冯璐
, 1 , 3 , * , 卜兆君
2 , 3 , 吴玉环
4 , 刘莎莎
2 , 3 , 刘超
2 , 3 1 滨州学院山东省黄河三角洲生态环境重点实验室, 山东滨州 256603 2 东北师范大学地理科学学院, 长白山地理过程与生态安全教育部重点实验室, 长春 130024 3 东北师范大学泥炭沼泽研究所, 国家环境保护湿地生态与植被恢复重点实验室, 长春 130024 4 杭州师范大学生命与环境科学学院, 杭州 310036 Characteristic environmental factors in peatlands facilitate the formation of persistent Sphagnum spore banks Lu FENG
, 1 , 3 , * , Zhao-Jun BU
2 , 3 , Yu-Huan WU
4 , Sha-Sha LIU
2 , 3 , Chao LIU
2 , 3 1 Shandong Key Laboratory of Eco-Environmental Science for the Yellow River Delta, Binzhou University, Binzhou, Shandong 256603, China 2 Key Laboratory of Geographical Processes and Ecological Security in Changbai Mountain, Ministry of Education, School of Geographical Sciences, Northeast Normal University, Changchun 130024, China 3 State Environmental Protection Key Laboratory of Wetland Ecology and Vegetation Restoration, Institute for Peat and Mire Research, Northeast Normal University, Changchun 130024, China 4 School of Life and Environment Science, Hangzhou Normal University, Hangzhou 310036, China 通讯作者: * fengl144@nenu.edu.cn 编委: 任海
责任编辑: 李敏, 赵航(实习)
收稿日期: 2019-04-15
修回日期: 2019-06-3
网络出版日期: 2019-06-20
基金资助: 国家自然科学基金 (41471043 )国家自然科学基金 (41871046 )国家自然科学基金 (41371103 )吉林省科技发展计划 (20190101025JH )山东省自然科学基金 (ZR2019PD008 )
Corresponding authors: * fengl144@nenu.edu.cn Received: 2019-04-15
Revised: 2019-06-3
Online: 2019-06-20
Fund supported: Supported by the National Natural Science Foundation of China (41471043 )Supported by the National Natural Science Foundation of China (41871046 )Supported by the National Natural Science Foundation of China (41371103 )Jilin Provincial Science and Technology Development Project (20190101025JH )the National Natural Science Foundation of Shandong Province (ZR2019PD008 )
摘要 研究泥炭地特征性环境因子——淹水、少氧和化感物质对泥炭藓孢子持久性的影响, 可深入理解泥炭地泥炭藓持久孢子库的形成机制, 为退化泥炭地泥炭藓地被恢复研究提供参考。该研究以藓丘种和丘间种两种泥炭藓的孢子为试验材料, 通过室内模拟控制实验的方法, 研究泥炭藓孢子在空气、超纯水、泥炭地地表水和泥炭藓沥出液中, 及3种速率充气下, 孢子萌发力持久性的变化。经充气处理后, 泥炭藓孢子持久性显著低于不充气处理。不充气时, 泥炭藓孢子在含有化感物质的泥炭地地表水和泥炭藓沥出液中保存, 持久性显著高于在超纯水中保存。通径分析结果显示, 溶解氧是影响泥炭地泥炭藓孢子持久性的主要因子和限制因子, 养分元素氮(TN)和磷(TP)的浓度为孢子持久性的负作用因子。研究结果表明, 泥炭藓孢子散布于苔藓地被基质或淹水的丘间生境中, 比暴露于空气或在无化感物质的水中, 能更好地维持萌发力。泥炭地中, 泥炭藓孢子和其他植物的繁殖体的超长寿命可能归因于少氧、养分贫乏和丰富的化感物质等泥炭地特征性环境因子。
关键词: 泥炭藓 ;
孢子持久性 ;
泥炭地 ;
溶解氧 ;
化感物质 Abstract Aims To test the effects of environmental factors in peatlands on the persistence of
Sphagnum spore germinability. The results may help to understand the mechanisms behind the formation of
Sphagnum spore banks in peatlands. They can also provide valuable insights for restoration of degraded peatlands.
Methods We determined the initial germination percentage in spores of two
Sphagnum species (hummock- forming
Sphagnum capillifolium and hollow-forming
S. flexuosum ) and then stored them for 60 days, either dry, in ultrapure water, peatland surface water or
Sphagnum water leachate. We varied oxygen concentration by injecting air at three concentrations during the storage experiment. After retrieval from experimental storage, spore germinability was assessed.
Important findings Spore germinability was lower after air-injection than under oxygen-deficiency. Spore germinability was higher after storage in the peatland surface and
Sphagnum leachate water, having high concentrations of allelochemicals, than in ultrapure water, under oxygen-deficiency. Path analysis showed that dissolved oxygen is the main factor negatively affecting
Sphagnum spore persistence in peatlands. Nitrogen (TN) and phosphorus (TP) also affect spore persistence negatively. These results indicate that once dispersed onto
Sphagnum substrates or waterlogged hollows,
Sphagnum spores can remain viable longer than when exposed to dry conditions or in water without allelochemicals. Extreme longevity of
Sphagnum spores and other plant propagules may be attributed to the oxygen-deficient, nutrient-poor and allelopathic substrates in peatlands.
Keywords: Sphagnum ;
spore persistence ;
peatland ;
dissolved oxygen ;
allelochemicals PDF (1113KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 冯璐, 卜兆君, 吴玉环, 刘莎莎, 刘超. 泥炭地特征性环境因子促进泥炭藓持久孢子库的形成. 植物生态学报, 2019, 43(6): 512-520. DOI:
10.17521/cjpe.2019.0082 FENG Lu, BU Zhao-Jun, WU Yu-Huan, LIU Sha-Sha, LIU Chao.
Characteristic environmental factors in peatlands facilitate the formation of persistent Sphagnum spore banks .
Chinese Journal of Plant Ecology , 2019, 43(6): 512-520. DOI:
10.17521/cjpe.2019.0082 因在植物种群的持续更新(
Saatkamp et al ., 2014 )和遗传多样性维持(
Tellier, 2019 )方面发挥重要作用, 植物持久有性繁殖体库(种子库和孢子库)备受****们关注。泥炭藓(
Sphagnum spp.)是北方泥炭地的主要造炭植物(
Turetsky, 2003 ), 以泥炭藓为优势植物的泥炭地生态系统, 长期积水, 氧和有效养分贫乏, 大量植物残体分解以及泥炭藓酸释放导致环境酸化, 次生代谢物质丰富, 泥炭地的这些特征性环境条件可能是诸多超长寿命的植物繁殖体(
Clymo & Duckett, 1986 ;
Shen-Miller et al ., 1995 ;
Sundberg & Rydin, 2000 ;
Bu et al ., 2017b )在此被发现的重要原因。
植物有性繁殖体的持久性因物种和环境条件的不同差异较大。由微生境差异引起的水位、pH、养分、离子浓度等环境因子的差异, 构成了泥炭地的环境梯度(
Wheeler & Proctor, 2000 ), 这种环境梯度均可能影响泥炭藓孢子的生活力、萌发和建植。如
Bu等(2017b) 在长白山哈泥泥炭地的研究发现, 水位较高的丘间生境中泥炭藓孢子的寿命更长。
Feng等(2017) 为期一年的埋藏实验发现, 间歇性淹水生境比从不淹水和持续淹水生境, 更适宜泥炭藓孢子萌发力的保持。泥炭藓孢子可以在添加动物粪便、桦树叶或泥炭藓灰分的泥炭中萌发(
Sundberg & Rydin, 2002 ), 表明泥炭地中养分较高的生境可能不利于泥炭藓持久孢子库的形成和稳定。然而, 尚无直接实验证据表明泥炭地的环境因子对地层中泥炭藓孢子寿命的影响。
苔藓植物通常生产较维管植物更为丰富的次生代谢物质, 其强烈的化感作用可以抑制微生物(
Montenegro et al ., 2009 )和维管植物种子的萌发和幼苗的生长(
Whitehead et al ., 2018 ), 如18种苔藓植物化感提取液均抑制
Fuchsia excorticata 种子的萌发和胚根的生长(
Michel et al ., 2011 )。苔藓地被基质的化感作用可能在泥炭藓持久孢子库的形成过程中产生关键作用, 然而受到的关注较少。已有研究发现, 金发藓(
Polytrichum strictum )地被基质的化感效应强于泥炭藓(
Sphagnum palustre )和中位泥炭藓(
Sphagnum magellanicum ), 可强烈抑制以上3种苔藓孢子的萌发(
Bu et al ., 2017a )。然而, 不萌发并不代表失去生活力, 这可能是植物分摊风险的“两头下注”策略(
Ooi et al ., 2009 ), 孢子萌发受到抑制, 却进入泥炭地地层中保持生活力, 是持久有性繁殖体库形成的前提。
对大部分植物种子而言, 环境中的氧含量直接影响其生活力和寿命。如4种十字花科植物的种子在不同气体和水环境中保存36年, 结果发现充满CO
2 的气体环境(缺氧)能提高种子的寿命(
González- Benito et al ., 2011 )。泥炭藓每年均产生大量孢子散布于泥炭地中, 随着泥炭的逐年累积, 在地层中有明显的垂直分布格局(
Bu et al ., 2017b )。在泥炭地地层中, 氧含量随着深度增加而减少(
Jauhiainen, 1998 ), 可能直接影响泥炭藓孢子的新陈代谢和老化速率, 进而影响其持久性。如
Sundberg和Rydin (2000)) 通过研究自然生境氧含量差异对泥炭藓孢子生活力持久性的影响, 发现孢子在少氧的丘间生境埋藏比在有氧的藓丘生境更快失去生活力。然而,
Feng等(2017) 模拟自然生境泥炭藓孢子不同埋藏深度的实验, 在埋藏1年后, 表层泥炭中(多氧环境)的孢子持久性高于埋藏在深层泥炭(少氧环境)的孢子。因此, 环境中氧含量对泥炭藓孢子持久性具体有何影响尚无一致结论。
本文基于泥炭藓存在超长持久孢子库的现状, 针对其形成机制存在的知识空白, 考虑到泥炭藓孢子散布生境的条件, 通过不同保存液处理, 模拟泥炭藓孢子在自然状态中可能散布的生境(有无水淹和有无化感物质), 并通过不同程度充气处理模拟环境的氧含量(少氧和多氧), 旨在回答: 1)是否化感物质利于泥炭藓孢子萌发力的保持, 是否总酚浓度直接影响孢子持久性? 2)基于
Feng等(2017) 的结论, 是否少氧环境更利于泥炭藓孢子萌发力的保持, 孢子持久性是否随着溶解氧浓度增加而降低? 研究结果有助于深入理解泥炭地泥炭藓持久孢子库的形成机制, 为退化泥炭地泥炭藓地被恢复研究提供参考。
1 材料和方法 1.1 孢蒴采集 分别于2014和2015年7月在长白山哈泥泥炭地(42.22° N, 126.52° E)采集尖叶泥炭藓(
Sphagnum capillifolium )和喙叶泥炭藓(
Sphagnum flexuosum )的成熟、饱满、未开裂孢蒴, 两种泥炭藓分别为典型的藓丘种和丘间种泥炭藓。将采集的两种泥炭藓的孢蒴连带其配子体, 放入塑料瓶中保持湿润, 用便携式自制冰盒带回实验室, 于4 ℃冰箱内黑暗保存备用。
1.2 实验设计 取两种泥炭藓的孢蒴各10个, 分别在烧杯中捣碎, 加入30 mL蒸馏水制成孢子悬浮液。吸取1 mL孢子悬液注入孔径7 μm的筛网制成的小包中, 滤除蒸馏水, 用19 mm长尾夹封口, 制成“孢子小包”。
本实验采用析因设计实验。将两种泥炭藓的“孢子小包”分别放入装有超纯水、泥炭地地表水和泥炭藓沥出液的烧杯中保存, 并通过充气泵注入空气, 根据自然状态下各溶液的溶解氧浓度和氧饱和状态下的溶解氧浓度, 设置3种不同程度充气处理, 分别为不充气、低速率充气(2.5 L·min
-1 )和高速率充气(4 L·min
-1 ), 并设置一组没有任何处理的在空气中保存。其中泥炭地地表水收集于哈泥泥炭地的水淹生境, 泥炭藓沥出液通过室内培养的中位泥炭藓的蒸馏水淋溶液获得, 具体方法为: 参照野外中位泥炭藓生长密度和夏季平均降水量, 将100 mL蒸馏水喷洒在装有20株中位泥炭藓的培养杯中, 过24 h后收集沥出液。所有处理中的水溶液均为100 mL, 每天由于蒸发损失的水分用超纯水补给, 每隔2日将所有溶液更新。实验处理进行60天。实验全程进行遮光处理, 防止孢子萌发影响实验结果。
1.3 测定方法 实验处理期间, 采用便携式多参数水质分析仪(HACH-HQ30d, Hach Company, Loveland, USA)测定不同处理的溶解氧浓度(
DO )、pH值和氧化还原电位(
Eh )。福林酚法(
Yu & Dahlgren, 2000 )测定超纯水、泥炭地地表水和泥炭藓沥出液的总酚浓度。使用流动元素分析仪(Smartchem 140, Westco Scientifiic, Brookfield, USA)测定上述3种保存液的总氮(TN)和总磷(TP)含量。使用火焰原子吸收分光光度计(Spectr AA 220 FS, Mulgrave, Victoria, Australia)测定上述3种保存液的K
+ 、Ca
2+ 、Na
+ 、Mg
2+ 浓度。
处理结束后, 将孢子接种到改良的Rudolph培养基(
Rudolph et al ., 1988 )上, 放入人工气候箱中进行培养。设置条件为白天16 h, 光照强度60 μmol·m
-2 ·s
-1 , 温度25 ℃; 夜晚8 h, 无光照, 温度20 ℃。在孢子培养的第3天进行第1次观察, 之后每4天观察一次孢子萌发情况。
1.4 数据分析 孢子持久性通过萌发率保持率(
GCR )计算, 公式如下:
(1) $GCR(\%)=(实验处理后的发芽率/厨师萌发率) ×100\%$ 所有数据用SPSS 19.0统计分析软件进行分析。对因变量孢子
GCR 数据进行正态检验, 服从正态分布可直接进行回归分析和通径分析(
杜家菊和陈志伟, 2010 ), 不符合正态分布则对数据进行标准化处理。双因素方差分析检验保存液类型、充气速率及二者交互作用对孢子持久性的影响。单因素方差分析比较不同处理中的
