薛晶月
, 1 , 2 , 王丽华
3 , 谢雨
1 , 高景
1 , 2 , 贺俊东
1 , 2 , 吴彦
, 1 , * 1 中国科学院成都生物研究所, 成都 610041 2 中国科学院大学, 北京 100049 3 阿坝师范学院, 四川汶川 623002 Effect of shrub coverage on grassland ecosystem carbon pool in southwestern China Jing-Yue XUE
, 1 , 2 , Li-Hua WANG
3 , Yu XIE
1 , Jing GAO
1 , 2 , Jun-Dong HE
1 , 2 , Yan WU
, 1 , * 1 Chengdu Institute of Biology, Chinese Academy of Sciences, Chengdu 610041, China 2 University of Chinese Academy of Sciences, Beijing 100049, China 3 Aba Teachers University, Wenchuan, Sichuan 623002, China 通讯作者: * wuyan@cib.ac.cn 编委: 黄建辉
责任编辑: 李敏
收稿日期: 2017-03-16
修回日期: 2019-04-9
网络出版日期: 2019-04-20
基金资助: 中国科学院战略性先导科技专项 (XDA05050307 )中国科学院战略性先导科技专项 (XDA05050404 )
Corresponding authors: * wuyan@cib.ac.cn Received: 2017-03-16
Revised: 2019-04-9
Online: 2019-04-20
Fund supported: Supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (XDA05050307 )Supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (XDA05050404 )
摘要 中国西南地区草地主要为暖性及热性草丛、灌草丛, 约占全国草地面积的1/10, 分析灌木植物盖度与草地碳库及其构成的关系对于准确评估尚处于次生演替阶段的南方草地碳储量具有重要意义。该研究基于野外实地调查, 将西南地区不同地貌类型的41个代表性草地样地依据灌木植物盖度划分为3种类型: 无灌木植物草地群落(灌木植物盖度为0)、低灌木植物盖度草地群落(灌木植物盖度0-10%)和高灌木植物盖度草地群落(灌木植物盖度10%-30%), 测定了群落地上、地下生物量和凋落物生物量以及植物和土壤碳含量, 计算碳密度。结果表明: 随着草地群落灌木植物盖度增大, 生态系统植被碳密度从0.304 kg·m
-2 增加到1.574 kg·m
-2 , 其中根系和凋落物碳库也呈增长趋势; 土壤碳密度从7.215 kg·m
-2 增加到9.735 kg·m
-2 , 生态系统碳密度从7.519 kg·m
-2 增加到11.309 kg·m
-2 。草地碳库构成中, 低灌木植物盖度草地群落的土壤碳库占生态系统碳库比例最小。草地群落灌木植物盖度增加改变了草地生态系统碳库构成并导致生态系统碳库增加, 建议在估算草地生态系统碳库时, 需要统筹考虑并兼顾南方地区草地群落灌木植物盖度变化。
关键词: 西南地区 ;
草地群落 ;
灌木植物盖度 ;
植被碳库 ;
土壤碳库 Abstract Aims The grassland in southwestern China is mainly warm and tropical grass and shrub grassland, accounting for approximately one-tenth of the national grassland area. Analysis of the relationship between shrub coverage and the grassland carbon pool and its composition is of great significance for accurately assessing carbon storage in the southwestern grassland, which grassland is still in the secondary succession stage.
Methods In a field investigation, 41 representative plots of different geomorphic types in southwestern China were divided into three types according to shrub coverage: no shrub grassland community (shrub coverage is 0), low shrub coverage grassland community (shrub coverage is 0-10%) and high shrub coverage grassland community (shrub coverage is 10%-30%). Carbon density was calculated by measuring the aboveground and underground biomass and litter biomass of grassland communities at different shrub coverage levels, as well as plant and soil carbon content.
Important findings The results showed that with increasing shrub coverage and species richness in grasslands, the ecosystem vegetation carbon density increased from 0.304 kg·m
-2 to 1.574 kg·m
-2 , and the roots and litter carbon density also showed growth trends. The soil carbon density increased from 7.215 kg·m
-2 to 9.735 kg·m
-2 , and the ecosystem carbon density increased from 7.519 kg·m
-2 to 11.309 kg·m
-2 , with increasing shrub coverage. Regarding the composition of the grassland carbon pool, the soil carbon pool of the low shrub coverage grassland accounted for the smallest proportion of the ecosystem carbon pool. In summary, the increase in shrub coverage changes the composition of the grassland ecosystem carbon pool and leads to an increase in the amount of ecosystem carbon. Therefore, when estimating the grassland ecosystem carbon pool, it is necessary to overall plan for and take into account the changes in grassland shrub coverage in southern China.
Keywords: southwestern China ;
grassland community ;
shrub coverage ;
vegetation carbon pool ;
soil carbon pool PDF (1105KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 薛晶月, 王丽华, 谢雨, 高景, 贺俊东, 吴彦. 西南地区草地群落灌木植物盖度对生态系统碳库的影响. 植物生态学报, 2019, 43(4): 365-373. DOI:
10.17521/cjpe.2017.0062 XUE Jing-Yue, WANG Li-Hua, XIE Yu, GAO Jing, HE Jun-Dong, WU Yan.
Effect of shrub coverage on grassland ecosystem carbon pool in southwestern China .
Chinese Journal of Plant Ecology , 2019, 43(4): 365-373. DOI:
10.17521/cjpe.2017.0062 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源。我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位。南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 )。目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 )。南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 )。因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要。
草地生态系统碳库由植被碳库和土壤碳库两部分构成。植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 )。西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 )。西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 )。但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 )。有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 )。在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 )。同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 )。然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性。
鉴于草地群落中灌木植物盖度对大尺度上的草地碳库估算具有重要意义, 本研究在我国西南地区选取了41个样地进行野外调查, 按照草地群落灌木植物盖度将草地分为3种类型, 实测不同灌木植物盖度情况下草地的地上、地下生物量和凋落物量, 同步测定植物和土壤碳含量、计算碳密度, 探讨西南地区草地群落灌木植物盖度变化对植被碳库、土壤碳库及其垂直分布的影响, 试图明确灌木植物盖度的变化对生态系统碳库的影响, 以期更好地理解草地生态系统碳循环过程, 为精确评估该类型生态系统碳库大小提供数据支撑。
1 材料和方法 1.1 野外调查 分别在我国西南地区的四川、重庆、云南、贵州等地选择丘陵地区、盆周山地、干热河谷地区、喀斯特石漠化区等典型地貌类型单元布置41个草地调查样地, 基本覆盖了我国西南地区的暖性草丛、暖性灌草丛、热性灌草丛、干热河谷灌丛等主要草地类型, 各样地的地理位置和基本信息见
表1 。
Table 1 表1 表1 西南地区草地调查样地的地理位置特征概况
Table 1
General information on the location and characteristics of the different investigation sites of grassland in southwestern China 地点 样地数 经度 纬度 海拔 地貌类型 样地编号 四川达州 2 107.60°-108.18° 31.86°-32.17° 1 055-1 060 盆周山地 Mountain area of basin 7, 14 四川绵阳 1 102.09° 26.93° 1 296 紫色丘陵区 Purple hilly area 8 四川雅安 3 102.49°-102.87° 29.29°-29.61° 1 171-1 592 盆周山地 Mountain area of basin 10, 15, 37 四川凉山 15 102.03°-103.77° 26.26°-29.08° 1 042-1 998 干热河谷及中山区 2, 4, 5, 6, 13, 19, 20, 21, 23, 24, 31, 32, 36, 39, 41 四川攀枝花 5 101.73°-101.85° 26.42°-26.82° 1 044-1 398 干热河谷 Dry-hot valley 1, 3, 9, 17, 18 云南曲靖 3 103.47°-104.27° 24.83°-26.77° 1 669-1 948 滇中丘陵区 Central Yunnan hilly area 33, 34, 35 云南红河 1 103.74° 24.36° 1 851 干热河谷 Dry-hot valley 25 云南昆明 1 102.41° 26.27° 1 184 滇中丘陵区 Central Yunnan hilly area 11 重庆 8 108.71°-109.14° 30.58°-32.06° 1 201-1 886 盆周山地 Mountain area of basin 16, 26, 27, 28, 29, 30, 38, 40 贵州黔南 2 105.08°-106.84° 25.28°-26.34° 1 469-1 504 喀斯特石漠化山区 12, 22
新窗口打开 |
下载CSV 在选择草地调查样地时, 尽量保证立地条件的一致性。各样地以中山地貌类型为主, 分布于1 042-1 998 m之间, 平均海拔1 500 m; 坡向大致以南坡(阳坡或半阳坡)为主, 坡度15°-25°, 坡位以坡中部为主。每个样地选择100 m × 100 m区域, 在对角线上设置一条100 m的调查样线, 样线上每隔10 m设置一个样方(共10个), 样方编号从1到10, 奇数位置设置5个1 m × 1 m的草本样方, 偶数位置设置5个5 m × 5 m的灌丛样方(内设1 m × 1 m的小样方调查草本层)。
在样地层面, 记录每个样地的经纬度、海拔、地形地貌、坡位、坡向、坡度以及群落的物种组成,测定每个物种的高度、盖度、密度。每个样地在中间位置挖一个大小为1.5 m × 0.5 m × 1 m (长×宽×深)的土壤剖面, 采用环刀法按照0-10、10-20、20-30、30-50、50-100 cm深度从上至下分层采集土壤样品, 每层取5个重复, 环刀的规格为高度5 cm、体积100 cm
3 。
在样方层面, 记录每个样方内的物种组成, 测定每个物种的高度、盖度、密度; 收割法测定样方中草本植物地上生物量, 收集凋落物并烘干; 样方中灌木则采用收割1/3-1/5的植株测定其生物量, 用电子天平(精度0.01 g)野外称取鲜质量, 室内测烘干后的质量。用土钻法采集每个样方中的土壤样品根系测定生物量, 每个样方一般取4-6钻混合, 用直径5 cm的土钻分0-10、10-20、20-30、30-50、50-100 cm 5个层次依次取样(土层深度不足100 cm则至母质层为止)、洗净收集根系, 室内烘干并折算为相应的根系分层生物量。
根据样方调查资料, 本试验41个样地共记录323种草本植物, 55种灌木植物。根据草地群落灌木植物盖度, 可将这41个样地分成3类: (1)无灌木植物草地群落(I), 指灌木植物盖度为0, 仅由草本植物组成的草本群落, 共18个样地。根据中国草地类型分类系统(
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 )划分草地类型, 草地类型为暖性草丛, 中禾草组, 白茅草地型。(2)低灌木植物盖度草地群落(II), 指灌木植物盖度0-10%的草本群落, 共18个样地。草地类型为热性灌草丛, 中禾草组, 具灌木的扭黄茅草地型。(3)高灌木植物盖度草地群落(III), 即灌木植物盖度10%-30%的草本群落, 共5个样地。草地类型为暖性灌草丛, 中禾草组, 主要为具灌木的荩草型。随着草地群落灌木植物盖度的增加, 草地类型从草丛变为灌草丛, 灌木种类和数量增多, 植物物种也更加丰富(
表2 )。
Table 2 表2 表2 西南地区不同灌木植物盖度样地的草地类型及主要物种
Table 2
Grassland types and main species in grassland with different shrub coverage levels in southwestern China 灌木植物盖度 Shrub coverage 样地编号 Plots No. 草地类型 Vegetation types 主要物种 Main species I 2, 5, 11, 13, 16, 20, 暖性草丛, 白茅型Imperata cylindrica grassland type 白茅、荩草、车前、老鹳草、小蓬草、天名精Imperata cylindrica, Arthraxon hispidus, Plantago asiatica, Geranium wilfordii, Conyza canadensis, Carpesium abrotanoides II 1, 3, 4, 8, 9, 10, 12, 14, 15, 17, 18, 19, 22, 25, 27, 29, 31, 32 热性灌草丛, 具灌木的扭黄茅型Heteropogon contortus grassland type 车桑子、川滇绣线菊、野蔷薇、马桑、扭黄茅、野古草属植物Dodonaea viscosa, Spiraea schneideriana, Rosa multiflora, Coriaria nepalensis, Heteropogon contortus, Arundinella spp. III 6, 7, 26, 28, 36 暖性灌草丛, 具灌木的荩草型Warm-temperate shrub tussock, Arthraxon hispidus grassland type 车桑子、川滇绣线菊、野蔷薇、火棘、蜡莲绣球、悬钩子属植物、车前、风轮菜、荩草、老鹳草Dodonaea viscosa, Spiraea schneideriana, Rosa multiflora, Pyracantha fortuneana, Hydrangea strigosa, Rubus ssp., Plantago asiatica,
I, II, and III represent no shrub grassland, low shrub coverage grassland, and high shrub coverage grassland, respectively. 表中I、II、III分别代表无灌木植物草地群落、低灌木植物盖度草地群落和高灌木植物盖度草地群落。 新窗口打开 |
下载CSV 1.2 样品处理 所有草本、灌木和凋落物生物量样品在65 ℃烘箱烘干至恒质量(
Perkins & Owens, 2003 ;
Nie et al ., 2016 ;
杨路存等, 2017 ), 测定生物量干质量; 土壤样品风干后过2 mm的土壤筛, 用静电吸附(经摩擦过的塑料卡片或玻璃棒)和手动方法去除根系。随后所有样品用球磨仪研磨后过100目筛, 得到的样品用于指标分析, 3次重复。植物和土壤样品碳含量用元素分析仪(Vario MAX, Elementar, Hanau, Germany)测定。
1.3 数据计算 西南地区草地生态系统植被碳库包括植物地上碳库、地下碳库和凋落物碳库, 计算公式(
Ravindranath & Ostwald, 2008 )为:
植被碳库:
(1) CD veg = C p × B p + C l × B l + Σ(C r × B r )i 土壤碳库(单位面积一定深度土层中土壤碳储量):
(2) CD soil = Σ(BD soil × C soil × T soil × (1 - G soil ))i (3) 草地生态系统碳库: CD eco = CD veg + CD soil 式中,
