 ,*, 任明迅,*海南大学热带农林学院 / 环南海陆域生物多样性研究中心, 海口 570228
,*, 任明迅,*海南大学热带农林学院 / 环南海陆域生物多样性研究中心, 海口 570228Difference in spatial distribution patterns and population structures of Rhododendron hainanense between both sides of riparian bends
CHEN Yi-Chao, ZHAO Ying, SONG Xi-Qiang,*, REN Ming-Xun,*Research Center for Terrestrial Biodiversity of the South China Sea, Institute of Tropical Agriculture and Forestry, Hainan University, Haikou 570228, China通讯作者:
编委: 曾波
责任编辑: 李敏
收稿日期:2018-02-20网络出版日期:2018-08-20
| 基金资助: |
Received:2018-02-20Online:2018-08-20
| Supported by: | SupportedbytheHainanProvincialInnovativeResearchTeamProgram( |

摘要
关键词:
Abstract
Methods Three nature reserves on Hainan Island were chosen as studied sites. In each site, two riparian bends were set as transects. The spatial distribution patterns and population dynamics of R. hainanense were compared for the convex versus concave banks by a method of “variance/mean method”.
Important findings (1) Rhododendron hainanense populations were at increasing stage on convex banks, but at declining stage on concave banks. (2) Rhododendron hainanense populations were at growing stage on both upstream and downstream of convex banks, but the ages of the individuals on concave banks were not continuous. (3) The number of individuals of each age-class declined away from the top of bend or river bank. (4) At scale of 2 m × 2 m, the individuals showed an aggregation distribution on convex banks and their downstream, but a random distribution on the upstream of convex banks, and on concave banks and their upstream and downstream. The fast flow and steep slopes may be the main reason for the fewer individuals and declining population on concave banks than on convex banks.
Keywords:
PDF (871KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
陈怡超, 赵莹, 宋希强, 任明迅. 海南杜鹃在河岸带弯道两侧的空间分布格局和年龄结构差异. 植物生态学报, 2018, 42(8): 841-849 doi:10.17521/cjpe.2018.0043
CHEN Yi-Chao, ZHAO Ying, SONG Xi-Qiang, REN Ming-Xun.
河岸带作为河流生态系统和陆地生态系统的过渡带, 拥有极高的空间异质性和复杂的生态过程, 孕育了丰富的物种多样性(Burt et al., 2010)。河岸带的群落结构与生态过程有别于河流与陆地生态系统(Naiman & Decamps, 1997), 特别是河道的地貌变化直接影响着河岸带的外貌结构与生态过程(Millar, 2000), 决定了河岸带生物多样性的形成与维持(Corenblit et al., 2007; Curran & Hession, 2013)。
河岸带的河道地貌变化最典型的表现之一, 就是形成河流弯道。天然河流的弯道一般占河流总长的80%以上。河水流经弯道, 受离心力作用, 表层水流偏向凹岸(河道外侧), 底部水流由凹岸流向凸岸(河道内侧), 造成河流凹岸水流快、易侵蚀; 凸岸水流慢、易堆积泥沙等显著差异。这些差异, 有可能导致凹岸和凸岸两岸的水土性质、植物繁殖体来源与聚集程度不同, 从而导致两岸的植被与植物生长出现差异(凃洋, 2017)。目前, 关于河岸带植被的研究集中于物种组成、水动力、地貌动力和地下水之间的相互作用及其建模(Solari et al., 2016), 需要进一步在更细微的局域尺度研究弯道凹岸和凸岸植物生长与种群动态的可能差异, 揭示河岸带生态过程与植物多样性形成及维持机制。
海南杜鹃(Rhododendron hainanense)是中国特有植物, 株高1-3 m, 花顶生、红色, 花期10月至翌年3月, 是杜鹃花属花期最晚、自然花期最长的一个物种(史佑海等, 2010), 极具园艺与园林开发应用价值。海南杜鹃仅分布在海南和广西海拔200-700 m的山区溪流沿岸(Fang et al., 2005), 但在溪流的平直段分布极少, 密集分布在溪流弯道, 并且在弯道内外两侧的分布具明显差异(陈怡超, 个人观察)。本研究针对海南杜鹃种群在河岸带弯道内外两侧的空间分布格局和种群年龄结构开展研究, 以揭示海南杜鹃在河岸带弯道凹岸和凸岸的分布特征与种群动态, 促进理解河岸带生态过程及生物多样性形成与维持机制。
1 研究区概况和研究方法
1.1 研究区概况
研究区位于海南省黎母山自然保护区(109.65°- 109.82° E, 19.12°-19.23° N)、吊罗山国家森林公园(109.72°-110.05° E, 18.72°-18.97° N)、尖峰岭国家森林公园(108.68°-109.20° E, 18.33°-18.95° N)。这3个地区都是我国原始热带雨林保护区, 生物多样性较高, 森林覆盖率达90%以上, 属于热带季风气候, 雨季和旱季分明, 夏秋多雨兼有台风。每个保护区内都有多条蜿蜒曲折的溪流, 溪流河岸带大多分布有海南杜鹃种群。1.2 研究方法
1.2.1 调查路线与数据采集在前期踏查的基础上, 于2017年8月在3个地区分别设置具有代表性, 未受人类破坏的溪流弯道样带, 调查溪流弯道的理化性状, 记录样带经纬度、海拔、河宽、溪流弯曲系数、弯顶位置等信息。以顺溪流方向为样带X轴, 距溪流河岸线的垂直距离为Y轴(图1)。样带长度为有海南杜鹃分布的弯道长度, 样带弯曲系数为样带实际长度与样带两端直线长度的比值(Crosato, 2008), 将溪流弯顶位置记为样带上的X值, 河宽为弯顶到对岸的距离。样带基本特征见表1。
图1
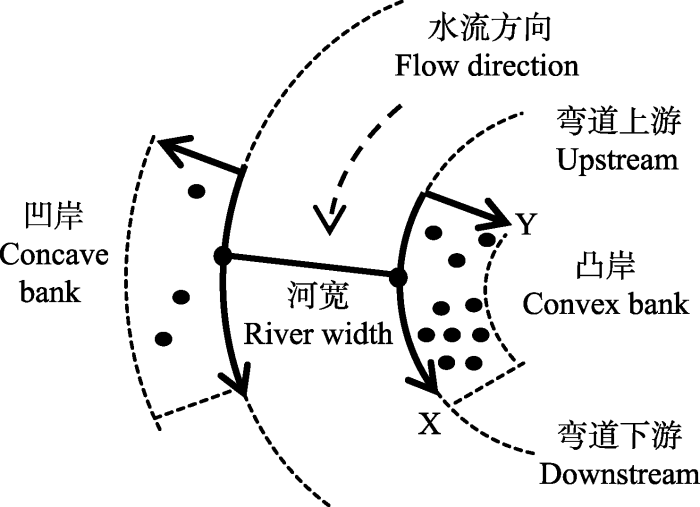 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1溪流弯道样带设置方法。
Fig. 1Transect setting method on stream bends.
Table 1
表1
表1河岸带弯道研究样带概况
Table 1
| 研究地点 Study site | 样带 Transect | 海拔 Altitude (m) | 样带长度 Transect length (m) | 弯曲系数 Transect sinuosity | 弯顶 Top of bend | 河宽 Stream width (m) | 郁闭度 Canopy density (%) |
|---|---|---|---|---|---|---|---|
| 黎母山 Mt. Limu | L1 | 577 | 50 | 1.04 | 20 | 6.5 | 90 |
| L2 | 573 | 16 | 1.07 | 8 | 10.5 | 85 | |
| 吊罗山 Mt. Diaoluo | D1 | 258 | 24 | 1.09 | 12 | 6.5 | 78 |
| D2 | 595 | 10 | 1.20 | 5 | 3.8 | 85 | |
| 尖峰岭 Mt. Jianfeng | J1 | 690 | 20 | 1.43 | 10 | 8 | 75 |
| J2 | 665 | 40 | 1.21 | 15 | 23 | 75 |
新窗口打开|下载CSV
对样带内所有海南杜鹃进行调查统计, 调查并记录样带内所有海南杜鹃的基径、冠幅、株高等指标, 并以平行河流方向为横坐标(x)、垂直于河流平行方向为纵坐标(y), 测定每株海南杜鹃的坐标(x, y)。如果一株海南杜鹃上有多个无性系分株, 则以其中基径最大的一株进行记录(康华靖等, 2007)。
1.2.2 空间分布格局及聚集强度
本研究采用可变尺度相邻格子法, 采用方差/平均值法在1 m× 1 m、2 m× 2 m、5 m × 5 m 3个尺度下(张文辉等, 2005; 赵正帅等, 2016), 对所有样带不同区域的海南杜鹃种群进行空间分布格局统计分析, 采用扩散系数(C, C值用t检验)、Cassie指数、平均拥挤度m*、丛生指数(I)、聚块性指数(m*/m)和负二项指数(K) 6个系数对海南杜鹃的空间分布格局类型和聚集强度进行判定和比较(何恒斌等, 2006; 康华靖等, 2007; 赵正帅等, 2016)。
1.2.3 种群结构划分
海南杜鹃属于常绿灌木, 分枝多, 有基部主干, 采用基径作为个体大小的指标研究种群结构(达良俊等, 2004; 张璞进等, 2017)。基径为海南杜鹃主干离地面10 cm处植株的直径(甄江红和刘果厚, 2008)。径级划分方法(李辛雷等, 2013)如下: I级(幼苗): 基径< 10 mm; II级(幼树): 基径10-20 mm; III级(小树): 基径20-30 mm; IV级(大树): 基径>30 mm。
1.2.4 数据处理
数据统计分析采用Microsoft Excel 2013软件完成。
2 结果分析
2.1 溪流弯道海南杜鹃空间分布格局与聚集强度
取样尺度是影响分布格局和聚集强度分析结果的重要因素(表2)。当尺度为1 m × 1 m时, 所有凸岸和凹岸的海南杜鹃种群都呈聚集分布; 在2 m × 2 m尺度下, 有从聚集分布向随机分布的变化趋势, 并且凹凸两岸表现出较大的差异; 在5 m × 5 m尺度下, 两岸的海南杜鹃种群都呈随机分布。Table 2
表2
表2不同研究尺度下的弯道两侧海南杜鹃种群的空间分布格局
Table 2
| 样带 Transect | 尺度 Scale (m) | 岸型 Bank type | 样本量 Sample size | 扩散系数 C | t检验 t test | Cassie指数 Ca | 平均拥挤度 m* | 丛生指数 I | 聚块性指数 m*/m | 负二项指数 K | 分布型 Pattern |
|---|---|---|---|---|---|---|---|---|---|---|---|
| L1 | 1 × 1 | 凹岸 Concave | 31 | 2.362 | 5.275** | 21.969 | 1.424 | 1.362 | 22.969 | 0.046 | C |
| 凸岸 Convex | 261 | 4.464 | 39.495** | 6.636 | 3.986 | 3.464 | 7.636 | 0.151 | C | ||
| 2 × 2 | 凹岸 Concave | 31 | 0.694 | -1.187 | -4.943 | -0.244 | -0.306 | -3.943 | -0.202 | R | |
| 凸岸 Convex | 261 | 1.549 | 6.258** | 1.051 | 1.071 | 0.549 | 2.051 | 0.951 | C | ||
| 5 × 5 | 凹岸 Concave | 31 | 0.338 | -2.563* | -10.675 | -0.600 | -0.662 | -9.675 | -0.094 | R | |
| 凸岸 Convex | 261 | 0.394 | -6.912** | -1.161 | -0.084 | -0.606 | -0.161 | -0.861 | R | ||
| L2 | 1 × 1 | 凹岸 Concave | 57 | 3.735 | 14.473** | 6.046 | 3.188 | 2.735 | 7.046 | 0.165 | C |
| 凸岸 Convex | 89 | 3.019 | 13.391** | 2.541 | 2.813 | 2.019 | 3.541 | 0.394 | C | ||
| 2 × 2 | 凹岸 Concave | 57 | 1.645 | 3.415** | 1.812 | 1.002 | 0.645 | 2.812 | 0.552 | C | |
| 凸岸 Convex | 89 | 1.982 | 6.512** | 1.412 | 1.677 | 0.982 | 2.412 | 0.708 | C | ||
| 5 × 5 | 凹岸 Concave | 57 | 0.803 | -1.044 | -0.692 | 0.088 | -0.197 | 0.308 | -1.445 | R | |
| 凸岸 Convex | 89 | 0.988 | -0.082 | -0.028 | 0.433 | -0.012 | 0.972 | -36.098 | R | ||
| D1 | 1 × 1 | 凸岸 Convex | 39 | 2.697 | 7.395** | 6.264 | 1.967 | 1.697 | 7.264 | 0.160 | C |
