌呼吸产生的CO2; 另一部分是异养呼吸, 主要是来自于凋落物分解和微生物降解土壤有机质产生的CO2 (Hanson et al., 2000; Bond-Lamberty et al., 2004).由于底物来源和对环境因子响应的差异, 土壤呼吸的自养和异养组分对氮沉降等全球变化的响应存在显著差别(Zhou et al., 2014; Wei et al., 2016).因此, 为了准确预测全球变化对土壤呼吸及生态系统碳循环的影响, 就需要区分土壤呼吸的不同组分, 并研究各组分对全球变化因子的响应规律(Wang et al., 2019c), 这对评估整个生态系统的碳收支平衡是非常重要的(Lovett et al., 2006; ArchMiller & Samuelson, 2016). ...
Contrasting effects of low and high nitrogen additions on soil CO2 flux components and ectomycorrhizal fungal sporocarp production in a boreal forest 1 2012
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Reduction of forest soil respiration in response to nitrogen deposition 5 2010
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... ; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... 氮是内蒙古半干旱草原生产力和碳交换过程的主要限制因子(Bai et al., 2010).本研究发现随着氮添加量的增加, Rs显著下降(图3), 相同的结果在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草原(Wei et al., 2018)中同样被报道.但是氮添加对Rs无显著影响(Fu et al., 2015)或有促进作用(Zhou et al., 2014, 2016)的结果也多次被报道.通过比较发现, 氮添加对Rs的正作用通常多出现在森林生态系统或短期的氮添加实验中(Zhou et al., 2014; Fu et al., 2015; Luo et al., 2016).而随着氮处理时间的延长,Rs对氮添加的正响应会逐渐减弱, 并转变成负响应(Zhou et al., 2014, 2016).这主要是因为在实验处理初期, 氮添加主要表现出对植物生长的促进作用, 但随着处理时间的延长, 氮添加所导致的土壤pH下降对植物和土壤微生物的负面作用逐渐增强(Janssens et al., 2010; Liang et al., 2019).本研究Rs的测定是在氮添加处理的第11和12年进行的, 历经了10年氮添加处理后, 土壤pH显著降低, 氮添加对Rs的抑制作用十分明显, 特别是当氮添加量大于10 g·m-2·a-1时, Rs显著降低了近22.9 %.Bai等(2010)在本区域的长期氮添加实验研究表明, 10.5 g·m-2·a-1氮添加量是影响内蒙古半干旱草原土壤氮有效性显著增加的关键阈值. ...
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
... ).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
Responses of soil respiration to N addition, burning and clipping in temperate semiarid grassland in northern China 2 2012
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... ; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Soil respiration, fine root production, and microbial biomass in cottonwood and loblolly pine plantations along a nitrogen fertilization gradient 1 2003
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
水、氮控制对短花针茅草原土壤呼吸的影响 1 2015
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
水、氮控制对短花针茅草原土壤呼吸的影响 1 2015
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Pathways regulating decreased soil respiration with nitrogen addition in a subtropical forest in China 1 2019
... 氮是内蒙古半干旱草原生产力和碳交换过程的主要限制因子(Bai et al., 2010).本研究发现随着氮添加量的增加, Rs显著下降(图3), 相同的结果在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草原(Wei et al., 2018)中同样被报道.但是氮添加对Rs无显著影响(Fu et al., 2015)或有促进作用(Zhou et al., 2014, 2016)的结果也多次被报道.通过比较发现, 氮添加对Rs的正作用通常多出现在森林生态系统或短期的氮添加实验中(Zhou et al., 2014; Fu et al., 2015; Luo et al., 2016).而随着氮处理时间的延长,Rs对氮添加的正响应会逐渐减弱, 并转变成负响应(Zhou et al., 2014, 2016).这主要是因为在实验处理初期, 氮添加主要表现出对植物生长的促进作用, 但随着处理时间的延长, 氮添加所导致的土壤pH下降对植物和土壤微生物的负面作用逐渐增强(Janssens et al., 2010; Liang et al., 2019).本研究Rs的测定是在氮添加处理的第11和12年进行的, 历经了10年氮添加处理后, 土壤pH显著降低, 氮添加对Rs的抑制作用十分明显, 特别是当氮添加量大于10 g·m-2·a-1时, Rs显著降低了近22.9 %.Bai等(2010)在本区域的长期氮添加实验研究表明, 10.5 g·m-2·a-1氮添加量是影响内蒙古半干旱草原土壤氮有效性显著增加的关键阈值. ...
Effects of water and nitrogen addition on ecosystem respiration across three types of steppe: the role of plant and microbial biomass 1 2018
... 草原生态系统是我国重要的生态系统类型, 占我国国土面积的近40%, 其中78%的草原位于北方干旱半干旱地区(陈佐忠和汪诗平, 2000), 对全球变化非常敏感(Niu et al., 2008; Liu et al., 2018).本研究基于建立在内蒙古半干旱草原的长期氮添加平台.自2008年平台建立以来, 已经进行了长达10年的氮添加处理.通过连续两年(2018-2019年)土壤呼吸及其组分的测量, 本研究对比了不同氮添加量和氮添加频率对草原土壤碳释放的影响, 阐明调控土壤呼吸及其组分响应的主要环境和生物因子.在长期氮添加背景下, 明确土壤呼吸及其组分对氮添加量和添加频率的响应规律, 对进一步理解氮沉降对草原生态系统土壤碳平衡与固碳潜力的影响有重要的意义. ...
Enhanced nitrogen deposition over China 1 2013
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Is net ecosystem production equal to ecosystem carbon accumulation? 1 2006
... 全球陆地生态系统近3/4的碳(C)储存在土壤中(Masciandaro et al., 2018), 储量高达1 395-2 011 Pg C, 约为大气碳储量的2-4倍(Scharlemann et al., 2014; Bond-Lamberty et al., 2018).作为陆地生态系统第二大碳通量, 土壤呼吸每年释放的CO2为68-98 Pg C, 约为每年化石燃料燃烧释放CO2的10倍(Raich & Schlesinger, 1992; Bond-Lamberty & Thomson, 2010).全球碳模型的预测表明, 由于全球变 化引发的土壤呼吸的微小变动都会显著影响大气 CO2浓度, 进而影响全球碳平衡(Cox et al., 2000; Zhu et al., 2016; Wang et al., 2019c).土壤呼吸根据CO2产生的来源不同主要分为两个组分, 一是自养呼吸, 主要是来自于植物根呼吸以及菌根真菌呼吸产生的CO2; 另一部分是异养呼吸, 主要是来自于凋落物分解和微生物降解土壤有机质产生的CO2 (Hanson et al., 2000; Bond-Lamberty et al., 2004).由于底物来源和对环境因子响应的差异, 土壤呼吸的自养和异养组分对氮沉降等全球变化的响应存在显著差别(Zhou et al., 2014; Wei et al., 2016).因此, 为了准确预测全球变化对土壤呼吸及生态系统碳循环的影响, 就需要区分土壤呼吸的不同组分, 并研究各组分对全球变化因子的响应规律(Wang et al., 2019c), 这对评估整个生态系统的碳收支平衡是非常重要的(Lovett et al., 2006; ArchMiller & Samuelson, 2016). ...
Atmospheric nitrogen deposition promotes carbon loss from peat bogs 1 2006
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
The responses of soil respiration to nitrogen addition in a temperate grassland in northern China 2 2016
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... 氮是内蒙古半干旱草原生产力和碳交换过程的主要限制因子(Bai et al., 2010).本研究发现随着氮添加量的增加, Rs显著下降(图3), 相同的结果在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草原(Wei et al., 2018)中同样被报道.但是氮添加对Rs无显著影响(Fu et al., 2015)或有促进作用(Zhou et al., 2014, 2016)的结果也多次被报道.通过比较发现, 氮添加对Rs的正作用通常多出现在森林生态系统或短期的氮添加实验中(Zhou et al., 2014; Fu et al., 2015; Luo et al., 2016).而随着氮处理时间的延长,Rs对氮添加的正响应会逐渐减弱, 并转变成负响应(Zhou et al., 2014, 2016).这主要是因为在实验处理初期, 氮添加主要表现出对植物生长的促进作用, 但随着处理时间的延长, 氮添加所导致的土壤pH下降对植物和土壤微生物的负面作用逐渐增强(Janssens et al., 2010; Liang et al., 2019).本研究Rs的测定是在氮添加处理的第11和12年进行的, 历经了10年氮添加处理后, 土壤pH显著降低, 氮添加对Rs的抑制作用十分明显, 特别是当氮添加量大于10 g·m-2·a-1时, Rs显著降低了近22.9 %.Bai等(2010)在本区域的长期氮添加实验研究表明, 10.5 g·m-2·a-1氮添加量是影响内蒙古半干旱草原土壤氮有效性显著增加的关键阈值. ...
Plant and soil microbe responses to light, warming and nitrogen addition in a temperate forest 1 2018
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
1 2018
... 全球陆地生态系统近3/4的碳(C)储存在土壤中(Masciandaro et al., 2018), 储量高达1 395-2 011 Pg C, 约为大气碳储量的2-4倍(Scharlemann et al., 2014; Bond-Lamberty et al., 2018).作为陆地生态系统第二大碳通量, 土壤呼吸每年释放的CO2为68-98 Pg C, 约为每年化石燃料燃烧释放CO2的10倍(Raich & Schlesinger, 1992; Bond-Lamberty & Thomson, 2010).全球碳模型的预测表明, 由于全球变 化引发的土壤呼吸的微小变动都会显著影响大气 CO2浓度, 进而影响全球碳平衡(Cox et al., 2000; Zhu et al., 2016; Wang et al., 2019c).土壤呼吸根据CO2产生的来源不同主要分为两个组分, 一是自养呼吸, 主要是来自于植物根呼吸以及菌根真菌呼吸产生的CO2; 另一部分是异养呼吸, 主要是来自于凋落物分解和微生物降解土壤有机质产生的CO2 (Hanson et al., 2000; Bond-Lamberty et al., 2004).由于底物来源和对环境因子响应的差异, 土壤呼吸的自养和异养组分对氮沉降等全球变化的响应存在显著差别(Zhou et al., 2014; Wei et al., 2016).因此, 为了准确预测全球变化对土壤呼吸及生态系统碳循环的影响, 就需要区分土壤呼吸的不同组分, 并研究各组分对全球变化因子的响应规律(Wang et al., 2019c), 这对评估整个生态系统的碳收支平衡是非常重要的(Lovett et al., 2006; ArchMiller & Samuelson, 2016). ...