C p 和
B p 表示地上活体植物碳含量(%)和生物量(g);
C l 和
B l 表示凋落物碳含量(%)和生物量(g);
C r 和
B r 表示根系碳含量(%)和生物量(g);
BD soil 表示土壤容重(g·cm
-3 );
C soil 表示土壤碳含量(%);
T soil 表示土层厚度(cm);
G soil 为砾石体积百分比;
i 表示土壤层。
1.4 数据分析 采用单因素方差分析方法对土壤层碳库、植被碳库, 以及植物地上部分、凋落物、根系碳库进行显著性检验。图表绘制使用Origin 9.0。统计分析使用SPSS 19.0统计软件, 显著性差异水平为
α = 0.05。
2 结果 2.1 不同灌木植物盖度草地群落的植被碳库 不同灌木植物盖度草地群落的植被碳库差异显著(
F = 11.459,
p < 0.001), 随着草地群落灌木植物盖度逐渐增大, 植被碳库逐渐增加, 且低灌木植物盖度草地群落和高灌木植物盖度草地群落的植被碳库显著大于无灌木植物草地群落, 但低灌木植物盖度草地群落和高灌木植物盖度草地群落的植被碳库差异不显著(
表3 ;
图1 )。就地上植物碳密度而言, 无灌木植物草地群落、低灌木植物盖度草地群落和高灌木植物盖度草地群落分别为0.057、0.089和0.073 kg·m
-2 , 彼此之间地上植物碳库差异显著(
F = 3.348,
p = 0.046)。其次, 凋落物碳密度分别为0.005、0.014和0.015 kg·m
-2 , 三者间凋落物碳库差异显著(
F = 9.223,
p = 0.001), 低灌木植物盖度草地群落和高灌木植物盖度草地群落的凋落物碳库显著大于无灌木植物草地群落。另外, 三者的地下植物碳密度分别为0.242、1.304和1.486 kg·m
-2 , 彼此之间差异显著(
F = 10.969,
p < 0.001), 表现为低灌木植物盖度草地群落和高灌木植物盖度草地群落的根系碳库显著大于无灌木植物草地群落(
图1 )。随着草地群落灌木植物盖度增加, 包括凋落物和地下碳库在内, 植被碳库有逐渐增加的趋势, 但这种变化随着灌木植物盖度增加, 趋势有所减弱, 表现为低灌木植物盖度草地群落和高灌木植物盖度草地群落的植被碳库差异不显著。
Table 3 表3 表3 西南地区不同灌木植物盖度草地生态系统碳库
Table 3
Carbon pool in grassland ecosystem with different shrub coverage in southwestern China 灌木植物 植被碳库-2 ) 土壤碳库-2 ) 生态系统碳库-2 ) 0-50 cm土层碳库-2 ) 植被碳库占生态 土壤碳库占生态 I 0.304 ± 0.036a 7.215 ± 0.507a 7.519 ± 0.505a 6.870 ± 0.468a 4.33 ± 0.79a 95.67 ± 0.79a II 1.407 ± 0.204b 6.031 ± 0.736a 7.438 ± 0.717a 5.485 ± 0.722a 19.70 ± 3.57b 80.30 ± 3.57b III 1.574 ± 0.675b 9.735 ± 0.444b 11.309 ± 1.011b 7.166 ± 0.374b 12.54 ± 4.36ab 87.46 ± 4.36ab F 11.459 1.610 5.734 1.287 9.053 9.053 p <0.001 0.213 0.007 0.288 0.001 0.001
I, II, and III represent no shrub grassland, low shrub coverage grassland, and high shrub coverage grassland, respectively. The data are the mean ± SE (I, II, n = 18; III, n = 5). Different lowercase letters indicate significant differences among sites with different shrub coverage levels (p < 0.05). I、II、III分别为无灌木植物草地群落、低灌木植物盖度草地群落和高灌木植物盖度草地群落。表中的数值为平均值±标准误差(I, II, n = 18; III, n = 5)。不同小写字母表示不同灌木植物盖度间差异显著(p < 0.05)。 新窗口打开 |
下载CSV 图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1西南地区不同灌木植物盖度草地群落的植被碳库。图中I、II、III分别代表无灌木植物草地群落、低灌木植物盖度草地群落和高灌木植物盖度草地群落, 数值为平均值±标准误差(I, II, n = 18; III, n = 5)。不同小写字母表示不同灌丛草地间差异显著(p < 0.05)。 Fig. 1Vegetation carbon pool of vegetation in grassland community with different shrub coverage in southwestern China. I, II, and III are the three different shrub coverage levels of the grassland community. I, II, and III represent no shrub grassland, low shrub coverage grassland, and high shrub coverage grassland, respectively. The data are the mean ± SE (I, II, n = 18; III, n = 5). Different lowercase letters indicate significant differences among sites with different shrub coverage of grassland levels (p < 0.05). 2.2 不同类型草地群落各土壤层碳库 从无灌木植物草地群落到高灌木植物盖度草地群落, 草地土壤碳库未呈现显著的变化趋势。就土壤碳库而言, 无灌木植物草地群落与低灌木植物盖度和高灌木植物盖度草地群落差异不显著(
F = 1.610,
p = 0.213)。这说明, 草地群落中灌木植物盖度的大小并没有对土壤碳库造成显著影响。在垂直剖面上, 高灌木植物盖度草地群落的土壤表层(0-10 cm)碳库显著大于低灌木植物盖度草地群落(
F = 4.475,
p = 0.018); 但在10-20 cm和30-50 cm土层, 不同灌木植物盖度草地群落土壤碳库无显著差异(
F = 1.914,
p = 0.161;
F = 0.093,
p = 0.912); 在20-30 cm土层无灌木植物草地群落碳库显著大于高灌木植物盖度草地群落(
F = 4.777,
p = 0.014), 低灌木植物盖度草地群落与二者差异均不显著; 在50-100 cm土层, 不同灌木植物盖度草地群落的土壤碳库差异显著(
F = 4.149,
p = 0.023), 从无灌木植物草地群落、低灌木植物盖度草地群落到高灌木植物盖度草地群落, 土壤碳库逐渐增加, 且高灌木植物盖度草地群落显著大于无灌木植物草地群落(
图2 )。总体来看, 一方面, 草地土壤碳库并没有因为灌木植物盖度增加发生显著变化; 另一方面, 不同灌木植物盖度草地群落土壤碳库的空间分层变化明显, 表层和底层灌木多的样地土壤碳相对较高, 而中间的过渡土层的土壤碳库则呈现多样性趋势。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2西南地区不同灌木植物盖度草地群落的土壤碳库。图中的数值为平均值±标准误差(I, II, n = 18; III, n = 5)。I、II、III分别代表无灌木植物草地群落、低灌木植物盖度草地群落和高灌木植物盖度草地群落。不同大写字母表示0-100 cm的土壤碳库差异显著(p < 0.05), 不同小写字母表示不同土层的碳库差异显著(p < 0.05)。 Fig. 2Soil carbon pool in grassland community with different shrub coverage in southwestern China. I, II, and III represent no shrub grassland, low shrub coverage grassland, and high shrub coverage grassland, respectively. The data are the means ± SE (I, II, n = 18; III, n = 5). Different uppercase letters indicate that the soil carbon pool at 0-100 cm was significantly different among different shrub coverage grassland communities (p < 0.05), and different lowercase letters indicate significant differences in the carbon pool at different soil layers (p < 0.05). 2.3 不同类别的草地生态系统碳库 从无灌木植物草地群落、低灌木植物盖度草地群落到高灌木植物盖度草地群落, 植被碳库和土壤碳库表现出了不同的变化趋势。植被碳库呈增加趋势, 土壤碳库先降低后增加, 生态系统碳库少量降低后显著增加。植被碳库在灌木从无到有的过程中发生了显著变化, 土壤碳库在草地群落灌木植物盖度从低到高的过程中显著增加。0-50 cm土层土壤碳库分别占1 m土壤碳库的95.22%、90.95%和77.28%, 50-100 cm土层碳库由4.78%增加到22.72%。说明随草地群落灌木植物盖度增加, 灌丛草地的土壤碳向深层分配, 这反映了灌木根系的贡献。植被碳库占草地生态系统碳库比例先增加后减少, 土壤碳库占生态系统碳库比例先降低后升高(
表3 )。
3 讨论 3.1 草地群落灌木植物盖度对植被碳库的影响 生物量作为碳库的重要组分, 是量化生态系统碳周转的重要指标(
Ferlan et al ., 2011 )。木本植物比草本植物具有更多的生物量(
Seastedt & Ramundo, 1990 ), 灌丛地上净初级生产力的增加导致地上碳储量的增加, 并使地下碳库有增加的潜力(
Norris et al ., 2001 ;
Smith & Johnson, 2003 )。因此, 在草地向灌丛和森林演替过程中, 草地群落灌木植物盖度的增大对植被碳库有重要影响。有研究表明, 中国草地植被碳密度平均值范围为0.216-1.148 kg·m
-2 (
Ni, 2002 ;
Fang et al ., 2007 ;
Piao et al ., 2007 )。利用遥感影像建立的植被生物量估算模型计算得到的中国草地植被生物量平均碳密度为0.315 kg·m
-2 (
朴世龙等, 2004 )。本研究中, 无灌木植物草地群落的植被层碳密度为0.304 kg·m
-2 , 与前人研究结果相近。然而低灌木植物盖度草地群落的植被碳密度为1.407 kg·m
-2 , 高灌木植物盖度草地群落的植被碳密度为1.574 kg·m
-2 , 有灌木植物草地群落碳密度远大于无灌木植物草地群落。低灌木植物盖度草地群落相对于无灌木植物草地群落既有草本层的凋落物, 同时也有灌木层的凋落物的贡献(
聂秀青等, 2018 ), 凋落物碳库增加。
Connin等(1997) 研究表明, 灌木相对于多年生草本植物具有更深广发达的根系, 地下根系生物量增加。因此, 灌木植物盖度增大, 植物地上生物量会有所增加, 进而影响草地群落的植被碳库, 忽略灌木植物盖度增加过程将导致对碳库的低估。
随着草地群落灌木植物盖度增加, 灌丛草地地上净初级生产力增加(
Reich et al ., 2001 ;
Hughes et al ., 2006 ;
Knapp et al ., 2008 ), 更多的生物量被分配到地上部分(
Jackson et al ., 1997 ;
Goodale & Davidson, 2002 ), 植被地上碳库增加。但当灌木植物盖度增加到一定程度后, 灌木植株可能会遮挡下层草本植物的光照, 减少草本植物生物量, 导致植物地上碳库反而会低于低灌木植物盖度草地群落。因此, 草地群落中的灌木植物盖度与植物地上碳储量之间存在一定的不确定性, 在大尺度的碳储量评估过程中需要给予高度重视。
3.2 草地群落灌木植物盖度对土壤碳库的影响 草地群落中灌木的增加对土壤碳库的影响目前还未有一致的认识, 土壤碳库与灌木植物盖度的关系也有很大的不确定性。例如,
Hughes等(2006) 的结果表明, 草地演替过程中土壤碳库没有显著改变;
万昊等(2015) 对黄土高原植被演替过程中土壤碳库的研究表明, 草地土壤碳储量高于灌木地; 而
Archer等(1995) 、
Brantley和Young (2010) 、
陈蕾伊等(2014) 则认为, 随着草地群落灌木植物盖度增加土壤碳库也增加。这种研究结果不一致的原因, 也许与气候条件有关, 例如
Jackson等(2002) 在干旱地区的研究也显示, 草地群落土壤碳库随灌木盖度的增加而增加; 但在年降水量较大的地区, 草地土壤碳库随着灌木盖度的增加而降低。灌木根系对土壤碳库的影响是多方面的(
Matamala & Schlesinger, 2003 ), 有研究认为, 灌木将更多的生物量分配到地上而减少地下生物量分配, 分解腐烂后返还给土壤的有机碳也相对较少(
Goodale & Davidson, 2002 ); 但也有研究显示, 灌木增加了凋落物及根系向土壤的营养输入, 根系生长有利于土壤有机质形成和积累(
Knapp et al ., 2008 )。
本研究中, 低灌木植物盖度草地群落土壤碳库(6.031 kg·m
-2 )小于无灌木植物草地群落(7.215 kg·m
-2 ), 究其原因可能是西南地区水热条件好, 灌木生长较为迅速, 在增加土壤的碳输入的同时(
Fang et al ., 2003 ), 也会增加土壤向大气的碳输出(
Feng et al ., 2008 ); 当然也有可能这些次生灌木尚处在幼龄期还不能对土壤碳库有显著的影响(
Alberti et al ., 2008 )。无灌木植物草地群落绝大部分土壤碳(95.22%)分布于0-50 cm土层, 表层草本植物根系是土壤有机碳密度较高的主要原因, 而灌木根系会延伸至较深土层(
万昊等, 2015 );
Hughes等(2006) 和
陈蕾伊等(2014) 发现, 灌丛草原与无灌丛草原相比, 表层土壤碳储量无显著差异, 而深层土壤碳储量增加。因此, 高灌木植物盖度草地群落比无灌木植物草地群落具有更高的土壤碳密度, 随着灌木植物盖度的增大, 草地生态系统倾向于增加土壤深层生物量以及土壤碳库。
3.3 草地群落灌木植物盖度对生态系统碳库的 影响 西南地区草地多属次生性质, 是以多年生草本植物为主体, 间混少量乔木或灌木的草地类型。在停止人为干扰、水土条件良好的地段, 经过一定时间的恢复后可能回到森林顶极群落(
吴征镒, 1980 )。在顺行演替过程中, 灌丛草地比无灌木植物草地群落维持了更高的物种丰富度(
Howard et al ., 2012 ), 草地向灌丛演替有利于草地群落增加生物量, 促进生态系统碳固定(
Pacala et al ., 2001 ); 随着灌木植物盖度增加, 草地向森林演替过程中生态系统固碳能力改变(
Post & Kwon, 2000 ;
Jackson et al ., 2002 ;
McKinley & Blair, 2008 )。有研究显示, 土壤有机碳密度在青藏高原高寒草地为6.5 kg·m
-2 、北方草地为8.5 kg·m
-2 (
Yang et al ., 2008 ), 而本研究实测的西南地区无灌木植物草地群落(7.215 kg·m
-2 )和低灌木植物盖度草地群落(6.031 kg·m
-2 )的土壤碳密度介于两者之间, 而高灌木植物盖度草地群落(9.735 kg·m
-2 )显著高于高寒草地和北方草地;
Ni (2002) 根据全球土壤碳氮数据库估算的中国草地土壤有机碳密度为13.2 kg·m
-2 , 显著高于本研究实测得到的西南地区草地的土壤平均碳密度。这表明以北方草地和高寒草地的数据为基础将会低估中国草地生态系统碳库, 以全球植被和土壤数据库为基础则会高估中国草地生态系统碳库。
4 结论 我国西南地区草地随着灌木郁闭度的增加, 草地生态系统碳库呈现增加趋势, 植被碳密度也逐渐增加。灌木植物盖度大于10%的草地群落碳库高达11.309 kg·m
-2 , 显著大于低灌木植物盖度草地群落和无灌木植物草地群落。但土壤碳库差异不显著且倾向于增加深层土壤碳密度。因此, 在估算草地生态系统碳库时, 建议统筹考虑并兼顾南方地区草地群落灌木植物盖度变化。
致谢 本文中云南和贵州的样方资料得到了中国科学院西双版纳植物园张一平老师和中国科学院植物研究所肖春旺老师的数据支持, 在此表示衷心感谢!感谢中国科学院成都生物研究所参与课题的所有老师和同学。
[1] Alberti G Peressotti A Piussi P Zerbi G 2008 ). Forest ecosystem carbon accumulation during a secondary succession in the Eastern Prealps of ItalyForestry , 81, 1 -11 . [本文引用: 1] [2] Animal Husbandry and Veterinary Department of the Ministry of Agriculture of the People’s Republic of China ( 1996 ). Rangeland Resources of China . China Science and Technology Press, Beijing. 250-277, 476 -489 . [本文引用: 2] [ 中华人民共和国农业部畜牧兽医司全国畜牧兽医总站 ( 1996 ). 中国草地资源 . 中国科学技术出版社, 北京. 250-277, 476 -489 .] [本文引用: 2] [3] Archer S Schimel DS Holland EA 1995 ). Mechanisms of shrubland expansion: Land use, climate or CO2 ?Climatic Change , 29, 91 -99 . [本文引用: 1] [4] Brantley ST Young DR 2010 ). Shrub expansion stimulates soil C and N storage along a coastal soil chronosequenceGlobal Change Biology , 16, 2052 -2061 . [本文引用: 1] [5] Cai WT Lai LM Li HY Zhou JH Guan TY Zhang XL Gao NN Zheng YR 2016 ). Progress of research on shrub encroachment in grasslandChinese Journal of Applied and Environmental Biology , 22, 531 -537 . [本文引用: 2] [ 蔡文涛 , 来利明 , 李贺祎 , 周继华 , 管天玉 , 张晓龙 , 高楠楠 , 郑元润 ( 2016 ). 草地灌丛化研究进展应用与环境生物学报 , 22, 531 -537 .] [本文引用: 2] [6] Chen LY Shen HH Fang JY 2014 ). Shrub-encroached grassland: A new vegetation typeChinese Journal of Nature , 36, 391 -396 . [ 陈蕾伊 , 沈海花 , 方精云 ( 2014 ). 