| 2 × 2 | 凸岸 Convex | 39 | 0.704 | -1.291 | -1.093 | -0.025 | -0.296 | -0.093 | -0.915 | R | |
| 5 × 5 | 凸岸 Convex | 39 | 0.307 | -3.021** | -5.332 | -0.563 | -0.693 | -4.332 | -0.188 | R | |
| D2 | 1 × 1 | 凹岸 Concave | 6 | 1.238 | 0.376 | 1.984 | 0.358 | 0.238 | 2.984 | 0.504 | C |
| 凸岸 Convex | 46 | 3.320 | 11.006** | 2.522 | 3.240 | 2.320 | 3.522 | 0.396 | C | ||
| 2 × 2 | 凹岸 Concave | 6 | 0.265 | -1.163 | -8.824 | -0.652 | -0.735 | -7.824 | -0.113 | R | |
| 凸岸 Convex | 46 | 1.050 | 0.237 | 0.078 | 0.689 | 0.050 | 1.078 | 12.811 | C | ||
| 5 × 5 | 凹岸 Concave | 6 | 0.027 | -1.539 | -8.111 | -0.853 | -0.973 | -7.111 | -0.123 | R | |
| 凸岸 Convex | 46 | 0.170 | -3.935** | -0.902 | 0.090 | -0.830 | 0.098 | -1.109 | R | ||
| J1 | 1 × 1 | 凸岸 Convex | 72 | 3.880 | 17.158** | 6.400 | 3.330 | 2.880 | 7.400 | 0.156 | C |
| 2 × 2 | 凸岸 Convex | 72 | 1.768 | 4.575** | 1.706 | 1.218 | 0.768 | 2.706 | 0.586 | C | |
| 5 × 5 | 凸岸 Convex | 72 | 0.678 | -1.918 | -0.894 | 0.038 | -0.322 | 0.106 | -1.118 | R | |
| J2 | 1 × 1 | 凸岸 Convex | 50 | 2.482 | 7.338** | 5.930 | 1.732 | 1.482 | 6.930 | 0.169 | C |
| 2 × 2 | 凸岸 Convex | 50 | 0.936 | -0.315 | -0.320 | 0.135 | -0.064 | 0.680 | -3.122 | R | |
| 5 × 5 | 凸岸 Convex | 50 | 0.173 | -4.092** | -3.307 | -0.577 | -0.827 | -2.307 | -0.302 | R |
新窗口打开|下载CSV
在2 m × 2 m的尺度下, 海南杜鹃种群在溪流弯道凸岸趋向呈聚集分布, 在凹岸则趋向呈随机分布(表2)。在调查的所有样带中, 除了D1和J2的凸岸的海南杜鹃种群呈随机分布, 其他溪流弯道凸岸的海南杜鹃都呈聚集分布; 除了L2凹岸的海南杜鹃种群呈聚集分布, 其他溪流弯道凹岸呈无分布或随机分布。参考所有聚集强度指标, 溪流弯道两侧海南杜鹃的个体数量和聚集强度都表现为: 凸岸>凹岸。各自然分布区的海南杜鹃种群聚集强度表现为: 黎母山>尖峰岭>吊罗山。
海南杜鹃在溪流弯道上游和下游的分布格局存在差异(表3)。在凸岸, 弯道下游的海南杜鹃种群基本呈聚集分布格局, 上游的海南杜鹃种群基本呈随机分布格局; 在凹岸, 弯道上下游基本呈无分布或随机分布。所有弯道两侧的海南杜鹃种群聚集强度基本表现为: 弯道下游>弯道上游。
Table 3
表3
表3弯道上下游海南杜鹃空间分布格局(尺度2 m × 2 m)
Table 3
| 样带 Transect | 岸型 Bank type | 上游/下游 Upstream/downstream | 扩散系数 C | t检验 t test | Cassie指数 Ca | 平均拥挤度 m* | 丛生指数 I | 聚块性指数 m*/m | 负二项指数 K | 分布型 Pattern |
|---|---|---|---|---|---|---|---|---|---|---|
| L1 | 凹岸 Concave | 上游 Upstream | 0.410 | -0.590 | -39.342 | -0.575 | -0.590 | -38.342 | -0.025 | R |
| 下游 Downstream | 0.702 | -1.096 | -3.196 | -0.205 | -0.298 | -2.196 | -0.313 | R | ||
| 凸岸 Convex | 上游 Upstream | 1.346 | 2.656** | 0.581 | 0.941 | 0.346 | 1.581 | 1.721 | C | |
| 下游 Downstream | 1.729 | 6.124** | 1.541 | 1.203 | 0.729 | 2.541 | 0.649 | C | ||
| L2 | 凹岸 Concave | 上游 Upstream | 0.447 | -0.874 | -7.368 | -0.478 | -0.553 | -6.368 | -0.136 | R |
| 下游 Downstream | 1.574 | 2.868** | 0.900 | 1.211 | 0.574 | 1.900 | 1.112 | C | ||
| 凸岸 Convex | 上游 Upstream | 1.154 | 0.409 | 0.659 | 0.389 | 0.154 | 1.659 | 1.518 | C | |
| 下游 Downstream | 1.837 | 5.056** | 0.724 | 1.993 | 0.837 | 1.724 | 1.382 | C | ||
| D1 | 凸岸 Convex | 上游 Upstream | 0.493 | -1.389 | -2.283 | -0.285 | -0.507 | -1.283 | -0.438 | R |
| 下游 Downstream | 0.759 | -0.780 | -0.788 | 0.065 | -0.241 | 0.212 | -1.270 | R | ||
| D2 | 凹岸 Concave | 上游 Upstream | 0.297 | -0.861 | -6.328 | -0.592 | -0.703 | -5.328 | -0.158 | R |
| 下游 Downstream | 0.219 | -0.552 | -14.063 | -0.726 | -0.781 | -13.063 | -0.071 | R | ||
| 凸岸 Convex | 上游 Upstream | 0.730 | -0.738 | -0.606 | 0.175 | -0.270 | 0.394 | -1.649 | R | |
| 下游 Downstream | 1.219 | 0.833 | 0.263 | 1.052 | 0.219 | 1.263 | 3.810 | C | ||
| J1 | 凸岸 Convex | 上游 Upstream | 0.861 | -0.340 | -0.853 | 0.024 | -0.139 | 0.147 | -1.172 | R |
| 下游 Downstream | 1.572 | 3.028** | 0.803 | 1.285 | 0.572 | 1.803 | 1.245 | C | ||
| J2 | 凸岸 Convex | 上游 Upstream | 0.483 | -0.896 | -7.091 | -0.444 | -0.517 | -6.091 | -0.141 | R |
| 下游 Downstream | 0.964 | -0.164 | -0.130 | 0.240 | -0.036 | 0.870 | -7.699 | R |
新窗口打开|下载CSV
河岸带不同发育阶段的海南杜鹃空间分布格局存在一定规律(表4)。幼苗更趋向于呈聚集分布格局, 龄级更大的海南杜鹃种群基本呈随机分布, 并且聚集强度随龄级的增加而减小。
Table 4
表4
表4不同龄级海南杜鹃空间分布格局(尺度2 m × 2 m)
Table 4
| 样带 Transect | 岸型 Bank type | 龄级 Age class | 扩散系数 C | t检验 t test | Cassie指数 Ca | 平均拥挤度 m* | 丛生指数 I | 聚块性指数 m*/m | 负二项指数 K | 分布型 Pattern |
|---|---|---|---|---|---|---|---|---|---|---|
| L1 | 凸岸 Convex | Ⅰ | 1.260 | 2.142* | 0.948 | 0.534 | 0.260 | 1.948 | 1.055 | C |
| Ⅱ | 0.705 | -1.723 | -2.141 | -0.157 | -0.295 | -1.141 | -0.467 | R | ||
| Ⅲ | 0.480 | -2.112* | -7.648 | -0.452 | -0.520 | -6.648 | -0.131 | R | ||
| Ⅳ | 0.498 | -1.588 | -11.960 | -0.460 | -0.502 | -10.960 | -0.084 | R | ||
| L2 | 凸岸 Convex | Ⅰ | 0.670 | -1.379 | -1.172 | -0.048 | -0.330 | -0.172 | -0.853 | R |
| Ⅱ | 0.928 | -0.274 | -0.307 | 0.162 | -0.072 | 0.693 | -3.253 | R | ||
| Ⅲ | 0.590 | -1.004 | -4.037 | -0.308 | -0.410 | -3.037 | -0.248 | R | ||
| Ⅳ | 0.451 | -1.228 | -6.390 | -0.463 | -0.549 | -5.390 | -0.157 | R | ||
| D1 | 凸岸 Convex | Ⅰ | 0.616 | -1.486 | -1.782 | -0.168 | -0.384 | -0.782 | -0.561 | R |
| Ⅱ | 0.386 | -0.971 | -14.743 | -0.573 | -0.614 | -13.743 | -0.068 | R | ||
| Ⅲ | 0.243 | -0.535 | -54.514 | -0.743 | -0.757 | -53.514 | -0.018 | R | ||
| Ⅳ | - | - | - | - | - | - | - | - | ||
| D2 | 凸岸 Convex | Ⅰ | 1.156 | 0.623 | 0.283 | 0.706 | 0.156 | 1.283 | 3.529 | C |
| Ⅱ | 0.259 | -1.386 | -5.558 | -0.608 | -0.741 | -4.558 | -0.180 | R | ||
| Ⅲ | 0.330 | -0.820 | -10.045 | -0.603 | -0.670 | -9.045 | -0.100 | R | ||
| Ⅳ | 0.250 | - | -45.000 | -0.733 | -0.750 | -44.000 | -0.022 | R | ||
| J1 | 凸岸 Convex | Ⅰ | 1.643 | 3.582** | 1.634 | 1.037 | 0.643 | 2.634 | 0.612 | C |
| Ⅱ | 0.370 | -1.261 | -11.206 | -0.574 | -0.630 | -10.206 | -0.089 | R | ||
| Ⅲ | - | - | - | - | - | - | - | - | ||
| Ⅳ | - | - | - | - | - | - | - | - | ||
| J2 | 凸岸 Convex | Ⅰ | 0.839 | -0.646 | -1.174 | -0.024 | -0.161 | -0.174 | -0.852 | R |
| Ⅱ | 0.733 | -0.706 | -4.271 | -0.204 | -0.267 | -3.271 | -0.234 | R | ||
| Ⅲ | 0.250 | - | -180.000 | -0.746 | -0.750 | -179.000 | -0.006 | R | ||
| Ⅳ | 0.250 | - | -180.000 | -0.746 | -0.750 | -179.000 | -0.006 | R |
新窗口打开|下载CSV
2.2 溪流弯道海南杜鹃种群结构
三个地区的海南杜鹃都在溪流弯道密集分布, 并且都为幼苗个体占优的增长型种群(图2), 个体数量随龄级增大逐渐减少(图2, 图3)。其中, 黎母山的海南杜鹃种群个体数量最多, 种群结构最完整, 而吊罗山和尖峰岭的小树和大树个体明显偏少。图2
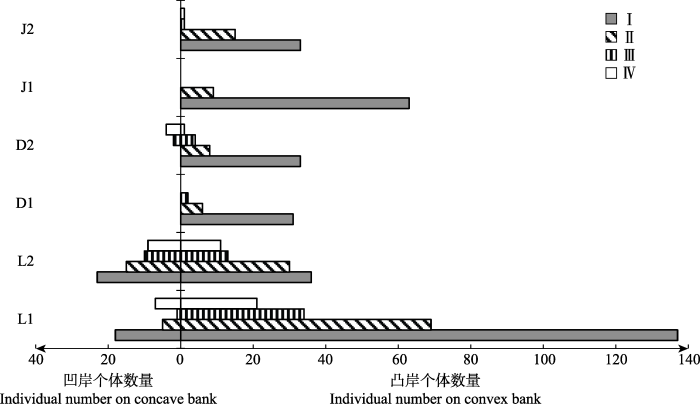 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2溪流弯道两侧海南杜鹃种群的年龄结构。D1, 吊罗山1号样带; D2, 吊罗山2号样带; J1, 尖峰岭1号样带; J2, 尖峰岭2号样带; L1, 黎母山1号样带; L2, 黎母山2号样带。I, 幼苗; II, 幼树; III, 小树; IV, 大树。
Fig. 2Age structures of the Rhododendron hainanense populations on both sides of stream bends. D1, No. 1 transect of Mt. Diaoluo; D2, No. 2 transect of Mt. Diaoluo; J1, No. 1 transect of Mt. Jianfeng; J2, No. 2 transect of Mt. Jianfeng; L1, No. 1 transect of Mt. Limu; L2, No. 2 transect of Mt. Limu. I, seedling; II, sapling; III, undershrub; IV, large shrub.
