Effects of vegetation restoration on soil organic carbon mineralization in the east of Hunan, China
GU Xiang1, ZHANG Shi-Ji1, LIU Zhao-Dan1, LI Lei-Da1, CHEN Jin-Lei1, WANG Liu-Fang1, FANG Xi,,1,2,3,*1 School of Life Science and Technology, Central South University of Forestry and Technology, Changsha 410004, China 2 National Engineering Laboratory of South China Forestry Ecology Applicable Technologies, Changsha 410004, China 3 Huitong National Field Station for Scientific Observation and Research of Chinese Fir Plantation Ecosystem in Hunan Province, Huitong, Hunan 438107, China
Supported by the National Forestry Public Welfare Industry Research Project..201504411 the National Natural Science Foundation of China.30771720 the National Natural Science Foundation of China.31170426
Abstract Aims Soil organic carbon (SOC) mineralization and its controlling factors are critical for understanding the mechanisms that regulate C storage and loss processes. Our objectives were to accurately assess the impacts of vegetation restoration on SOC mineralization and to illustrate the underlying mechanism of changes in SOC stability with vegetation restoration in the mid-subtropical region of China.Methods Four types of vegetation communities (Loropetalum chinense-Vaccinium bracteatum-Rhododendron mariesiiscrub-grass-land (LVR), Loropetalum chinense-Cunninghamia lanceolata-Quercus fabri shrubbery (LCQ), Pinus massoniana-Lithocarpus glaber-Loropetalum chinense coniferous-broad leaved mixed forest (PLL) and Lithocarpus glaber-Cleyera japonica-Cyclobalanopsis glauca evergreen broad-leaved forest (LAG) ) were selected for a successional sequence of the secondary forest in a hilly region of eastern Hunan Province, China. Soil samples (0-40 cm) were collected from four depths (0-10, 10-20, 20-30 and 30-40 cm). SOC mineralization was determined by the laboratory incubation method (alkali absorption method). Principal component analysis and stepwise regression were applied to analyze the relationships between SOC cumulative mineralization (Cm), the percentage of Cm to SOC content and vegetation factors and soil factors.Important findings 1) SOC mineralization rate displayed similar trends during the four restoration stages: during the early incubation period, SOC mineralization was high and decreased fast, and then the rate decreased slowly and tend to stabilize during the middle and late periods. The pattern was well fitted with the reciprocal equation. 2) SOC mineralization rate and Cm increased significantly with the vegetation restoration stage with the LAG having both the highest SOC mineralization rate and the highest Cm. In 0-40 cm soil layer, Cm in LAG soil was 359.06%-716.31%, 112.38%-232.61%, 94.40%-105.74% higher than that in LVR, LCQ, PLL soils, respectively. 3) The percentage of Cm to SOC content were 2.13%-4.99%, 3.42%-4.18%, 4.05%-4.64%, and 4.02%- 5.64% in 0-10, 10-20, 20-30, 30-40 cm soil layers, respectively. However, there was no significant difference in the percentage of Cm to SOC content among the different restoration stages. 4) Among different vegetation restoration stages, total nitrogen (TN) content and root biomass (RB) were the key factors affecting Cm. TN content and RB together explained 97.8% variations in Cm, of which TN content explained 96.9%. However, C:N was the dominant factor affecting the percentage of Cm to SOC content and it alone explained 49.4% variation. Vegetation restoration promoted SOC mineralization rate and reduced the percentage of Cm to SOC content in our study, which might be related to the changes in RB and soil nutrient during vegetation restoration. Keywords:mid-subtropical region of China;vegetation restoration;soil organic carbon mineralization;vegetation factor;soil factor
PDF (1856KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 本文引用格式 辜翔, 张仕吉, 刘兆丹, 李雷达, 陈金磊, 王留芳, 方晰. 湖南东部植被恢复对土壤有机碳矿化的影响. 植物生态学报[J], 2018, 42(12): 1211-1224 doi:10.17521/cjpe.2018.0202 GU Xiang. Effects of vegetation restoration on soil organic carbon mineralization in the east of Hunan, China. Chinese Journal of Plant Ecology[J], 2018, 42(12): 1211-1224 doi:10.17521/cjpe.2018.0202
土壤有机碳(SOC)矿化是土壤碳(C)输出的主要途径, 是大气中CO2的重要来源, 在维持全球C平衡中具有极其重要的作用(Saggar et al., 2001; Craine et al., 2010)。SOC矿化不仅反映SOC的稳定性和周转(周玉燕等, 2011; 黄宗胜等, 2012), 而且关系到土壤养分元素的释放与供应。因此, 研究SOC矿化特征及其影响因素是陆地生态系统C循环的研究热点, 有助于揭示SOC释放的驱动机制及其稳定性, 为深入研究土壤固C能力的影响机制和估算土壤C平衡提供基础数据, 而且对科学管理土壤养分, 有效缓解全球气候变暖具有重要意义(Zhu et al., 2017)。
Fig. 1The location and distribution of the different vegetation communities in the east of Hunan Province. LVR, Loropetalum chinense-Vaccinium bracteatum-Rhododendron mariesii scrub-?grass-?land; LCQ, Loropetalum chinense-Cunninghamia lanceolata-??Quercus fabri shrubbery; PLL, Pinus massoniana-?Lithocarpus glaber-Loropetalum chinense coniferous-broad leaved mixed forest; LAG, Lithocarpus glaber-Cleyera japonica-?Cyclobalanopsis glauca evergreen broad-leaved forest.
Table 1 Table 1Plant community biomass and nutrient characteristics of litter layer at different vegetation restoration stages in the east of Hunan Province (mean$\pm$SD)
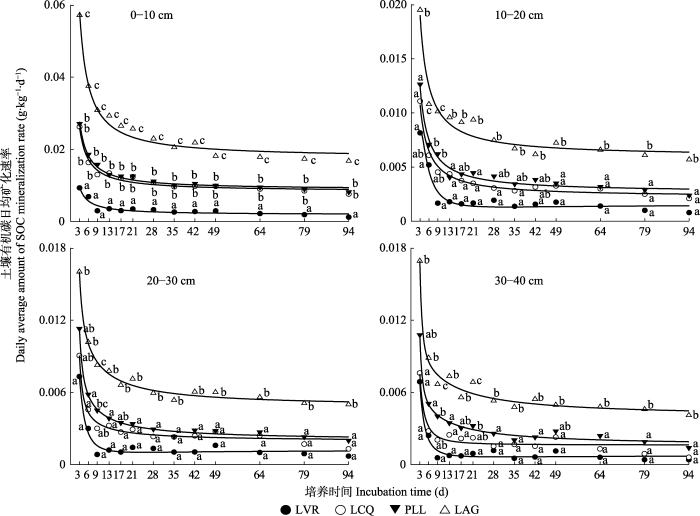
Fig. 2Soil organic carbon (SOC) mineralization rate at different vegetation restoration stages in the east of Hunan Province. LVR, Loropetalum chinense-Vaccinium bracteatum-Rhododendron mariesii scrub-grass-land; LCQ, Loropetalum chinense-Cunninghamia lanceolata-Quercus fabri shrubbery; PLL, Pinus massoniana-Lithocarpus glaber-Loropetalum chinense coniferous-broad leaved mixed forest; LAG, Lithocarpus glaber-Cleyera japonica-Cyclobalanopsis glauca evergreen broad-leaved forest. Different letters indicate significant differences among different vegetation restoration stages in the same soil layer at the same time of incubation (p < 0.05).
Table 3 表3 表3湖南东部不同植被恢复阶段土壤有机碳矿化速率拟合 Table 3Fitting of soil organic carbon (SOC) mineralization rate with incubation time using the reciprocal equations at different restoration stages in the east of Hunan Province
Fig. 3Soil organic carbon cumulative mineralization (Cm) at different vegetation restoration stages in the east of Hunan Province (mean + SD). LVR, Loropetalum chinense-Vaccinium bracteatum-Rhododendron mariesii scrub-grass-land; LCQ, Loropetalum chinense-Cunninghamia lanceolata-Quercus fabri shrubbery; PLL, Pinus massoniana-Lithocarpus glaber-?Loropetalum chinense coniferous-broad leaved mixed forest; LAG, Lithocarpus glaber-Cleyera japonica-Cyclobalanopsis glauca evergreen broad-leaved forest. Different capital letters represent significant differences among different vegetation restoration stages in the same soil layer (p < 0.05), and different lower letters indicate significant differences among different soil layers at the same vegetation restoration stage (p < 0.05).
Fig. 4The percentage of soil organic carbon cumulative mineralization to soil organic carbon content (Cm/SOC) at different vegetation restoration stages in the east of Hunan Province (mean + SD). LVR, Loropetalum chinense-Vaccinium bracteatum-Rhododendron mariesii scrub-grass-land; LCQ, Loropetalum chinense-Cunninghamia lanceolata-Quercus fabri shrubbery; PLL, Pinus massoniana-Lithocarpus glaber-?Loropetalum chinense coniferous-broad leaved mixed forest; LAG, Lithocarpus glaber-Cleyera japonica-Cyclobalanopsis glauca evergreen broad-leaved forest. Different capital letters represent significant differences among different vegetation restoration stages in the same soil layer (p < 0.05), and different lower letters indicate significant differences among different soil layers at the same vegetation restoration stage (p < 0.05).