Nitrogen addition reduces soil respiration in a mature tropical forest in southern China 2 2008
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... )或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe 1 2008
... 草原生态系统是我国重要的生态系统类型, 占我国国土面积的近40%, 其中78%的草原位于北方干旱半干旱地区(陈佐忠和汪诗平, 2000), 对全球变化非常敏感(Niu et al., 2008; Liu et al., 2018).本研究基于建立在内蒙古半干旱草原的长期氮添加平台.自2008年平台建立以来, 已经进行了长达10年的氮添加处理.通过连续两年(2018-2019年)土壤呼吸及其组分的测量, 本研究对比了不同氮添加量和氮添加频率对草原土壤碳释放的影响, 阐明调控土壤呼吸及其组分响应的主要环境和生物因子.在长期氮添加背景下, 明确土壤呼吸及其组分对氮添加量和添加频率的响应规律, 对进一步理解氮沉降对草原生态系统土壤碳平衡与固碳潜力的影响有重要的意义. ...
Nitrogen effects on net ecosystem carbon exchange in a temperate steppe 1 2010
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Global patterns of root dynamics under nitrogen enrichment 1 2017
... Rs对氮添加响应的强度和方向取决于Rh和Ra两个组分的响应, 本研究中氮添加显著降低了Rh, 对Ra并无显著影响.氮添加并没有影响Rh对Rs的贡献比例, Rh平均占Rs的62%, 这与本区域其他研究结果(Zhang et al., 2016a, 2019)一致.大部分陆地生态系统的生产力是受氮限制的, 通常氮添加会通过提高土壤氮可利用性而促进植物生长, 增加植物自养呼吸(Chen et al., 2015b).但土壤氮限制缓解的同时也会改变植物的地上地下碳分配策略, 减少植物对地下根系的碳分配比例, 该实验平台的研究表明在氮添加处理的第6和第7年, 氮添加使得ANPP增加了45%-53%, 但导致地下净初级生产力(BNPP)平均下降了40%-47% (Wang et al., 2019a).这可能是本研究中Ra对氮添加处理缺乏响应的原因.一项全球氮添加实验的meta分析表明, 氮添加会显著降低根系生物量和周转速率, 但对根系现存量没有影响(Peng et al., 2017), 由于Ra主要来自于现存根系呼吸, 这可以进一步解释为什么本研究中Ra对氮添加处理没有显著响应. ...
The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate 1 1992
... 全球陆地生态系统近3/4的碳(C)储存在土壤中(Masciandaro et al., 2018), 储量高达1 395-2 011 Pg C, 约为大气碳储量的2-4倍(Scharlemann et al., 2014; Bond-Lamberty et al., 2018).作为陆地生态系统第二大碳通量, 土壤呼吸每年释放的CO2为68-98 Pg C, 约为每年化石燃料燃烧释放CO2的10倍(Raich & Schlesinger, 1992; Bond-Lamberty & Thomson, 2010).全球碳模型的预测表明, 由于全球变 化引发的土壤呼吸的微小变动都会显著影响大气 CO2浓度, 进而影响全球碳平衡(Cox et al., 2000; Zhu et al., 2016; Wang et al., 2019c).土壤呼吸根据CO2产生的来源不同主要分为两个组分, 一是自养呼吸, 主要是来自于植物根呼吸以及菌根真菌呼吸产生的CO2; 另一部分是异养呼吸, 主要是来自于凋落物分解和微生物降解土壤有机质产生的CO2 (Hanson et al., 2000; Bond-Lamberty et al., 2004).由于底物来源和对环境因子响应的差异, 土壤呼吸的自养和异养组分对氮沉降等全球变化的响应存在显著差别(Zhou et al., 2014; Wei et al., 2016).因此, 为了准确预测全球变化对土壤呼吸及生态系统碳循环的影响, 就需要区分土壤呼吸的不同组分, 并研究各组分对全球变化因子的响应规律(Wang et al., 2019c), 这对评估整个生态系统的碳收支平衡是非常重要的(Lovett et al., 2006; ArchMiller & Samuelson, 2016). ...
Global soil carbon: understanding and managing the largest terrestrial carbon pool 1 2014
... 全球陆地生态系统近3/4的碳(C)储存在土壤中(Masciandaro et al., 2018), 储量高达1 395-2 011 Pg C, 约为大气碳储量的2-4倍(Scharlemann et al., 2014; Bond-Lamberty et al., 2018).作为陆地生态系统第二大碳通量, 土壤呼吸每年释放的CO2为68-98 Pg C, 约为每年化石燃料燃烧释放CO2的10倍(Raich & Schlesinger, 1992; Bond-Lamberty & Thomson, 2010).全球碳模型的预测表明, 由于全球变 化引发的土壤呼吸的微小变动都会显著影响大气 CO2浓度, 进而影响全球碳平衡(Cox et al., 2000; Zhu et al., 2016; Wang et al., 2019c).土壤呼吸根据CO2产生的来源不同主要分为两个组分, 一是自养呼吸, 主要是来自于植物根呼吸以及菌根真菌呼吸产生的CO2; 另一部分是异养呼吸, 主要是来自于凋落物分解和微生物降解土壤有机质产生的CO2 (Hanson et al., 2000; Bond-Lamberty et al., 2004).由于底物来源和对环境因子响应的差异, 土壤呼吸的自养和异养组分对氮沉降等全球变化的响应存在显著差别(Zhou et al., 2014; Wei et al., 2016).因此, 为了准确预测全球变化对土壤呼吸及生态系统碳循环的影响, 就需要区分土壤呼吸的不同组分, 并研究各组分对全球变化因子的响应规律(Wang et al., 2019c), 这对评估整个生态系统的碳收支平衡是非常重要的(Lovett et al., 2006; ArchMiller & Samuelson, 2016). ...
控制性增温和施氮对荒漠草原土壤呼吸的影响 1 2009
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
控制性增温和施氮对荒漠草原土壤呼吸的影响 1 2009
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Heterogeneity of grassland soil respiration: antagonistic effects of grazing and nitrogen addition 1 2019
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
A framework for assessing ecosystem dynamics in response to chronic resource alterations induced by global change 1 2009
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
A meta-analysis of 1,119 manipulative experiments on terrestrial carbon-cycling responses to global change 3 2019
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... ; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... ; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
A global analysis of soil acidification caused by nitrogen addition 1 2015
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
Global evidence on nitrogen saturation of terrestrial ecosystem net primary productivity 1 2016
... 大气氮沉降是持续而缓慢的氮添加过程, 而大多数模拟氮沉降实验研究是通过生长季少次脉冲式的氮添加处理进行的(Bai et al., 2015; Fang et al., 2017; Chen et al., 2018; Wang et al., 2019c), 不同的氮添加模式和频次会影响生态系统结构和功能对氮添加的响应(Tu et al., 2013; Zhang et al., 2014).本平台已有的研究结果表明, 在氮添加处理早期, 不同施氮频率对植物地上生产力并没有显著影响, 而对物种多样性、地下生产力和土壤微生物群落的影响是显著的(Zhang et al., 2016b; Wang et al., 2019a; Cao et al., 2020).Zhang等(2016b)对氮添加处理前5年的研究发现随着氮添加量的增加, 植物物种多样性显著降低, 而与HF (每年12次)相比, LF (每年2次)处理导致物种多样性丧失的速率更快, 这主要是因为LF处理下较高的氨态氮浓度限制了新物种的定植.Wang等(2019b)在氮添加处理的第5-7年的研究表明, 虽然不同施氮频率并未显著影响植被ANPP, 但LF显著降低了BNPP, 特别是在低氮添加量处理下.Cao等(2020)对施氮后第6和7年的研究表明, HF会显著减弱氮添加处理对植物和土壤细菌多样性的负作用.本研究在氮添加处理第11和12年进行测定, 我们并未发现不同施氮频率对土壤呼吸及其组分影响有显著差别, 而同期两种施氮频率下植物生产力、土壤微生物生物量、土壤细菌和真菌生物量以及F/B均没有显著的差异(附录I, 附录II).这表明随着氮添加处理时间的延长, 不同施氮频率对植物和微生物群落生产力和结构的影响逐渐减弱.因为随着土壤中可利用氮的不断增加, 造成土壤酸化和有毒离子的积累, 生态系统逐渐呈现出氮饱和的状态(Tian et al., 2016), 进而逐步削弱了不同施氮频率对生态系统影响的差别.本研究的HF处理虽然尽可能地提高了氮添加频次, 但与缓慢持续的大气氮沉降过程仍存在较大差别, 使得我们在研究生态系统结构和功能对大气氮沉降的响应过程中仍存在很大不确定性.随着氮添加手段的改进, 如采用具亲水性包衣的缓释尿素等, 可以更精确地模拟大气氮沉降特别是湿沉降的过程, 为准确模拟大气氮沉降对生态系统碳氮循环过程的影响提供了可能. ...