灌丛化草原: 一种新的植被景观自然杂志 , 36, 391 -396 .] [7] Connin SL Virginia R Chamberlain C 1997 ). Carbon isotopes reveal soil organic matter dynamics following arid land shrub expansionOecologia , 110, 374 -386 . [本文引用: 1] [8] Fang JY Geng XQ Zhao X Shen HH Hu HF 2018 ). How many areas of grasslands are there in China?Chinese Science Bulletin , 63, 1731 -1739 . [本文引用: 1] [ 方精云 , 耿晓庆 , 赵霞 , 沈海花 , 胡会峰 (2018 ). 我国草地面积有多大科学通报 , 63, 1731 -1739 .] [本文引用: 1] [9] Fang JY Guo ZD Piao SL Chen AP 2007 ). Terrestrial vegetation carbon sinks in China, 1981-2000Science in China Series D-Earth Sciences , 50, 1341 -1350 . [本文引用: 1] [10] Fang JY Piao SL Christopher BF Pan YD Guo QH Zhou LM Peng CH Tao S 2003 ). Increasing net primary production in China from 1982 to 1999Frontiers in Ecology and the Environment , 1, 293 -297 . [本文引用: 1] [11] Fang JY Yang YH Ma WH Mohammat A Shen HH 2010 ). Ecosystem carbon stocks and their changes in China’s grasslandsScientia Sinica (Vitae )757 -765 . [本文引用: 1] [ 方精云 , 杨元合 , 马文红 , 安尼瓦尔·买买提 , 沈海花 ( 2010 ). 中国草地生态系统碳库及其变化中国科学: 生命科学 , 53, 757 -765 .] [本文引用: 1] [12] Feng XJ Simpson AJ Wilson KP Williams DD Simpson AMJ 2008 ). Increased cuticular carbon sequestration and lignin oxidation in response to soil warmingNature Geoscience , 1, 836 -839 . [本文引用: 1] [13] Ferlan M Alberti G Eler K Batic F Peressotti A Miglietta F Zaldei A Simoncic P Vodnik D 2011 ). Comparing carbon fluxes between different stages of secondary succession of a karst grasslandAgriculture Ecosystems & Environment , 140, 199 -207 . [本文引用: 2] [14] Goodale CL Davidson EA 2002 ). Carbon cycle: Uncertain sinks in the shrubsNature , 418, 593 -594 . [本文引用: 2] [15] Howard KSC Eldridge DJ Soliveres S 2012 ). Positive effects of shrubs on plant species diversity do not change along a gradient in grazing pressure in an arid shrublandBasic and Applied Ecology , 13, 159 -168 . [本文引用: 1] [16] Huangfu JY Mao FX Lu XS 2012 ). Analysis of grassland and resources in southwest ChinaActa Prataculturae Sinica , 21(1 ), 75 -82 . DOI:
10.11686/cyxb20120110 Magsci [本文引用: 1] 根据中国草地资源调查资料(农业部畜牧兽医司,1996)、中国草业统计(2009)和相关文献,在实地调查的基础上,以中国西南地区的天然草地资源、冬闲田资源、林间草地为研究对象,分析了中国西南地区的草地资源状况。结果表明,1)西南地区天然草地总面积35.58×106 hm<sup>2</sup>,占该区域土地总面积的32.59%,可利用草地面积29.04×106hm<sup>2</sup>,各省可利用草地面积占天然草地面积比例都在70%以上;2)按照以服务畜牧业为主要原则,将天然草地资源划为4个区域:热带湿润灌草丛区、亚热带湿润灌草丛区、亚热带半湿润灌草丛区和高原温带湿润灌丛草甸区;3)西南地区岩溶面积达52.23×106hm<sup>2</sup>,占土地面积的29.50%,共有292 个岩溶县。石漠化面积达11.11×106hm<sup>2</sup>,占岩溶面积的21.27%,有88个石漠化严重县;4)西南地区农闲田可利用面积为961.46×104 hm<sup>2</sup>,农闲田已利用面积为193.11×104 hm<sup>2</sup>,占农闲田可利用面积的20.09%。冬闲田种草已利用面积共计118.58×104hm<sup>2</sup>,占农闲田可利用面积的12.33%,占农闲田已利用面积的61.40%。5)西南地区灌丛、疏林草地面积合计为1894.84×104hm<sup>2</sup>,占草地面积的29.81%,占可利用草地面积的36.06%。
[ 皇甫江云 , 毛凤显 , 卢欣石 ( 2012 ). 中国西南地区的草地资源分析草业学报 , 21(1 ), 75 -82 .] DOI:
10.11686/cyxb20120110 Magsci [本文引用: 1] 根据中国草地资源调查资料(农业部畜牧兽医司,1996)、中国草业统计(2009)和相关文献,在实地调查的基础上,以中国西南地区的天然草地资源、冬闲田资源、林间草地为研究对象,分析了中国西南地区的草地资源状况。结果表明,1)西南地区天然草地总面积35.58×106 hm<sup>2</sup>,占该区域土地总面积的32.59%,可利用草地面积29.04×106hm<sup>2</sup>,各省可利用草地面积占天然草地面积比例都在70%以上;2)按照以服务畜牧业为主要原则,将天然草地资源划为4个区域:热带湿润灌草丛区、亚热带湿润灌草丛区、亚热带半湿润灌草丛区和高原温带湿润灌丛草甸区;3)西南地区岩溶面积达52.23×106hm<sup>2</sup>,占土地面积的29.50%,共有292 个岩溶县。石漠化面积达11.11×106hm<sup>2</sup>,占岩溶面积的21.27%,有88个石漠化严重县;4)西南地区农闲田可利用面积为961.46×104 hm<sup>2</sup>,农闲田已利用面积为193.11×104 hm<sup>2</sup>,占农闲田可利用面积的20.09%。冬闲田种草已利用面积共计118.58×104hm<sup>2</sup>,占农闲田可利用面积的12.33%,占农闲田已利用面积的61.40%。5)西南地区灌丛、疏林草地面积合计为1894.84×104hm<sup>2</sup>,占草地面积的29.81%,占可利用草地面积的36.06%。
[17] Hughes RF Archer SR Asner GP Wessman CA McMurtry C Nelson J Ansley RJ 2006 ). Changes in aboveground primary production and carbon and nitrogen pools accompanying woody plant encroachment in a temperate savannaGlobal Change Biology , 12, 1733 -1747 . [本文引用: 3] [18] Jackson RB Banner JL Jobbagy EG Pockman WT Wall DH 2002 ). Ecosystem carbon loss with woody plant invasion of grasslandsNature , 418, 623 -626 . [本文引用: 2] [19] Jackson RB Mooney HA Schulze ED 1997 ). A global budget for fine root biomass, surface area, and nutrient contentsProceedings of the National Academy of Sciences of the United States of America , 94, 7362 -7366 . [本文引用: 1] [20] Knapp AK Briggs JM Collins SL Archer SR Bret-Harte MS Ewers BE Peters DP Young DR Shaver GR Pendall E Cleary MB 2008 ). Shrub encroachment in North American grasslands: Shifts in growth form dominance rapidly alters control of ecosystem carbon inputsGlobal Change Biology , 14, 615 -623 . [本文引用: 3] [21] Li B 1997 ). The rangeland degradation in North China and its preventive strategyScientia Agricultura Sinica , 30(6 ), 1 -9 . Magsci [本文引用: 1] 草地退化是在过牧、开垦等人为活动及不利自然因素影响下草地生态系统逆行演替的一种过程,是土地荒漠化的主要形式之一。本文在讨论了草地退化指标及其等级划分的基础上,编制了中国北方退化草地分布图。据此图计算,北方草地退化面积达137.77万km#+2,占该草地总面积的50.24%,近15年来以每年1.9%的速度在扩大退化。为此提出了防治对策及草地畜牧业可持续发展战略。
[ 李博 ( 1997 ). 中国北方草地退化及其防治对策中国农业科学 , 30(6 ), 1 -9 .] Magsci [本文引用: 1] 草地退化是在过牧、开垦等人为活动及不利自然因素影响下草地生态系统逆行演替的一种过程,是土地荒漠化的主要形式之一。本文在讨论了草地退化指标及其等级划分的基础上,编制了中国北方退化草地分布图。据此图计算,北方草地退化面积达137.77万km#+2,占该草地总面积的50.24%,近15年来以每年1.9%的速度在扩大退化。为此提出了防治对策及草地畜牧业可持续发展战略。
[22] Matamala R Schlesinger WH 2003 ). Impacts of fine root turnover on forest NPP and soil C sequestration potentialScience , 302, 1385 -1387 . [本文引用: 1] [23] McKinley DC Blair JM 2008 ). Woody plant encroachment by Juniperus virginiana in a mesic native grassland promotes rapid carbon and nitrogen accrualEcosystems 454 -468 . [本文引用: 1] [24] Ni J 2002 ). Carbon storage in grasslands of ChinaJournal of Arid Environments , 50, 205 -218 . [本文引用: 2] [25] Nie XQ Xiong F Li CB Yang LC Xiao YM Zhou GY 2018 ). Biomass allocation relationships in the grass layer in alpine shrubland ecosystems on the Tibetan PlateauActa Ecologica Sinica , 38, 6664 -6669 . [本文引用: 2] [ 聂秀青 , 熊丰 , 李长斌 , 杨路存 , 肖元明 , 周国英 (2018 ). 青藏高原高寒灌丛生态系统草本层生物量分配格局生态学报 , 38, 6664 -6669 .] [本文引用: 2] [26] Nie XQ Yang YH Yang LC Zhou GY 2016 ). Above- and belowground biomass allocation in shrub biomes across the northeast Tibetan PlateauPLOS ONE , 11, e0154251 . DOI: 10.1371/journal.pone.0154251 . [本文引用: 1] [27] Norris MD Blair JM Johnson LC McKane RB 2001 ). Assessing changes in biomass, productivity, and C and N stores following Juniperus virginiana forest expansion into tallgrass prairieCanadian Journal of Forest Research 1940 -1946 . [本文引用: 1] [28] Pacala SW Hurtt GC Baker D Peylin P Houghton RA Birdsey RA Heath L Sundquist ET Stallard RF Ciais P Moorcroft P Caspersen JP Shevliakova E Moore B Kohlmaier G Holland E Gloor M Harmon ME Fan S-M Sarmiento JL Goodale CL Schimel D Field CB 2001 ). Consistent land- and atmosphere-based U.S. carbon sink estimatesScience , 292, 2316 -2320 . [本文引用: 1] [29] Perkins SR Owens MK 2003 ). Growth and biomass allocation of shrub and grass seedlings in response to predicted changes in precipitation seasonalityPlant Ecology , 168, 107 -120 . [本文引用: 1] [30] Piao SL Fang JY Ciais P Peylin P Huang Y Sitch S Wang T 2009 ). The carbon balance of terrestrial ecosystems in ChinaNature , 458, 1009 -1013 . [本文引用: 1] [31] Piao SL Fang JY He JS Xiao Y 2004 ). Spatial distribution of grassland biomass in ChinaActa Phytoecologica Sinica , 28, 491 -498 . [本文引用: 1] [ 朴世龙 , 方精云 , 贺金生 , 肖玉 ( 2004 ). 中国草地植被生物量及其空间分布格局植物生态学报 , 28, 491 -498 .] [本文引用: 1] [32] Piao SL Fang JY Zhou LM Tan K Tao S 2007 ). Changes in biomass carbon stocks in China’s grasslands between 1982 and 1999Global Biogeochemical Cycles GB2002 . DOI: 10.1029/2005GB002634 . [本文引用: 1] [33] Post WM Kwon KC 2000 ). Soil carbon sequestration and land-use change: Processes and potentialGlobal Change Biology , 6, 317 -327 . [本文引用: 1] [34] Ravindranath NH Ostwald M 2008 ). Carbon Inventory Methods . Handbook for Greenhouse Gas Inventory, Carbon Mitigation and Roundwood Production Projects Springer,Heidelberg . 201 -214 . [本文引用: 1] [35] Reich PB Peterson DW Wedin DA Wrage K 2001 ). Fire and vegetation effects on productivity and nitrogen cycling across a forest-grassland continuumEcology , 82, 1703 -1719 . [本文引用: 1] [36] Ren JZ Zhang YJ 2002 ). Grassland resources in the south of China and its development strategyJournal of China Institute of Metrology , 13(3 ), 174 -180 . [本文引用: 2] [ 任继周 , 张英俊 ( 2002 ). 中国南方草地资源及其发展战略中国计量学院学报 , 13(3 ), 174 -180 .] [本文引用: 2] [37] Scurlock JMO Johnson K Olson RJ 2002 ). Estimating net primary productivity from grassland biomass dynamics measurementsGlobal Change Biology , 8, 736 -753 . [本文引用: 1] [38] Seastedt TR Ramundo RA 1990 ). The influence of fire on belowground processes of tallgrass prairiesFire in North American Tallgrass Prairies . University of Oklahoma Press, Norman, USA. [本文引用: 1] [39] Shen HH Zhu YK Zhao X Geng XQ Gao SQ Fang JY 2016 ). Analysis of current grassland resources in ChinaChinese Science Bulletin , 61, 139 -154 . [本文引用: 1] [ 沈海花 , 朱言坤 , 赵霞 , 耿晓庆 , 高树琴 , 方精云 ( 2016 ). 中国草地资源的现状分析科学通报 , 61, 139 -154 .] [本文引用: 1] [40] Smith DL Johnson LC 2003 ). Expansion of Juniperus virginiana L. in the Great Plains: Changes in soil organic carbon dynamicsGlobal Biogeochemical Cycles 1062 -1071 . [本文引用: 1] [41] Wan H Liu WG Wei J 2015 ). Effects of vegetation succession on carbon stock and δ 13 C in Loess PlateauChinese Journal of Ecology 100 -105 . [本文引用: 2] [ 万昊 , 刘卫国 , 魏杰 ( 2015 ). 黄土高原植被演替对土壤碳库及δ 13 C的影响 生态学杂志 , 34, 100 -105 .] [本文引用: 2] [42] Wang M Fu XQ Shi FS Wang YX Wang Q Lu T Wu Y 2014 ). Compensatory growth responding to clipping: A case study in a subtropical grassland northeast of ChongqingChinese Journal of Applied and Environmental Biology , 20, 474 -483 . [本文引用: 1] [ 王梅 , 付秀琴 , 石福孙 , 王彦星 , 王乾 , 卢涛 , 吴彦 ( 2014 ). 刈割对南方草地植物补偿性生长的影响——以渝东北部岐山草场为例应用与环境生物学报 , 20, 474 -483 .] [本文引用: 1] [43] Wu XD Song NP Pan J 2016 ). Effect of shrub (Caragana intermedia ) encroachment under different sandy habitats on content and distribution of soil organic carbon in desert grasslandTransactions of the Chinese Society of Agricultural Engineering 10 ), 115 -121 . Magsci [本文引用: 2] 为揭示不同沙地生境下灌丛化草地土壤有机碳及其组分的分布特征,以宁夏盐池县沙化草原不同沙地生境(丘间低地、固定沙地、半固定沙地、流动沙地)下不同发育期的柠条灌丛斑块及其对应的草地斑块为研究对象,对土壤颗粒组成、土壤有机碳(soil organic carbon,SOC)、颗粒有机碳(particulate organic carbon,POC)及易氧化有机碳含量(labile organic carbon,LOC)进行分析。结果表明:1)各沙地生境发育中期灌丛斑块的SOC、POC、LOC及POC/SOC极显著高于草地斑块(P<0.01),而LOC/SOC在灌丛和草地斑块间差异性不显著;2)从丘间低地、固定沙地、半固定沙地到流动沙地,灌丛斑块及其对应的草地斑块SOC、POC、LOC及LOC/SOC均在固定沙地出现峰值,然后呈下降趋势,表明固定沙地及不同沙地生境发育中期的灌丛斑块和草地斑块是SOC、LOC及POC变化的临界点;3)相关分析发现,发育中期2种斑块SOC与POC及LOC间及SOC与LOC/SOC间均呈极显著相关(P<0.01),但土壤黏粉粒与各组分有机碳及其分配比率间没有显著相关,因此LOC和POC可作为表征草地退化最敏感的指标。研究可为沙化草原土壤恢复研究提供理论依据。