DO 、pH值、
Eh 及主要元素和总酚浓度的差异。单因素方差分析和
t 检验分组比较不同处理对孢子持久性影响的差异(分组如下: 超纯水不充气、泥炭地地表水不充气和泥炭藓沥出液不充气, 超纯水不充气、低速率充气和高速率充气, 泥炭地地表水不充气、低速率充气和高速率充气, 泥炭藓沥出液不充气、低速率充气和高速率充气, 空气和超纯水不充气)。多重比较采用最小显著差异法(
LSD )检验(
α = 0.05)。通径分析方法比较各因子对孢子持久性的直接和间接影响的相对重要性。
2 结果和分析 2.1 保存液理化指标差 三种保存液的
DO 、pH值和
Eh 及主要元素和总酚浓度均有显著差异(
表1 ,
表2 )。充气显著影响
DO 、pH值和
Eh, 整体上
DO 和pH值随着充气速率增大而增加。在超纯水和泥炭地地表水中,
Eh 随着充气速率增大而增加, 但在泥炭藓沥出液中
Eh 随着充气速率增大而减小。泥炭藓沥出液的TN、TP、K
+ 和Na
+ 浓度显著高于泥炭地地表水, 但Ca
2+ 、Mg
2+ 和总酚浓度反之。
Table 1 表1 表1 不同保存液和不同充气速率处理中的溶解氧浓度(
DO )、pH值和氧化还原电位(
Eh )(平均值±标准误差,
n = 3)
Table 1
Dissolved oxygen concentration (DO ), pH value and redox potential (Eh ) in different water storage solutions with or without air injection (mean ± SE, n = 3) DO (mg·L-1 )pH Eh (mV)超纯水 不充气 Control 8.91 ± 0.02Ab 5.36 ± 0.04Bb 196.7 ± 3.9Bb 低速率充气 Low 8.84 ± 0.02c 5.24 ± 0.06b 191.0 ± 5.0b 高速率充气 High 9.09 ± 0.01a 5.55 ± 0.07a 206.2 ± 1.0a 泥炭地地表水 不充气 Control 6.91 ± 0.02Cc 5.80 ± 0.02A 181.2 ± 3.7Cc 低速率充气 Low 9.60 ± 0.04b 7.47 ± 0.06a 192.8 ± 1.4b 高速率充气 High 10.08 ± 0.03a 7.19 ± 0.01b 198.6 ± 1.6a 中位泥炭藓沥出液Sphagnum magellanicum 不充气 Control 7.52 ± 0.17Bc 4.96 ± 0.06Cc 247.1 ± 2.5Aa 低速率充气 Low 9.70 ± 0.02b 5.91 ± 0.09b 196.6 ± 5.8b 高速率充气 High 10.25 ± 0.02a 6.14 ± 0.01b 183.9 ± 0.4c
Different lowercase letters indicated significant differences among different air injection treatments for the same indicator (p < 0.05); different uppercase letters indicated significant differences among different storage solutions for the same indicator (p < 0.05). 不同小写字母表示同一指标在3种充气处理下差异显著(p < 0.05); 不同大写字母不同表示同一指标在各保存液间差异显著(p < 0.05)。 新窗口打开 |
下载CSV Table 2 表2 表2 三种保存液的主要化学元素和总酚浓度(mg·L
-1 )(平均值±标准误差,
n = 3)
Table 2
Main chemical elements and total phenolics concentration (mg·L-1 ) in three solutions (mean ± SE , n = 3) TN TP K+ Ca2+ Na+ Mg2+ 总酚 Phenolics 超纯水 Ultrapure water 0.00 ± 0.00c 0.00 ± 0.00c 0.00 ± 0.00c 0.05 ± 0.01c 0.00 ± 0.00c 0.02 ± 0.00c 0.00 ± 0.00c 泥炭地地表水 0.52 ± 0.05b 0.03 ± 0.01b 2.90 ± 0.33b 5.22 ± 0.70a 1.64 ± 0.16b 1.43 ± 0.12a 6.51 ± 0.05a 中位泥炭藓沥出液Sphagnum magellanicum 5.03 ± 0.31a 0.26 ± 0.04a 8.89 ± 1.13a 0.71 ± 0.49b 4.33 ± 0.18a 0.39 ± 0.06b 3.23 ± 0.09b
Different lowercase letters in each column indicates significant differences among different storage solution for the same indicator (p < 0.05). TN, total nitrogen; TP, total phosphorus. 每列不同小写字母表示不同保存液的同一指标间有显著差异(p < 0.05)。TN, 总氮; TP, 总磷。 新窗口打开 |
下载CSV 2.2 保存液类型和充气速率对泥炭藓孢子持久性的影响 保存液类型和充气速率对两种泥炭藓孢子
GCR 均有显著的交互作用(
表3 ), 均表现为当孢子保存于泥炭地地表水或泥炭藓沥出液中, 充气对孢子
GCR 有显著的负作用。
Table 3 表3 表3 保存液类型和充气速率及二者的交互作用对泥炭藓孢子持久性影响的双因素方差分析
Table 3
Two-way ANOVA on the effect of water type, air injection and the interaction between water type and air injection on spore persistence 来源 Source d.f. 尖叶泥炭藓Sphagnum 喙叶泥炭藓Sphagnum F p F p 保存液类型 Water type 2 3.74 0.044 3.33 0.059 充气速率 Air injection 2 16.31 0.000 2.90 0.081 保存液类型 × 充气速率 4 5.57 0.004 3.73 0.022
Bold values indicate significant effects (p < 0.05). 加粗表示影响显著(p < 0.05)。 新窗口打开 |
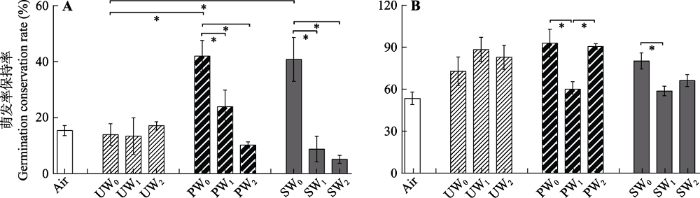
下载CSV 整体上, 不充气时(少氧条件下), 经泥炭地地表水和泥炭藓沥出液处理后的孢子
GCR (分别为67%和61%), 均比超纯水处理后的
GCR (44%)高。其中, 尖叶泥炭藓孢子经泥炭地地表水(
p = 0.016)和泥炭藓沥出液(
p = 0.019)保存后,
GCR 显著高于超纯水保存。两种泥炭藓孢子经无任何处理的空气中保存和超纯水不充气处理后, 萌发率无显著差异。
整体上, 充气处理后孢子
GCR 比不充气处理明显下降(从57%降至43%)。充气处理对超纯水中保存的两种泥炭藓孢子
GCR 均无显著影响。尖叶泥炭藓孢子, 在泥炭地地表水和泥炭藓沥出液中保存, 不充气处理的
GCR 分别显著高于低速率(
p = 0.035;
p = 0.005)和高速率充气(
p = 0.003;
p = 0.003)(
图1A )。喙叶泥炭藓孢子在泥炭地地表水中保存, 不充气(
p = 0.013)和高速率充气(
p = 0.018)处理后的
GCR 分别显著高于低速率充气; 在泥炭藓沥出液中保存, 不充气处理的
GCR 显著高于低速率充气(
p = 0.016) (
图1B )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1经3种保存液和3种程度充气处理后泥炭藓孢子的萌发率保持率(平均值±标准误差)。A, 尖叶泥炭藓(Sphagnum capillifolium )。B, 喙叶泥炭藓(Sphagnum flexuosum ); PW, 泥炭地表水; SW, 泥炭藓沥出液; UW, 超纯水。0, 不充气; 1, 低速率充气; 2, 高速率充气。*表示不同分组内(UW0 、PW0 和SW0 ; UW0 、UW1 和UW2 ; PW0 、PW1 和PW2 ; SW0 、SW1 和SW2 ; Air和UW0 )的处理间有显著差异(p < 0.05)。 Fig. 1Germination conservation rate of Sphagnum spores after three types of storage solutions and three levels of air injection. (mean ± SE ) A, S . capillifolium . B, S . flexuosum PW, peatland surface water; SW, Sphagnum leachate water; UW, ultrapurewater. 0, no air injection; 1, low air injection; 2, high air injection. * in each group (UW0 , PW0 and SW0 ; UW0 , UW1 and UW2 ; PW0 , PW1 and PW2 ; SW0 , SW1 and SW2 ; Air and UW0 ) indicated significant differences in one-way ANOVA (p < 0.05). 2.3 泥炭藓孢子持久性与保存液理化指标的相关性分析 尖叶泥炭藓和喙叶泥炭藓孢子
GCR 与
DO 均呈显著的负相关关系。尖叶泥炭藓孢子
GCR 与
Eh 呈显著正相关关系, 喙叶泥炭藓孢子
GCR 与TN、TP、K
+ 和Na
+ 浓度呈显著的负相关关系(
表4 )。
Table 4 表4 表4 保存液理化指标间及其与两种泥炭藓孢子持久性的相关关系
Table 4
Correlation analysis among water physicochemical indicators and between those indicators with sphagnum spore persistence 因子 Factor DO pH Eh TN TP K+ Ca2+ Na+ Mg2+ 总酚Phenolics DO pH 0.562 Eh -0.322 -0.401 TN 0.108 -0.183 0.364 TP 0.108 -0.175 0.362 1.000 K+ 0.096 -0.008 0.310 0.973 0.976 Ca2+ -0.074 0.765 -0.296 -0.307 -0.296 -0.080 Na+ 0.091 0.038 0.293 0.958 0.961 0.998 -0.022 Mg2+ -0.061 0.766 -0.254 -0.173 -0.161 0.058 0.990 0.116 总酚 0.047 0.767 -0.156 0.051 0.063 0.277 0.925 0.332 0.965 尖叶泥炭藓孢子持久性Sphagnum capillifolium GCR -0.777 -0.187 0.375 -0.126 -0.123 -0.057 0.310 -0.039 0.303 0.206 喙叶泥炭藓孢子持久性Sphagnum flexuosum GCR -0.402 -0.207 0.052 -0.382 -0.382 -0.365 0.146 -0.357 0.096 0.031
Bold correlation coefficients indicate significant correlation (p < 0.05). DO, dissolve oxygen; Eh, redox potential; GCR, germination conservation rate; TN, total nitrogen; TP, total phosphorus. 相关系数加粗表示显著相关(p < 0.05)。DO 。溶解氧; Eh, 氧化还原电位; GCR, 萌发率保持率; TN, 总氮; TP, 总磷。 新窗口打开 |
下载CSV 各理化指标中,
DO 与pH值, pH值与Ca
2+ 、Mg
2+ 、总酚浓度, TN与TP、K
+ 、Na
+ 浓度, TP与K
+ 、Na
+ 浓度, K
+ 与Na
+ 浓度, Ca
2+ 与Mg
2+ 、总酚浓度, Mg
2+ 与总酚浓度相关性较高(相关系数大于0.5)(
表4 )。
2.4 泥炭藓孢子持久性与保存液理化指标的多元回归分析 以保存液理化指标为自变量, 通过逐步回归建立与孢子
GCR 的最优回归方程: 尖叶泥炭藓孢子
GCR = 14.639 - 1.171
DO (
R 2 = 0.604,
p < 0.001), 方程筛选出
DO 对尖叶泥炭藓孢子
GCR 有显著影响, 由决定系数
R 2 可知其对尖叶泥炭藓孢子
GCR 的影响高达60.4%以上。喙叶泥炭藓孢子
GCR = 130.83 - 5.99
DO (
R 2 = 0.162,
p = 0.038), 方程筛选出
DO 对喙叶泥炭藓孢子
GCR 有显著影响, 但影响仅为16.2%。
2.5 泥炭藓孢子持久性与保存液理化指标的通径分析 保存液的理化指标对两种泥炭藓孢子
GCR 直接通径系数的绝对值由大到小依次为: 尖叶泥炭藓孢子:
DO 、pH值、Mg
2+ 浓度、Ca
2+ 浓度、总酚浓度、
Eh 、TN浓度、TP浓度、Na
+ 浓度、K
+ 浓度; 喙叶泥炭藓孢子:
DO 、TN浓度、TP浓度、K
+ 浓度、Na
+ 浓度、Ca
2+ 浓度、
Eh 、Mg
2+ 浓度、总酚浓度、pH值(
表5 )。
Table 5 表5 表5 两种泥炭藓孢子持久性与保存液理化指标的通径分析
Table 5
Path analysis of spore persistence and water physicochemical indicators 物种 因子 直接作用 间接作用 Indirect effect DO pH Eh TN TP K+ Ca2+ Na+ Mg2+ 总酚Phenolics 尖叶泥炭藓Sphagnum capillifolium DO -0.777 0.270 -0.068 -0.007 -0.007 0.003 -0.030 0.005 -0.025 0.018 pH 0.480 -0.437 -0.084 0.012 0.011 0.000 0.308 0.002 0.312 0.297 Eh 0.210 0.250 -0.192 -0.024 -0.022 0.008 -0.119 0.015 -0.103 -0.061 TN -0.067 -0.103 -0.088 0.076 -0.062 0.026 -0.124 0.049 -0.070 0.020 TP -0.062 -0.102 -0.084 0.076 -0.067 0.026 -0.119 0.049 -0.066 0.024 K+ 0.027 -0.091 -0.004 0.065 -0.065 -0.061 -0.032 0.051 0.024 0.107 Ca2+ 0.403 0.070 0.367 -0.062 0.021 0.018 -0.002 -0.001 0.403 0.358 Na+ 0.051 -0.087 0.018 0.062 -0.064 -0.060 0.027 -0.009 0.047 0.129 Mg2+ 0.407 0.058 0.367 -0.053 0.012 0.010 0.002 0.399 0.006 0.374 总酚Phenolics 0.387 -0.045 0.368 -0.033 -0.003 -0.004 0.007 0.373 0.017 0.393 喙叶泥炭藓Sphagnum flexuosum DO -0.402 0.014 0.029 -0.040 -0.040 -0.034 -0.009 -0.032 -0.005 0.003 pH 0.025 -0.226 0.036 0.068 0.065 0.003 0.098 -0.013 0.060 0.042 Eh -0.089 0.129 -0.010 -0.135 -0.135 -0.111 -0.038 -0.103 -0.020 -0.009 TN -0.372 -0.044 -0.005 -0.032 -0.372 -0.348 -0.039 -0.336 -0.013 0.003 TP -0.372 -0.043 -0.004 -0.032 -0.372 -0.349 -0.038 -0.337 -0.013 0.003 K+ -0.358 -0.038 0.000 -0.028 -0.362 -0.363 -0.010 -0.350 0.005 0.015 Ca2+ 0.128 0.030 0.019 0.026 0.114 0.110 0.029 0.008 0.077 0.051 Na+ -0.351 -0.037 0.001 -0.026 -0.356 -0.358 -0.357 -0.003 0.009 0.018 Mg2+ 0.078 0.024 0.019 0.023 0.064 0.060 -0.021 0.127 -0.041 0.053 总酚Phenolics 0.055 -0.019 0.019 0.014 -0.019 -0.023 -0.099 0.118 -0.117 0.075
