 ,1,2, 闫慧娟1,2, 卫腾宙1,2, 周文君1,2, 贾昕1,2,3, 查天山
,1,2, 闫慧娟1,2, 卫腾宙1,2, 周文君1,2, 贾昕1,2,3, 查天山 ,1,2,3,*
,1,2,3,*Relative changes of resource use efficiencies and their responses to environmental factors in Artemisia ordosica during growing season
Xin-Hao LI,1,2, Hui-Juan YAN1,2, Teng-Zhou WEI1,2, Wen-Jun ZHOU1,2, Xin JIA1,2,3, Tian-Shan ZHA,1,2,3,*通讯作者: 通信作者Corresponding author (tianshanzha@bjfu.edu.cn)
编委: 李敏
收稿日期:2019-08-7接受日期:2019-10-14网络出版日期:2019-10-20
| 基金资助: |
Received:2019-08-7Accepted:2019-10-14Online:2019-10-20
| Fund supported: |

摘要
为了探明西北半干旱区典型沙生植物油蒿(Artemisia ordosica)叶水平资源利用效率的相对变化及对环境因子的响应机制, 该研究于2018年5-10月, 使用LI-6400XT便携式光合仪测定了毛乌素沙地油蒿叶片的净光合速率(Pn)、蒸腾速率(E)、叶表面光合有效辐射(PARl)、叶表面温度(Tl)、叶表面相对湿度(RHl), 在实验室计算叶片单位面积氮含量(Narea), 分析了叶片氮利用效率(NUE)、水分利用效率(WUE)、光利用效率(LUE)与环境因子之间的关系及NUE、WUE、LUE之间的相对变化。研究结果表明, 在充足且稳定光强下油蒿的Pn主要受温度的影响, NUE、WUE与VPDl、Tl之间具有显著负相关关系, NUE、WUE与LUE间为正相关关系, NUE、WUE和LUE最大值分别发生在5、7和9月, 分别为9.43 μmol CO2·g-1·s-1、3.86 mmol·mol-1、0.04 mol·mol-1, 资源利用效率的变化主要受Pn的影响。温度通过影响植物N分配来改变Pn, 进而影响着资源利用效率, WUE与LUE显著正相关, 对构建荒漠区生态系统能量交换过程模型有重要意义。
关键词:
Abstract
Aims This study is to examine the relative changes in resource use efficiencies and their responses to the environmental variables of a typical desert plant Artemisia ordosica in the semi-arid area of Northwest China. Methods We measured the net photosynthetic rate (Pn), transpiration rate (E), leaf surface photosynthetically active radiation (PARl), leaf surface temperature (Tl), leaf surface relative humidity (RHl) of Artemisia ordosica from May to October, 2018 using a portable photosynthesis analyzer. We also analyzed nitrogen per leaf area (Narea) in the lab. We related nitrogen use efficiency (NUE), water use efficiency (WUE), light use efficiency (LUE) of the plant and their relative changes to the environmental variables. Important findings Temperature mainly affected the Pn of Artemisia ordosica under sufficient and stable light intensity. There was a significantly negative correlation between NUE, WUE and VPD1, Tl. NUE, WUE and LUE was positively correlated. The maximum NUE, WUE and LUE occurred in May, July and September, respectively, being 9.43 μmol CO2·g-1·s-1, 3.86 mmol·mol-1, and 0.04 mol·mol-1. The changes in resource use efficiencies were mainly affected by Pn. The results indicate that temperature affects resource use efficiencies by affecting the distribution of plant N and changes in Pn. WUE was significantly and positively correlated with LUE. These results may contribute to development of the energy exchange process model for the desert ecosystems.
Keywords:
PDF (1180KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
李鑫豪, 闫慧娟, 卫腾宙, 周文君, 贾昕, 查天山. 油蒿资源利用效率在生长季的相对变化及对环境因子的响应. 植物生态学报, 2019, 43(10): 889-898. DOI: 10.17521/cjpe.2019.0214
LI Xin-Hao, YAN Hui-Juan, WEI Teng-Zhou, ZHOU Wen-Jun, JIA Xin, ZHA Tian-Shan.
我国干旱半干旱地区占国土总面积的49%, 这一地区分布范围很广, 虽然植物稀少, 但基于如此广阔的分布面积, 其对碳循环和碳收支的贡献不容忽略(Chaves et al., 2004)。荒漠植物作为整个碳循环过程中的纽带, 对荒漠生态系统碳固定、水平衡以及养分动态等各方面都起着至关重要的作用(Bloom et al., 1985)。
由于全球气候变化, 增加了干旱半干旱地区极端天气的出现频率和强度(Schimel, 2010), 使得该地区的生态系统面临着急剧退化的严峻挑战(康博文等, 2010)。荒漠植物在干旱半干旱地区经常会遭受水分缺失、高温、高辐射等环境因子的胁迫(Porcar-Castell, 2011), 它们通过调整其形态、生理或生化特征来适应环境(Haque et al., 2014)。近年来, 植物的资源利用效率及与环境因子的关系一直是人们比较关注的问题, 其中植物叶片氮利用效率(NUE)、水分利用效率(WUE)及光利用效率(LUE)是许多生态系统生产模型中的重要参数, 是决定植物适应多变的环境条件, 协调自身碳水关系的关键因子。
植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(裴斌等, 2013)。植物所处的环境变化时, 光合作用也会受到不同程度的影响(张卫强等, 2006), 从而改变资源利用效率。例如, 刚毛柽柳(Tamarix hispida)在水分亏缺情况下通过降低蒸腾速率维持相对较高的光合速率, 从而有效提高了WUE, 以适应荒漠环境(孟鹏等, 2012); 银中杨(Populus alba ‘Berolinensis’)的光合速率随着温度的上升而降低(王晶英等, 2006)。植物的光合作用是控制植物与外界环境进行能量与物质交换的重要过程(王芸等, 2013), 控制着植物对CO2等物质的吸收, 对各资源利用效率有直接影响(王燕凌等, 2015)。因此, 了解植物光合作用与环境因子之间的相互关系可以有助于我们进一步了解荒漠植物的能量和水分交换动态过程(刘慧霞等, 2009)。
油蒿(Artemisia ordosica)为菊科蒿属, 半干旱区典型沙生植物, 近年来许多相关研究多是在控制试验条件下进行的(Reich et al., 1989; Chen et al., 2015), 影响因素较单一。本研究使用LI-6400XT便携式光合仪(LI-COR, Lincoln, USA), 进行原位测定, 分析油蒿叶片的NUE、WUE和LUE, 并结合同步监测的环境因子, 分析油蒿叶片生长季NUE、WUE和LUE的相对变化及其对环境因子的响应, 探明荒漠半干旱地区优势灌木种油蒿对毛乌素沙地干旱环境的适应机制, 旨在揭示: (1)油蒿的资源利用效率对环境因子的响应机制; (2)油蒿资源利用效率之间的相对变化。以期为构建生态系统能量交换过程模型提供依据。
1 材料和方法
1.1 样地概况
研究区位于宁夏盐池毛乌素沙地生态系统国家定位观测研究站, 实验站位于毛乌素沙地南缘, 37.07°-38.17° N, 106.5°-107.78° E, 海拔1 550 m。该区气候类型属于典型的中温带大陆性季风气候, 年降水量约287 mm, 且主要集中在7-9月, 降水年际变化大、保证率低, 但蒸发量却是降水量的6-7倍。年平均气温7.7 ℃, 极端最高值38.1 ℃, 极端最低值-29.6 ℃, 无霜期为128天, 冬春风沙天气较多。日照时间长, 温差大, 地区差异明显。研究区内以干旱区沙生植物为主, 优势物种为油蒿, 其他物种有花棒(Hedysarum scoparium)、杨柴(Hedysarum mongolicum)和北沙柳(Salix psammophila)等。油蒿, 又名黑沙蒿, 隶属菊科蒿属龙蒿亚属, 半灌木植物, 株高50-100 cm, 无明显主茎, 多分枝, 广泛分布于毛乌素沙地, 主要生长于固定半固定沙丘, 具有耐干旱、抗沙埋和耐贫瘠的特性。油蒿萌芽期在3月中下旬, 展叶期在4月初至5月初, 开花结实期在6月初至8月下旬, 落叶期为9月初至10月初。本研究样地内油蒿平均株高67.3 cm, 共27株。
1.2 研究方法
本实验监测点位于宁夏盐池荒漠生态系统定位研究站, 以毛乌素沙地的优势种典型沙生灌木油蒿为研究对象。在定位研究站内的通量塔周围, 选择一块地势平坦的油蒿样地(10 m × 10 m), 通过微气象观测系统对整个生长季的环境气象数据进行同步监测。1.2.1 气体交换参数的测定
