Impacts of nitrogen addition on plant phosphorus content in forest ecosystems and the underlying mechanisms
Chan-Ying FENG1, Cheng-Yang ZHENG,1,*, Di TIAN21 College of Urban and Environmental Sciences, Peking University, Beijing 100871, China 2 College of Life Sciences, Capital Normal University, Beijing 100048, China
Abstract Nitrogen (N) deposition has profound impacts on the phosphorus (P) cycling in forest ecosystems. Especially, the aggravated P limitation on tree growth under N addition has caused much attention to researchers. This article reviews the effects of N addition on plant P content in forest ecosystems. The result showed that N addition increased soil available P and facilitated the absorption of P by plants by promoting soil phosphatase activity, thereby increasing plant P content. Furthermore, changes in tree P content following N addition were also affected by species, life forms as well as experimental duration. Due to the inconsistency, the underlying mechanisms of changes in P content under N addition were further summarized as follows: 1) changes in soil available P content induced by exogenous N input affected the source of plant P; 2) N input affected the P uptake capacity of plants by affecting plant root exudates, mycorrhizal symbiosis and root morphological structure; 3) plant P utilization efficiency was also influenced with changes of P re-distribution and P re-absorption. Overall, for the changes in plant P under increasing exogenous N inputs, alterations of soil available P under N addition was the primary factor, while changes in plant P uptake capacity and P utilization efficiency ulteriorly regulated plant P content. Keywords:nitrogen deposition;forest ecosystems;plant phosphorus concentration;stoichiometry;underlying mechanisms
PDF (1464KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 冯婵莹, 郑成洋, 田地. 氮添加对森林植物磷含量的影响及其机制. 植物生态学报, 2019, 43(3): 185-196. DOI: 10.17521/cjpe.2018.0240 FENG Chan-Ying, ZHENG Cheng-Yang, TIAN Di. Impacts of nitrogen addition on plant phosphorus content in forest ecosystems and the underlying mechanisms. Chinese Journal of Plant Ecology, 2019, 43(3): 185-196. DOI: 10.17521/cjpe.2018.0240
AertsR ( 1996). Nutrient resorption from senescing leaves of perennials: Are there general patterns? Journal of Ecology, 84, 597-608. [本文引用: 1]
AllenMF, SwensonW, QuerejetaJI, EgertonwarburtonLM, TresederKK ( 2003). Ecology of mycorrhizae: A conceptual framework for complex interactions among plants and fungi Annual Review of Phytopathology, 41, 271-303. [本文引用: 1]
Alvarez-ClareS, MackMC, BrooksM ( 2013). A direct test of nitrogen and phosphorus limitation to net primary productivity in a lowland tropical wet forest Ecology, 94, 1540-1551. [本文引用: 2]
BünemannE, ObersonA, FrossardE ( 2011). Phosphorus in Action. Springer Berlin Heidelberg. [本文引用: 1]
CassonNJ, EimersMC, WatmoughSA ( 2012). An assessment of the nutrient status of sugar maple in Ontario: Indications of phosphorus limitation Environmental Monitoring and Assessment, 184, 5917-5927. [本文引用: 1]
Chapin IIIFS, MatsonPA, VitousekPM ( 2012). Principles of Terrestrial Ecosystem Ecology. Springer New York. [本文引用: 1]
Chapin IIIFS, ShaverGR, KedrowskiRA ( 1986). Environmental controls over carbon, nitrogen and phosphorus fractions in Eriophorum vaginatum in Alaskan tussock tundra Journal of Ecology, 74, 167-195. [本文引用: 1]
ChenFS, NiklasKJ, LiuY, FangXM, WanSZ, WangH ( 2015). Nitrogen and phosphorus additions alter nutrient dynamics but not resorption efficiencies of Chinese fir leaves and twigs differing in age Tree Physiology, 35, 1106-1117. [本文引用: 1]
ChenGT, PengY, ZhengJ, LiS, PengTC, QiuXR, TuLH ( 2017 a). Effects of short-term nitrogen addition on fine root biomass, lifespan and morphology of Castanopsis platyacantha in a subtropical secondary evergreen broad-leaved forest Chinese Journal of Plant Ecology, 41, 1041-1050. [本文引用: 1]
ChenGT, ZhengJ, PengTC, LiS, QiuXR, ChenYQ, MaHY, TuLH ( 2017 b). Fine root morphology and chemistry characteristics in different branch orders of Castanopsis platyacantha and their responses to nitrogen addition Chinese Journal of Applied Ecology, 28, 3461-3468. [本文引用: 1]
ChenWW, KouL, JiangL, GaoWL, YangH, WangHM, LiSG ( 2017). Short-term responses of foliar multi-element stoichiometry and nutrient resorption of slash pine to N addition in subtropical China Chinese Journal of Applied Ecology, 28, 1094-1102. [本文引用: 1]
CrousK, ósvaldssonA, EllsworthD ( 2015). Is phosphorus limiting in a mature Eucalyptus woodland? Phosphorus fertilisation stimulates stem growth Plant and Soil, 391, 293-305. [本文引用: 1]
CrowleyKF, McneilBE, LovettGM, CanhamCD, DriscollCT, RustadLE, DennyE, HallettRA, ArthurMA, BoggsJL, GoodaleCL, KahlJS, McNultySG, OllingerSV, PardoLH, SchabergPG, StoddardJL, WeandMP, WeathersKC ( 2012). Do nutrient limitation patterns shift from nitrogen toward phosphorus with increasing nitrogen deposition across the northeastern United States? Ecosystems, 15, 940-957. [本文引用: 2]
DaiKJ, HeF, GuanHL, ShenYX, ZhangGM ( 2006). Research advances in plant and its low-phosphorus environment inducement, adaptation and countermeasures Chinese Journal of Ecology, 25, 1580-1585. [本文引用: 1]
DengHJ, ZhangLN, ZhangGS, LinYM, WuCZ, HongW ( 2015). Effects of nitrogen deposition on leaf elements and their stoichiometric ratios in Schima superba and Pinus massoniana mixed forest Journal of Forest and Environment, 35, 118-124. [本文引用: 1]
DengMF, LiuLL, SunZZ, PiaoSL, MaYC, ChenYW, WangJ, QiaoCL, WangX, LiP ( 2016). Increased phosphate uptake but not resorption alleviates phosphorus deficiency induced by nitrogen deposition in temperate Larix principis-?rupprechtii plantations New Phytologist, 212, 1019-1029. [本文引用: 1]
DengQ, HuiD, DennisS, ReddyKC ( 2017). Responses of terrestrial ecosystem phosphorus cycling to nitrogen addition: A meta analysis Global Ecology and Biogeography, 26, 713-728. [本文引用: 3]
DevauN, Le CadreE, HinsingerP, JaillardB, GerardF ( 2009). Soil pH controls the environmental availability of phosphorus: Experimentaland mechanistic modelling approaches Applied Geochemistry, 24, 2163-2174. [本文引用: 1]
DiepenLTAV, LilleskovEA, PregitzerKS, MillerRM ( 2007). Decline of arbuscular mycorrhizal fungi in northern hardwood forests exposed to chronic nitrogen additions New Phytologist, 176, 175-183. [本文引用: 1]
DoddJC, BurtonCC, BurnsRG, JeffriesP ( 1987). Phosphatase activity associated with the roots and the rhizosphere of plants infected with vesicular-arbuscular mycorrhizal fungi New Phytologist, 107, 163-172. [本文引用: 1]
DuE ( 2016). Rise and fall of nitrogen deposition in the United States Proceedings of the National Academy of Sciences of the United States of America, 113, 3594-3595. [本文引用: 1]
ElserJJ, BrackenME, ClelandEE, GrunerDS, HarpoleWS, HillebrandH, NgaiJT, SeabloomEW, ShurinJB, SmithJE ( 2007). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems Ecology Letters, 10, 1135-1142. [本文引用: 1]
ElvirJA, RustadL, WiersmaGB, FernandezI, WhiteAS, WhiteGJ ( 2005). Eleven-year response of foliar chemistry to chronic nitrogen and sulfur additions at the Bear Brook Watershed in Maine Canadian Journal of Forest Research, 35, 1402-1410. [本文引用: 1]
EmmettBA ( 2007). Nitrogen saturation of terrestrial ecosystems: Some recent findings and their implications for our conceptual framework Water, Air, & Soil Pollution, 7, 99-109. [本文引用: 1]
FilippelliGM ( 2002). The global phosphorus cycle Reviews in Mineralogy & Geochemistry, 48, 391-425. [本文引用: 1]
FujitaY, VenterinkHO, van BodegomPM ( 2014). Low investment in sexual reproduction threatens plants adapted to phosphorus limitation Nature, 505, 82-86. [本文引用: 1]
GarrishV, CernusakLA, WinterK, TurnerBL ( 2010). Nitrogen to phosphorus ratio of plant biomass versus soil solution in a tropical pioneer tree, Ficus insipida Journal of Experimental Botany, 61, 3735-3748. [本文引用: 1]
GartenCT ( 1976). Correlations between concentrations of elements in plants Nature, 261, 686-688. [本文引用: 1]
GianinazziS, Gianinazzi-PearsonV, DexheimerJ ( 1979). Enzymatic studies on the metabolism of vesicular-arbuscular mycorrhiza. III. ultrastructural localization of acid and alkaline phosphatase in onion roots infected by glomus mosseae (nicol. & gerd.) New Phytologist, 82, 127-132. [本文引用: 1]
GundersenP, EmmettBA, Kj?naasOJ, KoopmansCJ, TietemaA ( 1998). Impact of nitrogen deposition on nitrogen cycling in forests: A synthesis of NITREX data Forest Ecology and Management, 101, 37-55. [本文引用: 1]