Bold coefficients of indirect effect indicate the factor has stronger indirect effect than direct effect. DO, dissolve oxygen; Eh, redox potential; TN, total nitrogen; TP, total phosphorus. 间接作用系数加粗表示该因子的间接作用大于直接作用. DO, 溶解氧; Eh, 氧化还原电位; TN, 总氮; TP, 总磷。 新窗口打开 |
下载CSV DO 对两种泥炭藓孢子
GCR 的直接作用最大, 并且均为负作用(
表5 )。pH值对尖叶泥炭藓孢子
GCR 的直接通径系数为0.480, 大于其所有间接通径系数, 表明pH值以直接作用为主。Ca
2+ 和Mg
2+ 浓度同样以直接作用为主。其他指标TN、TP、K
+ 、Na
+ 和总酚浓度均以间接作用为主。Ca
2+ 浓度对喙叶泥炭藓孢子
GCR 以直接作用为主(直接通径系数为0.128)。其他指标pH值、
Eh 、TN、TP、K
+ 、Na
+ 、Mg
2+ 和总酚浓度均以间接作用为主。
3 讨论和结论 本研究在实验室内模拟条件下完成, 尽管室内条件与自然环境有差异, 但实验所用的泥炭藓孢子、泥炭地地表水和制备沥出液的泥炭藓均采集于野外, 并且充气处理采用注入空气而不是纯氧, 所有实验条件最大限度地接近自然环境, 因此, 研究中各因子对泥炭藓孢子持久性的影响对于深刻认识泥炭藓持久孢子库的形成过程与机制有重要意义。在本试验的各因子中, 尖叶泥炭藓孢子持久性与
DO 和
Eh 显著相关, 其余指标与尖叶泥炭藓孢子持久性未达显著相关水平。喙叶泥炭藓孢子持久性与
DO 、TN、TP、K
+ 、Na
+ 浓度呈显著相关关系, 其余指标与喙叶泥炭藓孢子持久性未达显著相关水平。很多指标之间也存在显著相关关系。结合通径分析结果可知, 部分环境因子不仅直接影响孢子的持久性, 而且通过相互作用, 间接影响孢子持久性。因此, 简单的相关分析不能判定环境因子对泥炭藓孢子持久性的影响, 通过通径分析可区分各因子作用的大小(
Li et al ., 2018b ), 估计各因子的相对重要性, 提取关键影响因子。
苔藓植物作为从水生向陆生过渡的植物类群, 在进化过程中已具备了耐受干旱的能力(
Proctor et al ., 2007 )。因此, 两种泥炭藓孢子经空气和超纯水保存2个月后, 持久性无显著差异, 但空气保存后的萌发速度显著低于淹水保存, 表明孢子散布于淹水生境后, 时刻保持着快速萌发的能力。然而, 泥炭地正受到全球变化的威胁, 如全球变暖增加干旱频率, 导致泥炭地水位下降(
袁敏等, 2015 ), 很可能不利于地层中泥炭藓孢子的持久性。尖叶泥炭藓孢子经泥炭地地表水和泥炭藓沥出液保存后, 持久性显著高于超纯水保存, 表明纯水并不是决定孢子持久性的因素, 而水中的离子和化感物质可能是影响孢子持久性的关键因子。本研究结果与
Bu等(2017a) 苔藓化感作用减弱孢子萌发力的结论看似矛盾, 可能原因首先是实验物种的差异, 本文实验物种为尖叶泥炭藓和喙叶泥炭藓孢子, 而
Bu等(2017a) 实验物种为金发藓、泥炭藓和中位泥炭藓孢子, 本研究中尖叶泥炭藓孢子经3种保存液不充气处理后, 持久性之间有显著差异, 而喙叶泥炭藓孢子无显著差异, 可知不同种泥炭藓孢子对化感效应的响应不同。其次,
Bu等(2017a) 的研究中, 泥炭藓孢子经野外环境埋藏一个生长季, 可能遭受到紫外辐射和干旱等不利条件(
van Zanten, 1978 ), 导致孢子迅速老化, 持久性显著下降。此外, 由于其野外埋藏实验的苔藓基质化感物质浓度较高(
Mishler & Newton, 1988 ), 而本实验中的泥炭地地表水和泥炭藓沥出液分别经雨水和蒸馏水稀释, 降低了化感物质浓度。高浓度的化感物质虽然抑制孢子的萌发, 但很可能使其保持生活力, 而较低浓度的化感物质抑制微生物活性(
Verhoeven & Liefveld, 1997 ), 使孢子保持较高的萌发力。
成熟的泥炭藓孢子散布于潮湿的苔藓基质或积水的丘间生境中, 比散布于缺少化感物质的生境中能更好地维持萌发力。这与
Bu等(2017b) 对长白山哈泥泥炭地泥炭藓孢子寿命研究的结果一致, 即在少氧的丘间生境中泥炭藓孢子寿命更长。尽管通径分析结果显示, 总酚浓度并不是影响孢子持久性的主要因素, 但对两种泥炭藓孢子的持久性均表现为正作用, 并且通过Ca
2+ 浓度对其也有较大的间接正作用。此外, K
+ 和Na
+ 浓度通过总酚浓度对孢子的持久性也有间接的正作用。研究表明土荆芥(
Chenopodium ambrosioides )分泌的化感物质会增加受体植物细胞质中的Ca
2+ 累积(
Li et al ., 2018a )。环境中Na
+ 和K
+ 浓度增加, 促进
Salvia officinalis 分泌更多的酚类化感物质(
Taarit et al ., 2012 )。由此可推测, 若环境中或泥炭藓体内的Na
+ 和K
+ 浓度增加, 可能促进化感物质的分泌, 进而有助于泥炭藓孢子维持生活力和萌发力。本研究的泥炭地地表水中, Ca
2+ 和总酚浓度分别显著高于泥炭藓沥出液和超纯水, 泥炭地地表水中的高浓度酚类化合物促进Ca
2+ 的聚集, 虽抑制泥炭藓孢子的萌发(
刘礼洁等, 2019 ), 但却利于其保持生活力形成持久孢子库。因此也可解释丘间淹水生境埋藏的泥炭藓孢子寿命大于藓丘不淹水生境(
Bu et al ., 2017b )。
充气显著增加了保存液的
DO, 却降低了孢子持久性。由通径分析结果可知,
DO 是影响两种泥炭藓孢子持久性的最主要的作用因子。然而, 由保存液类型和充气速率对孢子持久性的交互作用可知, 充气在超纯水中对孢子持久性无显著影响, 但在泥炭地地表水和泥炭藓沥出液中, 却能显著降低孢子持久性。表明在本研究中
DO 不会直接氧化泥炭藓孢子, 加快其老化速率。虽然实验中总酚的浓度并未在充气条件下发生显著变化, 但其组成复杂(
H?ttenschwiler & Vitousek, 2000 ), 某些关键组分被氧化, 如泥炭藓酸(
Abbott et al ., 2013 ), 化感物质组分发生改变, 使原本富含化感物质的泥炭地地表水和泥炭藓沥出液的抑菌性下降, 提高了好氧细菌的活性(
Ke et al ., 2015 ), 进而加速孢子的分解, 不利于其保持萌发力。此外, 由多元回归分析结果可知,
DO 对两种泥炭藓孢子持久性的影响分别为60.4%和16.2%, 表明还有其他因素对孢子持久性有影响, 但未被考虑进来。因此, 泥炭地环境因子对泥炭藓孢子持久性的影响较为复杂, 若要深入研究泥炭地泥炭藓持久孢子库的形成机理, 需要考虑更多的影响因素, 例如环境温度、光照强度、化感物质各组分浓度和微生物活性的影响等。尤其在全球变暖的大背景下, 温度升高打破地层泥炭藓孢子休眠, 增加不必要的萌发, 减少泥炭藓孢子库储存(
Mcletchie, 1999 )。
N和P浓度是泥炭地植被生长的主要限制因子(
Aerts et al ., 1992 )。通过相关分析和通径分析结果可知, TN和TP浓度对两种泥炭藓孢子的持久性均表现为负作用, 尤其对喙叶泥炭藓孢子的持久性为直接的负作用。Ca
2+ 浓度对尖叶泥炭藓孢子的持久性有较大的间接负作用, 表明养分元素对泥炭藓孢子的持久性起到限制作用。
Boatman和Lark (1971) 研究认为泥炭藓孢子不能在野外条件下萌发建植是由于泥炭地养分的缺乏, 因此, 若雨养型泥炭地养分输入增加, 可能导致泥炭藓孢子不必要的萌发, 从而不利于泥炭藓孢子库的稳定。此外, 随着泥炭地中养分增加, 使原本养分较贫乏(
Song et al ., 2017 )的环境中微生物数量增加、活性增强(
Fierer et al ., 2009 ), 从而加速泥炭藓孢子的分解, 不利于其保持萌发力。因此可推测, 在全球变化大背景下, 大气氮沉降导致泥炭地养分输入增加, 将不利于泥炭藓持久孢子库的稳定。
在北方泥炭地, 成熟的泥炭藓孢子可能散布于不同的生境, 如干旱或潮湿的泥炭藓丛、间歇性淹水的丘间生境和持续淹水的低洼生境。在不淹水的泥炭藓地被基质内, 由于较高浓度的化感物质抑制孢子的萌发, 但却使其在短期内保持生活力。在间歇性淹水生境, 孢子保持较高的萌发力, 待到适宜的条件, 如光照、养分和温度, 即可萌发(
Feng et al ., 2017 )。在持续淹水生境, 泥炭藓孢子不能萌发, 但可长期保持较高的生活力(
Sundberg & Rydin, 2000 )。本研究表明, 溶解氧为影响泥炭地泥炭藓孢子持久性的主要因子, 同时也是限制因子, 化感物质浓度为正作用因子, 养分元素N和P浓度为负作用因子, 很可能与微生物多样性和活性增加有关(
Pinsonneault et al ., 2016 )。因此, 少氧、淹水、养分较贫乏的泥炭藓泥炭地, 是泥炭藓孢子长期保持生活力的理想场所, 在泥炭地发现泥炭藓长期持久孢子库(
Bu et al ., 2017b )及其他植物的超长寿命繁殖体也归因于此。
致谢 感谢东北师范大学马进泽博士在泥炭地地表水采集工作中给予的帮助。
[1] Abbott GD Swain EY Muhammad AB Allton K Belyea LR Laing CG Cowie GL 2013 ). Effect of water-table fluctuations on the degradation of Sphagnum phenols in surficial peatsGeochimica et Cosmochimica Acta 177 -191 . [本文引用: 1] [2] Aerts R Wallen B Malmer N 1992 ). Growth-limiting nutrients in Sphagnum -dominated bogs subject to low and high atmospheric nitrogen supplyJournal of Ecology 131 -140 . [本文引用: 1] [3] Boatman DJ Lark PM 1971 ). Inorganic nutrition of the protonemata of Sphagnum papillosum Lindb., S. magellanicum Brid. and S. cuspidatum EhrhNew Phytologist 1053 -1059 . [本文引用: 1] [4] Bu ZJ Li Z Liu LJ Sundberg S Feng YM Yang YH Liu S Song X Zhang XL 2017a ). Bryophyte spore germinability is inhibited by peatland substratesActa Oecologica, 78, 34 -40 . [本文引用: 4] [5] Bu ZJ Sundberg S Feng L Li HK Zhao HY Li HC 2017b ). The Methuselah of plant diaspores: Sphagnum spores can survive in nature for centuriesNew Phytologist 1398 -1402 . [本文引用: 6] [6] Clymo RS Duckett JG 1986 ). Regeneration of Sphagnum New Phytologist 589 -614 . [本文引用: 1] [7] Du JJ Chen ZW 2010 ). The methods of path analysis by SPSS linear regressionBulletin of Biology, 45(2 ), 4 -6 . DOI:
10.3969/j.issn.0006-3193.2010.02.002 URL [本文引用: 1] 由于通径分析可以将因变量与自变量的相互影响(相关系数)分解为直接影响(通径系数)和间接影响(间接通径系数),因此在遗传学等领域受到广泛的重视。目前在软件实现方法上,一方面缺乏必要的正态性检验,另一方面通径系数及间接相关系数计算步骤过于繁琐,限制通径分析的教学和使用。在应用中,我们注意到通过SPSS的线性回归"Linear"程序可以一次性获得计算通径系数的全部数据,从而简化通径分析的步骤。
[ 杜家菊 , 陈志伟 (2010 ). 使用SPSS线性回归实现通径分析的方法生物学通报 , 45(2 ), 4 -6 .] DOI:
10.3969/j.issn.0006-3193.2010.02.002 URL [本文引用: 1] 由于通径分析可以将因变量与自变量的相互影响(相关系数)分解为直接影响(通径系数)和间接影响(间接通径系数),因此在遗传学等领域受到广泛的重视。目前在软件实现方法上,一方面缺乏必要的正态性检验,另一方面通径系数及间接相关系数计算步骤过于繁琐,限制通径分析的教学和使用。在应用中,我们注意到通过SPSS的线性回归"Linear"程序可以一次性获得计算通径系数的全部数据,从而简化通径分析的步骤。
[8] Feng L Bu ZJ Mallik A Wang ZC Liu SS Wu YH 2017 ). Continuous waterlogging may not facilitate germinability maintenance of Sphagnum sporesWetlands 1015 -1022 . [本文引用: 4] [9] Fierer N Strickland MS Liptzin D Bradford MA Cleveland CC 2009 ). Global patterns in belowground communitiesEcology Letters, 12, 1238 -1249 . [本文引用: 1] [10] González-Benito ME Pérez-García F Tejeda G Gómez- Campo C 2011 ). Effect of the gaseous environment and water content on seed viability of four Brassicaceae species after 36 years storageSeed Science and Technology, 39, 443 -451 . [本文引用: 1] [11] H?ttenschwiler S Vitousek PM 2000 ). The role of polyphenols in terrestrial ecosystem nutrient cyclingTrends in Ecology & Evolution, 15, 238 -243 . [本文引用: 1] [12] Jauhiainen S 1998 ). Seed and spore banks of two boreal miresAnnales Botanici Fennici, 35, 197 -201 . [本文引用: 1] [13] Ke X Lu W Conrad R 2015 ). High oxygen concentration increases the abundance and activity of bacterial rather than archaeal nitrifiers in rice field soilMicrobial Ecology, 70, 961 -970 . [本文引用: 1] [14] Li J He Y Ma D He B Wang Y Chen B 2018a ). Volatile allelochemicals of Chenopodium ambrosioides L. induced mitochondrion-mediated Ca 2+ -dependent and caspase-?dependent apoptosis signaling pathways in receptor plant cellsPlant and Soil 297 -308 . [15] Li Y Rashid A Wang HJ Hu AY Lin LF Yu CP Chen M Sun Q 2018b ). Contribution of biotic and abiotic factors in the natural attenuation of sulfamethoxazole: A path analysis approachScience of the Total Environment, 633, 1217 -1226 . [本文引用: 1] [16] Liu LJ Bu ZJ Liu S Chen YD Feng L Fu B Yang YH Wang SZ 2019 ). Sand and dust deposition may retard the autogenic vegetation succession of peatlandsScientia Geographica Sinica, 39, 351 -358 . [本文引用: 1] [ 刘礼洁 , 卜兆君 , 刘霜 , 陈永达 , 冯璐 , 付彪 , 杨云荷 , 王升忠 (2019 ). 