Effects of grazing and simulated warming on plant community structure and productivity of alpine grassland in Northern Xizang, China
Hong-Bao WU1, Qing-Zhu GAO1, Hasbagan Ganjurjav,1,*, Yu LI2, Yu-Long YAN3, Guo-Zheng HU1, Xue-Xia WANG1, Jun YAN4, Shi-Cheng HE41Institute of Environment and Sustainable Development in Agriculture, Chinese Academy of Agricultural Sciences, Beijing 100081, China 2College of Environment, Beijing Normal University, Beijing 100875, China 3School of Life Sciences, Inner Mongolia University, Hohhot 0 10021, China (4) Nagqu Grassland Station, Nagqu, Xizang 852000, China
通讯作者: 通讯作者Author for correspondence (E-mail:jshe@pku.edu.cn)
the National Key R&D Program of China(2016YFC0502003) the National Natural Science Foundation of China(31570484) the National Natural Science Foundation of China( 31600366) the Central Public-interest Scientific Institution Basal Research Fund(BSRF201713)
Abstract Aims Climate change and grazing activities have important effects on species diversity and productivity of grassland ecosystems. The aim of this study is to reveal the differences in responses of species diversity and productivity to warming, grazing and their interactions in an alpine meadow ecosystem. Methods Warming experiment was established in 2011. In 2016, the grazing experiment and the combination of warming and grazing experiment were added. A two-year continuous field experiment was conducted (from 2016 to 2017) and plant community structure, community composition, productivity and species diversity were monitored. Twelve sites were sampled, and were divided into four treatments: control, warming, grazing and the combination of warming and grazing. All sampled species were classified into three functional groups: sedges, grasses and forbs. The species diversity and productivity were sampled in three different treatments. Important findings Results showed that warming and grazing had significant interaction on vegetation height and net primary productivity in alpine grassland. Under grazing treatment, warming had no significant effect on plant height, while warming significantly increased plant height without grazing. Under grazing treatment, the warming effect on net primary productivity significantly differed between 2016 and 2017. In 2016, warming had no significant effect on net primary productivity, while in 2017, warming significantly decreased net primary productivity. Warming and grazing had no significant interaction on species richness, coverage, species important value and species diversity in the alpine meadow. Under the treatments of warming and grazing, the total vegetation coverage decreased, and the proportion of forbs significantly increased. However, no significant effect of warming or grazing was found on species diversity. This study showed that warming and grazing significantly changed the community structure of alpine grassland. Therefore, with the projected climate change in the future, the intensification of grazing activities may lead to the decrease of alpine meadow productivity. Keywords:warming;grazing;the combination of warming and grazing;species diversity;net primary productivity
PDF (1386KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 吴红宝, 高清竹, 干珠扎布, 李钰, 闫玉龙, 胡国铮, 王学霞, 严俊, 何世丞. 放牧和模拟增温对藏北高寒草地植物群落特征及生产力的影响. 植物生态学报, 2019, 43(10): 853-862. DOI: 10.17521/cjpe.2018.0288 WU Hong-Bao, GAO Qing-Zhu, Ganjurjav Hasbagan, LI Yu, YAN Yu-Long, HU Guo-Zheng, WANG Xue-Xia, YAN Jun, HE Shi-Cheng. Effects of grazing and simulated warming on plant community structure and productivity of alpine grassland in Northern Xizang, China. Chinese Journal of Plant Ecology, 2019, 43(10): 853-862. DOI: 10.17521/cjpe.2018.0288
藏北高原位于青藏高原的腹地, 被称为“世界屋脊的屋脊”。该地区拥有42.1万km2高寒草地(高清竹等, 2006), 是西藏自治区主要的畜牧业生产基地和我国重要的生态屏障区, 是全球气候变化的启动区和敏感区, 是亚洲中部高寒环境中最为典型的一种生态系统。据报道, 青藏高原增温幅度高达每10年0.2 ℃ (Chen et al., 2013), 至2030年该地区将升温2.2-2.6 ℃ (National Development and Reform Commisssion, 2007), 显著高于全球和全国平均水平。由于过度放牧和草地不合理利用, 藏北高原草地退化面积达到58.2%, 重度退化和极重度退化草地面积分别占19.0%和6.5% (曹旭娟等, 2016)。近年来, 研究人员分别开展了增温和放牧对高寒草地植物生产力和物种多样性影响的研究(段敏杰等, 2010; Ganjurjav et al., 2016; Liu et al., 2018), 但增温和放牧的交互作用对物种多样性和生产力影响的研究鲜有报道。一些****通过刈割模拟放牧活动, 从而研究增温和放牧的交互作用对高寒草地群落结构及生产力的影响(Wang et al., 2012; Fu et al., 2015)。然而, 上述研究在很大程度上忽略了牲畜的践踏和采食作用对草地植物群落的影响。此外, 放牧与刈割的不同之处还在于对物种组成的影响, 放牧条件下牲畜对草地植物的采食具有物种间的选择喜好, 而刈割仅是对草地植物平均冠层高度进行统一收割(Klein et al., 2008; 段敏杰等, 2010; Wang et al., 2012)。因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据。
Fig. 3Mean air temperature, temperature and moisture of the soil (from April to August in 2016 and 2017) in control (CK), warming (W), grazing (G) and warming + grazing (WG) plots.
Table 1 表1 表1年份、增温、放牧及其交互作用对藏北高寒草地植物高度、盖度、物种重要值、净初级生产力及物种多样性影响的多因素方差分析结果(p值) Table 1Results (p value) of the three way ANOVA of year, warming, grazing and their interaction effects on height, coverage, species important value, net primary productivity and species diversity index of alpine grassland in Northern Xizang
因子 Factor
高度 Height
盖度 Coverage
物种重要值 Species important value
净初级生产力 Net primary productivity
物种多样性指数 Species diversity index
莎草类 Sedges
禾草类 Grasses
杂类草 Forbs
莎草类 Sedges
禾草类 Grasses
杂类草 Forbs
Species richness index
Shannon- Weiner index
Pielou index
Y
0.061
0.032
0.480
0.481
0.052
0.412
0.446
0.074
0.209
0.907
0.078
W
<0.001
<0.001
0.001
<0.001
0.055
0.773
0.174
0.618
1.000
0.543
0.331
G
<0.001
<0.001
<0.001
0.004
0.008
0.445
0.003
0.503
0.396
0.872
0.236
Y × W
0.065
0.055
0.396
0.444
0.716
0.191
0.199
0.158
0.668
0.523
0.511
Y × G
0.380
0.283
0.143
0.092
0.482
0.177
0.290
0.122
0.001
0.019
0.470
W × G
0.003
0.001
0.001
0.376
0.882
0.103
0.066
0.048
1.000
0.617
0.347
Y × W × G
0.903
0.103
0.587
0.926
0.169
0.201
0.708
0.145
0.209
0.332
0.904
G, 放牧; W, 增温; Y, 年份。加粗字体代表p < 0.05。 G, grazing; W, warming; Y, year. The bold numbers indicated p < 0.05.
Fig. 4Mean vegetation height (mean ± SE) in the control (CK), warming (W), grazing (G) and warming + grazing (WG) plots of alpine meadow in Northern Xizang. Different lowercase letters indicate significant differences in height in group plant (p < 0.05).
Table 2 表2 表2对照、增温、放牧及增温+放牧处理下藏北高寒草地群落物种组成及其重要值(2017年) Table 2Species composition and their important values of alpine grassland communities under treatments of control, warming, grazing and the combination of warming and grazing in Northern Xizang in 2017
植物类群 Plant group
物种 Species
CK
W
G
WG
莎草类 Sedges
高山嵩草 Kobresia pygmaea
0.420
0.354
0.475
0.448
青藏薹草 Carex moorcroftii
0.109
0.118
-
-
矮生嵩草 Kobresia humilis
0.028
-
-
-
禾草类 Grasses
紫花针茅 Stipa purpurea
-
-
0.113
0.179
草地早熟禾 Poa pratensis
0.197
0.238
-
-
杂类草 Forbs
垫状点地梅 Androsace tapete
0.003
0.002
-
-
星状雪兔子 Saussurea stella
-
-
0.064
0.033
矮火绒草 Leontopodium nanum
-
-
0.040
-
蓝花棘豆 Oxytropis caerulea
0.032
0.041
0.058
-
藏豆 Stracheya tibetica
-
0.019
-
0.064
蒲公英 Taraxacum mongolicum
-
-
0.014
0.025
肉果草 Lancea tibetica
0.052
0.034
-
0.007
短穗兔耳草 Lagotis brachystachya
0.008
0.030
0.034
0.024
二裂委陵菜 Potentilla bifurca
0.043
0.069
0.068
0.071
钉柱委陵菜 Potentilla saundersiana
0.047
0.061
0.073
0.075
多裂委陵菜 Potentilla multifida
0.047
0.034
0.061
0.074
山莓草 Sibbaldia procumbens
0.014
-
-
-
CK, 对照; W, 增温; G, 放牧; WG, 增温+放牧。“-”代表此物种在该处理中未出现。 CK, control; W, warming; G, grazing; WG, the combination of warming and grazing. “-” represent for this species did not appear in treatment.
Fig. 6Mean vegetation coverage and net primary production (mean + SE) in control (CK), warming (W), grazing (G) and warming + grazing (WG) plots of alpine grassland in Northern Xizang. Different lowercase letters indicate significant differences in coverage and net primary production in different treatments (p < 0.05).
Fig. 7Mean species diversity (mean + SE) in control (CK), warming (W), grazing (G) and warming + grazing (WG) plots of alpine grassland in Northern Xizang. Different lowercase letters indicate significant differences in species diversity in different treatments (p < 0.05)
Fig. 8Community structure, species diversity and productivity (mean + SE) in warming (W), grazing (G) and warming + grazing (WG) plots of alpine grassland in Northern Xizang. Different lowercase letters indicate differences in treatment effect (p < 0.05); **, indicate differences between treatment effect and zero (p < 0.01).
.BrownJH, ValoneTJ, CurtinCG (1997). Reorganization of an arid ecosystem in response to recent climate?change .Proceedings of the National Academy of Sciences of the United States of America, 94, 9729-9733. DOI:10.1073/pnas.94.18.9729URLPMID:11038570 [本文引用: 1] Natural ecosystems contain many individuals and species interacting with each other and with their abiotic environment. Such systems can be expected to exhibit complex dynamics in which small perturbations can be amplified to cause large changes. Here, we document the reorganization of an arid ecosystem that has occurred since the late 1970s. The density of woody shrubs increased 3-fold. Several previously common animal species went locally extinct, while other previously rare species increased. While these changes are symptomatic of desertification, they were not caused by livestock grazing or drought, the principal causes of historical desertification. The changes apparently were caused by a shift in regional climate: since 1977 winter precipitation throughout the region was substantially higher than average for this century. These changes illustrate the kinds of large, unexpected responses of complex natural ecosystems that can occur in response to both natural perturbations and human activities.
.CaoXJ, GanjurjavH, LiangY, GaoQZ, ZhangY, LiYE, WanYF, DanjiuLB (2016). Temporal and spatial distribution of grassland degradation in northern Tibet based on NDVI .Acta Prataculturae Sinica, 25(3), 1-8. DOI:10.11686/cyxb2015378URL [本文引用: 2] An expansive alpine grassland ecosystem dominates the landscape in Northern Tibet, and the status of this grassland has significant implications for the major Asian rivers, and for the climate and eco-environment of Tibet and the wider region. The harsh natural environment and fragile ecosystem, together with increasing frequency and intensity of human activities such as pastoralism, and climate change, have resulted in increasing reports of degradation of the grassland ecosystem. We used vegetation cover as main indicator of ecosystem health, and with remote sensing monitoring, calculated the grassland degradation index (GDI) and evaluated grassland degradation in Northern Tibet from 2000 to 2010. The analysis was based on the Normalized Differential Vegetation Index (NDVI) data from the Advanced Very High-Resolution Radiometer (AVHRR) with a spatial resolution of 8 km from 1981 to 1985 and from Moderate Resolution Imaging Spectroradiometer (MODIS) with a spatial resolution of 0.25 km of 2000-2010. Then, we have analyzed the temporal and spatial distribution of grassland degradation in Northern Tibet from 2000 to 2010. It was found that grassland degradation in Northern Tibet was still severe. The GDI was 1.97 in 2010, near to seriously degraded status. The extremely seriously degraded and seriously degraded categories occupied 6.5% and 19.0% of the study area, respectively. The moderately and slightly degraded grassland categories accounted for 15.1% and 12.1%, respectively. Undegraded grassland occupied 41.8% of the total grassland area in 2010. The extent of extremely seriously degraded and seriously degraded categories was greatly increased in the data from 2000-2010, compared with the period from 1981-1985. However, the increase in grassland degradation in Northern Tibet from 2000 to 2010 was slight, with the mean value of the GDI being 1.84 in 2010, representing a change from light degradation to moderate degradation in Northern Tibet from 2000 to 2010. [曹旭娟, 干珠扎布, 梁艳, 高清竹, 张勇, 李玉娥, 万运帆, 旦久罗布 (2016). 基于NDVI的藏北地区草地退化时空分布特征分析 草业学报, 25(3), 1-8.] DOI:10.11686/cyxb2015378URL [本文引用: 2] An expansive alpine grassland ecosystem dominates the landscape in Northern Tibet, and the status of this grassland has significant implications for the major Asian rivers, and for the climate and eco-environment of Tibet and the wider region. The harsh natural environment and fragile ecosystem, together with increasing frequency and intensity of human activities such as pastoralism, and climate change, have resulted in increasing reports of degradation of the grassland ecosystem. We used vegetation cover as main indicator of ecosystem health, and with remote sensing monitoring, calculated the grassland degradation index (GDI) and evaluated grassland degradation in Northern Tibet from 2000 to 2010. The analysis was based on the Normalized Differential Vegetation Index (NDVI) data from the Advanced Very High-Resolution Radiometer (AVHRR) with a spatial resolution of 8 km from 1981 to 1985 and from Moderate Resolution Imaging Spectroradiometer (MODIS) with a spatial resolution of 0.25 km of 2000-2010. Then, we have analyzed the temporal and spatial distribution of grassland degradation in Northern Tibet from 2000 to 2010. It was found that grassland degradation in Northern Tibet was still severe. The GDI was 1.97 in 2010, near to seriously degraded status. The extremely seriously degraded and seriously degraded categories occupied 6.5% and 19.0% of the study area, respectively. The moderately and slightly degraded grassland categories accounted for 15.1% and 12.1%, respectively. Undegraded grassland occupied 41.8% of the total grassland area in 2010. The extent of extremely seriously degraded and seriously degraded categories was greatly increased in the data from 2000-2010, compared with the period from 1981-1985. However, the increase in grassland degradation in Northern Tibet from 2000 to 2010 was slight, with the mean value of the GDI being 1.84 in 2010, representing a change from light degradation to moderate degradation in Northern Tibet from 2000 to 2010.
.ChenH, ZhuQA, PengCH, WuN, WangYF, FangXQ, GaoYH, ZhuD, YangG, TianJQ, KangXM, PiaoSL, OuyangH, XiangWH, LuoZB, JiangH, SongXZ, ZhangY, YuGR, ZhaoXQ, GongP, YaoTD, WuJH (2013). The impacts of climate change and human activities on biogeochemical cycles on the Qinghai-Tibetan Plateau .Global Change Biology, 19, 2940-2955. DOI:10.1111/gcb.12277URL [本文引用: 1] With a pace of about twice the observed rate of global warming, the temperature on the Qinghai-Tibetan Plateau (Earth's third pole') has increased by 0.2 degrees C per decade over the past 50years, which results in significant permafrost thawing and glacier retreat. Our review suggested that warming enhanced net primary production and soil respiration, decreased methane (CH4) emissions from wetlands and increased CH4 consumption of meadows, but might increase CH4 emissions from lakes. Warming-induced permafrost thawing and glaciers melting would also result in substantial emission of old carbon dioxide (CO2) and CH4. Nitrous oxide (N2O) emission was not stimulated by warming itself, but might be slightly enhanced by wetting. However, there are many uncertainties in such biogeochemical cycles under climate change. Human activities (e.g. grazing, land cover changes) further modified the biogeochemical cycles and amplified such uncertainties on the plateau. If the projected warming and wetting continues, the future biogeochemical cycles will be more complicated. So facing research in this field is an ongoing challenge of integrating field observations with process-based ecosystem models to predict the impacts of future climate change and human activities at various temporal and spatial scales. To reduce the uncertainties and to improve the precision of the predictions of the impacts of climate change and human activities on biogeochemical cycles, efforts should focus on conducting more field observation studies, integrating data within improved models, and developing new knowledge about coupling among carbon, nitrogen, and phosphorus biogeochemical cycles as well as about the role of microbes in these cycles.
