Effects of different disturbances on plant growth and content of main medicinal ingredients of rhizomatous clonal plant Glycyrrhiza uralensis in a natural population
Xue-Hua YE,,1,*, Jian-Guo XUE1, Xiu-Fang XIE2, Zhen-Ying HUANG11State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 2School of Medical Humanities, Capital Medical University, Beijing 100069, China
Supported by the Science and Technology Service Network Initiative Program of Chinese Academy of Sciences(KFJ-EW-STS-118) the National Key R&D Program of China(2016YFC050080502)
Abstract Aims Long-term enclosure and grazing prohibition protects Glycyrrhiza uralensis natural population from external disturbances, such as excessive excavation or overgrazing, but it does not prompt the rapid recovery of the damaged G. uralensis natural population. Appropriate external disturbance is always beneficial to the growth and reproduction of clonal plants such as G. uralensis due to plant clonality. But the roles and mechanisms of disturbances and plant clonality in the recovery of damaged G. uralensis natural populations are still not clear. Methods A natural population of G. uralensis was chosen in the present research, and a field experiment with both clipping (simulated grazing disturbance) and ramet digging out (simulated human excavation disturbance) treatments was conducted to test the effects of different disturbances on plant growth and the content of medicinal ingredients of G. uralensis in the natural population. Important findings The results showed that moderate disturbances, including clipping and ramet digging out treatments, had no significant effects on the ramet density, height and shoot biomass of individual ramets of G. uralensis in the natural population, suggesting the same compensation growth in G. uralensis. In contrast, G. uralensis showed different responses to severe disturbances, depending on the disturbance type. Significantly lower ramet density, height and shoot biomass of individual ramets were observed in G. uralensis under severe clipping treatment, indicating undercompensated growth; while significantly higher ramet height, shoot biomass of individual ramets and also total shoot biomass of plant population were observed under severe digging out treatment, suggesting super-compensatory growth in G. uralensis. Plant clonality, including clonal integration, clonal storage and selective placement of ramets, played important roles in the response of G. uralensis to external disturbance. The content of liquritin and glycyrrhizic acid in taproot of G. uralensis was improved to some extent by external disturbance, suggesting that disturbance at certain levels may improve the medicinal material quality of G. uralensis. Keywords:compensatory growth;disturbance;rhizome network;clonal plant;natural population;medicinal materials quality;Glycyrrhiza uralensis
PDF (1354KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 叶学华, 薛建国, 谢秀芳, 黄振英. 外部干扰对根茎型克隆植物甘草自然种群植株生长及主要药用成分含量的影响. 植物生态学报, 2020, 44(9): 951-961. DOI: 10.17521/cjpe.2020.0188 YE Xue-Hua, XUE Jian-Guo, XIE Xiu-Fang, HUANG Zhen-Ying. Effects of different disturbances on plant growth and content of main medicinal ingredients of rhizomatous clonal plant Glycyrrhiza uralensis in a natural population. Chinese Journal of Plant Ecology, 2020, 44(9): 951-961. DOI: 10.17521/cjpe.2020.0188
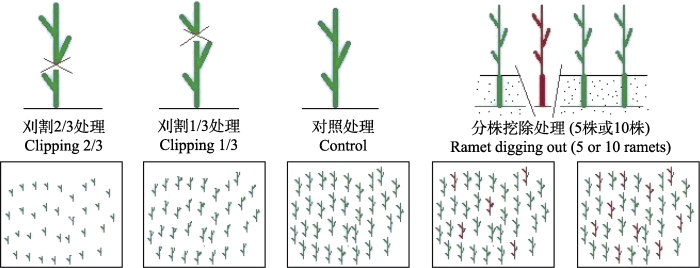
Fig. 1Experiments with both clipping (simulated grazing disturbance) and ramet digging out (simulated human excavation disturbance) treatments of Glycyrrhiza uralensis.
Table 1 表1 表1刈割和挖除干扰对甘草生长及其主要药用成分的影响 Table 1Effects of clipping and digging treatments on plant growth and content of main medicinal ingredients in the taproots of Glycyrrhiza uralensis
性状 Trait
刈割 Clipping
挖除 Digging
df
F
p
df
F
p
植物生长 Plant growth
分株高度 Height of ramets
2, 89
8.513
<0.001
2, 89
3.823
0.026
分株冠幅 Canopy of ramets
2, 89
3.113
0.049
2, 89
3.896
0.024
分株密度 Density of ramets
2, 17
5.051
0.021
2, 17
2.499
0.116
主根长 Taproot length
2, 17
0.296
0.748
2, 17
0.012
0.988
芦头直径 Diameter of reed head
2, 17
1.425
0.271
2, 17
1.806
0.198
分株生物量 Biomass of single ramet
地上生物量 Shoot biomass
2, 17
2.241
0.141
2, 17
3.867
0.044
主根生物量 Root biomass
2, 17
0.046
0.956
2, 17
0.672
0.525
主根药用成分含量 Medicinal ingredients
甘草苷含量 Liquritin content
2, 17
1.613
0.252
2, 17
2.964
0.103
甘草酸含量 Glycyrrhizic acid content
2, 17
1.722
0.233
2, 17
2.040
0.186
Bold data indicate significant effects of the treatments at the level of p < 0.05. 加粗数据表示差异显著(p < 0.05)。
Fig. 2Shoot (A, B) and taproot biomass (C, D) of individual ramets of Glycyrrhiza uralensis under different experimental treatments (mean ± SE). Different lowercase letters represent significant differences between different treatments (p < 0.05).
Fig. 3Ramet density (A, B), ramet height (C, D), ramet canopy (E, F) and taproot length (G, H) in Glycyrrhiza uralensis under different experimental treatments (mean ± SE). Different lowercase letters represent significant differences between different treatments (p < 0.05).