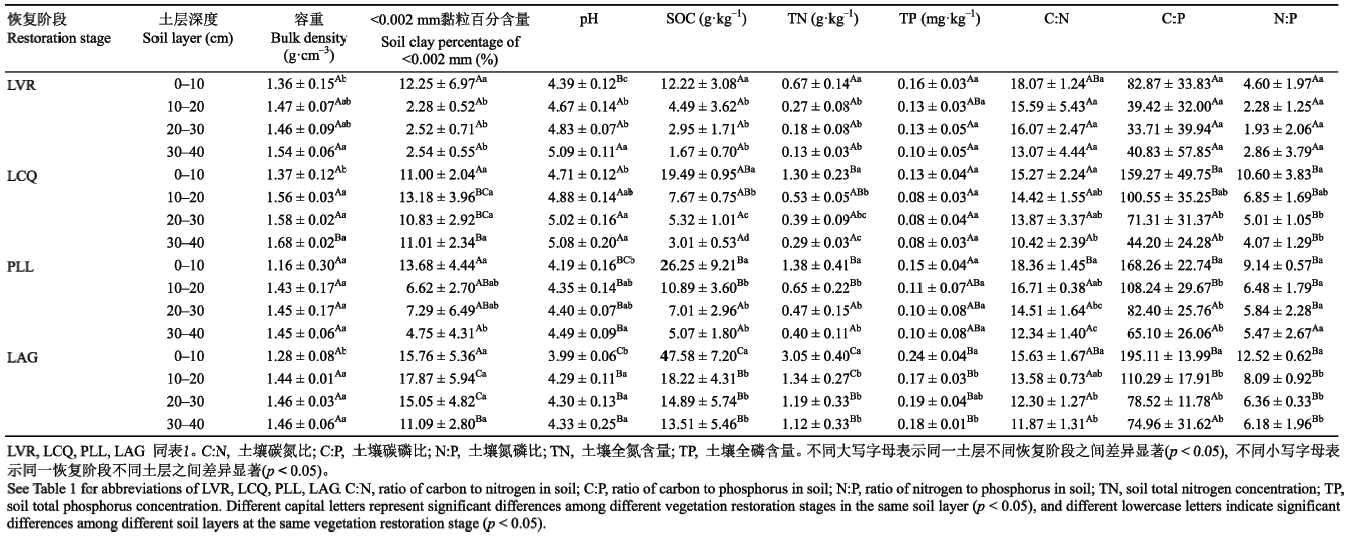
Table 4 表4 表4土壤有机碳累积矿化量(Cm)、土壤有机碳矿化率(Cm/SOC)与植被因子的Pearson相关系数 Table 4Pearson correlation coefficient between soil organic carbon (SOC) cumulative mineralization (Cm), the percentage of Cm to SOC content (Cm/SOC) and vegetation factors, respectively
Table 5 表5 表5土壤有机碳(SOC)累积矿化量(Cm)、土壤有机碳矿化率(Cm/SOC)与土壤因子的Pearson相关系数 Table 5Pearson correlation coefficient between soil organic carbon (SOC) cumulative mineralization (Cm), the percentage of Cm to SOC content (Cm/SOC) and soil factors, respectively
项目 Item
容重 Bulk density
<0.002 mm黏粒百分含量 Soil clay percentage of <0.002 mm
pH
SOC
TN
TP
C:N
C:P
N:P
Cm
-0.312
0.774**
-0.734**
0.971**
0.986**
0.595*
-0.315
0.653*
0.738**
Cm/SOC
0.154
-0.203
0.323
-0.320
-0.223
-0.172
-0.732**
-0.268
-0.096
C:N, ratio of carbon to nitrogen in soil; C:P, ratio of carbon to phosphorus in soil; N:P, ratio of nitrogen to phosphorus in soil; TN, soil total nitrogen concentration; TP, soil total phosphorus concentration. n = 13, * p < 0.05, ** p < 0.01. C:N, 土壤碳氮比; C:P, 土壤碳磷比; N:P, 土壤氮磷比; TN, 土壤全氮含量; TP, 土壤全磷含量。n = 13, * p < 0.05, ** p < 0.01。
Table 7 表7 表7土壤有机碳累积矿化量(Cm)和土壤有机碳矿化率(Cm/SOC)影响因子的逐步回归分析 Table 7The stepwise regression analysis for main influencing factors of soil organic carbon (SOC) cumulative mineralization (Cm) and the percentage of Cm to SOC content (Cm/SOC)
项目 Item
模型 Model
变量 Variable
回归方程 Regression equation
多元相关系数 Multiple correlation coefficient (R)
调整判定系数 Adjust R2
F
P
Cm
1
TN
Cm = 0.572 TN + 0.250
0.986
0.969
380.581
0.000
2
RB
Cm =0.474 TN + 0.000006 RB + 0.290
0.991
0.978
273.558
0.000
Cm/SOC
1
Soil C:N
Cm/SOC = -0.302 C:N +8.717
0.732
0.494
12.700
0.004
TN, soil total nitrogen concentration; RB, Root biomass; Soil C:N, ratio of carbon to nitrogen in soil. n = 13。TN, 土壤全氮含量; RB, 根系生物量; Soil C:N, 土壤碳氮比。
CraineJM, FiererN, MclauchlanKK ( 2010). Widespread coupling between the rate and temperature sensitivity of organic matter decay Nature Geoscience, 3, 854-857. DOI:10.1038/ngeo1009URL [本文引用: 1] Microbial breakdown of soil organic matter influences the potential for terrestrial ecosystems to sequester carbon, and the amount of carbon dioxide released to the atmosphere. Predicting the sensitivity of microbial decomposition to temperature change is therefore critical to predicting future atmospheric carbon dioxide concentrations and feedbacks to anthropogenic warming. According to enzyme kinetics, the more biogeochemically recalcitrant the organic matter, the greater the temperature sensitivity of microbial respiration. Here, we measured the temperature sensitivity of microbial respiration in soils from 28 sites in North America, ranging from Alaska to Puerto Rico, to test the generality of this principle. We show that the lower the rate of respiration at a reference temperature of 20 C-and thus the more biogeochemically recalcitrant the organic matter-the greater the temperature sensitivity of soil respiration. We compiled our findings with those from other studies, encapsulating a range of environments, and show that this relationship holds across multiple scales and soil types. Although physico-chemical protection of soil organic matter and substrate availability will also influence the temperature sensitivity of decomposition, we suggest that biogeochemically recalcitrant organic matter will respond the most sensitively to anticipated warming.
CuiJ, ZhangRJ, BuNS, ZhangHB, TangBP, LiZL, JiangLF, ChenJK, FangCM ( 2013). Changes in soil carbon sequestration and soil respiration following afforestation on paddy fields in north subtropical China Journal of Plant Ecology, 6, 240-252. DOI:10.1093/jpe/rts023URL [本文引用: 1] ABSTRACT AimsAlthough many studies have reported net gains of soil organic carbon (SOC) after afforestation on croplands, this is uncertain for Chinese paddy rice croplands. Here, we aimed to evaluate the effects of afforestation of paddy rice croplands on SOC sequestration and soil respiration (Rs). Such knowledge would improve our understanding of the effectiveness of various land use options on greenhouse gas mitigation in China.Methods The investigation was conducted on the Chongming Island, north subtropical China. Field sites were reclaimed from coastal salt marshes in the 1960s, and soils were homogeneous with simple land use histories. SOC stocks and Rs levels were monitored over one year in a paddy rice cropland, an evergreen and a deciduous broad-leaved plantation established on previous paddy fields and a reference fallow land site never cultivated. Laboratory incubation of soil under fast-changing temperatures was used to compare the temperature sensitivity (Q10) of SOC decomposition across land uses.Important FindingsAfter 15-20 years of afforestation on paddy fields, SOC concentration only slightly increased at the depth of 0-5cm but decreased in deeper layers, which resulted in a net loss of SOC stock in the top 40cm. Seasonal increase of SOC was observed during the rice-growing period in croplands but not in afforested soils, suggesting a stronger SOC sequestration by paddy rice cropping. However, SOC sequestered under cropping was more labile, as indicated by its higher contents of dissolved organic carbon and microbial biomass. Also, paddy soils had higher annual Rs than afforested soils; Rs abruptly increased after paddy fields were drained and plowed and remained distinctively high throughout the dry farming period. Laboratory incubation revealed that paddy soils had a much higher Q10 of SOC decomposition than afforested soils. Given that temperature was the primary controller of R s in this region, it was concluded that despite the stronger SOC sequestration by paddy rice cropping, its SOC was less stable than in afforested systems and might be more easily released into the atmosphere under global warming. 2012 The Author 2012. Published by Oxford University Press on behalf of the Institute of Botany, Chinese Academy of Sciences and the Botanical Society of China. All rights reserved.
FanYX, YangYS, GuoJF, YangZJ, ChenGS, XieJS, ZhongXJ, XuLL ( 2014). Changes in soil respiration and its temperature sensitivity at different successional stages of evergreen broadleaved forests in mid-subtropical China Chinese Journal of Plant Ecology, 38, 1155-1165. DOI:10.3724/SP.J.1258.2014.00111URL [本文引用: 2] 为探明中亚热带地区常绿阔叶林演替序列土壤呼吸(Rs)的变化趋势及其影响机制,在福建省建瓯市万木林自 然保护区选取演替时间分别为15年(演替初期)、47年(演替中期)和110年(演替后期)三个不同演替阶段,进行了为期1年的野外原位观测.结果发现: 演替初期、中期和后期的Rs分别为2.38、3.32和3.91 μmol·m-2·s-1,温度敏感性(Q10值)分别为2.64、1.97和1.79;与演替初期相比,演替后期的Rs显著增加64.29%,Q10值 显著降低32.30%;不同演替阶段Rs的季节变化模式相似,温度和含水量可分别解释季节变化的69.5%(初期)、81.9%(中期)和61.3%(后 期);回归分析发现,Rs与凋落物年归还量、细根生物量和土壤全氮和土壤有机质碳含量显著正相关.表明本研究区内植被演替促进了土壤碳排放,降低了土壤呼 吸的温度敏感性;土壤碳输入增加、养分含量的提高和细根生物量增大是中亚热带常绿阔叶林Rs随演替进程逐渐增大的主要原因. [ 范跃新, 杨玉盛, 郭剑芬, 杨智杰, 陈光水, 谢锦升, 钟小剑, 徐玲琳 ( 2014). 中亚热带常绿阔叶林不同演替阶段土壤呼吸及其温度敏感性的变化 植物生态学报, 38, 1155-1165.] DOI:10.3724/SP.J.1258.2014.00111URL [本文引用: 2] 为探明中亚热带地区常绿阔叶林演替序列土壤呼吸(Rs)的变化趋势及其影响机制,在福建省建瓯市万木林自 然保护区选取演替时间分别为15年(演替初期)、47年(演替中期)和110年(演替后期)三个不同演替阶段,进行了为期1年的野外原位观测.结果发现: 演替初期、中期和后期的Rs分别为2.38、3.32和3.91 μmol·m-2·s-1,温度敏感性(Q10值)分别为2.64、1.97和1.79;与演替初期相比,演替后期的Rs显著增加64.29%,Q10值 显著降低32.30%;不同演替阶段Rs的季节变化模式相似,温度和含水量可分别解释季节变化的69.5%(初期)、81.9%(中期)和61.3%(后 期);回归分析发现,Rs与凋落物年归还量、细根生物量和土壤全氮和土壤有机质碳含量显著正相关.表明本研究区内植被演替促进了土壤碳排放,降低了土壤呼 吸的温度敏感性;土壤碳输入增加、养分含量的提高和细根生物量增大是中亚热带常绿阔叶林Rs随演替进程逐渐增大的主要原因.