Nitrogen additions and microbial biomass: a meta-analysis of ecosystem studies 2 2008
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
Nitrogen addition stimulates different components of soil respiration in a subtropical bamboo ecosystem 1 2013
... 大气氮沉降是持续而缓慢的氮添加过程, 而大多数模拟氮沉降实验研究是通过生长季少次脉冲式的氮添加处理进行的(Bai et al., 2015; Fang et al., 2017; Chen et al., 2018; Wang et al., 2019c), 不同的氮添加模式和频次会影响生态系统结构和功能对氮添加的响应(Tu et al., 2013; Zhang et al., 2014).本平台已有的研究结果表明, 在氮添加处理早期, 不同施氮频率对植物地上生产力并没有显著影响, 而对物种多样性、地下生产力和土壤微生物群落的影响是显著的(Zhang et al., 2016b; Wang et al., 2019a; Cao et al., 2020).Zhang等(2016b)对氮添加处理前5年的研究发现随着氮添加量的增加, 植物物种多样性显著降低, 而与HF (每年12次)相比, LF (每年2次)处理导致物种多样性丧失的速率更快, 这主要是因为LF处理下较高的氨态氮浓度限制了新物种的定植.Wang等(2019b)在氮添加处理的第5-7年的研究表明, 虽然不同施氮频率并未显著影响植被ANPP, 但LF显著降低了BNPP, 特别是在低氮添加量处理下.Cao等(2020)对施氮后第6和7年的研究表明, HF会显著减弱氮添加处理对植物和土壤细菌多样性的负作用.本研究在氮添加处理第11和12年进行测定, 我们并未发现不同施氮频率对土壤呼吸及其组分影响有显著差别, 而同期两种施氮频率下植物生产力、土壤微生物生物量、土壤细菌和真菌生物量以及F/B均没有显著的差异(附录I, 附录II).这表明随着氮添加处理时间的延长, 不同施氮频率对植物和微生物群落生产力和结构的影响逐渐减弱.因为随着土壤中可利用氮的不断增加, 造成土壤酸化和有毒离子的积累, 生态系统逐渐呈现出氮饱和的状态(Tian et al., 2016), 进而逐步削弱了不同施氮频率对生态系统影响的差别.本研究的HF处理虽然尽可能地提高了氮添加频次, 但与缓慢持续的大气氮沉降过程仍存在较大差别, 使得我们在研究生态系统结构和功能对大气氮沉降的响应过程中仍存在很大不确定性.随着氮添加手段的改进, 如采用具亲水性包衣的缓释尿素等, 可以更精确地模拟大气氮沉降特别是湿沉降的过程, 为准确模拟大气氮沉降对生态系统碳氮循环过程的影响提供了可能. ...
Asymmetry in above- and belowground productivity responses to N addition in a semi-arid temperate steppe 6 2019
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
... Rs对氮添加响应的强度和方向取决于Rh和Ra两个组分的响应, 本研究中氮添加显著降低了Rh, 对Ra并无显著影响.氮添加并没有影响Rh对Rs的贡献比例, Rh平均占Rs的62%, 这与本区域其他研究结果(Zhang et al., 2016a, 2019)一致.大部分陆地生态系统的生产力是受氮限制的, 通常氮添加会通过提高土壤氮可利用性而促进植物生长, 增加植物自养呼吸(Chen et al., 2015b).但土壤氮限制缓解的同时也会改变植物的地上地下碳分配策略, 减少植物对地下根系的碳分配比例, 该实验平台的研究表明在氮添加处理的第6和第7年, 氮添加使得ANPP增加了45%-53%, 但导致地下净初级生产力(BNPP)平均下降了40%-47% (Wang et al., 2019a).这可能是本研究中Ra对氮添加处理缺乏响应的原因.一项全球氮添加实验的meta分析表明, 氮添加会显著降低根系生物量和周转速率, 但对根系现存量没有影响(Peng et al., 2017), 由于Ra主要来自于现存根系呼吸, 这可以进一步解释为什么本研究中Ra对氮添加处理没有显著响应. ...
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
... 大气氮沉降是持续而缓慢的氮添加过程, 而大多数模拟氮沉降实验研究是通过生长季少次脉冲式的氮添加处理进行的(Bai et al., 2015; Fang et al., 2017; Chen et al., 2018; Wang et al., 2019c), 不同的氮添加模式和频次会影响生态系统结构和功能对氮添加的响应(Tu et al., 2013; Zhang et al., 2014).本平台已有的研究结果表明, 在氮添加处理早期, 不同施氮频率对植物地上生产力并没有显著影响, 而对物种多样性、地下生产力和土壤微生物群落的影响是显著的(Zhang et al., 2016b; Wang et al., 2019a; Cao et al., 2020).Zhang等(2016b)对氮添加处理前5年的研究发现随着氮添加量的增加, 植物物种多样性显著降低, 而与HF (每年12次)相比, LF (每年2次)处理导致物种多样性丧失的速率更快, 这主要是因为LF处理下较高的氨态氮浓度限制了新物种的定植.Wang等(2019b)在氮添加处理的第5-7年的研究表明, 虽然不同施氮频率并未显著影响植被ANPP, 但LF显著降低了BNPP, 特别是在低氮添加量处理下.Cao等(2020)对施氮后第6和7年的研究表明, HF会显著减弱氮添加处理对植物和土壤细菌多样性的负作用.本研究在氮添加处理第11和12年进行测定, 我们并未发现不同施氮频率对土壤呼吸及其组分影响有显著差别, 而同期两种施氮频率下植物生产力、土壤微生物生物量、土壤细菌和真菌生物量以及F/B均没有显著的差异(附录I, 附录II).这表明随着氮添加处理时间的延长, 不同施氮频率对植物和微生物群落生产力和结构的影响逐渐减弱.因为随着土壤中可利用氮的不断增加, 造成土壤酸化和有毒离子的积累, 生态系统逐渐呈现出氮饱和的状态(Tian et al., 2016), 进而逐步削弱了不同施氮频率对生态系统影响的差别.本研究的HF处理虽然尽可能地提高了氮添加频次, 但与缓慢持续的大气氮沉降过程仍存在较大差别, 使得我们在研究生态系统结构和功能对大气氮沉降的响应过程中仍存在很大不确定性.随着氮添加手段的改进, 如采用具亲水性包衣的缓释尿素等, 可以更精确地模拟大气氮沉降特别是湿沉降的过程, 为准确模拟大气氮沉降对生态系统碳氮循环过程的影响提供了可能. ...
Responses of soil respiration to nitrogen addition in the Sanjiang Plain wetland, northeastern China 2 2019
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... 大气氮沉降是持续而缓慢的氮添加过程, 而大多数模拟氮沉降实验研究是通过生长季少次脉冲式的氮添加处理进行的(Bai et al., 2015; Fang et al., 2017; Chen et al., 2018; Wang et al., 2019c), 不同的氮添加模式和频次会影响生态系统结构和功能对氮添加的响应(Tu et al., 2013; Zhang et al., 2014).本平台已有的研究结果表明, 在氮添加处理早期, 不同施氮频率对植物地上生产力并没有显著影响, 而对物种多样性、地下生产力和土壤微生物群落的影响是显著的(Zhang et al., 2016b; Wang et al., 2019a; Cao et al., 2020).Zhang等(2016b)对氮添加处理前5年的研究发现随着氮添加量的增加, 植物物种多样性显著降低, 而与HF (每年12次)相比, LF (每年2次)处理导致物种多样性丧失的速率更快, 这主要是因为LF处理下较高的氨态氮浓度限制了新物种的定植.Wang等(2019b)在氮添加处理的第5-7年的研究表明, 虽然不同施氮频率并未显著影响植被ANPP, 但LF显著降低了BNPP, 特别是在低氮添加量处理下.Cao等(2020)对施氮后第6和7年的研究表明, HF会显著减弱氮添加处理对植物和土壤细菌多样性的负作用.本研究在氮添加处理第11和12年进行测定, 我们并未发现不同施氮频率对土壤呼吸及其组分影响有显著差别, 而同期两种施氮频率下植物生产力、土壤微生物生物量、土壤细菌和真菌生物量以及F/B均没有显著的差异(附录I, 附录II).这表明随着氮添加处理时间的延长, 不同施氮频率对植物和微生物群落生产力和结构的影响逐渐减弱.因为随着土壤中可利用氮的不断增加, 造成土壤酸化和有毒离子的积累, 生态系统逐渐呈现出氮饱和的状态(Tian et al., 2016), 进而逐步削弱了不同施氮频率对生态系统影响的差别.本研究的HF处理虽然尽可能地提高了氮添加频次, 但与缓慢持续的大气氮沉降过程仍存在较大差别, 使得我们在研究生态系统结构和功能对大气氮沉降的响应过程中仍存在很大不确定性.随着氮添加手段的改进, 如采用具亲水性包衣的缓释尿素等, 可以更精确地模拟大气氮沉降特别是湿沉降的过程, 为准确模拟大气氮沉降对生态系统碳氮循环过程的影响提供了可能. ...
Nitrogen addition reduces soil respiration but increases the relative contribution of heterotrophic component in an alpine meadow 8 2019
... 全球陆地生态系统近3/4的碳(C)储存在土壤中(Masciandaro et al., 2018), 储量高达1 395-2 011 Pg C, 约为大气碳储量的2-4倍(Scharlemann et al., 2014; Bond-Lamberty et al., 2018).作为陆地生态系统第二大碳通量, 土壤呼吸每年释放的CO2为68-98 Pg C, 约为每年化石燃料燃烧释放CO2的10倍(Raich & Schlesinger, 1992; Bond-Lamberty & Thomson, 2010).全球碳模型的预测表明, 由于全球变 化引发的土壤呼吸的微小变动都会显著影响大气 CO2浓度, 进而影响全球碳平衡(Cox et al., 2000; Zhu et al., 2016; Wang et al., 2019c).土壤呼吸根据CO2产生的来源不同主要分为两个组分, 一是自养呼吸, 主要是来自于植物根呼吸以及菌根真菌呼吸产生的CO2; 另一部分是异养呼吸, 主要是来自于凋落物分解和微生物降解土壤有机质产生的CO2 (Hanson et al., 2000; Bond-Lamberty et al., 2004).由于底物来源和对环境因子响应的差异, 土壤呼吸的自养和异养组分对氮沉降等全球变化的响应存在显著差别(Zhou et al., 2014; Wei et al., 2016).因此, 为了准确预测全球变化对土壤呼吸及生态系统碳循环的影响, 就需要区分土壤呼吸的不同组分, 并研究各组分对全球变化因子的响应规律(Wang et al., 2019c), 这对评估整个生态系统的碳收支平衡是非常重要的(Lovett et al., 2006; ArchMiller & Samuelson, 2016). ...
