 ,2*, 曾辉,1,2*1
,2*, 曾辉,1,2*1 2
Effects of nitrogen addition on the plant diversity and biomass of degraded grasslands of Nei Mongol, China
YANG Qian1,2, WANG Wei,2*, ZENG Hui,1,2*1 2
通讯作者: (Wang W,wangw@urban.pku.edu.cn; Zeng H,zengh@pkusz.edu.cn)
编委: 梁存柱
责任编辑: 王葳
| 基金资助: |
Online:2018-04-20
| Fund supported: |

摘要
人为干扰及气候变化导致内蒙古草地发生了大面积退化, 氮添加是促进退化草地生产力恢复的一项重要措施。该文基于2011年建立的氮肥添加实验平台, 以3个不同退化程度(中度退化、重度退化、极度退化)草地群落为研究对象, 设置对照、10、20、30、40和50 g·m -2·a -1 6种氮添加处理, 分析氮添加对退化草地恢复过程中群落多样性和生物量的影响。结果表明: (1)氮添加降低了中度、重度退化草地恢复进程中物种丰富度和多样性, 对极度退化草地恢复进程中物种丰富度和多样性无明显影响。(2)氮添加促进了3个不同退化程度草地恢复进程中群落地上生物量的增加。(3)氮添加显著增加了群落中禾草的地上生物量及其在群落地上生物量中所占的比例, 降低了杂类草在群落地上生物量中的比例, 但对杂类草地上生物量无显著影响。研究表明在利用施肥措施治理退化草地的过程中, 需要充分考虑草地退化程度以及由氮添加引起的群落多样性和生产力的改变对草地生态系统功能的影响。
关键词:
Abstract
Keywords:
PDF (6352KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
杨倩, 王娓, 曾辉. 氮添加对内蒙古退化草地植物群落多样性和生物量的影响. 植物生态学报[J], 2018, 42(4): 430-441 DOI:10.17521/cjpe.2017.0135
YANG Qian, WANG Wei, ZENG Hui.
草地是全球分布最广的一个植被类型(Scurlock & Hall, 1998), 其面积约占全球陆地总面积的25% (Xu et al., 2012)。草地生态系统在支撑畜牧业生产、防风固沙、水土保持、涵养水源、生物多样性保护, 以及陆地生态系统碳循环中占据着重要地位(沈海花等, 2016)。内蒙古草地是欧亚草原的重要组成部分(Bai et al., 2010), 是我国北方重要的生态安全屏障。但是近年来, 由于人为活动和气候变化等因素的影响, 内蒙古草地发生了大规模的退化(Quan et al., 2015), 致使草地生态系统生态服务功能降低, 如生物多样性减少、生产力水平降低、水土流失加剧(Asner et al., 2004; Kang et al., 2007; 刘碧荣等, 2015), 从而对区域环境、社会和经济的可持续发展构成了严重威胁。因此, 制定适宜的管理措施促进退化草地的恢复是草地生态学亟待解决的重点问题。
施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(李禄军等, 2010)。国内外广泛开展的草地施肥实验表明, 氮添加在促进植物地上部分生长、增加地上生物量(Bai et al., 2010; Stevens et al., 2015; Harpole et al., 2016; 李春丽等, 2016)的同时, 也会降低生态系统物种多样性(Stevens et al., 2004; Isbell et al., 2013; Niu et al., 2017)。Zhang等(2014)在内蒙古温带草原的研究表明, 氮添加降低了物种丰富度, 其中, 对于特定的氮添加速率, 低频率、大量施氮致使物种更多地丧失。Clark和Tilman (2008)在美国明尼苏达草原为期23年的施肥实验表明, 即使在低水平氮添加速率下, 群落中的稀有种仍会丧失。物种丧失降低了群落多样性, 并影响了生态系统稳定性(Yang et al., 2012; Song & Yu, 2015; 王晶等, 2016)和生态系统功能(Bai et al., 2010; Yang et al., 2012)。草地生态系统生产力的维持和可持续性在很大程度上依赖于物种多样性(Tilman et al., 1996; Fang et al., 2012)。因此, 了解氮添加对草地植物群落的影响规律, 揭示氮添加降低生物多样性的机制, 对于科学管理草地具有重要的意义。目前, 已有相关研究从土壤属性、种间竞争等方面对氮添加导致多样性降低的机制进行了探讨。例如: Zhang等(2014)研究表明, 氮添加导致物种丧失是由于土壤酸化和NH4+毒害的共同作用; Hautier等(2009)研究表明, 氮添加导致植物地上部分对光资源的竞争加剧, 从而使物种多样性降低; Rajaniemi等(2003)提出地下竞争假说来解释因氮添加而造成的物种多样性的下降。