图3
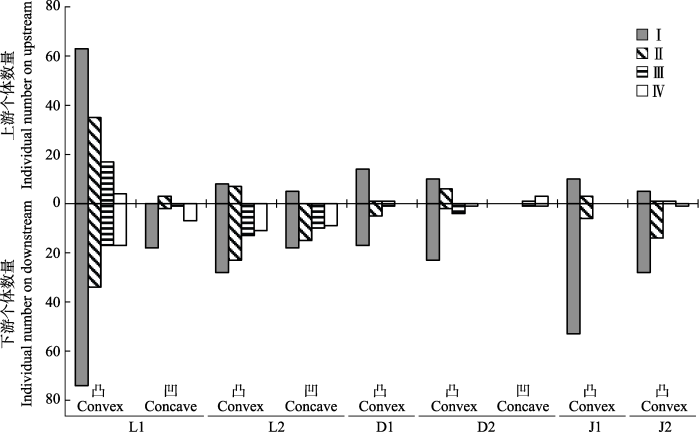 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3溪流弯道上下游海南杜鹃种群的年龄结构。D1, 吊罗山1号样带; D2, 吊罗山2号样带; J1, 尖峰岭1号样带; J2, 尖峰岭2号样带; L1, 黎母山1号样带; L2, 黎母山2号样带。I, 幼苗; II, 幼树; III, 小树; IV, 大树。
Fig. 3Age structures of the Rhododendron hainanense populations on upstream and downstream of stream bends. D1, No. 1 transect of Mt. Diaoluo; D2, No. 2 transect of Mt. Diaoluo; J1, No. 1 transect of Mt. Jianfeng; J2, No. 2 transect of Mt. Jianfeng; L1, No. 1 transect of Mt. Limu; L2, No. 2 transect of Mt. Limu. I, seedling; II, sapling; III, undershrub; IV, large shrub.
溪流弯道凸岸和凹岸的海南杜鹃种群结构存在明显差异(图2)。凸岸群体都为增长型, 凹岸只有L1、L2和D2有海南杜鹃分布, 其中L1和L2凹岸为稳定增长型, D2凹岸为衰退型。凸岸海南杜鹃幼苗数量远多于凹岸, 凹岸群体由于个体数较少而存在断龄现象。
溪流弯道上游和下游的海南杜鹃种群结构也存在明显差异(图3)。凸岸群体在弯道上下游都为增长型, 且下游各龄级海南杜鹃的个体数量基本都多于弯道上游, 种群结构更稳定。凹岸弯道上下游的海南杜鹃种群则出现严重断龄现象, 只有L2凹岸下游群体属于增长型, 其他凹岸上下游都为衰退型。
3 讨论
3.1 河岸带弯道海南杜鹃种群空间分布格局
在河道形态变化过程中, 河岸带植物和河岸之间有相互依存的关系(Corenblit et al., 2007; Camporeale et al., 2013; Curran & Hession, 2013)。河道的形态演化推动了植物群落的扩张(Bertoldi et al., 2011), 河岸两侧生态环境的异质性导致河岸带植被呈斑块分布(韩路等, 2013)。海南杜鹃在溪流弯道凸岸数量远多于凹岸, 可能与弯道两岸的基质与地形有关(Bendix & Hupp, 2000)。所有样带中, 除了黎母山L2的凹岸为沙质河岸, 其他样带的凹岸岸边都有大块裸露的岩石, 岩石间存在较大的空隙, 并且河岸较开阔, 起伏不平, 与溪流水面的垂直高差较大, 不利于植物固着生长; 而凸岸多为缓坡, 有大量小石头, 石头间充满了沙土, 植物生长茂密。弯道内外两岸地形地貌与基质的差异主要是由于河道水流造成的(Bertoldi et al., 2011)。当水流进入弯道上游河段, 离心力使凹岸水位比凸岸高, 形成水面横比降, 在弯道断面产生横向环流。弯曲河道中, 水流不断侵蚀凹岸的泥沙, 泥沙和较小的石块在横向环流的作用下从凹岸向凸岸转移, 并不断沉积形成浅滩(刘月琴, 2003); 而雨季洪水冲落的大岩石在离心力和重力的作用下更容易沉降在凹岸, 导致凸岸和凹岸的地貌特征存在明显差异。海南杜鹃性喜河岸带透水性良好的沙土或砂壤土, 溪流弯道凸岸的小石块与泥沙在混合沉降后, 适合海南杜鹃根系的生长。海南杜鹃繁密的根系和旺盛的分枝生长又反过来增强了河道的防侵蚀能力(Horvath, 2004; Tal & Paola, 2010; Gurnell et al., 2012)。
植物的繁殖体也更容易沉积和聚集在流速较慢、坡度较缓的弯道凸岸(Burkart, 2001; Merritt et al., 2010b)。室内初步实验发现, 海南杜鹃种子吸水后可沉入水中, 受水流搬运的方式与沙粒相似, 海南杜鹃种子可能在横向环流的作用下由凹岸向凸岸转移, 逐渐沉积或被原有植物根系拦截在凸岸附近及其下游一带, 导致海南杜鹃在溪流弯道凸岸形成聚集分布格局, 并且在凸岸下游的聚集强度最大。
海南杜鹃的花期正处海南岛的旱季, 果实与种子成熟时间是雨季和台风季节。细小的种子在风、雨、流水的作用下可以实现长距离的扩散, 特别是台风导致的溪流山洪成了海南杜鹃种子高效而快速的传播渠道。这可能是河岸带植物普遍的一种种子扩散适应对策(Drezner et al., 2001; Richardson et al., 2007; Greet et al., 2011; Solari et al., 2016)。
3.2 河岸带弯道海南杜鹃种群年龄结构
溪流弯道不同区域的海南杜鹃群体基本都是幼苗数量远多于龄级较大的个体, 这可能与洪水对海南杜鹃繁殖体的搬运作用有关。一般而言, 沙质土壤不利于植物根系深扎, 幼苗难以在暴雨和山洪中生存(达良俊等, 2004; 魏新增等, 2008; Balke et al., 2013)。在受水流冲刷最严重的凹岸, 海南杜鹃种群结构易呈单峰或间歇峰, 而凸岸下游受到的冲刷强度最小, 泥沙沉积量大, 使凸岸下游形成最适宜海南杜鹃等河岸带植物生长的环境。海南杜鹃种子在沉积后对凸岸的种群进行更新和补充, 导致凸岸弯顶偏下的海南杜鹃分布密集、呈稳定增长型, 成为该区域灌木层优势种。调查发现, 海南杜鹃在近弯顶位置易形成稳定的以幼苗和小树为主的增长型群体, 距离河岸或弯顶越远, 各龄级个体数量越呈下降趋势, 说明海南杜鹃生长与种子萌发需要较多的水分和光照条件。由于河岸带种内和种间竞争大, 而且年龄越大的个体要承受越多的洪水冲刷(康华靖等, 2007; 黄小波等, 2015), 导致海南杜鹃群体密度和聚集强度随龄级增加而减小。
汛期山洪对河岸的过度侵蚀或横向迁移可能会破坏现有的生态系统(Piégay et al., 2005; Bhuiyan et al., 2010), 但海南杜鹃的许多特性与生活策略有助于植株在激流中减少生物量损失并加快种群恢复。幼苗阶段的海南杜鹃枝条较软, 萌枝能力强, 山洪携带的石块撞击植株后留下的创伤处可萌生新枝及不定根(何东等, 2009; Catford & Jansson, 2014; Garssen et al., 2015)。特别是在经受大岩石的冲击和堆积的凹岸, 龄级较大的海南杜鹃植株萌发新枝与新根的比例较大(Merritt et al., 2010a; Balke et al., 2013); 而在植物生长较茂密的凸岸, 种子沉积较多, 实生苗的更新较多。
因此, 由于山区溪流水流较快, 弯道内外两侧基质与地形差别较大, 海南杜鹃及类似的河岸带植物在局域尺度上就出现了较明显的空间分化格局, 即在弯道内外两侧的植物生长、繁衍及种群年龄结构存在较大差异。这种局域尺度上的差异可能影响到了该植物的种群动态及河岸带的生态过程, 有可能进一步影响到局域物种相互作用与群落构建, 值得今后的河岸带生物多样性研究给予重视。

致谢
感谢海南大学热带农林学院向文倩、黎母山自然保护区护林员黎炳新、吊罗山自然保护区护林员梁宜文和尖峰岭自然保护区护林员苏红华在野外调查工作中给予的帮助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1111/1365-2664.12067URLPMID:23894211 [本文引用: 2]

1.Vegetated biogeomorphic systems (e.g. mangroves, salt marshes, dunes, riparian vegetation) have been intensively studied for the impact of the biota on sediment transport processes and the resulting self-organization of such landscapes. However, there is a lack of understanding of physical disturbance mechanisms that limit primary colonization in active sedimentary environments. 2.This study elucidates the effect of sediment disturbance during the seedling stage of pioneer vegetation, using mangroves as a model system. We performed mesocosm experiments that mimicked sediment disturbance as (i) accretion/burial of plants and (ii) erosion/excavation of plants of different magnitudes and temporal distribution in combination with water movement and inundation stress. 3.Cumulative sediment disturbance reduced seedling survival, with the faster-growingAvicennia albashowing less mortality than the slower-growingSonneratia alba. The presence of the additional stressors (inundation and water movement) predominantly reduced the survival ofS. alba. 4.Non-lethal accretion treatments increased shoot biomass of the seedlings, whereas non-lethal erosion treatments increased root biomass allocation. This morphological plasticity in combination with the abiotic disturbance history determined how much maximum erosion the seedlings were able to withstand. 5.Synthesis and applications. Seedling survival in dynamic sedimentary environments is determined by the frequency and magnitude of sediment accretion or erosion events, with non-lethal events causing feedbacks to seedling stability. Managers attempting restoration of mangroves, salt marshes, dunes and riparian vegetation should recognize sediment dynamics as a main bottleneck to primary colonization. The temporal distribution of erosion and accretion events has to be evaluated against the ability of the seedlings to outgrow or adjust to disturbances. Our results suggest that selecting fast-growing pioneer species and measures to enhance seedling growth or temporary reduction in sediment dynamics at the restoration site can aid restoration success for vegetated biogeomorphic ecosystems.
DOI:10.1002/(ISSN)1099-1085URL [本文引用: 1]
DOI:10.1002/esp.v36.11URL [本文引用: 2]
DOI:10.1061/(ASCE)HY.1943-7900.0000217URL [本文引用: 1]
The effectiveness of a novel approach of using vanes, installed at a low angle and attached to the bank, for bank protection and for the restoration of river meanders has been investigated in a laboratory study. Experiments were carried out in a large-scale meandering mobile-bed channel with graded sediment. The bed topography, three-dimensional flow pattern, and turbulence characteristics in the meandering channel with or without structures are analyzed. When a single or an array of such vanes is installed, the scour hole at the base of the outer bank is infilled and the thalweg is relocated toward the center of the river. The structures induce a secondary flow cell near the outer bank which counteracts the main spiral flow in the bend. In contrast to common spurs and bendway weirs, large-scale horizontal vortices are not generated behind the structures. Vanes which grade to the bed from bankfull level at the bank show better performance than low level ones, whereas multiple structures show positive effects as far downstream as the crossover section.