GüsewellS ( 2004). N:P ratios in terrestrial plants: Variation and functional significance New Phytologist, 164, 243-266. [本文引用: 1]
HedleyMJ, StewartJWB, ChauhanBS ( 1982). Changes in inorganic and organic soil phosphorus fractions induced by cultivation practices and by laboratory incubations Journal of the Soil Science Society of America, 46, 970-976. [本文引用: 1]
HeerwaardenLMV, ToetS, AertsR ( 2003). Nitrogen and phosphorus resorption efficiency and proficiency in six sub-arctic bog species after 4 years of nitrogen fertilization Journal of Ecology, 91, 1060-1070. [本文引用: 1]
HodgeA ( 2004). The plastic plant: Root responses to heterogeneous supplies of nutrients New Phytologist, 162, 9-24. [本文引用: 1]
HomeierJ, HertelD, CamenzindT, CumbicusNL, MaraunM, MartinsonGO, PomaM, RilligMC, SandmannD, ScheuS, VeldkampE, WilckeW, WullaertH, LeuschnerC ( 2012). Tropical andean forests are highly susceptible to nutrient inputs—Rapid effects of experimental N and P addition to an ecuadorian montane forest PLOS ONE, 7, e47128. DOI: 10.1371/journal.pone.?0047128. [本文引用: 1]
JakobsenI, AbbottLK, RobsonAD ( 2010). External hyphae of vesicular—Arbuscular mycorrhizal fungi associated with Trifolium subterraneum L New Phytologist, 120, 509-516. [本文引用: 1]
JansaJ, MozafarA, FrossardE ( 2005). Phosphorus acquisition strategies within arbuscular mycorrhizal fungal community of a single field site Plant and Soil, 276, 163-176. [本文引用: 1]
KoerselmanW ( 1996). The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation Journal of Applied Ecology, 33, 1441-1450. [本文引用: 2]
KouL, GuoDL, YangH, GaoWL, LiSG ( 2015). Growth, morphological traits and mycorrhizal colonization of fine roots respond differently to nitrogen addition in a slash pine plantation in subtropical China Plant and Soil, 391, 207-218. [本文引用: 2]
KouL, WangHM, GaoWL, ChenWW, YangH, LiSG ( 2017). Nitrogen addition regulates tradeoff between root capture and foliar resorption of nitrogen and phosphorus in a subtropical pine plantation Trees, 31, 77-91. [本文引用: 1]
LeeW, FennerM ( 1989). Mineral nutrient allocation in seeds and shoots of twelve Chionochloa species in relation to soil fertility Journal of Ecology, 77, 704-716. [本文引用: 1]
LehmannJ, MuraokaT, ZechW ( 2001). Root activity patterns in an Amazonian agroforest with fruit trees determined by 32P, 33P and 15N applications Agroforestry Systems, 52, 185-197. [本文引用: 1]
LiHY, ZhuYG, MarschnerP, SmithFA, SmithSE ( 2005). Wheat responses to arbuscular mycorrhizal fungi in a highly calcareous soil differ from those of clover, and change with plant development and P supply Plant and Soil, 277, 221-232. [本文引用: 1]
LiQK ( 1986). The progress of modern phosphate fertilizer research Progress in Soil Science, 14(2), 1-7. [本文引用: 1]
LiY, NiuSL, YuGR ( 2016). Aggravated phosphorus limitation on biomass production under increasing nitrogen loading: A meta-analysis Global Change Biology, 22, 934-943. [本文引用: 2]
LiY, TianD, YangH, NiuS ( 2018). Size-dependent nutrient limitation of tree growth from subtropical to cold temperate forests Functional Ecology, 32, 863-875. [本文引用: 1]
LiuJX, HuangWJ, ZhouGY, ZhangDQ, LiuSZ, LiYY ( 2013 a). Nitrogen to phosphorus ratios of tree species in response to elevated carbon dioxide and nitrogen addition in subtropical forests Global Change Biology, 19, 208-216. [本文引用: 1]
LiuL, ZhangT, GilliamFS, GundersenP, ZhangW, ChenH, MoJM ( 2013 b). Interactive effects of nitrogen and phosphorus on soil microbial communities in a tropical forest PLOS ONE, 8, e61188. DOI: 10.1371/journal.pone.0061188. [本文引用: 1]
LuXK, MoJM, GilliamFS, FangH, ZhuFF, FangYT, ZhangW, HuangJ ( 2012). Nitrogen addition shapes soil phosphorus availability in two reforested tropical forests in southern China Biotropica, 44, 302-311. [本文引用: 1]
LussenhopJ, TreonisA, CurtisPS, TeeriJA, VogelCS ( 1998). Response of soil biota to elevated atmospheric CO2 in poplar model systems Oecologia, 113, 247-251. [本文引用: 1]
MaYJ ( 2015). Effects of Fertilization on Nutrient Absorption Characteristics and the Law of C, N, P Ecological Stoichiometry of Cunninghamia lanceolata. Master degree dissertation, Northwest A & F University, Yangling Shaanxi. [本文引用: 1]
MarkleinAR, HoultonBZ ( 2012). Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems New Phytologist, 193, 696-704. [本文引用: 3]
MayorJR, WrightSJ, TurnerBL ( 2014). Species-specific responses of foliar nutrients to long-term nitrogen and phosphorus additions in a lowland tropical forest Journal of Ecology, 102, 36-44. [本文引用: 1]
MindenV, KleyerM ( 2014). Internal and external regulation of plant organ stoichiometry Plant Biology, 16, 897-907. [本文引用: 1]
MohrenGMJ, BurgerFW ( 1986). Phosphorus deficiency induced by nitrogen input in Douglas fir in the Netherlands Plant and Soil, 95, 191-200. [本文引用: 1]
MudgeSR, RaeAL, DiatloffE, SmithFW ( 2002). Expression analysis suggests novel roles for members of the Pht1 family of phosphate transporters in Arabidopsis Plant Journal, 31, 341-353. [本文引用: 1]
OlsenJK, BellLC ( 1990). A glasshouse evaluation of “critical” N and P concentrations and N:P ratios in various plant parts of six eucalypt species Australian Journal of Botany, 38, 669-678. [本文引用: 1]
PangL, ZhangY, ZhouZC, FengZP, ChuDY ( 2014). Effects of simulated nitrogen deposition on growth and phosphorus efficiency of Pinus massoniana under low phosphorus stress Journal of Applied Ecology, 25, 1275-1282. [本文引用: 2]
PangL, ZhouZC, ZhangY, FengZP ( 2016). Effects of atmospheric N sedimentation on growth and P efficiency of Pinus Massoniana mycorrhizal seedlings under low P stress Journal of Plant Nutrition and Fertilizer, 22, 225-235. [本文引用: 1]
PantHK, WarmanPR ( 2000). Enzymatic hydrolysis of soil organic phosphorus by immobilized phosphatases Biology and Fertility of Soils, 30, 306-311. [本文引用: 1]
PasqualiniD, UhlmannA, StürmerSL ( 2007). Arbuscular mycorrhizal fungal communities influence growth and phosphorus concentration of woody plants species from the Atlantic rain forest in South Brazil Forest Ecology and Management, 245, 148-155. [本文引用: 2]
RaghothamaKG ( 1999). Phosphate acquisition Annual Review of Plant Physiology and Plant Molecular Biology, 50, 665-693. [本文引用: 1]
ReedSC, YangX, ThorntonPE ( 2015). Incorporating phosphorus cycling into global modeling efforts: A worthwhile, tractable endeavor New Phytologist, 208, 324-329. [本文引用: 2]
SantiagoLS, WrightSJ, HarmsKE, YavittJB, KorineC, GarciaMN, TurnerBL ( 2012). Tropical tree seedling growth responses to nitrogen, phosphorus and potassium addition Journal of Ecology, 100, 309-316. [本文引用: 1]
SchreegLA, SantiagoLS, WrightSJ, TurnerBL ( 2014). Stem, root, and older leaf N:P ratios are more responsive indicators of soil nutrient availability than new foliage Ecology, 95, 2062-2068. [本文引用: 3]
SeeCR, YanaiRD, FiskMC, VadeboncoeurMA, QuinteroBA, FaheyTJ ( 2015). Soil nitrogen affects phosphorus recycling: Foliar resorption and plant-soil feedbacks in a northern hardwood forest Ecology, 96, 2488-2498. [本文引用: 1]
SongP, ZhangR, ZhangY, ZhouZC, FengZP ( 2016). Effects of simulated nitrogen deposition on fine root morphology, nitrogen and phosphorus efficiency of Pinus massoniana clone under phosphorus deficiency Chinese Journal of Plant Ecology, 40, 1136-1144. [本文引用: 1]
SternerRW, ElserJJ ( 2002).Ecological Stoichiometry:The Biology of Elements from Molecules to the Biosphere. Princeton University Press, Princeton, USA. [本文引用: 1]
TarafdarJC, ClaassenN ( 1998). Organic phosphorus compounds as a phosphorus source for higher plants through the activity of phosphatases produced by plant roots and microorganisms Biology and Fertility of Soils, 5, 308-312. [本文引用: 1]
TessierJT, RaynalDJ ( 2003). Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation Journal of Applied Ecology, 40, 523-534. [本文引用: 1]
TianD ( 2017). Effects of Nutrient Fertilization on the Main Processes of Carbon Cycling in Subtropical Forests. PhD dissertation, Peking University Beijing. [本文引用: 2]
TianD, JiangL, DuEZ, MaSH, ZengWJ, ZouAL, FengCY, XuLC, XingAJ, WangW, ZhengCY, JiCJ, ShenHH, FangJY ( 2018). Responses of forest ecosystems to increasing nitrogen deposition in China: A critical review Environmental Pollution, 243, 75-86. [本文引用: 4]
TianD, YanZB, NiklasKJ, HanWX, KattgeJ, ReichPB, LuoYK, ChenYH, TangZY, HuHF, WrightLJ, SchmidB, FangJY ( 2017). Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent National Science Review, 5, 728-739.