全球范围内, 有关氮添加对草地生态系统影响以及氮添加降低物种多样性机制的报道中, 只有少量实验是从退化草地恢复的角度出发(Mountford et al., 1993; Stevens, 2016)。相关研究表明, 退化草地在一定程度上受氮限制(Hooper & Johnson, 1999; 徐晓天, 2015), 草地施肥是促进退化草地恢复以及草地资源可持续利用的有效途径(张铜会等, 2008; 王晶等, 2016)。王晶等(2016)研究表明, 中高水平氮添加提高了轻度退化草地生产力, 对中度和重度退化草地恢复并无明显的促进作用; 且氮添加对3个不同退化程度草地的物种丰富度均无显著影响。Xu等(2015)在内蒙古5个不同退化梯度草地3年氮添加的实验表明, 氮添加对5个不同退化程度草地的物种丰富度均无显著影响, 而对地上生物量的影响与退化程度有关, 其中退化最严重的草地地上生物量增加最为明显。Bai等(2010)研究表明, 在成熟草地, 氮添加会增加地上生物量, 降低物种丰富度, 其中氮输入导致多年生禾草和多年生杂类草丧失, 提高了一年生草本的物种丰富度; 对于退化草地, 物种丰富度年际变化对氮添加的响应并不明显。由此可见, 在不同退化程度的草地, 氮添加对群落物种多样性和地上生物量影响的研究结论并不完全一致。
本研究在内蒙古设置中度、重度、极度3个不同退化程度草地氮添加实验, 拟解决以下3个科学问题: (1)氮添加如何影响不同退化草地恢复过程中物种多样性、植物功能群以及群落地上生物量? (2)在退化草地恢复过程中, 氮添加对草地多样性和生物量的影响是否随退化状况的不同而异? (3)氮添加对退化草地物种多样性影响的机制。本项研究可为退化草地恢复与科学管理提供理论依据。
1 材料和方法
1.1 研究地自然概况
本研究实验样地位于内蒙古自治区克什克腾旗南端的乌兰布统草原(图1)。研究区年平均气温-1.4 ℃, 年降水量350-400 mm。降水主要集中在5-8月, 约占全年降水量的76%, 其中7、8月份的降水量占全年降水量的51%, 10月到次年4月的降水形式主要是降雪。研究区主要土壤类型为栗钙土, 表层土壤质地主要为粉砂和砂(Liu et al., 2008)。研究区域主要物种有羊草(Leymus chinensis)、贝加尔针茅(Stipa baicalensis)、硬质早熟禾(Poa sphondylodes)、黄囊薹草(Carex korshinskyi)、腺毛委陵菜(Potentilla longifolia)、糙隐子草(Cleistogenes squarrosa)、冷蒿(Artemisia frigida)等。1.2 实验设计
以往的研究表明, 植被退化与土壤退化是密切关联的(van de Koppel et al., 1997), 植被退化表现为群落覆盖度下降和生产力降低(Su et al., 2005), 生态系统承载力、稳定性和服务功能下降, 优良牧草的种类和比例减少, 劣质牧草及有害毒草比例增加等(穆少杰等, 2017)。土壤退化主要表现为水土流失、土壤质地逆向改变、土壤肥力下降以及动物和微生物活性下降等(Su et al., 2005; 穆少杰等, 2017)。因此, 植被物种组成和土壤属性可以反映草地的退化状况。本研究将实验样地的植物划分为一年生植物、退化指示种和顶极种3个类群。顶极种指原始类型的建群种, 如我国北方草甸草原有羊草、贝加尔针茅等原始建群种, 并伴生无芒雀麦(Bromus inermis)、拂子茅(Calamagrostis epigeios)等中生优良牧草。退化指示种指冷蒿、糙隐子草、星毛委陵菜(Potentilla acaulis)、狼毒(Stellera chamaejasme)等适口性差或有毒植物, 这些物种随着退化程度增加而增加, 甚至会代替原来的优势植物而成为优势种(Liu et al., 2008)。一年生植物主要有藜(Chenopodium acuminatum)、狗尾草(Setaria viridis)、沙蓬(Agriophyllum squarrosum)等植物。根据3个植物类群的相对盖度(Liu et al., 2008)以及土壤质地和土壤养分含量, 建立草地退化评估体系。根据退化评估体系, 计算3个样地草地退化指数(GDI), 将实验样地划分为中度退化草地、重度退化草地和极度退化草地(表1)。GDI = (P1 × 1/3 + P2 × 2/3 + P3) × 1/3 + (STC × 1/2 + STN × 1/2) × 1/3 + (1/Sand) × 1/3
其中, P1、P2、P3分别代表一年生植物、退化指示种和顶极种的相对盖度, STC、STN分别代表土壤全碳含量和土壤全氮含量, Sand表示土壤砂粒含量。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1研究区域及样地分布图。EDG, 极度退化草地; MDG, 中度退化草地; SDG, 重度退化草地。
Fig. 1Study area and the spatial distribution of study sites. EDG, extremely degraded grassland; MDG, moderately degraded grassland; SDG, severely degraded grassland.