DOI:10.1046/j.1466-822x.2001.00270.xURL [本文引用: 1]
In Central European lowlands certain plant species grow mainly or exclusively in the corridors of large rivers. In German-speaking plant geography, they are known as 'Stromtalpflanzen'. The aim of this paper is to review the literature about definitions, explanations and species characteristics and to suggest future directions in research concerning this species group.
DOI:10.1002/eco.140URL [本文引用: 1]
Riparian zones have become increasingly important elements of river basin ecohydrology given their relevance of nutrient dynamics to water quality management. Most attention has been given to nitrogen cycling, denitrification in particular, but new processes have emerged as potentially important and need to be evaluated. Riparian zones are sensitive locations, so it is important to understand the impact of changes that can fundamentally alter their ecosystem processes and responses. Better understanding of how riparian buffering varies in time and space provides the basis for a catchment-scale approach, but it seems likely that low-order basins will remain the focus for research and management, given that low-order basins constitute the majority of basin area. Finally, it is argued that new ecohydrological indicators are needed that combine innovative biogeochemical parameters and landscape connectivity measures. Copyright 2010 John Wiley & Sons, Ltd.
DOI:10.1002/rog.20014URL [本文引用: 1]
[1] The study of river-riparian vegetation interactions is an important and intriguing research field in geophysics. Vegetation is an active element of the ecological dynamics of a floodplain which interacts with the fluvial processes and affects the flow field, sediment transport, and the morphology of the river. In turn, the river provides water, sediments, nutrients, and seeds to the nearby riparian vegetation, depending on the hydrological, hydraulic, and geomorphological characteristic of the stream. In the past, the study of this complex theme was approached in two different ways. On the one hand, the subject was faced from a mainly qualitative point of view by ecologists and biogeographers. Riparian vegetation dynamics and its spatial patterns have been described and demonstrated in detail, and the key role of several fluvial processes has been shown, but no mathematical models have been proposed. On the other hand, the quantitative approach to fluvial processes, which is typical of engineers, has led to the development of several morphodynamic models. However, the biological aspect has usually been neglected, and vegetation has only been considered as a static element. In recent years, different scientific communities (ranging from ecologists to biogeographers and from geomorphologists to hydrologists and fluvial engineers) have begun to collaborate and have proposed both semiquantitative and quantitative models of river-vegetation interconnections. These models demonstrate the importance of linking fluvial morphodynamics and riparian vegetation dynamics to understand the key processes that regulate a riparian environment in order to foresee the impact of anthropogenic actions and to carefully manage and rehabilitate riparian areas. In the first part of this work, we review the main interactions between rivers and riparian vegetation, and their possible modeling. In the second part, we discuss the semiquantitative and quantitative models which have been proposed to date, considering both multi- and single-thread rivers.
DOI:10.1111/nph.12951URLPMID:25130059 [本文引用: 1]
Contents
DOI:10.1016/j.earscirev.2007.05.004URL [本文引用: 2]
Until recently, one-way relationships between flow dynamics, geomorphology and plant ecology were considered dominantly when studying the functioning of river systems, whereby fluvial landforms and hydrogeomorphic processes drive the evolution of riparian plant communities. However, biological communities may significantly control geomorphic processes and have strong impacts on landform dynamics. In order to fully identify the processes linked to river dynamics (changes in time and space of fluvial landforms and associated plant communities), conceptual multidisciplinary progress is clearly needed. To understand the mutual interactions and feedbacks between fluvial landforms and vegetation community dynamics, this paper presents a detailed literature review of fluvial geomorphology, riparian plant ecology and hydraulic engineering knowledge. The historical and recent development of ecological plant succession theory toward the integration of hydrogeomorphic disturbances is discussed as well as the integration of vegetation within geomorphology as a significant landform control factor, incorporating both hydrogeomorphic controls on riparian vegetation dynamics and mechanical impacts of vegetation structures on flow properties and sediment dynamics. Recent progress in ecology, hydraulic engineering and fluvial geomorphology emphasises interdependence between biological and physical forms and processes. Based on this literature review, a ‘fluvial biogeomorphic succession’ concept is proposed to link fluvial landform and riparian vegetation community evolution within a bi-directional model. The succession of fluvial landforms and associated vegetation communities is composed of four main critical phases that represent a shift in the relative dominance of hydrogeomorphic and ecological processes as a response to biostabilisation and passive bioconstruction processes. The positive feedbacks associated with this shift lead to the development of characteristic biogeomorphic structures such as vegetated banks, islands or floodplains, which are moderated by the biogeomorphic functional roles of ‘ecosystem engineers’ that induce or reinforce the positive feedbacks. This fluvial biogeomorphic succession concept relates the natural Darwinian selection and ecological succession theories to fluvial geomorphology.
[本文引用: 1]
DOI:10.1016/j.jhydrol.2013.10.013URL [本文引用: 2]
Vegetation creates a complicated system of feedbacks and linkages across the fluvial system that is realized through river planform shape. Interactions occur among flow hydraulics, sediment deposition and erosion, and plant morphology, density, and biomechanics. Interest in the interactions and feedback loops between vegetation and the fluvial system has grown extensively in the past few years. This interest is partially driven by the popularity of stream restoration activities worldwide that include re-vegetation of stream banks and formation of an ecosystem that is intended to encourage the growth of aquatic macrophytes. We present a review of the research into the interactions and dependencies between vegetation and the fluvial system to identify hydraulic and sediment dynamics that are consistent around vegetation located in the channel, on channel banks, and over the floodplain. We illustrate process commonalities operating across spatial locations within the fluvial system and highlight some of the current research opportunities and challenges to encourage research collaborations between those working in areas of the fluvial system traditionally viewed as disparate.
DOI:10.17521/cjpe.2004.0054URL [本文引用: 2]
以浙江天童国家森林公园常绿阔叶林为对象 ,应用永久样方法和每木调查法调查了群落种类组成和结构 ,并对主要组成种的种群结构进行了分析。依据胸径级频率分布的形状 ,将各树种的种群结构归纳为 5种类型 ,并结合生物学、生态学特征 ,分别讨论了其更新类型 ,同时对调查群落所处的演替阶段进行了诊断。结果表明 :1)群落为 6种共优势群落 ,出现木本植物 6 9种。 2 )种群结构为单峰型的马尾松 (Pinusmassoniana)、枫香 (Liquidambarformosana)和檫木 (Sassafrastzumu)等为阳性高大乔木 ,无正常更新能力 ,更新类型是先锋群落优势种或是顶极性先锋种 ;间歇型的木荷 (Schimasuperba)、黑山山矾 (Symplocosheishanensis)和红楠 (Machilusthunbergii)等为具有耐荫性的常绿阔叶树种 ,更新具有波动性和机会性 ,是演替系列或顶极群落优势种 ;逆_J字型的米槠 (Castanopsiscarlesii)、栲树 (Castanopsisfargesii)和长叶石栎 (Lithocarpusharlandii)等为强耐荫性的常绿阔叶树种 ,通过幼苗库和根萌生枝进行更新 ,是顶极群落的优势种 ;浙江新木姜子 (Neolitseaauratavar.chekiangensis)、厚皮香 (Ternstroemiagymnanthera)等为L字型 ;连蕊茶 (Camelliafraternna)、老鼠矢 (Symplocosstellaris)等为单柱型 ,此二型是群落亚
DOI:10.17521/cjpe.2004.0054URL [本文引用: 2]
以浙江天童国家森林公园常绿阔叶林为对象 ,应用永久样方法和每木调查法调查了群落种类组成和结构 ,并对主要组成种的种群结构进行了分析。依据胸径级频率分布的形状 ,将各树种的种群结构归纳为 5种类型 ,并结合生物学、生态学特征 ,分别讨论了其更新类型 ,同时对调查群落所处的演替阶段进行了诊断。结果表明 :1)群落为 6种共优势群落 ,出现木本植物 6 9种。 2 )种群结构为单峰型的马尾松 (Pinusmassoniana)、枫香 (Liquidambarformosana)和檫木 (Sassafrastzumu)等为阳性高大乔木 ,无正常更新能力 ,更新类型是先锋群落优势种或是顶极性先锋种 ;间歇型的木荷 (Schimasuperba)、黑山山矾 (Symplocosheishanensis)和红楠 (Machilusthunbergii)等为具有耐荫性的常绿阔叶树种 ,更新具有波动性和机会性 ,是演替系列或顶极群落优势种 ;逆_J字型的米槠 (Castanopsiscarlesii)、栲树 (Castanopsisfargesii)和长叶石栎 (Lithocarpusharlandii)等为强耐荫性的常绿阔叶树种 ,通过幼苗库和根萌生枝进行更新 ,是顶极群落的优势种 ;浙江新木姜子 (Neolitseaauratavar.chekiangensis)、厚皮香 (Ternstroemiagymnanthera)等为L字型 ;连蕊茶 (Camelliafraternna)、老鼠矢 (Symplocosstellaris)等为单柱型 ,此二型是群落亚
DOI:10.1046/j.1466-822x.2001.00216.xURL [本文引用: 1]
This study focuses on the relationships between plant dispersal syndromes and plant distributions at the community scale. Species composition and cover are reported from 29 10 20-m vegetation plots along five topographic cross-sections in the riparian zone of the Hassayampa River Preserve, Arizona. We find that spatial patterns of dispersal guilds vary within the flood plain of this semiarid region river. Our main results are: (1) wind-dispersed species are fairly evenly distributed at all elevations and distances from the river, whereas cover of animal-dispersed species increases with elevation above, and at greater distances from, the river; (2) wind-dispersed species are proportionally more abundant in the pioneer Populus alix community, whereas plants in the late-seral Prosopis community are predominantly animal-dispersed; (3) most of the species classified as obligate-wetland and facultative-wetland are wind-dispersed, whereas facultative-upland and obligate-upland species are mostly animal-dispersed; and (4) there are significantly fewer wind-dispersed species in areas of high total vegetation cover. These results may reflect successional patterns resulting from periodic flooding. Low areas close to the river flood more frequently and with greater intensity than areas farther from the river. Many pioneer species that establish in disturbed areas are wind-dispersed. Over successional time, pioneer species cede to more drought tolerant species that are predominantly animal-dispersed.
[本文引用: 1]
DOI:10.1111/gcb.12921URLPMID:25752818 [本文引用: 1]
Abstract <p>A future higher risk of severe flooding of streams and rivers has been projected to change riparian plant community composition and species richness, but the extent and direction of the expected change remain uncertain. We conducted a meta-analysis to synthesize globally available experimental evidence and assess the effects of increased flooding on (1) riparian adult plant and seedling survival, (2) riparian plant biomass and (3) riparian plant species composition and richness. We evaluated which plant traits are of key importance for the response of riparian plant species to flooding. We identified and analysed 53 papers from ISI Web of Knowledge which presented quantitative experimental results on flooding treatments and corresponding control situations. Our meta-analysis demonstrated how longer duration of flooding, greater depth of flooding and, particularly, their combination reduce seedling survival of most riparian species. Plant height above water level, ability to elongate shoots and plasticity in root porosity were decisive for adult plant survival and growth during longer periods of flooding. Both ‘quiescence’ and ‘escape’ proved to be successful strategies promoting riparian plant survival, which was reflected in the wide variation in survival (full range between 0 and 100%) under fully submerged conditions, while plants that protrude above the water level (>2002cm) almost all survive. Our survey confirmed that the projected increase in the duration and depth of flooding periods is sufficient to result in species shifts. These shifts may lead to increased or decreased riparian species richness depending on the nutrient, climatic and hydrological status of the catchment. Species richness was generally reduced at flooded sites in nutrient-rich catchments and sites that previously experienced relatively stable hydrographs (e.g. rain-fed lowland streams). Species richness usually increased at sites in desert and semi-arid climate regions (e.g. intermittent streams).</p>
DOI:10.1111/j.1365-2427.2011.02564.xURL [本文引用: 1]
1. Whilst it is widely recognised that a natural flow regime is important for sustaining riverine ecosystems, the relative importance of the various components of flow regime for riparian vegetation dynamics is poorly understood. We sought to determine the current extent of knowledge on the importance of seasonal flow timing for riparian plants by conducting a systematic review of the literature using causal criteria analysis.2. Using a definition of iparian that included riverine, wetland and floodplain systems, we found sufficient evidence to provide strong support for the existence of causal relations between seasonal flow timing and a number of riparian plant processes, namely rates of waterborne dispersal (hydrochory), germination and growth, as well as riparian community composition. There was insufficient evidence to infer a causal relationship between flow timing and the reproduction or survival of riparian plants.3. Thus, we argue that seasonal flow timing is important for many of the processes that generate and sustain riparian vegetation communities. River regulation, and/or flow management aimed at restoring ecological values, should consider flow timing and its implication for riparian flora. Because of regulation, many of the rivers of south-eastern Australia have inverted seasonal flow patterns. Whilst direct evidence of the effects of this inversion on the flora of these rivers is lacking, the results of our causal analysis allow us to predict how these plant communities may have been affected.4. However, these predictions must be treated with caution because of the reliance of some of the causal analyses on wetland studies. For riverine flora, further research is particularly needed on the effects of seasonal flow timing on hydrochory, survival and reproduction.5. Causal criteria analysis provides a defensible and efficient means for assessing the extent of evidence for or against ecological hypotheses of this kind. In this case, systematic review of the literature provided strong evidence to support a number of causal links between seasonal flow timing and riparian vegetation dynamics, whilst also efficiently identifying knowledge gaps.