TibbettM, SandersFE ( 2002). Ectomycorrhizal symbiosis can enhance plant nutrition through improved access to discrete organic nutrient patches of high resource quality Annals of botany, 89, 783-789. [本文引用: 1]
TresederKK, VitousekPM ( 2001). Effects of soil nutrient availability on investment in acquisition of N and P in Hawaiian Rain Forests Ecology, 82, 946-954. [本文引用: 4]
TurnerBL ( 2010). Leaf nitrogen to phosphorus ratios of tropical trees: Experimental assessment of physiological and environmental controls New Phytologist, 185, 770-779. [本文引用: 1]
TurnerBL, BrenesarguedasT, ConditR ( 2018). Pervasive phosphorus limitation of tree species but not communities in tropical forests Nature, 555, 367-370. [本文引用: 1]
VanceCP ( 2001). Symbiotic nitrogen fixation and phosphorus acquisition: Plant nutrition in a world of decining renewable resources Plant Physiology, 127, 390-397. [本文引用: 1]
VanceCP, Uhde-Stone, Claudia, AllanDL ( 2010). Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource New Phytologist, 157, 423-447. [本文引用: 1]
VergutzL, ManzoniS, PorporatoA, NovaisRF, JacksonRB ( 2012). Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants Ecological Monographs, 82, 205-220. [本文引用: 1]
VitousekP ( 1982). Nutrient cycling and nutrient use efficiency The American Naturalist, 119, 553-572. [本文引用: 1]
VriesWD, DuE, Butterbach-BahlK ( 2014). Short and long-term impacts of nitrogen deposition on carbon sequestration by forest ecosystems Current Opinion in Environmental Sustainability, 9-10, 90-104. [本文引用: 1]
WaldnerP, MarchettoA, ThimonierA, SchmittM, RogoraM, GrankeO, MuesV, HansenK, KarlssonGP, ZlindraD, ClarkeN, VerstraetenA, LazdinsA, SchimmingC, IacobanC, LindroosA, VanguelovaE, BenhamS, MeesenburgH, NicolasM, KowalskaA, ApuhtinV, NapaU, LachmanovaZ, KristoefelF, BleekerA, IngerslevM, VesterdalL, MolinaJ, FischerU, SeidlingW, JonardM, O'DeaP, JohnsonJ, FischerR, LorenzM ( 2014). Detection of temporal trends in atmospheric deposition of inorganic nitrogen and sulphate to forests in Europe Atmospheric Environment, 95, 363-374. [本文引用: 1]
WangM, MurphyMT, MooreTR ( 2014). Nutrient resorption of two evergreen shrubs in response to long-term fertilization in a bog Oecologia, 174, 365-377. [本文引用: 1]
WangQSY, ZhengCY, ZhangXY, ZengFX, XingJ ( 2016). Impacts of nitrogen addition on foliar nitrogen and phosphorus stoichiometry in a subtropical evergreen broad-leaved forest in Mount Wuyi Chinese Journal of Plant Ecology, 40, 1124-1135. [本文引用: 1]
WeandMP, ArthurMA, LovettGM ( 2010). The phosphorus status of northern hardwoods differs by species but is unaffected by nitrogen fertilization Biogeochemistry, 97, 159-181. [本文引用: 1]
WurzburgerN, WrightSJ ( 2015). Fine-root responses to fertilization reveal multiple nutrient limitation in a lowland tropical forest Ecology, 96, 2137-2146. [本文引用: 1]
YanT, QuTT, SongHH, CiaisP, PiaoSL, SunZZ, ZengH ( 2018). Contrasting effects of N addition on the N and P status of understory vegetation in plantations of sapling and mature Larix principis-rupprechtii Journal of Plant Ecology, 11, 843-852. [本文引用: 1]
YanZB, GuanHY, HanWX, HanTS, GuoYL, FangJY ( 2016). Reproductive organ and young tissues show constrained elemental composition in Arabidopsis thaliana Annals of Botany, 117, 431-439. [本文引用: 2]
YangH ( 2018). Effects of nitrogen and phosphorus addition on leaf nutrient characteristics in a subtropical forest Trees, 32, 383-391. [本文引用: 1]
YangK, ZhuJJ, GuJC, YuLZ, WangZQ ( 2015). Changes in soil phosphorus fractions after 9 years of continuous nitrogen addition in a Larix gmelinii plantation Annals of Forest Science, 72, 435-442. [本文引用: 1]
YangX, TangZY, JiCJ, LiuHY, MaWH, MohhamotA, ShiZY, SunW, WangT, WangXP, WuX, YuSL, YueM, ZhengCY ( 2014). Scaling of nitrogen and phosphorus across plant organs in shrubland biomes across Northern China Scientific Reports, 4, 5448. DOI: 10.1038/srep05448. [本文引用: 1]
YuanZY, ChenHYH ( 2016). Negative effects of fertilization on plant nutrient resorption Ecology, 96, 373-380. [本文引用: 2]
ZhangQF, XieJS, LyuMK, XiongDC, WangJ, ChenYH, LiYQ, WangMK, YangYS ( 2017). Short-term effects of soil warming and nitrogen addition on the N:P stoichiometry of Cunninghamia lanceolata, in subtropical regions Plant and Soil, 411, 395-407. [本文引用: 3]
ZhangY, ZhouZC, YangQ ( 2013). Genetic variations in root morphology and phosphorus efficiency of Pinus massoniana, under heterogeneous and homogeneous low phosphorus conditions Plant and Soil, 364, 93-104. [本文引用: 1]
ZhaoY ( 2016). Effects of N Fertilization on C:N:P Ecological Stoichiometry of Chinese fir in Huitong. Master degree dissertation, Central South University of Forestry and Technology Changsha. [本文引用: 1]
ZhouLL, Addo-DansoSD, WuPF, LiSB, ZouXH, ZhangY, MaXQ ( 2016). Leaf resorption efficiency in relation to foliar and soil nutrient concentrations and stoichiometry of Cunninghamia lanceolata, with stand development in southern China Journal of Soils and Sediments, 16, 1448-1459. [本文引用: 1]
Nitrogen saturation in temperate forest ecosystems: Hypotheses revisited 1 1998
... 工业革命以来, 人类活动导致大气氮(N)沉降不断增加(Galloway et al., 2004), 中国平均N沉降速率从20世纪80年代的13.2 kg·hm-2·a-1增加到21世纪初的21.1 kg·hm-2·a-1 (Liu et al., 2013c).N沉降显著影响陆地生态系统的物种组成和生态过程(Vries et al., 2014), 其中最直接的是对生态系统N循环的影响.一般来说, 对于N限制的森林生态系统, 外源N输入导致土壤无机N含量升高, 土壤N可利用性增加, 植物可吸收N含量升高, 使得植物各器官N含量相应增加(Silver et al., 2005); 而对于非N限制的森林生态系统, 进一步的外源N输入可能导致生态系统N饱和, 使得N流失加剧的同时也会对生态系统其他化学元素循环过程产生深刻的影响(Aber et al., 1998; Emmett, 2007). ...
Nutrient resorption from senescing leaves of perennials: Are there general patterns? 1 1996
... 养分重吸收是指养分从植物衰老器官中转移到其他器官供其重复利用以提高养分利用效率的过程(Vitousek, 1982).这一机制可减少植物对外部养分吸收的依赖, 体现了植物对养分可利用性适应的保守策略(Aerts, 1996).大量研究认为土壤养分有效性与植物养分重吸收能力密切相关.由于N添加加剧了生态系统的P限制, 植物会通过提高对衰老器官中P养分的吸收能力而充分利用P养分, 以最大限度地缓解生长过程中的P限制.如Sullivan等(2014)在夏威夷的施肥实验研究发现, 在N丰富而P限制的森林中, 植物的P重吸收效率增加, 表明N添加条件下植物可通过增加P的重吸收调控自身P养分状况; See等(2015)在Bartlett森林的研究也认为土壤中N有效性的增加会导致凋落物中P含量的减少以实现植物对P的保留.但也有研究指出, N添加不会显著改变植物P重吸收, 表明N、P之间的相互关系对P养分重吸收过程的影响是复杂的(Yuan & Chen, 2016; Zhang et al., 2017), 不仅受外源养分可利用性的影响, 而且与植物养分吸收利用的生理过程(Hodge, 2004; Wang et al., 2014; Kou et al., 2017; Yang, 2018)、叶片生长与寿命(Zhou et al., 2016)、植物维持元素化学计量稳态的特性(Deng et al., 2016)、不同生活型对环境改变的适应(Vergutz et al., 2012)等多种因素密切相关(Tian et al., 2018).因此, N添加条件下植物改变P的重吸收能力只是调控植物内部P含量变化程度的选择性途径之一. ...
Ecology of mycorrhizae: A conceptual framework for complex interactions among plants and fungi 1 2003
... 植物可通过丛枝菌根真菌(AM真菌)和外生菌根真菌(EM真菌)共生而更有效地吸取利用土壤中的P.植物在与菌根共生过程中, 菌根真菌通过把菌丝延伸到植物根毛不能接触的土壤区域并分泌磷酸酶而水解、吸收植物根系不能利用的P养分(Allen et al., 2003; Jansa et al., 2005; Jakobsen et al., 2010).其中, AM真菌可以更有效地利用和矿化有机P, 因此, AM真菌共生对于植物P吸收更为有利(Tibbett & Sanders, 2002).已有研究认为施N可以影响植物菌根生长而改变植物对P的吸收.外源N输入条件下, 植物根系对菌根依赖作用增强, AM真菌的生物量增加, 促进了植物根系对土壤中P的吸收(Lussenhop et al., 1998; Kou et al., 2015; 庞丽等, 2016); 此外, Pasqualini等(2007)直接对植物接种AM真菌, 研究发现先锋树种的组织P含量显著增加, 表明N添加下植物P含量增加与AM真菌共生相关.然而, 也有研究认为N输入下植物菌根真菌生长受抑制或者不受显著影响, 如Liu等(2013b)对N添加下热带森林土壤微生物群落进行磷脂脂肪酸(phospholipids fatty acids, PLFAs)谱图分析, 结果表明N添加减少了AM菌根真菌的含量, 是施N样地植物P含量降低的一个主要原因. ...
A direct test of nitrogen and phosphorus limitation to net primary productivity in a lowland tropical wet forest 2 2013
... Mohren和Burger (1986)最早关注N沉降对森林植物P含量的影响, 研究自然N沉降下荷兰针叶林叶片的P含量变化, 发现外源N沉降使针叶P含量处于较低水平, N:P高达20.随后陆续有关于森林生态系统植物P含量对N沉降响应的研究, 有的研究发现植物叶片P含量增加(Liu et al., 2013a),有的没有显著变化(Weand et al., 2010),有的甚至减少(Tessier & Raynal, 2003; Yan et al., 2018).森林植物P含量对N沉降的响应会受到土壤类型及土壤发育阶段、植物本身遗传特性与环境适应性等多因素的影响.如不同气候区森林土壤养分状况存在差异, 热带和亚热带地区易受P限制, 而温带主要受N限制, 其对N沉降的响应敏感度存在差异(陈美领等, 2016; Yuan & Chen, 2016); 森林生态系统内部植物养分状况有很大差异, 甚至同一生态系统中不同物种或同一物种不同大小的个体受养分限制的程度也不相同, 进而对外源N养分输入的响应也就不同(Alvarez-Clare et al., 2013; Li et al., 2018). ...
... 植物叶片光合作用决定着整个植物体的生长速率, 且叶片对外部环境变化较为敏感.因此, 在大多森林养分添加实验中, 叶片经常被选取作为取样器官, 通过测定其N、P含量或计算N:P来指示植物的养分限制状况(Koerselman, 1996; Sullivan et al., 2014; Tian et al., 2017).森林植物叶片P含量对N添加的响应没有一致的研究结果, 研究结果表现为增加(Crowley et al., 2012; Mayor et al., 2014)、没有显著变化(Li et al., 2016)或减少(陈微微等, 2017; Deng et al., 2017).一般来说, 外源N输入增加条件下, 植物根系吸收大量的N并传输到叶片中, 促进光合作用和碳水化合物的累积; 同时, 为维持有机体内N、P的平衡, 植物会投资大量的N以促进土壤中磷酸酶的活性(Marklein & Houlton, 2012; 田地, 2017), 提高土壤中植物可利用性P含量, 并促进对P的吸收, 进而提高植物叶片的P含量.如在美国东北部, Crowley等(2012)研究森林植物养分含量在自然N沉降梯度(0-14 kg·hm-2)下的响应, 发现7种落叶树种的叶片P含量随着N沉降增加而增加.然而, 当植物生物量增长速率大于植物P元素吸收速率时, 就会产生“生物量稀释作用”, 植物叶片P含量表现为无显著变化甚至减少(Sterner & Elser, 2002; Yan et al., 2016; Zhang et al., 2017). ...