... ).因此, 为了准确预测全球变化对土壤呼吸及生态系统碳循环的影响, 就需要区分土壤呼吸的不同组分, 并研究各组分对全球变化因子的响应规律(Wang et al., 2019c), 这对评估整个生态系统的碳收支平衡是非常重要的(Lovett et al., 2006; ArchMiller & Samuelson, 2016). ...
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
... 土壤呼吸的测量在2018-2019年生长季进行, 这是长期氮添加实验平台氮添加处理的第11和12年.由于测定仪器限制, 本研究仅选取了6个区组(重复)下的6个氮添加水平(0、2、5、10、20、50 g·m-2·a-1)和2个施氮频率进行测量.采用深浅环法对土壤呼吸组分进行区分(Zhou et al., 2007), 此方法近年来已被广泛应用于草原土壤呼吸组分区分(Yan et al., 2010; Zeng et al., 2018; Wang et al., 2019c; Zhang et al., 2019).已有研究表明, 该区域草原近85%以上的根系分布在0-30 cm土层, 30 cm的深环可以阻止大部分植物根系长入(Zhang et al., 2019).为了减少深环中残留细根分解对测量的影响, 在测量开始前一年(2017年7月), 在每个处理样方内分别嵌入一个30 cm的深环和一个10 cm的浅环, 用于土壤呼吸及其组分的测量, 呼吸环高出地面约3 cm.土壤呼吸环质地为PVC, 直径为20 cm.土壤呼吸环的安装尽量选择植物间隙, 土壤环中的少量植物会在安装土壤环的时候就进行去除, 测量期间每1-2周检查并将土壤环中的植物地上部分剪除.其中浅环用于测量土壤总呼吸速率(Rs), 深环用于测量土壤异养呼吸速率(Rh), 二者的差值用于估算土壤自养呼吸速率(Ra)(Zhang et al., 2019).土壤呼吸速率的测量采用LI-8100土壤呼吸自动测量仪(LI-COR, Lincoln, USA).2018-2019年生长季5-9月进行测量, 每两周测量一次, 每次测量的时间控制在8:30-14:00之间.土壤呼吸测量的同时对深环和浅环0-10 cm土层土壤温度(ST)和土壤含水量(SWC)的进行测量.ST和SWC分别采用LI-8100附带的土壤热电偶探头(LI-8100-201, LI-COR, Lincoln, USA)和TDR-300 (Spectrum Technologies, Plainfield, USA)进行测量.根据以往的研究, 浅环对土壤温度和水分并没有显著影响, 可以采用环外土壤的测量数据, 但深环由于降低了土壤水分的侧向交换, 会显著影响环内的土壤温度和水分条件(Zhang et al., 2019).为了减少土壤温度和水分测量时探针插入对环内土壤的扰动, 在3个区组的处理样方中又嵌入一个30 cm的深环, 专门用于深环内土壤温度和水分的测量. ...
... 土壤微生物生物量(MB)和土壤微生物功能群组成采用磷脂脂肪酸(PLFA)方法测定(Frosteg?rd et al., 2011).根据细菌的特异性脂肪酸(i15:0、i16:0、i17:0、18:1ω7、cy17:0和cy19:0)和真菌的特异性脂肪酸(18:2ω6, 9)的含量代表土壤中细菌和真菌的生物量(Wang et al., 2019c), 并计算真菌/细菌比(F/B). ...
... 氮是内蒙古半干旱草原生产力和碳交换过程的主要限制因子(Bai et al., 2010).本研究发现随着氮添加量的增加, Rs显著下降(图3), 相同的结果在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草原(Wei et al., 2018)中同样被报道.但是氮添加对Rs无显著影响(Fu et al., 2015)或有促进作用(Zhou et al., 2014, 2016)的结果也多次被报道.通过比较发现, 氮添加对Rs的正作用通常多出现在森林生态系统或短期的氮添加实验中(Zhou et al., 2014; Fu et al., 2015; Luo et al., 2016).而随着氮处理时间的延长,Rs对氮添加的正响应会逐渐减弱, 并转变成负响应(Zhou et al., 2014, 2016).这主要是因为在实验处理初期, 氮添加主要表现出对植物生长的促进作用, 但随着处理时间的延长, 氮添加所导致的土壤pH下降对植物和土壤微生物的负面作用逐渐增强(Janssens et al., 2010; Liang et al., 2019).本研究Rs的测定是在氮添加处理的第11和12年进行的, 历经了10年氮添加处理后, 土壤pH显著降低, 氮添加对Rs的抑制作用十分明显, 特别是当氮添加量大于10 g·m-2·a-1时, Rs显著降低了近22.9 %.Bai等(2010)在本区域的长期氮添加实验研究表明, 10.5 g·m-2·a-1氮添加量是影响内蒙古半干旱草原土壤氮有效性显著增加的关键阈值. ...
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
... 大气氮沉降是持续而缓慢的氮添加过程, 而大多数模拟氮沉降实验研究是通过生长季少次脉冲式的氮添加处理进行的(Bai et al., 2015; Fang et al., 2017; Chen et al., 2018; Wang et al., 2019c), 不同的氮添加模式和频次会影响生态系统结构和功能对氮添加的响应(Tu et al., 2013; Zhang et al., 2014).本平台已有的研究结果表明, 在氮添加处理早期, 不同施氮频率对植物地上生产力并没有显著影响, 而对物种多样性、地下生产力和土壤微生物群落的影响是显著的(Zhang et al., 2016b; Wang et al., 2019a; Cao et al., 2020).Zhang等(2016b)对氮添加处理前5年的研究发现随着氮添加量的增加, 植物物种多样性显著降低, 而与HF (每年12次)相比, LF (每年2次)处理导致物种多样性丧失的速率更快, 这主要是因为LF处理下较高的氨态氮浓度限制了新物种的定植.Wang等(2019b)在氮添加处理的第5-7年的研究表明, 虽然不同施氮频率并未显著影响植被ANPP, 但LF显著降低了BNPP, 特别是在低氮添加量处理下.Cao等(2020)对施氮后第6和7年的研究表明, HF会显著减弱氮添加处理对植物和土壤细菌多样性的负作用.本研究在氮添加处理第11和12年进行测定, 我们并未发现不同施氮频率对土壤呼吸及其组分影响有显著差别, 而同期两种施氮频率下植物生产力、土壤微生物生物量、土壤细菌和真菌生物量以及F/B均没有显著的差异(附录I, 附录II).这表明随着氮添加处理时间的延长, 不同施氮频率对植物和微生物群落生产力和结构的影响逐渐减弱.因为随着土壤中可利用氮的不断增加, 造成土壤酸化和有毒离子的积累, 生态系统逐渐呈现出氮饱和的状态(Tian et al., 2016), 进而逐步削弱了不同施氮频率对生态系统影响的差别.本研究的HF处理虽然尽可能地提高了氮添加频次, 但与缓慢持续的大气氮沉降过程仍存在较大差别, 使得我们在研究生态系统结构和功能对大气氮沉降的响应过程中仍存在很大不确定性.随着氮添加手段的改进, 如采用具亲水性包衣的缓释尿素等, 可以更精确地模拟大气氮沉降特别是湿沉降的过程, 为准确模拟大气氮沉降对生态系统碳氮循环过程的影响提供了可能. ...
Effect of clipping on soil respiration components in temperate grassland of Loess Plateau 1 2016
... 全球陆地生态系统近3/4的碳(C)储存在土壤中(Masciandaro et al., 2018), 储量高达1 395-2 011 Pg C, 约为大气碳储量的2-4倍(Scharlemann et al., 2014; Bond-Lamberty et al., 2018).作为陆地生态系统第二大碳通量, 土壤呼吸每年释放的CO2为68-98 Pg C, 约为每年化石燃料燃烧释放CO2的10倍(Raich & Schlesinger, 1992; Bond-Lamberty & Thomson, 2010).全球碳模型的预测表明, 由于全球变 化引发的土壤呼吸的微小变动都会显著影响大气 CO2浓度, 进而影响全球碳平衡(Cox et al., 2000; Zhu et al., 2016; Wang et al., 2019c).土壤呼吸根据CO2产生的来源不同主要分为两个组分, 一是自养呼吸, 主要是来自于植物根呼吸以及菌根真菌呼吸产生的CO2; 另一部分是异养呼吸, 主要是来自于凋落物分解和微生物降解土壤有机质产生的CO2 (Hanson et al., 2000; Bond-Lamberty et al., 2004).由于底物来源和对环境因子响应的差异, 土壤呼吸的自养和异养组分对氮沉降等全球变化的响应存在显著差别(Zhou et al., 2014; Wei et al., 2016).因此, 为了准确预测全球变化对土壤呼吸及生态系统碳循环的影响, 就需要区分土壤呼吸的不同组分, 并研究各组分对全球变化因子的响应规律(Wang et al., 2019c), 这对评估整个生态系统的碳收支平衡是非常重要的(Lovett et al., 2006; ArchMiller & Samuelson, 2016). ...