DOI:10.1016/j.earscirev.2011.11.005URL [本文引用: 1]
The fluvial riparian and aquatic patch mosaic varies along rivers according to geomorphological setting, hydrological regime, sediment supply and surface鈥揼roundwater connectivity. This relation between physical processes and plants is not unidirectional. Once established, riparian and aquatic plants frequently act as physical ecosystem engineers by trapping and stabilising sediments, organic matter and the propagules of other plant species, modifying the local sedimentary and morphological environment by driving the development of landforms and associated habitats, and so facilitating the rapid establishment of other plants that can in turn reinforce the development of landforms such as river banks, vegetated islands and floodplains. This paper reviews knowledge on the hydrogeomorphological significance of riparian and aquatic vegetation with a particular emphasis on humid temperate, mixed load, gravel bed, floodplain rivers. First, we investigate how vegetation dynamics across river margins are governed by hydrological processes that can both promote riparian vegetation growth and disturb and destroy riparian and aquatic vegetation. We show, with some simple numerical modeling, that different combinations of moisture supply and flow disturbance have the potential to generate many different responses in the lateral distribution of vegetation biomass along river corridors. Second, building on the varied lateral biomass distributions that are primarily dictated by hydrological processes, we review research evaluating characteristic vegetation-mediated landform development. We investigate aquatic and riparian plants acting as physical ecosystem engineers by creating and modifying habitats in river systems with sufficient suspended sediment supply for habitat or landform building. These plants have a crucial impact on sediment stabilisation and pioneer landform building along the interface between plant (resistance) dominated and fluvial-disturbance (force) dominated zones of the river corridor. We present some examples of vegetation-mediated landforms along rivers with strongly contrasting hydrological regimes and thus lateral distributions of vegetation. Lastly, we present a conceptual synthetic model that links the development of pioneer landforms by engineering plants with river morphology and morphodynamics in humid temperate, mixed load, gravel bed, floodplain rivers. Drawing on four example rivers, we show how different plants and pioneer landforms act at the interface between the plant dominated and fluvial-disturbance dominated zones of the river corridor as river energy and vegetation colonisation and growth change.
[本文引用: 1]
[本文引用: 1]
DOI:10.3773/j.issn.1005-264x.2009.03.006URL [本文引用: 1]
以种群统计学和树木年代学的方法,从年龄结构、生命表、径向生长和干扰史4个方面,研究了神农架山地河岸带连香树(Cercidiphyllum japonicum)的种群结构与动态。结果显示:1)连香树种群内幼龄个体比较充足,但在长时段内更新时有断代,多个局域种群有明显可辨的同生群(年龄幅宽约为50a),其发生年代在20世纪20年代和70年代,高峰期则在40年代和90年代;2)萌枝更新非常突出,与种群年龄和群落盖度有显著相关(r〉0.6,p〈0.05);3)经20a以下和60~100a间两个死亡高峰以后,种群可达生活力最优阶段,180a以后出现衰老迹象,生存曲线总体上介于DeeveyⅡ和Ⅲ型之间;4)前30a连香树胸径的生长速率最快,经2次生长释放后,约在60a前后进入林冠层,90~120a间进入成熟期前,胸径分化最为剧烈;5)干扰高峰年(20世纪20年代、70年代和90年代)与种群更新的高峰年大致吻合。这些结果表明,个体生长发育特征与中长期的干扰对其种群结构与动态有重要影响。由于幼苗尤其是萌枝的的数量充足,连香树种群在中短期内可以维持。
DOI:10.3773/j.issn.1005-264x.2009.03.006URL [本文引用: 1]
以种群统计学和树木年代学的方法,从年龄结构、生命表、径向生长和干扰史4个方面,研究了神农架山地河岸带连香树(Cercidiphyllum japonicum)的种群结构与动态。结果显示:1)连香树种群内幼龄个体比较充足,但在长时段内更新时有断代,多个局域种群有明显可辨的同生群(年龄幅宽约为50a),其发生年代在20世纪20年代和70年代,高峰期则在40年代和90年代;2)萌枝更新非常突出,与种群年龄和群落盖度有显著相关(r〉0.6,p〈0.05);3)经20a以下和60~100a间两个死亡高峰以后,种群可达生活力最优阶段,180a以后出现衰老迹象,生存曲线总体上介于DeeveyⅡ和Ⅲ型之间;4)前30a连香树胸径的生长速率最快,经2次生长释放后,约在60a前后进入林冠层,90~120a间进入成熟期前,胸径分化最为剧烈;5)干扰高峰年(20世纪20年代、70年代和90年代)与种群更新的高峰年大致吻合。这些结果表明,个体生长发育特征与中长期的干扰对其种群结构与动态有重要影响。由于幼苗尤其是萌枝的的数量充足,连香树种群在中短期内可以维持。
DOI:10.3321/j.issn:1001-7488.2006.10.003URLMagsci [本文引用: 1]
<p>对沙冬青的株高和冠幅进行分级,研究磴口县3个沙冬青种群的大小结构和年龄结构,应用扩散系数<em>C</em>、负二次指数<em>K</em>、平均拥挤度<em>m</em><sup>*</sup>、丛生指数<em>I、Cassie</em>指数1/<em>K</em>、聚块性指数<em>m</em><sup>*</sup>/<em>m</em>和扩散型指数<em>I</em><sub>δ</sub>7种聚集度指标确定不同生境条件下沙冬青种群的空间分布格局类型和动态,考察沙冬青种群在不同尺度上的空间分布格局。结果表明不同生境条件下的沙冬青种群结构有差异,但都呈现衰退趋势。不同生境的沙冬青种群空间分布格局类型和聚集强度不同,聚集强度在不同尺度上表现出基本一致的变化趋势。样地2在25和100m<sup>2</sup>的范围内集群分布,样地3在150m<sup>2</sup>范围内集群分布,样地1在7个取样面积下,均成随机分布。不同发育阶段种群的分布格局为幼龄和老龄植株均成随机分布,中龄植株呈聚集分布。环境是沙冬青种群空间分布格局形成和发展的决定因子之一。</p>
DOI:10.3321/j.issn:1001-7488.2006.10.003URLMagsci [本文引用: 1]
<p>对沙冬青的株高和冠幅进行分级,研究磴口县3个沙冬青种群的大小结构和年龄结构,应用扩散系数<em>C</em>、负二次指数<em>K</em>、平均拥挤度<em>m</em><sup>*</sup>、丛生指数<em>I、Cassie</em>指数1/<em>K</em>、聚块性指数<em>m</em><sup>*</sup>/<em>m</em>和扩散型指数<em>I</em><sub>δ</sub>7种聚集度指标确定不同生境条件下沙冬青种群的空间分布格局类型和动态,考察沙冬青种群在不同尺度上的空间分布格局。结果表明不同生境条件下的沙冬青种群结构有差异,但都呈现衰退趋势。不同生境的沙冬青种群空间分布格局类型和聚集强度不同,聚集强度在不同尺度上表现出基本一致的变化趋势。样地2在25和100m<sup>2</sup>的范围内集群分布,样地3在150m<sup>2</sup>范围内集群分布,样地1在7个取样面积下,均成随机分布。不同发育阶段种群的分布格局为幼龄和老龄植株均成随机分布,中龄植株呈聚集分布。环境是沙冬青种群空间分布格局形成和发展的决定因子之一。</p>
DOI:10.1016/j.aquabot.2003.09.003URL [本文引用: 1]
Retention of coarse particulate organic matter (CPOM) in streams is affected by channel complexity, especially three-dimensional structures in stream channels. Much attention has focused on woody debris as retention structures, but macrophytes should function similarly. Retention and flow characteristics were measured in 5 m long reaches at four-replicate sites in the Breitenbach (Hessen, Germany). CPOM retention was measured by releasing chips of paper (6 mm diameter) as analog CPOM. Triplicate releases (200 chips each) were done before and after submerged, emergent and overhanging macrophytes were removed. Discharge ranged between 3 and 4.5 l s 611 during the experimental releases. Macrophyte biomass removed was 3.047, 3.012, 0.164, and 1.392 kg m 612 (fresh weight) respectively from sites 1 to 4. Instantaneous retention rates, calculated with an exponential decay function, were significantly higher ( P<0.001) with macrophytes than without macrophytes (0.8±0.2 versus 0.2±0.1). Velocity also was significantly reduced by macrophyte presence ( P<0.001). Macrophyte biomass was a significant predictor of retention ( P<0.001), whereas discharge was not ( P=0.3). Hence, retention appears to be positively related to macrophyte biomass. Macrophytes in the Breitenbach increase retention by forming effective sieve-like structures in the stream physically trapping CPOM and by reducing water velocity.
DOI:10.5846/stxb201406051159URL [本文引用: 1]
思茅松是云南省重要的材脂兼用树种,而择伐是思茅松天然林管理的有效手段,也是获取木材的重要途径.基于对云南省景谷县思茅松天然林的样地调查数据,采用种群径级结构代替年龄结构、Ripley的L函数点格局等方法,对思茅松天然林在不同强度择伐10a后的种群结构和空间分布格局及其不同发育阶段在不同尺度下的相互关系进行了系统研究和分析.结果表明:择伐10a后3种类型的思茅松自然种群的胸径、树高之间呈显著的正相关且可用幂指数模拟预测模型.未择伐的思茅松种群呈衰退型,轻度择伐(12.9%)后种群呈稳定型,中度择伐(29.3%)后的种群属增长型,适当的中度择伐有利于自然种群的更新;未择伐与中度择伐后的思茅松种群分布格局整体呈随机型,但后者较前者更趋向于均匀分布,轻度择伐后的思茅松种群分布格局随着尺度的增加由随机转向聚集分布.择伐对不同径级思茅松天然林种群空间分布格局有重要影响,未择伐的思茅松自然种群随着发育阶段的发展,多数尺度上由随机分布逐渐转变为聚集分布,至大树则又以随机分布为主;轻度择伐和中度择伐的思茅松自然种群则随着发育阶段的发展而由完全的随机分布逐渐转变为聚集分布,中度择伐的聚集分布则更为明显.思茅松不同发育阶段间在所有尺度上基本都呈显著正相关,表明思茅松种群种内竞争弱,有利于种群的维持.
