陈林
, , 王磊, 杨新国, 宋乃平
, * , 李月飞
* , 苏莹
* , 卞莹莹
* , 祝忠有
* , 孟文婷
* 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地, 银川 750021;宁夏大学西北土地退化与生态恢复国家重点实验室培育基地, 银川 750021
Reproductive characteristics of Artemisia scoparia and the analysis of the underlying soil drivers in a desert steppe of China CHEN Lin
, , WANG Lei, YANG Xin-Guo, SONG Nai-Ping
, * , LI Yue-Fei
* , SU Ying
* , BIAN Ying-Ying
* , ZHU Zhong-You
* , MENG Wen-Ting
* Breeding Base for State Key Laboratory of Land Degradation and Ecological Restoration in Northwest China, Ningxia University, Yinchuan 750021, China;and Key Laboratory for Restoration and Reconstruction of Degraded Ecosystem in Northwest China of Ministry of Education, Ningxia University, Yinchuan 750021, China
通讯作者: * songnp@163.com 编委: 王雷
责任编辑: 李敏
收稿日期: 2018-08-25
接受日期: 2018-12-14
网络出版日期: 2019-01-20
基金资助: 国家重点研发计划 .2016YFC0500709 宁夏高等学校一流学科建设(生态学)资助项目 .NXYLXK2017B06 宁夏自然科学基金 .2018AAC03037 第三批宁夏青年科技人才托举工程项目 .TJGC2018068
Received: 2018-08-25
Accepted: 2018-12-14
Online: 2019-01-20
Fund supported: Supported by the National Key R&D Program of China .2016YFC0500709 Ningxia University Top-ranking Discipline Construction Funding Project (Ecology) .NXYLXK2017B06 the Natural Science Foundation of Ningxia .2018AAC03037 The Third Batch of Ningxia Youth Talents Supporting Program .TJGC2018068
摘要 繁殖是植物生命活动的重要环节, 了解植物的繁殖特征是解释植物生态适应性和制定有效管理措施的重要依据。该研究以荒漠草原猪毛蒿(Artemisia scoparia )种群为研究对象, 通过测定不同土壤类型的理化性质和猪毛蒿的繁殖特征, 以期探讨影响其繁殖特征的主要土壤驱动因子。结果表明: 灰钙土、风沙土和基岩风化残积土的水溶性碳含量、全氮含量、全磷含量、全盐含量、土壤水分含量、土壤硬度存在显著差异。猪毛蒿平均个体大小、单株头状花序的平均质量和数量均以灰钙土生境下最大, 基岩风化残积土最小。繁殖分配在不同土壤类型下无显著差异, 但与单个头状花序质量、单株头状花序数量和质量间呈极显著正相关关系。单株头状花序数量与单个头状花序质量间呈负相关关系。在风沙土生境下, 单株头状花序质量主要受到土壤水溶性碳含量、 土壤水分含量以及pH值的共同影响; 单株头状花序数量受全盐含量的影响最大; 繁殖分配和单个头状花序质量主要受全碳含量的影响。灰钙土生境下, 单株头状花序质量与土壤水溶性碳含量、 土壤水分含量和有机碳含量呈正相关关系; 速效氮含量显著影响着单株头状花序数量。而基岩风化残积土生境下, 繁殖特征的变异主要受到土壤水溶性碳含量、 土壤硬度、 土壤水分含量、 全磷和速效磷含量的影响。综合分析发现, 土壤因子对猪毛蒿繁殖特征的影响程度不同, 其中单株头状花序数量和质量极显著地受到土壤水溶性碳含量和土壤水分含量的影响, 繁殖分配和单个头状花序质量与土壤水溶性碳含量、土壤水分含量呈负相关关系。因此, 土壤水溶性碳含量和土壤水分是荒漠草原地区影响猪毛蒿种群繁殖特征的主要土壤因子。 关键词: 荒漠草原 ;
猪毛蒿 ;
繁殖特征 ;
土壤因子 ;
冗余分析 Abstract Aims Artemisia scoparia and analyze its main soil driving factors in a desert steppe of China.Methods A. scoparia population in a desert steppe, and examined physicochemical properties of different soil types and reproductive characteristics of A. scoparia with the redundancy analysis.Important findings C ws ), total nitrogen content (TN ), total phosphorus content (TP ), total salt content (TS ), soil moisture (M s ) and soil hardness (SH ) of calcareous soil (SS), aeolian sand soil (ASS) and weathered residual soil (WB). The average mass (M a ), the number (N i ) of average individual head inflorescence, and the average individual size (S ai ) of a single plant were the largest in the SS habitat, followed by ASS and WB. No significant difference was observed in reproductive allocation (R a ) of A. scoparia in different soil types. An extremely significant positive correlation was observed between R a and the quality of single capitate inflorescence (M e ), N i and M a , respectively, while N i was negatively correlated with M e . The variation of the reproductive characteristics of A. scoparia was mainly affected by soil water soluble carbon content (C ws ), soil hardness (SH ), soil moisture (M s ), TP and available phosphorus content (AP ) in WB habitat. M a was mainly affected by C ws , M s and pH value; N i was mainly affected by total salt content (TS ); R a and M e were mainly affected by total carbon content (C t ) in the ASS habitat. M a was positively correlated with C ws , M s and C o , while the correlation between M a and C o was not significant; N i was significantly affected by available nitrogen content (AN ) in the SS habitat. A comprehensive analysis of the three soil types showed that the total explanatory amount of soil factors affecting the reproductive characteristics of A. scoparia was 30.74%. M a and N i were significantly affected by C ws and M s . R a and M e were negatively correlated with C ws and M s though they are not statistically significant. We conclude that C ws and M s are the main soil factors affecting the reproductive characteristics of A. scoparia populations in the desert grassland.Keywords: desert steppe ;
Artemisia scoparia ;
reproductive characteristic ;
soil factor ;
redundancy analysis PDF (14190KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 本文引用格式 陈林, 王磊, 杨新国, 宋乃平, 李月飞, 苏莹, 卞莹莹, 祝忠有, 孟文婷. 荒漠草原猪毛蒿种群繁殖特征的土壤驱动因子分析.
植物生态学报 [J], 2019, 43(1): 65-76 doi:10.17521/cjpe.2018.0211
CHEN Lin.
Reproductive characteristics of Artemisia scoparia and the analysis of the underlying soil drivers in a desert steppe of China .
Chinese Journal of Plant Ecology [J], 2019, 43(1): 65-76 doi:10.17521/cjpe.2018.0211
繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 )。不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 )。因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 )。同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a )。自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 )。
猪毛蒿(
Artemisia scoparia )是菊科蒿属草本, 是一种C
3 植物, 却呈现类似CAM植物的叶片特性(
杜华栋等, 2016 ), 广布于欧亚大陆温带至亚热带地区(
马德滋, 1986 ;
李克昌和郭思加, 2012 ), 在局部地区成为植物群落的优势种, 单个样地覆盖度可超过50% (
焦菊英等, 2008 )。目前, 国内外****对蒿属植物的研究多集中在植物生理、化学和药理学/配药学领域(
陈林等, 2017 ), 特别是猪毛蒿的化学成分及其医药用途方面是研究热点, 但较少考虑土壤因子对猪毛蒿繁殖特征的影响, 特别是荒漠草原异质生境条件下, 以不同土壤因子为驱动力, 对猪毛蒿繁殖特征及其影响因素的研究很少有报道。
在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响。宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 )。因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 )。而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 )。分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容。
但从不同角度研究同一个问题, 经常得出矛盾的结论(
Mayer et al ., 2011 )。所以从不同的因素中综合分析并找出影响植物繁殖特征的主要选择压力, 是摆在植物繁殖生态学家面前的挑战(
任明迅等, 2012 )。在众多的环境因子中, 土壤因子往往制约着荒漠植物生命过程, 起着关键作用(
霍举颂等, 2017 )。因此本研究基于对宁夏荒漠草原地区猪毛蒿种群繁殖特征的调查, 以土壤类型和驱动因子为切入点, 采用约束性排序中的冗余分析来探讨: (1)不同土壤类型对猪毛蒿的繁殖特征是否有影响; (2)哪些土壤因子对猪毛蒿繁殖特征起主要影响作用。通过此研究揭示异质生境下猪毛蒿繁殖特征与土壤驱动因子的关系及其机制, 以期为荒漠草原恢复和管理提供理论参考和科学依据。
1 研究方法 1.1 研究区概况和样地选择 本研究区位于宁夏中部干旱带的荒漠草原, 为鄂尔多斯高原缓坡起伏地带。干旱少雨, 蒸发量大, 冬春两季风大沙多, 属典型的中温带大陆性气候(
宋乃平等, 2018 )。近50年年平均气温为8.46 ℃, 年降水量为276.3 mm, 年日照时间为2 862.6 h。该区域植被主要受水分因素限制(
Ramakrishna et al ., 2003 ), 表现为物种多样性较低、群落结构简单、沙生特征明显, 地下水资源匮乏且植物常常无法利用(
刘小丹等, 2015 ;
Tang et al ., 2017 ;
Wang et al ., 2018 )。
采用典型样地调查方法, 样地设置主要考虑土壤类型和地形地貌等要素, 在宁夏中部荒漠草原区选取39个猪毛蒿群落样地(
图1 ; 附录I), 为减少抽样误差, 每个样地分别设置3-5个1 m × 1 m的样方, 样方间隔距离大于10 m, 共190个样方。其中, 风沙土生境105个样方, 灰钙土生境为65个, 基岩风化沉积土生境为20个。取样前1个月内无>5 mm单次降水且近7天内无降水事件发生。用GPS定位并记录采样点的经纬度和海拔等信息。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1宁夏荒漠草原猪毛蒿采集样地分布点。 Fig. 1Sampling sites of Artemisia scoparia in the desert steppe of Ningxia. 1.2 植株特征和繁殖特征测定 盖度测量选择常用的目测估算法; 高度和冠幅为样方内随机选取有代表性的3-5株, 用卷尺测量地上部分自然高度和冠幅后取其平均值。在样方内随机选取生长健康, 没有人为干扰和病虫害影响的个体, 挖取整株, 分株装于信封带回实验室。再次清理干净后, 测量其主根最长部分记为根长, 用游标卡尺测量其茎部直径记为茎粗, 把植株分为头状花序、根、茎和叶等部分, 分别装入信封并置于(65 ± 2) ℃烘箱中烘干至恒定质量, 然后用1/10000电子天平称取质量。地上生物量为头状花序、茎和叶生物量之和。总生物量为地上生物量和根生物量之和。根冠比为根生物量与地上生物量的比值。营养器官生物量为根、茎和叶生物量之和。
本研究中猪毛蒿个体大小以地上生物量干质量表示(
刘左军等, 2002 ;
何玉惠等, 2009 ); 繁殖器官大小以头状花序干质量表示。统计单株猪毛蒿的生物量、繁殖体数量、繁殖体质量, 并计算单个繁殖体质量, 单个繁殖体质量为单株繁殖体质量与单株繁殖体数量的比值。繁殖分配为繁殖器官干质量占总生物量干质量的百分比。
1.3 土壤因子测定 在所调查的植株样方内, 采用便携式土壤水分测量仪(ML3, Delta-T, Burwell, UK)和数显式土壤紧实度仪(SC-900, Spectrum Technologies, Aurora, USA)分别对0-10和10-20 cm土层土壤水分含量(
M s )和0-5、5-10和10-20 cm土壤硬度(
SH )进行测量。pH值采用电位法测定, 水土质量比为5:1; 全盐含量(
TS )通过电导率换算而成; 易氧化碳含量(
C eo )采用KMnO
4 氧化比色法测定; 用蒸馏水浸提0.45 μm滤膜过滤后采用K
2 CrO
7 法测定水溶性碳含量(
C ws ); 有机碳含量(
C o )采用K
2 CrO
7 氧化法测定; 全碳含量(
C t )采用元素分析仪(vario MACRO, Elementar, Langenselbold, Germany)测定; 全磷含量(
TP )采用钼锑抗比色法测定; 速效磷含量(
AP )采用0.5 mol·L
-1 NaHCO
3 浸提-钼锑抗比色法测定; 速效钾含量(
AK )采用乙酸铵浸提-火焰光度法测定; 全氮含量(
TN )采用凯氏定氮法测定; 碱解氮含量(
AN )采用碱解扩散法测定(
鲍士旦, 2000 )。
1.4 数据统计与分析 所有数据在Excel 2016和SPSS 21.0软件中进行统计、计算、分析和制图, 采取单因素方差分析和最小显著差异(
LSD )法分析异质生境土壤理化性质、猪毛蒿株高、茎粗、根长、根质量以及各生物量等方面的差异。0-20 cm土层土壤含水量为0-10和10-20 cm土层土壤含水量的加权平均值。0-20 cm土层土壤硬度为0-5、5-10和10-20 cm土层土壤硬度的加权平均值。变异系数为标准偏差与平均值之比。采用RStudio和Canoco 5.0软件绘制猪毛蒿繁殖特征和植株特征的相关关系图, 分析不同土壤类型下猪毛蒿繁殖特征和土壤因子之间的关系。将样地内猪毛蒿的繁殖特征指标设为响应变量, 包括单株头状花序数量、单个头状花序质量、单株头状花序质量和繁殖分配; 将土壤因子中的不同深度
M s 、不同深度
SH 、
C eo 、
C ws 、
C o 、
C t 、
AP 、
AK 、
AN 、
TN 、
TP 、pH值和
TS 等18个指标综合构建解释变量。选择用RDA或CCA分析: 需先做去趋势对应分析(DCA), 根据分析结果中Axis Lengths第一轴的大小, 如果大于4.0, 就应选CCA (基于单峰模型, 典范对应分析); 如果在3.0-4.0之间, 选RDA和CCA均可; 如果小于3.0, RDA的结果则更合理(基于线性模型, 冗余分析); 本操作在RStudio软件中执行。分析结果表明第一排序轴长小于3.0, 因此本项研究用RDA进行猪毛蒿种群繁殖特征与土壤因子的关系分析, 使用蒙特卡洛法, 用999个序列检验判断变量的解释能力。
2 结果和分析 2.1 土壤因子特性分析 从
表1 可以看出, 所调查样地中的3种土壤类型理化性质存在一定的差异。
C ws 、
TN 、
TP 、
M s 间存在极显著差异(
p < 0.01),
TS 和
SH (0-20 cm)存在显著差异(
p < 0.05)。其中, 风沙土中的
C ws 极显著低于灰钙土和基岩风化沉积土(
p < 0.01); 基岩风化沉积土中的
TN 极显著低于灰钙土和风沙土(
p < 0.01); 风沙土中的
TP 则极显著低于灰钙土和基岩风化沉积土(
p < 0.01); 表层
M s (0-20 cm)则是灰钙土中极显著高于基岩风化沉积土(
p < 0.01), 风沙土最低; 风沙土中的
TS 显著低于灰钙土和基岩风化沉积土(
p < 0.05);
SH (0-20 cm)则表现为灰钙土>基岩风化沉积土>风沙土。而
C eo 、
C o 、
C t 、
AP 、
AK 、
AN 、pH值、
SH (5 cm、10 cm和20 cm)间则无显著差异(
p > 0.05)。
Table 1 表1 表1 猪毛蒿种群不同生境中土壤特性的比较(平均值±标准误差)
Table 1
