 ,1,*, 邸楠1,*, 曹治国1,4,*, 刘金强1, 李豆豆1, 王烨1,2, 李广德3, 段劼1, 贾黎明1, 张瑞娜1
,1,*, 邸楠1,*, 曹治国1,4,*, 刘金强1, 李豆豆1, 王烨1,2, 李广德3, 段劼1, 贾黎明1, 张瑞娜1Characteristics and underlying mechanisms of plant deep soil water uptake and utilization: Implication for the cultivation of plantation trees
XI Ben-Ye,1,*, DI Nan1,*, CAO Zhi-Guo1,4,*, LIU Jin-Qiang1, LI Dou-Dou1, WANG Ye1,2, LI Guang-De3, DUAN Jie1, JIA Li-Ming1, ZHANG Rui-Na1通讯作者: E-mail:benyexi@bjfu.edu.cn
编委: 杜 盛
责任编辑: 李 敏
收稿日期:2018-04-11修回日期:2018-09-10网络出版日期:2018-09-20
| 基金资助: |
Received:2018-04-11Revised:2018-09-10Online:2018-09-20
| Fund supported: |

摘要
根系吸水是树木水分关系的重要环节, 在树木生理活动中发挥着至关重要的作用。深层土壤中的水资源含量一般相对较高, 常可为树木生长供给大量水分, 并在旱季保障其生存与正常生长。因此, 了解树木对深层土壤水的吸收利用特征与机制, 可帮助深入认识树木与环境的互作机制、树木的生长与生存策略、物种间的共存与竞争机制等内容, 同时还可帮助构建既能降低外部水资源投入, 又能避免水分生态环境负面效应的人工林绿色栽培制度。基于已有研究, 该文对树木吸收利用深层土壤水的特征与机制进行了综述。首先, 探讨了深层根系和深层土壤的界定, 指出对于除寒温带针叶林以外的其他主要森林植被类型, 可以1 m作为树木深根系和深土层的平均划分(参考)标准, 并明确了全球范围内树木深根系的成因。其次, 对已有研究中观察到的树木对深层土壤水的吸收利用特征及其影响因素进行了归纳与总结, 并从深根系性状调节、整株水力特性协调两方面探讨了树木高效吸收利用深层土壤水的机制, 如可通过深根系的空间、时间和效率调节策略来促进对深土层水分的吸收。最后, 提出了树木利用深土层水分对人工林培育的几点启示, 包括水分管理.中应使林木适度利用深层土壤水, 选用合适的灌水频率、合理的树种混交能促进深层土壤水分储库“缓冲”作用的发挥, 基于树木土壤水分利用深度的间伐木选择技术等, 并指出了该领域现有研究的不足以及今后的发展方向。
关键词:
Abstract
Root water uptake is an essential part of tree water relations and plays a crucial role in tree physiological activities. Water resource in deep soil is relatively abundant and can provide plenty of water to trees to guarantee their survival and healthy growth during dry seasons. Thus, a good comprehension of the characteristics and underlying mechanisms of deep soil water uptake and utilization by trees will deepen the understanding of the interaction between trees and the environment, tree survival and growth strategies, coexistence and competition among different species, etc. This knowledge is important in establishing green cultivation schemes for plantations, which depend less on the external water resources input and avoid the adverse effects on the water ecological environment. From existing studies, the characteristics and underlying mechanisms of deep water uptake and utilization by trees are reviewed. Firstly, the definition of deep roots and deep soil is discussed, and 1 m depth is recommended as the average (reference) definition standard in main forest vegetation types except the boreal forest. The reasons for the formation of deep tree roots around the globe were also determined. Secondly, the observed deep soil water uptake characteristics of trees and their influencing factors are summarized. Then, from the aspects of the adjustment of deep root traits and the coordination of hydraulic traits of different organs, the mechanisms of deep water uptake by trees are discussed. For example, the spatial, temporal and efficiency adjustment strategies of deep roots can be used to facilitate the absorption of deep soil water. Finally, some implications of deep soil water uptake for the cultivation of plantations are proposed, such as “for water management in plantations, trees should be induced to moderately utilize some deep soil water and an appropriate irrigation frequency should be selected”, “appropriate mixed planting of different tree species can facilitate the buffering effect of deep soil water storage”, “developing techniques of selecting trees for thinning based on the water uptake depths of different species”, etc. Deficiencies of existing studies and some future research directions were also pointed out.
Keywords:
PDF (1323KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
席本野, 邸楠, 曹治国, 刘金强, 李豆豆, 王烨, 李广德, 段劼, 贾黎明, 张瑞娜. 树木吸收利用深层土壤水的特征与机制: 对人工林培育的启示. 植物生态学报[J], 2018, 42(9): 885-905 DOI:10.17521/cjpe.2018.0083
XI Ben-Ye, DI Nan, CAO Zhi-Guo, LIU Jin-Qiang, LI Dou-Dou, WANG Ye, LI Guang-De, DUAN Jie, JIA Li-Ming, ZHANG Rui-Na.
我国木材资源对外依存度超过50%, 木材安全问题严重, 营造和培育速生丰产林缓解木材短缺势在必行。目前, 我国人工林面积已达6 933万hm2, 位居世界首位, 但人工林的质量和产量还较低, 远不能满足我国日益增长的木材需求, 亟须采取高效的定向集约培育措施来促进人工林生长。水分是树木生长的重要限制因子(Xi et al., 2016), 因此, 系统了解树木和水分的关系, 有助于制定出既能快速提高林木生长又能使水分利用效率最大化的高效集约栽培措施, 同时还能避免对水生态环境造成负面影响。
根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径。因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(Nepstad et al., 1994; Barbeta & Pe?uelas, 2017; Qi et al., 2018)。此外, 了解树木对深层土壤水的吸收利用特征还可帮助深入认识树木对干旱和气候变化的响应与适应机制(Nepstad et al., 1994; Broedel et al., 2017; Christina et al., 2017)、树木与环境的互作机制(Nepstad et al., 1994; 刘晓丽等, 2014)、树木的生长与生存策略(Xu & Li, 2008; Pinheiro et al., 2016; Zunzunegui et al., 2017)、物种间的共存与竞争机制(Barron-Gafford et al., 2017; Chitra-Tarak et al., 2018; de Deurwaerder et al., 2018; Guderle et al., 2018), 并进而为林地水分管理策略的制定(Adriano et al., 2017; Christina et al., 2017)、植被恢复(陈亚宁等, 2016; Sun et al., 2018)、生物多样性保护(Lindh et al., 2014)、林分结构配置与优化(Germon et al., 2017; 苗博等, 2017)、林木遗传育种(Pinheiro et al., 2016)、现有森林抚育措施优化(如间伐木的选择等)(Hardanto et al., 2017)、林地水文模型的优化和构建(Pinheiro et al., 2016; Fan et al., 2017)等提供依据。因此, 随着同位素、根系液流监测、土壤水分测定等技术的发展, 树木对深层土壤水分吸收利用的研究在近年来逐渐被广泛关注, 有必要对相关研究进行系统归纳与总结。
深层根系是植物借以吸收深层土壤水的唯一工具, 其在树木水分关系中的重要性早在近一个世纪以前就被人们所关注(Hayes & Stoeckler, 1935), 其对深层土壤养分吸收、土壤物理化学风化、碳地球化学循环、土壤动物和微生物群落等的影响也逐渐被了解(McCulley et al., 2004; Maeght et al., 2013; Billings et al., 2018)。但受限于研究工具和方法, 同时由于大家长期以来比较关注浅土层中的生物与非生物过程, 认为只通过研究浅土层中的根系即可对生态系统水平上根系系统的结构和功能作用有较好的了解, 而且浅土层中的各类数据资料较多亦较容易获取, 因此, 一直以来关于树木浅层根系的研究较多, 而对深层根系的研究则相对偏少(Stone & Kalisz, 1991; Canadell et al., 1996; Pierret et al., 2016)。Maeght等(2013)和Pierret等(2016)曾对深根系的研究方法、重要性及其功能进行了详细阐述, 但关于树木对深层土壤水的吸收利用特征与机制, 目前还未有对相关工作的系统综述。
因此, 本文拟依托现有研究, 对树木吸收利用深层土壤水的特征与机制进行综述。首先, 探讨深根系和深层土壤的界定, 并明确全球范围内树木深根系的成因。其次, 就现有研究中所观察到的树木对深土层水分的吸收利用特征及其影响因素进行归纳与总结, 并从深根系性状调节、整株水力特性协调两方面探讨树木对深层土壤水的高效吸收利用机制。最后, 基于上述总结, 探讨树木利用深层土壤水分的特征对人工林培育的启示, 并指出现有研究的不足之处和今后的发展方向。
1 深根系和深层土壤的定义
了解树木对深层土壤水的吸收利用特征, 需首先对深层土壤进行界定, 但深层土壤并非严格的科学概念, 而且全球不同地区土壤厚度差异很大(如土石山区与黄土高原)(Shangguan et al., 2017), 所以不同研究、不同领域对其界定不同, 无统一、明确的标准。已有的研究中深土层的界定通常比较经验化, 缺乏严格依据, 如林业上一般将50或60 cm以下定为深土层; 何春霞等(2016)在华北低丘山区将30-80 cm土层归为深层土壤; Brum等(2017)将巴西中部热带雨林中60-90 cm土层划为深土层; 袁国富等(2012)在黑河中游风沙侵蚀区的多枝柽柳(Tamarix ramosissima)人工林中则将2.7 m以下土层定为深土层。还有一部分研究是根据降水的入渗深度来进行定义, 如刘晓丽等(2013, 2014)根据降水入渗深度, 将黄土丘陵区密植枣林中2 m以下的土层划为深层土壤, 因为该土层不受降水影响, 只受根系吸水作用的影响。土壤是植物生长的介质, 故而深层土壤应是相对于植物而定义的概念, Schenk和Jackson (2005)也曾提出“深层土壤出现的地区”可类同于“深根系出现的地区”。因此, 从植物研究的角度考虑, 一种较合理的定义方法是将深根系分布的土层定义为深土层, 但这种方法需首先定义植物的深根系。然而, 自然界中的物种多种多样, 植株个体大小也在不断变化, 因此准确定义深根系非常困难, 长期以来难以达成共识(Maeght et al., 2013; Pierret et al., 2016)。Schenk和Jackson (2005)提出, 一个土壤剖面的2 m以下土层中如果包含有大量(至少5%)的根系, 则可认为该剖面具有深根系特征, 但并未明确给出深根系的定义和划分标准。基于全球475个根系剖面数据, Schenk和Jackson (2002a)估算得到15个陆地植被类型累积95%根系的平均分布深度(d95)为102 cm (29- 171 cm)(表1), 同时也发现, 所有根系剖面的取样深度中值为0.88 m。基于该结果, Maeght等(2013)首次明确提出: 一般而言, 可将生长在至少1 m以下土层中的根系定义为深层根系。目前, 关于1 m以下土层中根系动态的研究还很薄弱(Germon et al., 2016), 大家对其知之甚少; 然而, 这部分根系却具有非常重要的生态功能, 如从全球尺度来讲, 如果根系能够吸收1 m以下土层中的水分, 则植被的光合和蒸腾速率能分别提高16%和18% (Kleidon & Heimann, 1998); 因此, 从这个角度来看, 将1 m以下土层中的根系定义为深层根系具备一定合理性。Pierret等(2016)认为该定义虽过于简化, 但考虑到其简单、实用, 且兼顾了根系功能的生态意义, 因此建议仍以1 m为界来定义深根系。该划分标准在很多研究中也得到认可和应用(McCulley et al., 2004; Oliveira et al., 2005; Yang et al., 2017; Zunzunegui et al., 2017; Qi et al., 2018)。Fan等(2017)对全球不同气候、土壤和排水条件下超过1 000个植物物种的2 200个根系研究数据进行了分析, 发现目前观测到的根系最大分布深度为1 m左右的情况最多, 该结果对上述深根系划分方法在全球尺度上、针对所有陆地植被类型的一般适用性提供了一定支撑。Pierret等(2016)曾提出如果根据累积一定比例根系(长度或生物量)的分布深度来划分深根系, 则更加合理。该方法虽能针对不同地区的不同植物分别确定深根系划分标准, 但由于需要获取完整的根系剖面数据资料(取样至根系分布下限), 而且植物的根系分布深度又处在不断变化之中, 所以应用起来也存在较大困难; 而且如果划分标准不同, 各地区和植物间的深根系数据也很难统一起来进行对比分析。
Table 1
表1
表1全球不同植被类型累积95%根系的分布深度(d95) (数据源自Schenk & Jackson, 2002a)
Table 1
| 植被类型 Vegetation type | d95 (cm)* |
|---|---|
| 苔原 Tundra | 29 |
| 寒温带针叶林 Boreal forest | 58 |
| 寒温带森林(含人工林) Cool-temperate forest (including plantations) | 104 |
| 暖温带森林(含人工林) Warm-temperate forest (including plantations) | 121 |
| 森林草甸 Meadows in the forest zone | 40 |
| 草原 Prairie | 91 |
| 半荒漠草地 Semi-desert steppe | 120 |
| 温带稀树草原 Temperate savanna | 140 |
| 地中海灌丛/林地 Mediterranean shrubland/woodland | 171 |
| 半荒漠灌丛 Semi-desert shrubland | 131 |
| 荒漠 Desert | 112 |
| 干旱热带稀树草原 Dry tropical savannas | 144 |
| 湿润热带稀树草原 Humid tropical savannas | 94 |
| 热带半落叶和落叶林 Tropical semi-deciduous and deciduous forest | 95 |
| 热带常绿林 Tropical evergreen forest | 91 |
新窗口打开|下载CSV
综上所述, 就所有陆地植被类型平均而言, 可将1 m以下土层中的根系定义为深根系, 而该土层即为深土层。虽然其在特定气候、土壤和植被类型下的适用性有限, 但基于d95数据(表1; Schenk & Jackson, 2002a), 对于除寒温带针叶林以外的其他主要森林植被类型(寒温带森林(含人工林)、暖温带森林(含人工林)、热带半落叶和落叶林、热带常绿林), 仍建议以1 m作为树木深根系和深土层的平均划分(参考)标准, 以便于对树木的深根系研究进行标准化, 而且也有利于不同研究之间的横向对比。实际应用中, 如果这些森林植被类型所在地区(如山区)的土壤厚度小于1 m, 一般可认为树木在该地区较难产生深根系, 而且也不存在相对于树木而言的深土层。但需注意的是, 在很多地区, 即使土层较薄, 树木根系还是可以借助裂缝和缝隙直接生长至很深的风化或破碎的岩石层中, 吸收其中的水分(Schenk & Jackson, 2005; Fan et al., 2017; Rempe & Dietrich, 2018)。
2 树木深根系的成因
根系是树木吸收和传输土壤水分和养分资源的重要功能器官(Jackson et al. 1997; 王政权和郭大立, 2008; Ma et al., 2018), 其在深土层中的结构决定了树木对深层土壤资源的“觅取”范围和效率, 并进而对区域甚至全球水、碳、养分循环和气候产生重要影响(Nepstad et al., 1994; Jackson et al., 1997; Schenk & Jackson, 2002b; Lee et al., 2005; Fan et al., 2017)。然而, 深层根系虽然是一个全球普遍存在的重要植物功能性状(除了南极洲以外的其他大洲均有记载) (Canadell et al., 1996; Schenk & Jackson, 2005; Fan et al., 2017), 且能对树木功能的发挥产生重要影响, 但受限于研究方法和手段, 目前对深层根系及其功能与生态重要性的了解还十分有限(Pierret et al., 2016)。现有研究表明, 影响树木深根系产生的因子很多, 但归纳起来可分为植被因子(树种、生活型、植被类型、林龄、个体发育、物种竞争)、气候因子(降水量、潜在蒸散量)、土壤因子(土层厚度、土壤质地、土壤有机层厚度、土壤水分、土壤养分)和水文因子(地下水位), 且不同空间尺度上主要影响因子不同(Canadell et al., 1996; Schenk & Jackson, 2002a, 2002b, 2005; Fan et al., 2017)。
2.1 植被因子
树木根系的分布深度本质上由基因决定, 所以不同树种间的根系分布深度差异很大。例如, 对于金合欢属(Acacia, (12.85 ± 17.99) m)、桉属(Eucalyptus, (8.71 ± 8.75) m)、栎属(Quercus, (5.23 ± 5.79) m)、杨属(Populus, (3.00 ± 4.75) m)、松属(Pinus, (2.45 ± 3.92) m)、刺柏属(Juniperus, (2.41 ± 2.40) m)、槭属(Acer, (1.75 ± 1.26) m)、柳属(Salix, (1.35 ± 1.53) m)、冷杉属(Abies, (1.30 ± 0.67) m)、黄杉属(Pseudotsuga, (1.03 ± 0.37) m)、落叶松属(Larix, (0.90 ± 0.67) m)、水青冈属(Fagus, (0.83 ± 0.46) m)和云杉属(Picea, (0.74 ± 0.47) m)等主要人工林树种, 目前观测到的其平均根系深度最大可相差16倍(Fan et al., 2017)。全球范围内, 不同生活型和植被类型间, 树木根系分布深度也存在很大差异。平均而言, 常绿阔叶树((6.30 ± 10.17) m) >落叶阔叶树((3.71 ± 6.99) m) >常绿针叶树((1.79 ± 2.79) m) >落叶针叶树((0.78 ± 0.57) m) (Fan et al., 2017); 热带常绿林((7.3 ± 2.8) m) >温带针叶林((3.9 ± 0.4) m) >热带落叶林((3.7 ± 0.5) m) >温带落叶林((2.9 ± 0.2) m) >寒温带针叶林((2.0 ± 0.3) m) (Canadell et al., 1996)。
植物根系系统的大小(分布深度、水平延伸距离)与其地上部分的大小间存在显著正相关关系(Schenk & Jackson, 2002b), 因此, 在个体尺度上, 不同林龄和发育程度下树木的根系分布深度存在明显差异。一般而言, 随林龄增加, 根系分布逐渐加深(Christina et al., 2011; Laclau et al., 2013; Christina et al., 2017)。树木根系的发展在很大程度上也受个体发育控制, 生长速度越快, 越可能发展出深根系。如Christina等(2011)在巨尾桉(Eucalyptus grandis)纯林中观察到林木向上(树高)和向下(根深)的生长速度几乎相近, 林木栽植1.5年后, 树高和根深分别达到10.4和9.2 m, 而3.5年之后, 则达19.2和15.8 m。
在林分等小尺度上, 物种竞争能明显影响根系的向下生长。如果种内(如密度增大、树木个体增大等造成)和种间(如农林间作、树种混交等造成)竞争增强, 一般会促进树木深根系的发生和发展。如, Xi等(2013)发现, 在宽窄行栽植模式下的毛白杨(Populus tomentosa)人工林中, 随距树距离增加, 林木种内地下竞争会逐渐减弱, 从而可能引起根系分布的逐渐变浅; Mulia和Dupraz (2006)在法国南部胡桃(Juglans regca)纯林中观察到林木累积90%根系的分布深度为0.87 m, 但其与农作物间作后则达2 m以下。
2.2 气候因子
现有研究显示, 在全球大尺度上, 年降水量(MAP)和年潜在蒸散量(PET)可能是影响树木根系分布的两个重要气候因子(Schenk & Jackson, 2002a, 2005), 但对不同的地区、植被类型以及根系分布表征指标(d95、根系最大深度(dmax)), 二者的作用规律会发生变化。对于温带森林和寒温带针叶林, MAP和PET越大, 根系分布越深(d95变大), 且不受旱季长度影响; 对于热带森林, PET越大, 旱季越长, 根系分布越深, 而MAP越大则根系越浅(d95变小)?(Schenk & Jackson, 2002a)。Schenk和Jackson (2002b)的统计数据显示, 在水分限制性生态系统中(MAP ≤ 1 000 mm, MAP:PET < 1), 树木dmax不受MAP的影响, 但会随PET减小有轻微的增加趋势。Fan等(2017)针对云杉属、松属、栎属、杨属、桉属和金合欢属树种的分析也发现, 树木dmax不受MAP影响。由此可见, 全球尺度上, MAP和PET能对根系分布深度产生明显影响, 但对根系最大深度dmax的决定作用较弱。基于Schenk和Jackson (2005)利用包含MAP和PET的植物深根系概率预测气候模型的预测结果, 全球树木深根系出现概率最高的地区, 可能为赤道潮湿热带雨林附近具有季节性热带气候的区域, 如中美洲、巴西东南部、非洲刚果盆地北部和南部、印度大部、东南亚和澳大利亚北部。出现概率最低的区域可能是从北极圈到寒温带, 以及干旱地区和赤道热带雨林地区。
2.3 土壤因子
全球尺度上, 土壤是仅次于气候的树木根系分布深度的另一重要影响因子(Schenk & Jackson, 2002a)。根系对土壤环境非常敏感, 所以在小尺度上, 根系分布深度也受到土壤因子的较大影响。但由于详细土壤信息的获取非常困难, 因此, 已有的研究主要分析了土壤厚度、土壤质地、土壤有机层厚度和土壤水分对树木根系分布深度的影响。土壤厚度是树木以及其他植物最大根系分布深度的重要影响因子, 但很多情况下其并不能制约根系的最终分布深度(Fan et al., 2017), 已有很多研究发现根系可以借助岩石层中的缝隙扎根很深, 以吸收其中的水分和养分(Barbeta & Pe?uelas, 2017; Hasenmueller et al., 2017; Rempe & Dietrich, 2018)。例如, Rempe和Dietrich (2018)在加利福尼亚北部海岸山脉上的针阔常绿混交林中发现, 虽然土壤厚度只有30-75 cm, 但树木却能扎根至风化岩石层中(深度达16 m), 来吸收其中较土壤水更加丰富的稳定岩石水(rock moisture); Barbeta和Pe?uelas (2017)基于已有文献的统计, 估算出岩石层中水分对植物蒸腾的平均贡献达到约41%。
一般而言, 土壤质地因影响水分入渗深度和毛管水上升高度而能对树木及其他植物根系分布深度产生影响(Fan et al., 2017)。从暖温带到热带, 与中等质地土壤相比, 粗质地和细质地土壤上出现深根系(d95 > 2 m)的概率更大(Schenk & Jackson, 2005)。例如, Xu和Li (2008)在新疆古尔班通沙漠南缘, 发现粉壤土上梭梭(Haloxyolon ammodendron)根系的深度为3 m, 但在相距8 km的壤砂土上则达到10 m。然而, 依据Fan等(2017)对多种植物共计1 426个根系样本数据的分析结果, 可以看出土壤质地在全球尺度上对树木最大根系分布深度的影响可能较小。这主要是因为较粗质地土壤上的水分入渗虽然较深, 但质地较细土壤上的毛管上升水高度却较大, 而这二者的作用均会诱导深根系的产生(Fan et al., 2017)。
当土壤有机层较厚时, 土壤养分充足, 一般而言, 树木可能不需扎根很深便可获取充足的土壤养分。现有数据表明, 在寒温带针叶林、温带和热带森林中, d95均会随着土壤有机层厚度的减小而变深(Schenk & Jackson, 2002a)。然而, 在巴西南部, Christina等(2011)发现, 虽然对巨尾桉纯林进行了足量施肥, 但林木在栽植18个月后根系仍能长至9 m深。由此表明, 表层土壤养分的相对亏缺或胁迫并不是树木深根系产生的决定性因素。与此相似, 也有研究发现, 在干旱和半干旱地区, 即使浅土层养分状况较好, 但如果深土层中存在丰富的养分资源时(如可吸收的P、Ca2+、Mg2+等), 也会促使树木发展出较深的根系来吸收这些深层养分元素(McCulley et al., 2004)。
浅土层干旱虽不会决定深根系的发生, 但会促进深根系的发展。如Holloway等(2017)在意大利以黑杨(Populus nigra)为优势种的河岸林中发现, 湿润区域根系的最大分布深度约为1.75 m, 但在较干旱地点则可达2.35 m。同样, 深土层中的水分含量也会对树木深根系的发展产生一定的促进作用。Schenk和Jackson (2005)预测, 在全球尺度上, 如果2 m以下土层中的年水分储量超过0.15 m·a-1, 则出现深根系分布(d95 > 2 m)的概率会显著增大。
2.4 水文因子
虽然气候因子能对全球尺度上树木根系的分布深度产生明显影响, 但相同气候下却常出现较浅与较深根系分布的同时存在。这种现象可能由树种、林龄、土壤质地、种间竞争等因素引起(Mulia & Dupraz, 2006; Xu & Li, 2008; Fan et al., 2017), 但这些因素却不能解释在相同气候和土壤质地下, 为何林龄相同或相近的同一树种的纯林间根系分布仍可能会出现极大差异(Wagg, 1967; Sprackling & Read, 1979)。Fan等(2017)认为这种现象主要是由地下水位的差异引起, 因为501组根系剖面和地下水位数据显示, 植物根系的最大分布深度与地下水位间存在较强的线性相关关系, 而且具体到多个属的树种上这种规律也表现明显。地下水位之所以对根系最大分布深度起较强的控制作用, 主要是因为缺氧会限制根系的生长。但也有研究发现, 当地下水中有较多的溶解氧和养分时, 根系也会突破地下水的限制, 在地下水位以下生存(Naumburg et al., 2005; Pierret & Lacombe, 2018)。此外, 如果地下水位存在较大的季节性波动, 也常会出现根系分布在地下水位以下的情况, 而且也会促进深根系的产生, 如Di等(2018)在山东黄泛平原地下水位平均年变化为48-411 cm的立地上, 发现毛白杨根系分布深度仍可达到2.7 m。由此可见, 地下水位对根系分布深度的控制作用可能主要仅限于地下水位较稳定或变幅不大的情形, 而对于地下水位存在剧烈季节性波动的情形(全球广泛存在), 根系最大分布深度的决定因子目前尚不明确(Xi et al., 2018)。
较大的土壤机械阻力(如土壤容重、土壤硬盘等造成)、较低的氧气浓度和水分含量会限制根系的向下生长; 在寒冷地区, 较低的土壤温度和永久冻土层也会成为根系垂向延伸的限制因素(Stone & Kalisz, 1991)。因此, 为避开这些不利因素, 同时为降低在构建和维持根系以及吸收水分和养分资源时的能量消耗, 根系会尽可能在浅土层生长(Schenk, 2008)。但当气候干旱、物种土壤资源竞争等造成浅土层水分资源的相对亏缺, 林龄增大、大气蒸发潜力增加等造成蒸腾作用增强, 树木依靠浅层土壤中的水分吸收已不能满足自身蒸腾需求时, 其根系便会尽可能地向深土层生长来吸收更多的深层水分(Schenk, 2008)。因此, 觅取水资源以尽量满足自身蒸腾需求是树木根系选择向深土层生长的主要原动力。同时, 值得注意的是, 对深土层可能存在的丰富营养元素的需求也会促使深层根系的产生与发展(McCulley et al., 2004)。
综上所述, 基因决定了树木根系的潜在分布深度, 而气候(降水)、土壤(质地、土层厚度)、植被(物种地下竞争)和水文(地下水位)因子共同塑造的土壤水分剖面特征, 以及植被(林龄、个体发育)和气候(潜在蒸散量)因子共同影响的树木潜在蒸腾需求, 则主要决定了全球不同地区树木根系的实际分布深度。
全球范围内, 整体而言, 当地下水位较深, 远远超出根系可达范围时, 在干旱气候区, 降水较少且入渗较浅, 受降水入渗深度的限制, 根系分布也较浅; 在过湿气候区, 降水多而频繁, 浅土层始终持有充足水分, 树木不需产生深根便可获取足够的蒸腾用水, 因此根系分布也相对较浅; 但在季节性干旱气候区, 湿润季节较深的降水入渗会诱导深土层根系的增殖, 促进形成相对较深的根系分布。当地下水位相对较浅时, 在过湿气候区, 树木仍不需产生较深根系便可获取充足水分; 但在干旱和季节性干旱气候区, 毛管上升水会诱使树木形成较深根系, 以吸收深层土壤水和地下水来满足旱季较高的蒸腾所需(Fan et al., 2017)。
3 树木吸收利用深层土壤水的特征
3.1 树木对深层土壤水的直接吸收利用
从深层土壤中吸收水分运输至地上部分器官供给蒸腾和生长, 是树木对深层土壤水的最直接利用方式(Stone & Kalisz, 1991)。具备深根系是树木能够吸收利用深层土壤水的先决条件(Naumburg et al., 2005; Brum et al., 2017; Fan et al., 2017), 但其实际吸收利用特征则受到气候、土壤、地下水、植株生理活动等多种因子的影响和制约。潜在蒸散量和降水是影响树木深层根系吸水的重要气候因子。当潜在蒸散量增大时, 林木蒸腾量会相应变大, 其根系吸水范围会向更深的土层发展(Schenk, 2008; Nadezhdina et al., 2014; Guderle et al., 2018)。降水特征的变化能对深层根系吸水产生明显影响, 降水量减少一般会促进树木从深土层中觅取水分以满足蒸腾(Song et al., 2016; Broedel et al., 2017; Christina et al., 2018), 但树木若已对深层土壤水形成长期稳定的依赖性, 则其深层根系吸水特征对降水量的变化不敏感(Liu et al., 2018b)。如果降水量的季节分配发生变化, 树木的深层根系吸水量可能发生改变, 如数值模拟结果显示, 当更高比例的夏季降水量分配于冬季时, 美国东南部一火炬松(Pinus taeda)林分从1 m以下深土层的吸水量会更大(Qi et al., 2018)。降水频率对深层根系吸水比例的调控作用较弱, 但随降水频率降低(单次降水强度相应增大), 深层根系和深层土壤的树木干旱缓解能力会变弱(Qi et al., 2018)。
一般而言, 浅土层含水量较少而出现土壤干旱时, 会迫使树木转向吸收更多的深层土壤水并降低浅土层的快速干旱程度(Xi et al., 2013; Song et al., 2016)。但当树木蒸腾较大, 浅层根系吸水不能及时满足蒸腾需求时, 即使浅土层水分状况较好, 深层吸水比例仍会增加。土壤养分状况虽不能直接影响土壤水分吸收, 但可能会通过促进深根系的发育来影响树木深层吸水, 如Christina等(2018)发现如果对桉树人工林施钾肥, 则会提高林木对深层土壤水的吸收量。
地下水位变化会影响树木的深层根系吸水特征, 但不同树种响应不同。对于根系分布较深和依赖地下水的树种, 其根系吸水深度一般会随地下水位的降低而逐渐下移(Si et al., 2014; Chen et al., 2016), 且在干旱的季节和地区更为明显(Barbeta & Pe?uelas, 2017; Antunes et al., 2018), 这可能是下降的地下水位诱导了根系分布的变深而引起(Naumburg et al., 2005)。但对于根系分布较浅和不依赖地下水的树种, 这种根系吸水可塑性变化则较少发生(Antunes et al., 2018)。此外, 如果地下水中含盐量过高或酸性过强, 则会抑制树木对地下水的吸收利用(Steggles et al., 2016)。
树木根系吸水特征会因树体大小、健康状况以及根区位置的不同而发生改变。其中, 关于树体大小(个体发育)与根系吸水深度间的关系目前存在不一致的结果: 多数研究发现树体越大, 深层土壤水吸收比例会越大(Dawson & Ehleringer, 1991; Christina et al., 2017); 但也有研究发现较大的树木趋于在表土层吸收水分(Hardanto et al., 2017); 同时还有****在南美洲法属圭亚那的热带雨林中, 发现树木尺寸(胸径、树高)与平均根系吸水深度间的相关关系非常弱(Stahl et al., 2013)。随着树龄的增加、健康状况的恶化, 如果树体出现退化、老化, 则其主要水分来源会从深土层向浅土层转移, 且退化程度越高, 这种趋势越明显(Sun et al., 2018; 张欢等, 2018)。如在我国张北地区, 未退化的小叶杨(Populus simonii)林分主要利用320-400 cm土层中的水分, 而轻度、中度和重度退化林分的主要水源深度则分别在120-320、20-80和0-20 cm (张欢等, 2018)。此外, 在树木根区范围内, 距离树体越近, 根系可能分布越深(Xi et al., 2013), 所以深层土壤水的消耗量也会越多(Cubera & Moreno, 2007)。
同一树种对深层土壤水的吸收利用存在很强的时空变异性。具备深根系虽能保证树木同时利用不同土层的水分和(或)地下水, 但却存在明显的季节和年际变化: 湿润季节为高效利用降水会优先从浅土层吸收水分, 而干旱季节则会转向吸收更多的深层土壤水和(或)地下水(Xi et al., 2013; Song et al., 2016; Sun et al., 2018); 随林龄增长, 根系分布变深, 树木对深层水的消耗量一般会逐渐增加(刘晓丽等, 2014; Christina et al., 2017; Liu et al., 2018a)。不同地区的同一树种, 由于气候、土壤、林分特征、根系分布等诸多因子的差异, 往往具有不同的深层水分利用策略。例如, 旱季, 位于柬埔寨中部地区的橡胶树(Hevea brasiliensis)人工林的水分来源深度大于泰国东北部地区(更加湿润)(Kumagai et al., 2015); 在黑河下游额济纳三角洲地区相同地下水位下, 河边低地的胡杨(Populus euphratica)主要利用地下水和深层土壤水, 而沙丘和戈壁滩上胡杨的主要水源则为地下水和多个土层的水分(Si et al., 2014)。
同一地区不同树种对深层土壤水的吸收利用策略在时间和空间上常存在较大差异, 该现象在干旱(Chen et al., 2016)、半干旱(Liu et al., 2018a)、季节性干旱(Antunes et al., 2018)和湿润气候(Yang et al., 2017)条件下均存在。树木物候、耐旱性、资源空间大小、根系分布深度以及地下水位等的不同, 是造成这种差异的主要原因(Stratton et al., 2000; Stahl et al., 2013; Chen et al., 2016; Liu et al., 2018a)。
林分结构的差异和改变能引起树木根系吸水的可塑性变化。与纯林相比, 不同树种混交时, 会促使根系吸水模式可塑性较强的树种在旱季转向主要利用深层土壤水(Hardanto et al., 2017; Amazonas et al., 2018), 而且树种组成、混交方法的差异也会对深土层的水分消耗产生影响。例如, Amazonas等(2018)的数据显示, 与桉树(E. grandis × E. urophylla)纯林相比, 桉树与30个乡土树种的混交林中, 1 m以下土层的水分消耗量明显变小, 但桉树与30个乡土树种的混交林的深土层水分消耗量却略高于10个乡土先锋树种和30个乡土树种的混交林。林分密度对深层水分吸收利用的影响目前研究结果不一致。大多数研究显示, 随林分密度增大, 深土层含水率会降低, 即树木深层吸水量增加(Oliveira et al., 2005; Adane & Gates, 2015)。但在美国内布拉斯加州中南部, Adane和Gates (2015)却发现北美圆柏(Juniperus virginiana)纯林在间伐一半林木4年后, 1-3 m土层的含水率明显降低, 出现这种现象的原因可能是间伐后林冠降水截获减少和林下透光增强促进了林下植被生长和土壤蒸发, 从而导致土壤水分消耗量变大。
树木对深层土壤水的吸收利用, 从本质上讲, 主要是受到土壤水分垂直分布和植株蒸腾耗水的共同驱动, 因为二者分别决定了土壤中的水分供给能力和树木对水分的需求大小(Schenk, 2008; Fan et al., 2017)。因此, 上述因子之所以能对树木对深层土壤水的吸收利用产生影响, 主要源于能引起土壤水分分布和树木蒸腾耗水时空动态的改变。
综上所述, 整体而言, 与树木吸收利用浅土层水分的研究相比, 目前关于树木吸收利用深层水的研究还相对较少, 现有相关研究主要集中在我国西北、西非、美国西部、澳大利亚西部、伊比利亚半岛、巴西中部等干旱、半干旱、季节性干旱地区。此外, 现有的研究在测定树木对深层土壤水的利用时, 很少有直接测定至树木根区的下限, 即大多数研究对树木深层吸水模式和特征的了解并不“完整”。同时, 这些研究也大多是在1个生长季内的1个或几个时期开展, 而针对较长的时间尺度, 类似的试验研究极少, 目前只有Christina等(2017)在巴西研究了一个轮伐期内桉树人工林的深层吸水特征。
3.2 根系水力提升
水力提升(HL)是树木实现深层土壤水吸收利用的另一种方式, 同时也是除深根系和气孔调节之外的另一重要抗旱机制。该现象是指在浅土层与深土层存在水势梯度的情况下, 植物深层根系借助水势差在夜间将深土层中的水分运输至表层根系, 然后再释放至浅土层中以供白天蒸腾所需(Caldwell et al., 1998)。该现象最早发现于20世纪30年代, 但直到1987年才由Richards和Caldwell (1987)首次提出HL这一概念。据笔者统计, 水力提升现象目前已在全球40多个乔灌木树种中出现, 其发生区域覆盖干旱、半干旱、季节性干旱以及湿润气候区。国外对HL的研究较多, 已从简单的现象描述发展到定量评价、系统机理解释乃至与全球气候等结合探讨HL对气候变化以及森林动态的影响。国内对HL的研究起步较晚, 20世纪90年代中期开始至今, 相关的中文研究性论文仅有20多篇; 这些研究主要集中在西北干旱、半干旱地区和华北低丘山区的少数树种上, 如胡杨、多枝柽柳、胡桃等(袁国富等, 2012; 鱼腾飞等, 2014, 2017; 陈亚宁等, 2016; 何春霞等, 2016)。
现有研究证实, HL能对各类植物生态系统的水分循环产生重要影响。对于不同地区的不同生态系统, 通过HL作用可在干燥的浅土层中每天平均释放0.04-1.30 mm的水分, 从而使其水分含量平均提高28%-102%, 并供给2%-80%的蒸腾耗水(Neumann & Cardon, 2012; 苏华等, 2014)。虽然某些情况下HL的蒸腾贡献比例较小, 但若其发生在一些关键时期, 则会通过减缓表层根系空穴化的发生、避免表层根系死亡等方式, 对树木的水分关系产生重要影响。