沙尘沉降可能阻滞泥炭地植被的自发演替地理科学 , 39, 351 -358 .] [本文引用: 1] [17] McLetchie DN 1999 ). Dormancy/Nondormancy cycles in spores of the liverwort Sphaerocarpos texanus The Bryologist 15 -21 . [本文引用: 1] [18] Michel P Burritt DJ Lee WG 2011 ). Bryophytes display allelopathic interactions with tree species in native forest ecosystemsOikos, 120, 1272 -1280 . [本文引用: 1] [19] Mishler BD Newton AE 1988 ). Influences of mature plants and desiccation on germination of spores and gametophyticfragments of Tortula Journal of Bryology 327 -342 . [本文引用: 1] [20] Montenegro G Portaluppi MC Salas FA Díaz MF 2009 ). Biological properties of the Chilean native moss Sphagnum magellanicum Biological Research 233 -237 . [本文引用: 1] [21] Ooi MKJ Auld TD Denham AJ 2009 ). Climate change and bet-hedging: Interactions between increased soil temperatures and seed bank persistenceGlobal Change Biology, 15, 2375 -2386 . [本文引用: 1] [22] Pinsonneault AJ Moore TR Roulet NT 2016 ). Effects of long-term fertilization on peat stoichiometry and associated microbial enzyme activity in an ombrotrophic bogBiogeochemistry, 129, 149 -164 . [本文引用: 1] [23] Proctor MCF Oliver MJ Wood AJ Alpert P Stark LR Cleavitt NL Mishler BD 2007 ). Desiccation-tolerance in bryophytes: A reviewThe Bryologist, 110, 595 -621 . [本文引用: 1] [24] Rudolph H Kirchhoff M Gliesmann S 1988 ). Sphagnum culture techniques. In: Glime JM ed. Methods in BryologyProceedings of the Bryological Methods Workshop, Mainz, Hattori Botanical Laboratory, Nichinan, Japan. [本文引用: 1] [25] Saatkamp A Poschlod P Venable DL 2014 ). The functional role of soil seed banks in natural communitiesCABI , Wallingford. 263 -295 . [本文引用: 1] [26] Shen-Miller J Mudgett MB Schopf JW Clarke S Berger R 1995 ). Exceptional seed longevity and robust growth: Ancient Sacred Lotus from ChinaAmerican Journal of Botany, 82, 1367 -1380 . [本文引用: 1] [27] Song YY Song CC Meng HN Swarzenski CM Wang XW Tan WW 2017 ). Nitrogen additions affect litter quality and soil biochemical properties in a peatland of Northeast ChinaEcological Engineering, 100, 175 -185 . [本文引用: 1] [28] Sundberg S Rydin H 2000 ). Experimental evidence for a persistent spore bank in Sphagnum New Phytologist 105 -116 . [本文引用: 3] [29] Sundberg S Rydin H 2002 ). Habitat requirements for establishment of Sphagnum from sporesJournal of Ecology 268 -278 . [本文引用: 1] [30] Taarit MB Msaada K Hosni K Marzouk B 2012 ). Physiological changes, phenolic content and antioxidant activity of Salvia officinalis L. grown under saline conditionsJournal of the Science of Food and Agriculture 1614 -1619 . [本文引用: 1] [31] Tellier A 2019 ). Persistent seed banking as eco-evolutionary determinant of plant nucleotide diversity: Novel population genetics insightsNew Phytologist, 221, 725 -730 . [本文引用: 1] [32] Turetsky MR 2003 ). The role of bryophytes in carbon and nitrogen cyclingThe Bryologist, 106, 395 -409 . [本文引用: 1] [33] van Zanten BO 1978 ). Experimental studies on trans-oceanic long-range dispersal of moss spores in the Southern HemisphereJournal of the Hattori Botanical Laboratory, 44, 445 -482 . [本文引用: 1] [34] Verhoeven JTA Liefveld WM 1997 ). The ecological significance of organochemical compounds in Sphagnum Acta Botanica Neerlandica 117 -130 . [本文引用: 1] [35] Wheeler BD Proctor MCF 2000 ). Ecological gradients, subdivisions and terminology of north-west European miresJournal of Ecology, 88, 187 -203 . [本文引用: 1] [36] Whitehead J Wittemann M Cronberg N 2018 ). Allelopathy in bryophytes—A reviewLindbergia, 41, 01097 . DOI: 10.25227/linbg.01097 . [本文引用: 1] [37] Yu Z Dahlgren RA 2000 ). Evaluation of methods for measuring polyphenols in conifer foliageJournal of Chemical Ecology, 26, 2119 -2140 . [本文引用: 1] [38] Yuan M Bu ZJ Liu C Ma JZ Wang SZ 2015 ). Effects of water level and light intensity on capsule production dynamics of Sphagnum capillifolium Chinese Journal of Plant Ecology 501 -507 . [本文引用: 1] [ 袁敏 , 卜兆君 , 刘超 , 马进泽 , 王升忠 (2015 ). 水位与光强变化对尖叶泥炭藓孢蒴生产动态的影响植物生态学报 , 39, 501 -507 .] [本文引用: 1] Effect of water-table fluctuations on the degradation of
Sphagnum phenols in surficial peats
1
2013
... 充气显著增加了保存液的
DO, 却降低了孢子持久性.由通径分析结果可知,
DO 是影响两种泥炭藓孢子持久性的最主要的作用因子.然而, 由保存液类型和充气速率对孢子持久性的交互作用可知, 充气在超纯水中对孢子持久性无显著影响, 但在泥炭地地表水和泥炭藓沥出液中, 却能显著降低孢子持久性.表明在本研究中
DO 不会直接氧化泥炭藓孢子, 加快其老化速率.虽然实验中总酚的浓度并未在充气条件下发生显著变化, 但其组成复杂(
H?ttenschwiler & Vitousek, 2000 ), 某些关键组分被氧化, 如泥炭藓酸(
Abbott et al ., 2013 ), 化感物质组分发生改变, 使原本富含化感物质的泥炭地地表水和泥炭藓沥出液的抑菌性下降, 提高了好氧细菌的活性(
Ke et al ., 2015 ), 进而加速孢子的分解, 不利于其保持萌发力.此外, 由多元回归分析结果可知,
DO 对两种泥炭藓孢子持久性的影响分别为60.4%和16.2%, 表明还有其他因素对孢子持久性有影响, 但未被考虑进来.因此, 泥炭地环境因子对泥炭藓孢子持久性的影响较为复杂, 若要深入研究泥炭地泥炭藓持久孢子库的形成机理, 需要考虑更多的影响因素, 例如环境温度、光照强度、化感物质各组分浓度和微生物活性的影响等.尤其在全球变暖的大背景下, 温度升高打破地层泥炭藓孢子休眠, 增加不必要的萌发, 减少泥炭藓孢子库储存(
Mcletchie, 1999 ). ...
Growth-limiting nutrients in
Sphagnum -dominated bogs subject to low and high atmospheric nitrogen supply
1
1992
... N和P浓度是泥炭地植被生长的主要限制因子(
Aerts et al ., 1992 ).通过相关分析和通径分析结果可知, TN和TP浓度对两种泥炭藓孢子的持久性均表现为负作用, 尤其对喙叶泥炭藓孢子的持久性为直接的负作用.Ca
2+ 浓度对尖叶泥炭藓孢子的持久性有较大的间接负作用, 表明养分元素对泥炭藓孢子的持久性起到限制作用.
Boatman和Lark (1971) 研究认为泥炭藓孢子不能在野外条件下萌发建植是由于泥炭地养分的缺乏, 因此, 若雨养型泥炭地养分输入增加, 可能导致泥炭藓孢子不必要的萌发, 从而不利于泥炭藓孢子库的稳定.此外, 随着泥炭地中养分增加, 使原本养分较贫乏(
Song et al ., 2017 )的环境中微生物数量增加、活性增强(
Fierer et al ., 2009 ), 从而加速泥炭藓孢子的分解, 不利于其保持萌发力.因此可推测, 在全球变化大背景下, 大气氮沉降导致泥炭地养分输入增加, 将不利于泥炭藓持久孢子库的稳定. ...
Inorganic nutrition of the protonemata of
Sphagnum papillosum Lindb.,
S. magellanicum Brid. and
S. cuspidatum Ehrh
1
1971
... N和P浓度是泥炭地植被生长的主要限制因子(
Aerts et al ., 1992 ).通过相关分析和通径分析结果可知, TN和TP浓度对两种泥炭藓孢子的持久性均表现为负作用, 尤其对喙叶泥炭藓孢子的持久性为直接的负作用.Ca
2+ 浓度对尖叶泥炭藓孢子的持久性有较大的间接负作用, 表明养分元素对泥炭藓孢子的持久性起到限制作用.
Boatman和Lark (1971) 研究认为泥炭藓孢子不能在野外条件下萌发建植是由于泥炭地养分的缺乏, 因此, 若雨养型泥炭地养分输入增加, 可能导致泥炭藓孢子不必要的萌发, 从而不利于泥炭藓孢子库的稳定.此外, 随着泥炭地中养分增加, 使原本养分较贫乏(
Song et al ., 2017 )的环境中微生物数量增加、活性增强(
Fierer et al ., 2009 ), 从而加速泥炭藓孢子的分解, 不利于其保持萌发力.因此可推测, 在全球变化大背景下, 大气氮沉降导致泥炭地养分输入增加, 将不利于泥炭藓持久孢子库的稳定. ...
Bryophyte spore germinability is inhibited by peatland substrates
4
2017a
... 苔藓植物通常生产较维管植物更为丰富的次生代谢物质, 其强烈的化感作用可以抑制微生物(
Montenegro et al ., 2009 )和维管植物种子的萌发和幼苗的生长(
Whitehead et al ., 2018 ), 如18种苔藓植物化感提取液均抑制
Fuchsia excorticata 种子的萌发和胚根的生长(
Michel et al ., 2011 ).苔藓地被基质的化感作用可能在泥炭藓持久孢子库的形成过程中产生关键作用, 然而受到的关注较少.已有研究发现, 金发藓(
Polytrichum strictum )地被基质的化感效应强于泥炭藓(
Sphagnum palustre )和中位泥炭藓(
Sphagnum magellanicum ), 可强烈抑制以上3种苔藓孢子的萌发(
Bu et al ., 2017a ).然而, 不萌发并不代表失去生活力, 这可能是植物分摊风险的“两头下注”策略(
Ooi et al ., 2009 ), 孢子萌发受到抑制, 却进入泥炭地地层中保持生活力, 是持久有性繁殖体库形成的前提. ...
... 苔藓植物作为从水生向陆生过渡的植物类群, 在进化过程中已具备了耐受干旱的能力(
Proctor et al ., 2007 ).因此, 两种泥炭藓孢子经空气和超纯水保存2个月后, 持久性无显著差异, 但空气保存后的萌发速度显著低于淹水保存, 表明孢子散布于淹水生境后, 时刻保持着快速萌发的能力.然而, 泥炭地正受到全球变化的威胁, 如全球变暖增加干旱频率, 导致泥炭地水位下降(
袁敏等, 2015 ), 很可能不利于地层中泥炭藓孢子的持久性.尖叶泥炭藓孢子经泥炭地地表水和泥炭藓沥出液保存后, 持久性显著高于超纯水保存, 表明纯水并不是决定孢子持久性的因素, 而水中的离子和化感物质可能是影响孢子持久性的关键因子.本研究结果与
Bu等(2017a) 苔藓化感作用减弱孢子萌发力的结论看似矛盾, 可能原因首先是实验物种的差异, 本文实验物种为尖叶泥炭藓和喙叶泥炭藓孢子, 而
Bu等(2017a) 实验物种为金发藓、泥炭藓和中位泥炭藓孢子, 本研究中尖叶泥炭藓孢子经3种保存液不充气处理后, 持久性之间有显著差异, 而喙叶泥炭藓孢子无显著差异, 可知不同种泥炭藓孢子对化感效应的响应不同.其次,
Bu等(2017a) 的研究中, 泥炭藓孢子经野外环境埋藏一个生长季, 可能遭受到紫外辐射和干旱等不利条件(
van Zanten, 1978 ), 导致孢子迅速老化, 持久性显著下降.此外, 由于其野外埋藏实验的苔藓基质化感物质浓度较高(
Mishler & Newton, 1988 ), 而本实验中的泥炭地地表水和泥炭藓沥出液分别经雨水和蒸馏水稀释, 降低了化感物质浓度.高浓度的化感物质虽然抑制孢子的萌发, 但很可能使其保持生活力, 而较低浓度的化感物质抑制微生物活性(
Verhoeven & Liefveld, 1997 ), 使孢子保持较高的萌发力. ...
... 苔藓化感作用减弱孢子萌发力的结论看似矛盾, 可能原因首先是实验物种的差异, 本文实验物种为尖叶泥炭藓和喙叶泥炭藓孢子, 而
Bu等(2017a) 实验物种为金发藓、泥炭藓和中位泥炭藓孢子, 本研究中尖叶泥炭藓孢子经3种保存液不充气处理后, 持久性之间有显著差异, 而喙叶泥炭藓孢子无显著差异, 可知不同种泥炭藓孢子对化感效应的响应不同.其次,
Bu等(2017a) 的研究中, 泥炭藓孢子经野外环境埋藏一个生长季, 可能遭受到紫外辐射和干旱等不利条件(
van Zanten, 1978 ), 导致孢子迅速老化, 持久性显著下降.此外, 由于其野外埋藏实验的苔藓基质化感物质浓度较高(
Mishler & Newton, 1988 ), 而本实验中的泥炭地地表水和泥炭藓沥出液分别经雨水和蒸馏水稀释, 降低了化感物质浓度.高浓度的化感物质虽然抑制孢子的萌发, 但很可能使其保持生活力, 而较低浓度的化感物质抑制微生物活性(
Verhoeven & Liefveld, 1997 ), 使孢子保持较高的萌发力. ...
... 实验物种为金发藓、泥炭藓和中位泥炭藓孢子, 本研究中尖叶泥炭藓孢子经3种保存液不充气处理后, 持久性之间有显著差异, 而喙叶泥炭藓孢子无显著差异, 可知不同种泥炭藓孢子对化感效应的响应不同.其次,
Bu等(2017a) 的研究中, 泥炭藓孢子经野外环境埋藏一个生长季, 可能遭受到紫外辐射和干旱等不利条件(
van Zanten, 1978 ), 导致孢子迅速老化, 持久性显著下降.此外, 由于其野外埋藏实验的苔藓基质化感物质浓度较高(
Mishler & Newton, 1988 ), 而本实验中的泥炭地地表水和泥炭藓沥出液分别经雨水和蒸馏水稀释, 降低了化感物质浓度.高浓度的化感物质虽然抑制孢子的萌发, 但很可能使其保持生活力, 而较低浓度的化感物质抑制微生物活性(
Verhoeven & Liefveld, 1997 ), 使孢子保持较高的萌发力. ...
