Effects of Caragana microphylla encroachment on community structure and ecosystem function of a typical steppe
Wei DING1, Yu-Bing WANG2,3, Guan-Hai XIANG2,3, Yong-Gang CHI4, Shun-Bao LU,1,*, Shu-Xia ZHENG,2,*1College of Life Sciences, Jiangxi Normal University, Nanchang 330022, China 2State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 3University of Chinese Academy of Sciences, Beijing 100049, China 4College of Geography and Environmental Sciences, Zhejiang Normal University, Jinhua, Zhejiang 321004, China
National Key R&D Program of China(2016YFC0500801) National Natural Science Foundation of China(41671046) National Natural Science Foundation of China(31400393)
Abstract Aims Shrub encroachment is a critical ecological problem in arid and semi-arid ecosystems worldwide. The effects of shrub encroachment on ecosystem structure and function of grasslands are complicated and need to be explored in future studies. Our objective is to examine the effects and pathways of shrub encroachment on ecosystem structure and function in a typical steppe of the Inner Mongolia grassland. Methods Three grassland sites with different degrees of shrub encroachment (i.e. light, moderate, heavy) were selected in the Xilingol Nei Mongol, of which Caragana microphylla was the dominant shrub. Species richness and composition, aboveground net primary productivity (ANPP), soil property, and plant functional traits of dominant species were determined in this study. In addition, species diversity, functional attribute diversity, community-weighted mean traits, and vegetation leaf and soil carbon and nitrogen pools were further calculated. Important findings 1) The species richness, functional attribute diversity and community-weighted mean traits differed significantly among three grassland sites, and species diversity and functional diversity were relatively higher in the moderate shrub-encroachment site, indicating moderate shrub-encroachment favors biodiversity maintenance. 2) The aboveground net primary productivity of heavy shrub-encroachment grassland was significantly higher than those of light and moderate shrub-encroachment grasslands, which was mainly due to a shift in functional group composition, that is, the proportion of annuals and biennials to perennial grasses and forbs increased greatly with intensifying shrub encroachment. The vegetation leaf and soil carbon and nitrogen pools differed little among three sites. 3) Shrub encroachment did not directly affect ecosystem function, including ANPP, vegetation and soil nutrient pools, but it indirectly affected them through pathways of the shift in functional group composition and changes in soil property and functional diversity. Particularly, the shift in functional group composition and intensified soil drought and basification was separately important biotic and abiotic factors for variations in ecosystem function. Keywords:shrub encroachment;species diversity;functional diversity;plant functional trait;functional group composition;ecosystem function
PDF (1497KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 丁威, 王玉冰, 向官海, 迟永刚, 鲁顺保, 郑淑霞. 小叶锦鸡儿灌丛化对典型草原群落结构与生态系统功能的影响. 植物生态学报, 2020, 44(1): 33-43. DOI: 10.17521/cjpe.2019.0283 DING Wei, WANG Yu-Bing, XIANG Guan-Hai, CHI Yong-Gang, LU Shun-Bao, ZHENG Shu-Xia. Effects of Caragana microphylla encroachment on community structure and ecosystem function of a typical steppe. Chinese Journal of Plant Ecology, 2020, 44(1): 33-43. DOI: 10.17521/cjpe.2019.0283
Fig. 1Three grassland sites with light (A), moderate (B) and heavy (C) shrub encroachment by Caragana microphylla in Nei Mongol typical steppe.
Table 1 表1 表1内蒙古典型草原不同程度灌丛化草地的小叶锦鸡儿灌丛特征 Table 1Shrub characteristics of Caragana microphylla at light, moderate and heavy encroachment sites in Nei Mongol typical steppe
灌丛特征 Shrub characteristic
不同程度灌丛化 Degree of shrub encroachment
轻度 Light
中度 Moderate
重度 Heavy
数量 Number of bunches (No.·25 m-2)
2 ± 0.5b (45)
6 ± 0.6a (94)
5 ± 0.9a (80)
高度 Height (cm)
24.13 ± 1.45b (45)
25.71 ± 0.91b (94)
38.90 ± 2.23a (80)
冠幅 Crown (cm2)
8 505.6 ± 1 453.0ab (45)
6 907.3 ± 1 048.8b (94)
13 083.9 ± 2 289.0a (80)
冠幅面积比 Crown area ratio (%)
7.66 ± 1.81b (20)
12.99 ± 2.47ab (20)
20.93 ± 4.37a (20)
分蘖株数 Number of individuals (No.·25 m-2)
25 ± 6b (504)
32 ± 5ab (719)
47 ± 8a (942)
Shrub crown area ratio (%) is the proportion of crown area of C. microphylla to surveying quadrat of 5 m × 5 m. Data are mean ± SE. Number in parentheses is sampling size, and different lowercase letters in the same row indicate significant differences among sites (p < 0.05). 冠幅面积比为小叶锦鸡儿的冠幅面积占调查样方面积(5 m × 5 m)的比值(%), 表中数值为平均值±标准误差, 括号内的数值为样本数, 同一行不同小写字母表示各样地间差异显著(p < 0.05)。
Fig. 2Species richness and functional attribute diversity (FAD) at light, moderate and heavy encroachment sites in Nei Mongol typical steppe (mean + SE). FAD was calculated by plant height, plant biomass, stem:leaf biomass ratio and specific leaf area. Different lowercase letters indicate significant differences among sites (p < 0.05).
