<script type="text/javascript" src="https://cdn.bootcss.com/mathjax/2.7.2-beta.0/MathJax.js?config=TeX-AMS-MML_HTMLorMML"></script>
<script type='text/x-mathjax-config'>
MathJax.Hub.Config({
extensions: ["tex2jax.js"],
jax: ["input/TeX", "output/HTML-CSS"],
tex2jax: {inlineMath: [ ['$','$'], ["\\(","\\)"] ],displayMath: [ ['$$','$$'], ["\\[","\\]"] ],processEscapes: true},
"HTML-CSS": { availableFonts: ["TeX"] },
TeX: {equationNumbers: {autoNumber: ["none"], useLabelIds: true}},
"HTML-CSS": {linebreaks: {automatic: true}},
SVG: {linebreaks: {automatic: true}}
});
</script>
陈婵
1 , 张仕吉
1 , 李雷达
1 , 刘兆丹
1 , 陈金磊
1 , 辜翔
1 , 王留芳
1 , 方晰
, , 1 , 2 , * 1 中南林业科技大学生命科学与技术学院, 长沙 4100042 湖南会同杉木林生态系统国家野外科学观测研究站, 湖南会同 438107Carbon, nitrogen and phosphorus stoichiometry in leaf, litter and soil at different vegetation restoration stages in the mid-subtropical region of China Chan CHEN
1 , Shi-Ji ZHANG
1 , Lei-Da LI
1 , Zhao-Dan LIU
1 , Jin-Lei CHEN
1 , Xiang GU
1 , Liu-Fang WANG
1 , Xi FANG
, , 1 , 2 , * 1 School of Life Science and Technology, Central South University of Forestry and Technology, Changsha 410004, China 2 Huitong National Field Station for Scientific Observation and Research of Chinese Fir Plantation Ecosystem in Hunan Province, Huitong, Hunan 438107, China 通讯作者: * 方晰: ORCID: 0000-0003-1236-2335, (
fangxizhang@sina.com )
编委: 曾波
责任编辑: 李敏, 赵航(实习)
收稿日期: 2019-01-22
修回日期: 2019-08-6
网络出版日期: 2019-08-20
基金资助: 国家林业公益性行业科研专项 (201504411 )国家自然科学基金 (31670448 )
Corresponding authors: * fangxizhang@sina.com Received: 2019-01-22
Revised: 2019-08-6
Online: 2019-08-20
Fund supported: National Forestry Public Welfare Industry Research Project (201504411 )National Natural Science Foundation of China (31670448 )
摘要 为揭示植被恢复过程中生态系统的养分循环机制及植物的生存策略, 根据亚热带森林群落演替过程, 采用空间代替时间方法, 以湘中丘陵区地域相邻、环境条件基本一致的檵木(
Loropetalum chinensis ) +南烛(
Vaccinium bracteatu ) +杜鹃(
Rhododendron mariesii )灌草丛(LVR)、檵木+杉木(
Cunninghamia lanceolata ) +白栎(
Quercus fabri )灌木林(LCQ)、马尾松(
Pinus massoniana ) +柯(
Lithocarpus glaber ) +檵木针阔混交林(PLL)、柯+红淡比(
Cleyera japonica ) +青冈(
Cyclobalanopsis Glauca )常绿阔叶林(LCC)作为一个恢复系列, 设置固定样地, 采集植物叶片、未分解层凋落物和0-30 cm土壤样品, 测定有机碳(C)、全氮(N)、全磷(P)含量及其化学计量比, 运用异速生长关系、养分利用效率和再吸收效率分析植物对环境变化的响应和养分利用策略。结果表明: (1)随着植被恢复, 叶片C:N、C:P、N:P显著下降, 而叶片C、N、P含量和土壤C、N含量、C:P、N:P显著增加, 其中LCC植物叶片C、N含量, 土壤C、N含量及其N:P, PLL植物叶片P含量, 土壤C:P显著高于其他3个恢复阶段, 各恢复阶段植物叶片N:P > 20, 植物生长受P限制; 凋落物C、N、P含量及其化学计量比波动较大。(2)凋落物与叶片、土壤的化学计量特征之间的相关关系较弱, 叶片与土壤的化学计量特征之间具有显著相关关系, 其中叶片C、N、P含量与土壤C、N含量、C:N (除叶片C、N含量外)、C:P、N:P呈显著正相关关系; 叶片C:N与土壤C、N含量、C:P、N:P, 叶片C:P与土壤C含量、C:N、C:P, 叶片N:P与土壤C:N呈显著负相关关系。(3)植被恢复过程中, 叶片N、P之间具有显著异速生长关系, 异速生长指数为1.45, 叶片N、P的利用效率下降, 对N、P的再吸收效率增加, LCC叶片N利用效率最低, PLL叶片P利用效率最低而N、P再吸收效率最高。(4)叶片N含量内稳态弱, 而P含量具有较高的内稳态, 在土壤低P限制下植物能保持P平衡。植被恢复显著影响叶片、凋落物、土壤C、N、P含量及其化学计量比, 叶片与土壤之间C、N、P含量及化学计量比呈显著相关关系, 植物通过降低养分利用效率和提高养分再吸收效率适应土壤养分的变化, 叶片-凋落物-土壤系统的N、P循环随着植被恢复逐渐达到“化学计量平衡”。
关键词: 湘中丘陵区 ;
植被恢复 ;
碳氮磷化学计量比 ;
内稳态 ;
养分利用效率 ;
氮磷再吸收效率 Abstract Aims The aims of this study were to explore how vegetation restoration affects leaf, litter and soil C, N, P stoichiometry dynamics and nutrients cycling, and to characterize the homeostasis and nutrient use strategy of plants at different vegetation restoration stages in the mid-subtropical area of China.
Methods Four vegetation types representing the successional sequence in the secondary forests were selected using the “space for time substitution” approach in central hilly area of Hunan Province, China, which consists of
Loropetalum chinense +
Vaccinium bracteatum +
Rhododendron simsii scrub-grass-land (LVR),
Loropetalum chinense +
Cunninghamia lanceolata +
Quercus fabri shrubbery (LCQ),
Pinus massoniana +
Lithocarpus glaber +
Loropetalum chinense coniferous-broad leaved mixed forest (PLL), and
Lithocarpus glaber +
Cleyera japonica +
Cyclobalanopsis glauca evergreen broad-leaved forest (LCC). Permanent plots were established in each community. The organic carbon (C), total nitrogen (N) and total phosphorus (P) contents in leaf, undecomposed litter layer and 0-30 cm soil layer were quantified at each stage. The response and nutrient use strategy of plant to environmental changes were estimated by allometric growth, nutrient use efficiency and nutrient reabsorption efficiency.
Important findings 1) Along vegetation restoration, the leaf C:N, C:P ratios decreased significantly and the highest values were in LVR. Leaf C, N, P contents, soil C, N contents and soil C:N, C:P, N:P ratios increased significantly, in which leaf C, N contents and soil C, N contents, N:P in LCC were higher than those in LVR, LCQ and PLL, and leaf P content and soil C:N, C:P in PLL were higher than those in LVR, LCQ and LCC. Leaf N:P (>20) indicated that all restoration stages were P limited. C, N, P contents and their stoichiometry of litter fluctuated greatly. 2) The relationships between litter and leaf or soil nutrients and their stoichiometry were weak, and the significant correlations were found in the relationships between leaf and soil nutrients and their stoichiometry. Leaf C, N and P were positively correlated with soil C, N, C:N (except leaf C, N contents), C:P and N:P, while leaf C:N was negatively correlated with soil C, N, C:P and N:P, leaf C:P was negatively correlated with soil C content, C:N and C:P, and leaf N:P were negatively correlated with soil C:N. 3) During vegetation restoration, leaf N and P had significantly allometric growth relationship (
p < 0.01) with the allometric index being 1.45. The use efficiency of N and P nutrients in leaf showed decreasing trends and reabsorption efficiency showed increasing trends, and the lowest N use efficiency was observed in LCC and the lowest P use efficiency was in PLL, but the highest N, P reabsorption efficiency were both in PLL. 4) The leaf N content had weak homeostasis, and leaf P content had strong homeostasis to maintain P balance in plant under P limited in soil. Vegetation restoration had significant effects on leaf, litter and soil C, N, P contents and their stoichiometry. The C, N, P contents and their stoichiometry had significant correlations between leaf and soil. Plants could adapt to the shortage of soil nutrient supply mainly by reducing nutrient use efficiency and improving nutrient reabsorption capacity. The N and P cycles of the leaf-litter-soil system gradually reached the “stoichiometric equilibrium” during vegetation restoration.
Keywords: central hilly area of Hunan Province ;
vegetation restoration ;
carbon ;
nitrogen and phosphorus stoichiometry ratio ;
homeostasis ;
nutrient use efficiency ;
nitrogen and phosphorus reabsorption efficiency PDF (1358KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 陈婵, 张仕吉, 李雷达, 刘兆丹, 陈金磊, 辜翔, 王留芳, 方晰. 中亚热带植被恢复阶段植物叶片、凋落物、土壤碳氮磷化学计量特征. 植物生态学报, 2019, 43(8): 658-671. DOI:
10.17521/cjpe.2019.0018 CHEN Chan, ZHANG Shi-Ji, LI Lei-Da, LIU Zhao-Dan, CHEN Jin-Lei, GU Xiang, WANG Liu-Fang, FANG Xi.
Carbon, nitrogen and phosphorus stoichiometry in leaf, litter and soil at different vegetation restoration stages in the mid-subtropical region of China .
Chinese Journal of Plant Ecology , 2019, 43(8): 658-671. DOI:
10.17521/cjpe.2019.0018 植被恢复涉及植物群落与土壤环境的协调发展, 因其能改善脆弱或退化生态系统养分循环、土壤质量而受到广泛关注(
Xu et al ., 2019 )。随着植被恢复, 植物群落组成结构、土壤理化性质达到稳定状态(
O’Brien & Jastrow, 2013 ), 生态系统各组分(植物、凋落物、土壤)碳(C)、氮(N)、磷(P)含量及其循环规律调控着植物的生存发展和各种生态过程(
Reich et al ., 2006 ), 其化学计量比反映植物组成动态与土壤养分之间的平衡(
Zhang et al ., 2018 )。因此, 研究生态系统各组分C、N、P化学计量比随植被恢复的变化对理解各组分之间养分关系及分配特征十分必要, 有助于明确植被恢复过程中, 植物群落随土壤环境变化的发展方向(
Zhao et al ., 2015b ;
Ren et al ., 2016 )。
土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略。由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 )。随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b )。由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 )。基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 )。此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 )。随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a )。凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 )。植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比。同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 )。然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a )。
中国亚热带地区水热条件优越, 森林资源丰富, 是中国重要的生态屏障。但由于长期的人为干扰, 该地区地带性植被——常绿阔叶林遭到了严重的破坏。为保护生物多样性和提升森林生态系统服务, 促进森林植被恢复, 近20多年来, 中国政府实施了天然林保护、长江中上游防护林体系建设等系列林业生态工程, 该地区森林植被恢复迅速, 由于人为干扰强度不同, 形成了处于不同恢复阶段的多种次生林群落(
Xiang et al ., 2013 )。目前针对亚热带森林植被恢复过程中, 不同植物组织(
阎恩荣等, 2010 ;
Zeng et al ., 2017b )、土壤(
彭晓等, 2016 ;
Xu et al ., 2018 )的化学计量动态已有一些研究报道。然而, 将植物叶片-凋落物-土壤作为一个系统, 探讨其C、N、P化学计量动态及其各组分之间相互作用的研究鲜有报道, 植被恢复对整个生态系统养分运输和分配影响如何仍然不是很清楚。为此, 本研究根据亚热带森林群落演替进程, 在湘中丘陵区选取了4种处于不同恢复阶段(灌草丛、灌木林、针阔混交林、常绿阔叶林)的生态系统作为一个恢复序列, 研究植物叶片、凋落物、土壤C、N、P含量、化学计量比的变化及其相互之间的关联性, 探讨随植被恢复, 植物群落对土壤环境变化的响应及其养分利用策略, 剖析生态系统各组分之间养分动态平衡关系随着植被恢复的变化特征, 为亚热带森林植被恢复和养分管理提供科学依据。
1 材料和方法 1.1 研究地概况 研究地设置在湖南省中东部的长沙县(28.38°- 28.40° N, 113.28°-113.45° E), 地处幕连九山脉中支连云山山脉的余脉, 海拔为55-350 m, 属于典型的低山丘陵地貌。属于亚热带东南季风湿润气候, 季节变化明显, 年平均气温17.3 ℃, 7-8月极端高温39.8 ℃, 1月极端低温-10.3 ℃, 年降水量为1 412-1 559 mm, 主要分布在4-7月。土壤以板岩、页岩发育而成的红壤为主, 地带性植被为亚热带常绿阔叶林, 由于人为干扰(采伐、火烧、放牧)频繁, 强度各异, 原生地带性森林植被破坏严重。20世纪90年代初实施封山育林和退耕还林, 形成了处于不同恢复阶段的多种次生林群落(灌草丛、灌木林、马尾松(
Pinus massoniana )针阔混交林、常绿阔叶林)和以杉木(
Cunninghamia lanceolata )、马尾松、湿地松(
Pinus elliottii )为主的人工林群落, 为开展亚热带植被恢复研究提供了良好的场所。
1.2 样地设置和群落调查 采用空间代替时间的方法, 按照亚热带森林群落自然演替进程, 根据植被恢复程度及其群落树种组成, 选取地域相邻、环境条件(立地、气候和土壤)相似, 处于不同恢复阶段的4种生态系统: (1)檵木(
Loropetalum chinensis ) +南烛(
Vaccinium bracteatum ) +杜鹃(
Rhododendron mariesii )灌草丛(LVR): 1965年砍伐常绿阔叶林后, 炼山、人工整地, 1966年营造马尾松人工林, 无施肥历史, 1990年皆伐马尾松人工林。2012年, 停止各种人为干扰(砍杂、放牧), 让其自然恢复, 当前草本植物多且生长良好, 伴随分布许多幼小灌木植物, 属于恢复早期阶段。(2)檵木+杉木+白栎(
Quercus fabri )灌木林(LCQ): 1965年天然常绿阔叶林采伐后, 炼山、人工整地, 1966年营造杉木人工纯林, 无施肥历史, 1989年皆伐杉木人工纯林。2004年停止各种人为干扰, 让其自然恢复, 灌木植物发育良好, 但没有形成明显的乔木层, 草本植物稀少, 属于恢复中期第I阶段。(3)马尾松+柯(
Lithocarpus glaber ) +檵木针阔混交林(PLL): 20世纪60年代末, 天然常绿阔叶林皆伐后, 让其自然恢复, 目前已恢复为针阔混交林, 年龄为45-50年, 群落内幼苗幼树丰富, 植物密度较大, 但大直径个体所占比例相对较低, 属于恢复中期第II阶段。(4)柯+红淡比(
Cleyera japonica ) +青冈(
Cyclobalanopsis Glauca )常绿阔叶林(LCC): 长期以来无明显人为干扰, 保存比较完好, 群落结构相对稳定, 年龄为80-90年, 属于恢复后期(亚顶极)阶段。在LVR、LCQ各设置4块20 m × 20 m固定样地, 在PLL、LCC分别设置4块30 m × 30 m的固定样地。群落调查方法和4种群落的基本特征及其主要树种组成详见
辜翔等(2018) 的文献。
1.3 分析样品的采集 为了避免偶然性, 分别在2016年4、6、10月下旬采集植物叶片、凋落物、0-30 cm土层分析样品, 取3次采样测定的平均值作为最终的数据。在LVR固定样地, 沿着对角线均匀设置3个1 m × 1 m样方, 采集样方内所有灌木叶片、草本植物地上部分, 将同一样地3个样方植物叶片等比例混合为1个样品。根据群落调查结果(
辜翔等, 2018 ), 确定LCQ、PLL和LCC每块固定样地的优势树种, 对该固定样地的每种优势树种分别选取9株长势良好的样木。每种优势树种每次采集3株样木的叶片, 按上、中、下3个部位的东、西、南、北4个方向采集各株样木已完全成熟、无虫害的叶片, 草本植物地上部分采集同LVR, 再将同一样地内草本植物和优势树种叶片按生物量折算比例混合为1个样品。按
郑路和卢立华(2012) 的凋落物层分层标准, 在完成了草本植物地上部分样品采集后的3个1 m × 1 m的样方内采集未分解层的凋落物样品。完成凋落物采集后, 将样方内剩余凋落物清理干净, 用土钻(内径10 cm)取0-30 cm土层土壤样品, 将同一样地3个样方的凋落物、土壤分别混合为1个样品。所有的分析样品每次采样4个重复, 3次采样12个重复。
1.4 样品处理和分析 叶片、凋落物样品置于80 ℃恒温下烘干, 用植物粉碎机粉碎, 过0.25 mm筛后保存备用。土壤样品去除石砾、动植物残体后室内自然风干, 磨碎过0.25 mm筛后保存备用。
植物叶片、凋落物、土壤样品有机碳含量用重铬酸钾-浓硫酸滴定法测定, 全氮含量用K9840自动凯氏定氮仪(海能仪器公司, 济南)测定, 全磷含量用碱熔-钼锑抗比色法测定(
中国科学院南京土壤研究所, 1978 )。
1.5 数据统计分析 C、N、P含量用单位质量的含量(g·kg
-1 )表示, C:N、C:P、N:P为质量比。用Excel 2010统计各项指标的平均值和标准偏差, 用SigmaPlot 12.0制图。用SPSS 19.0单因素方差分析进行显著性(
p < 0.05)检验。用R 3.5.1中Corrplot包绘制叶片、凋落物、土壤C、N、P含量及其化学计量比的Pearson相关性图。用R 3.5.1中Smart包拟合叶片N、P含量的标准化主轴回归(SMA)关系, 即异速生长关系(
Warton et al ., 2012 ;
R Development Core Team, 2018 )。
(1)异速生长反映植物生长发育过程中有机体内两种属性(例如C、N、P含量)之间的变化规律, 常用公式(
Niklas, 1994 )如下:
(1) $y=a{{x}^{b}}$ 式中,
x、y 是生物属性,
a、b 均为常数。在实际研究中, 异速生长关系函数通常以对数形式表示, 公式(
韩文轩和方精云, 2008 )如下:
(2) $\log (y)=b\log (x)+\log (a)$ 式中, log(
a )为直线的截距,
b 为斜率, 即异速生长指数。
(2)养分利用效率(
NUE )反映植物对土壤养分(如N、P)的利用和适应状况。养分利用率高表明植物能通过较低的养分吸收量和存留量生产更多生物量, 是植物适应贫瘠土壤环境的重要竞争策略(
Grime, 2001 )。本研究采用Chapin指数(
Chapin III, 1980 )计算, 公式如下:
(3) $NUE=\frac{M}{{{A}_{i}}}=\frac{M}{M\times {{C}_{i}}}=\frac{1}{{{C}_{i}}}$ 式中,
NUE 为养分利用效率,
M 为植被生物量(kg·hm
-2 ),
A i 为植物
i 养分贮量(kg·hm
-2 ),
Ci 为植物
i 养分含量(g·kg
-1 )。
(3)养分再吸收效率(
NRE )是指养分从凋落物中转移并被新生组织再利用的效率, 叶片N、P的再吸收效率计算公式(
Milla et al ., 2005 )如下:
(4) $NRE=\frac{({{C}_{\text{L}}}-{{C}_{\text{F}}})}{{{C}_{\text{L}}}}\times 100$ 式中,
NRE 为养分再吸收效率(%),
C L 为植物叶片N (或P)含量(g·kg
-1 ),
C F 为凋落物N(或P)含量(g·kg
-1 )。
(4)植物内稳态反映植物随着环境变化保持自身化学组成稳定的能力(
Kooijman, 1995 ), 常用以下线性回归方程拟合:
(5) $y=kx+c$ 式中,
x 为土壤C、N、P含量(g·kg
-1 ),
y 为植物C、N、P含量(g·kg
-1 ),
k 为斜率,
c 为常数。当
k ≠ 0时, 表示植物体该养分含量内稳态弱, 随土壤环境变化而变化; 当
k = 0时, 则表示植物体该养分具有绝对的内稳态, 不受土壤环境变化的影响(
Sterner & Elser, 2002 )。
2 结果和分析 2.1 叶片、凋落物、土壤C、N、P含量及其化学计量比 植被恢复对植物叶片、凋落物、土壤C、N、P含量及其化学计量比影响显著(
图1 )。随着植被恢复, 叶片、凋落物、土壤C含量增加。LCC叶片C含量最高((435.81 ± 17.21) g·kg
-1 ), LVR最低((347.83 ± 28.11) g·kg
-1 ), 且LVR与LCQ、PLL、LCC差异显著(
p < 0.05); PLL凋落物C含量最高((507.10 ± 31.22) g·kg
-1 ), 与LVR ((387.08 ± 60.15) g·kg
-1 )、LCQ ((344.58 ± 67.00) g·kg
-1 )差异显著(
p < 0.05); 土壤C含量从LVR的(7.59 ± 2.57) g·kg
-1 增加到LCC的(23.99 ± 4.16) g·kg
-1 , 且不同恢复阶段之间差异显著(
p < 0.05)(
图1 A)。叶片、凋落物、土壤N含量总体上随着植被恢复而增加, 其中叶片、土壤N含量不同恢复阶段之间差异显著(
p < 0.05), LCC凋落物N含量与LVR、LCQ、PLL差异显著(
p < 0.05)(
图1 B)。从LVR到LCC, 叶片、凋落物、土壤P含量最大值分别出现在PLL ((0.74 ± 0.06) g·kg
-1 ), LCQ ((0.50 ± 0.10) g·kg
-1 ), LCC ((0.19 ± 0.02) g·kg
-1 ), 且叶片、凋落物、土壤P含量不同恢复阶段之间差异显著(
p < 0.05)(
图1 C)。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1中亚热带不同恢复阶段植物叶片、凋落物、土壤C (A)、N (B)、P (C)含量及C:N (D)、C:P (E)、N:P (F)(平均值±标准偏差, n = 12)。LCC, 柯+红淡比+青冈常绿阔叶林; LCQ, 檵木+杉木+白栎灌木林; LVR, 檵木+南烛+杜鹃灌草丛; PLL, 马尾松+柯+檵木针阔混交林。不同小写字母代表同一组分不同恢复阶段之间差异显著(p < 0.05)。 Fig. 1Contents of C (A), N (B) and P (C) and the C:N (D), C:P (E), N:P (F) in leaf, litter and soil at different restoration stages in the mid-subtropical region of China (mean ± SD , n = 12). LCC, Lithocarpus glaber + Cleyera japonica + Cyclobalanopsis glauca evergreen broad-leaved forest; LCQ, Loropetalum chinense + Cunninghamia lanceolata + Quercus fabri shrubbery; LVR, Loropetalum chinense + Vaccinium bracteatum + Rhododendron mariesii scrub-grass-land; PLL, Pinus massoniana + Lithocarpus glaber + Loropetalum chinense coniferous-broad leaved mixed forest. Different lowercase letters indicate significant differences at different restoration stages for the same component (p < 0.05). 叶片、凋落物、土壤C:N总体上随植被恢复呈下降趋势, LVR叶片C:N最大(42.06 ± 4.57), 显著高于LCQ、PLL、LCC (
p < 0.05); PLL凋落物(39.45 ± 10.14)和土壤(18.71 ± 2.35) C:N最大, 显著高于LVR、LCQ、LCC (
p < 0.05)(
图1D )。随着植被恢复, 叶片、凋落物C:P先下降后增加, LVR叶片(924.31 ± 333.04)最高, PLL最低(582.99 ± 65.84), 且LVR、LCQ、LCC与PLL之间, LVR与LCQ、LCC之间差异显著(
p < 0.05); PLL凋落物C:P最高(1 641.40 ± 308.75), LCQ最低(720.00 ± 218.04), PLL显著高于其他恢复阶段(
p < 0.05), LVR、LCC与LCQ之间差异显著(
p < 0.05); 土壤C:P总体上呈增加趋势, PLL (155.24 ± 46.85)显著高于LVR、LCQ (
p < 0.05)(
图1E )。随着植被恢复, 叶片、凋落物N:P的变化与其C:P基本一致, PLL叶片N:P最低(21.51 ± 1.04), 且与LVR、LCC差异显著(
p < 0.05); LCQ凋落物最低(30.56 ± 6.64), 但不同恢复阶段之间差异不显著(
p > 0.05); 土壤N:P呈增加趋势, 且LVR (4.51 ± 2.58)与LCQ、PLL、LCC差异显著(
p < 0.05)(
图1F )。
2.2 植物叶片、凋落物、土壤C、N、P及其化学计量比的相关关系 Pearson相关性分析结果(
图2 )表明, 叶片C含量与凋落物N含量, 叶片N含量与凋落物C、N含量、N:P, 叶片P含量与凋落物C含量、C:P、N:P呈显著正相关关系(
p < 0.05), 叶片C:N与凋落物C、N含量、N:P呈显著负相关关系(
p < 0.05)。叶片C、N、P含量与土壤C、N (除P外)含量、C:N (除C、N外)、C:P、N:P呈显著正相关关系(