HL的大小受多种环境因素影响。根系能触及地下水或毛管边缘水时, 会促进HL的发生或提高其水分提升量, 尤其是在深层土壤质地较粗、持水力较弱的情况下, 如Ryel等(2002)发现生长于粉壤立地上的Artemisia tridentata如果能触及地下水, 则在100 d的模拟干旱周期中, 其HL量平均每天可提高0.2 mm。土壤质地能明显影响HL的发生, 大田观察、室内控制试验以及模拟研究均表明当土壤砂性更强(质地变粗)时, HL发生的频率或大小会下降(Schippers et al., 1967; Yoder & Nowak, 1999; Siqueira et al., 2008; Wang et al., 2009)。其原因可能是当土壤逐渐干旱时, 砂性更强的立地上的土壤、根系以及根-土界面导水率会下降得更快或更难维持(Ryel et al., 2002)。土壤垂直剖面上水势梯度的存在是HL发生的必要条件, 因而土壤水分状况能对HL的大小产生影响。一般而言, 表层土壤较湿时, 表土层与深土层间的水势梯度较小, 不会诱发HL; 表层土壤过于干燥时, 由于对根系和土壤导水率影响较大, 因而也会限制HL的发生或降低其幅度; 只有当表土层干燥至较为适中的状态(约-0.4到-0.8 Mpa (Ishikawa & Bledsoe, 2000; Domec et al., 2004; Meinzer et al., 2004))而能与植物地上部分“竞争”水分时, HL才会发生并保持在较高的水平。此外, 在表层土壤较干旱时, 如果大气蒸发潜力较小, 树木蒸腾需求较弱, HL也会在白天发生; 但当夜间发生HL时, 树木夜间蒸腾和树体充水会对其水力提升产生抑制作用, 且后者的抑制作用高于前者(Yu et al., 2018)。
4 树木吸收利用深层土壤水的机制
树木根系主要分布于浅土层, 深层土壤中根系一般较少(Jackson et al., 1997; Schenk & Jackson, 2002a), 但其却在树木水分关系中发挥着非常重要的功能作用。因而, 长期以来, “为何少量的深根系却能在关键时期给树木供给大量的水分”成为众多****关注的科学问题。其中可能涉及很多机制, 本文主要从深根系性状调节和整株水力特性协调两方面进行论述。4.1 深根系性状调节
根系结构特征决定根系功能发挥, 树木实现对深层土壤水的直接利用首先需要在深土层中生长根系。但若想进一步通过水力提升来利用深层土壤水, 则还需在浅土层中分布根系, 使整个根系系统覆盖具有较大水分梯度差异的土壤区域(Neumann & Cardon, 2012)。根系具有很强的可塑性, 对土壤环境非常敏感(Prieto et al., 2015; Fan et al., 2017; Ma et al., 2018)。深土层的环境条件(氧气浓度、养分含量、碳氮比、温度变异性一般较低, 土壤容重和强度一般较大)与浅土层存在明显差异, 尤其是其中的水分含量一般较高且相对稳定。因此, 深根系经历水分亏缺或胁迫的概率相对较小, 故而其功能会与浅层根系出现分异, 可能会更加注重水分供给(Prieto et al., 2015; Wang et al., 2015)。所以, 在具备深根系的情况下, 树木为高效吸收利用深层土壤水, 会及时对其深层根系的结构、解剖、生理、化学、动态或形态等性状进行调节, 以充分发挥其水分供给功能。但对于自然界树木而言, 这种调节绝非只有一个方向或一条路径, 而是在不同气候、树种、土壤等条件下存在多种形式(图1), 即调节策略多样化。归纳起来, 这些调节大致集中于以下方面。
4.1.1 单位土体内深根系吸收面积
该方式是通过在单位土体内生长更多的根系, 增大根系吸水面积, 以提高树木深土层中的水分吸收量。土壤水分充足时, 根系吸收面积与吸水量成正比, 因此通过在单位体积土壤内增加吸收根的根长密度、根表面积密度、根系体积等, 来扩大根系在土壤中的吸水面积, 是提高深土层水分吸收的重要方式(McElrone et al., 2007)。为此, 树木可能会通过提高深土层中的根系分配比例(Adriano et al., 2017), 提高深根系生长速度或降低其周转速率(Germon et al., 2016; Lambais et al., 2017; Weemstra et al., 2017)来达到增大或维持较大深根系吸收面积的目的。由于深根系构建成本较高, 为实现“经济”平衡, 树木会通过减少深根系碳、氮含量, 降低其呼吸速率, 提高其木质素、纤维素含量, 延长根系寿命等方式降低其维持成本(Baddeley & Watson 2005; Prieto et al., 2015; Wang et al., 2015; Germon et al., 2016; Pierret et al., 2016)。
4.1.2 深根系分布范围
该方式是使根系在更大区域内吸收水分, 来提高深根系吸水量。扩大深层根系的分布深度, 不仅可增大其水养资源觅取范围, 而且也能减小根系个体间的资源竞争压力, 所以是树木提高深层水分吸收的最重要和最常见的一种方式。对于同一树种, 这种调节一般在较为干旱的地区比较明显(Ma et al., 2013), 而降水充足时则可能很弱(Laclau & Laclau 2009; Adriano et al., 2017)。此外, 深根系为拓展水分吸收范围, 也可能开采浅土层。如, Di等(2018)发现毛白杨垂直生长的骨骼根在扎至1 m以下土层后, 又突然斜向上生长到38 cm深的浅土层, 实现了对深层和浅层土壤水的同时吸收。
4.1.3 深根系水分吸收效率
该方式是通过提高单位长度或面积根系的水分吸收效率(或能力), 来增加根系从深土层吸收的水量, 而且其与增加单位土体根系吸收面积和根系分布范围相比, 可减少根系构建的碳成本投入。由于可缩短水分从土壤进入根系木质部的径向传输距离, 因而减小细根直径可能增大其水分吸收速度和效率, 并同时扩大其水分吸收范围(根系组织密度变化不大的情况下, 比根长增大)(Dhiman et al., 2017; Ma et al., 2018)。但很多研究发现深层细根的直径和比根长与浅层根系相比并未有显著差异(Adriano et al., 2017; Pinheiro et al., 2016; Kirfel et al., 2017), 可见树木可能较少通过调节根系形态来调控深根系吸水效率。
根系吸水过程中, 水分从根系表皮穿过皮层, 然后跨过内皮层进入木质部导管过程中, 可以通过共质体、质外体和跨膜等3种途径进行运输, 其中均涉及水分的跨膜运输(主要是跨膜途径), 而细胞膜上的水孔蛋白在水分子的跨膜中起主要控制作用(Gambetta et al., 2017)。因此, 调控深层根系水孔蛋白基因的表达, 增加或改变水孔蛋白的数量、状态(开闭)和传导率, 是提高深根系水分吸收效率和能力的重要途径, 其还可弥补根系表面积下降和缺氧对深层根系吸水的影响(McElrone et al., 2007; Comas et al., 2013; Johnson et al., 2014a; Gambetta et al., 2017)。例如, Johnson等(2014a)在美国德克萨斯州发现, 当干旱增强时, Quercus fusiformis深根系(约20 m深)中的水孔蛋白活性会上调, 使根系水力导度增加2.6倍, 从而保障林木旱季时吸收更多的深层土壤水。此外, 通过形成更多的菌根来扩大根系的吸收面积, 可能也是树木提高深根系水分吸收能力的一种方式(B?rja et al., 2017)。
4.1.4 深根系水分传输能力
根系系统水分供给量由其水分吸收和传输能力共同决定。为提高深层水吸收利用效率, 树木常提高深根系水分传输能力(轴向导水率), 将吸收进来的水分高效、快速地运至地上部分器官。如Pate等(1995)发现, Banksia prionotes垂直根位于深土层(130 cm)部分的比导水率较其浅土层(30 cm)部分高18倍。
为提高深根系水分传输能力, 树木会对其粗根以及细根(≤2 mm)中传输根(木质化、主要起传输和储藏作用的根系)(McCormack et al., 2015)的木质部结构进行调节。例如, 通过增大木质部平均导管直径、大径级导管数量、导管密度、导管长度、疏导组织占中柱面积的比例等来提高深根系导水率(Pate et al., 1995; McElrone et al., 2004; Johnson et al., 2014b; Kirfel et al., 2017)。这是因为根据泊肃叶定律, 水分传输管道(导管)中的水流量对管道半径极其敏感, 在驱动力压力梯度和液体黏度不变的情况下, 当管道半径加倍时, 其中的水流体积流速将增加16倍。深根系中还可能特化出一些因具有较大直径导管而具有超高导度的根系(high conductivity roots), 这种根系的导水率可较同径级根系的平均导水率高出50%以上(Kirfel et al., 2017)。此外, 树木还可通过增加深根系中具有相互连通性的导管的比例以及导管上纹孔的平均尺寸或数量, 来提高不同导管之间的水分传输, 从而增大根系导水率(Johnson et al., 2014b)。
深土层细根中的吸收根(非木质化、吸收水养资源的根系)(McCormack et al., 2015)在解剖结构上的变化目前还了解很少。但根据现有针对深土层中较细根系(如0.5-2 mm、1-2 mm) (Johnson et al., 2014b; Kirfel et al., 2017)以及浅土层不同深度(如0-10和20-30 cm)一级根(位于根系分支最末端的根系) (Wang et al., 2015)的研究结果, 可以推断深层吸收型细根很可能也会和粗根一样调节解剖结构(增大导管平均直径、大径级导管比例、中柱直径等), 来提高其导水率。
4.1.5 深根系物候
为保证深层根系能在旱季或表土层干燥时为地上部分器官及时供给充足水分, 树木会对深层根系的物候(存在明显季节性)进行调节, 以使其能适时、及时地激活和发挥水分供给功能: 在旱季树木对深层土壤水有较强需求时, 会快速启动深层根系的生长去觅取和吸收深层水源, 或进入休眠季地上部分器官不再生长时, 将更多的碳水化合物用于深根系生长, 以为次年吸收利用深层水做“材料准备” (Maeght et al., 2015; Germon et al., 2016; Lambais et al., 2017)。这可能是树木在平衡资源分配、协调各器官生长的同时, 为能高效利用深层水而采取的一种时间上的生态策略。例如, 在巴西桉树人工林中, 当干旱导致浅土层水分较少而浅层根系产量明显降低时, 4 m以下土层中的根系生长速度大幅快速升高(Lambais et al., 2017)。在法国杂交胡桃(Juglans regia × nigra)-小麦(Triticum turgidum subsp. durum)复合系统中, 从春季至夏初, 胡桃深层根系(深>2.5 m)生长的启动时间滞后于表层根系, 且在冬季落叶后, 深层根系出现第二个生长高峰(Germon et al., 2016)。
综上所述, 为提高对深层土壤水的吸收和利用, 树木可能存在多种类型的深根系性状调节策略。这些策略可简单归纳为: 在树木有深层土壤水需求的时间(调节深根系物候、动态), 从水资源更多的地方(调节深根系密度和分布范围), 将水分快速吸收(调节深根系吸收效率), 并高效运输(调节深根系传输能力)至地上部分器官(图1)。对于不同树种, 其在不同环境下可能采取其中的一种或多种策略来达到提高深土层水分吸收的目的。
图1
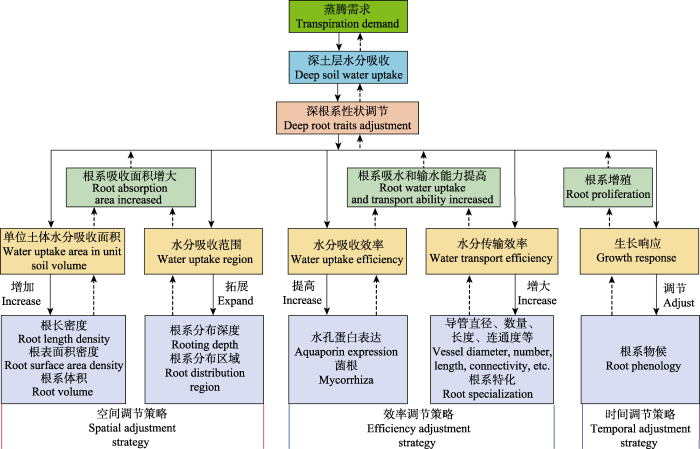 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1树木深根系为吸收深层土壤水而可能采取的潜在性状调节策略。
Fig. 1Potential deep tree root trait adjustment strategies for absorbing deep soil water.
然而, 需要注意的是, 真菌的侵染(形成菌根)可能会降低深根系性状可塑性的程度, 而且根系类型定义的不同(如以直径<0.5、1或2 mm等来定义细根)也可能使这种变化难以被发觉(Weemstra et al., 2017; Ma et al., 2018)。此外, 近年来, 有研究发现树木深根系与浅根系在营养元素的吸收上存在功能特化现象(da Silva et al., 2011; Bordron et al., 2018), 但在水分吸收上, 除已发现的“具有超高水力导度的特化根系”外(Kirfel et al., 2017), 是否还存在其他类似现象, 以及其对树木吸收深层土壤水的作用目前还尚不清楚。
4.2 整株水力特性协调
根系、茎干、枝条、叶片四个水力器官组成了植物的水分传输系统, 四者在保障整株水力路径中水分的传输上是功能统一的, 其中任一器官(尤其是叶片和根系)木质部水力导度下降或水力失调都会直接影响整株植物的水分传输效率, 从而影响叶片气体交换以及植物生存和生长(潘莹萍和陈亚鹏, 2014; Johnson et al., 2016; Brum et al., 2017; Wason et al., 2018)。因此, 为保障植株体内水分的高效运输和利用, 并防止空穴化引起的水力失败, 树木在吸收利用深层土壤水时, 必然要对地上与地下部分各器官的水力特性进行综合协调和权衡(Pate et al., 1995; McElrone et al., 2004, 2007)。然而, 目前单独研究根、茎、枝或叶水力性状的较多, 但运用联系的观点来综合研究四者水力特性间协调与权衡关系的还极少(Choat et al., 2012; 金鹰和王传宽, 2015; Johnson et al., 2016; Wason et al., 2018)。现有的少数研究结果显示, 树木在发展出较深的根系而能获取相对稳定的水源后, 为实现深层土壤水利用最大化, 会对叶片水力性状进行调节。例如, 会使树木的单位叶面积(“水分散失”面积)具有更大的根系表面积(“水分供给”面积), 从而使其在生长季维持相对稳定且更高的水分状况(较高的黎明前叶水势)和气孔导度, 以保持较大的蒸腾速率, 并使叶片相应具有较高的光合速率(Xu & Li, 2008; Bucci et al., 2009; Pinheiro et al., 2016; Brum et al., 2017)。由于深根系树木水分来源相对充足, 其叶片可能会暂时采取非等水调节策略(anisohydry), 降低气孔对叶片水分散失和传导的调控作用, 以使叶片能持续维持较高的气体交换, 但这也可能导致其叶水势在一天内出现更大变幅(Xu & Li, 2008; 罗丹丹等, 2017)。此外, 有研究发现深根系树种的叶片失膨点水势高于(绝对值更小)浅根系树种(Brum et al., 2017), 由此表明, 树木在发展出深根系后, 虽然水分来源更加充足和稳定, 但却可能面临更高的水力失败潜在风险, 即削弱叶片对水分胁迫的抵抗能力(Choat et al., 2012)。
树木通过深根系获取充足水分供给后, 其叶片与根系间的协调度可能也会提高, 从而保障叶片的水分需求信息能及时反馈和传递至根系, 然后使根系根据蒸腾需求通过生理调节策略来调控其水分吸收和供给量。例如, Xu和Li (2008)在新疆古尔班通古特沙漠南缘发现, 根深达10 m的梭梭的比叶导水率和最大蒸腾速率与根土界面的平均土壤水势(用黎明前叶水势表征)间有极高的显著相关关系, 而对于根深为3 m的梭梭, 这种关系非常弱。McElrone等(2007)在美国德克萨斯州爱德华兹高原针对两个树种均发现, 深层根系(20 m深)在树冠蒸腾发生变化时, 能快速地做出生理响应, 通过调控细根上水孔蛋白的活性, 来调节细根和粗根水力导度的大小, 实施“按需供水”。
各器官木质部的解剖结构及其相对变化特征, 决定了树木体内的水分传输效率(Schenk, 2008; Johnson et al., 2016)。当生长环境发生变化, 为维持生长和生存, 树木可能会改变其木质部结构以利于充足的水分供给, 但同时会尽可能降低其水力失败风险, 即在木质部水分传输效率与水力安全之间维持一种权衡(Schenk, 2008; Choat et al., 2012)。基于目前较少的研究结果, 可以推断, 不同树种的水力通路中(深根—浅根—茎干—枝条—叶片), 从下至上各器官木质部结构及其水力性状不存在统一的变化规律。如Kirfel等(2017)发现0-200 cm土层内, Fagus sylvatica根系木质部解剖结构(木质部空腔占比、导管密度、平均和最大导管直径等)和比导水率基本不受土层影响。但Pate等(1995)、McElrone等(2004)、Johnson等(2014b)和Wason等(2018)的研究却表明, 对于有些树种, 从其深根至浅根再到茎干和枝条, 木质部导管会逐渐变窄、变短, 其间的连通比例可能更小, 从而导致比导水率逐渐降低; 但导管结构的这种变化也使从下至上各器官木质部的导水率损失50%时对应的木质部水势(Ψ50)和导管纹孔进气压力值可能逐渐降低(绝对值更大), 从而致使导管抗空穴化的能力逐渐增强。此外, 对于不同树种, 根系深浅的不同会使其茎干(密度、导水率)和叶片(比叶面积)的水力结构产生差异(Bucci et al., 2009); 但对于同一树种, 根系变深而能持续获取稳定深层水后, 其茎干新生木质部的解剖结构可能会发生变化, 因为茎干支撑的蒸腾水流最大, 所以对生长环境的变化可能更敏感(Choat et al., 2012; Jasińska et al., 2015)。
5 树木吸收深层土壤水对人工林培育的启示
由上可知, 关于树木对深层土壤水的吸收利用特征, 目前在人工林和天然林中均已有一定研究, 但由于学科背景和关注点的不同, 将现有该领域的相关知识和信息应用于人工林(主要针对以生产纤维材、大径材等为目的的工业用材林)培育实践的工作却相对较少。因此, 基于现有研究成果, 针对从林木栽植到收获过程中涉及的水肥管理、间作、树体管理、林分结构调控、抚育采伐、低质低效林改造等主要培育技术环节, 从影响树木水分生理和深土层水分利用的角度出发, 提出以下几点可能会对人工林培育产生影响的启示(本文重点旨在提出相关理念, 而非制定具体技术措施; 此外, 一些众所周知的理念, 此处也不再赘述, 如树种选择技术中深根系树种可用于抗旱造林、提高造林成活率等)。5.1 水分管理中应使林木适度利用深层土壤水和选用合适的灌水频率
在干旱、半干旱和季节性干旱地区, 水分管理是人工林培育常采用的集约经营措施。现有的人工林水分管理模式大致分为从不灌溉(模式I)、栽植后前几年灌溉(模式II)、持续灌溉(模式III)等几类。模式I和II很难使人工林达到较高的生产力水平(Xi et al., 2014; 贺勇等, 2015; Zhu et al., 2017), 而且因为不灌溉和较少灌溉, 所以表面上最为“节水”。然而, 如前述, 由于这两种模式下浅层土壤长期比较干燥, 会促进深层根系生长, 因而可能会吸收和消耗大量的、“更加珍贵”的深层土壤水资源甚至地下水。所以, 可能会对深层土壤水分环境造成严重的负面影响(如土壤干层(Liu et al., 2018a)), 尤其是当降水不能对深层水资源进行有效补充时(Christina et al., 2017)。此外, 由于模式I和II下的林分可能对深层土壤水或地下水产生了长期的、较高的依赖性, 其水分胁迫抵抗能力会减弱, 因此一旦出现地下水位的剧烈下降或发生极端干旱事件, 则林分可能会更加脆弱, 死亡率更高(Xi et al., 2017; Chitra-Tarak et al., 2018; Sun et al., 2018)。模式III虽然能明显提高林木生长(Xi et al., 2014; 贺勇等, 2015; Zhu et al., 2017), 但会消耗大量的灌溉水资源, 并需要较高的成本投入。同时, 模式III还有可能造成林木根系分布深度较浅和范围较窄, 从而不利于林分的抗风性; 而且在一旦出现灌溉水资源短缺而不能及时供给水分时, 相对较小的根区范围可能会使林木遭受较大的死亡和枯梢风险。
因此, 如果对林木进行适当“亏缺”灌溉, 会促进土壤表层根系生长, 并能激发林木深层根系的适当发育, 使林木不仅能适度利用深层土壤水, 而且可充分利用降水资源。从而能有效地实现灌溉水资源的节约、降水水资源的高效利用与保存以及深层水资源的可持续利用, 并在大幅提高林木生长的同时, 降低对深土层水生态环境的负面影响。
如前所述, 降低降水频率而单次降水强度相应增大, 会减弱深层根系吸水对林木干旱的缓冲能力。因此, 对于漫灌、沟灌和畦灌等单次灌水定额较大而灌溉间隔期较长的灌溉方式, 应寻求最佳的灌水频率, 否则不仅会消耗大量灌溉水资源, 还可能使林分的抗旱能力减弱且长势不尽人意。在笔者团队的灌溉研究中, 就曾观察到在低频率和高灌水定额下, 树木的长势和水分状况反而相对较差。
5.2 施肥策略制定应考虑林木和深土层水分间的关系
施肥策略的差异会对林木的水分利用以及林木与深土层水分间的关系产生影响(Christina et al., 2018), 如果在制定具体施肥制度时能对相关信息予以充分考虑, 则有助于提高林木对干旱的抗性和应对气候变化的韧性。如Christina等(2018)在巴西桉树人工林研究中发现, 年降水量的下降会增加林木的吸水深度, 降低深土层中的储水量并造成地下水位下降, 但是不施钾肥的林分中的这种负面效应低于施钾肥林分; 由此表明, 制定合理的施肥制度, 有可能通过在雨季增加深层土壤储水量的方式, 在气候变化造成旱季延长时, 降低林木水分胁迫风险。此外, Christina等(2018)的研究表明, 采用较高施肥量的一个不利影响是在干旱时期可能会增加林分的水分胁迫程度, 因为高施肥量可能促进深根系生长, 加速深层水分消耗。因而, 在保证林木正常养分需求的情况下, 通过降低施肥量, 可以帮助缓解旱季树木的水分胁迫程度, 以及气候变化带来的干旱天气对林分生长的影响。这也为我们看待人工林培育中过量施肥可能带来的负面环境效应提供了除减小养分深层渗漏和地下水污染之外的新视角。
5.3 利用间作可实现生物灌溉
Sekiya等(2011)基于水力提升的原理, 设计了Brassica rapa var. peruviridis与多年生饲料植物(大黍(Panicum maximum)、高羊茅(Festuca arundinacea))间作的复合种植系统, 发现在将饲料植物地上部分割除后, 其根系系统能通过水力提升作用将深土层中的水分供给浅根系的B. rapa var. peruviridis利用, 使其保持正常的水分状况和产量。由此表明, 针对人工林, 或许也可采取类似的设计, 将浅根系树种与多年生饲料植物混交, 这样不仅可收获饲料, 而且还能将饲料植物的根系系统作为“灌溉工具”为目标树种供给水分, 实现“生物灌溉”, 降低人工灌溉成本。5.4 合理的修枝技术可改善深土层水分状况
如前所述, 树冠大小与根系分布深度间存在正相关关系, 蒸腾需求增大会促使林木吸收深土层水分, 这意味着对树冠尺寸进行控制, 可通过降低蒸腾来减缓深土层水分资源消耗。例如, 魏新光等(2014)在黄土丘陵区提出的节水型修剪技术, 通过控制树冠大小, 明显降低了枣树蒸腾量, 同时也使深土层水分消耗量相应降低。由此可见, 在人工林中, 进行适时、适量的修枝, 除了具有改良林木干形和材质等常规作用外, 还有可能改善深层水分状况, 从而提高林木抗旱能力, 促进其生长。5.5 合理的树种混交能促进深层土壤水分储库“缓冲”作用的发挥
合理的树种混交可促进林木对深层水资源的开采能力, 而且还可能通过水力提升作用将深土层中的水分转移至浅土层供混交的浅根系树种利用, 从而提高气候变化条件下混交林生态系统的稳定性和抗性。如, 在巴西的桉树与相思(Acacia mangium)混交林中, 0-17 m土层内单位面积土壤上的细根生物量远高于两个树种的纯林, 其细根比根长和比根表面积比相思纯林高110%-150%, 比桉树纯林高34%; 且混交林9-17 m土层内的相思和桉树细根比根长和比根表面积都显著高于各自纯林, 表明混交林深土层水资源的吸收利用能力远高于纯林, 从而使干旱条件下混交林中的深土层能更好地发挥其“水分缓冲库”的作用(Germon et al., 2017)。然而, 需要注意的是, 树种混交的这种作用是相对的。因为混交方式或树种不同, 深土层的水分消耗状态会存在很大差异, 这意味着树种混交后, 对于一部分树种, 深层水分状况可能较其纯林变好, 但对另一部分树种来讲, 却可能变差。例如, Amazonas等(2018)发现, 与和桉树混交相比(模式I), 30个乡土树种在与10个乡土先锋树种混交后(模式II), 1 m以下深土层中的水分略高, 但模式I的深土层水分却明显高于桉树纯林(模式III), 所以, 对于桉树而言, 模式I能改善深土层水分, 但对于30个乡土树种, 模式I却会加剧深土层水分消耗。因此, 在营造一些混交林时, 有必要考虑混交不同的伴生树种可能会对目标树种深土层水分环境造成的作用, 否则可能会给目标树种带来不利影响。
5.6 基于树木土壤水分利用深度的间伐木选择技术
在对人工林进行林分结构调控时(如抚育间伐、树种调控等), 间伐木的选择是关键。虽然多个研究表明, 个体较大的林木会更多地吸收深层土壤水(Dawson & Ehleringer, 1991; Christina et al., 2017), 但Hardanto等(2017)却在橡胶树(Hevea brasiliensis)纯林和混交林中发现, 个体较大的林木则更趋向于利用表土层水分。这表明, 除了能对林地地上空间环境因子(光照、温度、湿度等)产生影响外, 针对不同树种的林分, 间伐不同类型的林木还可能会对林地不同土层的水分产生明显作用, 进而影响保留木的根系吸水和林地水分循环。因此, 可考虑基于不同树种的土壤水分利用深度特征, 调整和完善现有的间伐木选择技术。此外, 虽然一般而言密度较大林分的深土层水分消耗会更多, 但Adane和Gates (2015)却发现北美圆柏纯林在间伐一半林木4年后, 深土层(1-3 m)的水分却反而更低, 且地下水的降水补给量只有微弱变化。虽然这种情况可能为个例, 但却反映出在通过间伐调控林分密度时, 间伐强度的确定需要将深土层水分的潜在变化方向考虑在内, 不然可能会使保留木的土壤水分环境恶化。
5.7 基于深土层水分状况改善的低质低效林改造技术
我国有许多低质低效人工林, 其中比较典型的是三北地区退化的杨树防护林。基于稳定同位素技术, Sun等(2018)和张欢等(2018)均研究发现, 张北地区的杨树防护林退化越严重, 其主要水分来源土层就越浅。那么是林分退化导致深土层水分利用减少, 还是深土层水分不足造成林木深层根系吸水变少, 从而加剧了林分退化, 目前尚不能完全确定。但可以肯定的是, 地下水位下降与张北地区杨树林的退化有相关关系(Sun et al., 2018)。这也间接反映出如果能对林分深土层的水分状况进行改善, 则很有可能避免或减缓类似的退化现象发生, 而且也有可能对已经退化的林分以不通过树种更换的方式实现改造和复壮。今后有必要探索一些低成本、可操作性强的深土层水分改良技术, 如通过整地或深层根系灌水器等技术实现。综上, 在人工林培育过程中, 应充分利用好“林木吸收利用深层土壤水分”这把“双刃剑”, 以制定出既能降低外部水资源投入, 又能避免水分生态环境负面效应, 同时还能提高气候变化下人工林生态系统抗性与韧性的绿色栽培制度。
6 现有研究的不足及今后的发展方向
目前关于树木吸收利用深层土壤水的特征与机制已有一定研究, 但总体而言, 该领域的相关工作还较少, 且存在诸多不足之处和待解决的问题, 今后应针对这些不足和问题加强相关研究, 具体包括以下方面。关于树木深根系成因。地理尺度上根系分布深度的水文调控理论尚不完善, 应加强植物根系分布深度数据搜集, 填补现有数据库中较多的地理空白区域(如我国华北平原等), 同时研究大幅变化的地下水位条件下根系分布深度的决定因子, 以完善相关理论。
关于树木对深层土壤水的吸收利用特征。1)缺乏对树木深层水长期利用规律的了解, 考虑到气候变化效应和人工林经营周期的长期性, 今后应采取定位连续观测或空间代时间的方法, 在更长的时间尺度上(如整个轮伐期)开展相关研究。2)缺少涵盖整个树木根系分布深度的“完整”水分利用数据资料, 不利于全面了解不同地区不同类型森林的深层水分关系, 今后应研发便于极深土层水分监测的工具和方法, 使土壤水分测定深度能容易地达到树木根区下限。3)浅土层资源有效性长期存在差异时, 对树木对深层水分利用的可塑性特征缺乏深入了解, 今后可通过人为调控浅土层水分、养分含量的方法开展研究, 相关知识将有助于优化人工林培育技术。
关于树木对深层土壤水的吸收利用机制。1)对树木深层根系系统的结构和动态缺乏全面、系统、深入的了解, 限制了对树木吸收利用深层水分特征和机理的深化认识与预测。今后应在不同类型深层根系(粗根、传输型和吸收型细根)的木质部解剖结构、共生菌依赖特性、功能性状间的异速变化关系、物候特征、功能与形态特化和可塑性等方面加强研究。2)对提高深层土壤水吸收利用效率, 树木不同器官间水力特性的相互协调策略还缺乏深入认识。今后应针对从深层细根至叶片整个水力通路, 系统研究木质部水分传输效率和水力安全性状的时空变化特征以及二者间的权衡关系。3)现有研究大多是利用根系深度存在自然差异的不同树种, 来间接了解“根系分布深度改变”时树木各器官水力特性的可塑性变化, 这无疑将无法排除树种因素的影响。因而, 今后可针对相同气候、立地和林龄下的同一树种, 通过人为调控根系深度来研究该科学问题。
由于研究难度大且在森林生理生态过程中扮演重要角色, 上层林冠被喻为“地球的第八大洲” (吴毅等, 2016)。根系作为植物“隐藏在地下的另一半”, 其深层部分的重要性虽早已被人知晓, 但其结构与功能却远未被探知, 考虑到其更大的研究难度, 深层根区完全可视为待被人类探索的“地球第九大洲”。基于全球森林资源保护与生产的需要, 建议今后加强对树木深层根系及其生理生态过程与机制的研究, 并更多地关注如何将相关知识成果有效地应用在人工林培育、生物多样性保护、植被恢复等领域。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 3]

Although impacts of land-use changes on groundwater recharge have been widely demonstrated across diverse environmental settings, most previous research has focused on the role of agriculture. This study investigates recharge impacts of tree plantations in a century-old experimental forest surrounded by mixed-grass prairie in the Northern High Plains (Nebraska National Forest), USA. Recharge was estimated using solute mass balance methods from unsaturated zone cores beneath 10 experimental plots with different vegetation and planting densities. Pine and cedar plantation plots had uniformly lower moisture contents and higher solute concentrations than grasslands. Cumulative solute concentrations were greatest beneath the plots with the highest planting densities (chloride concentrations 225-240 % and sulfate concentrations 175-230 % of the grassland plot). Estimated recharge rates beneath the dense plantations (4-10 mm yr) represent reductions of 86-94 % relative to the surrounding native grassland. Relationships between sulfate, chloride, and moisture content in the area's relatively homogenous sandy soils confirm that the unsaturated zone solute signals reflect partitioning between drainage and evapotranspiration in this setting. This study is among the first to explore afforestation impacts on recharge beneath sandy soils and sulfate as a tracer of deep drainage.
DOIURL [本文引用: 4]
Fine roots of orange trees reached a depth of approx. 602m in a sandy tropical soil. Root area index was more responsive to irrigation than leaf area index. Although the crucial role of deep rooting on
DOIURL [本文引用: 3]
Mixed forest plantations now emerge as an alternative to traditional plantations in the tropics and represent ecological gains associated with production, wood quality and nutrient cycling. Mixed plantations with higher diversity may also be advantageous concerning their use of soil water. To shed light onto water-related issues of mixing Eucalyptus and a high diversity of tropical native trees, we explored the following questions: What is the impact of high diversity mixed plantations of Eucalyptus intercropped with native trees on soil water? How does the mixture affect the physiology of water use in native trees? Firstly, we tested the hypothesis that stands of Eucalyptus mixed with a high diversity of native trees consume less water compared to Eucalyptus monocultures, by measuring the temporal dynamics of soil water. Secondly, we tested how mixing with Eucalyptus affects the hydraulic performance of fast- and slow-growing native species in these forestry systems. This is the first time a large experiment has been implemented to compare the effects of monospecific Eucalyptus plantations, native species mixtures and mixed plantations of Eucalyptus and native species on soil water dynamics under controlled conditions in terms of site, age, soil type, topography and climate. We found that high diversity mixed plantations of Eucalyptus and native trees use less soil water, than Eucalyptus monocultures. However, the soil under the mixtures was drier than in native species stands. The mixing with Eucalyptus affected the hydraulic performance of native species by decreasing the leaf water potential and stomatal conductance of the fast-growing species, suggesting that fast-growing species performance may be especially constrained by competition for water from Eucalyptus . These findings have important implications for forest management and ecological restoration in the tropics. They will help to further develop silvicultural options to adapt to climate change and improve plantation forestry by using mixed plantations for production purposes or rehabilitation of degraded lands.
DOIURL [本文引用: 3]
[本文引用: 1]
DOIURLPMID [本文引用: 4]
Water stored underground in the saturated and subsurface zones below the soil are important sources of water for plants in water-limited ecosystems. The presence of deep-rooted plants worldwide, however, suggests that the use of groundwater is not restricted to arid and seasonally dry ecosystems. We compiled the available data (71 species) on the relative contribution of groundwater to plant water estimated using stable isotopes and mixing models, which provided information about relative groundwater use, and analyzed their variation across different climates, seasons, plant types, edaphic conditions, and landscape positions. Plant use of groundwater was more likely at sites with a pronounced dry season, and represented on average 49 per cent of transpired water in dry seasons and 28 per cent in wet seasons. The relative contribution of groundwater to plant-water uptake was higher on rocky substrates (saprolite, fractured bedrock), which had reduced groundwater uptake when this source was deep belowground. In addition, we found that the connectivity between groundwater pools and plant water may be quantitatively larger and more widespread than reported by recent global estimations based on isotopic averaged values. Earth System Models should account for the feedbacks between transpiration and groundwater recharge.
DOIURL [本文引用: 1]
Abstract A long-standing ambition in ecosystem science has been to understand the relationship between ecosystem community composition, structure and function. Differential water use and hydraulic redistribution have been proposed as one mechanism that might allow for the coexistence of overstory woody plants and understory grasses. Here, we investigated how patterns of hydraulic redistribution influence overstory and understory ecophysiological function and how patterns vary across timescales of an individual precipitation event to an entire growing season. To this end, we linked measures of sap flux within lateral and tap roots, leaf-level photosynthesis, ecosystem-level carbon exchange and soil carbon dioxide efflux with local meteorology data. The hydraulic redistribution regime was characterized predominantly by hydraulic descent relative to hydraulic lift. We found only a competitive interaction between the overstory and understory, regardless of temporal time scale. Overstory trees used nearly all water lifted by the taproot to meet their own transpirational needs. Our work suggests that alleviating water stress is not the reason we find grasses growing in the understory of woody plants; rather, other stresses, such as excessive light and temperature, are being ameliorated. As such, both the two-layer model and stress gradient hypothesis need to be refined to account for this coexistence in drylands.