.NationalDevelopment and ReformCommission (2007). China’s National Climate Change Programme Environmental Policy Collection, 1-62. [本文引用: 1]
.DuanMJ, GaoQZ, WanYF, LiYE, GuoYQ, DanjiuLB, LuosangJC (2010). Effect of grazing on community characteristics and species diversity of Stipa purpurea alpine grassland in Northern Tibet. Acta Ecologica Sinica, 30, 3892-3900. [本文引用: 6]
.FuG, ShenZX, ZhangXZ (2018). Increased precipitation has stronger effects on plant production of an alpine meadow than does experimental warming in the Northern Tibetan Plateau . Agricultural and Forest Meteorology, 249, 11-21. DOI:10.1016/j.agrformet.2017.11.017URL [本文引用: 1]
.FuG, SunW, YuCQ, ZhangXZ, ShenZX, LiYL, YangPW, ZhouN (2015). Clipping alters the response of biomass production to experimental warming: A case study in an alpine meadow on the Tibetan Plateau, China .Journal of Mountain Science, 12, 935-942. DOI:10.1007/s11629-014-3035-zURL [本文引用: 1]
.FuG, ZhangXZ, ZhangYJ, ShiPL, LiYL, ZhouYT, YangPW, ShenZX (2013). Experimental warming does not enhance gross primary production and above-ground biomass in the alpine meadow of Tibet .Journal of Applied Remote Sensing, 7, 6451-6465. [本文引用: 1]
.GanjurjavH, GaoQZ, GornishES, SchwartzMW, LiangY, CaoXJ, ZhangWN, ZhangY, LiWH, WanYF, LiYE, DanjiuLB, GuoHB, LinED (2016). Differential response of alpine steppe and alpine meadow to climate warming in the central Qinghai-Tibetan Plateau .Agricultural and Forest Meteorology, 223, 233-240. DOI:10.1016/j.agrformet.2016.03.017URL [本文引用: 5]
.GanjurjavH, GuoYQ, GaoQZ, DuanMJ, WanYF, LiYE, DanjiuLB (2013). A study on optimal grazing rates in Stipa purpurea alpine grassland in Northern Tibet. Acta Prataculturae Sinica, 22(1), 130-137. DOI:10.11686/cyxb20130116URL [本文引用: 3]
.HoppingKA, KnappAK, DorjiT, KleinJA (2018). Warming and land use change concurrently erode ecosystem services in Tibet . Global Change Biology, 24, 5534-5548. DOI:10.1111/gcb.14417URLPMID:30086187 [本文引用: 2] Alpine meadows on the Tibetan Plateau comprise the largest alpine ecosystem in the world and provide critical ecosystem services, including forage production and carbon sequestration, on which people depend from local to global scales. However, the provision of these services may be threatened by climate warming combined with land use policies that are altering if and how pastoralists can continue to graze livestock, the dominant livelihood practice in this region for millennia. We synthesized findings from a climate warming and yak grazing experiment with landscape-level observations in central Tibet to gain insight into the trajectories of change that Tibet's alpine meadows will undergo in response to expected changes in climate and land use. We show that within 5?years, experimental warming drove an alpine community with intact, sedge-dominated turfs into a degraded state. With removal of livestock, consistent with policy intended to reverse degradation, a longer-term shift to a more shrub-dominated community will likely occur. Neither degraded nor shrub meadows produce forage or sequester carbon to the same degree as intact meadows, indicating that climate warming and drying will reduce the ability of Tibet's alpine meadows to provide key ecosystem services, and that livestock reduction policies intended to counteract trajectories of land degradation instead endanger contemporary livelihoods on the Tibetan Plateau.
.IPCC (2013). Climate Change 2013: The Physical Science Basis. Cambridge University Press, Cambridge, UK. 1535. [本文引用: 1]
.JiangYB, FanM, ZhangYJ (2017). Effect of short-term warming on plant community features of alpine meadow in Northern Tibet .Chinese Journal of Ecology, 36, 616-622. [本文引用: 1]
.KleinJA, HarteJ, ZhaoXQ (2005). Dynamic and complex microclimate responses to warming and grazing manipulations .Global Change Biology, 11, 1440-1451. DOI:10.1111/gcb.2005.11.issue-9URL [本文引用: 1]
.KleinJA, HarteJ, ZhaoXQ (2008). Decline in medicinal and forage species with warming is mediated by plant traits on the Tibetan Plateau .Ecosystems, 11, 775-789. DOI:10.1007/s10021-008-9160-1URL [本文引用: 1] Experimental studies of how global changes and human activities affect plant diversity often focus on broad measures of diversity and discuss the implications of these changes for ecosystem function. We examined how experimental warming and grazing affected species within plant groups of direct importance to Tibetan pastoralists: medicinal plants used by humans and palatable plants consumed by livestock. Warming resulted in species losses from both the medicinal and palatable plant groups; however, differential relative vulnerability to warming occurred. With respect to the percent of warming-induced species losses, the overall plant community lost 27%, medicinal plants lost 21%, and non-medicinal plants lost 40% of species. Losses of palatable and non-palatable species were similar to losses in the overall plant community. The deep-rootedness of medicinal plants resulted in lowered sensitivity to warming, whereas the shallow-rootedness of non-medicinal plants resulted in greater sensitivity to warming; the variable rooting depth of palatable and non-palatable plants resulted in an intermediate response to warming. Predicting the vulnerability of plant groups to human activities can be enhanced by knowledge of plant traits, their response to specific drivers, and their distribution within plant groups. Knowledge of the mechanisms through which a driver operates, and the evolutionary interaction of plants with that driver, will aid predictions. Future steps to protect ecosystem services furnished by medicinal and palatable plants will be required under the novel stress of a warmer climate. Grazing may be an important tool in maintaining some of these services under future warming.
.KurzI, O’ReillyCD, TunneyH (2006). Impact of cattle on soil physical properties and nutrient concentrations in overland flow from pasture in Ireland .Agriculture Ecosystems & Environment, 113, 378-390. [本文引用: 1]
.LiuHY, MiZR, LinL, WangYH, ZhangZH, ZhangFW, WangH, LiuLL, ZhuB, CaoGM, ZhaoXQ, SandersNJ, ClassenAT, ReichPB, HeJS (2018). Shifting plant species composition in response to climate change stabilizes grassland primary production .Proceedings of the National Academy of Sciences of the United States of America, 115, 4051-4056. DOI:10.1073/pnas.1700299114URLPMID:29666319 [本文引用: 5] The structure and function of alpine grassland ecosystems, including their extensive soil carbon stocks, are largely shaped by temperature. The Tibetan Plateau in particular has experienced significant warming over the past 50 y, and this warming trend is projected to intensify in the future. Such climate change will likely alter plant species composition and net primary production (NPP). Here we combined 32 y of observations and monitoring with a manipulative experiment of temperature and precipitation to explore the effects of changing climate on plant community structure and ecosystem function. First, long-term climate warming from 1983 to 2014, which occurred without systematic changes in precipitation, led to higher grass abundance and lower sedge abundance, but did not affect aboveground NPP. Second, an experimental warming experiment conducted over 4 y had no effects on any aspect of NPP, whereas drought manipulation (reducing precipitation by 50%), shifted NPP allocation belowground without affecting total NPP. Third, both experimental warming and drought treatments, supported by a meta-analysis at nine sites across the plateau, increased grass abundance at the expense of biomass of sedges and forbs. This shift in functional group composition led to deeper root systems, which may have enabled plant communities to acquire more water and thus stabilize ecosystem primary production even with a changing climate. Overall, our study demonstrates that shifting plant species composition in response to climate change may have stabilized primary production in this high-elevation ecosystem, but it also caused a shift from aboveground to belowground productivity.
.MarionB, BonisA, BouzilléJB (2010). How much does grazing- induced heterogeneity impact plant diversity in wet grasslands? écoscience, 17, 229-239. DOI:10.2980/17-3-3315URL [本文引用: 1]
.McNaughtonSJ (1979). Grazing as an optimization process: Grass-ungulate relationships in the Serengeti .The American Naturalist, 113, 691-703. DOI:10.1086/283426URL [本文引用: 1]
.NiuSL, XingXR, ZhangZ, XiaJY, ZhouXH, SongB, LiLH, WanSQ (2011). Water-use efficiency in response to climate change: From leaf to ecosystem in a temperate steppe .Global Change Biology, 17, 1073-1082. DOI:10.1111/gcb.2010.17.issue-2URL [本文引用: 1] Water-use efficiency (WUE) has been recognized as an important characteristic of ecosystem productivity, which links carbon (C) and water cycling. However, little is known about how WUE responds to climate change at different scales. Here, we investigated WUE at leaf, canopy, and ecosystem levels under increased precipitation and warming from 2005 to 2008 in a temperate steppe in Northern China. We measured gross ecosystem productivity (GEP), net ecosystem CO(2) exchange (NEE), evapotranspiration (ET), evaporation (E), canopy transpiration (T(c)), as well as leaf photosynthesis (P(max)) and transpiration (T(l)) of a dominant species to calculate canopy WUE (WUE(c)=GEP/T), ecosystem WUE (WUE(gep)=GEP/ET or WUE(nee)=NEE/ET) and leaf WUE (WUE(l)=P(max)/T(l)). The results showed that increased precipitation stimulated WUE(c), WUE(gep) and WUE(nee) by 17.1%, 10.2% and 12.6%, respectively, but decreased WUE(l) by 27.4%. Climate warming reduced canopy and ecosystem WUE over the 4 years but did not affect leaf level WUE. Across the 4 years and the measured plots, canopy and ecosystem WUE linearly increased, but leaf level WUE of the dominant species linearly decreased with increasing precipitation. The differential responses of canopy/ecosystem WUE and leaf WUE to climate change suggest that caution should be taken when upscaling WUE from leaf to larger scales. Our findings will also facilitate mechanistic understanding of the C-water relationships across different organism levels and in projecting the effects of climate warming and shifting precipitation regimes on productivity in arid and semiarid ecosystems.
.PolleyHW, DernerJD, JacksonRB, WilseyBJ, FayPA (2014). Impacts of climate change drivers on C4 grassland productivity: Scaling driver effects through the plant community .Journal of Experimental Botany, 65, 3415-3424. DOI:10.1093/jxb/eru009URL [本文引用: 1] Climate change drivers affect plant community productivity via three pathways: (i) direct effects of drivers on plants; (ii) the response of species abundances to drivers (community response); and (iii) the feedback effect of community change on productivity (community effect). The contribution of each pathway to driver-productivity relationships depends on functional traits of dominant species. We used data from three experiments in Texas, USA, to assess the role of community dynamics in the aboveground net primary productivity (ANPP) response of C-4 grasslands to two climate drivers applied singly: atmospheric CO2 enrichment and augmented summer precipitation. The ANPP-driver response differed among experiments because community responses and effects differed. ANPP increased by 80-120 g m(-2) per 100 mu l l(-1) rise in CO2 in separate experiments with pasture and tallgrass prairie assemblages. Augmenting ambient precipitation by 128 mm during one summer month each year increased ANPP more in native than in exotic communities in a third experiment. The community effect accounted for 21-38% of the ANPP CO2 response in the prairie experiment but little of the response in the pasture experiment. The community response to CO2 was linked to species traits associated with greater soil water from reduced transpiration (e. g. greater height). Community effects on the ANPP CO2 response and the greater ANPP response of native than exotic communities to augmented precipitation depended on species differences in transpiration efficiency. These results indicate that feedbacks from community change influenced ANPP-driver responses. However, the species traits that regulated community effects on ANPP differed from the traits that determined how communities responded to drivers.
.TilmanD, ReichPB, KnopsJ, WedinD, MielkeT, LehmanC (2001). Diversity and productivity in a long-term grassland experiment .Science, 294, 843-845. DOI:10.1126/science.1060391URLPMID:11679667 [本文引用: 1] Plant diversity and niche complementarity had progressively stronger effects on ecosystem functioning during a 7-year experiment, with 16-species plots attaining 2.7 times greater biomass than monocultures. Diversity effects were neither transients nor explained solely by a few productive or unviable species. Rather, many higher-diversity plots outperformed the best monoculture. These results help resolve debate over biodiversity and ecosystem functioning, show effects at higher than expected diversity levels, and demonstrate, for these ecosystems, that even the best-chosen monocultures cannot achieve greater productivity or carbon stores than higher-diversity sites.
.WangSP, DuanJC, XuGP, WangYF, ZhangZH, RuiYC, LuoCY, XuB, ZhuXX, ChangXF, CuiXY, NiuHS, ZhaoXQ, WangWY (2012). Effects of warming and grazing on soil N availability, species composition, and ANPP in an alpine meadow . Ecology, 93, 2365-2376. URL [本文引用: 5] Uncertainty about the effects of warming and grazing on soil nitrogen (N) availability, species composition, and aboveground net primary production (ANPP) limits our ability to predict how global carbon sequestration will vary under future warming with grazing in alpine regions. Through a controlled asymmetrical warming (1.2/1.7 degrees C during daytime/nighttime) with a grazing experiment from 2006 to 2010 in an alpine meadow, we found that warming alone and moderate grazing did not significantly affect soil net N mineralization. Although plant species richness significantly decreased by 10% due to warming after 2008, we caution that this may be due to the transient occurrence or disappearance of some rare plant species in all treatments. Warming significantly increased graminoid cover, except in 2009, and legume cover after 2008, but reduced non-legume forb cover in the community. Grazing significantly decreased cover of graminoids and legumes before 2009 but increased forb cover in 2010. Warming significantly increased ANPP regardless of grazing, whereas grazing reduced the response of ANPP to warming. N addition did not affect ANPP in both warming and grazing treatments. Our findings suggest that soil N availability does not determine ANPP under simulated warming and that heavy grazing rather than warming causes degradation of the alpine meadows.
.WangSP, LiYH, ChenZZ (1999). The optimal stocking rate on grazing system in Inner Mongolia steppe II. Based on relationship between stocking rate and aboveground net primary productivity .Acta Agrestia Sinica, 7, 192-197. [本文引用: 1]
.WangXM, ChenFH, DongZB (2006). The relative role of climatic and human factors in desertification in semiarid China .Global Environmental Change, 16, 48-57. DOI:10.1016/j.gloenvcha.2005.06.006URL [本文引用: 1]
.WuQ, HanGD, WangRZ, LiuF, QinJ (2016). Effects of simulated warming on grassland plants, soil and ecosystem carbon exchange .Chinese Journal of Grassland, 38, 105-114. [本文引用: 1]
.WuZT, DijkstraP, KochGW, Pe?uelasJ, HungateBA (2011). Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation .Global Change Biology, 17, 927-942. DOI:10.1111/j.1365-2486.2010.02302.xURL [本文引用: 1] Global mean temperature is predicted to increase by 2-7 degrees C and precipitation to change across the globe by the end of this century. To quantify climate effects on ecosystem processes, a number of climate change experiments have been established around the world in various ecosystems. Despite these efforts, general responses of terrestrial ecosystems to changes in temperature and precipitation, and especially to their combined effects, remain unclear. We used meta-analysis to synthesize ecosystem-level responses to warming, altered precipitation, and their combination. We focused on plant growth and ecosystem carbon (C) balance, including biomass, net primary production (NPP), respiration, net ecosystem exchange (NEE), and ecosystem photosynthesis, synthesizing results from 85 studies. We found that experimental warming and increased precipitation generally stimulated plant growth and ecosystem C fluxes, whereas decreased precipitation had the opposite effects. For example, warming significantly stimulated total NPP, increased ecosystem photosynthesis, and ecosystem respiration. Experimentally reduced precipitation suppressed aboveground NPP (ANPP) and NEE, whereas supplemental precipitation enhanced ANPP and NEE. Plant productivity and ecosystem C fluxes generally showed higher sensitivities to increased precipitation than to decreased precipitation. Interactive effects of warming and altered precipitation tended to be smaller than expected from additive, single-factor effects, though low statistical power limits the strength of these conclusions. New experiments with combined temperature and precipitation manipulations are needed to conclusively determine the importance of temperature-precipitation interactions on the C balance of terrestrial ecosystems under future climate conditions.