p < 0.05), 叶片C:N与土壤C、N含量、C:P、N:P, 叶片C:P与土壤C含量、C:N、C:P, 叶片N:P与土壤C:N呈显著负相关关系(
p < 0.05)。凋落物C含量与土壤C含量、C:P、N:P, 凋落物N含量与土壤N含量, 凋落物C:N与土壤C:P, 凋落物C:P与土壤C:N之间呈显著正相关关系(
p < 0.05), 而凋落物P含量与土壤C:N呈显著负相关关系(
p < 0.05)。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2植被恢复过程中植物叶片、凋落物、土壤C、N、P及其化学计量比的关系(n = 48)。 Fig. 2Relationships of leaf, litter, and soil C, N, P contents and stoichiometry during the restoration process (n = 48). 2.3 植物叶片的异速生长关系 不同恢复阶段N、P养分的协同作用随植物叶片的生长表现出不同的变化规律(
表1 )。LVR、PLL叶片N、P含量之间的主轴回归关系达到显著水平(
p < 0.01), 具有显著的异速生长关系(分别为
b = 3.81,
R 2 = 0.89;
b = 0.75,
R 2 = 0.83)。LCQ、LCC叶片N、P含量之间异速生长指数小于1, 且异速生长关系未达到显著水平(
p > 0.05)。整个恢复过程中, 叶片N、P养分的异速生长指数大于1, 且具有极显著的异速生长关系(
b = 1.45,
R 2 = 0.66,
p < 0.001)(
图3 )。
Table 1 表1 表1 不同恢复阶段植物叶片的N、P异速生长关系
Table 1
Allometric relationship of leaf N and P contents at each restoration stage logy vs. logx 恢复阶段 n 斜率b [95%置信区间]b [95% CI] 截距a [95%置信区间]a [95% CI ] 决定系数R 2 ) p logP vs. logN LVR 12 3.81 [2.83, 5.14] -4.28 [-5.45, -3.11] 0.89 <0.01 LCQ 12 0.54 [0.25, 1.17] -0.83 [-1.35, -0.31] 0.11 0.39 PLL 12 0.75 [0.52, 1.08] -1.03 [-1.37, -0.70] 0.83 <0.01 LCC 12 -0.96 [-2.07, -0.45] 1.00 [0.00, 2.00] 0.12 0.37
LVQ, LCQ, PLL, LCC see Fig. 1 . CI, confidence interval; Slope b, allometric index; n, the number of samples. Bold values indicate a significant major axis regression analysis relationship (p < 0.05). LVR, LCQ, PLL, LCC同图1 。斜率b 为异速生长指数, n 为样本数。加粗数据表示主轴回归关系达到显著水平(p < 0.05)。 新窗口打开 |
下载CSV 图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3植被恢复过程中植物叶片N、P养分的异速生长关系(n = 48)。 Fig. 3Allometric relationship of N and P contents of leaf during the restoration process (n = 48). 2.4 不同恢复阶段植物对N、P的利用效率和再吸收效率 植被恢复过程中, 植物叶片对N、P的利用效率具有不同的变化趋势(
图4A )。叶片对N的利用效率随植被恢复呈显著下降趋势, LCC降至57.48, 且LVR与LCQ、PLL、LCC之间, LCQ与LCC之间差异显著(
p < 0.05)。从LVR到PLL, 叶片对P的利用效率(1 362.85-2 735.59)呈下降趋势, LCC增高至LCQ水平, LVR显著高于其他恢复阶段(
p < 0.05), 叶片对P的利用效率高于对N的利用效率。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4植物叶片对N、P的利用效率(A)和再吸收效率(B)(n = 12)。LCC, LCQ, LVR, PLL同图1。NUE N , N的利用效率; NUE P , P的利用效率。不同小写字母代表不同恢复阶段之间的差异显著(p < 0.05)。 Fig. 4Utilization efficiency (A) and reabsorption efficiency (B) of N, P of leaf (n = 12). LCC, LCQ, LVR, PLL see Fig. 1. NUE N , nutrient use efficiency of N; NUE P , nutrient use efficiency of P. Different lowercase letters indicate significant differences at different restoration stages (p < 0.05). 植物叶片对N、P的再吸收效率随着植被恢复显著提高(
图4B ), PLL最高(N为14.79%, P为57.04%), LVR对N的再吸收效率最低, 为-24.00%, LCQ对P再吸收效率最低, 为18.60%。不同恢复阶段对N的再吸收效率显著低于对P的再吸收效率, N的再吸收效率最大值(14.79%)低于P的最小值(18.60%)。
2.5 土壤N、P含量与植物生长的关系 随着土壤N、P含量的变化, 叶片N、P含量呈不同的内稳态(
图5 )。随土壤N含量增加、P含量降低, 叶片P含量未出现大幅度变化, 线性回归斜率和相关系数趋近于0, 表明随着植被恢复叶片P含量内稳态较高, 不随着土壤N、P含量的变化而变化。叶片N含量与土壤N含量呈显著的线性正相关关系(
p < 0.001), 与土壤P含量回归斜率最大, 达14.88, 表明随着植被恢复, 叶片N的内稳态弱, 随土壤N、P含量的变化而变化, 特别是随土壤N含量的变化而显著变化。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5土壤N (A)、P (B)含量与植物叶片N、P含量的关系(n = 48)。***, p < 0.001。 Fig. 5Effects of soil N (A) and P (B) contents on the homeostasis of leaf N and P (n = 48). ***, p < 0.001. 3 讨论 3.1 植物叶片、凋落物、土壤C、N、P含量随植被恢复的变化 因植被恢复而形成不同功能性状的树种组成和结构(
Xu et al ., 2018 ), 导致植物养分吸收、凋落物数量和质量及其分解的差异, 进而影响土壤环境(
Yang & Luo, 2011 )。研究表明, 草本植物C含量低于灌木、乔木, 即灌木、乔木可以合成和积累更多的有机物, 形成更高的C含量(
He et al ., 2013 )。本研究中, 随着植被恢复, 叶片C含量增高, 与
He等(2013) 的研究结果一致, 与群落组成和结构随着植被恢复的变化有关。此外, 本研究中, 叶片对N、P的利用效率随植被恢复显著下降(
图4A ), 即N、P吸收量和存留量增高, 光合作用增强, 有利于干物质积累, 叶片C得到有效的积累。研究发现, 常绿乔木叶片寿命较长, 需要积累更多有机物质(如木质素等)构建保卫结构(
秦海等, 2010 ), 植物体保持较高的N、P含量, 有利于维持植被的生长代谢(
Zeng et al ., 2016 ), 乔木的生长相较于草本需要更多的N、P用于维持代谢所需。本研究中, 随着植被恢复, 叶片N含量显著增高。可能是由于随着植被恢复, 生物量增加促使植物需要更多富含N的物质(如酶、运输蛋白质和氨基酸)参与代谢活动, 加大叶片对N的吸收, 使得叶片N含量增加(
Qin et al ., 2016 )。而土壤N含量随植被恢复而增高, 能更大程度满足植物生长的需求, 也进一步解释了植物叶片N含量的变化(
Wang et al ., 2015 )。由于亚热带丘陵红壤区土壤严重缺P, 不同植被类型对P的再吸收利用效率不同。本研究中, PLL对P的再吸收利用效率最高(
图4B ), 其次是LCC, 这可能是PLL叶片P含量高于LCC, 而LCC高于LCQ、LVR的主要原因。随植被恢复, 植物叶片C、N、P含量呈递增趋势, 表明植被从灌草群落发展为灌木林、乔木林群落, 秉承了从资源保守策略转变为资源快速获取策略的特性(
Royer et al ., 2010 ), 更好地适应环境的变化。
凋落物养分含量明显受到群落优势植物及其凋落物组成的影响。研究发现, 林地凋落物C含量高于草地(
Paul et al ., 2002 ), 针叶林凋落物C含量高于阔叶林(
He et al ., 2013 )。本研究中, 随着植被恢复, 凋落物C含量显著提高, PLL最大, 与
Zeng等(2017a) 的研究结果一致, 是由于PLL以马尾松为优势树种, 凋落物主要以马尾松针叶为主, 而LVR以草本植物和小灌木为主, LCQ以落叶灌木为主, LCC以常绿阔叶树种为主所致。本研究中, 随着植被恢复, 凋落物N、P含量波动较大, 可能是由于不同恢复阶段植物对N、P的再吸收利用效率(
图4B )以及凋落物组成不同所致, 其中, PLL对N、P的再吸收利用效率最高, 其次LCC, 因而PLL凋落物N、P含量低于LCC。此外, 当凋落物N、P含量分别低于7、0.5 g·kg
-1 时, 表明凋落物N、P养分被叶片完全吸收; 而N、P含量分别高于10、0.8 g·kg
-1 时, 则表明没有完全吸收(
Killingbeck, 1996 ;
Yang et al ., 2018 )。本研究中, 凋落物N含量(12-18 g·kg
-1 )高于7 g·kg
-1 , P含量(0.31-0.49 g·kg
-1 )低于0.5 g·kg
-1 , 表明不同植被恢复阶段凋落物N均未被叶片完全吸收, 而P被完全吸收。可能是由于研究区林地土壤N较为丰富, 而P较为缺乏, 植物通过提高对P的再吸收利用以适应土壤P匮乏, 导致叶片对N的再吸收效率显著低于P再吸收效率。
森林恢复是决定土壤C、N、P含量变化的关键因素(
Xu et al ., 2018 )。有关本研究样地土壤C含量随着植被恢复而增加的机理已另有论文(
辜翔等, 2018 )详细讨论, 在这里不再重复。土壤N含量随植被恢复逐渐增加, 可能是受到凋落物养分释放的影响, 随着植被恢复, 凋落物C:N下降有利于N养分释放, 土壤N含量增加。而土壤P含量在恢复早期(LVR-LCQ)下降, 恢复后期(PLL-LCC)显著增加。经相关性分析表明, 土壤N、P含量与土壤C含量均呈显著的正相关关系(相关系数分别为0.917, 0.649,
p < 0.01,
n = 48)。表明植被恢复过程中, 土壤有机质的积累有利于土壤养分的提高(
Zhang et al ., 2019 )。此外, 可能是由于恢复早期植物对P的高需求和凋落物的低输入所致, LVR、LCQ植物对P的再吸收效率较高(
图4B ), 凋落物P归还低, 导致土壤P含量下降, 而恢复后期植被多样性和生物量增加(
辜翔等, 2018 ), 随凋落物P归还量增高, 土壤P含量有所提高。由于针叶树种叶片和凋落物酸度较高, 导致针叶林土壤P含量显著低于阔叶林(
Cao et al ., 2018 ), 因而PLL土壤P含量低于LCC。表明植被恢复形成不同的植物群落是影响土壤P含量的主要因素。
3.2 植物叶片、凋落物、土壤C、N、P化学计量比随植被恢复的变化 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 )。叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 )。植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 )。研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 )。本研究中, 叶片C:N、C:P总体上随植被恢复而下降。可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加。本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致。研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 )。但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 )。本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制。
本研究中, PLL凋落物C:N、C:P显著高于其他3个恢复阶段, 且随植被恢复的变化与C含量的变化一致, 表明是由于PLL凋落物以马尾松针叶为主, C含量较高所致。研究发现, 随着植被恢复, 凋落物N:P增加(
Zeng et al ., 2017b )。而本研究中, 随着植被恢复, 凋落物N:P有较大的波动, 其中PLL最高, 可能与凋落物P含量有关。随着植被恢复, 凋落物N:P的变化与P含量(
图1 )、P的再吸收效率(
图4B )的变化一致, 由于PLL对P的再吸收利用最高(
图4B ), 凋落物P含量下降, 导致凋落物N:P增高。研究表明, 当凋落物N:P > 25时, 其分解受到P限制(
Güsewell & Verhoeven, 2006 )。本研究中, 凋落物N:P (30-45)明显高于25, 表明本研究区凋落物分解受到P限制。
土壤C:N、C:P、N:P可作为土壤养分状况的良好指标(
Zhao et al ., 2015a ), 但因植物群落不同而异, 具有高度复杂性(
Zhang et al ., 2014 )。C、N、P不平衡性增加也会引起土壤C:N、C:P、N:P的变化(
Zhao et al ., 2015b )。本研究中, 土壤C:N (13-19)随着植被恢复先升高后下降, 与
Zhang等(2018) 的研究结果一致, 是由于从LVR到PLL土壤C含量增加幅度(121.66%)高于土壤N含量的增加幅度(79.75%), 导致土壤C:N升高; 而从PLL到LCC, 土壤C含量增加幅度(40.84%)低于土壤N含量的增加幅度(91.65%), 导致土壤C:N下降。这可以从植物对N的利用效率和再吸收效率来解释, 随着植被恢复, 植物对N的利用效率下降, 而恢复后期N的再吸收效率下降, 表明植物对土壤N的吸收量下降, 凋落物N含量较高, 有利于土壤N的积累。土壤C、N含量的变化是影响土壤C:P、N:P的主要因素(
Xu et al ., 2019 )。本研究中, 由于土壤P主要来源于母质的风化, 随植被恢复变化较小; 另一方面, 如前面分析, N的再吸收效率低于P, 凋落物的归还作用主要增加了土壤C、N的投入, 促进了土壤C、N的积累, 因而使得土壤C:P、N:P增加。在中国南方亚热带红壤丘陵区土壤较高的C:P、N:P也可以反映出林地土壤P的限制性(
Gao et al ., 2014 ;
Ren et al ., 2016 )。
3.3 叶片、凋落物、土壤C、N、P及化学计量比的关联性 限制性养分有效性、植物代谢速率和植物功能群共同影响叶片的化学计量模式, 因此, 不同区域植物叶片C、N、P化学计量比与环境的关系有所不同(
Zhao et al ., 2014 )。本研究中, 随着植被恢复, 土壤C含量增加, 叶片C含量增加, 与
Zhang等(2019) 的研究结果一致。同时, 土壤C含量与叶片N、P含量显著正相关, 表明随着植被恢复, 土壤有机质的积累和分解释放出更多可利用性N、P, 促进叶片对N、P的吸收。此外, 土壤N含量与叶片C、N含量显著正相关, 表明随植被恢复土壤N含量增加, 有利于植物的生长发育, 促进光合作用合成更多的有机物质。叶片N的变化决定着叶片C:N的变化。因此, 土壤C、N含量与叶片N的正相关性可以解释土壤C、N含量与叶片C:N的负相关性。本研究中, 土壤C:P、N:P与叶片P含量显著正相关, 与叶片C:P、N:P显著负相关, 可能是由于随着植被恢复, 土壤C:P、N:P增加, 植物受P的限制更为明显, 植物通过提高对P的吸收量(降低P的利用效率)和再吸收效率来维持正常生理活动, 这也是叶片P含量增加而C:P、N:P降低的原因之一。
凋落物是养分从植物到土壤的基本载体, 其动态交换可以实现并维持土壤养分和植物生长所需的元素比率之间的平衡(
Hessen et al ., 2004 )。凋落物养分含量及其化学计量比与土壤养分含量和化学计量比显著相关(
Yang et al ., 2018 ), 一方面, 表明凋落物分解促使有机物质向土壤释放N、P, 从而影响土壤C、N和P含量及其化学计量比(
Zhong et al ., 2013 ), 另一方面, 表明土壤养分含量的变化会影响植物对养分的利用策略, 进而影响凋落物养分含量及其化学计量比。本研究中, 土壤C、N含量与凋落物C、N含量显著正相关, 可能是随着植被恢复, 土壤C、N含量增加, 更能满足植物对养分的需求, 从而影响植物对养分的再吸收效率, 进而影响凋落物的养分含量。因此, 叶片N、P含量显著影响着凋落物养分含量。此外, 本研究中, 土壤C:N、C:P和N:P与凋落物C:N、C:P显著正相关, 与
汪宗飞和郑粉莉(2018) 、
张萍等(2018) 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 )。也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加。
3.4 植物叶片内稳态和养分利用策略对植被恢复的响应 研究表明, 土壤养分含量的变化直接影响植物养分吸收及其化学计量比(
Chen et al ., 2018 )。而植物通过改变化学计量比和采用适宜的养分利用策略适应土壤N、P供应的变化(
Sun et al ., 2018 )。植物N含量比P含量更稳定(
Yu et al ., 2010 )。本研究中, 随着植被恢复, 叶片N含量内稳态较弱, 受土壤N、P含量的变化影响较大, 叶片P含量具有较高的内稳态, 不随土壤N、P含量的变化而变化, 与
Yu等(2010) 的研究结果不同。可能是由于植物的限制性养分元素不同所致, 在
Yu等(2010) 的研究中, 植物主要受N限制, 而本研究中, 土壤P含量随植被恢复的变化小, 植物主要受P的限制, 在一定程度上证实了限制性养分元素稳定性假说“由于生理和养分平衡的制约, 限制性养分元素在植物体内的含量具有相对稳定性, 对环境变化的响应也较为稳定” (
Sterner & Elser, 2002 )。表明随着植被恢复, 在P缺乏的土壤环境中, 植物叶片P具有较强的内稳态。
研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 )。植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 )。凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 )。因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略。本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化。LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境。LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N。PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 )。PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求。LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 )。常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降。研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 )。本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性。
4 结论 植被恢复显著影响植物叶片、凋落物、土壤C、N、P含量及其化学计量比。不同组分C、N、P含量及其化学计量比随植被恢复的变化不完全一致, 叶片C:N、C:P显著下降, 叶片C、N、P含量和土壤C、N含量、C:N、C:P、N:P显著增加, 凋落物C、N、P含量及C:N、C:P、N:P波动较大。叶片与土壤间相关关系显著, 其中叶片C、N、P含量与土壤C、N含量、C:N (除C、N外)、C:P、N:P呈显著正相关关系, 叶片C:N与土壤C、N含量、C:P、N:P, 叶片C:P与土壤C含量、C:N、C:P, 叶片N:P与土壤C:N呈显著负相关关系, 而凋落物与叶片、土壤间的相关关系较弱。不同植被恢复阶段, 植物生长受P限制。叶片N含量内稳态较弱, P含量内稳态较高。叶片通过降低N、P的投入比例和利用效率维持体内平衡, 提高N、P再吸收效率以适应土壤环境随植被恢复的变化。研究结果对深入理解亚热带植物恢复过程中, 生态系统各组分(植物叶片、凋落物、土壤)之间的养分循环和植物的适应策略具有重要意义。
致谢 感谢中南林业科技大学生命科学与技术学院生态教研室项文化教授、博士生导师,雷丕锋博士、副教授在论文撰写过程中给予的帮助。感谢中南林业科技大学引进高层次人才科研启动基金项目(2014YJ019); 中南林业科技大学研究生科技创新基金项目(CX2017A07)的支持。
[1] Cao Y Chen YM 2017 ). Ecosystem C:N:P stoichiometry and carbon storage in plantations and a secondary forest on the Loess Plateau, ChinaEcological Engineering, 105, 125 -132 . [本文引用: 2] [2] Cao Y Zhang P Chen YM 2018 ). Soil C:N:P stoichiometry in plantations of N-fixing black locust and indigenous pine, and secondary oak forests in Northwest ChinaJournal of Soils and Sediments, 18, 1478 -1489 . [本文引用: 1] [3] Chapin III FS (1980 ). The mineral nutrition of wild plantsAnnual Review of Ecology and Systematics, 1, 233 -260 . [本文引用: 1] [4] Chen LL Deng Q Yuan ZY Mu XM Kallenbach RL 2018 ). Age-related C:N:P stoichiometry in two plantation forests in the Loess Plateau of ChinaEcological Engineering, 120, 14 -22 . [本文引用: 2] [5] Clinton PW Allen RB Davis MR 2002 ). Nitrogen storage and availability during stand development in a New Zealand Nothofagus forestCanadian Journal of Forest Research 344 -352 . [本文引用: 1] [6] Elser JJ Sterner RW Gorokhova E Fagan WF Markow TA Cotner JB Harrison JF Hobbie SE Odell GM Weider LW 2000 ). Biological stoichiometry from genes to ecosystemsEcology Letters, 3, 540 -550 . [本文引用: 1] [7] Gao Y He NP Yu GR Chen WL Wang QF 2014 ). Long-term effects of different land use types on C, N, and P stoichiometry and storage in subtropical ecosystems: A case study in ChinaEcological Engineering, 67, 171 -181 . [本文引用: 1] [8] Grime JP 2001 ). Plant Strategies, Vegetation Processes, and Ecosystem Properties. John Wiley & Sons, Chichester, UK . 13 -48 . [本文引用: 1] [9] Gu X Zhang SJ Liu ZD Li LD Chen JL Wang LF Fang X 2018 ). Effects of vegetation restoration on soil organic carbon concentration and density in the mid-subtropical region of ChinaChinese Journal of Plant Ecology, 42, 595 -608 . [本文引用: 4] [ 辜翔 , 张仕吉 , 刘兆丹 , 李雷达 , 陈金磊 , 王留芳 , 方晰 (2018 ). 中亚热带植被恢复对土壤有机碳含量、碳密度的影响植物生态学报 , 42, 595 -608 .] [本文引用: 4] [10] Güsewell S 2004 ). N:P ratios in terrestrial plants: Variation and functional significanceNew Phytologist, 164, 243 -266 . [本文引用: 1] [11] Güsewell S Koerselman W 2002 ). Variation in nitrogen and phosphorus concentrations of wetland plantsPerspectives in Plant Ecology, Evolution and Systematics, 5, 37 -61 . [本文引用: 1] [12] Güsewell S Verhoeven JTA 2006 ). Litter N:P ratios indicate whether N or P limits the decomposability of graminoid leaf litterPlant and Soil, 287, 131 -143 . [本文引用: 2] [13] Han WX Fang JY 2008 ). Review on the mechanism models of allometric scaling laws: 3/4 vs. 2/3 powerJournal of Plant Ecology (Chinese Version )951 -960 . [本文引用: 1] [ 韩文轩 , 方精云 (2008 ). 幂指数异速生长机制模型综述植物生态学报 , 32, 951 -960 .] [本文引用: 1] [14] He JS Wang L Flynn DFB Wang X Ma W Fang J 2008 ). Leaves nitrogen: Phosphorus stoichiometry across Chinese grassland biomesOecologia, 155, 301 -310 . [本文引用: 1] [15] He YJ Qin L Li ZY Liang XY Shao MX Tan L 2013 ). Carbon storage capacity of monoculture and mixed-?species plantations in subtropical ChinaForest Ecology and Management, 295, 193 -198 . [本文引用: 3] [16] Hector A Bagchi R 2007 ). Biodiversity and ecosystem multifunctionalityNature, 448, 188 -190 . [本文引用: 1] [17] Hedin LO 2004 ). Global organization of terrestrial plant-?nutrient interactionsProceedings of the National Academy of Sciences of the United States of America, 101, 10849 -10850 . [本文引用: 1] [18] Hessen DO ?gren GI Anderson TR Elser JJ de Ruiter PC 2004 ). Carbon sequestration in ecosystems: The role of stoichiometryEcology, 85, 1179 -1192 . [本文引用: 1] [19] Hu YF Shu XY He J Zhang YL Xiao HH Tang XY Gu YF Lan T Xia JG Ling J Chen GD Wang CQ Deng LJ Yuan S 2018 ). Storage of C, N, and P affected by afforestation with Salix cupularis in an alpine semiarid desert ecosystemLand Degradation & Development, 188 -198 . [本文引用: 1] [20] Institute of Soil Science, Chinese Academy of Sciences (1978 ). The Analysis of Soil Physical-Chemical Properties . Shanghai Scientific & Technical Publishers, Shanghai. [本文引用: 1] [ 中国科学院南京土壤研究所 (1978 ). 土壤理化分析 . 上海科技出版社, 上海.] [本文引用: 1] [21] Jiang LL He S Wu LF Yan YF Weng SF Liu J Wang WQ Zeng CS 2014 ). Characteristics of stoichiometric homeostasis of three plant species in wetlands in Minjiang EstuaryWetland Science, 12, 293 -298 . [本文引用: 1] [ 蒋利玲 , 何诗 , 吴丽凤 , 颜远烽 , 翁少峰 , 刘静 , 王维奇 , 曾从盛 (2014 ). 闽江河口湿地3种植物化学计量内稳性特征湿地科学 , 12, 293 -298 .] [本文引用: 1] [22] Killingbeck KT 1996 ). Nutrients in senesced leaves: Keys to the search for potential resorption and resorption proficiencyEcology, 77, 1716 -1727 . [本文引用: 1] [23] Koerselman W Meuleman AFM 1996 ). The vegetation N:P ratio: A new tool to detect the nature of nutrients limitationJournal of Applied Ecology, 33, 1441 -1450 . [本文引用: 1] [24] Kooijman SALM 1995 ). The stoichiometry of animal energeticsJournal of Theoretical Biology, 177, 139 -149 . [本文引用: 1] [25] Li CZ Zhao LH Sun PS Zhao FZ Kang D Yang GH Han XH Feng YZ Ren GX 2016 ). Deep soil C, N, and P stocks and stoichiometry in response to land use patterns in the Loess Hilly Region of ChinaPLOS ONE, 11, e0159075 . DOI: 10.1371/journal.pone.0159075 . [本文引用: 2] [26] Liu WD Su JR Li SF Zhang ZJ Li ZW 2010 ). Stoichiometry study of C, N and P in plant and soil at different successional stages of monsoon evergreen broad-leaved forest in Pu’er, Yunnan ProvinceActa Ecologica Sinica, 30, 6581 -6590 . [本文引用: 1] [ 刘万德 , 苏建荣 , 李帅锋 , 张志钧 , 李忠文 (2010 ). 