于5月1日至10月1日, 在通量塔附近选取一块大小为10 m × 10 m的样地, 从中选择5株长势均一、无病虫害的健康油蒿作为实验植株, 在每个月上中下旬选择晴朗无风的天气使用透明叶室在8:00- 11:00进行油蒿实时光合的测量。于每株油蒿向阳方向的靠下位置分别选取生长健康、无病虫害的叶片测定气体交换参数, 每株分别选取3片1-2年生小枝上的健康叶片, 当测量数值小数点后一位稳定时, 每个叶片记录20次读数, 测量指标有净光合速率(Pn)、蒸腾速率(E)、叶表面光合有效辐射(PARl)、叶表面温度(Tl)、叶表面相对湿度(RHl)等。
测定完气体交换参数后, 由于所测油蒿叶片面积小于叶室面积, 因此在进行叶片样品采集时, 选择生长良好、大小一致、没有遮阴的10片完全伸展、无病虫害的典型叶片, 将每个样株测定的叶片取回实验室进行拍照, 随后通过MATLAB计算其叶面积, 再计算气体交换值。将计算过叶面积的叶片在105 ℃下杀青20 min, 接着在70 ℃下持续烘干48 h至恒质量, 计算叶片干质量, 比叶面积(SLA)即为叶片面积与干质量的比值。将烘干后的叶片混合粉碎, 装入自封袋中, 用记号笔标明取样日期, 制成供试样品。由于实验站不能直接测量, 故带回北京委托中国科学研究院植物所测定。单位面积氮含量(Narea)即为氮含量与叶片面积之比。
1.2.2 光响应曲线和CO2响应曲线的测定
在每次气体交换参数测定时, 使用LI-6400-02红蓝光源叶室同时进行油蒿CO2响应曲线和光响应曲线的测量。于每株油蒿向阳方向的靠下位置分别选取生长健康、无病虫害的3片叶片测定光响应曲线和CO2响应曲线。
对油蒿进行光响应曲线的测定前要先对目标叶片进行充分的诱导。叶室温度设置为25 ℃, CO2浓度设置为环境中的浓度即可, 即400 μmol CO2·mol-1左右, 相对湿度设置为55%左右。光强梯度共设置18个点, 分别为2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、100、80、60、40、30、20、10和 0 μmol·m-2·s-1。进行测定时应在每个设置的光合有效辐射下适应3 min测定其光合速率。测定完成后通过模型拟合得到相关参数。
进行CO2响应曲线的测定时的温度、相对湿度的设定与光响应曲线测定相同。测定时的光强设置为饱和光强, CO2浓度梯度设定为50、60、80、100、200、300、400、600、800、1 000、1 200、1 400、1 450、1 500、1 550和1 600 μmol CO2·mol-1, 在每个CO2浓度下适应3 min后测定其光合速率。测定完成后通过模型拟合求得其最大羧化速率(Vcmax)和最大电子传递效率(Jmax), 分别用于光合系统中羧化系统和生物力能学系统的氮分配比例的计算。
1.2.3 环境因子的测定
环境因子由位于样地西北侧通量塔的微气象系统进行观察记录。其中, 光合有效辐射(PARa)由光合有效辐射传感器(PAR-LITE, Kipp and Zonen, Delft, the Netherlands)测得, 传感器布设在距地表5 m左右的通量塔上。空气温度(Ta)和空气相对湿度(RHa)由安装在地上6 m高度的传感器(HMP155A, Campbell Scientific, Logan, USA)测量。降水量数据由翻斗式雨量筒(TE-525 MM, Campbell Scientific, Logan, USA)测得, 雨量筒位于距通量塔50 m的地方。在距离通量塔5 m处的地方布设土壤水分探头(CS-616, Campbell Scientific, Logan, USA), 布设的深度分别为10 cm和30 cm; 通过CR3000数据采集器(CR3000, Campbell Scientific, Logan, USA)分别记录降水总量和土壤平均含水量, 每隔0.5 h记录一次数据。
1.3 数据的处理
1.3.1 最大羧化速率和最大电子传递速率的计算植物的最大羧化速率(Vcmax)和最大电子传递速率(Jmax)是通过LI-6400XT测定, 利用CO2气体小钢瓶进行CO2浓度控制, 测量植物叶片胞间CO2浓度响应曲线(A-Ci), 得到相关参数并进行计算。先通过CO2浓度响应曲线初始部分(外界CO2浓度在50-200 μmol·mol-1)的胞间CO2浓度(Ci)与A作拟合直线: A = k × Ci + i, 得到斜率k以及i。其中Ci为LI-6400直接测得; k即为羧化效率CE。然后通过Farquhar等方法计算Vcmax:
式中, Г*为无暗呼吸时的CO2补偿点, Kc为Michaelis-Menten RUBP (Ribulose-1,5-disphosphate, 1,5-二磷酸核酮糖)的羧化常量, 为469.83 μmol·mol-1, Ko为Michaelis-Menten RUBP的氧化常量, 为456.21 mmol·mol-1。O为植物细胞间的O2浓度, 为210 mmol·mol-1。
最大电子传递速率(Jmax)通过Loustau等(1999)提出的方法进行计算:
式中, Amax为最大净光合速率, Rd为暗呼吸速率。
1.3.2 叶片光合系统内氮分配组分比例计算
植物叶片光合系统内氮分配比例的各组分可利用Niinemets和Tenhunen (1997)提出的公式进行计算。在该方法中光合系统中的含氮物质可以按其在光合作用中的功能划分为三部分: Rubisco、生物力能学组分以及捕光组分, 其各部分氮含量占叶片总氮比例的计算方法如下:
羧化系统的分配比例:
生物力能学组分分配比例:
捕光系统分配比例:
光合系统氮分配比例:
式中, PC为光合系统中Rubisco组分内的氮占叶片总氮的比例; PB为光合系统生物力能学组分内的氮占叶片总氮的比例; PL为光合系统中捕光组分内的氮占叶片总氮的比例。植物叶片中分配给光合系统的氮占叶片总氮的比例(PP)即为以上3个比例之和。Narea为单位面积叶片氮含量, 即单位质量叶片氮含量与比叶面积的比值; Vcmax单位为μmol·m-2·s-1; Jmax单位为μmol·m-2·s-1; CC为叶片叶绿素浓度(mmol·g-1); Vcr是Rubisco酶的比活, 即单位Rubisco组分中的CO2固定活性(μmol CO2·g-1 Rubisco·s-1); Jmc为单位细胞色素f (Cytochrome f, Cyt f)的最大电子传递速率(μmol electrons·μmol-1 Cyt f·s-1)。Vcr和Jmc的计算方法如下(Harley et al., 1992):
式中, R为通用气体常数, 取8.314 J·K-1·mol-1; Tk是叶片温度(K), ΔHa、ΔHd、ΔS和c分别为活化能、去活化能、熵和比例常数, 计算Vcr时的值分别取74 kJ·mol-1、203 kJ·mol-1、645 J·K-1· mol-1和32.9; 计算Jmc时的值分别取24.1 kJ·mol-1、564.15 kJ·mol-1、1 810 J·K-1·mol-1和14.77。
CB为光合系统捕光组分中叶绿素与氮的比值(mmol Chl·(g·N)-1), 可以利用CB与比叶重(LMA)关系的线性公式来进行计算, 其相关系数较高(R2 = 0.97), 并且证明可以在多种植物类型中广泛应用(史作民等, 2014)。其公式为:
式中, LMA为单位面积叶片干质量(g·m-2)的值, 这样就可以通过LMA的值来计算出CB的值, 以便进一步计算PL。
1.3.3 资源利用效率的计算
氮利用效率计算公式为:
水分利用效率计算公式为:
光利用效率计算公式为:
因为每个月可用的数据点数量比较少, Pn与环境因子的关系描述为简单的线性回归关系, 并且在所有分析中没有包含交互条件, 参考已有的研究(Tarvainen et al., 2015), 建立的完整回归模型如下:
式中a, b, c和d是常数, Pn的单位为μmol·m-2·s-1, E的单位为mmol·m-2·s-1, PAR的单位是μmol·m-2·s-1, Narea的单位是g·m-2, T的单位是℃, intercept为残差。基于拟合的R2和Akaike信息标准(AIC)来评估模型的性能(Burnham & Anderson, 2002):
式中SSE是标准误差的平方和, n是样本容量, p是回归分析中使用的独立变量的数量。
R2描述了回归模型的整体拟合程度, 而AIC值用来评价多个回归模型各自的优良程度(Burnham et al., 2011), AIC值越小模型越优良。
所有分析均在Matlab 7.11.0中进行。
2 结果和分析
2.1 环境因子的动态变化
如图1A所示, 整个观测期内PARa平均值为403.89 μmol·m-2·s-1, 最大值为610.65 μmol·m-2·s-1 (第179天), 最小值为77.75 μmol·m-2·s-1 (第244天); 在图1B中, Ta的平均值为17.49 ℃, 最高气温为23.58 ℃ (第238天), 日平均最低值为5.44 ℃ (第125天); 在图1C中, VPDa的平均值为0.82 kPa, 最大值为2.04 kPa (第163天), 最小值为0.18 kPa (第130天); 由图1D可知, 生长季的总降水量为212.67 mm, 最大一次降水为21.9 mm (第180天)。土壤含水量随降水量变化而变化, 整个研究期的平均值为0.078 m3·m-3, 最大值为0.112 m3·m-3 (第243天), 最小值为0.52 m3·m-3 (第166天)。2.2 光合参数的生长季动态变化
在观测期间, 油蒿的气体交换参数如表1所示, 根据表1可计算光合系统氮分配比例, 结果如表2所示。由表1可见Narea在整个研究期内并无较大波动, 均维持在4 g·m-2上下, 光饱和点平均值为851.57 μmol·m-2·s-1。对T与Pp之间的关系作线性拟合, 结果如图2所示, T与Pp之间呈极显著负相关关系。图1
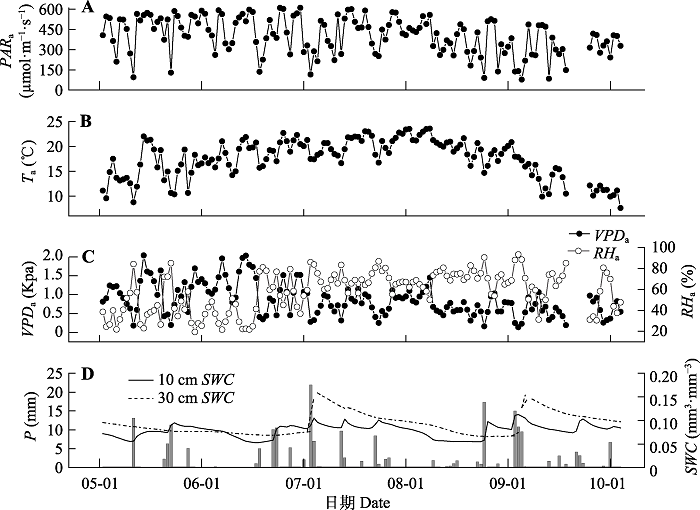 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1毛乌素沙地油蒿样地环境因子日均值的动态变化。P, 降水量; PARa, 光合有效辐射; RHa, 空气相对湿度; SWC, 土壤含水量; Ta, 空气温度; VPDa, 空气饱和水汽压差。
Fig. 1Dynamics in daily means of environmental variables from May 1 to October 1 in the sample plots of Artemisia ordosica in Mau Us Sandyland. P, precipitation; PARa, photosynthetically active radiation; RHa, air relative humidity; SWC, soil water content; Ta, air temperature; VPDa, air saturated water vapor pressure difference.
