Processes on reproductive ecology of plant species in the Qinghai-Xizang Plateau and adjacent highlands
Chan ZHANG1, Yu-Meng AN1, J?SCHKE Yun2, Lin-Lin WANG3, Zhi-Li ZHOU3, Li-Ping WANG3, Yong-Ping YANG,3,*, Yuan-Wen DUAN,,3,*1College of Life Sciences, Henan Normal University, Xinxiang, Henan 453007, China 2Senckenberg Natural History Museum, G?rlitz, 02826, Germany 3Kunming Institute of Botany, Chinese Academy of Sciences, Kunming 650201, China
Strategic Priority Research Program of Chinese Academy of Sciences(XDA20040000) Strategic Priority Research Program of Chinese Academy of Sciences(XDA20050000) National Natural Science Foundation of China(31500193) National Natural Science Foundation of China(31700368)
Abstract The Qinghai-Xizang Plateau and its adjacent highlands host a rich plant diversity, and understanding on how plant species reproduce successfully in those high mountain regions would be of great help for us to discover the evolution and adaptability of plants to the typical alpine environments. Here, we reviewed the progresses of plant reproductive ecology in the Qinghai-Xizang Plateau and its adjacent highlands, with emphases on resource allocation, reproductive mode, evolution of floral traits, etc. We also summarized the impacts of global climate changes on plant reproduction, and the applications of new technologies and methods in this research field to explore the potential mechanisms of plant evolution and adaptation. In the stressful environments of high mountain areas, with the increase of altitude, the reproductive allocation usually shows a trend of increase, and the proportion of resources invested in male function also increases. However, the specific pattern of resource allocation strongly depends on the mating system, individual size, life history, genetic variation and resource availability. In the context of resource and pollination limitation, plants will weight different reproductive modes. Clonal reproduction and self-pollination are conductive to reproductive assurance in populations with pollinator scarcity, while sexual reproduction and cross-pollination can improve seed quality and genetic diversity of offspring, which could benefit population survival in the potential climate flunctations. Therefore, the combinations of different reproductive modes and the generalized pollination networks may be the good choices to adapt to the stressful environments of highlands. The evolution of floral traits might be mainly driven by the selective pressure of local pollinators, but more and more attentions have been paid to the effects of exotic pollinators, herbivores, nectar robbers and abiotic environments such as temperature, precipitation and ultraviolet radiation on floral traits. In recent years, the Qinghai-Xizang Plateau has become one of research hotspots because of its fragility and sensitivity to climate change. Global climate change, characterized by global warming and increased nitrogen deposition, is directly or indirectly affecting the reproductive characteristics of plant species in the region. Climate change may cause shifts of phenology and distribution area of plants and pollinators, which leads to temporal decoupling and spatial mismatch between them. Plants can respond to climate change by adjusting floral traits (floral display, corolla structure, quantity and quality of reward), which may result in changes of type, amount and foraging behaviors of pollinators and affect reproductive success of plants at last. The applications of new technologies and methods could advance our knowledge on plant reproductive ecology. 3D printed flowers could control floral traits precisely and could be employed in examining the effects of subtle changes of floral traits on pollinator behaviors, which would be useful in understanding pollinator mediated selection on floral traits in combination with the uses of commercial pollinators. Importantly, with the development of high-throughput sequencing, the underlying mechanisms of important topics in plant reproductive ecology could be discovered more easily than before, especially in the field of floral evolution. As a conclusion, key issues that need additional attention in the current research as well as the fields worthy of further study are highlighted. Keywords:Qinghai-Xizang Plateau;reproductive allocation;sexual allocation;clonal reproduction;sexual reproduction;floral evolution
PDF (1352KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张婵, 安宇梦, Yun J?SCHKE, 王林林, 周知里, 王力平, 杨永平, 段元文. 青藏高原及周边高山地区的植物繁殖生态学研究进展. 植物生态学报, 2020, 44(1): 1-21. DOI: 10.17521/cjpe.2019.0296 ZHANG Chan, AN Yu-Meng, Yun J?SCHKE, WANG Lin-Lin, ZHOU Zhi-Li, WANG Li-Ping, YANG Yong-Ping, DUAN Yuan-Wen. Processes on reproductive ecology of plant species in the Qinghai-Xizang Plateau and adjacent highlands. Chinese Journal of Plant Ecology, 2020, 44(1): 1-21. DOI: 10.17521/cjpe.2019.0296
素有“世界屋脊”和地球“第三极”之称的青藏高原平均海拔超过4 000 m, 面积约2.5 × 106 km2, 是世界上面积最大、海拔最高的高原。本文所指的青藏高原及周边高山地区主要包括青海省和西藏自治区的全部以及四川省、云南省和甘肃省的部分地区, 邻国喜马拉雅高山地区的部分研究也有所涉及。青藏高原气候条件极具特色, 在植物生长季节, 主要受西南季风的控制, 降水及气温由东南向西北逐渐降低(An et al., 2001; Piao et al., 2003; 卢鹤立等, 2007)。受地形和水热条件的影响, 青藏高原植被类型呈现出明显的水平和垂直地带性变化特征。整体来看, 青藏高原东南部以森林群落为主, 逐渐向西北过渡为高寒草甸、高寒草原及荒漠。青藏高原部分地区的垂直地带性变化同样非常明显, 随海拔升高由森林植被逐渐过渡为灌丛、草甸、流石滩, 而这一特征主要体现在藏东南海拔跨度较大的高山地区(郑度和陈伟烈, 1981)。
青藏高原独特的地形和气候条件孕育了极为丰富的植物多样性资源, 其中高原东南部的东喜马拉雅-横断山脉地区不仅是我国特有植物种类最为丰富的地区, 而且是现代北温带和高山植物区系的重要分化和起源中心(Huang et al., 2016)。青藏高原隆升引起地理格局和自然环境的剧烈变化, 高山、峡谷等地理屏障阻碍了物种间基因流动, 提供了异质性生境和新的生态位, 从而驱动了植物的辐射演化和种类多样化(Yu et al., 2019)。青藏高原地区维管植物种数约占中国的1/2, 其中仅分布于该区域的特有种类占35%以上(孙航, 2013), 特有种子植物共计113科519属3 764种。分子钟的证据表明, 青藏高原一些大属类群都是伴随着高原隆升分化形成的(于海彬等, 2018)。进入第四纪更新世后(2.6-0.1 Ma), 冰期-间冰期的周期性气候波动进一步造就了青藏高原现今的植物多样性分布格局(Yu et al., 2019)。谱系地理学研究指出, 该地区高山植物群体演化历史主要分为两种模式: 一、冰期退却到高原边缘的避难所, 冰后期回迁到高原面, 其中横断山脉地区常作为主要的避难所, 这也是该地区植物多样性丰富的原因之一; 二、地理隔离使得高原内部也存在很多避难所(含微型避难所), 如三江源地区, 植物在冰后期发生局域性扩张, 使得这些地区的物种多样性也比较丰富(Qiu et al., 2011)。
Fig. 1Regression of reproductive allocation on elevation of 24 plant species in the Qinghai-Xizang Plateau and adjacent highlands. A, Annual and biennial species. B, Perennial species.
Fig. 2Proportion of flower color in plant communities of different regions. A, 107 plant species in subtropical (a.s.l. 900-2 000 m) and subalpine (a.s.l. 3 000-4 100 m) regions in the Himalayas of Nepal (Shrestha et al., 2014). B, 932 plant species in Hengduan Mountains, Southwestern China (average a.s.l. > 4 300 m)(Peng et al., 2014). C, 823 plant species in Hengduan Mountains, Southwestern China (average a.s.l. > 4 300 m)(Peng et al., 2012).
近年来, 以Illumina测序技术、454测序技术和ABI测序技术为代表的第二代测序技术以及以SMRT测序技术和单分子纳米孔测序技术为代表的第三代测序技术迅速发展, 利用高通量测序技术(第二代和第三代技术)能更全面、完整、快速地了解植物复杂的序列信息, 并且为全基因组测序工作尚未完成的非模式植物提供了研究的新途径, 从而极大地推动了植物分子生物学的研究进程(Belser et al., 2018; Song et al., 2018)。近年来, 高通量测序技术在植物繁殖生态学领域中的应用也越来越广泛。
高通量测序技术使植物花部特征演化的分子机制得以深入研究, 涉及花部特征演化的基因家族序列得以分析, 与花部特征演化密切相关的基因得以挖掘。采用BSA混池测序的方法, 发现LAR1基因对沟酸浆属(Mimulus)植物花色的演化有至关重要的作用(Yuan et al., 2016); 而actin基因的显性负效突变会引起该属植物花冠管宽度变化, 并导致熊蜂访花频率改变(Ding et al., 2017)。除了基因组测序外, 转录组测序同样在植物花部特征演化研究中扮演了重要的角色。在柳叶菜科克拉花属(Clarkia)不同种群斑点形成机制的研究中发现, CgMyb1基因是导致该植物花斑点形成的原因, 该基因启动子区序列的变化影响了花斑点的位置(Jiang & Rausher, 2018)。此外, 三代全长转录组数据为没有完成基因组测序的植物物种提供了相对高质量的参考序列, 为表型相关基因的挖掘工作提供了数据基础(Chao et al., 2019; Luo et al., 2019)。
AckerlyDD, JasieńskiM (1990). Size-dependent variation of gender in high density stands of the monoecious annual, Ambrosia artemisiifolia(Asteraceae) Oecologia, 82, 474-477.
AnZS, KutzbachJE, PrellWL, PorterSC (2001). Evolution of Asian monsoons and phased uplift of the Himalaya-?Tibetan Plateau since Late Miocene times Nature, 411, 62-66. [本文引用: 1]
BakerHG, BakerI (1983). Floral nectar constituents in relation to pollinator type In: Jones CE, Little RJ eds. Handbook of Experimental Pollination Biology. Van Nostrand Reinhold, New York.117-141. [本文引用: 1]
BazzazFA, AckerlyDD, ReekieEG (2000). Reproductive allocation in plants. In: Fenner M ed. Seeds: The Ecology of Regeneration in Plant Communities. CABI Publishing, Oxford. 1-29. [本文引用: 2]
BengtssonBO, CeplitisA (2000). The balance between sexual and asexual reproduction in plants living in variable environments Journal of Evolutionary Biology, 13, 415-422. DOI:10.1046/j.1420-9101.2000.00187.xURL [本文引用: 1]
BonserSP, AarssenLW (2009). Interpreting reproductive allometry: Individual strategies of allocation explain size- dependent reproduction in plant populations Perspectives in Plant Ecology, Evolution and Systematics, 11, 31-40. DOI:10.1016/j.ppees.2008.10.003URL [本文引用: 1]
BurkleL (2008). Bottom-up Effects of Nutrient Enrichment on Plants, Pollinators, and Their Interactions PhD dissertation, Dartmouth College, New Hampshire. [本文引用: 4]
CastroS, SilveiraP, NavarroL (2008). Effect of pollination on floral longevity and costs of delaying fertilization in the out-crossing Polygala vayredae Costa (Polygalaceae) Annals of Botany, 102, 1043-1048. DOI:10.1093/aob/mcn184URL [本文引用: 1]
CeulemansT, HulsmansE, VandenEW, HonnayO (2017). Nutrient enrichment is associated with altered nectar and pollen chemical composition in Succisa pratensis Moench and increased larval mortality of its pollinator Bombus terrestris L PLOS ONE, 12, e0175160. DOI: 10.1371/journal.pone.0175160. [本文引用: 2]
ChaoQ, GaoZF, ZhangD, ZhaoBG, DongFQ, FuCX, LiuLJ, WangBC (2019). The developmental dynamics of the populus stem transcriptome Plant Biotechnology Journal, 17, 206-219. [本文引用: 1]
CharlesworthD, CharlesworthB (1981). Allocation of resources to male and female functions in hermaphrodites Biological Journal of the Linnean Society, 15, 57-74. DOI:10.1111/j.1095-8312.1981.tb00748.xURL
CharnovE (1982). The Theory of Sex Allocation. Princeton University Press, Princeton. [本文引用: 1]
ChenZF, GaoJY (2011). Study on size-dependent sex allocation in Hedychium coccineum(Zingiberaceae) Journal of Yunnan University (Natural Sciences), 33, 594-603.