Fig. 3Community-weighted mean traits (CWM) at light, moderate and heavy encroachment sites in Nei Mongol typical steppe (mean + SE). PBCWM, community-weighted plant biomass; PHCWM, community-weighted plant height; SLACWM, community-weighted specific leaf area; SLRCWM, community-weighted stem:leaf ratio. Different lowercase letters indicate significant differences among sites (p < 0.05).
Fig. 4Aboveground net primary productivity (ANPP) and relative biomass (RAB) of different functional groups at light, moderate and heavy encroachment sites in Nei Mongol typical steppe (mean + SE). AB, annuals and biennials; PF, perennial forbs; PG, perennial graminoids. Different lowercase letters indicate significant differences among sites (p < 0.05).
Fig. 5Effects of shrub encroachment on C and N pools of vegetational leaf and soil in Nei Mongol typical steppe (mean + SE). No significant differences among sites (p > 0.05).
2.3 灌丛化对ANPP、植被和土壤养分库的影响程度与途径
SEM分析结果: c2 = 11.32, p = 0.66, RMSEA < 0.001, df = 14, n = 60。模型中功能群变化由AB与PG的生物量比值反映; 功能多样性由FAD反映。由于灌丛化对土壤水分和pH的影响方向不同, 随着灌丛化程度增加, 土壤含水量减少(r = -0.270, p < 0.05)而pH (r = 0.263, p < 0.05)升高, 即灌丛化导致土壤旱碱化加剧。植被养分库由叶片C和N库反映, 土壤养分库由土壤C和N库(0-20 cm)反映。实线和虚线分别表示变量之间作用关系显著(p < 0.05)和不显著(p > 0.05)。由SEM分析结果可知, 灌丛化对草原生态系统功能包括ANPP、植被和土壤养分库均没有直接的影响(p > 0.05)(图6)。灌丛化增加了草本群落物种丰富度(β = 0.36, p < 0.01)和功能多样性(β = 0.20, p < 0.05), 但降低了土壤含水量(r = -0.270, p < 0.05), 使土壤pH升高(r = 0.263, p < 0.05), 即灌丛化导致土壤旱碱化加剧(β = -0.32, p < 0.05)。另外, 灌丛化显著增加了一/二年生草本与多年生草本的生物量比值(AB/PG)(β = 0.32, p < 0.01), 即导致功能群组成发生变化, 促使一/二年生草本植物显著增加。灌丛化主要是通过影响物种丰富度进而间接影响功能多样性(β = 0.63, p < 0.001), 其直接效应和间接效应可解释功能多样性变化的53%。灌丛化对功能群组成的影响, 除了直接效应外, 还可通过土壤属性而间接影响(β = 0.43, p < 0.001), 二者共同解释功能群组成变化的37%。草本群落ANPP主要受功能群组成(β = 0.43, p < 0.001)、功能多样性(β = 0.30, p < 0.001)和土壤旱碱化(β = -0.26, p < 0.05)的影响, 三者能够解释ANPP变化的53%; ANPP提高主要是因为灌丛化导致一/二年生草本植物增加, 其次功能多样性也增加。植被养分库主要受ANPP (β = 0.85, p < 0.001)、功能群组成(β = -0.61, p < 0.001)和土壤旱碱化(β = -0.29, p < 0.05)的影响, 三者共同解释60%; 土壤养分库主要受功能群组成(β = 0.37, p < 0.05)和土壤旱碱化(β = 0.35, p < 0.05)的影响, 二者共同解释12%。因此, 灌丛化主要通过影响功能群组成、土壤理化性质和功能多样性, 间接地影响草原生态系统功能, 灌丛化导致功能群发生替代和土壤旱碱化是最重要的生物和非生物因素。
Fig. 6Structural equation model (SEM) analyses of direct and indirect effects of shrub encroachment on grassland ecosystem function, including aboveground net primary productivity (ANPP), vegetation and soil nutrient pools (C and N). Values associated with solid arrows are standardized path coefficients, indicating positive or negative effects. r2 values associated with response variables indicate the proportion of variation explained by relationships with other variables.