[ 吴旭东 , 宋乃平 , 潘军 ( 2016 ). 不同沙地生境下柠条灌丛化对草地土壤有机碳含量及分布的影响农业工程学报 , 32(10 ), 115 -121 .] Magsci [本文引用: 2] 为揭示不同沙地生境下灌丛化草地土壤有机碳及其组分的分布特征,以宁夏盐池县沙化草原不同沙地生境(丘间低地、固定沙地、半固定沙地、流动沙地)下不同发育期的柠条灌丛斑块及其对应的草地斑块为研究对象,对土壤颗粒组成、土壤有机碳(soil organic carbon,SOC)、颗粒有机碳(particulate organic carbon,POC)及易氧化有机碳含量(labile organic carbon,LOC)进行分析。结果表明:1)各沙地生境发育中期灌丛斑块的SOC、POC、LOC及POC/SOC极显著高于草地斑块(P<0.01),而LOC/SOC在灌丛和草地斑块间差异性不显著;2)从丘间低地、固定沙地、半固定沙地到流动沙地,灌丛斑块及其对应的草地斑块SOC、POC、LOC及LOC/SOC均在固定沙地出现峰值,然后呈下降趋势,表明固定沙地及不同沙地生境发育中期的灌丛斑块和草地斑块是SOC、LOC及POC变化的临界点;3)相关分析发现,发育中期2种斑块SOC与POC及LOC间及SOC与LOC/SOC间均呈极显著相关(P<0.01),但土壤黏粉粒与各组分有机碳及其分配比率间没有显著相关,因此LOC和POC可作为表征草地退化最敏感的指标。研究可为沙化草原土壤恢复研究提供理论依据。
[44] Wu ZY 1980 ). Vegetation of China . Science Press, Beijing. [本文引用: 1] [ 吴征镒 (1980 ). 中国植被 . 科学出版社, 北京.] [本文引用: 1] [45] Xie ZB Zhu JG Liu G Cadisch G Hasegawa T Chen CM Sun HF Tang HY Zeng Q 2007 ). Soil organic carbon stocks in China and changes from 1980s to 2000sGlobal Change Biology , 13, 1989 -2007 . [本文引用: 1] [46] Yang LC Li CB Ning Y Nie XQ Xu WH Zhou GY 2017 ). Carbon density and its spatial distribution in the Potentilla fruticosa dominated alpine shrub in Qinghai, ChinaChinese Journal of Plant Ecology 62 -70 . [本文引用: 1] [ 杨路存 , 李长斌 , 宁祎 , 聂秀青 , 徐文华 , 周国英 ( 2017 ). 青海高寒金露梅灌丛碳密度及其分配格局植物生态学报 , 41, 62 -70 .] [本文引用: 1] [47] Yang YH Fang JY Ma WH Smith P Mohammat A Wang SP Wang W 2010 ). Soil carbon stock and its changes in northern China’s grasslands from 1980s to 2000sGlobal Change Biology , 16, 3036 -3047 . [本文引用: 1] [48] Yang YH Fang JY Smith P Tang YH Chen AP Ji CJ Hu HF Rao S Tan K He JS 2009 ). Changes in topsoil carbon stock in the Tibetan grasslands between the 1980s and 2004Global Change Biology , 15, 2723 -2729 . [本文引用: 1] [49] Yang YH Fang JY Tang YH Ji CJ Zheng CY He JS Zhu BA 2008 ). Storage, patterns and controls of soil organic carbon in the Tibetan grasslandsGlobal Change Biology , 14, 1592 -1599 . [本文引用: 2] [50] Zhang Q Li JX Xu WT Xiong GM Xie ZQ 2017 ). Estimation of biomass allocation and carbon density of Rhododendron simsii shrubland in the subtropical mountainous areas of ChinaChinese Journal of Plant Ecology 43 -52 . [本文引用: 1] [ 张蔷 , 李家湘 , 徐文婷 , 熊高明 , 谢宗强 ( 2017 ). 中国亚热带山地杜鹃灌丛生物量分配及其碳密度估算植物生态学报 , 41, 43 -52 .] [本文引用: 1] [51] Zhang YQ Tang YH Jiang J Yang YH 2007 ). Characterizing the dynamics of soil organic carbon in grasslands on the Qinghai-Tibetan PlateauScience in China Series D: Earth Sciences , 50, 113 -120 . [本文引用: 2] [52] Zhao YN Du YY Ma YP Zhao YB Zhou YR Wang HM 2019 ). Soil organic carbon dynamics and their spatial prediction for anthropogenically introduced shrub encroachment in desert grassland of the Eastern Ningxia, ChinaChinese Journal of Applied Ecology , 30, 1927 -1935 . [本文引用: 1] [ 赵亚楠 , 杜艳艳 , 马彦平 , 赵延兵 , 周玉蓉 , 王红梅 ( 2019 ). 宁夏东部荒漠草原灌丛引入过程中土壤有机碳变化及其空间格局预测应用生态学报 , 30, 1927 -1935 ] [本文引用: 1] [53] Zhu CG Chen GC Zhou GY Han YJ Li JP 2007 ). Study on plant community diversity in valley shrub grassland of Qinghai Lake regionPratacultural Science , 24(3 ), 31 -35 . [本文引用: 1] [ 祝存冠 , 陈桂琛 , 周国英 , 韩友吉 , 李锦萍 ( 2007 ). 青海湖区河谷灌丛草地植被群落多样性研究草业科学 , 24(3 ), 31 -35 .] [本文引用: 1] Forest ecosystem carbon accumulation during a secondary succession in the Eastern Prealps of Italy
1
2008
... 本研究中, 低灌木植物盖度草地群落土壤碳库(6.031 kg·m
-2 )小于无灌木植物草地群落(7.215 kg·m
-2 ), 究其原因可能是西南地区水热条件好, 灌木生长较为迅速, 在增加土壤的碳输入的同时(
Fang et al ., 2003 ), 也会增加土壤向大气的碳输出(
Feng et al ., 2008 ); 当然也有可能这些次生灌木尚处在幼龄期还不能对土壤碳库有显著的影响(
Alberti et al ., 2008 ).无灌木植物草地群落绝大部分土壤碳(95.22%)分布于0-50 cm土层, 表层草本植物根系是土壤有机碳密度较高的主要原因, 而灌木根系会延伸至较深土层(
万昊等, 2015 );
Hughes等(2006) 和
陈蕾伊等(2014) 发现, 灌丛草原与无灌丛草原相比, 表层土壤碳储量无显著差异, 而深层土壤碳储量增加.因此, 高灌木植物盖度草地群落比无灌木植物草地群落具有更高的土壤碳密度, 随着灌木植物盖度的增大, 草地生态系统倾向于增加土壤深层生物量以及土壤碳库. ...
2
1996
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
... 根据样方调查资料, 本试验41个样地共记录323种草本植物, 55种灌木植物.根据草地群落灌木植物盖度, 可将这41个样地分成3类: (1)无灌木植物草地群落(I), 指灌木植物盖度为0, 仅由草本植物组成的草本群落, 共18个样地.根据中国草地类型分类系统(
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 )划分草地类型, 草地类型为暖性草丛, 中禾草组, 白茅草地型.(2)低灌木植物盖度草地群落(II), 指灌木植物盖度0-10%的草本群落, 共18个样地.草地类型为热性灌草丛, 中禾草组, 具灌木的扭黄茅草地型.(3)高灌木植物盖度草地群落(III), 即灌木植物盖度10%-30%的草本群落, 共5个样地.草地类型为暖性灌草丛, 中禾草组, 主要为具灌木的荩草型.随着草地群落灌木植物盖度的增加, 草地类型从草丛变为灌草丛, 灌木种类和数量增多, 植物物种也更加丰富(
表2 ). ...
2
1996
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
... 根据样方调查资料, 本试验41个样地共记录323种草本植物, 55种灌木植物.根据草地群落灌木植物盖度, 可将这41个样地分成3类: (1)无灌木植物草地群落(I), 指灌木植物盖度为0, 仅由草本植物组成的草本群落, 共18个样地.根据中国草地类型分类系统(
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 )划分草地类型, 草地类型为暖性草丛, 中禾草组, 白茅草地型.(2)低灌木植物盖度草地群落(II), 指灌木植物盖度0-10%的草本群落, 共18个样地.草地类型为热性灌草丛, 中禾草组, 具灌木的扭黄茅草地型.(3)高灌木植物盖度草地群落(III), 即灌木植物盖度10%-30%的草本群落, 共5个样地.草地类型为暖性灌草丛, 中禾草组, 主要为具灌木的荩草型.随着草地群落灌木植物盖度的增加, 草地类型从草丛变为灌草丛, 灌木种类和数量增多, 植物物种也更加丰富(
表2 ). ...
Mechanisms of shrubland expansion: Land use, climate or CO
2 ?
1
1995
... 草地群落中灌木的增加对土壤碳库的影响目前还未有一致的认识, 土壤碳库与灌木植物盖度的关系也有很大的不确定性.例如,
Hughes等(2006) 的结果表明, 草地演替过程中土壤碳库没有显著改变;
万昊等(2015) 对黄土高原植被演替过程中土壤碳库的研究表明, 草地土壤碳储量高于灌木地; 而
Archer等(1995) 、
Brantley和Young (2010) 、
陈蕾伊等(2014) 则认为, 随着草地群落灌木植物盖度增加土壤碳库也增加.这种研究结果不一致的原因, 也许与气候条件有关, 例如
Jackson等(2002) 在干旱地区的研究也显示, 草地群落土壤碳库随灌木盖度的增加而增加; 但在年降水量较大的地区, 草地土壤碳库随着灌木盖度的增加而降低.灌木根系对土壤碳库的影响是多方面的(
Matamala & Schlesinger, 2003 ), 有研究认为, 灌木将更多的生物量分配到地上而减少地下生物量分配, 分解腐烂后返还给土壤的有机碳也相对较少(
Goodale & Davidson, 2002 ); 但也有研究显示, 灌木增加了凋落物及根系向土壤的营养输入, 根系生长有利于土壤有机质形成和积累(
Knapp et al ., 2008 ). ...
Shrub expansion stimulates soil C and N storage along a coastal soil chronosequence
1
2010
... 草地群落中灌木的增加对土壤碳库的影响目前还未有一致的认识, 土壤碳库与灌木植物盖度的关系也有很大的不确定性.例如,
Hughes等(2006) 的结果表明, 草地演替过程中土壤碳库没有显著改变;
万昊等(2015) 对黄土高原植被演替过程中土壤碳库的研究表明, 草地土壤碳储量高于灌木地; 而
Archer等(1995) 、
Brantley和Young (2010) 、
陈蕾伊等(2014) 则认为, 随着草地群落灌木植物盖度增加土壤碳库也增加.这种研究结果不一致的原因, 也许与气候条件有关, 例如
Jackson等(2002) 在干旱地区的研究也显示, 草地群落土壤碳库随灌木盖度的增加而增加; 但在年降水量较大的地区, 草地土壤碳库随着灌木盖度的增加而降低.灌木根系对土壤碳库的影响是多方面的(
Matamala & Schlesinger, 2003 ), 有研究认为, 灌木将更多的生物量分配到地上而减少地下生物量分配, 分解腐烂后返还给土壤的有机碳也相对较少(
Goodale & Davidson, 2002 ); 但也有研究显示, 灌木增加了凋落物及根系向土壤的营养输入, 根系生长有利于土壤有机质形成和积累(
Knapp et al ., 2008 ). ...
草地灌丛化研究进展
2
2016
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
... ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
草地灌丛化研究进展
2
2016
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
... ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
灌丛化草原: 一种新的植被景观
2014
灌丛化草原: 一种新的植被景观
2014
Carbon isotopes reveal soil organic matter dynamics following arid land shrub expansion
1
1997
... 生物量作为碳库的重要组分, 是量化生态系统碳周转的重要指标(
Ferlan et al ., 2011 ).木本植物比草本植物具有更多的生物量(
Seastedt & Ramundo, 1990 ), 灌丛地上净初级生产力的增加导致地上碳储量的增加, 并使地下碳库有增加的潜力(
Norris et al ., 2001 ;
Smith & Johnson, 2003 ).因此, 在草地向灌丛和森林演替过程中, 草地群落灌木植物盖度的增大对植被碳库有重要影响.有研究表明, 中国草地植被碳密度平均值范围为0.216-1.148 kg·m
-2 (
Ni, 2002 ;
Fang et al ., 2007 ;
Piao et al ., 2007 ).利用遥感影像建立的植被生物量估算模型计算得到的中国草地植被生物量平均碳密度为0.315 kg·m
-2 (
朴世龙等, 2004 ).本研究中, 无灌木植物草地群落的植被层碳密度为0.304 kg·m
-2 , 与前人研究结果相近.然而低灌木植物盖度草地群落的植被碳密度为1.407 kg·m
-2 , 高灌木植物盖度草地群落的植被碳密度为1.574 kg·m
-2 , 有灌木植物草地群落碳密度远大于无灌木植物草地群落.低灌木植物盖度草地群落相对于无灌木植物草地群落既有草本层的凋落物, 同时也有灌木层的凋落物的贡献(
聂秀青等, 2018 ), 凋落物碳库增加.
Connin等(1997) 研究表明, 灌木相对于多年生草本植物具有更深广发达的根系, 地下根系生物量增加.因此, 灌木植物盖度增大, 植物地上生物量会有所增加, 进而影响草地群落的植被碳库, 忽略灌木植物盖度增加过程将导致对碳库的低估. ...
我国草地面积有多大
1
2018
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
我国草地面积有多大
1
2018
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
Terrestrial vegetation carbon sinks in China, 1981-2000
1
2007
... 生物量作为碳库的重要组分, 是量化生态系统碳周转的重要指标(
Ferlan et al ., 2011 ).木本植物比草本植物具有更多的生物量(
Seastedt & Ramundo, 1990 ), 灌丛地上净初级生产力的增加导致地上碳储量的增加, 并使地下碳库有增加的潜力(
Norris et al ., 2001 ;
Smith & Johnson, 2003 ).因此, 在草地向灌丛和森林演替过程中, 草地群落灌木植物盖度的增大对植被碳库有重要影响.有研究表明, 中国草地植被碳密度平均值范围为0.216-1.148 kg·m
-2 (
Ni, 2002 ;
Fang et al ., 2007 ;
Piao et al ., 2007 ).利用遥感影像建立的植被生物量估算模型计算得到的中国草地植被生物量平均碳密度为0.315 kg·m
-2 (
朴世龙等, 2004 ).本研究中, 无灌木植物草地群落的植被层碳密度为0.304 kg·m
-2 , 与前人研究结果相近.然而低灌木植物盖度草地群落的植被碳密度为1.407 kg·m
-2 , 高灌木植物盖度草地群落的植被碳密度为1.574 kg·m
-2 , 有灌木植物草地群落碳密度远大于无灌木植物草地群落.低灌木植物盖度草地群落相对于无灌木植物草地群落既有草本层的凋落物, 同时也有灌木层的凋落物的贡献(
聂秀青等, 2018 ), 凋落物碳库增加.
