 ,
, ,1,*, 冉飞1, 杨燕1, 张莉1,2, 肖瑶1,2
,1,*, 冉飞1, 杨燕1, 张莉1,2, 肖瑶1,2Flowering phenology and growth of typical shrub grass plants in response to simulated warmer and drier climate in early succession Taiga forests in the Da Hinggan Ling of northeast China
SONG Xiao-Yan1,2, WANG Gen-Xu,,1,*, RAN Fei1, YANG Yan1, ZHANG Li1,2, XIAO Yao1,2通讯作者: wanggx@imde.ac.cn
编委: 罗天祥
责任编辑: 李敏
收稿日期:2018-02-13修回日期:2018-05-09网络出版日期:2018-05-20
| 基金资助: |
Received:2018-02-13Revised:2018-05-09Online:2018-05-20
| Fund supported: |

摘要
为了揭示暖干化气候变化趋势对演替初期泰加林灌草层植物开花物候和生长的影响, 以东北大兴安岭演替初期泰加林灌草层(高度<50 cm)植物为研究对象, 根据植物生活史, 选择典型的早花植物圆锥薹草(Carex diandra)和水葡萄茶藨子(Ribes procumbens), 中花植物杜香(Ledum palustre)及晚花植物齿叶风毛菊(Saussurea neoserrata), 监测模拟增温和排水处理2年后其完整的开花物候及生长状况, 以期为认识气候变化下泰加林火后植被恢复演替提供依据。结果显示: 1)模拟增温和排水处理后, 早花植物开花温度限制的提早解除使其花期提前, 而水分限制使得晚花植物花期呈延后趋势, 这可能为中花植物提供更大的生态位空隙, 使其开花数量趋向于增加, 花期提前且呈延长趋势; 2)模拟增温和排水处理后, 圆锥薹草、杜香和齿叶风毛菊的盖度和频度均呈增加的趋势, 以杜香最显著, 而水葡萄茶藨子的盖度和频度均下降; 3)模拟增温与排水处理对植物开花物候的影响无显著交互作用, 但土壤水分降低影响部分植物生长对增温的响应。研究结果表明, 植物开花物候对暖干化气候的响应表现出明显的种间差异, 因植物开花功能群而异。早、中花植物花期趋于提前, 晚花期植物花期趋于延后, 一方面可能导致群落生态位变化, 对种间竞争产生潜在影响, 引发群落组成和结构改变; 另一方面可能成为群落调节其整体物候, 以适应气候变化的重要途径。
关键词:
Abstract
Keywords:
PDF (1057KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
宋小艳, 王根绪, 冉飞, 杨燕, 张莉, 肖瑶. 东北大兴安岭演替初期泰加林灌草层典型植物开花物候与生长对模拟暖干化气候的响应. 植物生态学报[J], 2018, 42(5): 539-549 DOI:10.17521/cjpe.2018.0042
SONG Xiao-Yan, WANG Gen-Xu, RAN Fei, YANG Yan, ZHANG Li, XIAO Yao.
气候变暖已成为不争的事实, 且在高纬度冻土区表现尤为明显(IPCC, 2013)。作为高纬度冻土区重要植被类型的泰加林, 其面积占全球森林面积的35%, 其土壤碳储量占到全球陆地生态系统有机碳储量的1/3, 在全球碳平衡中起到重要作用(Kim & Tanaka, 2003)。近30年来, 泰加林带正经历着2倍于全球平均水平的增温, 同时伴随着冻土融化、水分条件改变和火干扰增强等复杂的气候变化过程(Yu et al., 2008; Brown et al., 2015; Abbott et al., 2016)。泰加林巨大的森林面积和生态系统碳储量, 及其较高的敏感性, 使其对气候变化响应的研究成为生态领域关注的焦点。
物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (Walther et al., 2002; Badeck et al., 2004; Gugger et al., 2015)。物候与环境因子之间的关系具有重要的生态意义和进化意义。已有研究普遍表明, 气候变暖使得植物春季物候提前, 秋季物候延迟, 生长季时间延长(Badeck et al., 2004; Pe?uelas et al., 2009; Wang et al., 2014b; Meng et al., 2016)。但对增温后植物花期及繁殖速率变化的研究却未得出一致的结论(Amano et al., 2010; Wolkov ich et al., 2012; Dorji et al., 2013; Wang et al., 2014a)。该结论的不一致主要来源于以下3个方面: 首先, 不同物种繁殖物候对增温的响应(变化方向及敏感性)存在差异(Cleland et al., 2007, 2012; Sherry et al., 2007; Li et al., 2016; Meng et al., 2017); 其次, 增温过程是多因子耦合变化的过程, 繁殖物候对增温的响应可能受到其他共变因子的干扰(Jorgenson et al., 2001); 另外, 植被发育是一个连续的过程, 早期的发育阶段会对后期的发育阶段产生一定的限制作用(Post et al., 2008), 通过研究单个发育阶段来描述繁殖物候对气候变暖的响应存在局限性(Post et al., 2008; Wolkovich et al., 2012)。为此, 目前的研究多采用较完整的物候序列来反映繁殖物候的改变。但是, 对于增温造成的其他共变因子(如土壤水分降低)对繁殖物候的影响并未得到充分的认识, 使气候变化下植物繁殖物候的预测存在诸多不确定性(Forrest & Miller-Rushing, 2010)。
由于气候变暖使得火干扰对泰加林的影响进一步增强, 将会有更大比例的泰加林变成火烧迹地, 处于演替初级阶段(Yu et al., 2008; Abbott et al., 2016)。同时, 火烧迹地发生着普遍的冻土退化问题(Brown et al., 2015), 冻土融化后土壤水分的剧变强烈地影响着群落动态, 甚至影响泰加林的生态演替过程(王绪高等, 2004; 孙明学, 2011)。物候是一个对环境变化快速响应的指标(Badeck et al., 2004), 泰加林演替进程的改变可能首先体现在植被生长和繁殖物候上。因此, 演替初期泰加林植被生长和繁殖物候对气候变暖及土壤水分状态改变的响应研究具有重要的价值和意义, 有助于认识气候变化下泰加林的生态演替过程。本研究以演替初期泰加林为研究对象, 跟踪调查模拟增温和排水处理2年后灌草层4种典型物种的开花物候和生长性状, 旨在阐明以下3个问题: (1)暖干化气候变化趋势将对泰加林群落演替产生何种影响?(2)不同物种繁殖物候与生长对暖干化气候变化的响应是否存在种间差异?(3)排水处理是否影响植被繁殖物候与生长对气候变暖的响应?该研究将有助于更好地认识泰加林的生态演替过程, 同时弥补繁殖物候对增温及其共变环境因子响应研究的缺失, 为多年冻土区泰加林响应未来气候变暖的模型研究提供基础数据和理论依据, 也为人类应对未来气候变化提供参考。
1 材料和方法
1.1 研究区概况
研究区(51.89° N, 121.92° E, 海拔691 m)位于大兴安岭多年冻土区泰加林带, 距离内蒙古根河市区约175 km。年平均气温-3.6 ℃, 8月平均气温最高(18.1 ℃), 1月平均气温最低(-29.8 ℃)。年降水量约500 mm, 60%以上集中在6-8月。该区域受火干扰影响严重, 2008年的一场雷击大火, 导致约478 hm2的森林被烧毁, 植物全部死亡, 并且烧掉约5 cm厚的地被物。火干扰后自然恢复, 无人为干扰。研究样地位于该火烧迹地内, 2016年开展植物生长和物候调查时生态系统已自然恢复8年, 大量植被恢复。恢复植物主要包括: 较高大的灌木柴桦(Betula fruticosa)和辽东桤木(Alnus hirsuta), 平均高度分别为(1.7 ± 0.1) m和(2.2 ± 0.1) m, 平均盖度分别为21%和55%, 以及平均高度<50 cm的主要由杜香(Ledum palustre)、薹草属(Carex)、拂子茅属(Calamagrostis)及风毛菊属(Saussurea)植物组成的灌草层, 还有覆盖于部分阴湿地面的泥炭藓(Sphagnum)等。1.2 试验设计
模拟增温采用国际冻原计划所采用的被动式增温方法, 即开顶式增温罩(OTCs)。为了对样地内灌木进行更好的增温模拟, 本研究采用的OTCs高度较高, 约2 m, 底部和顶部均为4 m2的正方形。排水处理通过挖掘排水沟(3条)实现, 排水沟深度和宽度约40 cm, 长度约100 m。在200 m × 100 m的样地内按照裂区设计法布置各样方。首先将样地分为2个100 m × 100 m的区域, 随机选择其中一个区域进行排水处理, 形成排水与不排水两个小区, 排水与不排水区域之间距离约50 m。然后, 在排水和不排水两个小区内分别随机设置增温和不增温样方各8个, 样方大小50 cm × 50 cm, 距离3-5 m, 即总共形成4种处理(增温+排水, WD; 增温, W; 排水, D; 对照, CK), 32个样方。试验样地用围栏封育, 样方布置于2014年6月初完成。1.3 供试材料
根据样方内物种始花期的时间分布, 选择不同开花功能群(Sherry et al., 2007)的4种代表性植物为主要观测对象, 即早花植物圆锥薹草(Carex diandra)和水葡萄茶藨子(Ribes procumbens), 始花期分别为5月25日(儒略日(145 ± 2)天)和5月30日(儒略日(150 ± 1)天); 中花植物杜香, 始花期平均为6月27日(儒略日(178 ± 1)天); 晚花植物齿叶风毛菊(Saussurea neoserrata), 始花期平均为8月2日(儒略日(214 ± 4)天)。1.4 数据收集
样地内各气象要素, 由研究区中心设置的微型气象站提供。同时, 在排水和不排水区域分别随机选择一个OTCs, 在其内外布设数据采集器EM50 (Decagon Devices, Pullman, USA), 收集空气温度、湿度及太阳辐射强度等数据(距离地面约1 m)。每个样方内的土壤温度(5 cm)和土壤体积含水量(0-10 cm)由便携式仪器MST3000+ (STEP Systems GmbH, Essen, Germany)测量, 与物候观测同步进行。本研究采用固定样方, 实地观测植物开花物候。2016年5月至9月, 通过连续记录各样方内, 群落中目标物种处于各开花物候阶段的花的数量, 计算关键的生殖物候指标。在野外观测期间, 分别记录目标物种处于各物候阶段的花的数量和日期(以该年1月1日为第一天, 将记录日期转化为儒略日), 记录频度约1周一次, 开花集中期增加到1周2次。本研究观测的开花物候包括: (1)开花初始期(始花期), 即记录到第一朵花开花(非禾草状植物花萼和花瓣完全张开; 禾草状植物花药可见)的日期。(2)开花峰值期, 即记录到开花数量最大的日期。(3)开花结束期, 即记录到最后一朵花凋谢的日期。(4)开花持续时间, 即开花结束期与开花初始期的间隔时间。(5)最大开花数量, 即开花峰值期所记录到的开花数量。
在8月上旬, 群落地上净初级生产力达到峰值时, 调查目标物种的高度、盖度和频度(每个样方分为25个小格, 目标物种出现的小格数量所占比例)。在物候观测结束后, 将植物体齐地剪下, 分种收集所有地上部分。将收集到的各物种的地上部分装入牛皮纸袋, 带回实验室, 于干燥箱中65 ℃烘干48 h至恒质量, 然后称量, 记录质量。
1.5 数据分析
首先通过Excel整理数据并初步分析。然后在SPSS 19.0中利用广义线性混合模型(GLME)检验模拟增温和排水及其交互作用是否对各物候和生长指标产生显著影响。检验中以各物候和生长指标为响应变量, 增温与排水处理为固定效应, 样方所在的小区为随机效应。最后, 需要图形表示的结果在Origin 9.0中完成作图。2 结果
2.1 增温和排水处理后环境因子变化
本研究中, OTCs对其内空气温度具有明显的增温作用, 使得空气温度在排水与不排水区域内分别增加了0.99和0.74 ℃, 对土壤温度(p < 0.05)和土壤含水量(p < 0.05)均有显著的影响(表1)。排水对土壤环境因子的影响较大, 表现为土壤温度显著升高 (p < 0.01), 土壤水分显著降低(p < 0.01), 生长季平均水位降低(表1)。同时, 排水显著降低了土壤最大融化深度(p < 0.01; 表1)。Table 1
表1
表1模拟增温和排水处理对生长季主要环境因子的影响(平均值或平均值±标准误差)
Table 1
| 对照 Control | 增温 Warming | 排水 Drainage | 增温+排水 Warming + Drainage | |
|---|---|---|---|---|
| 空气温度 Air temperature (℃) | 7.62 | 8.36 | 7.69 | 8.68 |
| 土壤温度 Soil temperature at 5 cm (℃) W, D | 5.37 ± 0.14 | 5.38 ± 0.14 | 5.97 ± 0.04 | 5.91 ± 0.13 |
| 土壤水分 Soil moisture at 0-10 cm (%)W, D | 45.52 ± 2.31 | 38.16 ± 6.82 | 25.36 ± 2.47 | 19.52 ± 1.05 |
| 最大融化深度 Maximun thaw depth (cm)D | 62.4 ± 1.2 | 65.1 ± 1.2 | 57.5 ± 1.8 | 55.2 ± 1.6 |
| 生长季平均水位 Mean water table (cm) | -20.0 | ND | -26.3 | ND |
新窗口打开|下载CSV
2.2 植物开花物候对增温和排水处理的响应
对于早花植物圆锥薹草, 由于在CK和D样方内未监测到圆锥薹草开花, 因此该部分开花物候缺失(图1A)。而在W和WD处理下监测到了圆锥薹草开花, 说明模拟增温可能影响其开花。对于早花植物水葡萄茶藨子, 增温和排水处理后表现为开花初始期、结束期及峰值期均不同程度提前, 但花期持续时期没有显著变化(图1B)。具体而言, W、D和WD处理下水葡萄茶藨子的开花初始期比CK分别提前了1天、0天和3天; 开花峰值期比CK分别提前了7天、7天和9天; 开花结束期比CK分别提前了5天、5天和6天(图1)。图1
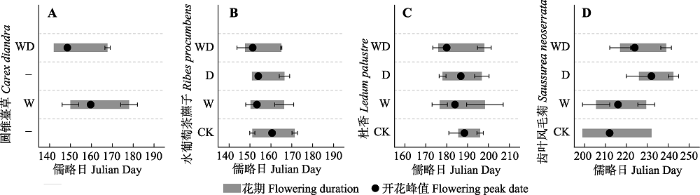 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1模拟增温和排水处理对灌草层代表性物种开花物候的影响(平均值±标准误差)。CK、W、D和WD分别代表对照、增温、排水和增温+排水处理。
Fig. 1Simulated warming and drainage effects on the flowering phenology of the typical shrub-grass species (mean ± SE). CK, control; W, warming; D, drainage; WD, warming + drainage.