GuX, FangX, XiangWH, LiSL, SunWJ ( 2013). Soil organic carbon and mineralizable organic carbon in four subtropical forests in hilly region of central Hunan Province, China Chinese Journal of Ecology, 32, 2687-2694. URL [本文引用: 1] 采用短期室内土壤培养法,比较研究了湘中丘陵区4种不同森林类型(杉木人工林、马尾松+石栎针阔混交林、南酸枣落叶阔叶林、石栎+青冈常绿阔叶林)土壤有机碳、可矿化有机碳的含量。结果表明,4种森林土壤有机碳含量、可矿化有机碳含量以及可矿化有机碳的分配比例均随土壤深度的增加而下降。同一土层有机碳含量从高至低的排序均表现为:石栎+青冈常绿阔叶林>马尾松+石栎针阔混交林>南酸枣落叶阔叶林>杉木人工林,0~30 cm土层,南酸枣落叶阔叶林、马尾松+石栎针阔混交林、石栎+青冈常绿阔叶林土壤有机碳含量比杉木人工林分别高出20.8%、29.7%和31.9%。可矿化有机碳含量及其分配比例从高至低的排序均表现为:石栎+青冈常绿阔叶林>南酸枣落叶阔叶林>马尾松+石栎针阔混交林>杉木人工林,0~30 cm土层,石栎+青冈常绿阔叶林、南酸枣落叶阔叶林、马尾松+石栎针阔混交林可矿化有机碳含量比杉木人工林分别高出51.6%、45.2%和41.0%,可矿化有机碳的分配比例比杉木人工林分别高出35.8%、29.3%和20.0%,4种森林土壤可矿化有机碳与总有机碳、土壤微生物生物量碳、全N、C/N之间均呈极显著相关性。 [ 辜翔, 方晰, 项文化, 李胜蓝, 孙伟军 ( 2013). 湘中丘陵区4种森林类型土壤有机碳和可矿化有机碳的比较 生态学杂志, 32, 2687-2694.] URL [本文引用: 1] 采用短期室内土壤培养法,比较研究了湘中丘陵区4种不同森林类型(杉木人工林、马尾松+石栎针阔混交林、南酸枣落叶阔叶林、石栎+青冈常绿阔叶林)土壤有机碳、可矿化有机碳的含量。结果表明,4种森林土壤有机碳含量、可矿化有机碳含量以及可矿化有机碳的分配比例均随土壤深度的增加而下降。同一土层有机碳含量从高至低的排序均表现为:石栎+青冈常绿阔叶林>马尾松+石栎针阔混交林>南酸枣落叶阔叶林>杉木人工林,0~30 cm土层,南酸枣落叶阔叶林、马尾松+石栎针阔混交林、石栎+青冈常绿阔叶林土壤有机碳含量比杉木人工林分别高出20.8%、29.7%和31.9%。可矿化有机碳含量及其分配比例从高至低的排序均表现为:石栎+青冈常绿阔叶林>南酸枣落叶阔叶林>马尾松+石栎针阔混交林>杉木人工林,0~30 cm土层,石栎+青冈常绿阔叶林、南酸枣落叶阔叶林、马尾松+石栎针阔混交林可矿化有机碳含量比杉木人工林分别高出51.6%、45.2%和41.0%,可矿化有机碳的分配比例比杉木人工林分别高出35.8%、29.3%和20.0%,4种森林土壤可矿化有机碳与总有机碳、土壤微生物生物量碳、全N、C/N之间均呈极显著相关性。
GuX, ZhangSJ, LiuZD, LiLD, ChenJL, WangLF, FangX ( 2018). Effects of vegetation restoration on soil organic carbon concentration and density in the mid-subtropical region of China Chinese Journal of Plant Ecology, 42, 595-608. [本文引用: 10]
HuYL, ZengDH, FanZP, AiGY ( 2007). Effects of degraded sandy grassland afforestation on soil quality in semiarid area of Northern China Chinese Journal of Applied Ecology, 18, 2391-2397. [本文引用: 1]
HuangZS, YuLF, FuYH ( 2012). Characteristics of soil mineralizable carbon pool in natural restoration process of karst forest vegetation Chinese Journal of Applied Ecology, 23, 2165-2170. URL [本文引用: 3] By the method of taking space instead of time, an incubation test was conducted to study the characteristics of soil mineralizable carbon pool during the natural restoration of Karst forest vegetation in Maolan Nature Reserve, Guizhou Province of Southwest China. It was observed that the contents of soil total organic carbon (TOC) and mineralizable carbon (MC) as well as the carbon mineralization rate decreased with increasing soil depth but increased with the process of vegetation restoration. The amount of cumulative released carbon and the carbon release rate increased with the process of restoration, but the release rate decreased with increasing incubation time. The soil MC/TOC increased with the restoration process but had less change with increasing soil depth, while the O decreased with increasing soil depth and through the process of restoration. The soil MC had a negative correlation with the existing litter amount (=-0.796) but positive correlation with the mass loss rate of the litter decomposition (=0.924). Soil habitat changed from strong interference at early stages to relative stability at late stages, and soil carbon sequestration changed from small capacity and strong potential at early stages to large capacity and weak potential at late stages. [ 黄宗胜, 喻理飞, 符裕红 ( 2012). 喀斯特森林植被自然恢复过程中土壤可矿化碳库特征 应用生态学报, 23, 2165-2170.] URL [本文引用: 3] By the method of taking space instead of time, an incubation test was conducted to study the characteristics of soil mineralizable carbon pool during the natural restoration of Karst forest vegetation in Maolan Nature Reserve, Guizhou Province of Southwest China. It was observed that the contents of soil total organic carbon (TOC) and mineralizable carbon (MC) as well as the carbon mineralization rate decreased with increasing soil depth but increased with the process of vegetation restoration. The amount of cumulative released carbon and the carbon release rate increased with the process of restoration, but the release rate decreased with increasing incubation time. The soil MC/TOC increased with the restoration process but had less change with increasing soil depth, while the O decreased with increasing soil depth and through the process of restoration. The soil MC had a negative correlation with the existing litter amount (=-0.796) but positive correlation with the mass loss rate of the litter decomposition (=0.924). Soil habitat changed from strong interference at early stages to relative stability at late stages, and soil carbon sequestration changed from small capacity and strong potential at early stages to large capacity and weak potential at late stages.
IyamuremyeF, GewinV, DickRP, DiackM, SeneM, BadianeA ( 2000). Carbon, nitrogen and phosphorus mineralization potential of native agroforestry plant residues in soils of Senegal Arid Soil Research and Rehabilitation, 14, 359-371. DOI:10.1080/08903060050136469URL [本文引用: 1] The objectives of this study were to investigate Piliostigma reticulatum (shrub) and Cordyla pinnata (tree) residues for chemical composition, and C, N, and P mineralizationin Senegalese soils. Soil samples (Sols ferrugineux tropicaux) were collected from a native agroforestry system beneath and outside a C. pinnata canopy. Soils were incubated for 11 weeks with P. reticulatum (leaves), C. pinnata (stem, leaves or stem + leaves), peanut (Arachis hypogea), or pearl millet (Pennisetum glaucum) residues. Nitrogen and P mineralization for the soil-plant mixtures was determined by periodic leaching with a 0.01 M CaCl 2 solution. An additional separate incubation was conducted to investigate C mineralization. The results showed that only peanut residues caused net N mineralization, while N immobilization occurred in the remaining treatments in both the soils derived from beneath or outside the canopy. This indicates that, at least in the short-term, these agroforestry residues would not likely be a source of N for crops. Net P mineralization varied among plant residues and soils sites but P. reticulatum amended soils increased soluble PO in both soils. This suggested that it could be useful for improving P avail4 ability. Peanut residues had the highest CO evolution in both soils suggesting a 2 probable relationship between C and N mineralization.
LiJY, DouXL, HuYH, GanDX, LiF ( 2018). Variation in soil organic carbon mineralization under various land cover types in urban areas Acta Ecologica Sinica, 38, 112-121. DOI:10.5846/stxb201612302718URL [本文引用: 1] 土壤有机碳(SOC)矿化是陆地生态系统碳循环的重要过程。因受到强烈的人为干扰,城市土壤生态服务功能严重退化,进而对城市土壤地球化学循环尤其是碳循环产生深刻的影响。以北京市奥林匹克森林公园的5种典型地表覆盖类型(草坪、灌木、行道树、植草砖、硬化地表)下土壤为研究对象,研究了城市不同地表覆盖类型下土壤有机碳矿化过程及固碳能力的差异。结果表明,城市5种地表覆盖类型下的土壤有机碳矿化趋势与自然生态系统中的土壤基本一致,都表现为前期矿化较为快速,后期明显减慢并且趋于平稳;不同地表覆盖类型下土壤的有机碳矿化作用有显著差异,灌木、行道树、植草砖覆盖下土壤有机碳矿化能力较强,硬化地表和草坪较弱,与土壤有机碳含量特征类似;一级动力学方程对各土样有机碳矿化过程的模拟结果较好,结果显示草坪覆盖下土壤固碳能力较强,灌木覆盖下次之,行道树、植草砖和硬化地表覆盖下较弱;土壤固碳能力的高低并不对应着土壤有机碳含量的高低,城市人为干扰和外源有机碳的输入对土壤有机碳储量影响较大;硬化地表下不同土层有机碳矿化作用无明显差异,而其他地表覆盖类型下的土壤有机碳矿化作用随土层加深显著减弱,特别是植草砖和行道树特征最为明显;各地表覆盖类型下土壤固碳能力随土层深度变化的规律不显著。城市土壤有机碳矿化的最主要限制因子是土壤有机碳的含量,土壤pH值、养分含量、粘粒含量等性质也通过影响土壤有机碳含量及微生物活动等对土壤有机碳矿化过程产生影响。 [ 李隽永, 窦晓琳, 胡印红, 甘德欣, 李锋 ( 2018). 城市不同地表覆盖类型下土壤有机碳矿化的差异 生态学报, 38, 112-121.] DOI:10.5846/stxb201612302718URL [本文引用: 1] 土壤有机碳(SOC)矿化是陆地生态系统碳循环的重要过程。因受到强烈的人为干扰,城市土壤生态服务功能严重退化,进而对城市土壤地球化学循环尤其是碳循环产生深刻的影响。以北京市奥林匹克森林公园的5种典型地表覆盖类型(草坪、灌木、行道树、植草砖、硬化地表)下土壤为研究对象,研究了城市不同地表覆盖类型下土壤有机碳矿化过程及固碳能力的差异。结果表明,城市5种地表覆盖类型下的土壤有机碳矿化趋势与自然生态系统中的土壤基本一致,都表现为前期矿化较为快速,后期明显减慢并且趋于平稳;不同地表覆盖类型下土壤的有机碳矿化作用有显著差异,灌木、行道树、植草砖覆盖下土壤有机碳矿化能力较强,硬化地表和草坪较弱,与土壤有机碳含量特征类似;一级动力学方程对各土样有机碳矿化过程的模拟结果较好,结果显示草坪覆盖下土壤固碳能力较强,灌木覆盖下次之,行道树、植草砖和硬化地表覆盖下较弱;土壤固碳能力的高低并不对应着土壤有机碳含量的高低,城市人为干扰和外源有机碳的输入对土壤有机碳储量影响较大;硬化地表下不同土层有机碳矿化作用无明显差异,而其他地表覆盖类型下的土壤有机碳矿化作用随土层加深显著减弱,特别是植草砖和行道树特征最为明显;各地表覆盖类型下土壤固碳能力随土层深度变化的规律不显著。城市土壤有机碳矿化的最主要限制因子是土壤有机碳的含量,土壤pH值、养分含量、粘粒含量等性质也通过影响土壤有机碳含量及微生物活动等对土壤有机碳矿化过程产生影响。
LiY, XuM, SunOJ, CuiW ( 2004). Effects of root and litter exclusion on soil CO2 efflux and microbial biomass in wet tropical forests Soil Biology & Biochemistry, 36, 2111-2114. DOI:10.1016/j.soilbio.2004.06.003URL [本文引用: 1] We examined the effects of root and litter exclusion on the rate of soil CO 2 efflux and microbial biomass at a soil depth of 25 cm in a secondary forest (dominated by Tabebuia heterophylla) and a pine ( Pinus caribaea) plantation in the Luquillo Experimental Forest in Puerto Rico. The experimental plots were initially established in 1990, when root, forest floor mass and new litterfall were excluded for 7 y since then. Soil respiration was significantly reduced in the litter and root exclusion plots in both the secondary forest and the pine plantation compared with the control. Root exclusion had a greater effect on soil CO 2 efflux than the litter exclusion in the plantation, whereas a reversed pattern was observed in the secondary forest. The reduction of microbial biomass in the root exclusion plot was greater in the secondary forest (59%) than in the plantation (31%), while there was no difference of the reduction in the litter exclusion plots between these forests. Our results suggest that above-ground input and roots (root litter and exudates) differentially affect soil CO 2 efflux under different vegetation types.