亚热带湿地松叶片多元素化学计量与养分回收对氮添加的短期响应 1 2017
... 植物叶片光合作用决定着整个植物体的生长速率, 且叶片对外部环境变化较为敏感.因此, 在大多森林养分添加实验中, 叶片经常被选取作为取样器官, 通过测定其N、P含量或计算N:P来指示植物的养分限制状况(Koerselman, 1996; Sullivan et al., 2014; Tian et al., 2017).森林植物叶片P含量对N添加的响应没有一致的研究结果, 研究结果表现为增加(Crowley et al., 2012; Mayor et al., 2014)、没有显著变化(Li et al., 2016)或减少(陈微微等, 2017; Deng et al., 2017).一般来说, 外源N输入增加条件下, 植物根系吸收大量的N并传输到叶片中, 促进光合作用和碳水化合物的累积; 同时, 为维持有机体内N、P的平衡, 植物会投资大量的N以促进土壤中磷酸酶的活性(Marklein & Houlton, 2012; 田地, 2017), 提高土壤中植物可利用性P含量, 并促进对P的吸收, 进而提高植物叶片的P含量.如在美国东北部, Crowley等(2012)研究森林植物养分含量在自然N沉降梯度(0-14 kg·hm-2)下的响应, 发现7种落叶树种的叶片P含量随着N沉降增加而增加.然而, 当植物生物量增长速率大于植物P元素吸收速率时, 就会产生“生物量稀释作用”, 植物叶片P含量表现为无显著变化甚至减少(Sterner & Elser, 2002; Yan et al., 2016; Zhang et al., 2017). ...
Is phosphorus limiting in a mature Eucalyptus woodland? Phosphorus fertilisation stimulates stem growth 1 2015
... N和磷(P)作为细胞的基本组成元素, 在核酸、蛋白质等生命物质的合成中起关键作用, 其含量平衡对森林植物生长发育至关重要(Garten, 1976; Chapin et al., 1986).Koerselman (1996)对欧洲沼泽生态系统植物的野外研究数据进行统计分析, 提出了植物N:P化学计量关系对植物生长的影响法则:当植物叶片N:P小于14时, 植物生长受N限制; 当N:P介于14-16时, 植物生长受N和P共同限制; 当N:P大于16时, 植物生长受P限制.后来Güsewell (2004)统计分析不同生态系统(沼泽、山地、森林和草地)植物N、P养分限制的N:P阈值, 认为判断植物N、P养分限制的普适N:P阈值分别为10和20.Elser等(2007)提出全球生态系统生产力普遍受到N限制, 同时P对陆地生态系统生产力的限制性同等重要(Vitousek et al., 2010; Harpole et al., 2011), 尤其是热带地区森林生态系统, N、P同时施加或施P对生态系统生产力有显著的促进作用(Santiago et al., 2012; Crous et al., 2015).植物N:P对生态系统生产力影响的耦合关系使植物P含量对外源N添加的响应受到广泛关注. ...
Do nutrient limitation patterns shift from nitrogen toward phosphorus with increasing nitrogen deposition across the northeastern United States? 2 2012
... 植物叶片光合作用决定着整个植物体的生长速率, 且叶片对外部环境变化较为敏感.因此, 在大多森林养分添加实验中, 叶片经常被选取作为取样器官, 通过测定其N、P含量或计算N:P来指示植物的养分限制状况(Koerselman, 1996; Sullivan et al., 2014; Tian et al., 2017).森林植物叶片P含量对N添加的响应没有一致的研究结果, 研究结果表现为增加(Crowley et al., 2012; Mayor et al., 2014)、没有显著变化(Li et al., 2016)或减少(陈微微等, 2017; Deng et al., 2017).一般来说, 外源N输入增加条件下, 植物根系吸收大量的N并传输到叶片中, 促进光合作用和碳水化合物的累积; 同时, 为维持有机体内N、P的平衡, 植物会投资大量的N以促进土壤中磷酸酶的活性(Marklein & Houlton, 2012; 田地, 2017), 提高土壤中植物可利用性P含量, 并促进对P的吸收, 进而提高植物叶片的P含量.如在美国东北部, Crowley等(2012)研究森林植物养分含量在自然N沉降梯度(0-14 kg·hm-2)下的响应, 发现7种落叶树种的叶片P含量随着N沉降增加而增加.然而, 当植物生物量增长速率大于植物P元素吸收速率时, 就会产生“生物量稀释作用”, 植物叶片P含量表现为无显著变化甚至减少(Sterner & Elser, 2002; Yan et al., 2016; Zhang et al., 2017). ...
... ), 提高土壤中植物可利用性P含量, 并促进对P的吸收, 进而提高植物叶片的P含量.如在美国东北部, Crowley等(2012)研究森林植物养分含量在自然N沉降梯度(0-14 kg·hm-2)下的响应, 发现7种落叶树种的叶片P含量随着N沉降增加而增加.然而, 当植物生物量增长速率大于植物P元素吸收速率时, 就会产生“生物量稀释作用”, 植物叶片P含量表现为无显著变化甚至减少(Sterner & Elser, 2002; Yan et al., 2016; Zhang et al., 2017). ...
Increased phosphate uptake but not resorption alleviates phosphorus deficiency induced by nitrogen deposition in temperate Larix principis-?rupprechtii plantations 1 2016
... 养分重吸收是指养分从植物衰老器官中转移到其他器官供其重复利用以提高养分利用效率的过程(Vitousek, 1982).这一机制可减少植物对外部养分吸收的依赖, 体现了植物对养分可利用性适应的保守策略(Aerts, 1996).大量研究认为土壤养分有效性与植物养分重吸收能力密切相关.由于N添加加剧了生态系统的P限制, 植物会通过提高对衰老器官中P养分的吸收能力而充分利用P养分, 以最大限度地缓解生长过程中的P限制.如Sullivan等(2014)在夏威夷的施肥实验研究发现, 在N丰富而P限制的森林中, 植物的P重吸收效率增加, 表明N添加条件下植物可通过增加P的重吸收调控自身P养分状况; See等(2015)在Bartlett森林的研究也认为土壤中N有效性的增加会导致凋落物中P含量的减少以实现植物对P的保留.但也有研究指出, N添加不会显著改变植物P重吸收, 表明N、P之间的相互关系对P养分重吸收过程的影响是复杂的(Yuan & Chen, 2016; Zhang et al., 2017), 不仅受外源养分可利用性的影响, 而且与植物养分吸收利用的生理过程(Hodge, 2004; Wang et al., 2014; Kou et al., 2017; Yang, 2018)、叶片生长与寿命(Zhou et al., 2016)、植物维持元素化学计量稳态的特性(Deng et al., 2016)、不同生活型对环境改变的适应(Vergutz et al., 2012)等多种因素密切相关(Tian et al., 2018).因此, N添加条件下植物改变P的重吸收能力只是调控植物内部P含量变化程度的选择性途径之一. ...
Responses of terrestrial ecosystem phosphorus cycling to nitrogen addition: A meta analysis 3 2017
... 植物叶片光合作用决定着整个植物体的生长速率, 且叶片对外部环境变化较为敏感.因此, 在大多森林养分添加实验中, 叶片经常被选取作为取样器官, 通过测定其N、P含量或计算N:P来指示植物的养分限制状况(Koerselman, 1996; Sullivan et al., 2014; Tian et al., 2017).森林植物叶片P含量对N添加的响应没有一致的研究结果, 研究结果表现为增加(Crowley et al., 2012; Mayor et al., 2014)、没有显著变化(Li et al., 2016)或减少(陈微微等, 2017; Deng et al., 2017).一般来说, 外源N输入增加条件下, 植物根系吸收大量的N并传输到叶片中, 促进光合作用和碳水化合物的累积; 同时, 为维持有机体内N、P的平衡, 植物会投资大量的N以促进土壤中磷酸酶的活性(Marklein & Houlton, 2012; 田地, 2017), 提高土壤中植物可利用性P含量, 并促进对P的吸收, 进而提高植物叶片的P含量.如在美国东北部, Crowley等(2012)研究森林植物养分含量在自然N沉降梯度(0-14 kg·hm-2)下的响应, 发现7种落叶树种的叶片P含量随着N沉降增加而增加.然而, 当植物生物量增长速率大于植物P元素吸收速率时, 就会产生“生物量稀释作用”, 植物叶片P含量表现为无显著变化甚至减少(Sterner & Elser, 2002; Yan et al., 2016; Zhang et al., 2017). ...
Correlations between concentrations of elements in plants 1 1976
... N和磷(P)作为细胞的基本组成元素, 在核酸、蛋白质等生命物质的合成中起关键作用, 其含量平衡对森林植物生长发育至关重要(Garten, 1976; Chapin et al., 1986).Koerselman (1996)对欧洲沼泽生态系统植物的野外研究数据进行统计分析, 提出了植物N:P化学计量关系对植物生长的影响法则:当植物叶片N:P小于14时, 植物生长受N限制; 当N:P介于14-16时, 植物生长受N和P共同限制; 当N:P大于16时, 植物生长受P限制.后来Güsewell (2004)统计分析不同生态系统(沼泽、山地、森林和草地)植物N、P养分限制的N:P阈值, 认为判断植物N、P养分限制的普适N:P阈值分别为10和20.Elser等(2007)提出全球生态系统生产力普遍受到N限制, 同时P对陆地生态系统生产力的限制性同等重要(Vitousek et al., 2010; Harpole et al., 2011), 尤其是热带地区森林生态系统, N、P同时施加或施P对生态系统生产力有显著的促进作用(Santiago et al., 2012; Crous et al., 2015).植物N:P对生态系统生产力影响的耦合关系使植物P含量对外源N添加的响应受到广泛关注. ...