对于中花植物杜香, 模拟增温和排水处理使其开花初始期及峰值期提前, 开花结束期延后, 表现为开花持续时间延长(图1C), 同时开花数量增加(图2)。具体而言, W、D和WD处理下杜香的开花初始期比CK分别提前了9天、8天和10天; 开花峰值期比CK分别提前了5天、2天和9天; 开花结束期比CK分别延后2天、1天和2天; 开花持续时间比CK分别延长了11天、8天和12天; 最大开花数量比CK分别增加了38朵、58朵和40朵。
图2
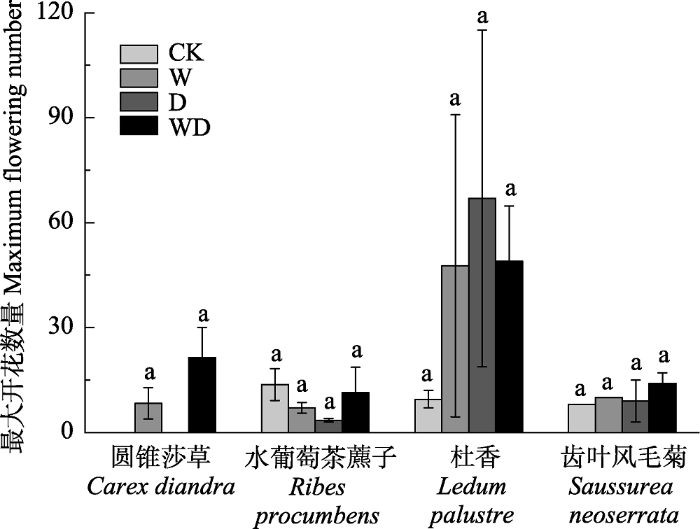 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2模拟增温和排水处理对灌草层代表性物种最大开花数量的影响(平均值±标准误差)。相同物种内不同小写字母表示处理间差异显著(p < 0.05)。CK、W、D和WD分别代表对照、增温、排水和增温+排水处理。
Fig. 2Simulated warming and drainage effects on maximum flowering number of typical shrub grass species (mean ± SE). Different small letters in the same species indicated significant difference among different treatments (p < 0.05). CK, control; W, warming; D, drainage; WD, warming + drainage.
对于晚花植物齿叶风毛菊, 模拟增温和排水处理使其开花初始期、结束期和峰值期都不同程度延后, 同时开花时间缩短(图1D)。具体而言, W、D和WD处理下齿叶风毛菊的开花初始期比CK分别延后了7天、27天和18天; 开花峰值期比CK分别延后了4天、20天和12天; 开花结束期比CK分别延后-3天、11天和7天; 而开花持续时间比CK分别缩短了9天、17天和11天(图1)。
但由于标准差较大, GLME仅检验到部分显著, 即排水处理使得圆锥薹草开花结束期显著提前(p < 0.01)及增温使得杜香开花峰值期显著提前(p < 0.05; 附录I; 图1)。模拟增温和排水处理对植物开花物候的影响没有显著的交互作用(附录I)。
Supplement I
附录I
附录I广义线性混合模型统计结果
Supplement I
| 物种 Species | n | 增温 Warming | 排水 Drainage | 增温 × 排水 Warming × Drainage | ||||
|---|---|---|---|---|---|---|---|---|
| 系数 Coefficient | 显著性 Significance | 系数 Coefficient | 显著性 Significance | 系数 Coefficient | 显著性 Significance | |||
| 圆锥薹草 Carex diandra | 高度 Height | 13 | 4.073 | 0.085 | 13.212 | 0.082 | -12.960 | 0.007 |
| 盖度 Coverage | 13 | -27.000 | 0.108 | -11.583 | 0.725 | 0.833 | 0.974 | |
| 频度 Frequency | 13 | -14.015 | 0.385 | -18.246 | 0.671 | -30.882 | 0.268 | |
| 地上生物量 Above-ground biomass | 13 | -2.692 | 0.362 | -1.731 | 0.774 | -0.322 | 0.934 | |
| 开花初始期 First flowering date | 7 | - | - | 8.000 | 0.210 | - | - | |
| 开花结束期 Last flowering date | 7 | - | - | 10.250 | 0.009 | - | - | |
| 开花峰值期 Flowering peak date | 7 | - | - | 11.167 | 0.076 | - | - | |
| 开花持续时间 Flowering duration | 7 | - | - | 2.250 | 0.727 | - | - | |
| 最大开花数量 Maximum flowering number | 7 | - | - | -13.167 | 0.496 | - | - | |
| 水葡萄茶藨子 Ribes procumbens | 高度 Height | 16 | 2.070 | 0.512 | -2.947 | 0.550 | 3.880 | 0.352 |
| 盖度 Coverage | 16 | -5.000 | 0.741 | -17.333 | 0.468 | 34.000 | 0.105 | |
| 频度 Frequency | 16 | -5.200 | 0.785 | -13.867 | 0.641 | 26.533 | 0.298 | |
| 地上生物量 Above-ground biomass | 16 | -6.507 | 0.263 | -8.738 | 0.299 | 10.792 | 0.138 | |
| 开花初始期 First flowering date | 15 | 1.727 | 0.449 | 1.382 | 0.799 | -2.524 | 0.604 | |
| 开花结束期 Last flowering date | 15 | 3.788 | 0.097 | 3.164 | 0.542 | 3.595 | 0.434 | |
| 开花峰值期 Flowering peak date | 15 | 5.576 | 0.104 | 4.327 | 0.581 | 4.571 | 0.512 | |
| 开花持续时间 Flowering duration | 15 | 2.061 | 0.418 | 1.782 | 0.768 | 6.119 | 0.247 | |
| 最大开花数量 Maximum flowering number | 15 | 1.424 | 0.813 | 3.073 | 0.832 | 14.548 | 0.252 | |
| 杜香 Ledum palustre | 高度 Height | 29 | 6.000 | 0.040 | 1.223 | 0.870 | -10.108 | 0.024 |
| 盖度 Coverage | 29 | 19.250 | 0.025 | -11.590 | 0.541 | -21.502 | 0.089 | |
| 频度 Frequency | 29 | 3.500 | 0.591 | -3.088 | 0.832 | -23.450 | 0.023 | |
| 地上生物量 Above-ground biomass | 29 | 12.999 | 0.249 | -20.981 | 0.579 | 5.287 | 0.824 | |
| 开花初始期 First flowering date | 16 | 4.088 | 0.177 | 3.514 | 0.642 | 6.933 | 0.241 | |
| 开花结束期 Last flowering date | 16 | -1.546 | 0.642 | -0.139 | 0.987 | -0.039 | 0.996 | |
| 开花峰值期 Flowering peak date | 16 | 6.097 | 0.043 | 3.042 | 0.671 | -3.051 | 0.457 | |
| 开花持续时间 Flowering duration | 16 | -5.634 | 0.243 | -3.653 | 0.763 | -7.315 | 0.470 | |
| 最大开花数量 Maximum flowering number | 16 | 0.838 | 0.982 | -24.736 | 0.796 | -52.318 | 0.493 | |
| 齿叶风毛菊 Saussurea neoserrata | 高度 Height | 11 | -9.412 | 0.614 | -12.858 | 0.759 | 5.733 | 0.828 |
| 盖度 Coverage | 11 | -1.914 | 0.671 | -1.052 | 0.918 | -1.315 | 0.837 | |
| 频度 Frequency | 11 | 4.000 | 0.757 | 4.667 | 0.866 | -18.667 | 0.336 | |
| 地上生物量 Above-ground biomass | 11 | -3.670 | 0.186 | -3.098 | 0.438 | 3.361 | 0.326 | |
| 开花初始期 First flowering date | 8 | 3.464 | 0.623 | -17.036 | 0.289 | -15.500 | 0.311 | |
| 开花结束期 Last flowering date | 8 | 3.143 | 0.447 | -9.857 | 0.287 | -1.000 | 0.916 | |
| 开花峰值期 Flowering peak date | 8 | 3.714 | 0.448 | -12.284 | 0.265 | -12.201 | 0.235 | |
| 开花持续时间 Flowering duration | 8 | -0.321 | 0.943 | 7.179 | 0.477 | 14.500 | 0.105 | |
| 最大开花数量 Maximum flowering number | 8 | -3.929 | 0.344 | -2.929 | 0.734 | 3.000 | 0.746 | |
新窗口打开|下载CSV
2.3 植物生长对增温和排水处理的响应
植物生长对增温和土壤水分降低具有敏感的响应, 且不同物种响应方式不同。增温和排水处理后圆锥薹草、杜香和齿叶风毛菊的盖度和频度均呈增加的趋势(图3)。GLME结果表明, 模拟增温显著增加了杜香的高度和盖度(附录I; 图3), 但排水处理影响模拟增温对杜香高度、频度以及圆锥薹草高度的作用, 表现出交互效应(附录I; 图3)。增温和排水处理后, 圆锥薹草、水葡萄茶藨子、杜香和齿叶风毛菊地上生物量均无明显的改变(附录I; 图3)。图3
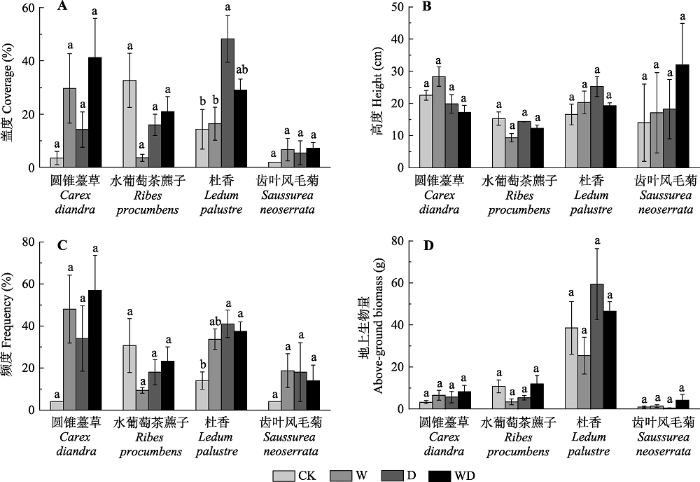 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3模拟增温和排水处理对灌草层代表性物种的盖度(A)、高度(B)、频度(C)及地上生物量(D)的影响(平均值±标准误差)。相同物种内不同小写字母表示处理间差异显著(p < 0.05)。CK、W、D和WD分别代表对照、增温、排水和增温+排水处理。
Fig. 3Simulated warming and drainage effects on coverage (A), height (B), frequency (C) and above-ground biomass (D) of the shrub-grass typical species (mean ± SE). Different small letters in the same species indicated significant difference among different treatments (p < 0.05). CK, control; W, warming; D, drainage; WD, warming + drainage.