Comparisons of soil properties in three soil types from the different habitats of Artemisia scoparia (mean ± SE ) 土壤特性 ASS SS WB ANOVA F p C eo (mg·g-1 )1.022 ± 0.760 1.088 ± 1.065 0.639 ± 0.337 2.192 0.115 C ws (mg·g-1 )0.048 ± 0.178 0.209 ± 0.439 0.297 ± 0.557 7.006 <0.001*** C o (%)0.449 ± 0.477 0.450 ± 0.187 0.424 ± 0.139 0.042 0.959 Ct (%)0.541 ± 0.242 0.605 ± 0.401 0.654 ± 0.268 1.594 0.206 AP (mg·kg-1 )2.776 ± 1.875 2.625 ± 1.854 1.755 ± 1.170 2.672 0.072 AK (mg·kg-1 )81.592 ± 37.170 71.040 ± 40.591 70.963 ± 40.242 1.668 0.192 AN (mg·kg-1 )230.668 ± 123.556 191.756 ± 54.719 237.687 ± 285.222 1.787 0.170 TN (%)0.034 ± 0.013 0.040 ± 0.013 0.027 ± 0.011 9.938 <0.001*** TP (%)0.029 ± 0.013 0.037 ± 0.010 0.038 ± 0.010 10.480 <0.001*** pH 9.076 ± 0.209 9.092 ± 0.165 9.090 ± 0.288 0.191 0.827 TS (g·kg-1 )0.265 ± 0.045 0.315 ± 0.198 0.317 ± 0.155 3.409 0.035* M s . 10 cm (%)7.591 ± 1.997 10.268 ± 3.757 8.990 ± 2.813 17.556 <0.001*** M s . 20 cm (%)7.309 ± 2.588 9.902 ± 4.008 8.768 ± 1.898 13.529 <0.001*** M s . 0-20 cm (%)7.489 ± 1.707 10.031 ± 3.274 8.747 ± 1.652 22.330 <0.001*** SH . 5 cm (kPa)1 003.436 ± 516.201 1 245.283 ± 796.110 1 060.000 ± 560.993 2.824 0.062 SH . 10 cm (kPa)1 377.634 ± 709.554 1 697.746 ± 1 196.433 1 611.250 ± 870.530 2.058 0.131 SH . 20 cm (kPa)2 032.554 ± 1303.646 2 465.967 ± 1 338.089 2 281.400 ± 1 124.882 2.140 0.121 SH . 0-20 cm (kPa)1 471.208 ± 678.777 1 807.194 ± 896.430 1 659.950 ± 726.002 3.714 0.026*
ASS, aeolian sand soil; SS, calcareous soil; WB, weathered residual soil. AK, available potassium content; AN, alkaline nitrogen content; AP, available phosphorus content; C eo , easily oxidized organic carbon content; C o , organic carbon content; C t , total carbon content; C ws , soil water soluble carbon content; M s , soil moisture content; SH, soil hardness; TN, total nitrogen content; TP, total phosphorus content; TS, total salt content。*, p < 0.05; **, p < 0.01; ***, p < 0.001. ASS, 风沙土; SS, 灰钙土; WB, 基岩风化沉积土。AK, 速效钾含量; AN, 碱解氮含量; AP, 速效磷含量; C eo , 易氧化碳含量; C o , 有机碳含量; C t , 全碳含量; C ws , 水溶性碳含量; M s , 土壤水分含量; SH, 土壤硬度; TN, 全氮含量; TP, 全磷含量; TS, 全盐含量。*, p < 0.05; **, p < 0.01; ***, p < 0.001。 新窗口打开 |
下载CSV 2.2 不同土壤类型下猪毛蒿的繁殖特征 猪毛蒿种群在荒漠草原普遍存在, 不同土壤类型下均有生长, 本研究中猪毛蒿个体大小以单株总生物量表示, 不同生境下的大小排序为: 灰钙土(10.60 g) >风沙土(7.07 g) >基岩风化残积土(1.28 g)。基岩风化残积土生境下猪毛蒿的繁殖分配(0.49 ± 0.08)略低于风沙土(0.54 ± 0.13)和灰钙土(0.52 ± 0.11), 但三者间均未达到显著差异(
p > 0.05)。灰钙土生境下单株头状花序数量的平均值(7 283.31个)远高于风沙土(4 700.94个)和基岩风化残积土(941.47个)(
p < 0.05), 风沙土中的变异系数高达307%, 单株头状花序数量最大值高达108 841个。基岩风化残积土生境下单株头状花序质量平均值(0.59 g)显著低于灰钙土(4.82 g)和风沙土生境(3.25 g) (
p < 0.05)。风沙土中单个头状花序质量(8.1 × 10
-4 g)和灰钙土(7.2 × 10
-4 g)、基岩风化残积土(6.1 × 10
-4 g)生境下差异不大(
p > 0.05), 最大值仅为42.2 × 10
-4 g。
Table 2 表2 表2 不同土壤类型下猪毛蒿的繁殖特征
Table 2
Reproductive characteristics of Artemisia scoparia 繁殖特征 土壤类型 最大值 最小值 平均值 标准偏差 变异系数 S ai ASS 157.89 0.16 7.07 19.75 2.79 SS 138.22 0.25 10.60 25.62 2.42 WB 4.02 0.29 1.28 1.23 0.95 R a ASS 0.78 0.05 0.54 0.13 0.24 SS 0.73 0.21 0.52 0.11 0.20 WB 0.61 0.33 0.49 0.08 0.14 N i ASS 108 841.00 107.00 4 700.94 14 557.55 3.07 SS 79 019.00 122.00 7 283.31 17 272.63 2.37 WB 3 414.00 202.00 941.47 988.64 1.05 M e ASS 42.2 1.8 8.1 4.7 5.5 SS 15.0 2.9 7.2 2.3 3.2 WB 7.7 4.7 6.1 1.0 1.6 M a ASS 71.25 0.06 3.25 9.14 2.79 SS 68.30 0.06 4.82 12.41 2.58 WB 2.13 0.14 0.59 0.63 1.06
ASS, aeolian sand soil; SS, calcareous soil; WB, weathered residual soil. M a , capitulum mass of each individual plant (g); M e , each acptitulum mass (g × 10-4 ); N i , capitulum number of each individual plant; R a , reproductive allocation; S ai , total biomass (g). ASS, 风沙土; SS, 灰钙土; WB, 基岩风化沉积土。M a , 单株头状花序质量(g); M e , 单个头状花序质量(g × 10-4 ); N i , 单株头状花序数量; R a , 繁殖分配; S ai , 个体大小, 以单株总生物量表示(g)。 新窗口打开 |
下载CSV 2.3 繁殖特征和植株性状的相关性分析 植物的生长与繁殖能力是植物适应环境的体现, 是统一的整体, 所以植株繁殖特征也必然与植株的各生长特征紧密联系(
刘志民等, 2003b )。猪毛蒿繁殖特征间的相关性分析表明, 繁殖分配与单个头状花序质量、单株头状花序数量与质量间呈极显著正相关关系(
r = 0.44,
r = 0.97,
p < 0.001), 单株头状花序数量与单个头状花序质量呈显著负相关关系(
r = -0.17,
p < 0.05)。繁殖分配与单株头状花序数量和单株头状花序质量间、单个头状花序质量与单株头状花序质量间均无相关关系(
p > 0.05)。
植株性状特征之间的相关性分析表明, 除了盖度与平均冠幅、平均高度与根长、根冠比与根生物量、根长、营养器官生物量、平均冠幅和茎粗间无显著相关关系(
p > 0.05)外, 所测定的其他植株特征之间均呈显著或极显著相关关系(
p < 0.05或
p < 0.001)。
猪毛蒿繁殖特征与植株性状特征之间的相关性分析表明, 繁殖分配与平均高度和茎粗呈极显著负相关关系(
r = -0.31,
p < 0.001;
r = -0.28,
p < 0.001)。单株头状花序数量与根冠比呈显著负相关关系(
r = -0.19,
p < 0.05), 与其他指标间均呈现极显著正相关关系(
p < 0.001)。单个头状花序质量仅与根长存在显著正相关关系(
r = 0.17,
p < 0.05), 而与其他指标均无相关关系(
p > 0.05)。单株头状花序质量与根冠比呈显著负相关关系(
r = -0.18,
p < 0.05), 与其他指标间呈极显著正相关关系(
p < 0.001)。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2猪毛蒿繁殖特征与植株性状特征间的Pearson相关性。AB, 地上总生物量(g); Cov , 盖度(%); CW, 平均冠幅(cm2 ); Hei, 平均高度(cm); M a , 单株头状花序质量(g); M e , 单个头状花序质量(g × 10-4 ); N i , 单株头状花序数量; R a , 繁殖分配; RB, 营养器官生物量(g); RL , 根长(cm); RS, 根冠比; RW, 根生物量(g); S ai , 总生物量(g); SB , 茎生物量(g); SD, 茎粗(mm)。*表示相关性水平; *, p < 0.05; **, p < 0.01; ***, p < 0.001。 Fig. 2Pearson’s correlation for reproductive characteristics and plant characteristics of Artemisia scoparia. AB, aboveground biomass (g); Cov, coverage (%); CW, crown width (cm2 ); Hei, average height (cm); M a , capitulum mass of each individual plant (g); M e , each acptitulum mass (g × 10-4 ); N i , capitulum number of each individual plant; R a , reproductive allocation; RB , reproductive biomass (g); RL, root length (cm); RS, root/shoot ratio; RW, root biomass (g); S ai , total biomass (g); SB, stem biomass (g); SD, stem diameter (mm). * meant the relevant level; *, p < 0.05; **, p < 0.01; ***, p < 0.001. 2.4 繁殖特征与土壤因子的关系 2.4.1 基于不同土壤类型的RDA分析
RDA排序图可以直观地表示不同土壤类型下猪毛蒿繁殖特征与土壤因子间的关系, 以及各土壤因子间的相互关系。带有空心箭头的黑色线段表示不同的土壤因子, 箭头所处的象限表示环境因子与排序轴间的正负相关性。箭头连线的长度表示该土壤因子对繁殖特征的影响程度, 连线越长, 说明相关性越大, 能解释影响程度的比例也越多。箭头连线和排序轴的夹角以及箭头连线之间的夹角表示相关性, 锐角表示成正相关关系, 钝角则表示成负相关关系。夹角越小, 相关性越高。
猪毛蒿繁殖特征与土壤因子关系的冗余分析结果显示, 在风沙土生境下, 各土壤因子对猪毛蒿繁殖特征影响的前两轴解释量为29.41%, 其中单株头状花序质量主要受到
C ws 、
M s 以及pH值的正向影响, 而与
SH 、
AN 、
AK 和
C eo 呈负相关关系。单株头状花序数量受
TS 的正向影响最大, 受
C t 和
C o 的负向影响较大(
图3A )。繁殖分配和单个头状花序质量主要受
C t 的影响。在灰钙土生境下, 土壤因子的前两轴解释量为64.39%, 单株头状花序质量与
C ws 、
M s 以及
C o 呈正相关关系(
图3B ), 说明随着
C ws 和不同土层
M s 的增加, 单株头状花序质量也越大。在基岩风化沉积土生境下, 猪毛蒿繁殖特征的变异主要受到
C ws 、
SH 、
M s 、
TP 、
AK 的影响, 其中99.62%的变异能被前两轴解释(
图3C )。单株头状花序质量主要受到
C ws 的影响, 其值越大, 单株头状花序质量越大。单株头状花序数量受
SH 的影响最大。单个头状花序质量和繁殖分配与
SH 、
TP 和
M s 等呈负相关, 说明这几个指标值越大, 单个头状花序质量和繁殖分配则越小。综合分析来看, 土壤因子对猪毛蒿繁殖特征影响的总解释量为30.74%, 单株头状花序数量和质量受
C ws 的影响最大, 其次为
M s , 与
TP 和pH值也呈正相关关系, 但影响较小, 而与
SH 和
C eo 几乎无关。繁殖分配和单个头状花序质量与
C ws 、
M s 呈负相关关系(
图3D )。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3猪毛蒿繁殖特征与土壤因子关系的冗余分析排序。A, 风沙土。B, 灰钙土。C, 基岩风化沉积土。D, 综合分析。M a , 单株头状花序质量(g); M e , 单个头状花序质量(g × 10-4 ); N i , 单株头状花序数量; R a , 繁殖分配。AK, 速效钾含量; AN, 碱解氮含量; AP, 速效磷含量; C eo , 易氧化碳含量; C o , 有机碳含量; C t , 全碳含量; C ws , 水溶性碳含量; M s . 10 cm, 0-10 cm土壤水分含量; M s . 20 cm, 10-20 cm土壤水分含量; M s . 0-20 cm, 0-20 cm土壤水分含量平均值; pH, pH值; SH . 5 cm, 0-5 cm土壤硬度; SH . 10 cm, 5-10 cm土壤硬度; SH . 20 cm, 10-20 cm土壤硬度; SH . 0-20 cm, 0-20 cm土壤硬度平均值; TN, 全氮含量; TP, 全磷含量; TS , 全盐含量。 Fig. 3Relationship between reproductive characters and soil factors in redundancy analyses. A, aeolian sand soil. B, calcareous soil. C, weathered residual soil. D, comprehensive analysis of three soil types. M a , capitulum mass of each individual plant (g); M e , each acptitulum mass (g × 10-4 ); N i , capitulum number of each individual plant; R a , reproductive allocation. AK, available potassium content; AN, available nitrogen content; AP, available phosphorus content; C eo , easily oxidized organic carbon content; C o , organic carbon content; C t , total carbon content; C ws , water soluble carbon content; M s . 10 cm, soil water content of 0-10 cm; M s . 20 cm, soil water content of 10-20 cm; M s . 0-20 cm, the average soil water content of 0-20 cm; pH, pH value; SH . 5 cm, soil hardness of 0-5 cm; SH . 10 cm, soil hardness of 5-10 cm; SH . 20 cm, soil hardness of 10-20 cm; SH . 0-20 cm, the average soil hardness of 0-20 cm; TN, total nitrogen content; TP, total phosphorus content; TS, total salt content. 2.4.2 土壤因子对繁殖特征变异的解释量
排序轴是反映综合生态梯度的抽象轴, 特征根值是反映相应的排序轴所集中的指标重要值矩阵中信息量的大小, 而相关系数表示的是各排序轴与真实环境梯度之间的相关性(
余敏等, 2013 )。因此进一步对每个繁殖特征进行Monte Carlo置换检验, 结果显示: 在基岩风化沉积土生境下, 对猪毛蒿繁殖特征的影响中,
C ws 的解释量高达71.7% (
p < 0.001), 其次是
TP (55.4%,
p < 0.001), 0-20 cm
M s 和
SH 的解释量分别为32.4%和43.6% (
p < 0.01), 而0-5、5-10和10-20 cm
SH 的解释量同样达到了显著或极显著水平(
p < 0.05或
p < 0.01)。
AP 的解释量也达到了21.9% (
p < 0.05)。在风沙土生境下, 各土壤因子的解释量均不高(<10%), 其中
C ws 、
TS 、
M s (0-20 cm)和
M s (0-10 cm)的解释量达到了显著水平或极显著水平(
p < 0.05或
p < 0.01)。在灰钙土生境下,
M s (0-20 cm)的解释量为47.9% (
p < 0.001), 0-10 cm和10-20 cm的
M s 解释量分别为43.2%和35.3%,
C ws 的解释量同样达到了极显著水平(22.1%,
p < 0.001),
AN 的解释量为10.0%, 达到了显著水平(
p < 0.05)。综合分析结果表明,
C ws 和
M s (0-20 cm)的解释量分别为19.9%和16.9%, 达到了极显著水平(
p < 0.001), 同样, 0-10 cm和10-20 cm
M s 的解释水平也达到了极显著水平(
p < 0.001), 说明0-20 cm土层的
M s 对猪毛蒿种群的繁殖特征有显著影响。而其他的土壤因子解释量均未能达到显著水平(
p > 0.05)。
Table 3 表3 表3 不同土壤类型下各土壤因子解释量和显著性检验结果
Table 3
Variance explained by soil indicators and significance of the test results 土壤因子 ASS SS WB 综合分析 Comprehensive analysis 解释量 F p 解释量 F p 解释量 F p 解释量 F p AK 0.10 < 0.1 0.817 6.02 3.7 0.053 3.10 0.6 0.468 1.45 2.6 0.101 AN 0.66 0.7 0.424 10.04 6.5 0.014 0.20 < 0.1 0.933 <0.01 < 0.1 0.969 AP 0.02 < 0.1 0.973 2.44 1.4 0.227 21.90 5.0 0.035 0.06 0.1 0.803 C eo 0.28 0.3 0.613 0.33 0.2 0.685 0.30 < 0.1 0.841 0.05 < 0.1 0.807 C o 1.34 1.3 0.247 4.17 2.5 0.112 2.70 0.5 0.487 0.28 0.5 0.468 pH 2.16 2.2 0.132 0.80 0.5 0.494 0.10 < 0.1 0.910 1.18 2.1 0.144 SH 0.25 0.2 0.644 0.55 0.3 0.590 43.60 13.9 0.002 0.17 0.3 0.593 SH . 5 cm1.57 1.6 0.208 1.93 1.1 0.282 20.90 4.8 0.038 0.27 0.5 0.499 SH . 10 cm0.36 0.4 0.572 0.12 < 0.1 0.849 29.00 7.3 0.014 0.31 0.6 0.466 SH. 20 cm0.11 0.1 0.783 5.25 3.2 0.071 37.30 10.7 0.004 0.05 < 0.1 0.820 M s 6.44 6.8 0.011 47.88 53.3 < 0.001 32.40 8.6 0.007 16.89 36.4 < 0.001 M s . 10 cm8.46 9.2 0.003 43.20 44.1 < 0.001 15.40 3.3 0.084 16.62 35.7 < 0.001 M s . 20 cm0.68 0.7 0.407 35.26 31.6 < 0.001 18.20 4.0 0.062 8.79 17.3 < 0.001 C t 2.81 2.9 0.087 0.71 0.4 0.484 3.00 0.6 0.471 0.26 0.5 0.497 TN 0.66 0.7 0.421 0.18 0.1 0.790 16.70 3.6 0.071 0.10 0.2 0.706 TP 0.13 0.1 0.769 0.98 0.6 0.467 55.40 22.3 < 0.001 1.32 2.4 0.122 TS 5.36 5.6 0.018 0.61 0.4 0.486 12.20 2.5 0.121 0.34 0.6 0.435 C ws 6.87 7.3 0.008 22.10 16.5 < 0.001 71.70 45.6 < 0.001 19.90 44.5 < 0.001