LiYM, GongL, XieLN ( 2017). Soil organic carbon content and carbon mineralization characteristics under different land use types in Northern Tarim Basin Bulletin of Soil and Water Conservation, 37(3), 216-221. DOI:10.13961/j.cnki.stbctb.2017.03.036URL [本文引用: 2] [目的]探讨土地利用方式对土壤有机碳含量及碳矿化的影响,为塔里木盆地北缘绿洲土壤生态系统的保护和恢复建设提供理论依据。[方法]基于野外采样和室内培养试验,分析土壤有机碳含量的基本特征,利用回归分析法拟合出土壤有机碳矿化动态变化过程。[结果]矿化累积释放的CO2含量大小依次为:果园〉棉田〉人工林〉弃耕地〉荒草地〉盐碱地〉沙地。不同土地利用方式土壤有机碳矿化反应趋势相同,1~6d为快速分解阶段,日均矿化量高但反应时间短,6~28d为缓慢分解阶段,动态变化与前者相反。有机碳矿化率大小依次为:沙地〉荒草地〉盐碱地〉弃耕地〉人工林〉棉田〉果园,沙地最高,达(10.36±0.24)%,表明沙地土壤有机碳稳定性最差,而果园具有较强的固定有机碳能力。[结论]土地利用方式对土壤有机碳矿化及其固碳能力均有显著影响。 [ 李杨梅, 贡璐, 解丽娜 ( 2017). 塔里木盆地北缘绿洲不同土地利用方式下土壤有机碳含量及其碳矿化特征 水土保持通报, 37(3), 216-221.] DOI:10.13961/j.cnki.stbctb.2017.03.036URL [本文引用: 2] [目的]探讨土地利用方式对土壤有机碳含量及碳矿化的影响,为塔里木盆地北缘绿洲土壤生态系统的保护和恢复建设提供理论依据。[方法]基于野外采样和室内培养试验,分析土壤有机碳含量的基本特征,利用回归分析法拟合出土壤有机碳矿化动态变化过程。[结果]矿化累积释放的CO2含量大小依次为:果园〉棉田〉人工林〉弃耕地〉荒草地〉盐碱地〉沙地。不同土地利用方式土壤有机碳矿化反应趋势相同,1~6d为快速分解阶段,日均矿化量高但反应时间短,6~28d为缓慢分解阶段,动态变化与前者相反。有机碳矿化率大小依次为:沙地〉荒草地〉盐碱地〉弃耕地〉人工林〉棉田〉果园,沙地最高,达(10.36±0.24)%,表明沙地土壤有机碳稳定性最差,而果园具有较强的固定有机碳能力。[结论]土地利用方式对土壤有机碳矿化及其固碳能力均有显著影响。
LiuS, YuanWY, LiXW, RongL, YangM, LiDH, WangQ ( 2008). Soil chemical characteristics and their effects on C:N ratio of rhizosphere and non-rizosphere of different herbages in triploid of Populous tomentosa woodland. Reseaech of Soil and Water Conservation, 15(2), 107-110. URLMagsci [本文引用: 1] 采用剥落分离法对三倍体毛白杨+黑麦草与三倍体毛白杨+自然草2种模式的根际与非根际土化学特性进行研究表明:两模式林地非根际土C/N值和pH值均大于其对应的根际土.自然草模式根际与非根际土C/N值大于黑麦草模式根际与非根际土,而pH值却相反.黑麦草模式根际(或非根际)土pH值大于的自然草模式根际(或非根际);两模式根际与非根际土N/P值(有效N与有效P的比值)接近14:1;两模式根际与非根际土速效K在4-5月均处于低谷,10-11月处于高峰;黑麦草模式根际土C/N值与速效K呈现较显著正相关,自然草模式根际土C/N值与N/P值呈现较显著正相关. [ 刘朔, 袁渭阳, 李贤伟, 荣丽, 杨渺, 李德会, 王巧 ( 2008). 不同草本层三倍体毛白杨根际与非根际土壤化学特性及其对C/N的影响 水土保持研究, 15(2), 107-110.] URLMagsci [本文引用: 1] 采用剥落分离法对三倍体毛白杨+黑麦草与三倍体毛白杨+自然草2种模式的根际与非根际土化学特性进行研究表明:两模式林地非根际土C/N值和pH值均大于其对应的根际土.自然草模式根际与非根际土C/N值大于黑麦草模式根际与非根际土,而pH值却相反.黑麦草模式根际(或非根际)土pH值大于的自然草模式根际(或非根际);两模式根际与非根际土N/P值(有效N与有效P的比值)接近14:1;两模式根际与非根际土速效K在4-5月均处于低谷,10-11月处于高峰;黑麦草模式根际土C/N值与速效K呈现较显著正相关,自然草模式根际土C/N值与N/P值呈现较显著正相关.
LiuYQ, WangF, KeGQ, WangYY, GuoSM, FanCF ( 2011). Effects of converting cultivated land into forest land on the characteristics of soil organic carbon in limestone mountain area in Ruichang, Jiangxi Chinese Journal of Applied Ecology, 22, 885-890. URL [本文引用: 1] 以江西瑞昌石灰岩山区退耕还林5年的林地为对象,研究了不同退耕还林模式(林苗一体化、药用纯林、多树种混交和竹阔混交)对土壤有机碳含量的影响.结果表明:与对照相比,退耕还林地0~20 cm土层的土壤有机碳、微生物生物量碳和可矿化碳含量分别增加24.4%、29%和18.4%,退耕还林显著提高了土壤有机碳含量,增强了土壤碳储能力;0~10 cm土层的土壤有机碳、微生物生物量碳和可矿化碳含量极显著高于10~20 cm土层,且退耕还林地土层间差异大于对照;4种退耕还林模式中,林苗一体化模式对土壤有机碳的影响较大. [ 刘苑秋, 王芳, 柯国庆, 王迎迎, 郭圣茂, 范承芳 ( 2011). 江西瑞昌石灰岩山区退耕还林对土壤有机碳的影响 应用生态学报, 22, 885-890.] URL [本文引用: 1] 以江西瑞昌石灰岩山区退耕还林5年的林地为对象,研究了不同退耕还林模式(林苗一体化、药用纯林、多树种混交和竹阔混交)对土壤有机碳含量的影响.结果表明:与对照相比,退耕还林地0~20 cm土层的土壤有机碳、微生物生物量碳和可矿化碳含量分别增加24.4%、29%和18.4%,退耕还林显著提高了土壤有机碳含量,增强了土壤碳储能力;0~10 cm土层的土壤有机碳、微生物生物量碳和可矿化碳含量极显著高于10~20 cm土层,且退耕还林地土层间差异大于对照;4种退耕还林模式中,林苗一体化模式对土壤有机碳的影响较大.
MilcuA, HeimA, EllisRJ, ScheuS, ManningP ( 2011). Identification of general patterns of nutrient and labile carbon control on soil carbon dynamics across a successional gradient Ecosystems, 14, 710-719. DOI:10.1007/s10021-011-9440-zURL [本文引用: 1] Carbon (C) inputs and nutrient availability are known to affect soil organic carbon (SOC) stocks. However, general rules regarding the operation of these factors across a range of soil nutrient availabilities and substrate qualities are unidentified. "Priming" (stimulated decomposition by labile C inputs) and 'preferential substrate utilization' (retarded decomposition due to shifts in community composition towards microbes that do not mineralize SOC) are two hypotheses to explain effects of labile C additions on SOC dynamics. For effects of nutrient additions (nitrogen and phosphorus) on SOC dynamics, the stoichiometric (faster decomposition of materials of low carbon-to-nutrient ratios) and 'microbial mining' (that is, reduced breakdown of recalcitrant C forms for nutrients under fertile conditions) hypotheses have been proposed. Using the natural gradient of soil nutrient availability and substrate quality of a chronosequence, combined with labile C and nutrient amendments, we explored the support for these contrasting hypotheses. Additions of labile C, nitrogen (N), phosphorus (P), and combinations of C and N and C and P were applied to three sites: 2-year fallow grassland, mature grassland and forest, and the effects of site and nutrient additions on litter decomposition and soil C dynamics were assessed. The response to C addition supported the preferential substrate hypothesis for easily degradable litter C and the priming hypothesis for SOC, but only in nitrogen-enriched soils of the forest site. Responses to N addition supported the microbial mining hypothesis irrespective of C substrate (litter or SOC), but only in the forest site. Further, P addition effects on SOC support the stoichiometric hypothesis; P availability appeared key to soil C release (priming) in the forest site if labile C and N is available. These results clearly link previously contrasting hypotheses of the factors controlling SOC with the natural gradient in litter quality and nutrient availability that exists in ecosystems at different successional stages. A holistic theory that incorporates this variability of responses, due to different mechanisms, depending on nutrient availability and substrate quality is essential for devising management strategies to safeguard soil C stocks.
PrihaO, GraystonSJ ( 2001). Microbial community structure and characteristics of the organic matter in soils under Pinus sylvestris, Picea abies and Betula pendula at two forest sites Biology & Fertility of Soils, 33, 17-24. [本文引用: 1]
QianHY, PanJJ, SunB ( 2013). The relative impact of land use and soil properties on sizes and turnover rates of soil organic carbon pools in subtropical China Soil Use & Management, 29, 510-518. DOI:10.1111/sum.12067URL [本文引用: 1] The dynamics of soil organic carbon (SOC) pools determine potential carbon sequestration and soil nutrient improvement. This study investigated the characteristics of SOC pools in five types of cultivated topsoils (0 15 cm) in subtropical China using laboratory incubation experiments under aerobic conditions. The sizes and turnover rates of the active, slow and resistant C pools were simulated using a first-order kinetic model. The relative influence of soil environmental properties on the dynamics of different SOC pools was evaluated by applying principal component analysis (PCA) and aggregated boosted trees (ABTs) analysis. The results show that there were significantly greater sizes of different SOC pools and lower turnover rates of slow C pool in two types of paddy soils than in upland soils. Land use exerted the most significant influence on the sizes of all SOC pools, followed by clay content and soil pH. The soil C/N ratio and pH were the major determinants for turnover rates of the active and slow C pools, followed by clay content which had more impact on the turnover rates of the active C pool than the slow C pool. It is concluded that soil type exerts a significant impact on the dynamics of SOC.
RezaMT, BeckerW, SachsenheimerK, MummeJ ( 2014). Hydrothermal carbonization (HTC): Near infrared spectroscopy and partial least-squares regression for determination of selective components in HTC solid and liquid products derived from maize silage Bioresource Technology, 161, 91-101. DOI:10.1016/j.biortech.2014.03.008URLPMID:24686376 [本文引用: 1] Near-infrared (NIR) spectroscopy was evaluated as a rapid method of predicting fiber components (hemicellulose, cellulose, lignin, and ash) and selective compounds of hydrochar and corresponding process liquor produced by hydrothermal carbonization (HTC) of maize silage. Several HTC reaction times and temperatures were applied and NIR spectra of both HTC solids and liquids were obtained and correlated with concentration determined from van-Soest fiber analysis, IC, and UHPLC. Partial least-squares regression was applied to calculate models for the prediction of selective substances. The model developed with the spectra had the best performance in 3–7 factors with a correlation coefficient, which varied between 0.9275–0.9880 and 0.9364–0.9957 for compounds in solid and liquid, respectively. Calculated root mean square errors of prediction (RMSEP) were 0.42–5.06mg/kg. The preliminary results indicate that NIR, a widely applied technique, might be applied to determine chemical compounds in HTC solid and liquid.