ArcherSR, SchimelDS, HollandEA (1995). Mechanisms of shrubland expansion: Land use, climate or CO2? Climatic Change, 29, 91-99. [本文引用: 2]
BargerNN, ArcherSR, CampbellJL, HuangCY, MortonJA, KnappAK (2011). Woody plant proliferation in North American drylands: A synthesis of impacts on ecosystem carbon balance Journal of Geophysical Research, 116, G00K07. DOI: 10.1029/2010JG001506. [本文引用: 3]
ChenLY, LiH, ZhangPJ, ZhaoX, ZhouLH, LiuTY, HuHF, BaiYF, ShenHH, FangJY (2015). Climate and native grassland vegetation as drivers of the community structures of shrub-encroached grasslands in Inner Mongolia, China Landscape Ecology, 30, 1627-1641. [本文引用: 2]
Darrouzet-NardiA, D’AntonioCM, DawsonTE (2006). Depth of water acquisition by invading shrubs and resident herbs in a Sierra Nevada meadow Plant and Soil, 285, 31-43. DOI:10.1007/s11104-005-4453-zURL [本文引用: 1]
EldridgeDJ, BowkerMA, MaestreFT, RogerE, ReynoldsJF, WhitfordWG (2011). Impacts of shrub encroachment on ecosystem structure and functioning: Towards a global synthesis Ecology Letters, 14, 709-722. DOI:10.1111/ele.2011.14.issue-7URL [本文引用: 1]
EldridgeDJ, SoliveresS, BowkerMA, ValJ (2013). Grazing dampens the positive effects of shrub encroachment on ecosystem functions in a semi-arid woodland Journal of Applied Ecology, 50, 1028-1038. [本文引用: 1]
GaoQ, LiuT (2015). Causes and consequences of shrub encroachment in arid and semiarid region: A disputable issue Arid Land Geography, 38, 1202-1212. [本文引用: 1]
GrossN, SudingKN, LavorelS, RoumetC (2007). Complementarity as a mechanism of coexistence between functional groups of grasses Journal of Ecology, 95, 1296-1305. [本文引用: 1]
GroverHD, MusickHB (1990). Shrubland encroachment in southern New Mexico, USA: An analysis of desertification processes in the American Southwest Climatic Change, 17, 305-330. [本文引用: 1]
HolzapfelC, MahallBE (1999). Bidirectional facilitation and interference between shrubs and annuals in the Mojave Desert Ecology, 80, 1747-1761. [本文引用: 1]
JacksonRB, BannerJL, JobbágyEG, PockmanWT, WallDH (2002). Ecosystem carbon loss with woody plant invasion of grasslands Nature, 418, 623-626. [本文引用: 1]
KnappAK, BriggsJM, CollinsSL, ArcherSR, Bret-HarteMS, EwersBE, PetersDP, YoungDR, ShaverGR, PendallE, ClearyMB (2008). Shrub encroachment in North American grasslands: Shifts in growth form dominance rapidly alters control of ecosystem carbon inputs Global Change Biology, 14, 615-623. [本文引用: 3]
LanZC, BaiYF (2012). Testing mechanisms of N-enrichment- induced species loss in a semiarid Inner Mongolia grassland: Critical thresholds and implications for long-term ecosystem responses Philosophical Transactions of the Royal Society B: Biological Sciences, 367, 3125-3134. [本文引用: 1]
LiXY, ZhangSY, PengHY, HuX, MaYJ (2013). Soil water and temperature dynamics in shrub-encroached grasslands and climatic implications: Results from Inner Mongolia steppe ecosystem of north China Agricultural and Forest Meteorology, 171-172, 20-30. DOI:10.1016/j.agrformet.2012.11.001URL [本文引用: 1]
Millennium Ecosystem Assessment (2005). Ecosystems and Human Well-being: Desertification Synthesis. World Resources Institute, Washington. [本文引用: 1]
MorganJA, MilchunasDG, LeCainDR, WestM, MosierAR (2007). Carbon dioxide enrichment alters plant community structure and accelerates shrub growth in the shortgrass steppe Proceedings of the National Academy of Sciences of the United States of America, 104, 14724-14729. [本文引用: 1]
ParizekB, RostagnoCM, SottiniR (2002). Soil erosion as affected by shrub encroachment in northeastern Patagonia Journal of Range Management, 55, 43-48. [本文引用: 2]
PengHY, LiXY, LiGY, ZhangZH, ZhangSY, LiL, ZhaoGQ, JiangZY, MaYJ (2013). Shrub encroachment with increasing anthropogenic disturbance in the semiarid Inner Mongolian grasslands of China Catena, 109, 39-48. [本文引用: 1]
PugnaireFI, ArmasC, MaestreFT (2011). Positive plant interactions in the Iberian Southeast: Mechanisms, environmental gradients, and ecosystem function Journal of Arid Environments, 75, 1310-1320. [本文引用: 2]