Fig. 4The content of liquritin (A, B) and glycyrrhizic acid (C, D) of taproot in Glycyrrhiza uralensis under different experimental treatments (mean ± SE). Different lowercase letters represent significant differences between different treatments (p < 0.05).
AdlerPB, HilleRisLambersJ ( 2008). The influence of climate and species composition on the population dynamics of ten prairie forbs Ecology, 89, 3049-3060. [本文引用: 1]
AlpertP ( 1999). Clonal integration in Fragaria chiloensis differs between populations: ramets from grassland are selfish Oecologia, 120, 69-76. [本文引用: 1]
AltesorA, LeoniE, GuidoA, ParueloJM ( 2017). Differential responses of three grasses to defoliation, water and light availability Plant Ecology, 218, 95-104. [本文引用: 2]
BachCE ( 2000). Effects of clonal integration on response to sand burial and defoliation by the dune plantIpomoea pes-caprae( Convolvulaceae) Australian Journal of Botany, 48, 159-166. [本文引用: 2]
BeyschlagW, WittlandM, JentschA, SteinleinT ( 2008). Soil crusts and disturbance benefit plant germination, establishhment and growth on nutrient deficient sand Basic and Applied Ecology, 9, 243-252. [本文引用: 1]
BullockJM, MortimerAM, BegonM ( 1994). Physiological integration among tillers of Holcus lanatus: age-dependence and responses to clipping and competition New Phytologist, 128, 737-747. [本文引用: 2]
ChaoMX, YeBP ( 2013). Influence of environmental abiotic factors on plant secondary metabolite biosynthesis Pharmaceutical Biotechnology, 20, 365-368.
ChenD, AliA, YongXH, LinCG, NiuXH, CaiAM, DongBC, ZhouZX, WangYJ, YuFH ( 2019). A multi-species comparison of selective placement patterns of ramets in invasive alien and native clonal plants to light, soil nutrient and water heterogeneity Science of the Total Environment, 657, 1568-1577. [本文引用: 2]
ChangCC, HalpernCB, AntosJA, AvolioML, BiswasA, CookJE, del MoralR, FischerDG, HolzA, PabstRJ, SwansonME, ZobelDB ( 2019). Testing conceptual models of early plant succession across a disturbance gradient Journal of Ecology, 107, 517-530. [本文引用: 1]
CharpentierA, StueferJF ( 1999). Functional specialization of ramets in Scirpus maritimus—Splitting the tasks of sexual reproduction, vegetative growth, and resource storage Plant Ecology, 141, 129-136. [本文引用: 1]
CheplickGP ( 2017). Responses of native plant populations on an unprotected beach to disturbance by storm-induced overwash events Plant Ecology, 218, 105-118. [本文引用: 1]
Chinese PharmacopoeiaCommission (2015). Pharmacopoeia of China. China Medical Science and Technology Press, Beijing. [本文引用: 1]
de KroonH, van GroenendaelJ ( 1997). The Ecology and Evolution of Clonal Plants Backhuys Publishers,Leiden.
DongBC, YuGL, GuoW, ZhangMX, DongM, YuFH ( 2010). How internode length, position and presence of leaves affect survival and growth of Alternanthera philoxeroides after fragmentation? Evolutionary Ecology, 24, 1447-1461.
DongM, AlatenB ( 1999). Clonal plasticity in response to rhizome severing and heterogeneous resource supply in the rhizomatous grass Psammochloa villosa in an Inner Mongolian dune, China Plant Ecology, 141, 53-58. [本文引用: 1]
Editorial Committee of Flora of China, the Chinese Academic of Science ( 1998). Flora of China: Volume 42(2). Science Press, Beijing. [本文引用: 1]
FerraroDO, OesterheldM ( 2002). Effect of defoliation on grass growth. A quantitative review Oikos, 98, 125-133.
FranklinDC, PriorLD, HogarthNJ, McMahonCR ( 2010). Bamboo, fire and flood: consequences of disturbance for the vegetative growth of a clumping, clonal plant Plant Ecology, 208, 319-332. [本文引用: 1]
GroganP, ZaminTJ ( 2018). Growth responses of the common arctic graminoid Eriophorum vaginatum to simulated grazing are independent of soil nitrogen availability Oecologia, 186, 151-162.
HerbenT, KlimesovaJ, ChytryM ( 2018). Effects of disturbance frequency and severity on plant traits: an assessment across a temperate flora Functional Ecology, 32, 799-808. [本文引用: 2]
HicksS, TurkingtonR ( 2000). Compensatory growth of three herbaceous perennial species: the effects of clipping and nutrient availability Canadian Journal of Botany, 78, 759-767. [本文引用: 1]
HolzapfelC, AlpertP ( 2003). Root cooperation in a clonal plant: connected strawberries segregate roots Oecologia, 134, 72-77. [本文引用: 2]
HuJF, ShenFJ ( 1996). A survey of the studies on chemical constituents of Glycyrrhiza Natural Product Research and Development, 8, 77-91. [本文引用: 1]
HuangLQ, GuoLP ( 2007). Secondary metabolites accumulating and geoherbs formation under environmental stress China Journal of Chinese Materia Medica, 32, 277-280. [本文引用: 1]
HuangMJ, WangWQ, WeiSL ( 2010). Investigation on medicinal plant resources of Glycyrrhiza uralensis in China and chemical assessment of its underground part China Journal of Chinese Materia Medica, 35, 947-952. [本文引用: 2]
Jane?ek?, Klime?ováJ ( 2014). Carbohydrate storage in meadow plants and its depletion after disturbance: Do roots and stem-derived organs differ in their roles? Oecologia, 175, 51-61.