Table 1
表1
表1实验样地的土壤和植被基本属性
Table 1
| 名称 Term | 极度退化草地 EDG | 重度退化草地 SDG | 中度退化草地 MDG |
|---|---|---|---|
| 物种组成 Species composition | 羊草、黄囊薹草、马唐、狗尾草、沙蓬藜等 Leymus chinensis; Carex korshinskyi; Digitaria sanguinalis; Setaria viridis; Agriophyllum squarrosum; Chenopodium acuminatum et al. | 拂子茅、贝加尔针茅、硬质早熟禾、黄囊薹草、腺毛委陵菜、星毛委陵菜、冷蒿、紫羊茅、冰草、狼毒、糙隐子草、阿尔泰狗娃花等 Calamagrostis epigeios; Stipa baicalensis; Poa sphondylodes; Carex korshinskyi; Potentilla longifolia; Potentilla acaulis; Artemisia frigida; Festuca rubra; Agropyron cristatum; Stellera chamaejasme; Cleistogenes squarrosa; Heteropappus altaicus et al. | 拂子茅、贝加尔针茅、羊草、硬质早熟禾、腺毛委陵菜、黄囊薹草、冷蒿、紫羊茅、冰草、糙隐子草、阴山胡枝子、阿尔泰狗娃花等 Calamagrostis epigeios; Stipa baicalensis; Leymus chinensis; Poa sphondylodes; Potentilla longifolia; Carex korshinskyi; Artemisia frigida; Festuca rubra; Agropyron cristatum; Cleistogenes squarrosa; Lespedeza inschanica; Heteropappus altaicus et al. |
| 顶极种相对盖度 Relative coverage of climax species (%) | 34.48 | 39.53 | 54.05 |
| 退化指示种相对盖度 Relative coverage of degradation indicators (%) | 34.48 | 32.56 | 29.73 |
| 一年生植物相对盖度 Relative coverage of annuals (%) | 31.04 | 27.91 | 16.22 |
| 土壤全碳 Soil total carbon (%) | 0.83 | 1.67 | 2.06 |
| 土壤全氮 Soil total nitrogen (%) | 0.06 | 0.14 | 0.17 |
| 砂粒含量 Sand content (%) | 66.10 | 57.30 | 58.40 |
| 草地退化指数 Grassland degradation index | 0.379 | 0.543 | 0.642 |
新窗口打开|下载CSV
复性实验样地于2011年春季建立并开始实施实验。每个退化样地采用完全随机设计方式进行布置, 共6个氮添加处理: 0 (对照)、10、20、30、40和50 g·m-2·a-1, 每个处理3个重复样方, 每个样方面积为6 m × 6 m, 共计18个样方, 每个样方之间设置1 m的缓冲区。3个退化梯度样地共计54个样方。
实验中所使用的氮肥为尿素, 每年施肥4次, 分别于5月、6月、7月、8月初进行, 每次施肥量为全年施肥总量的1/4。为保证施肥效果, 选择在阴雨天进行氮肥添加, 将预先称好的尿素均匀撒在样方内, 对照组不采取任何处理措施。
1.3 样品的采集与测定
于2016年生长季旺盛期(8月中下旬), 在每个实验样方内随机放置一个1 m × 1 m的样方, 进行群落调查, 为了避免边缘效应, 样方距边缘的距离大于50 cm, 然后测定样方内物种数, 以及每个物种的高度、盖度和个体数。地上生物量采用收获法测定。在每个实验样方内随机设置0.5 m × 0.5 m的小样方, 然后将样方内所有植物齐地收割, 带回实验室按照物种归类后装入信封, 置于65 ℃烘箱烘至恒质量。称量各物种的质量, 累加后计算单位面积内群落地上生物量。本研究根据3个不同退化程度草地物种组成, 将植物分为禾草和杂类草2个功能群, 计算各功能群生物量及其在群落总地上生物量中所占的比例。物种丰富度指单位面积(1 m2)样方内出现的物种数。物种多样性用Shannon-Wiener多样性指数(H)进行测度, 其计算公式为:
\(H=\sum^n_{i=1}\left(\frac{b_i}{\sum^n_{i=1}b_i}\times 100\right)\times ln\left(\frac{b_i}{\sum^n_{i=1}b_i}\times 100\right)\)
式中n是样方内物种数, bi是物种i的地上生物量(Zhang et al., 2014)。
通过计算生物量响应比率(处理组地上生物量与对照组地上生物量的比值), 来直接比较地上生物量对氮添加处理的响应。植物高度不对称性指数反映了物种对光的竞争能力, 使用株高的基尼系数(Gini)衡量功能群之间大小的不平等(Ren et al., 2010)。