RiffaldiR, SaviozziA, Levi-MinziR ( 1996). Carbon mineralization kinetics as in?uenced by soil properties Biological & Fertility of Soils, 22, 293-298. [本文引用: 1]
RossDJ, TateKR, ScottNA, WildeRH, RoddaNJ, TownsendJA ( 2002). Afforestation of pastures with Pinus radiata influences soil carbon and nitrogen pools and mineralisation and microbial properties Soil Research, 40, 1303-1318. DOI:10.1071/sr02020URL [本文引用: 1] In New Zealand, Pinus radiata D. Don is frequently planted on land under pasture primarily for production forestry, but with the added advantage of potentially offsetting carbon dioxide (CO2) emissions from energy and industrial sources. Conversion of pasture to P. radiata plantations can, however, result in lowered contents of soil carbon (C) at some sites. We here examine the effects of this land-use change on soil C and nitrogen (N) pools, and on microbial properties involved in the cycling of these nutrients, at 5 paired sites, each with an established pasture and P. radiata plantation. Four sites had first-rotation trees aged 12–30 years and the other site second-rotation trees aged 20 years. In mineral soil at 0–10 cm depth, total and microbial C and N, extractable C, CO2-C production, and, generally, net mineral-N production were lower under P. radiata than under pasture; differences were significant (P < 0.05), except for total and extractable C at 2 sites. Differences between these land uses were less distinct in soil at 10–30 cm depth. On an area basis, total C in 0–30 cm depth soil was lower under P. radiata than under pasture at most sites, but significantly lower at only one site. Total N, microbial C and N, and CO2-C and net mineral-N production were, however, again generally significantly lower under P. radiata. These ecosystem differences were less marked, although still present, except for CO2-C production, when forest litter (LFH material) was included in the area calculations. Overall, our study suggests that afforestation with P. radiata leads to a reduction in total N, microbial biomass, and microbial activity, but a less consistent effect on soil C storage after one rotation.
SaggarS, YeatesGW, ShepherdTG ( 2001). Cultivation effects on soil biological properties, microfauna and organic matter dynamics in Eutric Gleysol and Gleyic Luvisol soils in New Zealand Soil & Tillage Research, 8, 55-68. DOI:10.1016/S0167-1987(00)00184-7URL [本文引用: 1] Information on the influence of duration of cultivation on soil organic matter (SOM) dynamics is needed by researchers, policy makers and farmers for assessment of global ramifications of carbon (C) sequestration and agricultural sustainabilty. This paper examines the changes in microbial biomass carbon (MBC) and nitrogen (MBN), metabolic quotient ( qCO 2), microfaunal populations, and C and N mineralisation to assess the effect of cultivation on SOM dynamics. Cultivation, for up to 34 years, of soils previously under permanent ryegrass ( Lolium perenne L.) and clover ( Trifolium sp.) pastures, resulted in a 30–60% decline in concentration of soil organic C and N, and had a significant influence on soil biological parameters in two New Zealand (Eutric Gleysol, Kairanga silty clay loam and Gleyic Luvisol, Marton silt loam) soil types. Cultivated soils had consistently lower MBC and MBN contents, and higher qCO 2 than their counterparts in permanent pastures. Changes in the soil microfauna following cultivation were consistent with changes in microbial biomass. Both the amount and proportion of mineralisable N were also reduced with cultivation. In the pasture soils C mineralisation rates (under laboratory conditions) were twice those (ca. 15 mg (CO 2-C) kg 611 soil) in the 5–20 year cultivated soils (ca. 7 mg (CO 2-C) kg 611 soil), and were reduced to one-half (ca. 3.5 mg (CO 2-C) kg 611 soil) in the 34-year cultivated soil. Over 112 days, the soils lost approximately 2.4–5.0% of their total C, and 1.4–5.0% of total N was mineralised. The percentage C loss also differed with soil type, with Marton silt loam (260 g kg 611, clay) soil losing one and half times as much percentage C as Kairanga silty clay loam (420 g kg 611, clay) soil. Despite greater C mineralisation in soil from pasture, more C and N were conserved in it than in the cultivated soils. As annual C inputs in cultivated soils are smaller than the C decomposition, this would result in a negative soil C balance. The shifts in microbial biomass, its metabolic quotients and soil microfauna appear to be associated with differences in the quantity and ‘quality’ of inputs and SOM decomposition rates, and reflect the land-use change from pasture to continuous cultivation.
SchaeferDA, FengWT, ZouXM ( 2009). Plant carbon inputs and environmental factors strongly affect soil respiration in a subtropical forest of southwestern China Soil Biology & Biochemistry, 41, 1000-1007. DOI:10.1016/j.soilbio.2008.11.015URL [本文引用: 1] Soil respiration is a large component of global carbon fluxes, so it is important to explore how this carbon flux varies with environmental factors and carbon inputs from plants. As part of a long-term study on the chemical and biological effects of aboveground litterfall denial, root trenching and tree-stem girdling, we measured soil respiration for three years in plots where those treatments were applied singly and in combination. Tree-stem girdling terminates the flow of carbohydrates from canopy, but allows the roots to continue water and nutrient uptake. After carbon storage below the stem girdles is depleted, the girdled trees die. Root trenching immediately terminates root exudates as well as water and nutrient uptake. Excluding aboveground litterfall removes soil carbon inputs, but allows normal root functions to continue. We found that removing aboveground litterfall and the humus layer reduced soil respiration by more than the C input from litter, a respiration priming effect. When this treatment was combined with stem girdling, root trenching or those treatments in combination, the change in soil respiration was indistinguishable from the loss of litterfall C inputs. This suggests that litterfall priming occurs only when normal root processes persist. Soil respiration was significantly related to temperature in all treatment combinations, and to soil water content in all treatments except stem girdling alone, and girdling plus trenching. Aboveground litterfall was a significant predictor of soil respiration in control, stem-girdled, trenched and stem-girdled plus trenching treatments. Stem girdling significantly reduced soil respiration as a single factor, but root trenching did not. These results suggest that in addition to temperature, aboveground carbon inputs exert strong controls on forest soil respiration.
Scott-DentonLE, SparksKL, MonsonRK ( 2003). Spatial and temporal controls of soil respiration rate in a high- elevation, subalpine forest Soil Biology & Biochemistry, 35, 525-534. DOI:10.1016/S0038-0717(03)00007-5URL [本文引用: 1] We examined soil respiration to determine what measurable environmental variables can be used to predict variation in soil respiration rates, spatially and temporally, at a high-elevation, mixed conifer, subalpine forest site at the Niwot Ridge Ameriflux Site in Colorado. For three summers, soil respiration rates were measured using soil collars and a portable gas-exchange system. Transects of the collars were established to ensure spatial characterization of the litter-repleted areas beneath tree crowns and the litter-depleted open spaces between tree crowns. Soil temperature and soil moisture were both identified as important drivers of soil respiration rate, but were found to confound each other and to function as primary controls at different scales. Soil temperature represents a primary control seasonally, and soil moisture represents a primary control interannually. Spatially, organic layer thickness, ammonium concentration, water content, and the microbial and soil soluble carbon pools were found to predict variation from point to point. Soil microbial biomass strongly correlated to soil respiration rate, whereas root biomass was identified as a weak predictor of respiration rate and only when controlling for other variables. Spatial variation in soil respiration rate is highly determined by the depth of the soil organic horizon, which in this ecosystem varies predictably according to distance from trees. The conclusions that can be drawn from the study provide the foundation for the development of future models of soil respiration driven by fundamental variables of the climate and soil microenvironment.
ShaoYH, PanJJ, SunB ( 2005). Study of characteristics of soil organic carbon decompositions and carbon pool under different vegetations Journal of Soil and Water Conservation, 19(3), 24-28. DOI:10.3321/j.issn:1009-2242.2005.03.007URL [本文引用: 1] The characteristics of soil organic carbon (SOC)decompositions and the carbon pool sizes and turnover rates of active carbon pool,slow carbon pool and passive carbon pool under different vegetations and Chinese fir plantation of different forest ages are discussed. The results indicate that the general trend is there is a faster decomposition rate in the first incubation period than in the second period and in the surface soil than in the subsoil.In the surface soil,effect of decomposition rate on different vegetations is broad leaved forest>mature Chinese fir plantation and on Chinese fir plantation of different forest ages is midaged forest>mature forest>young forest.In the subsoil,there is a few differences on decomposition rate and culmulative amount of carbon mineralized under different vegetations;Effect of culmulative amount of soil organic carbon mineralized on different vegetations and Chinese fir plantation of different forest ages is the first month>the second month>the third month, the surface soil>the subsoil.In the surface soil,the effect is midaged Chinese fir plantation > broad leaved forest >mature Chinese fir plantation >young Chinese fir plantation;There is a different carbon pool sizes and turnover rate of activecarbon pool, slow carbon pool and passive carbon pool under different vegetations and Chinese fir plantation of different forest ages,the active pool(Ca)comprised 1.70% to 4.98% of the SOC in the surface and subsoil with an average field mean residence time(MRT) of 10 to 23 d,the slow pool comprised 16.21% to 49.52% of the SOC ,they had field MRT of 0.8 to 13.1 year,the passive pool comprised 46.22% to 80.50% with a assumed field MRT of 1 000 year. [ 邵月红, 潘剑君, 孙波 ( 2005). 不同森林植被下土壤有机碳的分解特征及碳库研究 水土保持学报, 19(3), 24-28.] DOI:10.3321/j.issn:1009-2242.2005.03.007URL [本文引用: 1] The characteristics of soil organic carbon (SOC)decompositions and the carbon pool sizes and turnover rates of active carbon pool,slow carbon pool and passive carbon pool under different vegetations and Chinese fir plantation of different forest ages are discussed. The results indicate that the general trend is there is a faster decomposition rate in the first incubation period than in the second period and in the surface soil than in the subsoil.In the surface soil,effect of decomposition rate on different vegetations is broad leaved forest>mature Chinese fir plantation and on Chinese fir plantation of different forest ages is midaged forest>mature forest>young forest.In the subsoil,there is a few differences on decomposition rate and culmulative amount of carbon mineralized under different vegetations;Effect of culmulative amount of soil organic carbon mineralized on different vegetations and Chinese fir plantation of different forest ages is the first month>the second month>the third month, the surface soil>the subsoil.In the surface soil,the effect is midaged Chinese fir plantation > broad leaved forest >mature Chinese fir plantation >young Chinese fir plantation;There is a different carbon pool sizes and turnover rate of activecarbon pool, slow carbon pool and passive carbon pool under different vegetations and Chinese fir plantation of different forest ages,the active pool(Ca)comprised 1.70% to 4.98% of the SOC in the surface and subsoil with an average field mean residence time(MRT) of 10 to 23 d,the slow pool comprised 16.21% to 49.52% of the SOC ,they had field MRT of 0.8 to 13.1 year,the passive pool comprised 46.22% to 80.50% with a assumed field MRT of 1 000 year.