Enzymatic studies on the metabolism of vesicular-arbuscular mycorrhiza. III. ultrastructural localization of acid and alkaline phosphatase in onion roots infected by glomus mosseae (nicol. & gerd.) 1 1979
Nitrogen and phosphorus resorption efficiency and proficiency in six sub-arctic bog species after 4 years of nitrogen fertilization 1 2003
... 从物种水平考虑, 森林植物叶片P含量对N添加的响应受种类、生活型、生长阶段的影响.在亚北极圈地区, Heerwaarden等(2003)研究常绿木本植物和落叶木本植物养分含量对N添加(NH4Cl, 10 g N·m-2·a-1)的响应.结果发现: 对于常绿木本植物Andromeda polifolia的叶片P含量没有表现出显著变化, 而一种岩高兰属植物Empetrum hermaphroditum表现为显著的负效应; 对于落叶木本植物, 笃斯越桔(Vaccinium uliginosum)和Betula nana均表现为显著的负效应.在亚热带常绿阔叶林中, 5年施N显著提高蕨类和草本植物的叶片P含量, 降低苔藓叶片P含量, 但对乔木和灌木叶片P含量无显著影响(王乔姝怡等, 2016).此外, 研究认为植物处于不同生长阶段时, 其P含量对N添加的响应也不一致: 幼苗阶段时植物P含量对施N响应更敏感(Tian et al., 2018).如马亚娟(2015)对3年生杉木(Cunninghamia lanceolata)幼苗进行低N处理(2-6 g N·pot-1), 结果发现施N显著降低杉木叶片P含量, 相对对照组下降了7.8%-24%; 对于杉木成熟植株, 低N处理(5 g·m-2·a-1)对叶片P含量影响不显著, 相对对照组仅下降了1.2% (赵月, 2016).原因是相对成熟植株, 幼苗植株处于生长旺盛期, 生物量增长速率高, 导致生物量“稀释作用”明显, 进而对各器官P含量产生显著影响(Chen et al., 2015; Zhang et al., 2017; Tian et al., 2018). ...
The plastic plant: Root responses to heterogeneous supplies of nutrients 1 2004
... 养分重吸收是指养分从植物衰老器官中转移到其他器官供其重复利用以提高养分利用效率的过程(Vitousek, 1982).这一机制可减少植物对外部养分吸收的依赖, 体现了植物对养分可利用性适应的保守策略(Aerts, 1996).大量研究认为土壤养分有效性与植物养分重吸收能力密切相关.由于N添加加剧了生态系统的P限制, 植物会通过提高对衰老器官中P养分的吸收能力而充分利用P养分, 以最大限度地缓解生长过程中的P限制.如Sullivan等(2014)在夏威夷的施肥实验研究发现, 在N丰富而P限制的森林中, 植物的P重吸收效率增加, 表明N添加条件下植物可通过增加P的重吸收调控自身P养分状况; See等(2015)在Bartlett森林的研究也认为土壤中N有效性的增加会导致凋落物中P含量的减少以实现植物对P的保留.但也有研究指出, N添加不会显著改变植物P重吸收, 表明N、P之间的相互关系对P养分重吸收过程的影响是复杂的(Yuan & Chen, 2016; Zhang et al., 2017), 不仅受外源养分可利用性的影响, 而且与植物养分吸收利用的生理过程(Hodge, 2004; Wang et al., 2014; Kou et al., 2017; Yang, 2018)、叶片生长与寿命(Zhou et al., 2016)、植物维持元素化学计量稳态的特性(Deng et al., 2016)、不同生活型对环境改变的适应(Vergutz et al., 2012)等多种因素密切相关(Tian et al., 2018).因此, N添加条件下植物改变P的重吸收能力只是调控植物内部P含量变化程度的选择性途径之一. ...
Tropical andean forests are highly susceptible to nutrient inputs—Rapid effects of experimental N and P addition to an ecuadorian montane forest 1 2012
External hyphae of vesicular—Arbuscular mycorrhizal fungi associated with Trifolium subterraneum L 1 2010
... 植物可通过丛枝菌根真菌(AM真菌)和外生菌根真菌(EM真菌)共生而更有效地吸取利用土壤中的P.植物在与菌根共生过程中, 菌根真菌通过把菌丝延伸到植物根毛不能接触的土壤区域并分泌磷酸酶而水解、吸收植物根系不能利用的P养分(Allen et al., 2003; Jansa et al., 2005; Jakobsen et al., 2010).其中, AM真菌可以更有效地利用和矿化有机P, 因此, AM真菌共生对于植物P吸收更为有利(Tibbett & Sanders, 2002).已有研究认为施N可以影响植物菌根生长而改变植物对P的吸收.外源N输入条件下, 植物根系对菌根依赖作用增强, AM真菌的生物量增加, 促进了植物根系对土壤中P的吸收(Lussenhop et al., 1998; Kou et al., 2015; 庞丽等, 2016); 此外, Pasqualini等(2007)直接对植物接种AM真菌, 研究发现先锋树种的组织P含量显著增加, 表明N添加下植物P含量增加与AM真菌共生相关.然而, 也有研究认为N输入下植物菌根真菌生长受抑制或者不受显著影响, 如Liu等(2013b)对N添加下热带森林土壤微生物群落进行磷脂脂肪酸(phospholipids fatty acids, PLFAs)谱图分析, 结果表明N添加减少了AM菌根真菌的含量, 是施N样地植物P含量降低的一个主要原因. ...
Phosphorus acquisition strategies within arbuscular mycorrhizal fungal community of a single field site 1 2005
... 植物可通过丛枝菌根真菌(AM真菌)和外生菌根真菌(EM真菌)共生而更有效地吸取利用土壤中的P.植物在与菌根共生过程中, 菌根真菌通过把菌丝延伸到植物根毛不能接触的土壤区域并分泌磷酸酶而水解、吸收植物根系不能利用的P养分(Allen et al., 2003; Jansa et al., 2005; Jakobsen et al., 2010).其中, AM真菌可以更有效地利用和矿化有机P, 因此, AM真菌共生对于植物P吸收更为有利(Tibbett & Sanders, 2002).已有研究认为施N可以影响植物菌根生长而改变植物对P的吸收.外源N输入条件下, 植物根系对菌根依赖作用增强, AM真菌的生物量增加, 促进了植物根系对土壤中P的吸收(Lussenhop et al., 1998; Kou et al., 2015; 庞丽等, 2016); 此外, Pasqualini等(2007)直接对植物接种AM真菌, 研究发现先锋树种的组织P含量显著增加, 表明N添加下植物P含量增加与AM真菌共生相关.然而, 也有研究认为N输入下植物菌根真菌生长受抑制或者不受显著影响, 如Liu等(2013b)对N添加下热带森林土壤微生物群落进行磷脂脂肪酸(phospholipids fatty acids, PLFAs)谱图分析, 结果表明N添加减少了AM菌根真菌的含量, 是施N样地植物P含量降低的一个主要原因. ...
The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation 2 1996
... N和磷(P)作为细胞的基本组成元素, 在核酸、蛋白质等生命物质的合成中起关键作用, 其含量平衡对森林植物生长发育至关重要(Garten, 1976; Chapin et al., 1986).Koerselman (1996)对欧洲沼泽生态系统植物的野外研究数据进行统计分析, 提出了植物N:P化学计量关系对植物生长的影响法则:当植物叶片N:P小于14时, 植物生长受N限制; 当N:P介于14-16时, 植物生长受N和P共同限制; 当N:P大于16时, 植物生长受P限制.后来Güsewell (2004)统计分析不同生态系统(沼泽、山地、森林和草地)植物N、P养分限制的N:P阈值, 认为判断植物N、P养分限制的普适N:P阈值分别为10和20.Elser等(2007)提出全球生态系统生产力普遍受到N限制, 同时P对陆地生态系统生产力的限制性同等重要(Vitousek et al., 2010; Harpole et al., 2011), 尤其是热带地区森林生态系统, N、P同时施加或施P对生态系统生产力有显著的促进作用(Santiago et al., 2012; Crous et al., 2015).植物N:P对生态系统生产力影响的耦合关系使植物P含量对外源N添加的响应受到广泛关注. ...
... 植物叶片光合作用决定着整个植物体的生长速率, 且叶片对外部环境变化较为敏感.因此, 在大多森林养分添加实验中, 叶片经常被选取作为取样器官, 通过测定其N、P含量或计算N:P来指示植物的养分限制状况(Koerselman, 1996; Sullivan et al., 2014; Tian et al., 2017).森林植物叶片P含量对N添加的响应没有一致的研究结果, 研究结果表现为增加(Crowley et al., 2012; Mayor et al., 2014)、没有显著变化(Li et al., 2016)或减少(陈微微等, 2017; Deng et al., 2017).一般来说, 外源N输入增加条件下, 植物根系吸收大量的N并传输到叶片中, 促进光合作用和碳水化合物的累积; 同时, 为维持有机体内N、P的平衡, 植物会投资大量的N以促进土壤中磷酸酶的活性(Marklein & Houlton, 2012; 田地, 2017), 提高土壤中植物可利用性P含量, 并促进对P的吸收, 进而提高植物叶片的P含量.如在美国东北部, Crowley等(2012)研究森林植物养分含量在自然N沉降梯度(0-14 kg·hm-2)下的响应, 发现7种落叶树种的叶片P含量随着N沉降增加而增加.然而, 当植物生物量增长速率大于植物P元素吸收速率时, 就会产生“生物量稀释作用”, 植物叶片P含量表现为无显著变化甚至减少(Sterner & Elser, 2002; Yan et al., 2016; Zhang et al., 2017). ...
Growth, morphological traits and mycorrhizal colonization of fine roots respond differently to nitrogen addition in a slash pine plantation in subtropical China 2 2015
... 植物可通过丛枝菌根真菌(AM真菌)和外生菌根真菌(EM真菌)共生而更有效地吸取利用土壤中的P.植物在与菌根共生过程中, 菌根真菌通过把菌丝延伸到植物根毛不能接触的土壤区域并分泌磷酸酶而水解、吸收植物根系不能利用的P养分(Allen et al., 2003; Jansa et al., 2005; Jakobsen et al., 2010).其中, AM真菌可以更有效地利用和矿化有机P, 因此, AM真菌共生对于植物P吸收更为有利(Tibbett & Sanders, 2002).已有研究认为施N可以影响植物菌根生长而改变植物对P的吸收.外源N输入条件下, 植物根系对菌根依赖作用增强, AM真菌的生物量增加, 促进了植物根系对土壤中P的吸收(Lussenhop et al., 1998; Kou et al., 2015; 庞丽等, 2016); 此外, Pasqualini等(2007)直接对植物接种AM真菌, 研究发现先锋树种的组织P含量显著增加, 表明N添加下植物P含量增加与AM真菌共生相关.然而, 也有研究认为N输入下植物菌根真菌生长受抑制或者不受显著影响, 如Liu等(2013b)对N添加下热带森林土壤微生物群落进行磷脂脂肪酸(phospholipids fatty acids, PLFAs)谱图分析, 结果表明N添加减少了AM菌根真菌的含量, 是施N样地植物P含量降低的一个主要原因. ...