JiangQ, PanZB, ZhangQY, LiM ( 2004). Countermeasures of Glycyrrhiza resource protection and sustainable utilization in Ningxia Resource Development & Market, 20, 128-130. [本文引用: 1]
KumordziBB, AubinI, CardouF, ShipleyB, ViolleC, JohnstoneJ, AnandM, ArsenaultA, BellFW, BergeronY, BoulangeatI, BrousseauM, de GradnpréL, DelagrangeS, FentonNJ, GravelD, MacdonaldSE, HamelB, HigelinM, HébertF, IsabelN, MallikA, McIntoshACS, McLarenJR, MessierC, MorrisD, ThiffaultN, TremblayJP, MunsonAD ( 2019). Geographic scale and disturbance influence intraspecific trait variability in leaves and roots of North American understorey plants Functional Ecology, 33, 1771-1784. DOI:10.1111/fec.v33.9URL
LiSZ, PenningsSC ( 2017). Timing of disturbance affects biomass and flowering of a saltmarsh plant and attack by stem-boring herbivores Ecosphere, 8, e01675. DOI: 10.1002/ecs2.1675. [本文引用: 3]
LiXB, ChenL, LiGQ, AnH ( 2013a). Influence of enclosure on Glyeyrrhiza uralensis community and distribution pattern in arid and semi-arid areas Acta Ecologica Sinica, 33, 3995-4001. [本文引用: 2]
LiXB, ChenL, LiGQ, AnH ( 2013b). Ecological distribution and propagative technique research of Glycyrrhiza resources in China Ecology and Environment, 22, 718-722. [本文引用: 2]
LiangB, YangAF, HuangFL, HuBZ ( 2006). Research advances on chemical constituents and pharmacological activities in Glycyrrhiza Journal of Northeast Agricultural University, 37, 115-119. [本文引用: 1]
LiuHD, YuFH, HeWM, ChuY, DongM ( 2009). Clonal integration improves compensatory growth in heavily grazed ramet populations of two inland-dune grasses Flora, 204, 298-305. [本文引用: 1]
LiuZR, ChengJ ( 2011). The correlation between the content of liquritin and glycyrrhizin acid in licorice and its growth factores Xinjiang Journal of Traditional Chinese Medicine, 29, 39-41. [本文引用: 1]
McNaughtonSJ ( 1983). Compensatory plant growth as a response to herbivory Oikos, 40, 329-336. [本文引用: 1]
McNaughtonSJ ( 1979). Grazing as an optimization process: grass-ungulate relationships in the Serengeti The American Naturalist, 113, 691-703. [本文引用: 1]
MeuriotF, Morvan-BertrandA, Noiraud-RomyN, DecauML, Escobar-GutiérrezAJ, GastalF, Prud’hommeMP ( 2018). Short-term effects of defoliation intensity on sugar remobilization and N fluxes in ryegrass Journal of Experimental Botany, 69, 3975-3986.
PitelkaLF, AshmunJW (1985). Physiology and integration of ramets in clonal plants//Jackson JBC, Buss LW, Cook RE. Population Biology and Evolution of Clonal Organisms. Yale University Press, New Haven. 399-436. [本文引用: 1]
PitelkaLF ( 1984). Application of the -3/2 power law to clonal herbs The American Naturalist, 123, 442-449.
QinRR, WangXL ( 2020). Effects of crown height on the compensatory growth of Italian ryegrass based on combined effects of stored organic matter and cytokinin Grassland Science, 66, 29-39. [本文引用: 2]
RenHY, TaubeFH, SteinCD, ZhangYJ, BaiYF, HuSJ ( 2018). Grazing weakens temporal stabilizing effects of diversity in the Eurasian steppe Ecology and Evolution, 8, 231-241. [本文引用: 1]
ReuschTBH ( 2006). Does disturbance enhance genotypic diversity in clonal organisms? A field test in the marine angiosperm Zostera marina Molecular Ecology, 15, 277-286. [本文引用: 1]
Saixiyala, YangD, ZhangSD, LiuGF, YangXJ, HuangZY, YeXH ( 2017). Facilitation by a spiny shrub on a rhizomatous clonal herbaceous in thicketization-grassland in Northern China: increased soil resources or shelter from herbivores Frontiers in Plant Science, 8, 809. DOI: 10.3389/fpls.2017.00809. [本文引用: 4]
ShangXH, YanYN, WangWQ ( 2010). Effect of exclosure on community structure, yield and quality of Glycyrrhiza uralensis root in G. uralensis community Journal of Agricultural University of Hebei, 33, 40-44. [本文引用: 3]
SuzukiJ, StueferJF ( 1999). On the ecological and evolutionary significance of storage in clonal plants Plant Species Biology, 14, 11-17. [本文引用: 2]
van StaalduinenMA, AntenNPR ( 2005). Differences in the compensatory growth of two co-occurring grass species in relation to water availability Oecologia, 146, 190-199. [本文引用: 2]
van GroenendaelJM, de KroonH ( 1990). Clonal Growth in Plants: Regulation and Function. SPB Academic Publishing, the Hague. [本文引用: 2]
WangJY, AbdullahI, XuTT, ZhuWY, GaoY, WangL ( 2019). Effects of mowing disturbance and competition on spatial expansion of the clonal plant Leymus chinensis into saline-alkali soil patches Environmental and Experimental Botany, 168, 103890. DOI: 10.1016/j.envexpbot.2019. 103890. [本文引用: 2]
WangYJ, ShiXP, ZhongZC . ( 2013). The relative importance of sexual reproduction and clonal propagation in rhizomatous herb Iris japonica Thunb. from two habitats of Jinyun Mountain, Southwest China Russian Journal of Ecology, 44, 199-206.