SongXZ, JiangH, YuSQ, MaYD, ZhouGM, DouRP, GuoPP ( 2009). Litter decomposition of dominant plant species in successional stages in mid-subtropical zone Chinese Journal of Applied Ecology, 20, 537-542. URL [本文引用: 1] 选择我国亚热带森林群落3个主要演替阶段的7个优势种(其中马尾松代表演替初期优势种,木荷 和香樟代表演替中期优势种,甜槠、小叶青冈栎、青冈和乐昌含笑代表演替后期优势种)的凋落物,采用网袋法进行分解试验。结果表明:马尾松凋落物分解得最 慢,年分解速率为0.51;木荷和香樟居中,分别为0.55和0.61;小叶青冈栎和乐昌含笑分解得最快,分别为0.89和1.12。沿着植被顺向演替的 梯度,凋落物分解速度呈现加快的趋势。分解速率同凋落物的初始P、N和木质素含量及木质素/N比值呈极显著相关(P〈0.01),同C/N比值有显著相关 关系(P〈0.05)。凋落物的P、N和木质素含量及木质素/N比值是预测凋落物分解快慢的良好指标。 [ 宋新章, 江洪, 余树全, 马元丹, 周国模, 窦荣鹏, 郭培培 ( 2009). 中亚热带森林群落不同演替阶段优势种凋落物分解试验 应用生态学报, 20, 537-542.] URL [本文引用: 1] 选择我国亚热带森林群落3个主要演替阶段的7个优势种(其中马尾松代表演替初期优势种,木荷 和香樟代表演替中期优势种,甜槠、小叶青冈栎、青冈和乐昌含笑代表演替后期优势种)的凋落物,采用网袋法进行分解试验。结果表明:马尾松凋落物分解得最 慢,年分解速率为0.51;木荷和香樟居中,分别为0.55和0.61;小叶青冈栎和乐昌含笑分解得最快,分别为0.89和1.12。沿着植被顺向演替的 梯度,凋落物分解速度呈现加快的趋势。分解速率同凋落物的初始P、N和木质素含量及木质素/N比值呈极显著相关(P〈0.01),同C/N比值有显著相关 关系(P〈0.05)。凋落物的P、N和木质素含量及木质素/N比值是预测凋落物分解快慢的良好指标。
SongY, ZhaoXZ, MaoZJ, SunT, HouLL ( 2013). SOC decomposition of four typical broad-leaved Korean pine communities in Xiaoxing ,an Mountain Acta Ecologica Sinica, 33, 443-453. DOI:10.5846/stxb201112021844URL [本文引用: 1] 土壤有机碳分解是陆地生态系统碳循环的重要组成部分。主要采用土壤有机碳释放速率的室内培养实验的方法,并根据三库一级动力学模型,对小兴安岭地区4种典型阔叶红松林的土壤有机碳分解特征及各组分含量进行研究。实验结果如下:(1)土壤有机碳的分解趋势表现为前期迅速,后期缓慢,并且土壤腐殖质层(A)大于淀积层(B);在4种阔叶红松林中,云冷杉红松林土壤有机碳的分解速率最大,枫桦红松林最小;土壤有机碳的分解速率与土壤总有机碳、活性碳及土壤的C/N呈显著的正相关关系(P〈0.05)。(2)在土壤A层和B层,4种阔叶红松林的活性碳分别占总有机碳的0.89%—1.78%和1.91%—2.87%,平均驻留时间为12—35 d和27—58 d。缓效性碳占总有机碳的22.58%—28.44%和23.87%—42.63%,平均驻留时间为4—19 a和18—37 a。惰性碳占总有机碳的69.98%—76.24%和54.50%—74.22%,平均驻留时间为173 a;土壤有机碳各组分含量及驻留时间的大小顺序均为:云冷杉红松林〉椴树红松林〉枫桦红松林〉蒙古栎红松林。 [ 宋媛, 赵溪竹, 毛子军, 孙涛, 侯玲玲 ( 2013). 小兴安岭4种典型阔叶红松林土壤有机碳分解特性 生态学报 , 33, 443-453.] DOI:10.5846/stxb201112021844URL [本文引用: 1] 土壤有机碳分解是陆地生态系统碳循环的重要组成部分。主要采用土壤有机碳释放速率的室内培养实验的方法,并根据三库一级动力学模型,对小兴安岭地区4种典型阔叶红松林的土壤有机碳分解特征及各组分含量进行研究。实验结果如下:(1)土壤有机碳的分解趋势表现为前期迅速,后期缓慢,并且土壤腐殖质层(A)大于淀积层(B);在4种阔叶红松林中,云冷杉红松林土壤有机碳的分解速率最大,枫桦红松林最小;土壤有机碳的分解速率与土壤总有机碳、活性碳及土壤的C/N呈显著的正相关关系(P〈0.05)。(2)在土壤A层和B层,4种阔叶红松林的活性碳分别占总有机碳的0.89%—1.78%和1.91%—2.87%,平均驻留时间为12—35 d和27—58 d。缓效性碳占总有机碳的22.58%—28.44%和23.87%—42.63%,平均驻留时间为4—19 a和18—37 a。惰性碳占总有机碳的69.98%—76.24%和54.50%—74.22%,平均驻留时间为173 a;土壤有机碳各组分含量及驻留时间的大小顺序均为:云冷杉红松林〉椴树红松林〉枫桦红松林〉蒙古栎红松林。
SunSQ, BhattiJS, JassalRS, ChangSX, ArevaloC, BlackTA, SiddersD ( 2015). Stand age and productivity control soil carbon dioxide efflux and organic carbon dynamics in poplar plantations Soil Science Society of America Journal, 79, 1638-1649. DOI:10.2136/sssaj2015.06.0233URL [本文引用: 2] Establishment of hybrid poplar (HP) plantations has been advocated as an effective method for sequestering CO2 from the atmosphere. However, how carbon (C) cycling in HP plantations changes during stand development under different soil fertility and climatic conditions is poorly understood. We studied the dynamics of soil respiration (R-s) and its heterotrophic (R-h) and autotrophic (R-a) components and soil organic C (SOC) content in HP plantations of different stand ages established on soils with different productivity (Class 1 vs. Class 3 soils) in Alberta (AB) and Ontario (ON), Canada. The Rs was the highest for AB-Class 1 (high-productivity) soils, followed by AB-Class 3 and ON-Class 3 (low-productivity) soils. Annual Rs, Rh, and R-h/R-s decreased in the first 3 yr in AB-Class 1 and 6 yr in AB-Class 3 soils after plantation establishment and increased thereafter, but they increased with stand age after plantation establishment in ON-Class 3 soils. The SOC content in the AB-Class 1, AB-Class 3, and ON-Class 3 sites decreased in the first 3, 5, and 1.5 yr, respectively, after plantation establishment and then increased, reaching the pre-establishment level after 6, 10, and 3 yr, respectively. From this first attempt to understand C cycling across multiple sites with different soil fertility levels, we conclude that stand age and soil productivity play a significant role in affecting C cycling, and such effects should be considered in regional and global C cycling models, especially when assessing the C sequestration potential of HP plantations.
SunWJ, FangX, XiangWH, ZhangSJ, LiSL ( 2013). Active pools of soil organic carbon in subtropical forests at different successional stages in Central Hunan, China Acta Ecologica Sinica, 33, 7765-7773. DOI:10.5846/stxb201301060046URL [本文引用: 1] 为了解天然次生林保护对土壤活 性有机碳库的影响,采用空间替代时间研究方法,对湘中丘陵区不同演替阶段4种林分类型(杉木人工林、马尾松+石栎针阔混交林、南酸枣落叶阔叶林、青冈+石 栎常绿阔叶林)土壤活性有机碳及其与土壤养分相关性进行研究。结果表明:1)各土层总有机碳(TOC)、微生物生物量碳(MBC)、水溶性有机碳 (DOC)、易氧化有机碳(EOC)含量均表现为:青冈+石栎常绿阔叶林南酸枣落叶阔叶林马尾松+石栎针阔混交林杉木人工林,在0—30cm土层,马尾 松+石栎针阔混交林、南酸枣落叶阔叶林、青冈+石栎常绿阔叶林TOC含量比杉木人工林分别高出13.40%、19.40%和29.91%,MBC含量分别 高出15.62%、32.89%和53.33%,DOC含量分别高出8.52%、8.75%和13.76%,EOC含量分别高出32.79%、 38.48%和78.30%;2)天然次生林各土层MBC占TOC的比率以南酸枣落叶阔叶林最高,青冈+石栎常绿阔叶林为其次,马尾松+石栎混交林最低, 均高于同一土层杉木人工林(除马尾松+石栎混交林15—30cm土层外),天然次生林各土层DOC占TOC的比率随着演替进展而下降,均低于同一土层杉木 人工林(除马尾松+石栎混交林0—15cm土层外),天然次生林各土层EOC占TOC的比率随着演替进展而增加,且均高于同一土层杉木人工林;3)土壤 MBC、DOC、EOC含量与TOC含量的相关性均达到极显著水平,且天然次生林土壤MBC、DOC、EOC含量与TOC含量的相关系数随着演替进展而增 高,均高于杉木人工林;4)4种林分土壤TOC、MBC、DOC、EOC含量与土壤全N、碱解N、全P、有效P、全K、速效K含量之间的相关性均达到显著 或极显著水平。 [ 孙伟军, 方晰, 项文化, 张仕吉, 李胜蓝 ( 2013). 湘中丘陵区不同演替阶段森林土壤活性有机碳库特征 生态学报, 33, 7765-7773.] DOI:10.5846/stxb201301060046URL [本文引用: 1] 为了解天然次生林保护对土壤活 性有机碳库的影响,采用空间替代时间研究方法,对湘中丘陵区不同演替阶段4种林分类型(杉木人工林、马尾松+石栎针阔混交林、南酸枣落叶阔叶林、青冈+石 栎常绿阔叶林)土壤活性有机碳及其与土壤养分相关性进行研究。结果表明:1)各土层总有机碳(TOC)、微生物生物量碳(MBC)、水溶性有机碳 (DOC)、易氧化有机碳(EOC)含量均表现为:青冈+石栎常绿阔叶林南酸枣落叶阔叶林马尾松+石栎针阔混交林杉木人工林,在0—30cm土层,马尾 松+石栎针阔混交林、南酸枣落叶阔叶林、青冈+石栎常绿阔叶林TOC含量比杉木人工林分别高出13.40%、19.40%和29.91%,MBC含量分别 高出15.62%、32.89%和53.33%,DOC含量分别高出8.52%、8.75%和13.76%,EOC含量分别高出32.79%、 38.48%和78.30%;2)天然次生林各土层MBC占TOC的比率以南酸枣落叶阔叶林最高,青冈+石栎常绿阔叶林为其次,马尾松+石栎混交林最低, 均高于同一土层杉木人工林(除马尾松+石栎混交林15—30cm土层外),天然次生林各土层DOC占TOC的比率随着演替进展而下降,均低于同一土层杉木 人工林(除马尾松+石栎混交林0—15cm土层外),天然次生林各土层EOC占TOC的比率随着演替进展而增加,且均高于同一土层杉木人工林;3)土壤 MBC、DOC、EOC含量与TOC含量的相关性均达到极显著水平,且天然次生林土壤MBC、DOC、EOC含量与TOC含量的相关系数随着演替进展而增 高,均高于杉木人工林;4)4种林分土壤TOC、MBC、DOC、EOC含量与土壤全N、碱解N、全P、有效P、全K、速效K含量之间的相关性均达到显著 或极显著水平。
WenQX ( 1984). The Determination Method of Soil Organic Matter. Chinese Agriculture Press, Beijing. 273-284. [本文引用: 1]