DOI:10.5846/stxb201406051159URL [本文引用: 1]
思茅松是云南省重要的材脂兼用树种,而择伐是思茅松天然林管理的有效手段,也是获取木材的重要途径.基于对云南省景谷县思茅松天然林的样地调查数据,采用种群径级结构代替年龄结构、Ripley的L函数点格局等方法,对思茅松天然林在不同强度择伐10a后的种群结构和空间分布格局及其不同发育阶段在不同尺度下的相互关系进行了系统研究和分析.结果表明:择伐10a后3种类型的思茅松自然种群的胸径、树高之间呈显著的正相关且可用幂指数模拟预测模型.未择伐的思茅松种群呈衰退型,轻度择伐(12.9%)后种群呈稳定型,中度择伐(29.3%)后的种群属增长型,适当的中度择伐有利于自然种群的更新;未择伐与中度择伐后的思茅松种群分布格局整体呈随机型,但后者较前者更趋向于均匀分布,轻度择伐后的思茅松种群分布格局随着尺度的增加由随机转向聚集分布.择伐对不同径级思茅松天然林种群空间分布格局有重要影响,未择伐的思茅松自然种群随着发育阶段的发展,多数尺度上由随机分布逐渐转变为聚集分布,至大树则又以随机分布为主;轻度择伐和中度择伐的思茅松自然种群则随着发育阶段的发展而由完全的随机分布逐渐转变为聚集分布,中度择伐的聚集分布则更为明显.思茅松不同发育阶段间在所有尺度上基本都呈显著正相关,表明思茅松种群种内竞争弱,有利于种群的维持.
DOI:10.3321/j.issn:1001-7488.2007.12.004URLMagsci [本文引用: 3]
<p>应用“相邻格子法”对不同生境下的香果树种群进行每木调查,获得野外资料。采用空间代替时间的方法,研究香果树的径级结构和分布格局,以及应用泊松方差/均值比、负二项式分布、Lloyd平均拥挤度指数和聚块指数4个指标分析其种群格局状况。结果表明:不同生境下的香果树径级结构具有较大的相似性——均缺乏幼苗和幼树,种群呈衰退趋势;不同生境下的香果树种群聚块规模为8~32m<sup>2</sup>;种群的空间分布格局主要为集群分布;格局动态明显呈现由集群分布到随机分布的扩散趋势。建议采取相应的措施保护母树,尤其是20年生以上的母树;还应加强对种子及幼苗生长动态变化的研究。</p>
DOI:10.3321/j.issn:1001-7488.2007.12.004URLMagsci [本文引用: 3]
<p>应用“相邻格子法”对不同生境下的香果树种群进行每木调查,获得野外资料。采用空间代替时间的方法,研究香果树的径级结构和分布格局,以及应用泊松方差/均值比、负二项式分布、Lloyd平均拥挤度指数和聚块指数4个指标分析其种群格局状况。结果表明:不同生境下的香果树径级结构具有较大的相似性——均缺乏幼苗和幼树,种群呈衰退趋势;不同生境下的香果树种群聚块规模为8~32m<sup>2</sup>;种群的空间分布格局主要为集群分布;格局动态明显呈现由集群分布到随机分布的扩散趋势。建议采取相应的措施保护母树,尤其是20年生以上的母树;还应加强对种子及幼苗生长动态变化的研究。</p>
[本文引用: 1]
[本文引用: 1]
DOI:10.3969/j.issn.1001-9235.2003.02.001URL [本文引用: 1]
对弯道水流、泥沙运动特性以及河弯演变规律在国内外的研究进展进行了概述 ,主要包括水面横比降、横向环流、纵向垂线平均流速、输沙特性以及最大冲深等内容。了解这些基本理论的研究现状 ,对江河治理、航运等方面的科学研究与设计有重要意义
DOI:10.3969/j.issn.1001-9235.2003.02.001URL [本文引用: 1]
对弯道水流、泥沙运动特性以及河弯演变规律在国内外的研究进展进行了概述 ,主要包括水面横比降、横向环流、纵向垂线平均流速、输沙特性以及最大冲深等内容。了解这些基本理论的研究现状 ,对江河治理、航运等方面的科学研究与设计有重要意义
DOI:10.1890/09-1533URL [本文引用: 1]
The spatial distribution and temporal availability of propagules fundamentally constrain plant community development. This study experimentally tested several hypotheses about the relative roles of wind and water dispersal in colonization and development of riparian communities along rivers. Through controlling the source of propagules (dispersed by wind, water, or both) reaching newly created, bare river margin sites, we isolated the relative roles of dispersal and other factors in plant community development over five years. Replicated treatments were established at 12 sites spanning 400 km along two adjacent rivers in northern Sweden, one fragmented by a series of dams, the other free-flowing. Bare river margins receiving only water-dispersed propagules had significantly higher species richness compared to plots receiving only wind-dispersed propagules during the initial two years of colonization. Species richness increased annually throughout the study along tranquil and turbulent reaches of the free-flowing river but reached an asymptote at comparatively low richness after a single year on the impounded river. Propagule source strongly influenced species richness during the initial establishment along both rivers, with richness being significantly higher in plots receiving water-dispersed seeds. This strong treatment effect continued to be important through time along the regulated river but diminished in importance along the free-flowing river where other factors such as soil moisture, light availability, and exposure of sites to fluvial disturbance overshadowed the influence of dispersal pathway in mediating species richness. This suggests that hydrochory (plant dispersal by water) may be more important for maintenance of diversity in regulated systems where long-distance dispersal is absent or negligible, but that the rich local propagule source along free-flowing rivers supports high species richness. The number of unique species was higher in water-dispersal plots along both the regulated and free-flowing rivers. This result suggests that hydrochory may contribute to temporal variability of sites, may enhance richness over time, and may have an important role in meta-population and meta-community dynamics of plant communities through long-distance (and local) dispersal and chance colonization. Our findings provide experimental evidence that water dispersal of plant propagules influences colonization dynamics and is important for long-term community development in riparian zones.
DOI:10.1111/j.1365-2427.2009.02206.xURL [本文引用: 1]
1. Riparian vegetation composition, structure and abundance are governed to a large degree by river flow regime and flow-mediated fluvial processes. Streamflow regime exerts selective pressures on riparian vegetation, resulting in adaptations (trait syndromes) to specific flow attributes. Widespread modification of flow regimes by humans has resulted in extensive alteration of riparian vegetation communities. Some of the negative effects of altered flow regimes on vegetation may be reversed by restoring components of the natural flow regime.2. Models have been developed that quantitatively relate components of the flow regime to attributes of riparian vegetation at the individual, population and community levels. Predictive models range from simple statistical relationships, to more complex stochastic matrix population models and dynamic simulation models. Of the dozens of predictive models reviewed here, most treat one or a few species, have many simplifying assumptions such as stable channel form, and do not specify the time-scale of response. In many cases, these models are very effective in developing alternative streamflow management plans for specific river reaches or segments but are not directly transferable to other rivers or other regions.3. A primary goal in riparian ecology is to develop general frameworks for prediction of vegetation response to changing environmental conditions. The development of riparian vegetation-flow response guilds offers a framework for transferring information from rivers where flow standards have been developed to maintain desirable vegetation attributes, to rivers with little or no existing information.4. We propose to organise riparian plants into non-phylogenetic groupings of species with shared traits that are related to components of hydrologic regime: life history, reproductive strategy, morphology, adaptations to fluvial disturbance and adaptations to water availability. Plants from any river or region may be grouped into these guilds and related to hydrologic attributes of a specific class of river using probabilistic response curves.5. Probabilistic models based on riparian response guilds enable prediction of the likelihood of change in each of the response guilds given projected changes in flow, and facilitate examination of trade-offs and risks associated with various flow management strategies. Riparian response guilds can be decomposed to the species level for individual projects or used to develop flow management guidelines for regional water management plans.
DOI:10.1029/1999WR900346URL [本文引用: 1]
Numerous studies have suggested the importance of bank vegetation as a control of channel patterns; however, to date there is no conclusive evidence that vegetation does represent a significant control. An analytical model is developed in order to assess the influence of bank vegetation on channel patterns of alluvial gravel-bed rivers. Three channel types are considered: meandering, wandering, and braided. Bank vegetation effects are quantified in terms of a friction angle '. A new theoretical meandering-braiding transition criterion is formulated that includes ', median grain diameter D, and bank-full discharge Q. The theoretical relation is tested against field data from 137 rivers and successfully discriminates between meandering and braided rivers. Wandering rivers show greater scatter. It is concluded that bank vegetation, as expressed in terms of ', does exert significant and quantifiable control on alluvial channel patterns. A simple planform stability diagram is developed to determine the sensitivity of gravel-bed rivers to changes in bank vegetation.
DOI:10.1146/annurev.ecolsys.28.1.621URL [本文引用: 1]
Riparian zones possess an unusually diverse array of species and environmental processes. The ecological diversity is related to variable flood regimes, geographically unique channel processes, altitudinal climate shifts, and upland influences on the fluvial corridor. The resulting dynamic environment supports a variety of life-history strategies, biogeochemical cycles and rates, and organisms adapted to disturbance regimes over broad spatial and temporal scales. Innovations in riparian zone management have been effective in ameliorating many ecological issues related to land use and environmental quality. Riparian zones play essential roles in water and landscape planning, in restoration of aquatic systems, and in catalyzing institutional and societal cooperation for these efforts.
DOI:10.1002/rra.881URL [本文引用: 1]
Traditional policies for managing river bank erosion are currently being reconsidered as a result of increased awareness regarding the unsustainable nature of some forms of bank protection, and the role played by bank erosion in providing ecosystem services and supporting geomorphological functions. River managers are therefore increasingly seeking to preserve bank erosion within a defined erodible corridor. This paper provides an overview of the erodible corridor concept, focusing on the provision of guidelines for applying the concept in practice. We argue that a nested approach is required to address management objectives across a range of scales (network scale, reach scale, local scale) and review the different geomorphic tools that are available to help managers define the extent and inner sensitivity of the erodible corridor. These tools include simple rules of thumb such as evaluation of the equilibrium meander amplitude, historical approaches based on overlays of historical channel position, and simulation modelling. The advantages and limitations of each of these tools are discussed. Copyright 2005 John Wiley & Sons, Ltd.
DOI:10.1111/j.1366-9516.2006.00314.xURL [本文引用: 1]
DOI:10.3969/j.issn.1000-2561.2010.04.009URL [本文引用: 1]
Based on the field investigation,9 species and 1 variety of Rhododendron species,classified to 4 subgenera of Rhododendron,Humenanthes,Tsutsusi and Rhodorustrum, were determined in Hainan. The morphological characters of each genus were described and a identification key to the species and variety was recommended. The landscape application and prospect of Rhododendron were also discussed.
DOI:10.3969/j.issn.1000-2561.2010.04.009URL [本文引用: 1]
Based on the field investigation,9 species and 1 variety of Rhododendron species,classified to 4 subgenera of Rhododendron,Humenanthes,Tsutsusi and Rhodorustrum, were determined in Hainan. The morphological characters of each genus were described and a identification key to the species and variety was recommended. The landscape application and prospect of Rhododendron were also discussed.
DOI:10.1002/rra.2910URL [本文引用: 2]
Riparian vegetation actively interacts with fluvial systems affecting river hydrodynamics, morphodynamics and groundwater. These interactions can be coupled because both vegetation and hydromorphology (i.e. the combined scientific study of hydrology and fluvial geomorphology) involve dynamic processes with similar temporal and spatial scales. To predict and assess the consequences of restoration measures, maintenance operations or human pressures in rivers, managers and planners may wish to model these interactions considering the different and interdisciplinary implications in the fields of ecology, geomorphology and hydrology. In this paper, we review models that are currently available and that incorporate the processes that relate riparian vegetation to hydromorphology. The models that are considered include those emphasizing hydraulic-geomorphological processes (such as flow resistance, sediment transport and bank dynamics) as well as those emphasizing ecological processes (seed dispersal, plant survival, growth, succession and mortality). Models interpreting the coupled evolution between riparian vegetation and river morphology and groundwater are also presented. The aim is to provide an overview of current modelling capabilities and limitations and to identify future modelling challenges. Copyright 2015 John Wiley & Sons, Ltd.