Table 1
表1
表1毛乌素沙地油蒿的气体交换参数
Table 1
| 日期 Date (Month-day) | E (mmol·m-2·s-1) | gs (mol·m2·s-1) | Narea (g·m-2) | SLA (cm2·g-1) | LSP (μmol·m-2·s-1) | Pn (μmol·m-2·s-1) | PAR (μmol·m-2·s-1) |
|---|---|---|---|---|---|---|---|
| 05-16 | 14.11 (1.6) | 0.51 (0.03) | 4.29 (1.20) | 61.30 (10.20) | 1 057 (95.1) | 28.78 (5.6) | 1 476.21 (96.7) |
| 05-22 | 10.60 (2.4) | 0.72 (0.11) | 4.16 (0.41) | 72.77 (12.10) | 1 034 (26.8) | 39.30 (5.1) | 1 787.40 (102.4) |
| 06-04 | 11.63 (1.9) | 0.38 (0.05) | 4.24 (0.57) | 43.22 (6.07) | 792 (54.7) | 22.40 (4.2) | 1 185.47 (123.7) |
| 06-13 | 12.84 (1.5) | 0.41 (0.06) | 4.36 (0.81) | 45.55 (5.47) | 784 (43.2) | 22.42 (4.1) | 1 357.43 (111.9) |
| 06-23 | 16.40 (3.4) | 0.53 (0.09) | 3.85 (0.73) | 61.98 (8.81) | 694 (41.9) | 24.30 (3.6) | 1 260.54 (98.6) |
| 07-05 | 17.93 (4.1) | 0.48 (0.08) | 3.70 (0.42) | 75.20 (7.34) | 1 119 (80.6) | 21.16 (2.7) | 1 662.20 (89.4) |
| 07-13 | 16.14 (1.9) | 0.52 (0.12) | 3.61 (0.64) | 60.50 (10.31) | 731 (45.7) | 19.77 (3.4) | 1 319.01 (127.5) |
| 07-23 | 6.68 (0.9) | 0.37 (0.06) | 3.83 (0.78) | 83.50 (14.81) | 1 234 (56.4) | 33.36 (4.1) | 1 152.30 (131.6) |
| 08-05 | 5.30 (0.8) | 0.36 (0.09) | 3.83 (0.17) | 60.84 (4.59) | 594 (12.9) | 10.70 (1.9) | 766.48 (234.8) |
| 08-14 | 8.80 (1.1) | 0.46 (0.10) | 4.04 (0.59) | 56.34 (4.96) | 779 (61.1) | 23.10 (5.1) | 927.46 (194.5) |
| 08-23 | 12.83 (5.4) | 0.49 (0.14) | 3.95 (0.88) | 58.53 (10.31) | 613 (31.8) | 11.94 (2.7) | 746.55 (84.9) |
| 09-02 | 14.20 (3.9) | 0.68 (0.22) | 3.91 (0.46) | 72.58 (11.62) | 847 (61.7) | 27.01 (4.1) | 1 148.57 (154.6) |
| 09-14 | 7.21 (2.1) | 0.63 (0.19) | 3.81 (0.53) | 68.65 (9.96) | 880 (31.1) | 23.34 (4.5) | 1 156.36 (164.8) |
| 09-23 | 5.68 (1.1) | 0.59 (0.18) | 4.12 (0.33) | 65.02 (8.13) | 764 (26.8) | 20.28 (4.2) | 1 192.83 (147.6) |
新窗口打开|下载CSV
Table 2
表2
表2毛乌素沙地油蒿光合系统氮分配参数
Table 2
| 日期 Date (Month-day) | Pc | Pb | Pl | Pp |
|---|---|---|---|---|
| 05-16 | 0.210 (0.06) | 0.015 (0.005) | 0.050 (0.010) | 0.275 (0.06) |
| 05-22 | 0.340 (0.06) | 0.025 (0.006) | 0.006 (0.021) | 0.347 (0.05) |
| 06-04 | 0.281 (0.02) | 0.014 (0.002) | 0.018 (0.015) | 0.301 (0.03) |
| 06-13 | 0.282 (0.04) | 0.016 (0.003) | 0.050 (0.010) | 0.297 (0.04) |
| 06-23 | 0.230 (0.07) | 0.019 (0.003) | 0.013 (0.021) | 0.248 (0.05) |
| 07-05 | 0.211 (0.06) | 0.030 (0.005) | 0.002 (0.012) | 0.253 (0.06) |
| 07-13 | 0.206 (0.04) | 0.020 (0.002) | 0.050 (0.012) | 0.276 (0.04) |
| 07-23 | 0.242 (0.10) | 0.029 (0.006) | 0.075 (0.026) | 0.350 (0.03) |
| 08-05 | 0.190 (0.01) | 0.023 (0.003) | 0.044 (0.011) | 0.253 (0.02) |
| 08-14 | 0.282 (0.03) | 0.029 (0.008) | 0.004 (0.011) | 0.311 (0.03) |
| 08-23 | 0.270 (0.03) | 0.015 (0.002) | 0.030 (0.006) | 0.291 (0.03) |
| 09-02 | 0.272 (0.06) | 0.018 (0.001) | 0.004 (0.012) | 0.282 (0.04) |
| 09-14 | 0.261 (0.04) | 0.015 (0.003) | 0.039 (0.010) | 0.308 (0.04) |
| 09-23 | 0.290 (0.04) | 0.022 (0.006) | 0.006 (0.009) | 0.318 (0.05) |
新窗口打开|下载CSV
2.3 资源利用效率的生长季动态变化
根据表1参数可计算NUE等, 结果见图3。整个观测期内NUE (图3A)的平均值为5.88 μmol CO2·g-1·s-1, 最大值为9.43 μmol CO2·g-1·s-1 (第145天), 最小值为2.80 μmol CO2·g-1·s-1 (第215天)。WUE (图3B)的平均值2.10 mmol·mol-1, 最大值3.86 mmol·mol-1 (第205天), 最小值为0.82 mmol·mol-1 (第185天)。LUE (图2C)的平均值0.019 2 mol·mol-1, 最大值为0.029 mol·mol-1 (第205天), 最小值为0.012 mol·mol-1 (第215天)。2.4 净光合作用的多重资源限制
在研究期间, 所有单参数模型(E、Tl、PARl、Narea)与Pn呈负相关关系(表3), Narea、E对Pn的预测能力非常差, Tl是叶水平Pn单参数的最佳预测指标; 双参数模型中E与Tl可以解释约42.8%的Pn变化; 三参数模型中E、Tl及PARl组成的模型最优。所有模型中实测值与直线拟合关系显著相关的有: 包含Tl的单参数模型; 包含E与Tl的双参数模型; 包含E、Tl及PARl的三参数模型。它们的AIC值在其同类型的模型中也最小, 模型最优。总体而言, 表现较好的模型都是将Tl与其他参数相结合。图2
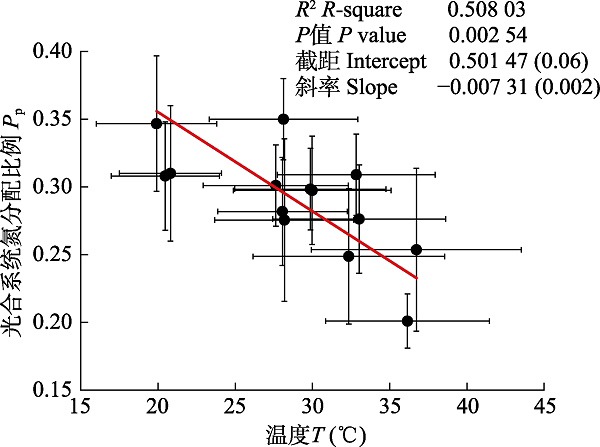 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2温度(T)对毛乌素沙地油蒿光合系统氮分配比例(Pp)的影响。
Fig. 2Effect of temperature (T) on nitrogen distribution allocation ratio of photosynthesis system(Pp) of Artemisia ordosica in Mau Us Sandyland.