[本文引用: 1]
DOIURL [本文引用: 1]
Functional specialization of fine roots was found for Eucalyptus grandis trees at harvesting age (662years) on tropical soils. Aiming to elucidate whether functional specialization is a ubiquitous feature of eucalypts, we focused on its changes with ontogeny, tree nutrient status and soil depth. We studied the potential uptake of N, K and Ca by 2-year-old E. grandis trees, as a function of soil depth and NPK fertilization. We injected NO 613 - 15 N, Rb + (K + analogue) and Sr 2+ (Ca 2+ analogue) tracers simultaneously in a solution at depths of 10, 50, 150 and 30062cm in a sandy Ferralsol soil. A complete randomized block design was set up with three replicates of paired trees per injection depth, in fertilized and non-fertilized plots. Recently expanded leaves were sampled at 7062days after tracer injection. Determination of foliar Rb, Sr concentrations and x( 15 N) allowed estimating the relative uptake potential (RUP) and the specific RUP (SRUP), defined as the ratio between RUP and fine root length density (RLD) in the corresponding soil layer. Various root traits were measured at each depth. Foliar N and K concentrations were higher in fertilized than in non-fertilized trees. The RUP of NO 3 – - 15 N decreased sharply with soil depth and the highest values of the SRUP of NO 3 – - 15 N were found at a depth of 5062cm. The RUP of Rb + and Sr 2+ did not change with soil depth, whilst the SRUP of Rb + and Sr 2+ were higher at the depth of 30062cm than in the topsoil, concomitant with an increase in root diameter and a decrease in root tissue density with depth. The SRUP of Rb + and Sr 2+ at a depth of 30062cm were on average 136 and 61% higher for fertilized trees than for non-fertilized trees, respectively. Fine roots of young E. grandis trees showed contrasting potential uptake rates with soil depth depending on the nutrient. Fertilization increased the uptake rate of Rb + and Sr 2+ by unit of root length in deep soil layers. Functional specialization of fine roots for cations of low mobility depending on depth previously shown at harvesting age also occurs in young E. grandis plantations and increases with fertilization application. This mechanism helps explaining very low amounts of cations lost by leaching in Eucalyptus plantations established in deep tropical soils, even in highly fertilized stands.
In:
DOIURL [本文引用: 1]
Global warming will most likely lead to increased drought stress in forest trees. We wanted to describe the adaptive responses of fine roots and fungal hyphae, at different soil depths, in a Norway...
DOIURL [本文引用: 2]
Abstract Understanding how Amazonian rainforests deal with extended droughts is critical in the face of changing climate. This research analyze the physical properties and the soil water dynamics of a deep soil profile in an area of primary forest in central Amazonia to elucidate these processes under drought and nondrought conditions. Physical soil properties derived from soil cores exhibited a distinctive layer between 480 and 8800002cm deep, characterized by higher microporosity and low plant water availability. In situ soil moisture measurements collected during the period from January 2003 through February 2006 and for depths ranging from 10 to 1,4300002cm suggest that, in the study site, the top 4800002cm of the soil profile satisfied most of the transpirational demands in normal climatological years. However, during exceptionally dry periods, such as the 2005 drought, root uptake occurs below 4800002cm. As concluded by previous studies, most of the uptake is concentrated in the first meter of the soil profile: More than 40% of the total demand for transpiration is supplied by the top meter of soil. Because deep root uptake occurred at greater depths than normal during the 2005 drought, our results suggest that this is a fundamental mechanism to cope with prolonged droughts.
DOIURL [本文引用: 5]
Background and aims The survival and coexistence of plants in water-limited environments are related to their ability to coordinate water acquisition and regulation of water loss. To assess the...
DOIURLPMID [本文引用: 2]
Adaptations of species to capture limiting resources is central for understanding structure and function of ecosystems. We studied the water economy of nine woody species differing in rooting depth in a Patagonian shrub steppe from southern Argentina to understand how soil water availability and rooting depth determine their hydraulic architecture. Soil water content and potentials, leaf water potentials ($\Psi _{Leaf} $) , hydraulic conductivity, wood density ($p_w $), rooting depth, and specific leaf area (SLA) were measured during two summers. Water potentials in the upper soil layers during a summer drought ranged from -2.3 to -3.6 MPa, increasing to -0.05 MPa below 150 cm. Predawn$\Psi _{Leaf} $was used as a surrogate of weighted mean soil water potential because no statistical differences in$\Psi _{Leap} $were observed between exposed and covered leaves. Species-specific differences in predawn$\Psi _{Leap} $were consistent with rooting depths. Predawn$\Psi _{Leap} $ranged from -4.0 MPa for shallow rooted shrubs to -1.0 MPa for deeprooted shrubs, suggesting that the roots of the latter have access to abundant moisture, whereas shallow-rooted shrubs are adapted to use water deposited mainly by small rainfall events. Wood density was a good predictor of hydraulic conductivity and SLA. Overall, we found that shallow rooted species had efficient water transport in terms of high specific and leaf specific hydraulic conductivity, low$p_w $, high SLA and a low minimum$\Psi _{Leap} $that exhibited strong seasonal changes, whereas deeply rooted shrubs maintained similar minimum$\Psi _{Leap} $throughout the year, had stems with high$p_w $and low hydraulic conductivity and leaves with low SLA. These two hydraulic syndromes were the extremes of a continuum with several species occupying different portions of a gradient in hydraulic characteristics. It appears that the marginal cost of having an extensive root system (e.g., high$p_w $and root hydraulic resistance) contributes to low growth rates of the deeply rooted species.
DOIURLPMID [本文引用: 1]
Hydraulic lift is the passive movement of water from roots into soil layers with lower water potential, while other parts of the root system in moister soil layers, usually at depth, are absorbing water. Here, we review the brief history of laboratory and field evidence supporting this phenomenon and discuss some of the consequences of this below-ground behavior for the ecology of plants. Hydraulic lift has been shown in a relatively small number of species (27 species of herbs, grasses, shrubs, and trees), but there is no fundamental reason why it should not be more common as long as active root systems are spanning a gradient in soil water potential ($\Psi _{\text{s}}$) and that the resistance to water loss from roots is low. While the majority of documented cases of hydraulic lift in the field are for semiarid and arid land species inhabiting desert and steppe environments, recent studies indicate that hydraulic lift is not restricted to these species or regions. Large quantities of water, amounting to an appreciable fraction of daily transpiration, are lifted at night. This temporary partial rehydration of upper soil layers provides a source of water, along with soil moisture deeper in the profile, for transpiration the following day and, under conditions of high atmospheric demand, can substantially facilitate water movement through the soil-plant-atmosphere system. Release of water into the upper soil layers has been shown to afford the opportunity for neighboring plants to utilize this source of water. Also, because soils tend to dry from the surface downward and nutrients are usually most plentiful in the upper soil layers, lifted water may provide moisture that facilitates favorable biogeochemical conditions for enhancing mineral nutrient availability, microbial processes, and the acquisition of nutrients by roots. Hydraulic lift may also prolong or enhance fine-root activity by keeping them hydrated. Such indirect benefits of hydraulic lift may have been the primary selective force in the evolution of this process. Alternatively, hydraulic lift may simply be the consequence of roots not possessing true rectifying properties (i.e., roots are leaky to water). Finally, the direction of water movement may also be downward or horizontal if the prevailing$\Psi _{\text{s}}$gradient so dictates, i.e., inverse, or lateral, hydraulic lift. Such downward movement through the root system may allow growth of roots in otherwise dry soil at depth, permitting the establishment of many phreatophytic species.
DOIURLPMID [本文引用: 4]
The depth at which plants are able to grow roots has important implications for the whole ecosystem hydrological balance, as well as for carbon and nutrient cycling. Here we summarize what we know about the maximum rooting depth of species belonging to the major terrestrial biomes. We found 290 observations of maximum rooting depth in the literature which covered 253 woody and herbaceous species. Maximum rooting depth ranged from 0.3 m for some tundra species to 68 m for Boscia albitrunca in the central Kalahari; 194 species had roots at least 2 m deep, 50 species had roots at a depth of 5 m or more, and 22 species had roots as deep as 10 m or more. The average for the globe was 4.6±0.5 m. Maximum rooting depth by biome was 2.0±0.3 m for boreal forest, 2.1±0.2 m for cropland, 9.5±2.4 m for desert, 5.2±0.8 m for sclerophyllous shrubland and forest, 3.9±0.4 m for temperate coniferous forest, 2.9±0.2 m for temperate deciduous forest, 2.6±0.2 m for temperate grassland, 3.7±0.5 m for tropical deciduous forest, 7.3±2.8 m for tropical evergreen forest, 15.0±5.4 m for tropical grassland/savanna, and 0.5±0.1 m for tundra. Grouping all the species across biomes (except croplands) by three basic functional groups: trees, shrubs, and herbaceous plants, the maximum rooting depth was 7.0±1.2 m for trees, 5.1±0.8 m for shrubs, and 2.6±0.1 m for herbaceous plants. These data show that deep root habits are quite common in woody and herbaceous species across most of the terrestrial biomes, far deeper than the traditional view has held up to now. This finding has important implications for a better understanding of ecosystem function and its application in developing ecosystem models.
DOIURL [本文引用: 5]
荒漠河岸林是长期适应干旱区内陆河流域环境的产物,具有独特的水分利用和生存策略.对黑河下游胡杨、柽柳等主要荒漠河岸林植物液流、水势、木质部导水率、根系水分再分配以及植物水分来源等的测试分析表明,在降水稀少、蒸发强烈的黑河下游,胡杨、柽柳主要利用的是深层土壤水和地下水,其中成熟胡杨主要吸收地下175~325 cm处土壤水和地下水供其蒸腾和生存所需;胡杨根系水力提升过程主要发生在10~70 cm土层,蒸散导致的土壤耗水量中有10%来自根系提升的水量;植物根和枝条木质部的初始导水率(Ks0)和最大导水率(Ks,max)相对较高,其中柽柳木质部的潜在导水能力比胡杨强;胡杨、柽柳液流速度呈单峰型,在午后达到峰值,黎明前水势无显著变化,反映未受到严重水分胁迫;黑河下游河道附近的地下水埋深大多在4m以内,基本可以满足荒漠河岸林植物的生长发育,黑河下游荒漠河岸林生态系统处于稳定和发展状态.
DOIURL [本文引用: 5]
荒漠河岸林是长期适应干旱区内陆河流域环境的产物,具有独特的水分利用和生存策略.对黑河下游胡杨、柽柳等主要荒漠河岸林植物液流、水势、木质部导水率、根系水分再分配以及植物水分来源等的测试分析表明,在降水稀少、蒸发强烈的黑河下游,胡杨、柽柳主要利用的是深层土壤水和地下水,其中成熟胡杨主要吸收地下175~325 cm处土壤水和地下水供其蒸腾和生存所需;胡杨根系水力提升过程主要发生在10~70 cm土层,蒸散导致的土壤耗水量中有10%来自根系提升的水量;植物根和枝条木质部的初始导水率(Ks0)和最大导水率(Ks,max)相对较高,其中柽柳木质部的潜在导水能力比胡杨强;胡杨、柽柳液流速度呈单峰型,在午后达到峰值,黎明前水势无显著变化,反映未受到严重水分胁迫;黑河下游河道附近的地下水埋深大多在4m以内,基本可以满足荒漠河岸林植物的生长发育,黑河下游荒漠河岸林生态系统处于稳定和发展状态.
DOIURL [本文引用: 2]
Background/Question/Methods There has been much interest in ecological processes, including maintenance of species diversity, turnover rates, growth rates and carbon fluxes, in tropical forests, especially in the light of climate change. Tropical forests encompass a wide range of climatic regimes from semi-arid woodland to aseasonal rainforests. Comparative studies of tropical forest types... [Show full abstract]
DOIURL [本文引用: 4]
DOIURL [本文引用: 2]
Whilst the relationships between growth strategies and leaf traits are well established in functional plant ecology, little attention has been paid to root traits in very deep soil layers. The objective of our study was to compare the vertical velocity of the above- and belowground exploration of the environment for one of the fastest-growing tree species. Fine roots were studied in a chronosequence of intensively-managed Eucalyptus plantations established on highly weathered soils. Here we show that the root front depth was accurately predicted at 85% of mean tree height for stands <20 m in height, in the absence of any physical or chemical barrier. Tree height and root front growth velocities peaked at 0.59 and 0.55 m month?1 respectively 9-10 months after planting, and decreased steadily thereafter. Fast root front displacement might provide a competitive advantage to fast-growing species in forests established on deep soils. Our study may contribute to the debate on the environmental impact of short-rotation plantation forests in the Tropics. (R sum d'auteur)
DOIURL [本文引用: 4]
Although large amounts of potassium (K) are applied in tropical crops and planted forests, little is known about the interaction between K nutrition and water supply regimes on water resources in tropical regions. This interaction is a major issue because climate change is expected to increase the length of drought periods in many tropical regions and soil water availability in deep soil layers is likely to have a major influence on tree growth during dry periods in tropical planted forests. A process-based model (MAESPA) was parameterized in a throughfall exclusion experiment in Brazil to gain insight into the combined effects of K deficiency and rainfall reduction (37% throughfall exclusion) on the water used by the trees, soil water storage and water table fluctuations over the first 4.562years after planting Eucalyptus grandis trees. A comparison of canopy transpiration in each plot with the values predicted for the same soil with the water content maintained at field capacity, made it possible to calculate a soil-driven tree water stress index for each treatment. Compared to K-fertilized trees with undisturbed rainfall (+K+W), canopy transpiration was 40% lower for K deficiency (61K+W), 20% lower for W deficit (+K61W) and 36% lower for combined K deficiency and W deficit (61K61W) on average. Water was withdrawn in deeper soil layers for 61W than for +W, particularly over dry seasons. Under contrasted K availability, water withdrawal was more superficial for 61K than for +K. Mean soil water content down to 1862m below surface (mbs) was 24% higher for 61K+W than for +K+W from 262years after planting (after canopy closure), while it was 24% lower for +K61W and 12% lower for 61K61W than for +K+W. The soil-driven tree water stress index was 166% higher over the first 4.562years after planting for 61W than for +W, 76% lower for 61K than for +K, and 14% lower for 61K61W than for +K+W. Over the study period, deep seepage was higher by 37162mm62yr 611 (+122%) for 61K than for +K and lower by 20062mm62yr 611 (6166%) for 61W than for +W. Deep seepage was lower by 44% for 61K61W than for +K+W. At the end of the study period, the model predicted a higher water table for 61K (1062mbs for 61K+W and 1662mbs for 61K61W) than for +K (1662mbs for +K+W and 1862mbs for +K61W). Our study suggests that flexible fertilization regimes could contribute to adjusting the local trade-off between wood production and demand for soil water resources in planted forests.
DOIURL [本文引用: 8]
Climate models predict that the frequency, intensity and duration of drought events will increase in tropical regions. Although water withdrawal from deep soil layers is generally considered to be an efficient adaptation to drought, there is little information on the role played by deep roots in tropical forests. Tropical Eucalyptus plantations managed in short rotation cycles are simple forest ecosystems that may provide an insight into the water use by trees in tropical forests. The contribution made by water withdrawn from deep soil layers to the water required for evapotranspiration was quantified daily from planting to harvesting age for a Eucalyptus grandis stand using a soil water transfer model coupled with an ecophysiological forest model (MAESPA). The model was parameterized using an extensive data set and validated using time series of the soil water content down to a depth of 1002m and water‐table level, as well as evapotranspiration measured using eddy covariance. Fast root growth after planting provided access to large quantities of water stored in deep soil layers over the first 202years. Eucalyptus roots reached the water‐table at a depth of 1202m after 202years. Although the mean water withdrawal from depths of over 1002m amounted to only 5% of canopy transpiration from planting to a harvesting age of 502years, the proportion of water taken up near the water‐table was much higher during dry periods. The water‐table rose from 18 to 1202m below‐ground over 202years after the harvest of the previous stand and then fell until harvesting age as evapotranspiration rates exceeded the annual rainfall. Deep rooting is an efficient strategy to increase the amount of water available for the trees, allowing the uptake of transient gravitational water and possibly giving access to a deep water‐table. Deep soil layers have an important buffer role for large amounts of water stored during the wet season that is taken up by trees during dry periods. Our study confirms that deep rooting could be a major mechanism explaining high transpiration rates throughout the year in many tropical forests.
DOIURLPMID [本文引用: 1]
Geneticists and breeders are positioned to breed plants with root traits that improve productivity under drought. However, a better understanding of root functional traits and how traits are related to whole plant strategies to increase crop productivity under different drought conditions is needed. Root traits associated with maintaining plant productivity under drought include small fine root diameters, long specific root length, and considerable root length density, especially at depths in soil with available water. In environments with late season water deficits, small xylem diameters in targeted seminal roots save soil water deep in the soil profile for use during crop maturation and result in improved yields. Capacity for deep root growth and large xylem diameters in deep roots may also improve root acquisition of water when ample water at depth is available. Xylem pit anatomy that makes xylem less eaky and prone to cavitation warrants further exploration holding promise that such traits may improve plant productivity in water-limited environments without negatively impacting yield under adequate water conditions. Rapid resumption of root growth following soil rewetting may improve plant productivity under episodic drought. Genetic control of many of these traits through breeding appears feasible. Several recent reviews have covered methods for screening root traits but an appreciation for the complexity of root systems (e.g., functional differences between fine and coarse roots) needs to be paired with these methods to successfully identify relevant traits for crop improvement. Screening of root traits at early stages in plant development can proxy traits at mature stages but verification is needed on a case by case basis that traits are linked to increased crop productivity under drought. Examples in lesquerella (Physaria) and rice (Oryza) show approaches to phenotyping of root traits and current understanding of root trait genetics for breeding.
DOIURL [本文引用: 1]
The spatial and temporal evolution of soil water content (θ) in Quercus ilex dehesas has been investigated to determine how trees modify the soil water dynamics and the nature of tree-grass interactions in terms of soil water use in these ecosystems. Soil physical parameters and θ were measured at different distances from the tree trunk (2–30 m) in the upper 300 cm of soil. θ was measured monthly by TDR during 2002–2005. Tree water potential was determined during the summers of 2004 and 2005. At deeper soil layers, mean θ values were higher beyond than beneath tree canopy during dry periods. θ depletion beyond tree canopy continued even in summer, when herbaceous plants dried up, suggesting that trees uptake water from the whole inter-tree space. Results have shown a high dependence of trees on deep water reserves throughout late spring and summer, which helps to avoid competition for water with herbaceous vegetation.
DOIURL [本文引用: 1]
1. Little is known about the role of deep roots in the nutrition of forest trees and their ability to provide a safety-net service taking up nutrients leached from the topsoil.2. To address this issue, we studied the potential uptake of N, K and Ca by Eucalyptus grandis trees (6 years of age 25 m mean height), in Brazil, as a function of soil depth, texture and water content. We injected -15N, Rb+ (analogue of K+) and Sr2+ (analogue of Ca2+) tracers simultaneously in a solution through plastic tubes at 10, 50, 150 and 300 cm in depth in a sandy and a clayey Ferralsol soil. A complete randomized design was set up with three replicates of paired trees per injection depth and soil type. Recently expanded leaves were sampled at various times after tracer injection in the summer, and the experiment was repeated in the winter. Soil water contents were continuously monitored at the different depths in the two soils.3. Determination of foliar Rb and Sr concentrations and 15N atom % made it possible to estimate the relative uptake potential (RUP) of tracer injections from the four soil depths and the specific RUP (SRUP), defined as RUP, per unit of fine root length density in the corresponding soil layer.4. The highest tracer uptake rates were found in the topsoil, but contrasting RUP distributions were observed for the three tracers. Whilst the RUP was higher for -15N than for Rb+ and Sr2+ in the upper 50 cm of soil, the highest SRUP values for Sr2+ and Rb+ were found at a depth of 300 cm in the sandy soil, as well as in the clayey soil when gravitational solutions reached that depth.5. Our results suggest that the fine roots of E. grandis trees exhibit contrasting potential uptake rates with depth depending on the nutrient. This functional specialization of roots might contribute to the high growth rates of E. grandis trees, efficiently providing the large amounts of nutrients required throughout the development of these fast-growing plantations.
DOIURL [本文引用: 2]
http://www.nature.com/doifinder/10.1038/350335a0
DOIURLPMID [本文引用: 1]
To date, reasons for the increase in liana abundance and biomass in the Neotropics are still unclear. One proposed hypothesis suggests that lianas, in comparison with trees, are more adaptable to drought conditions. Moreover, previous studies have assumed that lianas have a deeper root system, which provides access to deeper soil layers, thereby making them less susceptible to drought stress. The dual stable water isotope approach (未18O and 未2H) enables below-ground vegetation competition for water to be studied. Based on the occurrence of a natural gradient in soil water isotopic signatures, with enriched signatures in shallow soil relative to deep soil, the origin of vegetation water sources can be derived. Our study was performed on canopy trees and lianas reaching canopy level in tropical forests of French Guiana. Our results show liana xylem water isotopic signatures to be enriched in heavy isotopes in comparison with those from trees, indicating differences in water source depths and a more superficial root activity for lianas during the dry season. This enables them to efficiently capture dry season precipitation. Our study does not support the liana deep root water extraction hypothesis. Additionally, we provide new insights into water competition between tropical canopy lianas and trees. Results suggest that this competition is mitigated during the dry season due to water resource partitioning.
DOIURL [本文引用: 1]
Root-specific responses to stress are not well-known, and have been largely based on indirect measurements of bulk soil water extraction, which limits mechanistic modeling of root function.Here, we u
DOIURL [本文引用: 2]
Populus tomentosadevelopeda deep root system, wide-spreading and dense lateral roots, and structural roots exploring both deep and shallow soil resources to adapt to intense seasonal groundwater table
DOIURLPMID [本文引用: 1]
Hydraulic redistribution (HR), the passive movement of water via roots from moist to drier portions of the soil, occurs in many ecosystems, influencing both plant and ecosystem-water use. We examined the effects of HR on root hydraulic functioning during drought in young and old-growth Douglas-fir [Pseudotsuga menziesii (Mirb.) Franco] and ponderosa pine (Pinus ponderosa Dougl. Ex Laws) trees growing in four sites. During the 2002 growing season, in situ xylem embolism, water deficit and xylem vulnerability to embolism were measured on medium roots (2-4-mm diameter) collected at 20-30 cm depth. Soil water content and water potentials were monitored concurrently to determine the extent of HR. Additionally, the water potential and stomatal conductance$(g_{\rm{s}} )$of upper canopy leaves were measured throughout the growing season. In the site with young Douglas-fir trees, root embolism increased from 20 to 55 percent loss of conductivity (PLC) as the dry season progressed. In young ponderosa pine, root embolism increased from 45 to 75 PLC. In contrast, roots of old-growth Douglas-fir and ponderosa pine trees never experienced more than 30 and 40 PLC, respectively. HR kept soil water potential at 20-30 cm depth above -0.5 MPa in the old-growth Douglas-fir site and -1.8 MPa in the old-growth ponderosa pine site, which significantly reduced loss of shallow root function. In the young ponderosa pine stand, where little HR occurred, the water potential in the upper soil layers fell to about -2.8 MPa, which severely impaired root functioning and limited recovery when the fall rains returned. In both species, daily maximum$(g_{\rm{s}} )$decreased linearly with increasing root PLC, suggesting that root xylem embolism acted in concert with stomata to limit water loss, thereby maintaining minimum leaf water potential above critical values. HR appears to be an important mechanism for maintaining shallow root function during drought and preventing total stomatal closure.
DOIURLPMID [本文引用: 19]
Plant rooting depth affects ecosystem resilience to environmental stress such as drought. Deep roots connect deep soil/groundwater to the atmosphere, thus influencing the hydrologic cycle and climate. Deep roots enhance bedrock weathering, thus regulating the long-term carbon cycle. However, we know little about how deep roots go and why. Here,...
In:
DOIURL [本文引用: 2]
Water is one of the most critical resources limiting plant growth and crop productivity, and root water uptake is an important aspect of plant physiology governing plant water use and stress tolerance
DOIURL [本文引用: 5]
Background and Aims Fine roots play a major role in the global carbon cycle through respiration, exudation and decomposition processes, but their dynamics are poorly understood. Current
[本文引用: 2]
DOIURL [本文引用: 2]
react-text: 235 Phytomers are basic morphological units of plants. Knowledge of phytomer development is essential for understanding morphological plasticity, functional-structural modelling of plant growth, and the usage of leaf characteristics to indicate growth conditions at the time of production (e.g. stable isotope signals). Yet, systematic analysis on the process of phytomer development is unavailable... /react-text react-text: 236 /react-text [Show full abstract]
DOIURL [本文引用: 3]
Oil palm (Elaeis guineensis Jacq.) plantations cover large and continuously increasing areas of humid tropical lowlands. Landscapes dominated by oil palms usually consist of a mosaic of mono-cultural, homogeneous stands of varying age, which may be heterogeneous in their water use characteristics. However, studies on the water use characteristics of oil palms are still at an early stage and... [Show full abstract]
DOIURL [本文引用: 1]
Many areas in the world are characterized by shallow soils underlain by weathered bedrock, but root-rock interactions and their implications for regolith weathering are poorly understood. To test the role of tree roots in weathering bedrock, we excavated four pits along a catena in a shale-dominated catchment at the Susquehanna Shale Hills Critical Zone Observatory (SSHCZO) in central Pennsylvania. We measured a variety of biological, physical, and chemical properties including: 1) root density, distribution, and respiration, 2) soil gas, and 3) elemental compositions, mineralogy, and morphology of soil, rock, and rock fracture fill at ridge top, mid-slope, toe-slope, and valley floor sites. As expected, root density declined rapidly with depth; nevertheless, fine roots were present in rock fractures even in the deepest, least weathered shale sampled (~ 180 cm below the land surface). Root densities in shale fractures were comparable between the ridge top and mid-slope pits. However, they were significantly lower in the toe-slope, despite increasing rock fracture densities, which is likely due to a shallower water table depth at the downslope site. Average root respiration (per mass of dry root tissue) in rock fractures was comparable to rates in the soil. Thus, the total flux of CO 2 from root respiration tracked root densities, decreasing with depth. Potential microbial respiration, estimated as the laboratory C mineralization potential, was about an order of magnitude lower than measured root respiration in both the soil and shale fractures. Roots were only observed in large aperture (> 50 m) shale fractures that were filled with particulate material. The fill in these fractures was mineralogically and geochemically similar to the lowest soil horizons with respect to clay composition, element mobility, extractable dissolved organic C (DOC), inorganic N-species, and potentially mineralizable C and N, while total C and total N values for the fracture fill were similar to the shale bedrock. In the bulk soil, depletion profiles (Al, Fe, K, Mg, and Si) relative to unweathered shale reflected characteristic weathering of illite and vermiculized chlorite to kaolinite and are similar between soils and fracture fill. Such similarities indicate that the fracture coatings are likely the result of pedogenic processes that occur at depth in the fractures rather than translocation of soil particles downward into the fractures. Overall, our data suggest that roots and fill in shale fractures down to ~ 180 cm are qualitatively similar to those in surface soil horizons. Thus, the deepest manifestation of the chemical depletion profiles observed in the pits consists of the rock fracture fill, and this fill is present at low concentrations with similarly low concentrations of fine roots.
[本文引用: 1]
DOIURL [本文引用: 2]
了解林农复合系统的种间水分关系至关重要。该文通过稳定氘同位素研究了华北低丘山区核桃(Juglans regia)-菘蓝(Isatis tinctoria)/决明(Senna tora)复合系统各组分的水分来源,试图明确该果药复合系统的种间水分利用策略,为该区林农配置模式的选择提供理论依据。研究结果表明:果药复合系统的土壤含水量明显高于单作菘蓝和单作决明地块,在2012年、2013年上半年比单作菘蓝高26.74%和7.93%,在下半年比单作决明高17.39%和13.65%。在果药复合系统内部,土壤含水量以核桃树行中间位置的最低、树行北侧和树下最高。在各个土层深度,单作系统的土壤水氢稳定同位素比率(δD值)均比复合系统的高。在菘蓝生长时期的春旱期,复合系统中核桃的大部分水分来源于30–80 cm深层土壤水,表明此时期核桃表层根系活性不高;而决明生长时期正值雨季,此时核桃优先利用雨水补充的0–30 cm浅层土壤水、表层根系活性增强。在任何生长时期,菘蓝和决明85%以上的水分都来自浅层土壤水。在菘蓝苗期,其根系尚未扎入深层土壤中,单作菘蓝的水分完全来源于浅层土壤,而在2012年间作菘蓝却有5.7%的水分来自于深层土壤,在更为干旱的2013年该比例上升到9.7%,该结果证实了核桃在旱季存在"水力提升"作用,供浅根系作物吸收利用,并且越干旱,该水力提升作用越强。在华北低丘山区核桃-菘蓝/决明复合系统中,深根性核桃改善了复合系统的土壤水分状况,在旱季主要利用深层土壤水以避开与浅层作物的水分竞争、并能将深层土壤水提升至浅层土壤供菘蓝吸收利用,核桃与两种药材表现为水分互利关系,因而该模式适合在该地区发展。
DOIURL [本文引用: 2]
了解林农复合系统的种间水分关系至关重要。该文通过稳定氘同位素研究了华北低丘山区核桃(Juglans regia)-菘蓝(Isatis tinctoria)/决明(Senna tora)复合系统各组分的水分来源,试图明确该果药复合系统的种间水分利用策略,为该区林农配置模式的选择提供理论依据。研究结果表明:果药复合系统的土壤含水量明显高于单作菘蓝和单作决明地块,在2012年、2013年上半年比单作菘蓝高26.74%和7.93%,在下半年比单作决明高17.39%和13.65%。在果药复合系统内部,土壤含水量以核桃树行中间位置的最低、树行北侧和树下最高。在各个土层深度,单作系统的土壤水氢稳定同位素比率(δD值)均比复合系统的高。在菘蓝生长时期的春旱期,复合系统中核桃的大部分水分来源于30–80 cm深层土壤水,表明此时期核桃表层根系活性不高;而决明生长时期正值雨季,此时核桃优先利用雨水补充的0–30 cm浅层土壤水、表层根系活性增强。在任何生长时期,菘蓝和决明85%以上的水分都来自浅层土壤水。在菘蓝苗期,其根系尚未扎入深层土壤中,单作菘蓝的水分完全来源于浅层土壤,而在2012年间作菘蓝却有5.7%的水分来自于深层土壤,在更为干旱的2013年该比例上升到9.7%,该结果证实了核桃在旱季存在"水力提升"作用,供浅根系作物吸收利用,并且越干旱,该水力提升作用越强。在华北低丘山区核桃-菘蓝/决明复合系统中,深根性核桃改善了复合系统的土壤水分状况,在旱季主要利用深层土壤水以避开与浅层作物的水分竞争、并能将深层土壤水提升至浅层土壤供菘蓝吸收利用,核桃与两种药材表现为水分互利关系,因而该模式适合在该地区发展。
URL [本文引用: 2]
以北京永定河沙地107杨人工幼林为研究对象,比较滴灌和常规沟灌对杨树生长的影响。结果表明:滴灌栽培107杨,2、3年生幼龄林的蓄积量分别为13.37、41.15 m3/hm2,是沟灌的2.22、1.68倍。滴灌能明显促进107杨各器官的生长,增加生物产量,滴灌栽培使3年生杨树单株的干生物量为沟灌的2.42倍,并且滴灌还能提高107杨幼林体内含水率。滴灌栽培107杨幼林2、3年生的水分利用率分别是0.84和3.89 m3/hm2,分别是沟灌的1.91倍和2.84倍。滴灌能减少107杨幼林田间养分的流失,显著提高107杨幼林田间各养分利用率,同时对107杨单株植物各养分利用率也有一定提升。
URL [本文引用: 2]
以北京永定河沙地107杨人工幼林为研究对象,比较滴灌和常规沟灌对杨树生长的影响。结果表明:滴灌栽培107杨,2、3年生幼龄林的蓄积量分别为13.37、41.15 m3/hm2,是沟灌的2.22、1.68倍。滴灌能明显促进107杨各器官的生长,增加生物产量,滴灌栽培使3年生杨树单株的干生物量为沟灌的2.42倍,并且滴灌还能提高107杨幼林体内含水率。滴灌栽培107杨幼林2、3年生的水分利用率分别是0.84和3.89 m3/hm2,分别是沟灌的1.91倍和2.84倍。滴灌能减少107杨幼林田间养分的流失,显著提高107杨幼林田间各养分利用率,同时对107杨单株植物各养分利用率也有一定提升。
DOIURL [本文引用: 1]
Tree roots contribute to the resistance of riparian sediments to physical deformation and disintegration. Understanding reinforcement by roots requires information on root distributions within riparian soils and sediments. Continuous-depth models or curves have been proposed to describe vertical root density variations, providing useful indicators of the types of function that may be appropriate to riparian trees, but have generally been estimated for terrestrial species or broad vegetation types rather than riparian species or environments.We investigated vertical distributions of roots >0.165mm diameter of a single riparian tree species (Populus nigra L.) along the middle reaches of a single river (Tagliamento River, Italy), where Populus nigra dominates the riparian woodland. Root density (hundreds m612) and root area ratio (RAR in cm2 m612) were measured within 1065cm depth increments of 24 excavated bank profiles across nine sites. Sediment samples, extracted from distinct strata within the profiles, were analysed for moisture content, organic matter content and particle size. Statistical analyses identified two groups of wetter and drier profiles and five sediment types.Following loge-transformation of root density and RAR, linear regression analysis explored their variation with depth and, using dummy variables, any additional influence of moisture and sediment type. Significant linear regression relationships were estimated between both root density and RAR and depth which explained only 15% and 8% of the variance in the data. Incorporating moisture and then sediment characteristics into the analysis increased the variance explained in root density to 29% and 36% and in RAR to 14% and 26%.We conclude that riparian tree root density and RAR are highly spatially variable and are poorly explained by depth alone. Complex riparian sedimentary structures and moisture conditions are important influences on root distributions and so need to be incorporated into assessments of the contribution of roots to river bank reinforcement. Copyright 08 2016 John Wiley & Sons, Ltd.
DOIURLPMID [本文引用: 1]
In a 3-year study, seasonal and daily soil water fluctuations in a California blue oak woodland were investigated by measuring soil water potential ($\Psi _{\text{s}}$) at hourly intervals. Soil water potential remained relatively high well into the annual summer drought, with values above -0.5 MPa until June even in a dry year. As drought progressed,$\Psi _{\text{s}}$(at 25, 50, 75, and 100 cm depth) decreased to less than -3 MPa, providing evidence for continued blue oak root activity throughout the summer. We observed diurnal$\Psi _{\text{s}}$fluctuations (gradual increase at night and rapid decrease during daytime) characteristic of hydraulic lift, a process by which plant roots redistribute water from wet to dry soil layers. These diurnal fluctuations were observed at all four soil depths and began to appear when$\Psi _{\text{s}}$reached approximately -0.3 MPa. When$\Psi _{\text{s}}$reached approximately -3 MPa, fluctuations became "offset" from those typical of hydraulic lift. These offset fluctuations (apparent at low water potentials when temperature fluctuations were large) closely followed diurnal fluctuations in soil temperature. We propose that these offset patterns resulted from a combination of hydraulic lift cessation and an over-correction for temperature in the model used to calculate$\Psi _{\text{s}}$from raw sensor data. The appearance and disappearance of hydraulic lift fluctuations seemed to depend on$\Psi _{\text{s}}$. While soil temperatures and dates at which hydraulic lift appeared (and disappeared) were significantly different between wet and dry years,$\Psi _{\text{s}}$values associated with hydraulic lift appearance were not significantly different. Hydraulic lift occurred too late in summer to benefit annual forage grasses. However, water released by blue oak trees at night could slow the rate of soil water depletion and extend blue oaks' growing season.
DOIURLPMID [本文引用: 3]
Global biogeochemical models have improved dramatically in the last decade in their representation of the biosphere. Although leaf area data are an important input to such models and are readily available globally, global root distributions for modeling water and nutrient uptake and carbon cycling have not been available. This analysis provides global distributions for fine root biomass, length, and surface area with depth in the soil, and global estimates of nutrient pools in fine roots. Calculated root surface area is almost always greater than leaf area, more than an order of magnitude so in grasslands. The average C:N:P ratio in living fine roots is 450:11:1, and global fine root carbon is more than 5% of all carbon contained in the atmosphere. Assuming conservatively that fine roots turn over once per year, they represent 33% of global annual net primary productivity.