RatajczakZ, D?OdoricoP, NippertJB, CollinsSL, BrunsellNA, RaviS (2017). Changes in spatial variance during a grassland to shrubland state transition Journal of Ecology, 105, 750-760. [本文引用: 1]
RoscherC, WeigeltA, ProulxR, MarquardE, SchumacherJ, WeisserWW, SchmidB (2011). Identifying population- and community-level mechanisms of diversity-stability relationships in experimental grasslands Journal of Ecology, 99, 1460-1469. [本文引用: 1]
SeifanM, KadmonR (2006). Indirect effects of cattle grazing on shrub spatial pattern in a mediterranean scrub community Basic and Applied Ecology, 7, 496-506. DOI:10.1016/j.baae.2005.10.004URL [本文引用: 1]
ShackletonCM, ScholesRJ (2011). Above ground woody community attributes, biomass and carbon stocks along a rainfall gradient in the savannas of the central lowveld, South Africa South African Journal of Botany, 77, 184-192. DOI:10.1016/j.sajb.2010.07.014URL [本文引用: 1]
SoliveresS, EldridgeDJ (2014). Do changes in grazing pressure and the degree of shrub encroachment alter the effects of individual shrubs on understorey plant communities and soil function? Functional Ecology, 28, 530-537. [本文引用: 2]
ThompsonWA, EldridgeDJ (2005). Plant cover and composition in relation to density of Callitris glaucophylla(white cypress pine) along a rainfall gradient in eastern Australia Australian Journal of Botany, 53, 545. [本文引用: 1]
ThroopHL, ArcherSR, MongerHC, WaltmanS (2012). When bulk density methods matter: Implications for estimating soil organic carbon pools in rocky soils Journal of Arid Environments, 77, 66-71. [本文引用: 1]
van AukenOW (2009). Causes and consequences of woody plant encroachment into western North American grasslands Journal of Environmental Management, 90, 2931-2942. [本文引用: 2]
WalkerB, KinzigA, LangridgeJ (1999). Plant attribute diversity, resilience, and ecosystem function: The nature and significance of dominant and minor species Ecosystems, 2, 95-113. [本文引用: 1]
WanHW, PanQM, BaiYF (2013). China grassland biodiversity monitoring network: Indicators and implementation plan Biodiversity Science, 21, 639-650. [本文引用: 1]
XiongXG, HanXG (2006). Application of state and transition models to discussing the thicketization of steppe in Xilin River Basin, Inner Mongolia Acta Prataculturae Sinica, 15(2), 9-13. [本文引用: 1]
ZarovaliMP, YiakoulakiMD, PapanastasisVP (2007). Effects of shrub encroachment on herbage production and nutritive value in semi-arid Mediterranean grasslands Grass and Forage Science, 62, 355-363. DOI:10.1111/gfs.2007.62.issue-3URL [本文引用: 1]
ZhengSX, RenHY, LiWH, LanZC (2012). Scale-dependent effects of grazing on plant C:N:P stoichiometry and linkages to ecosystem functioning in the Inner Mongolia grassland PLOS ONE, 7, e51750. DOI: 10.1371/journal.pone.0051750. [本文引用: 1]
ZhouLH, ShenHH, ChenLY, LiH, ZhangPJ, ZhaoX, LiuTY, LiuSS, XingAJ, HuHF, FangJY (2019). Ecological consequences of shrub encroachment in the grasslands of northern China Landscape Ecology, 34, 119-130. [本文引用: 2]
Woody plant encroachment: Causes and consequences 2 2017
... 本研究结果显示, 不同程度灌丛化对草本群落植被和土壤养分库的影响不显著, 总体上植被叶片C、N库变化略大于土壤C、N库.有机C库反映了植物叶片、茎和根系的长期投入, 且有机C的含量会随着植物生产力的变化而变化, 生产力的变化代表着生态系统C吸收的变化, 是木本植物入侵改变生态系统C平衡的重要机制(Archer et al., 2017).已有研究表明灌木入侵对ANPP的影响与年降水量有关, 在半干旱区和半湿润区(年降水量> 336 mm), ANPP随灌木的入侵增加, 而在干旱地区(年降水量< 336 mm), ANPP则随灌木入侵而减少(Barger et al., 2011).本研究中, 随着灌丛化程度增加, 尽管ANPP增加, 然而植被C、N库却无明显变化, 其原因是功能群组成变化和土壤旱碱化对植被养分库的负效应(β值分别为-0.61和-0.29)中和了ANPP的正效应(β = 0.85).土壤容重和黏土含量是影响土壤有机C库的重要因素(Barger et al., 2011), 本研究中各样点的土壤容重差异不显著, 是导致土壤养分库变化不大的主要原因.由于土壤变化远远滞后于驱动它们的植被变化, 因而植被养分库变化略大于土壤养分库.由于灌丛化的成因较为复杂, 各驱动因子相互作用, 不同生态区域差异性较大, 灌木入侵对生态系统C库的影响可能是积极、消极或是中性的(Jackson et al., 2002; Knapp et al., 2008).造成地上和地下养分库响应不一致的原因, 可能有以下几点: (1)土壤质地、盐性以及地形坡度的不同使得地下养分库对地上变化的响应具有局部性(Barger et al., 2011; Throop et al., 2012); (2)灌丛景观的异质性、斑块大小和灌草性状的差异等导致灌丛化效应的不同, 以及土壤变化远远滞后于驱动它们的植被变化; (3)植物物种或功能群对地上、地下部分C分配的差异(Archer et al., 2017).因此, 灌丛化过程中地上、地下养分库如何变化有待于长时间尺度的研究. ...
... ); (2)灌丛景观的异质性、斑块大小和灌草性状的差异等导致灌丛化效应的不同, 以及土壤变化远远滞后于驱动它们的植被变化; (3)植物物种或功能群对地上、地下部分C分配的差异(Archer et al., 2017).因此, 灌丛化过程中地上、地下养分库如何变化有待于长时间尺度的研究. ...