云南普洱季风常绿阔叶林演替系列植物和土壤C、N、P化学计量特征生态学报 , 30, 6581 -6590 .] [本文引用: 1] [27] Liu Y Wu YX Han SJ Lin L 2009 ). Litterfall decomposition in four forest types in Changbai Mountains of ChinaChinese Journal of Ecology, 28, 400 -404 . [本文引用: 1] [ 刘颖 , 武耀祥 , 韩士杰 , 林鹿 (2009 ). 长白山四种森林类型凋落物分解动态生态学杂志 , 28, 400 -404 .] [本文引用: 1] [28] Milla R Castro-Díez P Maestro-Martínez M Montserrat-Martí G 2005 ). Does the gradualness of leaf shedding govern nutrient resorption from senescing leaves in Mediterranean woody plants?Plant and Soil, 278, 303 -313 . [本文引用: 1] [29] Mooshammer M Wanek W Schnecker J Wild B Leitner S Hofhansl F Bl?chl A H?mmerle I Frank AH Fuchslueger L Keiblinger KM Zechmeister Boltenstern S Richter A 2012 ). Stoichiometric controls of nitrogen and phosphorus cycling in decomposing beech leaf litterEcology, 93, 770 -782 . [本文引用: 1] [30] Nie LQ Wu Q Yao B Fu S Hu QW 2016 ). Leaf litter and soil carbon, nitrogen, and phosphorus stoichiometry of dominant plant species in the Poyang Lake wetlandActa Ecologica Sinica, 36, 1898 -1906 . [本文引用: 1] [ 聂兰琴 , 吴琴 , 尧波 , 付姗 , 胡启武 (2016 ). 鄱阳湖湿地优势植物叶片-凋落物-土壤碳氮磷化学计量特征生态学报 , 36, 1898 -1906 .] [本文引用: 1] [31] Niklas KJ 1994 ). Size-dependent variations in plant growth rates and the “3/4-power rule”American Journal of Botany, 81, 134 -144 . [本文引用: 1] [32] Ning QR Li SZ Jiang LC Zhao Y Liu R Zhang XY 2016 ). Foliar nutrient content and resorption efficiency of Pinus massoniana in the subtropical red soil erosion regionActa Ecologica Sinica, 3510 -3517 . [本文引用: 1] [ 宁秋蕊 , 李守中 , 姜良超 , 赵颖 , 刘溶 , 张欣影 (2016 ). 亚热带红壤侵蚀区马尾松针叶养分含量及再吸收特征生态学报 , 36, 3510 -3517 .] [本文引用: 1] [33] O’Brien SL Jastrow JD 2013 ). Physical and chemical protection in hierarchical soil aggregates regulates soil carbon and nitrogen recovery in restored perennial grasslandsSoil Biology & Biochemistry, 61, 1 -13 . [本文引用: 1] [34] Paul KI Polglase PJ Nyakuengama JG Khanna PK 2002 ). Change in soil carbon following afforestationForest Ecology and Management, 168, 241 -257 . [本文引用: 2] [35] Peng X Fang X Yu LH Xiang WH Huang ZH 2016 ). Change characteristic of soil C, N, P stoichiometric ratios in mid-subtropical forests restorationJournal of Central South University of Forestry & Technology, 36(11 ), 65 -72 . [本文引用: 1] [ 彭晓 , 方晰 , 喻林华 , 项文化 , 黄志宏 (2016 ). 中亚热带4种森林土壤碳、氮、磷化学计量特征中南林业科技大学学报 , 36(11 ), 65 -72 .] [本文引用: 1] [36] Qin H Li JX Gao SP Li C Li R Sheng XH 2010 ). Characteristics of leaf element contents for eight nutrients across 660 terrestrial plant species in ChinaActa Ecologica Sinica, 30, 1247 -1257 . [本文引用: 2] [ 秦海 , 李俊祥 , 高三平 , 李铖 , 李蓉 , 沈兴华 (2010 ). 中国660种陆生植物叶片8种元素含量特征生态学报 , 30, 1247 -1257 .] [本文引用: 2] [37] Qin J Xi WM Rahmlow A Kong HY Zhang Z Shangguan ZP 2016 ). Effects of forest plantation types on leaf traits of Ulmus pumila and Robinia pseudoacacia on the Loess Plateau, ChinaEcological Engineering 416 -425 . [本文引用: 1] [38] R Development Core Team 2018 ). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. [本文引用: 1] [39] Reich PB Tjoelker MG MacHado JL Oleksyn J 2006 ). Universal scaling of respiratory metabolism, size and nitrogen in plantsNature, 439, 457 -461 . [本文引用: 1] [40] Ren CJ Zhao FZ Kang D Yang GH Han XH Tong XG Feng YZ Ren GX 2016 ). Linkages of C:N:P stoichiometry and bacterial community in soil following afforestation of former farmlandForest Ecology and Management, 376, 59 -66 . [本文引用: 2] [41] Royer DL Miller IM Peppe DJ Hickey LJ 2010 ). Leaf economic traits from fossils support a weedy habit for early angiospermsAmerican Journal of Botany, 97, 438 -445 . [本文引用: 2] [42] Sardans J Alonso R Carnicer J Fernández-Martínez M Vivanco MG Pe?uelas J 2016 ). Factors influencing the foliar elemental composition and stoichiometry in forest trees in SpainPerspectives in Plant Ecology Evolution & Systematics, 18, 52 -69 . [本文引用: 1] [43] Schreeg LA Santiago LS Wright SJ Turner BL 2014 ). Stem, root, and older leaf N:P ratios are more responsive indicators of soil nutrient availability than new foliageEcology, 95, 2062 -2068 . [本文引用: 1] [44] Sterner RW Elser JJ 2002 ). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere . Princeton University Press, Princeton. [本文引用: 2] [45] Su B Han XG Huang JH Qu CM 2000 ). The nutrient use efficiency (NUE) of plants and it’s implications on the strategy of plant adaptation to nutrient-stressed environmentsActa Ecologica Sinica, 20, 335 -343 . [本文引用: 1] [ 苏波 , 韩兴国 , 黄建辉 , 渠春梅 (2000 ). 植物的养分利用效率(NUE)及植物对养分胁迫环境的适应策略生态学报 , 20, 335 -343 .] [本文引用: 1] [46] Sun X Shen Y Schuster MJ Searle EB Chen JH Yang GW Zhang YJ 2018 ). Initial responses of grass litter tissue chemistry and N:P stoichiometry to varied N and P input rates and ratios in Inner MongoliaAgriculture, Ecosystems & Environment, 252, 114 -125 . [本文引用: 1] [47] Tian LM Zhao L Wu XD Fang HB Zhao YH Hu GJ Yue GY Sheng Y Wu JC Chen J Wang ZW Li WP Zou DF Ping CL Shang W Zhao YG Zhang GL 2018 ). Soil moisture and texture primarily control the soil nutrient stoichiometry across the Tibetan grasslandScience of the Total Environment 192 -202 . [本文引用: 1] [48] Wang LL Zhao GX Li M Zhang MT Zhang LF Zhang XF An LZ Xu SJ 2015 ). C:N:P stoichiometry and leaf traits of halophytes in an arid saline environment, northwest ChinaPLOS ONE, 10, e0119935 . DOI: 10.1371/journal.pone.0119935 . [本文引用: 1] [49] Wang M Moore TR 2014 ). Carbon, nitrogen, phosphorus, and potassium stoichiometry in an ombrotrophic peatland reflects plant functional typeEcosystems, 17, 673 -684 . [本文引用: 1] [50] Wang ZF Zheng FL 2018 ). C, N, and P stoichiometric characteristics of Pinus tabulaeformis plantation in the Ziwuling Region of the Loess PlateauActa Ecologica Sinica, 6870 -6880 . [本文引用: 1] [ 汪宗飞 , 郑粉莉 (2018 ). 黄土高原子午岭地区人工油松林碳氮磷生态化学计量特征生态学报 , 38, 6870 -6880 .] [本文引用: 1] [51] Warton DI Duursma RA Falster DS Taskinen S 2012 ). SMART 3—An R package for estimation and inference about allometric linesMethods in Ecology and Evolution, 3, 257 -259 . [本文引用: 1] [52] Xiang WH Liu SH Lei XD Frank SC Tian DL Wang GJ Deng XW 2013 ). Secondary forest floristic composition, structure, and spatial pattern in subtropical ChinaJournal of Forest Research, 18, 111 -120 . [本文引用: 1] [53] Xiao HB Li ZW Dong YT Chang XF Deng L Huang JQ Nie XD Liu C Liu L Wang DY Liu QM Zhang YR 2017 ). Changes in microbial communities and respiration following the revegetation of eroded soilAgriculture Ecosystems & Environment, 246, 30 -37 . [本文引用: 1] [54] Xu CH Xiang WH Gou MM Chen L Lei PF Fang X Deng XW Ouyang S 2018 ). Effects of forest restoration on soil carbon, nitrogen, phosphorus, and their stoichiometry in Hunan, Southern ChinaSustainability, 10, 1874 . DOI:? 10.3390/su10061874 . [本文引用: 3] [55] Xu HW Qu Q Li P Guo ZQ Wulan E Xue S 2019 ). Stocks and stoichiometry of soil organic carbon, total nitrogen, and total phosphorus after vegetation restoration in the Loess Hilly Region, ChinaForests, 10, 27 . DOI: 10.3390/f10010027 . [本文引用: 2] [56] Yan ER Wang XH Guo M Zhong Q Zhou W 2010 ). C:N:P stoichiometry across evergreen broad-leaved forests, evergreen coniferous forests and deciduous broad-leaved forests in the Tiantong region, Zhejiang Province, eastern ChinaChinese Journal of Plant Ecology, 34, 48 -57 . [本文引用: 1] [ 阎恩荣 , 王希华 , 郭明 , 仲强 , 周武 (2010 ). 浙江天童常绿阔叶林、常绿针叶林与落叶阔叶林的C:N:P化学计量特征植物生态学报 , 34, 48 -57 .] [本文引用: 1] [57] Yang Y Liu BR An SS 2018 ). Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern ChinaCatena, 166, 328 -338 . [本文引用: 2] [58] Yang YH Luo YQ 2011 ). Carbon:nitrogen stoichiometry in forest ecosystems during stand developmentGlobal Ecology and Biogeography, 20, 354 -361 . [本文引用: 2] [59] Yu Q Chen QS Elser JJ He NP Wu HH Zhang GM Wu JG Bai YF Han XG 2010 ). Linking stoichiometric homoeostasis with ecosystem structure, functioning and stabilityEcology Letters, 13, 1390 -1399 . [本文引用: 3] [60] Yu Q Elser JJ He NP Wu HH Chen QS Zhang GM Han XG 2011 ). Stoichiometric homeostasis of vascular plants in the Inner Mongolia grasslandOecologia, 166, 1 -10 . [本文引用: 1] [61] Zeng QC Li X Dong YH An SS Darboux F 2016 ). Soil and plant components ecological stoichiometry in four steppe communities in the Loess Plateau of ChinaCatena, 147, 481 -488 . [本文引用: 1] [62] Zeng QC Liu Y Fang Y Ma RT Lal R An SS Huang YM 2017a ). Impact of vegetation restoration on plants and soil C:N:P stoichiometry on the Yunwu Mountain Reserve of ChinaEcological Engineering, 109, 92 -100 . [本文引用: 6] [63] Zeng YL Fang X Xiang WH Deng XW Peng CH 2017b ). Stoichiometric and nutrient resorption characteristics of dominant tree species in subtropical Chinese forestsEcology and Evolution, 7, 11033 -11043 . [本文引用: 2] [64] Zhang DJ Zhang J Yang WQ Wu FZ Huang YM 2014 ). Plant and soil seed bank diversity across a range of ages of Eucalyptus grandis plantations afforested on arable landsPlant and soil 307 -325 . [本文引用: 3] [65] Zhang GQ Zhang P Peng SZ Chen YM Cao Y 2017 ). The coupling of leaf, litter, and soil nutrients in warm temperate forests in northwestern ChinaScientific Reports, 7, 11754 . DOI: 10.1038/s41598-017-12199-5 . [本文引用: 1] [66] Zhang P Zhang GQ Zhao YP Peng SZ Chen YM Cao Y 2018 ). Ecological stoichiometry characteristics of leaf-litter-soil interactions in different forest types in the Loess hilly-gully region of ChinaActa Ecologica Sinica, 38, 5087 -5098 . [本文引用: 3] [ 张萍 , 章广琦 , 赵一娉 , 彭守璋 , 陈云明 , 曹扬 (2018 ). 黄土丘陵区不同森林类型叶片-凋落物-土壤生态化学计量特征生态学报 , 38, 5087 -5098 .] [本文引用: 3] [67] Zhang W Liu WC Xu MP Deng J Han XH Yang GH Feng YZ Ren GX 2019 ). Response of forest growth to C:N:P stoichiometry in plants and soils during Robinia pseudoacacia afforestation on the Loess Plateau, ChinaGeoderma 280 -289 . [本文引用: 3] [68] Zhang W Ren CJ Deng J Zhao FZ Yang GH Han XH Tong XG Feng YZ 2018 ). Plant functional composition and species diversity affect soil C, N, and P during secondary succession of abandoned farmland on the Loess PlateauEcological Engineering, 122, 91 -99 . [本文引用: 1] [69] Zhao FZ Kang D Han XH Yang GH Feng YZ Ren GX 2015a ). Soil stoichiometry and carbon storage in long-term afforestation soil affected by understory vegetation diversityEcological Engineering, 74, 415 -422 . [本文引用: 2] [70] Zhao FZ Sun J Ren CJ Kang D Deng J Han XH Yang GH Feng YZ Ren GX 2015b ). Land use change influences soil C, N and P stoichiometry under “Grain-to-Green Program” in ChinaScientific Reports, 5, 10195 . DOI: 10.1038/srep10195 . [本文引用: 4] [71] Zhao N He NP Wang QF Zhang XY Wang RL Xu ZW Yu GR 2014 ). The altitudinal patterns of leaf C:N:P stoichiometry are regulated by plant growth form, climate and soil on Changbai Mountain, ChinaPLOS ONE, 9, e95196 . DOI: 10.1371/journal.pone.0095196 . [本文引用: 1] [72] Zheng L Lu LH 2012 ). Standing crop and nutrient characteristics of forest floor litter in ChinaJournal of Northwest Forestry University, 27(1 ), 63 -69 . [本文引用: 1] [ 郑路 , 卢立华 (2012 ). 我国森林地表凋落物现存量及养分特征西北林学院学报 , 27(1 ), 63 -69 .] [本文引用: 1] [73] Zhong ZS Song XL Lu XG Xue ZS 2013 ). Ecological stoichiometry of carbon, nitrogen, and phosphorus in estuarine wetland soils: Influences of vegetation coverage, plant communities, geomorphology, and seawallsJournal of Soils and Sediments, 13, 1043 -1051 . [本文引用: 1] Ecosystem C:N:P stoichiometry and carbon storage in plantations and a secondary forest on the Loess Plateau, China
2
2017
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
Soil C:N:P stoichiometry in plantations of N-fixing black locust and indigenous pine, and secondary oak forests in Northwest China
1
2018
... 森林恢复是决定土壤C、N、P含量变化的关键因素(
Xu et al ., 2018 ).有关本研究样地土壤C含量随着植被恢复而增加的机理已另有论文(
辜翔等, 2018 )详细讨论, 在这里不再重复.土壤N含量随植被恢复逐渐增加, 可能是受到凋落物养分释放的影响, 随着植被恢复, 凋落物C:N下降有利于N养分释放, 土壤N含量增加.而土壤P含量在恢复早期(LVR-LCQ)下降, 恢复后期(PLL-LCC)显著增加.经相关性分析表明, 土壤N、P含量与土壤C含量均呈显著的正相关关系(相关系数分别为0.917, 0.649,
p < 0.01,
n = 48).表明植被恢复过程中, 土壤有机质的积累有利于土壤养分的提高(
Zhang et al ., 2019 ).此外, 可能是由于恢复早期植物对P的高需求和凋落物的低输入所致, LVR、LCQ植物对P的再吸收效率较高(
图4B ), 凋落物P归还低, 导致土壤P含量下降, 而恢复后期植被多样性和生物量增加(
辜翔等, 2018 ), 随凋落物P归还量增高, 土壤P含量有所提高.由于针叶树种叶片和凋落物酸度较高, 导致针叶林土壤P含量显著低于阔叶林(
Cao et al ., 2018 ), 因而PLL土壤P含量低于LCC.表明植被恢复形成不同的植物群落是影响土壤P含量的主要因素. ...
The mineral nutrition of wild plants
1
1980
... (2)养分利用效率(
NUE )反映植物对土壤养分(如N、P)的利用和适应状况.养分利用率高表明植物能通过较低的养分吸收量和存留量生产更多生物量, 是植物适应贫瘠土壤环境的重要竞争策略(
Grime, 2001 ).本研究采用Chapin指数(
Chapin III, 1980 )计算, 公式如下: ...
Age-related C:N:P stoichiometry in two plantation forests in the Loess Plateau of China
2
2018
... 凋落物是养分从植物到土壤的基本载体, 其动态交换可以实现并维持土壤养分和植物生长所需的元素比率之间的平衡(
Hessen et al ., 2004 ).凋落物养分含量及其化学计量比与土壤养分含量和化学计量比显著相关(
Yang et al ., 2018 ), 一方面, 表明凋落物分解促使有机物质向土壤释放N、P, 从而影响土壤C、N和P含量及其化学计量比(
Zhong et al ., 2013 ), 另一方面, 表明土壤养分含量的变化会影响植物对养分的利用策略, 进而影响凋落物养分含量及其化学计量比.本研究中, 土壤C、N含量与凋落物C、N含量显著正相关, 可能是随着植被恢复, 土壤C、N含量增加, 更能满足植物对养分的需求, 从而影响植物对养分的再吸收效率, 进而影响凋落物的养分含量.因此, 叶片N、P含量显著影响着凋落物养分含量.此外, 本研究中, 土壤C:N、C:P和N:P与凋落物C:N、C:P显著正相关, 与
汪宗飞和郑粉莉(2018) 、
张萍等(2018) 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 ).也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加. ...
... 研究表明, 土壤养分含量的变化直接影响植物养分吸收及其化学计量比(
Chen et al ., 2018 ).而植物通过改变化学计量比和采用适宜的养分利用策略适应土壤N、P供应的变化(
Sun et al ., 2018 ).植物N含量比P含量更稳定(
Yu et al ., 2010 ).本研究中, 随着植被恢复, 叶片N含量内稳态较弱, 受土壤N、P含量的变化影响较大, 叶片P含量具有较高的内稳态, 不随土壤N、P含量的变化而变化, 与
Yu等(2010) 的研究结果不同.可能是由于植物的限制性养分元素不同所致, 在
Yu等(2010) 的研究中, 植物主要受N限制, 而本研究中, 土壤P含量随植被恢复的变化小, 植物主要受P的限制, 在一定程度上证实了限制性养分元素稳定性假说“由于生理和养分平衡的制约, 限制性养分元素在植物体内的含量具有相对稳定性, 对环境变化的响应也较为稳定” (
Sterner & Elser, 2002 ).表明随着植被恢复, 在P缺乏的土壤环境中, 植物叶片P具有较强的内稳态. ...
Nitrogen storage and availability during stand development in a New Zealand
Nothofagus forest
1
2002
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
Biological stoichiometry from genes to ecosystems
1
2000
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
Long-term effects of different land use types on C, N, and P stoichiometry and storage in subtropical ecosystems: A case study in China
1
2014
... 土壤C:N、C:P、N:P可作为土壤养分状况的良好指标(
Zhao et al ., 2015a ), 但因植物群落不同而异, 具有高度复杂性(
Zhang et al ., 2014 ).C、N、P不平衡性增加也会引起土壤C:N、C:P、N:P的变化(
Zhao et al ., 2015b ).本研究中, 土壤C:N (13-19)随着植被恢复先升高后下降, 与
Zhang等(2018) 的研究结果一致, 是由于从LVR到PLL土壤C含量增加幅度(121.66%)高于土壤N含量的增加幅度(79.75%), 导致土壤C:N升高; 而从PLL到LCC, 土壤C含量增加幅度(40.84%)低于土壤N含量的增加幅度(91.65%), 导致土壤C:N下降.这可以从植物对N的利用效率和再吸收效率来解释, 随着植被恢复, 植物对N的利用效率下降, 而恢复后期N的再吸收效率下降, 表明植物对土壤N的吸收量下降, 凋落物N含量较高, 有利于土壤N的积累.土壤C、N含量的变化是影响土壤C:P、N:P的主要因素(
Xu et al ., 2019 ).本研究中, 由于土壤P主要来源于母质的风化, 随植被恢复变化较小; 另一方面, 如前面分析, N的再吸收效率低于P, 凋落物的归还作用主要增加了土壤C、N的投入, 促进了土壤C、N的积累, 因而使得土壤C:P、N:P增加.在中国南方亚热带红壤丘陵区土壤较高的C:P、N:P也可以反映出林地土壤P的限制性(
Gao et al ., 2014 ;
Ren et al ., 2016 ). ...