WangZW, LiLH, HanXG, DongM ( 2004). Do rhizome severing and shoot defoliation affect clonal growth of Leymus chinensis at ramet population level? Acta Oecologica, 26, 255-260. [本文引用: 1]
XieZC, ChuYK, ZhangWJ, LangDY, ZhangXH ( 2019). Bacillus pumilus alleviates drought stress and increases metabolite accumulation in Glycyrrhiza uralensis Fisch Environmental and Experimental Botany, 158, 99-106. [本文引用: 1]
YangLL, ChenJJ, YangTS, LiZ, HuWM, WangY ( 2013). Geographic distribution and resource survey of the wild medicinal plants Licorice ( Glycyrrhiza uralensis) in northwest China Chinese Wild Plant Resources, 32, 27-31. [本文引用: 1]
YangP, LiKC, XinJ, YunNQ, MaGZ, YouQ, ZhangZZ ( 2004). A study on the status and protective utilization of licorice resource in Ningxia Journal of Ningxia Agricultural College, 25, 17-20. [本文引用: 1]
YeXH, GaoSQ, LiuZL, ZhangYL, HuangZY, DongM ( 2015). Multiple adaptations to light and nutrient heterogeneity in the clonal plant Leymus secalinus with a combined growth form Flora, 213, 49-56.
YeXH, HuYK, LiuZL, GaoSQ, DongM ( 2013). Water heterogeneity affects water storage in two rhizomatous clonal plants Leymus secalinus and Calamagrostis pseudophragmites Chinese Journal of Plant Ecology, 37, 427-435.
YuFH, DongM, KrusiB ( 2004). Clonal integration helps Psammochloa villosa survive sand burial in an inland dune New Phytologist, 162, 697-704.
YuFH, WangN, HeWM, ChuY, DongM ( 2008). Adaptation of rhizome connections in drylands: increasing tolerance of clones to wind erosion Annals of Botany, 102, 571-577. [本文引用: 2]
ZhangCY, YangC, DongM ( 2002). Clonal integration and its ecological significance in Hedysarum laeave, a rhizomatous shrub in Mu Us Sandland Journal of Plant Research, 115, 113-118. [本文引用: 1]
ZhangDM, ZhaoWZ, LuoWC ( 2019). Effect of the population density on belowground bud bank of a rhizomatous clonal plant Leymus secalinus in Mu Us sandy land Journal of Plant Research, 132, 69-80.
ZhangSC, LongZP, QiGB, LiXY ( 2007). Research on grassland community management—A case study of Yanchi County Acta Agrestia Sinica, 15, 479-485. [本文引用: 2]
ZhuCG, ChenYN, LiWH, MaXD ( 2014). Effect of herbivory on the growth and photosynthesis of replanted Calligonum caput-medusae saplings in an infertile arid desert Plant Ecology, 215, 155-167. [本文引用: 1]
The influence of climate and species composition on the population dynamics of ten prairie forbs 1 2008
... 外部干扰是自然生态系统中植物所必须面对的一种重要生态过程.植物具有应对外部干扰并产生积极响应的机制, 比如: 叶片光合速率增加(Zhu et al., 2014), 贮藏器官(根、根状茎和匍匐茎等)碳水化合物的再分配(Oyarzabal & Oesterheld, 2009; Qin & Wang, 2020), 残余分生组织活性增强以加速叶的再生长和分蘖(Wang et al., 2013; Meuriot et al., 2018), 减少植物蒸腾面积以提高水分可利用性等(Ferraro & Oesterheld, 2002).通过这些机制, 植物受到外部干扰后的生长和繁殖能够得到维持, 甚至得以加强, 这种现象称为补偿性生长(McNaughton, 1983).不同植物种应对外部干扰的补偿效应不一样(Adler & HilleRisLambers, 2008; Altesor et al., 2017; Ran et al., 2018), 这种效应还受许多因素的影响, 如干扰的类型和程度(Hicks & Turkington, 2000; Oborny et al., 2000; Meuriot et al., 2018), 以及光照、温度和土壤养分等资源的可利用性(Oyarzabal & Oesterheld, 2009; Altesor et al., 2017; Grogan & Zamin, 2018).克隆植物特性, 如克隆整合或克隆储存, 能够促进受到外部干扰的克隆分株生长与恢复, 从而缓解外部干扰对克隆植物的影响, 也是克隆植物应对外部干扰的重要补偿性机制(Bullock et al., 1994; Bach, 2000; Zhang et al., 2002; Wang et al., 2004; Wang et al., 2019). ...
Clonal integration in Fragaria chiloensis differs between populations: ramets from grassland are selfish 1 1999
... 作为根茎型克隆植物, 甘草具有发达的地下根状茎, 能够形成巨大的地下根状茎网络.通过根状茎网络, 分散分布于较大空间的甘草相连分株之间可以进行资源的传输(即克隆整合; Alpert, 1999; Dong & Alaten, 1999)和新生分株生长位置的调整(即分株选择性放置; Ye et al., 2015; Saixiyala et al., 2017; Chen et al., 2019); 同时, 巨大的地下根状茎网络也具有储藏功能(即克隆储藏; Dong et al., 2010; 叶学华等, 2013), 不仅能够储藏碳水化合物, 而且能够“储藏”分生组织(芽; Suzuki & Stuefer, 1999; Zhang et al., 2019).当甘草受到动物采食或者人工采挖等外部干扰时, 这些克隆植物特性将帮助被干扰的甘草分株进行再生长或者促进新的分株生成.但克隆植物特性是否是甘草种群受到干扰后恢复生长的一种补偿机制, 还需要科学验证(Bullock et al., 1994; Bach, 2000). ...