CrudenRW, LyonDL (1985). Patterns of biomass allocation to male and female functions in plants with different mating systems Oecologia, 66, 299-306. DOI:10.1007/BF00379868URL [本文引用: 1]
DingBQ, MouFJ, SunW, ChenSL, PengFE, Bradshaw JrHD, YuanYW (2017). A dominant-negative actin mutation alters corolla tube width and pollinator visitation in Mimulus lewisii New Phytologist, 213, 1936-1944. DOI:10.1111/nph.2017.213.issue-4URL [本文引用: 1]
DorjiT, Totland?, MoeSR, HoppingKA, PanJB, KleinJA (2013). Plant functional traits mediate reproductive phenology and success in response to experimental warming and snow addition in Tibet Global Change Biology, 19, 459-472. [本文引用: 1]
DuanYW, DafniA, HouQZ, HeYP, LiuJQ (2010). Delayed selfing in an alpine biennial Gentianopsis paludosa(Gentianaceae) in the Qinghai-Tibetan Plateau Journal of Integrative Plant Biology, 52, 593-599. [本文引用: 1]
DuanYW, HeYP, LiuJQ (2005). Reproductive ecology of the Qinghai-Tibet Plateau endemic Gentiana straminea(Gentianaceae), a hermaphrodite perennial characterized by herkogamy and dichogamy Acta Oecologica, 27, 225-232. [本文引用: 1]
DuanYW, LiuJQ (2007). Pollinator shift and reproductive performance of the Qinghai-Tibetan Plateau endemic and endangered Swertia przewalskii(Gentianaceae) Biodiversity and Conservation, 16, 1839-1850. [本文引用: 1]
ElserJJ, BrackenMES, ClelandEE, GrunerDS, HarpoleWS, HillebrandH, NgaiJT, SeabloomEW, ShurinJB, SmithJE (2007). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems Ecology Letters, 10, 1135-1142. [本文引用: 1]
EmmettBA (2007). Nitrogen saturation of terrestrial ecosystems: Some recent findings and their implications for our conceptual framework Water, Air and Soil Pollution, 7, 99-109. [本文引用: 1]
FabbroT, K?rnerC (2004). Altitudinal differences in flower traits and reproductive allocation Flora, 199, 70-81. [本文引用: 2]
FanBL, MengJL, ZhaoZG, DuGZ (2008). Influence of altitude on reproductive traits and resource allocation of species of Ranunculaceae at east Qinghai Tibetan Plateau Acta Botanica Boreali-Occidentalia Sinica, 28, 805-811. [本文引用: 1]
FanDM, YangYP (2009). Altitudinal variations in flower and bulbil production of an alpine perennial, Polygonum viviparum L.(Polygonaceae) Plant Biology, 11, 493-497. [本文引用: 2]
FanDM, YangYP (2009). Reproductive performance in two populations of Polygonum viviparum(Polygonaceae) under different habitats Acta Botanica Yunnanica, 31, 153-157. [本文引用: 2]
FangQ, HuangSQ (2012). Relative stability of core groups in pollination networks in a biodiversity hotspot over four years PLOS ONE, 7, e32663. DOI: 10.1371/journal.pone.0032663. [本文引用: 2]
FangQ, HuangSQ (2013). A directed network analysis of heterospecific pollen transfer in a biodiverse community Ecology, 94, 1176-1185. DOI:10.1890/12-1634.1URL [本文引用: 2]
FangQ, HuangSQ (2016a). Plant-pollinator interactions in a biodiverse meadow are rather stable and tight for 3 consecutive years Integrative Zoology, 11, 199-206. DOI:10.1111/inz2.2016.11.issue-3URL [本文引用: 2]
FangQ, HuangSQ (2016b). A paradoxical mismatch between interspecific pollinator moves and heterospecific pollen receipt in a natural community Ecology, 97, 1970-1978. DOI:10.1002/ecy.1433URL [本文引用: 2]
FitterAH, FitterRSR (2002). Rapid changes in flowering time in British plants Science, 296, 1689-1691. [本文引用: 1]
GardenerMC, GillmanMP (2001). The effects of soil fertilizer on amino acids in the floral nectar of corncockle, Agrostemma githago(Caryophyllaceae) Oikos, 92, 101-106. [本文引用: 1]
GeblerM, Schoot UiterkampAJM, VisserC (2014). A global sustainability perspective on 3D printing technologies Energy Policy, 74, 158-167. DOI:10.1016/j.enpol.2014.08.033URL [本文引用: 1]
GijbelsP, CeulemansT, van den EndeW, HonnayO (2015). Experimental fertilization increases amino acid content in floral nectar, fruit set and degree of selfing in the orchid Gymnadenia conopsea Oecologia, 179, 785-795. DOI:10.1007/s00442-015-3381-8URL [本文引用: 1]
GijbelsP, van den EndeW, HonnayO (2014). Landscape scale variation in nectar amino acid and sugar composition in a Lepidoptera pollinated orchid species and its relation with fruit set Journal of Ecology, 102, 136-144. [本文引用: 1]
GongYB, HuangSQ (2009). Floral symmetry: Pollinator-?mediated stabilizing selection on flower size in bilateral species Proceedings of the Royal Society B, 276, 4013-4020. [本文引用: 1]
GongYB, HuangSQ (2011). Temporal stability of pollinator preference in an alpine plant community and its implications for the evolution of floral traits Oecologia, 166, 671-680. [本文引用: 2]
GuoH, MazerSJ, DuGZ (2010). Geographic variation in primary sex allocation per flower within and among 12 species of Pedicularis(Orobanchaceae): Proportional male investment increases with elevation American Journal of Botany, 97, 1334-1341. DOI:10.3732/ajb.0900301URL [本文引用: 1]
GuoW, WangLL, SunS, YangYP, DuanYW (2014). Sexual interference in two Chamerion species with contrasting modes of movement herkogamy Journal of Systematics and Evolution, 52, 355-362. DOI:10.1111/jse.12056URL [本文引用: 1] Evolution of herkogamy has been ascribed to preventing autonomous selfing within flowers, but this argument has received challenges due to the negative effects of sexual interference on reproductive success of plant species. We examined the effects of the two contrasting modes of herkogamy, detailed by the different patterns of style movement, on the foraging time of pollinators on flowers, pollen removal, and deposition in two Chamerion species. Our results suggested that the duration of foraging time of bumblebees was generally higher in C. angustifolium (L.) Holub than in C. conspersum (Hausskn.) Holub during the male-phase of flowers, but the reverse during the female-phase of flowers. As a result, stigma interfered with pollen removal more in C. conspersum than in C. angustifolium, based on examinations of pollen removal after one visit by a pollinator. However, this negative effect on total pollen removal was covered by the high pollinator diversity and the possible high visitation rate in the study sites. In contrast, we found that almost no difference existed in pollen deposition rate after one visit of a pollinator between the two Chamerion species, and emasculation did not increase the total pollen deposition in either species, indicating that anthers did not interfere with pollen deposition in the two Chamerion species. Our results suggested that, compared with C. conspersum, the derived mode of herkogamy in C. angustifolium reduced interference of stigmas in pollen removal. This might contribute to the wide distribution of C. angustifolium, to some degree, in environments of pollinator scarcity and low activity in newly colonized habitats.
HansenDM, van der NietT, JohnsonSD (2012). Floral signposts: Testing the significance of visual “nectar guides” for pollinator behaviour and plant fitness Proceedings of the Royal Society B, 279, 634-639. [本文引用: 1]
HeJD, XueJY, GaoJ, WangJN, WuY (2017). Adaptations of the floral characteristics and biomass allocation patterns of Gentiana hexaphylla to the altitudinal gradient of the eastern Qinghai-Tibet Plateau Journal of Mountain Science, 14, 1563-1576. DOI:10.1007/s11629-017-4424-xURL [本文引用: 1]
HeYP, DuanYW, FeiSM, LiuJQ, YangHL (2008). Resource allocation of Primula nutans population in the alpine wetland of the east Qinghai-Tibetan Plateau, China Chinese Journal of Applied and Environmental Biology, 14, 180-186.