Connin等(1997) 研究表明, 灌木相对于多年生草本植物具有更深广发达的根系, 地下根系生物量增加.因此, 灌木植物盖度增大, 植物地上生物量会有所增加, 进而影响草地群落的植被碳库, 忽略灌木植物盖度增加过程将导致对碳库的低估. ...
Increasing net primary production in China from 1982 to 1999
1
2003
... 本研究中, 低灌木植物盖度草地群落土壤碳库(6.031 kg·m
-2 )小于无灌木植物草地群落(7.215 kg·m
-2 ), 究其原因可能是西南地区水热条件好, 灌木生长较为迅速, 在增加土壤的碳输入的同时(
Fang et al ., 2003 ), 也会增加土壤向大气的碳输出(
Feng et al ., 2008 ); 当然也有可能这些次生灌木尚处在幼龄期还不能对土壤碳库有显著的影响(
Alberti et al ., 2008 ).无灌木植物草地群落绝大部分土壤碳(95.22%)分布于0-50 cm土层, 表层草本植物根系是土壤有机碳密度较高的主要原因, 而灌木根系会延伸至较深土层(
万昊等, 2015 );
Hughes等(2006) 和
陈蕾伊等(2014) 发现, 灌丛草原与无灌丛草原相比, 表层土壤碳储量无显著差异, 而深层土壤碳储量增加.因此, 高灌木植物盖度草地群落比无灌木植物草地群落具有更高的土壤碳密度, 随着灌木植物盖度的增大, 草地生态系统倾向于增加土壤深层生物量以及土壤碳库. ...
中国草地生态系统碳库及其变化
1
2010
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
中国草地生态系统碳库及其变化
1
2010
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
Increased cuticular carbon sequestration and lignin oxidation in response to soil warming
1
2008
... 本研究中, 低灌木植物盖度草地群落土壤碳库(6.031 kg·m
-2 )小于无灌木植物草地群落(7.215 kg·m
-2 ), 究其原因可能是西南地区水热条件好, 灌木生长较为迅速, 在增加土壤的碳输入的同时(
Fang et al ., 2003 ), 也会增加土壤向大气的碳输出(
Feng et al ., 2008 ); 当然也有可能这些次生灌木尚处在幼龄期还不能对土壤碳库有显著的影响(
Alberti et al ., 2008 ).无灌木植物草地群落绝大部分土壤碳(95.22%)分布于0-50 cm土层, 表层草本植物根系是土壤有机碳密度较高的主要原因, 而灌木根系会延伸至较深土层(
万昊等, 2015 );
Hughes等(2006) 和
陈蕾伊等(2014) 发现, 灌丛草原与无灌丛草原相比, 表层土壤碳储量无显著差异, 而深层土壤碳储量增加.因此, 高灌木植物盖度草地群落比无灌木植物草地群落具有更高的土壤碳密度, 随着灌木植物盖度的增大, 草地生态系统倾向于增加土壤深层生物量以及土壤碳库. ...
Comparing carbon fluxes between different stages of secondary succession of a karst grassland
2
2011
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
... 生物量作为碳库的重要组分, 是量化生态系统碳周转的重要指标(
Ferlan et al ., 2011 ).木本植物比草本植物具有更多的生物量(
Seastedt & Ramundo, 1990 ), 灌丛地上净初级生产力的增加导致地上碳储量的增加, 并使地下碳库有增加的潜力(
Norris et al ., 2001 ;
Smith & Johnson, 2003 ).因此, 在草地向灌丛和森林演替过程中, 草地群落灌木植物盖度的增大对植被碳库有重要影响.有研究表明, 中国草地植被碳密度平均值范围为0.216-1.148 kg·m
-2 (
Ni, 2002 ;
Fang et al ., 2007 ;
Piao et al ., 2007 ).利用遥感影像建立的植被生物量估算模型计算得到的中国草地植被生物量平均碳密度为0.315 kg·m
-2 (
朴世龙等, 2004 ).本研究中, 无灌木植物草地群落的植被层碳密度为0.304 kg·m
-2 , 与前人研究结果相近.然而低灌木植物盖度草地群落的植被碳密度为1.407 kg·m
-2 , 高灌木植物盖度草地群落的植被碳密度为1.574 kg·m
-2 , 有灌木植物草地群落碳密度远大于无灌木植物草地群落.低灌木植物盖度草地群落相对于无灌木植物草地群落既有草本层的凋落物, 同时也有灌木层的凋落物的贡献(
聂秀青等, 2018 ), 凋落物碳库增加.
Connin等(1997) 研究表明, 灌木相对于多年生草本植物具有更深广发达的根系, 地下根系生物量增加.因此, 灌木植物盖度增大, 植物地上生物量会有所增加, 进而影响草地群落的植被碳库, 忽略灌木植物盖度增加过程将导致对碳库的低估. ...
Carbon cycle: Uncertain sinks in the shrubs
2
2002
... 随着草地群落灌木植物盖度增加, 灌丛草地地上净初级生产力增加(
Reich et al ., 2001 ;
Hughes et al ., 2006 ;
Knapp et al ., 2008 ), 更多的生物量被分配到地上部分(
Jackson et al ., 1997 ;
Goodale & Davidson, 2002 ), 植被地上碳库增加.但当灌木植物盖度增加到一定程度后, 灌木植株可能会遮挡下层草本植物的光照, 减少草本植物生物量, 导致植物地上碳库反而会低于低灌木植物盖度草地群落.因此, 草地群落中的灌木植物盖度与植物地上碳储量之间存在一定的不确定性, 在大尺度的碳储量评估过程中需要给予高度重视. ...
... 草地群落中灌木的增加对土壤碳库的影响目前还未有一致的认识, 土壤碳库与灌木植物盖度的关系也有很大的不确定性.例如,
Hughes等(2006) 的结果表明, 草地演替过程中土壤碳库没有显著改变;
万昊等(2015) 对黄土高原植被演替过程中土壤碳库的研究表明, 草地土壤碳储量高于灌木地; 而
Archer等(1995) 、
Brantley和Young (2010) 、
陈蕾伊等(2014) 则认为, 随着草地群落灌木植物盖度增加土壤碳库也增加.这种研究结果不一致的原因, 也许与气候条件有关, 例如
Jackson等(2002) 在干旱地区的研究也显示, 草地群落土壤碳库随灌木盖度的增加而增加; 但在年降水量较大的地区, 草地土壤碳库随着灌木盖度的增加而降低.灌木根系对土壤碳库的影响是多方面的(
Matamala & Schlesinger, 2003 ), 有研究认为, 灌木将更多的生物量分配到地上而减少地下生物量分配, 分解腐烂后返还给土壤的有机碳也相对较少(
Goodale & Davidson, 2002 ); 但也有研究显示, 灌木增加了凋落物及根系向土壤的营养输入, 根系生长有利于土壤有机质形成和积累(
Knapp et al ., 2008 ). ...
Positive effects of shrubs on plant species diversity do not change along a gradient in grazing pressure in an arid shrubland
1
2012
... 西南地区草地多属次生性质, 是以多年生草本植物为主体, 间混少量乔木或灌木的草地类型.在停止人为干扰、水土条件良好的地段, 经过一定时间的恢复后可能回到森林顶极群落(
吴征镒, 1980 ).在顺行演替过程中, 灌丛草地比无灌木植物草地群落维持了更高的物种丰富度(
Howard et al ., 2012 ), 草地向灌丛演替有利于草地群落增加生物量, 促进生态系统碳固定(
Pacala et al ., 2001 ); 随着灌木植物盖度增加, 草地向森林演替过程中生态系统固碳能力改变(
Post & Kwon, 2000 ;
Jackson et al ., 2002 ;
McKinley & Blair, 2008 ).有研究显示, 土壤有机碳密度在青藏高原高寒草地为6.5 kg·m
-2 、北方草地为8.5 kg·m
-2 (
Yang et al ., 2008 ), 而本研究实测的西南地区无灌木植物草地群落(7.215 kg·m
-2 )和低灌木植物盖度草地群落(6.031 kg·m
-2 )的土壤碳密度介于两者之间, 而高灌木植物盖度草地群落(9.735 kg·m
-2 )显著高于高寒草地和北方草地;
Ni (2002) 根据全球土壤碳氮数据库估算的中国草地土壤有机碳密度为13.2 kg·m
-2 , 显著高于本研究实测得到的西南地区草地的土壤平均碳密度.这表明以北方草地和高寒草地的数据为基础将会低估中国草地生态系统碳库, 以全球植被和土壤数据库为基础则会高估中国草地生态系统碳库. ...
中国西南地区的草地资源分析
1
2012
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
中国西南地区的草地资源分析
1
2012
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
Changes in aboveground primary production and carbon and nitrogen pools accompanying woody plant encroachment in a temperate savanna
3
2006
... 随着草地群落灌木植物盖度增加, 灌丛草地地上净初级生产力增加(
Reich et al ., 2001 ;
Hughes et al ., 2006 ;
Knapp et al ., 2008 ), 更多的生物量被分配到地上部分(
Jackson et al ., 1997 ;
Goodale & Davidson, 2002 ), 植被地上碳库增加.但当灌木植物盖度增加到一定程度后, 灌木植株可能会遮挡下层草本植物的光照, 减少草本植物生物量, 导致植物地上碳库反而会低于低灌木植物盖度草地群落.因此, 草地群落中的灌木植物盖度与植物地上碳储量之间存在一定的不确定性, 在大尺度的碳储量评估过程中需要给予高度重视. ...
... 草地群落中灌木的增加对土壤碳库的影响目前还未有一致的认识, 土壤碳库与灌木植物盖度的关系也有很大的不确定性.例如,
Hughes等(2006) 的结果表明, 草地演替过程中土壤碳库没有显著改变;
万昊等(2015) 对黄土高原植被演替过程中土壤碳库的研究表明, 草地土壤碳储量高于灌木地; 而
Archer等(1995) 、
Brantley和Young (2010) 、
陈蕾伊等(2014) 则认为, 随着草地群落灌木植物盖度增加土壤碳库也增加.这种研究结果不一致的原因, 也许与气候条件有关, 例如
Jackson等(2002) 在干旱地区的研究也显示, 草地群落土壤碳库随灌木盖度的增加而增加; 但在年降水量较大的地区, 草地土壤碳库随着灌木盖度的增加而降低.灌木根系对土壤碳库的影响是多方面的(
Matamala & Schlesinger, 2003 ), 有研究认为, 灌木将更多的生物量分配到地上而减少地下生物量分配, 分解腐烂后返还给土壤的有机碳也相对较少(
Goodale & Davidson, 2002 ); 但也有研究显示, 灌木增加了凋落物及根系向土壤的营养输入, 根系生长有利于土壤有机质形成和积累(
Knapp et al ., 2008 ). ...
... 本研究中, 低灌木植物盖度草地群落土壤碳库(6.031 kg·m
-2 )小于无灌木植物草地群落(7.215 kg·m
-2 ), 究其原因可能是西南地区水热条件好, 灌木生长较为迅速, 在增加土壤的碳输入的同时(
Fang et al ., 2003 ), 也会增加土壤向大气的碳输出(
Feng et al ., 2008 ); 当然也有可能这些次生灌木尚处在幼龄期还不能对土壤碳库有显著的影响(
Alberti et al ., 2008 ).无灌木植物草地群落绝大部分土壤碳(95.22%)分布于0-50 cm土层, 表层草本植物根系是土壤有机碳密度较高的主要原因, 而灌木根系会延伸至较深土层(
万昊等, 2015 );
Hughes等(2006) 和
陈蕾伊等(2014) 发现, 灌丛草原与无灌丛草原相比, 表层土壤碳储量无显著差异, 而深层土壤碳储量增加.因此, 高灌木植物盖度草地群落比无灌木植物草地群落具有更高的土壤碳密度, 随着灌木植物盖度的增大, 草地生态系统倾向于增加土壤深层生物量以及土壤碳库. ...
Ecosystem carbon loss with woody plant invasion of grasslands
2
2002
... 草地群落中灌木的增加对土壤碳库的影响目前还未有一致的认识, 土壤碳库与灌木植物盖度的关系也有很大的不确定性.例如,
Hughes等(2006) 的结果表明, 草地演替过程中土壤碳库没有显著改变;
万昊等(2015) 对黄土高原植被演替过程中土壤碳库的研究表明, 草地土壤碳储量高于灌木地; 而
Archer等(1995) 、
Brantley和Young (2010) 、
陈蕾伊等(2014) 则认为, 随着草地群落灌木植物盖度增加土壤碳库也增加.这种研究结果不一致的原因, 也许与气候条件有关, 例如
Jackson等(2002) 在干旱地区的研究也显示, 草地群落土壤碳库随灌木盖度的增加而增加; 但在年降水量较大的地区, 草地土壤碳库随着灌木盖度的增加而降低.灌木根系对土壤碳库的影响是多方面的(
Matamala & Schlesinger, 2003 ), 有研究认为, 灌木将更多的生物量分配到地上而减少地下生物量分配, 分解腐烂后返还给土壤的有机碳也相对较少(
Goodale & Davidson, 2002 ); 但也有研究显示, 灌木增加了凋落物及根系向土壤的营养输入, 根系生长有利于土壤有机质形成和积累(
Knapp et al ., 2008 ). ...
... 西南地区草地多属次生性质, 是以多年生草本植物为主体, 间混少量乔木或灌木的草地类型.在停止人为干扰、水土条件良好的地段, 经过一定时间的恢复后可能回到森林顶极群落(
吴征镒, 1980 ).在顺行演替过程中, 灌丛草地比无灌木植物草地群落维持了更高的物种丰富度(
Howard et al ., 2012 ), 草地向灌丛演替有利于草地群落增加生物量, 促进生态系统碳固定(
Pacala et al ., 2001 ); 随着灌木植物盖度增加, 草地向森林演替过程中生态系统固碳能力改变(
Post & Kwon, 2000 ;
Jackson et al ., 2002 ;
McKinley & Blair, 2008 ).有研究显示, 土壤有机碳密度在青藏高原高寒草地为6.5 kg·m
-2 、北方草地为8.5 kg·m
-2 (
Yang et al ., 2008 ), 而本研究实测的西南地区无灌木植物草地群落(7.215 kg·m
-2 )和低灌木植物盖度草地群落(6.031 kg·m
-2 )的土壤碳密度介于两者之间, 而高灌木植物盖度草地群落(9.735 kg·m
-2 )显著高于高寒草地和北方草地;
Ni (2002) 根据全球土壤碳氮数据库估算的中国草地土壤有机碳密度为13.2 kg·m
-2 , 显著高于本研究实测得到的西南地区草地的土壤平均碳密度.这表明以北方草地和高寒草地的数据为基础将会低估中国草地生态系统碳库, 以全球植被和土壤数据库为基础则会高估中国草地生态系统碳库. ...
A global budget for fine root biomass, surface area, and nutrient contents
1
1997
... 随着草地群落灌木植物盖度增加, 灌丛草地地上净初级生产力增加(
Reich et al ., 2001 ;
Hughes et al ., 2006 ;
Knapp et al ., 2008 ), 更多的生物量被分配到地上部分(
Jackson et al ., 1997 ;
Goodale & Davidson, 2002 ), 植被地上碳库增加.但当灌木植物盖度增加到一定程度后, 灌木植株可能会遮挡下层草本植物的光照, 减少草本植物生物量, 导致植物地上碳库反而会低于低灌木植物盖度草地群落.因此, 草地群落中的灌木植物盖度与植物地上碳储量之间存在一定的不确定性, 在大尺度的碳储量评估过程中需要给予高度重视. ...