See Fig. 3 for soil indicator. 土壤因子同图3 。 新窗口打开 |
下载CSV 3 讨论和结论 植物种群的更新和个体生活史的完成均依赖于繁殖过程(
全杜娟等, 2012 ;
Larson & Funk, 2016 ), 植物的繁殖性状是植物资源投入-收益长期权衡的结果(
张大勇, 2004 ), 通过植株、花序等的资源分配可以很好地解释许多重要繁殖特征, 这在很大程度上协调了不同侧重点研究所得到的矛盾结论(
任明迅等, 2012 )。
3.1 异质生境对植物生长繁殖的影响 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 )。在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 )。反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 )。本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的。有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 )。薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 )。芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 )。而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 )。以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 )。
3.2 土壤因子对猪毛蒿繁殖特征的影响 C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分。植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源。土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 )。目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究。
有研究发现,
C ws 含量与降水量显著负相关, 是因为降水量的增加导致
C ws 淋溶、淋失作用加强, 从而
C ws 含量较低(
刘荣杰等, 2013 )。但本研究区降水较少, 降水的淋溶作用不足以使
C ws 发生淋失, 而
M s 相对较高的生境能促进枯枝落叶的分解, 进而增加了土壤中
C ws 的含量, 因此
C ws 与
M s 呈现出正相关关系。本研究区地下水位较低, 植物往往无法利用, 土壤中水分补给的主要来源是降水, 且具有很强的时空异质性和随机性, 导致该地区植物群落分布多呈斑块状(
宋乃平等, 2018 )。有研究结果表明荒漠植物数量特征的关键驱动力是
M s , 特别是表层
M s (0-20 cm)对荒漠植物群落数量特征起到非常重要的生态作用(
霍举颂等, 2017 )。本研究中, 单株头状花序质量和数量极显著地受到
M s 的影响,
C ws 与单株头状花序质量显著正相关, 这与有关****(
杜华栋等, 2016 )研究发现猪毛蒿叶片形态、解剖和生理指标的可塑性对立地
M s 和
C o 含量较为敏感的结果有相同之处。结合RDA分析图可知,
C ws 和
M s 是影响猪毛蒿繁殖特征的重要因子。
此外, 猪毛蒿虽然产生大量的头状花序, 即意味着开花数量较多, 但种子是否存在异型性及其在荒漠草原地区萌发率和种群动态如何, 这些都是今后该地区生态系统研究中值得考虑的重点内容。
致谢 感谢宁夏大学农学院本科生赵娟、王家锐、丁焜、王科鑫、王晓芳、贾静、马蓉、马静、王佳娟、张莹、禹小彦等同学在室内进行大量的猪毛蒿繁殖特征测定以及数据的录入和核对工作。
附录? 宁夏荒漠草原猪毛蒿样地信息 Supplement ? Basic information of the plots of
Artemisia scoparia in the desert steppe of Ningxia
http://www.plant-ecology.com/fileup/PDF/cjpe.2018.0211-S1.xlsx [1] Aysajan A 2018 ). Effect of habitat heterogeneity on floral trait differerntiation level in distylous species Primula nivalis . Acta Botanica Boreali-Occidentalia Sinica, 158 -165 . [本文引用: 1] [ 艾沙江·阿不都沙拉木 ( 2018 ). 异质生境对二型花柱植物雪地报春花性状分化的影响西北植物学报 , 38, 158 -165 .] [本文引用: 1] [2] Bao SD 2000 ). Soil Agricultural Chemistry Analysis . 3rd edn. China Agriculture Press, Beijing. [本文引用: 1] [ 鲍士旦 ( 2000 ). 土壤农化分析 . 第三版. 中国农业出版社, 北京.] [本文引用: 1] [3] Chen L Song NP Wang L Yang XG Li XB Su Y Li YF 2017 ). A bibliometric appraisal of research on Artemisia from 1986-2016. Acta Prataculturae Sinca, 223 -235 . [本文引用: 1] [ 陈林 , 宋乃平 , 王磊 , 杨新国 , 李学斌 , 苏莹 , 李月飞 ( 2017 ). 基于文献计量分析的蒿属植物研究进展草业学报 , 26, 223 -235 .] [本文引用: 1] [4] Chen T Wang YF 2014 ). Response of reproductive characteristics of Saussurea maceota Franch. to elevation at eastern Qinhai-Tibetan Plateau. Chinese Journal of Ecology, 3216 -3221 . [本文引用: 1] [ 陈亭 , 王一峰 ( 2014 ). 青藏高原东缘大耳叶风毛菊繁殖特征对海拔的响应生态学杂志 , 33, 3216 -3221 .] [本文引用: 1] [5] Chen Y Wang T Li PK Yao CL Yuan ZL Ye YZ 2016 ). Community characteristics and spatial distribution of dominant tree species in a deciduous broad-leaved forest of Muzhaling, Henan, ChinaChinese Journal of Plant Ecology, 40, 1179 -1188 . [本文引用: 1] [ 陈云 , 王婷 , 李培坤 , 姚成亮 , 袁志良 , 叶永忠 ( 2016 ). 河南木札岭温带落叶阔叶林群落特征及主要乔木空间分布格局植物生态学报 , 40, 1179 -1188 .] [本文引用: 1] [6] Dong M 1996 ). Plant clonal growth in heterogeneous habitats, risk-spreadingActa Phytoecologica Sinca, 20, 543 -548 . [本文引用: 1] [ 董鸣 ( 1996 ). 异质性生境中的植物克隆生长, 风险分摊植物生态学报 , 20, 543 -548 .] [本文引用: 1] [7] Du HD Jiao JY Kou M Miao F 2016 ). Adaptability of foliar morphological, anatomical, and physiological characteristics of the pioneer species Artemisia scoparia growing in a hilly-gully Loess Regin at different slope sites. Acta Ecologica Sinica, 2914 -2925 . [本文引用: 2] [ 杜华栋 , 焦菊英 , 寇萌 , 苗芳 ( 2016 ). 黄土高原先锋种猪毛蒿叶片形态解剖与生理特征对立地的适应性生态学报 , 36, 2914 -2925 .] [本文引用: 2] [8] Fang H Cheng S Lin E Yu G Niu S Wang Y Xu M Dang X Li L Wang L 2015 ). Elevated atmospheric carbon dioxide concentration stimulates soil microbial activity and impacts water-extractable organic carbon in an agricultural soilBiogeochemistry, 122, 253 -267 . DOI:
10.1007/s10533-014-0039-2 URL [本文引用: 1] [9] Flessa H Ludwig B Heil B Merbach W 2015 ). The origin of soil organic C, dissolved organic C and respiration in a long-term maize experiment in Halle, Germany, determined by 13 C natural abundance Journal of Plant Nutrition and Soil Science, 163, 157 -163 . [本文引用: 1] [10] Gélin U Wilson ME Cripps J Coulson G Festabianchet M 2016 ). Individual heterogeneity and offspring sex affect the growth-reproduction trade-off in a mammal with indeterminate growthOecologia, 180, 1127 -1135 . DOI:
10.1007/s00442-015-3531-z URL [本文引用: 1] [11] Gray AN Spies TA 1997 ). Microsite controls on tree seedling establishment in conifer forest canopy gapsEcology, 78, 2458 -2473 . DOI:
10.1890/0012-9658(1997)078[2458:MCOTSE]2.0.CO;2 URL [本文引用: 1] [12] He WM Zhong ZC 1997 ). The concept and research contents of plant propagation strategyJournal of Biology, 14, 1 -3 . [本文引用: 1] [ 何维明 , 钟章成 ( 1997 ). 植物繁殖对策的概念及其研究内容生物学杂志 , 14, 1 -3 .] [本文引用: 1] [13] He YH Zhao HL Liu XP Zhao XY Li YL Zhao W 2009 ). Reproductive allocation and its relationship with the size of corispermum elongatum in different sandy habitatsArid Zone Research, 26, 59 -64 . [本文引用: 1] [ 何玉惠 , 赵哈林 , 刘新平 , 赵学勇 , 李玉霖 , 赵玮 ( 2009 ). 不同类型沙地长穗虫实的繁殖分配及其与个体大小的关系干旱区研究 , 26, 59 -64 .] [本文引用: 1] [14] Huang ZH Lu QF Chen YZ 2017 ). Comparative study on reproductive success of Corydalis sheareri(Papaveraceae) between alkaline limestone soil and red soil habitats in a karst area. Biodiversity Science, 972 -980 . [本文引用: 1] [ 黄至欢 , 陆奇丰 , 陈颖卓 ( 2017 ). 地锦苗在石灰岩土壤和红壤生境中的繁殖成功的比较生物多样性 , 25, 972 -980 .] [本文引用: 1] [15] Huo JS Liu WG Liu JG Li HX Xu Y Maria N 2017 ). Driving forces of desert plant characteristics in a desert oasis transitional zone in Fukang, Xinjiang, ChinaActa Ecologica Sinica, 37, 8304 -8313 . [本文引用: 3] [ 霍举颂 , 刘卫国 , 刘建国 , 李宏侠 , 徐悦 , 玛丽娅·奴尔兰 ( 2017 ). 影响阜康荒漠-绿洲过渡带荒漠植物数量特征的土壤驱动力分析生态学报 , 37, 8304 -8313 .] [本文引用: 3] [16] Jiao DZ Jiang QX Cao R Yan QY Yang YF 2018 ). Quantitative characteristics and dynamics of the rhizome of Phragmites australis populations in heterogeneous habitats in the Zhalong Wetland. Acta Ecologica Sinica, 3432 -3440 . [本文引用: 1] [ 焦德志 , 姜秋旭 , 曹瑞 , 闫秋月 , 杨允菲 ( 2018 ). 扎龙湿地不同生境芦苇种群根茎数量特征及动态生态学报 , 38, 3432 -3440 .] [本文引用: 1] [17] Jiao JY Zhang ZG Jia YF Wang N Bai WJ 2008 ). Species camposition and classification of natural vegetation in the abandoned lands of the hilly-gullied region of North Shaanxi ProvinceActa Ecologica Snica, 28, 2981 -2997 . [本文引用: 1] [ 焦菊英 , 张振国 , 贾燕锋 , 王宁 , 白文娟 ( 2008 ). 陕北丘陵沟壑区撂荒地自然恢复植被的组成结构与数量分类生态学报 , 28, 2981 -2997 .] [本文引用: 1] [18] Johnson BG Verburg PS Arnone JA 2016 ). Plant species effects on soil nutrients and chemistry in arid ecological zonesOecologia, 182, 1 -19 . DOI:
10.1007/s00442-016-3559-8 URL [本文引用: 1] [19] Kendall BE Fox GA Fujiwara M Nogeire TM 2011 ). Demographic heterogeneity, cohort selection, and population growthEcology, 92, 1985 -1993 . DOI:
10.1890/11-0079.1 URL [本文引用: 1] [20] Koltes S 2006 ). Vegetation and fertilization effects on soil water soluble organic carbonCurrent Applied Physics, 6, 182 . DOI:
10.1016/j.cap.2005.07.036 URL [本文引用: 1] [21] L'Hérault V Franke A Lecomte N Alogut A Bêty J 2013 ). Landscape heterogeneity drives intra-population niche variation and reproduction in an arctic top predatorEcology & Evolution, 3, 2867 -2879 . [本文引用: 1] [22] Larson JE Funk JL 2016 ). Regeneration: An overlooked aspect of trait—Based plant community assembly modelsJournal of Ecology, 104, 1284 -1298 . DOI:
10.1111/1365-2745.12613 URL [本文引用: 1] [23] Li B Liu ZJ Zhao ZG Hu C Ren HM Wu GQ 2013 ). Influence of altitude on reproductive traits and reprodyctive allocation of different colours in Anemone obtusiloba populations. Acta Prataculturae Sinica, 10 -19 . [本文引用: 1] [ 李冰 , 刘左军 , 赵志刚 , 胡春 , 任红梅 , 伍国强 ( 2013 ). 海拔对钝裂银莲花不同花色居群间繁殖特征及繁殖分配的影响草业学报 , 22, 10 -19 .] [本文引用: 1] [24] Li KC Guo SJ 2012 ). Main Feed and Poisonous Plants in Ningxia . Sunshine Press, Yinchuan. [ 李克昌 , 郭思加 ( 2012 ). 宁夏主要饲用及有毒有害植物 . 阳光出版社, 银川.] [25] Liu F Wang SJ Luo HB Liu YS Liu HY 2008 ). Micro-habitats in karst forest ecosystem and variability of soilsActa Pedologica Sinica, 45, 1055 -1062 . [本文引用: 1] [ 刘方 , 王世杰 , 罗海波 , 刘元生 , 刘鸿雁 ( 2008 ). 喀斯特森林生态系统的小生境及其土壤异质性土壤学报 , 45, 1055 -1062 .] [本文引用: 1] [26] Liu FY Zhang ZX Wang XQ Li K Sun YY Zhang CH 2011 ). Effects of habitat heterogeneity on early growth of Quercus franchetii natural regeneration seedlings in the Jinsha River dry-hot valley. Chinese Journal of Applied and Environmental Biology, 338 -344 . [本文引用: 1] [ 刘方炎 , 张志翔 , 王小庆 , 李昆 , 孙永玉 , 张春华 ( 2011 ). 生境异质性对金沙江干热河谷锥连栎天然更新幼苗早期生长的影响应用与环境生物学报 , 17, 338 -344 .] [本文引用: 1] [27] Liu RJ Li ZC Wang B Wu YC Cheng CF Ma SJ Ge-Ri LT 2013 ). Seasonal dynamics of soil water-soluble organic carbon in secondary forests and Chinese fir plantattions in hilly region of northwest Zhejiang Province, East ChinaChinese Journal of Ecology, 32, 1385 -1390 . [本文引用: 1] [ 刘荣杰 , 李正才 , 王斌 , 吴亚丛 , 程彩芳 , 马少杰 , 格日乐图 ( 2013 ). 浙西北丘陵地区次生林与杉木林土壤水溶性有机碳季节动态生态学杂志 , 32, 1385 -1390 .] [本文引用: 1] [28] Liu XD Zhang KB Wang LL Yang XH 2015 ). How enclosure affects communitycharacteristics of the sandy grassland in semi-arid areas of northwestern ChinaJournal of Beijing Forestry University, 37(2 ), 48 -54 . [本文引用: 1] [ 刘小丹 , 张克斌 , 王黎黎 , 杨晓晖 ( 2015 ). 封育对半干旱区沙化草地群落特征的影响北京林业大学学报 , 37(2 ), 48 -54 .] [本文引用: 1] [29] Liu ZJ Du GZ Chen JK 2002 ). Size-dependent reproductive allocation of Ligularia virgaurea in different habitats. Acta Phytoecologica Sinica, 44 -50 . [本文引用: 1] [ 刘左军 , 杜国祯 , 陈家宽 ( 2002 ). 不同生境下黄帚橐吾(Ligularia virgaurea )个体大小依赖的繁殖分配植物生态学报 , 26, 44 -50 .] [本文引用: 1] [30] Liu ZM Jiang DM Gao HY Chang XL 2003 a). Relationships between plant reproductive strategy and disturbanceChinese Journal Apploed Ecology, 14, 418 -422 . [本文引用: 1] [ 刘志民 , 蒋德明 , 高红瑛 , 常学礼 ( 2003 a). 植物生活史繁殖对策与干扰关系的研究应用生态学报 , 14, 418 -422 .] [本文引用: 1] [31] Liu ZM Zhao XY Fan SX 2003 b). Grime’s CSR model and his philosphy of ecological researchAdvance in Earth Sciences, 18, 603 -608 . [本文引用: 1] [ 刘志民 , 赵晓英 , 范世香 ( 2003 b). Grime的植物对策思想和生态学研究理念地球科学进展 , 18, 603 -608 .] [本文引用: 1] [32] Ma DZ 1986 ). Flora of Ningxia . Ningxia People’s Press, Yinchuan. [本文引用: 1] [ 马德滋 ( 1986 ). 宁夏植物志 . 宁夏人民出版社, 银川.] [本文引用: 1] [33] Mayer C Adler L Armbruster WS Dafni A Eardley C Huang S-G Kevan PG Ollerton J Packer L Ssymank A Stout JC Potts SG 2011 ). Pollination ecology in the 21st Century: Key questions for future researchJournal of Pollination Ecology, 3, 8 -23 . DOI:
10.26786/1920-7603 URL [本文引用: 1] [34] Olmo M Lozano AM Barrón V Villar R 2016 ). Spatial heterogeneity of soil biochar content affects soil quality and wheat growth and yieldScience of the Total Environment, 562, 690 -700 . DOI:
10.1016/j.scitotenv.2016.04.089 URL [本文引用: 1] [35] Palmer MW Dixon PM 2010 ). Small-scale environmental heterogeneity and the analysis of species distributions along gradientsJournal of Vegetation Science, 1, 57 -65 . [本文引用: 1] [36] Quan DJ Wei Y Zhou XQ Yan C 2012 ). Growth dynamics, biommass allocation and ecological adaptation in Ceratocarpus arenarius L. Acta Ecologica Sinca, 3352 -3358 . [本文引用: 1] [ 全杜娟 , 魏岩 , 周晓青 , 严成 ( 2012 ). 角果藜的生长动态及其生殖配置生态学报 , 32, 3352 -3358 .] [本文引用: 1] [37] Ramakrishna RN Charles DK Hirofumi H William MJ Stephen CP Compton JT Ranga BM Steven WR 2003 ). Climate-driven increases in global terrestrial net primary production from 1982 to 1999Science, 300, 1560 -1563 . DOI:
10.1126/science.1082750 URL [本文引用: 1] [38] Ren MX Jiang XH Zhang DY 2012 ). Some important questions in plant reproductive ecologyBiodiversity Science, 20, 241 -249 . [本文引用: 2] [ 任明迅 , 姜新华 , 张大勇 ( 2012 ). 植物繁殖生态学的若干重要问题生物多样性 , 20, 241 -249 .] [本文引用: 2] [39] Scaglia B Adani F 2009 ). Biodegradability of soil water soluble organic carbon extracted from seven different soilsJournal of Environmental Sciences, 21, 641 -646 . DOI:
10.1016/S1001-0742(08)62319-0 URL [本文引用: 1] [40] Shen YF Tao WH Li SQ 2011 ). Effects of fertilization dissolved organic carbon and nitrogen in the ryegrass farmland soil in wind-water erosion crisscross region on the Loess PlateauJournal of Natural Resources, 26, 1387 -1393 . [本文引用: 1] [ 沈玉芳 , 陶武辉 , 李世清 ( 2011 ). 黄土高原水蚀风蚀交错区施肥对黑麦草农田土壤水溶性有机碳、氮的影响自然资源学报 , 26, 1387 -1393 .] [本文引用: 1] [41] Singh AK Rai A Pandey V Singh N 2017 ). Contribution of glomalin to dissolve organic carbon under different land uses and seasonality in dry tropicsJournal of Environmental Management, 192, 142 -149 . [本文引用: 1] [42] Song G An N Ning Z He Y Shi P Zhang J He N 2018 ). Climate warming impacts on soil organic carbon fractions and aggregate stability in a Tibetan alpine meadowSoil Biology & Biochemistry, 116, 224 -236 . [本文引用: 1] [43] Song NP Wang X Chen L Xue Y Chen J Sui JM Wang L Yang XG 2018 ). Co-existence mechanisms of plant species within “soil islands” habitat of desert steppeBiodiversity Science, 26, 667 -677 . [本文引用: 3] [ 宋乃平 , 王兴 , 陈林 , 薛毅 , 陈娟 , 随金明 , 王磊 , 杨新国 ( 2018 ). 荒漠草原“土岛”生境群落物种共存机制生物多样性 , 26, 667 -677 .] [本文引用: 3] [44] Tang Z Deng L An H Yan W Shangguan Z 2017 ). The effect of nitrogen addition on community structure and productivity in grasslands: A meta-analysisEcological Engineering, 99, 31 -38 . DOI:
10.1016/j.ecoleng.2016.11.039 URL [本文引用: 1] [45] Vavrek MC Mcgraw JB Yang HS 1997 ). Within-population variation in demography of taraxacum officinale: Season- and size-dependent survival, growth and reproductionJournal of Ecology, 85, 277 -287 . DOI:
10.2307/2960501 URL [本文引用: 1] [46] Wang HJ Chang SL Zhang YT Xie J He P Song CC Sun XJ 2016 ). Density-denpendent effects in Picea schrenkiana forests in Tianshan Mountains. Biodiversity Science, 252 -261 . [本文引用: 2] [ 王慧杰 , 常顺利 , 张毓涛 , 谢锦 , 何平 , 宋成程 , 孙雪娇 ( 2016 ). 天山雪岭云杉森林群落的密度制约效应生物多样性 , 24, 252 -261 .] [本文引用: 2] [47] Wang JC Shi X Zhang DY Yi LK 2009 ). The morphological plasticity of Eremosparton songoricum a long heterogeneous micro-habitats of continuous moisture gradient changes in sand dunes. Acta Ecologica Snica, 3641 -3648 . [本文引用: 1] [ 王建成 , 施翔 , 张道远 , 尹林克 ( 2009 ). 沙丘坡面异质性小生境中准噶尔无叶豆对水分条件变化的响应生态学报 , 29, 3641 -3648 .] [本文引用: 1] [48] Wang X Yang X Wang L Chen L Song N Gu J Xue Y 2018 ). A six-year grazing exclusion changed plant species diversity of a Stipa breviflora desert steppe community, northern China. PeerJ, e4359 . DOI: 10.7717/peerj.4359 . [本文引用: 1] [49] Wang XG Li CS Yong L Hua KK Zhou MH 2016 ). The impact of nitrogen amendment and crop growth on dissolved organic carbon in soil solutionJournal of Mountain Science, 13, 95 -103 . DOI:
10.1007/s11629-015-3556-0 URL [50] Wang YY Shi S Lin F Yuan ZQ Ye J Wang XG Hao ZQ 2014 ). Reproductive traits and their correlation among woody plants in a broadleaf-Korean pine (Pinus koraiensis ) mixed forest in Northeast China. Chinese Science Bulletin, 2407 -2415 . [本文引用: 1] [ 王芸芸 , 师帅 , 蔺菲 , 原作强 , 叶吉 , 王绪高 , 郝占庆 ( 2014 ). 长白山阔叶红松林木本植物繁殖特征及其关联性科学通报 , 59, 2407 -2415 .] [本文引用: 1] [51] Xiao H Xu CL Zhang DG Chai JL Pan TT Yu XJ 2018 ). Effects of simulated trampling and rainfall in sexual reproduction characteristics of Medicago ruthenica var. inschanica on alpine meadow. Chinese Journal of Ecology, 1976 -1982 . [本文引用: 1] [ 肖红 , 徐长林 , 张德罡 , 柴锦隆 , 潘涛涛 , 鱼小军 ( 2018 ). 模拟践踏和降水对高寒草甸阴山扁蓿豆有性繁殖特征的影响生态学杂志 , 37, 1976 -1982 .] [本文引用: 1] [52] Xiao L Liu GB Li P Xue S 2017 ). Effects of nitrogen addition and elevated CO2 concentration on soil dissolved organic carbon and nitrogen in rhizosphere and non-rhizosphere of Bothriochloa ischaemum . Chinese Journal of Applied Ecology, 64 -70 . [本文引用: 1] [ 肖列 , 刘国彬 , 李鹏 , 薛萐 ( 2017 ). 氮素添加和CO2 浓度升高对白羊草根际和非根际土壤水溶性有机碳、氮的影响应用生态学报 , 28, 64 -70 .] [本文引用: 1] [53] Xu GF Shen SC Zhang FD 2014 ). Adaptability and reproductive characteristics of Mikania micrantha H.B.K under different habitats. Ecology and Encironmental Sciences, 1258 -1264 . [本文引用: 1] [ 徐高峰 , 申时才 , 张付斗 ( 2014 ). 异质环境下入侵植物薇甘菊的适应性与繁殖特性生态环境学报 , 23, 1258 -1264 .] [本文引用: 1] [54] Xu Z 1999 ). Plant development and reproduction: Advances and prospectivesActa Botanica Sinica, 41, 909 -920 . [本文引用: 1] [55] Yu M Zhou ZY Kang FF Ouyang S Mi XC Sun JX 2013 ). Gradient analysis and environmental interpretation of understory herb-layer communities in Xiaoshegou of Lingkong Mountain, Shanxi, ChinaChinese Journal of Plant Ecology, 37, 373 -383 . [本文引用: 1] [ 余敏 , 周志勇 , 康峰峰 , 欧阳帅 , 米湘成 , 孙建新 ( 2013 ). 山西灵空山小蛇沟林下草本层植物群落梯度分析及环境解释植物生态学报 , 37, 373 -383 .] [本文引用: 1] [56] Yue YM Wang KL Xiong Y 2012 ). Feasibility of monitoring karst standing conditions with vegetation spectraSpectroscopy and Spectral Anaysis, 32, 1891 -1894 . [本文引用: 1] [ 岳跃民 , 王克林 , 熊鹰 ( 2012 ). 基于植被光谱监测喀斯特异质性生境可行性研究光谱学与光谱分析 , 32, 1891 -1894 .] [本文引用: 1] [57] Zhang DY 2004 ). Evolution of Plant Life History and Reproductive Ecology . Science Press, Beijing. [本文引用: 1] [ 张大勇 ( 2004 ). 植物生活史进化与繁殖生态学 . 科学出版社, 北京.] [本文引用: 1] [58] Zhang DY Jiang XH 2001 ). Mating system evoluion, resource allocation, and genetic diversity in plansActa Phytoecologica Sinica, 25, 130 -143 . [本文引用: 2] [ 张大勇 , 姜新华 ( 2001 ). 植物交配系统的进化、资源分配对策与遗传多样性植物生态学报 , 25, 130 -143 .] [本文引用: 2] [59] Zhang ZL Tuo F Shang H Zhang YM 2009 ). Differernce in growth status and ecological adaptability strategy of large-fruit seabuckthom under different site conditionsJournal of Southwest Forestry University (Natural Sciences )5 ), 12 -15 . [本文引用: 1] [ 张治来 , 拓飞 , 尚华 , 张永满 ( 2009 ). 不同立地类型大果沙棘生长及生态适应对策差异西南林业大学学报(自然科学) , 29(5 ), 12 -15 .] [本文引用: 1] [60] Zhao LC Zhao CZ Wang XP Wen J 2017 ). Interrelations between environmental factors and distribution of Tamarix gansuensis in Qinwangchuan wetland. Acta Ecologica Sinica, 3422 -3431 . [本文引用: 1] [ 赵连春 , 赵成章 , 王小鹏 , 文军 ( 2017 ). 秦王川湿地柽柳分布与环境因子的关系生态学报 , 38, 3422 -3431 .] [本文引用: 1] [61] Zhao RF Zhou HR Qian YB Zhang JJ 2006 ). Interrelations between plant communities and environmental factors of wetlands and surrounding lands in mid- and lower reaches of Tarim RiverChinese Journal of Applied Ecology, 17, 955 -960 . [本文引用: 1] [ 赵锐锋 , 周华荣 , 钱亦兵 , 张建军 ( 2006 ). 塔里木河中下游湿地及周边植物群落与环境因子的关系初探应用生态学报 , 17, 955 -960 .] [本文引用: 1] 异质生境对二型花柱植物雪地报春花性状分化的影响
1
2018
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
异质生境对二型花柱植物雪地报春花性状分化的影响
1
2018
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
1
2000
... 在所调查的植株样方内, 采用便携式土壤水分测量仪(ML3, Delta-T, Burwell, UK)和数显式土壤紧实度仪(SC-900, Spectrum Technologies, Aurora, USA)分别对0-10和10-20 cm土层土壤水分含量(
M s )和0-5、5-10和10-20 cm土壤硬度(
SH )进行测量.pH值采用电位法测定, 水土质量比为5:1; 全盐含量(
TS )通过电导率换算而成; 易氧化碳含量(
C eo )采用KMnO
4 氧化比色法测定; 用蒸馏水浸提0.45 μm滤膜过滤后采用K
2 CrO
7 法测定水溶性碳含量(
C ws ); 有机碳含量(
C o )采用K
2 CrO
7 氧化法测定; 全碳含量(
C t )采用元素分析仪(vario MACRO, Elementar, Langenselbold, Germany)测定; 全磷含量(
TP )采用钼锑抗比色法测定; 速效磷含量(
AP )采用0.5 mol·L
-1 NaHCO
3 浸提-钼锑抗比色法测定; 速效钾含量(
AK )采用乙酸铵浸提-火焰光度法测定; 全氮含量(
TN )采用凯氏定氮法测定; 碱解氮含量(
AN )采用碱解扩散法测定(
鲍士旦, 2000 ). ...