HeYP, DuanYW, LiuJQ, SmithWK (2006). Floral closure in response to temperature and pollination in Gentiana straminea Maxim.(Gentianaceae), an alpine perennial in the Qinghai-Tibetan Plateau Plant Systematics and Evolution, 256, 17-33. [本文引用: 2]
HeglandSJ, NielsenA, LázaroA, BjerknesAL, Totland? (2009). How does climate warming affect plant-pollinator interactions? Ecology Letters, 12, 184-195. [本文引用: 1]
HonnayO, BossuytB (2005). Prolonged clonal growth: Escape route or route to extinction Oikos, 108, 427-432. [本文引用: 1]
HuaiHY, WeiWH, ZhangYL, YangCX (2005). Characteristics of clonal growth and reproduction of Lagotis brachystachya at different altitudes Chinese Journal of Applied and Environmental Biology, 11, 18-22. [本文引用: 1]
HuangSQ, LuY, ChenYZ, LuoYB, DelphLF (2009). Parthenogenesis maintains male sterility in a gynodioecious orchid The American Naturalist, 174, 578-584. [本文引用: 2]
HuangSQ, TakahashiY, DafniA (2002). Why does the flower stalk of Pulsatilla cernua(Ranunculaceae) bend during anthesis American Journal of Botany, 89, 1599-1603. [本文引用: 2]
IgicB, LandeR, KohnJR (2008). Loss of self-incompatibility and its evolutionary consequences International Journal of Plant Sciences, 169, 93-104. [本文引用: 1]
InouyeDW (1983). The ecology of nectar robbing. In: Barbara B, Thomas E, Barbara B eds. The Biology of Nectaries. Columbia University Press, New York. 153-173. [本文引用: 1]
Intergovernmental Panel on Climate Change (IPCC) (2016). IPCC Database. http://www.ipcc.ch/.Cited: Aug 24, 2018. URL [本文引用: 1]
IwasaY, CohenD (1989). Optimal growth schedule of a perennial plant The American Naturalist, 133, 480-505. DOI:10.1086/284931URL [本文引用: 1]
JiangP, RausherM (2018). Two genetic changes in Cis-?regulatory elements caused evolution of petal spot position in Carkia Nature Plants, 4, 14-22. [本文引用: 1]
K?rnerC (2003). Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems Springer, Berlin. [本文引用: 2]
KoskiMH, AshmanTL (2015). Floral pigmentation patterns provide an example of Gloger’s rule in plants Nature Plants, 1, 14007. DOI: 10.1038/nplants.?2014.7. [本文引用: 1]
Koz?owskiJ (1992). Optimal allocation of resources to growth and reproduction: Implications for age and size at maturity Trends in Ecology and Evolution, 7, 15-19. [本文引用: 1]
LangDD, TangM, ZhouX (2018). Qualitative and quantitative molecular construction of plant-pollinator network: Application and prospective Biodiversity Science, 26, 445-456. DOI:10.17520/biods.2018058URL [本文引用: 2]
LeonardAS, PapajDR (2011). “X” marks the spot: The possible benefits of nectar guides to bees and plants Functional Ecology, 25, 1293-1301. DOI:10.1111/j.1365-2435.2011.01885.xURL [本文引用: 1]
LiB, LiuZJ, ZhaoZG, HuC, RenHM, WuGQ (2013). Influence of altitude on reproductive traits and reproductive allocation of different colours in Anemone obtusiloba populations Acta Prataculturae Sinica, 22(1), 10-19. DOI:10.11686/cyxb20130102URL [本文引用: 1]
LiJJ, YangC, LiuXL, ShaoXQ (2019). Inconsistent stoichiometry response of grasses and forbs to nitrogen and water additions in an alpine meadow of the Qinghai-Tibet Plateau Agriculture, Ecosystems & Environment, 279, 178-186. [本文引用: 1]
LiangY (2009). Reproductive Allocation Strategies of Gentiana in the Eastern of Qinghai-Tibet Plateau Master degree dissertation, Northwest Normal University, Lanzhou. [本文引用: 1]
LüCQ, TianHQ (2007). Spatial and temporal patterns of nitrogen deposition in China: Synthesis of observational data Journal of Geophysical Research Atmospheres, 112, D22S05. DOI: 10.1029/2006JD007990. [本文引用: 1]
LuHL, ShaoQQ, LiuJY, WangJB, ChenZQ (2007). Temporo-?spatial distribution of summer precipitation over Qinghai-?Tibet Plateau during the last 44 years Acta Geographica Sinica, 62, 946-958. [本文引用: 1]
LuNN, ZhaoZG (2014). Flower symmetry and flower size variability: An examination of Berg’s hypotheses in an alpine meadow Chinese Journal of Plant Ecology, 38, 460-467. DOI:10.3724/SP.J.1258.2014.00042URL [本文引用: 1]
LuoD, ZhouQ, WuYG, ChaiXT, LiuWX, WangYR, YangQC, WangZY, LiuZP (2019). Full-length transcript sequencing and comparative transcriptomic analysis to evaluate the contribution of osmotic and ionic stress components towards salinity tolerance in the roots of cultivated alfalfa (Medicago sativa L.) BMC Plant Biology, 19, 32. DOI: 10.1186/s12870-019-1630-4. DOI:10.1186/s12870-019-1630-4URL [本文引用: 1]
MaRJ, DuGZ, LuBR, ChenJK, SunK, HaraT, LiB (2006). Reproductive modes of three Ligularia weeds (Asteraceae) in grasslands in Qinghai-Tibet Plateau and their implications for grassland management Ecological Research, 21, 246-254. DOI:10.1007/s11284-005-0114-1URL [本文引用: 1] Ligularia virgaurea, L. sagitta and L. przewalskii are noxious weeds that are widely distributed in psychro-grasslands in the east of Qinghai-Tibet Plateau of China. However, little is known about reproductive modes and genetic diversity in the populations of L. virgaurea and L. sagitta under different environmental conditions, although such information is available for L. przewalskii. In this study, two experiments were conducted to examine the ex situ clonal growth of these weeds, and to determine the effects of reproductive modes on the diversity of their populations, using random amplified polymorphic DNA (RAPD) analysis. Transplanted seedlings of L.sagitta and L. virgaurea showed considerable clonal growth by forming rhizomes and ramets. The RAPD analysis showed that all individuals collected from natural populations were genetically different, indicating that no individuals originated from clonal growth. It is concluded that L. virgaurea and L. sagitta have the capacity for clonal growth, but the expression of their clonality is constrained by their growing conditions in natural grasslands. Sexual reproduction plays the major role in their population diversity in natural habitats, which requires that the control measures for these weeds should be targeted at the sexually reproducing populations.
MaYX, WangYF, LiYY, WangWY, KouJ (2017). Correlation between reproductive allocation and elevation of Saussurea graminea in eastern margin of the Tibetan Plateau Bulletin of Botanical Research, 37, 23-30. [本文引用: 1]
MalerbaR, NatteroJ (2012). Pollinator response to flower color polymorphism and floral display in a plant with a single-?locus floral color polymorphism: Consequences for plant reproduction Ecological Research, 27, 377-385. DOI:10.1007/s11284-011-0908-2URL [本文引用: 1]
MengJL (2010). Reproductive Strategies of Three Common Species of Ranunculaceae at Alpine Meadow in Qinghai Tibetan Plateau PhD dissertation, Lanzhou University, Lanzhou. [本文引用: 1]
MuJP, PengYH, XiXQ, WuXW, LiGY, NiklasKJ, SunSC (2015). Artificial asymmetric warming reduces nectar yield in a Tibetan alpine species of Asteraceae Annals of Botany, 116, 899-906. DOI:10.1093/aob/mcv042URL [本文引用: 2]
MuJP, ZengYL, WuQG, NiklasKJ, NiuKC (2016). Traditional grazing regimes promote biodiversity and increase nectar production in Tibetan alpine meadows Agriculture, Ecosystems & Environment, 233, 336-342. [本文引用: 1]
Mu?ozAA, Celedon-NeghmeC, CavieresLA, ArroyoMTK (2005). Bottom-up effects of nutrient availability on flower production, pollinator visitation, and seed output in a high-Andean shrub Oecologia, 143, 126-135. DOI:10.1007/s00442-004-1780-3URL
MunsonSM, LongAL (2017). Climate drives shifts in grass reproductive phenology across the western USA New Phytologist, 213, 1945-1955. DOI:10.1111/nph.2017.213.issue-4URL [本文引用: 2]
NazirR, ReshiZ, WafaiBA (2008). Reproductive ecology of medicinally important Kashmir Himalayan species of Digitalis L Plant Species Biology, 23, 59-70. DOI:10.1111/psb.2008.23.issue-2URL
NiuY, GongQB, PengDL, SunH, LiZM (2017). Function of male and hermaphroditic flowers and size-dependent gender diphasy of Lloydia oxycarpa(Liliaceae) from Hengduan Mountains Plant Diversity, 39, 187-193. DOI:10.1016/j.pld.2017.06.001URL
PengDL, OuXK, XuB, ZhangZQ, NiuY, LiZM, SunH (2014). Plant sexual systems correlated with morphological traits: Reflecting reproductive strategies of alpine plants Journal of Systematics and Evolution, 52, 368-377. DOI:10.1111/jse.v52.3URL [本文引用: 4]
PengDL, ZhangZQ, NiuY, YangY, SongB, SunH, LiZM (2012). Advances in the studies of reproductive strategies of alpine plants Biodiversity Science, 20, 286-299. DOI:10.3724/SP.J.1003.2012.14039URL [本文引用: 5]
PengDL, ZhangZQ, XuB, LiZM, SunH (2012). Patterns of flower morphology and sexual systems in the subnival belt of the Hengduan Mountains, SW China Alpine Botany, 122(2), 65-73. [本文引用: 3]
PiaoSL, FangJY, ChenAP (2003). Seasonal dynamics of terristrial net primary production in response to climate change in China Acta Botanica Sinica, 45, 269-275. [本文引用: 1]
PolichaT, DavisA, BarnadasM, DentingerBTM, RagusoRA, RoyBA (2016). Disentangling visual and olfactory signals in mushroom-mimicking Dracula orchids using realistic three-dimensional printed flowers New Phytologist, 210, 1058-1071. DOI:10.1111/nph.13855URL [本文引用: 1]
PottsJG (2015). Effects of floral symmetry on pollination in Bidens aristosa The Southwest Naturalist, 60, 370-373. DOI:10.1894/0038-4909-60.4.370URL [本文引用: 1]
QiGH (2018). Effect of Exotic Introduced Plants Azadirachta indica and Moringa Oleifera on Pollination Network of Dry-hot Valley in Honghe Area PhD dissertation, Chinese Academy of Forestry, Beijing. [本文引用: 1]
QiuYX, FuCX, ComesHP (2011). Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora Molecular Phylogenetics and Evolution, 59, 225-244. [本文引用: 1]
QuB (2017). Variation of Floral Traits and Reproductive Adaptation along Elevational Gradient of Clematis tangutica Master degree dissertation, Northwest Normal University, Lanzhou. [本文引用: 1]
ScavenVL, RaffertyNE (2013). Physiological effects of climate warming on flowering plants and insect pollinators and potential consequences for their interactions Current Zoology, 59, 418-426. DOI:10.1093/czoolo/59.3.418URL [本文引用: 1]
ShresthaM, DyerAG, BhattaraiP, BurdM (2014). Flower colour and phylogeny along an altitudinal gradient in the Himalayas of Nepal Journal of Ecology, 102, 126-135. [本文引用: 2]
SkogenKA, HolsingerKE, CardonZG (2011). Nitrogen deposition, competition and the decline of a regionally threatened legume, Desmodium cuspidatum Oecologia, 165, 261-269. DOI:10.1007/s00442-010-1818-7URL [本文引用: 1]
SmithJG, SconiersW, SpasojevicMJ, AshtonIW, SudingKN (2012). Phenological changes in alpine plants in response to increased snowpack, temperature, and nitrogen Arctic, Antarctic, and Alpine Research, 44, 135-142. [本文引用: 1]
SunF, ZhongZC (1997). Reproductive allocation in Gordonia acuminata in subtropical evergreen broad-leaved forest and analysis of its adaptation using gray rational degree Acta Phytoecologia Sinica, 21, 44-52. [本文引用: 2]
SunH (2013). Wild plant germplasm’s collection, investigation and preservation on the special environment from Qinghai-?Tibet Plateau Plant Diversity and Resources, 35, 217-220. [本文引用: 1]
SunH, NiuY, ChenYS, SongB, LiuCQ, PengDL, ChenJG, YangY (2014). Survival and reproduction of plant species in the Qinghai-Tibet Plateau Journal of Systematics and Evolution, 52, 378-396. [本文引用: 3]
SunJF, GongYB, RennerSS, HuangSQ (2008). Multifunctional bracts in the dove tree Davidia involucrata(Nyssaceae: Cornales): Rain protection and pollinator attraction The American Naturalist, 171, 119-124. DOI:10.1086/523953URL [本文引用: 1]
TakebayashiN, MorrellPL (2001). Is self-fertilization an evolutionary dead end? Revisiting an old hypothesis with genetic theories and a macroevolutionary approach American Journal of Botany, 88, 1143-1150. [本文引用: 1]
TorabinejadJ, CaldwellMM, FlintSD, DurhamS (1998). Susceptibility of pollen to UV-B radiation: An assay of 34 taxa American Journal of Botany, 85, 360-369. [本文引用: 1]
Totland? (2004). No evidence for a role of pollinator discrimination in causing selection on flower size through female reproduction Oikos, 106, 558-564. [本文引用: 1]
TrunschkeJ, St?cklinJ (2017). Plasticity of flower longevity in alpine plants is increased in populations from high elevation compared to low elevation populations Alpine Botany, 127(1), 41-51. DOI:10.1007/s00035-016-0176-4URL [本文引用: 2]
TuYL, WangLP, WangXL, WangLL, DuanYW (2019). Status of invasive plants on local pollination networks: A case study of Tagetes minuta in Tibet based on pollen grains from pollinators Biodiversity Science, 27, 306-313. DOI:10.17520/biods.2018269URL [本文引用: 1] Invasive plants that use local pollinators would better ensure their expansion in the new environment, but the role and status of the invasive plants on local pollination network is still unclear. In this paper, we analyzed the pollen grains of Tagetes minuta carried by pollinators and constructed a plant-pollinator network, to explore the roles and impacts of T. minuta in local pollination network. The results showed that pollination system of T. minuta was generalized, and there were 13 insect species visiting its flowers, 12 of which carried pollen grains of T. minuta, and the pollen grains of T. minuta accounted for 89.89%. Among the 12 pollinator species of T. minuta, one species of bees, two species of scorpion flies and one species of flies are the main pollinators. This study reveals that T. minuta has successfully used a variety of local pollinators for pollination in a relatively short period of time, indicating that T. minuta has strong adaptability and diffusion ability, and it is necessary to pay more attentions to the prevention and control of T. minuta in the future. [ 土艳丽, 王力平, 王喜龙, 王林林, 段元文 (2019). 利用昆虫携带的花粉初探西藏入侵植物印加孔雀草在当地传粉网络中的地位 生物多样性, 27, 306-313.] DOI:10.17520/biods.2018269URL [本文引用: 1] Invasive plants that use local pollinators would better ensure their expansion in the new environment, but the role and status of the invasive plants on local pollination network is still unclear. In this paper, we analyzed the pollen grains of Tagetes minuta carried by pollinators and constructed a plant-pollinator network, to explore the roles and impacts of T. minuta in local pollination network. The results showed that pollination system of T. minuta was generalized, and there were 13 insect species visiting its flowers, 12 of which carried pollen grains of T. minuta, and the pollen grains of T. minuta accounted for 89.89%. Among the 12 pollinator species of T. minuta, one species of bees, two species of scorpion flies and one species of flies are the main pollinators. This study reveals that T. minuta has successfully used a variety of local pollinators for pollination in a relatively short period of time, indicating that T. minuta has strong adaptability and diffusion ability, and it is necessary to pay more attentions to the prevention and control of T. minuta in the future.