图3
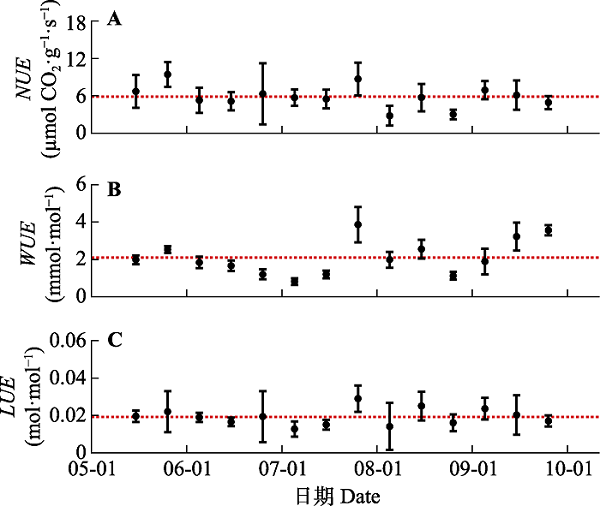 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3毛乌素沙地油蒿氮利用效率(NUE)、水利用效率(WUE)和光利用效率(LUE)在整个生长季的动态变化(平均值±标准偏差)。水平虚线为其对应利用效率的平均值。
Fig. 3Seasonal changes in nitrogen use efficiency (NUE), water use efficiency (WUE), light use efficiency (LUE) of Artemisia ordosica in Mau Us Sandyland (mean ± SD). The horizontal dashed lines in the figure are the average values of the corresponding resource use efficiency.
2.5 资源利用效率对环境因子的响应
环境因子可能直接对NUE、WUE及LUE造成影响, 对环境因子SWC、Tl、VPDl与NUE、WUE及LUE进行线性拟合, 结果见表4。其中10 cm处的土壤体积含水量(SWC10)和30 cm处的土壤体积含水量(SWC30)与NUE、WUE及LUE之间关系不显著, Tl、VPDl与NUE、WUE及LUE间均呈负相关关系, LUE 与Tl和VPDl之间关系不显著, Tl、VPDl与NUE、WUE之间关系显著, 其中WUE与Tl、VPDl之间的R2值 相比于NUE、LUE与Tl、VPDl之间的值大, 拟合性较好。Table 3
表3
表3毛乌素沙地油蒿光合速率与环境因子关系的模型拟合参数
Table 3
| a | b | c | d | i | SSE | R2 | p | AIC | |
|---|---|---|---|---|---|---|---|---|---|
| E | -0.004 5 (0.774 5) | 21.130 0 (9.446 3) | 347.4 | 0.083 3 | 0.998 9 | 50.95 | |||
| Tl | -0.567 3 (5.304 0) | 37.450 0 (2.940 0) | 224.7 | 0.353 1 | 0.025 1 | 44.86 | |||
| PARl | -0.007 6 (0.010 6) | 29.320 0 (11.833 7) | 288.6 | 0.169 2 | 0.144 0 | 48.36 | |||
| Narea | -0.125 7 (4.862 4) | 21.560 0 (18.830 0) | 347.3 | 0.000 3 | 0.956 0 | 50.96 | |||
| E × Tl | 0.373 2 (0.684 5) | -0.689 4 (0.529 1) | 36.698 2 (14.125 5) | 198.7 | 0.427 9 | 0.046 3 | 45.14 | ||
| E × PARl | 0.892 3 (0.979 6) | -0.017 8 (0.014 7) | 30.043 7 (10.716 9) | 211.4 | 0.391 5 | 0.065 1 | 46.01 | ||
| E × Narea | -0.006 7 (0.824 3) | -0.130 0 (5.157 6) | 21.650 4 (22.947 2) | 347.3 | 0.000 3 | 0.998 4 | 52.96 | ||
| Tl × PARl | -0.526 2 (0.648 0) | -0.001 3 (0.012 6) | 37.658 2 (15.047 4) | 223.7 | 0.356 1 | 0.088 8 | 46.80 | ||
| Tl × Narea | -0.581 4 (0.510 8) | -0.796 4 (4.135 7) | 40.900 1 (23.240 6) | 221.1 | 0.363 5 | 0.083 4 | 46.63 | ||
| Narea × PARl | -0.776 6 (4.740 7) | -0.008 0 (0.011 4) | 32.677 2 (23.971 2) | 285.2 | 0.178 9 | 0.338 2 | 52.20 | ||
| E × Tl × PARl | 0.824 8 (0.903 7) | -0.478 1 (0.581 3) | -0.011 3 (0.015 7) | 37.566 1 (13.437 9) | 158.2 | 0.544 5 | 0.041 8 | 43.95 | |
| E × Tl × Narea | 0.367 4 (0.722 6) | -0.699 8 (0.561 1) | -0.696 5 (4.138 5) | 39.723 7 (23.345 1) | 195.9 | 0.435 9 | 0.112 3 | 46.94 | |
| E × PARl × Narea | 0.909 9 (1.027 7) | -0.018 5 (0.015 7) | -1.043 5 (4.281 2) | 34.571 8 (21.699 6) | 205.3 | 0.408 9 | 0.138 7 | 47.60 | |
| Tl × PARl × Narea | -0.529 9 (0.681 9) | -0.001 7 (0.013 4) | -0.871 6 (4.416 7) | 41.487 4 (25.038 0) | 219.5 | 0.368 3 | 0.186 7 | 48.53 | |
| E × Tl × PARl × Narea | 0.843 1 (0.948 1) | -0.481 7 (0.608 5) | -0.012 0 (0.016 7) | -1.110 3 (3.935 1) | 42.441 8 (22.281 5) | 151.4 | 0.564 2 | 0.084 3 | 45.33 |
新窗口打开|下载CSV
Table 4
表4
表4毛乌素沙地油蒿氮利用效率(NUE)、水利用效率(WUE)和光利用效率(LUE)与各环境因子的相关关系
Table 4
| SWC10 | SWC30 | Tl | VPDl | ||
|---|---|---|---|---|---|
| NUE | R2 | 0.38 | 0.21 | -0.30 | -0.29 |
| p | 0.176 9 | 0.461 3 | 0.031 0 | 0.025 5 | |
| WUE | R2 | -0.08 | 0.07 | -0.42 | -0.31 |
| p | 0.772 0 | 0.804 6 | 0.010 9 | 0.036 9 | |
| LUE | R2 | 0.001 | 0.12 | -0.28 | -0.27 |
| p | 0.996 4 | 0.679 2 | 0.051 3 | 0.058 0 |
新窗口打开|下载CSV
3 讨论
3.1 净光合作用的多种资源限制
植物的净光合能力与PAR的关系非常密切(Medlyn et al., 2002; Han et al., 2004; Tarvainen et al., 2013), 当光照稳定且较高时, T成为主导因子, 因此, 植物净光合能力受到气候季节性变化的限制, 影响其资源利用效率。油蒿的Pn随着PAR的增加而降低(Wang et al., 2003)(表3), 由表1可知, 研究期内的LSP平均值为851 μmol·m-2·s-1, 而进行实时光合测量时PAR值基本都在LSP之上, 植物已经达到了光饱和状态, 所以随着PAR值的增加Pn反而会降低; Pn的变化也可以由VPD来解释, 高VPD使叶片水势和大气水汽之间的梯度变大, 造成植物E增大, 植物为了减少水分的耗散而关闭气孔, gs降低, 对CO2的供应造成影响, 随之使Pn降低。由于本研究测量时光照条件已超过油蒿的LSP (表1), 所以达到了光照稳定且饱和的状态, 且本研究在生长季持续测量, 在当地具有广泛的代表性, 所以不讨论当天气等环境因子变化时(如多云)的情况。综合Pn对环境因子的响应, 对Pn影响最大的是T (表3), T通过影响植物的N分配来对Pn产生作用从而表现出荒漠植物资源利用效率相对变化的特有模式。3.2 资源利用效率对环境因子的响应
油蒿NUE季节变异的影响因子有T与VPD, NUE与其都呈显著负相关关系, 与土壤含水量的相关关系不显著(表4)。NUE随着T与VPD的增加而下降, 可以从机理方面来解释(Troeng & Linder, 1982), 在生长季温度上升时相对应的空气也会变得更干燥, 同时也意味着水分的降低。在水分较低时, 植物会处于干旱胁迫的状态, 此时植物会在其叶片结构性状上做出一定的适应策略, 叶片会将更多的氮用于构造保卫结构来适应气候干旱对自身生活及生长所造成的影响, 比如将更多的氮用于形成不溶性蛋白纤维, 使自身细胞壁厚度和坚韧性增强, 为了防止气温的上升使自身受到损伤或失水过多, 叶片还会增加自身叶肉细胞的密度, 同时植物为了降低自身的水势, 还会在体内积累脯氨酸等可溶性化合物, 形成厚度大且面积小的叶片, 单位叶面积干物质量增加, 比叶面积变小。植物在进行氮分配时, 分配给非光合系统的氮的比例增加, 分配到光合系统的氮就会相应地减少, 因此随着T与VPD的升高, NUE会降低。在胁迫条件下的植物更倾向于将更多的氮分配到非光合组分中以增加植物的抗逆性, 保证其可以更顺利地完成生活史, 这与植物的生存策略有关, 与唐敬超等(2016)对格木(Erythrophleum fordii)和红锥(Castanopsis hystrix)的研究结果相一致。同时由图2可见, T与Pp呈极显著负相关关系, 证明在本研究中温度的升高影响了油蒿的光合系统氮分配, 从而影响了NUE。油蒿WUE季节变异的影响因子有T与VPD, WUE与其呈显著负相关关系, 与土壤含水量的相关关系不显著。T与VPD主要是通过影响gs进而影响WUE。温度的升高对植物的气孔有限制作用, 随着T的不断升高, 增大了叶片内外的水汽压差, 植物的蒸腾会变得强烈, 导致叶水势下降, 更加剧了蒸腾的变化, WUE也随之减小(阮成江和李代琼, 2001)。T的上升一般伴随着光照的增强, 随着PAR的增加, 强光与水分胁迫并存时, 会破坏光合作用固定CO2与吸收光能的平衡(Ma et al., 2018), 引发光抑制现象。随后气孔出于保护机制开始关闭, 因此蒸腾作用也逐渐削弱, 但是光合作用的下降程度大于蒸腾作用的程度, 因此WUE下降, 这与李建查等(2018)的研究结果一致。随着温度的升高, 叶片水势和大气水汽之间的梯度变大, 植物蒸腾作用不断加强, 直接导致WUE的下降, 且相比于非干旱时期, 干旱条件下的WUE变化更快, 可能是因为土壤的湿度条件降低了gs对VPD的敏感性(孙艳等, 2006)。
油蒿的LUE随着T与VPD的上升而逐渐下降, 主要是因为在生长季, 随着T的上升, 植物可利用的光能也不断增加, 但是油蒿生长在干旱地区, 光能不是其光合作用的主要限制因素, 植物体内的氮和水分不足以使光合强度随着PAR的增加一直增加下去, 所以随着PAR的上升, 光能对于植物来说处于富余状态, 超出了可利用范围, LUE反而会下降。
3.3 资源利用效率的相对变化