.XuMH, LiuM, XueX, ZhaiDT, PengF, YouQG, LiuY (2015). Effects of warming and clipping on the growth of aboveground vegetation in an alpine meadow .Ecology and Environmental Sciences, 24, 231-236. [本文引用: 1]
.YuCQ, HanFS, FuG (2019). Effects of 7 years experimental warming on soil bacterial and fungal community structure in the Northern Tibet alpine meadow at three elevations .Science of the Total Environment, 655, 814-822. DOI:10.1016/j.scitotenv.2018.11.309URLPMID:30481708 [本文引用: 1] A warming experiment was established along an altitudinal gradient (low elevation: 4313?m, mid-elevation: 4513?m and high elevation: 4693?m) in alpine meadows of the Northern Tibet to investigate the effects of warming on soil bacterial and fungal community structure. Elevation had significant effects on vegetation community coverage (CC), soil temperature (Ts) and pH, but not soil fungal diversity. Soil bacterial diversity at the high elevation was significantly lower than that at the low and mid-elevations, whereas there was no significant difference of soil bacterial diversity between the low and mid-elevations. After seven years of warming, soil fungal diversity was significantly increased at the mid-elevation but not the low and high elevations, and soil bacterial diversity was not significantly altered at the low, mid- and high elevations. Soil bacterial community structure was significantly altered at the low and mid-elevations but not the high elevation. Soil fungal community structure was significantly altered at all the three elevations. CC, Ts and pH significantly explained 20.55%, 5.30% and 12.61% of the variation of bacterial community structure, respectively. CC and Ts significantly explained 17.40% and 5.86% of the variation of fungal community structure, respectively. Therefore, responses of soil microbial community structure to warming may vary with elevation, which was mainly attributed to different vegetation coverage, soil temperature and/or pH conditions among the three elevations in this study alpine meadows.
.YuCQ, ShenZX, ZhangXZ, SunW, FuG (2014). Response of soil C and N, dissolved organic C and N, and inorganic N to short-term experimental warming in an alpine meadow on the Tibetan Plateau .The Scientific World Journal, 2014, 152576. DOI: 10.1155/2014/152576. DOI:10.1155/2014/152576URLPMID:24977179 [本文引用: 1] Although alpine meadows of Tibet are expected to be strongly affected by climatic warming, it remains unclear how soil organic C (SOC), total N (TN), ammonium N (NH4 (+)-N) , nitrate N (NO3 (+)-N), and dissolved organic C (DOC) and N (DON) respond to warming. This study aims to investigate the responses of these C and N pools to short-term experimental warming in an alpine meadow of Tibet. A warming experiment using open top chambers was conducted in an alpine meadow at three elevations (i.e., a low (4313 m), mid-(4513 m), and high (4693 m) elevation) in May 2010. Topsoil (0-20 cm depth) samples were collected in July-September 2011. Experimental warming increased soil temperature by ~1-1.4°C but decreased soil moisture by ~0.04 m(3) m(-3). Experimental warming had little effects on SOC, TN, DOC, and DON, which may be related to lower warming magnitude, the short period of warming treatment, and experimental warming-induced soil drying by decreasing soil microbial activity. Experimental warming decreased significantly inorganic N at the two lower elevations,but had negligible effect at the high elevation. Our findings suggested that the effects of short-term experimental warming on SOC, TN and dissolved organic matter were insignificant, only affecting inorganic forms.
.YuanJL, JiangXL, HuangWB, WangG (2004). Effects of grazing intensity and grazing season on plant species diversity in alpine meadow .Acta Prataculturae Sinica, 13(3), 16-21. [本文引用: 1]
.ZhaoW, ChenSP, LinGH (2008). Compensatory growth responses to clipping defoliation in Leymus chinensis (Poaceae) under nutrient addition and water deficiency conditions Plant Ecology,196, 85-99. DOI:10.1007/s11258-007-9336-3URL [本文引用: 1] Compensatory growth responses of Leymus chinensis, a dominant species in Inner Mongolia steppe, to clipping defoliation were evaluated in a pot-cultivated experiment under different nutrient (N and P) and water availability conditions. Leymus chinensis exhibited over-compensatory growth at the light and moderate clipping intensities (20% and 40% aerial mass removed) with a greater accumulated aboveground biomass, higher relative growth rate (RGR), more rhizomatic tillers and a stimulation of compensatory photosynthesis to the remnant leaves as compared with those of the unclipped plants. Intense clipping (80% aerial mass removed), which removed most of the aboveground tissues, greatly reduced the growth of aboveground biomass in comparison with that of the unclipped plants. Nitrogen addition only slightly improved the biomass production and RGR in light and moderately clipped plants, and it did not allow plants in the intense clipping condition to over-compensate. Phosphorus addition had no obvious influences on the growth and physiological responses to clipping defoliation. These results indicated that nutrient addition could not compensate for the negative effects of severe clipping on the defoliated grass. On the other hand, there were no distinct positive responses under water deficiency condition for L. chinensis at all clipping intensities with a significant reduction of aboveground and belowground biomass, lower RGR, fewer rhizomatic tillers, and a lower net photosynthetic rate than other wet treatments. Additionally, the chlorophyll contents of remnant leaves gradually increased with the increase of clipping intensities in each treatment. In conclusion, although L. chinensis could compensate for tissues removal by some morphological and physiological responses, intense clipping and drought can result in a significant decrease of biomass and growth rate, even under enriched nutrition conditions.
.ZhengW, DongQM, LiSX, LiHT, LiuY, YangSH (2012). Impact of grazing intensities on community biodiversity and productivity of alpine grassland in Qinghai Lake region .Acta Agrestia Sinica, 20, 1033-1038. [本文引用: 4]
.ZhouXH, WanSQ, LuoYQ (2007). Source components and interannual variability of soil CO2, efflux under experimental warming and clipping in a grassland ecosystem .Global Change Biology, 13, 761-775. DOI:10.1007/s10393-016-1183-zURLPMID:27660213 [本文引用: 1] Certain bat species serve as natural reservoirs for pathogens in several key viral families including henipa-, lyssa-, corona-, and filoviruses, which may pose serious threats to human health. The Common Vampire Bat (Desmodus rotundus), due to its abundance, sanguivorous feeding habit involving humans and domestic animals, and highly social behavioral ecology, may have an unusually high potential for interspecies disease transmission. Previous studies have investigated rabies dynamics in D. rotundus, yet the diversity of other viruses, bacteria, and other microbes that these bats may carry remains largely unknown. We screened 396 blood, urine, saliva, and fecal samples from D. rotundus captured in Guatemala for 13 viral families and genera. Positive results were found for rhabdovirus, adenovirus, and herpesvirus assays. We also screened these samples for Bartonella spp. and found that 38% of individuals tested positive. To characterize potential for interspecies transmission associated with feeding behavior, we also analyzed cytochrome B sequences from fecal samples to identify prey species and found that domestic cattle (Bos taurus) made up the majority of blood meals. Our findings suggest that the risk of pathogen spillover from Desmodus rotundus, including between domestic animal species, is possible and warrants further investigation to characterize this microbial diversity and expand our understanding of foraging ecology in their populations. Reorganization of an arid ecosystem in response to recent climate?change 1 1997
... 全球气候变化已成为世界各国政府和公众关注的焦点.IPCC第五次评估报告指出, 1880-2012年间全球地表平均温度上升0.85 ℃, 预计到21世纪末全球大气温度将增加1.5-4.8 ℃ (IPCC, 2013).草地是陆地生态系统的重要组成部分, 且极为脆弱, 对气候变化极其敏感.气候变化直接或间接地影响草地植物的群落结构与组成(Polley et al., 2014; Ganjurjav et al., 2016)、物种多样性及生产力(Wu et al., 2011; Wang et al., 2012).除气候变化外, 人类活动也是影响草地植物群落的重要因子.有研究认为, 人类活动对植物的影响大于环境因子, 甚至成为控制植物群落多样性的决定因素(Brown et al., 1997; Wang et al., 2006).放牧是草地生态系统最为重要的人类干扰因素, 植物物种多样性与放牧强度显著相关(段敏杰等, 2010; 郑伟等, 2012).放牧作为草地最普遍的利用方式, 牲畜通过采食和践踏行为对草地植物群落特征产生影响, 同时也会改变局地光、水和土壤养分利用率, 从而间接改变群落物种结构与分布(Zhou et al., 2007; Zhao et al., 2008). ...
基于NDVI的藏北地区草地退化时空分布特征分析 2 2016
... 藏北高原位于青藏高原的腹地, 被称为“世界屋脊的屋脊”.该地区拥有42.1万km2高寒草地(高清竹等, 2006), 是西藏自治区主要的畜牧业生产基地和我国重要的生态屏障区, 是全球气候变化的启动区和敏感区, 是亚洲中部高寒环境中最为典型的一种生态系统.据报道, 青藏高原增温幅度高达每10年0.2 ℃ (Chen et al., 2013), 至2030年该地区将升温2.2-2.6 ℃ (National Development and Reform Commisssion, 2007), 显著高于全球和全国平均水平.由于过度放牧和草地不合理利用, 藏北高原草地退化面积达到58.2%, 重度退化和极重度退化草地面积分别占19.0%和6.5% (曹旭娟等, 2016).近年来, 研究人员分别开展了增温和放牧对高寒草地植物生产力和物种多样性影响的研究(段敏杰等, 2010; Ganjurjav et al., 2016; Liu et al., 2018), 但增温和放牧的交互作用对物种多样性和生产力影响的研究鲜有报道.一些****通过刈割模拟放牧活动, 从而研究增温和放牧的交互作用对高寒草地群落结构及生产力的影响(Wang et al., 2012; Fu et al., 2015).然而, 上述研究在很大程度上忽略了牲畜的践踏和采食作用对草地植物群落的影响.此外, 放牧与刈割的不同之处还在于对物种组成的影响, 放牧条件下牲畜对草地植物的采食具有物种间的选择喜好, 而刈割仅是对草地植物平均冠层高度进行统一收割(Klein et al., 2008; 段敏杰等, 2010; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
... 放牧和增温会引起草地植物群落结构与物种多样性的改变, 进而对植物生产力产生重要影响(Wang et al., 2012; Hopping et al., 2018; Liu et al., 2018).植物生产力与物种多样性密切相关, 较高的物种多样性可以维持较高的植物生产力, 同时也可以加强群落生态系统的稳定性(Tilman et al., 2001; 马文静等, 2013).本研究表明, 由于放牧具有补偿效应(McNaughton, 1979), 在物种多样性增高2%时, 地上净初级生产力增加29%.通过对牲畜采食量分析后发现, 2年间杂草采食量仅占总量的21.8%, 而禾草采食量高达78.2%, 与前人的研究成果一致(段敏杰等, 2010; 郑伟等, 2012; 干珠扎布等, 2013).本研究中, 增温对净初级生产力无显著影响, 但2016和2017年增温对净初级生产力的影响存在年际差异(图6B), 这可能是由于2017年降水量高于2016年造成的(图1, 图3C).Fu等(2018)在藏北高原高寒草地的研究也表明, 年际间的降水差异是引起增温对生产力年际差异性影响的主要原因.此外, 有研究表明, 在不同的空气温度背景下, 增温对草地净初级生产力的影响有所不同(Liu et al., 2018).因此, 两年间的温度差异也可能引起生产力对增温的差异性响应. ...
基于NDVI的藏北地区草地退化时空分布特征分析 2 2016
... 藏北高原位于青藏高原的腹地, 被称为“世界屋脊的屋脊”.该地区拥有42.1万km2高寒草地(高清竹等, 2006), 是西藏自治区主要的畜牧业生产基地和我国重要的生态屏障区, 是全球气候变化的启动区和敏感区, 是亚洲中部高寒环境中最为典型的一种生态系统.据报道, 青藏高原增温幅度高达每10年0.2 ℃ (Chen et al., 2013), 至2030年该地区将升温2.2-2.6 ℃ (National Development and Reform Commisssion, 2007), 显著高于全球和全国平均水平.由于过度放牧和草地不合理利用, 藏北高原草地退化面积达到58.2%, 重度退化和极重度退化草地面积分别占19.0%和6.5% (曹旭娟等, 2016).近年来, 研究人员分别开展了增温和放牧对高寒草地植物生产力和物种多样性影响的研究(段敏杰等, 2010; Ganjurjav et al., 2016; Liu et al., 2018), 但增温和放牧的交互作用对物种多样性和生产力影响的研究鲜有报道.一些****通过刈割模拟放牧活动, 从而研究增温和放牧的交互作用对高寒草地群落结构及生产力的影响(Wang et al., 2012; Fu et al., 2015).然而, 上述研究在很大程度上忽略了牲畜的践踏和采食作用对草地植物群落的影响.此外, 放牧与刈割的不同之处还在于对物种组成的影响, 放牧条件下牲畜对草地植物的采食具有物种间的选择喜好, 而刈割仅是对草地植物平均冠层高度进行统一收割(Klein et al., 2008; 段敏杰等, 2010; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
... 放牧和增温会引起草地植物群落结构与物种多样性的改变, 进而对植物生产力产生重要影响(Wang et al., 2012; Hopping et al., 2018; Liu et al., 2018).植物生产力与物种多样性密切相关, 较高的物种多样性可以维持较高的植物生产力, 同时也可以加强群落生态系统的稳定性(Tilman et al., 2001; 马文静等, 2013).本研究表明, 由于放牧具有补偿效应(McNaughton, 1979), 在物种多样性增高2%时, 地上净初级生产力增加29%.通过对牲畜采食量分析后发现, 2年间杂草采食量仅占总量的21.8%, 而禾草采食量高达78.2%, 与前人的研究成果一致(段敏杰等, 2010; 郑伟等, 2012; 干珠扎布等, 2013).本研究中, 增温对净初级生产力无显著影响, 但2016和2017年增温对净初级生产力的影响存在年际差异(图6B), 这可能是由于2017年降水量高于2016年造成的(图1, 图3C).Fu等(2018)在藏北高原高寒草地的研究也表明, 年际间的降水差异是引起增温对生产力年际差异性影响的主要原因.此外, 有研究表明, 在不同的空气温度背景下, 增温对草地净初级生产力的影响有所不同(Liu et al., 2018).因此, 两年间的温度差异也可能引起生产力对增温的差异性响应. ...