Shrub encroachment in North American grasslands: Shifts in growth form dominance rapidly alters control of ecosystem carbon inputs
3
2008
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
... 随着草地群落灌木植物盖度增加, 灌丛草地地上净初级生产力增加(
Reich et al ., 2001 ;
Hughes et al ., 2006 ;
Knapp et al ., 2008 ), 更多的生物量被分配到地上部分(
Jackson et al ., 1997 ;
Goodale & Davidson, 2002 ), 植被地上碳库增加.但当灌木植物盖度增加到一定程度后, 灌木植株可能会遮挡下层草本植物的光照, 减少草本植物生物量, 导致植物地上碳库反而会低于低灌木植物盖度草地群落.因此, 草地群落中的灌木植物盖度与植物地上碳储量之间存在一定的不确定性, 在大尺度的碳储量评估过程中需要给予高度重视. ...
... 草地群落中灌木的增加对土壤碳库的影响目前还未有一致的认识, 土壤碳库与灌木植物盖度的关系也有很大的不确定性.例如,
Hughes等(2006) 的结果表明, 草地演替过程中土壤碳库没有显著改变;
万昊等(2015) 对黄土高原植被演替过程中土壤碳库的研究表明, 草地土壤碳储量高于灌木地; 而
Archer等(1995) 、
Brantley和Young (2010) 、
陈蕾伊等(2014) 则认为, 随着草地群落灌木植物盖度增加土壤碳库也增加.这种研究结果不一致的原因, 也许与气候条件有关, 例如
Jackson等(2002) 在干旱地区的研究也显示, 草地群落土壤碳库随灌木盖度的增加而增加; 但在年降水量较大的地区, 草地土壤碳库随着灌木盖度的增加而降低.灌木根系对土壤碳库的影响是多方面的(
Matamala & Schlesinger, 2003 ), 有研究认为, 灌木将更多的生物量分配到地上而减少地下生物量分配, 分解腐烂后返还给土壤的有机碳也相对较少(
Goodale & Davidson, 2002 ); 但也有研究显示, 灌木增加了凋落物及根系向土壤的营养输入, 根系生长有利于土壤有机质形成和积累(
Knapp et al ., 2008 ). ...
中国北方草地退化及其防治对策
1
1997
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
中国北方草地退化及其防治对策
1
1997
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
Impacts of fine root turnover on forest NPP and soil C sequestration potential
1
2003
... 草地群落中灌木的增加对土壤碳库的影响目前还未有一致的认识, 土壤碳库与灌木植物盖度的关系也有很大的不确定性.例如,
Hughes等(2006) 的结果表明, 草地演替过程中土壤碳库没有显著改变;
万昊等(2015) 对黄土高原植被演替过程中土壤碳库的研究表明, 草地土壤碳储量高于灌木地; 而
Archer等(1995) 、
Brantley和Young (2010) 、
陈蕾伊等(2014) 则认为, 随着草地群落灌木植物盖度增加土壤碳库也增加.这种研究结果不一致的原因, 也许与气候条件有关, 例如
Jackson等(2002) 在干旱地区的研究也显示, 草地群落土壤碳库随灌木盖度的增加而增加; 但在年降水量较大的地区, 草地土壤碳库随着灌木盖度的增加而降低.灌木根系对土壤碳库的影响是多方面的(
Matamala & Schlesinger, 2003 ), 有研究认为, 灌木将更多的生物量分配到地上而减少地下生物量分配, 分解腐烂后返还给土壤的有机碳也相对较少(
Goodale & Davidson, 2002 ); 但也有研究显示, 灌木增加了凋落物及根系向土壤的营养输入, 根系生长有利于土壤有机质形成和积累(
Knapp et al ., 2008 ). ...
Woody plant encroachment by
Juniperus virginiana in a mesic native grassland promotes rapid carbon and nitrogen accrual
1
2008
... 西南地区草地多属次生性质, 是以多年生草本植物为主体, 间混少量乔木或灌木的草地类型.在停止人为干扰、水土条件良好的地段, 经过一定时间的恢复后可能回到森林顶极群落(
吴征镒, 1980 ).在顺行演替过程中, 灌丛草地比无灌木植物草地群落维持了更高的物种丰富度(
Howard et al ., 2012 ), 草地向灌丛演替有利于草地群落增加生物量, 促进生态系统碳固定(
Pacala et al ., 2001 ); 随着灌木植物盖度增加, 草地向森林演替过程中生态系统固碳能力改变(
Post & Kwon, 2000 ;
Jackson et al ., 2002 ;
McKinley & Blair, 2008 ).有研究显示, 土壤有机碳密度在青藏高原高寒草地为6.5 kg·m
-2 、北方草地为8.5 kg·m
-2 (
Yang et al ., 2008 ), 而本研究实测的西南地区无灌木植物草地群落(7.215 kg·m
-2 )和低灌木植物盖度草地群落(6.031 kg·m
-2 )的土壤碳密度介于两者之间, 而高灌木植物盖度草地群落(9.735 kg·m
-2 )显著高于高寒草地和北方草地;
Ni (2002) 根据全球土壤碳氮数据库估算的中国草地土壤有机碳密度为13.2 kg·m
-2 , 显著高于本研究实测得到的西南地区草地的土壤平均碳密度.这表明以北方草地和高寒草地的数据为基础将会低估中国草地生态系统碳库, 以全球植被和土壤数据库为基础则会高估中国草地生态系统碳库. ...
Carbon storage in grasslands of China
2
2002
... 生物量作为碳库的重要组分, 是量化生态系统碳周转的重要指标(
Ferlan et al ., 2011 ).木本植物比草本植物具有更多的生物量(
Seastedt & Ramundo, 1990 ), 灌丛地上净初级生产力的增加导致地上碳储量的增加, 并使地下碳库有增加的潜力(
Norris et al ., 2001 ;
Smith & Johnson, 2003 ).因此, 在草地向灌丛和森林演替过程中, 草地群落灌木植物盖度的增大对植被碳库有重要影响.有研究表明, 中国草地植被碳密度平均值范围为0.216-1.148 kg·m
-2 (
Ni, 2002 ;
Fang et al ., 2007 ;
Piao et al ., 2007 ).利用遥感影像建立的植被生物量估算模型计算得到的中国草地植被生物量平均碳密度为0.315 kg·m
-2 (
朴世龙等, 2004 ).本研究中, 无灌木植物草地群落的植被层碳密度为0.304 kg·m
-2 , 与前人研究结果相近.然而低灌木植物盖度草地群落的植被碳密度为1.407 kg·m
-2 , 高灌木植物盖度草地群落的植被碳密度为1.574 kg·m
-2 , 有灌木植物草地群落碳密度远大于无灌木植物草地群落.低灌木植物盖度草地群落相对于无灌木植物草地群落既有草本层的凋落物, 同时也有灌木层的凋落物的贡献(
聂秀青等, 2018 ), 凋落物碳库增加.
Connin等(1997) 研究表明, 灌木相对于多年生草本植物具有更深广发达的根系, 地下根系生物量增加.因此, 灌木植物盖度增大, 植物地上生物量会有所增加, 进而影响草地群落的植被碳库, 忽略灌木植物盖度增加过程将导致对碳库的低估. ...
... 西南地区草地多属次生性质, 是以多年生草本植物为主体, 间混少量乔木或灌木的草地类型.在停止人为干扰、水土条件良好的地段, 经过一定时间的恢复后可能回到森林顶极群落(
吴征镒, 1980 ).在顺行演替过程中, 灌丛草地比无灌木植物草地群落维持了更高的物种丰富度(
Howard et al ., 2012 ), 草地向灌丛演替有利于草地群落增加生物量, 促进生态系统碳固定(
Pacala et al ., 2001 ); 随着灌木植物盖度增加, 草地向森林演替过程中生态系统固碳能力改变(
Post & Kwon, 2000 ;
Jackson et al ., 2002 ;
McKinley & Blair, 2008 ).有研究显示, 土壤有机碳密度在青藏高原高寒草地为6.5 kg·m
-2 、北方草地为8.5 kg·m
-2 (
Yang et al ., 2008 ), 而本研究实测的西南地区无灌木植物草地群落(7.215 kg·m
-2 )和低灌木植物盖度草地群落(6.031 kg·m
-2 )的土壤碳密度介于两者之间, 而高灌木植物盖度草地群落(9.735 kg·m
-2 )显著高于高寒草地和北方草地;
Ni (2002) 根据全球土壤碳氮数据库估算的中国草地土壤有机碳密度为13.2 kg·m
-2 , 显著高于本研究实测得到的西南地区草地的土壤平均碳密度.这表明以北方草地和高寒草地的数据为基础将会低估中国草地生态系统碳库, 以全球植被和土壤数据库为基础则会高估中国草地生态系统碳库. ...
青藏高原高寒灌丛生态系统草本层生物量分配格局
2
2018
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
... 生物量作为碳库的重要组分, 是量化生态系统碳周转的重要指标(
Ferlan et al ., 2011 ).木本植物比草本植物具有更多的生物量(
Seastedt & Ramundo, 1990 ), 灌丛地上净初级生产力的增加导致地上碳储量的增加, 并使地下碳库有增加的潜力(
Norris et al ., 2001 ;
Smith & Johnson, 2003 ).因此, 在草地向灌丛和森林演替过程中, 草地群落灌木植物盖度的增大对植被碳库有重要影响.有研究表明, 中国草地植被碳密度平均值范围为0.216-1.148 kg·m
-2 (
Ni, 2002 ;
Fang et al ., 2007 ;
Piao et al ., 2007 ).利用遥感影像建立的植被生物量估算模型计算得到的中国草地植被生物量平均碳密度为0.315 kg·m
-2 (
朴世龙等, 2004 ).本研究中, 无灌木植物草地群落的植被层碳密度为0.304 kg·m
-2 , 与前人研究结果相近.然而低灌木植物盖度草地群落的植被碳密度为1.407 kg·m
-2 , 高灌木植物盖度草地群落的植被碳密度为1.574 kg·m
-2 , 有灌木植物草地群落碳密度远大于无灌木植物草地群落.低灌木植物盖度草地群落相对于无灌木植物草地群落既有草本层的凋落物, 同时也有灌木层的凋落物的贡献(
聂秀青等, 2018 ), 凋落物碳库增加.
Connin等(1997) 研究表明, 灌木相对于多年生草本植物具有更深广发达的根系, 地下根系生物量增加.因此, 灌木植物盖度增大, 植物地上生物量会有所增加, 进而影响草地群落的植被碳库, 忽略灌木植物盖度增加过程将导致对碳库的低估. ...
青藏高原高寒灌丛生态系统草本层生物量分配格局
2
2018
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
... 生物量作为碳库的重要组分, 是量化生态系统碳周转的重要指标(
Ferlan et al ., 2011 ).木本植物比草本植物具有更多的生物量(
Seastedt & Ramundo, 1990 ), 灌丛地上净初级生产力的增加导致地上碳储量的增加, 并使地下碳库有增加的潜力(
Norris et al ., 2001 ;
Smith & Johnson, 2003 ).因此, 在草地向灌丛和森林演替过程中, 草地群落灌木植物盖度的增大对植被碳库有重要影响.有研究表明, 中国草地植被碳密度平均值范围为0.216-1.148 kg·m
-2 (
Ni, 2002 ;
Fang et al ., 2007 ;
Piao et al ., 2007 ).利用遥感影像建立的植被生物量估算模型计算得到的中国草地植被生物量平均碳密度为0.315 kg·m
-2 (
朴世龙等, 2004 ).本研究中, 无灌木植物草地群落的植被层碳密度为0.304 kg·m
-2 , 与前人研究结果相近.然而低灌木植物盖度草地群落的植被碳密度为1.407 kg·m
-2 , 高灌木植物盖度草地群落的植被碳密度为1.574 kg·m
-2 , 有灌木植物草地群落碳密度远大于无灌木植物草地群落.低灌木植物盖度草地群落相对于无灌木植物草地群落既有草本层的凋落物, 同时也有灌木层的凋落物的贡献(
聂秀青等, 2018 ), 凋落物碳库增加.
Connin等(1997) 研究表明, 灌木相对于多年生草本植物具有更深广发达的根系, 地下根系生物量增加.因此, 灌木植物盖度增大, 植物地上生物量会有所增加, 进而影响草地群落的植被碳库, 忽略灌木植物盖度增加过程将导致对碳库的低估. ...
Above- and belowground biomass allocation in shrub biomes across the northeast Tibetan Plateau
1
2016
... 所有草本、灌木和凋落物生物量样品在65 ℃烘箱烘干至恒质量(
Perkins & Owens, 2003 ;
Nie et al ., 2016 ;
杨路存等, 2017 ), 测定生物量干质量; 土壤样品风干后过2 mm的土壤筛, 用静电吸附(经摩擦过的塑料卡片或玻璃棒)和手动方法去除根系.随后所有样品用球磨仪研磨后过100目筛, 得到的样品用于指标分析, 3次重复.植物和土壤样品碳含量用元素分析仪(Vario MAX, Elementar, Hanau, Germany)测定. ...
Assessing changes in biomass, productivity, and C and N stores following
Juniperus virginiana forest expansion into tallgrass prairie
1
2001
... 生物量作为碳库的重要组分, 是量化生态系统碳周转的重要指标(
Ferlan et al ., 2011 ).木本植物比草本植物具有更多的生物量(
Seastedt & Ramundo, 1990 ), 灌丛地上净初级生产力的增加导致地上碳储量的增加, 并使地下碳库有增加的潜力(
Norris et al ., 2001 ;
Smith & Johnson, 2003 ).因此, 在草地向灌丛和森林演替过程中, 草地群落灌木植物盖度的增大对植被碳库有重要影响.有研究表明, 中国草地植被碳密度平均值范围为0.216-1.148 kg·m
-2 (
Ni, 2002 ;
Fang et al ., 2007 ;
Piao et al ., 2007 ).利用遥感影像建立的植被生物量估算模型计算得到的中国草地植被生物量平均碳密度为0.315 kg·m
-2 (
朴世龙等, 2004 ).本研究中, 无灌木植物草地群落的植被层碳密度为0.304 kg·m
-2 , 与前人研究结果相近.然而低灌木植物盖度草地群落的植被碳密度为1.407 kg·m
-2 , 高灌木植物盖度草地群落的植被碳密度为1.574 kg·m
-2 , 有灌木植物草地群落碳密度远大于无灌木植物草地群落.低灌木植物盖度草地群落相对于无灌木植物草地群落既有草本层的凋落物, 同时也有灌木层的凋落物的贡献(
聂秀青等, 2018 ), 凋落物碳库增加.
Connin等(1997) 研究表明, 灌木相对于多年生草本植物具有更深广发达的根系, 地下根系生物量增加.因此, 灌木植物盖度增大, 植物地上生物量会有所增加, 进而影响草地群落的植被碳库, 忽略灌木植物盖度增加过程将导致对碳库的低估. ...
Consistent land- and atmosphere-based U.S. carbon sink estimates
1
2001
... 西南地区草地多属次生性质, 是以多年生草本植物为主体, 间混少量乔木或灌木的草地类型.在停止人为干扰、水土条件良好的地段, 经过一定时间的恢复后可能回到森林顶极群落(
吴征镒, 1980 ).在顺行演替过程中, 灌丛草地比无灌木植物草地群落维持了更高的物种丰富度(
Howard et al ., 2012 ), 草地向灌丛演替有利于草地群落增加生物量, 促进生态系统碳固定(
Pacala et al ., 2001 ); 随着灌木植物盖度增加, 草地向森林演替过程中生态系统固碳能力改变(
Post & Kwon, 2000 ;
Jackson et al ., 2002 ;
McKinley & Blair, 2008 ).有研究显示, 土壤有机碳密度在青藏高原高寒草地为6.5 kg·m
-2 、北方草地为8.5 kg·m
-2 (
Yang et al ., 2008 ), 而本研究实测的西南地区无灌木植物草地群落(7.215 kg·m
-2 )和低灌木植物盖度草地群落(6.031 kg·m
-2 )的土壤碳密度介于两者之间, 而高灌木植物盖度草地群落(9.735 kg·m
-2 )显著高于高寒草地和北方草地;
Ni (2002) 根据全球土壤碳氮数据库估算的中国草地土壤有机碳密度为13.2 kg·m
-2 , 显著高于本研究实测得到的西南地区草地的土壤平均碳密度.这表明以北方草地和高寒草地的数据为基础将会低估中国草地生态系统碳库, 以全球植被和土壤数据库为基础则会高估中国草地生态系统碳库. ...