NUE、WUE和LUE之间的相对变化可能会由于观测时间尺度的不同而表现出不同的模式。在叶水平, 有研究(Cowan & Farquhar, 1977)认为由于温度的升高, 叶片为了减少水分蒸发而关闭气孔, 从而导致蒸腾降低, WUE增加, 而气孔的关闭同时造成CO2供应不足, Pn下降, LUE下降, 所以叶片尺度上的WUE与LUE呈负相关关系(Tarvainen et al., 2014)。在本研究中, 油蒿的NUE、WUE与LUE之间都呈正相关关系(图3A), NUE、WUE、LUE都同时增加和降低, 5到6月NUE较大, 7到9月WUE与LUE较大, 原因可能是7月前后测量时不同的环境因子对Pn的影响, 生长季T升高, 对Pp产生了影响, 从而影响Pn和NUE (图2)。较大NUE对应的LUE有大有小(图3B), 而Narea在一年中几乎没有变化, 也就是说在高光强与低光强条件下油蒿都能达到较大的光合强度, 光资源一直处于过量状态(表1), 而Pn的变化取决于T对植物N分配的影响, LUE与WUE之间为同增同降关系, 也意味着PAR与E是正比关系, 也证实了之前的研究结果(Ma et al., 2018)。本研究监测时间较短, 着重于NUE、WUE和LUE的季节性波动, 可能是时间尺度的不同造成NUE、WUE和LUE相对变化的不同模式(Reich et al., 1989), 强调了对植物进行长期监测的重要性。我们的结果表明, 当光照稳定且处于饱和状态时, 油蒿的Pn受T的影响最大, 二者呈负相关关系, NUE、WUE与T、VPD显著负相关, 生长季NUE、WUE及LUE之间呈正相关关系, 油蒿资源利用效率的相对变化取决于环境因子对Pn的影响, 对干旱区季风性气候区域的基于资源利用效率的碳通量建模具有重要意义。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
.
DOI:10.1146/annurev.es.16.110185.002051URL [本文引用: 1]
[本文引用: 1]
.
DOI:10.1007/s00265-010-1029-6URL [本文引用: 1]

We briefly outline the information-theoretic (I-T) approaches to valid inference including a review of some simple methods for making formal inference from all the hypotheses in the model set (multimodel inference). The I-T approaches can replace the usual t tests and ANOVA tables that are so inferentially limited, but still commonly used. The I-T methods are easy to compute and understand and provide formal measures of the strength of evidence for both the null and alternative hypotheses, given the data. We give an example to highlight the importance of deriving alternative hypotheses and representing these as probability models. Fifteen technical issues are addressed to clarify various points that have appeared incorrectly in the recent literature. We offer several remarks regarding the future of empirical science and data analysis under an I-T framework.
.
[本文引用: 1]
.
DOI:10.1016/j.jaridenv.2015.07.002URL [本文引用: 1]
[本文引用: 1]
DOI:10.1093/treephys/24.7.737URLPMID:15123445 [本文引用: 1]
We measured seasonal variation in area-based nitrogen concentration (N), maximum rate of carboxylation (Vcmax) and maximum rate of electron transport (Jmax) in 1-year-old needles along four first-order branches within a Pinus densiflora Sieb. & Zucc. crown, and analyzed their relationships to growth irradiance and temperature. Each leaf light environment was expressed as a ratio of the monthly mean of daily integrated photosynthetically active irradiance (Iint) for the particular needle to Iint above the canopy (Irel). Needle N decreased in the upper crown during the development of new needles, whereas it remained fairly constant in the lower crown, reflecting differences between upper and lower crown needles in their contribution to the nitrogen of new needles. Gradients of N within the crown were correlated with Irel in all seasons (r2 = 0.40-0.78). Seasonal variation in N was weakly correlated with mean daily air minimum temperatures. Both Vcmax and Jmax showed seasonal variation in all first-order branches, and decreased to their lowest values in winter. The gradients of Vcmax and Jmax within the crown were not correlated with Irel in some seasons, but were correlated with changes in N in most months (r2 = 0.33-0.75), except in the winter. Furthermore, the regression slope of the relationship between N and Vcmax and the temperature response of Vcmax and Jmax exhibited seasonal variation.
DOI:10.1016/j.envexpbot.2013.10.017URL [本文引用: 1]
The effect of heat stress on photosystem II (PS II) efficiency and post-stress recovery was studied in four wheat cultivars using chlorophyll fluorescence. The main aim was to examine the cultivar differences in relation to inhibition and recovery of PSII functionality after heat stress at different growth stages. The secondary aim was to investigate whether a pre-acclimation of plants to elevated temperature during the growth period induces a better tolerance to heat stress than for plants grown in ambient temperature or not. The plants were grown in two growth temperature conditions (15 degrees C and 25 degrees C) and subjected to heat stress (40 degrees C) for two days at early tillering and three days at anthesis and early grain development stages. The plants were returned to their original growth conditions after heat stress and recovery was observed for three days. The maximum photochemical efficiency (F-v/F-m) and the quantum yield of PSII (F-q'/F-m') were measured before, during and after the heat stress. The heat stress significantly inhibited the F-v/F-m and F-q'/F-m', in all wheat cultivars at all growth stages. There were significant differences in F-v/F-m among the cultivars at anthesis and at early grain development but not at early tillering stage. However, the cultivars differed significantly in F-q'/F-m' at all growth stages. At anthesis and early grain development, the cultivar C518 had the lowest reduction in F-v/F-m and F-q'/F-m' after heat stress and recovered fully after 72h in both growth conditions illustrating higher heat tolerance characteristics as compared to the other three cultivars. The largest decrease in F-v/F-m and F-q'/F-m', after heat stress occurred in the cultivar PWS7, which did not recover completely after 72 h. All cultivars grown at 25 degrees C had a slightly increased heat tolerance and better recovery compared to plants grown at 15 degrees C. The relative leaf chlorophyll content decreased significantly after heat stress in all cultivars at all growth stages. The elevated growth temperature (25 degrees C) accelerated plant growth resulting in early heading and reduced grain yield in comparison to ambient temperature (15 degrees C). (C) 2013 Elsevier B.V.
.
DOI:10.1111/pce.1992.15.issue-3URL [本文引用: 1]
URL [本文引用: 1]
URL [本文引用: 1]
.
[本文引用: 1]
[本文引用: 1]
.
[本文引用: 1]
[本文引用: 1]
.