DOIURL [本文引用: 1]
This study was performed on hybrid aspen saplings growing at the Free Air Humidity Manipulation site in Estonia. We investigated changes in wood anatomy and hydraulic conductivity in response to increased air humidity. Two hydraulic traits (specific conductivity and leaf-specific conductivity) and four anatomical traits of stem wood65–65relative vessel area (VA), vessel density (VD), pit area and pit aperture area65–65were influenced by the humidity manipulation. Stem hydraulic traits decreased in the apical direction, whereas branch hydraulic characteristics tended to be greatest in mid-canopy, associated with branch size. A reduction in VD due to increasing humidity was accompanied by a decrease in vessel lumen diameter, hydraulically weighted mean diameter (Dh), xylem vulnerability index and theoretical hydraulic conductivity. VA and Dh combined accounted for 87.4% of the total variation in kt of branches and 85.5% of that in stems across the treatments. Characters of branch vessels were more stable, and only the vessel-grouping index (the ratio of the total number of vessels to the total number of vessel groupings) was dependent on the interactive effect of the treatment and canopy position. Our results indicate that the increasing atmospheric humidity predicted for high latitudes will result in moderate changes in the structure and functioning of the hybrid aspen xylem.
DOIURL [本文引用: 1]
Leaf is the most important organ for carbon-water coupling of a plant because it is the primary medium for photosynthesis. It also acts as the hydraulic bottleneck and safety valve against hydraulic catastrophic dysfunctions. The leaf economics spectrum, which reflects the balance between investments and returns of leaf economic traits, provides a useful framework for examining species strategies as shaped by their evolutionary history. Changes in leaf hydraulic traits will influence leaf economic traits as well as plant survival and growth. Exploring trade-offs between leaf hydraulic and economic traits is thus of significance for modeling carbon-water relations, understanding the mechanisms of water/carbon investments, and extending the leaf economic spectrum. In this review, we first examined the trade-offs between leaf hydraulic and economic traits. Specially, we analyzed the relationships between leaf hydraulic conductivity and hydraulic vulnerability, water potential at the turgor loss point, water capacitance, safety margin, and leaf morphological, structural and functional traits. We then discussed potential mechanisms regulating leaf hydraulic and economic traits from leaf morphology, anatomy, venation, and stomatal functions. Finally, we proposed future research to: (1) develop an integrated whole-plant economics spectrum, including carbon-nitrogen-water resources and root-stem-leaf hydraulic transport system that will help revealing ecophysiological mechanisms of plant structure-functional coupling, carbon sequestration and water use; (2) explore a generalized trade-offs among leaf hydraulic safety, hydraulic efficiency and carbon fixation efficiency to advance our understanding of the relationships between biophysical structure and physiological metabolism in plant leaf construction under drought stress; and (3) explore the carbon-water metabolic relationship and coupling of water transport and growth rate for the metabolic theory and predictions at community scale.
DOIURL [本文引用: 1]
Leaf is the most important organ for carbon-water coupling of a plant because it is the primary medium for photosynthesis. It also acts as the hydraulic bottleneck and safety valve against hydraulic catastrophic dysfunctions. The leaf economics spectrum, which reflects the balance between investments and returns of leaf economic traits, provides a useful framework for examining species strategies as shaped by their evolutionary history. Changes in leaf hydraulic traits will influence leaf economic traits as well as plant survival and growth. Exploring trade-offs between leaf hydraulic and economic traits is thus of significance for modeling carbon-water relations, understanding the mechanisms of water/carbon investments, and extending the leaf economic spectrum. In this review, we first examined the trade-offs between leaf hydraulic and economic traits. Specially, we analyzed the relationships between leaf hydraulic conductivity and hydraulic vulnerability, water potential at the turgor loss point, water capacitance, safety margin, and leaf morphological, structural and functional traits. We then discussed potential mechanisms regulating leaf hydraulic and economic traits from leaf morphology, anatomy, venation, and stomatal functions. Finally, we proposed future research to: (1) develop an integrated whole-plant economics spectrum, including carbon-nitrogen-water resources and root-stem-leaf hydraulic transport system that will help revealing ecophysiological mechanisms of plant structure-functional coupling, carbon sequestration and water use; (2) explore a generalized trade-offs among leaf hydraulic safety, hydraulic efficiency and carbon fixation efficiency to advance our understanding of the relationships between biophysical structure and physiological metabolism in plant leaf construction under drought stress; and (3) explore the carbon-water metabolic relationship and coupling of water transport and growth rate for the metabolic theory and predictions at community scale.
DOIURL [本文引用: 1]
Key message Deep root hydraulic conductance is upregulated during severe drought and is associated with upregulation in aquaporin activity.
DOIURLPMID [本文引用: 4]
ABSTRACT Background and AimsDespite the importance of vessels in angiosperm roots for plant water transport, there is little research on the microanatomy of woody plant roots. Vessels in roots can be interconnected networks or nearly solitary, with few vessel-vessel connections. Species with few connections are common in arid habitats, presumably to isolate embolisms. In this study, measurements were made of root vessel pit sizes, vessel air-seeding pressures, pit membrane thicknesses and the degree of vessel interconnectedness in deep (approx. 20 m) and shallow (500 m) and more large pits than either shallow Q. fusiformis roots or S. lanuginosum roots. Vessel air-seeding pressures were approximately four times greater in Q. fusiformis than in S. lanuginosum and 1 3-1 9 times greater in shallow roots than in deep roots. Sideroxylon lanuginosum had 34-44 % of its vessels interconnected, whereas Q. fusiformis only had 1-6 % of its vessels connected. Vessel air-seeding pressures were unrelated to pit membrane thickness but showed a positive relationship with vessel interconnectedness.Conclusions These data support the hypothesis that species with more vessel-vessel integration are often less resistant to embolism than species with isolated vessels. This study also highlights the usefulness of tomography for vessel network analysis and the important role of 3-D xylem organization in plant hydraulic function.
DOIURLPMID [本文引用: 3]
Abstract Water transport from soils to the atmosphere is critical for plant growth and survival. However, we have a limited understanding about many portions of the whole-tree hydraulic pathway, because the vast majority of published information is on terminal branches. Our understanding of mature tree trunk hydraulic physiology, in particular, is limited. The hydraulic vulnerability segmentation hypothesis (HVSH) stipulates that distal portions of the plant (leaves, branches and roots) should be more vulnerable to embolism than trunks, which are nonredundant organs that require a massive carbon investment. In the current study, we compared vulnerability to loss of hydraulic function, leaf and xylem water potentials and the resulting hydraulic safety margins (in relation to the water potential causing 50% loss of hydraulic conductivity) in leaves, branches, trunks and roots of four angiosperms and four conifer tree species. Across all species, our results supported strongly the HVSH as leaves and roots were less resistant to embolism than branches or trunks. However, branches were consistently more resistant to embolism than any other portion of the plant, including trunks. Also, calculated whole-tree vulnerability to hydraulic dysfunction was much greater than vulnerability in branches. This was due to hydraulic dysfunction in roots and leaves at less negative water potentials than those causing branch or trunk dysfunction. Leaves and roots had narrow or negative hydraulic safety margins, but trunks and branches maintained positive safety margins. By using branch-based hydraulic information as a proxy for entire plants, much research has potentially overestimated embolism resistance, and possibly drought tolerance, for many species. This study highlights the necessity to reconsider past conclusions made about plant resistance to drought based on branch xylem only. This study also highlights the necessity for more research of whole-plant hydraulic physiology to better understand strategies of plant drought tolerance and the critical control points within the hydraulic pathway. The Author 2016. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
DOIURLPMID [本文引用: 6]
Despite their importance for water uptake and transport, the xylem anatomical and hydraulic properties of tree roots have only rarely been studied in the field. We measured mean vessel diameter (D), vessel density (VD), relative vessel lumen area (lumen area per xylem area) and derived potential hydraulic conductivity (Kp) in the xylem of 197 fine- to medium-diameter roots (1–10 mm) in the topsoil and subsoil (0–200 cm) of a mature European beech forest on sandy soil for examining the influence of root diameter and soil depth on xylem anatomical and derived hydraulic traits. All anatomical and functional traits showed strong dependence on root diameter and thus root age but no significant relation to soil depth. Averaged over topsoil and deep soil and variable flow path lengths in the roots,Dincreased linearly with root diameter from 6550 μm in the smallest diameter class (1–2 mm) to 6570 μm in 6–7 mm roots (corresponding to a mean root age of 6512 years), but remained invariant in roots >7 mm.Dnever exceeded 6582 μm in the 1–10 mm roots, probably in order to control the risk of frost- or drought-induced cavitation. This pattern was overlain by a high variability in xylem anatomy among similar-sized roots withKpshowing a higher variance component within than between root diameter classes. With 8% of the roots exceeding averageKpin their diameter class by 50–700%, we obtained evidence of the existence of ‘high-conductivity roots’ indicating functional differentiation among similar-sized roots. We conclude that the hydraulic properties of small to medium diameter roots of beech are mainly determined by root age, rendering root diameter a suitable predictor of hydraulic functioning, while soil depth – without referring to path length – had a negligible effect.
DOIURLPMID [本文引用: 1]
We outline a method of inferring rooting depth from a Terrestrial Biosphere Model by maximizing the benefit of the vegetation within the model. This corresponds to the evolutionary principle that vegetation has adapted to make best use of its local environment. We demonstrate this method with a simple coupled biosphere/soil hydrology model and find that deep rooted vegetation is predicted in most parts of the tropics. Even with a simple model like the one we use, it is possible to reproduce biome averages of observations fairly well. By using the optimized rooting depths global Annual Net Primary Production (and transpiration) increases substantially compared to a standard rooting depth of one meter, especially in tropical regions that have a dry season. The decreased river discharge due to the enhanced evaporation complies better with observations. We also found that the optimization process is primarily driven by the water deficit/surplus during the dry/wet season for humid and arid regions, respectively. Climate variability further enhances rooting depth estimates. In a sensitivity analysis where we simulate changes in the water use efficiency of the vegetation we find that vegetation with an optimized rooting depth is less vulnerable to variations in the forcing. We see the main application of this method in the modelling communities of land surface schemes of General Circulation Models and of global Terrestrial Biosphere Models. We conclude that in these models, the increased soil water storage is likely to have a significant impact on the simulated climate and the carbon budget, respectively. Also, effects of land use change like tropical deforestation are likely to be larger than previously thought.
DOIURL [本文引用: 1]
Delineating the characteristics of biosphere-atmosphere exchange in rubber (Hevea brasiliensisMüll. Arg.) plantations, which are rapidly expanding throughout mainland Southeast Asia, is important to understanding the impacts of the land-use change on environmental processes. In attempt to shed new light on the impacts of conversion to rubber, we have conducted eddy flux measurements over a 3-year period in two rubber plantation sites: (1) Som Sanuk, located in northeastern Thailand; and (2) Cambodian Rubber Research Institute (CRRI), located in central Cambodia. Both sites have a distinct dry season. We used a combination of actual evapotranspiration (ET) flux measurements and an inverted version of a simple 2-layerETmodel for estimating the mean canopy stomatal conductance (gs). The potential water balance (precipitation (P)61potential evaporation (ET_POT)) for each season (i.e., December–February: DJF, March–May: MAM, June–August: JJA, and September–November: SON) revealed when and how the water use is controlled. In the seasons when actual water balance (P61ET) was negative (DJF and MAM), the deficit was compensated with soil water from the previous season at depths of 0–2m (Thailand site) and 0–3m (Cambodia site). At both sites, the reference value ofgs(gsref) and the sensitivity ofgsto atmospheric demand (m) appeared to be less in DJF and MAM than each in the other two 3-month periods (seasons). On average, in a whole year,m/gsrefwas less in Thailand (<<0.6) than in Cambodia (near 0.6 for part of the year), suggesting that there was less sufficient stomatal regulation at the Thailand site, where there might be little risk of water stress-induced hydraulic failure because of its higher annual rainfall amount. In comparison, at CRRI where annualP61ET_POTwas negative, there was stricter stomatal regulation, preventing excessive xylem cavitation.
DOIURLPMID [本文引用: 1]
Although highly weathered soils cover considerable areas in tropical regions, little is known about exploration by roots in deep soil layers. Intensively managedEucalyptusplantations are simple forest ecosystems that can provide an insight into the belowground growth strategy of fast-growing tropical trees. Fast exploration of deep soil layers by eucalypt fine roots may contribute to achieving a gross primary production that is among the highest in the world for forests. Soil exploration by fine roots down to a depth of 10 m was studied throughout the complete cycle inEucalyptus grandisplantations managed in short rotation. Intersects of fine roots, less than 1 mm in diameter, and medium-sized roots, 1 3 mm in diameter, were counted on trench walls in a chronosequence of 1-, 2-, 3.5-, and 6-year-old plantations on a sandy soil, as well as in an adjacent 6-year-old stand growing in a clayey soil. Two soil profiles were studied down to a depth of 10 m in each stand (down to 6 m at ages 1 and 2 years) and 4 soil profiles down to 1.5 3.0 m deep. The root intersects were counted on 224 m2of trench walls in 15 pits. Monitoring the soil water content showed that, after clear-cutting, almost all the available water stored down to a depth of 7 m was taken up by tree roots within 1.1 year of planting. The soil space was explored intensively by fine roots down to a depth of 3 m from 1 year after planting, with an increase in anisotropy in the upper layers throughout the rotation. About 60% of fine root intersects were found at a depth of more than 1 m, irrespective of stand age. The root distribution was isotropic in deep soil layers and kriged maps showed fine root clumping. A considerable volume of soil was explored by fine roots in eucalypt plantations on deep tropical soils, which might prevent water and nutrient losses by deep drainage after canopy closure and contribute to maximizing resource uses.
DOIURL [本文引用: 1]
Sugarcane crops are managed over 8 million hectares in Brazil and future extensions might occur on less favorable lands where irrigation would be necessary to increase and stabilize yields. Root growth was studied by sequential soil coring under rainfed and irrigated conditions for one cultivar widely planted in Brazil. Root length densities (RLD) were measured 34, 49, 125, 179, 241 and 322 days after planting (DAP) down to a depth of 1 m. At the harvest (332 DAP), root intersects (a proxy for RLD) were counted on two vertical trench walls in each water supply regime, down to a depth of 6.0 m. The highest RLD in deep layers (below a depth of 0.6 m) were observed in the rainfed crop from 125 DAP onwards. By contrast, the highest RLD in the upper layers during dry periods were found in the irrigated crop. The maximum depth reached by roots at the harvest was little affected by irrigation: 4.70 m and 4.25 m in the rainfed and irrigated crop, respectively. About 50% of root intersects were observed below the depth of 1 m in the two water supply regimes. This pattern suggested a strong genetic control of root growth in deep soil layers. The total amount of root intersects 332 DAP was 49% higher in the rainfed crop than in the irrigated crop, and root distribution was more homogeneous. Mean root front velocity was about 0.5 cm day 611 the first 4 months after planting and increased thereafter up to the end of the harvest (1.86 cm day 611 and 1.75 cm day 611 on average in the rainfed and the irrigated crops, respectively). Our study pointed out the necessity to take into account the development of sugarcane roots in deep soil layers to improve our understanding of net primary production control by water availability.
DOIURL [本文引用: 3]
Background and aims While the role of deep roots in major ecosystem services has been shown for tropical forests, there have been few direct measurements of fine root dynamics at depths of more than...
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
Extreme temperatures and droughts in the wake of climate change potentially threaten plant diversity. A strategy that plants use to improve survival during seasonal drought is to establish deep roots, aptly named tap roots for their ability to tap into groundwater. Little is known, however, about the role of deep roots in maintaining plant diversity. Here, we extend an established model of plants canopies by Iwasa et al. (1985), in which plants of different heights compete for light, to allow strategic investments in an optional tap root. We investigate how emerging diversity varies with water table depth, soil water gradient and drought-induced mortality rate. Having a tap root enables plants to reach deep water, thus reducing mortality, but also carries a construction cost, thus inducing a tradeoff. We find (1) that tap roots maintain plant diversity under increasing drought mortality, (2) that tap roots evolve when ground water is accessible at low to intermediate depths, (3) no viable strategies at high drought mortality and deep water table, and (4) Red Queen evolutionary dynamics in mixed communities with and without tap root.
DOIURL [本文引用: 3]
Soil dry layer has occurred widely in artificial economic forest in Loess region, which constrained vegetation restoration and reconstruction. In order to investigate deep soil depletion (below 2m) in densely planted jujube (Ziziphus jujube Mill.) orchard, soil coring method (Luoyang shovel) was used to get soil water content from surface to the maximum fine rooting depth at 0.2m soil interval. The results showed that root water uptake had a tendency to extend gradually deep, deep soil water depletion was 0, 29.6, 149.9 and 155.7mm in 2, 4, 9 and 12 year-old jujube plantation, deep soil water supply was 203.7, 167.7, 35.5 and 29.7mm, respectively, annual deep soil water depletion accounted for 0,2.3%,5.1% and 3.9% of annual soil water supply. There was almost no available water in 2~4m soil layer when the jujube plantation developed for 9 years. Jujube water requirement could not be met by the existing rainfall and drip irrigation amount. Soil dry layer extent and depth can be evaluated more accurately with the maximum rainfall infiltration depth and maximum fine rooting depth as upper and lower boundaries. Soil water content could be increased through water catchment, water conservation and water supplement, which played an important role in alleviating jujube plantation deep soil drying.
DOIURL [本文引用: 3]
Soil dry layer has occurred widely in artificial economic forest in Loess region, which constrained vegetation restoration and reconstruction. In order to investigate deep soil depletion (below 2m) in densely planted jujube (Ziziphus jujube Mill.) orchard, soil coring method (Luoyang shovel) was used to get soil water content from surface to the maximum fine rooting depth at 0.2m soil interval. The results showed that root water uptake had a tendency to extend gradually deep, deep soil water depletion was 0, 29.6, 149.9 and 155.7mm in 2, 4, 9 and 12 year-old jujube plantation, deep soil water supply was 203.7, 167.7, 35.5 and 29.7mm, respectively, annual deep soil water depletion accounted for 0,2.3%,5.1% and 3.9% of annual soil water supply. There was almost no available water in 2~4m soil layer when the jujube plantation developed for 9 years. Jujube water requirement could not be met by the existing rainfall and drip irrigation amount. Soil dry layer extent and depth can be evaluated more accurately with the maximum rainfall infiltration depth and maximum fine rooting depth as upper and lower boundaries. Soil water content could be increased through water catchment, water conservation and water supplement, which played an important role in alleviating jujube plantation deep soil drying.
DOIURL [本文引用: 1]
In order to investigate the relationship between deep soil water (below 2m) vertical variation and root distribution in the nine-year-old dense jujube plantation in Loess hilly region. The soil core method and oven drying method were used to determine fine root dry weight density and soil water content. Sequential cluster analysis method was adopted to stratify the whole soil water profile into three layers, i.e. strong depletion layer (2.0~4.4m), less strong layer (4.4~5.0m) and weak depletion layer (5.0~7.0m) based on the combined analysis of soil water content and the root dry weight density. This stratification result could well reflect soil water characteristic absorbed by root. In the strong depletion layer, fine root dry weight density decreased fluctuantly with the increase of soil depth, but soil water content was low and stable. While in the less strong layer, fine root dry weight density decreased continuously, soil water content presented increasing tendency. The maximum fine root depth was 5m, there was no fine root in the weak depletion layer and soil water content increased obviously. This study determine the soil layer range of deep soil water depletion degree in densely jujube plantation. It shows that root distribution characteristic has an important effect on the deep soil water variation.
DOIURL [本文引用: 1]
In order to investigate the relationship between deep soil water (below 2m) vertical variation and root distribution in the nine-year-old dense jujube plantation in Loess hilly region. The soil core method and oven drying method were used to determine fine root dry weight density and soil water content. Sequential cluster analysis method was adopted to stratify the whole soil water profile into three layers, i.e. strong depletion layer (2.0~4.4m), less strong layer (4.4~5.0m) and weak depletion layer (5.0~7.0m) based on the combined analysis of soil water content and the root dry weight density. This stratification result could well reflect soil water characteristic absorbed by root. In the strong depletion layer, fine root dry weight density decreased fluctuantly with the increase of soil depth, but soil water content was low and stable. While in the less strong layer, fine root dry weight density decreased continuously, soil water content presented increasing tendency. The maximum fine root depth was 5m, there was no fine root in the weak depletion layer and soil water content increased obviously. This study determine the soil layer range of deep soil water depletion degree in densely jujube plantation. It shows that root distribution characteristic has an important effect on the deep soil water variation.
DOIURL [本文引用: 4]
Afforestation as an effective measure to control soil erosion has achieved remarkable effects in northern China. However, large scale of artificial afforestation can increase soil water consumption and induce soil desiccation in arid and semi-arid areas. This study analyzed the variations of soil water storage following the conversion of croplands into forests with different species and stand ages on the Loess Plateau. Three most common artificial forests dominated by Salix matsudana , Populus cathayana , and Sophora japonica with stand ages of 5, 10, and 1562years were investigated to determine the variations in soil water storage. The results showed that soil water storage decreased with increasing afforestation ages and soil depth. Salix matsudana mainly consumed shallow soil water (0–10062cm), P. cathayana mainly consumed deep soil water (100–15062cm), while S. japonica had relatively lower water consumption than the other two species. Converting cropland into forest resulted in a significant water deficit. Soil water deficit in the 0–10062cm soil profiles was significantly higher under S. matsudana than under the other two artificial forests. Severe soil water depletion and obvious soil desiccation occurred after 1262years of afforestation. Therefore, artificial forests with less water consumption, e.g. S. japonica , should be given priority in future afforestation practice. To maintain the sustainability of vegetation, changes in land-use patterns should be considered after 1262years of afforestation.
[本文引用: 1]
DOIURL [本文引用: 1]
水分是植物存活、生长和分布过程中的必需资源,阐明植物对干旱的应对和调节机制,是植物生理生态学和全球变化生态学的重要研究命题。植物对不同气候与土壤水分条件的长期适应会形成由一整套相关联的性状组成的水分调节策略,其中等水和非等水调节行为是两种典型的水分调节对策。区分并阐明植物的水分调节对策及其机制,不但在干旱地区植物育种、植被修复等实践中有广泛的应用前景,而且可为构建更精确的植被动态模型和预测气候变化情景下植被分布提供科学基础。该文首先阐述了等水和非等水调节行为的定义及3种定量分类方法:(1)基于气孔导度与叶水势的关系;(2)基于气孔导度与水汽压亏缺的关系;(3)基于黎明前叶水势与中午叶水势的关系。之后,从水力和碳经济性状两个方面比较分析了两种水分调节对策植物的种间差异。综合分析植物水分调节机制发现,水力信号与化学信号的相互作用是植物水分调节行为的主控因素。最后提出3个亟待开展研究的问题:(1)针对不同地区开展植物水分关系相关性状的测定,寻求可靠且普适的植物水分调节对策分类方法。(2)探索植物水分调节对策与水力、形态、结构、功能等性状之间的关联性,为改进植被动态模型提供可靠的参数。(3)加深理解不同时空尺度上植物水分调节过程,揭示植物对环境胁迫(尤其是干旱)的响应和适应机制。
DOIURL [本文引用: 1]
水分是植物存活、生长和分布过程中的必需资源,阐明植物对干旱的应对和调节机制,是植物生理生态学和全球变化生态学的重要研究命题。植物对不同气候与土壤水分条件的长期适应会形成由一整套相关联的性状组成的水分调节策略,其中等水和非等水调节行为是两种典型的水分调节对策。区分并阐明植物的水分调节对策及其机制,不但在干旱地区植物育种、植被修复等实践中有广泛的应用前景,而且可为构建更精确的植被动态模型和预测气候变化情景下植被分布提供科学基础。该文首先阐述了等水和非等水调节行为的定义及3种定量分类方法:(1)基于气孔导度与叶水势的关系;(2)基于气孔导度与水汽压亏缺的关系;(3)基于黎明前叶水势与中午叶水势的关系。之后,从水力和碳经济性状两个方面比较分析了两种水分调节对策植物的种间差异。综合分析植物水分调节机制发现,水力信号与化学信号的相互作用是植物水分调节行为的主控因素。最后提出3个亟待开展研究的问题:(1)针对不同地区开展植物水分关系相关性状的测定,寻求可靠且普适的植物水分调节对策分类方法。(2)探索植物水分调节对策与水力、形态、结构、功能等性状之间的关联性,为改进植被动态模型提供可靠的参数。(3)加深理解不同时空尺度上植物水分调节过程,揭示植物对环境胁迫(尤其是干旱)的响应和适应机制。
DOIURL [本文引用: 1]
Aimed to understand how soil water was depleted by deep roots, the effects of drip irrigation and stand age on the deep root distribution, rooting depth, and soil water profile dynamics were investigated in a jujube (Ziziphus jujube Mill. CV. Lizao) plantation.A soil coring method with a LuoYang shovel was used for sampling until no more roots were found.It showed that the maximum fine rooting depth (We found that the application of 33.3 mm of irrigation water (equivalent to 7 % of the local annual precipitation) could halve the maximum rooting depth, thereby reducing deep soil water depletion. Our results showed that a low-volume water supply reduced the maximum rooting depth in jujube and prevented the depletion of the deep soil water. Appropriate drip irrigation is an effective water management strategy for sustainable artificial forest development in semiarid regions.
DOIURLPMID [本文引用: 4]
Abstract Educational attainment for women of reproductive age is linked to reduced child and maternal mortality, lower fertility and improved reproductive health. Comparable analyses of attainment exist only at the national level, potentially obscuring patterns in subnational inequality. Evidence suggests that wide disparities between urban and rural populations exist, raising questions about where the majority of progress towards the education targets of the Sustainable Development Goals is occurring in African countries. Here we explore within-country inequalities by predicting years of schooling across five by five kilometre grids, generating estimates of average educational attainment by age and sex at subnational levels. Despite marked progress in attainment from 2000 to 2015 across Africa, substantial differences persist between locations and sexes. These differences have widened in many countries, particularly across the Sahel. These high-resolution, comparable estimates improve the ability of decision-makers to plan the precisely targeted interventions that will be necessary to deliver progress during the era of the Sustainable Development Goals.
[本文引用: 1]
DOIURLPMID [本文引用: 4]
The drivers underlying the development of deep root systems, whether genetic or environmental, are poorly understood but evidence has accumulated that deep rooting could be a more widespread and important trait among plants than commonly anticipated from their share of root biomass. Even though a distinct classification of "deep roots" is missing to date, deep roots provide important functions for individual plants such as nutrient and water uptake but can also shape plant communities by hydraulic lift (HL). Subterranean fauna and microbial communities are highly influenced by resources provided in the deep rhizosphere and deep roots can influence soil pedogenesis and carbon storage.Despite recent technological advances, the study of deep roots and their rhizosphere remains inherently time-consuming, technically demanding and costly, which explains why deep roots have yet to be given the attention they deserve. While state-of-the-art technologies are promising for laboratory studies involving relatively small soil volumes, they remain of limited use for the in situ observation of deep roots. Thus, basic techniques such as destructive sampling or observations at transparent interfaces with the soil (e.g., root windows) which have been known and used for decades to observe roots near the soil surface, must be adapted to the specific requirements of deep root observation. In this review, we successively address major physical, biogeochemical and ecological functions of deep roots to emphasize the significance of deep roots and to illustrate the yet limited knowledge. In the second part we describe the main methodological options to observe and measure deep roots, providing researchers interested in the field of deep root/rhizosphere studies with a comprehensive overview. Addressed methodologies are: excavations, trenches and soil coring approaches, minirhizotrons (MR), access shafts, caves and mines, and indirect approaches such as tracer-based techniques
DOIURLPMID [本文引用: 2]
Abstract Fine roots acquire essential soil resources and mediate biogeochemical cycling in terrestrial ecosystems. Estimates of carbon and nutrient allocation to build and maintain these structures remain uncertain because of the challenges of consistently measuring and interpreting fine-root systems. Traditionally, fine roots have been defined as all roots 0909¤ 2 mm in diameter, yet it is now recognized that this approach fails to capture the diversity of form and function observed among fine-root orders. Here, we demonstrate how order-based and functional classification frameworks improve our understanding of dynamic root processes in ecosystems dominated by perennial plants. In these frameworks, fine roots are either separated into individual root orders or functionally defined into a shorter-lived absorptive pool and a longer-lived transport fine-root pool. Using these frameworks, we estimate that fine-root production and turnover represent 22% of terrestrial net primary production globally - a c. 30% reduction from previous estimates assuming a single fine-root pool. Future work developing tools to rapidly differentiate functional fine-root classes, explicit incorporation of mycorrhizal fungi into fine-root studies, and wider adoption of a two-pool approach to model fine roots provide opportunities to better understand below-ground processes in the terrestrial biosphere. 0008 2015 The Authors. New Phytologist 0008 2015 New Phytologist Trust.
DOIURLPMID [本文引用: 4]
Explanations for the occurrence of deep-rooted plants in arid and semi-arid ecosystems have traditionally emphasized the uptake of relatively deep soil water. However, recent hydrologic data from arid systems show that soil water potentials at depth fluctuate little over long time periods, suggesting this water may be rarely utilized or replenished. In this study, we examine the distributions of root biomass, soil moisture and nutrient contents to 10-m depths at five semi-arid and arid sites across south-western USA. We couple these depth distributions with strontium (Sr) isotope data that show deep (>1 m) nutrient uptake is prevalent at four of the five sites. At all of the sites, the highest abundance of one or more of the measured nutrients occurred deep within the soil profile, particularly for P, Ca and Mg . Phosphate contents were greater at depth than in the top meter of soil at three of five sites. At Jornada, for example, the 2-3 m depth increment had twice the extractable P as the top meter of soil, despite the highest concentrations of P occurring at the surface. The prevalence of such deep resource pools, and our evidence for cation uptake from them, suggest nutrient uptake as a complementary explanation for the occurrence of deep-rooted plants in arid and semi-arid systems. We propose that hydraulic redistribution of shallow surface water to deep soil layers by roots may be the mechanism through which deep soil nutrients are mobilized and taken up by plants.
DOIURLPMID [本文引用: 4]
Although deep roots can contribute substantially to whole-tree water use, little is known about deep root functioning because of limited access for in situ measurements. We used a cave system on the Edwards Plateau of central Texas to investigate the physiology of water transport in roots at 18 20 m depth for two common tree species, Quercus fusiformis and Bumelia lanuginosa . Using sap flow and water potential measurements on deep roots, we found that calculated root hydraulic conductivity (RHC) fluctuated diurnally for both species and decreased under shading for B. lanuginosa . To assess whether these dynamic changes in RHC were regulated during initial water absorption by fine roots, we used an ultra-low flowmeter and hydroxyl radical inhibition to measure in situ fine root hydraulic conductivity (FRHC) and aquaporin contribution to FRHC (AQPC), respectively. During the summer, FRHC and AQPC were found to cycle diurnally in both species, with peaks corresponding to the period of highest transpirational demand at midday. During whole-tree shade treatments, B. lanuginosa FRHC ceased diurnal cycling and decreased by 75 and 35% at midday and midnight, respectively, while AQPC decreased by 41 and 30% during both time periods. A controlled growth-chamber study using hydroponically grown saplings confirmed daily cycling and shade-induced reductions in FRHC and AQPC. Winter measurements showed that the evergreen Q. fusiformis maintained high FRHC and AQPC throughout the year, while the deciduous B. lanuginosa ceased diurnal cycling and exhibited its lowest annual values for both parameters in winter. Adjustments in FRHC and AQPC to changing canopy water demands may help the trees maintain the use of reliable water resources from depth and contribute to the success of these species in this semi-arid environment.
DOIURL [本文引用: 3]
Summary 6168 To assess hydraulic architecture and limitations to water transport across whole trees, we compared xylem anatomy, vulnerability to cavitation (Ψ 50 ) and specific hydraulic conductivity ( K s ) of stems, shallow roots and deep roots (from caves to 20m depth) for four species: Juniperus ashei , Bumelia lanuginosa , Quercus fusiformis and Quercus sinuata . 6168 Mean, maximum and hydraulically weighted ( D h ) conduit diameters and K s were largest in deep roots, intermediate in shallow roots, and smallest in stems ( P< 0.05 for each). Mean vessel diameters of deep roots were 2.1–4.2-fold greater than in stems, and K s was seven to 38 times larger in the deep roots. 6168 Ψ 50 also increased from stems to roots with depth, as much as 24-fold from stems to deep roots in B.lanuginosa . For all species together, Ψ 50 was positively correlated with both D h and K s , suggesting a potential trade-off exists between conducting efficiency and safety. 6168 The anatomical and hydraulic differences documented here suggest that the structure of deep roots minimizes flow resistance and maximizes deep water uptake.
DOIURLPMID [本文引用: 1]
We used concurrent measurements of soil content and soil potential ((soil)) to assess the effects of (soil) on and hydraulic redistribution (HR) of soil by roots during seasonal drought cycles at six sites characterized by differences in the types and amounts of woody vegetation and in climate. The six sites included a semi-arid old-ponderosa pine (Dougl. ex P. Laws & C. Laws) forest, a moist old-((Mirb.) Franco) forest, a 24-year-old forest and three Brazilian savanna sites differing in tree density. At all of the sites, HR was confined largely to the upper 60 cm of soil. There was a common threshold relationship between the relative magnitude of HR and (soil) among the six study sites. Below a threshold (soil) of about -0.4 MPa, overnight recharge of soil storage increased sharply, and reached a maximum value of 80-90% over a range of (soil) from ~ -1.2 to -1.5 MPa. Although amounts of hydraulically redistributed to the upper 60 cm of soil were relatively small (0 to 0.4 mm day(-1)), they greatly reduced the rates of seasonal decline in (soil). The effectiveness of HR in delaying soil drying diminished with increasing sapwood area per ground area. The relationship between soil utilization and (soil) in the 20-60-cm layer was nearly identical for all six sites. Soil utilization varied with a surrogate measure of rhizosphere conductance in a similar manner at all six sites. The similarities in relationships between (soil) and HR, soil utilization and relative rhizosphere conductance among the six sites, suggests that, despite probable differences in maximum rooting depth and density, there was a convergence in biophysical controls on soil utilization and redistribution in the upper soil layers where the density of finer roots is greatest.
DOIURL [本文引用: 1]
利用稳定氢同位素和热扩散技术研究张北防护林杨树的水分来源和蒸腾耗水,分析确定未退化与退化杨树的水分关系差异.结果表明:在生长季节中退化杨树主要利用0~30 cm土壤水分,未退化杨树主要利用30~80 cm土壤水分,两者的水分来源不同.旱季时,未退化杨树利用深层土壤水分和地下水的比例明显高于退化杨树.雨季中,杨树对0~30 cm土壤水分的利用比例增加,退化杨树增加幅度明显高于未退化杨树,对30~180 cm土壤水分的利用比例均减少.未退化杨树的液流速率大于退化杨树,不同天气中液流速率表现出相似的变化趋势,但未退化杨树液流的启动时间比退化杨树早.相关分析表明,未退化和退化杨树液流速率与土壤温度、风速、太阳辐射、相对湿度、空气温度均呈极显著的相关关系.退化杨树液流速率与土壤温度和空气相对湿度呈极显著负相关,与其他因素呈显著正相关,而未退化杨树仅与空气相对湿度呈极显著负相关,与其他因素均呈显著正相关关系,表明退化和未退化杨树蒸腾耗水易受环境条件的影响.退化杨树液流日累计量明显小于未退化杨树,表明其蒸腾耗水量较少;退化杨树水分来源浅,蒸腾耗水的减少并不能阻止林分退化.
DOIURL [本文引用: 1]
利用稳定氢同位素和热扩散技术研究张北防护林杨树的水分来源和蒸腾耗水,分析确定未退化与退化杨树的水分关系差异.结果表明:在生长季节中退化杨树主要利用0~30 cm土壤水分,未退化杨树主要利用30~80 cm土壤水分,两者的水分来源不同.旱季时,未退化杨树利用深层土壤水分和地下水的比例明显高于退化杨树.雨季中,杨树对0~30 cm土壤水分的利用比例增加,退化杨树增加幅度明显高于未退化杨树,对30~180 cm土壤水分的利用比例均减少.未退化杨树的液流速率大于退化杨树,不同天气中液流速率表现出相似的变化趋势,但未退化杨树液流的启动时间比退化杨树早.相关分析表明,未退化和退化杨树液流速率与土壤温度、风速、太阳辐射、相对湿度、空气温度均呈极显著的相关关系.退化杨树液流速率与土壤温度和空气相对湿度呈极显著负相关,与其他因素呈显著正相关,而未退化杨树仅与空气相对湿度呈极显著负相关,与其他因素均呈显著正相关关系,表明退化和未退化杨树蒸腾耗水易受环境条件的影响.退化杨树液流日累计量明显小于未退化杨树,表明其蒸腾耗水量较少;退化杨树水分来源浅,蒸腾耗水的减少并不能阻止林分退化.