The Methuselah of plant diaspores:
Sphagnum spores can survive in nature for centuries
6
2017b
... 因在植物种群的持续更新(
Saatkamp et al ., 2014 )和遗传多样性维持(
Tellier, 2019 )方面发挥重要作用, 植物持久有性繁殖体库(种子库和孢子库)备受****们关注.泥炭藓(
Sphagnum spp.)是北方泥炭地的主要造炭植物(
Turetsky, 2003 ), 以泥炭藓为优势植物的泥炭地生态系统, 长期积水, 氧和有效养分贫乏, 大量植物残体分解以及泥炭藓酸释放导致环境酸化, 次生代谢物质丰富, 泥炭地的这些特征性环境条件可能是诸多超长寿命的植物繁殖体(
Clymo & Duckett, 1986 ;
Shen-Miller et al ., 1995 ;
Sundberg & Rydin, 2000 ;
Bu et al ., 2017b )在此被发现的重要原因. ...
... 植物有性繁殖体的持久性因物种和环境条件的不同差异较大.由微生境差异引起的水位、pH、养分、离子浓度等环境因子的差异, 构成了泥炭地的环境梯度(
Wheeler & Proctor, 2000 ), 这种环境梯度均可能影响泥炭藓孢子的生活力、萌发和建植.如
Bu等(2017b) 在长白山哈泥泥炭地的研究发现, 水位较高的丘间生境中泥炭藓孢子的寿命更长.
Feng等(2017) 为期一年的埋藏实验发现, 间歇性淹水生境比从不淹水和持续淹水生境, 更适宜泥炭藓孢子萌发力的保持.泥炭藓孢子可以在添加动物粪便、桦树叶或泥炭藓灰分的泥炭中萌发(
Sundberg & Rydin, 2002 ), 表明泥炭地中养分较高的生境可能不利于泥炭藓持久孢子库的形成和稳定.然而, 尚无直接实验证据表明泥炭地的环境因子对地层中泥炭藓孢子寿命的影响. ...
... 对大部分植物种子而言, 环境中的氧含量直接影响其生活力和寿命.如4种十字花科植物的种子在不同气体和水环境中保存36年, 结果发现充满CO
2 的气体环境(缺氧)能提高种子的寿命(
González- Benito et al ., 2011 ).泥炭藓每年均产生大量孢子散布于泥炭地中, 随着泥炭的逐年累积, 在地层中有明显的垂直分布格局(
Bu et al ., 2017b ).在泥炭地地层中, 氧含量随着深度增加而减少(
Jauhiainen, 1998 ), 可能直接影响泥炭藓孢子的新陈代谢和老化速率, 进而影响其持久性.如
Sundberg和Rydin (2000)) 通过研究自然生境氧含量差异对泥炭藓孢子生活力持久性的影响, 发现孢子在少氧的丘间生境埋藏比在有氧的藓丘生境更快失去生活力.然而,
Feng等(2017) 模拟自然生境泥炭藓孢子不同埋藏深度的实验, 在埋藏1年后, 表层泥炭中(多氧环境)的孢子持久性高于埋藏在深层泥炭(少氧环境)的孢子.因此, 环境中氧含量对泥炭藓孢子持久性具体有何影响尚无一致结论. ...
... 成熟的泥炭藓孢子散布于潮湿的苔藓基质或积水的丘间生境中, 比散布于缺少化感物质的生境中能更好地维持萌发力.这与
Bu等(2017b) 对长白山哈泥泥炭地泥炭藓孢子寿命研究的结果一致, 即在少氧的丘间生境中泥炭藓孢子寿命更长.尽管通径分析结果显示, 总酚浓度并不是影响孢子持久性的主要因素, 但对两种泥炭藓孢子的持久性均表现为正作用, 并且通过Ca
2+ 浓度对其也有较大的间接正作用.此外, K
+ 和Na
+ 浓度通过总酚浓度对孢子的持久性也有间接的正作用.研究表明土荆芥(
Chenopodium ambrosioides )分泌的化感物质会增加受体植物细胞质中的Ca
2+ 累积(
Li et al ., 2018a ).环境中Na
+ 和K
+ 浓度增加, 促进
Salvia officinalis 分泌更多的酚类化感物质(
Taarit et al ., 2012 ).由此可推测, 若环境中或泥炭藓体内的Na
+ 和K
+ 浓度增加, 可能促进化感物质的分泌, 进而有助于泥炭藓孢子维持生活力和萌发力.本研究的泥炭地地表水中, Ca
2+ 和总酚浓度分别显著高于泥炭藓沥出液和超纯水, 泥炭地地表水中的高浓度酚类化合物促进Ca
2+ 的聚集, 虽抑制泥炭藓孢子的萌发(
刘礼洁等, 2019 ), 但却利于其保持生活力形成持久孢子库.因此也可解释丘间淹水生境埋藏的泥炭藓孢子寿命大于藓丘不淹水生境(
Bu et al ., 2017b ). ...
... ), 但却利于其保持生活力形成持久孢子库.因此也可解释丘间淹水生境埋藏的泥炭藓孢子寿命大于藓丘不淹水生境(
Bu et al ., 2017b ). ...
... 在北方泥炭地, 成熟的泥炭藓孢子可能散布于不同的生境, 如干旱或潮湿的泥炭藓丛、间歇性淹水的丘间生境和持续淹水的低洼生境.在不淹水的泥炭藓地被基质内, 由于较高浓度的化感物质抑制孢子的萌发, 但却使其在短期内保持生活力.在间歇性淹水生境, 孢子保持较高的萌发力, 待到适宜的条件, 如光照、养分和温度, 即可萌发(
Feng et al ., 2017 ).在持续淹水生境, 泥炭藓孢子不能萌发, 但可长期保持较高的生活力(
Sundberg & Rydin, 2000 ).本研究表明, 溶解氧为影响泥炭地泥炭藓孢子持久性的主要因子, 同时也是限制因子, 化感物质浓度为正作用因子, 养分元素N和P浓度为负作用因子, 很可能与微生物多样性和活性增加有关(
Pinsonneault et al ., 2016 ).因此, 少氧、淹水、养分较贫乏的泥炭藓泥炭地, 是泥炭藓孢子长期保持生活力的理想场所, 在泥炭地发现泥炭藓长期持久孢子库(
Bu et al ., 2017b )及其他植物的超长寿命繁殖体也归因于此. ...
Regeneration of
Sphagnum 1
1986
... 因在植物种群的持续更新(
Saatkamp et al ., 2014 )和遗传多样性维持(
Tellier, 2019 )方面发挥重要作用, 植物持久有性繁殖体库(种子库和孢子库)备受****们关注.泥炭藓(
Sphagnum spp.)是北方泥炭地的主要造炭植物(
Turetsky, 2003 ), 以泥炭藓为优势植物的泥炭地生态系统, 长期积水, 氧和有效养分贫乏, 大量植物残体分解以及泥炭藓酸释放导致环境酸化, 次生代谢物质丰富, 泥炭地的这些特征性环境条件可能是诸多超长寿命的植物繁殖体(
Clymo & Duckett, 1986 ;
Shen-Miller et al ., 1995 ;
Sundberg & Rydin, 2000 ;
Bu et al ., 2017b )在此被发现的重要原因. ...
使用SPSS线性回归实现通径分析的方法
1
2010
... 所有数据用SPSS 19.0统计分析软件进行分析.对因变量孢子
GCR 数据进行正态检验, 服从正态分布可直接进行回归分析和通径分析(
杜家菊和陈志伟, 2010 ), 不符合正态分布则对数据进行标准化处理.双因素方差分析检验保存液类型、充气速率及二者交互作用对孢子持久性的影响.单因素方差分析比较不同处理中的
DO 、pH值、
Eh 及主要元素和总酚浓度的差异.单因素方差分析和
t 检验分组比较不同处理对孢子持久性影响的差异(分组如下: 超纯水不充气、泥炭地地表水不充气和泥炭藓沥出液不充气, 超纯水不充气、低速率充气和高速率充气, 泥炭地地表水不充气、低速率充气和高速率充气, 泥炭藓沥出液不充气、低速率充气和高速率充气, 空气和超纯水不充气).多重比较采用最小显著差异法(
LSD )检验(
α = 0.05).通径分析方法比较各因子对孢子持久性的直接和间接影响的相对重要性. ...
使用SPSS线性回归实现通径分析的方法
1
2010
... 所有数据用SPSS 19.0统计分析软件进行分析.对因变量孢子
GCR 数据进行正态检验, 服从正态分布可直接进行回归分析和通径分析(
杜家菊和陈志伟, 2010 ), 不符合正态分布则对数据进行标准化处理.双因素方差分析检验保存液类型、充气速率及二者交互作用对孢子持久性的影响.单因素方差分析比较不同处理中的
DO 、pH值、
Eh 及主要元素和总酚浓度的差异.单因素方差分析和
t 检验分组比较不同处理对孢子持久性影响的差异(分组如下: 超纯水不充气、泥炭地地表水不充气和泥炭藓沥出液不充气, 超纯水不充气、低速率充气和高速率充气, 泥炭地地表水不充气、低速率充气和高速率充气, 泥炭藓沥出液不充气、低速率充气和高速率充气, 空气和超纯水不充气).多重比较采用最小显著差异法(
LSD )检验(
α = 0.05).通径分析方法比较各因子对孢子持久性的直接和间接影响的相对重要性. ...
Continuous waterlogging may not facilitate germinability maintenance of
Sphagnum spores
4
2017
... 植物有性繁殖体的持久性因物种和环境条件的不同差异较大.由微生境差异引起的水位、pH、养分、离子浓度等环境因子的差异, 构成了泥炭地的环境梯度(
Wheeler & Proctor, 2000 ), 这种环境梯度均可能影响泥炭藓孢子的生活力、萌发和建植.如
Bu等(2017b) 在长白山哈泥泥炭地的研究发现, 水位较高的丘间生境中泥炭藓孢子的寿命更长.
Feng等(2017) 为期一年的埋藏实验发现, 间歇性淹水生境比从不淹水和持续淹水生境, 更适宜泥炭藓孢子萌发力的保持.泥炭藓孢子可以在添加动物粪便、桦树叶或泥炭藓灰分的泥炭中萌发(
Sundberg & Rydin, 2002 ), 表明泥炭地中养分较高的生境可能不利于泥炭藓持久孢子库的形成和稳定.然而, 尚无直接实验证据表明泥炭地的环境因子对地层中泥炭藓孢子寿命的影响. ...
... 对大部分植物种子而言, 环境中的氧含量直接影响其生活力和寿命.如4种十字花科植物的种子在不同气体和水环境中保存36年, 结果发现充满CO
2 的气体环境(缺氧)能提高种子的寿命(
González- Benito et al ., 2011 ).泥炭藓每年均产生大量孢子散布于泥炭地中, 随着泥炭的逐年累积, 在地层中有明显的垂直分布格局(
Bu et al ., 2017b ).在泥炭地地层中, 氧含量随着深度增加而减少(
Jauhiainen, 1998 ), 可能直接影响泥炭藓孢子的新陈代谢和老化速率, 进而影响其持久性.如
Sundberg和Rydin (2000)) 通过研究自然生境氧含量差异对泥炭藓孢子生活力持久性的影响, 发现孢子在少氧的丘间生境埋藏比在有氧的藓丘生境更快失去生活力.然而,
Feng等(2017) 模拟自然生境泥炭藓孢子不同埋藏深度的实验, 在埋藏1年后, 表层泥炭中(多氧环境)的孢子持久性高于埋藏在深层泥炭(少氧环境)的孢子.因此, 环境中氧含量对泥炭藓孢子持久性具体有何影响尚无一致结论. ...
... 本文基于泥炭藓存在超长持久孢子库的现状, 针对其形成机制存在的知识空白, 考虑到泥炭藓孢子散布生境的条件, 通过不同保存液处理, 模拟泥炭藓孢子在自然状态中可能散布的生境(有无水淹和有无化感物质), 并通过不同程度充气处理模拟环境的氧含量(少氧和多氧), 旨在回答: 1)是否化感物质利于泥炭藓孢子萌发力的保持, 是否总酚浓度直接影响孢子持久性? 2)基于
Feng等(2017) 的结论, 是否少氧环境更利于泥炭藓孢子萌发力的保持, 孢子持久性是否随着溶解氧浓度增加而降低? 研究结果有助于深入理解泥炭地泥炭藓持久孢子库的形成机制, 为退化泥炭地泥炭藓地被恢复研究提供参考. ...
... 在北方泥炭地, 成熟的泥炭藓孢子可能散布于不同的生境, 如干旱或潮湿的泥炭藓丛、间歇性淹水的丘间生境和持续淹水的低洼生境.在不淹水的泥炭藓地被基质内, 由于较高浓度的化感物质抑制孢子的萌发, 但却使其在短期内保持生活力.在间歇性淹水生境, 孢子保持较高的萌发力, 待到适宜的条件, 如光照、养分和温度, 即可萌发(
Feng et al ., 2017 ).在持续淹水生境, 泥炭藓孢子不能萌发, 但可长期保持较高的生活力(
Sundberg & Rydin, 2000 ).本研究表明, 溶解氧为影响泥炭地泥炭藓孢子持久性的主要因子, 同时也是限制因子, 化感物质浓度为正作用因子, 养分元素N和P浓度为负作用因子, 很可能与微生物多样性和活性增加有关(
Pinsonneault et al ., 2016 ).因此, 少氧、淹水、养分较贫乏的泥炭藓泥炭地, 是泥炭藓孢子长期保持生活力的理想场所, 在泥炭地发现泥炭藓长期持久孢子库(
Bu et al ., 2017b )及其他植物的超长寿命繁殖体也归因于此. ...
Global patterns in belowground communities
1
2009
... N和P浓度是泥炭地植被生长的主要限制因子(
Aerts et al ., 1992 ).通过相关分析和通径分析结果可知, TN和TP浓度对两种泥炭藓孢子的持久性均表现为负作用, 尤其对喙叶泥炭藓孢子的持久性为直接的负作用.Ca
2+ 浓度对尖叶泥炭藓孢子的持久性有较大的间接负作用, 表明养分元素对泥炭藓孢子的持久性起到限制作用.
Boatman和Lark (1971) 研究认为泥炭藓孢子不能在野外条件下萌发建植是由于泥炭地养分的缺乏, 因此, 若雨养型泥炭地养分输入增加, 可能导致泥炭藓孢子不必要的萌发, 从而不利于泥炭藓孢子库的稳定.此外, 随着泥炭地中养分增加, 使原本养分较贫乏(
Song et al ., 2017 )的环境中微生物数量增加、活性增强(
Fierer et al ., 2009 ), 从而加速泥炭藓孢子的分解, 不利于其保持萌发力.因此可推测, 在全球变化大背景下, 大气氮沉降导致泥炭地养分输入增加, 将不利于泥炭藓持久孢子库的稳定. ...