1
2000
... 在所调查的植株样方内, 采用便携式土壤水分测量仪(ML3, Delta-T, Burwell, UK)和数显式土壤紧实度仪(SC-900, Spectrum Technologies, Aurora, USA)分别对0-10和10-20 cm土层土壤水分含量(
M s )和0-5、5-10和10-20 cm土壤硬度(
SH )进行测量.pH值采用电位法测定, 水土质量比为5:1; 全盐含量(
TS )通过电导率换算而成; 易氧化碳含量(
C eo )采用KMnO
4 氧化比色法测定; 用蒸馏水浸提0.45 μm滤膜过滤后采用K
2 CrO
7 法测定水溶性碳含量(
C ws ); 有机碳含量(
C o )采用K
2 CrO
7 氧化法测定; 全碳含量(
C t )采用元素分析仪(vario MACRO, Elementar, Langenselbold, Germany)测定; 全磷含量(
TP )采用钼锑抗比色法测定; 速效磷含量(
AP )采用0.5 mol·L
-1 NaHCO
3 浸提-钼锑抗比色法测定; 速效钾含量(
AK )采用乙酸铵浸提-火焰光度法测定; 全氮含量(
TN )采用凯氏定氮法测定; 碱解氮含量(
AN )采用碱解扩散法测定(
鲍士旦, 2000 ). ...
基于文献计量分析的蒿属植物研究进展
1
2017
... 猪毛蒿(
Artemisia scoparia )是菊科蒿属草本, 是一种C
3 植物, 却呈现类似CAM植物的叶片特性(
杜华栋等, 2016 ), 广布于欧亚大陆温带至亚热带地区(
马德滋, 1986 ;
李克昌和郭思加, 2012 ), 在局部地区成为植物群落的优势种, 单个样地覆盖度可超过50% (
焦菊英等, 2008 ).目前, 国内外****对蒿属植物的研究多集中在植物生理、化学和药理学/配药学领域(
陈林等, 2017 ), 特别是猪毛蒿的化学成分及其医药用途方面是研究热点, 但较少考虑土壤因子对猪毛蒿繁殖特征的影响, 特别是荒漠草原异质生境条件下, 以不同土壤因子为驱动力, 对猪毛蒿繁殖特征及其影响因素的研究很少有报道. ...
基于文献计量分析的蒿属植物研究进展
1
2017
... 猪毛蒿(
Artemisia scoparia )是菊科蒿属草本, 是一种C
3 植物, 却呈现类似CAM植物的叶片特性(
杜华栋等, 2016 ), 广布于欧亚大陆温带至亚热带地区(
马德滋, 1986 ;
李克昌和郭思加, 2012 ), 在局部地区成为植物群落的优势种, 单个样地覆盖度可超过50% (
焦菊英等, 2008 ).目前, 国内外****对蒿属植物的研究多集中在植物生理、化学和药理学/配药学领域(
陈林等, 2017 ), 特别是猪毛蒿的化学成分及其医药用途方面是研究热点, 但较少考虑土壤因子对猪毛蒿繁殖特征的影响, 特别是荒漠草原异质生境条件下, 以不同土壤因子为驱动力, 对猪毛蒿繁殖特征及其影响因素的研究很少有报道. ...
青藏高原东缘大耳叶风毛菊繁殖特征对海拔的响应
1
2014
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
青藏高原东缘大耳叶风毛菊繁殖特征对海拔的响应
1
2014
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
河南木札岭温带落叶阔叶林群落特征及主要乔木空间分布格局
1
2016
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
河南木札岭温带落叶阔叶林群落特征及主要乔木空间分布格局
1
2016
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
异质性生境中的植物克隆生长, 风险分摊
1
1996
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
异质性生境中的植物克隆生长, 风险分摊
1
1996
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
黄土高原先锋种猪毛蒿叶片形态解剖与生理特征对立地的适应性
2
2016
... 猪毛蒿(
Artemisia scoparia )是菊科蒿属草本, 是一种C
3 植物, 却呈现类似CAM植物的叶片特性(
杜华栋等, 2016 ), 广布于欧亚大陆温带至亚热带地区(
马德滋, 1986 ;
李克昌和郭思加, 2012 ), 在局部地区成为植物群落的优势种, 单个样地覆盖度可超过50% (
焦菊英等, 2008 ).目前, 国内外****对蒿属植物的研究多集中在植物生理、化学和药理学/配药学领域(
陈林等, 2017 ), 特别是猪毛蒿的化学成分及其医药用途方面是研究热点, 但较少考虑土壤因子对猪毛蒿繁殖特征的影响, 特别是荒漠草原异质生境条件下, 以不同土壤因子为驱动力, 对猪毛蒿繁殖特征及其影响因素的研究很少有报道. ...
... 有研究发现,
C ws 含量与降水量显著负相关, 是因为降水量的增加导致
C ws 淋溶、淋失作用加强, 从而
C ws 含量较低(
刘荣杰等, 2013 ).但本研究区降水较少, 降水的淋溶作用不足以使
C ws 发生淋失, 而
M s 相对较高的生境能促进枯枝落叶的分解, 进而增加了土壤中
C ws 的含量, 因此
C ws 与
M s 呈现出正相关关系.本研究区地下水位较低, 植物往往无法利用, 土壤中水分补给的主要来源是降水, 且具有很强的时空异质性和随机性, 导致该地区植物群落分布多呈斑块状(
宋乃平等, 2018 ).有研究结果表明荒漠植物数量特征的关键驱动力是
M s , 特别是表层
M s (0-20 cm)对荒漠植物群落数量特征起到非常重要的生态作用(
霍举颂等, 2017 ).本研究中, 单株头状花序质量和数量极显著地受到
M s 的影响,
C ws 与单株头状花序质量显著正相关, 这与有关****(
杜华栋等, 2016 )研究发现猪毛蒿叶片形态、解剖和生理指标的可塑性对立地
M s 和
C o 含量较为敏感的结果有相同之处.结合RDA分析图可知,
C ws 和
M s 是影响猪毛蒿繁殖特征的重要因子. ...
黄土高原先锋种猪毛蒿叶片形态解剖与生理特征对立地的适应性
2
2016
... 猪毛蒿(
Artemisia scoparia )是菊科蒿属草本, 是一种C
3 植物, 却呈现类似CAM植物的叶片特性(
杜华栋等, 2016 ), 广布于欧亚大陆温带至亚热带地区(
马德滋, 1986 ;
李克昌和郭思加, 2012 ), 在局部地区成为植物群落的优势种, 单个样地覆盖度可超过50% (
焦菊英等, 2008 ).目前, 国内外****对蒿属植物的研究多集中在植物生理、化学和药理学/配药学领域(
陈林等, 2017 ), 特别是猪毛蒿的化学成分及其医药用途方面是研究热点, 但较少考虑土壤因子对猪毛蒿繁殖特征的影响, 特别是荒漠草原异质生境条件下, 以不同土壤因子为驱动力, 对猪毛蒿繁殖特征及其影响因素的研究很少有报道. ...
... 有研究发现,
C ws 含量与降水量显著负相关, 是因为降水量的增加导致
C ws 淋溶、淋失作用加强, 从而
C ws 含量较低(
刘荣杰等, 2013 ).但本研究区降水较少, 降水的淋溶作用不足以使
C ws 发生淋失, 而
M s 相对较高的生境能促进枯枝落叶的分解, 进而增加了土壤中
C ws 的含量, 因此
C ws 与
M s 呈现出正相关关系.本研究区地下水位较低, 植物往往无法利用, 土壤中水分补给的主要来源是降水, 且具有很强的时空异质性和随机性, 导致该地区植物群落分布多呈斑块状(
宋乃平等, 2018 ).有研究结果表明荒漠植物数量特征的关键驱动力是
M s , 特别是表层
M s (0-20 cm)对荒漠植物群落数量特征起到非常重要的生态作用(
霍举颂等, 2017 ).本研究中, 单株头状花序质量和数量极显著地受到
M s 的影响,
C ws 与单株头状花序质量显著正相关, 这与有关****(
杜华栋等, 2016 )研究发现猪毛蒿叶片形态、解剖和生理指标的可塑性对立地
M s 和
C o 含量较为敏感的结果有相同之处.结合RDA分析图可知,
C ws 和
M s 是影响猪毛蒿繁殖特征的重要因子. ...
Elevated atmospheric carbon dioxide concentration stimulates soil microbial activity and impacts water-extractable organic carbon in an agricultural soil
1
2015
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
The origin of soil organic C, dissolved organic C and respiration in a long-term maize experiment in Halle, Germany, determined by
13 C natural abundance
1
2015
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
Individual heterogeneity and offspring sex affect the growth-reproduction trade-off in a mammal with indeterminate growth
1
2016
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
Microsite controls on tree seedling establishment in conifer forest canopy gaps
1
1997
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
植物繁殖对策的概念及其研究内容
1
1997
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
植物繁殖对策的概念及其研究内容
1
1997
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
不同类型沙地长穗虫实的繁殖分配及其与个体大小的关系
1
2009
... 本研究中猪毛蒿个体大小以地上生物量干质量表示(
刘左军等, 2002 ;
何玉惠等, 2009 ); 繁殖器官大小以头状花序干质量表示.统计单株猪毛蒿的生物量、繁殖体数量、繁殖体质量, 并计算单个繁殖体质量, 单个繁殖体质量为单株繁殖体质量与单株繁殖体数量的比值.繁殖分配为繁殖器官干质量占总生物量干质量的百分比. ...
不同类型沙地长穗虫实的繁殖分配及其与个体大小的关系
1
2009
... 本研究中猪毛蒿个体大小以地上生物量干质量表示(
刘左军等, 2002 ;
何玉惠等, 2009 ); 繁殖器官大小以头状花序干质量表示.统计单株猪毛蒿的生物量、繁殖体数量、繁殖体质量, 并计算单个繁殖体质量, 单个繁殖体质量为单株繁殖体质量与单株繁殖体数量的比值.繁殖分配为繁殖器官干质量占总生物量干质量的百分比. ...
地锦苗在石灰岩土壤和红壤生境中的繁殖成功的比较
1
2017
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
地锦苗在石灰岩土壤和红壤生境中的繁殖成功的比较
1
2017
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
影响阜康荒漠-绿洲过渡带荒漠植物数量特征的土壤驱动力分析
3
2017
... 但从不同角度研究同一个问题, 经常得出矛盾的结论(
Mayer et al ., 2011 ).所以从不同的因素中综合分析并找出影响植物繁殖特征的主要选择压力, 是摆在植物繁殖生态学家面前的挑战(
任明迅等, 2012 ).在众多的环境因子中, 土壤因子往往制约着荒漠植物生命过程, 起着关键作用(
霍举颂等, 2017 ).因此本研究基于对宁夏荒漠草原地区猪毛蒿种群繁殖特征的调查, 以土壤类型和驱动因子为切入点, 采用约束性排序中的冗余分析来探讨: (1)不同土壤类型对猪毛蒿的繁殖特征是否有影响; (2)哪些土壤因子对猪毛蒿繁殖特征起主要影响作用.通过此研究揭示异质生境下猪毛蒿繁殖特征与土壤驱动因子的关系及其机制, 以期为荒漠草原恢复和管理提供理论参考和科学依据. ...