The impacts of climate change and human activities on biogeochemical cycles on the Qinghai-Tibetan Plateau 1 2013
... 藏北高原位于青藏高原的腹地, 被称为“世界屋脊的屋脊”.该地区拥有42.1万km2高寒草地(高清竹等, 2006), 是西藏自治区主要的畜牧业生产基地和我国重要的生态屏障区, 是全球气候变化的启动区和敏感区, 是亚洲中部高寒环境中最为典型的一种生态系统.据报道, 青藏高原增温幅度高达每10年0.2 ℃ (Chen et al., 2013), 至2030年该地区将升温2.2-2.6 ℃ (National Development and Reform Commisssion, 2007), 显著高于全球和全国平均水平.由于过度放牧和草地不合理利用, 藏北高原草地退化面积达到58.2%, 重度退化和极重度退化草地面积分别占19.0%和6.5% (曹旭娟等, 2016).近年来, 研究人员分别开展了增温和放牧对高寒草地植物生产力和物种多样性影响的研究(段敏杰等, 2010; Ganjurjav et al., 2016; Liu et al., 2018), 但增温和放牧的交互作用对物种多样性和生产力影响的研究鲜有报道.一些****通过刈割模拟放牧活动, 从而研究增温和放牧的交互作用对高寒草地群落结构及生产力的影响(Wang et al., 2012; Fu et al., 2015).然而, 上述研究在很大程度上忽略了牲畜的践踏和采食作用对草地植物群落的影响.此外, 放牧与刈割的不同之处还在于对物种组成的影响, 放牧条件下牲畜对草地植物的采食具有物种间的选择喜好, 而刈割仅是对草地植物平均冠层高度进行统一收割(Klein et al., 2008; 段敏杰等, 2010; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
China’s National Climate Change Programme 1 2007
... 藏北高原位于青藏高原的腹地, 被称为“世界屋脊的屋脊”.该地区拥有42.1万km2高寒草地(高清竹等, 2006), 是西藏自治区主要的畜牧业生产基地和我国重要的生态屏障区, 是全球气候变化的启动区和敏感区, 是亚洲中部高寒环境中最为典型的一种生态系统.据报道, 青藏高原增温幅度高达每10年0.2 ℃ (Chen et al., 2013), 至2030年该地区将升温2.2-2.6 ℃ (National Development and Reform Commisssion, 2007), 显著高于全球和全国平均水平.由于过度放牧和草地不合理利用, 藏北高原草地退化面积达到58.2%, 重度退化和极重度退化草地面积分别占19.0%和6.5% (曹旭娟等, 2016).近年来, 研究人员分别开展了增温和放牧对高寒草地植物生产力和物种多样性影响的研究(段敏杰等, 2010; Ganjurjav et al., 2016; Liu et al., 2018), 但增温和放牧的交互作用对物种多样性和生产力影响的研究鲜有报道.一些****通过刈割模拟放牧活动, 从而研究增温和放牧的交互作用对高寒草地群落结构及生产力的影响(Wang et al., 2012; Fu et al., 2015).然而, 上述研究在很大程度上忽略了牲畜的践踏和采食作用对草地植物群落的影响.此外, 放牧与刈割的不同之处还在于对物种组成的影响, 放牧条件下牲畜对草地植物的采食具有物种间的选择喜好, 而刈割仅是对草地植物平均冠层高度进行统一收割(Klein et al., 2008; 段敏杰等, 2010; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
放牧对藏北紫花针茅高寒草原植物群落特征的影响 6 2010
... 全球气候变化已成为世界各国政府和公众关注的焦点.IPCC第五次评估报告指出, 1880-2012年间全球地表平均温度上升0.85 ℃, 预计到21世纪末全球大气温度将增加1.5-4.8 ℃ (IPCC, 2013).草地是陆地生态系统的重要组成部分, 且极为脆弱, 对气候变化极其敏感.气候变化直接或间接地影响草地植物的群落结构与组成(Polley et al., 2014; Ganjurjav et al., 2016)、物种多样性及生产力(Wu et al., 2011; Wang et al., 2012).除气候变化外, 人类活动也是影响草地植物群落的重要因子.有研究认为, 人类活动对植物的影响大于环境因子, 甚至成为控制植物群落多样性的决定因素(Brown et al., 1997; Wang et al., 2006).放牧是草地生态系统最为重要的人类干扰因素, 植物物种多样性与放牧强度显著相关(段敏杰等, 2010; 郑伟等, 2012).放牧作为草地最普遍的利用方式, 牲畜通过采食和践踏行为对草地植物群落特征产生影响, 同时也会改变局地光、水和土壤养分利用率, 从而间接改变群落物种结构与分布(Zhou et al., 2007; Zhao et al., 2008). ...
... 藏北高原位于青藏高原的腹地, 被称为“世界屋脊的屋脊”.该地区拥有42.1万km2高寒草地(高清竹等, 2006), 是西藏自治区主要的畜牧业生产基地和我国重要的生态屏障区, 是全球气候变化的启动区和敏感区, 是亚洲中部高寒环境中最为典型的一种生态系统.据报道, 青藏高原增温幅度高达每10年0.2 ℃ (Chen et al., 2013), 至2030年该地区将升温2.2-2.6 ℃ (National Development and Reform Commisssion, 2007), 显著高于全球和全国平均水平.由于过度放牧和草地不合理利用, 藏北高原草地退化面积达到58.2%, 重度退化和极重度退化草地面积分别占19.0%和6.5% (曹旭娟等, 2016).近年来, 研究人员分别开展了增温和放牧对高寒草地植物生产力和物种多样性影响的研究(段敏杰等, 2010; Ganjurjav et al., 2016; Liu et al., 2018), 但增温和放牧的交互作用对物种多样性和生产力影响的研究鲜有报道.一些****通过刈割模拟放牧活动, 从而研究增温和放牧的交互作用对高寒草地群落结构及生产力的影响(Wang et al., 2012; Fu et al., 2015).然而, 上述研究在很大程度上忽略了牲畜的践踏和采食作用对草地植物群落的影响.此外, 放牧与刈割的不同之处还在于对物种组成的影响, 放牧条件下牲畜对草地植物的采食具有物种间的选择喜好, 而刈割仅是对草地植物平均冠层高度进行统一收割(Klein et al., 2008; 段敏杰等, 2010; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
... ; 段敏杰等, 2010; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
... 放牧和增温对高寒草地植物群落结构具有重要影响(段敏杰等, 2010; Ganjurjav et al., 2016).不同功能群植物高度、盖度及优势度的变化, 主要是受放牧过程中牲畜选择性采食的影响.可食性较高的禾草类和莎草类被家畜优先采食, 导致其高度、盖度及优势度降低, 而可食性较低的杂类草物种比例增加(段敏杰等, 2010; 何婷, 2015).这与本研究中放牧显著降低植物高度, 增加杂类草物种比例的结论一致.增温一方面满足植物生长的热量需求, 另一方面却加速土壤水分蒸发, 降低土壤水分可利用率, 影响群落的物种组成(Ganjurjav et al., 2016).本研究表明, 增温显著改变高寒草地植物群落特征, 植物高度增加82.3%, 盖度降低29.5%.OTC的阻挡作用使热量不易散失满足了植物生长发育的热量需求, 同时增温也改变了土壤无机氮的含量(Yu et al., 2014, 2019), 进而满足植物生长的个体发育, 抑制植物的分蘖繁殖(武倩等, 2016)导致盖度降低(Hopping et al., 2018).此外, OTC增温的同时加剧温室内土壤水分的蒸发, 降低其土壤湿度(Liu et al., 2018), 改变了温室内植物生长的微气候环境, 改变了植物对土壤水分和养分的吸收(Klein et al., 2005), 进而影响植物群落组成.草地生态系统作为陆地生态系统的重要组成部分, 受气候变化和人类活动的双重影响.本研究表明, 放牧和模拟增温对高寒草地植物高度具有显著交互作用.在放牧条件下, 增温对植物高度无显著影响; 然而, 在不放牧条件下, 增温显著增加了植物高度. ...
... ).不同功能群植物高度、盖度及优势度的变化, 主要是受放牧过程中牲畜选择性采食的影响.可食性较高的禾草类和莎草类被家畜优先采食, 导致其高度、盖度及优势度降低, 而可食性较低的杂类草物种比例增加(段敏杰等, 2010; 何婷, 2015).这与本研究中放牧显著降低植物高度, 增加杂类草物种比例的结论一致.增温一方面满足植物生长的热量需求, 另一方面却加速土壤水分蒸发, 降低土壤水分可利用率, 影响群落的物种组成(Ganjurjav et al., 2016).本研究表明, 增温显著改变高寒草地植物群落特征, 植物高度增加82.3%, 盖度降低29.5%.OTC的阻挡作用使热量不易散失满足了植物生长发育的热量需求, 同时增温也改变了土壤无机氮的含量(Yu et al., 2014, 2019), 进而满足植物生长的个体发育, 抑制植物的分蘖繁殖(武倩等, 2016)导致盖度降低(Hopping et al., 2018).此外, OTC增温的同时加剧温室内土壤水分的蒸发, 降低其土壤湿度(Liu et al., 2018), 改变了温室内植物生长的微气候环境, 改变了植物对土壤水分和养分的吸收(Klein et al., 2005), 进而影响植物群落组成.草地生态系统作为陆地生态系统的重要组成部分, 受气候变化和人类活动的双重影响.本研究表明, 放牧和模拟增温对高寒草地植物高度具有显著交互作用.在放牧条件下, 增温对植物高度无显著影响; 然而, 在不放牧条件下, 增温显著增加了植物高度. ...
... 放牧和增温会引起草地植物群落结构与物种多样性的改变, 进而对植物生产力产生重要影响(Wang et al., 2012; Hopping et al., 2018; Liu et al., 2018).植物生产力与物种多样性密切相关, 较高的物种多样性可以维持较高的植物生产力, 同时也可以加强群落生态系统的稳定性(Tilman et al., 2001; 马文静等, 2013).本研究表明, 由于放牧具有补偿效应(McNaughton, 1979), 在物种多样性增高2%时, 地上净初级生产力增加29%.通过对牲畜采食量分析后发现, 2年间杂草采食量仅占总量的21.8%, 而禾草采食量高达78.2%, 与前人的研究成果一致(段敏杰等, 2010; 郑伟等, 2012; 干珠扎布等, 2013).本研究中, 增温对净初级生产力无显著影响, 但2016和2017年增温对净初级生产力的影响存在年际差异(图6B), 这可能是由于2017年降水量高于2016年造成的(图1, 图3C).Fu等(2018)在藏北高原高寒草地的研究也表明, 年际间的降水差异是引起增温对生产力年际差异性影响的主要原因.此外, 有研究表明, 在不同的空气温度背景下, 增温对草地净初级生产力的影响有所不同(Liu et al., 2018).因此, 两年间的温度差异也可能引起生产力对增温的差异性响应. ...
放牧对藏北紫花针茅高寒草原植物群落特征的影响 6 2010
... 全球气候变化已成为世界各国政府和公众关注的焦点.IPCC第五次评估报告指出, 1880-2012年间全球地表平均温度上升0.85 ℃, 预计到21世纪末全球大气温度将增加1.5-4.8 ℃ (IPCC, 2013).草地是陆地生态系统的重要组成部分, 且极为脆弱, 对气候变化极其敏感.气候变化直接或间接地影响草地植物的群落结构与组成(Polley et al., 2014; Ganjurjav et al., 2016)、物种多样性及生产力(Wu et al., 2011; Wang et al., 2012).除气候变化外, 人类活动也是影响草地植物群落的重要因子.有研究认为, 人类活动对植物的影响大于环境因子, 甚至成为控制植物群落多样性的决定因素(Brown et al., 1997; Wang et al., 2006).放牧是草地生态系统最为重要的人类干扰因素, 植物物种多样性与放牧强度显著相关(段敏杰等, 2010; 郑伟等, 2012).放牧作为草地最普遍的利用方式, 牲畜通过采食和践踏行为对草地植物群落特征产生影响, 同时也会改变局地光、水和土壤养分利用率, 从而间接改变群落物种结构与分布(Zhou et al., 2007; Zhao et al., 2008). ...
... 藏北高原位于青藏高原的腹地, 被称为“世界屋脊的屋脊”.该地区拥有42.1万km2高寒草地(高清竹等, 2006), 是西藏自治区主要的畜牧业生产基地和我国重要的生态屏障区, 是全球气候变化的启动区和敏感区, 是亚洲中部高寒环境中最为典型的一种生态系统.据报道, 青藏高原增温幅度高达每10年0.2 ℃ (Chen et al., 2013), 至2030年该地区将升温2.2-2.6 ℃ (National Development and Reform Commisssion, 2007), 显著高于全球和全国平均水平.由于过度放牧和草地不合理利用, 藏北高原草地退化面积达到58.2%, 重度退化和极重度退化草地面积分别占19.0%和6.5% (曹旭娟等, 2016).近年来, 研究人员分别开展了增温和放牧对高寒草地植物生产力和物种多样性影响的研究(段敏杰等, 2010; Ganjurjav et al., 2016; Liu et al., 2018), 但增温和放牧的交互作用对物种多样性和生产力影响的研究鲜有报道.一些****通过刈割模拟放牧活动, 从而研究增温和放牧的交互作用对高寒草地群落结构及生产力的影响(Wang et al., 2012; Fu et al., 2015).然而, 上述研究在很大程度上忽略了牲畜的践踏和采食作用对草地植物群落的影响.此外, 放牧与刈割的不同之处还在于对物种组成的影响, 放牧条件下牲畜对草地植物的采食具有物种间的选择喜好, 而刈割仅是对草地植物平均冠层高度进行统一收割(Klein et al., 2008; 段敏杰等, 2010; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
... ; 段敏杰等, 2010; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
... 放牧和增温对高寒草地植物群落结构具有重要影响(段敏杰等, 2010; Ganjurjav et al., 2016).不同功能群植物高度、盖度及优势度的变化, 主要是受放牧过程中牲畜选择性采食的影响.可食性较高的禾草类和莎草类被家畜优先采食, 导致其高度、盖度及优势度降低, 而可食性较低的杂类草物种比例增加(段敏杰等, 2010; 何婷, 2015).这与本研究中放牧显著降低植物高度, 增加杂类草物种比例的结论一致.增温一方面满足植物生长的热量需求, 另一方面却加速土壤水分蒸发, 降低土壤水分可利用率, 影响群落的物种组成(Ganjurjav et al., 2016).本研究表明, 增温显著改变高寒草地植物群落特征, 植物高度增加82.3%, 盖度降低29.5%.OTC的阻挡作用使热量不易散失满足了植物生长发育的热量需求, 同时增温也改变了土壤无机氮的含量(Yu et al., 2014, 2019), 进而满足植物生长的个体发育, 抑制植物的分蘖繁殖(武倩等, 2016)导致盖度降低(Hopping et al., 2018).此外, OTC增温的同时加剧温室内土壤水分的蒸发, 降低其土壤湿度(Liu et al., 2018), 改变了温室内植物生长的微气候环境, 改变了植物对土壤水分和养分的吸收(Klein et al., 2005), 进而影响植物群落组成.草地生态系统作为陆地生态系统的重要组成部分, 受气候变化和人类活动的双重影响.本研究表明, 放牧和模拟增温对高寒草地植物高度具有显著交互作用.在放牧条件下, 增温对植物高度无显著影响; 然而, 在不放牧条件下, 增温显著增加了植物高度. ...
... ).不同功能群植物高度、盖度及优势度的变化, 主要是受放牧过程中牲畜选择性采食的影响.可食性较高的禾草类和莎草类被家畜优先采食, 导致其高度、盖度及优势度降低, 而可食性较低的杂类草物种比例增加(段敏杰等, 2010; 何婷, 2015).这与本研究中放牧显著降低植物高度, 增加杂类草物种比例的结论一致.增温一方面满足植物生长的热量需求, 另一方面却加速土壤水分蒸发, 降低土壤水分可利用率, 影响群落的物种组成(Ganjurjav et al., 2016).本研究表明, 增温显著改变高寒草地植物群落特征, 植物高度增加82.3%, 盖度降低29.5%.OTC的阻挡作用使热量不易散失满足了植物生长发育的热量需求, 同时增温也改变了土壤无机氮的含量(Yu et al., 2014, 2019), 进而满足植物生长的个体发育, 抑制植物的分蘖繁殖(武倩等, 2016)导致盖度降低(Hopping et al., 2018).此外, OTC增温的同时加剧温室内土壤水分的蒸发, 降低其土壤湿度(Liu et al., 2018), 改变了温室内植物生长的微气候环境, 改变了植物对土壤水分和养分的吸收(Klein et al., 2005), 进而影响植物群落组成.草地生态系统作为陆地生态系统的重要组成部分, 受气候变化和人类活动的双重影响.本研究表明, 放牧和模拟增温对高寒草地植物高度具有显著交互作用.在放牧条件下, 增温对植物高度无显著影响; 然而, 在不放牧条件下, 增温显著增加了植物高度. ...