Differential responses of three grasses to defoliation, water and light availability 2 2017
... 外部干扰是自然生态系统中植物所必须面对的一种重要生态过程.植物具有应对外部干扰并产生积极响应的机制, 比如: 叶片光合速率增加(Zhu et al., 2014), 贮藏器官(根、根状茎和匍匐茎等)碳水化合物的再分配(Oyarzabal & Oesterheld, 2009; Qin & Wang, 2020), 残余分生组织活性增强以加速叶的再生长和分蘖(Wang et al., 2013; Meuriot et al., 2018), 减少植物蒸腾面积以提高水分可利用性等(Ferraro & Oesterheld, 2002).通过这些机制, 植物受到外部干扰后的生长和繁殖能够得到维持, 甚至得以加强, 这种现象称为补偿性生长(McNaughton, 1983).不同植物种应对外部干扰的补偿效应不一样(Adler & HilleRisLambers, 2008; Altesor et al., 2017; Ran et al., 2018), 这种效应还受许多因素的影响, 如干扰的类型和程度(Hicks & Turkington, 2000; Oborny et al., 2000; Meuriot et al., 2018), 以及光照、温度和土壤养分等资源的可利用性(Oyarzabal & Oesterheld, 2009; Altesor et al., 2017; Grogan & Zamin, 2018).克隆植物特性, 如克隆整合或克隆储存, 能够促进受到外部干扰的克隆分株生长与恢复, 从而缓解外部干扰对克隆植物的影响, 也是克隆植物应对外部干扰的重要补偿性机制(Bullock et al., 1994; Bach, 2000; Zhang et al., 2002; Wang et al., 2004; Wang et al., 2019). ...
... ; Altesor et al., 2017; Grogan & Zamin, 2018).克隆植物特性, 如克隆整合或克隆储存, 能够促进受到外部干扰的克隆分株生长与恢复, 从而缓解外部干扰对克隆植物的影响, 也是克隆植物应对外部干扰的重要补偿性机制(Bullock et al., 1994; Bach, 2000; Zhang et al., 2002; Wang et al., 2004; Wang et al., 2019). ...
Effects of clonal integration on response to sand burial and defoliation by the dune plantIpomoea pes-caprae( Convolvulaceae) 2 2000
... 作为根茎型克隆植物, 甘草具有发达的地下根状茎, 能够形成巨大的地下根状茎网络.通过根状茎网络, 分散分布于较大空间的甘草相连分株之间可以进行资源的传输(即克隆整合; Alpert, 1999; Dong & Alaten, 1999)和新生分株生长位置的调整(即分株选择性放置; Ye et al., 2015; Saixiyala et al., 2017; Chen et al., 2019); 同时, 巨大的地下根状茎网络也具有储藏功能(即克隆储藏; Dong et al., 2010; 叶学华等, 2013), 不仅能够储藏碳水化合物, 而且能够“储藏”分生组织(芽; Suzuki & Stuefer, 1999; Zhang et al., 2019).当甘草受到动物采食或者人工采挖等外部干扰时, 这些克隆植物特性将帮助被干扰的甘草分株进行再生长或者促进新的分株生成.但克隆植物特性是否是甘草种群受到干扰后恢复生长的一种补偿机制, 还需要科学验证(Bullock et al., 1994; Bach, 2000). ...
... 外部干扰是自然生态系统中植物所必须面对的一种重要生态过程.植物具有应对外部干扰并产生积极响应的机制, 比如: 叶片光合速率增加(Zhu et al., 2014), 贮藏器官(根、根状茎和匍匐茎等)碳水化合物的再分配(Oyarzabal & Oesterheld, 2009; Qin & Wang, 2020), 残余分生组织活性增强以加速叶的再生长和分蘖(Wang et al., 2013; Meuriot et al., 2018), 减少植物蒸腾面积以提高水分可利用性等(Ferraro & Oesterheld, 2002).通过这些机制, 植物受到外部干扰后的生长和繁殖能够得到维持, 甚至得以加强, 这种现象称为补偿性生长(McNaughton, 1983).不同植物种应对外部干扰的补偿效应不一样(Adler & HilleRisLambers, 2008; Altesor et al., 2017; Ran et al., 2018), 这种效应还受许多因素的影响, 如干扰的类型和程度(Hicks & Turkington, 2000; Oborny et al., 2000; Meuriot et al., 2018), 以及光照、温度和土壤养分等资源的可利用性(Oyarzabal & Oesterheld, 2009; Altesor et al., 2017; Grogan & Zamin, 2018).克隆植物特性, 如克隆整合或克隆储存, 能够促进受到外部干扰的克隆分株生长与恢复, 从而缓解外部干扰对克隆植物的影响, 也是克隆植物应对外部干扰的重要补偿性机制(Bullock et al., 1994; Bach, 2000; Zhang et al., 2002; Wang et al., 2004; Wang et al., 2019). ...
Soil crusts and disturbance benefit plant germination, establishhment and growth on nutrient deficient sand 1 2008
... 干扰作为自然界中普遍存在的一种生态学过程, 直接影响植物个体生长、光合产物和次生化合物含量、地下根系、种群空间分布格局、种群或群落生产力、种间关系、群落演替等各个方面(Beyschlag et al., 2008; Franklin et al., 2010; Cheplick, 2017; Li & Pennings, 2017; Herben et al., 2018; Chang et al., 2019; Kumordzi et al., 2019).克隆植物, 特别是游击型克隆植物, 其分株在一定时间段往往通过连接结构(即间隔子)连接在一起形成克隆网络, 并通过克隆网络实现相连分株之间资源和信息的共享(de Kroon & van Groenendael, 1997; 董鸣, 2001).克隆网络的存在显著提高了克隆植物应对外部干扰的能力.例如: Yu等(2004, 2008)发现根茎相连显著提高了根茎型克隆植物沙鞭(Psammochloa villosa)应对沙埋和风蚀干扰的能力; Liu等(2009)研究发现根茎相连促进了遭受采食的根茎型克隆植物沙鞭和甘蒙雀麦(Bromus korotkiji)的分株生长, 尤其是在采食强度很高(去除90%地上枝叶)的情况下.通过克隆网络, 克隆植物可以控制新生分株数量增加以最大限度避免自疏(van Groenendael & de Kroon, 1990), 促进相连分株之间根系的分离以削弱种内竞争(Holzaptel & Alpert, 2003), 从而有利于克隆植物种群的稳定与维持. ...