Plant Strategies, Vegetation Processes, and Ecosystem Properties. 1
2001
... (2)养分利用效率(
NUE )反映植物对土壤养分(如N、P)的利用和适应状况.养分利用率高表明植物能通过较低的养分吸收量和存留量生产更多生物量, 是植物适应贫瘠土壤环境的重要竞争策略(
Grime, 2001 ).本研究采用Chapin指数(
Chapin III, 1980 )计算, 公式如下: ...
中亚热带植被恢复对土壤有机碳含量、碳密度的影响
4
2018
... 采用空间代替时间的方法, 按照亚热带森林群落自然演替进程, 根据植被恢复程度及其群落树种组成, 选取地域相邻、环境条件(立地、气候和土壤)相似, 处于不同恢复阶段的4种生态系统: (1)檵木(
Loropetalum chinensis ) +南烛(
Vaccinium bracteatum ) +杜鹃(
Rhododendron mariesii )灌草丛(LVR): 1965年砍伐常绿阔叶林后, 炼山、人工整地, 1966年营造马尾松人工林, 无施肥历史, 1990年皆伐马尾松人工林.2012年, 停止各种人为干扰(砍杂、放牧), 让其自然恢复, 当前草本植物多且生长良好, 伴随分布许多幼小灌木植物, 属于恢复早期阶段.(2)檵木+杉木+白栎(
Quercus fabri )灌木林(LCQ): 1965年天然常绿阔叶林采伐后, 炼山、人工整地, 1966年营造杉木人工纯林, 无施肥历史, 1989年皆伐杉木人工纯林.2004年停止各种人为干扰, 让其自然恢复, 灌木植物发育良好, 但没有形成明显的乔木层, 草本植物稀少, 属于恢复中期第I阶段.(3)马尾松+柯(
Lithocarpus glaber ) +檵木针阔混交林(PLL): 20世纪60年代末, 天然常绿阔叶林皆伐后, 让其自然恢复, 目前已恢复为针阔混交林, 年龄为45-50年, 群落内幼苗幼树丰富, 植物密度较大, 但大直径个体所占比例相对较低, 属于恢复中期第II阶段.(4)柯+红淡比(
Cleyera japonica ) +青冈(
Cyclobalanopsis Glauca )常绿阔叶林(LCC): 长期以来无明显人为干扰, 保存比较完好, 群落结构相对稳定, 年龄为80-90年, 属于恢复后期(亚顶极)阶段.在LVR、LCQ各设置4块20 m × 20 m固定样地, 在PLL、LCC分别设置4块30 m × 30 m的固定样地.群落调查方法和4种群落的基本特征及其主要树种组成详见
辜翔等(2018) 的文献. ...
... 为了避免偶然性, 分别在2016年4、6、10月下旬采集植物叶片、凋落物、0-30 cm土层分析样品, 取3次采样测定的平均值作为最终的数据.在LVR固定样地, 沿着对角线均匀设置3个1 m × 1 m样方, 采集样方内所有灌木叶片、草本植物地上部分, 将同一样地3个样方植物叶片等比例混合为1个样品.根据群落调查结果(
辜翔等, 2018 ), 确定LCQ、PLL和LCC每块固定样地的优势树种, 对该固定样地的每种优势树种分别选取9株长势良好的样木.每种优势树种每次采集3株样木的叶片, 按上、中、下3个部位的东、西、南、北4个方向采集各株样木已完全成熟、无虫害的叶片, 草本植物地上部分采集同LVR, 再将同一样地内草本植物和优势树种叶片按生物量折算比例混合为1个样品.按
郑路和卢立华(2012) 的凋落物层分层标准, 在完成了草本植物地上部分样品采集后的3个1 m × 1 m的样方内采集未分解层的凋落物样品.完成凋落物采集后, 将样方内剩余凋落物清理干净, 用土钻(内径10 cm)取0-30 cm土层土壤样品, 将同一样地3个样方的凋落物、土壤分别混合为1个样品.所有的分析样品每次采样4个重复, 3次采样12个重复. ...
... 森林恢复是决定土壤C、N、P含量变化的关键因素(
Xu et al ., 2018 ).有关本研究样地土壤C含量随着植被恢复而增加的机理已另有论文(
辜翔等, 2018 )详细讨论, 在这里不再重复.土壤N含量随植被恢复逐渐增加, 可能是受到凋落物养分释放的影响, 随着植被恢复, 凋落物C:N下降有利于N养分释放, 土壤N含量增加.而土壤P含量在恢复早期(LVR-LCQ)下降, 恢复后期(PLL-LCC)显著增加.经相关性分析表明, 土壤N、P含量与土壤C含量均呈显著的正相关关系(相关系数分别为0.917, 0.649,
p < 0.01,
n = 48).表明植被恢复过程中, 土壤有机质的积累有利于土壤养分的提高(
Zhang et al ., 2019 ).此外, 可能是由于恢复早期植物对P的高需求和凋落物的低输入所致, LVR、LCQ植物对P的再吸收效率较高(
图4B ), 凋落物P归还低, 导致土壤P含量下降, 而恢复后期植被多样性和生物量增加(
辜翔等, 2018 ), 随凋落物P归还量增高, 土壤P含量有所提高.由于针叶树种叶片和凋落物酸度较高, 导致针叶林土壤P含量显著低于阔叶林(
Cao et al ., 2018 ), 因而PLL土壤P含量低于LCC.表明植被恢复形成不同的植物群落是影响土壤P含量的主要因素. ...
... ), 凋落物P归还低, 导致土壤P含量下降, 而恢复后期植被多样性和生物量增加(
辜翔等, 2018 ), 随凋落物P归还量增高, 土壤P含量有所提高.由于针叶树种叶片和凋落物酸度较高, 导致针叶林土壤P含量显著低于阔叶林(
Cao et al ., 2018 ), 因而PLL土壤P含量低于LCC.表明植被恢复形成不同的植物群落是影响土壤P含量的主要因素. ...
中亚热带植被恢复对土壤有机碳含量、碳密度的影响
4
2018
... 采用空间代替时间的方法, 按照亚热带森林群落自然演替进程, 根据植被恢复程度及其群落树种组成, 选取地域相邻、环境条件(立地、气候和土壤)相似, 处于不同恢复阶段的4种生态系统: (1)檵木(
Loropetalum chinensis ) +南烛(
Vaccinium bracteatum ) +杜鹃(
Rhododendron mariesii )灌草丛(LVR): 1965年砍伐常绿阔叶林后, 炼山、人工整地, 1966年营造马尾松人工林, 无施肥历史, 1990年皆伐马尾松人工林.2012年, 停止各种人为干扰(砍杂、放牧), 让其自然恢复, 当前草本植物多且生长良好, 伴随分布许多幼小灌木植物, 属于恢复早期阶段.(2)檵木+杉木+白栎(
Quercus fabri )灌木林(LCQ): 1965年天然常绿阔叶林采伐后, 炼山、人工整地, 1966年营造杉木人工纯林, 无施肥历史, 1989年皆伐杉木人工纯林.2004年停止各种人为干扰, 让其自然恢复, 灌木植物发育良好, 但没有形成明显的乔木层, 草本植物稀少, 属于恢复中期第I阶段.(3)马尾松+柯(
Lithocarpus glaber ) +檵木针阔混交林(PLL): 20世纪60年代末, 天然常绿阔叶林皆伐后, 让其自然恢复, 目前已恢复为针阔混交林, 年龄为45-50年, 群落内幼苗幼树丰富, 植物密度较大, 但大直径个体所占比例相对较低, 属于恢复中期第II阶段.(4)柯+红淡比(
Cleyera japonica ) +青冈(
Cyclobalanopsis Glauca )常绿阔叶林(LCC): 长期以来无明显人为干扰, 保存比较完好, 群落结构相对稳定, 年龄为80-90年, 属于恢复后期(亚顶极)阶段.在LVR、LCQ各设置4块20 m × 20 m固定样地, 在PLL、LCC分别设置4块30 m × 30 m的固定样地.群落调查方法和4种群落的基本特征及其主要树种组成详见
辜翔等(2018) 的文献. ...
... 为了避免偶然性, 分别在2016年4、6、10月下旬采集植物叶片、凋落物、0-30 cm土层分析样品, 取3次采样测定的平均值作为最终的数据.在LVR固定样地, 沿着对角线均匀设置3个1 m × 1 m样方, 采集样方内所有灌木叶片、草本植物地上部分, 将同一样地3个样方植物叶片等比例混合为1个样品.根据群落调查结果(
辜翔等, 2018 ), 确定LCQ、PLL和LCC每块固定样地的优势树种, 对该固定样地的每种优势树种分别选取9株长势良好的样木.每种优势树种每次采集3株样木的叶片, 按上、中、下3个部位的东、西、南、北4个方向采集各株样木已完全成熟、无虫害的叶片, 草本植物地上部分采集同LVR, 再将同一样地内草本植物和优势树种叶片按生物量折算比例混合为1个样品.按
郑路和卢立华(2012) 的凋落物层分层标准, 在完成了草本植物地上部分样品采集后的3个1 m × 1 m的样方内采集未分解层的凋落物样品.完成凋落物采集后, 将样方内剩余凋落物清理干净, 用土钻(内径10 cm)取0-30 cm土层土壤样品, 将同一样地3个样方的凋落物、土壤分别混合为1个样品.所有的分析样品每次采样4个重复, 3次采样12个重复. ...
... 森林恢复是决定土壤C、N、P含量变化的关键因素(
Xu et al ., 2018 ).有关本研究样地土壤C含量随着植被恢复而增加的机理已另有论文(
辜翔等, 2018 )详细讨论, 在这里不再重复.土壤N含量随植被恢复逐渐增加, 可能是受到凋落物养分释放的影响, 随着植被恢复, 凋落物C:N下降有利于N养分释放, 土壤N含量增加.而土壤P含量在恢复早期(LVR-LCQ)下降, 恢复后期(PLL-LCC)显著增加.经相关性分析表明, 土壤N、P含量与土壤C含量均呈显著的正相关关系(相关系数分别为0.917, 0.649,
p < 0.01,
n = 48).表明植被恢复过程中, 土壤有机质的积累有利于土壤养分的提高(
Zhang et al ., 2019 ).此外, 可能是由于恢复早期植物对P的高需求和凋落物的低输入所致, LVR、LCQ植物对P的再吸收效率较高(
图4B ), 凋落物P归还低, 导致土壤P含量下降, 而恢复后期植被多样性和生物量增加(
辜翔等, 2018 ), 随凋落物P归还量增高, 土壤P含量有所提高.由于针叶树种叶片和凋落物酸度较高, 导致针叶林土壤P含量显著低于阔叶林(
Cao et al ., 2018 ), 因而PLL土壤P含量低于LCC.表明植被恢复形成不同的植物群落是影响土壤P含量的主要因素. ...
... ), 凋落物P归还低, 导致土壤P含量下降, 而恢复后期植被多样性和生物量增加(
辜翔等, 2018 ), 随凋落物P归还量增高, 土壤P含量有所提高.由于针叶树种叶片和凋落物酸度较高, 导致针叶林土壤P含量显著低于阔叶林(
Cao et al ., 2018 ), 因而PLL土壤P含量低于LCC.表明植被恢复形成不同的植物群落是影响土壤P含量的主要因素. ...
N:P ratios in terrestrial plants: Variation and functional significance
1
2004
... 研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 ).植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 ).凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 ).因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略.本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化.LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境.LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N.PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 ).PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求.LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 ).常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降.研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 ).本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性. ...
Variation in nitrogen and phosphorus concentrations of wetland plants
1
2002
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
Litter N:P ratios indicate whether N or P limits the decomposability of graminoid leaf litter
2
2006
... 本研究中, PLL凋落物C:N、C:P显著高于其他3个恢复阶段, 且随植被恢复的变化与C含量的变化一致, 表明是由于PLL凋落物以马尾松针叶为主, C含量较高所致.研究发现, 随着植被恢复, 凋落物N:P增加(
Zeng et al ., 2017b ).而本研究中, 随着植被恢复, 凋落物N:P有较大的波动, 其中PLL最高, 可能与凋落物P含量有关.随着植被恢复, 凋落物N:P的变化与P含量(
图1 )、P的再吸收效率(
图4B )的变化一致, 由于PLL对P的再吸收利用最高(
图4B ), 凋落物P含量下降, 导致凋落物N:P增高.研究表明, 当凋落物N:P > 25时, 其分解受到P限制(
Güsewell & Verhoeven, 2006 ).本研究中, 凋落物N:P (30-45)明显高于25, 表明本研究区凋落物分解受到P限制. ...
... 凋落物是养分从植物到土壤的基本载体, 其动态交换可以实现并维持土壤养分和植物生长所需的元素比率之间的平衡(
Hessen et al ., 2004 ).凋落物养分含量及其化学计量比与土壤养分含量和化学计量比显著相关(
Yang et al ., 2018 ), 一方面, 表明凋落物分解促使有机物质向土壤释放N、P, 从而影响土壤C、N和P含量及其化学计量比(
Zhong et al ., 2013 ), 另一方面, 表明土壤养分含量的变化会影响植物对养分的利用策略, 进而影响凋落物养分含量及其化学计量比.本研究中, 土壤C、N含量与凋落物C、N含量显著正相关, 可能是随着植被恢复, 土壤C、N含量增加, 更能满足植物对养分的需求, 从而影响植物对养分的再吸收效率, 进而影响凋落物的养分含量.因此, 叶片N、P含量显著影响着凋落物养分含量.此外, 本研究中, 土壤C:N、C:P和N:P与凋落物C:N、C:P显著正相关, 与
汪宗飞和郑粉莉(2018) 、
张萍等(2018) 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 ).也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加. ...
幂指数异速生长机制模型综述
1
2008
... 式中,
x、y 是生物属性,
a、b 均为常数.在实际研究中, 异速生长关系函数通常以对数形式表示, 公式(
韩文轩和方精云, 2008 )如下: ...
幂指数异速生长机制模型综述
1
2008
... 式中,
x、y 是生物属性,
a、b 均为常数.在实际研究中, 异速生长关系函数通常以对数形式表示, 公式(
韩文轩和方精云, 2008 )如下: ...
Leaves nitrogen: Phosphorus stoichiometry across Chinese grassland biomes
1
2008
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
Carbon storage capacity of monoculture and mixed-?species plantations in subtropical China
3
2013
... 因植被恢复而形成不同功能性状的树种组成和结构(
Xu et al ., 2018 ), 导致植物养分吸收、凋落物数量和质量及其分解的差异, 进而影响土壤环境(
Yang & Luo, 2011 ).研究表明, 草本植物C含量低于灌木、乔木, 即灌木、乔木可以合成和积累更多的有机物, 形成更高的C含量(
He et al ., 2013 ).本研究中, 随着植被恢复, 叶片C含量增高, 与
He等(2013) 的研究结果一致, 与群落组成和结构随着植被恢复的变化有关.此外, 本研究中, 叶片对N、P的利用效率随植被恢复显著下降(
图4A ), 即N、P吸收量和存留量增高, 光合作用增强, 有利于干物质积累, 叶片C得到有效的积累.研究发现, 常绿乔木叶片寿命较长, 需要积累更多有机物质(如木质素等)构建保卫结构(
秦海等, 2010 ), 植物体保持较高的N、P含量, 有利于维持植被的生长代谢(
Zeng et al ., 2016 ), 乔木的生长相较于草本需要更多的N、P用于维持代谢所需.本研究中, 随着植被恢复, 叶片N含量显著增高.可能是由于随着植被恢复, 生物量增加促使植物需要更多富含N的物质(如酶、运输蛋白质和氨基酸)参与代谢活动, 加大叶片对N的吸收, 使得叶片N含量增加(
Qin et al ., 2016 ).而土壤N含量随植被恢复而增高, 能更大程度满足植物生长的需求, 也进一步解释了植物叶片N含量的变化(
Wang et al ., 2015 ).由于亚热带丘陵红壤区土壤严重缺P, 不同植被类型对P的再吸收利用效率不同.本研究中, PLL对P的再吸收利用效率最高(
图4B ), 其次是LCC, 这可能是PLL叶片P含量高于LCC, 而LCC高于LCQ、LVR的主要原因.随植被恢复, 植物叶片C、N、P含量呈递增趋势, 表明植被从灌草群落发展为灌木林、乔木林群落, 秉承了从资源保守策略转变为资源快速获取策略的特性(
Royer et al ., 2010 ), 更好地适应环境的变化. ...
... ).本研究中, 随着植被恢复, 叶片C含量增高, 与
He等(2013) 的研究结果一致, 与群落组成和结构随着植被恢复的变化有关.此外, 本研究中, 叶片对N、P的利用效率随植被恢复显著下降(
图4A ), 即N、P吸收量和存留量增高, 光合作用增强, 有利于干物质积累, 叶片C得到有效的积累.研究发现, 常绿乔木叶片寿命较长, 需要积累更多有机物质(如木质素等)构建保卫结构(
秦海等, 2010 ), 植物体保持较高的N、P含量, 有利于维持植被的生长代谢(
Zeng et al ., 2016 ), 乔木的生长相较于草本需要更多的N、P用于维持代谢所需.本研究中, 随着植被恢复, 叶片N含量显著增高.可能是由于随着植被恢复, 生物量增加促使植物需要更多富含N的物质(如酶、运输蛋白质和氨基酸)参与代谢活动, 加大叶片对N的吸收, 使得叶片N含量增加(
Qin et al ., 2016 ).而土壤N含量随植被恢复而增高, 能更大程度满足植物生长的需求, 也进一步解释了植物叶片N含量的变化(
Wang et al ., 2015 ).由于亚热带丘陵红壤区土壤严重缺P, 不同植被类型对P的再吸收利用效率不同.本研究中, PLL对P的再吸收利用效率最高(
图4B ), 其次是LCC, 这可能是PLL叶片P含量高于LCC, 而LCC高于LCQ、LVR的主要原因.随植被恢复, 植物叶片C、N、P含量呈递增趋势, 表明植被从灌草群落发展为灌木林、乔木林群落, 秉承了从资源保守策略转变为资源快速获取策略的特性(
Royer et al ., 2010 ), 更好地适应环境的变化. ...
... 凋落物养分含量明显受到群落优势植物及其凋落物组成的影响.研究发现, 林地凋落物C含量高于草地(
Paul et al ., 2002 ), 针叶林凋落物C含量高于阔叶林(
He et al ., 2013 ).本研究中, 随着植被恢复, 凋落物C含量显著提高, PLL最大, 与
Zeng等(2017a) 的研究结果一致, 是由于PLL以马尾松为优势树种, 凋落物主要以马尾松针叶为主, 而LVR以草本植物和小灌木为主, LCQ以落叶灌木为主, LCC以常绿阔叶树种为主所致.本研究中, 随着植被恢复, 凋落物N、P含量波动较大, 可能是由于不同恢复阶段植物对N、P的再吸收利用效率(
图4B )以及凋落物组成不同所致, 其中, PLL对N、P的再吸收利用效率最高, 其次LCC, 因而PLL凋落物N、P含量低于LCC.此外, 当凋落物N、P含量分别低于7、0.5 g·kg
-1 时, 表明凋落物N、P养分被叶片完全吸收; 而N、P含量分别高于10、0.8 g·kg
-1 时, 则表明没有完全吸收(
Killingbeck, 1996 ;
Yang et al ., 2018 ).本研究中, 凋落物N含量(12-18 g·kg
-1 )高于7 g·kg
-1 , P含量(0.31-0.49 g·kg
-1 )低于0.5 g·kg
-1 , 表明不同植被恢复阶段凋落物N均未被叶片完全吸收, 而P被完全吸收.可能是由于研究区林地土壤N较为丰富, 而P较为缺乏, 植物通过提高对P的再吸收利用以适应土壤P匮乏, 导致叶片对N的再吸收效率显著低于P再吸收效率. ...
Biodiversity and ecosystem multifunctionality
1
2007
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
Global organization of terrestrial plant-?nutrient interactions
1
2004
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
Carbon sequestration in ecosystems: The role of stoichiometry
1
2004
... 凋落物是养分从植物到土壤的基本载体, 其动态交换可以实现并维持土壤养分和植物生长所需的元素比率之间的平衡(
Hessen et al ., 2004 ).凋落物养分含量及其化学计量比与土壤养分含量和化学计量比显著相关(
Yang et al ., 2018 ), 一方面, 表明凋落物分解促使有机物质向土壤释放N、P, 从而影响土壤C、N和P含量及其化学计量比(
Zhong et al ., 2013 ), 另一方面, 表明土壤养分含量的变化会影响植物对养分的利用策略, 进而影响凋落物养分含量及其化学计量比.本研究中, 土壤C、N含量与凋落物C、N含量显著正相关, 可能是随着植被恢复, 土壤C、N含量增加, 更能满足植物对养分的需求, 从而影响植物对养分的再吸收效率, 进而影响凋落物的养分含量.因此, 叶片N、P含量显著影响着凋落物养分含量.此外, 本研究中, 土壤C:N、C:P和N:P与凋落物C:N、C:P显著正相关, 与
汪宗飞和郑粉莉(2018) 、
张萍等(2018) 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 ).也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加. ...
Storage of C, N, and P affected by afforestation with
Salix cupularis in an alpine semiarid desert ecosystem
1
2018
... 研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 ).植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 ).凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 ).因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略.本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化.LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境.LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N.PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 ).PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求.LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 ).常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降.研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 ).本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性. ...
1
1978
... 植物叶片、凋落物、土壤样品有机碳含量用重铬酸钾-浓硫酸滴定法测定, 全氮含量用K9840自动凯氏定氮仪(海能仪器公司, 济南)测定, 全磷含量用碱熔-钼锑抗比色法测定(
中国科学院南京土壤研究所, 1978 ). ...
1
1978
... 植物叶片、凋落物、土壤样品有机碳含量用重铬酸钾-浓硫酸滴定法测定, 全氮含量用K9840自动凯氏定氮仪(海能仪器公司, 济南)测定, 全磷含量用碱熔-钼锑抗比色法测定(
中国科学院南京土壤研究所, 1978 ). ...
闽江河口湿地3种植物化学计量内稳性特征
1
2014
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
闽江河口湿地3种植物化学计量内稳性特征
1
2014
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
Nutrients in senesced leaves: Keys to the search for potential resorption and resorption proficiency
1
1996
... 凋落物养分含量明显受到群落优势植物及其凋落物组成的影响.研究发现, 林地凋落物C含量高于草地(
Paul et al ., 2002 ), 针叶林凋落物C含量高于阔叶林(
He et al ., 2013 ).本研究中, 随着植被恢复, 凋落物C含量显著提高, PLL最大, 与
Zeng等(2017a) 的研究结果一致, 是由于PLL以马尾松为优势树种, 凋落物主要以马尾松针叶为主, 而LVR以草本植物和小灌木为主, LCQ以落叶灌木为主, LCC以常绿阔叶树种为主所致.本研究中, 随着植被恢复, 凋落物N、P含量波动较大, 可能是由于不同恢复阶段植物对N、P的再吸收利用效率(
图4B )以及凋落物组成不同所致, 其中, PLL对N、P的再吸收利用效率最高, 其次LCC, 因而PLL凋落物N、P含量低于LCC.此外, 当凋落物N、P含量分别低于7、0.5 g·kg
-1 时, 表明凋落物N、P养分被叶片完全吸收; 而N、P含量分别高于10、0.8 g·kg
-1 时, 则表明没有完全吸收(
Killingbeck, 1996 ;
Yang et al ., 2018 ).本研究中, 凋落物N含量(12-18 g·kg
-1 )高于7 g·kg
-1 , P含量(0.31-0.49 g·kg
-1 )低于0.5 g·kg
-1 , 表明不同植被恢复阶段凋落物N均未被叶片完全吸收, 而P被完全吸收.可能是由于研究区林地土壤N较为丰富, 而P较为缺乏, 植物通过提高对P的再吸收利用以适应土壤P匮乏, 导致叶片对N的再吸收效率显著低于P再吸收效率. ...