DOI:10.1093/treephys/19.11.707URLPMID:12651309 [本文引用: 1]
We analyzed processes limiting photosynthesis in two-year-old, container-grown Pinus pinaster Ait. seedlings subjected to phosphorus (P) deficiency. After withholding P for 3 months, seedlings were supplied P at four relative addition rates (0, 0.005, 0.01 and 0.02 day(-1)) in a nutrient recycling system. At Weeks 12 and 22, responses of photosynthesis to CO(2) and irradiance were measured and the following parameters derived: maximal velocity of carboxylation by Rubisco, V(m); apparent quantum efficiency of electron transport, alpha maximal electron transport rate, J(m); stomatal conductance and relative stomatal limitation of photosynthesis. At Week 22, these measurements were combined with concurrent measurements of chlorophyll fluorescence to determine the quantum yield of PSII, and a theoretical partitioning of total light-driven linear electron flow between fractions used to regenerate carboxylated and oxygenated RuBP. After 12 weeks of treatment, needle P concentrations ranged from 0.04 to 0.15 x 10(-2) g g(DW) (-1), and then remained constant until Week 22. Values of J(m), alpha and V(m) increased with increasing needle P concentration (from 30 to 133 &mgr;mol m(-2) s(-1), 0.02 to 0.25 mol mol(-1) and 13 to 78 &mgr;mol CO(2) m(-2) s(-1) at the lowest and highest needle P concentrations, respectively). Under ambient conditions, net assimilation rates in P-deficient seedlings were limited by V(m) under saturating irradiance, and by J(m) under limiting irradiance, but not by triose-P regeneration. There was no detectable change in the partitioning of total light-driven linear electron flow between the fractions used for carboxylation and oxygenation. Predawn photochemical efficiency of PSII was significantly reduced in seedlings with low P concentrations. Although stomatal conductance tended to decrease with decreasing needle P concentration, relative stomatal limitation was not significantly affected. At Week 22, there was an attenuation of the effects of P nutrition on V(m) and an increase in alpha and J(m) that was probably related to cessation of growth and the seasonal decline in natural irradiance.
.
DOI:10.1016/j.agrformet.2018.09.004URL [本文引用: 2]
.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.5846/stxbURL [本文引用: 1]
DOI:10.5846/stxbURL [本文引用: 1]
DOI:10.1111/j.1399-3054.2011.01488.xURL [本文引用: 1]
Partitioning of excitation energy between photochemical quenching (PQ) and non-photochemical quenching (NPQ) processes is constantly adjusted in the leaf in order to preserve the photosynthetic energy balance. Adjustments in PQ and NPQ often result from a combination of different temporal components that can be simplified into reversible and sustained components. While reversible PQ and NPQ are relatively well understood, the controls behind the sustained components of PQ and NPQ, or the interaction between sustained and reversible NPQ, remain elusive. In this study, I used a full year of high-resolution chlorophyll fluorescence (ChlF) data obtained with a Monitoring-PAM fluorometer (Walz, Effeltrich, Germany) in needles of boreal Pinus sylvestris in situ to quantitatively analyse the dynamics and interaction between temporal components of NPQ and PQ and their control by the environment. To enable the estimation of sustained and reversible components of PQ and NPQ, a number of key ChlF parameters were reviewed and adapted to the analysis of long-term monitoring data. Overall, NPQ was drastically enhanced during winter via the accumulation of sustained NPQ in a process regulated by air temperature. Reversible NPQ retained some functionality even at temperatures well below zero and was not inhibited by the presence of sustained NPQ per se but by low temperatures alone. This suggests that temporal NPQ components co-operate in an additive rather than complementary fashion, conferring additional flexibility to the photoprotective role of NPQ. Finally, the potential of the sustained photochemical quenching parameter (qL(s)) to track photoinhibition in situ was discussed.
DOI:10.1093/treephys/5.2.173URLPMID:14972985 [本文引用: 2]
Photosynthetic utilization of water and nitrogen in Ulmus americana L. seedlings was tightly linked with the relative availability of each resource. During periodic drying cycles, water use efficiency increased as predawn water potential fell from -0.5 to -2.0 MPa. During the later stages of such drying cycles, the relative contribution of stomatal limitations to the total net photosynthetic limitation appeared to be at its greatest, whereas biochemical limitations were predominant in well-watered plants grown under low nitrogen (N) availability. For any level of leaf water status, water use efficiency of photosynthesis (WUE) was always greater in plants with high leaf N content than in plants with low leaf N content. Photosynthetic nitrogen use efficiency (PNUE) was always greater in plants with low leaf N content than in plants with high leaf N content, for any level of water status. In combined N treatments and predawn water status classes, there was a significant inverse relationship between PNUE and WUE.
.
[本文引用: 1]
[本文引用: 1]
.
DOI:10.1126/science.1184946URLPMID:20093461 [本文引用: 1]
.
[本文引用: 1]
[本文引用: 1]
.
URL [本文引用: 1]
In this study,cucumber leaves were under high temperature and strong light stress after two days of spraying with 50~400 μmol·L-1 salicylic acid (SA) solution.The measurements showed that SA pretreatment could inhibit the decrease of intrinsic photochemical efficiency (Fv/Fm),quantum yield of PSII(ΦPSⅡ),maximum fluorescence value(Fm),and photochemical quenching (qP) by 16.1%~30.2%,11.9%~33.0%,7.2%~41.0% and 27.2%~160.8%,respectively,and promote the increase of non-photochemical quenching (NPQ) by 13.1%~62.9%,but had little effect on initial fluorescence value (Fo).SA pretreatment could also inhibit the decrease of xanthophyll cycle pool in cucumber leaves,and increase the deepoxidation extent of xanthophyll cycle (A+Z)/(V+A+Z),which was 29.5% and 24.6% higher than control,respectively.It was suggested that SA could protect the photosynthetic apparatus of cucumber leaves under high temperature and strong light through promoting non-radiative energy dissipation.
URL [本文引用: 1]
In this study,cucumber leaves were under high temperature and strong light stress after two days of spraying with 50~400 μmol·L-1 salicylic acid (SA) solution.The measurements showed that SA pretreatment could inhibit the decrease of intrinsic photochemical efficiency (Fv/Fm),quantum yield of PSII(ΦPSⅡ),maximum fluorescence value(Fm),and photochemical quenching (qP) by 16.1%~30.2%,11.9%~33.0%,7.2%~41.0% and 27.2%~160.8%,respectively,and promote the increase of non-photochemical quenching (NPQ) by 13.1%~62.9%,but had little effect on initial fluorescence value (Fo).SA pretreatment could also inhibit the decrease of xanthophyll cycle pool in cucumber leaves,and increase the deepoxidation extent of xanthophyll cycle (A+Z)/(V+A+Z),which was 29.5% and 24.6% higher than control,respectively.It was suggested that SA could protect the photosynthetic apparatus of cucumber leaves under high temperature and strong light through promoting non-radiative energy dissipation.
.
URL [本文引用: 1]
Leaf photosynthetic characteristics of five tree species, i.e. Dalbergia odorifera, Acrocarpus fraxinifolius, Erythrophleum fordii, Castanopsis hystrix and Betula alnoides seedlings were studied in this paper. The results showed that D. odorifera, B. alnoides and A. fraxinifolius had higher light saturation point (LSP) and light compensation point (LCP) than C. hystrix and E. fordii, so that the former group could make better use of highintensity illumination, while the later group could tolerate shading. High utilization capacity of CO2 (i.e. lower carbon dioxide compensation point (CCP) and higher carboxylation efficiency (CE)) enabled D. odorifera with high photosynthetic capacity. Balanced supply capacity of CO2 (i.e. stomatal conductance (gs) and intercellular CO2 concentration (Ci)) and utilization capacity of CO2 enabled A. fraxinifolius with high photosynthetic capacity too. Although B. alnoides had high capacity of supplying CO2 to photosynthetic apparatus, low capacity of using CO2 limited its photosynthetic capacity. E. fordii and C. hystrix had low supply and utilization capacity of CO2, resulting in their lower photosynthetic capacity than the other tree species.
URL [本文引用: 1]
Leaf photosynthetic characteristics of five tree species, i.e. Dalbergia odorifera, Acrocarpus fraxinifolius, Erythrophleum fordii, Castanopsis hystrix and Betula alnoides seedlings were studied in this paper. The results showed that D. odorifera, B. alnoides and A. fraxinifolius had higher light saturation point (LSP) and light compensation point (LCP) than C. hystrix and E. fordii, so that the former group could make better use of highintensity illumination, while the later group could tolerate shading. High utilization capacity of CO2 (i.e. lower carbon dioxide compensation point (CCP) and higher carboxylation efficiency (CE)) enabled D. odorifera with high photosynthetic capacity. Balanced supply capacity of CO2 (i.e. stomatal conductance (gs) and intercellular CO2 concentration (Ci)) and utilization capacity of CO2 enabled A. fraxinifolius with high photosynthetic capacity too. Although B. alnoides had high capacity of supplying CO2 to photosynthetic apparatus, low capacity of using CO2 limited its photosynthetic capacity. E. fordii and C. hystrix had low supply and utilization capacity of CO2, resulting in their lower photosynthetic capacity than the other tree species.
.