Effect of the gaseous environment and water content on seed viability of four Brassicaceae species after 36 years storage
1
2011
... 对大部分植物种子而言, 环境中的氧含量直接影响其生活力和寿命.如4种十字花科植物的种子在不同气体和水环境中保存36年, 结果发现充满CO
2 的气体环境(缺氧)能提高种子的寿命(
González- Benito et al ., 2011 ).泥炭藓每年均产生大量孢子散布于泥炭地中, 随着泥炭的逐年累积, 在地层中有明显的垂直分布格局(
Bu et al ., 2017b ).在泥炭地地层中, 氧含量随着深度增加而减少(
Jauhiainen, 1998 ), 可能直接影响泥炭藓孢子的新陈代谢和老化速率, 进而影响其持久性.如
Sundberg和Rydin (2000)) 通过研究自然生境氧含量差异对泥炭藓孢子生活力持久性的影响, 发现孢子在少氧的丘间生境埋藏比在有氧的藓丘生境更快失去生活力.然而,
Feng等(2017) 模拟自然生境泥炭藓孢子不同埋藏深度的实验, 在埋藏1年后, 表层泥炭中(多氧环境)的孢子持久性高于埋藏在深层泥炭(少氧环境)的孢子.因此, 环境中氧含量对泥炭藓孢子持久性具体有何影响尚无一致结论. ...
The role of polyphenols in terrestrial ecosystem nutrient cycling
1
2000
... 充气显著增加了保存液的
DO, 却降低了孢子持久性.由通径分析结果可知,
DO 是影响两种泥炭藓孢子持久性的最主要的作用因子.然而, 由保存液类型和充气速率对孢子持久性的交互作用可知, 充气在超纯水中对孢子持久性无显著影响, 但在泥炭地地表水和泥炭藓沥出液中, 却能显著降低孢子持久性.表明在本研究中
DO 不会直接氧化泥炭藓孢子, 加快其老化速率.虽然实验中总酚的浓度并未在充气条件下发生显著变化, 但其组成复杂(
H?ttenschwiler & Vitousek, 2000 ), 某些关键组分被氧化, 如泥炭藓酸(
Abbott et al ., 2013 ), 化感物质组分发生改变, 使原本富含化感物质的泥炭地地表水和泥炭藓沥出液的抑菌性下降, 提高了好氧细菌的活性(
Ke et al ., 2015 ), 进而加速孢子的分解, 不利于其保持萌发力.此外, 由多元回归分析结果可知,
DO 对两种泥炭藓孢子持久性的影响分别为60.4%和16.2%, 表明还有其他因素对孢子持久性有影响, 但未被考虑进来.因此, 泥炭地环境因子对泥炭藓孢子持久性的影响较为复杂, 若要深入研究泥炭地泥炭藓持久孢子库的形成机理, 需要考虑更多的影响因素, 例如环境温度、光照强度、化感物质各组分浓度和微生物活性的影响等.尤其在全球变暖的大背景下, 温度升高打破地层泥炭藓孢子休眠, 增加不必要的萌发, 减少泥炭藓孢子库储存(
Mcletchie, 1999 ). ...
Seed and spore banks of two boreal mires
1
1998
... 对大部分植物种子而言, 环境中的氧含量直接影响其生活力和寿命.如4种十字花科植物的种子在不同气体和水环境中保存36年, 结果发现充满CO
2 的气体环境(缺氧)能提高种子的寿命(
González- Benito et al ., 2011 ).泥炭藓每年均产生大量孢子散布于泥炭地中, 随着泥炭的逐年累积, 在地层中有明显的垂直分布格局(
Bu et al ., 2017b ).在泥炭地地层中, 氧含量随着深度增加而减少(
Jauhiainen, 1998 ), 可能直接影响泥炭藓孢子的新陈代谢和老化速率, 进而影响其持久性.如
Sundberg和Rydin (2000)) 通过研究自然生境氧含量差异对泥炭藓孢子生活力持久性的影响, 发现孢子在少氧的丘间生境埋藏比在有氧的藓丘生境更快失去生活力.然而,
Feng等(2017) 模拟自然生境泥炭藓孢子不同埋藏深度的实验, 在埋藏1年后, 表层泥炭中(多氧环境)的孢子持久性高于埋藏在深层泥炭(少氧环境)的孢子.因此, 环境中氧含量对泥炭藓孢子持久性具体有何影响尚无一致结论. ...
High oxygen concentration increases the abundance and activity of bacterial rather than archaeal nitrifiers in rice field soil
1
2015
... 充气显著增加了保存液的
DO, 却降低了孢子持久性.由通径分析结果可知,
DO 是影响两种泥炭藓孢子持久性的最主要的作用因子.然而, 由保存液类型和充气速率对孢子持久性的交互作用可知, 充气在超纯水中对孢子持久性无显著影响, 但在泥炭地地表水和泥炭藓沥出液中, 却能显著降低孢子持久性.表明在本研究中
DO 不会直接氧化泥炭藓孢子, 加快其老化速率.虽然实验中总酚的浓度并未在充气条件下发生显著变化, 但其组成复杂(
H?ttenschwiler & Vitousek, 2000 ), 某些关键组分被氧化, 如泥炭藓酸(
Abbott et al ., 2013 ), 化感物质组分发生改变, 使原本富含化感物质的泥炭地地表水和泥炭藓沥出液的抑菌性下降, 提高了好氧细菌的活性(
Ke et al ., 2015 ), 进而加速孢子的分解, 不利于其保持萌发力.此外, 由多元回归分析结果可知,
DO 对两种泥炭藓孢子持久性的影响分别为60.4%和16.2%, 表明还有其他因素对孢子持久性有影响, 但未被考虑进来.因此, 泥炭地环境因子对泥炭藓孢子持久性的影响较为复杂, 若要深入研究泥炭地泥炭藓持久孢子库的形成机理, 需要考虑更多的影响因素, 例如环境温度、光照强度、化感物质各组分浓度和微生物活性的影响等.尤其在全球变暖的大背景下, 温度升高打破地层泥炭藓孢子休眠, 增加不必要的萌发, 减少泥炭藓孢子库储存(
Mcletchie, 1999 ). ...
Volatile allelochemicals of
Chenopodium ambrosioides L. induced mitochondrion-mediated Ca
2+ -dependent and caspase-?dependent apoptosis signaling pathways in receptor plant cells
2018a
Contribution of biotic and abiotic factors in the natural attenuation of sulfamethoxazole: A path analysis approach
1
2018b
... 本研究在实验室内模拟条件下完成, 尽管室内条件与自然环境有差异, 但实验所用的泥炭藓孢子、泥炭地地表水和制备沥出液的泥炭藓均采集于野外, 并且充气处理采用注入空气而不是纯氧, 所有实验条件最大限度地接近自然环境, 因此, 研究中各因子对泥炭藓孢子持久性的影响对于深刻认识泥炭藓持久孢子库的形成过程与机制有重要意义.在本试验的各因子中, 尖叶泥炭藓孢子持久性与
DO 和
Eh 显著相关, 其余指标与尖叶泥炭藓孢子持久性未达显著相关水平.喙叶泥炭藓孢子持久性与
DO 、TN、TP、K
+ 、Na
+ 浓度呈显著相关关系, 其余指标与喙叶泥炭藓孢子持久性未达显著相关水平.很多指标之间也存在显著相关关系.结合通径分析结果可知, 部分环境因子不仅直接影响孢子的持久性, 而且通过相互作用, 间接影响孢子持久性.因此, 简单的相关分析不能判定环境因子对泥炭藓孢子持久性的影响, 通过通径分析可区分各因子作用的大小(
Li et al ., 2018b ), 估计各因子的相对重要性, 提取关键影响因子. ...
沙尘沉降可能阻滞泥炭地植被的自发演替
1
2019
... 成熟的泥炭藓孢子散布于潮湿的苔藓基质或积水的丘间生境中, 比散布于缺少化感物质的生境中能更好地维持萌发力.这与
Bu等(2017b) 对长白山哈泥泥炭地泥炭藓孢子寿命研究的结果一致, 即在少氧的丘间生境中泥炭藓孢子寿命更长.尽管通径分析结果显示, 总酚浓度并不是影响孢子持久性的主要因素, 但对两种泥炭藓孢子的持久性均表现为正作用, 并且通过Ca
2+ 浓度对其也有较大的间接正作用.此外, K
+ 和Na
+ 浓度通过总酚浓度对孢子的持久性也有间接的正作用.研究表明土荆芥(
Chenopodium ambrosioides )分泌的化感物质会增加受体植物细胞质中的Ca
2+ 累积(
Li et al ., 2018a ).环境中Na
+ 和K
+ 浓度增加, 促进
Salvia officinalis 分泌更多的酚类化感物质(
Taarit et al ., 2012 ).由此可推测, 若环境中或泥炭藓体内的Na
+ 和K
+ 浓度增加, 可能促进化感物质的分泌, 进而有助于泥炭藓孢子维持生活力和萌发力.本研究的泥炭地地表水中, Ca
2+ 和总酚浓度分别显著高于泥炭藓沥出液和超纯水, 泥炭地地表水中的高浓度酚类化合物促进Ca
2+ 的聚集, 虽抑制泥炭藓孢子的萌发(
刘礼洁等, 2019 ), 但却利于其保持生活力形成持久孢子库.因此也可解释丘间淹水生境埋藏的泥炭藓孢子寿命大于藓丘不淹水生境(
Bu et al ., 2017b ). ...
沙尘沉降可能阻滞泥炭地植被的自发演替
1
2019
... 成熟的泥炭藓孢子散布于潮湿的苔藓基质或积水的丘间生境中, 比散布于缺少化感物质的生境中能更好地维持萌发力.这与
Bu等(2017b) 对长白山哈泥泥炭地泥炭藓孢子寿命研究的结果一致, 即在少氧的丘间生境中泥炭藓孢子寿命更长.尽管通径分析结果显示, 总酚浓度并不是影响孢子持久性的主要因素, 但对两种泥炭藓孢子的持久性均表现为正作用, 并且通过Ca
2+ 浓度对其也有较大的间接正作用.此外, K
+ 和Na
+ 浓度通过总酚浓度对孢子的持久性也有间接的正作用.研究表明土荆芥(
Chenopodium ambrosioides )分泌的化感物质会增加受体植物细胞质中的Ca
2+ 累积(
Li et al ., 2018a ).环境中Na
+ 和K
+ 浓度增加, 促进
Salvia officinalis 分泌更多的酚类化感物质(
Taarit et al ., 2012 ).由此可推测, 若环境中或泥炭藓体内的Na
+ 和K
+ 浓度增加, 可能促进化感物质的分泌, 进而有助于泥炭藓孢子维持生活力和萌发力.本研究的泥炭地地表水中, Ca
2+ 和总酚浓度分别显著高于泥炭藓沥出液和超纯水, 泥炭地地表水中的高浓度酚类化合物促进Ca
2+ 的聚集, 虽抑制泥炭藓孢子的萌发(
刘礼洁等, 2019 ), 但却利于其保持生活力形成持久孢子库.因此也可解释丘间淹水生境埋藏的泥炭藓孢子寿命大于藓丘不淹水生境(
Bu et al ., 2017b ). ...
Dormancy/Nondormancy cycles in spores of the liverwort
Sphaerocarpos texanus 1
1999
... 充气显著增加了保存液的
DO, 却降低了孢子持久性.由通径分析结果可知,
DO 是影响两种泥炭藓孢子持久性的最主要的作用因子.然而, 由保存液类型和充气速率对孢子持久性的交互作用可知, 充气在超纯水中对孢子持久性无显著影响, 但在泥炭地地表水和泥炭藓沥出液中, 却能显著降低孢子持久性.表明在本研究中
DO 不会直接氧化泥炭藓孢子, 加快其老化速率.虽然实验中总酚的浓度并未在充气条件下发生显著变化, 但其组成复杂(
H?ttenschwiler & Vitousek, 2000 ), 某些关键组分被氧化, 如泥炭藓酸(
Abbott et al ., 2013 ), 化感物质组分发生改变, 使原本富含化感物质的泥炭地地表水和泥炭藓沥出液的抑菌性下降, 提高了好氧细菌的活性(
Ke et al ., 2015 ), 进而加速孢子的分解, 不利于其保持萌发力.此外, 由多元回归分析结果可知,
DO 对两种泥炭藓孢子持久性的影响分别为60.4%和16.2%, 表明还有其他因素对孢子持久性有影响, 但未被考虑进来.因此, 泥炭地环境因子对泥炭藓孢子持久性的影响较为复杂, 若要深入研究泥炭地泥炭藓持久孢子库的形成机理, 需要考虑更多的影响因素, 例如环境温度、光照强度、化感物质各组分浓度和微生物活性的影响等.尤其在全球变暖的大背景下, 温度升高打破地层泥炭藓孢子休眠, 增加不必要的萌发, 减少泥炭藓孢子库储存(
Mcletchie, 1999 ). ...
Bryophytes display allelopathic interactions with tree species in native forest ecosystems
1
2011
... 苔藓植物通常生产较维管植物更为丰富的次生代谢物质, 其强烈的化感作用可以抑制微生物(
Montenegro et al ., 2009 )和维管植物种子的萌发和幼苗的生长(
Whitehead et al ., 2018 ), 如18种苔藓植物化感提取液均抑制
Fuchsia excorticata 种子的萌发和胚根的生长(
Michel et al ., 2011 ).苔藓地被基质的化感作用可能在泥炭藓持久孢子库的形成过程中产生关键作用, 然而受到的关注较少.已有研究发现, 金发藓(
Polytrichum strictum )地被基质的化感效应强于泥炭藓(
Sphagnum palustre )和中位泥炭藓(
Sphagnum magellanicum ), 可强烈抑制以上3种苔藓孢子的萌发(
Bu et al ., 2017a ).然而, 不萌发并不代表失去生活力, 这可能是植物分摊风险的“两头下注”策略(
Ooi et al ., 2009 ), 孢子萌发受到抑制, 却进入泥炭地地层中保持生活力, 是持久有性繁殖体库形成的前提. ...
Influences of mature plants and desiccation on germination of spores and gametophyticfragments of
Tortula 1
1988
... 苔藓植物作为从水生向陆生过渡的植物类群, 在进化过程中已具备了耐受干旱的能力(
Proctor et al ., 2007 ).因此, 两种泥炭藓孢子经空气和超纯水保存2个月后, 持久性无显著差异, 但空气保存后的萌发速度显著低于淹水保存, 表明孢子散布于淹水生境后, 时刻保持着快速萌发的能力.然而, 泥炭地正受到全球变化的威胁, 如全球变暖增加干旱频率, 导致泥炭地水位下降(
袁敏等, 2015 ), 很可能不利于地层中泥炭藓孢子的持久性.尖叶泥炭藓孢子经泥炭地地表水和泥炭藓沥出液保存后, 持久性显著高于超纯水保存, 表明纯水并不是决定孢子持久性的因素, 而水中的离子和化感物质可能是影响孢子持久性的关键因子.本研究结果与
Bu等(2017a) 苔藓化感作用减弱孢子萌发力的结论看似矛盾, 可能原因首先是实验物种的差异, 本文实验物种为尖叶泥炭藓和喙叶泥炭藓孢子, 而
Bu等(2017a) 实验物种为金发藓、泥炭藓和中位泥炭藓孢子, 本研究中尖叶泥炭藓孢子经3种保存液不充气处理后, 持久性之间有显著差异, 而喙叶泥炭藓孢子无显著差异, 可知不同种泥炭藓孢子对化感效应的响应不同.其次,
Bu等(2017a) 的研究中, 泥炭藓孢子经野外环境埋藏一个生长季, 可能遭受到紫外辐射和干旱等不利条件(
van Zanten, 1978 ), 导致孢子迅速老化, 持久性显著下降.此外, 由于其野外埋藏实验的苔藓基质化感物质浓度较高(
Mishler & Newton, 1988 ), 而本实验中的泥炭地地表水和泥炭藓沥出液分别经雨水和蒸馏水稀释, 降低了化感物质浓度.高浓度的化感物质虽然抑制孢子的萌发, 但很可能使其保持生活力, 而较低浓度的化感物质抑制微生物活性(
Verhoeven & Liefveld, 1997 ), 使孢子保持较高的萌发力. ...