The vegetation N:P ratio: A new tool to detect the nature of nutrients limitation
1
1996
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
The stoichiometry of animal energetics
1
1995
... (4)植物内稳态反映植物随着环境变化保持自身化学组成稳定的能力(
Kooijman, 1995 ), 常用以下线性回归方程拟合: ...
Deep soil C, N, and P stocks and stoichiometry in response to land use patterns in the Loess Hilly Region of China
2
2016
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
... ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
云南普洱季风常绿阔叶林演替系列植物和土壤C、N、P化学计量特征
1
2010
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
云南普洱季风常绿阔叶林演替系列植物和土壤C、N、P化学计量特征
1
2010
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
长白山四种森林类型凋落物分解动态
1
2009
... 研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 ).植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 ).凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 ).因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略.本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化.LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境.LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N.PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 ).PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求.LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 ).常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降.研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 ).本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性. ...
长白山四种森林类型凋落物分解动态
1
2009
... 研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 ).植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 ).凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 ).因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略.本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化.LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境.LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N.PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 ).PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求.LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 ).常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降.研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 ).本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性. ...
Does the gradualness of leaf shedding govern nutrient resorption from senescing leaves in Mediterranean woody plants?
1
2005
... (3)养分再吸收效率(
NRE )是指养分从凋落物中转移并被新生组织再利用的效率, 叶片N、P的再吸收效率计算公式(
Milla et al ., 2005 )如下: ...
Stoichiometric controls of nitrogen and phosphorus cycling in decomposing beech leaf litter
1
2012
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
鄱阳湖湿地优势植物叶片-凋落物-土壤碳氮磷化学计量特征
1
2016
... 凋落物是养分从植物到土壤的基本载体, 其动态交换可以实现并维持土壤养分和植物生长所需的元素比率之间的平衡(
Hessen et al ., 2004 ).凋落物养分含量及其化学计量比与土壤养分含量和化学计量比显著相关(
Yang et al ., 2018 ), 一方面, 表明凋落物分解促使有机物质向土壤释放N、P, 从而影响土壤C、N和P含量及其化学计量比(
Zhong et al ., 2013 ), 另一方面, 表明土壤养分含量的变化会影响植物对养分的利用策略, 进而影响凋落物养分含量及其化学计量比.本研究中, 土壤C、N含量与凋落物C、N含量显著正相关, 可能是随着植被恢复, 土壤C、N含量增加, 更能满足植物对养分的需求, 从而影响植物对养分的再吸收效率, 进而影响凋落物的养分含量.因此, 叶片N、P含量显著影响着凋落物养分含量.此外, 本研究中, 土壤C:N、C:P和N:P与凋落物C:N、C:P显著正相关, 与
汪宗飞和郑粉莉(2018) 、
张萍等(2018) 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 ).也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加. ...
鄱阳湖湿地优势植物叶片-凋落物-土壤碳氮磷化学计量特征
1
2016
... 凋落物是养分从植物到土壤的基本载体, 其动态交换可以实现并维持土壤养分和植物生长所需的元素比率之间的平衡(
Hessen et al ., 2004 ).凋落物养分含量及其化学计量比与土壤养分含量和化学计量比显著相关(
Yang et al ., 2018 ), 一方面, 表明凋落物分解促使有机物质向土壤释放N、P, 从而影响土壤C、N和P含量及其化学计量比(
Zhong et al ., 2013 ), 另一方面, 表明土壤养分含量的变化会影响植物对养分的利用策略, 进而影响凋落物养分含量及其化学计量比.本研究中, 土壤C、N含量与凋落物C、N含量显著正相关, 可能是随着植被恢复, 土壤C、N含量增加, 更能满足植物对养分的需求, 从而影响植物对养分的再吸收效率, 进而影响凋落物的养分含量.因此, 叶片N、P含量显著影响着凋落物养分含量.此外, 本研究中, 土壤C:N、C:P和N:P与凋落物C:N、C:P显著正相关, 与
汪宗飞和郑粉莉(2018) 、
张萍等(2018) 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 ).也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加. ...
Size-dependent variations in plant growth rates and the “3/4-power rule”
1
1994
... (1)异速生长反映植物生长发育过程中有机体内两种属性(例如C、N、P含量)之间的变化规律, 常用公式(
Niklas, 1994 )如下: ...
亚热带红壤侵蚀区马尾松针叶养分含量及再吸收特征
1
2016
... 研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 ).植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 ).凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 ).因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略.本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化.LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境.LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N.PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 ).PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求.LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 ).常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降.研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 ).本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性. ...
亚热带红壤侵蚀区马尾松针叶养分含量及再吸收特征
1
2016
... 研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 ).植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 ).凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 ).因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略.本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化.LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境.LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N.PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 ).PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求.LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 ).常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降.研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 ).本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性. ...
Physical and chemical protection in hierarchical soil aggregates regulates soil carbon and nitrogen recovery in restored perennial grasslands
1
2013
... 植被恢复涉及植物群落与土壤环境的协调发展, 因其能改善脆弱或退化生态系统养分循环、土壤质量而受到广泛关注(
Xu et al ., 2019 ).随着植被恢复, 植物群落组成结构、土壤理化性质达到稳定状态(
O’Brien & Jastrow, 2013 ), 生态系统各组分(植物、凋落物、土壤)碳(C)、氮(N)、磷(P)含量及其循环规律调控着植物的生存发展和各种生态过程(
Reich et al ., 2006 ), 其化学计量比反映植物组成动态与土壤养分之间的平衡(
Zhang et al ., 2018 ).因此, 研究生态系统各组分C、N、P化学计量比随植被恢复的变化对理解各组分之间养分关系及分配特征十分必要, 有助于明确植被恢复过程中, 植物群落随土壤环境变化的发展方向(
Zhao et al ., 2015b ;
Ren et al ., 2016 ). ...
Change in soil carbon following afforestation
2
2002
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
... 凋落物养分含量明显受到群落优势植物及其凋落物组成的影响.研究发现, 林地凋落物C含量高于草地(
Paul et al ., 2002 ), 针叶林凋落物C含量高于阔叶林(
He et al ., 2013 ).本研究中, 随着植被恢复, 凋落物C含量显著提高, PLL最大, 与
Zeng等(2017a) 的研究结果一致, 是由于PLL以马尾松为优势树种, 凋落物主要以马尾松针叶为主, 而LVR以草本植物和小灌木为主, LCQ以落叶灌木为主, LCC以常绿阔叶树种为主所致.本研究中, 随着植被恢复, 凋落物N、P含量波动较大, 可能是由于不同恢复阶段植物对N、P的再吸收利用效率(
图4B )以及凋落物组成不同所致, 其中, PLL对N、P的再吸收利用效率最高, 其次LCC, 因而PLL凋落物N、P含量低于LCC.此外, 当凋落物N、P含量分别低于7、0.5 g·kg
-1 时, 表明凋落物N、P养分被叶片完全吸收; 而N、P含量分别高于10、0.8 g·kg
-1 时, 则表明没有完全吸收(
Killingbeck, 1996 ;
Yang et al ., 2018 ).本研究中, 凋落物N含量(12-18 g·kg
-1 )高于7 g·kg
-1 , P含量(0.31-0.49 g·kg
-1 )低于0.5 g·kg
-1 , 表明不同植被恢复阶段凋落物N均未被叶片完全吸收, 而P被完全吸收.可能是由于研究区林地土壤N较为丰富, 而P较为缺乏, 植物通过提高对P的再吸收利用以适应土壤P匮乏, 导致叶片对N的再吸收效率显著低于P再吸收效率. ...
中亚热带4种森林土壤碳、氮、磷化学计量特征
1
2016
... 中国亚热带地区水热条件优越, 森林资源丰富, 是中国重要的生态屏障.但由于长期的人为干扰, 该地区地带性植被——常绿阔叶林遭到了严重的破坏.为保护生物多样性和提升森林生态系统服务, 促进森林植被恢复, 近20多年来, 中国政府实施了天然林保护、长江中上游防护林体系建设等系列林业生态工程, 该地区森林植被恢复迅速, 由于人为干扰强度不同, 形成了处于不同恢复阶段的多种次生林群落(
Xiang et al ., 2013 ).目前针对亚热带森林植被恢复过程中, 不同植物组织(
阎恩荣等, 2010 ;
Zeng et al ., 2017b )、土壤(
彭晓等, 2016 ;
Xu et al ., 2018 )的化学计量动态已有一些研究报道.然而, 将植物叶片-凋落物-土壤作为一个系统, 探讨其C、N、P化学计量动态及其各组分之间相互作用的研究鲜有报道, 植被恢复对整个生态系统养分运输和分配影响如何仍然不是很清楚.为此, 本研究根据亚热带森林群落演替进程, 在湘中丘陵区选取了4种处于不同恢复阶段(灌草丛、灌木林、针阔混交林、常绿阔叶林)的生态系统作为一个恢复序列, 研究植物叶片、凋落物、土壤C、N、P含量、化学计量比的变化及其相互之间的关联性, 探讨随植被恢复, 植物群落对土壤环境变化的响应及其养分利用策略, 剖析生态系统各组分之间养分动态平衡关系随着植被恢复的变化特征, 为亚热带森林植被恢复和养分管理提供科学依据. ...
中亚热带4种森林土壤碳、氮、磷化学计量特征
1
2016
... 中国亚热带地区水热条件优越, 森林资源丰富, 是中国重要的生态屏障.但由于长期的人为干扰, 该地区地带性植被——常绿阔叶林遭到了严重的破坏.为保护生物多样性和提升森林生态系统服务, 促进森林植被恢复, 近20多年来, 中国政府实施了天然林保护、长江中上游防护林体系建设等系列林业生态工程, 该地区森林植被恢复迅速, 由于人为干扰强度不同, 形成了处于不同恢复阶段的多种次生林群落(
Xiang et al ., 2013 ).目前针对亚热带森林植被恢复过程中, 不同植物组织(
阎恩荣等, 2010 ;
Zeng et al ., 2017b )、土壤(
彭晓等, 2016 ;
Xu et al ., 2018 )的化学计量动态已有一些研究报道.然而, 将植物叶片-凋落物-土壤作为一个系统, 探讨其C、N、P化学计量动态及其各组分之间相互作用的研究鲜有报道, 植被恢复对整个生态系统养分运输和分配影响如何仍然不是很清楚.为此, 本研究根据亚热带森林群落演替进程, 在湘中丘陵区选取了4种处于不同恢复阶段(灌草丛、灌木林、针阔混交林、常绿阔叶林)的生态系统作为一个恢复序列, 研究植物叶片、凋落物、土壤C、N、P含量、化学计量比的变化及其相互之间的关联性, 探讨随植被恢复, 植物群落对土壤环境变化的响应及其养分利用策略, 剖析生态系统各组分之间养分动态平衡关系随着植被恢复的变化特征, 为亚热带森林植被恢复和养分管理提供科学依据. ...
中国660种陆生植物叶片8种元素含量特征
2
2010
... 因植被恢复而形成不同功能性状的树种组成和结构(
Xu et al ., 2018 ), 导致植物养分吸收、凋落物数量和质量及其分解的差异, 进而影响土壤环境(
Yang & Luo, 2011 ).研究表明, 草本植物C含量低于灌木、乔木, 即灌木、乔木可以合成和积累更多的有机物, 形成更高的C含量(
He et al ., 2013 ).本研究中, 随着植被恢复, 叶片C含量增高, 与
He等(2013) 的研究结果一致, 与群落组成和结构随着植被恢复的变化有关.此外, 本研究中, 叶片对N、P的利用效率随植被恢复显著下降(
图4A ), 即N、P吸收量和存留量增高, 光合作用增强, 有利于干物质积累, 叶片C得到有效的积累.研究发现, 常绿乔木叶片寿命较长, 需要积累更多有机物质(如木质素等)构建保卫结构(
秦海等, 2010 ), 植物体保持较高的N、P含量, 有利于维持植被的生长代谢(
Zeng et al ., 2016 ), 乔木的生长相较于草本需要更多的N、P用于维持代谢所需.本研究中, 随着植被恢复, 叶片N含量显著增高.可能是由于随着植被恢复, 生物量增加促使植物需要更多富含N的物质(如酶、运输蛋白质和氨基酸)参与代谢活动, 加大叶片对N的吸收, 使得叶片N含量增加(
Qin et al ., 2016 ).而土壤N含量随植被恢复而增高, 能更大程度满足植物生长的需求, 也进一步解释了植物叶片N含量的变化(
Wang et al ., 2015 ).由于亚热带丘陵红壤区土壤严重缺P, 不同植被类型对P的再吸收利用效率不同.本研究中, PLL对P的再吸收利用效率最高(
图4B ), 其次是LCC, 这可能是PLL叶片P含量高于LCC, 而LCC高于LCQ、LVR的主要原因.随植被恢复, 植物叶片C、N、P含量呈递增趋势, 表明植被从灌草群落发展为灌木林、乔木林群落, 秉承了从资源保守策略转变为资源快速获取策略的特性(
Royer et al ., 2010 ), 更好地适应环境的变化. ...
... 研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 ).植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 ).凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 ).因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略.本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化.LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境.LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N.PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 ).PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求.LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 ).常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降.研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 ).本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性. ...
中国660种陆生植物叶片8种元素含量特征
2
2010
... 因植被恢复而形成不同功能性状的树种组成和结构(
Xu et al ., 2018 ), 导致植物养分吸收、凋落物数量和质量及其分解的差异, 进而影响土壤环境(
Yang & Luo, 2011 ).研究表明, 草本植物C含量低于灌木、乔木, 即灌木、乔木可以合成和积累更多的有机物, 形成更高的C含量(
He et al ., 2013 ).本研究中, 随着植被恢复, 叶片C含量增高, 与
He等(2013) 的研究结果一致, 与群落组成和结构随着植被恢复的变化有关.此外, 本研究中, 叶片对N、P的利用效率随植被恢复显著下降(
图4A ), 即N、P吸收量和存留量增高, 光合作用增强, 有利于干物质积累, 叶片C得到有效的积累.研究发现, 常绿乔木叶片寿命较长, 需要积累更多有机物质(如木质素等)构建保卫结构(
秦海等, 2010 ), 植物体保持较高的N、P含量, 有利于维持植被的生长代谢(
Zeng et al ., 2016 ), 乔木的生长相较于草本需要更多的N、P用于维持代谢所需.本研究中, 随着植被恢复, 叶片N含量显著增高.可能是由于随着植被恢复, 生物量增加促使植物需要更多富含N的物质(如酶、运输蛋白质和氨基酸)参与代谢活动, 加大叶片对N的吸收, 使得叶片N含量增加(
Qin et al ., 2016 ).而土壤N含量随植被恢复而增高, 能更大程度满足植物生长的需求, 也进一步解释了植物叶片N含量的变化(
Wang et al ., 2015 ).由于亚热带丘陵红壤区土壤严重缺P, 不同植被类型对P的再吸收利用效率不同.本研究中, PLL对P的再吸收利用效率最高(
图4B ), 其次是LCC, 这可能是PLL叶片P含量高于LCC, 而LCC高于LCQ、LVR的主要原因.随植被恢复, 植物叶片C、N、P含量呈递增趋势, 表明植被从灌草群落发展为灌木林、乔木林群落, 秉承了从资源保守策略转变为资源快速获取策略的特性(
Royer et al ., 2010 ), 更好地适应环境的变化. ...
... 研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 ).植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 ).凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 ).因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略.本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化.LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境.LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N.PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 ).PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求.LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 ).常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降.研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 ).本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性. ...
Effects of forest plantation types on leaf traits of
Ulmus pumila and
Robinia pseudoacacia on the Loess Plateau, China
1
2016
... 因植被恢复而形成不同功能性状的树种组成和结构(
Xu et al ., 2018 ), 导致植物养分吸收、凋落物数量和质量及其分解的差异, 进而影响土壤环境(
Yang & Luo, 2011 ).研究表明, 草本植物C含量低于灌木、乔木, 即灌木、乔木可以合成和积累更多的有机物, 形成更高的C含量(
He et al ., 2013 ).本研究中, 随着植被恢复, 叶片C含量增高, 与
He等(2013) 的研究结果一致, 与群落组成和结构随着植被恢复的变化有关.此外, 本研究中, 叶片对N、P的利用效率随植被恢复显著下降(
图4A ), 即N、P吸收量和存留量增高, 光合作用增强, 有利于干物质积累, 叶片C得到有效的积累.研究发现, 常绿乔木叶片寿命较长, 需要积累更多有机物质(如木质素等)构建保卫结构(
秦海等, 2010 ), 植物体保持较高的N、P含量, 有利于维持植被的生长代谢(
Zeng et al ., 2016 ), 乔木的生长相较于草本需要更多的N、P用于维持代谢所需.本研究中, 随着植被恢复, 叶片N含量显著增高.可能是由于随着植被恢复, 生物量增加促使植物需要更多富含N的物质(如酶、运输蛋白质和氨基酸)参与代谢活动, 加大叶片对N的吸收, 使得叶片N含量增加(
Qin et al ., 2016 ).而土壤N含量随植被恢复而增高, 能更大程度满足植物生长的需求, 也进一步解释了植物叶片N含量的变化(
Wang et al ., 2015 ).由于亚热带丘陵红壤区土壤严重缺P, 不同植被类型对P的再吸收利用效率不同.本研究中, PLL对P的再吸收利用效率最高(
图4B ), 其次是LCC, 这可能是PLL叶片P含量高于LCC, 而LCC高于LCQ、LVR的主要原因.随植被恢复, 植物叶片C、N、P含量呈递增趋势, 表明植被从灌草群落发展为灌木林、乔木林群落, 秉承了从资源保守策略转变为资源快速获取策略的特性(
Royer et al ., 2010 ), 更好地适应环境的变化. ...
R: A Language and Environment for Statistical Computing. 1
2018
... C、N、P含量用单位质量的含量(g·kg
-1 )表示, C:N、C:P、N:P为质量比.用Excel 2010统计各项指标的平均值和标准偏差, 用SigmaPlot 12.0制图.用SPSS 19.0单因素方差分析进行显著性(
p < 0.05)检验.用R 3.5.1中Corrplot包绘制叶片、凋落物、土壤C、N、P含量及其化学计量比的Pearson相关性图.用R 3.5.1中Smart包拟合叶片N、P含量的标准化主轴回归(SMA)关系, 即异速生长关系(
Warton et al ., 2012 ;
R Development Core Team, 2018 ). ...
Universal scaling of respiratory metabolism, size and nitrogen in plants
1
2006
... 植被恢复涉及植物群落与土壤环境的协调发展, 因其能改善脆弱或退化生态系统养分循环、土壤质量而受到广泛关注(
Xu et al ., 2019 ).随着植被恢复, 植物群落组成结构、土壤理化性质达到稳定状态(
O’Brien & Jastrow, 2013 ), 生态系统各组分(植物、凋落物、土壤)碳(C)、氮(N)、磷(P)含量及其循环规律调控着植物的生存发展和各种生态过程(
Reich et al ., 2006 ), 其化学计量比反映植物组成动态与土壤养分之间的平衡(
Zhang et al ., 2018 ).因此, 研究生态系统各组分C、N、P化学计量比随植被恢复的变化对理解各组分之间养分关系及分配特征十分必要, 有助于明确植被恢复过程中, 植物群落随土壤环境变化的发展方向(
Zhao et al ., 2015b ;
Ren et al ., 2016 ). ...
Linkages of C:N:P stoichiometry and bacterial community in soil following afforestation of former farmland
2
2016
... 植被恢复涉及植物群落与土壤环境的协调发展, 因其能改善脆弱或退化生态系统养分循环、土壤质量而受到广泛关注(
Xu et al ., 2019 ).随着植被恢复, 植物群落组成结构、土壤理化性质达到稳定状态(
O’Brien & Jastrow, 2013 ), 生态系统各组分(植物、凋落物、土壤)碳(C)、氮(N)、磷(P)含量及其循环规律调控着植物的生存发展和各种生态过程(
Reich et al ., 2006 ), 其化学计量比反映植物组成动态与土壤养分之间的平衡(
Zhang et al ., 2018 ).因此, 研究生态系统各组分C、N、P化学计量比随植被恢复的变化对理解各组分之间养分关系及分配特征十分必要, 有助于明确植被恢复过程中, 植物群落随土壤环境变化的发展方向(
Zhao et al ., 2015b ;
Ren et al ., 2016 ). ...
... 土壤C:N、C:P、N:P可作为土壤养分状况的良好指标(
Zhao et al ., 2015a ), 但因植物群落不同而异, 具有高度复杂性(
Zhang et al ., 2014 ).C、N、P不平衡性增加也会引起土壤C:N、C:P、N:P的变化(
Zhao et al ., 2015b ).本研究中, 土壤C:N (13-19)随着植被恢复先升高后下降, 与
Zhang等(2018) 的研究结果一致, 是由于从LVR到PLL土壤C含量增加幅度(121.66%)高于土壤N含量的增加幅度(79.75%), 导致土壤C:N升高; 而从PLL到LCC, 土壤C含量增加幅度(40.84%)低于土壤N含量的增加幅度(91.65%), 导致土壤C:N下降.这可以从植物对N的利用效率和再吸收效率来解释, 随着植被恢复, 植物对N的利用效率下降, 而恢复后期N的再吸收效率下降, 表明植物对土壤N的吸收量下降, 凋落物N含量较高, 有利于土壤N的积累.土壤C、N含量的变化是影响土壤C:P、N:P的主要因素(
Xu et al ., 2019 ).本研究中, 由于土壤P主要来源于母质的风化, 随植被恢复变化较小; 另一方面, 如前面分析, N的再吸收效率低于P, 凋落物的归还作用主要增加了土壤C、N的投入, 促进了土壤C、N的积累, 因而使得土壤C:P、N:P增加.在中国南方亚热带红壤丘陵区土壤较高的C:P、N:P也可以反映出林地土壤P的限制性(
Gao et al ., 2014 ;
Ren et al ., 2016 ). ...
Leaf economic traits from fossils support a weedy habit for early angiosperms
2
2010
... 因植被恢复而形成不同功能性状的树种组成和结构(
Xu et al ., 2018 ), 导致植物养分吸收、凋落物数量和质量及其分解的差异, 进而影响土壤环境(
Yang & Luo, 2011 ).研究表明, 草本植物C含量低于灌木、乔木, 即灌木、乔木可以合成和积累更多的有机物, 形成更高的C含量(
He et al ., 2013 ).本研究中, 随着植被恢复, 叶片C含量增高, 与
He等(2013) 的研究结果一致, 与群落组成和结构随着植被恢复的变化有关.此外, 本研究中, 叶片对N、P的利用效率随植被恢复显著下降(
图4A ), 即N、P吸收量和存留量增高, 光合作用增强, 有利于干物质积累, 叶片C得到有效的积累.研究发现, 常绿乔木叶片寿命较长, 需要积累更多有机物质(如木质素等)构建保卫结构(
秦海等, 2010 ), 植物体保持较高的N、P含量, 有利于维持植被的生长代谢(
Zeng et al ., 2016 ), 乔木的生长相较于草本需要更多的N、P用于维持代谢所需.本研究中, 随着植被恢复, 叶片N含量显著增高.可能是由于随着植被恢复, 生物量增加促使植物需要更多富含N的物质(如酶、运输蛋白质和氨基酸)参与代谢活动, 加大叶片对N的吸收, 使得叶片N含量增加(
Qin et al ., 2016 ).而土壤N含量随植被恢复而增高, 能更大程度满足植物生长的需求, 也进一步解释了植物叶片N含量的变化(
Wang et al ., 2015 ).由于亚热带丘陵红壤区土壤严重缺P, 不同植被类型对P的再吸收利用效率不同.本研究中, PLL对P的再吸收利用效率最高(
图4B ), 其次是LCC, 这可能是PLL叶片P含量高于LCC, 而LCC高于LCQ、LVR的主要原因.随植被恢复, 植物叶片C、N、P含量呈递增趋势, 表明植被从灌草群落发展为灌木林、乔木林群落, 秉承了从资源保守策略转变为资源快速获取策略的特性(
Royer et al ., 2010 ), 更好地适应环境的变化. ...