Mechanisms of shrubland expansion: Land use, climate or CO2? 2 1995
... 灌丛化, 即草原生态系统中原生灌木/木本植物的植株密度、盖度和生物量增加的现象(Archer et al., 1995), 是世界范围内干旱半干旱地区发生的一种重要的土地覆盖变化(Knapp et al., 2008; Maestre et al., 2009; Li et al., 2013).在过去150多年里, 全球草原区域有10%-20%地区发生了灌丛化, 使得超过20亿的人口受其影响, 严重威胁了草原地区的经济生产和生态安全(Parizek et al., 2002; van Auken, 2009).导致草原灌丛化的原因有很多, 包括: 气候变化如大气CO2浓度上升和季节性干旱、火烧程度和过度放牧等(Neilson, 1986; Archer et al., 1995; Huxman et al., 2005; Seifan & Kadmon, 2006; Morgan et al., 2007; Shackleton & Scholes, 2011).我国灌丛化草原具有分布广、面积大的特点, 对我国北方区域尺度上灌丛化实地调查发现, 降水是影响灌丛盖度和斑块大小的主要因素, 而温度是影响灌丛株高和斑块密度的首要因素; 干旱高温区域灌丛盖度低, 灌丛斑块小而多, 而湿润低温区域灌丛盖度高, 灌丛斑块大而少(Chen et al., 2015).在大的区域尺度上, 灌丛化通过降低草本的丰富度、多度和地上生物量从而使景观异质性降低, 然而土壤碳(C)、氮(N)含量也降低(Zhou et al., 2019).在内蒙古草原, 约有5.1 × 106 hm2草原出现了灌丛化现象, 以小叶锦鸡儿(Caragana microphylla)灌丛化最为典型(熊小刚和韩兴国, 2006). ...
... ; Archer et al., 1995; Huxman et al., 2005; Seifan & Kadmon, 2006; Morgan et al., 2007; Shackleton & Scholes, 2011).我国灌丛化草原具有分布广、面积大的特点, 对我国北方区域尺度上灌丛化实地调查发现, 降水是影响灌丛盖度和斑块大小的主要因素, 而温度是影响灌丛株高和斑块密度的首要因素; 干旱高温区域灌丛盖度低, 灌丛斑块小而多, 而湿润低温区域灌丛盖度高, 灌丛斑块大而少(Chen et al., 2015).在大的区域尺度上, 灌丛化通过降低草本的丰富度、多度和地上生物量从而使景观异质性降低, 然而土壤碳(C)、氮(N)含量也降低(Zhou et al., 2019).在内蒙古草原, 约有5.1 × 106 hm2草原出现了灌丛化现象, 以小叶锦鸡儿(Caragana microphylla)灌丛化最为典型(熊小刚和韩兴国, 2006). ...
Woody plant proliferation in North American drylands: A synthesis of impacts on ecosystem carbon balance 3 2011
... 本研究结果显示, 不同程度灌丛化对草本群落植被和土壤养分库的影响不显著, 总体上植被叶片C、N库变化略大于土壤C、N库.有机C库反映了植物叶片、茎和根系的长期投入, 且有机C的含量会随着植物生产力的变化而变化, 生产力的变化代表着生态系统C吸收的变化, 是木本植物入侵改变生态系统C平衡的重要机制(Archer et al., 2017).已有研究表明灌木入侵对ANPP的影响与年降水量有关, 在半干旱区和半湿润区(年降水量> 336 mm), ANPP随灌木的入侵增加, 而在干旱地区(年降水量< 336 mm), ANPP则随灌木入侵而减少(Barger et al., 2011).本研究中, 随着灌丛化程度增加, 尽管ANPP增加, 然而植被C、N库却无明显变化, 其原因是功能群组成变化和土壤旱碱化对植被养分库的负效应(β值分别为-0.61和-0.29)中和了ANPP的正效应(β = 0.85).土壤容重和黏土含量是影响土壤有机C库的重要因素(Barger et al., 2011), 本研究中各样点的土壤容重差异不显著, 是导致土壤养分库变化不大的主要原因.由于土壤变化远远滞后于驱动它们的植被变化, 因而植被养分库变化略大于土壤养分库.由于灌丛化的成因较为复杂, 各驱动因子相互作用, 不同生态区域差异性较大, 灌木入侵对生态系统C库的影响可能是积极、消极或是中性的(Jackson et al., 2002; Knapp et al., 2008).造成地上和地下养分库响应不一致的原因, 可能有以下几点: (1)土壤质地、盐性以及地形坡度的不同使得地下养分库对地上变化的响应具有局部性(Barger et al., 2011; Throop et al., 2012); (2)灌丛景观的异质性、斑块大小和灌草性状的差异等导致灌丛化效应的不同, 以及土壤变化远远滞后于驱动它们的植被变化; (3)植物物种或功能群对地上、地下部分C分配的差异(Archer et al., 2017).因此, 灌丛化过程中地上、地下养分库如何变化有待于长时间尺度的研究. ...
... = 0.85).土壤容重和黏土含量是影响土壤有机C库的重要因素(Barger et al., 2011), 本研究中各样点的土壤容重差异不显著, 是导致土壤养分库变化不大的主要原因.由于土壤变化远远滞后于驱动它们的植被变化, 因而植被养分库变化略大于土壤养分库.由于灌丛化的成因较为复杂, 各驱动因子相互作用, 不同生态区域差异性较大, 灌木入侵对生态系统C库的影响可能是积极、消极或是中性的(Jackson et al., 2002; Knapp et al., 2008).造成地上和地下养分库响应不一致的原因, 可能有以下几点: (1)土壤质地、盐性以及地形坡度的不同使得地下养分库对地上变化的响应具有局部性(Barger et al., 2011; Throop et al., 2012); (2)灌丛景观的异质性、斑块大小和灌草性状的差异等导致灌丛化效应的不同, 以及土壤变化远远滞后于驱动它们的植被变化; (3)植物物种或功能群对地上、地下部分C分配的差异(Archer et al., 2017).因此, 灌丛化过程中地上、地下养分库如何变化有待于长时间尺度的研究. ...