Nitrogen addition decreased soil respiration and its components in a longterm fenced grassland on the Loess Plateau 2 2018
... 氮是内蒙古半干旱草原生产力和碳交换过程的主要限制因子(Bai et al., 2010).本研究发现随着氮添加量的增加, Rs显著下降(图3), 相同的结果在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草原(Wei et al., 2018)中同样被报道.但是氮添加对Rs无显著影响(Fu et al., 2015)或有促进作用(Zhou et al., 2014, 2016)的结果也多次被报道.通过比较发现, 氮添加对Rs的正作用通常多出现在森林生态系统或短期的氮添加实验中(Zhou et al., 2014; Fu et al., 2015; Luo et al., 2016).而随着氮处理时间的延长,Rs对氮添加的正响应会逐渐减弱, 并转变成负响应(Zhou et al., 2014, 2016).这主要是因为在实验处理初期, 氮添加主要表现出对植物生长的促进作用, 但随着处理时间的延长, 氮添加所导致的土壤pH下降对植物和土壤微生物的负面作用逐渐增强(Janssens et al., 2010; Liang et al., 2019).本研究Rs的测定是在氮添加处理的第11和12年进行的, 历经了10年氮添加处理后, 土壤pH显著降低, 氮添加对Rs的抑制作用十分明显, 特别是当氮添加量大于10 g·m-2·a-1时, Rs显著降低了近22.9 %.Bai等(2010)在本区域的长期氮添加实验研究表明, 10.5 g·m-2·a-1氮添加量是影响内蒙古半干旱草原土壤氮有效性显著增加的关键阈值. ...
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
Soil acidification reduces the effects of short-term nutrient enrichment on plant and soil biota and their interactions in grasslands 1 2020
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Differential responses of auto- and heterotrophic soil respiration to water and nitrogen addition in a semiarid temperate steppe 2 2010
... 土壤呼吸的测量在2018-2019年生长季进行, 这是长期氮添加实验平台氮添加处理的第11和12年.由于测定仪器限制, 本研究仅选取了6个区组(重复)下的6个氮添加水平(0、2、5、10、20、50 g·m-2·a-1)和2个施氮频率进行测量.采用深浅环法对土壤呼吸组分进行区分(Zhou et al., 2007), 此方法近年来已被广泛应用于草原土壤呼吸组分区分(Yan et al., 2010; Zeng et al., 2018; Wang et al., 2019c; Zhang et al., 2019).已有研究表明, 该区域草原近85%以上的根系分布在0-30 cm土层, 30 cm的深环可以阻止大部分植物根系长入(Zhang et al., 2019).为了减少深环中残留细根分解对测量的影响, 在测量开始前一年(2017年7月), 在每个处理样方内分别嵌入一个30 cm的深环和一个10 cm的浅环, 用于土壤呼吸及其组分的测量, 呼吸环高出地面约3 cm.土壤呼吸环质地为PVC, 直径为20 cm.土壤呼吸环的安装尽量选择植物间隙, 土壤环中的少量植物会在安装土壤环的时候就进行去除, 测量期间每1-2周检查并将土壤环中的植物地上部分剪除.其中浅环用于测量土壤总呼吸速率(Rs), 深环用于测量土壤异养呼吸速率(Rh), 二者的差值用于估算土壤自养呼吸速率(Ra)(Zhang et al., 2019).土壤呼吸速率的测量采用LI-8100土壤呼吸自动测量仪(LI-COR, Lincoln, USA).2018-2019年生长季5-9月进行测量, 每两周测量一次, 每次测量的时间控制在8:30-14:00之间.土壤呼吸测量的同时对深环和浅环0-10 cm土层土壤温度(ST)和土壤含水量(SWC)的进行测量.ST和SWC分别采用LI-8100附带的土壤热电偶探头(LI-8100-201, LI-COR, Lincoln, USA)和TDR-300 (Spectrum Technologies, Plainfield, USA)进行测量.根据以往的研究, 浅环对土壤温度和水分并没有显著影响, 可以采用环外土壤的测量数据, 但深环由于降低了土壤水分的侧向交换, 会显著影响环内的土壤温度和水分条件(Zhang et al., 2019).为了减少土壤温度和水分测量时探针插入对环内土壤的扰动, 在3个区组的处理样方中又嵌入一个30 cm的深环, 专门用于深环内土壤温度和水分的测量. ...
Response of soil respiration to nitrogen addition in two subtropical forest types 1 2020
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
氮添加对内蒙古退化草地植物群落多样性和生物量的影响 1 2018
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
氮添加对内蒙古退化草地植物群落多样性和生物量的影响 1 2018
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
Soil respiration and its autotrophic and heterotrophic components in response to nitrogen addition among different degraded temperate grasslands 1 2018
... 土壤呼吸的测量在2018-2019年生长季进行, 这是长期氮添加实验平台氮添加处理的第11和12年.由于测定仪器限制, 本研究仅选取了6个区组(重复)下的6个氮添加水平(0、2、5、10、20、50 g·m-2·a-1)和2个施氮频率进行测量.采用深浅环法对土壤呼吸组分进行区分(Zhou et al., 2007), 此方法近年来已被广泛应用于草原土壤呼吸组分区分(Yan et al., 2010; Zeng et al., 2018; Wang et al., 2019c; Zhang et al., 2019).已有研究表明, 该区域草原近85%以上的根系分布在0-30 cm土层, 30 cm的深环可以阻止大部分植物根系长入(Zhang et al., 2019).为了减少深环中残留细根分解对测量的影响, 在测量开始前一年(2017年7月), 在每个处理样方内分别嵌入一个30 cm的深环和一个10 cm的浅环, 用于土壤呼吸及其组分的测量, 呼吸环高出地面约3 cm.土壤呼吸环质地为PVC, 直径为20 cm.土壤呼吸环的安装尽量选择植物间隙, 土壤环中的少量植物会在安装土壤环的时候就进行去除, 测量期间每1-2周检查并将土壤环中的植物地上部分剪除.其中浅环用于测量土壤总呼吸速率(Rs), 深环用于测量土壤异养呼吸速率(Rh), 二者的差值用于估算土壤自养呼吸速率(Ra)(Zhang et al., 2019).土壤呼吸速率的测量采用LI-8100土壤呼吸自动测量仪(LI-COR, Lincoln, USA).2018-2019年生长季5-9月进行测量, 每两周测量一次, 每次测量的时间控制在8:30-14:00之间.土壤呼吸测量的同时对深环和浅环0-10 cm土层土壤温度(ST)和土壤含水量(SWC)的进行测量.ST和SWC分别采用LI-8100附带的土壤热电偶探头(LI-8100-201, LI-COR, Lincoln, USA)和TDR-300 (Spectrum Technologies, Plainfield, USA)进行测量.根据以往的研究, 浅环对土壤温度和水分并没有显著影响, 可以采用环外土壤的测量数据, 但深环由于降低了土壤水分的侧向交换, 会显著影响环内的土壤温度和水分条件(Zhang et al., 2019).为了减少土壤温度和水分测量时探针插入对环内土壤的扰动, 在3个区组的处理样方中又嵌入一个30 cm的深环, 专门用于深环内土壤温度和水分的测量. ...
Arbuscular mycorrhizal fungi regulate soil respiration and its response to precipitation change in a semiarid steppe 1 2016
... Rs对氮添加响应的强度和方向取决于Rh和Ra两个组分的响应, 本研究中氮添加显著降低了Rh, 对Ra并无显著影响.氮添加并没有影响Rh对Rs的贡献比例, Rh平均占Rs的62%, 这与本区域其他研究结果(Zhang et al., 2016a, 2019)一致.大部分陆地生态系统的生产力是受氮限制的, 通常氮添加会通过提高土壤氮可利用性而促进植物生长, 增加植物自养呼吸(Chen et al., 2015b).但土壤氮限制缓解的同时也会改变植物的地上地下碳分配策略, 减少植物对地下根系的碳分配比例, 该实验平台的研究表明在氮添加处理的第6和第7年, 氮添加使得ANPP增加了45%-53%, 但导致地下净初级生产力(BNPP)平均下降了40%-47% (Wang et al., 2019a).这可能是本研究中Ra对氮添加处理缺乏响应的原因.一项全球氮添加实验的meta分析表明, 氮添加会显著降低根系生物量和周转速率, 但对根系现存量没有影响(Peng et al., 2017), 由于Ra主要来自于现存根系呼吸, 这可以进一步解释为什么本研究中Ra对氮添加处理没有显著响应. ...
Changing precipitation exerts greater influence on soil heterotrophic than autotrophic respiration in a semiarid steppe 7 2019
... 土壤呼吸的测量在2018-2019年生长季进行, 这是长期氮添加实验平台氮添加处理的第11和12年.由于测定仪器限制, 本研究仅选取了6个区组(重复)下的6个氮添加水平(0、2、5、10、20、50 g·m-2·a-1)和2个施氮频率进行测量.采用深浅环法对土壤呼吸组分进行区分(Zhou et al., 2007), 此方法近年来已被广泛应用于草原土壤呼吸组分区分(Yan et al., 2010; Zeng et al., 2018; Wang et al., 2019c; Zhang et al., 2019).已有研究表明, 该区域草原近85%以上的根系分布在0-30 cm土层, 30 cm的深环可以阻止大部分植物根系长入(Zhang et al., 2019).为了减少深环中残留细根分解对测量的影响, 在测量开始前一年(2017年7月), 在每个处理样方内分别嵌入一个30 cm的深环和一个10 cm的浅环, 用于土壤呼吸及其组分的测量, 呼吸环高出地面约3 cm.土壤呼吸环质地为PVC, 直径为20 cm.土壤呼吸环的安装尽量选择植物间隙, 土壤环中的少量植物会在安装土壤环的时候就进行去除, 测量期间每1-2周检查并将土壤环中的植物地上部分剪除.其中浅环用于测量土壤总呼吸速率(Rs), 深环用于测量土壤异养呼吸速率(Rh), 二者的差值用于估算土壤自养呼吸速率(Ra)(Zhang et al., 2019).土壤呼吸速率的测量采用LI-8100土壤呼吸自动测量仪(LI-COR, Lincoln, USA).2018-2019年生长季5-9月进行测量, 每两周测量一次, 每次测量的时间控制在8:30-14:00之间.土壤呼吸测量的同时对深环和浅环0-10 cm土层土壤温度(ST)和土壤含水量(SWC)的进行测量.ST和SWC分别采用LI-8100附带的土壤热电偶探头(LI-8100-201, LI-COR, Lincoln, USA)和TDR-300 (Spectrum Technologies, Plainfield, USA)进行测量.根据以往的研究, 浅环对土壤温度和水分并没有显著影响, 可以采用环外土壤的测量数据, 但深环由于降低了土壤水分的侧向交换, 会显著影响环内的土壤温度和水分条件(Zhang et al., 2019).为了减少土壤温度和水分测量时探针插入对环内土壤的扰动, 在3个区组的处理样方中又嵌入一个30 cm的深环, 专门用于深环内土壤温度和水分的测量. ...