Growth and biomass allocation of shrub and grass seedlings in response to predicted changes in precipitation seasonality
1
2003
... 所有草本、灌木和凋落物生物量样品在65 ℃烘箱烘干至恒质量(
Perkins & Owens, 2003 ;
Nie et al ., 2016 ;
杨路存等, 2017 ), 测定生物量干质量; 土壤样品风干后过2 mm的土壤筛, 用静电吸附(经摩擦过的塑料卡片或玻璃棒)和手动方法去除根系.随后所有样品用球磨仪研磨后过100目筛, 得到的样品用于指标分析, 3次重复.植物和土壤样品碳含量用元素分析仪(Vario MAX, Elementar, Hanau, Germany)测定. ...
The carbon balance of terrestrial ecosystems in China
1
2009
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
中国草地植被生物量及其空间分布格局
1
2004
... 生物量作为碳库的重要组分, 是量化生态系统碳周转的重要指标(
Ferlan et al ., 2011 ).木本植物比草本植物具有更多的生物量(
Seastedt & Ramundo, 1990 ), 灌丛地上净初级生产力的增加导致地上碳储量的增加, 并使地下碳库有增加的潜力(
Norris et al ., 2001 ;
Smith & Johnson, 2003 ).因此, 在草地向灌丛和森林演替过程中, 草地群落灌木植物盖度的增大对植被碳库有重要影响.有研究表明, 中国草地植被碳密度平均值范围为0.216-1.148 kg·m
-2 (
Ni, 2002 ;
Fang et al ., 2007 ;
Piao et al ., 2007 ).利用遥感影像建立的植被生物量估算模型计算得到的中国草地植被生物量平均碳密度为0.315 kg·m
-2 (
朴世龙等, 2004 ).本研究中, 无灌木植物草地群落的植被层碳密度为0.304 kg·m
-2 , 与前人研究结果相近.然而低灌木植物盖度草地群落的植被碳密度为1.407 kg·m
-2 , 高灌木植物盖度草地群落的植被碳密度为1.574 kg·m
-2 , 有灌木植物草地群落碳密度远大于无灌木植物草地群落.低灌木植物盖度草地群落相对于无灌木植物草地群落既有草本层的凋落物, 同时也有灌木层的凋落物的贡献(
聂秀青等, 2018 ), 凋落物碳库增加.
Connin等(1997) 研究表明, 灌木相对于多年生草本植物具有更深广发达的根系, 地下根系生物量增加.因此, 灌木植物盖度增大, 植物地上生物量会有所增加, 进而影响草地群落的植被碳库, 忽略灌木植物盖度增加过程将导致对碳库的低估. ...
中国草地植被生物量及其空间分布格局
1
2004
... 生物量作为碳库的重要组分, 是量化生态系统碳周转的重要指标(
Ferlan et al ., 2011 ).木本植物比草本植物具有更多的生物量(
Seastedt & Ramundo, 1990 ), 灌丛地上净初级生产力的增加导致地上碳储量的增加, 并使地下碳库有增加的潜力(
Norris et al ., 2001 ;
Smith & Johnson, 2003 ).因此, 在草地向灌丛和森林演替过程中, 草地群落灌木植物盖度的增大对植被碳库有重要影响.有研究表明, 中国草地植被碳密度平均值范围为0.216-1.148 kg·m
-2 (
Ni, 2002 ;
Fang et al ., 2007 ;
Piao et al ., 2007 ).利用遥感影像建立的植被生物量估算模型计算得到的中国草地植被生物量平均碳密度为0.315 kg·m
-2 (
朴世龙等, 2004 ).本研究中, 无灌木植物草地群落的植被层碳密度为0.304 kg·m
-2 , 与前人研究结果相近.然而低灌木植物盖度草地群落的植被碳密度为1.407 kg·m
-2 , 高灌木植物盖度草地群落的植被碳密度为1.574 kg·m
-2 , 有灌木植物草地群落碳密度远大于无灌木植物草地群落.低灌木植物盖度草地群落相对于无灌木植物草地群落既有草本层的凋落物, 同时也有灌木层的凋落物的贡献(
聂秀青等, 2018 ), 凋落物碳库增加.
Connin等(1997) 研究表明, 灌木相对于多年生草本植物具有更深广发达的根系, 地下根系生物量增加.因此, 灌木植物盖度增大, 植物地上生物量会有所增加, 进而影响草地群落的植被碳库, 忽略灌木植物盖度增加过程将导致对碳库的低估. ...
Changes in biomass carbon stocks in China’s grasslands between 1982 and 1999
1
2007
... 生物量作为碳库的重要组分, 是量化生态系统碳周转的重要指标(
Ferlan et al ., 2011 ).木本植物比草本植物具有更多的生物量(
Seastedt & Ramundo, 1990 ), 灌丛地上净初级生产力的增加导致地上碳储量的增加, 并使地下碳库有增加的潜力(
Norris et al ., 2001 ;
Smith & Johnson, 2003 ).因此, 在草地向灌丛和森林演替过程中, 草地群落灌木植物盖度的增大对植被碳库有重要影响.有研究表明, 中国草地植被碳密度平均值范围为0.216-1.148 kg·m
-2 (
Ni, 2002 ;
Fang et al ., 2007 ;
Piao et al ., 2007 ).利用遥感影像建立的植被生物量估算模型计算得到的中国草地植被生物量平均碳密度为0.315 kg·m
-2 (
朴世龙等, 2004 ).本研究中, 无灌木植物草地群落的植被层碳密度为0.304 kg·m
-2 , 与前人研究结果相近.然而低灌木植物盖度草地群落的植被碳密度为1.407 kg·m
-2 , 高灌木植物盖度草地群落的植被碳密度为1.574 kg·m
-2 , 有灌木植物草地群落碳密度远大于无灌木植物草地群落.低灌木植物盖度草地群落相对于无灌木植物草地群落既有草本层的凋落物, 同时也有灌木层的凋落物的贡献(
聂秀青等, 2018 ), 凋落物碳库增加.
Connin等(1997) 研究表明, 灌木相对于多年生草本植物具有更深广发达的根系, 地下根系生物量增加.因此, 灌木植物盖度增大, 植物地上生物量会有所增加, 进而影响草地群落的植被碳库, 忽略灌木植物盖度增加过程将导致对碳库的低估. ...
Soil carbon sequestration and land-use change: Processes and potential
1
2000
... 西南地区草地多属次生性质, 是以多年生草本植物为主体, 间混少量乔木或灌木的草地类型.在停止人为干扰、水土条件良好的地段, 经过一定时间的恢复后可能回到森林顶极群落(
吴征镒, 1980 ).在顺行演替过程中, 灌丛草地比无灌木植物草地群落维持了更高的物种丰富度(
Howard et al ., 2012 ), 草地向灌丛演替有利于草地群落增加生物量, 促进生态系统碳固定(
Pacala et al ., 2001 ); 随着灌木植物盖度增加, 草地向森林演替过程中生态系统固碳能力改变(
Post & Kwon, 2000 ;
Jackson et al ., 2002 ;
McKinley & Blair, 2008 ).有研究显示, 土壤有机碳密度在青藏高原高寒草地为6.5 kg·m
-2 、北方草地为8.5 kg·m
-2 (
Yang et al ., 2008 ), 而本研究实测的西南地区无灌木植物草地群落(7.215 kg·m
-2 )和低灌木植物盖度草地群落(6.031 kg·m
-2 )的土壤碳密度介于两者之间, 而高灌木植物盖度草地群落(9.735 kg·m
-2 )显著高于高寒草地和北方草地;
Ni (2002) 根据全球土壤碳氮数据库估算的中国草地土壤有机碳密度为13.2 kg·m
-2 , 显著高于本研究实测得到的西南地区草地的土壤平均碳密度.这表明以北方草地和高寒草地的数据为基础将会低估中国草地生态系统碳库, 以全球植被和土壤数据库为基础则会高估中国草地生态系统碳库. ...
Carbon Inventory Methods .
Handbook for Greenhouse Gas Inventory, Carbon Mitigation and Roundwood Production Projects 1
2008
... 西南地区草地生态系统植被碳库包括植物地上碳库、地下碳库和凋落物碳库, 计算公式(
Ravindranath & Ostwald, 2008 )为: ...
Fire and vegetation effects on productivity and nitrogen cycling across a forest-grassland continuum
1
2001
... 随着草地群落灌木植物盖度增加, 灌丛草地地上净初级生产力增加(
Reich et al ., 2001 ;
Hughes et al ., 2006 ;
Knapp et al ., 2008 ), 更多的生物量被分配到地上部分(
Jackson et al ., 1997 ;
Goodale & Davidson, 2002 ), 植被地上碳库增加.但当灌木植物盖度增加到一定程度后, 灌木植株可能会遮挡下层草本植物的光照, 减少草本植物生物量, 导致植物地上碳库反而会低于低灌木植物盖度草地群落.因此, 草地群落中的灌木植物盖度与植物地上碳储量之间存在一定的不确定性, 在大尺度的碳储量评估过程中需要给予高度重视. ...
中国南方草地资源及其发展战略
2
2002
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
... ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
中国南方草地资源及其发展战略
2
2002
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
... ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
Estimating net primary productivity from grassland biomass dynamics measurements
1
2002
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
The influence of fire on belowground processes of tallgrass prairies
1
1990
... 生物量作为碳库的重要组分, 是量化生态系统碳周转的重要指标(
Ferlan et al ., 2011 ).木本植物比草本植物具有更多的生物量(
Seastedt & Ramundo, 1990 ), 灌丛地上净初级生产力的增加导致地上碳储量的增加, 并使地下碳库有增加的潜力(
Norris et al ., 2001 ;
Smith & Johnson, 2003 ).因此, 在草地向灌丛和森林演替过程中, 草地群落灌木植物盖度的增大对植被碳库有重要影响.有研究表明, 中国草地植被碳密度平均值范围为0.216-1.148 kg·m
-2 (
Ni, 2002 ;
Fang et al ., 2007 ;
Piao et al ., 2007 ).利用遥感影像建立的植被生物量估算模型计算得到的中国草地植被生物量平均碳密度为0.315 kg·m
-2 (
朴世龙等, 2004 ).本研究中, 无灌木植物草地群落的植被层碳密度为0.304 kg·m
-2 , 与前人研究结果相近.然而低灌木植物盖度草地群落的植被碳密度为1.407 kg·m
-2 , 高灌木植物盖度草地群落的植被碳密度为1.574 kg·m
-2 , 有灌木植物草地群落碳密度远大于无灌木植物草地群落.低灌木植物盖度草地群落相对于无灌木植物草地群落既有草本层的凋落物, 同时也有灌木层的凋落物的贡献(
聂秀青等, 2018 ), 凋落物碳库增加.
Connin等(1997) 研究表明, 灌木相对于多年生草本植物具有更深广发达的根系, 地下根系生物量增加.因此, 灌木植物盖度增大, 植物地上生物量会有所增加, 进而影响草地群落的植被碳库, 忽略灌木植物盖度增加过程将导致对碳库的低估. ...
中国草地资源的现状分析
1
2016
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
中国草地资源的现状分析
1
2016
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
Expansion of
Juniperus virginiana L. in the Great Plains: Changes in soil organic carbon dynamics
1
2003
... 生物量作为碳库的重要组分, 是量化生态系统碳周转的重要指标(
Ferlan et al ., 2011 ).木本植物比草本植物具有更多的生物量(
Seastedt & Ramundo, 1990 ), 灌丛地上净初级生产力的增加导致地上碳储量的增加, 并使地下碳库有增加的潜力(
Norris et al ., 2001 ;
Smith & Johnson, 2003 ).因此, 在草地向灌丛和森林演替过程中, 草地群落灌木植物盖度的增大对植被碳库有重要影响.有研究表明, 中国草地植被碳密度平均值范围为0.216-1.148 kg·m
-2 (
Ni, 2002 ;
Fang et al ., 2007 ;
Piao et al ., 2007 ).利用遥感影像建立的植被生物量估算模型计算得到的中国草地植被生物量平均碳密度为0.315 kg·m
-2 (
朴世龙等, 2004 ).本研究中, 无灌木植物草地群落的植被层碳密度为0.304 kg·m
-2 , 与前人研究结果相近.然而低灌木植物盖度草地群落的植被碳密度为1.407 kg·m
-2 , 高灌木植物盖度草地群落的植被碳密度为1.574 kg·m
-2 , 有灌木植物草地群落碳密度远大于无灌木植物草地群落.低灌木植物盖度草地群落相对于无灌木植物草地群落既有草本层的凋落物, 同时也有灌木层的凋落物的贡献(
聂秀青等, 2018 ), 凋落物碳库增加.
Connin等(1997) 研究表明, 灌木相对于多年生草本植物具有更深广发达的根系, 地下根系生物量增加.因此, 灌木植物盖度增大, 植物地上生物量会有所增加, 进而影响草地群落的植被碳库, 忽略灌木植物盖度增加过程将导致对碳库的低估. ...
黄土高原植被演替对土壤碳库及
δ 13 C的影响
2
2015
... 草地群落中灌木的增加对土壤碳库的影响目前还未有一致的认识, 土壤碳库与灌木植物盖度的关系也有很大的不确定性.例如,
Hughes等(2006) 的结果表明, 草地演替过程中土壤碳库没有显著改变;
万昊等(2015) 对黄土高原植被演替过程中土壤碳库的研究表明, 草地土壤碳储量高于灌木地; 而
Archer等(1995) 、
Brantley和Young (2010) 、
陈蕾伊等(2014) 则认为, 随着草地群落灌木植物盖度增加土壤碳库也增加.这种研究结果不一致的原因, 也许与气候条件有关, 例如
Jackson等(2002) 在干旱地区的研究也显示, 草地群落土壤碳库随灌木盖度的增加而增加; 但在年降水量较大的地区, 草地土壤碳库随着灌木盖度的增加而降低.灌木根系对土壤碳库的影响是多方面的(
Matamala & Schlesinger, 2003 ), 有研究认为, 灌木将更多的生物量分配到地上而减少地下生物量分配, 分解腐烂后返还给土壤的有机碳也相对较少(
Goodale & Davidson, 2002 ); 但也有研究显示, 灌木增加了凋落物及根系向土壤的营养输入, 根系生长有利于土壤有机质形成和积累(
Knapp et al ., 2008 ). ...
... 本研究中, 低灌木植物盖度草地群落土壤碳库(6.031 kg·m
-2 )小于无灌木植物草地群落(7.215 kg·m
-2 ), 究其原因可能是西南地区水热条件好, 灌木生长较为迅速, 在增加土壤的碳输入的同时(
Fang et al ., 2003 ), 也会增加土壤向大气的碳输出(
Feng et al ., 2008 ); 当然也有可能这些次生灌木尚处在幼龄期还不能对土壤碳库有显著的影响(
Alberti et al ., 2008 ).无灌木植物草地群落绝大部分土壤碳(95.22%)分布于0-50 cm土层, 表层草本植物根系是土壤有机碳密度较高的主要原因, 而灌木根系会延伸至较深土层(
万昊等, 2015 );
Hughes等(2006) 和
陈蕾伊等(2014) 发现, 灌丛草原与无灌丛草原相比, 表层土壤碳储量无显著差异, 而深层土壤碳储量增加.因此, 高灌木植物盖度草地群落比无灌木植物草地群落具有更高的土壤碳密度, 随着灌木植物盖度的增大, 草地生态系统倾向于增加土壤深层生物量以及土壤碳库. ...