Physiological integration among tillers of Holcus lanatus: age-dependence and responses to clipping and competition 2 1994
... 作为根茎型克隆植物, 甘草具有发达的地下根状茎, 能够形成巨大的地下根状茎网络.通过根状茎网络, 分散分布于较大空间的甘草相连分株之间可以进行资源的传输(即克隆整合; Alpert, 1999; Dong & Alaten, 1999)和新生分株生长位置的调整(即分株选择性放置; Ye et al., 2015; Saixiyala et al., 2017; Chen et al., 2019); 同时, 巨大的地下根状茎网络也具有储藏功能(即克隆储藏; Dong et al., 2010; 叶学华等, 2013), 不仅能够储藏碳水化合物, 而且能够“储藏”分生组织(芽; Suzuki & Stuefer, 1999; Zhang et al., 2019).当甘草受到动物采食或者人工采挖等外部干扰时, 这些克隆植物特性将帮助被干扰的甘草分株进行再生长或者促进新的分株生成.但克隆植物特性是否是甘草种群受到干扰后恢复生长的一种补偿机制, 还需要科学验证(Bullock et al., 1994; Bach, 2000). ...
... 外部干扰是自然生态系统中植物所必须面对的一种重要生态过程.植物具有应对外部干扰并产生积极响应的机制, 比如: 叶片光合速率增加(Zhu et al., 2014), 贮藏器官(根、根状茎和匍匐茎等)碳水化合物的再分配(Oyarzabal & Oesterheld, 2009; Qin & Wang, 2020), 残余分生组织活性增强以加速叶的再生长和分蘖(Wang et al., 2013; Meuriot et al., 2018), 减少植物蒸腾面积以提高水分可利用性等(Ferraro & Oesterheld, 2002).通过这些机制, 植物受到外部干扰后的生长和繁殖能够得到维持, 甚至得以加强, 这种现象称为补偿性生长(McNaughton, 1983).不同植物种应对外部干扰的补偿效应不一样(Adler & HilleRisLambers, 2008; Altesor et al., 2017; Ran et al., 2018), 这种效应还受许多因素的影响, 如干扰的类型和程度(Hicks & Turkington, 2000; Oborny et al., 2000; Meuriot et al., 2018), 以及光照、温度和土壤养分等资源的可利用性(Oyarzabal & Oesterheld, 2009; Altesor et al., 2017; Grogan & Zamin, 2018).克隆植物特性, 如克隆整合或克隆储存, 能够促进受到外部干扰的克隆分株生长与恢复, 从而缓解外部干扰对克隆植物的影响, 也是克隆植物应对外部干扰的重要补偿性机制(Bullock et al., 1994; Bach, 2000; Zhang et al., 2002; Wang et al., 2004; Wang et al., 2019). ...
环境非生物因子对植物次生代谢产物合成的影响 2013
环境非生物因子对植物次生代谢产物合成的影响 2013
A multi-species comparison of selective placement patterns of ramets in invasive alien and native clonal plants to light, soil nutrient and water heterogeneity 2 2019
Clonal plasticity in response to rhizome severing and heterogeneous resource supply in the rhizomatous grass Psammochloa villosa in an Inner Mongolian dune, China 1 1999
... 作为根茎型克隆植物, 甘草具有发达的地下根状茎, 能够形成巨大的地下根状茎网络.通过根状茎网络, 分散分布于较大空间的甘草相连分株之间可以进行资源的传输(即克隆整合; Alpert, 1999; Dong & Alaten, 1999)和新生分株生长位置的调整(即分株选择性放置; Ye et al., 2015; Saixiyala et al., 2017; Chen et al., 2019); 同时, 巨大的地下根状茎网络也具有储藏功能(即克隆储藏; Dong et al., 2010; 叶学华等, 2013), 不仅能够储藏碳水化合物, 而且能够“储藏”分生组织(芽; Suzuki & Stuefer, 1999; Zhang et al., 2019).当甘草受到动物采食或者人工采挖等外部干扰时, 这些克隆植物特性将帮助被干扰的甘草分株进行再生长或者促进新的分株生成.但克隆植物特性是否是甘草种群受到干扰后恢复生长的一种补偿机制, 还需要科学验证(Bullock et al., 1994; Bach, 2000). ...
Facilitation by a spiny shrub on a rhizomatous clonal herbaceous in thicketization-grassland in Northern China: increased soil resources or shelter from herbivores 4 2017
... 作为根茎型克隆植物, 甘草具有发达的地下根状茎, 能够形成巨大的地下根状茎网络.通过根状茎网络, 分散分布于较大空间的甘草相连分株之间可以进行资源的传输(即克隆整合; Alpert, 1999; Dong & Alaten, 1999)和新生分株生长位置的调整(即分株选择性放置; Ye et al., 2015; Saixiyala et al., 2017; Chen et al., 2019); 同时, 巨大的地下根状茎网络也具有储藏功能(即克隆储藏; Dong et al., 2010; 叶学华等, 2013), 不仅能够储藏碳水化合物, 而且能够“储藏”分生组织(芽; Suzuki & Stuefer, 1999; Zhang et al., 2019).当甘草受到动物采食或者人工采挖等外部干扰时, 这些克隆植物特性将帮助被干扰的甘草分株进行再生长或者促进新的分株生成.但克隆植物特性是否是甘草种群受到干扰后恢复生长的一种补偿机制, 还需要科学验证(Bullock et al., 1994; Bach, 2000). ...