... ).造成地上和地下养分库响应不一致的原因, 可能有以下几点: (1)土壤质地、盐性以及地形坡度的不同使得地下养分库对地上变化的响应具有局部性(Barger et al., 2011; Throop et al., 2012); (2)灌丛景观的异质性、斑块大小和灌草性状的差异等导致灌丛化效应的不同, 以及土壤变化远远滞后于驱动它们的植被变化; (3)植物物种或功能群对地上、地下部分C分配的差异(Archer et al., 2017).因此, 灌丛化过程中地上、地下养分库如何变化有待于长时间尺度的研究. ...
Climate and native grassland vegetation as drivers of the community structures of shrub-encroached grasslands in Inner Mongolia, China 2 2015
... 灌丛化, 即草原生态系统中原生灌木/木本植物的植株密度、盖度和生物量增加的现象(Archer et al., 1995), 是世界范围内干旱半干旱地区发生的一种重要的土地覆盖变化(Knapp et al., 2008; Maestre et al., 2009; Li et al., 2013).在过去150多年里, 全球草原区域有10%-20%地区发生了灌丛化, 使得超过20亿的人口受其影响, 严重威胁了草原地区的经济生产和生态安全(Parizek et al., 2002; van Auken, 2009).导致草原灌丛化的原因有很多, 包括: 气候变化如大气CO2浓度上升和季节性干旱、火烧程度和过度放牧等(Neilson, 1986; Archer et al., 1995; Huxman et al., 2005; Seifan & Kadmon, 2006; Morgan et al., 2007; Shackleton & Scholes, 2011).我国灌丛化草原具有分布广、面积大的特点, 对我国北方区域尺度上灌丛化实地调查发现, 降水是影响灌丛盖度和斑块大小的主要因素, 而温度是影响灌丛株高和斑块密度的首要因素; 干旱高温区域灌丛盖度低, 灌丛斑块小而多, 而湿润低温区域灌丛盖度高, 灌丛斑块大而少(Chen et al., 2015).在大的区域尺度上, 灌丛化通过降低草本的丰富度、多度和地上生物量从而使景观异质性降低, 然而土壤碳(C)、氮(N)含量也降低(Zhou et al., 2019).在内蒙古草原, 约有5.1 × 106 hm2草原出现了灌丛化现象, 以小叶锦鸡儿(Caragana microphylla)灌丛化最为典型(熊小刚和韩兴国, 2006). ...
... 本研究结果显示, 不同程度灌丛化对草本群落植被和土壤养分库的影响不显著, 总体上植被叶片C、N库变化略大于土壤C、N库.有机C库反映了植物叶片、茎和根系的长期投入, 且有机C的含量会随着植物生产力的变化而变化, 生产力的变化代表着生态系统C吸收的变化, 是木本植物入侵改变生态系统C平衡的重要机制(Archer et al., 2017).已有研究表明灌木入侵对ANPP的影响与年降水量有关, 在半干旱区和半湿润区(年降水量> 336 mm), ANPP随灌木的入侵增加, 而在干旱地区(年降水量< 336 mm), ANPP则随灌木入侵而减少(Barger et al., 2011).本研究中, 随着灌丛化程度增加, 尽管ANPP增加, 然而植被C、N库却无明显变化, 其原因是功能群组成变化和土壤旱碱化对植被养分库的负效应(β值分别为-0.61和-0.29)中和了ANPP的正效应(β = 0.85).土壤容重和黏土含量是影响土壤有机C库的重要因素(Barger et al., 2011), 本研究中各样点的土壤容重差异不显著, 是导致土壤养分库变化不大的主要原因.由于土壤变化远远滞后于驱动它们的植被变化, 因而植被养分库变化略大于土壤养分库.由于灌丛化的成因较为复杂, 各驱动因子相互作用, 不同生态区域差异性较大, 灌木入侵对生态系统C库的影响可能是积极、消极或是中性的(Jackson et al., 2002; Knapp et al., 2008).造成地上和地下养分库响应不一致的原因, 可能有以下几点: (1)土壤质地、盐性以及地形坡度的不同使得地下养分库对地上变化的响应具有局部性(Barger et al., 2011; Throop et al., 2012); (2)灌丛景观的异质性、斑块大小和灌草性状的差异等导致灌丛化效应的不同, 以及土壤变化远远滞后于驱动它们的植被变化; (3)植物物种或功能群对地上、地下部分C分配的差异(Archer et al., 2017).因此, 灌丛化过程中地上、地下养分库如何变化有待于长时间尺度的研究. ...