... 研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 ).植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 ).凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 ).因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略.本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化.LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境.LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N.PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 ).PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求.LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 ).常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降.研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 ).本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性. ...
Factors influencing the foliar elemental composition and stoichiometry in forest trees in Spain
1
2016
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
Stem, root, and older leaf N:P ratios are more responsive indicators of soil nutrient availability than new foliage
1
2014
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
2
2002
... 式中,
x 为土壤C、N、P含量(g·kg
-1 ),
y 为植物C、N、P含量(g·kg
-1 ),
k 为斜率,
c 为常数.当
k ≠ 0时, 表示植物体该养分含量内稳态弱, 随土壤环境变化而变化; 当
k = 0时, 则表示植物体该养分具有绝对的内稳态, 不受土壤环境变化的影响(
Sterner & Elser, 2002 ). ...
... 研究表明, 土壤养分含量的变化直接影响植物养分吸收及其化学计量比(
Chen et al ., 2018 ).而植物通过改变化学计量比和采用适宜的养分利用策略适应土壤N、P供应的变化(
Sun et al ., 2018 ).植物N含量比P含量更稳定(
Yu et al ., 2010 ).本研究中, 随着植被恢复, 叶片N含量内稳态较弱, 受土壤N、P含量的变化影响较大, 叶片P含量具有较高的内稳态, 不随土壤N、P含量的变化而变化, 与
Yu等(2010) 的研究结果不同.可能是由于植物的限制性养分元素不同所致, 在
Yu等(2010) 的研究中, 植物主要受N限制, 而本研究中, 土壤P含量随植被恢复的变化小, 植物主要受P的限制, 在一定程度上证实了限制性养分元素稳定性假说“由于生理和养分平衡的制约, 限制性养分元素在植物体内的含量具有相对稳定性, 对环境变化的响应也较为稳定” (
Sterner & Elser, 2002 ).表明随着植被恢复, 在P缺乏的土壤环境中, 植物叶片P具有较强的内稳态. ...
植物的养分利用效率(NUE)及植物对养分胁迫环境的适应策略
1
2000
... 研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 ).植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 ).凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 ).因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略.本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化.LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境.LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N.PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 ).PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求.LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 ).常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降.研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 ).本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性. ...
植物的养分利用效率(NUE)及植物对养分胁迫环境的适应策略
1
2000
... 研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 ).植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 ).凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 ).因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略.本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化.LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境.LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N.PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 ).PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求.LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 ).常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降.研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 ).本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性. ...
Initial responses of grass litter tissue chemistry and N:P stoichiometry to varied N and P input rates and ratios in Inner Mongolia
1
2018
... 研究表明, 土壤养分含量的变化直接影响植物养分吸收及其化学计量比(
Chen et al ., 2018 ).而植物通过改变化学计量比和采用适宜的养分利用策略适应土壤N、P供应的变化(
Sun et al ., 2018 ).植物N含量比P含量更稳定(
Yu et al ., 2010 ).本研究中, 随着植被恢复, 叶片N含量内稳态较弱, 受土壤N、P含量的变化影响较大, 叶片P含量具有较高的内稳态, 不随土壤N、P含量的变化而变化, 与
Yu等(2010) 的研究结果不同.可能是由于植物的限制性养分元素不同所致, 在
Yu等(2010) 的研究中, 植物主要受N限制, 而本研究中, 土壤P含量随植被恢复的变化小, 植物主要受P的限制, 在一定程度上证实了限制性养分元素稳定性假说“由于生理和养分平衡的制约, 限制性养分元素在植物体内的含量具有相对稳定性, 对环境变化的响应也较为稳定” (
Sterner & Elser, 2002 ).表明随着植被恢复, 在P缺乏的土壤环境中, 植物叶片P具有较强的内稳态. ...
Soil moisture and texture primarily control the soil nutrient stoichiometry across the Tibetan grassland
1
2018
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
C:N:P stoichiometry and leaf traits of halophytes in an arid saline environment, northwest China
1
2015
... 因植被恢复而形成不同功能性状的树种组成和结构(
Xu et al ., 2018 ), 导致植物养分吸收、凋落物数量和质量及其分解的差异, 进而影响土壤环境(
Yang & Luo, 2011 ).研究表明, 草本植物C含量低于灌木、乔木, 即灌木、乔木可以合成和积累更多的有机物, 形成更高的C含量(
He et al ., 2013 ).本研究中, 随着植被恢复, 叶片C含量增高, 与
He等(2013) 的研究结果一致, 与群落组成和结构随着植被恢复的变化有关.此外, 本研究中, 叶片对N、P的利用效率随植被恢复显著下降(
图4A ), 即N、P吸收量和存留量增高, 光合作用增强, 有利于干物质积累, 叶片C得到有效的积累.研究发现, 常绿乔木叶片寿命较长, 需要积累更多有机物质(如木质素等)构建保卫结构(
秦海等, 2010 ), 植物体保持较高的N、P含量, 有利于维持植被的生长代谢(
Zeng et al ., 2016 ), 乔木的生长相较于草本需要更多的N、P用于维持代谢所需.本研究中, 随着植被恢复, 叶片N含量显著增高.可能是由于随着植被恢复, 生物量增加促使植物需要更多富含N的物质(如酶、运输蛋白质和氨基酸)参与代谢活动, 加大叶片对N的吸收, 使得叶片N含量增加(
Qin et al ., 2016 ).而土壤N含量随植被恢复而增高, 能更大程度满足植物生长的需求, 也进一步解释了植物叶片N含量的变化(
Wang et al ., 2015 ).由于亚热带丘陵红壤区土壤严重缺P, 不同植被类型对P的再吸收利用效率不同.本研究中, PLL对P的再吸收利用效率最高(
图4B ), 其次是LCC, 这可能是PLL叶片P含量高于LCC, 而LCC高于LCQ、LVR的主要原因.随植被恢复, 植物叶片C、N、P含量呈递增趋势, 表明植被从灌草群落发展为灌木林、乔木林群落, 秉承了从资源保守策略转变为资源快速获取策略的特性(
Royer et al ., 2010 ), 更好地适应环境的变化. ...
Carbon, nitrogen, phosphorus, and potassium stoichiometry in an ombrotrophic peatland reflects plant functional type
1
2014
... 研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 ).植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 ).凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 ).因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略.本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化.LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境.LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N.PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 ).PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求.LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 ).常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降.研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 ).本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性. ...
黄土高原子午岭地区人工油松林碳氮磷生态化学计量特征
1
2018
... 凋落物是养分从植物到土壤的基本载体, 其动态交换可以实现并维持土壤养分和植物生长所需的元素比率之间的平衡(
Hessen et al ., 2004 ).凋落物养分含量及其化学计量比与土壤养分含量和化学计量比显著相关(
Yang et al ., 2018 ), 一方面, 表明凋落物分解促使有机物质向土壤释放N、P, 从而影响土壤C、N和P含量及其化学计量比(
Zhong et al ., 2013 ), 另一方面, 表明土壤养分含量的变化会影响植物对养分的利用策略, 进而影响凋落物养分含量及其化学计量比.本研究中, 土壤C、N含量与凋落物C、N含量显著正相关, 可能是随着植被恢复, 土壤C、N含量增加, 更能满足植物对养分的需求, 从而影响植物对养分的再吸收效率, 进而影响凋落物的养分含量.因此, 叶片N、P含量显著影响着凋落物养分含量.此外, 本研究中, 土壤C:N、C:P和N:P与凋落物C:N、C:P显著正相关, 与
汪宗飞和郑粉莉(2018) 、
张萍等(2018) 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 ).也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加. ...
黄土高原子午岭地区人工油松林碳氮磷生态化学计量特征
1
2018
... 凋落物是养分从植物到土壤的基本载体, 其动态交换可以实现并维持土壤养分和植物生长所需的元素比率之间的平衡(
Hessen et al ., 2004 ).凋落物养分含量及其化学计量比与土壤养分含量和化学计量比显著相关(
Yang et al ., 2018 ), 一方面, 表明凋落物分解促使有机物质向土壤释放N、P, 从而影响土壤C、N和P含量及其化学计量比(
Zhong et al ., 2013 ), 另一方面, 表明土壤养分含量的变化会影响植物对养分的利用策略, 进而影响凋落物养分含量及其化学计量比.本研究中, 土壤C、N含量与凋落物C、N含量显著正相关, 可能是随着植被恢复, 土壤C、N含量增加, 更能满足植物对养分的需求, 从而影响植物对养分的再吸收效率, 进而影响凋落物的养分含量.因此, 叶片N、P含量显著影响着凋落物养分含量.此外, 本研究中, 土壤C:N、C:P和N:P与凋落物C:N、C:P显著正相关, 与
汪宗飞和郑粉莉(2018) 、
张萍等(2018) 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 ).也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加. ...
SMART 3—An R package for estimation and inference about allometric lines
1
2012
... C、N、P含量用单位质量的含量(g·kg
-1 )表示, C:N、C:P、N:P为质量比.用Excel 2010统计各项指标的平均值和标准偏差, 用SigmaPlot 12.0制图.用SPSS 19.0单因素方差分析进行显著性(
p < 0.05)检验.用R 3.5.1中Corrplot包绘制叶片、凋落物、土壤C、N、P含量及其化学计量比的Pearson相关性图.用R 3.5.1中Smart包拟合叶片N、P含量的标准化主轴回归(SMA)关系, 即异速生长关系(
Warton et al ., 2012 ;
R Development Core Team, 2018 ). ...
Secondary forest floristic composition, structure, and spatial pattern in subtropical China
1
2013
... 中国亚热带地区水热条件优越, 森林资源丰富, 是中国重要的生态屏障.但由于长期的人为干扰, 该地区地带性植被——常绿阔叶林遭到了严重的破坏.为保护生物多样性和提升森林生态系统服务, 促进森林植被恢复, 近20多年来, 中国政府实施了天然林保护、长江中上游防护林体系建设等系列林业生态工程, 该地区森林植被恢复迅速, 由于人为干扰强度不同, 形成了处于不同恢复阶段的多种次生林群落(
Xiang et al ., 2013 ).目前针对亚热带森林植被恢复过程中, 不同植物组织(
阎恩荣等, 2010 ;
Zeng et al ., 2017b )、土壤(
彭晓等, 2016 ;
Xu et al ., 2018 )的化学计量动态已有一些研究报道.然而, 将植物叶片-凋落物-土壤作为一个系统, 探讨其C、N、P化学计量动态及其各组分之间相互作用的研究鲜有报道, 植被恢复对整个生态系统养分运输和分配影响如何仍然不是很清楚.为此, 本研究根据亚热带森林群落演替进程, 在湘中丘陵区选取了4种处于不同恢复阶段(灌草丛、灌木林、针阔混交林、常绿阔叶林)的生态系统作为一个恢复序列, 研究植物叶片、凋落物、土壤C、N、P含量、化学计量比的变化及其相互之间的关联性, 探讨随植被恢复, 植物群落对土壤环境变化的响应及其养分利用策略, 剖析生态系统各组分之间养分动态平衡关系随着植被恢复的变化特征, 为亚热带森林植被恢复和养分管理提供科学依据. ...
Changes in microbial communities and respiration following the revegetation of eroded soil
1
2017
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
Effects of forest restoration on soil carbon, nitrogen, phosphorus, and their stoichiometry in Hunan, Southern China
3
2018
... 中国亚热带地区水热条件优越, 森林资源丰富, 是中国重要的生态屏障.但由于长期的人为干扰, 该地区地带性植被——常绿阔叶林遭到了严重的破坏.为保护生物多样性和提升森林生态系统服务, 促进森林植被恢复, 近20多年来, 中国政府实施了天然林保护、长江中上游防护林体系建设等系列林业生态工程, 该地区森林植被恢复迅速, 由于人为干扰强度不同, 形成了处于不同恢复阶段的多种次生林群落(
Xiang et al ., 2013 ).目前针对亚热带森林植被恢复过程中, 不同植物组织(
阎恩荣等, 2010 ;
Zeng et al ., 2017b )、土壤(
彭晓等, 2016 ;
Xu et al ., 2018 )的化学计量动态已有一些研究报道.然而, 将植物叶片-凋落物-土壤作为一个系统, 探讨其C、N、P化学计量动态及其各组分之间相互作用的研究鲜有报道, 植被恢复对整个生态系统养分运输和分配影响如何仍然不是很清楚.为此, 本研究根据亚热带森林群落演替进程, 在湘中丘陵区选取了4种处于不同恢复阶段(灌草丛、灌木林、针阔混交林、常绿阔叶林)的生态系统作为一个恢复序列, 研究植物叶片、凋落物、土壤C、N、P含量、化学计量比的变化及其相互之间的关联性, 探讨随植被恢复, 植物群落对土壤环境变化的响应及其养分利用策略, 剖析生态系统各组分之间养分动态平衡关系随着植被恢复的变化特征, 为亚热带森林植被恢复和养分管理提供科学依据. ...
... 因植被恢复而形成不同功能性状的树种组成和结构(
Xu et al ., 2018 ), 导致植物养分吸收、凋落物数量和质量及其分解的差异, 进而影响土壤环境(
Yang & Luo, 2011 ).研究表明, 草本植物C含量低于灌木、乔木, 即灌木、乔木可以合成和积累更多的有机物, 形成更高的C含量(
He et al ., 2013 ).本研究中, 随着植被恢复, 叶片C含量增高, 与
He等(2013) 的研究结果一致, 与群落组成和结构随着植被恢复的变化有关.此外, 本研究中, 叶片对N、P的利用效率随植被恢复显著下降(
图4A ), 即N、P吸收量和存留量增高, 光合作用增强, 有利于干物质积累, 叶片C得到有效的积累.研究发现, 常绿乔木叶片寿命较长, 需要积累更多有机物质(如木质素等)构建保卫结构(
秦海等, 2010 ), 植物体保持较高的N、P含量, 有利于维持植被的生长代谢(
Zeng et al ., 2016 ), 乔木的生长相较于草本需要更多的N、P用于维持代谢所需.本研究中, 随着植被恢复, 叶片N含量显著增高.可能是由于随着植被恢复, 生物量增加促使植物需要更多富含N的物质(如酶、运输蛋白质和氨基酸)参与代谢活动, 加大叶片对N的吸收, 使得叶片N含量增加(
Qin et al ., 2016 ).而土壤N含量随植被恢复而增高, 能更大程度满足植物生长的需求, 也进一步解释了植物叶片N含量的变化(
Wang et al ., 2015 ).由于亚热带丘陵红壤区土壤严重缺P, 不同植被类型对P的再吸收利用效率不同.本研究中, PLL对P的再吸收利用效率最高(
图4B ), 其次是LCC, 这可能是PLL叶片P含量高于LCC, 而LCC高于LCQ、LVR的主要原因.随植被恢复, 植物叶片C、N、P含量呈递增趋势, 表明植被从灌草群落发展为灌木林、乔木林群落, 秉承了从资源保守策略转变为资源快速获取策略的特性(
Royer et al ., 2010 ), 更好地适应环境的变化. ...
... 森林恢复是决定土壤C、N、P含量变化的关键因素(
Xu et al ., 2018 ).有关本研究样地土壤C含量随着植被恢复而增加的机理已另有论文(
辜翔等, 2018 )详细讨论, 在这里不再重复.土壤N含量随植被恢复逐渐增加, 可能是受到凋落物养分释放的影响, 随着植被恢复, 凋落物C:N下降有利于N养分释放, 土壤N含量增加.而土壤P含量在恢复早期(LVR-LCQ)下降, 恢复后期(PLL-LCC)显著增加.经相关性分析表明, 土壤N、P含量与土壤C含量均呈显著的正相关关系(相关系数分别为0.917, 0.649,
p < 0.01,
n = 48).表明植被恢复过程中, 土壤有机质的积累有利于土壤养分的提高(
Zhang et al ., 2019 ).此外, 可能是由于恢复早期植物对P的高需求和凋落物的低输入所致, LVR、LCQ植物对P的再吸收效率较高(
图4B ), 凋落物P归还低, 导致土壤P含量下降, 而恢复后期植被多样性和生物量增加(
辜翔等, 2018 ), 随凋落物P归还量增高, 土壤P含量有所提高.由于针叶树种叶片和凋落物酸度较高, 导致针叶林土壤P含量显著低于阔叶林(
Cao et al ., 2018 ), 因而PLL土壤P含量低于LCC.表明植被恢复形成不同的植物群落是影响土壤P含量的主要因素. ...
Stocks and stoichiometry of soil organic carbon, total nitrogen, and total phosphorus after vegetation restoration in the Loess Hilly Region, China
2
2019
... 植被恢复涉及植物群落与土壤环境的协调发展, 因其能改善脆弱或退化生态系统养分循环、土壤质量而受到广泛关注(
Xu et al ., 2019 ).随着植被恢复, 植物群落组成结构、土壤理化性质达到稳定状态(
O’Brien & Jastrow, 2013 ), 生态系统各组分(植物、凋落物、土壤)碳(C)、氮(N)、磷(P)含量及其循环规律调控着植物的生存发展和各种生态过程(
Reich et al ., 2006 ), 其化学计量比反映植物组成动态与土壤养分之间的平衡(
Zhang et al ., 2018 ).因此, 研究生态系统各组分C、N、P化学计量比随植被恢复的变化对理解各组分之间养分关系及分配特征十分必要, 有助于明确植被恢复过程中, 植物群落随土壤环境变化的发展方向(
Zhao et al ., 2015b ;
Ren et al ., 2016 ). ...
... 土壤C:N、C:P、N:P可作为土壤养分状况的良好指标(
Zhao et al ., 2015a ), 但因植物群落不同而异, 具有高度复杂性(
Zhang et al ., 2014 ).C、N、P不平衡性增加也会引起土壤C:N、C:P、N:P的变化(
Zhao et al ., 2015b ).本研究中, 土壤C:N (13-19)随着植被恢复先升高后下降, 与
Zhang等(2018) 的研究结果一致, 是由于从LVR到PLL土壤C含量增加幅度(121.66%)高于土壤N含量的增加幅度(79.75%), 导致土壤C:N升高; 而从PLL到LCC, 土壤C含量增加幅度(40.84%)低于土壤N含量的增加幅度(91.65%), 导致土壤C:N下降.这可以从植物对N的利用效率和再吸收效率来解释, 随着植被恢复, 植物对N的利用效率下降, 而恢复后期N的再吸收效率下降, 表明植物对土壤N的吸收量下降, 凋落物N含量较高, 有利于土壤N的积累.土壤C、N含量的变化是影响土壤C:P、N:P的主要因素(
Xu et al ., 2019 ).本研究中, 由于土壤P主要来源于母质的风化, 随植被恢复变化较小; 另一方面, 如前面分析, N的再吸收效率低于P, 凋落物的归还作用主要增加了土壤C、N的投入, 促进了土壤C、N的积累, 因而使得土壤C:P、N:P增加.在中国南方亚热带红壤丘陵区土壤较高的C:P、N:P也可以反映出林地土壤P的限制性(
Gao et al ., 2014 ;
Ren et al ., 2016 ). ...
浙江天童常绿阔叶林、常绿针叶林与落叶阔叶林的C:N:P化学计量特征
1
2010
... 中国亚热带地区水热条件优越, 森林资源丰富, 是中国重要的生态屏障.但由于长期的人为干扰, 该地区地带性植被——常绿阔叶林遭到了严重的破坏.为保护生物多样性和提升森林生态系统服务, 促进森林植被恢复, 近20多年来, 中国政府实施了天然林保护、长江中上游防护林体系建设等系列林业生态工程, 该地区森林植被恢复迅速, 由于人为干扰强度不同, 形成了处于不同恢复阶段的多种次生林群落(
Xiang et al ., 2013 ).目前针对亚热带森林植被恢复过程中, 不同植物组织(
阎恩荣等, 2010 ;
Zeng et al ., 2017b )、土壤(
彭晓等, 2016 ;
Xu et al ., 2018 )的化学计量动态已有一些研究报道.然而, 将植物叶片-凋落物-土壤作为一个系统, 探讨其C、N、P化学计量动态及其各组分之间相互作用的研究鲜有报道, 植被恢复对整个生态系统养分运输和分配影响如何仍然不是很清楚.为此, 本研究根据亚热带森林群落演替进程, 在湘中丘陵区选取了4种处于不同恢复阶段(灌草丛、灌木林、针阔混交林、常绿阔叶林)的生态系统作为一个恢复序列, 研究植物叶片、凋落物、土壤C、N、P含量、化学计量比的变化及其相互之间的关联性, 探讨随植被恢复, 植物群落对土壤环境变化的响应及其养分利用策略, 剖析生态系统各组分之间养分动态平衡关系随着植被恢复的变化特征, 为亚热带森林植被恢复和养分管理提供科学依据. ...
浙江天童常绿阔叶林、常绿针叶林与落叶阔叶林的C:N:P化学计量特征
1
2010
... 中国亚热带地区水热条件优越, 森林资源丰富, 是中国重要的生态屏障.但由于长期的人为干扰, 该地区地带性植被——常绿阔叶林遭到了严重的破坏.为保护生物多样性和提升森林生态系统服务, 促进森林植被恢复, 近20多年来, 中国政府实施了天然林保护、长江中上游防护林体系建设等系列林业生态工程, 该地区森林植被恢复迅速, 由于人为干扰强度不同, 形成了处于不同恢复阶段的多种次生林群落(
Xiang et al ., 2013 ).目前针对亚热带森林植被恢复过程中, 不同植物组织(
阎恩荣等, 2010 ;
Zeng et al ., 2017b )、土壤(
彭晓等, 2016 ;
Xu et al ., 2018 )的化学计量动态已有一些研究报道.然而, 将植物叶片-凋落物-土壤作为一个系统, 探讨其C、N、P化学计量动态及其各组分之间相互作用的研究鲜有报道, 植被恢复对整个生态系统养分运输和分配影响如何仍然不是很清楚.为此, 本研究根据亚热带森林群落演替进程, 在湘中丘陵区选取了4种处于不同恢复阶段(灌草丛、灌木林、针阔混交林、常绿阔叶林)的生态系统作为一个恢复序列, 研究植物叶片、凋落物、土壤C、N、P含量、化学计量比的变化及其相互之间的关联性, 探讨随植被恢复, 植物群落对土壤环境变化的响应及其养分利用策略, 剖析生态系统各组分之间养分动态平衡关系随着植被恢复的变化特征, 为亚热带森林植被恢复和养分管理提供科学依据. ...