WuJG, ZhangXQ, WangYH, XuDY ( 2002). The effects of land use changes on the distribution of soil organic carbon in physical fraction of soil Scientia Silvae Science, 38(4), 19-29. DOI:10.11707/j.1001-7488.20020404URLMagsci [本文引用: 1] <p>根据宁夏固原地区近25a的历史资料,本文将土地利用类型分为:天然次生林(山杨、辽东栎、灌木)、次生林砍伐后形成的牧草地、农田及农田或牧草地形成的人工林(13a、18a、25a华北落叶松) ,并应用土壤有机质物理分组方法对这些类型0~110cm土层土壤物理组分中的有机碳(轻组有机碳与颗粒有机碳)含量、分配比例及非保护性有机碳分配比例进行了研究。结果表明:轻组有机碳分配比例,几种天然次生林比人工林、牧草地和农田都显著低,而人工林、牧草地与农田之间的差异却不显著;轻组有机碳含量,农田和牧草地比天然次生林分别低46%和14%以上,人工林比农田和牧草地分别高100%和27%以上;颗粒有机碳分配比例,天然次生林比人工林和农田低,牧草地与天然次生林的差异不显著;颗粒有机碳含量方面,农田和牧草地比天然次生林分别低38%和25%以上,而人工林比农田和牧草地分别高79%和47%以上;非保护性库中有机碳分配比例方面,天然次生林比人工林、农田低。从以上结果可以看出,天然林变成农田使土壤有机碳稳定性降低,而变成牧草地则影响较小。在农田中造林,对土壤有机碳的稳定性影响较小,而在牧草地中造林则使土壤有机碳稳定性降低。土地利用变化主要对0~50cm土层内的土壤物理组分有机碳产生影响,其含量及分配比例总体上随土层深度增加而降低,并在土壤剖面中分布表现为表层组(0~10cm)、主根系层组、次根系层组、根系底层组(90cm以下)几个土壤层次,各个土壤层组内差异较小,层间差异较大。</p> [ 吴建国, 张小全, 王彦辉, 徐德应 ( 2002). 土地利用变化对土壤物理组分中有机碳分配的影响 林业科学, 38(4), 19-29.] DOI:10.11707/j.1001-7488.20020404URLMagsci [本文引用: 1] <p>根据宁夏固原地区近25a的历史资料,本文将土地利用类型分为:天然次生林(山杨、辽东栎、灌木)、次生林砍伐后形成的牧草地、农田及农田或牧草地形成的人工林(13a、18a、25a华北落叶松) ,并应用土壤有机质物理分组方法对这些类型0~110cm土层土壤物理组分中的有机碳(轻组有机碳与颗粒有机碳)含量、分配比例及非保护性有机碳分配比例进行了研究。结果表明:轻组有机碳分配比例,几种天然次生林比人工林、牧草地和农田都显著低,而人工林、牧草地与农田之间的差异却不显著;轻组有机碳含量,农田和牧草地比天然次生林分别低46%和14%以上,人工林比农田和牧草地分别高100%和27%以上;颗粒有机碳分配比例,天然次生林比人工林和农田低,牧草地与天然次生林的差异不显著;颗粒有机碳含量方面,农田和牧草地比天然次生林分别低38%和25%以上,而人工林比农田和牧草地分别高79%和47%以上;非保护性库中有机碳分配比例方面,天然次生林比人工林、农田低。从以上结果可以看出,天然林变成农田使土壤有机碳稳定性降低,而变成牧草地则影响较小。在农田中造林,对土壤有机碳的稳定性影响较小,而在牧草地中造林则使土壤有机碳稳定性降低。土地利用变化主要对0~50cm土层内的土壤物理组分有机碳产生影响,其含量及分配比例总体上随土层深度增加而降低,并在土壤剖面中分布表现为表层组(0~10cm)、主根系层组、次根系层组、根系底层组(90cm以下)几个土壤层次,各个土壤层组内差异较小,层间差异较大。</p>
XuX, ShiZ, LiDJ, ReyA, RuanHH, CraineJ, LiangJY, ZhouJZ, LuoYQ ( 2016). Soil properties control decomposition of soil organic carbon: Results from data-assimilation analysis Geoderma, 262, 235-242. DOI:10.1016/j.geoderma.2015.08.038URL [本文引用: 1] Soil properties, such as clay content, are hypothesized to control decomposition of soil organic carbon (SOC). However, these hypotheses of soil property-C decomposition relationships have not been explicitly tested at large spatial scales. Here, we used a data-assimilation approach to evaluate the roles of soil properties and environmental factors in regulating decomposition of SOC. A three-pool (active, slow, and passive) C-cycling model was optimally fitted with 376 published laboratory incubation data from soils acquired from 73 sites with mean annual temperature ranging from 610215 to 2602°C. Our results showed that soil physical and chemical properties regulated decomposition rates of the active and the slow C pools. Decomposition rates were lower for soils with high clay content, high field water holding capacity (WHC), and high C:N ratio. Multifactor regression and structural equation modeling (SEM) analyses showed that clay content was the most important variable in regulating decomposition of SOC. In contrast to the active and slow C pools, soil properties or environmental factors had little effect on the decomposition of the passive C pool. Our results show inverse soil property-C decomposition relationships and quantitatively evaluate the essential roles of soil texture (clay content) in controlling decomposition of SOC at a large spatial scale. The results may help model development and projection of changes in terrestrial C sequestration in the future.
YanYP, CaoJH, LiangY, WangP, WangH ( 2012). Organic carbon storage and decomposition rate of three subcategories of calcareous soil in karst area Earth and Environment, 40(2), 219-226. URL [本文引用: 1] 应用土壤培养法,比较分析了桂林毛村典型岩溶区黑色石灰土、棕色 石灰土、红色石灰土三种亚类石灰土在25℃和70%田间饱和含水量条件下培养90d有机碳矿化速率的差异。结果显示:各亚类土壤有机碳矿化速率和累积释放 CO2-C量总体上都随土层加深而递减。0~20cm至20~40cm层递减幅度最大。各亚类石灰土有机碳矿化速率和累计释放量的大小顺序为:黑色石灰 土〉棕色石灰土〉红色石灰土,黑色石灰土的矿化速率远远大于棕色石灰土和红色石灰土,其中0~20cm土层差异最大。土壤有机碳矿化速率和有机碳含量呈正 相关。黑色石灰土土壤有机碳矿化释放的CO2-C分配比例最高,达到3.33%,其次是红色石灰土,比例为2.92%,旱地棕色石灰土矿化比例最低,为 1.90%,说明桂林毛村典型岩溶区黑色石灰土和红色石灰土有机碳稳定性较弱,旱地棕色石灰土具有较强的固定有机碳能力。 [ 严毅萍, 曹建华, 梁毅, 王培, 王宏 ( 2012). 桂林毛村岩溶区三种亚类石灰土有机碳矿化研究 地球与环境, 40(2), 219-226.] URL [本文引用: 1] 应用土壤培养法,比较分析了桂林毛村典型岩溶区黑色石灰土、棕色 石灰土、红色石灰土三种亚类石灰土在25℃和70%田间饱和含水量条件下培养90d有机碳矿化速率的差异。结果显示:各亚类土壤有机碳矿化速率和累积释放 CO2-C量总体上都随土层加深而递减。0~20cm至20~40cm层递减幅度最大。各亚类石灰土有机碳矿化速率和累计释放量的大小顺序为:黑色石灰 土〉棕色石灰土〉红色石灰土,黑色石灰土的矿化速率远远大于棕色石灰土和红色石灰土,其中0~20cm土层差异最大。土壤有机碳矿化速率和有机碳含量呈正 相关。黑色石灰土土壤有机碳矿化释放的CO2-C分配比例最高,达到3.33%,其次是红色石灰土,比例为2.92%,旱地棕色石灰土矿化比例最低,为 1.90%,说明桂林毛村典型岩溶区黑色石灰土和红色石灰土有机碳稳定性较弱,旱地棕色石灰土具有较强的固定有机碳能力。
ZhangH, LüMK, JiangJ, PuXT, WangEX, QiuX, XieJS ( 2016). Effects of vegetation restoration on topsoil and subsoil organic carbon mineralization in red soil erosion area Journal of Soil and Water Conservation, 30(1), 244-249. [本文引用: 4]
ZhangL, ZhangDL, MaoZJ ( 2017). Characteristic mineralization of soil organic carbon in different successional series of broadleaved Korean pine forests in the temperate zone in China, Acta Ecology Sinica, 37, 6370-6378. DOI:10.5846/stxb201607111415URL [本文引用: 3] 土壤有机碳矿化与陆地生态系统碳循环和全球气候变化关系密切,为准确评估中国温带小兴安岭阔叶红松林不同演替系列土壤有机碳矿化特征及变化规律.以年代序列法代替群落次生演替过程,采用室内恒温培养(碱液吸收法)测定阔叶红松林不同演替系列(中生演替系列、湿生演替系列、旱生演替系列)6种群落类型土壤有机碳矿化量和矿化速率.3个演替系列土壤有机碳含量均表现出一致的剖面变化特征,随着土层深度的加深有机碳矿化量逐渐降少.且不同演替系列土层间有机碳矿化量不同,中生演替系列原始阔叶红松林土壤有机碳累计矿化量最大,其次为旱生演替系列,湿生演替系列最小.3个演替系列土壤有机碳矿化速率随时间变化呈现基本一致的趋势,即培养前期快速下降、后期逐渐趋于平稳.3个演替系列6种群落类型土壤有机碳矿化差异显著,表现为原始阔叶红松林>白桦次生林>云冷杉红松林>红松枫桦次生林>蒙古栎红松林>蒙古栎、黑桦次生林.阔叶红松林不同演替系列土壤有机矿化采用非线性指数拟合效果较好.阔叶红松林不同演替系列土壤有机碳矿化与土壤全氮、凋落物量显著正相关,与土壤含水率、容重、土壤酸碱度显著负相关.不同演替系列群落的演替历史、土壤质地和养分状况等生态因子是导致阔叶红松林不同演替系列土壤有机碳矿化差异的原因. [ 张玲, 张东来, 毛子军 ( 2017). 中国温带阔叶红松林不同演替系列土壤有机碳矿化特征 生态学报, 37, 6370-6378.] DOI:10.5846/stxb201607111415URL [本文引用: 3] 土壤有机碳矿化与陆地生态系统碳循环和全球气候变化关系密切,为准确评估中国温带小兴安岭阔叶红松林不同演替系列土壤有机碳矿化特征及变化规律.以年代序列法代替群落次生演替过程,采用室内恒温培养(碱液吸收法)测定阔叶红松林不同演替系列(中生演替系列、湿生演替系列、旱生演替系列)6种群落类型土壤有机碳矿化量和矿化速率.3个演替系列土壤有机碳含量均表现出一致的剖面变化特征,随着土层深度的加深有机碳矿化量逐渐降少.且不同演替系列土层间有机碳矿化量不同,中生演替系列原始阔叶红松林土壤有机碳累计矿化量最大,其次为旱生演替系列,湿生演替系列最小.3个演替系列土壤有机碳矿化速率随时间变化呈现基本一致的趋势,即培养前期快速下降、后期逐渐趋于平稳.3个演替系列6种群落类型土壤有机碳矿化差异显著,表现为原始阔叶红松林>白桦次生林>云冷杉红松林>红松枫桦次生林>蒙古栎红松林>蒙古栎、黑桦次生林.阔叶红松林不同演替系列土壤有机矿化采用非线性指数拟合效果较好.阔叶红松林不同演替系列土壤有机碳矿化与土壤全氮、凋落物量显著正相关,与土壤含水率、容重、土壤酸碱度显著负相关.不同演替系列群落的演替历史、土壤质地和养分状况等生态因子是导致阔叶红松林不同演替系列土壤有机碳矿化差异的原因.
ZhaoM, ZhouJ, KalbitzbK ( 2008). Carbon mineralization and properties of water-extractable organic carbon in soils of the south Loess Plateau in China European Journal of Soil Biology, 44, 158-165. DOI:10.1016/j.ejsobi.2007.09.007URL [本文引用: 1] C02d) and the highest C content (2.3%) of the studied soils. In the soils exposed to intensive crop production without regular addition of organic manure we found the largest C mineralization (0.8502mg02C02g02C02d) and the lowest contents of organic C (0.9%). Addition of organic manure over a time scale of millennia resulted in high organic C contents (1.8%) and small C mineralization (0.5502mg02C02g02C02d). The content of WEOC reflected differences in C mineralization between the soils quite well and the two variables correlated significantly. Water-extractable organic C decreased during C mineralization from the soil illustrating its mainly labile character. Carbon mineralization from soils was particularly large in soils with small specific UV absorbance of WEOC. We conclude that amounts and properties of WEOC reflected differences in the stability of soil organic C. Both afforestation of arable land and the long-term addition of organic manure may contribute to C accumulation and stabilization in these soils.