DOI:10.1002/esp.1908URL [本文引用: 1]
A series of laboratory experiments demonstrates that riparian vegetation can cause a braided channel to self-organize to, and maintain, a dynamic, single-thread channel. The initial condition for the experiments was steady-state braiding in non-cohesive sand under uniform discharge. From here, an experiment consisted of repeated cycles alternating a short duration high flow with a long duration low flow, and uniform dispersal of alfalfa seeds over the bed at the end of each high flow. Plants established on freshly deposited bars and areas of braidplain that were unoccupied during low flow. The presence of the plants had the effect of progressively focusing the high flow so that a single dominant channel developed. The single-thread channel self-adjusted to carry the high flow. Vegetation also slowed the rate of bank erosion. Matching of deposition along the point bar with erosion along the outer bend enabled the channel to develop sinuosity and migrate laterally while suppressing channel splitting and the creation of new channel width. The experimental channels spontaneously reproduced many of the mechanisms by which natural meandering channels migrate and maintain a single dominant channel, in particular bend growth and channel cutoff. In contrast with the braided system, where channel switching is a nearly continuous process, vegetation maintained a coherent channel until wholesale diversion of flow via cutoff and/or avulsion occurred, by which point the previous channel tended to be highly unfavorable for flow. Thus vegetation discouraged the coexistence of multiple channels. Varying discharge was key to allowing expression of feedbacks between the plants and the flow and promoting the transition from braiding to a single-thread channel that was then dynamically maintained. Copyright 2010 John Wiley & Sons, Ltd.
[本文引用: 1]
[本文引用: 1]
DOI:10.3773/j.issn.1005-264x.2008.04.011URL [本文引用: 1]
东亚特有珍稀濒危植物领春木(Euptelea pleiospermum)在神农架地区集中分布于河岸带中。为初步探讨领春木在河岸带集中分布的成因和维持机制,在神农架地区4条河流——沿渡河、香溪河、南河和堵河的河岸带,共设置T30个样方(20m×30m),从种群的大小级结构、静态生命表、存活曲线、空间分布格局及其动态5个方面分析了神农架地区河岸带中领春木种群的数量特征与空间分布格局。结果表明:1)幼龄期个体缺乏,中龄期个体相对丰富,老龄期个体数量稀少。纺锤型的大小级结构表明种群属于衰退型,但是以萌蘖为主要更新方式使其种群在较长一段时间内得以维持;2)现存的Ⅰ、Ⅱ级个体数少,导致静态生命表中Ⅰ、Ⅱ级个体死亡率出现负值,自疏现象造成第Ⅳ级个体出现死亡率高峰,由于接近实际寿命,在第Ⅶ级死亡率达到最高;3)虽然其幼苗存活率较低,但由于其幼树的存活率较高,其存活曲线接近DeeveyⅠ型,表明该地区河岸带的环境条件较适宜领春木种群的生长;4)空间分布格局总体为聚集分布,这与大多数珍稀植物种群一致。人为干扰和自然环境影响其分布格局,使种群由聚集分布向随机分布发展。因此使领春木种群在河岸带中沿海拔呈现“一带多岛”现象;5)从幼龄期到中龄期再到老龄期,不同发育阶段的领春木种群的分布格局由聚集分布逐渐变为随机分布。但由于分布格局受种群本身生物学特性、自然环境因素(如坡向和海拔)和人为干扰的影响,领春木在不同河岸带中的分布格局有所差异。
DOI:10.3773/j.issn.1005-264x.2008.04.011URL [本文引用: 1]
东亚特有珍稀濒危植物领春木(Euptelea pleiospermum)在神农架地区集中分布于河岸带中。为初步探讨领春木在河岸带集中分布的成因和维持机制,在神农架地区4条河流——沿渡河、香溪河、南河和堵河的河岸带,共设置T30个样方(20m×30m),从种群的大小级结构、静态生命表、存活曲线、空间分布格局及其动态5个方面分析了神农架地区河岸带中领春木种群的数量特征与空间分布格局。结果表明:1)幼龄期个体缺乏,中龄期个体相对丰富,老龄期个体数量稀少。纺锤型的大小级结构表明种群属于衰退型,但是以萌蘖为主要更新方式使其种群在较长一段时间内得以维持;2)现存的Ⅰ、Ⅱ级个体数少,导致静态生命表中Ⅰ、Ⅱ级个体死亡率出现负值,自疏现象造成第Ⅳ级个体出现死亡率高峰,由于接近实际寿命,在第Ⅶ级死亡率达到最高;3)虽然其幼苗存活率较低,但由于其幼树的存活率较高,其存活曲线接近DeeveyⅠ型,表明该地区河岸带的环境条件较适宜领春木种群的生长;4)空间分布格局总体为聚集分布,这与大多数珍稀植物种群一致。人为干扰和自然环境影响其分布格局,使种群由聚集分布向随机分布发展。因此使领春木种群在河岸带中沿海拔呈现“一带多岛”现象;5)从幼龄期到中龄期再到老龄期,不同发育阶段的领春木种群的分布格局由聚集分布逐渐变为随机分布。但由于分布格局受种群本身生物学特性、自然环境因素(如坡向和海拔)和人为干扰的影响,领春木在不同河岸带中的分布格局有所差异。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.13287/j.1001-9332.201602.021URL [本文引用: 2]
以叶尔羌河下游阿瓦提县天然胡杨和灰叶胡杨混交林、塔里木河上游农一师16团天然灰叶胡杨林和塔里木河中游轮台县天然胡杨林为对象,研究胡杨和灰叶胡杨克隆分株种群的空间分布格局.结果表明:8种不同取样尺度(5 m×5 m、5 m×10 m、5 m×15 m、10 m× 10m、10 m×15 m、15 m×15 m、15 m×20m、20 m×20 m)下,3个研究区2物种克隆分株种群的空间分布格局均表现为集群分布,表明集群分布是研究区胡杨和灰叶胡杨克隆分株种群空间分布格局的基本属性;在5 m×5 m尺度上,3个研究区2物种克隆分株种群均表现出负二项参数最小、Cassie指标和聚块性指标最大、聚集强度最大的分布特征.
DOI:10.13287/j.1001-9332.201602.021URL [本文引用: 2]
以叶尔羌河下游阿瓦提县天然胡杨和灰叶胡杨混交林、塔里木河上游农一师16团天然灰叶胡杨林和塔里木河中游轮台县天然胡杨林为对象,研究胡杨和灰叶胡杨克隆分株种群的空间分布格局.结果表明:8种不同取样尺度(5 m×5 m、5 m×10 m、5 m×15 m、10 m× 10m、10 m×15 m、15 m×15 m、15 m×20m、20 m×20 m)下,3个研究区2物种克隆分株种群的空间分布格局均表现为集群分布,表明集群分布是研究区胡杨和灰叶胡杨克隆分株种群空间分布格局的基本属性;在5 m×5 m尺度上,3个研究区2物种克隆分株种群均表现出负二项参数最小、Cassie指标和聚块性指标最大、聚集强度最大的分布特征.
[本文引用: 1]
[本文引用: 1]
Seedling establishment in a dynamic sedimentary environment: A conceptual framework using mangroves
2
2013
... 溪流弯道不同区域的海南杜鹃群体基本都是幼苗数量远多于龄级较大的个体, 这可能与洪水对海南杜鹃繁殖体的搬运作用有关.一般而言, 沙质土壤不利于植物根系深扎, 幼苗难以在暴雨和山洪中生存(
... 汛期山洪对河岸的过度侵蚀或横向迁移可能会破坏现有的生态系统(
Hydrological and geomorphic impacts on riparian plant communities
1
2000
... 在河道形态变化过程中, 河岸带植物和河岸之间有相互依存的关系(
Interactions between river flows and colonizing vegetation on a braided river: Exploring spatial and temporal dynamics in riparian vegetation cover using satellite data
2
2011
... 在河道形态变化过程中, 河岸带植物和河岸之间有相互依存的关系(
... 弯道内外两岸地形地貌与基质的差异主要是由于河道水流造成的(
Bank-attached vanes for bank erosion control and restoration of river meanders
1
2010
... 汛期山洪对河岸的过度侵蚀或横向迁移可能会破坏现有的生态系统(
River corridor plants (Stromtalpflanzen) in Central European lowland: A review of a poorly understood plant distribution pattern
1
2001
... 植物的繁殖体也更容易沉积和聚集在流速较慢、坡度较缓的弯道凸岸(
What do we still need to know about the ecohydrology of riparian zones?
1
2010
... 河岸带作为河流生态系统和陆地生态系统的过渡带, 拥有极高的空间异质性和复杂的生态过程, 孕育了丰富的物种多样性(
Modeling the interaction between river morphodynamics and riparian vegetation
1
2013
... 在河道形态变化过程中, 河岸带植物和河岸之间有相互依存的关系(
Drowned, buried and carried away: Effects of plant traits on the distribution of native and alien species in riparian ecosystems
1
2014
... 汛期山洪对河岸的过度侵蚀或横向迁移可能会破坏现有的生态系统(
Reciprocal interactions and adjustments between fluvial landforms and vegetation dynamics in river corridors: A review of complementary approaches
2
2007
... 河岸带作为河流生态系统和陆地生态系统的过渡带, 拥有极高的空间异质性和复杂的生态过程, 孕育了丰富的物种多样性(
... 在河道形态变化过程中, 河岸带植物和河岸之间有相互依存的关系(
1
2008
... 在前期踏查的基础上, 于2017年8月在3个地区分别设置具有代表性, 未受人类破坏的溪流弯道样带, 调查溪流弯道的理化性状, 记录样带经纬度、海拔、河宽、溪流弯曲系数、弯顶位置等信息.以顺溪流方向为样带X轴, 距溪流河岸线的垂直距离为Y轴(
Vegetative impacts on hydraulics and sediment processes across the fluvial system
2
2013
... 河岸带作为河流生态系统和陆地生态系统的过渡带, 拥有极高的空间异质性和复杂的生态过程, 孕育了丰富的物种多样性(
... 在河道形态变化过程中, 河岸带植物和河岸之间有相互依存的关系(
浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型
2
2004
... 海南杜鹃属于常绿灌木, 分枝多, 有基部主干, 采用基径作为个体大小的指标研究种群结构(
... 溪流弯道不同区域的海南杜鹃群体基本都是幼苗数量远多于龄级较大的个体, 这可能与洪水对海南杜鹃繁殖体的搬运作用有关.一般而言, 沙质土壤不利于植物根系深扎, 幼苗难以在暴雨和山洪中生存(
浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型
2
2004
... 海南杜鹃属于常绿灌木, 分枝多, 有基部主干, 采用基径作为个体大小的指标研究种群结构(