WangLL, ZhangC, YangML, ZhangGP, ZhangZQ, YangYP, DuanYW (2017). Intensified wind pollination mediated by pollen dimorphism after range expansion in an ambophilous biennial Aconitum gymnandrum Ecology and Evolution, 7, 541-549. DOI:10.1002/ece3.2017.7.issue-2URL [本文引用: 1]
WangLP, WangLL, HeZR, YangYP, DuanYW (2019). Application of 3D printing technology in plant reproductive ecology Guihaia, 39, 279-284. [本文引用: 1]
WangQ (2013). Biological Effects of Experimental Warming on Pollination in Impatiens Oxyanthera (Balsaminaceae) PhD dissertation, University of Chinese Academy of Sciences, Beijing. [本文引用: 1]
WangY, LuoJ, XuJC (2009). A comparative study on the pollination mechanism of Anisodus luridus and A. carniolicoides(Solanaceae) Acta Botanica Yunnanica, 31, 211-218. [本文引用: 1]
WangYX, LiuZJ, ZhaoZG, HouM, ZhangXR, LüWL (2018). Responses of floral longevity to pollination environments in 11 species from two alpine meadows Biodiversity Science, 26, 510-518. [本文引用: 1]
WebbCJ (1999). Empirical studies: Evolution and maintenance of dimorphic breeding systems In: Geber MA, Dawson TE, Delph LF eds. Gender and Sexual Dimorphism in Flowering Plants. Springer, Berlin. 61-95. [本文引用: 1]
WeinerJ, CampbellLG, PinoJ, EcharteL (2009). The allometry of reproduction within plant populations Journal of Ecology, 97, 1220-1233. [本文引用: 1]
WuY, LiuYR, PengH, YangY, LiuGL, CaoGX, ZhangQ (2015). Pollination ecology of alpine herb Meconopsis integrifolia at different altitudes Chinese Journal of Plant Ecology, 39, 1-13. [本文引用: 1]
XiaJY, WanSQ (2008). Global response patterns of terrestrial plant species to nitrogen addition New Phytologist, 179, 428-439. [本文引用: 1]
XiaoYA, ZhangSS, YanXH, DongM (2015). New advances in effects of global warming on plant-pollinator networks Acta Ecologica Sinica, 35, 3871-3880. [本文引用: 2]
XuB, WangJN, ShiFS, GaoJ, WuN (2013). Adaptation of biomass allocation patterns of wild Fritillaria unibracteata to alpine environment in the eastern Qinghai-Xizang Plateau Chinese Journal of Plant Ecology, 37, 187-196. [本文引用: 1]
XueJ, HeJ, WangL, GaoJ, WuY (2018). Plant traits and biomass allocation of Gentiana hexaphylla on different slope aspects at the eastern margin of Qinghai-Tibet Plateau Applied Ecology and Environmental Research, 16, 1835-1853. [本文引用: 1]
YangJQ, FanYL, JiangXF, LiQJ, ZhuXF (2018a). Correlation between the timing of autonomous selfing and floral traits: A comparative study from three selfing Gentianopsis species (Gentianaceae) Scientific Reports, 8, 3634. DOI: 10.1038/s41598-018-21930-9. [本文引用: 1]
YangJY (2018). Interspecific Relationships in Alpine Meadow Plant Communities: Pollinator-mediated Interactions and the Effect of Grazing Master degree dissertation, Lanzhou University, Lanzhou. [本文引用: 2]
YangML, WangLL, ZhangGP, MengLH, YangYP, DuanYW (2018b). Equipped for migrations across high latitude regions? Reduced spur length and outcrossing rate in a biennial Halenia elliptica(Gentianaceae) with mixed mating system along a latitude gradient Frontiers in Genetics, 9, 223. DOI: 10.3389/fgene.2018.00223. [本文引用: 1]
YangY, K?rnerC, SunH (2008). The ecological significance of pubescence in Saussurea medusa, a high-elevation Himalayan “woolly plant” Arctic, Antarctic, and Alpine Research, 40, 250-255. [本文引用: 1]
YangY, SunH (2009). The bracts of Saussurea velutina(Asteraceae) protect inflorescences from fluctuating weather at high elevations of the Hengduan mountains, southwestern China Arctic, Antarctic, and Alpine Research, 41, 515-521. [本文引用: 2]
YuHB, DeaneDC, SuiXH, FangSQ, ChuCJ, LiuY, HeFL (2019). Testing multiple hypotheses for the high endemic plant diversity of the Tibetan Plateau Global Ecology and Biogeography, 28, 131-144. DOI:10.1111/geb.v28.2URL [本文引用: 1]
YuHB, ZhangYL, LiuLS, ChenC, QiW (2018). Floristic characteristics and diversity patterns of seed plants endemic to the Tibetan Plateau Biodiversity Science, 26, 130-137. [本文引用: 1]
YuQ, JiaDR, TianB, YangYP, DuanYW (2016). Changes of flowering phenology and flower size in rosaceous plants from a biodiversity hotspot in the past century Scientific Reports, 6, 28302. DOI: 10.1038/srep28302. [本文引用: 2]
YuanYW, RebochoAB, SagawaJM, StanleyLE, Bradshaw JrHD (2016). Competition between anthocyanin and flavonol biosynthesis produces spatial pattern variation of floral pigments between Mimulus species Proceedings of the National Academy of Sciences of the United States of America, 113, 2448-2453. [本文引用: 2]
ZhangC, VereeckenNJ, WangLL, TianB, DafniA, YangYP, DuanYW (2017). Are nectar guide colour changes a reliable signal to pollinators that enhances reproductive success? Plant Ecology & Diversity, 10, 89-96. [本文引用: 1]
ZhangC, WangLL, YangYP, DuanYW (2015). Flower evolution of alpine forbs in the open top chambers (OTCs) from the Qinghai-Tibet Plateau Scientific Reports, 5, 10254. DOI: 10.1038/srep10254. [本文引用: 1]
ZhangC, ZhouGY, YangYP, DuanYW (2014b). Better than nothing: Evolution of autonomous selfing under strong inbreeding depression in an alpine annual from the Qinghai- Tibet Plateau Journal of Systematics and Evolution, 52, 363-367. [本文引用: 1]
ZhangDY (2004). Evolution of Plant Life History and Reproductive Ecology. Science Press, Beijing. [本文引用: 2]
ZhangFP, CaiXH, WangH, RenZX, Larson-RabinZ, LiDZ (2012). Dark purple nectar as a foraging signal in a bird-pollinated Himalayan plant New Phytologist, 193, 188-195. [本文引用: 1]
ZhangFP, YangQY, ZhangSB (2016). Dual effect of phenolic nectar on three floral visitors of Elsholtzia rugulosa(Lamiaceae) in SW China PLOS ONE, 11, e0154381. DOI: 10.1371/journal.pone.0154381. [本文引用: 1]
ZhangGP (2017). The Research of Flower Traits Evolution in Apline Plant Based on Community Phylogeny Master degree dissertation, Yunnan Normal University, Kunming. [本文引用: 1]
ZhangLC, WangXJ, DuGZ (2011a). Primary floral allocation per flower in 12 Pedicularis(Orobanchaceae) species: Significant effect of two distinct rewarding types for pollinators Journal of Plant Research, 124, 655-661.
ZhangN (2006). Effects of UV-B Radiation Photosynthesis Growth and Reproduction of Three Annuals in Alpine Meadow PhD dissertation, Lanzhou University, Lanzhou. [本文引用: 1]
ZhangS, AiHL, YuWB, WangH, LiDZ (2010). Flower heliotropism of Anemone rivularis(Ranunculaceae) in the Himalayas: Effects on floral temperature and reproductive fitness Plant Ecology, 209, 301-312. [本文引用: 1]
ZhangYW, WangY, GuoYH (2006). The effects of nectar robbing on plant reproduction and evolution Journal of Plant Ecology (Chinese Version), 30, 695-702. [本文引用: 1]
ZhangZQ, KressWJ, XieWJ, RenPY, GaoJY, LiQJ (2011b). Reproductive biology of two Himalayan alpine gingers (Roscoea spp., Zingiberaceae) in China: Pollination syndrome and compensatory floral mechanisms Plant Biology, 13, 582-589. [本文引用: 1]
ZhangZQ, ZhangYH, SunH (2011c). The reproductive biology of Stellera chamaejasme(Thymelaeaceae): A self-?incompatible weed with specialized flowers Flora, 206, 567-574. [本文引用: 1]
ZhaoBB, NiuKC, DuGZ (2009). The effect of grazing on above-ground biomass allocation of 27 plant species in an alpine meadow plant community in Qinghai-Tibetan Plateau Acta Ecologica Sinica, 29, 1596-1606. [本文引用: 1]
ZhaoF, YangYP (2008). Reproductive allocation in a dioecious perennial Oxyria sinensis(Polygonaceae) along altitudinal gradients Journal of Systematics and Evolution, 46, 830-835. [本文引用: 1]
ZhaoYH (2016). Effects of Altitude and Species Phylogeny on Pollination Network in Himalaya-Hengduan Mountains PhD dissertation, University of Chinese Academy of Sciences, Beijing. [本文引用: 3]
ZhaoYH, LázaroA, RenZX, ZhouW, LiHD, TaoZB, XuK, WuZK, WolfeLM, LiDZ, WangH (2019). The topological differences between visitation and pollen transport networks: A comparison in species rich communities of the Himalaya-Hengduan Mountains Oikos, 128, 551-562. [本文引用: 1]
ZhaoZG, DuGZ (2003). Characteristics of the mating system and strategies for resource allocation in Ranunculaceae Journal of Lanzhou University (Natural Sciences), 39, 70-74. [本文引用: 1]
ZhaoZG, DuGZ, ZhouXH, WangMT, RenQJ (2006). Variations with altitude in reproductive traits and resource allocation of three Tibetan species of Ranunculaceae Australian Journal of Botany, 54, 691-700. DOI:10.1071/BT05015URL [本文引用: 1]
ZhaoZG, LiuZJ, ConnerJK (2015). Plasticity of floral sex allocation within inflorescences of hermaphrodite Aconitum gymnandrum Journal of Plant Ecology, 8, 130-135. DOI:10.1093/jpe/rtv019URL Aims Sex allocation in plants is often plastic, enabling individuals to adjust to variable environments. However, the predicted male-biased sex allocation in response to low resource conditions has rarely been experimentally tested in hermaphroditic plants. In particular, it is unknown whether distal flowers in linear inflorescences show a larger shift to male allocation relative to basal flowers when resources are reduced. In this study, we measure position-dependent plasticity of floral sex allocation within racemes of Aconitum gymnandrum in response to reduced resource availability. Methods Using a defoliation treatment in the field applied to potted plants from a nested half-sibling design, we examined the effects of the treatment, flower position, family and their interactions. Important findings Allocation to male function increased with more distal flower position, while female allocation either did not change with position or declined at the most distal flowers. Defoliation significantly reduced the mass of both the androecium and gynoecium, but not anther number or carpel number. Gynoecial mass declined more strongly with defoliation than did androecial mass, resulting in a significant increase in the androecium/gynoecium ratio as predicted by sex allocation theory. Plastic responses of androecium mass and gynoecium mass were affected by flower position, with less mass lost in basal flowers, but similar plastic magnitude in both sexual traits across flower position lead to consistent variation in the androecium/gynoecium ratio along the inflorescence. A significant treatment*paternal family interaction for the androecium/gynoecium ratio is evidence for additive genetic variation for plastic floral sex allocation, which means that further evolution of allocation can occur.