Differences in the compensatory growth of two co-occurring grass species in relation to water availability 2 2005
... 外部干扰是自然生态系统中植物所必须面对的一种重要生态过程.植物具有应对外部干扰并产生积极响应的机制, 比如: 叶片光合速率增加(Zhu et al., 2014), 贮藏器官(根、根状茎和匍匐茎等)碳水化合物的再分配(Oyarzabal & Oesterheld, 2009; Qin & Wang, 2020), 残余分生组织活性增强以加速叶的再生长和分蘖(Wang et al., 2013; Meuriot et al., 2018), 减少植物蒸腾面积以提高水分可利用性等(Ferraro & Oesterheld, 2002).通过这些机制, 植物受到外部干扰后的生长和繁殖能够得到维持, 甚至得以加强, 这种现象称为补偿性生长(McNaughton, 1983).不同植物种应对外部干扰的补偿效应不一样(Adler & HilleRisLambers, 2008; Altesor et al., 2017; Ran et al., 2018), 这种效应还受许多因素的影响, 如干扰的类型和程度(Hicks & Turkington, 2000; Oborny et al., 2000; Meuriot et al., 2018), 以及光照、温度和土壤养分等资源的可利用性(Oyarzabal & Oesterheld, 2009; Altesor et al., 2017; Grogan & Zamin, 2018).克隆植物特性, 如克隆整合或克隆储存, 能够促进受到外部干扰的克隆分株生长与恢复, 从而缓解外部干扰对克隆植物的影响, 也是克隆植物应对外部干扰的重要补偿性机制(Bullock et al., 1994; Bach, 2000; Zhang et al., 2002; Wang et al., 2004; Wang et al., 2019). ...
... ; Meuriot et al., 2018), 以及光照、温度和土壤养分等资源的可利用性(Oyarzabal & Oesterheld, 2009; Altesor et al., 2017; Grogan & Zamin, 2018).克隆植物特性, 如克隆整合或克隆储存, 能够促进受到外部干扰的克隆分株生长与恢复, 从而缓解外部干扰对克隆植物的影响, 也是克隆植物应对外部干扰的重要补偿性机制(Bullock et al., 1994; Bach, 2000; Zhang et al., 2002; Wang et al., 2004; Wang et al., 2019). ...
2 1990
... 干扰作为自然界中普遍存在的一种生态学过程, 直接影响植物个体生长、光合产物和次生化合物含量、地下根系、种群空间分布格局、种群或群落生产力、种间关系、群落演替等各个方面(Beyschlag et al., 2008; Franklin et al., 2010; Cheplick, 2017; Li & Pennings, 2017; Herben et al., 2018; Chang et al., 2019; Kumordzi et al., 2019).克隆植物, 特别是游击型克隆植物, 其分株在一定时间段往往通过连接结构(即间隔子)连接在一起形成克隆网络, 并通过克隆网络实现相连分株之间资源和信息的共享(de Kroon & van Groenendael, 1997; 董鸣, 2001).克隆网络的存在显著提高了克隆植物应对外部干扰的能力.例如: Yu等(2004, 2008)发现根茎相连显著提高了根茎型克隆植物沙鞭(Psammochloa villosa)应对沙埋和风蚀干扰的能力; Liu等(2009)研究发现根茎相连促进了遭受采食的根茎型克隆植物沙鞭和甘蒙雀麦(Bromus korotkiji)的分株生长, 尤其是在采食强度很高(去除90%地上枝叶)的情况下.通过克隆网络, 克隆植物可以控制新生分株数量增加以最大限度避免自疏(van Groenendael & de Kroon, 1990), 促进相连分株之间根系的分离以削弱种内竞争(Holzaptel & Alpert, 2003), 从而有利于克隆植物种群的稳定与维持. ...
... 外部干扰是自然生态系统中植物所必须面对的一种重要生态过程.植物具有应对外部干扰并产生积极响应的机制, 比如: 叶片光合速率增加(Zhu et al., 2014), 贮藏器官(根、根状茎和匍匐茎等)碳水化合物的再分配(Oyarzabal & Oesterheld, 2009; Qin & Wang, 2020), 残余分生组织活性增强以加速叶的再生长和分蘖(Wang et al., 2013; Meuriot et al., 2018), 减少植物蒸腾面积以提高水分可利用性等(Ferraro & Oesterheld, 2002).通过这些机制, 植物受到外部干扰后的生长和繁殖能够得到维持, 甚至得以加强, 这种现象称为补偿性生长(McNaughton, 1983).不同植物种应对外部干扰的补偿效应不一样(Adler & HilleRisLambers, 2008; Altesor et al., 2017; Ran et al., 2018), 这种效应还受许多因素的影响, 如干扰的类型和程度(Hicks & Turkington, 2000; Oborny et al., 2000; Meuriot et al., 2018), 以及光照、温度和土壤养分等资源的可利用性(Oyarzabal & Oesterheld, 2009; Altesor et al., 2017; Grogan & Zamin, 2018).克隆植物特性, 如克隆整合或克隆储存, 能够促进受到外部干扰的克隆分株生长与恢复, 从而缓解外部干扰对克隆植物的影响, 也是克隆植物应对外部干扰的重要补偿性机制(Bullock et al., 1994; Bach, 2000; Zhang et al., 2002; Wang et al., 2004; Wang et al., 2019). ...