3 讨论
3.1 植物开花物候对模拟暖干化气候的响应
不同物种具有不同的繁殖物候期, 这是物种长期进化过程中不断适应环境的结果, 也是植物降低种间竞争并维持群落水平物种共存的重要机制(Dudgeon et al., 1999; Cleland et al., 2006)。本研究发现, 不同物种的开花物候对气候变化(增温及土壤水分降低)的响应也表现出明显的种间差异, 尤其体现在不同开花功能群间。这与Sherry等(2007)和Meng等(2016, 2017)的研究结果一致。对于早花植物, 模拟增温及排水处理使其开花初始期、峰值期和结束期呈提前趋势。增温使早花期植物始花期提前得到了较为普遍的验证(Sherry et al., 2007; 吴瑞芬等, 2009; CaraDonna et al., 2014), 它是植物适应性进化的结果(Anderson et al., 2012)。植物始花期受控于其前期温度积累(吴瑞芬等, 2009; Wang et al., 2014a),增温使其温度积累提前达到要求从而提前植物始花期。同时, Schemske等(1978)认为, 因为温带森林较低的早春季节温度对植物生长和昆虫活动产生限制作用, 温度成为植物繁殖物候的重要限制因素, 增温对温度限制的解除也可能促使植物始花期提前。此外, 在受冻土强烈影响的该区域, 气候变化还可能通过加速春季冻土融化进程, 而影响早花植物始花期(Dorji et al., 2013)。本研究中, 由于开花结束期也相应地提前, 植物花期时长并没有发生显著改变。也就是说, 早花植物的花期产生了一个整体的向前推移, 但持续时间保持不变。由于普遍的认识是早花植物比晚花植物对气候变暖更加敏感(Meng et al., 2016), 因此对早花植物的关注较多, 而对晚花植物开花物候的增温响应还缺乏足够的认识。但就植物群落而言, 各种植物开花次第进行且相互间存在重叠, 各物种开花物候组成了群落开花物候, 这是一个复杂的过程, 只关注始花期较早的植物对气候变化的响应不能科学地反映植物物候的变化。因此, 本研究观测了晚花植物齿叶风毛菊对气候变化的响应。与Sherry等(2007)的研究结果类似, 本研究结果表明, 增温及排水条件下, 晚花植物齿叶风毛菊呈始花期延后, 花期缩短趋势。这可能是由于晚花植物始花期在盛夏, 不再像早花植物受低温限制, 增温反而可能导致繁殖限制, 从而使其开花初始期延后。虽然研究表明, 降雨增加对植物开花和结果物候无显著影响(Sherry et al., 2007), 但是增雨对物候的影响与其发生的时间和量有关系(叶鑫等, 2014)。本研究中, 模拟增温和排水处理下, 齿叶风毛菊在大量降雨到来, 土壤水分充足后进入花期, 说明模拟增温和排水处理导致的土壤含水量降低(表1), 可能使盛夏开花的齿叶风毛菊因水分限制而推迟开花。植物物候是一系列循环发生的连续的季节性生物事件, 各物候阶段存在此消彼长的密切联系(Post et al., 2008)。晚花植物的齿叶风毛菊更长的营养生长时间及花蕾阶段可使其花期延后。同时, 增温和排水处理缩短了晚花植物齿叶风毛菊的开花持续时间, 这可能对其遗传变异及进化适应产生选择压力(Sherry et al., 2007)。
植物生长变化是群落响应气候变化的又一重要途径(Pe?uelas et al., 2007)。本研究发现植物生长对模拟增温和排水处理具有敏感的响应, 且不同物种间响应方式存在差异。增温和排水处理后圆锥薹草、杜香和齿叶风毛菊的盖度和频度都呈增加的趋势, 其中杜香的高度和盖度在增温后显著增加(图3; 附录I)。但对于水葡萄茶藨子, 增温及排水处理后其高度、盖度和频度均呈下降趋势(图3)。不同物种的生长对气候变化的响应与其生长特性及其对环境的适应性有关(Pe?uelas et al., 2007)。水葡萄茶藨子多生长于林下阴湿处及河岸旁, 是对水分要求较高的喜阴植物。因此, 模拟增温和排水处理后土壤水分的降低对水葡萄茶藨子生长产生不利的影响。相反, 圆锥薹草、杜香和齿叶风毛菊则表现出对模拟增温和排水处理的积极响应。由于不利的环境条件及群落种间竞争的压力, 增温及排水处理后水葡萄茶藨子的各性状特征表现为下降趋势。就本研究中选取的4种代表性物种来推测, 演替初期泰加林群落应对短期的暖干化气候变化, 将可能会通过物种间对生境的适应能力, 改变种间竞争关系, 从而影响群落组成和结构(Pe?uelas et al., 2002)。
3.3 排水处理对植物开花物候和生长的影响
温度可以直接控制生物物候事件开始的时间, 被称为生物原始的“生物钟” (Ausín et al., 2005; Forrest & Miller-Rushing, 2010)。在众多影响物候的环境因子中, 温度所起的作用最大(Wolkovich et al., 2012)。但是, 本研究中排水处理(D)对植物开花物候的影响表现出与模拟增温(W)相似的效果, 说明排水处理会在一定程度上影响植物开花物候。研究表明, 土壤水分降低对植物开花物候具有复杂的影响(Galen et al., 1999; Pe?uelas & Filella, 2001; Wang et al., 2014b)。干旱一般会通过缩短植物花期来减少因开花而增加的水分丧失量(Pe?uelas & Filella, 2001)。同时, 低水分状况可以减缓花芽和结实的过程(Galen et al., 1999), 但也研究得出相反的结果(Wang et al., 2014b)。此外, 本研究中, 排水处理使得土壤表层温度有明显的升高, 这可能对植物根系物候产生影响(Sullivan & Welker, 2005; Steinaker et al., 2010)。有研究发现, 浅层根系与地上物候保持较好的同步性(Brouwer, 1983), 土壤水分降低则有可能通过根系物候对地上物候产生影响。本研究中, 模拟增温和排水处理对植物开花物候的影响并未表现出显著的交互效应, 说明土壤水分降低并不会影响植物开花物候对模拟增温的响应。该结果与本研究样地较高的土壤水分有关, 在受水分限制的生态系统则可能出现不同的结果, 这有待进一步探索。土壤水分作为植物生命活动的重要基础, 对植物生长产生重要的直接影响。在本研究中, 土壤水分充足, 模拟土壤水分降低对圆锥薹草、杜香和齿叶风毛菊的生长(高度、盖度、频度)产生一定的促进作用, 对水葡萄茶藨子产生一定的抑制作用, 但均未达显著水平。然而, 模拟土壤水分降低显著影响模拟增温对杜香高度、频度以及圆锥薹草高度的作用(附录I; 图3)。该结果说明, 模拟增温对杜香和圆锥薹草生长的促进作用可能受到土壤水分降低的限制。
3.4 模拟暖干化气候条件下的群落演替倾向
本研究中, 杜香是灌草层的优势物种, 其生物量占群落总生物量(高度<50 cm的灌草层)的(46.0 ± 4.2)%。在增温和土壤水分降低的气候条件下, 属于中花植物的杜香开花峰值期显著提前, 且花期呈延长趋势, 开花数量呈增加趋势。更长的花期使得杜香的花期与其传粉动物的活动期不一致的可能性降低, 增加了杜香的繁殖成功率。延长的花期和增加的开花数量还可以增加配型, 使基因多样化, 利于杜香的进化。同时, 植物个体开花物候对环境变化的响应与其生长性状紧密相连。有研究表明, 在增温条件下, 物候提前较多的物种, 在丰富度和生态幅上有较大的提高, 反之, 其性状特征(生物量、相对盖度、开花数目)则相对降低(Hulme, 2011)。与此一致, 本研究中模拟增温使杜香开花峰值期显著提前, 虽然其生物量没有显著变化, 但作为主要性状特征的高度和盖度均显著提高(附录I; 图3)。综上, 说明在短期增温条件下, 杜香可以通过调整开花物候和生长性状, 使其生态幅增加, 而在群落中的优势地位更加明显。根据孙明学(2011)的研究, 在大兴安岭泰加林的火后演替过程中, 起初灌木层中频度高的物种很少, 随着时间的推移, 灌木层中植被对资源和空间的竞争将日益加剧, 随着乔木层林冠逐渐恢复, 灌木层利用资源和空间的压力也急剧上升, 从而又对一些阳性灌木进行淘汰, 最终使灌木层物种数量减少, 使优势灌木种的频度显著提高。同样, 草本层也经历这样的过程, 随着演替时间的推移, 草本层物种数量大幅下降, 从而使高频物种占据优势。因此, 火后灌草层植被的演替过程, 是一个优势物种逐渐淘汰其他物种, 最终占据主导地位的过程(王绪高等, 2004; 孙明学, 2011)。短期来看, 模拟增温和土壤水分降低促使作为优势物种的杜香更具繁殖优势, 且占有更宽的生态幅, 使其在灌草层的优势地位更加明显, 这与泰加林火后恢复中灌草层的演替路径非常一致(王绪高等, 2004; 孙明学, 2011)。因此, 我们可以在一定程度上推测, 短期内增温和土壤水分降低可能加速演替初期泰加林灌草层的火后演替进程。由于高频物种所占比例的增加是有利于群落稳定发展的, 能为乔木的更新创造良好的生态环境(孙明学, 2011)。因此, 增温和土壤水分降低可能形成有利于泰加林恢复的生态环境, 加快泰加林恢复更新。
4 结论和展望
未来暖干化气候条件将改变植物开花物候, 且这种改变因植物开花功能群而异。早花植物由于开花温度限制的提早解除而花期提前; 晚花植物由于开花受水分限制, 花期呈延后趋势; 早花植物和晚花植物开花物候的反方向变化可能为中花植物提供更大的生态位空隙, 使其开花数量趋向于增加, 花期提前且呈延长趋势。暖干化气候变化导致的开花物候改变可能为物种入侵或对群落结构变化提供生态位间隙, 从而可能对种间竞争产生潜在影响, 引发群落组成改变, 对生态和进化产生深远影响。同时, 群落通过不同开花功能群植物对气候变化的响应不同, 而调节群落整体物候变化, 可能是群落适应气候变化的重要途径。开展群落物候对气候变化的响应研究是气候变化下物候研究的重要方向。本研究只观测了4种代表性植物的生长与开花物候, 可能无法表征群落整体的响应。但在一定程度上, 我们可以推测短期的暖干化气候变化可能加速演替初期泰加林灌草层的火后演替进程, 促进泰加林的恢复更新。另外, 本研究区域水分充足, 土壤水分降低仅影响了部分物种生长对增温的响应, 并未影响植物开花物候对增温的响应。但在受水分限制的生态系统中, 土壤水分降低是否影响植物物候对增温的响应仍是需要明确的重要问题。
致谢
感谢中国科学院沈阳应用生态研究所胡远满研究员、黄彦青研究员、布仁仓研究员、常禹研究员和郭金停博士等在植物鉴定工作中给予的帮助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 2]

As the permafrost region warms, its large organic carbon pool will be increasingly vulnerable to decomposition, combustion, and hydrologic export. Models predict that some portion of this release will be offset by increased production of Arctic and boreal biomass; however, the lack of robust estimates of net carbon balance increases the risk of further overshooting international emissions targets. Precise empirical or model-based assessments of the critical factors driving carbon balance are unlikely in the near future, so to address this gap, we present estimates from 98 permafrost-region experts of the response of biomass, wild75re, and hydrologic carbon 75ux to climate change. Results suggest that contrary to model projections, total permafrost-region biomass could decrease due to water stress and disturbance, factors that are not adequately incorporated in current models. Assessments indicate that end-of-the-century organic carbon release from Arctic rivers and collapsing coastlines could increase by 75% while carbon loss via burning could increase four-fold. Experts identi75ed water balance, shifts in vegetation community, and permafrost degradation as the key sources of uncertainty in predicting future system response. In combination with previous 75ndings, results suggest the permafrost region will become a carbon source to the atmosphere by 2100 regardless of warming scenario but that 65%–85% of permafrost carbon release can still be avoided if human emissions are actively reduced.