Biological properties of the Chilean native moss
Sphagnum magellanicum 1
2009
... 苔藓植物通常生产较维管植物更为丰富的次生代谢物质, 其强烈的化感作用可以抑制微生物(
Montenegro et al ., 2009 )和维管植物种子的萌发和幼苗的生长(
Whitehead et al ., 2018 ), 如18种苔藓植物化感提取液均抑制
Fuchsia excorticata 种子的萌发和胚根的生长(
Michel et al ., 2011 ).苔藓地被基质的化感作用可能在泥炭藓持久孢子库的形成过程中产生关键作用, 然而受到的关注较少.已有研究发现, 金发藓(
Polytrichum strictum )地被基质的化感效应强于泥炭藓(
Sphagnum palustre )和中位泥炭藓(
Sphagnum magellanicum ), 可强烈抑制以上3种苔藓孢子的萌发(
Bu et al ., 2017a ).然而, 不萌发并不代表失去生活力, 这可能是植物分摊风险的“两头下注”策略(
Ooi et al ., 2009 ), 孢子萌发受到抑制, 却进入泥炭地地层中保持生活力, 是持久有性繁殖体库形成的前提. ...
Climate change and bet-hedging: Interactions between increased soil temperatures and seed bank persistence
1
2009
... 苔藓植物通常生产较维管植物更为丰富的次生代谢物质, 其强烈的化感作用可以抑制微生物(
Montenegro et al ., 2009 )和维管植物种子的萌发和幼苗的生长(
Whitehead et al ., 2018 ), 如18种苔藓植物化感提取液均抑制
Fuchsia excorticata 种子的萌发和胚根的生长(
Michel et al ., 2011 ).苔藓地被基质的化感作用可能在泥炭藓持久孢子库的形成过程中产生关键作用, 然而受到的关注较少.已有研究发现, 金发藓(
Polytrichum strictum )地被基质的化感效应强于泥炭藓(
Sphagnum palustre )和中位泥炭藓(
Sphagnum magellanicum ), 可强烈抑制以上3种苔藓孢子的萌发(
Bu et al ., 2017a ).然而, 不萌发并不代表失去生活力, 这可能是植物分摊风险的“两头下注”策略(
Ooi et al ., 2009 ), 孢子萌发受到抑制, 却进入泥炭地地层中保持生活力, 是持久有性繁殖体库形成的前提. ...
Effects of long-term fertilization on peat stoichiometry and associated microbial enzyme activity in an ombrotrophic bog
1
2016
... 在北方泥炭地, 成熟的泥炭藓孢子可能散布于不同的生境, 如干旱或潮湿的泥炭藓丛、间歇性淹水的丘间生境和持续淹水的低洼生境.在不淹水的泥炭藓地被基质内, 由于较高浓度的化感物质抑制孢子的萌发, 但却使其在短期内保持生活力.在间歇性淹水生境, 孢子保持较高的萌发力, 待到适宜的条件, 如光照、养分和温度, 即可萌发(
Feng et al ., 2017 ).在持续淹水生境, 泥炭藓孢子不能萌发, 但可长期保持较高的生活力(
Sundberg & Rydin, 2000 ).本研究表明, 溶解氧为影响泥炭地泥炭藓孢子持久性的主要因子, 同时也是限制因子, 化感物质浓度为正作用因子, 养分元素N和P浓度为负作用因子, 很可能与微生物多样性和活性增加有关(
Pinsonneault et al ., 2016 ).因此, 少氧、淹水、养分较贫乏的泥炭藓泥炭地, 是泥炭藓孢子长期保持生活力的理想场所, 在泥炭地发现泥炭藓长期持久孢子库(
Bu et al ., 2017b )及其他植物的超长寿命繁殖体也归因于此. ...
Desiccation-tolerance in bryophytes: A review
1
2007
... 苔藓植物作为从水生向陆生过渡的植物类群, 在进化过程中已具备了耐受干旱的能力(
Proctor et al ., 2007 ).因此, 两种泥炭藓孢子经空气和超纯水保存2个月后, 持久性无显著差异, 但空气保存后的萌发速度显著低于淹水保存, 表明孢子散布于淹水生境后, 时刻保持着快速萌发的能力.然而, 泥炭地正受到全球变化的威胁, 如全球变暖增加干旱频率, 导致泥炭地水位下降(
袁敏等, 2015 ), 很可能不利于地层中泥炭藓孢子的持久性.尖叶泥炭藓孢子经泥炭地地表水和泥炭藓沥出液保存后, 持久性显著高于超纯水保存, 表明纯水并不是决定孢子持久性的因素, 而水中的离子和化感物质可能是影响孢子持久性的关键因子.本研究结果与
Bu等(2017a) 苔藓化感作用减弱孢子萌发力的结论看似矛盾, 可能原因首先是实验物种的差异, 本文实验物种为尖叶泥炭藓和喙叶泥炭藓孢子, 而
Bu等(2017a) 实验物种为金发藓、泥炭藓和中位泥炭藓孢子, 本研究中尖叶泥炭藓孢子经3种保存液不充气处理后, 持久性之间有显著差异, 而喙叶泥炭藓孢子无显著差异, 可知不同种泥炭藓孢子对化感效应的响应不同.其次,
Bu等(2017a) 的研究中, 泥炭藓孢子经野外环境埋藏一个生长季, 可能遭受到紫外辐射和干旱等不利条件(
van Zanten, 1978 ), 导致孢子迅速老化, 持久性显著下降.此外, 由于其野外埋藏实验的苔藓基质化感物质浓度较高(
Mishler & Newton, 1988 ), 而本实验中的泥炭地地表水和泥炭藓沥出液分别经雨水和蒸馏水稀释, 降低了化感物质浓度.高浓度的化感物质虽然抑制孢子的萌发, 但很可能使其保持生活力, 而较低浓度的化感物质抑制微生物活性(
Verhoeven & Liefveld, 1997 ), 使孢子保持较高的萌发力. ...
Sphagnum culture techniques. In: Glime JM ed. Methods in Bryology
1
1988
... 处理结束后, 将孢子接种到改良的Rudolph培养基(
Rudolph et al ., 1988 )上, 放入人工气候箱中进行培养.设置条件为白天16 h, 光照强度60 μmol·m
-2 ·s
-1 , 温度25 ℃; 夜晚8 h, 无光照, 温度20 ℃.在孢子培养的第3天进行第1次观察, 之后每4天观察一次孢子萌发情况. ...
The functional role of soil seed banks in natural communities
1
2014
... 因在植物种群的持续更新(
Saatkamp et al ., 2014 )和遗传多样性维持(
Tellier, 2019 )方面发挥重要作用, 植物持久有性繁殖体库(种子库和孢子库)备受****们关注.泥炭藓(
Sphagnum spp.)是北方泥炭地的主要造炭植物(
Turetsky, 2003 ), 以泥炭藓为优势植物的泥炭地生态系统, 长期积水, 氧和有效养分贫乏, 大量植物残体分解以及泥炭藓酸释放导致环境酸化, 次生代谢物质丰富, 泥炭地的这些特征性环境条件可能是诸多超长寿命的植物繁殖体(
Clymo & Duckett, 1986 ;
Shen-Miller et al ., 1995 ;
Sundberg & Rydin, 2000 ;
Bu et al ., 2017b )在此被发现的重要原因. ...
Exceptional seed longevity and robust growth: Ancient Sacred Lotus from China
1
1995
... 因在植物种群的持续更新(
Saatkamp et al ., 2014 )和遗传多样性维持(
Tellier, 2019 )方面发挥重要作用, 植物持久有性繁殖体库(种子库和孢子库)备受****们关注.泥炭藓(
Sphagnum spp.)是北方泥炭地的主要造炭植物(
Turetsky, 2003 ), 以泥炭藓为优势植物的泥炭地生态系统, 长期积水, 氧和有效养分贫乏, 大量植物残体分解以及泥炭藓酸释放导致环境酸化, 次生代谢物质丰富, 泥炭地的这些特征性环境条件可能是诸多超长寿命的植物繁殖体(
Clymo & Duckett, 1986 ;
Shen-Miller et al ., 1995 ;
Sundberg & Rydin, 2000 ;
Bu et al ., 2017b )在此被发现的重要原因. ...
Nitrogen additions affect litter quality and soil biochemical properties in a peatland of Northeast China
1
2017
... N和P浓度是泥炭地植被生长的主要限制因子(
Aerts et al ., 1992 ).通过相关分析和通径分析结果可知, TN和TP浓度对两种泥炭藓孢子的持久性均表现为负作用, 尤其对喙叶泥炭藓孢子的持久性为直接的负作用.Ca
2+ 浓度对尖叶泥炭藓孢子的持久性有较大的间接负作用, 表明养分元素对泥炭藓孢子的持久性起到限制作用.
Boatman和Lark (1971) 研究认为泥炭藓孢子不能在野外条件下萌发建植是由于泥炭地养分的缺乏, 因此, 若雨养型泥炭地养分输入增加, 可能导致泥炭藓孢子不必要的萌发, 从而不利于泥炭藓孢子库的稳定.此外, 随着泥炭地中养分增加, 使原本养分较贫乏(
Song et al ., 2017 )的环境中微生物数量增加、活性增强(
Fierer et al ., 2009 ), 从而加速泥炭藓孢子的分解, 不利于其保持萌发力.因此可推测, 在全球变化大背景下, 大气氮沉降导致泥炭地养分输入增加, 将不利于泥炭藓持久孢子库的稳定. ...
Experimental evidence for a persistent spore bank in
Sphagnum 3
2000
... 因在植物种群的持续更新(
Saatkamp et al ., 2014 )和遗传多样性维持(
Tellier, 2019 )方面发挥重要作用, 植物持久有性繁殖体库(种子库和孢子库)备受****们关注.泥炭藓(
Sphagnum spp.)是北方泥炭地的主要造炭植物(
Turetsky, 2003 ), 以泥炭藓为优势植物的泥炭地生态系统, 长期积水, 氧和有效养分贫乏, 大量植物残体分解以及泥炭藓酸释放导致环境酸化, 次生代谢物质丰富, 泥炭地的这些特征性环境条件可能是诸多超长寿命的植物繁殖体(
Clymo & Duckett, 1986 ;
Shen-Miller et al ., 1995 ;
Sundberg & Rydin, 2000 ;
Bu et al ., 2017b )在此被发现的重要原因. ...
... 对大部分植物种子而言, 环境中的氧含量直接影响其生活力和寿命.如4种十字花科植物的种子在不同气体和水环境中保存36年, 结果发现充满CO
2 的气体环境(缺氧)能提高种子的寿命(
González- Benito et al ., 2011 ).泥炭藓每年均产生大量孢子散布于泥炭地中, 随着泥炭的逐年累积, 在地层中有明显的垂直分布格局(
Bu et al ., 2017b ).在泥炭地地层中, 氧含量随着深度增加而减少(
Jauhiainen, 1998 ), 可能直接影响泥炭藓孢子的新陈代谢和老化速率, 进而影响其持久性.如
Sundberg和Rydin (2000)) 通过研究自然生境氧含量差异对泥炭藓孢子生活力持久性的影响, 发现孢子在少氧的丘间生境埋藏比在有氧的藓丘生境更快失去生活力.然而,
Feng等(2017) 模拟自然生境泥炭藓孢子不同埋藏深度的实验, 在埋藏1年后, 表层泥炭中(多氧环境)的孢子持久性高于埋藏在深层泥炭(少氧环境)的孢子.因此, 环境中氧含量对泥炭藓孢子持久性具体有何影响尚无一致结论. ...
... 在北方泥炭地, 成熟的泥炭藓孢子可能散布于不同的生境, 如干旱或潮湿的泥炭藓丛、间歇性淹水的丘间生境和持续淹水的低洼生境.在不淹水的泥炭藓地被基质内, 由于较高浓度的化感物质抑制孢子的萌发, 但却使其在短期内保持生活力.在间歇性淹水生境, 孢子保持较高的萌发力, 待到适宜的条件, 如光照、养分和温度, 即可萌发(
Feng et al ., 2017 ).在持续淹水生境, 泥炭藓孢子不能萌发, 但可长期保持较高的生活力(
Sundberg & Rydin, 2000 ).本研究表明, 溶解氧为影响泥炭地泥炭藓孢子持久性的主要因子, 同时也是限制因子, 化感物质浓度为正作用因子, 养分元素N和P浓度为负作用因子, 很可能与微生物多样性和活性增加有关(
Pinsonneault et al ., 2016 ).因此, 少氧、淹水、养分较贫乏的泥炭藓泥炭地, 是泥炭藓孢子长期保持生活力的理想场所, 在泥炭地发现泥炭藓长期持久孢子库(
Bu et al ., 2017b )及其他植物的超长寿命繁殖体也归因于此. ...
Habitat requirements for establishment of
Sphagnum from spores
1
2002
... 植物有性繁殖体的持久性因物种和环境条件的不同差异较大.由微生境差异引起的水位、pH、养分、离子浓度等环境因子的差异, 构成了泥炭地的环境梯度(
Wheeler & Proctor, 2000 ), 这种环境梯度均可能影响泥炭藓孢子的生活力、萌发和建植.如
Bu等(2017b) 在长白山哈泥泥炭地的研究发现, 水位较高的丘间生境中泥炭藓孢子的寿命更长.
Feng等(2017) 为期一年的埋藏实验发现, 间歇性淹水生境比从不淹水和持续淹水生境, 更适宜泥炭藓孢子萌发力的保持.泥炭藓孢子可以在添加动物粪便、桦树叶或泥炭藓灰分的泥炭中萌发(
Sundberg & Rydin, 2002 ), 表明泥炭地中养分较高的生境可能不利于泥炭藓持久孢子库的形成和稳定.然而, 尚无直接实验证据表明泥炭地的环境因子对地层中泥炭藓孢子寿命的影响. ...