Nitrogen addition regulates tradeoff between root capture and foliar resorption of nitrogen and phosphorus in a subtropical pine plantation 1 2017
... 养分重吸收是指养分从植物衰老器官中转移到其他器官供其重复利用以提高养分利用效率的过程(Vitousek, 1982).这一机制可减少植物对外部养分吸收的依赖, 体现了植物对养分可利用性适应的保守策略(Aerts, 1996).大量研究认为土壤养分有效性与植物养分重吸收能力密切相关.由于N添加加剧了生态系统的P限制, 植物会通过提高对衰老器官中P养分的吸收能力而充分利用P养分, 以最大限度地缓解生长过程中的P限制.如Sullivan等(2014)在夏威夷的施肥实验研究发现, 在N丰富而P限制的森林中, 植物的P重吸收效率增加, 表明N添加条件下植物可通过增加P的重吸收调控自身P养分状况; See等(2015)在Bartlett森林的研究也认为土壤中N有效性的增加会导致凋落物中P含量的减少以实现植物对P的保留.但也有研究指出, N添加不会显著改变植物P重吸收, 表明N、P之间的相互关系对P养分重吸收过程的影响是复杂的(Yuan & Chen, 2016; Zhang et al., 2017), 不仅受外源养分可利用性的影响, 而且与植物养分吸收利用的生理过程(Hodge, 2004; Wang et al., 2014; Kou et al., 2017; Yang, 2018)、叶片生长与寿命(Zhou et al., 2016)、植物维持元素化学计量稳态的特性(Deng et al., 2016)、不同生活型对环境改变的适应(Vergutz et al., 2012)等多种因素密切相关(Tian et al., 2018).因此, N添加条件下植物改变P的重吸收能力只是调控植物内部P含量变化程度的选择性途径之一. ...
Mineral nutrient allocation in seeds and shoots of twelve Chionochloa species in relation to soil fertility 1 1989
... 植物的N、P养分含量及其比值用于指示植物N、P养分有效性及限制情况, 这一方法最早应用于海藻和浮游植物的研究, 随后其有效性在草本植物的研究中得到验证, 由此森林生态学家广泛地把树木叶片的N、P养分含量及N:P作为植物N、P养分有效性及限制情况的判断标准(Schreeg et al., 2014).从已有研究看, N添加对森林植物P含量的影响多集中在对植物叶片的分析, 较少研究关注植物其他器官如枝、茎和根等的响应.然而, 已有的关于森林植物枝、茎和根的P含量对N添加响应的研究认为, 枝、茎和根等营养器官的P含量更能指示土壤的P养分有效性(Minden & Kleyer, 2014; Schreeg et al., 2014; Yang et al., 2014).因此, 分析叶片P含量的同时, 植物枝、茎和根等营养器官中的P含量变化不可忽视, 而仅仅把叶片P含量及其N:P的变化作为森林植物P养分限制的标准, 这一方法是否继续适用于未来N沉降的研究中, 需要开展更多关于植物不同器官P含量对N添加响应的研究进一步验证.此外, 植物的生殖器官(花、果实和种子)决定植物的生殖过程, 其养分(如P)含量等在植物种子扩散、幼苗育成过程中起关键作用(Lee & Fenner, 1989; Fujita et al., 2014).因此, 开展关于森林植物生殖器官P含量对N添加的响应研究有利于更好地理解N沉降对植物生殖过程的影响.为了更精确地了解N添加对植物P养分的影响, 除了综合考虑植物各营养器官和生殖器官的P含量变化以外, 还可结合同位素示踪法研究植物不同器官间P的转化和迁移过程, 如Lehmann等(2001)通过32P同位素示踪的方法研究不同果树根系对P养分的吸收活性, 发现不同果树的P吸收能力不同, 且果树根系从土壤中吸收32P后, 32P在果树体内迁移至叶片, 且在36天后叶片32P达到峰值. ...
Root activity patterns in an Amazonian agroforest with fruit trees determined by 32P, 33P and 15N applications 1 2001
... 植物的N、P养分含量及其比值用于指示植物N、P养分有效性及限制情况, 这一方法最早应用于海藻和浮游植物的研究, 随后其有效性在草本植物的研究中得到验证, 由此森林生态学家广泛地把树木叶片的N、P养分含量及N:P作为植物N、P养分有效性及限制情况的判断标准(Schreeg et al., 2014).从已有研究看, N添加对森林植物P含量的影响多集中在对植物叶片的分析, 较少研究关注植物其他器官如枝、茎和根等的响应.然而, 已有的关于森林植物枝、茎和根的P含量对N添加响应的研究认为, 枝、茎和根等营养器官的P含量更能指示土壤的P养分有效性(Minden & Kleyer, 2014; Schreeg et al., 2014; Yang et al., 2014).因此, 分析叶片P含量的同时, 植物枝、茎和根等营养器官中的P含量变化不可忽视, 而仅仅把叶片P含量及其N:P的变化作为森林植物P养分限制的标准, 这一方法是否继续适用于未来N沉降的研究中, 需要开展更多关于植物不同器官P含量对N添加响应的研究进一步验证.此外, 植物的生殖器官(花、果实和种子)决定植物的生殖过程, 其养分(如P)含量等在植物种子扩散、幼苗育成过程中起关键作用(Lee & Fenner, 1989; Fujita et al., 2014).因此, 开展关于森林植物生殖器官P含量对N添加的响应研究有利于更好地理解N沉降对植物生殖过程的影响.为了更精确地了解N添加对植物P养分的影响, 除了综合考虑植物各营养器官和生殖器官的P含量变化以外, 还可结合同位素示踪法研究植物不同器官间P的转化和迁移过程, 如Lehmann等(2001)通过32P同位素示踪的方法研究不同果树根系对P养分的吸收活性, 发现不同果树的P吸收能力不同, 且果树根系从土壤中吸收32P后, 32P在果树体内迁移至叶片, 且在36天后叶片32P达到峰值. ...
Wheat responses to arbuscular mycorrhizal fungi in a highly calcareous soil differ from those of clover, and change with plant development and P supply 1 2005
... 菌根真菌生长对施N的响应差异可能源于施N下不同程度的土壤酸化作用和植物不同的养分状态.长期施N或者过量施N容易导致土壤酸化而不利于菌根生长(Diepen et al., 2007).同时, 菌根真菌生长需要大量的碳投资, 而在施N条件下, 当植物从菌根真菌获得的P吸收的收益不足以弥补菌根真菌生长发育耗费碳供给的代价时, 生态系统会更倾向于其他途径如投资N生成磷酸酶以获取P, 由此菌根真菌共生就会呈现出对施N的负响应(Li et al., 2005).如Treseder和Vitousek (2001)在对夏威夷几种不同类型的森林生态系统的施N实验研究发现, 施N抑制或不显著影响植物菌根真菌生物量, 但显著增加了植物根际土壤磷酸酶的活性. ...
Arbuscular mycorrhizal fungal communities influence growth and phosphorus concentration of woody plants species from the Atlantic rain forest in South Brazil 2 2007
... 植物可通过丛枝菌根真菌(AM真菌)和外生菌根真菌(EM真菌)共生而更有效地吸取利用土壤中的P.植物在与菌根共生过程中, 菌根真菌通过把菌丝延伸到植物根毛不能接触的土壤区域并分泌磷酸酶而水解、吸收植物根系不能利用的P养分(Allen et al., 2003; Jansa et al., 2005; Jakobsen et al., 2010).其中, AM真菌可以更有效地利用和矿化有机P, 因此, AM真菌共生对于植物P吸收更为有利(Tibbett & Sanders, 2002).已有研究认为施N可以影响植物菌根生长而改变植物对P的吸收.外源N输入条件下, 植物根系对菌根依赖作用增强, AM真菌的生物量增加, 促进了植物根系对土壤中P的吸收(Lussenhop et al., 1998; Kou et al., 2015; 庞丽等, 2016); 此外, Pasqualini等(2007)直接对植物接种AM真菌, 研究发现先锋树种的组织P含量显著增加, 表明N添加下植物P含量增加与AM真菌共生相关.然而, 也有研究认为N输入下植物菌根真菌生长受抑制或者不受显著影响, 如Liu等(2013b)对N添加下热带森林土壤微生物群落进行磷脂脂肪酸(phospholipids fatty acids, PLFAs)谱图分析, 结果表明N添加减少了AM菌根真菌的含量, 是施N样地植物P含量降低的一个主要原因. ...
... 植物叶片光合作用决定着整个植物体的生长速率, 且叶片对外部环境变化较为敏感.因此, 在大多森林养分添加实验中, 叶片经常被选取作为取样器官, 通过测定其N、P含量或计算N:P来指示植物的养分限制状况(Koerselman, 1996; Sullivan et al., 2014; Tian et al., 2017).森林植物叶片P含量对N添加的响应没有一致的研究结果, 研究结果表现为增加(Crowley et al., 2012; Mayor et al., 2014)、没有显著变化(Li et al., 2016)或减少(陈微微等, 2017; Deng et al., 2017).一般来说, 外源N输入增加条件下, 植物根系吸收大量的N并传输到叶片中, 促进光合作用和碳水化合物的累积; 同时, 为维持有机体内N、P的平衡, 植物会投资大量的N以促进土壤中磷酸酶的活性(Marklein & Houlton, 2012; 田地, 2017), 提高土壤中植物可利用性P含量, 并促进对P的吸收, 进而提高植物叶片的P含量.如在美国东北部, Crowley等(2012)研究森林植物养分含量在自然N沉降梯度(0-14 kg·hm-2)下的响应, 发现7种落叶树种的叶片P含量随着N沉降增加而增加.然而, 当植物生物量增长速率大于植物P元素吸收速率时, 就会产生“生物量稀释作用”, 植物叶片P含量表现为无显著变化甚至减少(Sterner & Elser, 2002; Yan et al., 2016; Zhang et al., 2017). ...
Assessing nutrient limitation in complex forested ecosystems: Alternatives to large-scale fertilization experiments 2 2014
... 植物叶片光合作用决定着整个植物体的生长速率, 且叶片对外部环境变化较为敏感.因此, 在大多森林养分添加实验中, 叶片经常被选取作为取样器官, 通过测定其N、P含量或计算N:P来指示植物的养分限制状况(Koerselman, 1996; Sullivan et al., 2014; Tian et al., 2017).森林植物叶片P含量对N添加的响应没有一致的研究结果, 研究结果表现为增加(Crowley et al., 2012; Mayor et al., 2014)、没有显著变化(Li et al., 2016)或减少(陈微微等, 2017; Deng et al., 2017).一般来说, 外源N输入增加条件下, 植物根系吸收大量的N并传输到叶片中, 促进光合作用和碳水化合物的累积; 同时, 为维持有机体内N、P的平衡, 植物会投资大量的N以促进土壤中磷酸酶的活性(Marklein & Houlton, 2012; 田地, 2017), 提高土壤中植物可利用性P含量, 并促进对P的吸收, 进而提高植物叶片的P含量.如在美国东北部, Crowley等(2012)研究森林植物养分含量在自然N沉降梯度(0-14 kg·hm-2)下的响应, 发现7种落叶树种的叶片P含量随着N沉降增加而增加.然而, 当植物生物量增长速率大于植物P元素吸收速率时, 就会产生“生物量稀释作用”, 植物叶片P含量表现为无显著变化甚至减少(Sterner & Elser, 2002; Yan et al., 2016; Zhang et al., 2017). ...