Ecological stoichiometry in leaves, roots, litters and soil among different plant communities in a desertified region of Northern China
2
2018
... 凋落物养分含量明显受到群落优势植物及其凋落物组成的影响.研究发现, 林地凋落物C含量高于草地(
Paul et al ., 2002 ), 针叶林凋落物C含量高于阔叶林(
He et al ., 2013 ).本研究中, 随着植被恢复, 凋落物C含量显著提高, PLL最大, 与
Zeng等(2017a) 的研究结果一致, 是由于PLL以马尾松为优势树种, 凋落物主要以马尾松针叶为主, 而LVR以草本植物和小灌木为主, LCQ以落叶灌木为主, LCC以常绿阔叶树种为主所致.本研究中, 随着植被恢复, 凋落物N、P含量波动较大, 可能是由于不同恢复阶段植物对N、P的再吸收利用效率(
图4B )以及凋落物组成不同所致, 其中, PLL对N、P的再吸收利用效率最高, 其次LCC, 因而PLL凋落物N、P含量低于LCC.此外, 当凋落物N、P含量分别低于7、0.5 g·kg
-1 时, 表明凋落物N、P养分被叶片完全吸收; 而N、P含量分别高于10、0.8 g·kg
-1 时, 则表明没有完全吸收(
Killingbeck, 1996 ;
Yang et al ., 2018 ).本研究中, 凋落物N含量(12-18 g·kg
-1 )高于7 g·kg
-1 , P含量(0.31-0.49 g·kg
-1 )低于0.5 g·kg
-1 , 表明不同植被恢复阶段凋落物N均未被叶片完全吸收, 而P被完全吸收.可能是由于研究区林地土壤N较为丰富, 而P较为缺乏, 植物通过提高对P的再吸收利用以适应土壤P匮乏, 导致叶片对N的再吸收效率显著低于P再吸收效率. ...
... 凋落物是养分从植物到土壤的基本载体, 其动态交换可以实现并维持土壤养分和植物生长所需的元素比率之间的平衡(
Hessen et al ., 2004 ).凋落物养分含量及其化学计量比与土壤养分含量和化学计量比显著相关(
Yang et al ., 2018 ), 一方面, 表明凋落物分解促使有机物质向土壤释放N、P, 从而影响土壤C、N和P含量及其化学计量比(
Zhong et al ., 2013 ), 另一方面, 表明土壤养分含量的变化会影响植物对养分的利用策略, 进而影响凋落物养分含量及其化学计量比.本研究中, 土壤C、N含量与凋落物C、N含量显著正相关, 可能是随着植被恢复, 土壤C、N含量增加, 更能满足植物对养分的需求, 从而影响植物对养分的再吸收效率, 进而影响凋落物的养分含量.因此, 叶片N、P含量显著影响着凋落物养分含量.此外, 本研究中, 土壤C:N、C:P和N:P与凋落物C:N、C:P显著正相关, 与
汪宗飞和郑粉莉(2018) 、
张萍等(2018) 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 ).也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加. ...
Carbon:nitrogen stoichiometry in forest ecosystems during stand development
2
2011
... 因植被恢复而形成不同功能性状的树种组成和结构(
Xu et al ., 2018 ), 导致植物养分吸收、凋落物数量和质量及其分解的差异, 进而影响土壤环境(
Yang & Luo, 2011 ).研究表明, 草本植物C含量低于灌木、乔木, 即灌木、乔木可以合成和积累更多的有机物, 形成更高的C含量(
He et al ., 2013 ).本研究中, 随着植被恢复, 叶片C含量增高, 与
He等(2013) 的研究结果一致, 与群落组成和结构随着植被恢复的变化有关.此外, 本研究中, 叶片对N、P的利用效率随植被恢复显著下降(
图4A ), 即N、P吸收量和存留量增高, 光合作用增强, 有利于干物质积累, 叶片C得到有效的积累.研究发现, 常绿乔木叶片寿命较长, 需要积累更多有机物质(如木质素等)构建保卫结构(
秦海等, 2010 ), 植物体保持较高的N、P含量, 有利于维持植被的生长代谢(
Zeng et al ., 2016 ), 乔木的生长相较于草本需要更多的N、P用于维持代谢所需.本研究中, 随着植被恢复, 叶片N含量显著增高.可能是由于随着植被恢复, 生物量增加促使植物需要更多富含N的物质(如酶、运输蛋白质和氨基酸)参与代谢活动, 加大叶片对N的吸收, 使得叶片N含量增加(
Qin et al ., 2016 ).而土壤N含量随植被恢复而增高, 能更大程度满足植物生长的需求, 也进一步解释了植物叶片N含量的变化(
Wang et al ., 2015 ).由于亚热带丘陵红壤区土壤严重缺P, 不同植被类型对P的再吸收利用效率不同.本研究中, PLL对P的再吸收利用效率最高(
图4B ), 其次是LCC, 这可能是PLL叶片P含量高于LCC, 而LCC高于LCQ、LVR的主要原因.随植被恢复, 植物叶片C、N、P含量呈递增趋势, 表明植被从灌草群落发展为灌木林、乔木林群落, 秉承了从资源保守策略转变为资源快速获取策略的特性(
Royer et al ., 2010 ), 更好地适应环境的变化. ...
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability
3
2010
... 研究表明, 土壤养分含量的变化直接影响植物养分吸收及其化学计量比(
Chen et al ., 2018 ).而植物通过改变化学计量比和采用适宜的养分利用策略适应土壤N、P供应的变化(
Sun et al ., 2018 ).植物N含量比P含量更稳定(
Yu et al ., 2010 ).本研究中, 随着植被恢复, 叶片N含量内稳态较弱, 受土壤N、P含量的变化影响较大, 叶片P含量具有较高的内稳态, 不随土壤N、P含量的变化而变化, 与
Yu等(2010) 的研究结果不同.可能是由于植物的限制性养分元素不同所致, 在
Yu等(2010) 的研究中, 植物主要受N限制, 而本研究中, 土壤P含量随植被恢复的变化小, 植物主要受P的限制, 在一定程度上证实了限制性养分元素稳定性假说“由于生理和养分平衡的制约, 限制性养分元素在植物体内的含量具有相对稳定性, 对环境变化的响应也较为稳定” (
Sterner & Elser, 2002 ).表明随着植被恢复, 在P缺乏的土壤环境中, 植物叶片P具有较强的内稳态. ...
... ).本研究中, 随着植被恢复, 叶片N含量内稳态较弱, 受土壤N、P含量的变化影响较大, 叶片P含量具有较高的内稳态, 不随土壤N、P含量的变化而变化, 与
Yu等(2010) 的研究结果不同.可能是由于植物的限制性养分元素不同所致, 在
Yu等(2010) 的研究中, 植物主要受N限制, 而本研究中, 土壤P含量随植被恢复的变化小, 植物主要受P的限制, 在一定程度上证实了限制性养分元素稳定性假说“由于生理和养分平衡的制约, 限制性养分元素在植物体内的含量具有相对稳定性, 对环境变化的响应也较为稳定” (
Sterner & Elser, 2002 ).表明随着植被恢复, 在P缺乏的土壤环境中, 植物叶片P具有较强的内稳态. ...
... 的研究结果不同.可能是由于植物的限制性养分元素不同所致, 在
Yu等(2010) 的研究中, 植物主要受N限制, 而本研究中, 土壤P含量随植被恢复的变化小, 植物主要受P的限制, 在一定程度上证实了限制性养分元素稳定性假说“由于生理和养分平衡的制约, 限制性养分元素在植物体内的含量具有相对稳定性, 对环境变化的响应也较为稳定” (
Sterner & Elser, 2002 ).表明随着植被恢复, 在P缺乏的土壤环境中, 植物叶片P具有较强的内稳态. ...
Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland
1
2011
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
Soil and plant components ecological stoichiometry in four steppe communities in the Loess Plateau of China
1
2016
... 因植被恢复而形成不同功能性状的树种组成和结构(
Xu et al ., 2018 ), 导致植物养分吸收、凋落物数量和质量及其分解的差异, 进而影响土壤环境(
Yang & Luo, 2011 ).研究表明, 草本植物C含量低于灌木、乔木, 即灌木、乔木可以合成和积累更多的有机物, 形成更高的C含量(
He et al ., 2013 ).本研究中, 随着植被恢复, 叶片C含量增高, 与
He等(2013) 的研究结果一致, 与群落组成和结构随着植被恢复的变化有关.此外, 本研究中, 叶片对N、P的利用效率随植被恢复显著下降(
图4A ), 即N、P吸收量和存留量增高, 光合作用增强, 有利于干物质积累, 叶片C得到有效的积累.研究发现, 常绿乔木叶片寿命较长, 需要积累更多有机物质(如木质素等)构建保卫结构(
秦海等, 2010 ), 植物体保持较高的N、P含量, 有利于维持植被的生长代谢(
Zeng et al ., 2016 ), 乔木的生长相较于草本需要更多的N、P用于维持代谢所需.本研究中, 随着植被恢复, 叶片N含量显著增高.可能是由于随着植被恢复, 生物量增加促使植物需要更多富含N的物质(如酶、运输蛋白质和氨基酸)参与代谢活动, 加大叶片对N的吸收, 使得叶片N含量增加(
Qin et al ., 2016 ).而土壤N含量随植被恢复而增高, 能更大程度满足植物生长的需求, 也进一步解释了植物叶片N含量的变化(
Wang et al ., 2015 ).由于亚热带丘陵红壤区土壤严重缺P, 不同植被类型对P的再吸收利用效率不同.本研究中, PLL对P的再吸收利用效率最高(
图4B ), 其次是LCC, 这可能是PLL叶片P含量高于LCC, 而LCC高于LCQ、LVR的主要原因.随植被恢复, 植物叶片C、N、P含量呈递增趋势, 表明植被从灌草群落发展为灌木林、乔木林群落, 秉承了从资源保守策略转变为资源快速获取策略的特性(
Royer et al ., 2010 ), 更好地适应环境的变化. ...
Impact of vegetation restoration on plants and soil C:N:P stoichiometry on the Yunwu Mountain Reserve of China
6
2017a
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
... ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
... ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
... 凋落物养分含量明显受到群落优势植物及其凋落物组成的影响.研究发现, 林地凋落物C含量高于草地(
Paul et al ., 2002 ), 针叶林凋落物C含量高于阔叶林(
He et al ., 2013 ).本研究中, 随着植被恢复, 凋落物C含量显著提高, PLL最大, 与
Zeng等(2017a) 的研究结果一致, 是由于PLL以马尾松为优势树种, 凋落物主要以马尾松针叶为主, 而LVR以草本植物和小灌木为主, LCQ以落叶灌木为主, LCC以常绿阔叶树种为主所致.本研究中, 随着植被恢复, 凋落物N、P含量波动较大, 可能是由于不同恢复阶段植物对N、P的再吸收利用效率(
图4B )以及凋落物组成不同所致, 其中, PLL对N、P的再吸收利用效率最高, 其次LCC, 因而PLL凋落物N、P含量低于LCC.此外, 当凋落物N、P含量分别低于7、0.5 g·kg
-1 时, 表明凋落物N、P养分被叶片完全吸收; 而N、P含量分别高于10、0.8 g·kg
-1 时, 则表明没有完全吸收(
Killingbeck, 1996 ;
Yang et al ., 2018 ).本研究中, 凋落物N含量(12-18 g·kg
-1 )高于7 g·kg
-1 , P含量(0.31-0.49 g·kg
-1 )低于0.5 g·kg
-1 , 表明不同植被恢复阶段凋落物N均未被叶片完全吸收, 而P被完全吸收.可能是由于研究区林地土壤N较为丰富, 而P较为缺乏, 植物通过提高对P的再吸收利用以适应土壤P匮乏, 导致叶片对N的再吸收效率显著低于P再吸收效率. ...
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
... 研究表明, 植物按照某种比例吸收和利用各种养分, 并在体内保持彼此相对平衡以适应土壤环境的变化(
秦海等, 2010 ).植物通过调整自身的适应性策略, 以高的养分吸收适应生境的变化(
宁秋蕊等, 2016 ).凋落物养分含量是平衡生态系统养分循环的关键环节, 养分再吸收对养分利用策略、植物生长和植物个体水平的竞争能力具有重要意义(
Wang & Moore, 2014 ).因此, 植物的异速生长关系、养分利用效率和养分再吸收效率可以反映植物对环境变化的适应策略.本研究发现, 植物通过调节叶片对N的吸收适应土壤随植被恢复的变化.LVR叶片N、P养分的投入比例为3.81:1, 是由于LVR以草本植物为主, 而生长迅速的草本植物叶片化学计量比具有更大的灵活性(
Güsewell, 2004 ), 通过增加N的投入适应P缺乏的土壤环境.LVR叶片N的再吸收效率最小且为负值, 是由于草本植物生长过程中需要大量的N, 但因其生长期短, 土壤N能满足其生长, 不需要从凋落物再吸收N.PLL叶片N、P的投入降低为0.75:1, N、P的再吸收效率最高, 是由于PLL凋落物分解较慢, N、P释放效率低(
刘颖等, 2009 ).PLL叶片P养分利用效率最低, 表明植物对P的吸收量和存留量高, 即植物通过提高养分吸收量以适应土壤P的缺乏, 从凋落物中再吸收N、P以满足对养分的需求.LCQ、LCC的N、P异速生长关系不显著, 表明植物生长过程中不存在N、P按比例消耗, 可能是由于灌木、常绿阔叶乔木具有快速的资源获取策略, 随土壤的变化调整养分消耗比例, 能更容易适应变化的环境(
Royer et al ., 2010 ).常绿阔叶乔木主要通过维持叶片的寿命和降低养分含量以减少养分的损失, 不是通过高的利用效率和再吸收率(
苏波等, 2000 ), 因此LCC的N、P利用效率较低, N再吸收效率趋近于0, P再吸收效率下降.研究发现, 在低P胁迫下, P的利用效率和再吸收效率高于N (
Zeng et al ., 2017a ), 本研究也得到类似的结果, 表明在P限制条件下, 植物通过加强对土壤P的吸收或再吸收, 形成自我调节机制(
Hu et al ., 2018 ).本研究中, 植被恢复过程中按1.45:1的N、P比例投入叶片生长, 此外随植被恢复N、P的养分利用效率显著下降, 而再吸收效率增高, 表明随着植物恢复, 植物叶片、凋落物、土壤之间的N、P循环逐渐达到“化学计量平衡”, 反映生态系统养分循环过程的协调性. ...
Stoichiometric and nutrient resorption characteristics of dominant tree species in subtropical Chinese forests
2
2017b
... 中国亚热带地区水热条件优越, 森林资源丰富, 是中国重要的生态屏障.但由于长期的人为干扰, 该地区地带性植被——常绿阔叶林遭到了严重的破坏.为保护生物多样性和提升森林生态系统服务, 促进森林植被恢复, 近20多年来, 中国政府实施了天然林保护、长江中上游防护林体系建设等系列林业生态工程, 该地区森林植被恢复迅速, 由于人为干扰强度不同, 形成了处于不同恢复阶段的多种次生林群落(
Xiang et al ., 2013 ).目前针对亚热带森林植被恢复过程中, 不同植物组织(
阎恩荣等, 2010 ;
Zeng et al ., 2017b )、土壤(
彭晓等, 2016 ;
Xu et al ., 2018 )的化学计量动态已有一些研究报道.然而, 将植物叶片-凋落物-土壤作为一个系统, 探讨其C、N、P化学计量动态及其各组分之间相互作用的研究鲜有报道, 植被恢复对整个生态系统养分运输和分配影响如何仍然不是很清楚.为此, 本研究根据亚热带森林群落演替进程, 在湘中丘陵区选取了4种处于不同恢复阶段(灌草丛、灌木林、针阔混交林、常绿阔叶林)的生态系统作为一个恢复序列, 研究植物叶片、凋落物、土壤C、N、P含量、化学计量比的变化及其相互之间的关联性, 探讨随植被恢复, 植物群落对土壤环境变化的响应及其养分利用策略, 剖析生态系统各组分之间养分动态平衡关系随着植被恢复的变化特征, 为亚热带森林植被恢复和养分管理提供科学依据. ...
... 本研究中, PLL凋落物C:N、C:P显著高于其他3个恢复阶段, 且随植被恢复的变化与C含量的变化一致, 表明是由于PLL凋落物以马尾松针叶为主, C含量较高所致.研究发现, 随着植被恢复, 凋落物N:P增加(
Zeng et al ., 2017b ).而本研究中, 随着植被恢复, 凋落物N:P有较大的波动, 其中PLL最高, 可能与凋落物P含量有关.随着植被恢复, 凋落物N:P的变化与P含量(
图1 )、P的再吸收效率(
图4B )的变化一致, 由于PLL对P的再吸收利用最高(
图4B ), 凋落物P含量下降, 导致凋落物N:P增高.研究表明, 当凋落物N:P > 25时, 其分解受到P限制(
Güsewell & Verhoeven, 2006 ).本研究中, 凋落物N:P (30-45)明显高于25, 表明本研究区凋落物分解受到P限制. ...
Plant and soil seed bank diversity across a range of ages of
Eucalyptus grandis plantations afforested on arable lands
3
2014
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
... )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
... 土壤C:N、C:P、N:P可作为土壤养分状况的良好指标(
Zhao et al ., 2015a ), 但因植物群落不同而异, 具有高度复杂性(
Zhang et al ., 2014 ).C、N、P不平衡性增加也会引起土壤C:N、C:P、N:P的变化(
Zhao et al ., 2015b ).本研究中, 土壤C:N (13-19)随着植被恢复先升高后下降, 与
Zhang等(2018) 的研究结果一致, 是由于从LVR到PLL土壤C含量增加幅度(121.66%)高于土壤N含量的增加幅度(79.75%), 导致土壤C:N升高; 而从PLL到LCC, 土壤C含量增加幅度(40.84%)低于土壤N含量的增加幅度(91.65%), 导致土壤C:N下降.这可以从植物对N的利用效率和再吸收效率来解释, 随着植被恢复, 植物对N的利用效率下降, 而恢复后期N的再吸收效率下降, 表明植物对土壤N的吸收量下降, 凋落物N含量较高, 有利于土壤N的积累.土壤C、N含量的变化是影响土壤C:P、N:P的主要因素(
Xu et al ., 2019 ).本研究中, 由于土壤P主要来源于母质的风化, 随植被恢复变化较小; 另一方面, 如前面分析, N的再吸收效率低于P, 凋落物的归还作用主要增加了土壤C、N的投入, 促进了土壤C、N的积累, 因而使得土壤C:P、N:P增加.在中国南方亚热带红壤丘陵区土壤较高的C:P、N:P也可以反映出林地土壤P的限制性(
Gao et al ., 2014 ;
Ren et al ., 2016 ). ...
The coupling of leaf, litter, and soil nutrients in warm temperate forests in northwestern China
1
2017
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
黄土丘陵区不同森林类型叶片-凋落物-土壤生态化学计量特征
3
2018
... 土壤C:N、C:P、N:P可作为土壤养分状况的良好指标(
Zhao et al ., 2015a ), 但因植物群落不同而异, 具有高度复杂性(
Zhang et al ., 2014 ).C、N、P不平衡性增加也会引起土壤C:N、C:P、N:P的变化(
Zhao et al ., 2015b ).本研究中, 土壤C:N (13-19)随着植被恢复先升高后下降, 与
Zhang等(2018) 的研究结果一致, 是由于从LVR到PLL土壤C含量增加幅度(121.66%)高于土壤N含量的增加幅度(79.75%), 导致土壤C:N升高; 而从PLL到LCC, 土壤C含量增加幅度(40.84%)低于土壤N含量的增加幅度(91.65%), 导致土壤C:N下降.这可以从植物对N的利用效率和再吸收效率来解释, 随着植被恢复, 植物对N的利用效率下降, 而恢复后期N的再吸收效率下降, 表明植物对土壤N的吸收量下降, 凋落物N含量较高, 有利于土壤N的积累.土壤C、N含量的变化是影响土壤C:P、N:P的主要因素(
Xu et al ., 2019 ).本研究中, 由于土壤P主要来源于母质的风化, 随植被恢复变化较小; 另一方面, 如前面分析, N的再吸收效率低于P, 凋落物的归还作用主要增加了土壤C、N的投入, 促进了土壤C、N的积累, 因而使得土壤C:P、N:P增加.在中国南方亚热带红壤丘陵区土壤较高的C:P、N:P也可以反映出林地土壤P的限制性(
Gao et al ., 2014 ;
Ren et al ., 2016 ). ...
... 凋落物是养分从植物到土壤的基本载体, 其动态交换可以实现并维持土壤养分和植物生长所需的元素比率之间的平衡(
Hessen et al ., 2004 ).凋落物养分含量及其化学计量比与土壤养分含量和化学计量比显著相关(
Yang et al ., 2018 ), 一方面, 表明凋落物分解促使有机物质向土壤释放N、P, 从而影响土壤C、N和P含量及其化学计量比(
Zhong et al ., 2013 ), 另一方面, 表明土壤养分含量的变化会影响植物对养分的利用策略, 进而影响凋落物养分含量及其化学计量比.本研究中, 土壤C、N含量与凋落物C、N含量显著正相关, 可能是随着植被恢复, 土壤C、N含量增加, 更能满足植物对养分的需求, 从而影响植物对养分的再吸收效率, 进而影响凋落物的养分含量.因此, 叶片N、P含量显著影响着凋落物养分含量.此外, 本研究中, 土壤C:N、C:P和N:P与凋落物C:N、C:P显著正相关, 与
汪宗飞和郑粉莉(2018) 、
张萍等(2018) 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 ).也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加. ...
... 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 ).也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加. ...
黄土丘陵区不同森林类型叶片-凋落物-土壤生态化学计量特征
3
2018
... 土壤C:N、C:P、N:P可作为土壤养分状况的良好指标(
Zhao et al ., 2015a ), 但因植物群落不同而异, 具有高度复杂性(
Zhang et al ., 2014 ).C、N、P不平衡性增加也会引起土壤C:N、C:P、N:P的变化(
Zhao et al ., 2015b ).本研究中, 土壤C:N (13-19)随着植被恢复先升高后下降, 与
Zhang等(2018) 的研究结果一致, 是由于从LVR到PLL土壤C含量增加幅度(121.66%)高于土壤N含量的增加幅度(79.75%), 导致土壤C:N升高; 而从PLL到LCC, 土壤C含量增加幅度(40.84%)低于土壤N含量的增加幅度(91.65%), 导致土壤C:N下降.这可以从植物对N的利用效率和再吸收效率来解释, 随着植被恢复, 植物对N的利用效率下降, 而恢复后期N的再吸收效率下降, 表明植物对土壤N的吸收量下降, 凋落物N含量较高, 有利于土壤N的积累.土壤C、N含量的变化是影响土壤C:P、N:P的主要因素(
Xu et al ., 2019 ).本研究中, 由于土壤P主要来源于母质的风化, 随植被恢复变化较小; 另一方面, 如前面分析, N的再吸收效率低于P, 凋落物的归还作用主要增加了土壤C、N的投入, 促进了土壤C、N的积累, 因而使得土壤C:P、N:P增加.在中国南方亚热带红壤丘陵区土壤较高的C:P、N:P也可以反映出林地土壤P的限制性(
Gao et al ., 2014 ;
Ren et al ., 2016 ). ...
... 凋落物是养分从植物到土壤的基本载体, 其动态交换可以实现并维持土壤养分和植物生长所需的元素比率之间的平衡(
Hessen et al ., 2004 ).凋落物养分含量及其化学计量比与土壤养分含量和化学计量比显著相关(
Yang et al ., 2018 ), 一方面, 表明凋落物分解促使有机物质向土壤释放N、P, 从而影响土壤C、N和P含量及其化学计量比(
Zhong et al ., 2013 ), 另一方面, 表明土壤养分含量的变化会影响植物对养分的利用策略, 进而影响凋落物养分含量及其化学计量比.本研究中, 土壤C、N含量与凋落物C、N含量显著正相关, 可能是随着植被恢复, 土壤C、N含量增加, 更能满足植物对养分的需求, 从而影响植物对养分的再吸收效率, 进而影响凋落物的养分含量.因此, 叶片N、P含量显著影响着凋落物养分含量.此外, 本研究中, 土壤C:N、C:P和N:P与凋落物C:N、C:P显著正相关, 与
汪宗飞和郑粉莉(2018) 、
张萍等(2018) 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 ).也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加. ...