$G=\frac{\sum\limits_{i=1}^{n}{\sum\limits_{j=1}^{n}{|{{X}_{i}}-{{X}_{j}}|}}}{2{{n}^{2}}\bar{X}}$
上式中, Xi和Xj分别表示功能群i和j植株的平均高度, n表示功能群数量, Xˉ表示所有功能群植株的平均高度。
1.4 统计分析
采用Microsoft Excel 2007整理数据, 统计分析均在SPSS 19.0中完成。用双因素方差分析将退化梯度和氮添加处理作为固定因子, 分析各因子单独及交互作用对群落物种丰富度、物种多样性、地上生物量、各功能群地上生物量及其在群落地上生物量中所占的比例的影响。同一退化类型处理采用单因素方差分析, Duncan’s多重比较的方法进行差异显著性检验(p = 0.05)。以氮添加量为连续变量, 对物种丰富度和地上生物量进行回归参数估计, 以此确定氮添加量与物种丰富度、多样性和地上生物量之间的一般关系。作图在SigmaPlot 10.0中完成。2 结果和分析
2.1 不同退化草地恢复进程中氮添加对群落物种丰富度和多样性的影响
氮添加和退化梯度对群落物种丰富度和多样性有显著影响, 氮添加和退化类型的交互作用对物种丰富度有显著影响, 对多样性无显著影响(表2)。氮添加显著降低了中度和重度退化草地恢复进程中群落物种丰富度和多样性, 但对极度退化草地恢复进程中群落物种丰富度和多样性无显著性影响(图2)。其中, 在中度退化草地, 施氮量为40、50 g·m-2·a-1, 物种丰富度明显降低; 在重度退化草地施氮量为30 g·m-2·a-1, 物种丰富度与对照组相比明显降低(图2A)。中度退化草地施氮量为40 g·m-2·a-1, 多样性降低, 重度退化草地施氮量为20 g·m-2·a-1, 多样性降低(图2B)。Table 2
表2
表2氮添加处理、退化类型以及它们的交互作用对群落物种丰富度、多样性和地上生物量的双因素方差分析
Table 2
| 响应 Response | 名称 Term | df | F | p |
|---|---|---|---|---|
| 物种丰富度 Species richness | DT | 2 | 40.64 | <0.001 |
| N | 5 | 8.50 | <0.001 | |
| DT × N | 10 | 2.27 | 0.035 | |
| Shannon-Wiener指数 Shannon-Wiener index | DT | 2 | 12.84 | <0.001 |
| N | 5 | 5.06 | 0.001 | |
| DT × N | 10 | 1.50 | 0.179 | |
| 地上生物量 Aboveground biomass | DT | 2 | 42.08 | <0.001 |
| N | 5 | 9.53 | <0.001 | |
| DT×N | 10 | 1.30 | 0.264 |
新窗口打开|下载CSV
图2
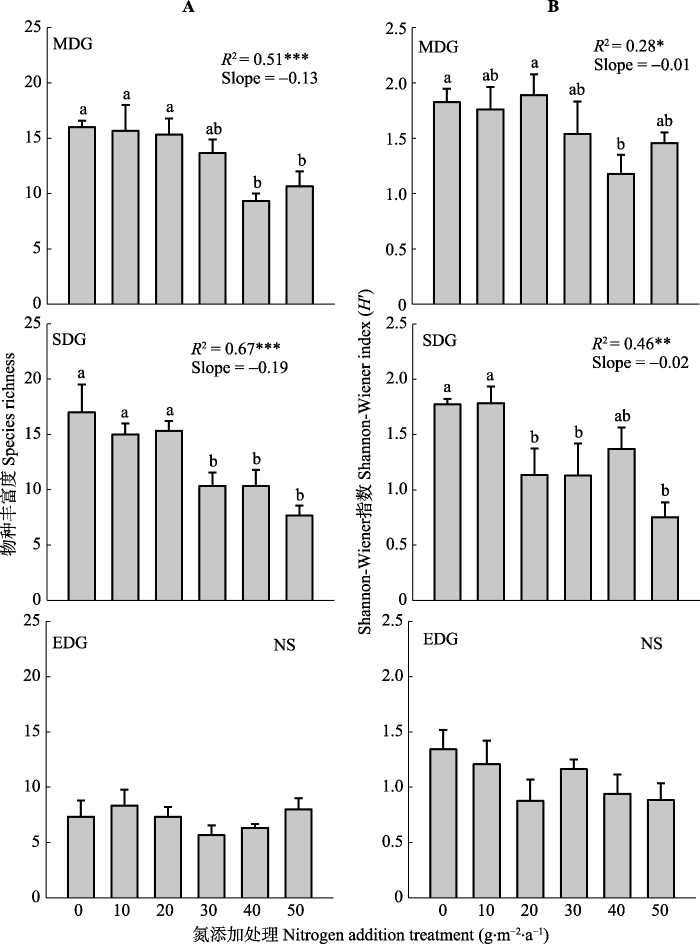 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2氮添加对不同退化草地恢复进程中物种丰富度(A)、物种多样性(B)的影响(平均值±标准误差)。单因素方差分析, Duncan’s多重比较进行处理间差异分析, 不同字母表示差异显著(p < 0.05)。每个样地, 以氮添加量为连续变量, 对物种丰富度进行回归参数估计(物种丰富度=截距+斜率×氮添加量)。NS, p > 0.05; *, p < 0.05; **, p < 0.01; ***, p < 0.001。EDG, 极度退化草地; MDG, 中度退化草地; SDG, 重度退化草地。
Fig. 2Effects of nitrogen addition on plant species richness (A) and species diversity (B) at different degraded grasslands (mean ± SE). The letters indicted significant differences in Duncan’s multiple (p < 0.05) range tests based on one-way ANOVA; NS indicates non-significant (p > 0.05). For each site, regression confidents were estimated based on linear models with nitrogen treatment as the independent variables (species richness = Intercept + slope × nitrogen addition amount). NS, p > 0.05; *, p < 0.05; **, p < 0.01; ***, p < 0.001. EDG, extremely degraded grassland; MDG, moderately degraded grassland; SDG, severely degraded grassland.