DOIURL
DOIURLPMID [本文引用: 1]
Abstract Widespread concerns about global biodiversity loss have led to a growing demand for indices of biodiversity status. Today, climate change is among the most serious threats to global biodiversity. Although many studies have revealed phenological responses to climate change, no long-term community-level indices have been developed. We derived a 250-year index of first flowering dates for 405 plant species in the UK for assessing the impact of climate change on plant communities. The estimated community-level index in the most recent 25 years was 2.2-12.7 days earlier than any other consecutive 25-year period since 1760. The index was closely correlated with February-April mean Central England Temperature, with flowering 5.0 days earlier for every 1 degrees C increase in temperature. The index was relatively sensitive to the number of species, not records per species, included in the model. Our results demonstrate how multi-species, multiple-site phenological events can be integrated to obtain indices showing trends for each species and across species. This index should play an important role in monitoring the impact of climate change on biodiversity. Furthermore, this approach can be extended to incorporate data from other taxa and countries for evaluating cross-taxa and cross-country phenological responses to climate change.
DOIURLPMID [本文引用: 1]
Anthropogenic climate change has already altered the timing of major life-history transitions, such as the initiation of reproduction. Both phenotypic plasticity and adaptive evolution can underlie rapid phenological shifts in response to climate change, but their relative contributions are poorly understood. Here, we combine a continuous 38 year field survey with quantitative genetic field experiments to assess adaptation in the context of climate change. We focused on Boechera stricta (Brassicaeae), a mustard native to the US Rocky Mountains. Flowering phenology advanced significantly from 1973 to 2011, and was strongly associated with warmer temperatures and earlier snowmelt dates. Strong directional selection favoured earlier flowering in contemporary environments (2010-2011). Climate change could drive this directional selection, and promote even earlier flowering as temperatures continue to increase. Our quantitative genetic analyses predict a response to selection of 0.2 to 0.5 days acceleration in flowering per generation, which could account for more than 20 per cent of the phenological change observed in the long-term dataset. However, the strength of directional selection and the predicted evolutionary response are likely much greater now than even 30 years ago because of rapidly changing climatic conditions. We predict that adaptation will likely be necessary for long-term in situ persistence in the context of climate change.
DOIURL [本文引用: 1]
DOIURL [本文引用: 3]
Climate change effects on seasonal activity in terrestrial ecosystems are significant and well documented, especially in the middle and higher latitudes. Temperature is a main driver of many plant developmental processes, and in many cases higher temperatures have been shown to speed up plant development and lead to earlier switching to the next ontogenetic stage. Qualitatively consistent advancement of vegetation activity in spring has been documented using three independent methods, based on ground observations, remote sensing, and analysis of the atmospheric CO 2 signal. However, estimates of the trends for advancement obtained using the same method differ substantially. We propose that a high fraction of this uncertainty is related to the time frame analysed and changes in trends at decadal time scales. Furthermore, the correlation between estimates of the initiation of spring activity derived from ground observations and remote sensing at interannual time scales is often weak. We propose that this is caused by qualitative differences in the traits observed using the two methods, as well as the mixture of different ecosystems and species within the satellite scenes.
URL [本文引用: 1]
DOIURL [本文引用: 2]
Abstract We examined the effects of fire disturbance on permafrost degradation and thaw settlement across a series of wildfires (from ~1930 to 2010) in the forested areas of collapse-scar bog complexes in the Tanana Flats lowland of interior Alaska. Field measurements were combined with numerical modeling of soil thermal dynamics to assess the roles of fire severity and climate history in postfire permafrost dynamics. Field-based calculations of potential thaw settlement following the loss of remaining ice-rich permafrost averaged 0.665m. This subsidence would cause the surface elevations of forests to drop on average 0.165m below the surface water level of adjacent collapse-scar features. Up to 0.565m of thaw settlement was documented after recent fires, causing water impoundment and further thawing along forest margins. Substantial heterogeneity in soil properties (organic layer thickness, texture, moisture, and ice content) was attributed to differing site histories, which resulted in distinct soil thermal regimes by soil type. Model simulations showed increasing vulnerability of permafrost to deep thawing and thaw settlement with increased fire severity (i.e., reduced organic layer thickness). However, the thresholds of fire severity that triggered permafrost destabilization varied temporally in response to climate. Simulated permafrost dynamics underscore the importance of multiyear to multidecadal fluctuations in air temperature and snow depth in mediating the effects of fire on permafrost. Our results suggest that permafrost is becoming increasingly vulnerable to substantial thaw and collapse after moderate to high-severity fire, and the ability of permafrost to recover is diminishing as the climate continues to warm.
DOIURL [本文引用: 1]
Phenology--the timing of biological events--is highly sensitive to climate change. However, our general understanding of how phenology responds to climate change is based almost solely on incomplete assessments of phenology (such as first date of flowering) rather than on entire phenological distributions. Using a uniquely comprehensive 39-y flowering phenology dataset from the Colorado Rocky Mountains that contains more than 2 million flower counts, we reveal a diversity of species-level phenological shifts that bring into question the accuracy of previous estimates of long-term phenological change. For 60 species, we show that first, peak, and last flowering rarely shift uniformly and instead usually shift independently of one another, resulting in a diversity of phenological changes through time. Shifts in the timing of first flowering on average overestimate the magnitude of shifts in the timing of peak flowering, fail to predict shifts in the timing of last flowering, and underrepresent the number of species changing phenology in this plant community. Ultimately, this diversity of species-level phenological shifts contributes to altered coflowering patterns within the community, a redistribution of floral abundance across the season, and an expansion of the flowering season by more than I mo during the course of our study period. These results demonstrate the substantial reshaping of ecological communities that can be attributed to shifts in phenology.
DOIURLPMID [本文引用: 1]
Abstract Earlier spring phenology observed in many plant species in recent decades provides compelling evidence that species are already responding to the rising global temperatures associated with anthropogenic climate change. There is great variability among species, however, in their phenological sensitivity to temperature. Species that do not phenologically "track" climate change may be at a disadvantage if their growth becomes limited by missed interactions with mutualists, or a shorter growing season relative to earlier-active competitors. Here, we set out to test the hypothesis that phenological sensitivity could be used to predict species performance in a warming climate, by synthesizing results across terrestrial warming experiments. We assembled data for 57 species across 24 studies where flowering or vegetative phenology was matched with a measure of species performance. Performance metrics included biomass, percent cover, number of flowers, or individual growth. We found that species that advanced their phenology with warming also increased their performance, whereas those that did not advance tended to decline in performance with warming. This indicates that species that cannot phenologically "track" climate may be at increased risk with future climate change, and it suggests that phenological monitoring may provide an important tool for setting future conservation priorities.
DOIURLPMID [本文引用: 1]
Shifting plant phenology (i.e., timing of flowering and other developmental events) in recent decades establishes that species and ecosystems are already responding to global environmental change. Earlier flowering and an extended period of active plant growth across much of the northern hemisphere have been interpreted as responses to warming. However, several kinds of environmental change have the potential to influence the phenology of flowering and primary production. Here, we report shifts in phenology of flowering and canopy greenness (Normalized Difference Vegetation Index) in response to four experimentally simulated global changes: warming, elevated CO , nitrogen (N) deposition, and increased precipitation. Consistent with previous observations, warming accelerated both flowering and greening of the canopy, but phenological responses to the other global change treatments were diverse. Elevated CO and N addition delayed flowering in grasses, but slightly accelerated flowering in forbs. The opposing responses of these two important functional groups decreased their phenological complementarity and potentially increased competition for limiting soil resources. At the ecosystem level, timing of canopy greenness mirrored the flowering phenology of the grasses, which dominate primary production in this system. Elevated CO delayed greening, whereas N addition dampened the acceleration of greening caused by warming. Increased precipitation had no consistent impacts on phenology. This diversity of phenological changes, between plant functional groups and in response to multiple environmental changes, helps explain the diversity in large-scale observations and indicates that changing temperature is only one of several factors reshaping the seasonality of ecosystem processes.
DOIURLPMID [本文引用: 1]
Plants are finely tuned to the seasonality of their environment, and shifts in the timing of plant activity (i.e. phenology) provide some of the most compelling evidence that species and ecosystems are being influenced by global environmental change. Researchers across disciplines have observed shifting phenology at multiple scales, including earlier spring flowering in individual plants and an earlier spring green-up of the land surface revealed in satellite images. Experimental and modeling approaches have sought to identify the mechanisms causing these shifts, as well as to make predictions regarding the consequences. Here, we discuss recent advances in several fields that have enabled scaling between species responses to recent climatic changes and shifts in ecosystem productivity, with implications for global carbon cycling.
DOIURLPMID [本文引用: 2]
Abstract Global climate change is predicted to have large impacts on the phenology and reproduction of alpine plants, which will have important implications for plant demography and community interactions, trophic dynamics, ecosystem energy balance, and human livelihoods. In this article we report results of a 3-year, fully factorial experimental study exploring how warming, snow addition, and their combination affect reproductive phenology, effort, and success of four alpine plant species belonging to three different life forms in a semiarid, alpine meadow ecosystem on the central Tibetan Plateau. Our results indicate that warming and snow addition change reproductive phenology and success, but responses are not uniform across species. Moreover, traits associated with resource acquisition, such as rooting depth and life history (early vs. late flowering), mediate plant phenology, and reproductive responses to changing climatic conditions. Specifically, we found that warming delayed the reproductive phenology and decreased number of inflorescences of Kobresia pygmaea C. B. Clarke, a shallow-rooted, early-flowering plant, which may be mainly constrained by upper-soil moisture availability. Because K. pygmaea is the dominant species in the alpine meadow ecosystem, these results may have important implications for ecosystem dynamics and for pastoralists and wildlife in the region. 2012 Blackwell Publishing Ltd.
DOIURL [本文引用: 1]
The lower intertidal zone (0.0 to + 1.0 m mean low water [MLW]) ofky shores in New England is a space-limited community occupied by two similar rhodophyte seaweeds, Chondrus crispus and Mastocarpus stellatus, that overlap broadly in their use of three essential resources: space, light, and nutrients. C. crispus coexists primarily with the prostrate-crust generation of M. stellatus lower on the shore (less than +0.35 m MLW) and with the alternate upright-frond generation higher on the shore (greater than +0.35 m MLW). Our objectives were to determine (1) whether these two species compete and (2) if so, what process(es) enable their coexistence. Upright fronds of M. stellatus transplanted to the lowest intertidal zone (less than +0.25 m MLW) where C. crispus predominates grew faster and showed higher survivorship after 2 yr than those transplanted to areas where M. stellatus predominates. However, the failure of upright fronds of M. stellatus to consistently recruit limits their abundance in the lowest zone and reduces the frequency of preemptive competition by C. crispus. Moreover, when M. stellatus was grown in combination with fronds of C. crispus in this zone, the dominant competitor C. crispus suppressed the growth and reproductive output of M. stellatus fronds. Neither species was significantly consumed by littorinid gastropods in field and laboratory experiments, indicating that herbivory does not control patterns of coexistence. In the low zone, coexistence between C. crispus and M. stellatus appears to be mediated primarily by disturbances from winter storm waves that remove large, fast-growing C. crispus fronds and limit its abundance. Coexistence of C. crispus fronds and M. stellatus crusts in this zone may also result from their different patterns of resource use. In contrast to the low zone, the slow growth of C. crispus in the mid-low zone (approximately +0.5 m MLW) prevented the overgrowth of fronds of M. stellatus and, hence, prevented competition from
DOIURLPMID [本文引用: 2]
Phenology affects nearly all aspects of ecology and evolution. Virtually all biological phenomena-from individual physiology to interspecific relationships to global nutrient fluxes-have annual cycles and are influenced by the timing of abiotic events. Recent years have seen a surge of interest in this topic, as an increasing number of studies document phenological responses to climate change. Much recent research has addressed the genetic controls on phenology, modelling techniques and ecosystem-level and evolutionary consequences of phenological change. To date, however, these efforts have tended to proceed independently. Here, we bring together some of these disparate lines of inquiry to clarify vocabulary, facilitate comparisons among habitat types and promote the integration of ideas and methodologies across different disciplines and scales. We discuss the relationship between phenology and life history, the distinction between organismal- and population-level perspectives on phenology and the influence of phenology on evolutionary processes, communities and ecosystems. Future work should focus on linking ecological and physiological aspects of phenology, understanding the demographic effects of phenological change and explicitly accounting for seasonality and phenology in forecasts of ecological and evolutionary responses to climate change.