Effects of mowing disturbance and competition on spatial expansion of the clonal plant Leymus chinensis into saline-alkali soil patches 2 2019
... 外部干扰是自然生态系统中植物所必须面对的一种重要生态过程.植物具有应对外部干扰并产生积极响应的机制, 比如: 叶片光合速率增加(Zhu et al., 2014), 贮藏器官(根、根状茎和匍匐茎等)碳水化合物的再分配(Oyarzabal & Oesterheld, 2009; Qin & Wang, 2020), 残余分生组织活性增强以加速叶的再生长和分蘖(Wang et al., 2013; Meuriot et al., 2018), 减少植物蒸腾面积以提高水分可利用性等(Ferraro & Oesterheld, 2002).通过这些机制, 植物受到外部干扰后的生长和繁殖能够得到维持, 甚至得以加强, 这种现象称为补偿性生长(McNaughton, 1983).不同植物种应对外部干扰的补偿效应不一样(Adler & HilleRisLambers, 2008; Altesor et al., 2017; Ran et al., 2018), 这种效应还受许多因素的影响, 如干扰的类型和程度(Hicks & Turkington, 2000; Oborny et al., 2000; Meuriot et al., 2018), 以及光照、温度和土壤养分等资源的可利用性(Oyarzabal & Oesterheld, 2009; Altesor et al., 2017; Grogan & Zamin, 2018).克隆植物特性, 如克隆整合或克隆储存, 能够促进受到外部干扰的克隆分株生长与恢复, 从而缓解外部干扰对克隆植物的影响, 也是克隆植物应对外部干扰的重要补偿性机制(Bullock et al., 1994; Bach, 2000; Zhang et al., 2002; Wang et al., 2004; Wang et al., 2019). ...
... ), 以及光照、温度和土壤养分等资源的可利用性(Oyarzabal & Oesterheld, 2009; Altesor et al., 2017; Grogan & Zamin, 2018).克隆植物特性, 如克隆整合或克隆储存, 能够促进受到外部干扰的克隆分株生长与恢复, 从而缓解外部干扰对克隆植物的影响, 也是克隆植物应对外部干扰的重要补偿性机制(Bullock et al., 1994; Bach, 2000; Zhang et al., 2002; Wang et al., 2004; Wang et al., 2019). ...
The relative importance of sexual reproduction and clonal propagation in rhizomatous herb Iris japonica Thunb. from two habitats of Jinyun Mountain, Southwest China 2013
Do rhizome severing and shoot defoliation affect clonal growth of Leymus chinensis at ramet population level? 1 2004
 ,
, ,1,*, 薛建国1, 谢秀芳2, 黄振英1
,1,*, 薛建国1, 谢秀芳2, 黄振英1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT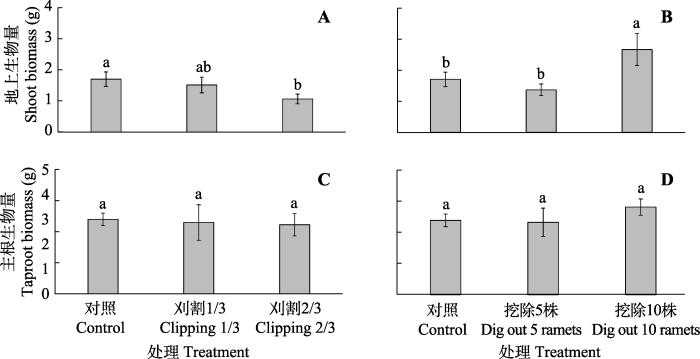 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT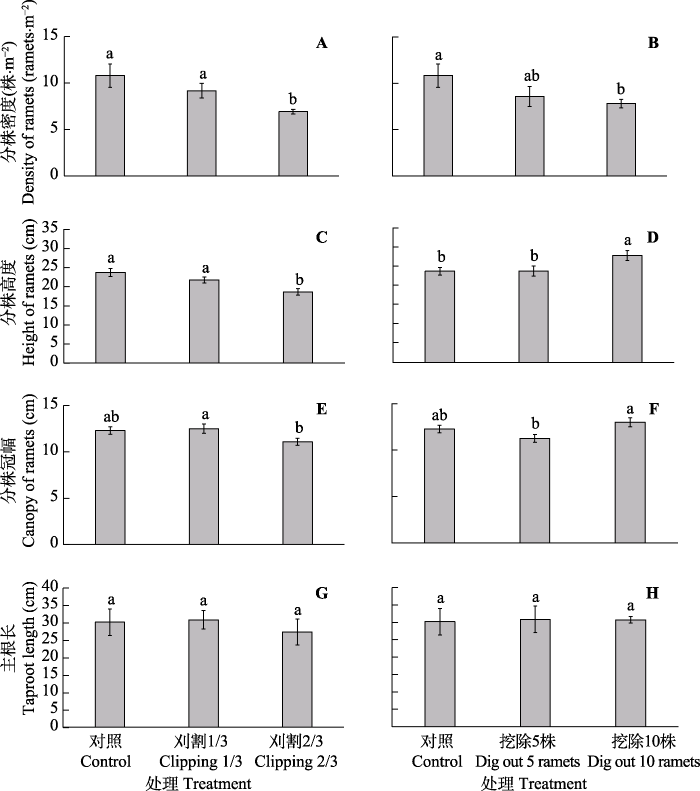 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT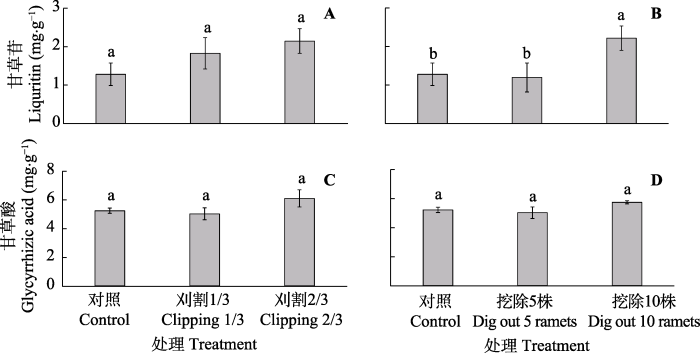 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}