... Rs对氮添加响应的强度和方向取决于Rh和Ra两个组分的响应, 本研究中氮添加显著降低了Rh, 对Ra并无显著影响.氮添加并没有影响Rh对Rs的贡献比例, Rh平均占Rs的62%, 这与本区域其他研究结果(Zhang et al., 2016a, 2019)一致.大部分陆地生态系统的生产力是受氮限制的, 通常氮添加会通过提高土壤氮可利用性而促进植物生长, 增加植物自养呼吸(Chen et al., 2015b).但土壤氮限制缓解的同时也会改变植物的地上地下碳分配策略, 减少植物对地下根系的碳分配比例, 该实验平台的研究表明在氮添加处理的第6和第7年, 氮添加使得ANPP增加了45%-53%, 但导致地下净初级生产力(BNPP)平均下降了40%-47% (Wang et al., 2019a).这可能是本研究中Ra对氮添加处理缺乏响应的原因.一项全球氮添加实验的meta分析表明, 氮添加会显著降低根系生物量和周转速率, 但对根系现存量没有影响(Peng et al., 2017), 由于Ra主要来自于现存根系呼吸, 这可以进一步解释为什么本研究中Ra对氮添加处理没有显著响应. ...
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
Rapid plant species loss at high rates and at low frequency of N addition in temperate steppe 4 2014
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
... 本研究样地位于内蒙古锡林郭勒盟的内蒙古草原生态系统国家定位研究站(116.70° E, 43.38° N, 海拔1 200 m).该地区属于温带半干旱大陆性气候, 长期(1983-2012年)的气象数据表明, 该区域的年平均气温为0.9 ℃, 月平均气温变化从最冷月(1月) -21.7 ℃到最暖月(7月) 19.7 ℃.年降水量为355.3 mm, 其中60%-80%的降水量集中在生长季(5-8月)(Zhang et al., 2014).该区域主要土壤类型为栗钙土.地带性植被为温带半干旱典型草原, 优势物种为大针茅(Stipa grandis)、羊草(Leymus chinensis)等.该区域大气氮沉降的背景值为1.8 g·m-2·a-1, 其中干、湿氮沉降分别为1.2和0.6 g·m-2·a-1 (Giese et al., 2013). ...
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
... 氮是内蒙古半干旱草原生产力和碳交换过程的主要限制因子(Bai et al., 2010).本研究发现随着氮添加量的增加, Rs显著下降(图3), 相同的结果在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草原(Wei et al., 2018)中同样被报道.但是氮添加对Rs无显著影响(Fu et al., 2015)或有促进作用(Zhou et al., 2014, 2016)的结果也多次被报道.通过比较发现, 氮添加对Rs的正作用通常多出现在森林生态系统或短期的氮添加实验中(Zhou et al., 2014; Fu et al., 2015; Luo et al., 2016).而随着氮处理时间的延长,Rs对氮添加的正响应会逐渐减弱, 并转变成负响应(Zhou et al., 2014, 2016).这主要是因为在实验处理初期, 氮添加主要表现出对植物生长的促进作用, 但随着处理时间的延长, 氮添加所导致的土壤pH下降对植物和土壤微生物的负面作用逐渐增强(Janssens et al., 2010; Liang et al., 2019).本研究Rs的测定是在氮添加处理的第11和12年进行的, 历经了10年氮添加处理后, 土壤pH显著降低, 氮添加对Rs的抑制作用十分明显, 特别是当氮添加量大于10 g·m-2·a-1时, Rs显著降低了近22.9 %.Bai等(2010)在本区域的长期氮添加实验研究表明, 10.5 g·m-2·a-1氮添加量是影响内蒙古半干旱草原土壤氮有效性显著增加的关键阈值. ...
... 大气氮沉降对生态系统的影响是缓慢而持续的(Galloway et al., 2008; Smith et al., 2009).目前绝大多数的研究是采用每年1-2次施氮的方式模拟氮沉降(Clark & Tilman, 2008; Fornara & Tilman, 2012; Bai et al., 2015 ; Chen et al., 2018).虽然少数研究已经增加了氮添加的次数如每年4-6次施氮(Fang et al., 2017; Ma et al., 2018; Wang et al., 2019c), 但是这种脉冲式的氮添加方式并不能真实模拟大气氮沉降过程, 并使得研究结果可能出现偏差.Zhang等(2014)的研究表明, 相比于每年12次施氮, 每年2次氮添加会导致植物丰富度下降得更加显著, 从而高估了大气氮沉降对植物群落的影响.同时Wang等(2019a)发现虽然不同施氮频率并未显著影响植被地上生产力, 但低频氮添加显著降低了地下生产力.周纪东等(2016)发现不同的氮添加频率对温带半干旱草原土壤有机碳含量没有显著影响.而关于土壤微生物群落的研究表明, 相比于12次施氮, 每年2次氮添加会导致细菌多样性显著下降(Cao et al., 2020).考虑到施氮频率对土壤微生物的影响, 那么高频氮添加是否会减弱氮对土壤呼吸及其组分的影响呢?目前关于不同氮添加频率对土壤呼吸影响的差异鲜有报道.因此, 有必要通过对比不同氮添加频率处理对生态系统影响的差异, 来评估以往的低频氮添加实验在研究生态系统结构和功能对氮沉降响应效应时所造成的潜在误差. ...
... 氮是内蒙古半干旱草原生产力和碳交换过程的主要限制因子(Bai et al., 2010).本研究发现随着氮添加量的增加, Rs显著下降(图3), 相同的结果在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草原(Wei et al., 2018)中同样被报道.但是氮添加对Rs无显著影响(Fu et al., 2015)或有促进作用(Zhou et al., 2014, 2016)的结果也多次被报道.通过比较发现, 氮添加对Rs的正作用通常多出现在森林生态系统或短期的氮添加实验中(Zhou et al., 2014; Fu et al., 2015; Luo et al., 2016).而随着氮处理时间的延长,Rs对氮添加的正响应会逐渐减弱, 并转变成负响应(Zhou et al., 2014, 2016).这主要是因为在实验处理初期, 氮添加主要表现出对植物生长的促进作用, 但随着处理时间的延长, 氮添加所导致的土壤pH下降对植物和土壤微生物的负面作用逐渐增强(Janssens et al., 2010; Liang et al., 2019).本研究Rs的测定是在氮添加处理的第11和12年进行的, 历经了10年氮添加处理后, 土壤pH显著降低, 氮添加对Rs的抑制作用十分明显, 特别是当氮添加量大于10 g·m-2·a-1时, Rs显著降低了近22.9 %.Bai等(2010)在本区域的长期氮添加实验研究表明, 10.5 g·m-2·a-1氮添加量是影响内蒙古半干旱草原土壤氮有效性显著增加的关键阈值. ...
Interactive effects of global change factors on soil respiration and its components: a meta-analysis 2016
Different responses of soil respiration and its components to nitrogen addition among biomes: a meta-analysis 7 2014
... 全球陆地生态系统近3/4的碳(C)储存在土壤中(Masciandaro et al., 2018), 储量高达1 395-2 011 Pg C, 约为大气碳储量的2-4倍(Scharlemann et al., 2014; Bond-Lamberty et al., 2018).作为陆地生态系统第二大碳通量, 土壤呼吸每年释放的CO2为68-98 Pg C, 约为每年化石燃料燃烧释放CO2的10倍(Raich & Schlesinger, 1992; Bond-Lamberty & Thomson, 2010).全球碳模型的预测表明, 由于全球变 化引发的土壤呼吸的微小变动都会显著影响大气 CO2浓度, 进而影响全球碳平衡(Cox et al., 2000; Zhu et al., 2016; Wang et al., 2019c).土壤呼吸根据CO2产生的来源不同主要分为两个组分, 一是自养呼吸, 主要是来自于植物根呼吸以及菌根真菌呼吸产生的CO2; 另一部分是异养呼吸, 主要是来自于凋落物分解和微生物降解土壤有机质产生的CO2 (Hanson et al., 2000; Bond-Lamberty et al., 2004).由于底物来源和对环境因子响应的差异, 土壤呼吸的自养和异养组分对氮沉降等全球变化的响应存在显著差别(Zhou et al., 2014; Wei et al., 2016).因此, 为了准确预测全球变化对土壤呼吸及生态系统碳循环的影响, 就需要区分土壤呼吸的不同组分, 并研究各组分对全球变化因子的响应规律(Wang et al., 2019c), 这对评估整个生态系统的碳收支平衡是非常重要的(Lovett et al., 2006; ArchMiller & Samuelson, 2016). ...