[本文引用: 2]
DOIURLPMID [本文引用: 1]
BACKGROUND AND AIMS: Recent global changes, particularly warming and drought, have had worldwide repercussions on the timing of flowering events for many plant species. Phenological shifts have also been reported in alpine environments, where short growing seasons and low temperatures make reproduction particularly challenging, requiring fine-tuning to environmental cues. However, it remains unclear if species from such habitats, with their specific adaptations, harbour the same potential for phenological plasticity as species from less demanding habitats. METHODS: Fourteen congeneric species pairs originating from mid and high elevation were reciprocally transplanted to common gardens at 1050 and 2000 a.s.l. that mimic prospective climates and natural field conditions. A drought treatment was implemented to assess the combined effects of temperature and precipitation changes on the onset and duration of reproductive phenophases. A phenotypic plasticity index was calculated to evaluate if mid- and high-elevation species harbour the same potential for plasticity in reproductive phenology. KEY RESULTS: Transplantations resulted in considerable shifts in reproductive phenology, with highly advanced initiation and shortened phenophases at the lower (and warmer) site for both mid- and high-elevation species. Drought stress amplified these responses and induced even further advances and shortening of phenophases, a response consistent with an 'escape strategy'. The observed phenological shifts were generally smaller in number of days for high-elevation species and resulted in a smaller phenotypic plasticity index, relative to their mid-elevation congeners. CONCLUSIONS: While mid- and high-elevation species seem to adequately shift their reproductive phenology to track ongoing climate changes, high-elevation species were less capable of doing so and appeared more genetically constrained to their specific adaptations to an extreme environment (i.e. a short, cold growing season).
DOIURLPMID [本文引用: 1]
61 Plant phenology is particularly sensitive to climate and a key indicator of environmental change. Globally, first flowering dates (FFDs) have advanced by several days per decade in response to recent climate warming, but, while earlier flowering should allow plant distributions to increase, a link between FFD and range changes has not been observed. 61 Here I show for 347 species that the extent to which FFD has responded to climate warming is linked to the degree to which their relative distributions have changed over 30 yr across the British Isles. 61 Native plant species whose phenology did not track climate change declined in distribution, whereas species that became more widespread all exhibited earlier flowering. In contrast, alien neophytes showed both a stronger phenological response to warming and a more marked increase in distribution, but no link between the two. 61 These trends were consistent both for relative changes in the national distribution and for local abundance. At the national scale, the more recently an alien species became established in Britain, the more likely it was to increase in distribution irrespective of FFD, suggesting that recent changes in alien species distributions are decoupled from climate and driven by other factors.
Cambridge University Press, New York.
DOIURL [本文引用: 2]
Climate models have continued to be developed and improved since the AR4, and many models have been extended into Earth System models by including the representation of biogeochemical cycles important to climate change. These models allow for policy-relevant calculations such as the carbon dioxide (CO2) emissions compatible with a specified climate stabilization target. In addition, the range of climate variables and processes that have been evaluated has greatly expanded, and differences between models and observations are increasingly quantified using erformance metrics . In this chapter, model evaluation covers simulation of the mean climate, of historical climate change, of variability on multiple time scales and of regional modes of variability. This evaluation is based on recent internationally coordinated model experiments, including simulations of historic and paleo climate, specialized experiments designed to provide insight into key climate processes and feedbacks and regional climate downscaling. Figure 9.44 provides an overview of model capabilities as assessed in this chapter, including improvements, or lack thereof, relative to models assessed in the AR4. The chapter concludes with an assessment of recent work connecting model performance to the detection and attribution of climate change as well as to future projections.
DOIURL [本文引用: 1]
DOIURL
DOIURL [本文引用: 1]
We tested whether the flowering times of animal-pollinated plants are influenced by phylogenetic membership and by life form (e.g., annual, perennial, etc.). We analyzed existing data sets on 2298 animal-pollinated angiosperms of North and South Carolina and 1575 animal-pollinated angiosperms of temperate Japan, and also analyzed a null model based upon the Carolina flora. Our analyses of the complete data sets and of subsets including only the largest families showed that: (1) the floras of the Carolinas and temperate Japan have bimodal distributions of flowering times, with peaks in spring and late summer; (2) within each of these floras, families differ very significantly in flowering time; (3) for most families, flowering times in the Carolinas and in Japan are statistically indistinguishable; (4) the sequences of flowering, ordered by family, are also statistically indistinguishable in the two floras; (5) within each flora, skewness of flowering time differs markedly among families; (6) for a typical family, the skewness of flowering time is the same in the two floras; (7) there is a significant negative correlation between skewness and mean family flowering time; and (8) life forms differ in flowering time, though less markedly than families. These results demonstrate that phylogenetic membership and life form strongly influence a species' flowering time. We argue that seasonal limitations of flowering times are caused by phylogenetic constraints, which may not have changed for millions of years. This study does not provide the degree of resolution needed to determine whether or not there is natural selection for alteration of flowering times in these floras, whether by competition or other mechanisms. However, our results do suggest that competition for pollination does not push species' flowering times beyond seasonal boundaries imposed by phylogenetic constraints. The effects of competition on flowering time are probably limited to small and temporary readjustments of the phenological relationships of competitors. Studies of the flowering times of animal-pollinated plants must consider the strong limits to seasonal displacement imposed by phylogeny and life form, and the probable existence of many alternative modes of escape from competition besides seasonal shifts. We consider the implications of recent paleobotanical studies that suggest that temperate plant communities may not have been sufficiently stable through time for plants to have achieved competitive equilibria. Phylogenetic and life history constraints are likely to influence the evolution of many community characters besides flowering time. The methods we have used and the conclusions we have drawn are relevant to a wide variety of ecological characters in plants and animals.
DOIURLPMID [本文引用: 2]
Abstract Organisms' life cycles consist of hierarchical stages, from a single phenological stage (for example, flowering within a season), to vegetative and reproductive phases, to the total lifespan of the individual. Yet phenological events are typically studied in isolation, limiting our understanding of life history responses to climate change. Here, we reciprocally transfer plant communities along an elevation gradient to investigate plastic changes in the duration of sequential phenological events for six alpine species. We show that prolonged flowering leads to longer reproductive phases and activity periods when plants are moved to warmer locations. In contrast, shorter post-fruiting leaf and flowering stages led to shorter vegetative and reproductive phases, respectively, which resulted in shorter activity periods when plants were moved to cooler conditions. Therefore, phenological responses to warming and cooling do not simply mirror one another in the opposite direction, and low temperature may limit reproductive allocation in the alpine region.
DOIURL [本文引用: 2]
Change in individual species phenology is often unsuitable for predicting change in community phenology because of different responses of different species to temperature change. However, few studies have observed community phenological sequences in the field. Here we explore the changes in timing and duration of the community phenological sequence (i.e. onset of leaf-out (OLO), first flower bud (FB), first flowering (FF), first fruiting-set (FFS), post-fruiting vegetation (OPFV), first leaf-coloring (FLC) and complete leaf-coloring (CLC)) along an elevation gradient from 3200 to 3800m in an alpine meadow on the Tibetan plateau. Our results indicate that OLO and FFS significantly advanced and other timings of phenological events significantly delayed at 3200m compared with higher elevations (3600 and 3800m). The flowering duration of the community was shortest and other phenological durations (except budding stage and post-fruiting vegetation stage) were longest at 3200m. The duration of the growing season decreased as elevation increased, and the ratio of the durations of the reproductive period and growing season was smallest at 3200m. There were negative correlations between the proportion of early-spring flowering functional group plants and FB, and the durations of leafing and post-fruiting vegetation of the community. Positive correlations were found between the proportion of mid-summer flowering functional group plants in the community and these variables. There were significant negative correlations between flowering duration of the community and annual mean air temperature and soil moisture. Therefore, our results suggest that different community compositions might respond differently to climate change.
DOIURLPMID [本文引用: 1]
Our ability to predict how temperature modifies phenology at the community scale is limited by our lack of understanding of responses by functional groups of flowering plants. These responses differ among species with different life histories. We performed a reciprocal transplant experiment along four elevation gradients (e.g., 3,200, 3,400, 3,600 and 3,800 m) to investigate the effects of warming (transferred downward) and cooling (transferred upward) on plant flowering functional groups (FFGs) and community phenological sequences (i. e., seven phenological events). Warming significantly decreased early-spring-flowering (ESF) plant coverage and increased mid-summer-flowering plant (MSF) coverage, while cooling had the opposite effect. All community phenological events were advanced by warming and delayed by cooling except for the date of complete leaf-coloring, which showed the opposite response. Warming and cooling could cause greater advance or delay in early-season phenological events of the community through increased coverage of MSF species, and warming could delay late-season phenological events of the community by increased coverage of ESF species. These results suggested that coverage change of FFGs in the community induced by temperature change could mediate the responses of the community phenological events to temperature change in the future. The response of phenological events to temperature change at the species level may not be sufficient to predict phenological responses at the community-level due to phenological compensation between species in the community.
DOIURLPMID [本文引用: 2]
Climate change is already altering the magnitude and/or frequency of extreme events which will in turn affect plant fitness more than any change in the average. Although the fingerprint of anthropogenic warming in recent phenological records is well understood, the impacts of extreme events have been largely neglected. Thus, the temperature response of European phenological records to warm and cold spells was studied using the COST725 database. We restricted our analysis to the period 1951–2004 due to better spatial coverage. Warm and cold spells were identified using monthly mean ENSEMBLES temperature data on a 0.5° grid for Europe. Their phenological impact was assessed as anomalies from maps displaying mean onsets for 1930–1939. Our results clearly exhibit continental cold spells predominating in the period 1951–1988, especially during the growing season, whereas the period from 1989 onwards was mainly characterised by warm spells in all seasons. The impacts of these warm/cold spells on the onset of phenological seasons differed strongly depending on species, phase and timing. “False” phases such as the sowing of winter cereals hardly reacted to summer warm/cold spells; only the sowing of summer cereals mirrored spring temperature warm/cold spells. The heading dates of winter cereals did not reveal any consistent results probably due to fewer warm/cold spells identified in the relevant late spring months. Apple flowering and the harvest of winter cereals were the best indicators of warm/cold spells in early spring and summer, also being spatially coherent with the patterns of warm/cold spells.
DOIURLPMID [本文引用: 1]
ABSTRACT Animal and plant life cycles are increasingly shown to depend on temperature trends and patterns. In their Perspective, Pe uelas and Filella review the evidence that global warming during the 20th century has affected the growth period of plants and the development and behavior of animals from insects to birds. The authors warn that changes in the interdependence between species could have unpredictable consequences for ecosystems, that the lengthening of the plant growing season contributes to the global increased carbon fixation, and that changes in phenology may affect not only ecosystems but also agriculture and sanitation.
DOIURL [本文引用: 2]
Abstract The available data on climate over the past century indicate that the earth is warming. Important biological effects, including changes of plant and animal life cycle events, have already been reported. However, evidence of such effects is still scarce and has been mostly limited to northern latitudes. Here we provide the first long-term (1952鈥2000) evidence of altered life cycles for some of the most abundant Mediterranean plants and birds, and one butterfly species. Average annual temperatures in the study area (Cardedeu, NE Spain) have increased by 1.4 C over the observation period while precipitation remained unchanged. A conservative linear treatment of the data shows that leaves unfold on average 16days earlier, leaves fall on average 13days later, and plants flower on average 6days earlier than in 1952. Fruiting occurs on average 9days earlier than in 1974. Butterflies appear 11days earlier, but spring migratory birds arrive 15days later than in 1952. The stronger changes both in temperature and in phenophases timing occurred in the last 25years. There are no significant relationships among changes in phenophases and the average date for each phenophase and species. There are not either significant differences among species with different Raunkiaer life-forms or different origin (native, exotic or agricultural). However, there is a wide range of phenological alterations among the different species, which may alter their competitive ability, and thus, their ecology and conservation, and the structure and functioning of ecosystems. Moreover, the lengthening of plant growing season in this and other northern hemisphere regions may contribute to a global increase in biospheric activity.
DOIURL [本文引用: 1]
DOIURLPMID [本文引用: 3]
A longer growing season as a result of climate change will in turn affect climate through biogeochemical and biophysical effects.
DOIURLPMID
Abstract Climatic warming is associated with organisms breeding earlier in the season than is typical for their species. In some species, however, response to warming is more complex than a simple advance in the timing of all life history events preceding reproduction. Disparities in the extent to which different components of the reproductive phenology of organisms vary with climatic warming indicate that not all life history events are equally responsive to environmental variation. Here, we propose that our understanding of phenological response to climate change can be improved by considering entire sequences of events comprising the aggregate life histories of organisms preceding reproduction. We present results of a two-year warming experiment conducted on 33 individuals of three plant species inhabiting a low-arctic site. Analysis of phenological sequences of three key events for each species revealed how the aggregate life histories preceding reproduction responded to warming, and which individual events exerted the greatest influence on aggregate life history variation. For alpine chickweed (Cerastium alpinum), warming elicited a shortening of the duration of the emergence stage by 2.5 days on average, but the aggregate life history did not differ between warmed and ambient plots. For gray willow (Salix glauca), however, all phenological events monitored occurred earlier on warmed than on ambient plots, and warming reduced the aggregate life history of this species by 22 days on average. Similarly, in dwarf birch (Betula nana), warming advanced flower bud set, blooming, and fruit set and reduced the aggregate life history by 27 days on average. Our approach provides important insight into life history responses of many organisms to climate change and other forms of environmental variation. Such insight may be compromised by considering changes in individual phenological events in isolation.