ZhaoZG, MengJL, FanBL, DuGZ (2008). Size-dependent sex allocation in Aconitum gymnandrum(Ranunculaceae): Physiological basis and effects of maternal family and environment Plant Biology, 10, 694-703.
ZhengD, ChenWL (1981). A preliminary study on the vertical belts of vegetation of the eastern Himalayas Bulletin of Botany, 23, 228-234. [本文引用: 1]
ZhuJT (2016). Effects of experimental warming on plant reproductive phenology in Xizang alpine meadow Chinese Journal of Plant Ecology, 40, 1028-1036. [本文引用: 2]
Chromosome-?scale assemblies of plant genomes using nanopore long reads and optical maps 1 2018
... 近年来, 以Illumina测序技术、454测序技术和ABI测序技术为代表的第二代测序技术以及以SMRT测序技术和单分子纳米孔测序技术为代表的第三代测序技术迅速发展, 利用高通量测序技术(第二代和第三代技术)能更全面、完整、快速地了解植物复杂的序列信息, 并且为全基因组测序工作尚未完成的非模式植物提供了研究的新途径, 从而极大地推动了植物分子生物学的研究进程(Belser et al., 2018; Song et al., 2018).近年来, 高通量测序技术在植物繁殖生态学领域中的应用也越来越广泛. ...
The balance between sexual and asexual reproduction in plants living in variable environments 1 2000
Effect of pollination on floral longevity and costs of delaying fertilization in the out-crossing Polygala vayredae Costa (Polygalaceae) 1 2008
... 选择异交繁殖的高山植物往往借助延长柱头可授性(吴云等, 2015)和花寿命(Zhang et al., 2011b)以及大而艳丽的花展示(Peng et al., 2012)来增加授粉概率, 但是这不但需要增加花开放的资源投入, 还可能要付出一定的适合度代价(Castro et al., 2008).因此, 高山植物还会采取其他一些积极的策略来吸引昆虫, 如紫草科疏花软紫草(Arnebia szechenyi)已授粉花的花冠宿存并保持一定的颜色, 能增加整个植物的花展示面积, 为未授粉的花吸引传粉者(Zhang et al., 2017).另外, 泛化的传粉媒介也是高山植物对传粉者缺乏的适应机制, 例如露蕊乌头主要通过熊蜂传粉结实, 但是从冰期避难所向外扩张的过程中, 伴随着熊蜂访花频率的降低, 边缘种群表现出更加明显的风媒传粉特征, 并起到一定的繁殖保障作用(Wang et al., 2017).因此, 在高山地区拥有风媒和虫媒混合传粉系统的风虫媒(ambophily)植物更有优势(彭德力等, 2012). ...
Nutrient enrichment is associated with altered nectar and pollen chemical composition in Succisa pratensis Moench and increased larval mortality of its pollinator Bombus terrestris L 2 2017
... 在许多陆地生态系统中, 养分是限制初级生产力, 影响群落组成的重要因子(Elser et al., 2007).其中, 氮是限制植物生长最为重要的一个因子, 大气氮沉降通过影响土壤养分作用于植物光合作用、生长和繁殖, 从而改变群落中物种的组成和功能(Xia & Wan, 2008).氮添加通常会提高群落中禾草类植物的比例, 这可能是由于在高氮环境中禾草具有更强的竞争力(Li et al., 2019).另外, 群落中固氮植物(如豆科植物)的比例在氮添加后往往会减少, 因为当氮不再是限制性资源时, 它们将在竞争中失去优势(Emmett, 2007; Skogen et al., 2011).更为重要的是, 氮沉降通常导致植物多样性降低从而减少传粉者的食物来源, 这是当今农业过量施肥造成全球传粉者种类和数量急剧减少的主要原因(Ceulemans et al., 2017). ...
Delayed selfing in an alpine biennial Gentianopsis paludosa(Gentianaceae) in the Qinghai-Tibetan Plateau 1 2010
... 但是, 自交容易导致近交衰退(inbreeding depression)以及种群遗传多样性的丧失, 特别是在高山严酷环境下, 近交衰退更加严重, 因而不能作为长期的繁殖策略; 而且, 自交谱系不能转化为异交, 被认为是“进化的死胡同” (Takebayashi & Morrell, 2001).因此, 也有一些高山植物具有自交不亲和基础上的专性异交系统, 例如, 瑞香科瑞香狼毒(Stellera chamaejasme)(Zhang et al., 2011c).此外, 很多高山植物进化出雌雄异熟和雌雄异位等花部特征来促进异交并避免雌雄功能相互干扰, 如龙胆科麻花艽(Duan et al., 2005)和柳叶菜科柳兰(Chamerion angustifolium)(Guo et al., 2014)等.但更多的植物则是选择异交为主、自交为辅的混合型交配系统, 这似乎是应对高山环境传粉限制的最优选择, 异交的繁育系统能够提高种子的质量和后代的遗传多样性, 同时自交亲和机制为自花授粉创造了前提条件, 起到了双重繁殖保障作用(彭德力等, 2012).例如, 龙胆科椭圆叶花锚(Halenia elliptica)属于典型的混合交配系统, 低纬度种群比高纬度种群具有更高的访花频率和异花传粉概率(Yang et al., 2018b).在延迟自交(delayed selfing)这种特殊的交配系统中, 自花授粉一直延迟到异交机会丧失时才发生, 既避免了自交与异交竞争造成的花粉和种子折损, 又能保证产生后代, 对植物的雄性功能和雌性功能均有益(Duan et al., 2010).对香格里拉高山草甸3种扁蕾属(Gentianopsis)植物的交配系统研究发现, 具有延迟自交机制的大花扁蕾(G. grandis)显著提高了异交概率(Yang et al., 2018a). ...
Reproductive ecology of the Qinghai-Tibet Plateau endemic Gentiana straminea(Gentianaceae), a hermaphrodite perennial characterized by herkogamy and dichogamy 1 2005
... 但是, 自交容易导致近交衰退(inbreeding depression)以及种群遗传多样性的丧失, 特别是在高山严酷环境下, 近交衰退更加严重, 因而不能作为长期的繁殖策略; 而且, 自交谱系不能转化为异交, 被认为是“进化的死胡同” (Takebayashi & Morrell, 2001).因此, 也有一些高山植物具有自交不亲和基础上的专性异交系统, 例如, 瑞香科瑞香狼毒(Stellera chamaejasme)(Zhang et al., 2011c).此外, 很多高山植物进化出雌雄异熟和雌雄异位等花部特征来促进异交并避免雌雄功能相互干扰, 如龙胆科麻花艽(Duan et al., 2005)和柳叶菜科柳兰(Chamerion angustifolium)(Guo et al., 2014)等.但更多的植物则是选择异交为主、自交为辅的混合型交配系统, 这似乎是应对高山环境传粉限制的最优选择, 异交的繁育系统能够提高种子的质量和后代的遗传多样性, 同时自交亲和机制为自花授粉创造了前提条件, 起到了双重繁殖保障作用(彭德力等, 2012).例如, 龙胆科椭圆叶花锚(Halenia elliptica)属于典型的混合交配系统, 低纬度种群比高纬度种群具有更高的访花频率和异花传粉概率(Yang et al., 2018b).在延迟自交(delayed selfing)这种特殊的交配系统中, 自花授粉一直延迟到异交机会丧失时才发生, 既避免了自交与异交竞争造成的花粉和种子折损, 又能保证产生后代, 对植物的雄性功能和雌性功能均有益(Duan et al., 2010).对香格里拉高山草甸3种扁蕾属(Gentianopsis)植物的交配系统研究发现, 具有延迟自交机制的大花扁蕾(G. grandis)显著提高了异交概率(Yang et al., 2018a). ...
Pollinator shift and reproductive performance of the Qinghai-Tibetan Plateau endemic and endangered Swertia przewalskii(Gentianaceae) 1 2007
Geographic variation in primary sex allocation per flower within and among 12 species of Pedicularis(Orobanchaceae): Proportional male investment increases with elevation 1 2010
Adaptations of the floral characteristics and biomass allocation patterns of Gentiana hexaphylla to the altitudinal gradient of the eastern Qinghai-Tibet Plateau 1 2017
Floral closure in response to temperature and pollination in Gentiana straminea Maxim.(Gentianaceae), an alpine perennial in the Qinghai-Tibetan Plateau 2 2006
... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
... ; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
How does climate warming affect plant-pollinator interactions? 1 2009
Warming, CO2, and nitrogen deposition interactively affect a plant-pollinator mutuallism 3 2012
... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
... ).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
... 除了传粉者的选择压力, 植食动物、盗蜜行为以及非生物环境在花部特征演化中同样具有重要作用.大而鲜艳的花在吸引传粉者的同时也会引起啃食者的注意, 因而植物处于一个两难的境地(黄双全, 2014).对兰科雌花两性花异株植物缘毛鸟足兰(Satyrium nepalense var. ciliatum)的研究表明, 两性花个体花大, 吸引熊蜂访花的比例高, 但同时被昆虫啃食的比例也显著高于雌性个体, 说明花部特征受到啃食者的选择(Huang et al., 2009).通过对比未放牧、轻度放牧、适度放牧和过度放牧样地钝苞雪莲的花部特征及其花蜜量发现, 轻度和适度放牧条件下, 由于植株间对光的竞争强度减弱, 花及花序数量增加, 花蜜量增多, 对传粉者的吸引力增加(Mu et al., 2016).盗蜜(nectar robbing)特指访花者不是从花冠口进入, 而是直接在花冠上打洞并从中取食花蜜的行为(Inouye, 1983).盗蜜行为对宿主植物的适合度有影响, 因而也可以作为一种选择压力对花部进化起作用(张彦文等, 2006). ...
Why does the flower stalk of Pulsatilla cernua(Ranunculaceae) bend during anthesis 2 2002
... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
... ), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
Loss of self-incompatibility and its evolutionary consequences 1 2008
Full-length transcript sequencing and comparative transcriptomic analysis to evaluate the contribution of osmotic and ionic stress components towards salinity tolerance in the roots of cultivated alfalfa (Medicago sativa L.) 1 2019
... 高通量测序技术使植物花部特征演化的分子机制得以深入研究, 涉及花部特征演化的基因家族序列得以分析, 与花部特征演化密切相关的基因得以挖掘.采用BSA混池测序的方法, 发现LAR1基因对沟酸浆属(Mimulus)植物花色的演化有至关重要的作用(Yuan et al., 2016); 而actin基因的显性负效突变会引起该属植物花冠管宽度变化, 并导致熊蜂访花频率改变(Ding et al., 2017).除了基因组测序外, 转录组测序同样在植物花部特征演化研究中扮演了重要的角色.在柳叶菜科克拉花属(Clarkia)不同种群斑点形成机制的研究中发现, CgMyb1基因是导致该植物花斑点形成的原因, 该基因启动子区序列的变化影响了花斑点的位置(Jiang & Rausher, 2018).此外, 三代全长转录组数据为没有完成基因组测序的植物物种提供了相对高质量的参考序列, 为表型相关基因的挖掘工作提供了数据基础(Chao et al., 2019; Luo et al., 2019). ...