... 养分重吸收是指养分从植物衰老器官中转移到其他器官供其重复利用以提高养分利用效率的过程(Vitousek, 1982).这一机制可减少植物对外部养分吸收的依赖, 体现了植物对养分可利用性适应的保守策略(Aerts, 1996).大量研究认为土壤养分有效性与植物养分重吸收能力密切相关.由于N添加加剧了生态系统的P限制, 植物会通过提高对衰老器官中P养分的吸收能力而充分利用P养分, 以最大限度地缓解生长过程中的P限制.如Sullivan等(2014)在夏威夷的施肥实验研究发现, 在N丰富而P限制的森林中, 植物的P重吸收效率增加, 表明N添加条件下植物可通过增加P的重吸收调控自身P养分状况; See等(2015)在Bartlett森林的研究也认为土壤中N有效性的增加会导致凋落物中P含量的减少以实现植物对P的保留.但也有研究指出, N添加不会显著改变植物P重吸收, 表明N、P之间的相互关系对P养分重吸收过程的影响是复杂的(Yuan & Chen, 2016; Zhang et al., 2017), 不仅受外源养分可利用性的影响, 而且与植物养分吸收利用的生理过程(Hodge, 2004; Wang et al., 2014; Kou et al., 2017; Yang, 2018)、叶片生长与寿命(Zhou et al., 2016)、植物维持元素化学计量稳态的特性(Deng et al., 2016)、不同生活型对环境改变的适应(Vergutz et al., 2012)等多种因素密切相关(Tian et al., 2018).因此, N添加条件下植物改变P的重吸收能力只是调控植物内部P含量变化程度的选择性途径之一. ...
Organic phosphorus compounds as a phosphorus source for higher plants through the activity of phosphatases produced by plant roots and microorganisms 1 1998
Contrasting effects of N addition on the N and P status of understory vegetation in plantations of sapling and mature Larix principis-rupprechtii 1 2018
... Mohren和Burger (1986)最早关注N沉降对森林植物P含量的影响, 研究自然N沉降下荷兰针叶林叶片的P含量变化, 发现外源N沉降使针叶P含量处于较低水平, N:P高达20.随后陆续有关于森林生态系统植物P含量对N沉降响应的研究, 有的研究发现植物叶片P含量增加(Liu et al., 2013a),有的没有显著变化(Weand et al., 2010),有的甚至减少(Tessier & Raynal, 2003; Yan et al., 2018).森林植物P含量对N沉降的响应会受到土壤类型及土壤发育阶段、植物本身遗传特性与环境适应性等多因素的影响.如不同气候区森林土壤养分状况存在差异, 热带和亚热带地区易受P限制, 而温带主要受N限制, 其对N沉降的响应敏感度存在差异(陈美领等, 2016; Yuan & Chen, 2016); 森林生态系统内部植物养分状况有很大差异, 甚至同一生态系统中不同物种或同一物种不同大小的个体受养分限制的程度也不相同, 进而对外源N养分输入的响应也就不同(Alvarez-Clare et al., 2013; Li et al., 2018). ...
Reproductive organ and young tissues show constrained elemental composition in Arabidopsis thaliana 2 2016
... 植物叶片光合作用决定着整个植物体的生长速率, 且叶片对外部环境变化较为敏感.因此, 在大多森林养分添加实验中, 叶片经常被选取作为取样器官, 通过测定其N、P含量或计算N:P来指示植物的养分限制状况(Koerselman, 1996; Sullivan et al., 2014; Tian et al., 2017).森林植物叶片P含量对N添加的响应没有一致的研究结果, 研究结果表现为增加(Crowley et al., 2012; Mayor et al., 2014)、没有显著变化(Li et al., 2016)或减少(陈微微等, 2017; Deng et al., 2017).一般来说, 外源N输入增加条件下, 植物根系吸收大量的N并传输到叶片中, 促进光合作用和碳水化合物的累积; 同时, 为维持有机体内N、P的平衡, 植物会投资大量的N以促进土壤中磷酸酶的活性(Marklein & Houlton, 2012; 田地, 2017), 提高土壤中植物可利用性P含量, 并促进对P的吸收, 进而提高植物叶片的P含量.如在美国东北部, Crowley等(2012)研究森林植物养分含量在自然N沉降梯度(0-14 kg·hm-2)下的响应, 发现7种落叶树种的叶片P含量随着N沉降增加而增加.然而, 当植物生物量增长速率大于植物P元素吸收速率时, 就会产生“生物量稀释作用”, 植物叶片P含量表现为无显著变化甚至减少(Sterner & Elser, 2002; Yan et al., 2016; Zhang et al., 2017). ...
Scaling of nitrogen and phosphorus across plant organs in shrubland biomes across Northern China 1 2014
... 植物的N、P养分含量及其比值用于指示植物N、P养分有效性及限制情况, 这一方法最早应用于海藻和浮游植物的研究, 随后其有效性在草本植物的研究中得到验证, 由此森林生态学家广泛地把树木叶片的N、P养分含量及N:P作为植物N、P养分有效性及限制情况的判断标准(Schreeg et al., 2014).从已有研究看, N添加对森林植物P含量的影响多集中在对植物叶片的分析, 较少研究关注植物其他器官如枝、茎和根等的响应.然而, 已有的关于森林植物枝、茎和根的P含量对N添加响应的研究认为, 枝、茎和根等营养器官的P含量更能指示土壤的P养分有效性(Minden & Kleyer, 2014; Schreeg et al., 2014; Yang et al., 2014).因此, 分析叶片P含量的同时, 植物枝、茎和根等营养器官中的P含量变化不可忽视, 而仅仅把叶片P含量及其N:P的变化作为森林植物P养分限制的标准, 这一方法是否继续适用于未来N沉降的研究中, 需要开展更多关于植物不同器官P含量对N添加响应的研究进一步验证.此外, 植物的生殖器官(花、果实和种子)决定植物的生殖过程, 其养分(如P)含量等在植物种子扩散、幼苗育成过程中起关键作用(Lee & Fenner, 1989; Fujita et al., 2014).因此, 开展关于森林植物生殖器官P含量对N添加的响应研究有利于更好地理解N沉降对植物生殖过程的影响.为了更精确地了解N添加对植物P养分的影响, 除了综合考虑植物各营养器官和生殖器官的P含量变化以外, 还可结合同位素示踪法研究植物不同器官间P的转化和迁移过程, 如Lehmann等(2001)通过32P同位素示踪的方法研究不同果树根系对P养分的吸收活性, 发现不同果树的P吸收能力不同, 且果树根系从土壤中吸收32P后, 32P在果树体内迁移至叶片, 且在36天后叶片32P达到峰值. ...
Negative effects of fertilization on plant nutrient resorption 2 2016
... Mohren和Burger (1986)最早关注N沉降对森林植物P含量的影响, 研究自然N沉降下荷兰针叶林叶片的P含量变化, 发现外源N沉降使针叶P含量处于较低水平, N:P高达20.随后陆续有关于森林生态系统植物P含量对N沉降响应的研究, 有的研究发现植物叶片P含量增加(Liu et al., 2013a),有的没有显著变化(Weand et al., 2010),有的甚至减少(Tessier & Raynal, 2003; Yan et al., 2018).森林植物P含量对N沉降的响应会受到土壤类型及土壤发育阶段、植物本身遗传特性与环境适应性等多因素的影响.如不同气候区森林土壤养分状况存在差异, 热带和亚热带地区易受P限制, 而温带主要受N限制, 其对N沉降的响应敏感度存在差异(陈美领等, 2016; Yuan & Chen, 2016); 森林生态系统内部植物养分状况有很大差异, 甚至同一生态系统中不同物种或同一物种不同大小的个体受养分限制的程度也不相同, 进而对外源N养分输入的响应也就不同(Alvarez-Clare et al., 2013; Li et al., 2018). ...
... 养分重吸收是指养分从植物衰老器官中转移到其他器官供其重复利用以提高养分利用效率的过程(Vitousek, 1982).这一机制可减少植物对外部养分吸收的依赖, 体现了植物对养分可利用性适应的保守策略(Aerts, 1996).大量研究认为土壤养分有效性与植物养分重吸收能力密切相关.由于N添加加剧了生态系统的P限制, 植物会通过提高对衰老器官中P养分的吸收能力而充分利用P养分, 以最大限度地缓解生长过程中的P限制.如Sullivan等(2014)在夏威夷的施肥实验研究发现, 在N丰富而P限制的森林中, 植物的P重吸收效率增加, 表明N添加条件下植物可通过增加P的重吸收调控自身P养分状况; See等(2015)在Bartlett森林的研究也认为土壤中N有效性的增加会导致凋落物中P含量的减少以实现植物对P的保留.但也有研究指出, N添加不会显著改变植物P重吸收, 表明N、P之间的相互关系对P养分重吸收过程的影响是复杂的(Yuan & Chen, 2016; Zhang et al., 2017), 不仅受外源养分可利用性的影响, 而且与植物养分吸收利用的生理过程(Hodge, 2004; Wang et al., 2014; Kou et al., 2017; Yang, 2018)、叶片生长与寿命(Zhou et al., 2016)、植物维持元素化学计量稳态的特性(Deng et al., 2016)、不同生活型对环境改变的适应(Vergutz et al., 2012)等多种因素密切相关(Tian et al., 2018).因此, N添加条件下植物改变P的重吸收能力只是调控植物内部P含量变化程度的选择性途径之一. ...