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
... 有研究发现,
C ws 含量与降水量显著负相关, 是因为降水量的增加导致
C ws 淋溶、淋失作用加强, 从而
C ws 含量较低(
刘荣杰等, 2013 ).但本研究区降水较少, 降水的淋溶作用不足以使
C ws 发生淋失, 而
M s 相对较高的生境能促进枯枝落叶的分解, 进而增加了土壤中
C ws 的含量, 因此
C ws 与
M s 呈现出正相关关系.本研究区地下水位较低, 植物往往无法利用, 土壤中水分补给的主要来源是降水, 且具有很强的时空异质性和随机性, 导致该地区植物群落分布多呈斑块状(
宋乃平等, 2018 ).有研究结果表明荒漠植物数量特征的关键驱动力是
M s , 特别是表层
M s (0-20 cm)对荒漠植物群落数量特征起到非常重要的生态作用(
霍举颂等, 2017 ).本研究中, 单株头状花序质量和数量极显著地受到
M s 的影响,
C ws 与单株头状花序质量显著正相关, 这与有关****(
杜华栋等, 2016 )研究发现猪毛蒿叶片形态、解剖和生理指标的可塑性对立地
M s 和
C o 含量较为敏感的结果有相同之处.结合RDA分析图可知,
C ws 和
M s 是影响猪毛蒿繁殖特征的重要因子. ...
影响阜康荒漠-绿洲过渡带荒漠植物数量特征的土壤驱动力分析
3
2017
... 但从不同角度研究同一个问题, 经常得出矛盾的结论(
Mayer et al ., 2011 ).所以从不同的因素中综合分析并找出影响植物繁殖特征的主要选择压力, 是摆在植物繁殖生态学家面前的挑战(
任明迅等, 2012 ).在众多的环境因子中, 土壤因子往往制约着荒漠植物生命过程, 起着关键作用(
霍举颂等, 2017 ).因此本研究基于对宁夏荒漠草原地区猪毛蒿种群繁殖特征的调查, 以土壤类型和驱动因子为切入点, 采用约束性排序中的冗余分析来探讨: (1)不同土壤类型对猪毛蒿的繁殖特征是否有影响; (2)哪些土壤因子对猪毛蒿繁殖特征起主要影响作用.通过此研究揭示异质生境下猪毛蒿繁殖特征与土壤驱动因子的关系及其机制, 以期为荒漠草原恢复和管理提供理论参考和科学依据. ...
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
... 有研究发现,
C ws 含量与降水量显著负相关, 是因为降水量的增加导致
C ws 淋溶、淋失作用加强, 从而
C ws 含量较低(
刘荣杰等, 2013 ).但本研究区降水较少, 降水的淋溶作用不足以使
C ws 发生淋失, 而
M s 相对较高的生境能促进枯枝落叶的分解, 进而增加了土壤中
C ws 的含量, 因此
C ws 与
M s 呈现出正相关关系.本研究区地下水位较低, 植物往往无法利用, 土壤中水分补给的主要来源是降水, 且具有很强的时空异质性和随机性, 导致该地区植物群落分布多呈斑块状(
宋乃平等, 2018 ).有研究结果表明荒漠植物数量特征的关键驱动力是
M s , 特别是表层
M s (0-20 cm)对荒漠植物群落数量特征起到非常重要的生态作用(
霍举颂等, 2017 ).本研究中, 单株头状花序质量和数量极显著地受到
M s 的影响,
C ws 与单株头状花序质量显著正相关, 这与有关****(
杜华栋等, 2016 )研究发现猪毛蒿叶片形态、解剖和生理指标的可塑性对立地
M s 和
C o 含量较为敏感的结果有相同之处.结合RDA分析图可知,
C ws 和
M s 是影响猪毛蒿繁殖特征的重要因子. ...
扎龙湿地不同生境芦苇种群根茎数量特征及动态
1
2018
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
扎龙湿地不同生境芦苇种群根茎数量特征及动态
1
2018
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
陕北丘陵沟壑区撂荒地自然恢复植被的组成结构与数量分类
1
2008
... 猪毛蒿(
Artemisia scoparia )是菊科蒿属草本, 是一种C
3 植物, 却呈现类似CAM植物的叶片特性(
杜华栋等, 2016 ), 广布于欧亚大陆温带至亚热带地区(
马德滋, 1986 ;
李克昌和郭思加, 2012 ), 在局部地区成为植物群落的优势种, 单个样地覆盖度可超过50% (
焦菊英等, 2008 ).目前, 国内外****对蒿属植物的研究多集中在植物生理、化学和药理学/配药学领域(
陈林等, 2017 ), 特别是猪毛蒿的化学成分及其医药用途方面是研究热点, 但较少考虑土壤因子对猪毛蒿繁殖特征的影响, 特别是荒漠草原异质生境条件下, 以不同土壤因子为驱动力, 对猪毛蒿繁殖特征及其影响因素的研究很少有报道. ...
陕北丘陵沟壑区撂荒地自然恢复植被的组成结构与数量分类
1
2008
... 猪毛蒿(
Artemisia scoparia )是菊科蒿属草本, 是一种C
3 植物, 却呈现类似CAM植物的叶片特性(
杜华栋等, 2016 ), 广布于欧亚大陆温带至亚热带地区(
马德滋, 1986 ;
李克昌和郭思加, 2012 ), 在局部地区成为植物群落的优势种, 单个样地覆盖度可超过50% (
焦菊英等, 2008 ).目前, 国内外****对蒿属植物的研究多集中在植物生理、化学和药理学/配药学领域(
陈林等, 2017 ), 特别是猪毛蒿的化学成分及其医药用途方面是研究热点, 但较少考虑土壤因子对猪毛蒿繁殖特征的影响, 特别是荒漠草原异质生境条件下, 以不同土壤因子为驱动力, 对猪毛蒿繁殖特征及其影响因素的研究很少有报道. ...
Plant species effects on soil nutrients and chemistry in arid ecological zones
1
2016
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
Demographic heterogeneity, cohort selection, and population growth
1
2011
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
Vegetation and fertilization effects on soil water soluble organic carbon
1
2006
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
Landscape heterogeneity drives intra-population niche variation and reproduction in an arctic top predator
1
2013
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
Regeneration: An overlooked aspect of trait—Based plant community assembly models
1
2016
... 植物种群的更新和个体生活史的完成均依赖于繁殖过程(
全杜娟等, 2012 ;
Larson & Funk, 2016 ), 植物的繁殖性状是植物资源投入-收益长期权衡的结果(
张大勇, 2004 ), 通过植株、花序等的资源分配可以很好地解释许多重要繁殖特征, 这在很大程度上协调了不同侧重点研究所得到的矛盾结论(
任明迅等, 2012 ). ...
海拔对钝裂银莲花不同花色居群间繁殖特征及繁殖分配的影响
1
2013
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
海拔对钝裂银莲花不同花色居群间繁殖特征及繁殖分配的影响
1
2013
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
0
2012
0
2012
喀斯特森林生态系统的小生境及其土壤异质性
1
2008
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
喀斯特森林生态系统的小生境及其土壤异质性
1
2008
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
生境异质性对金沙江干热河谷锥连栎天然更新幼苗早期生长的影响
1
2011
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
生境异质性对金沙江干热河谷锥连栎天然更新幼苗早期生长的影响
1
2011
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
浙西北丘陵地区次生林与杉木林土壤水溶性有机碳季节动态
1
2013
... 有研究发现,
C ws 含量与降水量显著负相关, 是因为降水量的增加导致
C ws 淋溶、淋失作用加强, 从而
C ws 含量较低(
刘荣杰等, 2013 ).但本研究区降水较少, 降水的淋溶作用不足以使
C ws 发生淋失, 而
M s 相对较高的生境能促进枯枝落叶的分解, 进而增加了土壤中
C ws 的含量, 因此
C ws 与
M s 呈现出正相关关系.本研究区地下水位较低, 植物往往无法利用, 土壤中水分补给的主要来源是降水, 且具有很强的时空异质性和随机性, 导致该地区植物群落分布多呈斑块状(
宋乃平等, 2018 ).有研究结果表明荒漠植物数量特征的关键驱动力是
M s , 特别是表层
M s (0-20 cm)对荒漠植物群落数量特征起到非常重要的生态作用(
霍举颂等, 2017 ).本研究中, 单株头状花序质量和数量极显著地受到
M s 的影响,
C ws 与单株头状花序质量显著正相关, 这与有关****(
杜华栋等, 2016 )研究发现猪毛蒿叶片形态、解剖和生理指标的可塑性对立地
M s 和
C o 含量较为敏感的结果有相同之处.结合RDA分析图可知,
C ws 和
M s 是影响猪毛蒿繁殖特征的重要因子. ...
浙西北丘陵地区次生林与杉木林土壤水溶性有机碳季节动态
1
2013
... 有研究发现,
C ws 含量与降水量显著负相关, 是因为降水量的增加导致
C ws 淋溶、淋失作用加强, 从而
C ws 含量较低(
刘荣杰等, 2013 ).但本研究区降水较少, 降水的淋溶作用不足以使
C ws 发生淋失, 而
M s 相对较高的生境能促进枯枝落叶的分解, 进而增加了土壤中
C ws 的含量, 因此
C ws 与
M s 呈现出正相关关系.本研究区地下水位较低, 植物往往无法利用, 土壤中水分补给的主要来源是降水, 且具有很强的时空异质性和随机性, 导致该地区植物群落分布多呈斑块状(
宋乃平等, 2018 ).有研究结果表明荒漠植物数量特征的关键驱动力是
M s , 特别是表层
M s (0-20 cm)对荒漠植物群落数量特征起到非常重要的生态作用(
霍举颂等, 2017 ).本研究中, 单株头状花序质量和数量极显著地受到
M s 的影响,
C ws 与单株头状花序质量显著正相关, 这与有关****(
杜华栋等, 2016 )研究发现猪毛蒿叶片形态、解剖和生理指标的可塑性对立地
M s 和
C o 含量较为敏感的结果有相同之处.结合RDA分析图可知,
C ws 和
M s 是影响猪毛蒿繁殖特征的重要因子. ...
封育对半干旱区沙化草地群落特征的影响
1
2015
... 本研究区位于宁夏中部干旱带的荒漠草原, 为鄂尔多斯高原缓坡起伏地带.干旱少雨, 蒸发量大, 冬春两季风大沙多, 属典型的中温带大陆性气候(
宋乃平等, 2018 ).近50年年平均气温为8.46 ℃, 年降水量为276.3 mm, 年日照时间为2 862.6 h.该区域植被主要受水分因素限制(
Ramakrishna et al ., 2003 ), 表现为物种多样性较低、群落结构简单、沙生特征明显, 地下水资源匮乏且植物常常无法利用(
刘小丹等, 2015 ;
Tang et al ., 2017 ;
Wang et al ., 2018 ). ...
封育对半干旱区沙化草地群落特征的影响
1
2015
... 本研究区位于宁夏中部干旱带的荒漠草原, 为鄂尔多斯高原缓坡起伏地带.干旱少雨, 蒸发量大, 冬春两季风大沙多, 属典型的中温带大陆性气候(
宋乃平等, 2018 ).近50年年平均气温为8.46 ℃, 年降水量为276.3 mm, 年日照时间为2 862.6 h.该区域植被主要受水分因素限制(
Ramakrishna et al ., 2003 ), 表现为物种多样性较低、群落结构简单、沙生特征明显, 地下水资源匮乏且植物常常无法利用(
刘小丹等, 2015 ;
Tang et al ., 2017 ;
Wang et al ., 2018 ). ...
不同生境下黄帚橐吾(
Ligularia virgaurea )个体大小依赖的繁殖分配
1
2002
... 本研究中猪毛蒿个体大小以地上生物量干质量表示(
刘左军等, 2002 ;
何玉惠等, 2009 ); 繁殖器官大小以头状花序干质量表示.统计单株猪毛蒿的生物量、繁殖体数量、繁殖体质量, 并计算单个繁殖体质量, 单个繁殖体质量为单株繁殖体质量与单株繁殖体数量的比值.繁殖分配为繁殖器官干质量占总生物量干质量的百分比. ...
不同生境下黄帚橐吾(
Ligularia virgaurea )个体大小依赖的繁殖分配
1
2002
... 本研究中猪毛蒿个体大小以地上生物量干质量表示(
刘左军等, 2002 ;
何玉惠等, 2009 ); 繁殖器官大小以头状花序干质量表示.统计单株猪毛蒿的生物量、繁殖体数量、繁殖体质量, 并计算单个繁殖体质量, 单个繁殖体质量为单株繁殖体质量与单株繁殖体数量的比值.繁殖分配为繁殖器官干质量占总生物量干质量的百分比. ...
植物生活史繁殖对策与干扰关系的研究
1
2003
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
植物生活史繁殖对策与干扰关系的研究
1
2003
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
Grime的植物对策思想和生态学研究理念
1
2003
... 植物的生长与繁殖能力是植物适应环境的体现, 是统一的整体, 所以植株繁殖特征也必然与植株的各生长特征紧密联系(
刘志民等, 2003b ).猪毛蒿繁殖特征间的相关性分析表明, 繁殖分配与单个头状花序质量、单株头状花序数量与质量间呈极显著正相关关系(
r = 0.44,
r = 0.97,
p < 0.001), 单株头状花序数量与单个头状花序质量呈显著负相关关系(
r = -0.17,
p < 0.05).繁殖分配与单株头状花序数量和单株头状花序质量间、单个头状花序质量与单株头状花序质量间均无相关关系(
p > 0.05). ...
Grime的植物对策思想和生态学研究理念
1
2003
... 植物的生长与繁殖能力是植物适应环境的体现, 是统一的整体, 所以植株繁殖特征也必然与植株的各生长特征紧密联系(
刘志民等, 2003b ).猪毛蒿繁殖特征间的相关性分析表明, 繁殖分配与单个头状花序质量、单株头状花序数量与质量间呈极显著正相关关系(
r = 0.44,
r = 0.97,
p < 0.001), 单株头状花序数量与单个头状花序质量呈显著负相关关系(
r = -0.17,
p < 0.05).繁殖分配与单株头状花序数量和单株头状花序质量间、单个头状花序质量与单株头状花序质量间均无相关关系(
p > 0.05). ...