... 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 ).也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加. ...
Response of forest growth to C:N:P stoichiometry in plants and soils during
Robinia pseudoacacia afforestation on the Loess Plateau, China
3
2019
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
... 森林恢复是决定土壤C、N、P含量变化的关键因素(
Xu et al ., 2018 ).有关本研究样地土壤C含量随着植被恢复而增加的机理已另有论文(
辜翔等, 2018 )详细讨论, 在这里不再重复.土壤N含量随植被恢复逐渐增加, 可能是受到凋落物养分释放的影响, 随着植被恢复, 凋落物C:N下降有利于N养分释放, 土壤N含量增加.而土壤P含量在恢复早期(LVR-LCQ)下降, 恢复后期(PLL-LCC)显著增加.经相关性分析表明, 土壤N、P含量与土壤C含量均呈显著的正相关关系(相关系数分别为0.917, 0.649,
p < 0.01,
n = 48).表明植被恢复过程中, 土壤有机质的积累有利于土壤养分的提高(
Zhang et al ., 2019 ).此外, 可能是由于恢复早期植物对P的高需求和凋落物的低输入所致, LVR、LCQ植物对P的再吸收效率较高(
图4B ), 凋落物P归还低, 导致土壤P含量下降, 而恢复后期植被多样性和生物量增加(
辜翔等, 2018 ), 随凋落物P归还量增高, 土壤P含量有所提高.由于针叶树种叶片和凋落物酸度较高, 导致针叶林土壤P含量显著低于阔叶林(
Cao et al ., 2018 ), 因而PLL土壤P含量低于LCC.表明植被恢复形成不同的植物群落是影响土壤P含量的主要因素. ...
... 限制性养分有效性、植物代谢速率和植物功能群共同影响叶片的化学计量模式, 因此, 不同区域植物叶片C、N、P化学计量比与环境的关系有所不同(
Zhao et al ., 2014 ).本研究中, 随着植被恢复, 土壤C含量增加, 叶片C含量增加, 与
Zhang等(2019) 的研究结果一致.同时, 土壤C含量与叶片N、P含量显著正相关, 表明随着植被恢复, 土壤有机质的积累和分解释放出更多可利用性N、P, 促进叶片对N、P的吸收.此外, 土壤N含量与叶片C、N含量显著正相关, 表明随植被恢复土壤N含量增加, 有利于植物的生长发育, 促进光合作用合成更多的有机物质.叶片N的变化决定着叶片C:N的变化.因此, 土壤C、N含量与叶片N的正相关性可以解释土壤C、N含量与叶片C:N的负相关性.本研究中, 土壤C:P、N:P与叶片P含量显著正相关, 与叶片C:P、N:P显著负相关, 可能是由于随着植被恢复, 土壤C:P、N:P增加, 植物受P的限制更为明显, 植物通过提高对P的吸收量(降低P的利用效率)和再吸收效率来维持正常生理活动, 这也是叶片P含量增加而C:P、N:P降低的原因之一. ...
Plant functional composition and species diversity affect soil C, N, and P during secondary succession of abandoned farmland on the Loess Plateau
1
2018
... 植被恢复涉及植物群落与土壤环境的协调发展, 因其能改善脆弱或退化生态系统养分循环、土壤质量而受到广泛关注(
Xu et al ., 2019 ).随着植被恢复, 植物群落组成结构、土壤理化性质达到稳定状态(
O’Brien & Jastrow, 2013 ), 生态系统各组分(植物、凋落物、土壤)碳(C)、氮(N)、磷(P)含量及其循环规律调控着植物的生存发展和各种生态过程(
Reich et al ., 2006 ), 其化学计量比反映植物组成动态与土壤养分之间的平衡(
Zhang et al ., 2018 ).因此, 研究生态系统各组分C、N、P化学计量比随植被恢复的变化对理解各组分之间养分关系及分配特征十分必要, 有助于明确植被恢复过程中, 植物群落随土壤环境变化的发展方向(
Zhao et al ., 2015b ;
Ren et al ., 2016 ). ...
Soil stoichiometry and carbon storage in long-term afforestation soil affected by understory vegetation diversity
2
2015a
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
... 土壤C:N、C:P、N:P可作为土壤养分状况的良好指标(
Zhao et al ., 2015a ), 但因植物群落不同而异, 具有高度复杂性(
Zhang et al ., 2014 ).C、N、P不平衡性增加也会引起土壤C:N、C:P、N:P的变化(
Zhao et al ., 2015b ).本研究中, 土壤C:N (13-19)随着植被恢复先升高后下降, 与
Zhang等(2018) 的研究结果一致, 是由于从LVR到PLL土壤C含量增加幅度(121.66%)高于土壤N含量的增加幅度(79.75%), 导致土壤C:N升高; 而从PLL到LCC, 土壤C含量增加幅度(40.84%)低于土壤N含量的增加幅度(91.65%), 导致土壤C:N下降.这可以从植物对N的利用效率和再吸收效率来解释, 随着植被恢复, 植物对N的利用效率下降, 而恢复后期N的再吸收效率下降, 表明植物对土壤N的吸收量下降, 凋落物N含量较高, 有利于土壤N的积累.土壤C、N含量的变化是影响土壤C:P、N:P的主要因素(
Xu et al ., 2019 ).本研究中, 由于土壤P主要来源于母质的风化, 随植被恢复变化较小; 另一方面, 如前面分析, N的再吸收效率低于P, 凋落物的归还作用主要增加了土壤C、N的投入, 促进了土壤C、N的积累, 因而使得土壤C:P、N:P增加.在中国南方亚热带红壤丘陵区土壤较高的C:P、N:P也可以反映出林地土壤P的限制性(
Gao et al ., 2014 ;
Ren et al ., 2016 ). ...
Land use change influences soil C, N and P stoichiometry under “Grain-to-Green Program” in China
4
2015b
... 植被恢复涉及植物群落与土壤环境的协调发展, 因其能改善脆弱或退化生态系统养分循环、土壤质量而受到广泛关注(
Xu et al ., 2019 ).随着植被恢复, 植物群落组成结构、土壤理化性质达到稳定状态(
O’Brien & Jastrow, 2013 ), 生态系统各组分(植物、凋落物、土壤)碳(C)、氮(N)、磷(P)含量及其循环规律调控着植物的生存发展和各种生态过程(
Reich et al ., 2006 ), 其化学计量比反映植物组成动态与土壤养分之间的平衡(
Zhang et al ., 2018 ).因此, 研究生态系统各组分C、N、P化学计量比随植被恢复的变化对理解各组分之间养分关系及分配特征十分必要, 有助于明确植被恢复过程中, 植物群落随土壤环境变化的发展方向(
Zhao et al ., 2015b ;
Ren et al ., 2016 ). ...
... 土壤是植物营养物质的主要来源, 其C、N、P含量的变化影响着植物养分吸收利用策略.由于地形(
Tian et al ., 2018 )、土地利用类型(
Li et al ., 2016 )和植被类型(
Zhang et al ., 2014 )的影响, 土壤C、N、P含量及其化学计量比在不同时空尺度上存在异质性, 特别是植被盖度、植物类型及其群落组成显著影响土壤C、N、P含量及其化学计量比(
Zhang et al ., 2014 ).随着植被恢复, 植被类型及其群落组成结构变化有利于生物量的积累, 增加土壤有机物质的输入, 促进土壤养分的积累(
Xiao et al ., 2017 ), 从而提高土壤C、N、P含量和改变其化学计量比(
Zhao et al ., 2015b ).由于木本植物有更多的凋落物归还土壤, 且凋落物C、N含量较高(
Paul et al ., 2002 ;
Zhao et al ., 2015a ), 因而林地土壤C、N含量及其C:P、N:P均高于草地(
Li et al ., 2016 ).基于生长速率假说, C:P、N:P低的植物生长迅速, 对土壤资源竞争能力高于C:P、N:P高的植物, 特别是在贫瘠的土壤环境中(
Zhang et al ., 2019 ).此外, 植物养分元素的内稳态可反映各养分元素的平衡状态, 植物对环境变化的响应策略(
Yu et al ., 2011 ).随着植被恢复, 土壤养分含量改变, 植物养分表现出内稳态, 即植物养分不随土壤环境变化而变化, 多数显著的内稳态关系主要体现在植物叶片N和土壤N之间(
蒋利玲等, 2014 ;
Zeng et al ., 2017a ).凋落物作为连接植物与土壤的“纽带”, 其养分特性与植物、土壤密切相关(
Cao & Chen, 2017 ).植物对落叶前N的再吸收效率随着正向演替而降低, 表明恢复早期在缺N土壤中, 植物通过提高N的再吸收效率以满足其对N的需求(
Zeng et al ., 2017a ), 从而影响凋落物养分含量及其化学计量比.同时, 凋落物化学计量比影响着凋落物分解和养分释放的速率, 进而影响土壤养分的有效性和C固定(
Mooshammer et al ., 2012 ;
Zhang et al ., 2017 ).然而近年来, 有关化学计量学的研究主要集中在单一器官或组分, 仅有少量研究关注生态系统水平(
He et al ., 2008 ), 在植被恢复演替方面, 当前的研究也主要集中在各组分(叶片、凋落物、土壤) C、N、P化学计量比的变化特征, 涉及随着植被恢复, 叶片-凋落物-土壤系统化学计量比之间协同作用变化的研究仍比较少见, 限制了人们对陆地生态系统养分元素地球化学循环的理解(
Zeng et al ., 2017a ). ...
... 叶片是光合作用的核心器官, 因而叶片的化学计量比能代表整株的状况(
Sardans et al ., 2016 ).叶片C:N、C:P表征植物同化C的能力、植物的生长速率和营养利用效率, 较低的C:N、C:P对应较高的生长速率(
刘万德等, 2010 ).植被类型是影响植物C:N的主要因素(
Schreeg et al ., 2014 ).研究发现, 叶片C:N随植被恢复有3种不同的变化: 下降(
Zhao et al ., 2015b ;
Cao & Chen, 2017 )、急剧增加(
Yang & Luo, 2011 )、没有变化(
Clinton et al ., 2002 ).本研究中, 叶片C:N、C:P总体上随植被恢复而下降.可能是由于植物C同化和营养元素吸收的差异, 通常认为C不是植物生长的限制因子, 植物C:N、C:P主要受N、P含量的影响(
Hedin, 2004 ), 叶片N、P含量随植被恢复显著增加, 使得叶片C:N、C:P下降, 而LCC叶片P含量略有所降低, 因而其C:P增高, 表明植物的生长速率随着植被恢复而增加.本研究中, 叶片N:P随着植被恢复先下降再增加, 与
Zeng等(2017a) 的研究一致, 可能是随着植被恢复, 叶片N、P含量增加, 但从LVR到PLL, 叶片P含量增加幅度高于N, 而到LCC, 叶片P含量下降所致.研究表明, 当叶片N:P > 16, 植物生长受P限制; 当N:P < 14, 受N限制; 当14 < N:P < 16, 受N、P共同限制(
Koerselman & Meuleman, 1996 ).但也有研究发现, 养分限制中N没有最大值(
Güsewell & Koerselman, 2002 ), 当P < 1 g·kg
-1 时只有P限制作用(
Hector & Bagchi, 2007 ).本研究中, 叶片N:P (20-28)远远高于全球水平(13.8和12.7)(
Elser et al ., 2000 ), 是由于本研究林地土壤N含量相对高, 而P含量相对低, 也表明本研究区植物生长主要受到P的限制. ...
... 土壤C:N、C:P、N:P可作为土壤养分状况的良好指标(
Zhao et al ., 2015a ), 但因植物群落不同而异, 具有高度复杂性(
Zhang et al ., 2014 ).C、N、P不平衡性增加也会引起土壤C:N、C:P、N:P的变化(
Zhao et al ., 2015b ).本研究中, 土壤C:N (13-19)随着植被恢复先升高后下降, 与
Zhang等(2018) 的研究结果一致, 是由于从LVR到PLL土壤C含量增加幅度(121.66%)高于土壤N含量的增加幅度(79.75%), 导致土壤C:N升高; 而从PLL到LCC, 土壤C含量增加幅度(40.84%)低于土壤N含量的增加幅度(91.65%), 导致土壤C:N下降.这可以从植物对N的利用效率和再吸收效率来解释, 随着植被恢复, 植物对N的利用效率下降, 而恢复后期N的再吸收效率下降, 表明植物对土壤N的吸收量下降, 凋落物N含量较高, 有利于土壤N的积累.土壤C、N含量的变化是影响土壤C:P、N:P的主要因素(
Xu et al ., 2019 ).本研究中, 由于土壤P主要来源于母质的风化, 随植被恢复变化较小; 另一方面, 如前面分析, N的再吸收效率低于P, 凋落物的归还作用主要增加了土壤C、N的投入, 促进了土壤C、N的积累, 因而使得土壤C:P、N:P增加.在中国南方亚热带红壤丘陵区土壤较高的C:P、N:P也可以反映出林地土壤P的限制性(
Gao et al ., 2014 ;
Ren et al ., 2016 ). ...
The altitudinal patterns of leaf C:N:P stoichiometry are regulated by plant growth form, climate and soil on Changbai Mountain, China
1
2014
... 限制性养分有效性、植物代谢速率和植物功能群共同影响叶片的化学计量模式, 因此, 不同区域植物叶片C、N、P化学计量比与环境的关系有所不同(
Zhao et al ., 2014 ).本研究中, 随着植被恢复, 土壤C含量增加, 叶片C含量增加, 与
Zhang等(2019) 的研究结果一致.同时, 土壤C含量与叶片N、P含量显著正相关, 表明随着植被恢复, 土壤有机质的积累和分解释放出更多可利用性N、P, 促进叶片对N、P的吸收.此外, 土壤N含量与叶片C、N含量显著正相关, 表明随植被恢复土壤N含量增加, 有利于植物的生长发育, 促进光合作用合成更多的有机物质.叶片N的变化决定着叶片C:N的变化.因此, 土壤C、N含量与叶片N的正相关性可以解释土壤C、N含量与叶片C:N的负相关性.本研究中, 土壤C:P、N:P与叶片P含量显著正相关, 与叶片C:P、N:P显著负相关, 可能是由于随着植被恢复, 土壤C:P、N:P增加, 植物受P的限制更为明显, 植物通过提高对P的吸收量(降低P的利用效率)和再吸收效率来维持正常生理活动, 这也是叶片P含量增加而C:P、N:P降低的原因之一. ...
我国森林地表凋落物现存量及养分特征
1
2012
... 为了避免偶然性, 分别在2016年4、6、10月下旬采集植物叶片、凋落物、0-30 cm土层分析样品, 取3次采样测定的平均值作为最终的数据.在LVR固定样地, 沿着对角线均匀设置3个1 m × 1 m样方, 采集样方内所有灌木叶片、草本植物地上部分, 将同一样地3个样方植物叶片等比例混合为1个样品.根据群落调查结果(
辜翔等, 2018 ), 确定LCQ、PLL和LCC每块固定样地的优势树种, 对该固定样地的每种优势树种分别选取9株长势良好的样木.每种优势树种每次采集3株样木的叶片, 按上、中、下3个部位的东、西、南、北4个方向采集各株样木已完全成熟、无虫害的叶片, 草本植物地上部分采集同LVR, 再将同一样地内草本植物和优势树种叶片按生物量折算比例混合为1个样品.按
郑路和卢立华(2012) 的凋落物层分层标准, 在完成了草本植物地上部分样品采集后的3个1 m × 1 m的样方内采集未分解层的凋落物样品.完成凋落物采集后, 将样方内剩余凋落物清理干净, 用土钻(内径10 cm)取0-30 cm土层土壤样品, 将同一样地3个样方的凋落物、土壤分别混合为1个样品.所有的分析样品每次采样4个重复, 3次采样12个重复. ...
我国森林地表凋落物现存量及养分特征
1
2012
... 为了避免偶然性, 分别在2016年4、6、10月下旬采集植物叶片、凋落物、0-30 cm土层分析样品, 取3次采样测定的平均值作为最终的数据.在LVR固定样地, 沿着对角线均匀设置3个1 m × 1 m样方, 采集样方内所有灌木叶片、草本植物地上部分, 将同一样地3个样方植物叶片等比例混合为1个样品.根据群落调查结果(
辜翔等, 2018 ), 确定LCQ、PLL和LCC每块固定样地的优势树种, 对该固定样地的每种优势树种分别选取9株长势良好的样木.每种优势树种每次采集3株样木的叶片, 按上、中、下3个部位的东、西、南、北4个方向采集各株样木已完全成熟、无虫害的叶片, 草本植物地上部分采集同LVR, 再将同一样地内草本植物和优势树种叶片按生物量折算比例混合为1个样品.按
郑路和卢立华(2012) 的凋落物层分层标准, 在完成了草本植物地上部分样品采集后的3个1 m × 1 m的样方内采集未分解层的凋落物样品.完成凋落物采集后, 将样方内剩余凋落物清理干净, 用土钻(内径10 cm)取0-30 cm土层土壤样品, 将同一样地3个样方的凋落物、土壤分别混合为1个样品.所有的分析样品每次采样4个重复, 3次采样12个重复. ...
Ecological stoichiometry of carbon, nitrogen, and phosphorus in estuarine wetland soils: Influences of vegetation coverage, plant communities, geomorphology, and seawalls
1
2013
... 凋落物是养分从植物到土壤的基本载体, 其动态交换可以实现并维持土壤养分和植物生长所需的元素比率之间的平衡(
Hessen et al ., 2004 ).凋落物养分含量及其化学计量比与土壤养分含量和化学计量比显著相关(
Yang et al ., 2018 ), 一方面, 表明凋落物分解促使有机物质向土壤释放N、P, 从而影响土壤C、N和P含量及其化学计量比(
Zhong et al ., 2013 ), 另一方面, 表明土壤养分含量的变化会影响植物对养分的利用策略, 进而影响凋落物养分含量及其化学计量比.本研究中, 土壤C、N含量与凋落物C、N含量显著正相关, 可能是随着植被恢复, 土壤C、N含量增加, 更能满足植物对养分的需求, 从而影响植物对养分的再吸收效率, 进而影响凋落物的养分含量.因此, 叶片N、P含量显著影响着凋落物养分含量.此外, 本研究中, 土壤C:N、C:P和N:P与凋落物C:N、C:P显著正相关, 与
汪宗飞和郑粉莉(2018) 、
张萍等(2018) 的研究结果略有不同, 可能是由于凋落物P含量与土壤P含量的关系因研究区域和物种不同而存在差异, P含量在凋落物和土壤之间为显著负相关关系(
张萍等, 2018 )或无相关性(
聂兰琴等, 2016 ).也由于植物对养分的需求和归还是由土壤养分含量和限制性元素共同控制(
Güsewell & Verhoeven, 2006 ;
Chen et al ., 2018 ), 从叶片N、P含量及其化学计量比和养分再吸收效率可知, 土壤C:N、C:P和N:P高, 表明土壤可利用性N、P含量低, 植物为了维持生长, 在组织凋落前转移了大部分养分, 凋落物养分含量下降, 凋落物C:N、C:P增加. ...
 ,
, ,1,2,*
,1,2,*
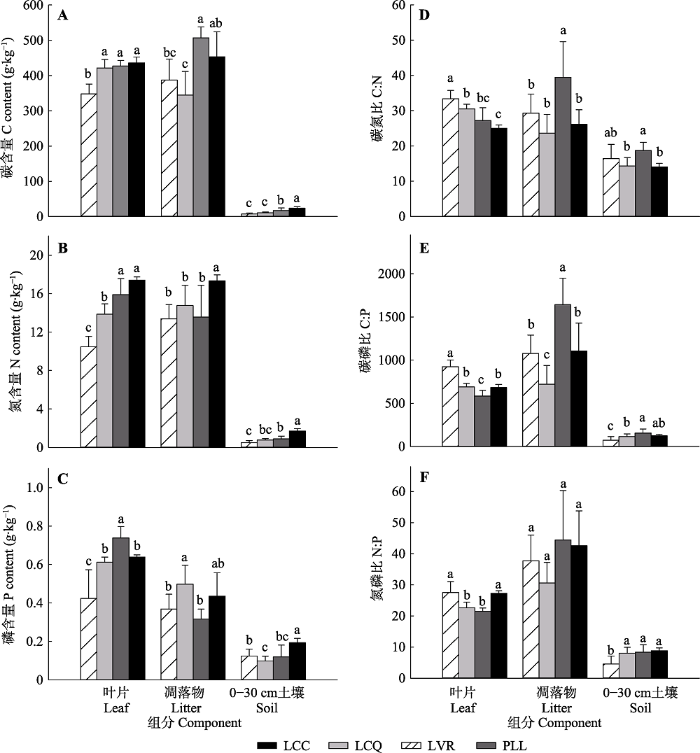 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT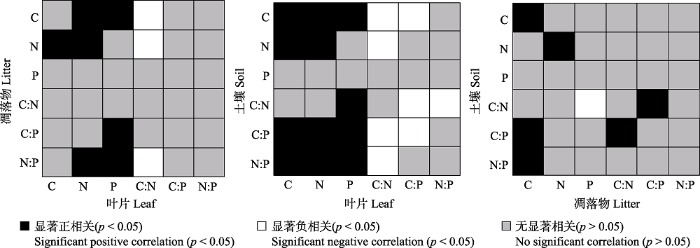 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT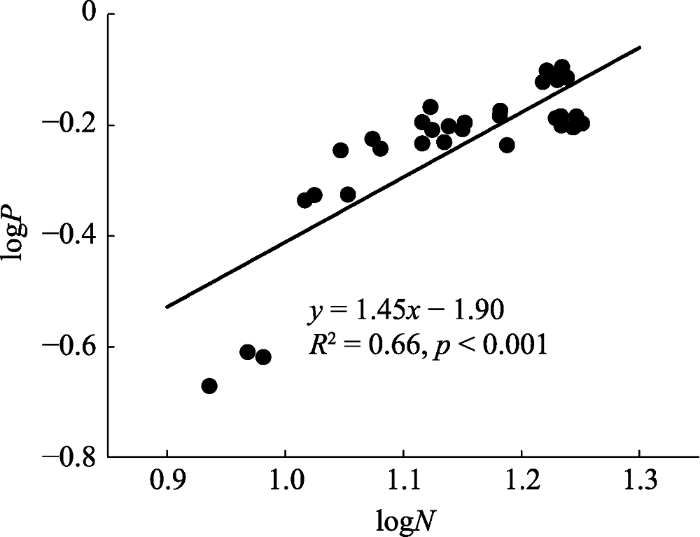 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT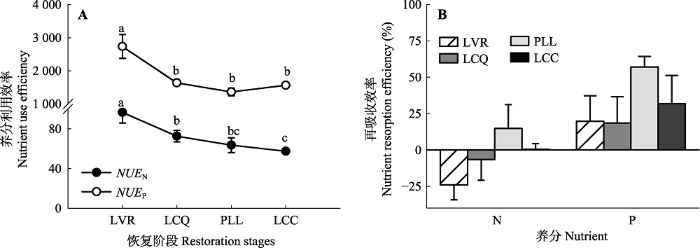 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT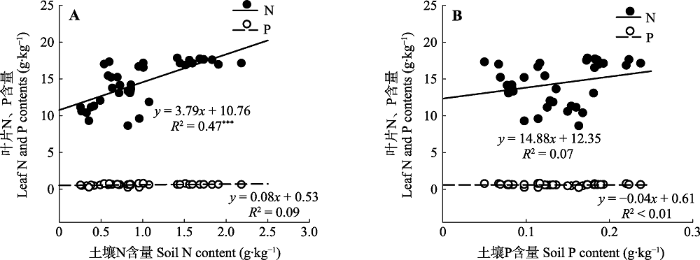 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}