2.2 不同退化草地恢复进程中氮添加对群落地上生物量的影响
氮添加和退化梯度对群落地上生物量有显著的影响, 但它们的交互作用并不显著(表2)。在中度退化草地恢复过程中, 氮添加促进了地上生物量的增加, 增加了34.4%-114.4%, 重度退化草地地上生物量增加了46.1%-170.4%, 极度退化草地地上生物量增加了4.3%-233.7% (图3A)。3个退化草地地上生物量只有在较高的施氮量下产生明显的响应。与对照相比, 在中度退化草地, 氮添加量≥30 g·m-2·a-1, 地上生物量明显增加; 重度退化草地, 氮添加量≥20 g·m-2·a-1, 地上生物量明显增加; 而在极度退化草地, 氮添加量为40 g·m-2·a-1, 与对照相比地上生物量明显增加(图3A)。在中度和极度退化草地, 地上生物量响应比率随氮添加呈现先增加后降低的趋势, 其中两个退化样地均是在氮添加量达到40 g·m-2·a-1后, 地上生物量响应比率降低(图3B)。重度退化草地地上生物量响应比率随氮添加量增加呈现先增加后基本保持不变的趋势(图3B)。图3
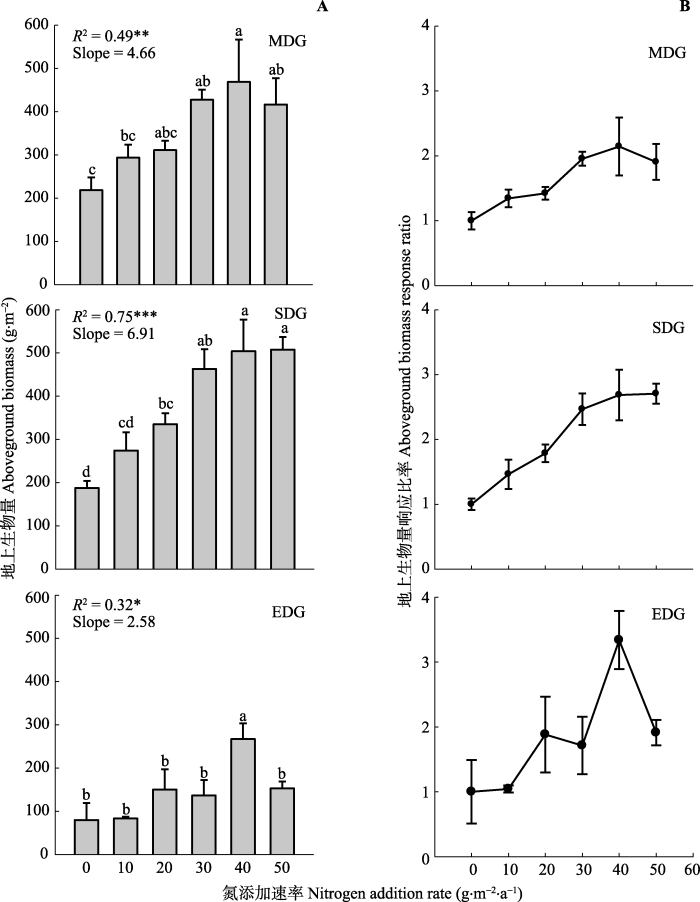 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3氮添加对不同程度退化草地恢复进程中地上生物量(A)、地上生物量响应比率(B)的影响(平均值±标准误差)。单因素方差分析、Duncan’s多重比较进行处理间差异分析, 不同字母表示差异显著(p < 0.05)。每个样地, 以氮添加量为连续变量, 对地上生物量进行回归参数估计(地上生物量=截距+斜率×氮添加量)。*, p < 0.05; **, p < 0.01; ***, p < 0.001。EDG, 极度退化草地; MDG, 中度退化草地; SDG, 重度退化草地。
Fig. 3Effects of nitrogen addition on aboveground biomass (A) and aboveground biomass response ratio (B) at different degraded communities (mean ± SE). The letters indicate significantly different in Duncan’s multiple (p < 0.05) range tests from one-way ANOVA. For each site, regression coefficients were estimated by using a linear model with N treatment as the independent variable (aboveground biomass = intercept + slope × nitrogen addition amount). *, p < 0.05; **, p < 0.01; ***, p < 0.001. EDG, extremely degraded grassland; MDG, moderately degraded grassland; SDG, severely degraded grassland.