1
1986
... 猪毛蒿(
Artemisia scoparia )是菊科蒿属草本, 是一种C
3 植物, 却呈现类似CAM植物的叶片特性(
杜华栋等, 2016 ), 广布于欧亚大陆温带至亚热带地区(
马德滋, 1986 ;
李克昌和郭思加, 2012 ), 在局部地区成为植物群落的优势种, 单个样地覆盖度可超过50% (
焦菊英等, 2008 ).目前, 国内外****对蒿属植物的研究多集中在植物生理、化学和药理学/配药学领域(
陈林等, 2017 ), 特别是猪毛蒿的化学成分及其医药用途方面是研究热点, 但较少考虑土壤因子对猪毛蒿繁殖特征的影响, 特别是荒漠草原异质生境条件下, 以不同土壤因子为驱动力, 对猪毛蒿繁殖特征及其影响因素的研究很少有报道. ...
1
1986
... 猪毛蒿(
Artemisia scoparia )是菊科蒿属草本, 是一种C
3 植物, 却呈现类似CAM植物的叶片特性(
杜华栋等, 2016 ), 广布于欧亚大陆温带至亚热带地区(
马德滋, 1986 ;
李克昌和郭思加, 2012 ), 在局部地区成为植物群落的优势种, 单个样地覆盖度可超过50% (
焦菊英等, 2008 ).目前, 国内外****对蒿属植物的研究多集中在植物生理、化学和药理学/配药学领域(
陈林等, 2017 ), 特别是猪毛蒿的化学成分及其医药用途方面是研究热点, 但较少考虑土壤因子对猪毛蒿繁殖特征的影响, 特别是荒漠草原异质生境条件下, 以不同土壤因子为驱动力, 对猪毛蒿繁殖特征及其影响因素的研究很少有报道. ...
Pollination ecology in the 21st Century: Key questions for future research
1
2011
... 但从不同角度研究同一个问题, 经常得出矛盾的结论(
Mayer et al ., 2011 ).所以从不同的因素中综合分析并找出影响植物繁殖特征的主要选择压力, 是摆在植物繁殖生态学家面前的挑战(
任明迅等, 2012 ).在众多的环境因子中, 土壤因子往往制约着荒漠植物生命过程, 起着关键作用(
霍举颂等, 2017 ).因此本研究基于对宁夏荒漠草原地区猪毛蒿种群繁殖特征的调查, 以土壤类型和驱动因子为切入点, 采用约束性排序中的冗余分析来探讨: (1)不同土壤类型对猪毛蒿的繁殖特征是否有影响; (2)哪些土壤因子对猪毛蒿繁殖特征起主要影响作用.通过此研究揭示异质生境下猪毛蒿繁殖特征与土壤驱动因子的关系及其机制, 以期为荒漠草原恢复和管理提供理论参考和科学依据. ...
Spatial heterogeneity of soil biochar content affects soil quality and wheat growth and yield
1
2016
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
Small-scale environmental heterogeneity and the analysis of species distributions along gradients
1
2010
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
角果藜的生长动态及其生殖配置
1
2012
... 植物种群的更新和个体生活史的完成均依赖于繁殖过程(
全杜娟等, 2012 ;
Larson & Funk, 2016 ), 植物的繁殖性状是植物资源投入-收益长期权衡的结果(
张大勇, 2004 ), 通过植株、花序等的资源分配可以很好地解释许多重要繁殖特征, 这在很大程度上协调了不同侧重点研究所得到的矛盾结论(
任明迅等, 2012 ). ...
角果藜的生长动态及其生殖配置
1
2012
... 植物种群的更新和个体生活史的完成均依赖于繁殖过程(
全杜娟等, 2012 ;
Larson & Funk, 2016 ), 植物的繁殖性状是植物资源投入-收益长期权衡的结果(
张大勇, 2004 ), 通过植株、花序等的资源分配可以很好地解释许多重要繁殖特征, 这在很大程度上协调了不同侧重点研究所得到的矛盾结论(
任明迅等, 2012 ). ...
Climate-driven increases in global terrestrial net primary production from 1982 to 1999
1
2003
... 本研究区位于宁夏中部干旱带的荒漠草原, 为鄂尔多斯高原缓坡起伏地带.干旱少雨, 蒸发量大, 冬春两季风大沙多, 属典型的中温带大陆性气候(
宋乃平等, 2018 ).近50年年平均气温为8.46 ℃, 年降水量为276.3 mm, 年日照时间为2 862.6 h.该区域植被主要受水分因素限制(
Ramakrishna et al ., 2003 ), 表现为物种多样性较低、群落结构简单、沙生特征明显, 地下水资源匮乏且植物常常无法利用(
刘小丹等, 2015 ;
Tang et al ., 2017 ;
Wang et al ., 2018 ). ...
植物繁殖生态学的若干重要问题
2
2012
... 但从不同角度研究同一个问题, 经常得出矛盾的结论(
Mayer et al ., 2011 ).所以从不同的因素中综合分析并找出影响植物繁殖特征的主要选择压力, 是摆在植物繁殖生态学家面前的挑战(
任明迅等, 2012 ).在众多的环境因子中, 土壤因子往往制约着荒漠植物生命过程, 起着关键作用(
霍举颂等, 2017 ).因此本研究基于对宁夏荒漠草原地区猪毛蒿种群繁殖特征的调查, 以土壤类型和驱动因子为切入点, 采用约束性排序中的冗余分析来探讨: (1)不同土壤类型对猪毛蒿的繁殖特征是否有影响; (2)哪些土壤因子对猪毛蒿繁殖特征起主要影响作用.通过此研究揭示异质生境下猪毛蒿繁殖特征与土壤驱动因子的关系及其机制, 以期为荒漠草原恢复和管理提供理论参考和科学依据. ...
... 植物种群的更新和个体生活史的完成均依赖于繁殖过程(
全杜娟等, 2012 ;
Larson & Funk, 2016 ), 植物的繁殖性状是植物资源投入-收益长期权衡的结果(
张大勇, 2004 ), 通过植株、花序等的资源分配可以很好地解释许多重要繁殖特征, 这在很大程度上协调了不同侧重点研究所得到的矛盾结论(
任明迅等, 2012 ). ...
植物繁殖生态学的若干重要问题
2
2012
... 但从不同角度研究同一个问题, 经常得出矛盾的结论(
Mayer et al ., 2011 ).所以从不同的因素中综合分析并找出影响植物繁殖特征的主要选择压力, 是摆在植物繁殖生态学家面前的挑战(
任明迅等, 2012 ).在众多的环境因子中, 土壤因子往往制约着荒漠植物生命过程, 起着关键作用(
霍举颂等, 2017 ).因此本研究基于对宁夏荒漠草原地区猪毛蒿种群繁殖特征的调查, 以土壤类型和驱动因子为切入点, 采用约束性排序中的冗余分析来探讨: (1)不同土壤类型对猪毛蒿的繁殖特征是否有影响; (2)哪些土壤因子对猪毛蒿繁殖特征起主要影响作用.通过此研究揭示异质生境下猪毛蒿繁殖特征与土壤驱动因子的关系及其机制, 以期为荒漠草原恢复和管理提供理论参考和科学依据. ...
... 植物种群的更新和个体生活史的完成均依赖于繁殖过程(
全杜娟等, 2012 ;
Larson & Funk, 2016 ), 植物的繁殖性状是植物资源投入-收益长期权衡的结果(
张大勇, 2004 ), 通过植株、花序等的资源分配可以很好地解释许多重要繁殖特征, 这在很大程度上协调了不同侧重点研究所得到的矛盾结论(
任明迅等, 2012 ). ...
Biodegradability of soil water soluble organic carbon extracted from seven different soils
1
2009
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
黄土高原水蚀风蚀交错区施肥对黑麦草农田土壤水溶性有机碳、氮的影响
1
2011
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
黄土高原水蚀风蚀交错区施肥对黑麦草农田土壤水溶性有机碳、氮的影响
1
2011
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
Contribution of glomalin to dissolve organic carbon under different land uses and seasonality in dry tropics
1
2017
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
Climate warming impacts on soil organic carbon fractions and aggregate stability in a Tibetan alpine meadow
1
2018
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
荒漠草原“土岛”生境群落物种共存机制
3
2018
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
... 本研究区位于宁夏中部干旱带的荒漠草原, 为鄂尔多斯高原缓坡起伏地带.干旱少雨, 蒸发量大, 冬春两季风大沙多, 属典型的中温带大陆性气候(
宋乃平等, 2018 ).近50年年平均气温为8.46 ℃, 年降水量为276.3 mm, 年日照时间为2 862.6 h.该区域植被主要受水分因素限制(
Ramakrishna et al ., 2003 ), 表现为物种多样性较低、群落结构简单、沙生特征明显, 地下水资源匮乏且植物常常无法利用(
刘小丹等, 2015 ;
Tang et al ., 2017 ;
Wang et al ., 2018 ). ...
... 有研究发现,
C ws 含量与降水量显著负相关, 是因为降水量的增加导致
C ws 淋溶、淋失作用加强, 从而
C ws 含量较低(
刘荣杰等, 2013 ).但本研究区降水较少, 降水的淋溶作用不足以使
C ws 发生淋失, 而
M s 相对较高的生境能促进枯枝落叶的分解, 进而增加了土壤中
C ws 的含量, 因此
C ws 与
M s 呈现出正相关关系.本研究区地下水位较低, 植物往往无法利用, 土壤中水分补给的主要来源是降水, 且具有很强的时空异质性和随机性, 导致该地区植物群落分布多呈斑块状(
宋乃平等, 2018 ).有研究结果表明荒漠植物数量特征的关键驱动力是
M s , 特别是表层
M s (0-20 cm)对荒漠植物群落数量特征起到非常重要的生态作用(
霍举颂等, 2017 ).本研究中, 单株头状花序质量和数量极显著地受到
M s 的影响,
C ws 与单株头状花序质量显著正相关, 这与有关****(
杜华栋等, 2016 )研究发现猪毛蒿叶片形态、解剖和生理指标的可塑性对立地
M s 和
C o 含量较为敏感的结果有相同之处.结合RDA分析图可知,
C ws 和
M s 是影响猪毛蒿繁殖特征的重要因子. ...
荒漠草原“土岛”生境群落物种共存机制
3
2018
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
... 本研究区位于宁夏中部干旱带的荒漠草原, 为鄂尔多斯高原缓坡起伏地带.干旱少雨, 蒸发量大, 冬春两季风大沙多, 属典型的中温带大陆性气候(
宋乃平等, 2018 ).近50年年平均气温为8.46 ℃, 年降水量为276.3 mm, 年日照时间为2 862.6 h.该区域植被主要受水分因素限制(
Ramakrishna et al ., 2003 ), 表现为物种多样性较低、群落结构简单、沙生特征明显, 地下水资源匮乏且植物常常无法利用(
刘小丹等, 2015 ;
Tang et al ., 2017 ;
Wang et al ., 2018 ). ...
... 有研究发现,
C ws 含量与降水量显著负相关, 是因为降水量的增加导致
C ws 淋溶、淋失作用加强, 从而
C ws 含量较低(
刘荣杰等, 2013 ).但本研究区降水较少, 降水的淋溶作用不足以使
C ws 发生淋失, 而
M s 相对较高的生境能促进枯枝落叶的分解, 进而增加了土壤中
C ws 的含量, 因此
C ws 与
M s 呈现出正相关关系.本研究区地下水位较低, 植物往往无法利用, 土壤中水分补给的主要来源是降水, 且具有很强的时空异质性和随机性, 导致该地区植物群落分布多呈斑块状(
宋乃平等, 2018 ).有研究结果表明荒漠植物数量特征的关键驱动力是
M s , 特别是表层
M s (0-20 cm)对荒漠植物群落数量特征起到非常重要的生态作用(
霍举颂等, 2017 ).本研究中, 单株头状花序质量和数量极显著地受到
M s 的影响,
C ws 与单株头状花序质量显著正相关, 这与有关****(
杜华栋等, 2016 )研究发现猪毛蒿叶片形态、解剖和生理指标的可塑性对立地
M s 和
C o 含量较为敏感的结果有相同之处.结合RDA分析图可知,
C ws 和
M s 是影响猪毛蒿繁殖特征的重要因子. ...
The effect of nitrogen addition on community structure and productivity in grasslands: A meta-analysis
1
2017
... 本研究区位于宁夏中部干旱带的荒漠草原, 为鄂尔多斯高原缓坡起伏地带.干旱少雨, 蒸发量大, 冬春两季风大沙多, 属典型的中温带大陆性气候(
宋乃平等, 2018 ).近50年年平均气温为8.46 ℃, 年降水量为276.3 mm, 年日照时间为2 862.6 h.该区域植被主要受水分因素限制(
Ramakrishna et al ., 2003 ), 表现为物种多样性较低、群落结构简单、沙生特征明显, 地下水资源匮乏且植物常常无法利用(
刘小丹等, 2015 ;
Tang et al ., 2017 ;
Wang et al ., 2018 ). ...
Within-population variation in demography of taraxacum officinale: Season- and size-dependent survival, growth and reproduction
1
1997
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
天山雪岭云杉森林群落的密度制约效应
2
2016
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
天山雪岭云杉森林群落的密度制约效应
2
2016
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
沙丘坡面异质性小生境中准噶尔无叶豆对水分条件变化的响应
1
2009
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
沙丘坡面异质性小生境中准噶尔无叶豆对水分条件变化的响应
1
2009
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
A six-year grazing exclusion changed plant species diversity of a
Stipa breviflora desert steppe community, northern China.
1
2018
... 本研究区位于宁夏中部干旱带的荒漠草原, 为鄂尔多斯高原缓坡起伏地带.干旱少雨, 蒸发量大, 冬春两季风大沙多, 属典型的中温带大陆性气候(
宋乃平等, 2018 ).近50年年平均气温为8.46 ℃, 年降水量为276.3 mm, 年日照时间为2 862.6 h.该区域植被主要受水分因素限制(
Ramakrishna et al ., 2003 ), 表现为物种多样性较低、群落结构简单、沙生特征明显, 地下水资源匮乏且植物常常无法利用(
刘小丹等, 2015 ;
Tang et al ., 2017 ;
Wang et al ., 2018 ). ...
The impact of nitrogen amendment and crop growth on dissolved organic carbon in soil solution
0
2016
长白山阔叶红松林木本植物繁殖特征及其关联性
1
2014
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
长白山阔叶红松林木本植物繁殖特征及其关联性
1
2014
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
模拟践踏和降水对高寒草甸阴山扁蓿豆有性繁殖特征的影响
1
2018
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
模拟践踏和降水对高寒草甸阴山扁蓿豆有性繁殖特征的影响
1
2018
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
氮素添加和CO
2 浓度升高对白羊草根际和非根际土壤水溶性有机碳、氮的影响
1
2017
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
氮素添加和CO
2 浓度升高对白羊草根际和非根际土壤水溶性有机碳、氮的影响
1
2017
...