DOI:10.1007/s00442-013-2703-yURL [本文引用: 1]
The sensitivity of carbon (C) assimilation to within-canopy nitrogen (N) allocation and of stomatal conductance (g (s)) to environmental variables were investigated along a vertical canopy gradient in a fertile Norway spruce [Picea abies (L.) Karst.] stand. Maximum rates of ribulose bisphosphate-saturated carboxylation (V (cmax)) and electron transport (J (max)) exhibited weak relationships with needle N content. Using these relationships together with a combined stomatal-photosynthesis model, it was found that the sensitivity of C assimilation of 12 1-year old shoots to within-canopy N allocation pattern was very weak. Modelled C assimilation based on optimal compared to observed N allocation pattern increased by only 1-2 %, and altering total needle N content by +/- 30 % resulted in a 2-4 % change in modelled C assimilation. C assimilation was more sensitive to water use and changed by 8-12 % in response to +/- 30 % altered stomatal conductance. No indications of significant limitations of photosynthesis by other nutrients or non-optimal within-canopy allocation of water were detected. The sensitivity of g (s) to photosynthetic photon flux density (PPFD) was found to be stronger in the lower canopy, while no significant within-canopy variation was observed in light-saturated g (s) or stomatal sensitivity to vapour pressure deficit (VPD). The results of this study show that, at this N rich site, photosynthesis integrated for shoots at different canopy positions is only marginally affected by N allocation pattern and that increased stand-scale N availability would only be truly beneficial to canopy photosynthesis if it resulted in increased leaf area.
.
DOI:10.1093/treephys/tpu036URL [本文引用: 1]
Stem CO2 efflux is known to vary seasonally and vertically along tree stems. However, annual tree- and stand-scale efflux estimates are commonly based on measurements made only a few times a year, during daytime and at breast height. In this study, the effect of these simplifying assumptions on annual efflux estimates and their influence on the estimates of the importance of stems in stand-scale carbon cycling are evaluated. In order to assess the strength of seasonal, diurnal and along-stem variability in CO2 efflux, half-hourly measurements were carried out at three heights on three mature Norway spruce (Picea abies (L.) Karst.) trees over a period of 3 years. Making the common assumption of breast height efflux rates being representative of the entire stem was found to result in underestimations of 10-17% in the annual tree-scale CO2 efflux. Upscaling using only daytime measurements from breast height increased the underestimation to 15-20%. Furthermore, the results show that the strength of the vertical gradient varies seasonally, being strongest in the early summer and non-existent during the cool months. The observed seasonality in the vertical CO2 efflux gradient could not be explained by variation in stem temperature, temperature response of the CO2 efflux (Q(10)), outer-bark permeability, CO2 transport in the xylem or CO2 release from the phloem. However, the estimated CO2 concentration immediately beneath the bark was considerably higher in the upper stem during the main period of diameter growth, coinciding with the strongest vertical efflux gradient. These results suggest that higher growth rates in the upper stem are the main cause for the observed vertical variation in the stem CO2 effluxes. Furthermore, the results indicate that accounting for the vertical efflux variation is essential for assessments of the importance of stems in stand-scale carbon cycling.
.
DOI:10.1111/pce.12565URLPMID:25944258 [本文引用: 1]
Previous leaf-scale studies of carbon assimilation describe short-term resource-use efficiency (RUE) trade-offs where high use efficiency of one resource requires low RUE of another. However, varying resource availabilities may cause long-term RUE trade-offs to differ from the short-term patterns. This may have important implications for understanding canopy-scale resource use and allocation. We used continuous gas exchange measurements collected at five levels within a Norway spruce, Picea abies (L.) karst., canopy over 3 years to assess seasonal differences in the interactions between shoot-scale resource availability (light, water and nitrogen), net photosynthesis (An ) and the use efficiencies of light (LUE), water (WUE) and nitrogen (NUE) for carbon assimilation. The continuous data set was used to develop and evaluate multiple regression models for predicting monthly shoot-scale An . These models showed that shoot-scale An was strongly dependent on light availability and was generally well described with simple one- or two-parameter models. WUE peaked in spring, NUE in summer and LUE in autumn. However, the relative importance of LUE for carbon assimilation increased with canopy depth at all times. Our results suggest that accounting for seasonal and within-canopy trade-offs may be important for RUE-based modelling of canopy carbon uptake.
.
DOI:10.1111/ppl.1982.54.issue-1URL [本文引用: 1]
URL [本文引用: 1]
On the base of survey and analysis of conditions of a expressway roadside slope in central Zhejiang Province,and according to the layer attitude of rocks,its differentiation and attitude of slope,the slope was divided into several sub areas,and the stability of sub areas were evaluated in the paper.Slope fixation and slope revegetation technologies were designed.This paper enumerates,analyses and evaluates practical engineering conditions to demonstrate slope maintenance technology,which have important value for the roadside design.
URL [本文引用: 1]
On the base of survey and analysis of conditions of a expressway roadside slope in central Zhejiang Province,and according to the layer attitude of rocks,its differentiation and attitude of slope,the slope was divided into several sub areas,and the stability of sub areas were evaluated in the paper.Slope fixation and slope revegetation technologies were designed.This paper enumerates,analyses and evaluates practical engineering conditions to demonstrate slope maintenance technology,which have important value for the roadside design.
.
DOI:10.1093/aob/mcg110URLPMID:12740213 [本文引用: 1]
An automatic gas exchange system was used to continuously measure water and carbon fluxes of attached shoots of Scots pine trees (Pinus sylvestris L.) grown in environment-controlled chambers for a 3-year period (1998-2000) and exposed to either normal ambient conditions (CON), elevated CO2 (+350 micro mol mol-1; EC), elevated temperature (+2-6 degrees C; ET) or a combination of EC and ET (ECT). EC treatment enhanced the mean daily total carbon flux per unit projected needle area (Fc.d) by 17-21 %, depending on the year. This corresponds to a 16-24 % increase in light-use efficiency (LUE) based on incident photosynthetically active radiation. The EC treatment reduced the mean daily total water flux (Fw.d) by 1-12 %, corresponding to a 13-35 % increase in water-use efficiency (WUE). The ET treatment increased Fc.d by 10-18 %, resulting in an 8-19 % increase in LUE, and Fw.d by 48-74 %, resulting in a reduction of WUE by 19-34 %. There was no interaction between CO2 and temperature elevation in connection with either carbon or water fluxes, as the carbon flux responded similarly in both ECT and EC, while the water flux in the ECT treatment was similar to that in ET. Regressions indicated that the increase in maximum LUE was greater with increasing air temperature, whereas changes in WUE were related only to high vapour pressure deficit. Furthermore, changes in LUE and WUE caused by ECT treatment displayed strong diurnal and seasonal variation.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