2.3 氮添加对植物功能群地上生物量的影响
氮添加和退化梯度对禾草地上生物量有显著的影响, 但它们的交互作用并不显著; 退化梯度对杂类草地上生物量有显著的影响, 氮添加及氮添加与退化程度的交互作用对杂类草地上生物量影响不显著(表3)。其中, 在3个不同退化草地恢复过程中, 氮添加均显著增加了禾草地上生物量。与对照组相比, 中度、重度和极度退化草地禾草地上生物量分别增加了3-10倍、2-5倍和2-10倍(图4)。对于杂类草来说, 与对照组相比, 3个不同退化程度草地杂类草地上生物量对氮添加的响应不明显(图4)。Table 3
表3
表3氮添加处理、退化类型以及两者的交互作用对不同功能群地上生物量及其在群落地上生物量中所占比例(%)的双因素方差分析
Table 3
| 响应 Response | 名称 Term | df | F | p |
|---|---|---|---|---|
| 禾草生物量 Grass biomass | DT | 2 | 11.70 | <0.001 |
| N | 5 | 11.76 | <0.001 | |
| DT × N | 10 | 0.72 | 0.696 | |
| 禾草百分比 Grass percentage (%) | DT | 2 | 12.70 | <0.001 |
| N | 5 | 7.34 | <0.001 | |
| DT × N | 10 | 0.81 | 0.616 | |
| 杂类草生物量 Forb biomass | DT | 2 | 28.28 | <0.001 |
| N | 5 | 1.13 | 0.358 | |
| DT × N | 10 | 1.35 | 0.238 | |
| 杂类草百分比 Forb percentage (%) | DT | 2 | 12.35 | <0.001 |
| N | 5 | 7.01 | <0.001 | |
| DT × N | 10 | 0.80 | 0.738 |
新窗口打开|下载CSV
图4
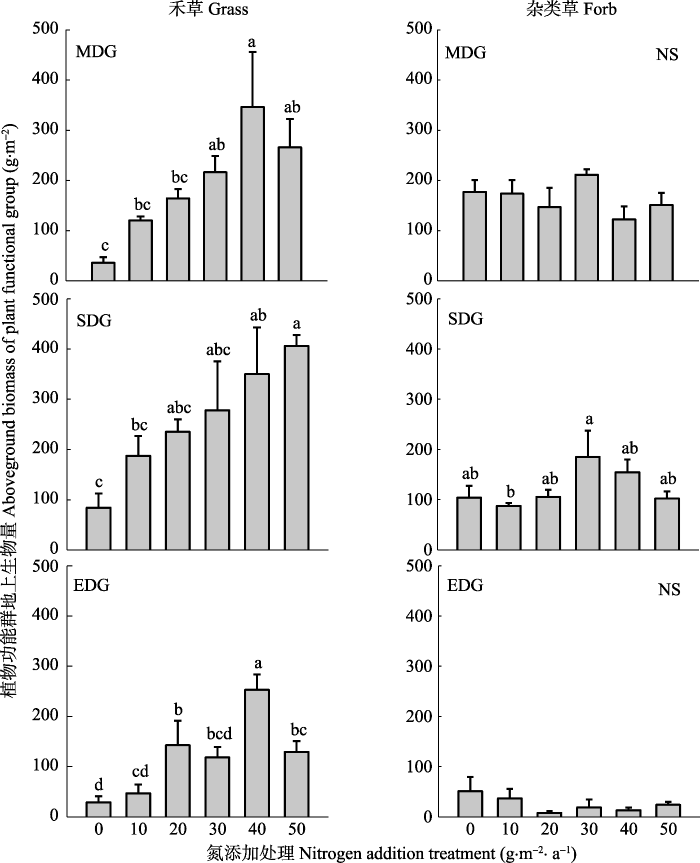 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4氮添加对不同程度退化草地恢复进程中功能群地上生物量的影响(平均值±标准误差)。单因素方差分析、Duncan’s多重比较进行处理间差异分析, 不同字母表示差异显著(p < 0.05), NS表示差异不显著。EDG, 极度退化草地; MDG, 中度退化草地; SDG, 重度退化草地。
Fig. 4Change in aboveground biomass (mean ± SE) with nitrogen addition on aboveground biomass by plant function groups under three levels of degraded grassland. The different letters indicate significant differences in Duncan’s multiple (p < 0.05) range tests from one-way ANOVA, NS indicates non-significant (p > 0.05). EDG, extremely degraded grassland; MDG, moderately degraded grassland; SDG; severely degraded grassland.