... 放牧和增温会引起草地植物群落结构与物种多样性的改变, 进而对植物生产力产生重要影响(Wang et al., 2012; Hopping et al., 2018; Liu et al., 2018).植物生产力与物种多样性密切相关, 较高的物种多样性可以维持较高的植物生产力, 同时也可以加强群落生态系统的稳定性(Tilman et al., 2001; 马文静等, 2013).本研究表明, 由于放牧具有补偿效应(McNaughton, 1979), 在物种多样性增高2%时, 地上净初级生产力增加29%.通过对牲畜采食量分析后发现, 2年间杂草采食量仅占总量的21.8%, 而禾草采食量高达78.2%, 与前人的研究成果一致(段敏杰等, 2010; 郑伟等, 2012; 干珠扎布等, 2013).本研究中, 增温对净初级生产力无显著影响, 但2016和2017年增温对净初级生产力的影响存在年际差异(图6B), 这可能是由于2017年降水量高于2016年造成的(图1, 图3C).Fu等(2018)在藏北高原高寒草地的研究也表明, 年际间的降水差异是引起增温对生产力年际差异性影响的主要原因.此外, 有研究表明, 在不同的空气温度背景下, 增温对草地净初级生产力的影响有所不同(Liu et al., 2018).因此, 两年间的温度差异也可能引起生产力对增温的差异性响应. ...
Increased precipitation has stronger effects on plant production of an alpine meadow than does experimental warming in the Northern Tibetan Plateau 1 2018
... 放牧和增温会引起草地植物群落结构与物种多样性的改变, 进而对植物生产力产生重要影响(Wang et al., 2012; Hopping et al., 2018; Liu et al., 2018).植物生产力与物种多样性密切相关, 较高的物种多样性可以维持较高的植物生产力, 同时也可以加强群落生态系统的稳定性(Tilman et al., 2001; 马文静等, 2013).本研究表明, 由于放牧具有补偿效应(McNaughton, 1979), 在物种多样性增高2%时, 地上净初级生产力增加29%.通过对牲畜采食量分析后发现, 2年间杂草采食量仅占总量的21.8%, 而禾草采食量高达78.2%, 与前人的研究成果一致(段敏杰等, 2010; 郑伟等, 2012; 干珠扎布等, 2013).本研究中, 增温对净初级生产力无显著影响, 但2016和2017年增温对净初级生产力的影响存在年际差异(图6B), 这可能是由于2017年降水量高于2016年造成的(图1, 图3C).Fu等(2018)在藏北高原高寒草地的研究也表明, 年际间的降水差异是引起增温对生产力年际差异性影响的主要原因.此外, 有研究表明, 在不同的空气温度背景下, 增温对草地净初级生产力的影响有所不同(Liu et al., 2018).因此, 两年间的温度差异也可能引起生产力对增温的差异性响应. ...
Clipping alters the response of biomass production to experimental warming: A case study in an alpine meadow on the Tibetan Plateau, China 1 2015
... 藏北高原位于青藏高原的腹地, 被称为“世界屋脊的屋脊”.该地区拥有42.1万km2高寒草地(高清竹等, 2006), 是西藏自治区主要的畜牧业生产基地和我国重要的生态屏障区, 是全球气候变化的启动区和敏感区, 是亚洲中部高寒环境中最为典型的一种生态系统.据报道, 青藏高原增温幅度高达每10年0.2 ℃ (Chen et al., 2013), 至2030年该地区将升温2.2-2.6 ℃ (National Development and Reform Commisssion, 2007), 显著高于全球和全国平均水平.由于过度放牧和草地不合理利用, 藏北高原草地退化面积达到58.2%, 重度退化和极重度退化草地面积分别占19.0%和6.5% (曹旭娟等, 2016).近年来, 研究人员分别开展了增温和放牧对高寒草地植物生产力和物种多样性影响的研究(段敏杰等, 2010; Ganjurjav et al., 2016; Liu et al., 2018), 但增温和放牧的交互作用对物种多样性和生产力影响的研究鲜有报道.一些****通过刈割模拟放牧活动, 从而研究增温和放牧的交互作用对高寒草地群落结构及生产力的影响(Wang et al., 2012; Fu et al., 2015).然而, 上述研究在很大程度上忽略了牲畜的践踏和采食作用对草地植物群落的影响.此外, 放牧与刈割的不同之处还在于对物种组成的影响, 放牧条件下牲畜对草地植物的采食具有物种间的选择喜好, 而刈割仅是对草地植物平均冠层高度进行统一收割(Klein et al., 2008; 段敏杰等, 2010; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
Experimental warming does not enhance gross primary production and above-ground biomass in the alpine meadow of Tibet 1 2013
... 本研究表明, 模拟增温和放牧对高寒草地植物净初级生产力具有显著交互作用.在放牧条件下, 增温对净初级生产力的影响存在年际差异, 2016年增温对生产力无显著影响, 2017年增温显著降低植物净初级生产力.增温显著增加高寒草地植物的高度, 但也加剧高寒草地土壤水分的蒸发速率, 降低其生态系统水分可利用率(Niu et al., 2011; Ganjurjav et al., 2016), 从而降低高寒草地植物生产力(Fu et al., 2013).总体而言, 增温对草地生产力无显著影响(图7B), 这与Liu等(2018)在青藏高原的研究结果相似.除采食作用外, 牲畜的践踏行为也是造成草地生产力急剧减少的关键因子(干珠扎布等, 2013).牲畜践踏不仅直接影响地上植物的生长发育,改变植物群落结构,加剧物种间竞争, 还会引起土壤结构改变和养分失衡(Kurz et al., 2006), 如加剧土壤板结,容重加大,降低土壤通透性等, 导致土壤含水量下降, 进而调控土壤有机质分解和养分转化(干珠扎布等, 2013; 何婷, 2015).由此可知, 放牧降低生产力对增温的响应程度(Wang et al., 2012), 两者具有显著交互作用. ...
Differential response of alpine steppe and alpine meadow to climate warming in the central Qinghai-Tibetan Plateau 5 2016
... 全球气候变化已成为世界各国政府和公众关注的焦点.IPCC第五次评估报告指出, 1880-2012年间全球地表平均温度上升0.85 ℃, 预计到21世纪末全球大气温度将增加1.5-4.8 ℃ (IPCC, 2013).草地是陆地生态系统的重要组成部分, 且极为脆弱, 对气候变化极其敏感.气候变化直接或间接地影响草地植物的群落结构与组成(Polley et al., 2014; Ganjurjav et al., 2016)、物种多样性及生产力(Wu et al., 2011; Wang et al., 2012).除气候变化外, 人类活动也是影响草地植物群落的重要因子.有研究认为, 人类活动对植物的影响大于环境因子, 甚至成为控制植物群落多样性的决定因素(Brown et al., 1997; Wang et al., 2006).放牧是草地生态系统最为重要的人类干扰因素, 植物物种多样性与放牧强度显著相关(段敏杰等, 2010; 郑伟等, 2012).放牧作为草地最普遍的利用方式, 牲畜通过采食和践踏行为对草地植物群落特征产生影响, 同时也会改变局地光、水和土壤养分利用率, 从而间接改变群落物种结构与分布(Zhou et al., 2007; Zhao et al., 2008). ...
... 藏北高原位于青藏高原的腹地, 被称为“世界屋脊的屋脊”.该地区拥有42.1万km2高寒草地(高清竹等, 2006), 是西藏自治区主要的畜牧业生产基地和我国重要的生态屏障区, 是全球气候变化的启动区和敏感区, 是亚洲中部高寒环境中最为典型的一种生态系统.据报道, 青藏高原增温幅度高达每10年0.2 ℃ (Chen et al., 2013), 至2030年该地区将升温2.2-2.6 ℃ (National Development and Reform Commisssion, 2007), 显著高于全球和全国平均水平.由于过度放牧和草地不合理利用, 藏北高原草地退化面积达到58.2%, 重度退化和极重度退化草地面积分别占19.0%和6.5% (曹旭娟等, 2016).近年来, 研究人员分别开展了增温和放牧对高寒草地植物生产力和物种多样性影响的研究(段敏杰等, 2010; Ganjurjav et al., 2016; Liu et al., 2018), 但增温和放牧的交互作用对物种多样性和生产力影响的研究鲜有报道.一些****通过刈割模拟放牧活动, 从而研究增温和放牧的交互作用对高寒草地群落结构及生产力的影响(Wang et al., 2012; Fu et al., 2015).然而, 上述研究在很大程度上忽略了牲畜的践踏和采食作用对草地植物群落的影响.此外, 放牧与刈割的不同之处还在于对物种组成的影响, 放牧条件下牲畜对草地植物的采食具有物种间的选择喜好, 而刈割仅是对草地植物平均冠层高度进行统一收割(Klein et al., 2008; 段敏杰等, 2010; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
... 放牧和增温对高寒草地植物群落结构具有重要影响(段敏杰等, 2010; Ganjurjav et al., 2016).不同功能群植物高度、盖度及优势度的变化, 主要是受放牧过程中牲畜选择性采食的影响.可食性较高的禾草类和莎草类被家畜优先采食, 导致其高度、盖度及优势度降低, 而可食性较低的杂类草物种比例增加(段敏杰等, 2010; 何婷, 2015).这与本研究中放牧显著降低植物高度, 增加杂类草物种比例的结论一致.增温一方面满足植物生长的热量需求, 另一方面却加速土壤水分蒸发, 降低土壤水分可利用率, 影响群落的物种组成(Ganjurjav et al., 2016).本研究表明, 增温显著改变高寒草地植物群落特征, 植物高度增加82.3%, 盖度降低29.5%.OTC的阻挡作用使热量不易散失满足了植物生长发育的热量需求, 同时增温也改变了土壤无机氮的含量(Yu et al., 2014, 2019), 进而满足植物生长的个体发育, 抑制植物的分蘖繁殖(武倩等, 2016)导致盖度降低(Hopping et al., 2018).此外, OTC增温的同时加剧温室内土壤水分的蒸发, 降低其土壤湿度(Liu et al., 2018), 改变了温室内植物生长的微气候环境, 改变了植物对土壤水分和养分的吸收(Klein et al., 2005), 进而影响植物群落组成.草地生态系统作为陆地生态系统的重要组成部分, 受气候变化和人类活动的双重影响.本研究表明, 放牧和模拟增温对高寒草地植物高度具有显著交互作用.在放牧条件下, 增温对植物高度无显著影响; 然而, 在不放牧条件下, 增温显著增加了植物高度. ...
... ).这与本研究中放牧显著降低植物高度, 增加杂类草物种比例的结论一致.增温一方面满足植物生长的热量需求, 另一方面却加速土壤水分蒸发, 降低土壤水分可利用率, 影响群落的物种组成(Ganjurjav et al., 2016).本研究表明, 增温显著改变高寒草地植物群落特征, 植物高度增加82.3%, 盖度降低29.5%.OTC的阻挡作用使热量不易散失满足了植物生长发育的热量需求, 同时增温也改变了土壤无机氮的含量(Yu et al., 2014, 2019), 进而满足植物生长的个体发育, 抑制植物的分蘖繁殖(武倩等, 2016)导致盖度降低(Hopping et al., 2018).此外, OTC增温的同时加剧温室内土壤水分的蒸发, 降低其土壤湿度(Liu et al., 2018), 改变了温室内植物生长的微气候环境, 改变了植物对土壤水分和养分的吸收(Klein et al., 2005), 进而影响植物群落组成.草地生态系统作为陆地生态系统的重要组成部分, 受气候变化和人类活动的双重影响.本研究表明, 放牧和模拟增温对高寒草地植物高度具有显著交互作用.在放牧条件下, 增温对植物高度无显著影响; 然而, 在不放牧条件下, 增温显著增加了植物高度. ...
... 本研究表明, 模拟增温和放牧对高寒草地植物净初级生产力具有显著交互作用.在放牧条件下, 增温对净初级生产力的影响存在年际差异, 2016年增温对生产力无显著影响, 2017年增温显著降低植物净初级生产力.增温显著增加高寒草地植物的高度, 但也加剧高寒草地土壤水分的蒸发速率, 降低其生态系统水分可利用率(Niu et al., 2011; Ganjurjav et al., 2016), 从而降低高寒草地植物生产力(Fu et al., 2013).总体而言, 增温对草地生产力无显著影响(图7B), 这与Liu等(2018)在青藏高原的研究结果相似.除采食作用外, 牲畜的践踏行为也是造成草地生产力急剧减少的关键因子(干珠扎布等, 2013).牲畜践踏不仅直接影响地上植物的生长发育,改变植物群落结构,加剧物种间竞争, 还会引起土壤结构改变和养分失衡(Kurz et al., 2006), 如加剧土壤板结,容重加大,降低土壤通透性等, 导致土壤含水量下降, 进而调控土壤有机质分解和养分转化(干珠扎布等, 2013; 何婷, 2015).由此可知, 放牧降低生产力对增温的响应程度(Wang et al., 2012), 两者具有显著交互作用. ...
藏北紫花针茅高寒草原适宜放牧率研究 3 2013
... 放牧和增温会引起草地植物群落结构与物种多样性的改变, 进而对植物生产力产生重要影响(Wang et al., 2012; Hopping et al., 2018; Liu et al., 2018).植物生产力与物种多样性密切相关, 较高的物种多样性可以维持较高的植物生产力, 同时也可以加强群落生态系统的稳定性(Tilman et al., 2001; 马文静等, 2013).本研究表明, 由于放牧具有补偿效应(McNaughton, 1979), 在物种多样性增高2%时, 地上净初级生产力增加29%.通过对牲畜采食量分析后发现, 2年间杂草采食量仅占总量的21.8%, 而禾草采食量高达78.2%, 与前人的研究成果一致(段敏杰等, 2010; 郑伟等, 2012; 干珠扎布等, 2013).本研究中, 增温对净初级生产力无显著影响, 但2016和2017年增温对净初级生产力的影响存在年际差异(图6B), 这可能是由于2017年降水量高于2016年造成的(图1, 图3C).Fu等(2018)在藏北高原高寒草地的研究也表明, 年际间的降水差异是引起增温对生产力年际差异性影响的主要原因.此外, 有研究表明, 在不同的空气温度背景下, 增温对草地净初级生产力的影响有所不同(Liu et al., 2018).因此, 两年间的温度差异也可能引起生产力对增温的差异性响应. ...
... 本研究表明, 模拟增温和放牧对高寒草地植物净初级生产力具有显著交互作用.在放牧条件下, 增温对净初级生产力的影响存在年际差异, 2016年增温对生产力无显著影响, 2017年增温显著降低植物净初级生产力.增温显著增加高寒草地植物的高度, 但也加剧高寒草地土壤水分的蒸发速率, 降低其生态系统水分可利用率(Niu et al., 2011; Ganjurjav et al., 2016), 从而降低高寒草地植物生产力(Fu et al., 2013).总体而言, 增温对草地生产力无显著影响(图7B), 这与Liu等(2018)在青藏高原的研究结果相似.除采食作用外, 牲畜的践踏行为也是造成草地生产力急剧减少的关键因子(干珠扎布等, 2013).牲畜践踏不仅直接影响地上植物的生长发育,改变植物群落结构,加剧物种间竞争, 还会引起土壤结构改变和养分失衡(Kurz et al., 2006), 如加剧土壤板结,容重加大,降低土壤通透性等, 导致土壤含水量下降, 进而调控土壤有机质分解和养分转化(干珠扎布等, 2013; 何婷, 2015).由此可知, 放牧降低生产力对增温的响应程度(Wang et al., 2012), 两者具有显著交互作用. ...