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... 氮是内蒙古半干旱草原生产力和碳交换过程的主要限制因子(Bai et al., 2010).本研究发现随着氮添加量的增加, Rs显著下降(图3), 相同的结果在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草原(Wei et al., 2018)中同样被报道.但是氮添加对Rs无显著影响(Fu et al., 2015)或有促进作用(Zhou et al., 2014, 2016)的结果也多次被报道.通过比较发现, 氮添加对Rs的正作用通常多出现在森林生态系统或短期的氮添加实验中(Zhou et al., 2014; Fu et al., 2015; Luo et al., 2016).而随着氮处理时间的延长,Rs对氮添加的正响应会逐渐减弱, 并转变成负响应(Zhou et al., 2014, 2016).这主要是因为在实验处理初期, 氮添加主要表现出对植物生长的促进作用, 但随着处理时间的延长, 氮添加所导致的土壤pH下降对植物和土壤微生物的负面作用逐渐增强(Janssens et al., 2010; Liang et al., 2019).本研究Rs的测定是在氮添加处理的第11和12年进行的, 历经了10年氮添加处理后, 土壤pH显著降低, 氮添加对Rs的抑制作用十分明显, 特别是当氮添加量大于10 g·m-2·a-1时, Rs显著降低了近22.9 %.Bai等(2010)在本区域的长期氮添加实验研究表明, 10.5 g·m-2·a-1氮添加量是影响内蒙古半干旱草原土壤氮有效性显著增加的关键阈值. ...
... 的正作用通常多出现在森林生态系统或短期的氮添加实验中(Zhou et al., 2014; Fu et al., 2015; Luo et al., 2016).而随着氮处理时间的延长,Rs对氮添加的正响应会逐渐减弱, 并转变成负响应(Zhou et al., 2014, 2016).这主要是因为在实验处理初期, 氮添加主要表现出对植物生长的促进作用, 但随着处理时间的延长, 氮添加所导致的土壤pH下降对植物和土壤微生物的负面作用逐渐增强(Janssens et al., 2010; Liang et al., 2019).本研究Rs的测定是在氮添加处理的第11和12年进行的, 历经了10年氮添加处理后, 土壤pH显著降低, 氮添加对Rs的抑制作用十分明显, 特别是当氮添加量大于10 g·m-2·a-1时, Rs显著降低了近22.9 %.Bai等(2010)在本区域的长期氮添加实验研究表明, 10.5 g·m-2·a-1氮添加量是影响内蒙古半干旱草原土壤氮有效性显著增加的关键阈值. ...
... 对氮添加的正响应会逐渐减弱, 并转变成负响应(Zhou et al., 2014, 2016).这主要是因为在实验处理初期, 氮添加主要表现出对植物生长的促进作用, 但随着处理时间的延长, 氮添加所导致的土壤pH下降对植物和土壤微生物的负面作用逐渐增强(Janssens et al., 2010; Liang et al., 2019).本研究Rs的测定是在氮添加处理的第11和12年进行的, 历经了10年氮添加处理后, 土壤pH显著降低, 氮添加对Rs的抑制作用十分明显, 特别是当氮添加量大于10 g·m-2·a-1时, Rs显著降低了近22.9 %.Bai等(2010)在本区域的长期氮添加实验研究表明, 10.5 g·m-2·a-1氮添加量是影响内蒙古半干旱草原土壤氮有效性显著增加的关键阈值. ...
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
Source components and interannual variability of soil CO2 efflux under experimental warming and clipping in a grassland ecosystem 1 2007
... 土壤呼吸的测量在2018-2019年生长季进行, 这是长期氮添加实验平台氮添加处理的第11和12年.由于测定仪器限制, 本研究仅选取了6个区组(重复)下的6个氮添加水平(0、2、5、10、20、50 g·m-2·a-1)和2个施氮频率进行测量.采用深浅环法对土壤呼吸组分进行区分(Zhou et al., 2007), 此方法近年来已被广泛应用于草原土壤呼吸组分区分(Yan et al., 2010; Zeng et al., 2018; Wang et al., 2019c; Zhang et al., 2019).已有研究表明, 该区域草原近85%以上的根系分布在0-30 cm土层, 30 cm的深环可以阻止大部分植物根系长入(Zhang et al., 2019).为了减少深环中残留细根分解对测量的影响, 在测量开始前一年(2017年7月), 在每个处理样方内分别嵌入一个30 cm的深环和一个10 cm的浅环, 用于土壤呼吸及其组分的测量, 呼吸环高出地面约3 cm.土壤呼吸环质地为PVC, 直径为20 cm.土壤呼吸环的安装尽量选择植物间隙, 土壤环中的少量植物会在安装土壤环的时候就进行去除, 测量期间每1-2周检查并将土壤环中的植物地上部分剪除.其中浅环用于测量土壤总呼吸速率(Rs), 深环用于测量土壤异养呼吸速率(Rh), 二者的差值用于估算土壤自养呼吸速率(Ra)(Zhang et al., 2019).土壤呼吸速率的测量采用LI-8100土壤呼吸自动测量仪(LI-COR, Lincoln, USA).2018-2019年生长季5-9月进行测量, 每两周测量一次, 每次测量的时间控制在8:30-14:00之间.土壤呼吸测量的同时对深环和浅环0-10 cm土层土壤温度(ST)和土壤含水量(SWC)的进行测量.ST和SWC分别采用LI-8100附带的土壤热电偶探头(LI-8100-201, LI-COR, Lincoln, USA)和TDR-300 (Spectrum Technologies, Plainfield, USA)进行测量.根据以往的研究, 浅环对土壤温度和水分并没有显著影响, 可以采用环外土壤的测量数据, 但深环由于降低了土壤水分的侧向交换, 会显著影响环内的土壤温度和水分条件(Zhang et al., 2019).为了减少土壤温度和水分测量时探针插入对环内土壤的扰动, 在3个区组的处理样方中又嵌入一个30 cm的深环, 专门用于深环内土壤温度和水分的测量. ...
Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality 3 2020
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... , 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
Patterns and mechanisms of responses by soil microbial communities to nitrogen addition 2 2017
... 工业革命以来, 人类活动特别是化石燃料的燃烧和氮肥的大量使用, 导致了全球CO2浓度和大气氮沉降速率急剧上升(Galloway et al., 2008; Mo et al., 2008).从1980到2010年, 我国大气氮沉降量持续增加了近60%, 达到近0.8 g·m-2 (Liu et al., 2013).预计到21世纪末, 全球大气氮沉降量将是目前的2.5倍(Galloway et al., 2004; Luo et al., 2016).大气氮沉降已经对生态系统的结构和功能产生了显著影响(Song et al., 2019; Chen et al., 2020).已有的氮添加实验表明氮添加可以显著促进植被生产力(杨倩等, 2018; Song et al., 2019; Wang et al., 2019a), 但降低了土壤微生物生物量及其活性(Xiao et al., 2020; Zhou et al., 2020).植物和微生物对氮沉降的不同响应将直接导致土壤呼吸及其自养和异养组分对氮沉降响应的差异(Janssens et al., 2010; Zhou et al., 2017, 2020).虽然通过氮添加实验, 研究者们已经对生态系统碳交换过程对氮沉降的响应方面进行了大量研究(Luca et al., 2006; Niu et al., 2010; Jia et al., 2012; Shi et al., 2019; Bai et al., 2020), 但关于氮沉降对土壤呼吸及其组分的影响仍存在争议.不同研究表明, 氮添加可以提高(Craine et al., 2001; Hasselquist et al., 2012; Jia et al., 2012; Wang et al., 2019b)或抑制(Mo et al., 2008; Janssens et al., 2010; 李寅龙等, 2015; Chen et al., 2016; Song et al., 2019; Yan et al., 2020)土壤呼吸或对其没有影响(Lee & Jose, 2003; Allison et al., 2008; 珊丹等, 2009).Zhou等(2014)整合分析了54个草地氮添加实验的结果, 发现氮添加对土壤总呼吸表现为促进作用, 平均提高7.8%, 这主要是由于氮添加对自养呼吸组分的提高, 而氮添加对异养呼吸组分均表现出抑制作用.此外, 随着氮添加时间的延长, 土壤呼吸对氮添加响应的强度显著下降, 这一趋势在森林生态系统中表现得最为明显(Zhou et al., 2014).这是因为在实验处理初期, 氮添加显著促进植物生长并增加土壤碳输入, 促进土壤自养呼吸, 并使得土壤总呼吸表现出增加趋势(Fang et al., 2018); 而在长期氮添加情况下, 由于土壤pH等条件的改变, 导致土壤微生物群落结构发生变化, 进而对土壤异养呼吸的抑制作用增加(Treseder, 2008).但目前由于在自然生态系统中持续10年以上的长期氮添加平台数据仍然十分有限, 使得我们还难以对草原生态系统土壤呼吸及其组分对长期氮添加的响应进行评估. ...