氮添加和退化梯度对禾草、杂类草地上生物量在群落地上生物量中的比例有显著影响, 但交互作用并不显著(表3)。在3个不同退化程度草地, 氮添加显著增加了禾草的比例。其中, 在中度和极度退化草地, 氮添加量达到30 g·m-2·a-1, 禾草比例明显增加; 在重度退化草地, 氮添加量为50 g·m-2·a-1, 禾草比例明显增加, 增加了35.8% (图5)。氮添加对杂类草比例的影响与禾草截然相反, 在3个不同退化程度草地, 氮添加降低了杂类草比例(图5)。
图5
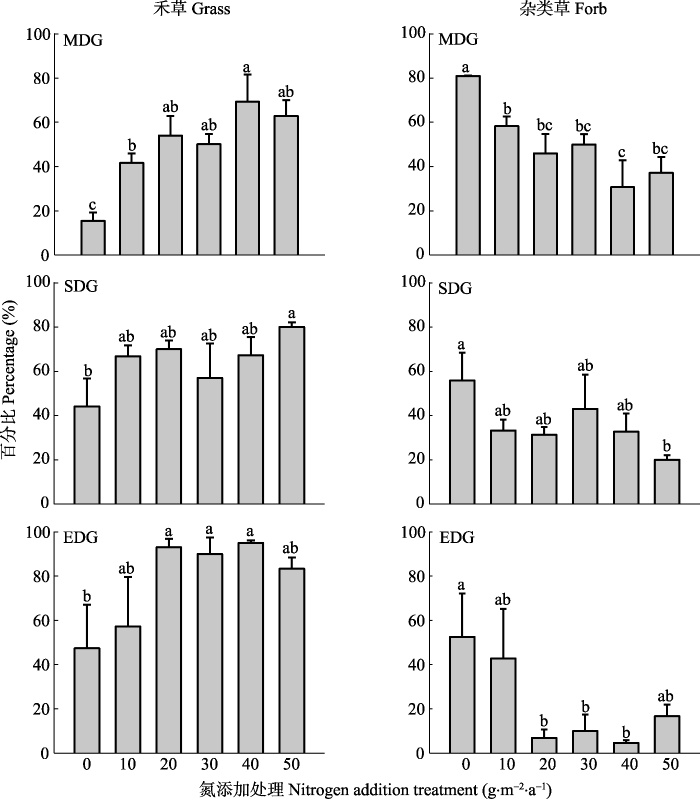 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5氮添加对不同程度退化草地恢复进程中功能群地上生物量在群落地上总生物量中所占比例的影响(平均值±标准误差)。单因素方差分析、Duncan’s多重比较进行处理间差异分析, 不同字母表示差异显著(p < 0.05)。EDG, 极度退化草地; MDG, 中度退化草地; SDG, 重度退化草地。
Fig. 5Proportion of aboveground biomass in the total biomass varied with nitrogen addition, plant functional groups (grass vs forb), and degradation level (mean ± SE). The different letters indicate the significant difference from the Duncan’s multiple (p < 0.05) range tests (one-way ANOVA). EDG, extremely degraded grassland; MDG, moderately degraded grassland; SDG, severely degraded grassland.
3 讨论
3.1 光资源竞争是氮添加对物种多样性影响的重要机制
本研究中, 氮添加降低了群落物种丰富度和多样性, 这与前人的研究结果(Fang et al., 2012; Tianet al., 2015, 2016; Niu et al., 2017)一致, 但其影响在不同程度退化草地有所差异。我们的实验结果表明: 在中度和重度退化草地, 与对照组相比, 氮添加增加了基尼系数; 而在极度退化草地, 氮添加处理下的基尼系数与对照组相比, 没有增加(图6)。即在中度和重度退化草地, 氮添加致使禾草和杂类草植株高度不对称增加, 处于群落上层的禾草(如羊草、贝加尔针茅等)对群落下层的杂类草(如星毛委陵菜等)的遮阴作用增强, 致使一些光竞争能力处于弱势的物种丧失, 从而降低了物种多样性(李禄军等, 2010)。而在极度退化草地, 功能群之间植株的高度不对称性没有增加, 植株之间对光资源的获取较为一致, 因此, 物种多样性没有发生显著变化。氮添加促进草地地上部分生长, 中度、重度退化草地植被
覆盖度增加(约60%), 群落下层物种之间光资源竞争加剧(Hautier et al., 2009; Ren et al., 2010), 致使一些物种丧失, 从而降低了物种多样性; 而在极度退化草地, 虽然氮添加会促进地上部分生长, 但由于草地退化极其严重, 群落的盖度依然很低(约40%), 植物生长受光限制的影响有限, 因而物种多样性没有明显改变。因此, 光资源竞争可能是氮添加影响物种多样性的重要机制。
图6
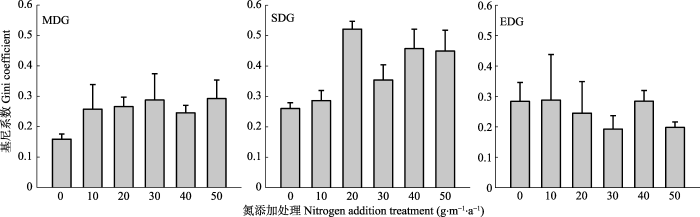 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6氮添加对不同程度退化草地基尼系数(基于功能群植物高度不对称)的影响(平均值±标准误差)。EDG, 极度退化草地; MDG, 中度退化草地; SDG, 重度退化草地。
Fig. 6Change in Gini coefficiesed on the asymmetry of plant functional groups height with nitrogen addition rate at grasslands under three different levels of degradations (mean ± SE). EDG, extremely degraded grassland; MDG, moderately degraded grassland; SDG, severely degraded grassland.