... 藏北高原位于青藏高原的腹地, 被称为“世界屋脊的屋脊”.该地区拥有42.1万km2高寒草地(高清竹等, 2006), 是西藏自治区主要的畜牧业生产基地和我国重要的生态屏障区, 是全球气候变化的启动区和敏感区, 是亚洲中部高寒环境中最为典型的一种生态系统.据报道, 青藏高原增温幅度高达每10年0.2 ℃ (Chen et al., 2013), 至2030年该地区将升温2.2-2.6 ℃ (National Development and Reform Commisssion, 2007), 显著高于全球和全国平均水平.由于过度放牧和草地不合理利用, 藏北高原草地退化面积达到58.2%, 重度退化和极重度退化草地面积分别占19.0%和6.5% (曹旭娟等, 2016).近年来, 研究人员分别开展了增温和放牧对高寒草地植物生产力和物种多样性影响的研究(段敏杰等, 2010; Ganjurjav et al., 2016; Liu et al., 2018), 但增温和放牧的交互作用对物种多样性和生产力影响的研究鲜有报道.一些****通过刈割模拟放牧活动, 从而研究增温和放牧的交互作用对高寒草地群落结构及生产力的影响(Wang et al., 2012; Fu et al., 2015).然而, 上述研究在很大程度上忽略了牲畜的践踏和采食作用对草地植物群落的影响.此外, 放牧与刈割的不同之处还在于对物种组成的影响, 放牧条件下牲畜对草地植物的采食具有物种间的选择喜好, 而刈割仅是对草地植物平均冠层高度进行统一收割(Klein et al., 2008; 段敏杰等, 2010; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
1 2006
... 藏北高原位于青藏高原的腹地, 被称为“世界屋脊的屋脊”.该地区拥有42.1万km2高寒草地(高清竹等, 2006), 是西藏自治区主要的畜牧业生产基地和我国重要的生态屏障区, 是全球气候变化的启动区和敏感区, 是亚洲中部高寒环境中最为典型的一种生态系统.据报道, 青藏高原增温幅度高达每10年0.2 ℃ (Chen et al., 2013), 至2030年该地区将升温2.2-2.6 ℃ (National Development and Reform Commisssion, 2007), 显著高于全球和全国平均水平.由于过度放牧和草地不合理利用, 藏北高原草地退化面积达到58.2%, 重度退化和极重度退化草地面积分别占19.0%和6.5% (曹旭娟等, 2016).近年来, 研究人员分别开展了增温和放牧对高寒草地植物生产力和物种多样性影响的研究(段敏杰等, 2010; Ganjurjav et al., 2016; Liu et al., 2018), 但增温和放牧的交互作用对物种多样性和生产力影响的研究鲜有报道.一些****通过刈割模拟放牧活动, 从而研究增温和放牧的交互作用对高寒草地群落结构及生产力的影响(Wang et al., 2012; Fu et al., 2015).然而, 上述研究在很大程度上忽略了牲畜的践踏和采食作用对草地植物群落的影响.此外, 放牧与刈割的不同之处还在于对物种组成的影响, 放牧条件下牲畜对草地植物的采食具有物种间的选择喜好, 而刈割仅是对草地植物平均冠层高度进行统一收割(Klein et al., 2008; 段敏杰等, 2010; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
放牧率对川西北高寒草地植物多样性和生产力的影响 2 2015
... 放牧和增温对高寒草地植物群落结构具有重要影响(段敏杰等, 2010; Ganjurjav et al., 2016).不同功能群植物高度、盖度及优势度的变化, 主要是受放牧过程中牲畜选择性采食的影响.可食性较高的禾草类和莎草类被家畜优先采食, 导致其高度、盖度及优势度降低, 而可食性较低的杂类草物种比例增加(段敏杰等, 2010; 何婷, 2015).这与本研究中放牧显著降低植物高度, 增加杂类草物种比例的结论一致.增温一方面满足植物生长的热量需求, 另一方面却加速土壤水分蒸发, 降低土壤水分可利用率, 影响群落的物种组成(Ganjurjav et al., 2016).本研究表明, 增温显著改变高寒草地植物群落特征, 植物高度增加82.3%, 盖度降低29.5%.OTC的阻挡作用使热量不易散失满足了植物生长发育的热量需求, 同时增温也改变了土壤无机氮的含量(Yu et al., 2014, 2019), 进而满足植物生长的个体发育, 抑制植物的分蘖繁殖(武倩等, 2016)导致盖度降低(Hopping et al., 2018).此外, OTC增温的同时加剧温室内土壤水分的蒸发, 降低其土壤湿度(Liu et al., 2018), 改变了温室内植物生长的微气候环境, 改变了植物对土壤水分和养分的吸收(Klein et al., 2005), 进而影响植物群落组成.草地生态系统作为陆地生态系统的重要组成部分, 受气候变化和人类活动的双重影响.本研究表明, 放牧和模拟增温对高寒草地植物高度具有显著交互作用.在放牧条件下, 增温对植物高度无显著影响; 然而, 在不放牧条件下, 增温显著增加了植物高度. ...
... 本研究表明, 模拟增温和放牧对高寒草地植物净初级生产力具有显著交互作用.在放牧条件下, 增温对净初级生产力的影响存在年际差异, 2016年增温对生产力无显著影响, 2017年增温显著降低植物净初级生产力.增温显著增加高寒草地植物的高度, 但也加剧高寒草地土壤水分的蒸发速率, 降低其生态系统水分可利用率(Niu et al., 2011; Ganjurjav et al., 2016), 从而降低高寒草地植物生产力(Fu et al., 2013).总体而言, 增温对草地生产力无显著影响(图7B), 这与Liu等(2018)在青藏高原的研究结果相似.除采食作用外, 牲畜的践踏行为也是造成草地生产力急剧减少的关键因子(干珠扎布等, 2013).牲畜践踏不仅直接影响地上植物的生长发育,改变植物群落结构,加剧物种间竞争, 还会引起土壤结构改变和养分失衡(Kurz et al., 2006), 如加剧土壤板结,容重加大,降低土壤通透性等, 导致土壤含水量下降, 进而调控土壤有机质分解和养分转化(干珠扎布等, 2013; 何婷, 2015).由此可知, 放牧降低生产力对增温的响应程度(Wang et al., 2012), 两者具有显著交互作用. ...
放牧率对川西北高寒草地植物多样性和生产力的影响 2 2015
... 放牧和增温对高寒草地植物群落结构具有重要影响(段敏杰等, 2010; Ganjurjav et al., 2016).不同功能群植物高度、盖度及优势度的变化, 主要是受放牧过程中牲畜选择性采食的影响.可食性较高的禾草类和莎草类被家畜优先采食, 导致其高度、盖度及优势度降低, 而可食性较低的杂类草物种比例增加(段敏杰等, 2010; 何婷, 2015).这与本研究中放牧显著降低植物高度, 增加杂类草物种比例的结论一致.增温一方面满足植物生长的热量需求, 另一方面却加速土壤水分蒸发, 降低土壤水分可利用率, 影响群落的物种组成(Ganjurjav et al., 2016).本研究表明, 增温显著改变高寒草地植物群落特征, 植物高度增加82.3%, 盖度降低29.5%.OTC的阻挡作用使热量不易散失满足了植物生长发育的热量需求, 同时增温也改变了土壤无机氮的含量(Yu et al., 2014, 2019), 进而满足植物生长的个体发育, 抑制植物的分蘖繁殖(武倩等, 2016)导致盖度降低(Hopping et al., 2018).此外, OTC增温的同时加剧温室内土壤水分的蒸发, 降低其土壤湿度(Liu et al., 2018), 改变了温室内植物生长的微气候环境, 改变了植物对土壤水分和养分的吸收(Klein et al., 2005), 进而影响植物群落组成.草地生态系统作为陆地生态系统的重要组成部分, 受气候变化和人类活动的双重影响.本研究表明, 放牧和模拟增温对高寒草地植物高度具有显著交互作用.在放牧条件下, 增温对植物高度无显著影响; 然而, 在不放牧条件下, 增温显著增加了植物高度. ...
... 本研究表明, 模拟增温和放牧对高寒草地植物净初级生产力具有显著交互作用.在放牧条件下, 增温对净初级生产力的影响存在年际差异, 2016年增温对生产力无显著影响, 2017年增温显著降低植物净初级生产力.增温显著增加高寒草地植物的高度, 但也加剧高寒草地土壤水分的蒸发速率, 降低其生态系统水分可利用率(Niu et al., 2011; Ganjurjav et al., 2016), 从而降低高寒草地植物生产力(Fu et al., 2013).总体而言, 增温对草地生产力无显著影响(图7B), 这与Liu等(2018)在青藏高原的研究结果相似.除采食作用外, 牲畜的践踏行为也是造成草地生产力急剧减少的关键因子(干珠扎布等, 2013).牲畜践踏不仅直接影响地上植物的生长发育,改变植物群落结构,加剧物种间竞争, 还会引起土壤结构改变和养分失衡(Kurz et al., 2006), 如加剧土壤板结,容重加大,降低土壤通透性等, 导致土壤含水量下降, 进而调控土壤有机质分解和养分转化(干珠扎布等, 2013; 何婷, 2015).由此可知, 放牧降低生产力对增温的响应程度(Wang et al., 2012), 两者具有显著交互作用. ...
Warming and land use change concurrently erode ecosystem services in Tibet 2 2018
... 放牧和增温对高寒草地植物群落结构具有重要影响(段敏杰等, 2010; Ganjurjav et al., 2016).不同功能群植物高度、盖度及优势度的变化, 主要是受放牧过程中牲畜选择性采食的影响.可食性较高的禾草类和莎草类被家畜优先采食, 导致其高度、盖度及优势度降低, 而可食性较低的杂类草物种比例增加(段敏杰等, 2010; 何婷, 2015).这与本研究中放牧显著降低植物高度, 增加杂类草物种比例的结论一致.增温一方面满足植物生长的热量需求, 另一方面却加速土壤水分蒸发, 降低土壤水分可利用率, 影响群落的物种组成(Ganjurjav et al., 2016).本研究表明, 增温显著改变高寒草地植物群落特征, 植物高度增加82.3%, 盖度降低29.5%.OTC的阻挡作用使热量不易散失满足了植物生长发育的热量需求, 同时增温也改变了土壤无机氮的含量(Yu et al., 2014, 2019), 进而满足植物生长的个体发育, 抑制植物的分蘖繁殖(武倩等, 2016)导致盖度降低(Hopping et al., 2018).此外, OTC增温的同时加剧温室内土壤水分的蒸发, 降低其土壤湿度(Liu et al., 2018), 改变了温室内植物生长的微气候环境, 改变了植物对土壤水分和养分的吸收(Klein et al., 2005), 进而影响植物群落组成.草地生态系统作为陆地生态系统的重要组成部分, 受气候变化和人类活动的双重影响.本研究表明, 放牧和模拟增温对高寒草地植物高度具有显著交互作用.在放牧条件下, 增温对植物高度无显著影响; 然而, 在不放牧条件下, 增温显著增加了植物高度. ...
... 放牧和增温会引起草地植物群落结构与物种多样性的改变, 进而对植物生产力产生重要影响(Wang et al., 2012; Hopping et al., 2018; Liu et al., 2018).植物生产力与物种多样性密切相关, 较高的物种多样性可以维持较高的植物生产力, 同时也可以加强群落生态系统的稳定性(Tilman et al., 2001; 马文静等, 2013).本研究表明, 由于放牧具有补偿效应(McNaughton, 1979), 在物种多样性增高2%时, 地上净初级生产力增加29%.通过对牲畜采食量分析后发现, 2年间杂草采食量仅占总量的21.8%, 而禾草采食量高达78.2%, 与前人的研究成果一致(段敏杰等, 2010; 郑伟等, 2012; 干珠扎布等, 2013).本研究中, 增温对净初级生产力无显著影响, 但2016和2017年增温对净初级生产力的影响存在年际差异(图6B), 这可能是由于2017年降水量高于2016年造成的(图1, 图3C).Fu等(2018)在藏北高原高寒草地的研究也表明, 年际间的降水差异是引起增温对生产力年际差异性影响的主要原因.此外, 有研究表明, 在不同的空气温度背景下, 增温对草地净初级生产力的影响有所不同(Liu et al., 2018).因此, 两年间的温度差异也可能引起生产力对增温的差异性响应. ...
1
... 全球气候变化已成为世界各国政府和公众关注的焦点.IPCC第五次评估报告指出, 1880-2012年间全球地表平均温度上升0.85 ℃, 预计到21世纪末全球大气温度将增加1.5-4.8 ℃ (IPCC, 2013).草地是陆地生态系统的重要组成部分, 且极为脆弱, 对气候变化极其敏感.气候变化直接或间接地影响草地植物的群落结构与组成(Polley et al., 2014; Ganjurjav et al., 2016)、物种多样性及生产力(Wu et al., 2011; Wang et al., 2012).除气候变化外, 人类活动也是影响草地植物群落的重要因子.有研究认为, 人类活动对植物的影响大于环境因子, 甚至成为控制植物群落多样性的决定因素(Brown et al., 1997; Wang et al., 2006).放牧是草地生态系统最为重要的人类干扰因素, 植物物种多样性与放牧强度显著相关(段敏杰等, 2010; 郑伟等, 2012).放牧作为草地最普遍的利用方式, 牲畜通过采食和践踏行为对草地植物群落特征产生影响, 同时也会改变局地光、水和土壤养分利用率, 从而间接改变群落物种结构与分布(Zhou et al., 2007; Zhao et al., 2008). ...
Grazing as an optimization process: Grass-ungulate relationships in the Serengeti 1 1979
... 放牧和增温会引起草地植物群落结构与物种多样性的改变, 进而对植物生产力产生重要影响(Wang et al., 2012; Hopping et al., 2018; Liu et al., 2018).植物生产力与物种多样性密切相关, 较高的物种多样性可以维持较高的植物生产力, 同时也可以加强群落生态系统的稳定性(Tilman et al., 2001; 马文静等, 2013).本研究表明, 由于放牧具有补偿效应(McNaughton, 1979), 在物种多样性增高2%时, 地上净初级生产力增加29%.通过对牲畜采食量分析后发现, 2年间杂草采食量仅占总量的21.8%, 而禾草采食量高达78.2%, 与前人的研究成果一致(段敏杰等, 2010; 郑伟等, 2012; 干珠扎布等, 2013).本研究中, 增温对净初级生产力无显著影响, 但2016和2017年增温对净初级生产力的影响存在年际差异(图6B), 这可能是由于2017年降水量高于2016年造成的(图1, 图3C).Fu等(2018)在藏北高原高寒草地的研究也表明, 年际间的降水差异是引起增温对生产力年际差异性影响的主要原因.此外, 有研究表明, 在不同的空气温度背景下, 增温对草地净初级生产力的影响有所不同(Liu et al., 2018).因此, 两年间的温度差异也可能引起生产力对增温的差异性响应. ...
Water-use efficiency in response to climate change: From leaf to ecosystem in a temperate steppe 1 2011
... 本研究表明, 模拟增温和放牧对高寒草地植物净初级生产力具有显著交互作用.在放牧条件下, 增温对净初级生产力的影响存在年际差异, 2016年增温对生产力无显著影响, 2017年增温显著降低植物净初级生产力.增温显著增加高寒草地植物的高度, 但也加剧高寒草地土壤水分的蒸发速率, 降低其生态系统水分可利用率(Niu et al., 2011; Ganjurjav et al., 2016), 从而降低高寒草地植物生产力(Fu et al., 2013).总体而言, 增温对草地生产力无显著影响(图7B), 这与Liu等(2018)在青藏高原的研究结果相似.除采食作用外, 牲畜的践踏行为也是造成草地生产力急剧减少的关键因子(干珠扎布等, 2013).牲畜践踏不仅直接影响地上植物的生长发育,改变植物群落结构,加剧物种间竞争, 还会引起土壤结构改变和养分失衡(Kurz et al., 2006), 如加剧土壤板结,容重加大,降低土壤通透性等, 导致土壤含水量下降, 进而调控土壤有机质分解和养分转化(干珠扎布等, 2013; 何婷, 2015).由此可知, 放牧降低生产力对增温的响应程度(Wang et al., 2012), 两者具有显著交互作用. ...