DOIURL [本文引用: 6]
Fecundity characteristics, phenology, and behavior of insect flower-visitors were studied for 7 early flowering woodland herbs: Claytonia virginica, Dentaria laciniata, Dicentra canadensis, Dicentra cucullaria, Erythronium albidum, Isopyrum biternatum, and Sanguinaria canadensis. Sanguinaria canadensis is facultatively autogamous, the Dicentras are obligate outcrossers, and the remainder are self-compatible, at least within a stem. All are insect pollinated except sometimes S. canadensis. The numbers of ovules per flower and flowers per stem tended to be inversely correlated, and large-seeded species (S. canadensis, E. albidum, I. biternatum) had lower numbers of potential seeds per stem than did small-seeded species. Flowering of all species typically occurred during the first prolonged period of weather suitable for pollinator activity and ceased by the time the canopy closed. Annual differences in flowering times were associated with differences in average temperatures (i.e., early blooming in a warm, early spring), but cumulative degree-hours or degree-days of air or soil temperatures were not well correlated with flowering times. Other constraints on flowering phenology are discussed, including the predictability of suitable conditions, a proposed "fail-safe" mechanism that may assure flowering before canopy closure even if temperatures are abnomally low, and the importance of nontemperature factors in defining suitable conditions. Flowering time was not very finely tuned to the temperature regime and pollinator activity; flowers blooming during the flowering peak often had low seed production and the fertilization rate of most species was low. Evidence that seed production may have been pollinator limited for several species was obtained by comparing the success of hand pollination and of natural pollination, rarity of certain specialized pollinators, and estimates of the abortion rates of fertilized ovules. We suggest that flowering in early spring is a high-risk option in terms of insect-mediated sexual reproduction. Certain flower-visiting insects favored D. laciniata out of proportion to its abundance, but no effect on seed set of other species was detectable. Honeybees were abundant and active flower visitors with the potential for disrupting ecological/evolutionary relationships between native insects and flowers.
DOIURL [本文引用: 3]
DOIURL [本文引用: 1]
Abstract Phenology is central to understanding vegetation response to climate change, as well as vegetation effects on plant resources, but most temporal production data is based on shoots, especially those of trees. In contrast, most production in temperate and colder regions is belowground, and is frequently dominated by grasses. We report root and shoot phenology in 7-year old monocultures of 10 dominant species (five woody species, five grasses) in southern Canada. Woody shoot production was greatest about 8 weeks before the peak of root production, whereas grass shoot maxima preceded root maxima by 2 4 weeks. Over the growing season, woody root, and grass root and shoot production increased significantly with soil temperature. In contrast, the timing of woody shoot production was not related to soil temperature ( r =0.01). The duration of root production was significantly greater than that of shoot production (grasses: 22%, woody species: 54%). Woody species produced cooler and moister soils than grasses, but growth forms did not affect seasonal patterns of soil conditions. Although woody shoots are the current benchmark for phenology studies, the other three components examined here (woody plant roots, grass shoots and roots) differed greatly in peak production time, as well as production duration. These results highlight that shoot and root phenology is not coincident, and further, that major plant growth forms differ in their timing of above- and belowground production. Thus, considering total plant phenology instead of only tree shoot phenology should provide a better understanding of ecosystem response to climate change.
DOIURLPMID [本文引用: 4]
We examined the effects of passive open-top warming chambers on Eriophorum vaginatum production near Toolik Lake, Alaska, USA. During the 2002 growing season, chamber warming was consistent with the magnitude and seasonality observed in recent decades throughout northwestern North America. Leaf-growth rates were higher in late May and early June; maximum growth rates in each leaf cohort occurred earlier and peak biomass was observed 2002days earlier within the chambers. Consequently, plants within the chambers maintained more live leaf biomass during the period of highest photosynthetically active radiation. Annual leaf production within the chambers (21±202mg02tiller) was not significantly different than under ambient conditions (17±202mg02tiller) ( P =0.2256) despite higher early-season growth rates. Root growth began earlier; growth rates were higher in late May and early June, and maximum growth rates occurred earlier within the chambers. Therefore, plants within the chambers maintained greater root biomass during what earlier studies have identified as a period of relatively high nutrient availability. Annual root production within the chambers (191±4202g02m 612 ) was not significantly different than under ambient conditions (119±4802g02m 612 ) ( P =0.1979), although there was a trend toward higher production within the chambers. The tendency toward higher root production within the chambers is consistent with previous laboratory experiments and with the predictions of biomass allocation theory.
URL [本文引用: 2]
本文以“3S”技术和数学统计软件为主要技术手段,根据1974-2004年30多年来发生的各类火灾的基础数据,绘制30年塔河林业局森林火灾火烧迹地分布图,结合该地区地理信息系统,将火点分布图与不同类型专题图相叠加,分析了各影响因子与塔河林业局林火的关系,并以该地区的不同时期发生不同火烧强度的火烧迹地为研究对象,研究不同林火强度对不同林型下植被更新及土壤环境的影响,并根据不同经营目标,选择合适的森林植被更新恢复模式,缩短火后森林生态系统恢复时间,提高森林恢复质量。研究结果如下: (1)方差分析结果表明,火烧年限对土壤物理性质如含水率,容重和孔隙度影响显著,而火烧强度对土壤含水率和容重的影响无明显的规律性,但各火烧迹地数据随火烧强度呈现出的变化趋势表明火烧可以使土壤结构变紧实。 (2)重度火烧对土壤有机质和速效磷含量的增加有一定的促进作用,并使土壤中水解氮的含量呈增加趋势,但随着年限的增加,速效磷的含量下降,速效钾的含量呈不规律变化;中度的火烧有利于土壤中全氮的增加,轻度火烧和中度火烧使土壤中水解氮的含量有所下降,土壤速效钾含量呈现逐年增加的趋势。经过不同年限和不同强度火烧后的铜、锌、铁、锰含量呈现出无规律的变化。但土壤中的铜和铁存在着一定的相关性,其含量经过不同强度的火烧后在各年限上所表现出来的变化趋势非常相似。 (3)不同林型乔木更新能力由大到小的顺序为:草类落叶松林落叶松白桦林杜香落叶松林杜鹃落叶松林坡地落叶松林杜鹃樟子松林;不同火烧强度对不同树种的更新有很大影响,大多数重火烧迹地将在很长一段时间内形成以杨桦为主的阔叶林;中度火烧初期更新最初形成以杨桦为优势树种的阔叶林,后逐渐演变为针阔混交林;轻度火烧有利于针叶林的更新。 (4)不同火烧强度对植被更新数量的影响在各林分类型有所不同。杜鹃落叶松林不同火烧强度下与为火烧前植被种类数量相差不大;落叶松白桦林林型与坡地落叶松林型火烧后植物种类明显增加;各林分类型中乔木层和灌木层盖度随火后恢复时间的推移呈升高的趋势,而草本层盖度则随时间的推移而呈下降的趋势;随着火烧强度的增大,高频物种所占的比例降低,植物分布的均匀性显著下降。 (5)不同火烧迹地应采取不同的森林的恢复途径:经营条件和立地条件较好的区域应该采用人工植苗更新;经营条件差,火烧面积较大的区域,森林更新恢复很大程度上要依赖于天然更新程度。 (6)针叶林遭轻度火烧后,林内环境变化不大;在中度火烧迹地上,针叶林的天然更新情况是比较差的,经过很长时间的恢复阔叶树将会占有很大的比例;重度火烧后的森林,必须加大人工促进更新的力度,缩短植被恢复所需要的时间。 (7)根据塔河林业局火烧迹地上5个主要林型:杜鹃落叶松林、草类落叶松林、杜香落叶松林、杜鹃樟子松林和落叶松白桦林林型特点,分析不同林分类型在不同火烧强度下植被及土壤理化性质的变化情况,提出了相应的更新措施和技术。 (8)火烧迹地上落叶松和樟子松人工更新现状调查结果表明,人工更新比天然更新对土壤结构、植被状况的改造具有更明显的效果,植被恢复时间更短。以商品林和生态公益林为不同的培育目标,提出塔河林业局火烧迹地人工更新技术,并提出了落叶松和樟子松两个人工更新主要树种的四种更新模式。 本文可从理论上完善国内在林火对植被恢复和土壤环境影响等内容。在实践中,可为合理有效的确定火后森林植被恢复模式,加速火后被破坏森林的恢复,制定有效的森林经营措施对策,提供可靠的科学依据。
URL [本文引用: 2]
本文以“3S”技术和数学统计软件为主要技术手段,根据1974-2004年30多年来发生的各类火灾的基础数据,绘制30年塔河林业局森林火灾火烧迹地分布图,结合该地区地理信息系统,将火点分布图与不同类型专题图相叠加,分析了各影响因子与塔河林业局林火的关系,并以该地区的不同时期发生不同火烧强度的火烧迹地为研究对象,研究不同林火强度对不同林型下植被更新及土壤环境的影响,并根据不同经营目标,选择合适的森林植被更新恢复模式,缩短火后森林生态系统恢复时间,提高森林恢复质量。研究结果如下: (1)方差分析结果表明,火烧年限对土壤物理性质如含水率,容重和孔隙度影响显著,而火烧强度对土壤含水率和容重的影响无明显的规律性,但各火烧迹地数据随火烧强度呈现出的变化趋势表明火烧可以使土壤结构变紧实。 (2)重度火烧对土壤有机质和速效磷含量的增加有一定的促进作用,并使土壤中水解氮的含量呈增加趋势,但随着年限的增加,速效磷的含量下降,速效钾的含量呈不规律变化;中度的火烧有利于土壤中全氮的增加,轻度火烧和中度火烧使土壤中水解氮的含量有所下降,土壤速效钾含量呈现逐年增加的趋势。经过不同年限和不同强度火烧后的铜、锌、铁、锰含量呈现出无规律的变化。但土壤中的铜和铁存在着一定的相关性,其含量经过不同强度的火烧后在各年限上所表现出来的变化趋势非常相似。 (3)不同林型乔木更新能力由大到小的顺序为:草类落叶松林落叶松白桦林杜香落叶松林杜鹃落叶松林坡地落叶松林杜鹃樟子松林;不同火烧强度对不同树种的更新有很大影响,大多数重火烧迹地将在很长一段时间内形成以杨桦为主的阔叶林;中度火烧初期更新最初形成以杨桦为优势树种的阔叶林,后逐渐演变为针阔混交林;轻度火烧有利于针叶林的更新。 (4)不同火烧强度对植被更新数量的影响在各林分类型有所不同。杜鹃落叶松林不同火烧强度下与为火烧前植被种类数量相差不大;落叶松白桦林林型与坡地落叶松林型火烧后植物种类明显增加;各林分类型中乔木层和灌木层盖度随火后恢复时间的推移呈升高的趋势,而草本层盖度则随时间的推移而呈下降的趋势;随着火烧强度的增大,高频物种所占的比例降低,植物分布的均匀性显著下降。 (5)不同火烧迹地应采取不同的森林的恢复途径:经营条件和立地条件较好的区域应该采用人工植苗更新;经营条件差,火烧面积较大的区域,森林更新恢复很大程度上要依赖于天然更新程度。 (6)针叶林遭轻度火烧后,林内环境变化不大;在中度火烧迹地上,针叶林的天然更新情况是比较差的,经过很长时间的恢复阔叶树将会占有很大的比例;重度火烧后的森林,必须加大人工促进更新的力度,缩短植被恢复所需要的时间。 (7)根据塔河林业局火烧迹地上5个主要林型:杜鹃落叶松林、草类落叶松林、杜香落叶松林、杜鹃樟子松林和落叶松白桦林林型特点,分析不同林分类型在不同火烧强度下植被及土壤理化性质的变化情况,提出了相应的更新措施和技术。 (8)火烧迹地上落叶松和樟子松人工更新现状调查结果表明,人工更新比天然更新对土壤结构、植被状况的改造具有更明显的效果,植被恢复时间更短。以商品林和生态公益林为不同的培育目标,提出塔河林业局火烧迹地人工更新技术,并提出了落叶松和樟子松两个人工更新主要树种的四种更新模式。 本文可从理论上完善国内在林火对植被恢复和土壤环境影响等内容。在实践中,可为合理有效的确定火后森林植被恢复模式,加速火后被破坏森林的恢复,制定有效的森林经营措施对策,提供可靠的科学依据。
DOIURLPMID
Abstract There is now ample evidence of the ecological impacts of recent climate change, from polar terrestrial to tropical marine environments. The responses of both flora and fauna span an array of ecosystems and organizational hierarchies, from the species to the community levels. Despite continued uncertainty as to community and ecosystem trajectories under global change, our review exposes a coherent pattern of ecological change across systems. Although we are only at an early stage in the projected trends of global warming, ecological responses to recent climate change are already clearly visible.