Physiological changes, phenolic content and antioxidant activity of
Salvia officinalis L. grown under saline conditions
1
2012
... 成熟的泥炭藓孢子散布于潮湿的苔藓基质或积水的丘间生境中, 比散布于缺少化感物质的生境中能更好地维持萌发力.这与
Bu等(2017b) 对长白山哈泥泥炭地泥炭藓孢子寿命研究的结果一致, 即在少氧的丘间生境中泥炭藓孢子寿命更长.尽管通径分析结果显示, 总酚浓度并不是影响孢子持久性的主要因素, 但对两种泥炭藓孢子的持久性均表现为正作用, 并且通过Ca
2+ 浓度对其也有较大的间接正作用.此外, K
+ 和Na
+ 浓度通过总酚浓度对孢子的持久性也有间接的正作用.研究表明土荆芥(
Chenopodium ambrosioides )分泌的化感物质会增加受体植物细胞质中的Ca
2+ 累积(
Li et al ., 2018a ).环境中Na
+ 和K
+ 浓度增加, 促进
Salvia officinalis 分泌更多的酚类化感物质(
Taarit et al ., 2012 ).由此可推测, 若环境中或泥炭藓体内的Na
+ 和K
+ 浓度增加, 可能促进化感物质的分泌, 进而有助于泥炭藓孢子维持生活力和萌发力.本研究的泥炭地地表水中, Ca
2+ 和总酚浓度分别显著高于泥炭藓沥出液和超纯水, 泥炭地地表水中的高浓度酚类化合物促进Ca
2+ 的聚集, 虽抑制泥炭藓孢子的萌发(
刘礼洁等, 2019 ), 但却利于其保持生活力形成持久孢子库.因此也可解释丘间淹水生境埋藏的泥炭藓孢子寿命大于藓丘不淹水生境(
Bu et al ., 2017b ). ...
Persistent seed banking as eco-evolutionary determinant of plant nucleotide diversity: Novel population genetics insights
1
2019
... 因在植物种群的持续更新(
Saatkamp et al ., 2014 )和遗传多样性维持(
Tellier, 2019 )方面发挥重要作用, 植物持久有性繁殖体库(种子库和孢子库)备受****们关注.泥炭藓(
Sphagnum spp.)是北方泥炭地的主要造炭植物(
Turetsky, 2003 ), 以泥炭藓为优势植物的泥炭地生态系统, 长期积水, 氧和有效养分贫乏, 大量植物残体分解以及泥炭藓酸释放导致环境酸化, 次生代谢物质丰富, 泥炭地的这些特征性环境条件可能是诸多超长寿命的植物繁殖体(
Clymo & Duckett, 1986 ;
Shen-Miller et al ., 1995 ;
Sundberg & Rydin, 2000 ;
Bu et al ., 2017b )在此被发现的重要原因. ...
The role of bryophytes in carbon and nitrogen cycling
1
2003
... 因在植物种群的持续更新(
Saatkamp et al ., 2014 )和遗传多样性维持(
Tellier, 2019 )方面发挥重要作用, 植物持久有性繁殖体库(种子库和孢子库)备受****们关注.泥炭藓(
Sphagnum spp.)是北方泥炭地的主要造炭植物(
Turetsky, 2003 ), 以泥炭藓为优势植物的泥炭地生态系统, 长期积水, 氧和有效养分贫乏, 大量植物残体分解以及泥炭藓酸释放导致环境酸化, 次生代谢物质丰富, 泥炭地的这些特征性环境条件可能是诸多超长寿命的植物繁殖体(
Clymo & Duckett, 1986 ;
Shen-Miller et al ., 1995 ;
Sundberg & Rydin, 2000 ;
Bu et al ., 2017b )在此被发现的重要原因. ...
Experimental studies on trans-oceanic long-range dispersal of moss spores in the Southern Hemisphere
1
1978
... 苔藓植物作为从水生向陆生过渡的植物类群, 在进化过程中已具备了耐受干旱的能力(
Proctor et al ., 2007 ).因此, 两种泥炭藓孢子经空气和超纯水保存2个月后, 持久性无显著差异, 但空气保存后的萌发速度显著低于淹水保存, 表明孢子散布于淹水生境后, 时刻保持着快速萌发的能力.然而, 泥炭地正受到全球变化的威胁, 如全球变暖增加干旱频率, 导致泥炭地水位下降(
袁敏等, 2015 ), 很可能不利于地层中泥炭藓孢子的持久性.尖叶泥炭藓孢子经泥炭地地表水和泥炭藓沥出液保存后, 持久性显著高于超纯水保存, 表明纯水并不是决定孢子持久性的因素, 而水中的离子和化感物质可能是影响孢子持久性的关键因子.本研究结果与
Bu等(2017a) 苔藓化感作用减弱孢子萌发力的结论看似矛盾, 可能原因首先是实验物种的差异, 本文实验物种为尖叶泥炭藓和喙叶泥炭藓孢子, 而
Bu等(2017a) 实验物种为金发藓、泥炭藓和中位泥炭藓孢子, 本研究中尖叶泥炭藓孢子经3种保存液不充气处理后, 持久性之间有显著差异, 而喙叶泥炭藓孢子无显著差异, 可知不同种泥炭藓孢子对化感效应的响应不同.其次,
Bu等(2017a) 的研究中, 泥炭藓孢子经野外环境埋藏一个生长季, 可能遭受到紫外辐射和干旱等不利条件(
van Zanten, 1978 ), 导致孢子迅速老化, 持久性显著下降.此外, 由于其野外埋藏实验的苔藓基质化感物质浓度较高(
Mishler & Newton, 1988 ), 而本实验中的泥炭地地表水和泥炭藓沥出液分别经雨水和蒸馏水稀释, 降低了化感物质浓度.高浓度的化感物质虽然抑制孢子的萌发, 但很可能使其保持生活力, 而较低浓度的化感物质抑制微生物活性(
Verhoeven & Liefveld, 1997 ), 使孢子保持较高的萌发力. ...
The ecological significance of organochemical compounds in
Sphagnum 1
1997
... 苔藓植物作为从水生向陆生过渡的植物类群, 在进化过程中已具备了耐受干旱的能力(
Proctor et al ., 2007 ).因此, 两种泥炭藓孢子经空气和超纯水保存2个月后, 持久性无显著差异, 但空气保存后的萌发速度显著低于淹水保存, 表明孢子散布于淹水生境后, 时刻保持着快速萌发的能力.然而, 泥炭地正受到全球变化的威胁, 如全球变暖增加干旱频率, 导致泥炭地水位下降(
袁敏等, 2015 ), 很可能不利于地层中泥炭藓孢子的持久性.尖叶泥炭藓孢子经泥炭地地表水和泥炭藓沥出液保存后, 持久性显著高于超纯水保存, 表明纯水并不是决定孢子持久性的因素, 而水中的离子和化感物质可能是影响孢子持久性的关键因子.本研究结果与
Bu等(2017a) 苔藓化感作用减弱孢子萌发力的结论看似矛盾, 可能原因首先是实验物种的差异, 本文实验物种为尖叶泥炭藓和喙叶泥炭藓孢子, 而
Bu等(2017a) 实验物种为金发藓、泥炭藓和中位泥炭藓孢子, 本研究中尖叶泥炭藓孢子经3种保存液不充气处理后, 持久性之间有显著差异, 而喙叶泥炭藓孢子无显著差异, 可知不同种泥炭藓孢子对化感效应的响应不同.其次,
Bu等(2017a) 的研究中, 泥炭藓孢子经野外环境埋藏一个生长季, 可能遭受到紫外辐射和干旱等不利条件(
van Zanten, 1978 ), 导致孢子迅速老化, 持久性显著下降.此外, 由于其野外埋藏实验的苔藓基质化感物质浓度较高(
Mishler & Newton, 1988 ), 而本实验中的泥炭地地表水和泥炭藓沥出液分别经雨水和蒸馏水稀释, 降低了化感物质浓度.高浓度的化感物质虽然抑制孢子的萌发, 但很可能使其保持生活力, 而较低浓度的化感物质抑制微生物活性(
Verhoeven & Liefveld, 1997 ), 使孢子保持较高的萌发力. ...
Ecological gradients, subdivisions and terminology of north-west European mires
1
2000
... 植物有性繁殖体的持久性因物种和环境条件的不同差异较大.由微生境差异引起的水位、pH、养分、离子浓度等环境因子的差异, 构成了泥炭地的环境梯度(
Wheeler & Proctor, 2000 ), 这种环境梯度均可能影响泥炭藓孢子的生活力、萌发和建植.如
Bu等(2017b) 在长白山哈泥泥炭地的研究发现, 水位较高的丘间生境中泥炭藓孢子的寿命更长.
Feng等(2017) 为期一年的埋藏实验发现, 间歇性淹水生境比从不淹水和持续淹水生境, 更适宜泥炭藓孢子萌发力的保持.泥炭藓孢子可以在添加动物粪便、桦树叶或泥炭藓灰分的泥炭中萌发(
Sundberg & Rydin, 2002 ), 表明泥炭地中养分较高的生境可能不利于泥炭藓持久孢子库的形成和稳定.然而, 尚无直接实验证据表明泥炭地的环境因子对地层中泥炭藓孢子寿命的影响. ...
Allelopathy in bryophytes—A review
1
2018
... 苔藓植物通常生产较维管植物更为丰富的次生代谢物质, 其强烈的化感作用可以抑制微生物(
Montenegro et al ., 2009 )和维管植物种子的萌发和幼苗的生长(
Whitehead et al ., 2018 ), 如18种苔藓植物化感提取液均抑制
Fuchsia excorticata 种子的萌发和胚根的生长(
Michel et al ., 2011 ).苔藓地被基质的化感作用可能在泥炭藓持久孢子库的形成过程中产生关键作用, 然而受到的关注较少.已有研究发现, 金发藓(
Polytrichum strictum )地被基质的化感效应强于泥炭藓(
Sphagnum palustre )和中位泥炭藓(
Sphagnum magellanicum ), 可强烈抑制以上3种苔藓孢子的萌发(
Bu et al ., 2017a ).然而, 不萌发并不代表失去生活力, 这可能是植物分摊风险的“两头下注”策略(
Ooi et al ., 2009 ), 孢子萌发受到抑制, 却进入泥炭地地层中保持生活力, 是持久有性繁殖体库形成的前提. ...
Evaluation of methods for measuring polyphenols in conifer foliage
1
2000
... 实验处理期间, 采用便携式多参数水质分析仪(HACH-HQ30d, Hach Company, Loveland, USA)测定不同处理的溶解氧浓度(
DO )、pH值和氧化还原电位(
Eh ).福林酚法(
Yu & Dahlgren, 2000 )测定超纯水、泥炭地地表水和泥炭藓沥出液的总酚浓度.使用流动元素分析仪(Smartchem 140, Westco Scientifiic, Brookfield, USA)测定上述3种保存液的总氮(TN)和总磷(TP)含量.使用火焰原子吸收分光光度计(Spectr AA 220 FS, Mulgrave, Victoria, Australia)测定上述3种保存液的K
+ 、Ca
2+ 、Na
+ 、Mg
2+ 浓度. ...
水位与光强变化对尖叶泥炭藓孢蒴生产动态的影响
1
2015
... 苔藓植物作为从水生向陆生过渡的植物类群, 在进化过程中已具备了耐受干旱的能力(
Proctor et al ., 2007 ).因此, 两种泥炭藓孢子经空气和超纯水保存2个月后, 持久性无显著差异, 但空气保存后的萌发速度显著低于淹水保存, 表明孢子散布于淹水生境后, 时刻保持着快速萌发的能力.然而, 泥炭地正受到全球变化的威胁, 如全球变暖增加干旱频率, 导致泥炭地水位下降(
袁敏等, 2015 ), 很可能不利于地层中泥炭藓孢子的持久性.尖叶泥炭藓孢子经泥炭地地表水和泥炭藓沥出液保存后, 持久性显著高于超纯水保存, 表明纯水并不是决定孢子持久性的因素, 而水中的离子和化感物质可能是影响孢子持久性的关键因子.本研究结果与
Bu等(2017a) 苔藓化感作用减弱孢子萌发力的结论看似矛盾, 可能原因首先是实验物种的差异, 本文实验物种为尖叶泥炭藓和喙叶泥炭藓孢子, 而
Bu等(2017a) 实验物种为金发藓、泥炭藓和中位泥炭藓孢子, 本研究中尖叶泥炭藓孢子经3种保存液不充气处理后, 持久性之间有显著差异, 而喙叶泥炭藓孢子无显著差异, 可知不同种泥炭藓孢子对化感效应的响应不同.其次,
Bu等(2017a) 的研究中, 泥炭藓孢子经野外环境埋藏一个生长季, 可能遭受到紫外辐射和干旱等不利条件(
van Zanten, 1978 ), 导致孢子迅速老化, 持久性显著下降.此外, 由于其野外埋藏实验的苔藓基质化感物质浓度较高(
Mishler & Newton, 1988 ), 而本实验中的泥炭地地表水和泥炭藓沥出液分别经雨水和蒸馏水稀释, 降低了化感物质浓度.高浓度的化感物质虽然抑制孢子的萌发, 但很可能使其保持生活力, 而较低浓度的化感物质抑制微生物活性(
Verhoeven & Liefveld, 1997 ), 使孢子保持较高的萌发力. ...
水位与光强变化对尖叶泥炭藓孢蒴生产动态的影响
1
2015
... 苔藓植物作为从水生向陆生过渡的植物类群, 在进化过程中已具备了耐受干旱的能力(
Proctor et al ., 2007 ).因此, 两种泥炭藓孢子经空气和超纯水保存2个月后, 持久性无显著差异, 但空气保存后的萌发速度显著低于淹水保存, 表明孢子散布于淹水生境后, 时刻保持着快速萌发的能力.然而, 泥炭地正受到全球变化的威胁, 如全球变暖增加干旱频率, 导致泥炭地水位下降(
袁敏等, 2015 ), 很可能不利于地层中泥炭藓孢子的持久性.尖叶泥炭藓孢子经泥炭地地表水和泥炭藓沥出液保存后, 持久性显著高于超纯水保存, 表明纯水并不是决定孢子持久性的因素, 而水中的离子和化感物质可能是影响孢子持久性的关键因子.本研究结果与
Bu等(2017a) 苔藓化感作用减弱孢子萌发力的结论看似矛盾, 可能原因首先是实验物种的差异, 本文实验物种为尖叶泥炭藓和喙叶泥炭藓孢子, 而
Bu等(2017a) 实验物种为金发藓、泥炭藓和中位泥炭藓孢子, 本研究中尖叶泥炭藓孢子经3种保存液不充气处理后, 持久性之间有显著差异, 而喙叶泥炭藓孢子无显著差异, 可知不同种泥炭藓孢子对化感效应的响应不同.其次,
Bu等(2017a) 的研究中, 泥炭藓孢子经野外环境埋藏一个生长季, 可能遭受到紫外辐射和干旱等不利条件(
van Zanten, 1978 ), 导致孢子迅速老化, 持久性显著下降.此外, 由于其野外埋藏实验的苔藓基质化感物质浓度较高(
Mishler & Newton, 1988 ), 而本实验中的泥炭地地表水和泥炭藓沥出液分别经雨水和蒸馏水稀释, 降低了化感物质浓度.高浓度的化感物质虽然抑制孢子的萌发, 但很可能使其保持生活力, 而较低浓度的化感物质抑制微生物活性(
Verhoeven & Liefveld, 1997 ), 使孢子保持较高的萌发力. ...
 ,1,3,*, 卜兆君2,3, 吴玉环4, 刘莎莎2,3, 刘超2,3
,1,3,*, 卜兆君2,3, 吴玉环4, 刘莎莎2,3, 刘超2,3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}