Testing mechanisms of N-enrichment- induced species loss in a semiarid Inner Mongolia grassland: Critical thresholds and implications for long-term ecosystem responses 1 2012
Assessing functional diversity in the field— Methodology matters! 1 2008
... 统计分析采用SPSS 20.0软件, 通过单因素方差分析和Duncan’s多重比较方法, 分析不同灌丛化样地小叶锦鸡儿灌丛特征、草本群落物种丰富度、功能多样性、初级生产力、植被叶片和土壤C、N库的差异性.采用R软件FDiversity程序包计算功能多样性, 功能多样性指标用功能性状多样性指数(FAD)(Walker et al., 1999)和群落加权性状平均值(CWM)(Lavorel et al., 2008)表示.FAD和CWM均是利用样方内4个优势物种的功能性状(PH、PB、SLR和SLA)和相对生物量进行计算.具体计算公式如下: ...
Soil water and temperature dynamics in shrub-encroached grasslands and climatic implications: Results from Inner Mongolia steppe ecosystem of north China 1 2013
... 灌丛化, 即草原生态系统中原生灌木/木本植物的植株密度、盖度和生物量增加的现象(Archer et al., 1995), 是世界范围内干旱半干旱地区发生的一种重要的土地覆盖变化(Knapp et al., 2008; Maestre et al., 2009; Li et al., 2013).在过去150多年里, 全球草原区域有10%-20%地区发生了灌丛化, 使得超过20亿的人口受其影响, 严重威胁了草原地区的经济生产和生态安全(Parizek et al., 2002; van Auken, 2009).导致草原灌丛化的原因有很多, 包括: 气候变化如大气CO2浓度上升和季节性干旱、火烧程度和过度放牧等(Neilson, 1986; Archer et al., 1995; Huxman et al., 2005; Seifan & Kadmon, 2006; Morgan et al., 2007; Shackleton & Scholes, 2011).我国灌丛化草原具有分布广、面积大的特点, 对我国北方区域尺度上灌丛化实地调查发现, 降水是影响灌丛盖度和斑块大小的主要因素, 而温度是影响灌丛株高和斑块密度的首要因素; 干旱高温区域灌丛盖度低, 灌丛斑块小而多, 而湿润低温区域灌丛盖度高, 灌丛斑块大而少(Chen et al., 2015).在大的区域尺度上, 灌丛化通过降低草本的丰富度、多度和地上生物量从而使景观异质性降低, 然而土壤碳(C)、氮(N)含量也降低(Zhou et al., 2019).在内蒙古草原, 约有5.1 × 106 hm2草原出现了灌丛化现象, 以小叶锦鸡儿(Caragana microphylla)灌丛化最为典型(熊小刚和韩兴国, 2006). ...
Shrub encroachment can reverse desertification in semi-arid Mediterranean grasslands 5 2009
... 灌丛化, 即草原生态系统中原生灌木/木本植物的植株密度、盖度和生物量增加的现象(Archer et al., 1995), 是世界范围内干旱半干旱地区发生的一种重要的土地覆盖变化(Knapp et al., 2008; Maestre et al., 2009; Li et al., 2013).在过去150多年里, 全球草原区域有10%-20%地区发生了灌丛化, 使得超过20亿的人口受其影响, 严重威胁了草原地区的经济生产和生态安全(Parizek et al., 2002; van Auken, 2009).导致草原灌丛化的原因有很多, 包括: 气候变化如大气CO2浓度上升和季节性干旱、火烧程度和过度放牧等(Neilson, 1986; Archer et al., 1995; Huxman et al., 2005; Seifan & Kadmon, 2006; Morgan et al., 2007; Shackleton & Scholes, 2011).我国灌丛化草原具有分布广、面积大的特点, 对我国北方区域尺度上灌丛化实地调查发现, 降水是影响灌丛盖度和斑块大小的主要因素, 而温度是影响灌丛株高和斑块密度的首要因素; 干旱高温区域灌丛盖度低, 灌丛斑块小而多, 而湿润低温区域灌丛盖度高, 灌丛斑块大而少(Chen et al., 2015).在大的区域尺度上, 灌丛化通过降低草本的丰富度、多度和地上生物量从而使景观异质性降低, 然而土壤碳(C)、氮(N)含量也降低(Zhou et al., 2019).在内蒙古草原, 约有5.1 × 106 hm2草原出现了灌丛化现象, 以小叶锦鸡儿(Caragana microphylla)灌丛化最为典型(熊小刚和韩兴国, 2006). ...
Indirect effects of cattle grazing on shrub spatial pattern in a mediterranean scrub community 1 2006
... 灌丛化, 即草原生态系统中原生灌木/木本植物的植株密度、盖度和生物量增加的现象(Archer et al., 1995), 是世界范围内干旱半干旱地区发生的一种重要的土地覆盖变化(Knapp et al., 2008; Maestre et al., 2009; Li et al., 2013).在过去150多年里, 全球草原区域有10%-20%地区发生了灌丛化, 使得超过20亿的人口受其影响, 严重威胁了草原地区的经济生产和生态安全(Parizek et al., 2002; van Auken, 2009).导致草原灌丛化的原因有很多, 包括: 气候变化如大气CO2浓度上升和季节性干旱、火烧程度和过度放牧等(Neilson, 1986; Archer et al., 1995; Huxman et al., 2005; Seifan & Kadmon, 2006; Morgan et al., 2007; Shackleton & Scholes, 2011).我国灌丛化草原具有分布广、面积大的特点, 对我国北方区域尺度上灌丛化实地调查发现, 降水是影响灌丛盖度和斑块大小的主要因素, 而温度是影响灌丛株高和斑块密度的首要因素; 干旱高温区域灌丛盖度低, 灌丛斑块小而多, 而湿润低温区域灌丛盖度高, 灌丛斑块大而少(Chen et al., 2015).在大的区域尺度上, 灌丛化通过降低草本的丰富度、多度和地上生物量从而使景观异质性降低, 然而土壤碳(C)、氮(N)含量也降低(Zhou et al., 2019).在内蒙古草原, 约有5.1 × 106 hm2草原出现了灌丛化现象, 以小叶锦鸡儿(Caragana microphylla)灌丛化最为典型(熊小刚和韩兴国, 2006). ...