DOIURL [本文引用: 2]
Abstract Understanding how flowering phenology responds to warming and cooling (i.e., symmetric or asymmetric response) is needed to predict the response of flowering phenology to future climate change that will happen with the occurrence of warm and cold years superimposed upon a long-term trend. A three-year reciprocal translocation experiment was performed along an elevation gradient from 3200 m to 3800 m in the Tibetan Plateau for six alpine plants. Transplanting to lower elevation (warming) advanced the first flowering date (FFD) and transplanting to higher elevation (cooling) had the opposite effect. The FFD of early spring flowering plants (ESF) was four times less sensitive to warming than to cooling (by 612.1 d/°C and 8.4 d/°C, respectively), while midsummer flowering plants (MSF) were about twice as sensitive to warming than to cooling (618.0 d/°C and 4.9 d/°C, respectively). Compared with pooled warming and cooling data, warming alone significantly underpredicted 3.1 d/°C for ESF and overestimated 1.7 d/°C for MSF. These results suggest that future empirical and experimental studies should consider nonlinear temperature responses that can cause such warming–cooling asymmetries as well as differing life strategies (ESF vs. MSF) among plant species.
DOIURL [本文引用: 6]
Previous studies have focused on the effects of increased temperatures on a single green-up and/or flowering event, but less is known about how acceleration of spring phenology may change subsequent phenological events. We present results of a field experiment to test the hypotheses that (1) the timing of phenological events does not necessarily delay as elevation increases; (2) changes in the timing of a sequence of phenological events will be consistent for all phenological events along the elevation gradient; and thus (3) change in the timing of phenological events does not affect the duration of the entire reproductive stage in the alpine region. The experiment was conducted along an elevation gradient from 3200 to 380002m using two early-spring flowering (ESF) sedges and four mid-summer flowering (MSF) plants (two forbs and two grasses). Generally, our results only supported the first hypothesis. Lower elevation delayed the starting dates of all phenological events for ESF plants at 320002m compared with other elevations, whereas the opposite trend was observed for MSF-grasses. MSF-forbs had the earliest leaf-out at 320002m and the earliest first flowering at 360002m, and onset of fruit-set advanced with increasing elevation. The entire reproductive duration was shortened with increasing elevation for MSF-forbs, whereas it was the shortest for ESF at 360002m and for MSF-grasses at 3200 and/or 380002m. Individual reproductive stages had independent responses to climate change. The duration of the entire growing season for ESF plants decreased as elevation increased. For MSF-forbs, it was longest at 320002m and shortest at 340002m, while for MSF-grasses it was shortest at 320002m and at 380002m. Reproduction was compressed into shorter time periods only for MSF-forbs at 3600 and 380002m. Therefore, reproduction is not tightly integrated across the life cycle, and earlier reproductive development induced by warmer spring temperatures did not consistently advance flowering and fruiting times and their durations for the alpine plants studied. The effects of climate change on the timing and duration of phenological events were species-specific. Selection for changes in the timing and duration of individual phenological stages in response to climate change due to evolutionary adaptation should be taken into account.
URL [本文引用: 2]
采用由植被空间序列推断时间系列的方法 ,分析大兴安岭呼中林区近 2 0年来不同火烧迹地植被变化情况。结果表明 ,森林火后初始 ,植物尤其是草本物种迅速增多 ,火烧迹地的植被主要由草本所占据 ,但随着时间的推移 ,草本物种不仅在数量上有明显减少 ,在物种组成上有很大的变化 ,而且盖度也逐渐减少。灌木及乔木物种由于种类较少 ,在种类及组成上演替初期变化不如草本明显 ,但是盖度却逐渐增多。在本地区火后植被次生演替过程中 ,高频物种所占的比例较高且呈上升趋势 ,这明显有悖于Raunkiaer频度定律关于在相对稳定的群落中高频物种所占的比例应相对较低的规律。另外 ,不同火烧迹地间灌木的物种相似度明显高于草本的物种相似度 ,火烧年份相近的火烧迹地上的草本物种相似度要明显高于火烧年份远的草本物种相似度 ,但灌木间的物种相似度却没有出现这种现象 ,没有明显的规律。
URL [本文引用: 2]
采用由植被空间序列推断时间系列的方法 ,分析大兴安岭呼中林区近 2 0年来不同火烧迹地植被变化情况。结果表明 ,森林火后初始 ,植物尤其是草本物种迅速增多 ,火烧迹地的植被主要由草本所占据 ,但随着时间的推移 ,草本物种不仅在数量上有明显减少 ,在物种组成上有很大的变化 ,而且盖度也逐渐减少。灌木及乔木物种由于种类较少 ,在种类及组成上演替初期变化不如草本明显 ,但是盖度却逐渐增多。在本地区火后植被次生演替过程中 ,高频物种所占的比例较高且呈上升趋势 ,这明显有悖于Raunkiaer频度定律关于在相对稳定的群落中高频物种所占的比例应相对较低的规律。另外 ,不同火烧迹地间灌木的物种相似度明显高于草本的物种相似度 ,火烧年份相近的火烧迹地上的草本物种相似度要明显高于火烧年份远的草本物种相似度 ,但灌木间的物种相似度却没有出现这种现象 ,没有明显的规律。
DOIURLPMID [本文引用: 3]
Abstract Warming experiments are increasingly relied on to estimate plant responses to global climate change. For experiments to provide meaningful predictions of future responses, they should reflect the empirical record of responses to temperature variability and recent warming, including advances in the timing of flowering and leafing. We compared phenology (the timing of recurring life history events) in observational studies and warming experiments spanning four continents and 1,634 plant species using a common measure of temperature sensitivity (change in days per degree Celsius). We show that warming experiments underpredict advances in the timing of flowering and leafing by 8.5-fold and 4.0-fold, respectively, compared with long-term observations. For species that were common to both study types, the experimental results did not match the observational data in sign or magnitude. The observational data also showed that species that flower earliest in the spring have the highest temperature sensitivities, but this trend was not reflected in the experimental data. These significant mismatches seem to be unrelated to the study length or to the degree of manipulated warming in experiments. The discrepancy between experiments and observations, however, could arise from complex interactions among multiple drivers in the observational data, or it could arise from remediable artefacts in the experiments that result in lower irradiance and drier soils, thus dampening the phenological responses to manipulated warming. Our results introduce uncertainty into ecosystem models that are informed solely by experiments and suggest that responses to climate change that are predicted using such models should be re-evaluated.
URL [本文引用: 1]
为了解气候和物候变化规律,指导农业生产和环境监测,用线性倾向估计法分析了1982-2006年内蒙古 地区草本植物春季物候及其前期温度的变化趋势,并分区域分析了植物春季物候与温度的关系,通过逐步回归分别建立了中西部和东部地区植物始花期的温度回归模 型,通过模型对未来气候变化情景下内蒙古地区草本植物始花期变化进行了预估.结果表明:20世纪80年代以来植物始花期变化为提前趋势,温度变化为增温趋 势,春季变暖比冬季明显;温度和始花期的变化趋势均有明显的地域特征,中西部地区增温趋势和植物始花期提前趋势均大于东部地区,春季温度和植物始花期在两 区域平均变化趋势均显著,冬季温度在中西部地区变化显著,而在东部地区变化不显著;植物始花期与其前期温度呈明显的负相关,春季温度是影响开花的主要因 子,未来如温度上升1 ℃,始花期提早3.1~5.0 d.
URL [本文引用: 1]
为了解气候和物候变化规律,指导农业生产和环境监测,用线性倾向估计法分析了1982-2006年内蒙古 地区草本植物春季物候及其前期温度的变化趋势,并分区域分析了植物春季物候与温度的关系,通过逐步回归分别建立了中西部和东部地区植物始花期的温度回归模 型,通过模型对未来气候变化情景下内蒙古地区草本植物始花期变化进行了预估.结果表明:20世纪80年代以来植物始花期变化为提前趋势,温度变化为增温趋 势,春季变暖比冬季明显;温度和始花期的变化趋势均有明显的地域特征,中西部地区增温趋势和植物始花期提前趋势均大于东部地区,春季温度和植物始花期在两 区域平均变化趋势均显著,冬季温度在中西部地区变化显著,而在东部地区变化不显著;植物始花期与其前期温度呈明显的负相关,春季温度是影响开花的主要因 子,未来如温度上升1 ℃,始花期提早3.1~5.0 d.
DOIURL [本文引用: 2]
植物物候特征对环境条件的季节和年际变化具有较强的指示作用,因此研究植物物候特征对环境条件变化的响应,对理解植物和环境之间的相互作用关系、植物的适应机制和生存策略,以及应对全球变化都具有重要的意义。该研究基于2009-2011年高寒矮生嵩草(Kobresiahumilis)草甸养分水分控制实验的植物物候观测数据资料,采用巢式方差分析、物候指数和聚类分析方法,开展了高寒矮生嵩草草甸主要植物物候特征对养分和水分添加的响应研究。结果表明:(1)养分添加处理之间植物返青期和枯黄期均无显著差异,但养分添加中氮磷处理对主要物种作用较明显,使莎草科、禾本科、杂类草主要代表植物的返青期和枯黄期推迟。(2)增雪处理效应明显,主要优势物种无论是何种养分添加,在增雪处理后均表现出花期物候提前的趋势(p〈0.01),同时增雪处理使杂类草植物返青期显著提前p〈0.05)。增水处理对植物的作用效果并不一致,其中垂穗披碱草(Elymusnutans)和双柱头蔗草(Scirpusdistigmaticus)的枯黄期显著推迟0〈0.05),而杂类草枯黄期提前。(3)养分添加后,不同物种的物候特征表现出显著差异(p〈0.01),例如雪白委陵菜(Potentillanivea)枯黄期显著推迟p〈0.05),而双柱头蔗草的枯黄期显著提前p〈0.05),但物种对养分添加响应的差异以植物类群为单位,禾本科植物表现为返青期推迟,而莎草科植物表现为返青期提前。(4)矮生嵩草草甸主要植物营养生长期与果后营养期持续天数之间呈负相关关系,主要植物物候特征经聚类分析可以分为3个类群,3个类群经氮磷钾、钾和氮钾三个养分添加处理后植物物候特征变化较大。研究表明,高寒矮生嵩草草甸植物物候特征在物种水平响应和水分添加后的响应表现出较大差异,
DOIURL [本文引用: 2]
植物物候特征对环境条件的季节和年际变化具有较强的指示作用,因此研究植物物候特征对环境条件变化的响应,对理解植物和环境之间的相互作用关系、植物的适应机制和生存策略,以及应对全球变化都具有重要的意义。该研究基于2009-2011年高寒矮生嵩草(Kobresiahumilis)草甸养分水分控制实验的植物物候观测数据资料,采用巢式方差分析、物候指数和聚类分析方法,开展了高寒矮生嵩草草甸主要植物物候特征对养分和水分添加的响应研究。结果表明:(1)养分添加处理之间植物返青期和枯黄期均无显著差异,但养分添加中氮磷处理对主要物种作用较明显,使莎草科、禾本科、杂类草主要代表植物的返青期和枯黄期推迟。(2)增雪处理效应明显,主要优势物种无论是何种养分添加,在增雪处理后均表现出花期物候提前的趋势(p〈0.01),同时增雪处理使杂类草植物返青期显著提前p〈0.05)。增水处理对植物的作用效果并不一致,其中垂穗披碱草(Elymusnutans)和双柱头蔗草(Scirpusdistigmaticus)的枯黄期显著推迟0〈0.05),而杂类草枯黄期提前。(3)养分添加后,不同物种的物候特征表现出显著差异(p〈0.01),例如雪白委陵菜(Potentillanivea)枯黄期显著推迟p〈0.05),而双柱头蔗草的枯黄期显著提前p〈0.05),但物种对养分添加响应的差异以植物类群为单位,禾本科植物表现为返青期推迟,而莎草科植物表现为返青期提前。(4)矮生嵩草草甸主要植物营养生长期与果后营养期持续天数之间呈负相关关系,主要植物物候特征经聚类分析可以分为3个类群,3个类群经氮磷钾、钾和氮钾三个养分添加处理后植物物候特征变化较大。研究表明,高寒矮生嵩草草甸植物物候特征在物种水平响应和水分添加后的响应表现出较大差异,
DOIURL
Understanding both historic and current fire regimes is indispensable to sustainable forest landscape management. In this paper, we use a spatially explicit landscape simulation model, LANDIS, to simulate historic and current fire regimes in the Great Xing n Mountains, in northeastern China. We analyzed fire frequency, fire size, fire intensity, and spatial pattern of burnt patches. Our simulated results show that fire frequency under the current fire scenario is lower than under the historic fire scenario; total area burnt is larger with lower fire intensity under the historic fire scenario, and smaller with higher fire intensity under the current fire scenario. We also found most areas were burned by high intensity fires under the current fire scenario, but by low to moderate fires under the historic fire scenario. Burnt patches exhibit a different pattern between the two simulation scenarios. Large patches burnt by high intensity class fires dominate the landscape under the current fire scenario, and under historic fire scenario, patches burnt by low to moderate fire intensity fires have relatively larger size than those burnt by high intensity fires. Based on these simulated results, we suggest that prescribed burning or coarse woody debris reduction should be incorporated into forest management plans in this region, especially on north-facing slopes. Tree planting may be a better management option on these severely burned areas whereas prescribed burning after small area selective cutting, retaining dispersed seed trees, may be a sound forest management alternative in areas except for the severely burned patches.