Impacts of climate change drivers on C4 grassland productivity: Scaling driver effects through the plant community 1 2014
... 全球气候变化已成为世界各国政府和公众关注的焦点.IPCC第五次评估报告指出, 1880-2012年间全球地表平均温度上升0.85 ℃, 预计到21世纪末全球大气温度将增加1.5-4.8 ℃ (IPCC, 2013).草地是陆地生态系统的重要组成部分, 且极为脆弱, 对气候变化极其敏感.气候变化直接或间接地影响草地植物的群落结构与组成(Polley et al., 2014; Ganjurjav et al., 2016)、物种多样性及生产力(Wu et al., 2011; Wang et al., 2012).除气候变化外, 人类活动也是影响草地植物群落的重要因子.有研究认为, 人类活动对植物的影响大于环境因子, 甚至成为控制植物群落多样性的决定因素(Brown et al., 1997; Wang et al., 2006).放牧是草地生态系统最为重要的人类干扰因素, 植物物种多样性与放牧强度显著相关(段敏杰等, 2010; 郑伟等, 2012).放牧作为草地最普遍的利用方式, 牲畜通过采食和践踏行为对草地植物群落特征产生影响, 同时也会改变局地光、水和土壤养分利用率, 从而间接改变群落物种结构与分布(Zhou et al., 2007; Zhao et al., 2008). ...
Diversity and productivity in a long-term grassland experiment 1 2001
... 放牧和增温会引起草地植物群落结构与物种多样性的改变, 进而对植物生产力产生重要影响(Wang et al., 2012; Hopping et al., 2018; Liu et al., 2018).植物生产力与物种多样性密切相关, 较高的物种多样性可以维持较高的植物生产力, 同时也可以加强群落生态系统的稳定性(Tilman et al., 2001; 马文静等, 2013).本研究表明, 由于放牧具有补偿效应(McNaughton, 1979), 在物种多样性增高2%时, 地上净初级生产力增加29%.通过对牲畜采食量分析后发现, 2年间杂草采食量仅占总量的21.8%, 而禾草采食量高达78.2%, 与前人的研究成果一致(段敏杰等, 2010; 郑伟等, 2012; 干珠扎布等, 2013).本研究中, 增温对净初级生产力无显著影响, 但2016和2017年增温对净初级生产力的影响存在年际差异(图6B), 这可能是由于2017年降水量高于2016年造成的(图1, 图3C).Fu等(2018)在藏北高原高寒草地的研究也表明, 年际间的降水差异是引起增温对生产力年际差异性影响的主要原因.此外, 有研究表明, 在不同的空气温度背景下, 增温对草地净初级生产力的影响有所不同(Liu et al., 2018).因此, 两年间的温度差异也可能引起生产力对增温的差异性响应. ...
Effects of warming and grazing on soil N availability, species composition, and ANPP in an alpine meadow 5 2012
... 全球气候变化已成为世界各国政府和公众关注的焦点.IPCC第五次评估报告指出, 1880-2012年间全球地表平均温度上升0.85 ℃, 预计到21世纪末全球大气温度将增加1.5-4.8 ℃ (IPCC, 2013).草地是陆地生态系统的重要组成部分, 且极为脆弱, 对气候变化极其敏感.气候变化直接或间接地影响草地植物的群落结构与组成(Polley et al., 2014; Ganjurjav et al., 2016)、物种多样性及生产力(Wu et al., 2011; Wang et al., 2012).除气候变化外, 人类活动也是影响草地植物群落的重要因子.有研究认为, 人类活动对植物的影响大于环境因子, 甚至成为控制植物群落多样性的决定因素(Brown et al., 1997; Wang et al., 2006).放牧是草地生态系统最为重要的人类干扰因素, 植物物种多样性与放牧强度显著相关(段敏杰等, 2010; 郑伟等, 2012).放牧作为草地最普遍的利用方式, 牲畜通过采食和践踏行为对草地植物群落特征产生影响, 同时也会改变局地光、水和土壤养分利用率, 从而间接改变群落物种结构与分布(Zhou et al., 2007; Zhao et al., 2008). ...
... 藏北高原位于青藏高原的腹地, 被称为“世界屋脊的屋脊”.该地区拥有42.1万km2高寒草地(高清竹等, 2006), 是西藏自治区主要的畜牧业生产基地和我国重要的生态屏障区, 是全球气候变化的启动区和敏感区, 是亚洲中部高寒环境中最为典型的一种生态系统.据报道, 青藏高原增温幅度高达每10年0.2 ℃ (Chen et al., 2013), 至2030年该地区将升温2.2-2.6 ℃ (National Development and Reform Commisssion, 2007), 显著高于全球和全国平均水平.由于过度放牧和草地不合理利用, 藏北高原草地退化面积达到58.2%, 重度退化和极重度退化草地面积分别占19.0%和6.5% (曹旭娟等, 2016).近年来, 研究人员分别开展了增温和放牧对高寒草地植物生产力和物种多样性影响的研究(段敏杰等, 2010; Ganjurjav et al., 2016; Liu et al., 2018), 但增温和放牧的交互作用对物种多样性和生产力影响的研究鲜有报道.一些****通过刈割模拟放牧活动, 从而研究增温和放牧的交互作用对高寒草地群落结构及生产力的影响(Wang et al., 2012; Fu et al., 2015).然而, 上述研究在很大程度上忽略了牲畜的践踏和采食作用对草地植物群落的影响.此外, 放牧与刈割的不同之处还在于对物种组成的影响, 放牧条件下牲畜对草地植物的采食具有物种间的选择喜好, 而刈割仅是对草地植物平均冠层高度进行统一收割(Klein et al., 2008; 段敏杰等, 2010; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
... ; Wang et al., 2012).因此, 本文通过2年野外模拟增温、放牧及二者的交互作用试验对藏北高寒草地植物群落物种多样性及生产力开展研究, 探讨草地植物群落物种多样性与生产力对不同处理的响应机制, 旨在为藏北地区高寒草地合理利用、保护物种多样性及维持草地生态系统可持续健康发展提供理论依据. ...
... 放牧和增温会引起草地植物群落结构与物种多样性的改变, 进而对植物生产力产生重要影响(Wang et al., 2012; Hopping et al., 2018; Liu et al., 2018).植物生产力与物种多样性密切相关, 较高的物种多样性可以维持较高的植物生产力, 同时也可以加强群落生态系统的稳定性(Tilman et al., 2001; 马文静等, 2013).本研究表明, 由于放牧具有补偿效应(McNaughton, 1979), 在物种多样性增高2%时, 地上净初级生产力增加29%.通过对牲畜采食量分析后发现, 2年间杂草采食量仅占总量的21.8%, 而禾草采食量高达78.2%, 与前人的研究成果一致(段敏杰等, 2010; 郑伟等, 2012; 干珠扎布等, 2013).本研究中, 增温对净初级生产力无显著影响, 但2016和2017年增温对净初级生产力的影响存在年际差异(图6B), 这可能是由于2017年降水量高于2016年造成的(图1, 图3C).Fu等(2018)在藏北高原高寒草地的研究也表明, 年际间的降水差异是引起增温对生产力年际差异性影响的主要原因.此外, 有研究表明, 在不同的空气温度背景下, 增温对草地净初级生产力的影响有所不同(Liu et al., 2018).因此, 两年间的温度差异也可能引起生产力对增温的差异性响应. ...
... 本研究表明, 模拟增温和放牧对高寒草地植物净初级生产力具有显著交互作用.在放牧条件下, 增温对净初级生产力的影响存在年际差异, 2016年增温对生产力无显著影响, 2017年增温显著降低植物净初级生产力.增温显著增加高寒草地植物的高度, 但也加剧高寒草地土壤水分的蒸发速率, 降低其生态系统水分可利用率(Niu et al., 2011; Ganjurjav et al., 2016), 从而降低高寒草地植物生产力(Fu et al., 2013).总体而言, 增温对草地生产力无显著影响(图7B), 这与Liu等(2018)在青藏高原的研究结果相似.除采食作用外, 牲畜的践踏行为也是造成草地生产力急剧减少的关键因子(干珠扎布等, 2013).牲畜践踏不仅直接影响地上植物的生长发育,改变植物群落结构,加剧物种间竞争, 还会引起土壤结构改变和养分失衡(Kurz et al., 2006), 如加剧土壤板结,容重加大,降低土壤通透性等, 导致土壤含水量下降, 进而调控土壤有机质分解和养分转化(干珠扎布等, 2013; 何婷, 2015).由此可知, 放牧降低生产力对增温的响应程度(Wang et al., 2012), 两者具有显著交互作用. ...
The relative role of climatic and human factors in desertification in semiarid China 1 2006
... 全球气候变化已成为世界各国政府和公众关注的焦点.IPCC第五次评估报告指出, 1880-2012年间全球地表平均温度上升0.85 ℃, 预计到21世纪末全球大气温度将增加1.5-4.8 ℃ (IPCC, 2013).草地是陆地生态系统的重要组成部分, 且极为脆弱, 对气候变化极其敏感.气候变化直接或间接地影响草地植物的群落结构与组成(Polley et al., 2014; Ganjurjav et al., 2016)、物种多样性及生产力(Wu et al., 2011; Wang et al., 2012).除气候变化外, 人类活动也是影响草地植物群落的重要因子.有研究认为, 人类活动对植物的影响大于环境因子, 甚至成为控制植物群落多样性的决定因素(Brown et al., 1997; Wang et al., 2006).放牧是草地生态系统最为重要的人类干扰因素, 植物物种多样性与放牧强度显著相关(段敏杰等, 2010; 郑伟等, 2012).放牧作为草地最普遍的利用方式, 牲畜通过采食和践踏行为对草地植物群落特征产生影响, 同时也会改变局地光、水和土壤养分利用率, 从而间接改变群落物种结构与分布(Zhou et al., 2007; Zhao et al., 2008). ...
模拟增温对草地植物、土壤和生态系统碳交换的影响 1 2016
... 放牧和增温对高寒草地植物群落结构具有重要影响(段敏杰等, 2010; Ganjurjav et al., 2016).不同功能群植物高度、盖度及优势度的变化, 主要是受放牧过程中牲畜选择性采食的影响.可食性较高的禾草类和莎草类被家畜优先采食, 导致其高度、盖度及优势度降低, 而可食性较低的杂类草物种比例增加(段敏杰等, 2010; 何婷, 2015).这与本研究中放牧显著降低植物高度, 增加杂类草物种比例的结论一致.增温一方面满足植物生长的热量需求, 另一方面却加速土壤水分蒸发, 降低土壤水分可利用率, 影响群落的物种组成(Ganjurjav et al., 2016).本研究表明, 增温显著改变高寒草地植物群落特征, 植物高度增加82.3%, 盖度降低29.5%.OTC的阻挡作用使热量不易散失满足了植物生长发育的热量需求, 同时增温也改变了土壤无机氮的含量(Yu et al., 2014, 2019), 进而满足植物生长的个体发育, 抑制植物的分蘖繁殖(武倩等, 2016)导致盖度降低(Hopping et al., 2018).此外, OTC增温的同时加剧温室内土壤水分的蒸发, 降低其土壤湿度(Liu et al., 2018), 改变了温室内植物生长的微气候环境, 改变了植物对土壤水分和养分的吸收(Klein et al., 2005), 进而影响植物群落组成.草地生态系统作为陆地生态系统的重要组成部分, 受气候变化和人类活动的双重影响.本研究表明, 放牧和模拟增温对高寒草地植物高度具有显著交互作用.在放牧条件下, 增温对植物高度无显著影响; 然而, 在不放牧条件下, 增温显著增加了植物高度. ...
模拟增温对草地植物、土壤和生态系统碳交换的影响 1 2016
... 放牧和增温对高寒草地植物群落结构具有重要影响(段敏杰等, 2010; Ganjurjav et al., 2016).不同功能群植物高度、盖度及优势度的变化, 主要是受放牧过程中牲畜选择性采食的影响.可食性较高的禾草类和莎草类被家畜优先采食, 导致其高度、盖度及优势度降低, 而可食性较低的杂类草物种比例增加(段敏杰等, 2010; 何婷, 2015).这与本研究中放牧显著降低植物高度, 增加杂类草物种比例的结论一致.增温一方面满足植物生长的热量需求, 另一方面却加速土壤水分蒸发, 降低土壤水分可利用率, 影响群落的物种组成(Ganjurjav et al., 2016).本研究表明, 增温显著改变高寒草地植物群落特征, 植物高度增加82.3%, 盖度降低29.5%.OTC的阻挡作用使热量不易散失满足了植物生长发育的热量需求, 同时增温也改变了土壤无机氮的含量(Yu et al., 2014, 2019), 进而满足植物生长的个体发育, 抑制植物的分蘖繁殖(武倩等, 2016)导致盖度降低(Hopping et al., 2018).此外, OTC增温的同时加剧温室内土壤水分的蒸发, 降低其土壤湿度(Liu et al., 2018), 改变了温室内植物生长的微气候环境, 改变了植物对土壤水分和养分的吸收(Klein et al., 2005), 进而影响植物群落组成.草地生态系统作为陆地生态系统的重要组成部分, 受气候变化和人类活动的双重影响.本研究表明, 放牧和模拟增温对高寒草地植物高度具有显著交互作用.在放牧条件下, 增温对植物高度无显著影响; 然而, 在不放牧条件下, 增温显著增加了植物高度. ...
Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation 1 2011
... 全球气候变化已成为世界各国政府和公众关注的焦点.IPCC第五次评估报告指出, 1880-2012年间全球地表平均温度上升0.85 ℃, 预计到21世纪末全球大气温度将增加1.5-4.8 ℃ (IPCC, 2013).草地是陆地生态系统的重要组成部分, 且极为脆弱, 对气候变化极其敏感.气候变化直接或间接地影响草地植物的群落结构与组成(Polley et al., 2014; Ganjurjav et al., 2016)、物种多样性及生产力(Wu et al., 2011; Wang et al., 2012).除气候变化外, 人类活动也是影响草地植物群落的重要因子.有研究认为, 人类活动对植物的影响大于环境因子, 甚至成为控制植物群落多样性的决定因素(Brown et al., 1997; Wang et al., 2006).放牧是草地生态系统最为重要的人类干扰因素, 植物物种多样性与放牧强度显著相关(段敏杰等, 2010; 郑伟等, 2012).放牧作为草地最普遍的利用方式, 牲畜通过采食和践踏行为对草地植物群落特征产生影响, 同时也会改变局地光、水和土壤养分利用率, 从而间接改变群落物种结构与分布(Zhou et al., 2007; Zhao et al., 2008). ...
Effects of 7 years experimental warming on soil bacterial and fungal community structure in the Northern Tibet alpine meadow at three elevations 1 2019
... 放牧和增温对高寒草地植物群落结构具有重要影响(段敏杰等, 2010; Ganjurjav et al., 2016).不同功能群植物高度、盖度及优势度的变化, 主要是受放牧过程中牲畜选择性采食的影响.可食性较高的禾草类和莎草类被家畜优先采食, 导致其高度、盖度及优势度降低, 而可食性较低的杂类草物种比例增加(段敏杰等, 2010; 何婷, 2015).这与本研究中放牧显著降低植物高度, 增加杂类草物种比例的结论一致.增温一方面满足植物生长的热量需求, 另一方面却加速土壤水分蒸发, 降低土壤水分可利用率, 影响群落的物种组成(Ganjurjav et al., 2016).本研究表明, 增温显著改变高寒草地植物群落特征, 植物高度增加82.3%, 盖度降低29.5%.OTC的阻挡作用使热量不易散失满足了植物生长发育的热量需求, 同时增温也改变了土壤无机氮的含量(Yu et al., 2014, 2019), 进而满足植物生长的个体发育, 抑制植物的分蘖繁殖(武倩等, 2016)导致盖度降低(Hopping et al., 2018).此外, OTC增温的同时加剧温室内土壤水分的蒸发, 降低其土壤湿度(Liu et al., 2018), 改变了温室内植物生长的微气候环境, 改变了植物对土壤水分和养分的吸收(Klein et al., 2005), 进而影响植物群落组成.草地生态系统作为陆地生态系统的重要组成部分, 受气候变化和人类活动的双重影响.本研究表明, 放牧和模拟增温对高寒草地植物高度具有显著交互作用.在放牧条件下, 增温对植物高度无显著影响; 然而, 在不放牧条件下, 增温显著增加了植物高度. ...