C o 会对荒漠植物产生重要影响(
霍举颂等, 2017 ), 而
C ws 是最活跃的碳组分(
沈玉芳等, 2011 ), 它是能溶解于水, 具有不同大小和结构, 活性较高的有机分子混合体(
Flessa et al ., 2015 ), 可在土壤中迅速转化成其他组分, 为植物提供养分.植物枯枝落叶分解和根系分泌物是
C ws 的重要来源(
肖列等, 2017 ), 而
C ws 又是土壤微生物主要的能量来源.土壤湿度的不同会造成土壤微生物活性的差异, 从而土壤微生物对
C ws 的消耗也不一样, 导致
C ws 含量出现变化(
Fang et al ., 2015 ;
Wang et al ., 2016 ).目前, 对于影响
C ws 的因素和原因的研究相对较多(
Koltes, 2006 ;
Scaglia & Adani, 2009 ;
Singh et al ., 2017 ;
Song et al ., 2018 ), 而
C ws 是如何影响植物生长性状, 特别是如何影响植物繁殖性状的机理机制还有待进一步深入研究. ...
异质环境下入侵植物薇甘菊的适应性与繁殖特性
1
2014
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
异质环境下入侵植物薇甘菊的适应性与繁殖特性
1
2014
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
Plant development and reproduction: Advances and prospectives
1
1999
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
山西灵空山小蛇沟林下草本层植物群落梯度分析及环境解释
1
2013
... 排序轴是反映综合生态梯度的抽象轴, 特征根值是反映相应的排序轴所集中的指标重要值矩阵中信息量的大小, 而相关系数表示的是各排序轴与真实环境梯度之间的相关性(
余敏等, 2013 ).因此进一步对每个繁殖特征进行Monte Carlo置换检验, 结果显示: 在基岩风化沉积土生境下, 对猪毛蒿繁殖特征的影响中,
C ws 的解释量高达71.7% (
p < 0.001), 其次是
TP (55.4%,
p < 0.001), 0-20 cm
M s 和
SH 的解释量分别为32.4%和43.6% (
p < 0.01), 而0-5、5-10和10-20 cm
SH 的解释量同样达到了显著或极显著水平(
p < 0.05或
p < 0.01).
AP 的解释量也达到了21.9% (
p < 0.05).在风沙土生境下, 各土壤因子的解释量均不高(<10%), 其中
C ws 、
TS 、
M s (0-20 cm)和
M s (0-10 cm)的解释量达到了显著水平或极显著水平(
p < 0.05或
p < 0.01).在灰钙土生境下,
M s (0-20 cm)的解释量为47.9% (
p < 0.001), 0-10 cm和10-20 cm的
M s 解释量分别为43.2%和35.3%,
C ws 的解释量同样达到了极显著水平(22.1%,
p < 0.001),
AN 的解释量为10.0%, 达到了显著水平(
p < 0.05).综合分析结果表明,
C ws 和
M s (0-20 cm)的解释量分别为19.9%和16.9%, 达到了极显著水平(
p < 0.001), 同样, 0-10 cm和10-20 cm
M s 的解释水平也达到了极显著水平(
p < 0.001), 说明0-20 cm土层的
M s 对猪毛蒿种群的繁殖特征有显著影响.而其他的土壤因子解释量均未能达到显著水平(
p > 0.05). ...
山西灵空山小蛇沟林下草本层植物群落梯度分析及环境解释
1
2013
... 排序轴是反映综合生态梯度的抽象轴, 特征根值是反映相应的排序轴所集中的指标重要值矩阵中信息量的大小, 而相关系数表示的是各排序轴与真实环境梯度之间的相关性(
余敏等, 2013 ).因此进一步对每个繁殖特征进行Monte Carlo置换检验, 结果显示: 在基岩风化沉积土生境下, 对猪毛蒿繁殖特征的影响中,
C ws 的解释量高达71.7% (
p < 0.001), 其次是
TP (55.4%,
p < 0.001), 0-20 cm
M s 和
SH 的解释量分别为32.4%和43.6% (
p < 0.01), 而0-5、5-10和10-20 cm
SH 的解释量同样达到了显著或极显著水平(
p < 0.05或
p < 0.01).
AP 的解释量也达到了21.9% (
p < 0.05).在风沙土生境下, 各土壤因子的解释量均不高(<10%), 其中
C ws 、
TS 、
M s (0-20 cm)和
M s (0-10 cm)的解释量达到了显著水平或极显著水平(
p < 0.05或
p < 0.01).在灰钙土生境下,
M s (0-20 cm)的解释量为47.9% (
p < 0.001), 0-10 cm和10-20 cm的
M s 解释量分别为43.2%和35.3%,
C ws 的解释量同样达到了极显著水平(22.1%,
p < 0.001),
AN 的解释量为10.0%, 达到了显著水平(
p < 0.05).综合分析结果表明,
C ws 和
M s (0-20 cm)的解释量分别为19.9%和16.9%, 达到了极显著水平(
p < 0.001), 同样, 0-10 cm和10-20 cm
M s 的解释水平也达到了极显著水平(
p < 0.001), 说明0-20 cm土层的
M s 对猪毛蒿种群的繁殖特征有显著影响.而其他的土壤因子解释量均未能达到显著水平(
p > 0.05). ...
基于植被光谱监测喀斯特异质性生境可行性研究
1
2012
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
基于植被光谱监测喀斯特异质性生境可行性研究
1
2012
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
1
2004
... 植物种群的更新和个体生活史的完成均依赖于繁殖过程(
全杜娟等, 2012 ;
Larson & Funk, 2016 ), 植物的繁殖性状是植物资源投入-收益长期权衡的结果(
张大勇, 2004 ), 通过植株、花序等的资源分配可以很好地解释许多重要繁殖特征, 这在很大程度上协调了不同侧重点研究所得到的矛盾结论(
任明迅等, 2012 ). ...
1
2004
... 植物种群的更新和个体生活史的完成均依赖于繁殖过程(
全杜娟等, 2012 ;
Larson & Funk, 2016 ), 植物的繁殖性状是植物资源投入-收益长期权衡的结果(
张大勇, 2004 ), 通过植株、花序等的资源分配可以很好地解释许多重要繁殖特征, 这在很大程度上协调了不同侧重点研究所得到的矛盾结论(
任明迅等, 2012 ). ...
植物交配系统的进化、资源分配对策与遗传多样性
2
2001
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
植物交配系统的进化、资源分配对策与遗传多样性
2
2001
... 繁殖不仅是种群形成、发展和进化的核心问题, 也是生物群落和生态系统演替的基础(
陈亭和王一峰, 2014 ).不同种类的植物采取不同的繁殖方式以维持其种群的延续, 并形成多样化的分布格局(
L'Hérault et al ., 2013 ;
王芸芸等, 2014 ).因此, 繁殖特征一定程度上反映了植物适应环境变化的策略(
Gélin et al ., 2016 ;
肖红等, 2018 ).同时, 了解植物的繁殖特征也是制定有效管理措施的重要依据(
Kendall et al ., 2011 ), 特别是植物自然繁殖更新能力作为干旱区植被管理和恢复实践的重要研究内容, 一直受到人们的关注(
Vavrek et al ., 1997 ;
刘志民等, 2003a ).自20世纪80年代以来, 对植物繁殖适应性的研究逐渐增多, 繁殖生态学成了国际生态学领域的一个新热点(
Xu, 1999 ;
张大勇和姜新华, 2001 ). ...
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
不同立地类型大果沙棘生长及生态适应对策差异
1
2009
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
不同立地类型大果沙棘生长及生态适应对策差异
1
2009
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
秦王川湿地柽柳分布与环境因子的关系
1
2017
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
秦王川湿地柽柳分布与环境因子的关系
1
2017
... 在自然界, 植物所处的生境条件在时间和空间上都是变化的, 不仅在大的空间尺度上存在, 在小尺度上也被探测到(
董鸣, 1996 ), 甚至10 cm尺度上微生境的异质性也有****进行了研究(
Gray & Spies, 1997 ), 均发现生境的异质性对植物的生长、定居、存活、分布等产生影响.宁夏中部荒漠草原相对海拔变化较小, 无明显的气候垂直分布区域, 海拔差异不会对气压、气温的差异等造成很大影响, 该区域土壤水分、养分状况主要受地形起伏、地面高度、土壤类型等差异的影响形成异质性环境, 进而造成植被空间分布格局的差异(
刘方等, 2008 ;
赵连春等, 2017 ), 植被类型往往表现为斑块性(
宋乃平等, 2018 ).因此, 生境异质性是影响植物的重要因素(
Palmer & Dixon, 2010 ;
陈云等, 2016 ;
王慧杰等, 2016 ).而植物的繁殖性状同时受到内在遗传和外在环境因子的共同影响(
李冰等, 2013 ), 外在环境的作用对植物的影响更显著(
张大勇和姜新华, 2001 ).分析影响植物繁殖特征的众多因素并找出主要影响因子, 是需要重点研究的内容. ...
塔里木河中下游湿地及周边植物群落与环境因子的关系初探
1
2006
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
塔里木河中下游湿地及周边植物群落与环境因子的关系初探
1
2006
... 生境的异质性在不同植物群落中普遍存在, 直接影响植物的生长发育(
Olmo et al ., 2016 ).在相似的气候条件下, 土壤理化性质的空间差异决定了植被分布的空间差异(
赵锐锋等, 2006 ), 也制约了干旱地区植物生命过程, 对植物种类、生长、分布及数量特征产生重要影响(
岳跃民等, 2012 ;
Johnson et al ., 2016 ).反过来, 生活在不同环境条件和资源状况中的植物往往具有与之相对应的形态和生理特点(
王建成等, 2009 ), 植物种群的繁殖特征往往也表现出明显差异(
张治来等, 2009 ).本研究中, 猪毛蒿的个体大小、单株头状花序数量、单个头状花序质量、单株头状花序质量在异质生境条件下均表现出一定的差异, 说明不同土壤类型和
M s 条件对其生长和繁殖特征的影响较为明显, 灰钙土生境下猪毛蒿个体较大, 且单株头状花序质量较大、数量较多, 但繁殖分配和单个头状花序质量却不是最大的.有研究发现, 生境异质性可以影响群落中植被恢复和补充的各个阶段, 尤其是幼苗建成过程中的早期生长阶段(
刘方炎等, 2011 ), 甚至对花期物候和坐果率都有显著影响(
黄至欢等, 2017 ).薇甘菊(
Mikania micrantha )叶片功能性状、开花结实和繁殖分配对异质环境条件也具有不同的适应对策(
徐高峰等, 2014 ).芦苇(
Phragmites australis )种群在根茎长度、生物量、干物质贮量等数量特征上在生境间的差异均表现出明显的土壤因子环境效应(
焦德志等, 2018 ).而异质生境中长花柱花性状的变化是导致雪地报春(
Primula nivalis )花性状分化的主要因素(
艾沙江·阿不都沙拉木, 2018 ).以上研究结果均说明植物会对异质生境产生相适应的对策, 因此植物适应的对象是生存环境, 但最终目标是物种的持续生存和繁衍(
何维明和钟章成, 1997 ). ...
 ,, 王磊, 杨新国, 宋乃平
,, 王磊, 杨新国, 宋乃平 ,*, 李月飞*, 苏莹*, 卞莹莹*, 祝忠有*, 孟文婷*宁夏大学西北土地退化与生态恢复国家重点实验室培育基地, 银川 750021;宁夏大学西北土地退化与生态恢复国家重点实验室培育基地, 银川 750021
,*, 李月飞*, 苏莹*, 卞莹莹*, 祝忠有*, 孟文婷*宁夏大学西北土地退化与生态恢复国家重点实验室培育基地, 银川 750021;宁夏大学西北土地退化与生态恢复国家重点实验室培育基地, 银川 750021
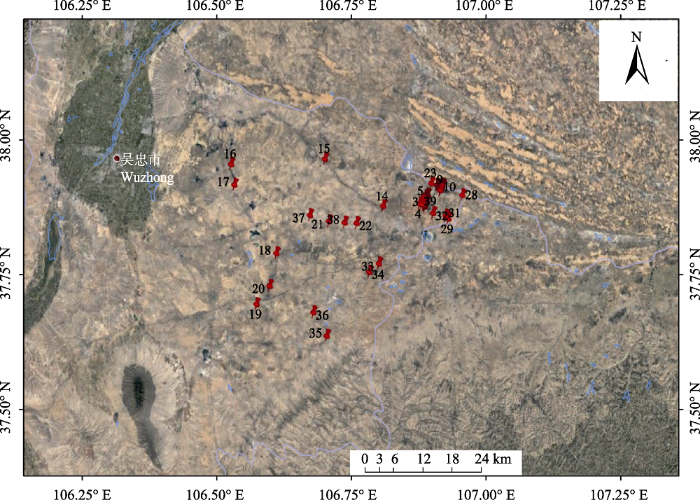 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT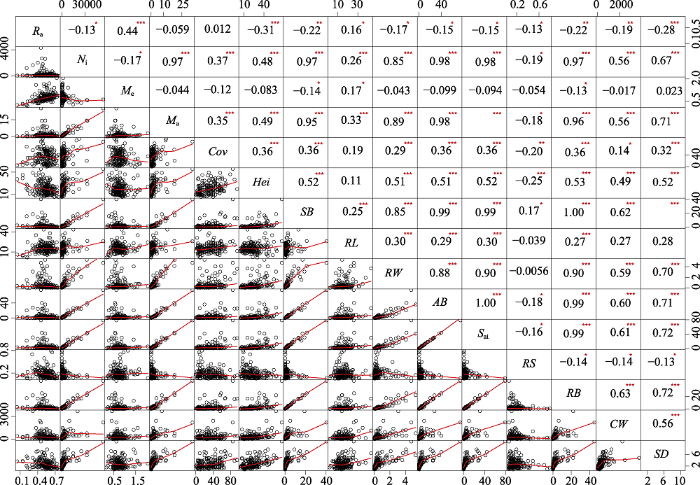 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT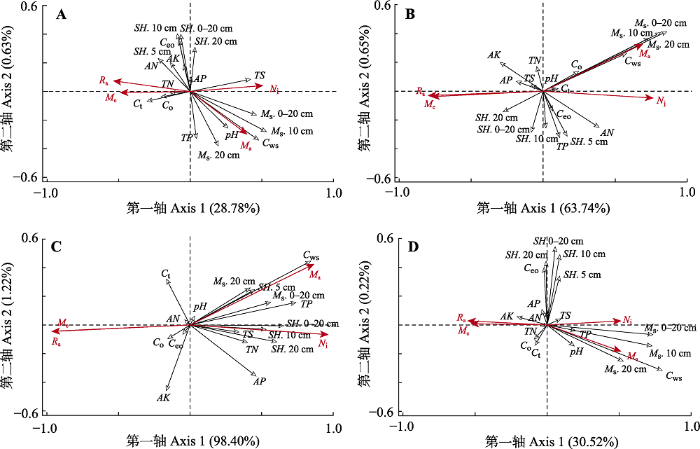 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}