Biomass offsets little or none of permafrost carbon release from soils, streams, and wildfire: An expert assessment
2
2016
... 气候变暖已成为不争的事实, 且在高纬度冻土区表现尤为明显(
... 由于气候变暖使得火干扰对泰加林的影响进一步增强, 将会有更大比例的泰加林变成火烧迹地, 处于演替初级阶段(
Community assembly, niche conservatism, and adaptive evolution in changing environments
2003
A 250-year index of first flowering dates and its response to temperature changes
1
2010
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
Phenotypic plasticity and adaptive evolution contribute to advancing flowering phenology in response to climate change
1
2012
... 不同物种具有不同的繁殖物候期, 这是物种长期进化过程中不断适应环境的结果, 也是植物降低种间竞争并维持群落水平物种共存的重要机制(
Environmental regulation of flowering
1
2005
... 温度可以直接控制生物物候事件开始的时间, 被称为生物原始的“生物钟” (
Responses of spring phenology to climate change
3
2004
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... ).物候与环境因子之间的关系具有重要的生态意义和进化意义.已有研究普遍表明, 气候变暖使得植物春季物候提前, 秋季物候延迟, 生长季时间延长(
... 由于气候变暖使得火干扰对泰加林的影响进一步增强, 将会有更大比例的泰加林变成火烧迹地, 处于演替初级阶段(
Functional equilibrium: Sense or nonsense
1
1983
... 温度可以直接控制生物物候事件开始的时间, 被称为生物原始的“生物钟” (
Interactive effects of wildfire and climate on permafrost degradation in Alaskan lowland forests
2
2015
... 气候变暖已成为不争的事实, 且在高纬度冻土区表现尤为明显(
... 由于气候变暖使得火干扰对泰加林的影响进一步增强, 将会有更大比例的泰加林变成火烧迹地, 处于演替初级阶段(
Shifts in flowering phenology reshape a subalpine plant community
1
2014
... 不同物种具有不同的繁殖物候期, 这是物种长期进化过程中不断适应环境的结果, 也是植物降低种间竞争并维持群落水平物种共存的重要机制(
Phenological tracking enables positive species responses to climate change
1
2012
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
Diverse responses of phenology to global changes in a grassland ecosystem
1
2006
... 不同物种具有不同的繁殖物候期, 这是物种长期进化过程中不断适应环境的结果, 也是植物降低种间竞争并维持群落水平物种共存的重要机制(
Shifting plant phenology in response to global change
1
2007
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
Plant functional traits mediate reproductive phenology and success in response to experimental warming and snow addition in Tibet
2
2013
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... 不同物种具有不同的繁殖物候期, 这是物种长期进化过程中不断适应环境的结果, 也是植物降低种间竞争并维持群落水平物种共存的重要机制(
Coexistence of similar species in a space-limited intertidal zone
1
1999
... 不同物种具有不同的繁殖物候期, 这是物种长期进化过程中不断适应环境的结果, 也是植物降低种间竞争并维持群落水平物种共存的重要机制(
Toward a synthetic understanding of the role of phenology in ecology and evolution
2
2010
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... 温度可以直接控制生物物候事件开始的时间, 被称为生物原始的“生物钟” (
Are flowers physiolosical sinks or faucets? Costs ans correlates of water use by flowers of Polemonium viscosum
2
1999
... 温度可以直接控制生物物候事件开始的时间, 被称为生物原始的“生物钟” (
... ).同时, 低水分状况可以减缓花芽和结实的过程(
Lower plasticity exhibited by high-versus mid-elevation species in their phenological responses to manipulated temperature and drought
1
2015
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
Contrasting impacts of climate-driven flowering phenology on changes in alien and native plant species distributions
1
2011
... 本研究中, 杜香是灌草层的优势物种, 其生物量占群落总生物量(高度<50 cm的灌草层)的(46.0 ± 4.2)%.在增温和土壤水分降低的气候条件下, 属于中花植物的杜香开花峰值期显著提前, 且花期呈延长趋势, 开花数量呈增加趋势.更长的花期使得杜香的花期与其传粉动物的活动期不一致的可能性降低, 增加了杜香的繁殖成功率.延长的花期和增加的开花数量还可以增加配型, 使基因多样化, 利于杜香的进化.同时, 植物个体开花物候对环境变化的响应与其生长性状紧密相连.有研究表明, 在增温条件下, 物候提前较多的物种, 在丰富度和生态幅上有较大的提高, 反之, 其性状特征(生物量、相对盖度、开花数目)则相对降低(
The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change
2
2013
... 气候变暖已成为不争的事实, 且在高纬度冻土区表现尤为明显(
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
Permafrost degradation and ecological changes associated with a warming climate in central Alaska
1
2001
... 气候变暖已成为不争的事实, 且在高纬度冻土区表现尤为明显(
Effect of forest fire on the fluxes of CO2, CH4 and N2O in boreal forest soils, interior Alaska
2003
Constraints and competition in the evolution of flowering phenology
1
1986
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
Responses of sequential and hierarchical phenological events to warming and cooling in alpine meadows
2
2016
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... 由于普遍的认识是早花植物比晚花植物对气候变暖更加敏感(
Changes in phenological sequences of alpine communities across a natural elevation gradient
2
2016
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... 不同物种具有不同的繁殖物候期, 这是物种长期进化过程中不断适应环境的结果, 也是植物降低种间竞争并维持群落水平物种共存的重要机制(
Changes in flowering functional group affect responses of community phenological sequences to temperature change
1
2017
... 不同物种具有不同的繁殖物候期, 这是物种长期进化过程中不断适应环境的结果, 也是植物降低种间竞争并维持群落水平物种共存的重要机制(
Effects of recent warm and cold spells on European plant phenology
2
2011
... 温度可以直接控制生物物候事件开始的时间, 被称为生物原始的“生物钟” (
... ).干旱一般会通过缩短植物花期来减少因开花而增加的水分丧失量(
Phenology: Reapones to a warming world
1
2001
... 植物生长变化是群落响应气候变化的又一重要途径(
Changed plant and animal life cycles from 1952 to 2000 in the Mediterranean region
2
2002
... 植物生长变化是群落响应气候变化的又一重要途径(
... ).不同物种的生长对气候变化的响应与其生长特性及其对环境的适应性有关(
Response of plant species richness and primary productivity in shrublands along a north-south gradient in Europe to seven years of experimental warming and drought: Reductions in primary productivity in the heat and drought year of 2003
1
2007
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
Phenology feedbacks on climate change
3
2009
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... ), 通过研究单个发育阶段来描述繁殖物候对气候变暖的响应存在局限性(
... 由于普遍的认识是早花植物比晚花植物对气候变暖更加敏感(
Phenological sequences reveal aggregate life history response to climatic warming
2008
Flowering ecology of some spring woodland herbs
6
1978
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... 根据样方内物种始花期的时间分布, 选择不同开花功能群(
... 不同物种具有不同的繁殖物候期, 这是物种长期进化过程中不断适应环境的结果, 也是植物降低种间竞争并维持群落水平物种共存的重要机制(
... ),增温使其温度积累提前达到要求从而提前植物始花期.同时,
... 由于普遍的认识是早花植物比晚花植物对气候变暖更加敏感(
... ).晚花植物的齿叶风毛菊更长的营养生长时间及花蕾阶段可使其花期延后.同时, 增温和排水处理缩短了晚花植物齿叶风毛菊的开花持续时间, 这可能对其遗传变异及进化适应产生选择压力(
Divergence of reproductive phenology under climate warming
3
2007
... 不同物种具有不同的繁殖物候期, 这是物种长期进化过程中不断适应环境的结果, 也是植物降低种间竞争并维持群落水平物种共存的重要机制(
... 由于普遍的认识是早花植物比晚花植物对气候变暖更加敏感(
... 温度可以直接控制生物物候事件开始的时间, 被称为生物原始的“生物钟” (
Asynchronicity in root and shoot phenology in grass and woody plants
1
2010
... 温度可以直接控制生物物候事件开始的时间, 被称为生物原始的“生物钟” (
Warming chambers stimulate early season growth of an arctic sedge: Results of a minirhizotron field study
4
2005
... 由于气候变暖使得火干扰对泰加林的影响进一步增强, 将会有更大比例的泰加林变成火烧迹地, 处于演替初级阶段(
... 根据
... ;
... ).因此, 我们可以在一定程度上推测, 短期内增温和土壤水分降低可能加速演替初期泰加林灌草层的火后演替进程.由于高频物种所占比例的增加是有利于群落稳定发展的, 能为乔木的更新创造良好的生态环境(
塔河林区林火对土壤性质与植被恢复的影响
2
2011
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... 根据
塔河林区林火对土壤性质与植被恢复的影响
2
2011
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... 根据
Ecological responses to recent climate change
2002
Asymmetric sensitivity of first flowering date to warming and cooling in alpine plants
2
2014
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... 不同物种具有不同的繁殖物候期, 这是物种长期进化过程中不断适应环境的结果, 也是植物降低种间竞争并维持群落水平物种共存的重要机制(
Timing and duration of phenological sequences of alpine plants along an elevation gradient on the Tibetan Plateau
6
2014
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... 由于气候变暖使得火干扰对泰加林的影响进一步增强, 将会有更大比例的泰加林变成火烧迹地, 处于演替初级阶段(
... 温度可以直接控制生物物候事件开始的时间, 被称为生物原始的“生物钟” (
... ), 但也研究得出相反的结果(
... 根据
... ).短期来看, 模拟增温和土壤水分降低促使作为优势物种的杜香更具繁殖优势, 且占有更宽的生态幅, 使其在灌草层的优势地位更加明显, 这与泰加林火后恢复中灌草层的演替路径非常一致(
大兴安岭北坡落叶松林火后植被演替过程研究
2
2004
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... 温度可以直接控制生物物候事件开始的时间, 被称为生物原始的“生物钟” (
大兴安岭北坡落叶松林火后植被演替过程研究
2
2004
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... 温度可以直接控制生物物候事件开始的时间, 被称为生物原始的“生物钟” (
Warming experiments underpredict plant phenological responses to climate change
3
2012
... 物候是研究生态系统响应气候变化机制的重要途径, 且对气候变化响应迅速, 被称为气候变化的“指纹” (
... 不同物种具有不同的繁殖物候期, 这是物种长期进化过程中不断适应环境的结果, 也是植物降低种间竞争并维持群落水平物种共存的重要机制(
... ).植物始花期受控于其前期温度积累(
内蒙古典型草本植物春季物候变化及其对气候变暖的响应
1
2009
... 由于普遍的认识是早花植物比晚花植物对气候变暖更加敏感(
内蒙古典型草本植物春季物候变化及其对气候变暖的响应
1
2009
... 由于普遍的认识是早花植物比晚花植物对气候变暖更加敏感(
高寒矮生嵩草草甸主要植物物候特征对养分和水分添加的响应
2
2014
... 气候变暖已成为不争的事实, 且在高纬度冻土区表现尤为明显(
... 由于气候变暖使得火干扰对泰加林的影响进一步增强, 将会有更大比例的泰加林变成火烧迹地, 处于演替初级阶段(
高寒矮生嵩草草甸主要植物物候特征对养分和水分添加的响应
2
2014
... 气候变暖已成为不争的事实, 且在高纬度冻土区表现尤为明显(
... 由于气候变暖使得火干扰对泰加林的影响进一步增强, 将会有更大比例的泰加林变成火烧迹地, 处于演替初级阶段(
Historic and current fire regimes in the Great Xing’an Mountains, northeastern China: Implications for long-term forest management
2008

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}