Short-term effects of soil warming and nitrogen addition on the N:P stoichiometry of Cunninghamia lanceolata, in subtropical regions 3 2017
... 植物叶片光合作用决定着整个植物体的生长速率, 且叶片对外部环境变化较为敏感.因此, 在大多森林养分添加实验中, 叶片经常被选取作为取样器官, 通过测定其N、P含量或计算N:P来指示植物的养分限制状况(Koerselman, 1996; Sullivan et al., 2014; Tian et al., 2017).森林植物叶片P含量对N添加的响应没有一致的研究结果, 研究结果表现为增加(Crowley et al., 2012; Mayor et al., 2014)、没有显著变化(Li et al., 2016)或减少(陈微微等, 2017; Deng et al., 2017).一般来说, 外源N输入增加条件下, 植物根系吸收大量的N并传输到叶片中, 促进光合作用和碳水化合物的累积; 同时, 为维持有机体内N、P的平衡, 植物会投资大量的N以促进土壤中磷酸酶的活性(Marklein & Houlton, 2012; 田地, 2017), 提高土壤中植物可利用性P含量, 并促进对P的吸收, 进而提高植物叶片的P含量.如在美国东北部, Crowley等(2012)研究森林植物养分含量在自然N沉降梯度(0-14 kg·hm-2)下的响应, 发现7种落叶树种的叶片P含量随着N沉降增加而增加.然而, 当植物生物量增长速率大于植物P元素吸收速率时, 就会产生“生物量稀释作用”, 植物叶片P含量表现为无显著变化甚至减少(Sterner & Elser, 2002; Yan et al., 2016; Zhang et al., 2017). ...
... 从物种水平考虑, 森林植物叶片P含量对N添加的响应受种类、生活型、生长阶段的影响.在亚北极圈地区, Heerwaarden等(2003)研究常绿木本植物和落叶木本植物养分含量对N添加(NH4Cl, 10 g N·m-2·a-1)的响应.结果发现: 对于常绿木本植物Andromeda polifolia的叶片P含量没有表现出显著变化, 而一种岩高兰属植物Empetrum hermaphroditum表现为显著的负效应; 对于落叶木本植物, 笃斯越桔(Vaccinium uliginosum)和Betula nana均表现为显著的负效应.在亚热带常绿阔叶林中, 5年施N显著提高蕨类和草本植物的叶片P含量, 降低苔藓叶片P含量, 但对乔木和灌木叶片P含量无显著影响(王乔姝怡等, 2016).此外, 研究认为植物处于不同生长阶段时, 其P含量对N添加的响应也不一致: 幼苗阶段时植物P含量对施N响应更敏感(Tian et al., 2018).如马亚娟(2015)对3年生杉木(Cunninghamia lanceolata)幼苗进行低N处理(2-6 g N·pot-1), 结果发现施N显著降低杉木叶片P含量, 相对对照组下降了7.8%-24%; 对于杉木成熟植株, 低N处理(5 g·m-2·a-1)对叶片P含量影响不显著, 相对对照组仅下降了1.2% (赵月, 2016).原因是相对成熟植株, 幼苗植株处于生长旺盛期, 生物量增长速率高, 导致生物量“稀释作用”明显, 进而对各器官P含量产生显著影响(Chen et al., 2015; Zhang et al., 2017; Tian et al., 2018). ...
... 养分重吸收是指养分从植物衰老器官中转移到其他器官供其重复利用以提高养分利用效率的过程(Vitousek, 1982).这一机制可减少植物对外部养分吸收的依赖, 体现了植物对养分可利用性适应的保守策略(Aerts, 1996).大量研究认为土壤养分有效性与植物养分重吸收能力密切相关.由于N添加加剧了生态系统的P限制, 植物会通过提高对衰老器官中P养分的吸收能力而充分利用P养分, 以最大限度地缓解生长过程中的P限制.如Sullivan等(2014)在夏威夷的施肥实验研究发现, 在N丰富而P限制的森林中, 植物的P重吸收效率增加, 表明N添加条件下植物可通过增加P的重吸收调控自身P养分状况; See等(2015)在Bartlett森林的研究也认为土壤中N有效性的增加会导致凋落物中P含量的减少以实现植物对P的保留.但也有研究指出, N添加不会显著改变植物P重吸收, 表明N、P之间的相互关系对P养分重吸收过程的影响是复杂的(Yuan & Chen, 2016; Zhang et al., 2017), 不仅受外源养分可利用性的影响, 而且与植物养分吸收利用的生理过程(Hodge, 2004; Wang et al., 2014; Kou et al., 2017; Yang, 2018)、叶片生长与寿命(Zhou et al., 2016)、植物维持元素化学计量稳态的特性(Deng et al., 2016)、不同生活型对环境改变的适应(Vergutz et al., 2012)等多种因素密切相关(Tian et al., 2018).因此, N添加条件下植物改变P的重吸收能力只是调控植物内部P含量变化程度的选择性途径之一. ...
Genetic variations in root morphology and phosphorus efficiency of Pinus massoniana, under heterogeneous and homogeneous low phosphorus conditions 1 2013
... 从物种水平考虑, 森林植物叶片P含量对N添加的响应受种类、生活型、生长阶段的影响.在亚北极圈地区, Heerwaarden等(2003)研究常绿木本植物和落叶木本植物养分含量对N添加(NH4Cl, 10 g N·m-2·a-1)的响应.结果发现: 对于常绿木本植物Andromeda polifolia的叶片P含量没有表现出显著变化, 而一种岩高兰属植物Empetrum hermaphroditum表现为显著的负效应; 对于落叶木本植物, 笃斯越桔(Vaccinium uliginosum)和Betula nana均表现为显著的负效应.在亚热带常绿阔叶林中, 5年施N显著提高蕨类和草本植物的叶片P含量, 降低苔藓叶片P含量, 但对乔木和灌木叶片P含量无显著影响(王乔姝怡等, 2016).此外, 研究认为植物处于不同生长阶段时, 其P含量对N添加的响应也不一致: 幼苗阶段时植物P含量对施N响应更敏感(Tian et al., 2018).如马亚娟(2015)对3年生杉木(Cunninghamia lanceolata)幼苗进行低N处理(2-6 g N·pot-1), 结果发现施N显著降低杉木叶片P含量, 相对对照组下降了7.8%-24%; 对于杉木成熟植株, 低N处理(5 g·m-2·a-1)对叶片P含量影响不显著, 相对对照组仅下降了1.2% (赵月, 2016).原因是相对成熟植株, 幼苗植株处于生长旺盛期, 生物量增长速率高, 导致生物量“稀释作用”明显, 进而对各器官P含量产生显著影响(Chen et al., 2015; Zhang et al., 2017; Tian et al., 2018). ...
施N肥对会同杉木人工林C:N:P生态化学计量特征的影响 1 2016
... 从物种水平考虑, 森林植物叶片P含量对N添加的响应受种类、生活型、生长阶段的影响.在亚北极圈地区, Heerwaarden等(2003)研究常绿木本植物和落叶木本植物养分含量对N添加(NH4Cl, 10 g N·m-2·a-1)的响应.结果发现: 对于常绿木本植物Andromeda polifolia的叶片P含量没有表现出显著变化, 而一种岩高兰属植物Empetrum hermaphroditum表现为显著的负效应; 对于落叶木本植物, 笃斯越桔(Vaccinium uliginosum)和Betula nana均表现为显著的负效应.在亚热带常绿阔叶林中, 5年施N显著提高蕨类和草本植物的叶片P含量, 降低苔藓叶片P含量, 但对乔木和灌木叶片P含量无显著影响(王乔姝怡等, 2016).此外, 研究认为植物处于不同生长阶段时, 其P含量对N添加的响应也不一致: 幼苗阶段时植物P含量对施N响应更敏感(Tian et al., 2018).如马亚娟(2015)对3年生杉木(Cunninghamia lanceolata)幼苗进行低N处理(2-6 g N·pot-1), 结果发现施N显著降低杉木叶片P含量, 相对对照组下降了7.8%-24%; 对于杉木成熟植株, 低N处理(5 g·m-2·a-1)对叶片P含量影响不显著, 相对对照组仅下降了1.2% (赵月, 2016).原因是相对成熟植株, 幼苗植株处于生长旺盛期, 生物量增长速率高, 导致生物量“稀释作用”明显, 进而对各器官P含量产生显著影响(Chen et al., 2015; Zhang et al., 2017; Tian et al., 2018). ...
Leaf resorption efficiency in relation to foliar and soil nutrient concentrations and stoichiometry of Cunninghamia lanceolata, with stand development in southern China 1 2016
... 养分重吸收是指养分从植物衰老器官中转移到其他器官供其重复利用以提高养分利用效率的过程(Vitousek, 1982).这一机制可减少植物对外部养分吸收的依赖, 体现了植物对养分可利用性适应的保守策略(Aerts, 1996).大量研究认为土壤养分有效性与植物养分重吸收能力密切相关.由于N添加加剧了生态系统的P限制, 植物会通过提高对衰老器官中P养分的吸收能力而充分利用P养分, 以最大限度地缓解生长过程中的P限制.如Sullivan等(2014)在夏威夷的施肥实验研究发现, 在N丰富而P限制的森林中, 植物的P重吸收效率增加, 表明N添加条件下植物可通过增加P的重吸收调控自身P养分状况; See等(2015)在Bartlett森林的研究也认为土壤中N有效性的增加会导致凋落物中P含量的减少以实现植物对P的保留.但也有研究指出, N添加不会显著改变植物P重吸收, 表明N、P之间的相互关系对P养分重吸收过程的影响是复杂的(Yuan & Chen, 2016; Zhang et al., 2017), 不仅受外源养分可利用性的影响, 而且与植物养分吸收利用的生理过程(Hodge, 2004; Wang et al., 2014; Kou et al., 2017; Yang, 2018)、叶片生长与寿命(Zhou et al., 2016)、植物维持元素化学计量稳态的特性(Deng et al., 2016)、不同生活型对环境改变的适应(Vergutz et al., 2012)等多种因素密切相关(Tian et al., 2018).因此, N添加条件下植物改变P的重吸收能力只是调控植物内部P含量变化程度的选择性途径之一. ...
 ,1,*, 田地21
,1,*, 田地21 
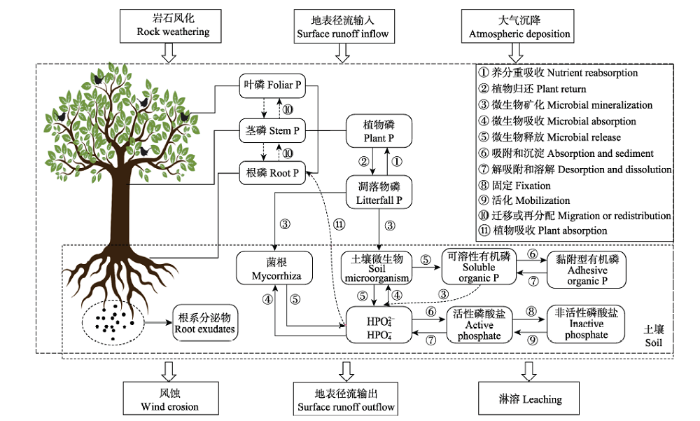 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}