... 溪流弯道不同区域的海南杜鹃群体基本都是幼苗数量远多于龄级较大的个体, 这可能与洪水对海南杜鹃繁殖体的搬运作用有关.一般而言, 沙质土壤不利于植物根系深扎, 幼苗难以在暴雨和山洪中生存(
Plant distribution and dispersal mechanisms at the Hassayampa River Preserve, Arizona, USA
1
2001
... 海南杜鹃的花期正处海南岛的旱季, 果实与种子成熟时间是雨季和台风季节.细小的种子在风、雨、流水的作用下可以实现长距离的扩散, 特别是台风导致的溪流山洪成了海南杜鹃种子高效而快速的传播渠道.这可能是河岸带植物普遍的一种种子扩散适应对策(
1
2005
... 海南杜鹃(Rhododendron hainanense)是中国特有植物, 株高1-3 m, 花顶生、红色, 花期10月至翌年3月, 是杜鹃花属花期最晚、自然花期最长的一个物种(
Riparian plant community responses to increased flooding: A meta-analysis
1
2015
... 汛期山洪对河岸的过度侵蚀或横向迁移可能会破坏现有的生态系统(
The importance of seasonal flow timing for riparian vegetation dynamics: A systematic review using causal criteria analysis
1
2011
... 海南杜鹃的花期正处海南岛的旱季, 果实与种子成熟时间是雨季和台风季节.细小的种子在风、雨、流水的作用下可以实现长距离的扩散, 特别是台风导致的溪流山洪成了海南杜鹃种子高效而快速的传播渠道.这可能是河岸带植物普遍的一种种子扩散适应对策(
Changing river channels: The roles of hydrological processes, plants and pioneer fluvial landforms in humid temperate, mixed load, gravel bed rivers
1
2012
... 弯道内外两岸地形地貌与基质的差异主要是由于河道水流造成的(
河岸带生态学研究进展与展望
1
2013
... 在河道形态变化过程中, 河岸带植物和河岸之间有相互依存的关系(
河岸带生态学研究进展与展望
1
2013
... 在河道形态变化过程中, 河岸带植物和河岸之间有相互依存的关系(
神农架山地河岸带连香树的种群结构与动态
1
2009
... 汛期山洪对河岸的过度侵蚀或横向迁移可能会破坏现有的生态系统(
神农架山地河岸带连香树的种群结构与动态
1
2009
... 汛期山洪对河岸的过度侵蚀或横向迁移可能会破坏现有的生态系统(
磴口县沙冬青种群结构和空间分布格局的研究
1
2006
... 本研究采用可变尺度相邻格子法, 采用方差/平均值法在1 m× 1 m、2 m× 2 m、5 m × 5 m 3个尺度下(
磴口县沙冬青种群结构和空间分布格局的研究
1
2006
... 本研究采用可变尺度相邻格子法, 采用方差/平均值法在1 m× 1 m、2 m× 2 m、5 m × 5 m 3个尺度下(
Retention of particulate matter by macrophytes in a first-order stream
1
2004
... 弯道内外两岸地形地貌与基质的差异主要是由于河道水流造成的(
择伐对思茅松自然种群结构和空间分布格局的影响
1
2015
... 海南杜鹃种子在沉积后对凸岸的种群进行更新和补充, 导致凸岸弯顶偏下的海南杜鹃分布密集、呈稳定增长型, 成为该区域灌木层优势种.调查发现, 海南杜鹃在近弯顶位置易形成稳定的以幼苗和小树为主的增长型群体, 距离河岸或弯顶越远, 各龄级个体数量越呈下降趋势, 说明海南杜鹃生长与种子萌发需要较多的水分和光照条件.由于河岸带种内和种间竞争大, 而且年龄越大的个体要承受越多的洪水冲刷(
择伐对思茅松自然种群结构和空间分布格局的影响
1
2015
... 海南杜鹃种子在沉积后对凸岸的种群进行更新和补充, 导致凸岸弯顶偏下的海南杜鹃分布密集、呈稳定增长型, 成为该区域灌木层优势种.调查发现, 海南杜鹃在近弯顶位置易形成稳定的以幼苗和小树为主的增长型群体, 距离河岸或弯顶越远, 各龄级个体数量越呈下降趋势, 说明海南杜鹃生长与种子萌发需要较多的水分和光照条件.由于河岸带种内和种间竞争大, 而且年龄越大的个体要承受越多的洪水冲刷(
不同生境香果树种群的径级结构与分布格局
3
2007
... 对样带内所有海南杜鹃进行调查统计, 调查并记录样带内所有海南杜鹃的基径、冠幅、株高等指标, 并以平行河流方向为横坐标(x)、垂直于河流平行方向为纵坐标(y), 测定每株海南杜鹃的坐标(x, y).如果一株海南杜鹃上有多个无性系分株, 则以其中基径最大的一株进行记录(
... 本研究采用可变尺度相邻格子法, 采用方差/平均值法在1 m× 1 m、2 m× 2 m、5 m × 5 m 3个尺度下(
... 海南杜鹃种子在沉积后对凸岸的种群进行更新和补充, 导致凸岸弯顶偏下的海南杜鹃分布密集、呈稳定增长型, 成为该区域灌木层优势种.调查发现, 海南杜鹃在近弯顶位置易形成稳定的以幼苗和小树为主的增长型群体, 距离河岸或弯顶越远, 各龄级个体数量越呈下降趋势, 说明海南杜鹃生长与种子萌发需要较多的水分和光照条件.由于河岸带种内和种间竞争大, 而且年龄越大的个体要承受越多的洪水冲刷(
不同生境香果树种群的径级结构与分布格局
3
2007
... 对样带内所有海南杜鹃进行调查统计, 调查并记录样带内所有海南杜鹃的基径、冠幅、株高等指标, 并以平行河流方向为横坐标(x)、垂直于河流平行方向为纵坐标(y), 测定每株海南杜鹃的坐标(x, y).如果一株海南杜鹃上有多个无性系分株, 则以其中基径最大的一株进行记录(
... 本研究采用可变尺度相邻格子法, 采用方差/平均值法在1 m× 1 m、2 m× 2 m、5 m × 5 m 3个尺度下(
... 海南杜鹃种子在沉积后对凸岸的种群进行更新和补充, 导致凸岸弯顶偏下的海南杜鹃分布密集、呈稳定增长型, 成为该区域灌木层优势种.调查发现, 海南杜鹃在近弯顶位置易形成稳定的以幼苗和小树为主的增长型群体, 距离河岸或弯顶越远, 各龄级个体数量越呈下降趋势, 说明海南杜鹃生长与种子萌发需要较多的水分和光照条件.由于河岸带种内和种间竞争大, 而且年龄越大的个体要承受越多的洪水冲刷(
广东鹅凰嶂自然保护区杜鹃红山茶种群结构与空间分布格局
1
2013
... 海南杜鹃属于常绿灌木, 分枝多, 有基部主干, 采用基径作为个体大小的指标研究种群结构(
广东鹅凰嶂自然保护区杜鹃红山茶种群结构与空间分布格局
1
2013
... 海南杜鹃属于常绿灌木, 分枝多, 有基部主干, 采用基径作为个体大小的指标研究种群结构(
弯曲型河流基本特性研究进展
1
2003
... 弯道内外两岸地形地貌与基质的差异主要是由于河道水流造成的(
弯曲型河流基本特性研究进展
1
2003
... 弯道内外两岸地形地貌与基质的差异主要是由于河道水流造成的(
Consequences of propagule dispersal and river fragmentation for riparian plant community diversity and turnover
1
2010
... 汛期山洪对河岸的过度侵蚀或横向迁移可能会破坏现有的生态系统(
Theory, methods and tools for determining environmental flows for riparian vegetation: Riparian vegetation- flow response guilds
1
2010
... 植物的繁殖体也更容易沉积和聚集在流速较慢、坡度较缓的弯道凸岸(
Influence of bank vegetation on alluvial channel patterns
1
2000
... 河岸带作为河流生态系统和陆地生态系统的过渡带, 拥有极高的空间异质性和复杂的生态过程, 孕育了丰富的物种多样性(
The ecology of interfaces: Riparian zones
1
1997
... 河岸带作为河流生态系统和陆地生态系统的过渡带, 拥有极高的空间异质性和复杂的生态过程, 孕育了丰富的物种多样性(
A review of techniques available for delimiting the erodible river corridor: A sustainable approach to managing bank erosion
1
2005
... 汛期山洪对河岸的过度侵蚀或横向迁移可能会破坏现有的生态系统(
Riparian vegetation: Degradation, alien plant invasions, and restoration prospects
1
2007
... 海南杜鹃的花期正处海南岛的旱季, 果实与种子成熟时间是雨季和台风季节.细小的种子在风、雨、流水的作用下可以实现长距离的扩散, 特别是台风导致的溪流山洪成了海南杜鹃种子高效而快速的传播渠道.这可能是河岸带植物普遍的一种种子扩散适应对策(
海南野生杜鹃花属植物种质资源调查研究
1
2010
... 海南杜鹃(Rhododendron hainanense)是中国特有植物, 株高1-3 m, 花顶生、红色, 花期10月至翌年3月, 是杜鹃花属花期最晚、自然花期最长的一个物种(
海南野生杜鹃花属植物种质资源调查研究
1
2010
... 海南杜鹃(Rhododendron hainanense)是中国特有植物, 株高1-3 m, 花顶生、红色, 花期10月至翌年3月, 是杜鹃花属花期最晚、自然花期最长的一个物种(
Advances on modelling riparian vegetation—Hydromorphology interactions
2
2016
... 河岸带的河道地貌变化最典型的表现之一, 就是形成河流弯道.天然河流的弯道一般占河流总长的80%以上.河水流经弯道, 受离心力作用, 表层水流偏向凹岸(河道外侧), 底部水流由凹岸流向凸岸(河道内侧), 造成河流凹岸水流快、易侵蚀; 凸岸水流慢、易堆积泥沙等显著差异.这些差异, 有可能导致凹岸和凸岸两岸的水土性质、植物繁殖体来源与聚集程度不同, 从而导致两岸的植被与植物生长出现差异(
... 海南杜鹃的花期正处海南岛的旱季, 果实与种子成熟时间是雨季和台风季节.细小的种子在风、雨、流水的作用下可以实现长距离的扩散, 特别是台风导致的溪流山洪成了海南杜鹃种子高效而快速的传播渠道.这可能是河岸带植物普遍的一种种子扩散适应对策(
Effects of vegetation on channel morphodynamics: Results and insights from laboratory experiments
1
2010
... 弯道内外两岸地形地貌与基质的差异主要是由于河道水流造成的(
1
2017
... 河岸带的河道地貌变化最典型的表现之一, 就是形成河流弯道.天然河流的弯道一般占河流总长的80%以上.河水流经弯道, 受离心力作用, 表层水流偏向凹岸(河道外侧), 底部水流由凹岸流向凸岸(河道内侧), 造成河流凹岸水流快、易侵蚀; 凸岸水流慢、易堆积泥沙等显著差异.这些差异, 有可能导致凹岸和凸岸两岸的水土性质、植物繁殖体来源与聚集程度不同, 从而导致两岸的植被与植物生长出现差异(
1
2017
... 河岸带的河道地貌变化最典型的表现之一, 就是形成河流弯道.天然河流的弯道一般占河流总长的80%以上.河水流经弯道, 受离心力作用, 表层水流偏向凹岸(河道外侧), 底部水流由凹岸流向凸岸(河道内侧), 造成河流凹岸水流快、易侵蚀; 凸岸水流慢、易堆积泥沙等显著差异.这些差异, 有可能导致凹岸和凸岸两岸的水土性质、植物繁殖体来源与聚集程度不同, 从而导致两岸的植被与植物生长出现差异(
神农架地区河岸带中领春木种群数量特征与空间分布格局
1
2008
... 溪流弯道不同区域的海南杜鹃群体基本都是幼苗数量远多于龄级较大的个体, 这可能与洪水对海南杜鹃繁殖体的搬运作用有关.一般而言, 沙质土壤不利于植物根系深扎, 幼苗难以在暴雨和山洪中生存(
神农架地区河岸带中领春木种群数量特征与空间分布格局
1
2008
... 溪流弯道不同区域的海南杜鹃群体基本都是幼苗数量远多于龄级较大的个体, 这可能与洪水对海南杜鹃繁殖体的搬运作用有关.一般而言, 沙质土壤不利于植物根系深扎, 幼苗难以在暴雨和山洪中生存(
内蒙古灌丛化草原毛刺锦鸡儿种群结构和空间分布格局
1
2017
... 海南杜鹃属于常绿灌木, 分枝多, 有基部主干, 采用基径作为个体大小的指标研究种群结构(
内蒙古灌丛化草原毛刺锦鸡儿种群结构和空间分布格局
1
2017
... 海南杜鹃属于常绿灌木, 分枝多, 有基部主干, 采用基径作为个体大小的指标研究种群结构(
太白山太白红杉种群空间分布格局研究
1
2005
... 本研究采用可变尺度相邻格子法, 采用方差/平均值法在1 m× 1 m、2 m× 2 m、5 m × 5 m 3个尺度下(
太白山太白红杉种群空间分布格局研究
1
2005
... 本研究采用可变尺度相邻格子法, 采用方差/平均值法在1 m× 1 m、2 m× 2 m、5 m × 5 m 3个尺度下(
塔里木河流域胡杨和灰叶胡杨克隆分株空间分布格局
2
2016
... 本研究采用可变尺度相邻格子法, 采用方差/平均值法在1 m× 1 m、2 m× 2 m、5 m × 5 m 3个尺度下(
... ;
塔里木河流域胡杨和灰叶胡杨克隆分株空间分布格局
2
2016
... 本研究采用可变尺度相邻格子法, 采用方差/平均值法在1 m× 1 m、2 m× 2 m、5 m × 5 m 3个尺度下(
... ;
不同生境条件下四合木(Tetraena mongolica Maxim.)种群结构特征
1
2008
... 海南杜鹃属于常绿灌木, 分枝多, 有基部主干, 采用基径作为个体大小的指标研究种群结构(
不同生境条件下四合木(Tetraena mongolica Maxim.)种群结构特征
1
2008
... 海南杜鹃属于常绿灌木, 分枝多, 有基部主干, 采用基径作为个体大小的指标研究种群结构(
Copyright © 2021 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}