DOIURL [本文引用: 3]
The spatial distribution of fine roots of two deciduous tree species was investigated in contrasting growing conditions in southern France. Hybrid walnut trees (Juglans regia脳nigra cv. NG23) and hybrid poplars (Populus euramericana cv. I214) were both cultivated with or without annual winter intercrops for 10 years on deep alluvial soils. Soil samples for measuring the fine root distribution of both trees and crops were obtained by soil coring down to 3-m depth at several distances and orientations from the tree trunk. The distribution of live fine roots from walnut and poplar trees was patchy and sometimes unexpected. In the tree-only stands, fine root profiles followed the expected pattern, as fine root density decreased with increasing depth and distance from the tree trunk. However, many fine root profiles under intercropped trees were uniform with depth, and some inverse profiles were observed. These distributions may result from a high degree of plasticity of tree root systems to sense and adapt to fluctuating and heterogeneous soil conditions. The distortion of the tree root system was more pronounced for the walnut trees that only partially explored the soil volume: in the tree-only stand, the walnut rooting pattern was very superficial, but in the intercropped stand walnut trees developed a deep and dense fine root network below the crop rooting zone. The larger poplars explored the whole available soil volume, but the intercrop significantly displaced the root density from the topsoil to layers below 1 m depth. Most tree root growth models assume a decreasing fine root density with depth and distance from the tree stem. These models would not predict correctly tree ree and tree nderstorey competition for water and nutrients in 3D heterogeneous soil conditions that prevail under low-density tree stands. To account for the integrated response of tree root systems to such transient gradients in soils, we need a dynamic model that would allow for both genotypic plasticity and transient environmental local soil conditions.
DOIURL [本文引用: 1]
Abstract Hydraulic redistribution plays a relevant role in the water relations of trees in climates with alternation between warm/dry and cold/wet periods. We aim to illustrate the ability of Mediterranean deep-rooted rainfed olive trees to maintain transpiration during the hot dry season and to redistribute soil water through roots, tending to temporarily homogenize soil moisture vertically. Sap flow was monitored by the heat field deformation method for 2·565years in the stem, lignotuber, medium and shallow roots of an olive tree tracing the long-term variations in the patterns of transpiration, water uptake and hydraulic redistribution. During the same period, soil water content and meteorological data were measured and related to sap flow. Results show that hydraulic redistribution within the rhizosphere buffers the seasonal and long-term water deficits. Under high evaporative demand during the dry summer, the deeper roots connected to stem through the lignotuber uptake water supporting transpiration needs and reducing the intensive drying of the upper soil layers. Copyright 08 2014 John Wiley & Sons, Ltd.
DOIURLPMID [本文引用: 3]
Abstract Although changes in depth to groundwater occur naturally, anthropogenic alterations may exacerbate these fluctuations and, thus, affect vegetation reliant on groundwater. These effects include changes in physiology, structure, and community dynamics, particularly in arid regions where groundwater can be an important water source for many plants. To properly manage ecosystems subject to changes in depth to groundwater, plant responses to both rising and falling groundwater tables must be understood. However, most research has focused exclusively on riparian ecosystems, ignoring regions where groundwater is available to a wider range of species. Here, we review responses of riparian and other species to changes in groundwater levels in arid environments. Although decreasing water tables often result in plant water stress and reduced live biomass, the converse is not necessarily true for rising water tables. Initially, rising water tables kill flooded roots because most species cannot tolerate the associated low oxygen levels. Thus, flooded plants can also experience water stress. Ultimately, individual species responses to either scenario depend on drought and flooding tolerance and the change in root system size and water uptake capacity. However, additional environmental and biological factors can play important roles in the severity of vegetation response to altered groundwater tables. Using the reviewed information, we created two conceptual models to highlight vegetation dynamics in areas with groundwater fluctuations. These models use flow charts to identify key vegetation and ecosystem properties and their responses to changes in groundwater tables to predict community responses. We then incorporated key concepts from these models into EDYS, a comprehensive ecosystem model, to highlight the potential complexity of predicting community change under different fluctuating groundwater scenarios. Such models provide a valuable tool for managing vegetation and groundwater use in areas where groundwater is important to both plants and humans, particularly in the context of climate change.
[本文引用: 4]
DOIURLPMID [本文引用: 2]
Summary337I.Introduction337II.Synthesis of the magnitudes of HR across ecosystems338III.Hydraulic redistribution models339IV.Methodological considerations affecting the magnitude of HR344V.Site characteristics affecting the magnitude of HR346VI.Plant characteristics affecting the magnitude of HR347VII.Conclusions349 Acknowledgements350 References350SummaryHydraulic redistribution (HR) 090009 the movement of water from moist to dry soil through plant roots 090009 occurs worldwide within a range of different ecosystems and plant species. The proposed ecological and hydrologic impacts of HR include increasing dry-season transpiration and photosynthetic rates, prolonging the life span of fine roots and maintaining root090009soil contact in dry soils, and moving rainwater down into deeper soil layers where it does not evaporate. In this review, we compile estimates of the magnitude of HR from ecosystems around the world, using representative empirical and modeling studies from which we could extract amounts of water redistributed by plant root systems. The reported average magnitude of HR varies by nearly two orders of magnitude across ecosystems, from 0.04 to 1.3 mm H2O d0908081 in the empirical literature, and from 0.1 to 3.23 mm H2O d0908081 in the modeling literature. Using these synthesized data, along with other published studies, we examine this variation in the magnitude of upward and downward HR, considering effects of plant, soil and ecosystem characteristics, as well as effects of methodological details (in both empirical and modeling studies) on estimates of HR. We take both ecological and hydrologic perspectives.
DOIURL [本文引用: 2]
1. Water is a key resource in tropical savannas. Changes in vegetation structure due to land-use change and increased fire frequency may affect the availability of water and the flux of water through these ecosystems. 2. We compared the seasonal soil moisture dynamics of two adjacent savanna ecosystems with contrasting tree densities in central Brazil. Our goal was to investigate the influence of tree density on deep water uptake, soil water dynamics and evapotranspiration. 3. Soil water was measured using a depth of 7·5 m beneath the tree-dominated cerrado denso ecosystem and to 4 m beneath the grass-dominated campo sujo ecosystem. Plant-available water (PAW) throughout the cerrado denso soil profile ranged from 293 mm at the end of the dry season to 689 mm during the wet season. In the grass-dominated site, PAW in the profile ranged from 155 to 362 mm. 4. During the dry season, ≈82% of the water used in cerrado denso and 67% in campo sujo was extracted from the profile below 1 m. The tree-dominated cerrado denso used 137 mm more water than the grass-dominated campo sujo. Significant deep soil water uptake was also observed during the wet season of 1998, when rainfall was below average. 5. Evapotranspiration (ET) rates (estimated as change in soil moisture over time) were higher in cerrado denso than in campo sujo during both seasons. Estimated ET ranged from 1·4 mm day-1during the dry season to 5·8 mm day-1for the wet season in cerrado denso, and from 0·9 mm day-1in the dry season to 4·5 mm day-1in early wet season in campo sujo. The differences in PAW and ET rates between the two ecosystems are associated not only with differences in root distribution, but also with differences in tree densities and the phenology of full-leaf canopies. 6. Our results suggest that deep-rooted plants may contribute significantly to the water balance of cerrado ecosystems, and that the hydrological cycle of this biome could change as woody vegetation is replaced by exotic grasses and agricultural crops.
URL [本文引用: 1]
叶片水力性状表征了叶片为适应外在环境而形成的水分传输方面的生存策略.叶片水力性状会限制整个植株的水 分传输,并影响植物的气体交换及其对干旱的响应,因此关于叶片水力性状的研究已成为植物水分关系领域的研究热点之一.本文概括了叶片水力性状的基本指标 (包括叶片整体水力导度(Kleaf)、叶片木质部水力导度(Kxylem)、叶片木质部外水力导度(Kout-xylem)等)和叶片水力导度的5种主 要测量方法;总结了叶脉网络结构和环境因素对叶片水力性状的影响、叶片水力性状与叶片功能指标(气孔导度、叶片水势、叶片最大光合速率)的匹配与权衡关 系,以及叶片水力性状与植物抗旱性关系的最新研究进展;对今后叶片水力性状的研究提出了两点建议:1)将叶片水力性状与气体交换和叶解剖结构等相结合,构 建叶片碳-水耦合模型,揭示叶片应对外界环境变化而采取的生态策略,以及植物的水-碳投资机理;2)开展植株各部分(根-茎-叶)间水分传输的交互作用研 究,筛选出水力系统高效安全的物种.
URL [本文引用: 1]
叶片水力性状表征了叶片为适应外在环境而形成的水分传输方面的生存策略.叶片水力性状会限制整个植株的水 分传输,并影响植物的气体交换及其对干旱的响应,因此关于叶片水力性状的研究已成为植物水分关系领域的研究热点之一.本文概括了叶片水力性状的基本指标 (包括叶片整体水力导度(Kleaf)、叶片木质部水力导度(Kxylem)、叶片木质部外水力导度(Kout-xylem)等)和叶片水力导度的5种主 要测量方法;总结了叶脉网络结构和环境因素对叶片水力性状的影响、叶片水力性状与叶片功能指标(气孔导度、叶片水势、叶片最大光合速率)的匹配与权衡关 系,以及叶片水力性状与植物抗旱性关系的最新研究进展;对今后叶片水力性状的研究提出了两点建议:1)将叶片水力性状与气体交换和叶解剖结构等相结合,构 建叶片碳-水耦合模型,揭示叶片应对外界环境变化而采取的生态策略,以及植物的水-碳投资机理;2)开展植株各部分(根-茎-叶)间水分传输的交互作用研 究,筛选出水力系统高效安全的物种.
DOIURL [本文引用: 4]
Specific hydraulic conductivities (Ks) of trunks (stem), superficial lateral roots, and deeply penetrating sinker (tap) roots were examined in variously-aged tree or shrub species of Proteaceae (Banksia prionotes, Banksia ilicifolia and Adenanthos cygnorum) in native habitat on deep sand overlying water tables located at 1.8-3.8 m depth. Ks values (on the basis of organ transectional area) for sinker roots (range from 30 to 780 10-3 m2 MPa-1 s-1) were consistently greater than in associated laterals (2 to 50 10-3 m2 MPa-1 s-1), and much greater again than in trunks (0.5 to 9 10-3 m2 MPa-1 s-1). Wood was essentially diffuseporous and mean lengths of vessels in the trunk, laterals and sinker roots of B. prionotes were 4 cm, approximately 75 cm and greater than 1.5 m, respectively. A 5-20-fold increase in Ks from top to base of a sinker was associated with progressive increases in mean radii of conducting elements, proportional areas of stele occupied by xylem conduits and percentage of organ fresh weight displaceable by mild vacuum extraction of water from vessel lumina. Resulting from these changes, conductance (Kh) of a sinker was relatively constant along its length, or even increased with depth, despite a 10-15-fold downward attenuation in transectional area over the 1.3-1.9 m length of root studied. Correlation plots of mean vessel radius and Ks (lumen area basis) grouped data for laterals separately from that of sinkers and showed interspecific differences in Ks values for similar range classes of lumen radius. Increases in conduit flow with increasing radius were less than predicted from the HagenPoiseuille equation.
[本文引用: 1]
DOIURLPMID [本文引用: 7]
Abstract BACKGROUND: Deep roots are a common trait among a wide range of plant species and biomes, and are pivotal to the very existence of ecosystem services such as pedogenesis, groundwater and streamflow regulation, soil carbon sequestration and moisture content in the lower troposphere. Notwithstanding the growing realization of the functional significance of deep roots across disciplines such as soil science, agronomy, hydrology, ecophysiology or climatology, research efforts allocated to the study of deep roots remain incommensurate with those devoted to shallow roots. This is due in part to the fact that, despite technological advances, observing and measuring deep roots remains challenging. SCOPE: Here, other reasons that explain why there are still so many fundamental unresolved questions related to deep roots are discussed. These include the fact that a number of hypotheses and models that are widely considered as verified and sufficiently robust are only partly supported by data. Evidence has accumulated that deep rooting could be a more widespread and important trait among plants than usually considered based on the share of biomass that it represents. Examples that indicate that plant roots have different structures and play different roles with respect to major biochemical cycles depending on their position within the soil profile are also examined and discussed. CONCLUSIONS: Current knowledge gaps are identified and new lines of research for improving our understanding of the processes that drive deep root growth and functioning are proposed. This ultimately leads to a reflection on an alternative paradigm that could be used in the future as a unifying framework to describe and analyse deep rooting. Despite the many hurdles that pave the way to a practical understanding of deep rooting functions, it is anticipated that, in the relatively near future, increased knowledge about the deep rooting traits of a variety of plants and crops will have direct and tangible influence on how we manage natural and cultivated ecosystems. The Author 2016. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
DOIURL [本文引用: 5]
Although pioneer studies showed several decades ago that deep rooting is common in tropical forests, direct measurements of fine root distributions over the entire soil profile explored by the roots are still scarce. Our study aimed to compare, 2years after planting, fine root traits ofEucalyptustrees planted from cuttings and from seedlings in order to assess whether the propagation mode has an influence on the capacity of the trees to explore very deep soils. Soils cores were sampled down to a depth of 13.5m at the peak of leaf area index (LAI), 2years after planting, under threeEucalyptusclones (belonging to speciesE. saligna,E. grandis E. urophylla,E. grandis E. camaldulensis) and underE. grandisseedlings in the same Ferralsol soil. LAI was estimated using allometric equations based on destructive sampling of eight trees per genotype. All the genotypes exhibited fine root densities roughly constant between the depths of 0.25 and 6.00m. Changes in fine root traits (diameter, specific root length and specific root area) were low between the topsoil and the root front. The ratios between mean tree height and root front depth ranged from 0.8 to 1.2 for the four genotypes. Although tree vertical extension was roughly symmetric above and belowground for all the genotypes, the depth of the root front ranged from 8.0m for the seedlings and theE. grandis E. urophyllaclone to 11.5m for theE. salignaclone. Soil water content profiles suggested that the four genotypes had the capacity to withdraw water down to a depth of 8 10m over the first 2years after planting. Total fine root length ranged from 3.3 to 6.0km per m2of soil depending on the genotype. The root area/leaf area ratio ranged from 1.3 to 3.2 and was negatively correlated with LAI across the four genotypes. This pattern suggests that the genotypes more conservative for water use (with a low LAI) invest more in fine root area relative to leaf area than genotypes adapted to wet regions (with a high LAI). The velocity of downward movement of the root front might be a relevant criterion in the last stage of the breeding programs to select clones with a fast exploration of deep soil layers in drought prone regions.
DOIURL [本文引用: 3]
Summary There is a fundamental trade-off between leaf traits associated with either resource acquisition or resource conservation. This gradient of trait variation, called the economics spectrum , also applies to fine roots, but whether it is consistent for coarse roots or at the plant community level remains untested. We measured a set of morphological and chemical root traits at a community level (functional parameters; FP) in 20 plant communities located along land-use intensity gradients and across three climatic zones (tropical, mediterranean and montane). We hypothesized (i) the existence of a root economics spectrum in plant communities consistent within root types (fine, < 202mm; coarse, 2–502mm), (ii) that variations in root FP occur with soil depths (top 2002cm of soil and 100–15002cm deep) and (iii) along land-use gradients. Root FP covaried, in line with the resource acquisition–conservation trade-off, from communities with root FP associated with resource acquisition (e.g. high specific root length, SRL; thin diameters and low root dry matter contents, RDMC) to root FP associated with resource conservation (e.g. low SRL, thick diameters and high RDMC). This pattern was consistent for both fine and coarse roots indicating a strong consistency of a trade-off between resource acquisition and conservation for plant roots. Roots had different suites of traits at different depths, suggesting a disparity in root function and exploitation capacities. Shallow, fine roots were thinner, richer in nitrogen and with lower lignin concentrations associated with greater exploitation capacities compared to deep, fine roots. Shallow, coarse roots were richer in nitrogen, carbon and soluble concentrations than deep, coarse roots. Fine root parameters of highly disturbed, herbaceous-dominated plant communities in poorer soils were associated with foraging strategies, that is greater SRL and lower RDMC and lignin concentration than those from less disturbed communities. Coarse roots, however, were less sensitive to the land-use gradient. Synthesis . This study demonstrates the existence of a general trade-off in root construction at a community level, which operates within all root types, suggesting that all plant tissues are controlled by the trade-off between resource acquisition and conservation.
DOIURL [本文引用: 4]
Abstract Forests in the Southeast U.S. are predicted to experience future changes in seasonal patterns of precipitation inputs as well as more variable precipitation events. These climate change induced alterations could increase drought and lower soil water availability. Drought could alter rooting patterns and increase the importance of deep roots that access subsurface water resources. To address plant response to drought in both deep rooting and soil water utilization as well as soil drainage, we utilize a throughfall reduction experiment in a loblolly pine plantation of the Southeast U.S. to calibrate and validate a hydrological model. The model was accurately calibrated against field measured soil moisture data under ambient rainfall and validated using 30% throughfall reduction data. Using this model we then tested these scenarios: (1) evenly reduced precipitation; (2) less precipitation in summer, more in winter; (3) same total amount of precipitation with less frequent but heavier storms; and (4) shallower rooting depth under the above three scenarios. When less precipitation was received, drainage decreased proportionally much faster than evapotranspiration implying plants will acquire water first to the detriment of drainage. When precipitation was reduced by more than 30%, plants relied on stored soil water to satisfy evapotranspiration suggesting 30% may be a threshold that if sustained over the long-term would deplete plant available soil water. Under the third scenario, evapotranspiration and drainage decreased, while surface runoff increased. Changes in root biomass measured before and four years after the throughfall reduction experiment were not detected among treatments. Model simulations, however, indicated gains in evapotranspiration with deeper roots under evenly reduced precipitation and seasonal precipitation redistribution scenarios, but not when precipitation frequency was adjusted. Deep soil and deep rooting can provide an important buffer capacity when precipitation alone cannot satisfy the evapotranspirational demand of forests. How this buffering capacity will persist in the face of changing precipitation inputs, however, will depend less on seasonal redistribution than on the magnitude of reductions and changes in rainfall frequency.
DOIURLPMID [本文引用: 3]
Recent theory and field observations suggest that a systematically varying weathering zone, that can be tens of meters thick, commonly develops in the bedrock underlying hillslopes. Weathering turns otherwise poorly conductive bedrock into a dynamic water storage reservoir. Infiltrating precipitation typically will pass through unsaturated weathered bedrock before reaching groundwater and running off to streams. This invisible and difficult to access unsaturated zone is virtually unexplored compared with the surface soil mantle. We have proposed the term ock moisture to describe the exchangeable water stored in the unsaturated zone in weathered bedrock, purposely choosing a term parallel to, but distinct from, soil moisture, because weathered bedrock is a distinctly different material that is distributed across landscapes independently of soil thickness. Here, we report a multiyear intensive campaign of quantifying rock moisture across a hillslope underlain by a thick weathered bedrock zone using repeat neutron probe measurements in a suite of boreholes. Rock moisture storage accumulates in the wet season, reaches a characteristic upper value, and rapidly passes any additional rainfall downward to groundwater. Hence, rock moisture storage mediates the initiation and magnitude of recharge and runoff. In the dry season, rock moisture storage is gradually depleted by trees for transpiration, leading to a common lower value at the end of the dry season. Up to 27% of the annual rainfall is seasonally stored as rock moisture. Significant rock moisture storage is likely common, and yet it is missing from hydrologic and land-surface models used to predict regional and global climate.
DOIURLPMID [本文引用: 1]
Diel soil water potential fluctuations reflected daytime depletion and nocturnal resupply of water in upper soil layers. Transpiration suppression experiments demonstrated that water absorption by roots caused the daytime depletion. The soil water potential data and experimental results suggest that at night water absorbed from moist soil by deeper roots is transported to and lost from roots into drier upper soil layers. The deeper roots appear to absorb and transport water both day and night. Implications for the efficiency of deep roots and water storage, nutrient uptake and water parasitism in upper soil layers are discussed.
DOIURLPMID [本文引用: 2]
The significance of soil water redistribution facilitated by roots (an extension of "hydraulic lift", here termed hydraulic redistribution) was assessed for a stand of Artemisia tridentata using measurements and a simulation model. The model incorporated water movement within the soil via unsaturated flow and hydraulic redistribution and soil water loss from transpiration. The model used Buckingham-Darcy's law for unsaturated flow while hydraulic redistribution was developed as a function of the distribution of active roots, root conductance for water, and relative soil-root (rhizosphere) conductance for water. Simulations were conducted to compare model predictions with time courses of soil water potential at several depths, and to evaluate the importance of root distribution, soil hydraulic conductance and root xylem conductance on transpiration rates and the dynamics of soil water. The model was able to effectively predict soil water potential during a summer drying cycle, and the rapid redistribution of water down to 1.5 m into the soil column after rainfall events. Results of simulations indicated that hydraulic redistribution could increase whole canopy transpiration over a 100-day drying cycle. While the increase was only 3.5% over the entire 100-day period, hydraulic redistribution increased transpiration up to 20.5% for some days. The presence of high soil water content within the lower rooting zone appears to be necessary for sizeable increases in transpiration due to hydraulic redistribution. Simulation results also indicated that root distributions with roots concentrated in shallow soil layers experienced the greatest increase in transpiration due to hydraulic redistribution. This redistribution had much less effect on transpiration with more uniform root distributions, higher soil hydraulic conductivity and lower root conductivity. Simulation results indicated that redistribution of water by roots can be an important component in soil water dynamics, and the model presented here provides a useful approach to incorporating hydraulic redistribution into larger models of soil processes.
DOIURL [本文引用: 6]
The factors that shape vertical root distributions in different soils and under different climates and vegetation are poorly understood. This makes it difficult to parameterize root profiles in vegetation-, hydrology, biogeochemistry-, or global circulation models. Recently, it has been proposed that vertical root distributions in the vadose zone could be predicted from soil water infiltration and extraction patterns as a function of climatic variability, soil, and vegetation characteristics. A number of ecological factors favor shallow over deep roots, suggesting that root profiles of plant communities may tend to be as shallow as possible and as deep as needed to fulfill evapotranspirational demands. To test this hypothesis, a stochastic, one-dimensional soil water infiltration and extraction model (SWIEM) was developed that simulates soil water infiltration through 600 discrete soil layers to a depth of 6 m. Water input is simulated in Monte Carlo fashion based on site-specific long-term precipitation data. Water extraction proceeds from the top down, with extraction depths determined by potential evapotranspiration ( PET) and the vertical distribution of soil water. The resulting shallowest possible water extraction profile was tested against nine measured root profiles from long-term ecological research sites in different biomes. Two other approaches, based on mean root distributions for biomes and an empirical regression model, were also compared to the observed root distributions. Soil water extraction patterns predicted by the SWIEM model matched observed vertical root distributions better than the other two approaches. These findings show that vertical root distributions in different biomes tend to approach the shallowest possible shape, thereby creating a useful null model for future research on root distributions and a promising tool for parameterization of global models.
DOIURL [本文引用: 10]
Studies in global plant biogeography have almost exclusively analyzed relationships of abiotic and biotic factors with the distribution and structure of vegetation aboveground. The goal of this study was to extend such analyses to the belowground structure of vegetation by determining the biotic and abiotic factors that influence vertical root distributions in the soil, including soil, climate, and plant properties. The analysis used a database of vertical root profiles from the literature with 475 profiles from 209 geographic locations. Since most profiles were not sampled to the maximum rooting depth, several techniques were used to estimate the amount of roots at greater depths, to a maximum of 3 m in some systems. The accuracy of extrapolations was tested using a subset of deeply (>2 m) sampled or completely sampled profiles. Vertical root distributions for each profile were characterized by the interpolated 50% and 95% rooting depths (the depths above which 50% or 95% of all roots were located). General linear models incorporating plant life-form dominance, climate, and soil variables explained as much as 50% of the variance in rooting depths for various biomes and life-forms. Annual potential evapotranspiration (PET) and precipitation together accounted for the largest proportion of the variance (12-16% globally and 38% in some systems). Mean 95% rooting depths increased with decreasing latitude from 80 to 30 but showed no clear trend in the tropics. Annual PET, annual precipitation, and length of the warm season were all positively correlated with rooting depths. Rooting depths in tropical vegetation were only weakly correlated with climatic variables but were strongly correlated with sampling depths, suggesting that even after extrapolation, sampling depths there were often insufficient to characterize root profiles. Globally, >90% of all profiles had at least 50% of all roots in the upper 0.3 m of the soil profile (including organic horizons) and 95% of all roots in the upper 2 m. Deeper rooting depths were mainly found in water-limited ecosystems. Deeper 95% rooting depths were also found for shrublands compared to grasslands, in sandy soils vs. clay or loam soils, and in systems with relatively shallow organic horizons compared with deeper organic horizons.
DOIURL [本文引用: 3]
1 In water-limited environments, the availability of water and nutrients to plants depends on environmental conditions, sizes and shapes of their root systems, and root competition. The goal of this study was to predict root system sizes and shapes for different plant growth forms using data on above-ground plant sizes, climate and soil texture. 2 A new data set of > 1300 records of root system sizes for individual plants was collected from the literature for deserts, scrublands, grasslands and savannas with 1000 mm mean annual precipitation (MAP). Maximum rooting depths, maximum lateral root spreads and their ratios were measured. 3 Root system sizes differed among growth forms and increased with above-ground size: annuals < perennial forbs = grasses < semi-shrubs < shrubs < trees. Stem succulents were as shallowly rooted as annuals but had lateral root spreads similar to shrubs. 4 Absolute rooting depths increased with MAP in all growth forms except shrubs and trees, but were not strongly related to potential evapotranspiration (PET). Except in trees, root systems tended to be shallower and wider in dry and hot climates and deeper and narrower in cold and wet climates. Shrubs were more shallowly rooted under climates with summer than winter precipitation regimes. 5 Relative to above-ground plant sizes, root system sizes decreased with increasing PET for all growth forms, but decreased with increasing MAP only for herbaceous plants. Thus relative rooting depths tended to increase with aridity, although absolute rooting depths decreased with aridity. 6 Using an independent data set of 20 test locations, rooting depths were predicted from MAP using regression models for three broad growth forms. The models succeeded in explaining 62% of the observed variance in median rooting depths. 7 Based on the data analysed here, Walter's two-layer model of soil depth partitioning between woody and herbaceous plants appears to be most appropriate in drier regimes (< 500 mm MAP) and in systems with substantial winter precipitation.
DOIURL [本文引用: 9]
Global data sets of rooting depths, long-term means of monthly precipitation and potential evapotranspiration, and soil texture were used to predict the probability of deep rooting around the globe. Rooting depths were considered to be deep if 5% or more of all roots in a profile were located below 2 m depth. Spatially explicit global predictions were based on empirical models relating the probability of deep rooting with climate and soil texture. Based on these predictions, deep roots are most likely to occur in seasonally dry, semiarid to humid tropical regions under savanna or thorn-scrub vegetation or under seasonally dry semideciduous to evergreen forests. Deep roots are least likely to occur in arctic, boreal or cool-temperate regions and in per-humid climates such as equatorial rain forests. Under warm-temperate to tropical climates, rooting depths are more likely to be deep in coarse-textured and fine-textured soil than in soils of medium texture. These observations agree well with predictions based on a previously published conceptual model, according to which water infiltration depths and evaporative demand are the main climatic factors to determine vertical root distributions on a global scale.
DOIURL [本文引用: 1]
A device was designed which used gypsum and electrical conductivity as a means of detecting and quantitizing the amount of water emanating from underground plant parts under natural conditions. Use of this device with the Common French Bean demonstrated that considerable amounts of moisture emanated from the underground part of the stem. The amount of water emanated was found to be influenced by environmental conditions, differences in soil type and degree of soil compaction. When plants were grown in an environment of approximately 100 per cent relative air humidity and at a temperature of 28 to 30°C, 115 to 125 08l of water emanated from an 11 mm length of stem in a 24-hour period, in contrast to 70 to 80 08l of water emanating from plants grown at 50 to 60 per cent relative air humidity and at a temperature of 18 to 20°C. Water emanation was more rapid in a compacted soil than loose soil and also appeared to be dependent on the amount of sand in a soil. Whereas 115 to 125 08l of water emanated in a silt loam soil, only 35 to 45 08l emanated in a loamy sand soil and less than 10 08l in washed sea sand during a 24-hour period. The sensitivity of the conductivity unit to changes in moisture and the utility of its design enables its employment under varied environmental conditions and offers a technique for plant pathologists to study the effect of both the below-and aboveground environments on moisture emanation from underground plant parts and the resultant effect on the associated microflora.
DOIURL [本文引用: 1]
When a plant encounters spatially heterogeneous soil moisture within its root system, usually drier surface and moister subsurface soils, water can move between these layers through the root system, a plant process known as hydraulic lift or redistribution. The water thus transferred is available not only for the plant itself but also for its neighbors. We examined application of this process as a possible biological irrigation tool. As 'donors', we used perennial forage plants with their shoots removed to minimize the effect of lightinterception by them on the 'receiver' plants growing alongside them. In a horizontally split-root experiment, where an upper container was filled with sand and a lower one with water, superior donor species could maintain the upper sand in a fully hydrated condition for several weeks, increasing stomatal conductance in the receivers. The effects were also confirmed in a waterlimited agricultural field, as significant differences were found in canopy temperature and yield in neighboring crop plants in the presence or absence of donor root systems. These results suggest that deep-rooting associate plants with their shoots removed function as an irrigation tool and improve crop production in water-scarce environments.
DOIURL [本文引用: 1]
Abstract Depth to bedrock serves as the lower boundary of land surface models, which controls hydrologic and biogeochemical processes. This paper presents a framework for global estimation of depth to bedrock (DTB). Observations were extracted from a global compilation of soil profile data (ca. 1,30,000 locations) and borehole data (ca. 1.6 million locations). Additional pseudo???observations generated by expert knowledge were added to fill in large sampling gaps. The model training points were then overlaid on a stack of 155 covariates including DEM???based hydrological and morphological derivatives, lithologic units, MODIS surface reflectance bands and vegetation indices derived from the MODIS land products. Global spatial prediction models were developed using random forest and Gradient Boosting Tree algorithms. The final predictions were generated at the spatial resolution of 250 m as an ensemble prediction of the two independently fitted models. The 10???fold cross???validation shows that the models explain 59% for absolute DTB and 34% for censored DTB (depths deep than 200 cm are predicted as 200 cm). The model for occurrence of R horizon (bedrock) within 200 cm does a good job. Visual comparisons of predictions in the study areas where more detailed maps of depth to bedrock exist show that there is a general match with spatial patterns from similar local studies. Limitation of the data set and extrapolation in data spare areas should not be ignored in applications. To improve accuracy of spatial prediction, more borehole drilling logs will need to be added to supplement the existing training points in under???represented areas.
[本文引用: 2]
DOIURL [本文引用: 1]
Estimation of water uptake by plants and subsequent water stress are complicated by the need to resolve the soil-plant hydrodynamics at scales ranging from millimeters to meters. Using a simplified homogenization technique, the three-dimensional (3-D) soil water movement dynamics can be reduced to solving two 1-D coupled Richards' equations, one for the radial water movement toward rootlets (mesoscale, important for diurnal cycle) and a second for vertical water motion (macroscale, relevant to interstorm timescales). This approach allows explicit simulation of known features of root uptake such as diurnal hysteresis in canopy conductance, hydraulic lift, and compensatory root water uptake during extended drying cycles. A simple scaling analysis suggests that the effectiveness of the hydraulic lift is mainly controlled by the root vertical distribution, while the soil moisture levels at which hydraulic lift is most effective is dictated by soil hydraulic properties and surrogates for atmospheric water vapor demand.
[本文引用: 3]
[本文引用: 1]
DOIURLPMID [本文引用: 2]
Though the root biomass of tropical rainforest trees is concentrated in the upper soil layers, soil water uptake by deep roots has been shown to contribute to tree transpiration. A precise evaluation of the relationship between tree dimensions and depth of water uptake would be useful in tree-based modelling approaches designed to anticipate the response of tropical rainforest ecosystems to future changes in environmental conditions. We used an innovative dual-isotope labelling approach (deuterium in surface soil and oxygen at 120-cm depth) coupled with a modelling approach to investigate the role of tree dimensions in soil water uptake in a tropical rainforest exposed to seasonal drought. We studied 65 trees of varying diameter and height and with a wide range of predawn leaf water potential (I-pd) values. We confirmed that about half of the studied trees relied on soil water below 100-cm depth during dry periods. I-pd was negatively correlated with depth of water extraction and can be taken as a rough proxy of this depth. Some trees showed considerable plasticity in their depth of water uptake, exhibiting an efficient adaptive strategy for water and nutrient resource acquisition. We did not find a strong relationship between tree dimensions and depth of water uptake. While tall trees preferentially extract water from layers below 100-cm depth, shorter trees show broad variations in mean depth of water uptake. This precludes the use of tree dimensions to parameterize functional models.
DOIURL [本文引用: 1]
Knowledge regarding the use of groundwater by plants has implications for successful mine rehabilitation and revegetation programs in water-limited environments. In this study, we combined several approaches to investigate water sources used by Acacia papyrocarpa (Western myall) in the far west of South Australia, including stable isotopes, water potential, groundwater and soil chemistry, and root mapping techniques. Plant δ18O signatures and water potentials were compared against a range of possible sources: rainwater, surface soil water (≤102m depth), and deep groundwater (>2002m depth). Our aim was to determine whether groundwater contributed to the mix of waters used by A.02papyrocarpa.Overall, we found that trees did not source surface soil water (≤102m), and probably sourced deep soil water (i.e. >102m) rather than deep groundwater. Groundwater, however, could not be dismissed as a potential source, as root mapping showed tree roots were capable of reaching groundwater at depths >2002m, and isotope results indicated a potential contribution by groundwater to tree water use. However, low osmotic potentials and/or high acidity levels were shown to pose likely barriers to groundwater uptake, at least at the time of sampling. We conclude that because groundwater salinity and acidity are spatially variable in this region, plants with extensive root systems may be able to utilize zones of groundwater with lower salinity and pH levels. Overall, this study contributes to our limited understanding of groundwater use by trees occurring in water-limited environments where groundwater is extremely deep (>2002m depth).
DOIURL [本文引用: 3]
Abstract Based on literature reports, personal communications and our own observations, maximum vertical radial root extents were tabulated for various woody species, primarily forest trees and shrubs, and horticultural trees. Data were summarized for 49 families, 96 genera and 211 species, as well as for forest stands composed of mixtures of species. These data demonstrate the inherent capability of many species to develop deep or far-reaching roots in the absence of restrictive soil or substrate characteristics. These data also suggest that extensive roots may play a more important role in uptake of water and nutrients than indicated by their density alone, and that actual, rather than assumed, root extent must be evaluated on a site-specific basis to provide realistic estimates of ecosystem properties and processes.
DOIURLPMID [本文引用: 1]
Lowland dry forests are unique in Hawaii for their high diversity of tree species compared with wet forests. We characterized spatial and temporal partitioning of soil water resources among seven indigenous and one invasive dry forest species to determine whether the degree of partitioning was consistent with the relatively high species richness in these forests. Patterns of water utilization were inferred from stable hydrogen isotope ratios (δD) of soil and xylem water, zones of soil water depletion, plant water status, leaf phenology, and spatial patterns of species distribution. Soil water δD values ranged from -20‰ near the surface to -48‰ at 130 cm depth. Metrosideros polymorpha, an evergreen species, and Reynoldsia sandwicensis, a drought-deciduous species, had xylem sap δD values of about -52‰, and appeared to obtain their water largely from deeper soil layers. The remaining six species had xylem δD values ranging from -33 to -42‰, and apparently obtained water from shallower soil layers. Xylem water δD values were negatively correlated with minimum annual leaf water potential and positively correlated with leaf solute content, an integrated measure of leaf water deficit. Seasonal patterns of leaf production ranged from dry season deciduous at one extreme to evergreen with near constant leaf expansion rates at the other. Species tapping water more actively from deeper soil layers tended to exhibit larger seasonality of leaf production than species relying on shallower soil water sources. Individuals of Myoporum sandwicense were more spatially isolated than would be expected by chance. Even though this species apparently extracted water primarily from shallow soil layers, as indicated by its xylem δD values, its nearly constant growth rates across all seasons may have been the result of a larger volume of soil water available per individual. The two dominant species, Diospyros sandwicensis and Nestegis sandwicensis, exhibited low leaf water potentials during the dry season and apparently drew water mostly from the upper portion of the soil profile, which may have allowed them to exploit light precipitation events more effectively than the more deeply rooted species. Character displacement in spatial and temporal patterns of soil water uptake was consistent with the relatively high diversity of woody species in Hawaiian dry forests.
DOIURL [本文引用: 1]
Hydraulic redistribution (HR), one of the common bio-physical processes, plays key roles in mediating plant drought stress, regulating plant interspecific relations and community composition, as well as in influencing water and carbon balance of ecosystems. Great improvements have been achieved in HR research with fast development of isotopic labelling and tracing technologies. This paper summarizes the effects of HR on nutrient cycling of soil-plant system, based on studies over the past decade. HR increases soil water content in dry soil layers, thus helping to prevent embolism in roots, increasing the survival rate and the growth rate of fine roots and improving microbial activities. Such effects improve plant nutrient uptake, i.e., hydraulic lift promotes nitrogen uptake from upper soil layers and hydraulic descent promotes phosphorus uptake from deep soil layers. HR may facilitate nutrient exchange between upper and lower soil nutrient pools, improve nutrient flows and regulate the N:P ratio in both plants and soil. These effects may ultimately affect global ecosystems. Under the global change scenarios (e.g. nitrogen deposition), it is necessary to further explore the effects of HR on biogeochemical cycles. HR should be taken into account when using ecosystem models for future predictions.