Resource limitation in plants—An economic analogy
1
1985
... 我国干旱半干旱地区占国土总面积的49%, 这一地区分布范围很广, 虽然植物稀少, 但基于如此广阔的分布面积, 其对碳循环和碳收支的贡献不容忽略(
1
... 式中a, b, c和d是常数, Pn的单位为μmol·m-2·s-1, E的单位为mmol·m-2·s-1, PAR的单位是μmol·m-2·s-1, Narea的单位是g·m-2, T的单位是℃, intercept为残差.基于拟合的R2和Akaike信息标准(AIC)来评估模型的性能(
AIC model selection and multimodel inference in behavioral ecology: Some background, observations, and comparisons
1
2011
... R2描述了回归模型的整体拟合程度, 而AIC值用来评价多个回归模型各自的优良程度(
Water use efficiency and photosynthesis
1
2004
... 我国干旱半干旱地区占国土总面积的49%, 这一地区分布范围很广, 虽然植物稀少, 但基于如此广阔的分布面积, 其对碳循环和碳收支的贡献不容忽略(
Leaf nitrogen is closely coupled to phenophases in a desert shrub ecosystem in China
1
2015
... 油蒿(Artemisia ordosica)为菊科蒿属, 半干旱区典型沙生植物, 近年来许多相关研究多是在控制试验条件下进行的(
1
1977
... NUE、WUE和LUE之间的相对变化可能会由于观测时间尺度的不同而表现出不同的模式.在叶水平, 有研究(
Spatial and seasonal variability of temperature responses of biochemical photosynthesis parameters and leaf nitrogen content within a Pinus densiflora crown
1
2004
... 植物的净光合能力与PAR的关系非常密切(
Heat stress and recovery of photosystem II efficiency in wheat (Triticum aestivum L.) cultivars acclimated to different growth temperatures
1
2014
... 由于全球气候变化, 增加了干旱半干旱地区极端天气的出现频率和强度(
Modelling photosynthesis of cotton grown in elevated CO2
1
1992
... 式中, PC为光合系统中Rubisco组分内的氮占叶片总氮的比例; PB为光合系统生物力能学组分内的氮占叶片总氮的比例; PL为光合系统中捕光组分内的氮占叶片总氮的比例.植物叶片中分配给光合系统的氮占叶片总氮的比例(PP)即为以上3个比例之和.Narea为单位面积叶片氮含量, 即单位质量叶片氮含量与比叶面积的比值; Vcmax单位为μmol·m-2·s-1; Jmax单位为μmol·m-2·s-1; CC为叶片叶绿素浓度(mmol·g-1); Vcr是Rubisco酶的比活, 即单位Rubisco组分中的CO2固定活性(μmol CO2·g-1 Rubisco·s-1); Jmc为单位细胞色素f (Cytochrome f, Cyt f)的最大电子传递速率(μmol electrons·μmol-1 Cyt f·s-1).Vcr和Jmc的计算方法如下(
陕北毛乌素沙漠黑沙蒿根系分布特征研究
1
2010
... 由于全球气候变化, 增加了干旱半干旱地区极端天气的出现频率和强度(
陕北毛乌素沙漠黑沙蒿根系分布特征研究
1
2010
... 由于全球气候变化, 增加了干旱半干旱地区极端天气的出现频率和强度(
干热河谷不同土壤水分下甜玉米灌浆期光合作用光响应特征
1
2018
... 油蒿WUE季节变异的影响因子有T与VPD, WUE与其呈显著负相关关系, 与土壤含水量的相关关系不显著.T与VPD主要是通过影响gs进而影响WUE.温度的升高对植物的气孔有限制作用, 随着T的不断升高, 增大了叶片内外的水汽压差, 植物的蒸腾会变得强烈, 导致叶水势下降, 更加剧了蒸腾的变化, WUE也随之减小(
干热河谷不同土壤水分下甜玉米灌浆期光合作用光响应特征
1
2018
... 油蒿WUE季节变异的影响因子有T与VPD, WUE与其呈显著负相关关系, 与土壤含水量的相关关系不显著.T与VPD主要是通过影响gs进而影响WUE.温度的升高对植物的气孔有限制作用, 随着T的不断升高, 增大了叶片内外的水汽压差, 植物的蒸腾会变得强烈, 导致叶水势下降, 更加剧了蒸腾的变化, WUE也随之减小(
不同土壤水分条件下硅对紫花苜蓿水分利用效率及产量构成要素的影响
1
2009
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(
不同土壤水分条件下硅对紫花苜蓿水分利用效率及产量构成要素的影响
1
2009
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(
Photosynthetic responses to phosphorus nutrition in two-year- old maritime pine seedlings
1
1999
... 最大电子传递速率(Jmax)通过
Energy and water vapor exchange over a young plantation in northern China
2
2018
... 油蒿WUE季节变异的影响因子有T与VPD, WUE与其呈显著负相关关系, 与土壤含水量的相关关系不显著.T与VPD主要是通过影响gs进而影响WUE.温度的升高对植物的气孔有限制作用, 随着T的不断升高, 增大了叶片内外的水汽压差, 植物的蒸腾会变得强烈, 导致叶水势下降, 更加剧了蒸腾的变化, WUE也随之减小(
... NUE、WUE和LUE之间的相对变化可能会由于观测时间尺度的不同而表现出不同的模式.在叶水平, 有研究(
Temperature response of parameters of a biochemically based model of photosynthesis. II. A review of experimental data
1
2002
... 植物的净光合能力与PAR的关系非常密切(
彰武松、樟子松光合生产与蒸腾耗水特性
1
2012
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(
彰武松、樟子松光合生产与蒸腾耗水特性
1
2012
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(
A model separating leaf structural and physiological effects on carbon gain along light gradients for the shade-tolerant species Acer saccharum.
1
1997
... 植物叶片光合系统内氮分配比例的各组分可利用
土壤干旱胁迫对沙棘叶片光合作用和抗氧化酶活性的影响
1
2013
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(
土壤干旱胁迫对沙棘叶片光合作用和抗氧化酶活性的影响
1
2013
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(
A high-resolution portrait of the annual dynamics of photochemical and non-photochemical quenching in needles of Pinus sylvestris.
1
2011
... 由于全球气候变化, 增加了干旱半干旱地区极端天气的出现频率和强度(
Response of Ulmus americana seedlings to varying nitrogen and water status. 2. Water and nitrogen use efficiency in photosynthesis.
2
1989
... 油蒿(Artemisia ordosica)为菊科蒿属, 半干旱区典型沙生植物, 近年来许多相关研究多是在控制试验条件下进行的(
... NUE、WUE和LUE之间的相对变化可能会由于观测时间尺度的不同而表现出不同的模式.在叶水平, 有研究(
黄土丘陵区沙棘气孔导度及其影响因子
1
2001
... 油蒿WUE季节变异的影响因子有T与VPD, WUE与其呈显著负相关关系, 与土壤含水量的相关关系不显著.T与VPD主要是通过影响gs进而影响WUE.温度的升高对植物的气孔有限制作用, 随着T的不断升高, 增大了叶片内外的水汽压差, 植物的蒸腾会变得强烈, 导致叶水势下降, 更加剧了蒸腾的变化, WUE也随之减小(
黄土丘陵区沙棘气孔导度及其影响因子
1
2001
... 油蒿WUE季节变异的影响因子有T与VPD, WUE与其呈显著负相关关系, 与土壤含水量的相关关系不显著.T与VPD主要是通过影响gs进而影响WUE.温度的升高对植物的气孔有限制作用, 随着T的不断升高, 增大了叶片内外的水汽压差, 植物的蒸腾会变得强烈, 导致叶水势下降, 更加剧了蒸腾的变化, WUE也随之减小(
Drylands in the earth system
1
2010
... 由于全球气候变化, 增加了干旱半干旱地区极端天气的出现频率和强度(
植物叶片氮分配及其影响因子研究进展
1
2015
... CB为光合系统捕光组分中叶绿素与氮的比值(mmol Chl·(g·N)-1), 可以利用CB与比叶重(LMA)关系的线性公式来进行计算, 其相关系数较高(R2 = 0.97), 并且证明可以在多种植物类型中广泛应用(
植物叶片氮分配及其影响因子研究进展
1
2015
... CB为光合系统捕光组分中叶绿素与氮的比值(mmol Chl·(g·N)-1), 可以利用CB与比叶重(LMA)关系的线性公式来进行计算, 其相关系数较高(R2 = 0.97), 并且证明可以在多种植物类型中广泛应用(
高温强光下水杨酸对黄瓜叶片叶绿素荧光和叶黄素循环的影响
1
2006
... 油蒿WUE季节变异的影响因子有T与VPD, WUE与其呈显著负相关关系, 与土壤含水量的相关关系不显著.T与VPD主要是通过影响gs进而影响WUE.温度的升高对植物的气孔有限制作用, 随着T的不断升高, 增大了叶片内外的水汽压差, 植物的蒸腾会变得强烈, 导致叶水势下降, 更加剧了蒸腾的变化, WUE也随之减小(
高温强光下水杨酸对黄瓜叶片叶绿素荧光和叶黄素循环的影响
1
2006
... 油蒿WUE季节变异的影响因子有T与VPD, WUE与其呈显著负相关关系, 与土壤含水量的相关关系不显著.T与VPD主要是通过影响gs进而影响WUE.温度的升高对植物的气孔有限制作用, 随着T的不断升高, 增大了叶片内外的水汽压差, 植物的蒸腾会变得强烈, 导致叶水势下降, 更加剧了蒸腾的变化, WUE也随之减小(
南亚热带五种树种幼苗光合特征
1
2016
... 油蒿NUE季节变异的影响因子有T与VPD, NUE与其都呈显著负相关关系, 与土壤含水量的相关关系不显著(
南亚热带五种树种幼苗光合特征
1
2016
... 油蒿NUE季节变异的影响因子有T与VPD, NUE与其都呈显著负相关关系, 与土壤含水量的相关关系不显著(
Weak vertical canopy gradients of photosynthetic capacities and stomatal responses in a fertile Norway spruce stand
1
2013
... 植物的净光合能力与PAR的关系非常密切(
Vertical gradients and seasonal variation in stem CO2 efflux within a Norway spruce stand
1
2014
... NUE、WUE和LUE之间的相对变化可能会由于观测时间尺度的不同而表现出不同的模式.在叶水平, 有研究(
Seasonal and within-canopy variation in shoot-scale resource-use efficiency trade-offs in a Norway spruce stand
1
2015
... 因为每个月可用的数据点数量比较少, Pn与环境因子的关系描述为简单的线性回归关系, 并且在所有分析中没有包含交互条件, 参考已有的研究(
Gas exchange in a 20-year-old stand of scots pine. I. Net photosynthesis of current and one-year-old shoots within and between seasons
1
1982
... 油蒿NUE季节变异的影响因子有T与VPD, NUE与其都呈显著负相关关系, 与土壤含水量的相关关系不显著(
银中杨光合作用和蒸腾作用对土壤干旱的响应
1
2006
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(
银中杨光合作用和蒸腾作用对土壤干旱的响应
1
2006
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(
Light and water- use efficiencies of pine shoots exposed to elevated carbon dioxide and temperature
1
2003
... 植物的净光合能力与PAR的关系非常密切(
塔里木河下游刚毛柽柳光合作用、蒸腾作用及水分利用效率特性研究
1
2015
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(
塔里木河下游刚毛柽柳光合作用、蒸腾作用及水分利用效率特性研究
1
2015
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(
荒漠植物白麻气孔导度特征及其影响因子研究
1
2013
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(
荒漠植物白麻气孔导度特征及其影响因子研究
1
2013
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(
光辐射强度对侧柏油松幼苗光合特性与水分利用效率的影响
1
2006
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(
光辐射强度对侧柏油松幼苗光合特性与水分利用效率的影响
1
2006
... 植物叶片氮(N)含量、水分及光条件在干旱地区是影响植物光合蒸腾过程、制约植物生长发育的关键因素(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}