Reproductive modes of three Ligularia weeds (Asteraceae) in grasslands in Qinghai-Tibet Plateau and their implications for grassland management 1 2006
Pollinator response to flower color polymorphism and floral display in a plant with a single-?locus floral color polymorphism: Consequences for plant reproduction 1 2012
Proportion of flower color in plant communities of different regions. A, 107 plant species in subtropical (a.s.l. 900-2 000 m) and subalpine (a.s.l. 3 000-4 100 m) regions in the Himalayas of Nepal (<xref ref-type="bibr" rid="b103">Shrestha <i>et al</i>., 2014</xref>). B, 932 plant species in Hengduan Mountains, Southwestern China (average a.s.l. > 4 300 m)(<xref ref-type="bibr" rid="b92">Peng <i>et al</i>., 2014</xref>). C, 823 plant species in Hengduan Mountains, Southwestern China (average a.s.l. > 4 300 m)(<xref ref-type="bibr" rid="b94">Peng <i>et al</i>., 2012</xref>).Fig. 2 值得注意的是, 蜜蜂养殖业的发展向自然生态系统中释放了大量的人工饲养蜜蜂, 进而对本土植物的花部特征演化产生了一定的选择压力.对青藏高原地区菊科钝苞雪莲(Saussurea nigrescens)的研究发现, 养蜂场附近的植株花蜜分泌量减少了60%以上, 并将结余的资源用于生产更多的花和种子(Mu et al., 2014).外来传粉者可能通过影响植物花部特征的进化而进一步作用于本地传粉者的访花行为和种群动态, 这对当地传粉网络稳定性的影响亟待深入研究.但当本地传粉昆虫缺乏时, 人工饲养的蜜蜂可协助本地植物完成传粉过程(Duan & Liu, 2007). ...
... ). B, 932 plant species in Hengduan Mountains, Southwestern China (average a.s.l. > 4 300 m)(Peng et al., 2014). C, 823 plant species in Hengduan Mountains, Southwestern China (average a.s.l. > 4 300 m)(Peng et al., 2012). Fig. 2 值得注意的是, 蜜蜂养殖业的发展向自然生态系统中释放了大量的人工饲养蜜蜂, 进而对本土植物的花部特征演化产生了一定的选择压力.对青藏高原地区菊科钝苞雪莲(Saussurea nigrescens)的研究发现, 养蜂场附近的植株花蜜分泌量减少了60%以上, 并将结余的资源用于生产更多的花和种子(Mu et al., 2014).外来传粉者可能通过影响植物花部特征的进化而进一步作用于本地传粉者的访花行为和种群动态, 这对当地传粉网络稳定性的影响亟待深入研究.但当本地传粉昆虫缺乏时, 人工饲养的蜜蜂可协助本地植物完成传粉过程(Duan & Liu, 2007). ...
Proportion of flower color in plant communities of different regions. A, 107 plant species in subtropical (a.s.l. 900-2 000 m) and subalpine (a.s.l. 3 000-4 100 m) regions in the Himalayas of Nepal (<xref ref-type="bibr" rid="b103">Shrestha <i>et al</i>., 2014</xref>). B, 932 plant species in Hengduan Mountains, Southwestern China (average a.s.l. > 4 300 m)(<xref ref-type="bibr" rid="b92">Peng <i>et al</i>., 2014</xref>). C, 823 plant species in Hengduan Mountains, Southwestern China (average a.s.l. > 4 300 m)(<xref ref-type="bibr" rid="b94">Peng <i>et al</i>., 2012</xref>).Fig. 2 值得注意的是, 蜜蜂养殖业的发展向自然生态系统中释放了大量的人工饲养蜜蜂, 进而对本土植物的花部特征演化产生了一定的选择压力.对青藏高原地区菊科钝苞雪莲(Saussurea nigrescens)的研究发现, 养蜂场附近的植株花蜜分泌量减少了60%以上, 并将结余的资源用于生产更多的花和种子(Mu et al., 2014).外来传粉者可能通过影响植物花部特征的进化而进一步作用于本地传粉者的访花行为和种群动态, 这对当地传粉网络稳定性的影响亟待深入研究.但当本地传粉昆虫缺乏时, 人工饲养的蜜蜂可协助本地植物完成传粉过程(Duan & Liu, 2007). ...
... ). C, 823 plant species in Hengduan Mountains, Southwestern China (average a.s.l. > 4 300 m)(Peng et al., 2012). Fig. 2 值得注意的是, 蜜蜂养殖业的发展向自然生态系统中释放了大量的人工饲养蜜蜂, 进而对本土植物的花部特征演化产生了一定的选择压力.对青藏高原地区菊科钝苞雪莲(Saussurea nigrescens)的研究发现, 养蜂场附近的植株花蜜分泌量减少了60%以上, 并将结余的资源用于生产更多的花和种子(Mu et al., 2014).外来传粉者可能通过影响植物花部特征的进化而进一步作用于本地传粉者的访花行为和种群动态, 这对当地传粉网络稳定性的影响亟待深入研究.但当本地传粉昆虫缺乏时, 人工饲养的蜜蜂可协助本地植物完成传粉过程(Duan & Liu, 2007). ...
Seasonal dynamics of terristrial net primary production in response to climate change in China 1 2003
... 素有“世界屋脊”和地球“第三极”之称的青藏高原平均海拔超过4 000 m, 面积约2.5 × 106 km2, 是世界上面积最大、海拔最高的高原.本文所指的青藏高原及周边高山地区主要包括青海省和西藏自治区的全部以及四川省、云南省和甘肃省的部分地区, 邻国喜马拉雅高山地区的部分研究也有所涉及.青藏高原气候条件极具特色, 在植物生长季节, 主要受西南季风的控制, 降水及气温由东南向西北逐渐降低(An et al., 2001; Piao et al., 2003; 卢鹤立等, 2007).受地形和水热条件的影响, 青藏高原植被类型呈现出明显的水平和垂直地带性变化特征.整体来看, 青藏高原东南部以森林群落为主, 逐渐向西北过渡为高寒草甸、高寒草原及荒漠.青藏高原部分地区的垂直地带性变化同样非常明显, 随海拔升高由森林植被逐渐过渡为灌丛、草甸、流石滩, 而这一特征主要体现在藏东南海拔跨度较大的高山地区(郑度和陈伟烈, 1981). ...
Disentangling visual and olfactory signals in mushroom-mimicking Dracula orchids using realistic three-dimensional printed flowers 1 2016
Plant molecular phylogeography in China and adjacent regions: Tracing the genetic imprints of Quaternary climate and environmental change in the world’s most diverse temperate flora 1 2011
... 青藏高原独特的地形和气候条件孕育了极为丰富的植物多样性资源, 其中高原东南部的东喜马拉雅-横断山脉地区不仅是我国特有植物种类最为丰富的地区, 而且是现代北温带和高山植物区系的重要分化和起源中心(Huang et al., 2016).青藏高原隆升引起地理格局和自然环境的剧烈变化, 高山、峡谷等地理屏障阻碍了物种间基因流动, 提供了异质性生境和新的生态位, 从而驱动了植物的辐射演化和种类多样化(Yu et al., 2019).青藏高原地区维管植物种数约占中国的1/2, 其中仅分布于该区域的特有种类占35%以上(孙航, 2013), 特有种子植物共计113科519属3 764种.分子钟的证据表明, 青藏高原一些大属类群都是伴随着高原隆升分化形成的(于海彬等, 2018).进入第四纪更新世后(2.6-0.1 Ma), 冰期-间冰期的周期性气候波动进一步造就了青藏高原现今的植物多样性分布格局(Yu et al., 2019).谱系地理学研究指出, 该地区高山植物群体演化历史主要分为两种模式: 一、冰期退却到高原边缘的避难所, 冰后期回迁到高原面, 其中横断山脉地区常作为主要的避难所, 这也是该地区植物多样性丰富的原因之一; 二、地理隔离使得高原内部也存在很多避难所(含微型避难所), 如三江源地区, 植物在冰后期发生局域性扩张, 使得这些地区的物种多样性也比较丰富(Qiu et al., 2011). ...
甘青铁线莲花部特征的海拔变异及其繁殖适应 1 2017
... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
甘青铁线莲花部特征的海拔变异及其繁殖适应 1 2017
... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
Physiological effects of climate warming on flowering plants and insect pollinators and potential consequences for their interactions 1 2013
... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
Proportion of flower color in plant communities of different regions. A, 107 plant species in subtropical (a.s.l. 900-2 000 m) and subalpine (a.s.l. 3 000-4 100 m) regions in the Himalayas of Nepal (<xref ref-type="bibr" rid="b103">Shrestha <i>et al</i>., 2014</xref>). B, 932 plant species in Hengduan Mountains, Southwestern China (average a.s.l. > 4 300 m)(<xref ref-type="bibr" rid="b92">Peng <i>et al</i>., 2014</xref>). C, 823 plant species in Hengduan Mountains, Southwestern China (average a.s.l. > 4 300 m)(<xref ref-type="bibr" rid="b94">Peng <i>et al</i>., 2012</xref>).Fig. 2 值得注意的是, 蜜蜂养殖业的发展向自然生态系统中释放了大量的人工饲养蜜蜂, 进而对本土植物的花部特征演化产生了一定的选择压力.对青藏高原地区菊科钝苞雪莲(Saussurea nigrescens)的研究发现, 养蜂场附近的植株花蜜分泌量减少了60%以上, 并将结余的资源用于生产更多的花和种子(Mu et al., 2014).外来传粉者可能通过影响植物花部特征的进化而进一步作用于本地传粉者的访花行为和种群动态, 这对当地传粉网络稳定性的影响亟待深入研究.但当本地传粉昆虫缺乏时, 人工饲养的蜜蜂可协助本地植物完成传粉过程(Duan & Liu, 2007). ...
... Proportion of flower color in plant communities of different regions. A, 107 plant species in subtropical (a.s.l. 900-2 000 m) and subalpine (a.s.l. 3 000-4 100 m) regions in the Himalayas of Nepal (Shrestha et al., 2014). B, 932 plant species in Hengduan Mountains, Southwestern China (average a.s.l. > 4 300 m)(Peng et al., 2014). C, 823 plant species in Hengduan Mountains, Southwestern China (average a.s.l. > 4 300 m)(Peng et al., 2012). Fig. 2 值得注意的是, 蜜蜂养殖业的发展向自然生态系统中释放了大量的人工饲养蜜蜂, 进而对本土植物的花部特征演化产生了一定的选择压力.对青藏高原地区菊科钝苞雪莲(Saussurea nigrescens)的研究发现, 养蜂场附近的植株花蜜分泌量减少了60%以上, 并将结余的资源用于生产更多的花和种子(Mu et al., 2014).外来传粉者可能通过影响植物花部特征的进化而进一步作用于本地传粉者的访花行为和种群动态, 这对当地传粉网络稳定性的影响亟待深入研究.但当本地传粉昆虫缺乏时, 人工饲养的蜜蜂可协助本地植物完成传粉过程(Duan & Liu, 2007). ...