DOIURL [本文引用: 1]
Hydraulic redistribution (HR), one of the common bio-physical processes, plays key roles in mediating plant drought stress, regulating plant interspecific relations and community composition, as well as in influencing water and carbon balance of ecosystems. Great improvements have been achieved in HR research with fast development of isotopic labelling and tracing technologies. This paper summarizes the effects of HR on nutrient cycling of soil-plant system, based on studies over the past decade. HR increases soil water content in dry soil layers, thus helping to prevent embolism in roots, increasing the survival rate and the growth rate of fine roots and improving microbial activities. Such effects improve plant nutrient uptake, i.e., hydraulic lift promotes nitrogen uptake from upper soil layers and hydraulic descent promotes phosphorus uptake from deep soil layers. HR may facilitate nutrient exchange between upper and lower soil nutrient pools, improve nutrient flows and regulate the N:P ratio in both plants and soil. These effects may ultimately affect global ecosystems. Under the global change scenarios (e.g. nitrogen deposition), it is necessary to further explore the effects of HR on biogeochemical cycles. HR should be taken into account when using ecosystem models for future predictions.
DOIURL [本文引用: 6]
[本文引用: 1]
DOIURL [本文引用: 1]
Hydraulic lift can be a desirable characteristic for cotton growing in areas that experience frequent drying of the topsoil. This study employed specialized soil columns to test if cotton can hydraulically lift water and whether this hydraulically lifted water can facilitate P acquisition from the dry topsoil of a clay and a sandy soil. The role of subsoil P placement on cotton growth was also tested. By using a Theta probe, hydraulic lift was detected in the clay soil for 7 days after water was withheld from the topsoil. However, in the sandy soil, a significant overnight increase in soil water was only detected for one day for the treatment with subsoil P application. Where P was applied to the topsoil, and water was withheld, cotton roots showed significant root P uptake from the sandy soil, but negligible P uptake from the clay soil. Irrespective of topsoil watering, the higher root length density in the subsoil of the clay soil accounted for the pronounced response to deep P placement. This study demonstrated that soil texture affects the amount and longevity of hydraulic lift from cotton roots. Hydraulic lift detected in the clay soil did not aid P acquisition from dry surface soil by cotton plants.
DOIURLPMID [本文引用: 3]
Abstract Root traits in morphology, chemistry and anatomy are important to root physiological functions, but the differences between shallow and deep roots have rarely been studied in woody plants. Here, we selected three temperate hardwood species, Juglans mandshurica Maxim., Fraxinus mandschurica Rupr. and Phellodendron amurense Rupr., in plantations in northeastern China and measured morphological, anatomical and chemical traits of root tips (i.e., the first-order roots) at surface (0-10 m) and subsurface (20-30 m) soil layers. The objectives of this study were to identify how those traits changed with soil depth and to reveal potential functional differences. The results showed that root diameters in deep root tips were greater in J. mandshurica and F. mandschurica, but smaller in P. amurense. However, root stele diameter and the ratio of stele to root diameter in the subsurface layer were consistently greater in all three species, which may enhance their abilities to penetrate into soil. All deep roots exhibited lower tissue nitrogen concentration and respiration rate, which were possibly caused by lower nutrient availability in the subsurface soil layer. Significant differences between shallow and deep roots were observed in xylem structure, with deep roots having thicker stele, wider maximum conduit and greater number of conduits per stele. Compared with shallow roots, the theoretical hydraulic conductivities in deep roots were enhanced by 133% (J. mandshurica), 78% (F. mandschurica) and 217% (P. amurense), respectively, indicating higher efficiency of transportation. Our results suggest that trees' root tip anatomical structure and physiological activity vary substantially with soil environment. The Author 2015. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
[本文引用: 1]
[本文引用: 1]
DOIURLPMID [本文引用: 3]
Root systems perform the crucial task of absorbing water from the soil to meet the demands of a transpiring canopy. Roots are thought to operate like electrical fuses, which break when carrying an excessive load under conditions of drought stress. Yet the exact site and sequence of this dysfunction in roots remains elusive. Using in vivo X-ray computed microtomography (microCT), we found that... [Show full abstract]
[本文引用: 2]
DOIURL [本文引用: 1]
In order to alleviate the soil drying of the jujube forest in semiarid loess area, we took the gradient control on jujube tree specifications by pruning, using the TDP tree rod liquid flow monitoring system to monitor the jujube tree transpiration and CNC100 neutron moisture meter to monitor soil moisture. It was found that the pruning on jujube had a significant influence on the transpiration and water consumption at different time scales through the research for two consecutive years. Pruning can significantly reduce the tree transpiration in flowering period of fruit and fruit enlargement period of jujube, and reduce the consumption of soil moisture. Compared with the control treatments, the tree transpiration with mild, moderate and severe pruning of the whole growth period decreased by 31.7%, 52.0% and 68.6%, respectively. The relationship between tree transpiration and specifications were analyzed by linear and nonlinear analysis method, and then a mathematical model of natural precipitation and trim production target was built. The suitable target yield for the sustainable management of jujube forest in northern Shaanxi was 1.2 104kg/hm2, and the tree index for water-saving trim regulation was the crown and the new length. The new mode was formed that the production of jujube was determined by soil moisture and jujube specifications were determined by production.
DOIURL [本文引用: 1]
In order to alleviate the soil drying of the jujube forest in semiarid loess area, we took the gradient control on jujube tree specifications by pruning, using the TDP tree rod liquid flow monitoring system to monitor the jujube tree transpiration and CNC100 neutron moisture meter to monitor soil moisture. It was found that the pruning on jujube had a significant influence on the transpiration and water consumption at different time scales through the research for two consecutive years. Pruning can significantly reduce the tree transpiration in flowering period of fruit and fruit enlargement period of jujube, and reduce the consumption of soil moisture. Compared with the control treatments, the tree transpiration with mild, moderate and severe pruning of the whole growth period decreased by 31.7%, 52.0% and 68.6%, respectively. The relationship between tree transpiration and specifications were analyzed by linear and nonlinear analysis method, and then a mathematical model of natural precipitation and trim production target was built. The suitable target yield for the sustainable management of jujube forest in northern Shaanxi was 1.2 104kg/hm2, and the tree index for water-saving trim regulation was the crown and the new length. The new mode was formed that the production of jujube was determined by soil moisture and jujube specifications were determined by production.
DOIURL [本文引用: 1]
林冠是生物圈中物种最丰富却最鲜为人知的生境之一。它在森林与大气的物质、能量交换过程中发挥着至关重要的作用。但因林冠调查技术的限制,林冠及生存在其中的附生植物在生态系统中的功能尚未得到足够的重视。塔吊在三维空间中作业具有"全方位、高精度、非破坏、可重复"的特征。林冠塔吊已成为当前林冠学研究的标志,并为林冠附生植物研究提供了契机。国际上,欧美国家利用塔吊技术对林冠层附生植物多样性与空间分布等进行了大量的研究,取得了丰硕的成果。该文介绍了塔吊的构造、林冠塔吊建设历史和站点分布及国际林冠研究组织等概况,并对依托塔吊开展的附生植物研究进展进行了评述。此外,还简要介绍了我国塔吊建设与林冠生态学发展情况。在系统分析国内外附生植物研究现状基础上,从附生植物多样性、附生植物空间格局与维持机制、生态适应性、与林冠动物的关系以及附生植物对气候变化的响应等5个方面对今后基于林冠塔吊开展附生植物研究进行了展望。
DOIURL [本文引用: 1]
林冠是生物圈中物种最丰富却最鲜为人知的生境之一。它在森林与大气的物质、能量交换过程中发挥着至关重要的作用。但因林冠调查技术的限制,林冠及生存在其中的附生植物在生态系统中的功能尚未得到足够的重视。塔吊在三维空间中作业具有"全方位、高精度、非破坏、可重复"的特征。林冠塔吊已成为当前林冠学研究的标志,并为林冠附生植物研究提供了契机。国际上,欧美国家利用塔吊技术对林冠层附生植物多样性与空间分布等进行了大量的研究,取得了丰硕的成果。该文介绍了塔吊的构造、林冠塔吊建设历史和站点分布及国际林冠研究组织等概况,并对依托塔吊开展的附生植物研究进展进行了评述。此外,还简要介绍了我国塔吊建设与林冠生态学发展情况。在系统分析国内外附生植物研究现状基础上,从附生植物多样性、附生植物空间格局与维持机制、生态适应性、与林冠动物的关系以及附生植物对气候变化的响应等5个方面对今后基于林冠塔吊开展附生植物研究进行了展望。
DOIURL [本文引用: 1]
To establish the methodological basis for developing optimal irrigation strategies for increasing the productivity of triploidPopulus tomentosaplantations using modelling methods, the accuracy of HYDRUS models for simulating one-dimensional (HYDRUS-1D) soil water dynamics under rainfed natural conditions (NC), and two-dimensional soil water dynamics (HYDRUS (2D/3D)) under subsurface drip irrigated (SDI) conditions was evaluated using field data. The relationship between tree growth and soil water availability (rθ) at different depths, which has not been thoroughly investigated in poplar plantations, was also examined. In general, the average soil water content (θ) in different soil layers predicted by both HYDRUS models and theθwithin the two-dimensional domain around drippers predicted by HYDRUS (2D/3D) agreed well with the observed values. Under both treatments, therθincreased with depth and was most variable in the surface 30cm soil. The amount of variation in basal area at breast height (ABH) growth explained byrθin various soil layers ranged widely, suggesting that soil water at different soil depths made different contributions to the variation in growth. The proportion of variation in ABH growth explained by averagerθwas highest (R2=0.709) in the 0–30cm layer, and decreased with increasing integrated depth of the root-zone. Tree growth was unconstrained when therθof the 0–30cm layer was above 0.7. Based on these results, it can be concluded that HYDRUS-1D and HYDRUS (2D/3D) can be used as tools to accurately simulate long-term soil water dynamics inP. tomentosaplantations, at least in sites with similar characteristics to ours. HYDRUS modeling can be used to assess the impacts ofrθon productivity of matureP. tomentosaplantations. This study also shows that monitoring soil moisture of the surface soil provides a robust means for predicting tree growth ofP. tomentosaplantations at sites with similar soil to ours.
[本文引用: 1]
DOIURL [本文引用: 1]
To help achieve precise irrigation inPopulus tomentosaplantations, the stand water use characteristics of a matureP. tomentosaplantation under well-watered drip irrigated conditions were investigated over two growing seasons (April–October) in 2010 and 2011. Crop coefficient models for predicting stand water use were constructed and tested. The quantitative responses of stand water use to groundwater level (GWL) under different irrigation conditions and soil water availability (rθ) in different soil layers, which have not been thoroughly examined in poplar plantations, were also investigated. The stand evapotranspiration (ETa) was dominated by soil evaporation (Es) before late April and after the middle of September, but transpiration (Tr) became the dominating component ofETabetween late April and mid-September accounting for 77%. The mean dailyTr,EsandETafor non-rainfall periods were 2.67, 1.04 and 3.71mmd611, respectively. The relative mean absolute error of the crop coefficient models used to predictETa(13%) andTr(16%) in the non-irrigated treatment (CK) during periods with no water stress were small, suggesting these models can be used to accurately predict stand water use ofP. tomentosaunder well-watered conditions. Fractional transpiration rate ofP. tomentosawas significantly (p<0.0001) correlated torθof different soil layers within 0–70cm depth, but the same relationship was not detected forrθbelow 70cm depth. The proportion of variation inTrexplained byrθwas highest (R2=0.630) in the 0–30cm layer. Tree transpiration was unconstrained when therθof the 0–30cm layer was above 0.6, but ifrθin the surface 30cm soil was not maintained above 0.6, there was a reduction of water uptake and transpiration inP. tomentosa.Significant (p<0.0001) correlation was found between fractional transpiration rate and GWL in the CK treatment, and as the GWL decreased below 300cm depth,TrofP. tomentosadeclined gradually. Whereas, similar phenomenon was not observed in the irrigation treatment. This study therefore indicates (1) that the shallow soil layers should be the key soil zone for irrigation water management in plantations ofP. tomentosaand similar tree species located on sites similar to those in our study, and (2) as the GWL declines below 300cm depth, irrigation should be applied inP. tomentosaplantations to maintain water uptake and tree growth.
DOIURL [本文引用: 2]
In order to find the optimal subsurface drip irrigation (SDI) scheduling for mature triploid Populus tomentosa plantations in the North China Plain, a field experiment was conducted in 2010 and 2011 to investigate the effects of SDI at different soil water potential (SWP) thresholds on the growth and transpiration of a P. tomentosa plantation when it was six and seven years old. The experiment included three SWP treatments, which initiated irrigation when the SWP at 20 cm depth and 10 cm distance from a drip emitter reached 09080825 (T25), 09080850 (T50) and 09080875 (T75) kPa, respectively. A control non-irrigation treatment (CK) was also included. Long-term SWP, soil water content (SWC), transpiration, tree growth, meteorological factors and groundwater level were monitored. Results showed that SDI influenced the SWC only in 009000980 cm of soil. From April to July, on average, the cumulative stand-level transpiration on a ground area basis (E) and growth of basal area at breast height accounted for 81% and 93% of their corresponding whole-season values, while the cumulative reference crop potential evapotranspiration (ET0) was 43% higher than the rainfall. In contrast, from August to October, the growth rate of P. tomentosa was very slow, while the cumulative rainfall was 36% higher than ET0 and the average groundwater level was relatively high (125 cm). Relative to CK, the E under the SWP treatments was significantly (P < 0.05) increased by 2009000973%. Decreasing the SWP irrigation threshold from 09080825 to 09080850 kPa significantly reduced E by 31% (P < 0.05), but decreasing the threshold from 09080850 to 09080875 kPa did not further reduce P. tomentosa E. Relative to CK, T25, T50 and T75 increased the annual above-ground dry biomass (ADB) increment by 54% (P < 0.05), 34% (P < 0.05) and 24% (P > 0.05) in 2010, and by 28% (P > 0.05), 29% (P > 0.05) and 32% (P < 0.05) in 2011, respectively. However, no significant difference in ADB increment was detected among the SWP treatments. Based on these results, it can be concluded that when planting P. tomentosa at sites with similar characteristics to ours in the North China Plain: (1) SDI could be promoted in the cultivation of P. tomentosa to improve tree growth; (2) a range of 09080850 to 09080875 kPa at a depth of 20 cm and 10 cm distant from a drip emitter is recommended as the irrigation threshold for scheduling SDI in P. tomentosa plantations and (3) irrigation should be applied between April and July, while drainage should be implemented between August and October.
DOIURL [本文引用: 4]
The form and water uptake characteristics of the fine root system in a 5-year-old triploid Populus tomentosa plantation were investigated to make recommendations related to irrigation water management for P. tomentosa plantation. Fine roots for analysis were collected from 2106 soil cores taken around eight trees. Soil moisture, trunk sap flow and evaporation were measured concurrently for four months in two experimental plots using time-domain reflectometry, thermal dissipation sensors and micro-lysimeters, respectively. Nearly half (44%) of fine roots corresponded to 0.2-0.5 mm diameter. Generally, lateral root distribution was even, however, the vertical root profile showed an unusual pattern (nearly an 'S' shape). Dense fine roots occurred in surface soil and nearly one third (28%) of total fine roots occurred below 100 cm depth, indicating the plantation had developed a dimorphic root system. With increasing distance from the tree, root distribution tended to be shallower. Mean fine root diameter was significantly larger (P < 0.05) below 120 cm, probably due to soil texture change or/and anoxia. Root water uptake in the 0-20 cm layer contributed 58% of that within the 0-90 cm soil layer, suggesting surface roots played the major water uptake role in shallow soil (<90 cm). On average, P. tomentosa extracted 57% of transpired water from deep soil (>90 cm), implying deep roots can contribute significantly to the water relations of mature P. tomentosa plantations. This functional significance of deep roots might be determined by their high length density and relatively large diameter. Based on these results, three irrigation management strategies were recommended: (1) irrigation schedules should be devised based on periodic measurement of the depth to water table; (2) water should be mainly provided to and maintained in the surface 40 cm soil; and (3) water should be applied to the zone within 1 m from the tree. (c) 2012 Elsevier B.V. All rights reserved.
DOIURL [本文引用: 6]
An experiment was conducted on Haloxylon ammodendron C.A. Mey, a small xeric tree. Soil water content, soil evaporation, leaf water potential, leaf transpiration rate and stomatal conductance were measured at the two sites that contrast in soil texture: sandy and heavy textured, 8 km apart on the southern periphery of Gurbantonggut Desert, Central Asia, during the 2005 and 2006 growing seasons. Leaf specific hydraulic conductance was calculated from the measurements, and root distributions of plants grown at the two sites were quantified by whole-root system excavation. In general, plants grown in sandy soil experienced better water status than in heavy textured soil. Low soil evaporation loss is not the main reason for this better plant water status at sandy site. Plants in sandy soil developed much deeper root systems, larger root surface areas and higher root: leaf surface area ratio than in heavy textured soil, which facilitated plants acquiring more water and surviving the prolonged drought period. Plants growing at light textured sites should have an advantage in acclimatising to the changed water conditions of the future. Plants at the more sandy sites have a larger buffering capacity to excessive variation in ambient conditions.
[本文引用: 2]
DOIURL [本文引用: 1]
Hydraulic lift was investigated among native plants in the Mojave Desert using in situ thermocouple psychrometers. Night lighting and day shading experiments were used to verify the phenomenon. Hydraulic lift was detected for all species examined: five shrub species with different rooting depths and leaf phenologies and one perennial grass species. This study was the first to document hydraulic lift for a CAM species, Yucca schidigera. The pattern of diel flux in soil water potential for the CAM species was temporally opposite to that of C 3 species: for the CAM plant, soil water potential increased in shallow soils during the day when the plant was not transpiring and decreased at night when transpiration began. Because CAM plants transport water to shallow soils during the day when surrounding C 3 and C 4 plants transpire, CAM species that hydraulically lift water may influence water relations of surrounding species to a greater extent than hydraulically lifting C 3 or C 4 species. A strong, negative relationship between the percent sand in the study site soils at the 0.35 m soil depth and the frequency that hydraulic lift was observed at that depth suggests that the occurrence of hydraulic lift is negatively influenced by coarse-textured soils, perhaps due to less root鈥搒oil contact in sandy soils relative to finer-textured soils. Differences in soil texture among study sites may explain, in part, differences in the frequency that hydraulic lift was detected among these species. Further investigations are needed to elucidate species versus soil texture effects on hydraulic lift.
DOIURL [本文引用: 1]
During the night, plant water loss can occur either through the roots, as hydraulic redistribution (HR), or through the leaves via the stoma, as nocturnal transpiration (En), which was methodologically difficult to separate from stem refilling (Re). While HR andEnhave been reported across a range of species, ecosystem, and climate zone, there is little understanding on the interactions betweenEnand/orReand HR. As water movement at night occurs via gradients of water potential, it is expected that during periods of high atmospheric vapor pressure deficit (VPD), water loss viaEnwill override water loss via HR. To test this hypothesis, sap flow in stems and roots ofPopulus euphraticaOliv. trees, growing in a riparian zone in a hyperarid climate, was measured once in a year. Nocturnal stem sap flow was separated intoEnandReusing the “forecasted refilling” method. Substantial nocturnal sap flow (38% of 24‐hr flux on average) was observed and positively correlated with VPD; however, the strength of the correlation was lower (R2=02.55) than diurnal sap flow (Ed) (R2=02.72), suggesting that nocturnal stem sap flow was attributed to both water loss through the canopy and replenishment of water in stem tissues. Partitioning of nocturnal sap flow shows thatReconstituted approximately 80%, andEn~20%, of nocturnal sap flow. The amount of root sap flow attributed to redistribution was negatively related toEd(R2=02.69) and the amount of acropetally sap flow in stems,Re(R2=02.41) andEn(R2=02.14). It was suggested that the magnitude of HR is more strongly depressed byRethat was recharge to the water loss viaEdthan byEn. It was consistent with whole‐tree water balance theory, that the nighttime upward sap flow to xylem, stem refilling and transpiration, may depress hydraulic redistribution of roots.
DOIURL [本文引用: 1]
作为一种重要的生态过程,阐明水力再分配的模式、大小和影响因子 是理解其生态水文效应的前提.选取黑河下游荒漠河岸林主要建群种胡杨为研究对象,采用热比率法辨识胡杨根系水力再分配模式,量化水力再分配大小和影响因 子.结果表明:胡杨不仅具有向上的水力提升,在降水条件下,也具有逆向水力提升,即水力下传或叶片吸收.同时,在河道灌水条件下也具有侧向再分配功能.胡 杨单个根系水力再分配的大小在0.16~0.26 kg/d之间,平均为0.21 kg/d,占第2天蒸腾的比例为38.75%.相关分析和逐步回归分析表明:水力再分配大小与水汽压差、气温、土壤含水量呈显著正相关,而与相对湿度呈显 著负相关,其中水汽压差与土壤含水量是影响胡杨根系水力再分配大小的主导因子.
DOIURL [本文引用: 1]
作为一种重要的生态过程,阐明水力再分配的模式、大小和影响因子 是理解其生态水文效应的前提.选取黑河下游荒漠河岸林主要建群种胡杨为研究对象,采用热比率法辨识胡杨根系水力再分配模式,量化水力再分配大小和影响因 子.结果表明:胡杨不仅具有向上的水力提升,在降水条件下,也具有逆向水力提升,即水力下传或叶片吸收.同时,在河道灌水条件下也具有侧向再分配功能.胡 杨单个根系水力再分配的大小在0.16~0.26 kg/d之间,平均为0.21 kg/d,占第2天蒸腾的比例为38.75%.相关分析和逐步回归分析表明:水力再分配大小与水汽压差、气温、土壤含水量呈显著正相关,而与相对湿度呈显 著负相关,其中水汽压差与土壤含水量是影响胡杨根系水力再分配大小的主导因子.
DOIURL [本文引用: 1]
准确量化植物根系水力提升(HL)及其生态.水文效应对于陆地生态系统水分循环和全球变化研究具有重要意义。基于2011—2012年黑河下游柽柳林地土壤含水量和涡度协方差观测资料,通过将土壤体积含水量分割为HL和水分损失量(WD),结合涡度协方差测定的潜热通量计算的蒸散量(ET),首次定量黑河下游柽柳根系舭及其对Er的贡献。据估算,柽柳根系HL主要发生在20—60(3111深度,生长季HL大小在0-1.4mm/d之间变化,平均为0.22mm/d,WD在0—0.76mm/d之间变化,平均为0.23mm/d,HL与WD的年内变化存在同步性,且HL与WD处于正平衡状态,表明HL通过将深层吸收的土壤水或地下水释放在根系吸收层以供植物蒸腾消耗外,还有剩余水分留存在该层内。生长季ET在0.31—5.38mm/d之间变化,平均为2.82mm/d,但值得注意的是,HL与ET的年内变化存在时间滞后性,HL在5月最高,但E丁在7月最大。HL对ET的贡献率在0.06%-108.25%之间变化。平均为19.25%,比例高于100%的时段主要在生长初期,也就是说在蒸散最大的夏季,HL是相对较小的,其原因可能有2个:一是HL受到夏季深层土壤干化的抑制,二是HL受到夏季夜间蒸腾的抑制,究竟是何种原因还有待进一步研究。
DOIURL [本文引用: 1]
准确量化植物根系水力提升(HL)及其生态.水文效应对于陆地生态系统水分循环和全球变化研究具有重要意义。基于2011—2012年黑河下游柽柳林地土壤含水量和涡度协方差观测资料,通过将土壤体积含水量分割为HL和水分损失量(WD),结合涡度协方差测定的潜热通量计算的蒸散量(ET),首次定量黑河下游柽柳根系舭及其对Er的贡献。据估算,柽柳根系HL主要发生在20—60(3111深度,生长季HL大小在0-1.4mm/d之间变化,平均为0.22mm/d,WD在0—0.76mm/d之间变化,平均为0.23mm/d,HL与WD的年内变化存在同步性,且HL与WD处于正平衡状态,表明HL通过将深层吸收的土壤水或地下水释放在根系吸收层以供植物蒸腾消耗外,还有剩余水分留存在该层内。生长季ET在0.31—5.38mm/d之间变化,平均为2.82mm/d,但值得注意的是,HL与ET的年内变化存在时间滞后性,HL在5月最高,但E丁在7月最大。HL对ET的贡献率在0.06%-108.25%之间变化。平均为19.25%,比例高于100%的时段主要在生长初期,也就是说在蒸散最大的夏季,HL是相对较小的,其原因可能有2个:一是HL受到夏季深层土壤干化的抑制,二是HL受到夏季夜间蒸腾的抑制,究竟是何种原因还有待进一步研究。
DOIURL [本文引用: 2]
AimsMethodsImportant findings The soil water profile can be divided into three layers: shallow relatively wet layer (0.2–1.7 m), middle relatively dry layer (1.7–2.7 m), and deep available water layer. In the shallow relatively wet layer, the soil water content showed obvious diurnal variation, decreased in daytime and increased at night. At the same time, no similar variation of soil water content in the other two layers was observed. Concurrent measurement and comparison of the plant stem water potential and the soil water potential in the shallow relatively wet layer suggested that a positive water potential gradient in root-soil interface would exist at night, which is the physical basis of water efflux from root to soil. The root hydraulic lift of T. ramosissima is the main reason that the shallow soil layer was relatively wet under the arid climate. Further root sampling determined that fine roots in the shallow soil layer were greatly developed, which indicated that the hydraulically lifted water maintained the development of shallow fine roots and ensured root activity. The great development of shallow roots is a probable explanation for the sand-fixation function of T. ramosissima. It was estimated that the percentage of hydraulically lifted water to the daily evapotranspiration in T. ramosissima stand was about 5%–8%. Water from the deep layer accounted for most water consumption of T. ramosissima, which is jointly controlled by soil texture, root uptake and the groundwater table.
DOIURL [本文引用: 2]
AimsMethodsImportant findings The soil water profile can be divided into three layers: shallow relatively wet layer (0.2–1.7 m), middle relatively dry layer (1.7–2.7 m), and deep available water layer. In the shallow relatively wet layer, the soil water content showed obvious diurnal variation, decreased in daytime and increased at night. At the same time, no similar variation of soil water content in the other two layers was observed. Concurrent measurement and comparison of the plant stem water potential and the soil water potential in the shallow relatively wet layer suggested that a positive water potential gradient in root-soil interface would exist at night, which is the physical basis of water efflux from root to soil. The root hydraulic lift of T. ramosissima is the main reason that the shallow soil layer was relatively wet under the arid climate. Further root sampling determined that fine roots in the shallow soil layer were greatly developed, which indicated that the hydraulically lifted water maintained the development of shallow fine roots and ensured root activity. The great development of shallow roots is a probable explanation for the sand-fixation function of T. ramosissima. It was estimated that the percentage of hydraulically lifted water to the daily evapotranspiration in T. ramosissima stand was about 5%–8%. Water from the deep layer accounted for most water consumption of T. ramosissima, which is jointly controlled by soil texture, root uptake and the groundwater table.
DOIURL [本文引用: 3]
在张北地区,以杨树为主的防护林出现不同程度的退化现象,水分是干旱区植物生存的主要限制因子,为揭示水分与防护林退化的关系,本文基于稳定氢氧同位素技术,通过对比小叶杨枝条水与潜在水源的同位素值,探究不同退化程度下小叶杨的水分来源及其对各水源的利用比率.结果表明: 不同退化程度的杨树林水分来源不同,随退化程度的加深,小叶杨的水分来源从深层逐渐向表层转移.无退化的小叶杨主要利用320~400 cm的土壤水,利用率为25.1%;轻度退化的小叶杨主要利用120~180、180~240和240~320 cm的土壤水,对这3层的利用率总和将近50.0%,而对其他土壤水利用较少;中度退化的小叶杨主要利用20~40、40~60和60~80 cm的土壤水,对这3层土壤水每层利用率在17.5%~20.9%范围,对120 cm及其以下的土壤水利用率均低于10.0%;而重度退化的小叶杨主要利用0~20 cm的表层土壤水分,利用率为30.4%,明显高于其他水源.杨树防护林在衰退过程中的水分来源逐渐变浅,而林地浅层土壤较低的土壤含水量无法满足杨树的正常水分需求,加速了杨树林的退化和死亡.
DOIURL [本文引用: 3]
在张北地区,以杨树为主的防护林出现不同程度的退化现象,水分是干旱区植物生存的主要限制因子,为揭示水分与防护林退化的关系,本文基于稳定氢氧同位素技术,通过对比小叶杨枝条水与潜在水源的同位素值,探究不同退化程度下小叶杨的水分来源及其对各水源的利用比率.结果表明: 不同退化程度的杨树林水分来源不同,随退化程度的加深,小叶杨的水分来源从深层逐渐向表层转移.无退化的小叶杨主要利用320~400 cm的土壤水,利用率为25.1%;轻度退化的小叶杨主要利用120~180、180~240和240~320 cm的土壤水,对这3层的利用率总和将近50.0%,而对其他土壤水利用较少;中度退化的小叶杨主要利用20~40、40~60和60~80 cm的土壤水,对这3层土壤水每层利用率在17.5%~20.9%范围,对120 cm及其以下的土壤水利用率均低于10.0%;而重度退化的小叶杨主要利用0~20 cm的表层土壤水分,利用率为30.4%,明显高于其他水源.杨树防护林在衰退过程中的水分来源逐渐变浅,而林地浅层土壤较低的土壤含水量无法满足杨树的正常水分需求,加速了杨树林的退化和死亡.