黄土高原植被演替对土壤碳库及
δ 13 C的影响
2
2015
... 草地群落中灌木的增加对土壤碳库的影响目前还未有一致的认识, 土壤碳库与灌木植物盖度的关系也有很大的不确定性.例如,
Hughes等(2006) 的结果表明, 草地演替过程中土壤碳库没有显著改变;
万昊等(2015) 对黄土高原植被演替过程中土壤碳库的研究表明, 草地土壤碳储量高于灌木地; 而
Archer等(1995) 、
Brantley和Young (2010) 、
陈蕾伊等(2014) 则认为, 随着草地群落灌木植物盖度增加土壤碳库也增加.这种研究结果不一致的原因, 也许与气候条件有关, 例如
Jackson等(2002) 在干旱地区的研究也显示, 草地群落土壤碳库随灌木盖度的增加而增加; 但在年降水量较大的地区, 草地土壤碳库随着灌木盖度的增加而降低.灌木根系对土壤碳库的影响是多方面的(
Matamala & Schlesinger, 2003 ), 有研究认为, 灌木将更多的生物量分配到地上而减少地下生物量分配, 分解腐烂后返还给土壤的有机碳也相对较少(
Goodale & Davidson, 2002 ); 但也有研究显示, 灌木增加了凋落物及根系向土壤的营养输入, 根系生长有利于土壤有机质形成和积累(
Knapp et al ., 2008 ). ...
... 本研究中, 低灌木植物盖度草地群落土壤碳库(6.031 kg·m
-2 )小于无灌木植物草地群落(7.215 kg·m
-2 ), 究其原因可能是西南地区水热条件好, 灌木生长较为迅速, 在增加土壤的碳输入的同时(
Fang et al ., 2003 ), 也会增加土壤向大气的碳输出(
Feng et al ., 2008 ); 当然也有可能这些次生灌木尚处在幼龄期还不能对土壤碳库有显著的影响(
Alberti et al ., 2008 ).无灌木植物草地群落绝大部分土壤碳(95.22%)分布于0-50 cm土层, 表层草本植物根系是土壤有机碳密度较高的主要原因, 而灌木根系会延伸至较深土层(
万昊等, 2015 );
Hughes等(2006) 和
陈蕾伊等(2014) 发现, 灌丛草原与无灌丛草原相比, 表层土壤碳储量无显著差异, 而深层土壤碳储量增加.因此, 高灌木植物盖度草地群落比无灌木植物草地群落具有更高的土壤碳密度, 随着灌木植物盖度的增大, 草地生态系统倾向于增加土壤深层生物量以及土壤碳库. ...
刈割对南方草地植物补偿性生长的影响——以渝东北部岐山草场为例
1
2014
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
刈割对南方草地植物补偿性生长的影响——以渝东北部岐山草场为例
1
2014
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
不同沙地生境下柠条灌丛化对草地土壤有机碳含量及分布的影响
2
2016
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
... ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
不同沙地生境下柠条灌丛化对草地土壤有机碳含量及分布的影响
2
2016
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
... ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
1
1980
... 西南地区草地多属次生性质, 是以多年生草本植物为主体, 间混少量乔木或灌木的草地类型.在停止人为干扰、水土条件良好的地段, 经过一定时间的恢复后可能回到森林顶极群落(
吴征镒, 1980 ).在顺行演替过程中, 灌丛草地比无灌木植物草地群落维持了更高的物种丰富度(
Howard et al ., 2012 ), 草地向灌丛演替有利于草地群落增加生物量, 促进生态系统碳固定(
Pacala et al ., 2001 ); 随着灌木植物盖度增加, 草地向森林演替过程中生态系统固碳能力改变(
Post & Kwon, 2000 ;
Jackson et al ., 2002 ;
McKinley & Blair, 2008 ).有研究显示, 土壤有机碳密度在青藏高原高寒草地为6.5 kg·m
-2 、北方草地为8.5 kg·m
-2 (
Yang et al ., 2008 ), 而本研究实测的西南地区无灌木植物草地群落(7.215 kg·m
-2 )和低灌木植物盖度草地群落(6.031 kg·m
-2 )的土壤碳密度介于两者之间, 而高灌木植物盖度草地群落(9.735 kg·m
-2 )显著高于高寒草地和北方草地;
Ni (2002) 根据全球土壤碳氮数据库估算的中国草地土壤有机碳密度为13.2 kg·m
-2 , 显著高于本研究实测得到的西南地区草地的土壤平均碳密度.这表明以北方草地和高寒草地的数据为基础将会低估中国草地生态系统碳库, 以全球植被和土壤数据库为基础则会高估中国草地生态系统碳库. ...
1
1980
... 西南地区草地多属次生性质, 是以多年生草本植物为主体, 间混少量乔木或灌木的草地类型.在停止人为干扰、水土条件良好的地段, 经过一定时间的恢复后可能回到森林顶极群落(
吴征镒, 1980 ).在顺行演替过程中, 灌丛草地比无灌木植物草地群落维持了更高的物种丰富度(
Howard et al ., 2012 ), 草地向灌丛演替有利于草地群落增加生物量, 促进生态系统碳固定(
Pacala et al ., 2001 ); 随着灌木植物盖度增加, 草地向森林演替过程中生态系统固碳能力改变(
Post & Kwon, 2000 ;
Jackson et al ., 2002 ;
McKinley & Blair, 2008 ).有研究显示, 土壤有机碳密度在青藏高原高寒草地为6.5 kg·m
-2 、北方草地为8.5 kg·m
-2 (
Yang et al ., 2008 ), 而本研究实测的西南地区无灌木植物草地群落(7.215 kg·m
-2 )和低灌木植物盖度草地群落(6.031 kg·m
-2 )的土壤碳密度介于两者之间, 而高灌木植物盖度草地群落(9.735 kg·m
-2 )显著高于高寒草地和北方草地;
Ni (2002) 根据全球土壤碳氮数据库估算的中国草地土壤有机碳密度为13.2 kg·m
-2 , 显著高于本研究实测得到的西南地区草地的土壤平均碳密度.这表明以北方草地和高寒草地的数据为基础将会低估中国草地生态系统碳库, 以全球植被和土壤数据库为基础则会高估中国草地生态系统碳库. ...
Soil organic carbon stocks in China and changes from 1980s to 2000s
1
2007
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
青海高寒金露梅灌丛碳密度及其分配格局
1
2017
... 所有草本、灌木和凋落物生物量样品在65 ℃烘箱烘干至恒质量(
Perkins & Owens, 2003 ;
Nie et al ., 2016 ;
杨路存等, 2017 ), 测定生物量干质量; 土壤样品风干后过2 mm的土壤筛, 用静电吸附(经摩擦过的塑料卡片或玻璃棒)和手动方法去除根系.随后所有样品用球磨仪研磨后过100目筛, 得到的样品用于指标分析, 3次重复.植物和土壤样品碳含量用元素分析仪(Vario MAX, Elementar, Hanau, Germany)测定. ...
青海高寒金露梅灌丛碳密度及其分配格局
1
2017
... 所有草本、灌木和凋落物生物量样品在65 ℃烘箱烘干至恒质量(
Perkins & Owens, 2003 ;
Nie et al ., 2016 ;
杨路存等, 2017 ), 测定生物量干质量; 土壤样品风干后过2 mm的土壤筛, 用静电吸附(经摩擦过的塑料卡片或玻璃棒)和手动方法去除根系.随后所有样品用球磨仪研磨后过100目筛, 得到的样品用于指标分析, 3次重复.植物和土壤样品碳含量用元素分析仪(Vario MAX, Elementar, Hanau, Germany)测定. ...
Soil carbon stock and its changes in northern China’s grasslands from 1980s to 2000s
1
2010
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
Changes in topsoil carbon stock in the Tibetan grasslands between the 1980s and 2004
1
2009
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
Storage, patterns and controls of soil organic carbon in the Tibetan grasslands
2
2008
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
... 西南地区草地多属次生性质, 是以多年生草本植物为主体, 间混少量乔木或灌木的草地类型.在停止人为干扰、水土条件良好的地段, 经过一定时间的恢复后可能回到森林顶极群落(
吴征镒, 1980 ).在顺行演替过程中, 灌丛草地比无灌木植物草地群落维持了更高的物种丰富度(
Howard et al ., 2012 ), 草地向灌丛演替有利于草地群落增加生物量, 促进生态系统碳固定(
Pacala et al ., 2001 ); 随着灌木植物盖度增加, 草地向森林演替过程中生态系统固碳能力改变(
Post & Kwon, 2000 ;
Jackson et al ., 2002 ;
McKinley & Blair, 2008 ).有研究显示, 土壤有机碳密度在青藏高原高寒草地为6.5 kg·m
-2 、北方草地为8.5 kg·m
-2 (
Yang et al ., 2008 ), 而本研究实测的西南地区无灌木植物草地群落(7.215 kg·m
-2 )和低灌木植物盖度草地群落(6.031 kg·m
-2 )的土壤碳密度介于两者之间, 而高灌木植物盖度草地群落(9.735 kg·m
-2 )显著高于高寒草地和北方草地;
Ni (2002) 根据全球土壤碳氮数据库估算的中国草地土壤有机碳密度为13.2 kg·m
-2 , 显著高于本研究实测得到的西南地区草地的土壤平均碳密度.这表明以北方草地和高寒草地的数据为基础将会低估中国草地生态系统碳库, 以全球植被和土壤数据库为基础则会高估中国草地生态系统碳库. ...
中国亚热带山地杜鹃灌丛生物量分配及其碳密度估算
1
2017
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
中国亚热带山地杜鹃灌丛生物量分配及其碳密度估算
1
2017
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
Characterizing the dynamics of soil organic carbon in grasslands on the Qinghai-Tibetan Plateau
2
2007
... 草地是陆地生态系统的重要组成部分, 对陆地乃至全球碳循环影响巨大(
Scurlock et al ., 2002 ), 也是我国重要的国土资源.我国草地面积占国土面积的41.7%, 占世界草地面积的12.5%, 位居世界第二位.南方天然草地资源, 尤其是大面积连片草地, 主要分布在西南地区(
任继周和张英俊, 2002 ), 西南地区草地面积占全国草地面积的8.13% (
中华人民共和国农业部畜牧兽医司全国畜牧兽医总站, 1996 ;
沈海花等, 2016 ).目前, 针对中国草地生态系统碳库及其动态变化的相关研究较多, 但主要集中于青藏高原和北方草地(
Zhang et al ., 2007 ;
Yang et al ., 2008 ;
方精云等, 2010 ).南方草地95%以上为零星天然草场(
任继周和张英俊, 2002 ;
皇甫江云等, 2012 ), 南方草地主要以草山草坡为主, 多属于次生植被类型, 处于草丛和灌草丛经灌丛向森林演替的过渡阶段, 受制于地形起伏而缺乏对碳库的精确评估(
方精云等, 2018 ).因此, 考虑草地群落灌木植物盖度的作用对于精确估算该地区草地碳库大小十分必要. ...
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
宁夏东部荒漠草原灌丛引入过程中土壤有机碳变化及其空间格局预测
1
2019
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
宁夏东部荒漠草原灌丛引入过程中土壤有机碳变化及其空间格局预测
1
2019
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
青海湖区河谷灌丛草地植被群落多样性研究
1
2007
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
青海湖区河谷灌丛草地植被群落多样性研究
1
2007
... 草地生态系统碳库由植被碳库和土壤碳库两部分构成.植被碳库包括植物地上、地下部分和凋落物, 其大小受到植物群落结构的显著影响(
聂秀青等, 2018 ).西南地区水热条件较好, 植物生长期长, 草地以暖性、热性草丛和灌草丛为主(
李博, 1997 ;
王梅等, 2014 ).西南地区温暖湿润的气候条件有利于草地群落中灌木植物的生长扩张(
蔡文涛等, 2016 ;
张蔷等, 2017 ), 灌木植物的增加会改变植物群落结构, 引起土壤理化性质和微生物群落的改变, 进而可能影响生态系统碳库的动态变化(
祝存冠等, 2007 ;
吴旭东等, 2016 ).但是, 草地群落灌木植物盖度增加对生态系统碳库的影响仍然存在不确定性(
Xie et al ., 2007 ;
Zhang et al ., 2007 ;
Piao et al ., 2009 ;
Yang et al ., 2009 ,
2010 ).有研究表明, 灌丛斑块土壤有机碳含量显著高于对应的草地斑块(
吴旭东等, 2016 ), 但也有研究表明灌木植物的引入并没有显著改变土壤有机碳含量(
赵亚楠等, 2019 ).在干旱地区灌木入侵会导致草地生态系统退化, 降低生物多样性而降低净初级生产力; 而在湿润地区, 草地灌丛化可通过增大叶面积指数而增加净初级生产力(
Knapp et al ., 2008 ;
蔡文涛等, 2016 ).同时, 植物碳分配、根深度以及土壤氮循环过程都将影响生态系统碳储量(
Ferlan et al ., 2011 ).然而, 草地群落中灌木植物盖度增加对生态系统总体碳库影响的相关研究鲜见, 这将增大对草地生态系统碳库估算的不确定性. ...
 ,1,2, 王丽华3, 谢雨1, 高景1,2, 贺俊东1,2, 吴彦
,1,2, 王丽华3, 谢雨1, 高景1,2, 贺俊东1,2, 吴彦 ,1,*
,1,*
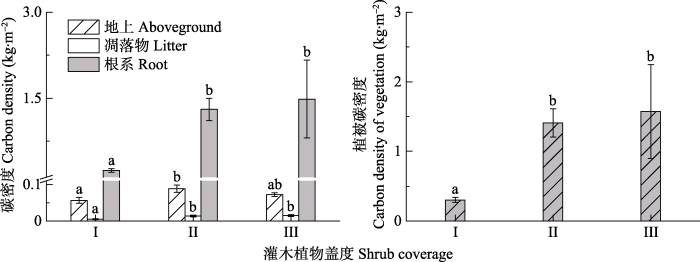 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT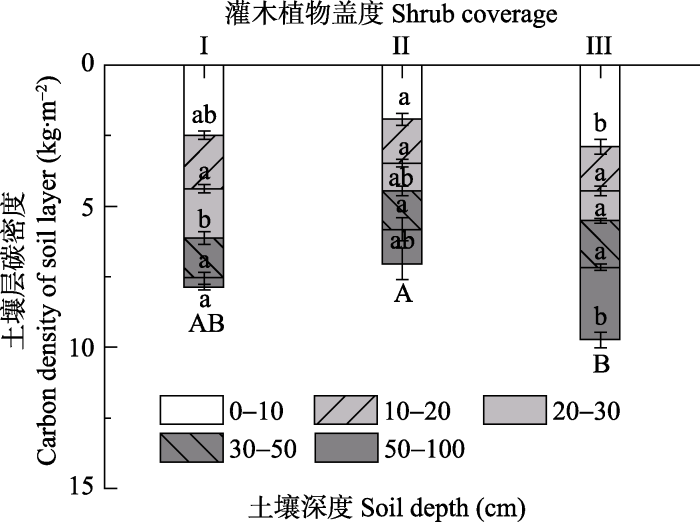 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}