2014). Response of soil C and N, dissolved organic C and N, and inorganic N to short-term experimental warming in an alpine meadow on the Tibetan Plateau 1 2014
... 放牧和增温对高寒草地植物群落结构具有重要影响(段敏杰等, 2010; Ganjurjav et al., 2016).不同功能群植物高度、盖度及优势度的变化, 主要是受放牧过程中牲畜选择性采食的影响.可食性较高的禾草类和莎草类被家畜优先采食, 导致其高度、盖度及优势度降低, 而可食性较低的杂类草物种比例增加(段敏杰等, 2010; 何婷, 2015).这与本研究中放牧显著降低植物高度, 增加杂类草物种比例的结论一致.增温一方面满足植物生长的热量需求, 另一方面却加速土壤水分蒸发, 降低土壤水分可利用率, 影响群落的物种组成(Ganjurjav et al., 2016).本研究表明, 增温显著改变高寒草地植物群落特征, 植物高度增加82.3%, 盖度降低29.5%.OTC的阻挡作用使热量不易散失满足了植物生长发育的热量需求, 同时增温也改变了土壤无机氮的含量(Yu et al., 2014, 2019), 进而满足植物生长的个体发育, 抑制植物的分蘖繁殖(武倩等, 2016)导致盖度降低(Hopping et al., 2018).此外, OTC增温的同时加剧温室内土壤水分的蒸发, 降低其土壤湿度(Liu et al., 2018), 改变了温室内植物生长的微气候环境, 改变了植物对土壤水分和养分的吸收(Klein et al., 2005), 进而影响植物群落组成.草地生态系统作为陆地生态系统的重要组成部分, 受气候变化和人类活动的双重影响.本研究表明, 放牧和模拟增温对高寒草地植物高度具有显著交互作用.在放牧条件下, 增温对植物高度无显著影响; 然而, 在不放牧条件下, 增温显著增加了植物高度. ...
Compensatory growth responses to clipping defoliation in Leymus chinensis (Poaceae) under nutrient addition and water deficiency conditions 1 2008
... 全球气候变化已成为世界各国政府和公众关注的焦点.IPCC第五次评估报告指出, 1880-2012年间全球地表平均温度上升0.85 ℃, 预计到21世纪末全球大气温度将增加1.5-4.8 ℃ (IPCC, 2013).草地是陆地生态系统的重要组成部分, 且极为脆弱, 对气候变化极其敏感.气候变化直接或间接地影响草地植物的群落结构与组成(Polley et al., 2014; Ganjurjav et al., 2016)、物种多样性及生产力(Wu et al., 2011; Wang et al., 2012).除气候变化外, 人类活动也是影响草地植物群落的重要因子.有研究认为, 人类活动对植物的影响大于环境因子, 甚至成为控制植物群落多样性的决定因素(Brown et al., 1997; Wang et al., 2006).放牧是草地生态系统最为重要的人类干扰因素, 植物物种多样性与放牧强度显著相关(段敏杰等, 2010; 郑伟等, 2012).放牧作为草地最普遍的利用方式, 牲畜通过采食和践踏行为对草地植物群落特征产生影响, 同时也会改变局地光、水和土壤养分利用率, 从而间接改变群落物种结构与分布(Zhou et al., 2007; Zhao et al., 2008). ...
放牧强度对环青海湖高寒草原群落物种多样性和生产力的影响 4 2012
... 全球气候变化已成为世界各国政府和公众关注的焦点.IPCC第五次评估报告指出, 1880-2012年间全球地表平均温度上升0.85 ℃, 预计到21世纪末全球大气温度将增加1.5-4.8 ℃ (IPCC, 2013).草地是陆地生态系统的重要组成部分, 且极为脆弱, 对气候变化极其敏感.气候变化直接或间接地影响草地植物的群落结构与组成(Polley et al., 2014; Ganjurjav et al., 2016)、物种多样性及生产力(Wu et al., 2011; Wang et al., 2012).除气候变化外, 人类活动也是影响草地植物群落的重要因子.有研究认为, 人类活动对植物的影响大于环境因子, 甚至成为控制植物群落多样性的决定因素(Brown et al., 1997; Wang et al., 2006).放牧是草地生态系统最为重要的人类干扰因素, 植物物种多样性与放牧强度显著相关(段敏杰等, 2010; 郑伟等, 2012).放牧作为草地最普遍的利用方式, 牲畜通过采食和践踏行为对草地植物群落特征产生影响, 同时也会改变局地光、水和土壤养分利用率, 从而间接改变群落物种结构与分布(Zhou et al., 2007; Zhao et al., 2008). ...
... 放牧和增温会引起草地植物群落结构与物种多样性的改变, 进而对植物生产力产生重要影响(Wang et al., 2012; Hopping et al., 2018; Liu et al., 2018).植物生产力与物种多样性密切相关, 较高的物种多样性可以维持较高的植物生产力, 同时也可以加强群落生态系统的稳定性(Tilman et al., 2001; 马文静等, 2013).本研究表明, 由于放牧具有补偿效应(McNaughton, 1979), 在物种多样性增高2%时, 地上净初级生产力增加29%.通过对牲畜采食量分析后发现, 2年间杂草采食量仅占总量的21.8%, 而禾草采食量高达78.2%, 与前人的研究成果一致(段敏杰等, 2010; 郑伟等, 2012; 干珠扎布等, 2013).本研究中, 增温对净初级生产力无显著影响, 但2016和2017年增温对净初级生产力的影响存在年际差异(图6B), 这可能是由于2017年降水量高于2016年造成的(图1, 图3C).Fu等(2018)在藏北高原高寒草地的研究也表明, 年际间的降水差异是引起增温对生产力年际差异性影响的主要原因.此外, 有研究表明, 在不同的空气温度背景下, 增温对草地净初级生产力的影响有所不同(Liu et al., 2018).因此, 两年间的温度差异也可能引起生产力对增温的差异性响应. ...
放牧强度对环青海湖高寒草原群落物种多样性和生产力的影响 4 2012
... 全球气候变化已成为世界各国政府和公众关注的焦点.IPCC第五次评估报告指出, 1880-2012年间全球地表平均温度上升0.85 ℃, 预计到21世纪末全球大气温度将增加1.5-4.8 ℃ (IPCC, 2013).草地是陆地生态系统的重要组成部分, 且极为脆弱, 对气候变化极其敏感.气候变化直接或间接地影响草地植物的群落结构与组成(Polley et al., 2014; Ganjurjav et al., 2016)、物种多样性及生产力(Wu et al., 2011; Wang et al., 2012).除气候变化外, 人类活动也是影响草地植物群落的重要因子.有研究认为, 人类活动对植物的影响大于环境因子, 甚至成为控制植物群落多样性的决定因素(Brown et al., 1997; Wang et al., 2006).放牧是草地生态系统最为重要的人类干扰因素, 植物物种多样性与放牧强度显著相关(段敏杰等, 2010; 郑伟等, 2012).放牧作为草地最普遍的利用方式, 牲畜通过采食和践踏行为对草地植物群落特征产生影响, 同时也会改变局地光、水和土壤养分利用率, 从而间接改变群落物种结构与分布(Zhou et al., 2007; Zhao et al., 2008). ...
... 放牧和增温会引起草地植物群落结构与物种多样性的改变, 进而对植物生产力产生重要影响(Wang et al., 2012; Hopping et al., 2018; Liu et al., 2018).植物生产力与物种多样性密切相关, 较高的物种多样性可以维持较高的植物生产力, 同时也可以加强群落生态系统的稳定性(Tilman et al., 2001; 马文静等, 2013).本研究表明, 由于放牧具有补偿效应(McNaughton, 1979), 在物种多样性增高2%时, 地上净初级生产力增加29%.通过对牲畜采食量分析后发现, 2年间杂草采食量仅占总量的21.8%, 而禾草采食量高达78.2%, 与前人的研究成果一致(段敏杰等, 2010; 郑伟等, 2012; 干珠扎布等, 2013).本研究中, 增温对净初级生产力无显著影响, 但2016和2017年增温对净初级生产力的影响存在年际差异(图6B), 这可能是由于2017年降水量高于2016年造成的(图1, 图3C).Fu等(2018)在藏北高原高寒草地的研究也表明, 年际间的降水差异是引起增温对生产力年际差异性影响的主要原因.此外, 有研究表明, 在不同的空气温度背景下, 增温对草地净初级生产力的影响有所不同(Liu et al., 2018).因此, 两年间的温度差异也可能引起生产力对增温的差异性响应. ...
Source components and interannual variability of soil CO2, efflux under experimental warming and clipping in a grassland ecosystem 1 2007
... 全球气候变化已成为世界各国政府和公众关注的焦点.IPCC第五次评估报告指出, 1880-2012年间全球地表平均温度上升0.85 ℃, 预计到21世纪末全球大气温度将增加1.5-4.8 ℃ (IPCC, 2013).草地是陆地生态系统的重要组成部分, 且极为脆弱, 对气候变化极其敏感.气候变化直接或间接地影响草地植物的群落结构与组成(Polley et al., 2014; Ganjurjav et al., 2016)、物种多样性及生产力(Wu et al., 2011; Wang et al., 2012).除气候变化外, 人类活动也是影响草地植物群落的重要因子.有研究认为, 人类活动对植物的影响大于环境因子, 甚至成为控制植物群落多样性的决定因素(Brown et al., 1997; Wang et al., 2006).放牧是草地生态系统最为重要的人类干扰因素, 植物物种多样性与放牧强度显著相关(段敏杰等, 2010; 郑伟等, 2012).放牧作为草地最普遍的利用方式, 牲畜通过采食和践踏行为对草地植物群落特征产生影响, 同时也会改变局地光、水和土壤养分利用率, 从而间接改变群落物种结构与分布(Zhou et al., 2007; Zhao et al., 2008). ...
 ,1,*, 李钰2, 闫玉龙3, 胡国铮1, 王学霞1, 严俊4, 何世丞4
,1,*, 李钰2, 闫玉龙3, 胡国铮1, 王学霞1, 严俊4, 何世丞4
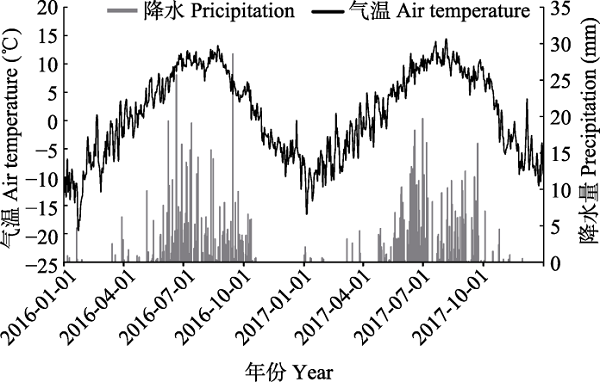 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT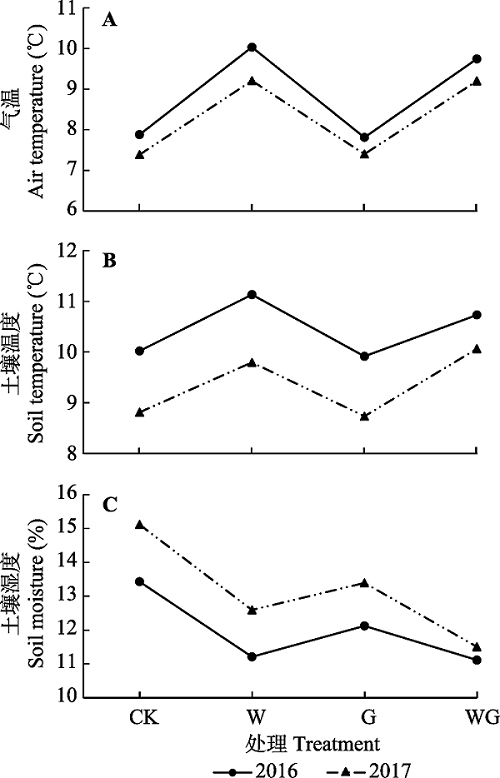 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT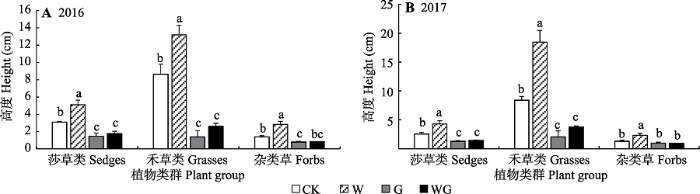 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT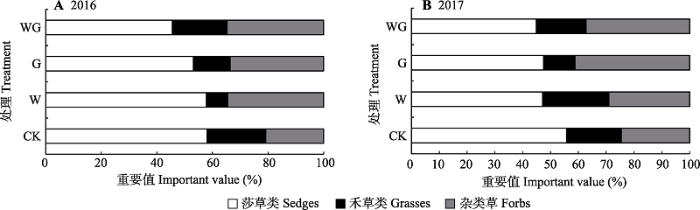 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT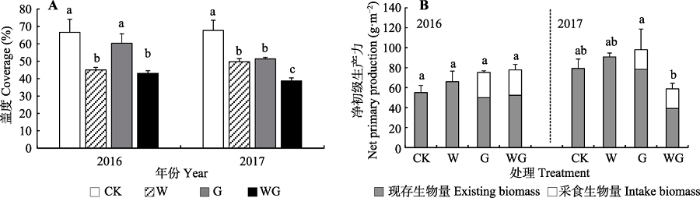 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT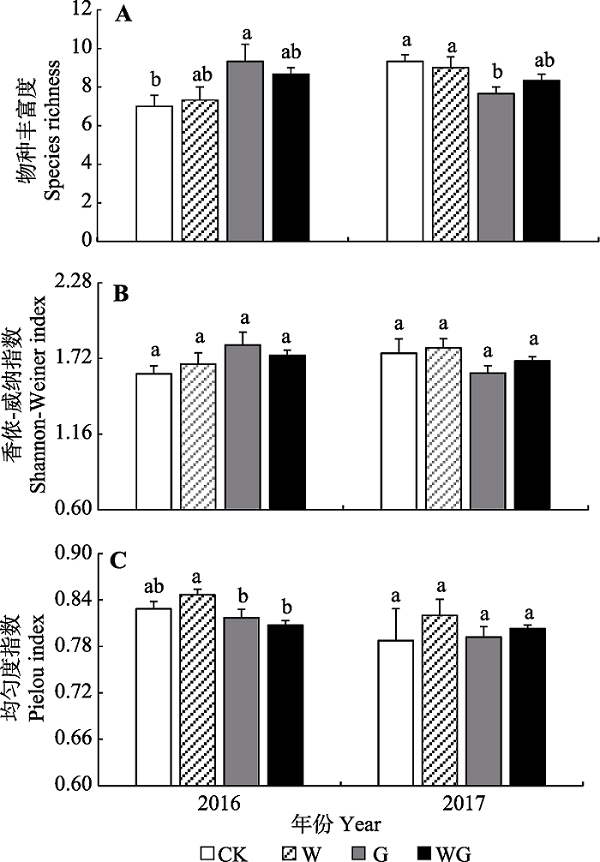 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT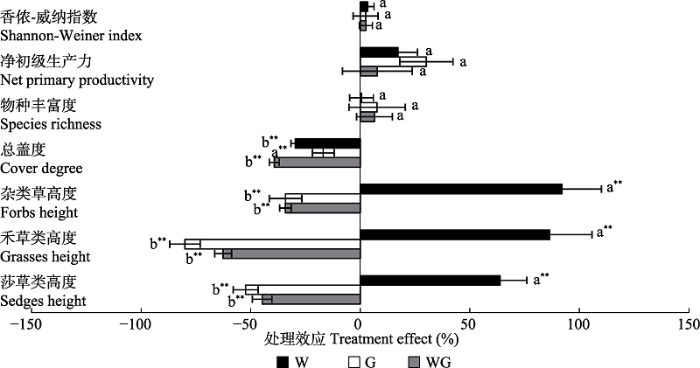 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}