[本文引用: 2]
[本文引用: 2]
Determining the impacts of experimental forest plantation on groundwater recharge in the Nebraska Sand Hills (USA) using chloride and sulfate
3
2015
... 林分结构的差异和改变能引起树木根系吸水的可塑性变化.与纯林相比, 不同树种混交时, 会促使根系吸水模式可塑性较强的树种在旱季转向主要利用深层土壤水(
... ).但在美国内布拉斯加州中南部,
... 此外, 虽然一般而言密度较大林分的深土层水分消耗会更多,
Deep rooting of rainfed and irrigated orange trees in Brazil
4
2017
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 该方式是通过在单位土体内生长更多的根系, 增大根系吸水面积, 以提高树木深土层中的水分吸收量.土壤水分充足时, 根系吸收面积与吸水量成正比, 因此通过在单位体积土壤内增加吸收根的根长密度、根表面积密度、根系体积等, 来扩大根系在土壤中的吸水面积, 是提高深土层水分吸收的重要方式(
... 该方式是使根系在更大区域内吸收水分, 来提高深根系吸水量.扩大深层根系的分布深度, 不仅可增大其水养资源觅取范围, 而且也能减小根系个体间的资源竞争压力, 所以是树木提高深层水分吸收的最重要和最常见的一种方式.对于同一树种, 这种调节一般在较为干旱的地区比较明显(
... 该方式是通过提高单位长度或面积根系的水分吸收效率(或能力), 来增加根系从深土层吸收的水量, 而且其与增加单位土体根系吸收面积和根系分布范围相比, 可减少根系构建的碳成本投入.由于可缩短水分从土壤进入根系木质部的径向传输距离, 因而减小细根直径可能增大其水分吸收速度和效率, 并同时扩大其水分吸收范围(根系组织密度变化不大的情况下, 比根长增大)(
Combining Eucalyptus wood production with the recovery of native tree diversity in mixed plantings: Implications for water use and availability
3
2018
... 林分结构的差异和改变能引起树木根系吸水的可塑性变化.与纯林相比, 不同树种混交时, 会促使根系吸水模式可塑性较强的树种在旱季转向主要利用深层土壤水(
... ), 而且树种组成、混交方法的差异也会对深土层的水分消耗产生影响.例如,
... 然而, 需要注意的是, 树种混交的这种作用是相对的.因为混交方式或树种不同, 深土层的水分消耗状态会存在很大差异, 这意味着树种混交后, 对于一部分树种, 深层水分状况可能较其纯林变好, 但对另一部分树种来讲, 却可能变差.例如,
Contrasting plant water-use responses to groundwater depth in coastal dune ecosystems
3
2018
... 地下水位变化会影响树木的深层根系吸水特征, 但不同树种响应不同.对于根系分布较深和依赖地下水的树种, 其根系吸水深度一般会随地下水位的降低而逐渐下移(
... ).但对于根系分布较浅和不依赖地下水的树种, 这种根系吸水可塑性变化则较少发生(
... 同一地区不同树种对深层土壤水的吸收利用策略在时间和空间上常存在较大差异, 该现象在干旱(
Influences of root diameter, tree age, soil depth and season on fine root survivorship in Prunus avium
1
2005
... 该方式是通过在单位土体内生长更多的根系, 增大根系吸水面积, 以提高树木深土层中的水分吸收量.土壤水分充足时, 根系吸收面积与吸水量成正比, 因此通过在单位体积土壤内增加吸收根的根长密度、根表面积密度、根系体积等, 来扩大根系在土壤中的吸水面积, 是提高深土层水分吸收的重要方式(
Relative contribution of groundwater to plant transpiration estimated with stable isotopes
4
2017
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 土壤厚度是树木以及其他植物最大根系分布深度的重要影响因子, 但很多情况下其并不能制约根系的最终分布深度(
... 在加利福尼亚北部海岸山脉上的针阔常绿混交林中发现, 虽然土壤厚度只有30-75 cm, 但树木却能扎根至风化岩石层中(深度达16 m), 来吸收其中较土壤水更加丰富的稳定岩石水(rock moisture);
... 地下水位变化会影响树木的深层根系吸水特征, 但不同树种响应不同.对于根系分布较深和依赖地下水的树种, 其根系吸水深度一般会随地下水位的降低而逐渐下移(
Impacts of hydraulic redistribution on grass-tree competition vs facilitation in a semi-arid savanna
1
2017
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
Loss of deep roots limits biogenic agents of soil development that are only partially restored by decades of forest regeneration
1
2018
... 深层根系是植物借以吸收深层土壤水的唯一工具, 其在树木水分关系中的重要性早在近一个世纪以前就被人们所关注(
Fertilization increases the functional specialization of fine roots in deep soil layers for young Eucalyptus grandis trees
1
2018
... 然而, 需要注意的是, 真菌的侵染(形成菌根)可能会降低深根系性状可塑性的程度, 而且根系类型定义的不同(如以直径<0.5、1或2 mm等来定义细根)也可能使这种变化难以被发觉(
Norway spruce fine roots and fungal hyphae grow deeper in forest soils after extended drought
1
2017
... 根系吸水过程中, 水分从根系表皮穿过皮层, 然后跨过内皮层进入木质部导管过程中, 可以通过共质体、质外体和跨膜等3种途径进行运输, 其中均涉及水分的跨膜运输(主要是跨膜途径), 而细胞膜上的水孔蛋白在水分子的跨膜中起主要控制作用(
Deep soil water dynamics in an undisturbed primary forest in central Amazonia: Differences between normal years, and the 2005 drought
2
2017
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 潜在蒸散量和降水是影响树木深层根系吸水的重要气候因子.当潜在蒸散量增大时, 林木蒸腾量会相应变大, 其根系吸水范围会向更深的土层发展(
Coordination of rooting depth and leaf hydraulic traits defines drought-related strategies in the campos rupestres, a tropical montane biodiversity hotspot
5
2017
... 了解树木对深层土壤水的吸收利用特征, 需首先对深层土壤进行界定, 但深层土壤并非严格的科学概念, 而且全球不同地区土壤厚度差异很大(如土石山区与黄土高原)(
... 从深层土壤中吸收水分运输至地上部分器官供给蒸腾和生长, 是树木对深层土壤水的最直接利用方式(
... 根系、茎干、枝条、叶片四个水力器官组成了植物的水分传输系统, 四者在保障整株水力路径中水分的传输上是功能统一的, 其中任一器官(尤其是叶片和根系)木质部水力导度下降或水力失调都会直接影响整株植物的水分传输效率, 从而影响叶片气体交换以及植物生存和生长(
... 现有的少数研究结果显示, 树木在发展出较深的根系而能获取相对稳定的水源后, 为实现深层土壤水利用最大化, 会对叶片水力性状进行调节.例如, 会使树木的单位叶面积(“水分散失”面积)具有更大的根系表面积(“水分供给”面积), 从而使其在生长季维持相对稳定且更高的水分状况(较高的黎明前叶水势)和气孔导度, 以保持较大的蒸腾速率, 并使叶片相应具有较高的光合速率(
... ).此外, 有研究发现深根系树种的叶片失膨点水势高于(绝对值更小)浅根系树种(
Soil water availability and rooting depth as determinants of hydraulic architecture of Patagonian woody species
2
2009
... 现有的少数研究结果显示, 树木在发展出较深的根系而能获取相对稳定的水源后, 为实现深层土壤水利用最大化, 会对叶片水力性状进行调节.例如, 会使树木的单位叶面积(“水分散失”面积)具有更大的根系表面积(“水分供给”面积), 从而使其在生长季维持相对稳定且更高的水分状况(较高的黎明前叶水势)和气孔导度, 以保持较大的蒸腾速率, 并使叶片相应具有较高的光合速率(
... 各器官木质部的解剖结构及其相对变化特征, 决定了树木体内的水分传输效率(
Hydraulic lift: Consequences of water efflux from the roots of plants
1
1998
... 水力提升(HL)是树木实现深层土壤水吸收利用的另一种方式, 同时也是除深根系和气孔调节之外的另一重要抗旱机制.该现象是指在浅土层与深土层存在水势梯度的情况下, 植物深层根系借助水势差在夜间将深土层中的水分运输至表层根系, 然后再释放至浅土层中以供白天蒸腾所需(
Maximum rooting depth of vegetation types at the global scale
4
1996
... 深层根系是植物借以吸收深层土壤水的唯一工具, 其在树木水分关系中的重要性早在近一个世纪以前就被人们所关注(
... 根系是树木吸收和传输土壤水分和养分资源的重要功能器官(
... 现有研究表明, 影响树木深根系产生的因子很多, 但归纳起来可分为植被因子(树种、生活型、植被类型、林龄、个体发育、物种竞争)、气候因子(降水量、潜在蒸散量)、土壤因子(土层厚度、土壤质地、土壤有机层厚度、土壤水分、土壤养分)和水文因子(地下水位), 且不同空间尺度上主要影响因子不同(
... 全球范围内, 不同生活型和植被类型间, 树木根系分布深度也存在很大差异.平均而言, 常绿阔叶树((6.30 ± 10.17) m) >落叶阔叶树((3.71 ± 6.99) m) >常绿针叶树((1.79 ± 2.79) m) >落叶针叶树((0.78 ± 0.57) m) (
黑河下游荒漠河岸林植物水分传输观测试验研究
5
2016
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 地下水位变化会影响树木的深层根系吸水特征, 但不同树种响应不同.对于根系分布较深和依赖地下水的树种, 其根系吸水深度一般会随地下水位的降低而逐渐下移(
... 同一地区不同树种对深层土壤水的吸收利用策略在时间和空间上常存在较大差异, 该现象在干旱(
... ;
... 国外对HL的研究较多, 已从简单的现象描述发展到定量评价、系统机理解释乃至与全球气候等结合探讨HL对气候变化以及森林动态的影响.国内对HL的研究起步较晚, 20世纪90年代中期开始至今, 相关的中文研究性论文仅有20多篇; 这些研究主要集中在西北干旱、半干旱地区和华北低丘山区的少数树种上, 如胡杨、多枝柽柳、胡桃等(
黑河下游荒漠河岸林植物水分传输观测试验研究
5
2016
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 地下水位变化会影响树木的深层根系吸水特征, 但不同树种响应不同.对于根系分布较深和依赖地下水的树种, 其根系吸水深度一般会随地下水位的降低而逐渐下移(
... 同一地区不同树种对深层土壤水的吸收利用策略在时间和空间上常存在较大差异, 该现象在干旱(
... ;
... 国外对HL的研究较多, 已从简单的现象描述发展到定量评价、系统机理解释乃至与全球气候等结合探讨HL对气候变化以及森林动态的影响.国内对HL的研究起步较晚, 20世纪90年代中期开始至今, 相关的中文研究性论文仅有20多篇; 这些研究主要集中在西北干旱、半干旱地区和华北低丘山区的少数树种上, 如胡杨、多枝柽柳、胡桃等(
The effects of groundwater depth on water uptake of Populus euphratica and Tamarix ramosissima in the hyperarid region of Northwestern China
2016
The roots of the drought: Hydrology and water uptake strategies mediate forest-wide demographic response to precipitation
2
2018
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 模式I和II很难使人工林达到较高的生产力水平(
Global convergence in the vulnerability of forests to drought
4
2012
... 根系、茎干、枝条、叶片四个水力器官组成了植物的水分传输系统, 四者在保障整株水力路径中水分的传输上是功能统一的, 其中任一器官(尤其是叶片和根系)木质部水力导度下降或水力失调都会直接影响整株植物的水分传输效率, 从而影响叶片气体交换以及植物生存和生长(
... 现有的少数研究结果显示, 树木在发展出较深的根系而能获取相对稳定的水源后, 为实现深层土壤水利用最大化, 会对叶片水力性状进行调节.例如, 会使树木的单位叶面积(“水分散失”面积)具有更大的根系表面积(“水分供给”面积), 从而使其在生长季维持相对稳定且更高的水分状况(较高的黎明前叶水势)和气孔导度, 以保持较大的蒸腾速率, 并使叶片相应具有较高的光合速率(
... 各器官木质部的解剖结构及其相对变化特征, 决定了树木体内的水分传输效率(
... ); 但对于同一树种, 根系变深而能持续获取稳定深层水后, 其茎干新生木质部的解剖结构可能会发生变化, 因为茎干支撑的蒸腾水流最大, 所以对生长环境的变化可能更敏感(
Almost symmetrical vertical growth rates above and below ground in one the world’s most productive forests
2
2011
... 植物根系系统的大小(分布深度、水平延伸距离)与其地上部分的大小间存在显著正相关关系(
... 当土壤有机层较厚时, 土壤养分充足, 一般而言, 树木可能不需扎根很深便可获取充足的土壤养分.现有数据表明, 在寒温带针叶林、温带和热带森林中, d95均会随着土壤有机层厚度的减小而变深(
Simulating the effects of different potassium and water supply regimes on soil water content and water table depth over a rotation of a tropical Eucalyptus grandis plantation
4
2018
... 潜在蒸散量和降水是影响树木深层根系吸水的重要气候因子.当潜在蒸散量增大时, 林木蒸腾量会相应变大, 其根系吸水范围会向更深的土层发展(
... 施肥策略的差异会对林木的水分利用以及林木与深土层水分间的关系产生影响(
... ), 如果在制定具体施肥制度时能对相关信息予以充分考虑, 则有助于提高林木对干旱的抗性和应对气候变化的韧性.如
... 此外,
Importance of deep water uptake in tropical eucalypt forest
8
2017
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... ;
... 植物根系系统的大小(分布深度、水平延伸距离)与其地上部分的大小间存在显著正相关关系(
... 树木根系吸水特征会因树体大小、健康状况以及根区位置的不同而发生改变.其中, 关于树体大小(个体发育)与根系吸水深度间的关系目前存在不一致的结果: 多数研究发现树体越大, 深层土壤水吸收比例会越大(
... 同一树种对深层土壤水的吸收利用存在很强的时空变异性.具备深根系虽能保证树木同时利用不同土层的水分和(或)地下水, 但却存在明显的季节和年际变化: 湿润季节为高效利用降水会优先从浅土层吸收水分, 而干旱季节则会转向吸收更多的深层土壤水和(或)地下水(
... 综上所述, 整体而言, 与树木吸收利用浅土层水分的研究相比, 目前关于树木吸收利用深层水的研究还相对较少, 现有相关研究主要集中在我国西北、西非、美国西部、澳大利亚西部、伊比利亚半岛、巴西中部等干旱、半干旱、季节性干旱地区.此外, 现有的研究在测定树木对深层土壤水的利用时, 很少有直接测定至树木根区的下限, 即大多数研究对树木深层吸水模式和特征的了解并不“完整”.同时, 这些研究也大多是在1个生长季内的1个或几个时期开展, 而针对较长的时间尺度, 类似的试验研究极少, 目前只有
... 模式I和II很难使人工林达到较高的生产力水平(
... 在对人工林进行林分结构调控时(如抚育间伐、树种调控等), 间伐木的选择是关键.虽然多个研究表明, 个体较大的林木会更多地吸收深层土壤水(
Root traits contributing to plant productivity under drought
1
2013
... 根系吸水过程中, 水分从根系表皮穿过皮层, 然后跨过内皮层进入木质部导管过程中, 可以通过共质体、质外体和跨膜等3种途径进行运输, 其中均涉及水分的跨膜运输(主要是跨膜途径), 而细胞膜上的水孔蛋白在水分子的跨膜中起主要控制作用(
Effects of single Quercus ilex trees upon spatial and seasonal changes in soil water content in dehesas of central western Spain
1
2007
... 树木根系吸水特征会因树体大小、健康状况以及根区位置的不同而发生改变.其中, 关于树体大小(个体发育)与根系吸水深度间的关系目前存在不一致的结果: 多数研究发现树体越大, 深层土壤水吸收比例会越大(
Functional specialization of Eucalyptus fine roots: Contrasting potential uptake rates for nitrogen, potassium and calcium tracers at varying soil depths
1
2011
... 然而, 需要注意的是, 真菌的侵染(形成菌根)可能会降低深根系性状可塑性的程度, 而且根系类型定义的不同(如以直径<0.5、1或2 mm等来定义细根)也可能使这种变化难以被发觉(
Streamside trees that do not use stream water
2
1991
... 树木根系吸水特征会因树体大小、健康状况以及根区位置的不同而发生改变.其中, 关于树体大小(个体发育)与根系吸水深度间的关系目前存在不一致的结果: 多数研究发现树体越大, 深层土壤水吸收比例会越大(
... 在对人工林进行林分结构调控时(如抚育间伐、树种调控等), 间伐木的选择是关键.虽然多个研究表明, 个体较大的林木会更多地吸收深层土壤水(
Liana and tree below-ground water competition— Evidence for water resource partitioning during the dry season
1
2018
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
Quantifying root water extraction after drought recovery using sub-mm in situ empirical data
1
2017
... 该方式是通过提高单位长度或面积根系的水分吸收效率(或能力), 来增加根系从深土层吸收的水量, 而且其与增加单位土体根系吸收面积和根系分布范围相比, 可减少根系构建的碳成本投入.由于可缩短水分从土壤进入根系木质部的径向传输距离, 因而减小细根直径可能增大其水分吸收速度和效率, 并同时扩大其水分吸收范围(根系组织密度变化不大的情况下, 比根长增大)(
Root-system characteristics of plantation-grown Populus tomentosa adapted to seasonal fluctuation in the groundwater table
2
2018
... 地下水位之所以对根系最大分布深度起较强的控制作用, 主要是因为缺氧会限制根系的生长.但也有研究发现, 当地下水中有较多的溶解氧和养分时, 根系也会突破地下水的限制, 在地下水位以下生存(
... 该方式是使根系在更大区域内吸收水分, 来提高深根系吸水量.扩大深层根系的分布深度, 不仅可增大其水养资源觅取范围, 而且也能减小根系个体间的资源竞争压力, 所以是树木提高深层水分吸收的最重要和最常见的一种方式.对于同一树种, 这种调节一般在较为干旱的地区比较明显(
Native root xylem embolism and stomatal closure in stands of Douglas-fir and ponderosa pine: Mitigation by hydraulic redistribution
1
2004
... HL的大小受多种环境因素影响.根系能触及地下水或毛管边缘水时, 会促进HL的发生或提高其水分提升量, 尤其是在深层土壤质地较粗、持水力较弱的情况下, 如
Hydrologic regulation of plant rooting depth
19
2017
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 土壤是植物生长的介质, 故而深层土壤应是相对于植物而定义的概念,
... 综上所述, 就所有陆地植被类型平均而言, 可将1 m以下土层中的根系定义为深根系, 而该土层即为深土层.虽然其在特定气候、土壤和植被类型下的适用性有限, 但基于d95数据(
... 根系是树木吸收和传输土壤水分和养分资源的重要功能器官(
... ;
... 现有研究表明, 影响树木深根系产生的因子很多, 但归纳起来可分为植被因子(树种、生活型、植被类型、林龄、个体发育、物种竞争)、气候因子(降水量、潜在蒸散量)、土壤因子(土层厚度、土壤质地、土壤有机层厚度、土壤水分、土壤养分)和水文因子(地下水位), 且不同空间尺度上主要影响因子不同(
... 树木根系的分布深度本质上由基因决定, 所以不同树种间的根系分布深度差异很大.例如, 对于金合欢属(Acacia, (12.85 ± 17.99) m)、桉属(Eucalyptus, (8.71 ± 8.75) m)、栎属(Quercus, (5.23 ± 5.79) m)、杨属(Populus, (3.00 ± 4.75) m)、松属(Pinus, (2.45 ± 3.92) m)、刺柏属(Juniperus, (2.41 ± 2.40) m)、槭属(Acer, (1.75 ± 1.26) m)、柳属(Salix, (1.35 ± 1.53) m)、冷杉属(Abies, (1.30 ± 0.67) m)、黄杉属(Pseudotsuga, (1.03 ± 0.37) m)、落叶松属(Larix, (0.90 ± 0.67) m)、水青冈属(Fagus, (0.83 ± 0.46) m)和云杉属(Picea, (0.74 ± 0.47) m)等主要人工林树种, 目前观测到的其平均根系深度最大可相差16倍(
... 全球范围内, 不同生活型和植被类型间, 树木根系分布深度也存在很大差异.平均而言, 常绿阔叶树((6.30 ± 10.17) m) >落叶阔叶树((3.71 ± 6.99) m) >常绿针叶树((1.79 ± 2.79) m) >落叶针叶树((0.78 ± 0.57) m) (
... 现有研究显示, 在全球大尺度上, 年降水量(MAP)和年潜在蒸散量(PET)可能是影响树木根系分布的两个重要气候因子(
... 土壤厚度是树木以及其他植物最大根系分布深度的重要影响因子, 但很多情况下其并不能制约根系的最终分布深度(
... 一般而言, 土壤质地因影响水分入渗深度和毛管水上升高度而能对树木及其他植物根系分布深度产生影响(
... )根系的深度为3 m, 但在相距8 km的壤砂土上则达到10 m.然而, 依据
... 对多种植物共计1 426个根系样本数据的分析结果, 可以看出土壤质地在全球尺度上对树木最大根系分布深度的影响可能较小.这主要是因为较粗质地土壤上的水分入渗虽然较深, 但质地较细土壤上的毛管上升水高度却较大, 而这二者的作用均会诱导深根系的产生(
... 虽然气候因子能对全球尺度上树木根系的分布深度产生明显影响, 但相同气候下却常出现较浅与较深根系分布的同时存在.这种现象可能由树种、林龄、土壤质地、种间竞争等因素引起(
... ).
... 全球范围内, 整体而言, 当地下水位较深, 远远超出根系可达范围时, 在干旱气候区, 降水较少且入渗较浅, 受降水入渗深度的限制, 根系分布也较浅; 在过湿气候区, 降水多而频繁, 浅土层始终持有充足水分, 树木不需产生深根便可获取足够的蒸腾用水, 因此根系分布也相对较浅; 但在季节性干旱气候区, 湿润季节较深的降水入渗会诱导深土层根系的增殖, 促进形成相对较深的根系分布.当地下水位相对较浅时, 在过湿气候区, 树木仍不需产生较深根系便可获取充足水分; 但在干旱和季节性干旱气候区, 毛管上升水会诱使树木形成较深根系, 以吸收深层土壤水和地下水来满足旱季较高的蒸腾所需(
... 从深层土壤中吸收水分运输至地上部分器官供给蒸腾和生长, 是树木对深层土壤水的最直接利用方式(
... 树木对深层土壤水的吸收利用, 从本质上讲, 主要是受到土壤水分垂直分布和植株蒸腾耗水的共同驱动, 因为二者分别决定了土壤中的水分供给能力和树木对水分的需求大小(
... 根系具有很强的可塑性, 对土壤环境非常敏感(
Aquaporin and root water uptake
2
2017
... 根系吸水过程中, 水分从根系表皮穿过皮层, 然后跨过内皮层进入木质部导管过程中, 可以通过共质体、质外体和跨膜等3种途径进行运输, 其中均涉及水分的跨膜运输(主要是跨膜途径), 而细胞膜上的水孔蛋白在水分子的跨膜中起主要控制作用(
... ;
Unexpected phenology and lifespan of shallow and deep fine roots of walnut trees grown in a silvoarable Mediterranean agroforestry system
5
2016
... 土壤是植物生长的介质, 故而深层土壤应是相对于植物而定义的概念,
... 该方式是通过在单位土体内生长更多的根系, 增大根系吸水面积, 以提高树木深土层中的水分吸收量.土壤水分充足时, 根系吸收面积与吸水量成正比, 因此通过在单位体积土壤内增加吸收根的根长密度、根表面积密度、根系体积等, 来扩大根系在土壤中的吸水面积, 是提高深土层水分吸收的重要方式(
... ;
... 为保证深层根系能在旱季或表土层干燥时为地上部分器官及时供给充足水分, 树木会对深层根系的物候(存在明显季节性)进行调节, 以使其能适时、及时地激活和发挥水分供给功能: 在旱季树木对深层土壤水有较强需求时, 会快速启动深层根系的生长去觅取和吸收深层水源, 或进入休眠季地上部分器官不再生长时, 将更多的碳水化合物用于深根系生长, 以为次年吸收利用深层水做“材料准备” (
... )复合系统中, 从春季至夏初, 胡桃深层根系(深>2.5 m)生长的启动时间滞后于表层根系, 且在冬季落叶后, 深层根系出现第二个生长高峰(
Consequences of mixing Acacia mangium and Eucalyptus grandis on soil exploration by fine-roots down to a depth of 17 m
2
2017
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 合理的树种混交可促进林木对深层水资源的开采能力, 而且还可能通过水力提升作用将深土层中的水分转移至浅土层供混交的浅根系树种利用, 从而提高气候变化条件下混交林生态系统的稳定性和抗性.如, 在巴西的桉树与相思(Acacia mangium)混交林中, 0-17 m土层内单位面积土壤上的细根生物量远高于两个树种的纯林, 其细根比根长和比根表面积比相思纯林高110%-150%, 比桉树纯林高34%; 且混交林9-17 m土层内的相思和桉树细根比根长和比根表面积都显著高于各自纯林, 表明混交林深土层水资源的吸收利用能力远高于纯林, 从而使干旱条件下混交林中的深土层能更好地发挥其“水分缓冲库”的作用(
Dynamic niche partitioning in root water uptake facilitates efficient water use in more diverse grassland plant communities
2
2018
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 潜在蒸散量和降水是影响树木深层根系吸水的重要气候因子.当潜在蒸散量增大时, 林木蒸腾量会相应变大, 其根系吸水范围会向更深的土层发展(
Tree soil water uptake and transpiration in mono-cultural and jungle rubber stands of Sumatra
3
2017
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 树木根系吸水特征会因树体大小、健康状况以及根区位置的不同而发生改变.其中, 关于树体大小(个体发育)与根系吸水深度间的关系目前存在不一致的结果: 多数研究发现树体越大, 深层土壤水吸收比例会越大(
... 林分结构的差异和改变能引起树木根系吸水的可塑性变化.与纯林相比, 不同树种混交时, 会促使根系吸水模式可塑性较强的树种在旱季转向主要利用深层土壤水(
Weathering of rock to regolith: The activity of deep roots in bedrock fractures
1
2017
... 土壤厚度是树木以及其他植物最大根系分布深度的重要影响因子, 但很多情况下其并不能制约根系的最终分布深度(
Possibilities of Shelterbelt Planting in the Plains Regions
1
1935
... 深层根系是植物借以吸收深层土壤水的唯一工具, 其在树木水分关系中的重要性早在近一个世纪以前就被人们所关注(
华北低丘山区果药复合系统种间水分利用策略
2
2016
... 了解树木对深层土壤水的吸收利用特征, 需首先对深层土壤进行界定, 但深层土壤并非严格的科学概念, 而且全球不同地区土壤厚度差异很大(如土石山区与黄土高原)(
... 国外对HL的研究较多, 已从简单的现象描述发展到定量评价、系统机理解释乃至与全球气候等结合探讨HL对气候变化以及森林动态的影响.国内对HL的研究起步较晚, 20世纪90年代中期开始至今, 相关的中文研究性论文仅有20多篇; 这些研究主要集中在西北干旱、半干旱地区和华北低丘山区的少数树种上, 如胡杨、多枝柽柳、胡桃等(
华北低丘山区果药复合系统种间水分利用策略
2
2016
... 了解树木对深层土壤水的吸收利用特征, 需首先对深层土壤进行界定, 但深层土壤并非严格的科学概念, 而且全球不同地区土壤厚度差异很大(如土石山区与黄土高原)(
... 国外对HL的研究较多, 已从简单的现象描述发展到定量评价、系统机理解释乃至与全球气候等结合探讨HL对气候变化以及森林动态的影响.国内对HL的研究起步较晚, 20世纪90年代中期开始至今, 相关的中文研究性论文仅有20多篇; 这些研究主要集中在西北干旱、半干旱地区和华北低丘山区的少数树种上, 如胡杨、多枝柽柳、胡桃等(
地面滴灌对107杨幼林生长和水肥利用的影响
2
2015
... 模式I和II很难使人工林达到较高的生产力水平(
... ;
地面滴灌对107杨幼林生长和水肥利用的影响
2
2015
... 模式I和II很难使人工林达到较高的生产力水平(
... ;
Physical environmental controls on riparian root profiles associated with black poplar ( Populus nigra L.) along the Tagliamento River, Italy
1
2017
... 浅土层干旱虽不会决定深根系的发生, 但会促进深根系的发展.如
Seasonal and diurnal patterns of soil water potential in the rhizosphere of blue oaks: Evidence for hydraulic lift
1
2000
... HL的大小受多种环境因素影响.根系能触及地下水或毛管边缘水时, 会促进HL的发生或提高其水分提升量, 尤其是在深层土壤质地较粗、持水力较弱的情况下, 如
A global budget for fine root biomass, surface area, and nutrient contents
3
1997
... 根系是树木吸收和传输土壤水分和养分资源的重要功能器官(
... ;
... 树木根系主要分布于浅土层, 深层土壤中根系一般较少(
Impact of elevated atmospheric humidity on anatomical and hydraulic traits of xylem in hybrid aspen
1
2015
... 各器官木质部的解剖结构及其相对变化特征, 决定了树木体内的水分传输效率(
植物叶片水力与经济性状权衡关系的研究进展
1
2015
... 根系、茎干、枝条、叶片四个水力器官组成了植物的水分传输系统, 四者在保障整株水力路径中水分的传输上是功能统一的, 其中任一器官(尤其是叶片和根系)木质部水力导度下降或水力失调都会直接影响整株植物的水分传输效率, 从而影响叶片气体交换以及植物生存和生长(
植物叶片水力与经济性状权衡关系的研究进展
1
2015
... 根系、茎干、枝条、叶片四个水力器官组成了植物的水分传输系统, 四者在保障整株水力路径中水分的传输上是功能统一的, 其中任一器官(尤其是叶片和根系)木质部水力导度下降或水力失调都会直接影响整株植物的水分传输效率, 从而影响叶片气体交换以及植物生存和生长(
a). Role of aquaporin activity in regulating deep and shallow root hydraulic conductance during extreme drought
1
2014
... 根系吸水过程中, 水分从根系表皮穿过皮层, 然后跨过内皮层进入木质部导管过程中, 可以通过共质体、质外体和跨膜等3种途径进行运输, 其中均涉及水分的跨膜运输(主要是跨膜途径), 而细胞膜上的水孔蛋白在水分子的跨膜中起主要控制作用(
Contrasting hydraulic architecture and function in deep and shallow roots of tree species from a semi-arid habitat
4
2014b
... 为提高深根系水分传输能力, 树木会对其粗根以及细根(≤2 mm)中传输根(木质化、主要起传输和储藏作用的根系)(
... ).此外, 树木还可通过增加深根系中具有相互连通性的导管的比例以及导管上纹孔的平均尺寸或数量, 来提高不同导管之间的水分传输, 从而增大根系导水率(
... 深土层细根中的吸收根(非木质化、吸收水养资源的根系)(
... 各器官木质部的解剖结构及其相对变化特征, 决定了树木体内的水分传输效率(
A test of the hydraulic vulnerability segmentation hypothesis in angiosperm and conifer tree species
3
2016
... 根系、茎干、枝条、叶片四个水力器官组成了植物的水分传输系统, 四者在保障整株水力路径中水分的传输上是功能统一的, 其中任一器官(尤其是叶片和根系)木质部水力导度下降或水力失调都会直接影响整株植物的水分传输效率, 从而影响叶片气体交换以及植物生存和生长(
... ;
... 各器官木质部的解剖结构及其相对变化特征, 决定了树木体内的水分传输效率(
Influence of root diameter and soil depth on the xylem anatomy of fine- to medium-sized roots of mature beech trees in the top- and subsoil
6
2017
... 该方式是通过提高单位长度或面积根系的水分吸收效率(或能力), 来增加根系从深土层吸收的水量, 而且其与增加单位土体根系吸收面积和根系分布范围相比, 可减少根系构建的碳成本投入.由于可缩短水分从土壤进入根系木质部的径向传输距离, 因而减小细根直径可能增大其水分吸收速度和效率, 并同时扩大其水分吸收范围(根系组织密度变化不大的情况下, 比根长增大)(
... 为提高深根系水分传输能力, 树木会对其粗根以及细根(≤2 mm)中传输根(木质化、主要起传输和储藏作用的根系)(
... ).这是因为根据泊肃叶定律, 水分传输管道(导管)中的水流量对管道半径极其敏感, 在驱动力压力梯度和液体黏度不变的情况下, 当管道半径加倍时, 其中的水流体积流速将增加16倍.深根系中还可能特化出一些因具有较大直径导管而具有超高导度的根系(high conductivity roots), 这种根系的导水率可较同径级根系的平均导水率高出50%以上(
... 深土层细根中的吸收根(非木质化、吸收水养资源的根系)(
... 然而, 需要注意的是, 真菌的侵染(形成菌根)可能会降低深根系性状可塑性的程度, 而且根系类型定义的不同(如以直径<0.5、1或2 mm等来定义细根)也可能使这种变化难以被发觉(
... 各器官木质部的解剖结构及其相对变化特征, 决定了树木体内的水分传输效率(
A method of determining rooting depth from a terrestrial biosphere model and its impacts on the global water and carbon cycle
1
1998
... 土壤是植物生长的介质, 故而深层土壤应是相对于植物而定义的概念,
How do rubber ( Hevea brasiliensis) plantations behave under seasonal water stress in northeastern Thailand and central Cambodia
1
2015
... 同一树种对深层土壤水的吸收利用存在很强的时空变异性.具备深根系虽能保证树木同时利用不同土层的水分和(或)地下水, 但却存在明显的季节和年际变化: 湿润季节为高效利用降水会优先从浅土层吸收水分, 而干旱季节则会转向吸收更多的深层土壤水和(或)地下水(
Dynamics of soil exploration by fine roots down to a depth of 10 m throughout the entire rotation in Eucalyptus grandisplantations
1
2013
... 植物根系系统的大小(分布深度、水平延伸距离)与其地上部分的大小间存在显著正相关关系(
Growth of the whole root system for a plant crop of sugarcane under rainfed and irrigated environments in Brazil
1
2009
... 该方式是使根系在更大区域内吸收水分, 来提高深根系吸水量.扩大深层根系的分布深度, 不仅可增大其水养资源觅取范围, 而且也能减小根系个体间的资源竞争压力, 所以是树木提高深层水分吸收的最重要和最常见的一种方式.对于同一树种, 这种调节一般在较为干旱的地区比较明显(
Contrasting phenology of Eucalyptus grandisfine roots in upper and very deep soil layers in Brazil
3
2017
... 该方式是通过在单位土体内生长更多的根系, 增大根系吸水面积, 以提高树木深土层中的水分吸收量.土壤水分充足时, 根系吸收面积与吸水量成正比, 因此通过在单位体积土壤内增加吸收根的根长密度、根表面积密度、根系体积等, 来扩大根系在土壤中的吸水面积, 是提高深土层水分吸收的重要方式(
... 为保证深层根系能在旱季或表土层干燥时为地上部分器官及时供给充足水分, 树木会对深层根系的物候(存在明显季节性)进行调节, 以使其能适时、及时地激活和发挥水分供给功能: 在旱季树木对深层土壤水有较强需求时, 会快速启动深层根系的生长去觅取和吸收深层水源, 或进入休眠季地上部分器官不再生长时, 将更多的碳水化合物用于深根系生长, 以为次年吸收利用深层水做“材料准备” (
... ).这可能是树木在平衡资源分配、协调各器官生长的同时, 为能高效利用深层水而采取的一种时间上的生态策略.例如, 在巴西桉树人工林中, 当干旱导致浅土层水分较少而浅层根系产量明显降低时, 4 m以下土层中的根系生长速度大幅快速升高(
Root functioning modifies seasonal climate
1
2005
... 根系是树木吸收和传输土壤水分和养分资源的重要功能器官(
Plant diversity and drought: The role of deep roots
1
2014
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
黄土半干旱区枣林深层土壤水分消耗特征
3
2014
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 了解树木对深层土壤水的吸收利用特征, 需首先对深层土壤进行界定, 但深层土壤并非严格的科学概念, 而且全球不同地区土壤厚度差异很大(如土石山区与黄土高原)(
... 同一树种对深层土壤水的吸收利用存在很强的时空变异性.具备深根系虽能保证树木同时利用不同土层的水分和(或)地下水, 但却存在明显的季节和年际变化: 湿润季节为高效利用降水会优先从浅土层吸收水分, 而干旱季节则会转向吸收更多的深层土壤水和(或)地下水(
黄土半干旱区枣林深层土壤水分消耗特征
3
2014
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 了解树木对深层土壤水的吸收利用特征, 需首先对深层土壤进行界定, 但深层土壤并非严格的科学概念, 而且全球不同地区土壤厚度差异很大(如土石山区与黄土高原)(
... 同一树种对深层土壤水的吸收利用存在很强的时空变异性.具备深根系虽能保证树木同时利用不同土层的水分和(或)地下水, 但却存在明显的季节和年际变化: 湿润季节为高效利用降水会优先从浅土层吸收水分, 而干旱季节则会转向吸收更多的深层土壤水和(或)地下水(
密植枣林地深层土壤水分垂直变化与根系分布关系
1
2013
... 了解树木对深层土壤水的吸收利用特征, 需首先对深层土壤进行界定, 但深层土壤并非严格的科学概念, 而且全球不同地区土壤厚度差异很大(如土石山区与黄土高原)(
密植枣林地深层土壤水分垂直变化与根系分布关系
1
2013
... 了解树木对深层土壤水的吸收利用特征, 需首先对深层土壤进行界定, 但深层土壤并非严格的科学概念, 而且全球不同地区土壤厚度差异很大(如土石山区与黄土高原)(
Soil water depletion patterns of artificial forest species and ages on the Loess Plateau (China)
4
2018a
... 同一树种对深层土壤水的吸收利用存在很强的时空变异性.具备深根系虽能保证树木同时利用不同土层的水分和(或)地下水, 但却存在明显的季节和年际变化: 湿润季节为高效利用降水会优先从浅土层吸收水分, 而干旱季节则会转向吸收更多的深层土壤水和(或)地下水(
... 同一地区不同树种对深层土壤水的吸收利用策略在时间和空间上常存在较大差异, 该现象在干旱(
... ;
... 模式I和II很难使人工林达到较高的生产力水平(
Water use by broadleaved tree species in response to changes in precipitation in a mountainous area of Beijing
1
2018b
... 潜在蒸散量和降水是影响树木深层根系吸水的重要气候因子.当潜在蒸散量增大时, 林木蒸腾量会相应变大, 其根系吸水范围会向更深的土层发展(
植物水分调节对策: 等水与非等水行为
1
2017
... 现有的少数研究结果显示, 树木在发展出较深的根系而能获取相对稳定的水源后, 为实现深层土壤水利用最大化, 会对叶片水力性状进行调节.例如, 会使树木的单位叶面积(“水分散失”面积)具有更大的根系表面积(“水分供给”面积), 从而使其在生长季维持相对稳定且更高的水分状况(较高的黎明前叶水势)和气孔导度, 以保持较大的蒸腾速率, 并使叶片相应具有较高的光合速率(
植物水分调节对策: 等水与非等水行为
1
2017
... 现有的少数研究结果显示, 树木在发展出较深的根系而能获取相对稳定的水源后, 为实现深层土壤水利用最大化, 会对叶片水力性状进行调节.例如, 会使树木的单位叶面积(“水分散失”面积)具有更大的根系表面积(“水分供给”面积), 从而使其在生长季维持相对稳定且更高的水分状况(较高的黎明前叶水势)和气孔导度, 以保持较大的蒸腾速率, 并使叶片相应具有较高的光合速率(
Effects of drip irrigation on the deep root distribution, rooting depth, and soil water profile of jujube in a semiarid region
1
2013
... 该方式是使根系在更大区域内吸收水分, 来提高深根系吸水量.扩大深层根系的分布深度, 不仅可增大其水养资源觅取范围, 而且也能减小根系个体间的资源竞争压力, 所以是树木提高深层水分吸收的最重要和最常见的一种方式.对于同一树种, 这种调节一般在较为干旱的地区比较明显(
Evolutionary history resolves global organization of root functional roots
4
2018
... 根系是树木吸收和传输土壤水分和养分资源的重要功能器官(
... 根系具有很强的可塑性, 对土壤环境非常敏感(
... 该方式是通过提高单位长度或面积根系的水分吸收效率(或能力), 来增加根系从深土层吸收的水量, 而且其与增加单位土体根系吸收面积和根系分布范围相比, 可减少根系构建的碳成本投入.由于可缩短水分从土壤进入根系木质部的径向传输距离, 因而减小细根直径可能增大其水分吸收速度和效率, 并同时扩大其水分吸收范围(根系组织密度变化不大的情况下, 比根长增大)(
... 然而, 需要注意的是, 真菌的侵染(形成菌根)可能会降低深根系性状可塑性的程度, 而且根系类型定义的不同(如以直径<0.5、1或2 mm等来定义细根)也可能使这种变化难以被发觉(
Seasonal patterns of fine root production and turnover in a mature rubber tree ( Hevea brasiliensisMüll. Arg.) stand—Differentiation with soil depth and implications for soil carbon stocks
1
2015
... 为保证深层根系能在旱季或表土层干燥时为地上部分器官及时供给充足水分, 树木会对深层根系的物候(存在明显季节性)进行调节, 以使其能适时、及时地激活和发挥水分供给功能: 在旱季树木对深层土壤水有较强需求时, 会快速启动深层根系的生长去觅取和吸收深层水源, 或进入休眠季地上部分器官不再生长时, 将更多的碳水化合物用于深根系生长, 以为次年吸收利用深层水做“材料准备” (
How to study deep roots and why it matters
4
2013
... 深层根系是植物借以吸收深层土壤水的唯一工具, 其在树木水分关系中的重要性早在近一个世纪以前就被人们所关注(
... ).
... 土壤是植物生长的介质, 故而深层土壤应是相对于植物而定义的概念,
... ), 同时也发现, 所有根系剖面的取样深度中值为0.88 m.基于该结果,
Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes
2
2015
... 为提高深根系水分传输能力, 树木会对其粗根以及细根(≤2 mm)中传输根(木质化、主要起传输和储藏作用的根系)(
... 深土层细根中的吸收根(非木质化、吸收水养资源的根系)(
Nutrient uptake as a contributing explanation for deep rooting in arid and semi-arid ecosystems
4
2004
... 深层根系是植物借以吸收深层土壤水的唯一工具, 其在树木水分关系中的重要性早在近一个世纪以前就被人们所关注(
... 土壤是植物生长的介质, 故而深层土壤应是相对于植物而定义的概念,
... 当土壤有机层较厚时, 土壤养分充足, 一般而言, 树木可能不需扎根很深便可获取充足的土壤养分.现有数据表明, 在寒温带针叶林、温带和热带森林中, d95均会随着土壤有机层厚度的减小而变深(
... 较大的土壤机械阻力(如土壤容重、土壤硬盘等造成)、较低的氧气浓度和水分含量会限制根系的向下生长; 在寒冷地区, 较低的土壤温度和永久冻土层也会成为根系垂向延伸的限制因素(
Aquaporin-mediated changes in hydraulic conductivity of deep tree roots accessed via caves
4
2007
... 该方式是通过在单位土体内生长更多的根系, 增大根系吸水面积, 以提高树木深土层中的水分吸收量.土壤水分充足时, 根系吸收面积与吸水量成正比, 因此通过在单位体积土壤内增加吸收根的根长密度、根表面积密度、根系体积等, 来扩大根系在土壤中的吸水面积, 是提高深土层水分吸收的重要方式(
... 根系吸水过程中, 水分从根系表皮穿过皮层, 然后跨过内皮层进入木质部导管过程中, 可以通过共质体、质外体和跨膜等3种途径进行运输, 其中均涉及水分的跨膜运输(主要是跨膜途径), 而细胞膜上的水孔蛋白在水分子的跨膜中起主要控制作用(
... 根系、茎干、枝条、叶片四个水力器官组成了植物的水分传输系统, 四者在保障整株水力路径中水分的传输上是功能统一的, 其中任一器官(尤其是叶片和根系)木质部水力导度下降或水力失调都会直接影响整株植物的水分传输效率, 从而影响叶片气体交换以及植物生存和生长(
... 树木通过深根系获取充足水分供给后, 其叶片与根系间的协调度可能也会提高, 从而保障叶片的水分需求信息能及时反馈和传递至根系, 然后使根系根据蒸腾需求通过生理调节策略来调控其水分吸收和供给量.例如,
Variation in xylem structure and function in stems and roots of trees to 20 m depth
3
2004
... 为提高深根系水分传输能力, 树木会对其粗根以及细根(≤2 mm)中传输根(木质化、主要起传输和储藏作用的根系)(
... 根系、茎干、枝条、叶片四个水力器官组成了植物的水分传输系统, 四者在保障整株水力路径中水分的传输上是功能统一的, 其中任一器官(尤其是叶片和根系)木质部水力导度下降或水力失调都会直接影响整株植物的水分传输效率, 从而影响叶片气体交换以及植物生存和生长(
... 各器官木质部的解剖结构及其相对变化特征, 决定了树木体内的水分传输效率(
Converging patterns of uptake and hydraulic redistribution of soil water in contrasting woody vegetation types
1
2004
... HL的大小受多种环境因素影响.根系能触及地下水或毛管边缘水时, 会促进HL的发生或提高其水分提升量, 尤其是在深层土壤质地较粗、持水力较弱的情况下, 如
基于稳定同位素和热扩散技术的张北杨树水分关系差异
1
2017
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
基于稳定同位素和热扩散技术的张北杨树水分关系差异
1
2017
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
Unusual fine root distributions of two deciduous tree species in southern France: What consequences for modeling of tree root dynamics?