Nitrogen deposition, competition and the decline of a regionally threatened legume, Desmodium cuspidatum 1 2011
... 在许多陆地生态系统中, 养分是限制初级生产力, 影响群落组成的重要因子(Elser et al., 2007).其中, 氮是限制植物生长最为重要的一个因子, 大气氮沉降通过影响土壤养分作用于植物光合作用、生长和繁殖, 从而改变群落中物种的组成和功能(Xia & Wan, 2008).氮添加通常会提高群落中禾草类植物的比例, 这可能是由于在高氮环境中禾草具有更强的竞争力(Li et al., 2019).另外, 群落中固氮植物(如豆科植物)的比例在氮添加后往往会减少, 因为当氮不再是限制性资源时, 它们将在竞争中失去优势(Emmett, 2007; Skogen et al., 2011).更为重要的是, 氮沉降通常导致植物多样性降低从而减少传粉者的食物来源, 这是当今农业过量施肥造成全球传粉者种类和数量急剧减少的主要原因(Ceulemans et al., 2017). ...
Phenological changes in alpine plants in response to increased snowpack, temperature, and nitrogen 1 2012
OsFTIP7 determines auxin-mediated anther dehiscence in rice 1 2018
... 近年来, 以Illumina测序技术、454测序技术和ABI测序技术为代表的第二代测序技术以及以SMRT测序技术和单分子纳米孔测序技术为代表的第三代测序技术迅速发展, 利用高通量测序技术(第二代和第三代技术)能更全面、完整、快速地了解植物复杂的序列信息, 并且为全基因组测序工作尚未完成的非模式植物提供了研究的新途径, 从而极大地推动了植物分子生物学的研究进程(Belser et al., 2018; Song et al., 2018).近年来, 高通量测序技术在植物繁殖生态学领域中的应用也越来越广泛. ...
四川大头茶繁殖分配及其环境适应性的关联度研究 2 1997
... 青藏高原独特的地形和气候条件孕育了极为丰富的植物多样性资源, 其中高原东南部的东喜马拉雅-横断山脉地区不仅是我国特有植物种类最为丰富的地区, 而且是现代北温带和高山植物区系的重要分化和起源中心(Huang et al., 2016).青藏高原隆升引起地理格局和自然环境的剧烈变化, 高山、峡谷等地理屏障阻碍了物种间基因流动, 提供了异质性生境和新的生态位, 从而驱动了植物的辐射演化和种类多样化(Yu et al., 2019).青藏高原地区维管植物种数约占中国的1/2, 其中仅分布于该区域的特有种类占35%以上(孙航, 2013), 特有种子植物共计113科519属3 764种.分子钟的证据表明, 青藏高原一些大属类群都是伴随着高原隆升分化形成的(于海彬等, 2018).进入第四纪更新世后(2.6-0.1 Ma), 冰期-间冰期的周期性气候波动进一步造就了青藏高原现今的植物多样性分布格局(Yu et al., 2019).谱系地理学研究指出, 该地区高山植物群体演化历史主要分为两种模式: 一、冰期退却到高原边缘的避难所, 冰后期回迁到高原面, 其中横断山脉地区常作为主要的避难所, 这也是该地区植物多样性丰富的原因之一; 二、地理隔离使得高原内部也存在很多避难所(含微型避难所), 如三江源地区, 植物在冰后期发生局域性扩张, 使得这些地区的物种多样性也比较丰富(Qiu et al., 2011). ...
Correlation between the timing of autonomous selfing and floral traits: A comparative study from three selfing Gentianopsis species (Gentianaceae) 1 2018a
... 但是, 自交容易导致近交衰退(inbreeding depression)以及种群遗传多样性的丧失, 特别是在高山严酷环境下, 近交衰退更加严重, 因而不能作为长期的繁殖策略; 而且, 自交谱系不能转化为异交, 被认为是“进化的死胡同” (Takebayashi & Morrell, 2001).因此, 也有一些高山植物具有自交不亲和基础上的专性异交系统, 例如, 瑞香科瑞香狼毒(Stellera chamaejasme)(Zhang et al., 2011c).此外, 很多高山植物进化出雌雄异熟和雌雄异位等花部特征来促进异交并避免雌雄功能相互干扰, 如龙胆科麻花艽(Duan et al., 2005)和柳叶菜科柳兰(Chamerion angustifolium)(Guo et al., 2014)等.但更多的植物则是选择异交为主、自交为辅的混合型交配系统, 这似乎是应对高山环境传粉限制的最优选择, 异交的繁育系统能够提高种子的质量和后代的遗传多样性, 同时自交亲和机制为自花授粉创造了前提条件, 起到了双重繁殖保障作用(彭德力等, 2012).例如, 龙胆科椭圆叶花锚(Halenia elliptica)属于典型的混合交配系统, 低纬度种群比高纬度种群具有更高的访花频率和异花传粉概率(Yang et al., 2018b).在延迟自交(delayed selfing)这种特殊的交配系统中, 自花授粉一直延迟到异交机会丧失时才发生, 既避免了自交与异交竞争造成的花粉和种子折损, 又能保证产生后代, 对植物的雄性功能和雌性功能均有益(Duan et al., 2010).对香格里拉高山草甸3种扁蕾属(Gentianopsis)植物的交配系统研究发现, 具有延迟自交机制的大花扁蕾(G. grandis)显著提高了异交概率(Yang et al., 2018a). ...
Equipped for migrations across high latitude regions? Reduced spur length and outcrossing rate in a biennial Halenia elliptica(Gentianaceae) with mixed mating system along a latitude gradient 1 2018b
... 但是, 自交容易导致近交衰退(inbreeding depression)以及种群遗传多样性的丧失, 特别是在高山严酷环境下, 近交衰退更加严重, 因而不能作为长期的繁殖策略; 而且, 自交谱系不能转化为异交, 被认为是“进化的死胡同” (Takebayashi & Morrell, 2001).因此, 也有一些高山植物具有自交不亲和基础上的专性异交系统, 例如, 瑞香科瑞香狼毒(Stellera chamaejasme)(Zhang et al., 2011c).此外, 很多高山植物进化出雌雄异熟和雌雄异位等花部特征来促进异交并避免雌雄功能相互干扰, 如龙胆科麻花艽(Duan et al., 2005)和柳叶菜科柳兰(Chamerion angustifolium)(Guo et al., 2014)等.但更多的植物则是选择异交为主、自交为辅的混合型交配系统, 这似乎是应对高山环境传粉限制的最优选择, 异交的繁育系统能够提高种子的质量和后代的遗传多样性, 同时自交亲和机制为自花授粉创造了前提条件, 起到了双重繁殖保障作用(彭德力等, 2012).例如, 龙胆科椭圆叶花锚(Halenia elliptica)属于典型的混合交配系统, 低纬度种群比高纬度种群具有更高的访花频率和异花传粉概率(Yang et al., 2018b).在延迟自交(delayed selfing)这种特殊的交配系统中, 自花授粉一直延迟到异交机会丧失时才发生, 既避免了自交与异交竞争造成的花粉和种子折损, 又能保证产生后代, 对植物的雄性功能和雌性功能均有益(Duan et al., 2010).对香格里拉高山草甸3种扁蕾属(Gentianopsis)植物的交配系统研究发现, 具有延迟自交机制的大花扁蕾(G. grandis)显著提高了异交概率(Yang et al., 2018a). ...
The ecological significance of pubescence in Saussurea medusa, a high-elevation Himalayan “woolly plant” 1 2008
... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
The bracts of Saussurea velutina(Asteraceae) protect inflorescences from fluctuating weather at high elevations of the Hengduan mountains, southwestern China 2 2009
... 青藏高原独特的地形和气候条件孕育了极为丰富的植物多样性资源, 其中高原东南部的东喜马拉雅-横断山脉地区不仅是我国特有植物种类最为丰富的地区, 而且是现代北温带和高山植物区系的重要分化和起源中心(Huang et al., 2016).青藏高原隆升引起地理格局和自然环境的剧烈变化, 高山、峡谷等地理屏障阻碍了物种间基因流动, 提供了异质性生境和新的生态位, 从而驱动了植物的辐射演化和种类多样化(Yu et al., 2019).青藏高原地区维管植物种数约占中国的1/2, 其中仅分布于该区域的特有种类占35%以上(孙航, 2013), 特有种子植物共计113科519属3 764种.分子钟的证据表明, 青藏高原一些大属类群都是伴随着高原隆升分化形成的(于海彬等, 2018).进入第四纪更新世后(2.6-0.1 Ma), 冰期-间冰期的周期性气候波动进一步造就了青藏高原现今的植物多样性分布格局(Yu et al., 2019).谱系地理学研究指出, 该地区高山植物群体演化历史主要分为两种模式: 一、冰期退却到高原边缘的避难所, 冰后期回迁到高原面, 其中横断山脉地区常作为主要的避难所, 这也是该地区植物多样性丰富的原因之一; 二、地理隔离使得高原内部也存在很多避难所(含微型避难所), 如三江源地区, 植物在冰后期发生局域性扩张, 使得这些地区的物种多样性也比较丰富(Qiu et al., 2011). ...
... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
Testing multiple hypotheses for the high endemic plant diversity of the Tibetan Plateau 1 2019
... 青藏高原独特的地形和气候条件孕育了极为丰富的植物多样性资源, 其中高原东南部的东喜马拉雅-横断山脉地区不仅是我国特有植物种类最为丰富的地区, 而且是现代北温带和高山植物区系的重要分化和起源中心(Huang et al., 2016).青藏高原隆升引起地理格局和自然环境的剧烈变化, 高山、峡谷等地理屏障阻碍了物种间基因流动, 提供了异质性生境和新的生态位, 从而驱动了植物的辐射演化和种类多样化(Yu et al., 2019).青藏高原地区维管植物种数约占中国的1/2, 其中仅分布于该区域的特有种类占35%以上(孙航, 2013), 特有种子植物共计113科519属3 764种.分子钟的证据表明, 青藏高原一些大属类群都是伴随着高原隆升分化形成的(于海彬等, 2018).进入第四纪更新世后(2.6-0.1 Ma), 冰期-间冰期的周期性气候波动进一步造就了青藏高原现今的植物多样性分布格局(Yu et al., 2019).谱系地理学研究指出, 该地区高山植物群体演化历史主要分为两种模式: 一、冰期退却到高原边缘的避难所, 冰后期回迁到高原面, 其中横断山脉地区常作为主要的避难所, 这也是该地区植物多样性丰富的原因之一; 二、地理隔离使得高原内部也存在很多避难所(含微型避难所), 如三江源地区, 植物在冰后期发生局域性扩张, 使得这些地区的物种多样性也比较丰富(Qiu et al., 2011). ...
青藏高原特有种子植物区系特征及多样性分布格局 1 2018
... 青藏高原独特的地形和气候条件孕育了极为丰富的植物多样性资源, 其中高原东南部的东喜马拉雅-横断山脉地区不仅是我国特有植物种类最为丰富的地区, 而且是现代北温带和高山植物区系的重要分化和起源中心(Huang et al., 2016).青藏高原隆升引起地理格局和自然环境的剧烈变化, 高山、峡谷等地理屏障阻碍了物种间基因流动, 提供了异质性生境和新的生态位, 从而驱动了植物的辐射演化和种类多样化(Yu et al., 2019).青藏高原地区维管植物种数约占中国的1/2, 其中仅分布于该区域的特有种类占35%以上(孙航, 2013), 特有种子植物共计113科519属3 764种.分子钟的证据表明, 青藏高原一些大属类群都是伴随着高原隆升分化形成的(于海彬等, 2018).进入第四纪更新世后(2.6-0.1 Ma), 冰期-间冰期的周期性气候波动进一步造就了青藏高原现今的植物多样性分布格局(Yu et al., 2019).谱系地理学研究指出, 该地区高山植物群体演化历史主要分为两种模式: 一、冰期退却到高原边缘的避难所, 冰后期回迁到高原面, 其中横断山脉地区常作为主要的避难所, 这也是该地区植物多样性丰富的原因之一; 二、地理隔离使得高原内部也存在很多避难所(含微型避难所), 如三江源地区, 植物在冰后期发生局域性扩张, 使得这些地区的物种多样性也比较丰富(Qiu et al., 2011). ...