Above ground woody community attributes, biomass and carbon stocks along a rainfall gradient in the savannas of the central lowveld, South Africa 1 2011
... 灌丛化, 即草原生态系统中原生灌木/木本植物的植株密度、盖度和生物量增加的现象(Archer et al., 1995), 是世界范围内干旱半干旱地区发生的一种重要的土地覆盖变化(Knapp et al., 2008; Maestre et al., 2009; Li et al., 2013).在过去150多年里, 全球草原区域有10%-20%地区发生了灌丛化, 使得超过20亿的人口受其影响, 严重威胁了草原地区的经济生产和生态安全(Parizek et al., 2002; van Auken, 2009).导致草原灌丛化的原因有很多, 包括: 气候变化如大气CO2浓度上升和季节性干旱、火烧程度和过度放牧等(Neilson, 1986; Archer et al., 1995; Huxman et al., 2005; Seifan & Kadmon, 2006; Morgan et al., 2007; Shackleton & Scholes, 2011).我国灌丛化草原具有分布广、面积大的特点, 对我国北方区域尺度上灌丛化实地调查发现, 降水是影响灌丛盖度和斑块大小的主要因素, 而温度是影响灌丛株高和斑块密度的首要因素; 干旱高温区域灌丛盖度低, 灌丛斑块小而多, 而湿润低温区域灌丛盖度高, 灌丛斑块大而少(Chen et al., 2015).在大的区域尺度上, 灌丛化通过降低草本的丰富度、多度和地上生物量从而使景观异质性降低, 然而土壤碳(C)、氮(N)含量也降低(Zhou et al., 2019).在内蒙古草原, 约有5.1 × 106 hm2草原出现了灌丛化现象, 以小叶锦鸡儿(Caragana microphylla)灌丛化最为典型(熊小刚和韩兴国, 2006). ...
Do changes in grazing pressure and the degree of shrub encroachment alter the effects of individual shrubs on understorey plant communities and soil function? 2 2014
 ,1,*, 郑淑霞
,1,*, 郑淑霞
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT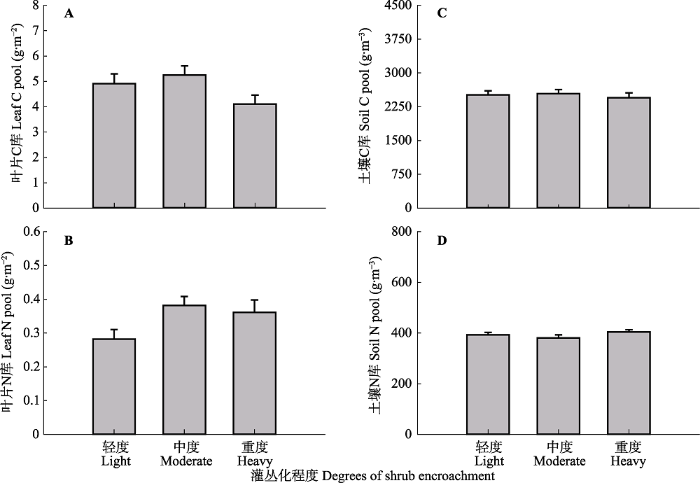 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT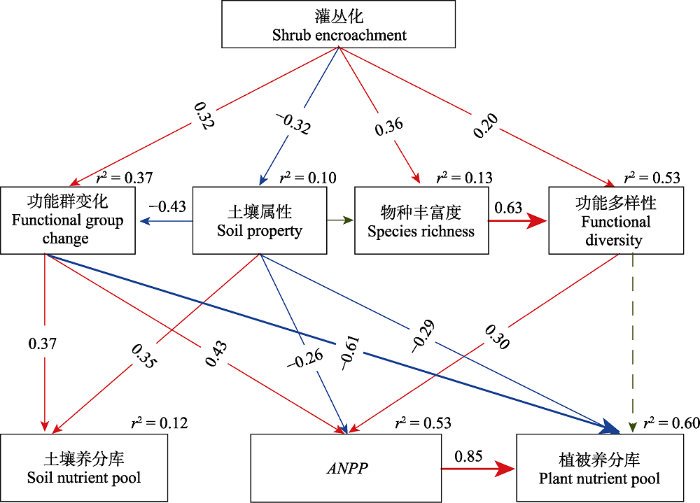 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}