3
2006
... 植物根系系统的大小(分布深度、水平延伸距离)与其地上部分的大小间存在显著正相关关系(
... 在林分等小尺度上, 物种竞争能明显影响根系的向下生长.如果种内(如密度增大、树木个体增大等造成)和种间(如农林间作、树种混交等造成)竞争增强, 一般会促进树木深根系的发生和发展.如,
... 虽然气候因子能对全球尺度上树木根系的分布深度产生明显影响, 但相同气候下却常出现较浅与较深根系分布的同时存在.这种现象可能由树种、林龄、土壤质地、种间竞争等因素引起(
Water uptake and hydraulic redistribution under a seasonal climate: Long-term study in a rainfed olive orchard
1
2014
... 潜在蒸散量和降水是影响树木深层根系吸水的重要气候因子.当潜在蒸散量增大时, 林木蒸腾量会相应变大, 其根系吸水范围会向更深的土层发展(
Phreatophytic vegetation and groundwater fluctuations: A review of current research and application of ecosystem response modeling with an emphasis on great basin vegetation
3
2005
... 地下水位之所以对根系最大分布深度起较强的控制作用, 主要是因为缺氧会限制根系的生长.但也有研究发现, 当地下水中有较多的溶解氧和养分时, 根系也会突破地下水的限制, 在地下水位以下生存(
... 从深层土壤中吸收水分运输至地上部分器官供给蒸腾和生长, 是树木对深层土壤水的最直接利用方式(
... 地下水位变化会影响树木的深层根系吸水特征, 但不同树种响应不同.对于根系分布较深和依赖地下水的树种, 其根系吸水深度一般会随地下水位的降低而逐渐下移(
The role of deep roots in the hydrological and carbon cycles of Amazonian forests and pastures
4
1994
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... ).此外, 了解树木对深层土壤水的吸收利用特征还可帮助深入认识树木对干旱和气候变化的响应与适应机制(
... )、树木与环境的互作机制(
... 根系是树木吸收和传输土壤水分和养分资源的重要功能器官(
The magnitude of hydraulic redistribution by plant roots: A review and synthesis of empirical and modeling studies
2
2012
... 现有研究证实, HL能对各类植物生态系统的水分循环产生重要影响.对于不同地区的不同生态系统, 通过HL作用可在干燥的浅土层中每天平均释放0.04-1.30 mm的水分, 从而使其水分含量平均提高28%-102%, 并供给2%-80%的蒸腾耗水(
... 根系结构特征决定根系功能发挥, 树木实现对深层土壤水的直接利用首先需要在深土层中生长根系.但若想进一步通过水力提升来利用深层土壤水, 则还需在浅土层中分布根系, 使整个根系系统覆盖具有较大水分梯度差异的土壤区域(
Deep root function in soil water dynamics in cerrado savannas of central Brazil
2
2005
... 土壤是植物生长的介质, 故而深层土壤应是相对于植物而定义的概念,
... 林分结构的差异和改变能引起树木根系吸水的可塑性变化.与纯林相比, 不同树种混交时, 会促使根系吸水模式可塑性较强的树种在旱季转向主要利用深层土壤水(
叶片水力性状研究进展
1
2014
... 根系、茎干、枝条、叶片四个水力器官组成了植物的水分传输系统, 四者在保障整株水力路径中水分的传输上是功能统一的, 其中任一器官(尤其是叶片和根系)木质部水力导度下降或水力失调都会直接影响整株植物的水分传输效率, 从而影响叶片气体交换以及植物生存和生长(
叶片水力性状研究进展
1
2014
... 根系、茎干、枝条、叶片四个水力器官组成了植物的水分传输系统, 四者在保障整株水力路径中水分的传输上是功能统一的, 其中任一器官(尤其是叶片和根系)木质部水力导度下降或水力失调都会直接影响整株植物的水分传输效率, 从而影响叶片气体交换以及植物生存和生长(
Hydraulic architecture and xylem structure of the dimorphic roots systems of South-West Australian species of Proteaceae
4
1995
... 根系系统水分供给量由其水分吸收和传输能力共同决定.为提高深层水吸收利用效率, 树木常提高深根系水分传输能力(轴向导水率), 将吸收进来的水分高效、快速地运至地上部分器官.如
... 为提高深根系水分传输能力, 树木会对其粗根以及细根(≤2 mm)中传输根(木质化、主要起传输和储藏作用的根系)(
... 根系、茎干、枝条、叶片四个水力器官组成了植物的水分传输系统, 四者在保障整株水力路径中水分的传输上是功能统一的, 其中任一器官(尤其是叶片和根系)木质部水力导度下降或水力失调都会直接影响整株植物的水分传输效率, 从而影响叶片气体交换以及植物生存和生长(
... 各器官木质部的解剖结构及其相对变化特征, 决定了树木体内的水分传输效率(
Hydrologic regulation of plant rooting depth: Breakthrough or observational conundrum?
1
2018
... 地下水位之所以对根系最大分布深度起较强的控制作用, 主要是因为缺氧会限制根系的生长.但也有研究发现, 当地下水中有较多的溶解氧和养分时, 根系也会突破地下水的限制, 在地下水位以下生存(
Understanding deep roots and their functions in ecosystems: An advocacy for more unconventional research
7
2016
... 深层根系是植物借以吸收深层土壤水的唯一工具, 其在树木水分关系中的重要性早在近一个世纪以前就被人们所关注(
... 和
... 土壤是植物生长的介质, 故而深层土壤应是相对于植物而定义的概念,
... ); 因此, 从这个角度来看, 将1 m以下土层中的根系定义为深层根系具备一定合理性.
... 对全球不同气候、土壤和排水条件下超过1 000个植物物种的2 200个根系研究数据进行了分析, 发现目前观测到的根系最大分布深度为1 m左右的情况最多, 该结果对上述深根系划分方法在全球尺度上、针对所有陆地植被类型的一般适用性提供了一定支撑.
... 根系是树木吸收和传输土壤水分和养分资源的重要功能器官(
... 该方式是通过在单位土体内生长更多的根系, 增大根系吸水面积, 以提高树木深土层中的水分吸收量.土壤水分充足时, 根系吸收面积与吸水量成正比, 因此通过在单位体积土壤内增加吸收根的根长密度、根表面积密度、根系体积等, 来扩大根系在土壤中的吸水面积, 是提高深土层水分吸收的重要方式(
A fast exploration of very deep soil layers by Eucalyptusseedlings and clones in Brazil
5
2016
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... )、林木遗传育种(
... )、林地水文模型的优化和构建(
... 该方式是通过提高单位长度或面积根系的水分吸收效率(或能力), 来增加根系从深土层吸收的水量, 而且其与增加单位土体根系吸收面积和根系分布范围相比, 可减少根系构建的碳成本投入.由于可缩短水分从土壤进入根系木质部的径向传输距离, 因而减小细根直径可能增大其水分吸收速度和效率, 并同时扩大其水分吸收范围(根系组织密度变化不大的情况下, 比根长增大)(
... 现有的少数研究结果显示, 树木在发展出较深的根系而能获取相对稳定的水源后, 为实现深层土壤水利用最大化, 会对叶片水力性状进行调节.例如, 会使树木的单位叶面积(“水分散失”面积)具有更大的根系表面积(“水分供给”面积), 从而使其在生长季维持相对稳定且更高的水分状况(较高的黎明前叶水势)和气孔导度, 以保持较大的蒸腾速率, 并使叶片相应具有较高的光合速率(
Root functional parameters along a land-use gradient: Evidence of a community-?level economics spectrum
3
2015
... 根系具有很强的可塑性, 对土壤环境非常敏感(
... ).深土层的环境条件(氧气浓度、养分含量、碳氮比、温度变异性一般较低, 土壤容重和强度一般较大)与浅土层存在明显差异, 尤其是其中的水分含量一般较高且相对稳定.因此, 深根系经历水分亏缺或胁迫的概率相对较小, 故而其功能会与浅层根系出现分异, 可能会更加注重水分供给(
... 该方式是通过在单位土体内生长更多的根系, 增大根系吸水面积, 以提高树木深土层中的水分吸收量.土壤水分充足时, 根系吸收面积与吸水量成正比, 因此通过在单位体积土壤内增加吸收根的根长密度、根表面积密度、根系体积等, 来扩大根系在土壤中的吸水面积, 是提高深土层水分吸收的重要方式(
Modeling the effect of changing precipitation inputs on deep soil water utilization
4
2018
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 土壤是植物生长的介质, 故而深层土壤应是相对于植物而定义的概念,
... 潜在蒸散量和降水是影响树木深层根系吸水的重要气候因子.当潜在蒸散量增大时, 林木蒸腾量会相应变大, 其根系吸水范围会向更深的土层发展(
... ).降水频率对深层根系吸水比例的调控作用较弱, 但随降水频率降低(单次降水强度相应增大), 深层根系和深层土壤的树木干旱缓解能力会变弱(
Direct observations of rock moisture, a hidden component of the hydrologic cycle
3
2018
... 综上所述, 就所有陆地植被类型平均而言, 可将1 m以下土层中的根系定义为深根系, 而该土层即为深土层.虽然其在特定气候、土壤和植被类型下的适用性有限, 但基于d95数据(
... 土壤厚度是树木以及其他植物最大根系分布深度的重要影响因子, 但很多情况下其并不能制约根系的最终分布深度(
... ).例如,
Hydraulic lift: Substantial nocturnal water transport between soil layers by Artem isiatridentate roots
1
1987
... 水力提升(HL)是树木实现深层土壤水吸收利用的另一种方式, 同时也是除深根系和气孔调节之外的另一重要抗旱机制.该现象是指在浅土层与深土层存在水势梯度的情况下, 植物深层根系借助水势差在夜间将深土层中的水分运输至表层根系, 然后再释放至浅土层中以供白天蒸腾所需(
Hydraulic redistribution in a stand of Artemisia tridentate: Evaluation of benefits to transpiration assessed with a simulation model
2
2002
... HL的大小受多种环境因素影响.根系能触及地下水或毛管边缘水时, 会促进HL的发生或提高其水分提升量, 尤其是在深层土壤质地较粗、持水力较弱的情况下, 如
... ).其原因可能是当土壤逐渐干旱时, 砂性更强的立地上的土壤、根系以及根-土界面导水率会下降得更快或更难维持(
The shallowest possible water extraction profile: A null model for global root distribution
6
2008
... 较大的土壤机械阻力(如土壤容重、土壤硬盘等造成)、较低的氧气浓度和水分含量会限制根系的向下生长; 在寒冷地区, 较低的土壤温度和永久冻土层也会成为根系垂向延伸的限制因素(
... ).但当气候干旱、物种土壤资源竞争等造成浅土层水分资源的相对亏缺, 林龄增大、大气蒸发潜力增加等造成蒸腾作用增强, 树木依靠浅层土壤中的水分吸收已不能满足自身蒸腾需求时, 其根系便会尽可能地向深土层生长来吸收更多的深层水分(
... 潜在蒸散量和降水是影响树木深层根系吸水的重要气候因子.当潜在蒸散量增大时, 林木蒸腾量会相应变大, 其根系吸水范围会向更深的土层发展(
... 树木对深层土壤水的吸收利用, 从本质上讲, 主要是受到土壤水分垂直分布和植株蒸腾耗水的共同驱动, 因为二者分别决定了土壤中的水分供给能力和树木对水分的需求大小(
... 各器官木质部的解剖结构及其相对变化特征, 决定了树木体内的水分传输效率(
... ).当生长环境发生变化, 为维持生长和生存, 树木可能会改变其木质部结构以利于充足的水分供给, 但同时会尽可能降低其水力失败风险, 即在木质部水分传输效率与水力安全之间维持一种权衡(
The global biogeography of roots
10
2002a
... 土壤是植物生长的介质, 故而深层土壤应是相对于植物而定义的概念,
... 全球不同植被类型累积95%根系的分布深度(d95) (
... The estimated depths of 95% cumulative roots (d95) for global vegetation types (
... 综上所述, 就所有陆地植被类型平均而言, 可将1 m以下土层中的根系定义为深根系, 而该土层即为深土层.虽然其在特定气候、土壤和植被类型下的适用性有限, 但基于d95数据(
... 现有研究表明, 影响树木深根系产生的因子很多, 但归纳起来可分为植被因子(树种、生活型、植被类型、林龄、个体发育、物种竞争)、气候因子(降水量、潜在蒸散量)、土壤因子(土层厚度、土壤质地、土壤有机层厚度、土壤水分、土壤养分)和水文因子(地下水位), 且不同空间尺度上主要影响因子不同(
... 现有研究显示, 在全球大尺度上, 年降水量(MAP)和年潜在蒸散量(PET)可能是影响树木根系分布的两个重要气候因子(
... 变小)?(
... 全球尺度上, 土壤是仅次于气候的树木根系分布深度的另一重要影响因子(
... 当土壤有机层较厚时, 土壤养分充足, 一般而言, 树木可能不需扎根很深便可获取充足的土壤养分.现有数据表明, 在寒温带针叶林、温带和热带森林中, d95均会随着土壤有机层厚度的减小而变深(
... 树木根系主要分布于浅土层, 深层土壤中根系一般较少(
Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems
3
2002b
... 根系是树木吸收和传输土壤水分和养分资源的重要功能器官(
... 现有研究表明, 影响树木深根系产生的因子很多, 但归纳起来可分为植被因子(树种、生活型、植被类型、林龄、个体发育、物种竞争)、气候因子(降水量、潜在蒸散量)、土壤因子(土层厚度、土壤质地、土壤有机层厚度、土壤水分、土壤养分)和水文因子(地下水位), 且不同空间尺度上主要影响因子不同(
... 现有研究显示, 在全球大尺度上, 年降水量(MAP)和年潜在蒸散量(PET)可能是影响树木根系分布的两个重要气候因子(
Mapping the global distribution of deep roots in relation to climate and soil characteristics
9
2005
... 土壤是植物生长的介质, 故而深层土壤应是相对于植物而定义的概念,
... ).
... 综上所述, 就所有陆地植被类型平均而言, 可将1 m以下土层中的根系定义为深根系, 而该土层即为深土层.虽然其在特定气候、土壤和植被类型下的适用性有限, 但基于d95数据(
... 根系是树木吸收和传输土壤水分和养分资源的重要功能器官(
... 现有研究表明, 影响树木深根系产生的因子很多, 但归纳起来可分为植被因子(树种、生活型、植被类型、林龄、个体发育、物种竞争)、气候因子(降水量、潜在蒸散量)、土壤因子(土层厚度、土壤质地、土壤有机层厚度、土壤水分、土壤养分)和水文因子(地下水位), 且不同空间尺度上主要影响因子不同(
... 现有研究显示, 在全球大尺度上, 年降水量(MAP)和年潜在蒸散量(PET)可能是影响树木根系分布的两个重要气候因子(
... 基于
... 一般而言, 土壤质地因影响水分入渗深度和毛管水上升高度而能对树木及其他植物根系分布深度产生影响(
... 浅土层干旱虽不会决定深根系的发生, 但会促进深根系的发展.如
Emanation of water from underground plant parts
1
1967
... HL的大小受多种环境因素影响.根系能触及地下水或毛管边缘水时, 会促进HL的发生或提高其水分提升量, 尤其是在深层土壤质地较粗、持水力较弱的情况下, 如
Applying hydraulic lift in an agroecosystem: Forage plants with shoots removed supply water to neighboring vegetable crops
1
2011
...
Mapping the global depth to bedrock for land surface modeling
1
2017
... 了解树木对深层土壤水的吸收利用特征, 需首先对深层土壤进行界定, 但深层土壤并非严格的科学概念, 而且全球不同地区土壤厚度差异很大(如土石山区与黄土高原)(
Water sources of desert riparian Populus euphraticaforests
2
2014
... 地下水位变化会影响树木的深层根系吸水特征, 但不同树种响应不同.对于根系分布较深和依赖地下水的树种, 其根系吸水深度一般会随地下水位的降低而逐渐下移(
... 同一树种对深层土壤水的吸收利用存在很强的时空变异性.具备深根系虽能保证树木同时利用不同土层的水分和(或)地下水, 但却存在明显的季节和年际变化: 湿润季节为高效利用降水会优先从浅土层吸收水分, 而干旱季节则会转向吸收更多的深层土壤水和(或)地下水(
Onset of water stress, hysteresis in plant conductance, and hydraulic lift: Scaling soil water dynamics from millimeters to meters
1
2008
... HL的大小受多种环境因素影响.根系能触及地下水或毛管边缘水时, 会促进HL的发生或提高其水分提升量, 尤其是在深层土壤质地较粗、持水力较弱的情况下, 如
Sources of water used by Pinus sylvestrisvar
3
2016
... 潜在蒸散量和降水是影响树木深层根系吸水的重要气候因子.当潜在蒸散量增大时, 林木蒸腾量会相应变大, 其根系吸水范围会向更深的土层发展(
... 一般而言, 浅土层含水量较少而出现土壤干旱时, 会迫使树木转向吸收更多的深层土壤水并降低浅土层的快速干旱程度(
... 同一树种对深层土壤水的吸收利用存在很强的时空变异性.具备深根系虽能保证树木同时利用不同土层的水分和(或)地下水, 但却存在明显的季节和年际变化: 湿润季节为高效利用降水会优先从浅土层吸收水分, 而干旱季节则会转向吸收更多的深层土壤水和(或)地下水(
Tree Root Systems in Eastern Nebraska
1
1979
... 虽然气候因子能对全球尺度上树木根系的分布深度产生明显影响, 但相同气候下却常出现较浅与较深根系分布的同时存在.这种现象可能由树种、林龄、土壤质地、种间竞争等因素引起(
Depth of soil water uptake by tropical rainforest trees during dry periods: Does tree dimension matter?
2
2013
... 树木根系吸水特征会因树体大小、健康状况以及根区位置的不同而发生改变.其中, 关于树体大小(个体发育)与根系吸水深度间的关系目前存在不一致的结果: 多数研究发现树体越大, 深层土壤水吸收比例会越大(
... 同一地区不同树种对深层土壤水的吸收利用策略在时间和空间上常存在较大差异, 该现象在干旱(
The potential for deep groundwater use by Acacia papyrocarpa( Western myall) in a water-limited environment
1
2016
... 地下水位变化会影响树木的深层根系吸水特征, 但不同树种响应不同.对于根系分布较深和依赖地下水的树种, 其根系吸水深度一般会随地下水位的降低而逐渐下移(
On the maximum extent of tree roots
3
1991
... 深层根系是植物借以吸收深层土壤水的唯一工具, 其在树木水分关系中的重要性早在近一个世纪以前就被人们所关注(
... 较大的土壤机械阻力(如土壤容重、土壤硬盘等造成)、较低的氧气浓度和水分含量会限制根系的向下生长; 在寒冷地区, 较低的土壤温度和永久冻土层也会成为根系垂向延伸的限制因素(
... 从深层土壤中吸收水分运输至地上部分器官供给蒸腾和生长, 是树木对深层土壤水的最直接利用方式(
Temporal and spatial partitioning of water resources among eight woody species in a Hawaiian dry forest
1
2000
... 同一地区不同树种对深层土壤水的吸收利用策略在时间和空间上常存在较大差异, 该现象在干旱(
水分再分配对土壤-植物系统养分循环的生态意义
1
2014
... 现有研究证实, HL能对各类植物生态系统的水分循环产生重要影响.对于不同地区的不同生态系统, 通过HL作用可在干燥的浅土层中每天平均释放0.04-1.30 mm的水分, 从而使其水分含量平均提高28%-102%, 并供给2%-80%的蒸腾耗水(
水分再分配对土壤-植物系统养分循环的生态意义
1
2014
... 现有研究证实, HL能对各类植物生态系统的水分循环产生重要影响.对于不同地区的不同生态系统, 通过HL作用可在干燥的浅土层中每天平均释放0.04-1.30 mm的水分, 从而使其水分含量平均提高28%-102%, 并供给2%-80%的蒸腾耗水(
Stable isotope analysis reveals prolonged drought stress in poplar plantation mortality of the three-north shelter forest in Northern China
6
2018
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 树木根系吸水特征会因树体大小、健康状况以及根区位置的不同而发生改变.其中, 关于树体大小(个体发育)与根系吸水深度间的关系目前存在不一致的结果: 多数研究发现树体越大, 深层土壤水吸收比例会越大(
... 同一树种对深层土壤水的吸收利用存在很强的时空变异性.具备深根系虽能保证树木同时利用不同土层的水分和(或)地下水, 但却存在明显的季节和年际变化: 湿润季节为高效利用降水会优先从浅土层吸收水分, 而干旱季节则会转向吸收更多的深层土壤水和(或)地下水(
... 模式I和II很难使人工林达到较高的生产力水平(
... 我国有许多低质低效人工林, 其中比较典型的是三北地区退化的杨树防护林.基于稳定同位素技术,
... 均研究发现, 张北地区的杨树防护林退化越严重, 其主要水分来源土层就越浅.那么是林分退化导致深土层水分利用减少, 还是深土层水分不足造成林木深层根系吸水变少, 从而加剧了林分退化, 目前尚不能完全确定.但可以肯定的是, 地下水位下降与张北地区杨树林的退化有相关关系(
Origin and Development of White Spruce Root-Forms. Queen’s Printer and Controller of Stationery, Ottawa, Ontario, Canada
1
1967
... 虽然气候因子能对全球尺度上树木根系的分布深度产生明显影响, 但相同气候下却常出现较浅与较深根系分布的同时存在.这种现象可能由树种、林龄、土壤质地、种间竞争等因素引起(
The role of hydraulic lift and subsoil P placement in P uptake of cotton ( Gossypium hirsutum L.)
1
2009
... HL的大小受多种环境因素影响.根系能触及地下水或毛管边缘水时, 会促进HL的发生或提高其水分提升量, 尤其是在深层土壤质地较粗、持水力较弱的情况下, 如
Root tip morphology, anatomy, chemistry and potential hydraulic conductivity vary with soil depth in three temperate hardwood species
3
2015
... 根系具有很强的可塑性, 对土壤环境非常敏感(
... 该方式是通过在单位土体内生长更多的根系, 增大根系吸水面积, 以提高树木深土层中的水分吸收量.土壤水分充足时, 根系吸收面积与吸水量成正比, 因此通过在单位体积土壤内增加吸收根的根长密度、根表面积密度、根系体积等, 来扩大根系在土壤中的吸水面积, 是提高深土层水分吸收的重要方式(
... 深土层细根中的吸收根(非木质化、吸收水养资源的根系)(
根系生态学
1
2008
... 根系是树木吸收和传输土壤水分和养分资源的重要功能器官(
根系生态学
1
2008
... 根系是树木吸收和传输土壤水分和养分资源的重要功能器官(
Hydraulic safety margins and air-seeding thresholds in roots, trunks, branches and petioles of four northern hardwood trees
3
2018
... 根系、茎干、枝条、叶片四个水力器官组成了植物的水分传输系统, 四者在保障整株水力路径中水分的传输上是功能统一的, 其中任一器官(尤其是叶片和根系)木质部水力导度下降或水力失调都会直接影响整株植物的水分传输效率, 从而影响叶片气体交换以及植物生存和生长(
... ;
... 各器官木质部的解剖结构及其相对变化特征, 决定了树木体内的水分传输效率(
Fine-root trait plasticity of beech ( Fagus sylvatica) and spruce( Picea abies) forests on two contrasting soils
2
2017
... 该方式是通过在单位土体内生长更多的根系, 增大根系吸水面积, 以提高树木深土层中的水分吸收量.土壤水分充足时, 根系吸收面积与吸水量成正比, 因此通过在单位体积土壤内增加吸收根的根长密度、根表面积密度、根系体积等, 来扩大根系在土壤中的吸水面积, 是提高深土层水分吸收的重要方式(
... 然而, 需要注意的是, 真菌的侵染(形成菌根)可能会降低深根系性状可塑性的程度, 而且根系类型定义的不同(如以直径<0.5、1或2 mm等来定义细根)也可能使这种变化难以被发觉(
修剪对黄土丘陵区枣树蒸腾的调控作用
1
2014
... 如前所述, 树冠大小与根系分布深度间存在正相关关系, 蒸腾需求增大会促使林木吸收深土层水分, 这意味着对树冠尺寸进行控制, 可通过降低蒸腾来减缓深土层水分资源消耗.例如,
修剪对黄土丘陵区枣树蒸腾的调控作用
1
2014
... 如前所述, 树冠大小与根系分布深度间存在正相关关系, 蒸腾需求增大会促使林木吸收深土层水分, 这意味着对树冠尺寸进行控制, 可通过降低蒸腾来减缓深土层水分资源消耗.例如,
基于林冠塔吊的附生植物生态学研究进展
1
2016
... 由于研究难度大且在森林生理生态过程中扮演重要角色, 上层林冠被喻为“地球的第八大洲” (
基于林冠塔吊的附生植物生态学研究进展
1
2016
... 由于研究难度大且在森林生理生态过程中扮演重要角色, 上层林冠被喻为“地球的第八大洲” (
Modeling growth response to soil water availability simulated by HYDRUS for a mature triploid Populus tomentosaplantation located on the North China Plain
1
2016
... 我国木材资源对外依存度超过50%, 木材安全问题严重, 营造和培育速生丰产林缓解木材短缺势在必行.目前, 我国人工林面积已达6 933万hm2, 位居世界首位, 但人工林的质量和产量还较低, 远不能满足我国日益增长的木材需求, 亟须采取高效的定向集约培育措施来促进人工林生长.水分是树木生长的重要限制因子(
Hydrologic regulation of plant rooting depth: Pay attention to the widespread scenario with intense seasonal groundwater table fluctuation
1
2018
... 地下水位之所以对根系最大分布深度起较强的控制作用, 主要是因为缺氧会限制根系的生长.但也有研究发现, 当地下水中有较多的溶解氧和养分时, 根系也会突破地下水的限制, 在地下水位以下生存(
Modeling stand water use response to soil water availability and groundwater level for a mature Populus tomentosa plantation located on the North China Plain
1
2017
... 模式I和II很难使人工林达到较高的生产力水平(
The effect of subsurface irrigation at different soil water potential thresholds on the growth and transpiration of Populus tomentosain the North China Plain
2
2014
... 模式I和II很难使人工林达到较高的生产力水平(
... ).模式III虽然能明显提高林木生长(
Characteristics of fine root system and water uptake in a triploid Populus tomentosa plantation in the North China Plain: Implications for irrigation water management
4
2013
... 在林分等小尺度上, 物种竞争能明显影响根系的向下生长.如果种内(如密度增大、树木个体增大等造成)和种间(如农林间作、树种混交等造成)竞争增强, 一般会促进树木深根系的发生和发展.如,
... 一般而言, 浅土层含水量较少而出现土壤干旱时, 会迫使树木转向吸收更多的深层土壤水并降低浅土层的快速干旱程度(
... 树木根系吸水特征会因树体大小、健康状况以及根区位置的不同而发生改变.其中, 关于树体大小(个体发育)与根系吸水深度间的关系目前存在不一致的结果: 多数研究发现树体越大, 深层土壤水吸收比例会越大(
... 同一树种对深层土壤水的吸收利用存在很强的时空变异性.具备深根系虽能保证树木同时利用不同土层的水分和(或)地下水, 但却存在明显的季节和年际变化: 湿润季节为高效利用降水会优先从浅土层吸收水分, 而干旱季节则会转向吸收更多的深层土壤水和(或)地下水(
Rooting depth and leaf hydraulic conductance in the xeric tree Haloxyolon ammodendron growing at sites of contrasting soil texture
6
2008
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 一般而言, 土壤质地因影响水分入渗深度和毛管水上升高度而能对树木及其他植物根系分布深度产生影响(
... 虽然气候因子能对全球尺度上树木根系的分布深度产生明显影响, 但相同气候下却常出现较浅与较深根系分布的同时存在.这种现象可能由树种、林龄、土壤质地、种间竞争等因素引起(
... 现有的少数研究结果显示, 树木在发展出较深的根系而能获取相对稳定的水源后, 为实现深层土壤水利用最大化, 会对叶片水力性状进行调节.例如, 会使树木的单位叶面积(“水分散失”面积)具有更大的根系表面积(“水分供给”面积), 从而使其在生长季维持相对稳定且更高的水分状况(较高的黎明前叶水势)和气孔导度, 以保持较大的蒸腾速率, 并使叶片相应具有较高的光合速率(
... ).由于深根系树木水分来源相对充足, 其叶片可能会暂时采取非等水调节策略(anisohydry), 降低气孔对叶片水分散失和传导的调控作用, 以使叶片能持续维持较高的气体交换, 但这也可能导致其叶水势在一天内出现更大变幅(
... 树木通过深根系获取充足水分供给后, 其叶片与根系间的协调度可能也会提高, 从而保障叶片的水分需求信息能及时反馈和传递至根系, 然后使根系根据蒸腾需求通过生理调节策略来调控其水分吸收和供给量.例如,
Deep soil water extraction helps to drought avoidance but shallow soil water uptake during dry season controls the inter-?annual variation in tree growth in four subtropical plantations
2
2017
... 土壤是植物生长的介质, 故而深层土壤应是相对于植物而定义的概念,
... 同一地区不同树种对深层土壤水的吸收利用策略在时间和空间上常存在较大差异, 该现象在干旱(
Hydraulic lift among native plant species in the Mojave Desert
1
1999
... HL的大小受多种环境因素影响.根系能触及地下水或毛管边缘水时, 会促进HL的发生或提高其水分提升量, 尤其是在深层土壤质地较粗、持水力较弱的情况下, 如
Depressed hydraulic redistribution of roots more by stem refilling than by nocturnal transpiration for Populus euphraticaOliv. in situ measurement
1
2018
... HL的大小受多种环境因素影响.根系能触及地下水或毛管边缘水时, 会促进HL的发生或提高其水分提升量, 尤其是在深层土壤质地较粗、持水力较弱的情况下, 如
胡杨根系水力再分配的模式、大小及其影响因子
1
2014
... 国外对HL的研究较多, 已从简单的现象描述发展到定量评价、系统机理解释乃至与全球气候等结合探讨HL对气候变化以及森林动态的影响.国内对HL的研究起步较晚, 20世纪90年代中期开始至今, 相关的中文研究性论文仅有20多篇; 这些研究主要集中在西北干旱、半干旱地区和华北低丘山区的少数树种上, 如胡杨、多枝柽柳、胡桃等(
胡杨根系水力再分配的模式、大小及其影响因子
1
2014
... 国外对HL的研究较多, 已从简单的现象描述发展到定量评价、系统机理解释乃至与全球气候等结合探讨HL对气候变化以及森林动态的影响.国内对HL的研究起步较晚, 20世纪90年代中期开始至今, 相关的中文研究性论文仅有20多篇; 这些研究主要集中在西北干旱、半干旱地区和华北低丘山区的少数树种上, 如胡杨、多枝柽柳、胡桃等(
黑河下游柽柳根系水力提升对林分蒸散的贡献
1
2017
... 国外对HL的研究较多, 已从简单的现象描述发展到定量评价、系统机理解释乃至与全球气候等结合探讨HL对气候变化以及森林动态的影响.国内对HL的研究起步较晚, 20世纪90年代中期开始至今, 相关的中文研究性论文仅有20多篇; 这些研究主要集中在西北干旱、半干旱地区和华北低丘山区的少数树种上, 如胡杨、多枝柽柳、胡桃等(
黑河下游柽柳根系水力提升对林分蒸散的贡献
1
2017
... 国外对HL的研究较多, 已从简单的现象描述发展到定量评价、系统机理解释乃至与全球气候等结合探讨HL对气候变化以及森林动态的影响.国内对HL的研究起步较晚, 20世纪90年代中期开始至今, 相关的中文研究性论文仅有20多篇; 这些研究主要集中在西北干旱、半干旱地区和华北低丘山区的少数树种上, 如胡杨、多枝柽柳、胡桃等(
沙丘多枝柽柳灌丛根层土壤含水量变化特征与根系水力提升证据
2
2012
... 了解树木对深层土壤水的吸收利用特征, 需首先对深层土壤进行界定, 但深层土壤并非严格的科学概念, 而且全球不同地区土壤厚度差异很大(如土石山区与黄土高原)(
... 国外对HL的研究较多, 已从简单的现象描述发展到定量评价、系统机理解释乃至与全球气候等结合探讨HL对气候变化以及森林动态的影响.国内对HL的研究起步较晚, 20世纪90年代中期开始至今, 相关的中文研究性论文仅有20多篇; 这些研究主要集中在西北干旱、半干旱地区和华北低丘山区的少数树种上, 如胡杨、多枝柽柳、胡桃等(
沙丘多枝柽柳灌丛根层土壤含水量变化特征与根系水力提升证据
2
2012
... 了解树木对深层土壤水的吸收利用特征, 需首先对深层土壤进行界定, 但深层土壤并非严格的科学概念, 而且全球不同地区土壤厚度差异很大(如土石山区与黄土高原)(
... 国外对HL的研究较多, 已从简单的现象描述发展到定量评价、系统机理解释乃至与全球气候等结合探讨HL对气候变化以及森林动态的影响.国内对HL的研究起步较晚, 20世纪90年代中期开始至今, 相关的中文研究性论文仅有20多篇; 这些研究主要集中在西北干旱、半干旱地区和华北低丘山区的少数树种上, 如胡杨、多枝柽柳、胡桃等(
张北地区退化杨树防护林的水分利用特征
3
2018
... 树木根系吸水特征会因树体大小、健康状况以及根区位置的不同而发生改变.其中, 关于树体大小(个体发育)与根系吸水深度间的关系目前存在不一致的结果: 多数研究发现树体越大, 深层土壤水吸收比例会越大(
... )林分主要利用320-400 cm土层中的水分, 而轻度、中度和重度退化林分的主要水源深度则分别在120-320、20-80和0-20 cm (
... 我国有许多低质低效人工林, 其中比较典型的是三北地区退化的杨树防护林.基于稳定同位素技术,
张北地区退化杨树防护林的水分利用特征
3
2018
... 树木根系吸水特征会因树体大小、健康状况以及根区位置的不同而发生改变.其中, 关于树体大小(个体发育)与根系吸水深度间的关系目前存在不一致的结果: 多数研究发现树体越大, 深层土壤水吸收比例会越大(
... )林分主要利用320-400 cm土层中的水分, 而轻度、中度和重度退化林分的主要水源深度则分别在120-320、20-80和0-20 cm (
... 我国有许多低质低效人工林, 其中比较典型的是三北地区退化的杨树防护林.基于稳定同位素技术,
Effects of soil water and nitrogen on the stand volume of four hybrid Populus tomentosa clones
2
2017
... 模式I和II很难使人工林达到较高的生产力水平(
... ;
Reliance on deep soil water in the tree species Argania spinose
2
2017
... 根系吸水在树木生理活动中发挥着至关重要的作用, 是树木水分来源的主要途径.因此, 构建基于树木水分关系的人工林高效集约栽培措施, 需充分摸清树木对土壤水尤其是深层土壤水的吸收利用特征, 因为大多情况下深层土壤中的水资源含量相对较高, 作为水分“储备库”与“缓冲库”, 常可为树木生长供给大量水分, 且在旱季保障树木的生存与正常生长(
... 土壤是植物生长的介质, 故而深层土壤应是相对于植物而定义的概念,

{kind=link}
{kind=link}