青藏高原特有种子植物区系特征及多样性分布格局 1 2018
... 青藏高原独特的地形和气候条件孕育了极为丰富的植物多样性资源, 其中高原东南部的东喜马拉雅-横断山脉地区不仅是我国特有植物种类最为丰富的地区, 而且是现代北温带和高山植物区系的重要分化和起源中心(Huang et al., 2016).青藏高原隆升引起地理格局和自然环境的剧烈变化, 高山、峡谷等地理屏障阻碍了物种间基因流动, 提供了异质性生境和新的生态位, 从而驱动了植物的辐射演化和种类多样化(Yu et al., 2019).青藏高原地区维管植物种数约占中国的1/2, 其中仅分布于该区域的特有种类占35%以上(孙航, 2013), 特有种子植物共计113科519属3 764种.分子钟的证据表明, 青藏高原一些大属类群都是伴随着高原隆升分化形成的(于海彬等, 2018).进入第四纪更新世后(2.6-0.1 Ma), 冰期-间冰期的周期性气候波动进一步造就了青藏高原现今的植物多样性分布格局(Yu et al., 2019).谱系地理学研究指出, 该地区高山植物群体演化历史主要分为两种模式: 一、冰期退却到高原边缘的避难所, 冰后期回迁到高原面, 其中横断山脉地区常作为主要的避难所, 这也是该地区植物多样性丰富的原因之一; 二、地理隔离使得高原内部也存在很多避难所(含微型避难所), 如三江源地区, 植物在冰后期发生局域性扩张, 使得这些地区的物种多样性也比较丰富(Qiu et al., 2011). ...
Changes of flowering phenology and flower size in rosaceous plants from a biodiversity hotspot in the past century 2 2016
... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
基于群落系统发育探讨高山植物的花部特征演化 1 2017
... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
Primary floral allocation per flower in 12 Pedicularis(Orobanchaceae) species: Significant effect of two distinct rewarding types for pollinators 2011a
Flower heliotropism of Anemone rivularis(Ranunculaceae) in the Himalayas: Effects on floral temperature and reproductive fitness 1 2010
... 低温胁迫、暴雨频发以及强烈的紫外辐射是高山环境的典型特征.为应对这一系列极端环境, 高山植物往往会增加对支持及保护结构的投资, 如毛茛科甘青铁线莲(Clematis tangutica)在高海拔地区萼片十分发达, 以此来抵御外界不利因素对雌雄配子体的伤害(屈博, 2017).温度是植物生长与繁殖过程中重要的环境因子, 花发育、花蜜分泌等过程均受温度影响, 特别是在受到低温胁迫的高山植物带, 温度在花的形态、颜色和报酬等特征的进化过程中发挥着重要作用(Scaven & Rafferty, 2013).例如, “温室植物” (green house plant)菊科毡毛雪莲(Saussurea velutina)的苞片对花序具有保温作用, 有助于其发育(Yang & Sun, 2009); “棉毛植物” (cotton plant)菊科水母雪兔子(Saussurea medusa)的绒毛苞片除了具有保温效应, 还具有防水以及反射瞬时极端辐射的作用(Yang et al., 2008).此外, 在存在热量限制的高山地区, 向日运动(Zhang et al., 2010)及花闭合现象(He et al., 2006)也都是对低温环境较为常见的适应机制.大量研究发现, 花数量、花大小和花的对称性等特征均与温度紧密相关(Hoover et al., 2012; Yu et al., 2016).对青藏高原嵩草草甸64种开花植物花大小与花温度的测量结果表明, 花温度积累与花大小呈正相关关系, 且两侧对称花的温度积累比辐射对称花更为迅速(张国鹏, 2017).此外, 温度还可能通过影响植株的花蜜量、花蜜浓度和花蜜成分, 改变对传粉者的吸引力, 甚至影响传粉者的寿命(Hoover et al., 2012; Mu et al., 2015).高山带降水事件频发, 雨水会冲刷掉部分花粉、降低花粉的活性并稀释花蜜浓度(Huang et al., 2002; He et al., 2006), 因此一些植物进化出特殊的花部结构(比如下垂的花冠或者起遮盖作用的苞片等)来防止花粉受到雨水的影响(Huang et al., 2002; Sun et al., 2008).紫外辐射也会严重降低花粉的活性(Torabinejad et al., 1998).对横断山区的42种高山植物两种不同花部结构特征(即花粉暴露型与花粉遮盖型)研究表明, 花部具有的保护结构能有效地阻挡紫外辐射对花粉活力的影响(Zhang et al., 2014a).另外, 一些植物还演化出能够吸收紫外光的花部区域, 从而保护花粉等繁殖器官免受紫外辐射的伤害(Koski & Ashman, 2015).以上结果均表明紫外辐射在高山植物花部特征的进化过程中起到了非常重要的作用. ...
盗蜜行为在植物繁殖生态学中的意义 1 2006
... 除了传粉者的选择压力, 植食动物、盗蜜行为以及非生物环境在花部特征演化中同样具有重要作用.大而鲜艳的花在吸引传粉者的同时也会引起啃食者的注意, 因而植物处于一个两难的境地(黄双全, 2014).对兰科雌花两性花异株植物缘毛鸟足兰(Satyrium nepalense var. ciliatum)的研究表明, 两性花个体花大, 吸引熊蜂访花的比例高, 但同时被昆虫啃食的比例也显著高于雌性个体, 说明花部特征受到啃食者的选择(Huang et al., 2009).通过对比未放牧、轻度放牧、适度放牧和过度放牧样地钝苞雪莲的花部特征及其花蜜量发现, 轻度和适度放牧条件下, 由于植株间对光的竞争强度减弱, 花及花序数量增加, 花蜜量增多, 对传粉者的吸引力增加(Mu et al., 2016).盗蜜(nectar robbing)特指访花者不是从花冠口进入, 而是直接在花冠上打洞并从中取食花蜜的行为(Inouye, 1983).盗蜜行为对宿主植物的适合度有影响, 因而也可以作为一种选择压力对花部进化起作用(张彦文等, 2006). ...
盗蜜行为在植物繁殖生态学中的意义 1 2006
... 除了传粉者的选择压力, 植食动物、盗蜜行为以及非生物环境在花部特征演化中同样具有重要作用.大而鲜艳的花在吸引传粉者的同时也会引起啃食者的注意, 因而植物处于一个两难的境地(黄双全, 2014).对兰科雌花两性花异株植物缘毛鸟足兰(Satyrium nepalense var. ciliatum)的研究表明, 两性花个体花大, 吸引熊蜂访花的比例高, 但同时被昆虫啃食的比例也显著高于雌性个体, 说明花部特征受到啃食者的选择(Huang et al., 2009).通过对比未放牧、轻度放牧、适度放牧和过度放牧样地钝苞雪莲的花部特征及其花蜜量发现, 轻度和适度放牧条件下, 由于植株间对光的竞争强度减弱, 花及花序数量增加, 花蜜量增多, 对传粉者的吸引力增加(Mu et al., 2016).盗蜜(nectar robbing)特指访花者不是从花冠口进入, 而是直接在花冠上打洞并从中取食花蜜的行为(Inouye, 1983).盗蜜行为对宿主植物的适合度有影响, 因而也可以作为一种选择压力对花部进化起作用(张彦文等, 2006). ...
Reproductive biology of two Himalayan alpine gingers (Roscoea spp., Zingiberaceae) in China: Pollination syndrome and compensatory floral mechanisms 1 2011b
... 选择异交繁殖的高山植物往往借助延长柱头可授性(吴云等, 2015)和花寿命(Zhang et al., 2011b)以及大而艳丽的花展示(Peng et al., 2012)来增加授粉概率, 但是这不但需要增加花开放的资源投入, 还可能要付出一定的适合度代价(Castro et al., 2008).因此, 高山植物还会采取其他一些积极的策略来吸引昆虫, 如紫草科疏花软紫草(Arnebia szechenyi)已授粉花的花冠宿存并保持一定的颜色, 能增加整个植物的花展示面积, 为未授粉的花吸引传粉者(Zhang et al., 2017).另外, 泛化的传粉媒介也是高山植物对传粉者缺乏的适应机制, 例如露蕊乌头主要通过熊蜂传粉结实, 但是从冰期避难所向外扩张的过程中, 伴随着熊蜂访花频率的降低, 边缘种群表现出更加明显的风媒传粉特征, 并起到一定的繁殖保障作用(Wang et al., 2017).因此, 在高山地区拥有风媒和虫媒混合传粉系统的风虫媒(ambophily)植物更有优势(彭德力等, 2012). ...
The reproductive biology of Stellera chamaejasme(Thymelaeaceae): A self-?incompatible weed with specialized flowers 1 2011c
... 但是, 自交容易导致近交衰退(inbreeding depression)以及种群遗传多样性的丧失, 特别是在高山严酷环境下, 近交衰退更加严重, 因而不能作为长期的繁殖策略; 而且, 自交谱系不能转化为异交, 被认为是“进化的死胡同” (Takebayashi & Morrell, 2001).因此, 也有一些高山植物具有自交不亲和基础上的专性异交系统, 例如, 瑞香科瑞香狼毒(Stellera chamaejasme)(Zhang et al., 2011c).此外, 很多高山植物进化出雌雄异熟和雌雄异位等花部特征来促进异交并避免雌雄功能相互干扰, 如龙胆科麻花艽(Duan et al., 2005)和柳叶菜科柳兰(Chamerion angustifolium)(Guo et al., 2014)等.但更多的植物则是选择异交为主、自交为辅的混合型交配系统, 这似乎是应对高山环境传粉限制的最优选择, 异交的繁育系统能够提高种子的质量和后代的遗传多样性, 同时自交亲和机制为自花授粉创造了前提条件, 起到了双重繁殖保障作用(彭德力等, 2012).例如, 龙胆科椭圆叶花锚(Halenia elliptica)属于典型的混合交配系统, 低纬度种群比高纬度种群具有更高的访花频率和异花传粉概率(Yang et al., 2018b).在延迟自交(delayed selfing)这种特殊的交配系统中, 自花授粉一直延迟到异交机会丧失时才发生, 既避免了自交与异交竞争造成的花粉和种子折损, 又能保证产生后代, 对植物的雄性功能和雌性功能均有益(Duan et al., 2010).对香格里拉高山草甸3种扁蕾属(Gentianopsis)植物的交配系统研究发现, 具有延迟自交机制的大花扁蕾(G. grandis)显著提高了异交概率(Yang et al., 2018a). ...
The topological differences between visitation and pollen transport networks: A comparison in species rich communities of the Himalaya-Hengduan Mountains 1 2019
 ,3,*, 段元文
,3,*, 段元文 ,
,
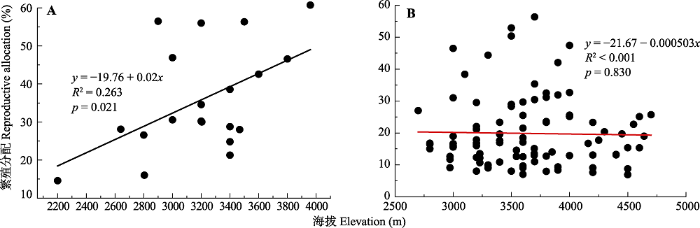 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT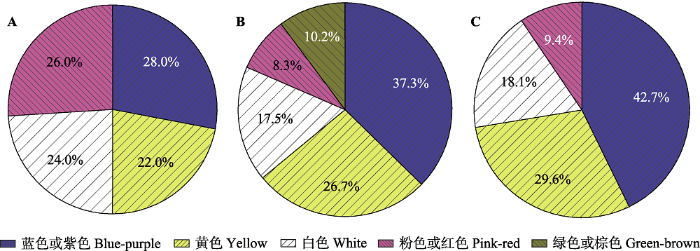 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}