... 本研究中Rs对氮添加的响应主要取决于其Rh的变化, 随着氮添加量的增加, Rh显著降低(图3).在高寒草甸(Wang et al., 2019c)和黄土高原半干旱草地(Wei et al., 2018)也有同样发现.本研究发现Rh对氮添加量非常敏感, 在低浓度的氮添加量(2 g·m-2·a-1)处理下就出现显著的下降, 而这一氮添加量接近当地大气氮沉降量1.8 g·m-2·a-1 (Giese et al., 2013).Rh对氮添加的负响应主要有以下几个原因.首先, 地下碳分配比例的下降减少了根际分泌物, 降低了土壤有机质的降解(Janssens et al., 2010).根际是植物-微生物碳交换的重要界面, 而根际分泌物是微生物分解底物的重要来源.地下碳输入量的下降会显著降低土壤微生物有机碳降解速率.本研究中土壤呼吸测定年份并没有进行BNPP的测量, 但氮添加处理早期的数据已表明植物将更多的碳分配至地上, BNPP显著降低(Wang et al., 2019a).因此本研究中土壤呼吸及其组分与ANPP之间显著的负相关关系可能是由于氮沉降促进植物地上生长, 而减少向地下的碳分配, 抑制植物根系生长, 进而导致土壤呼吸速率降低(图5).第二, 氮添加显著降低了土壤微生物生物量和活性, 改变土壤微生物群落结构和功能.多个氮添加实验集成分析研究均表明, 土壤氮有效性的提高显著降低了土壤微生物生物量(Janssens et al., 2010; Zhou et al., 2017).这一方面是因为氮供给增加使得土壤氮同化所需能量降低(Bowden et al., 2004), 另一方面是因为氮添加导致土壤pH值降低, 有害金属离子富集(Tian & Niu, 2015), 限制了微生物生长和活性(Treseder, 2008; Chen et al., 2019).氮添加不仅降低土壤微生物生物量, 还会改变微生物群落结构(Chen et al., 2015a; Bowden et al., 2019; Zhou et al., 2020).本研究中, 长期氮添加导致土壤pH显著下降, 由对照处理下的7.55下降至高氮添加处理下的4.72, 降低了近37.5%.氮添加不但显著降低了土壤细菌和真菌的生物量, 同时还降低了微生物的F/B (附录I).而土壤总呼吸及其异养组分均与土壤pH、土壤微生物生物量和F/B表现出显著的线性正相关关系(图4, 图5).第三, 氮添加通过改变植被生产力和群落结构, 影响土壤微环境.本研究表明氮添加显著增加了植被高度和生物量, 并通过降低地表辐射强度和提高植物蒸腾作用, 降低了表层土壤温度和含水量(图1).但在高氮添加处理下, 土壤含水量出现了显著回升, 这主要是大量的氮输入提高了植物盖度, 降低了土壤温度, 使得土壤水分蒸发作用下降.与以往研究(Zhou et al., 2014)相似, 在本研究中土壤温度和水分是调控土壤呼吸及其组分变化的重要环境因子, 随着土壤温度和含水量的增加, Rs和Rh均显著提高.此外, 氮添加显著降低了Rs和Rh的水分敏感性, 这主要与氮添加导致的土壤微生物生物量的下降有关.降低的微生物生物量及其活性限制了Rs和Rh对土壤水分变化的响应(Zhang et al., 2019). ...
Divergent effects of nitrogen addition on soil respiration in a semiarid grassland 1 2016
... 全球陆地生态系统近3/4的碳(C)储存在土壤中(Masciandaro et al., 2018), 储量高达1 395-2 011 Pg C, 约为大气碳储量的2-4倍(Scharlemann et al., 2014; Bond-Lamberty et al., 2018).作为陆地生态系统第二大碳通量, 土壤呼吸每年释放的CO2为68-98 Pg C, 约为每年化石燃料燃烧释放CO2的10倍(Raich & Schlesinger, 1992; Bond-Lamberty & Thomson, 2010).全球碳模型的预测表明, 由于全球变 化引发的土壤呼吸的微小变动都会显著影响大气 CO2浓度, 进而影响全球碳平衡(Cox et al., 2000; Zhu et al., 2016; Wang et al., 2019c).土壤呼吸根据CO2产生的来源不同主要分为两个组分, 一是自养呼吸, 主要是来自于植物根呼吸以及菌根真菌呼吸产生的CO2; 另一部分是异养呼吸, 主要是来自于凋落物分解和微生物降解土壤有机质产生的CO2 (Hanson et al., 2000; Bond-Lamberty et al., 2004).由于底物来源和对环境因子响应的差异, 土壤呼吸的自养和异养组分对氮沉降等全球变化的响应存在显著差别(Zhou et al., 2014; Wei et al., 2016).因此, 为了准确预测全球变化对土壤呼吸及生态系统碳循环的影响, 就需要区分土壤呼吸的不同组分, 并研究各组分对全球变化因子的响应规律(Wang et al., 2019c), 这对评估整个生态系统的碳收支平衡是非常重要的(Lovett et al., 2006; ArchMiller & Samuelson, 2016). ...
 ,
, ,1,2,*
,1,2,*
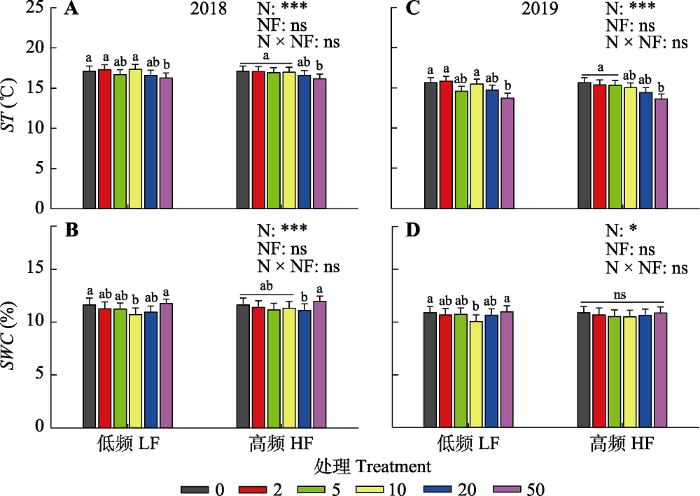 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT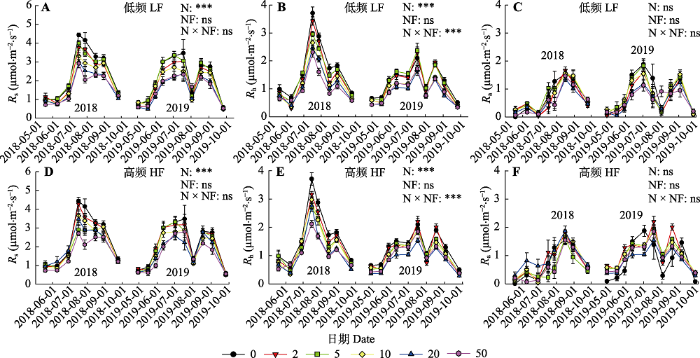 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT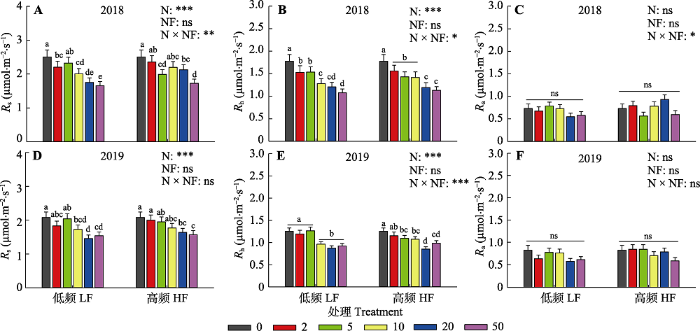 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT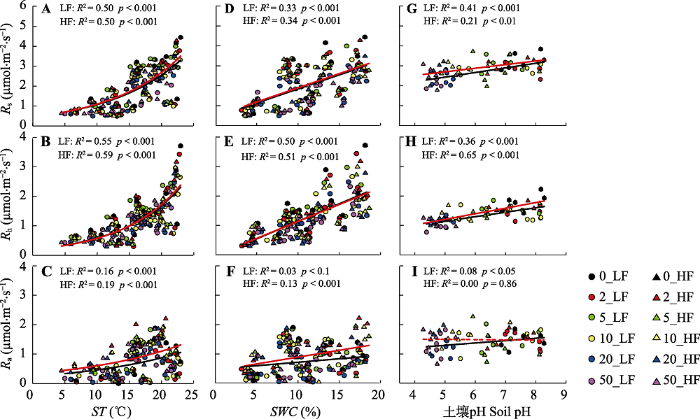 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT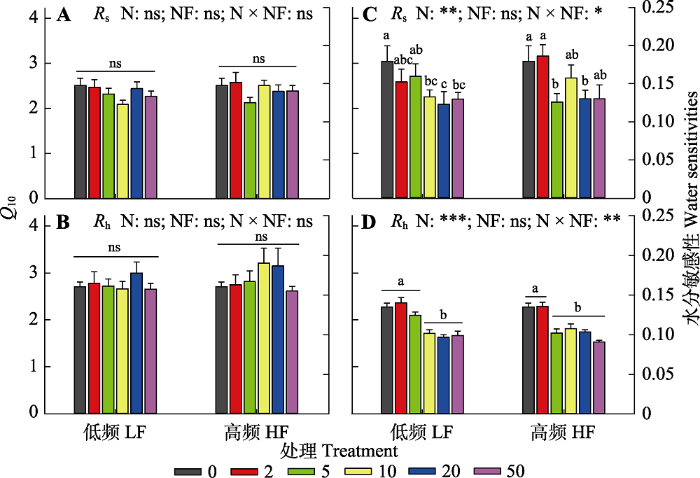 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT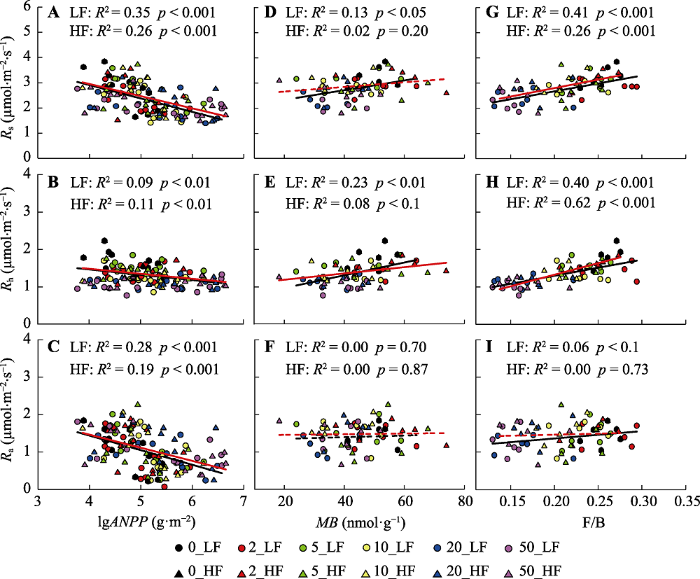 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}