ZhouYY, JiaXH, ZhaoX, ChenGX, GaoYH, WuP ( 2011). Soil carbon mineralization potential under different configuration of vegetation Chinese Journal of Ecology, 30, 2442-2448. URL [本文引用: 2] 通过实验室培养的方法,研究了沙坡头地区不同植被配置区(纯柠条、纯油蒿、柠条油蒿混交林)土壤碳矿化潜力及不同凋落物在土壤中的分解。结果表明,通过103d的室内培养,相同配置不同处理的土壤碳矿化差异不显著,灌丛密度高的植被配置碳矿化量高。不同生境土壤CO2-C的释放在灌丛下不加凋落物处理的情况下,单行油蒿与双行柠条样地间碳矿化存在显著性差异(P=0.047),其他各样地及各处理差异均不显著。总的来说,柠条样地土壤碳矿化潜力高于其他2种样地。凋落物分解速率在不同生境土壤最初的分解速率均为1年生草本混合样油蒿柠条,在培养40d之后,3种凋落物的分解速率趋于一致,这与草本较高的C、N、C/N比有关。另外,柠条样地的土壤加油蒿凋落物比在柠条样地的土壤中加柠条凋落物有较高的初始碳矿化潜力,这除与油蒿凋落物较易分解有关外还与不同生境土壤养分的可利用性有关,但在培养过程中出现拐点,这与柠条较高的C、N、C/N有关。在不同生境的土壤中,碳矿化潜力均为灌丛下灌丛外,灌木的存在使更多的有机质和养分积聚在灌丛下,形成灌丛肥岛,对碳的固存具有显著贡献。 [ 周玉燕, 贾晓红, 赵昕, 陈国雄, 高艳红, 吴攀 ( 2011). 不同植被配置下土壤碳矿化潜力 生态学杂志, 30, 2442-2448.] URL [本文引用: 2] 通过实验室培养的方法,研究了沙坡头地区不同植被配置区(纯柠条、纯油蒿、柠条油蒿混交林)土壤碳矿化潜力及不同凋落物在土壤中的分解。结果表明,通过103d的室内培养,相同配置不同处理的土壤碳矿化差异不显著,灌丛密度高的植被配置碳矿化量高。不同生境土壤CO2-C的释放在灌丛下不加凋落物处理的情况下,单行油蒿与双行柠条样地间碳矿化存在显著性差异(P=0.047),其他各样地及各处理差异均不显著。总的来说,柠条样地土壤碳矿化潜力高于其他2种样地。凋落物分解速率在不同生境土壤最初的分解速率均为1年生草本混合样油蒿柠条,在培养40d之后,3种凋落物的分解速率趋于一致,这与草本较高的C、N、C/N比有关。另外,柠条样地的土壤加油蒿凋落物比在柠条样地的土壤中加柠条凋落物有较高的初始碳矿化潜力,这除与油蒿凋落物较易分解有关外还与不同生境土壤养分的可利用性有关,但在培养过程中出现拐点,这与柠条较高的C、N、C/N有关。在不同生境的土壤中,碳矿化潜力均为灌丛下灌丛外,灌木的存在使更多的有机质和养分积聚在灌丛下,形成灌丛肥岛,对碳的固存具有显著贡献。
ZhuRH, ZhengZC, LiTX, ZhangXZ, HeSQ, WangYD, LiuT, LiW ( 2017). Dynamics of soil organic carbon mineralization in tea plantations converted from farmland at Western Sichuan, China PLOS ONE, 12(9), e0185271. DOI: 10.1371/journal.pone.0185271. DOI:10.1371/journal.pone.0185271URLPMID:5607202 [本文引用: 2] Climate warming and land use change are some of the drivers affecting soil organic carbon (SOC) dynamics. The Grain for Green Project, local natural resources, and geographical conditions have resulted in farmland conversion into tea plantations in the hilly region of Western Sichuan. However, the effect of such land conversion on SOC mineralization remains unknown. In order to understand the temperature sensitivity of SOC decomposition in tea plantations converted from farmland, this study considered the different years (i.e., 2–3, 9–10, and 16–17 years) of tea plantations converted from farmland as the study site, and soil was incubated for 28 days at 15°C, 25°C, and 35°C to measure the soil respiration rate, amount, and temperature coefficient (Q10). Temperature and land use type interactively affected the SOC mineralization rate, and the cumulative amount of SOC mineralization in all the plots was the largest at 35°C. SOC mineralization was greater and more sensitive to temperature changes in the farmland than in the tea plantations. Compared with the control, tea plantation soils showed lower SOC mineralization rate and cumulative mineralization amount. The 16–17-year-old tea plantation with a low SOC mineralization amount and high SOC content revealed the benefits of carbon sequestration enhancement obtained by converting farmland into tea plantations. The first-order kinetic equation described SOC mineralization dynamics well. Farmland conversion into tea plantations appeared to reduce the potentially mineralizable carbon pool, and the age of tea plantations also had an effect on the SOC mineralization and sequestration. The relatively weak SOC mineralization temperature sensitivity of the tea plantation soils suggested that the SOC pool of the tea plantation soils was less vulnerable to warming than that of the control soils. Widespread coupling between the rate and temperature sensitivity of organic matter decay 1 2010
... 土壤有机碳(SOC)矿化是土壤碳(C)输出的主要途径, 是大气中CO2的重要来源, 在维持全球C平衡中具有极其重要的作用(Saggar et al., 2001; Craine et al., 2010).SOC矿化不仅反映SOC的稳定性和周转(周玉燕等, 2011; 黄宗胜等, 2012), 而且关系到土壤养分元素的释放与供应.因此, 研究SOC矿化特征及其影响因素是陆地生态系统C循环的研究热点, 有助于揭示SOC释放的驱动机制及其稳定性, 为深入研究土壤固C能力的影响机制和估算土壤C平衡提供基础数据, 而且对科学管理土壤养分, 有效缓解全球气候变暖具有重要意义(Zhu et al., 2017). ...
Changes in soil carbon sequestration and soil respiration following afforestation on paddy fields in north subtropical China 1 2013
Microbial community structure and characteristics of the organic matter in soils under Pinus sylvestris, Picea abies and Betula pendula at two forest sites 1 2001
Hydrothermal carbonization (HTC): Near infrared spectroscopy and partial least-squares regression for determination of selective components in HTC solid and liquid products derived from maize silage 1 2014
... LC、LN、LP含量的测定详见辜翔等(2018)的文献, LL含量测定采用Van-Soest法(Reza et al., 2014)测定.每个分析样品平行测定2次, 取平均值作为该样品的最终测定结果.4种植被恢复阶段凋落物层养分特征如表1所示. ...
Carbon mineralization kinetics as in?uenced by soil properties 1 1996
Cultivation effects on soil biological properties, microfauna and organic matter dynamics in Eutric Gleysol and Gleyic Luvisol soils in New Zealand 1 2001
... 土壤有机碳(SOC)矿化是土壤碳(C)输出的主要途径, 是大气中CO2的重要来源, 在维持全球C平衡中具有极其重要的作用(Saggar et al., 2001; Craine et al., 2010).SOC矿化不仅反映SOC的稳定性和周转(周玉燕等, 2011; 黄宗胜等, 2012), 而且关系到土壤养分元素的释放与供应.因此, 研究SOC矿化特征及其影响因素是陆地生态系统C循环的研究热点, 有助于揭示SOC释放的驱动机制及其稳定性, 为深入研究土壤固C能力的影响机制和估算土壤C平衡提供基础数据, 而且对科学管理土壤养分, 有效缓解全球气候变暖具有重要意义(Zhu et al., 2017). ...
Plant carbon inputs and environmental factors strongly affect soil respiration in a subtropical forest of southwestern China 1 2009
 ,
, ,1,2,3,*
,1,2,3,*
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT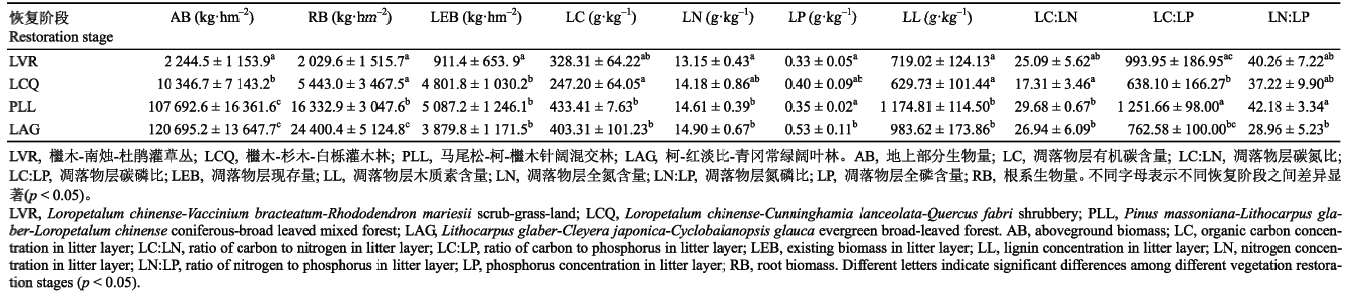
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT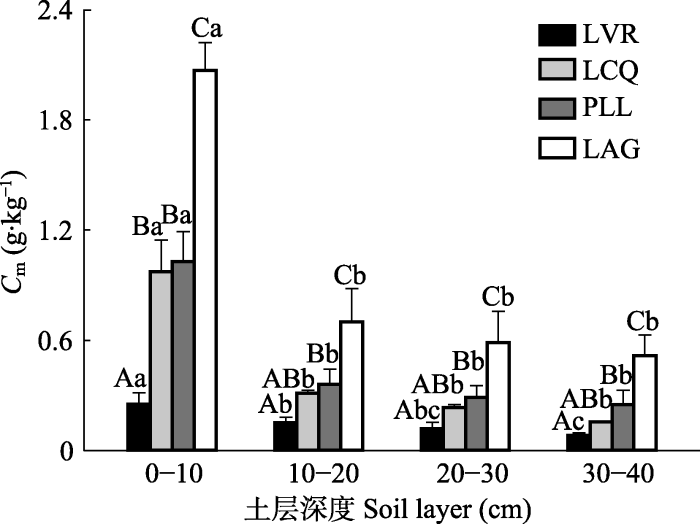 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT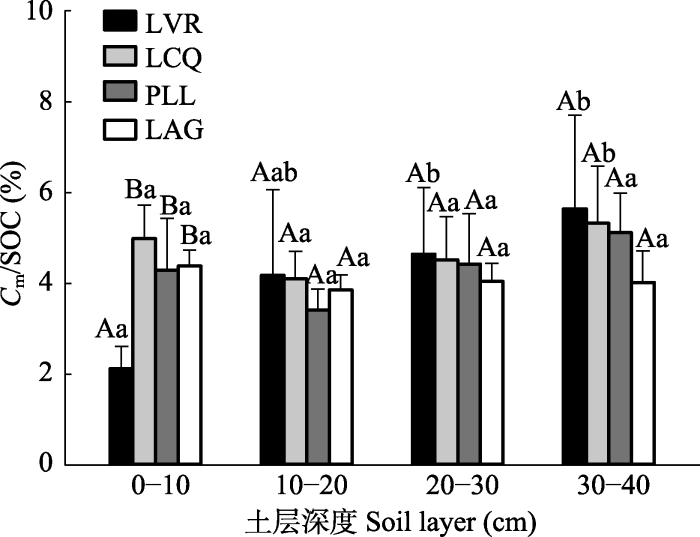 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}