3.2 氮添加促进了不同退化草地群落地上生物量增加
氮添加促进了群落地上生物量增加, 这与他人的研究结果(Bai et al., 2008; Zhang et al., 2015)一致,但3个不同退化程度草地对氮添加的响应存在差异。对于地上生物量响应比率, 中度、极度退化草地的地上生物量响应比率呈先增加后降低的趋势, 重度退化草地呈先增加后基本保持不变的趋势。一些研究表明: 退化草地生态系统受氮限制(徐晓天, 2015), 氮添加促进了土壤可利用氮含量的增加(刘碧荣等, 2015), 解除了植物生长受氮限制的状态, 促进了植物地上部分生长, 增加了群落地上生物量(Bai et al., 2010; Lü et al., 2014; Zhang et al., 2014), 不同退化程度草地土壤性质特别是水分条件的差异可能是造成群落生物量对氮添加的响应有所差异的主要原因。氮添加促进植物群落地上生物量增加存在一个饱和临界值。当施氮速率低于饱和临界值, 地上生物量随氮添加而增加; 反之, 氮添加速率高于饱和临界值则会对植物生长产生毒害作用, 地上生物量随着氮添加会减少。Bai等(2010)在内蒙古典型草原的研究结果显示, 当施氮量为10.5 g·m-2·a-1时, 地上生物量增加趋于饱和。本研究中20 g·m-2·a-1与30 g·m-2·a-1的施氮量对草地地上生物量的促进作用基本一致, 说明本研究地的氮饱和临界值可能高于10.5 g·m-2·a-1, 这可能与草地退化有关, 也可能与当地气候条件等有关。
3.3 氮添加显著增加了禾草的地上生物量
研究结果显示, 不同退化程度的草地群落氮添加均显著增加了禾草的地上生物量及其在群落地上生物量中所占比例, 而对杂类草地上生物量没有显著影响, 但显著降低了其在地上生物量中所占的比例。杨晓霞等(2014)在青藏高原高寒草甸的研究同样发现, 氮添加促进了禾草生物量及比例的增加, 对杂类草地上生物量影响不明显但降低了其比例。Xu等(2015)的研究结果表明, 氮添加对禾草的地上生物量影响比杂类草更为明显。同时该地区之前的研究结果表明, 氮添加促使禾草地上生物量增加, 但降低了杂类草地上生物量(Song et al., 2011; He et al., 2016; Tian et al., 2016)。禾草根系比较发达(例如羊草、根茎型禾草), 具有更高的氮和水分利用效率(Song et al., 2011; He et al., 2016), 因此在施氮条件下, 禾草比杂类草更具有竞争优势, 使其地上部分快速生长, 生物量明显增加。另外, 禾草株型比较高大, 一般处于群落上层, 可以获得更丰富的光资源(Hautier et al., 2009), 合成的光合产物增加, 增加了地上生物量。总体来说氮添加促进禾草地上生物量显著增加, 且群落地上生物量增加主要是由于禾草地上生物量增加所引起的。杂类草(如星毛委陵菜等)一般处于群落下层, 在资源竞争中处于劣势。虽然氮添加可以缓解植物生长所受的养分限制, 但是由于处于群落上层的禾草具有更高的水氮利用效率(Song et al., 2011; He et al., 2016), 对氮添加响应更为明显(Hautier et al., 2009; 李春丽等, 2016), 禾草迅速生长使得其对下层杂类草的生长抑制作用增强, 杂类草在获取水分和养分资源中处于不利位置, 导致其生物量对氮添加响应不明显。另外, 禾草快速生长使其对低矮杂类草遮阴作用增强, 杂类草生长受光限制在一定程度上也抵消了氮添加的积极作用(李春丽等, 2016), 由于禾草地上生物量增加显著提高了群落地上生物量, 而杂类草地上生物量没有因施氮而发生明显改变, 因此杂类草所占比例降低。中度和重度退化草地中禾草的遮阴作用较强, 光资源可能是限制杂类草生长的重要因子; 而在极度退化草地, 禾草生长截取了多数的水分和养分, 杂类草对养分和水分资源的竞争也有可能是导致其生长受限的重要原因。
4 结论
在退化草地恢复过程中, 氮添加对群落多样性与群落地上生物量的影响与草地的退化程度有关。氮添加降低了中度、重度退化草地的物种丰富度和物种多样性, 对极度退化草地的物种丰富度和物种多样性无明显影响; 氮添加促进退化草地群落地上生物量增加, 特别是显著增加了群落中禾草地上生物量及其比例; 氮添加对杂类草地上生物量无影响, 但降低了杂类草在群落地上生物量所占比例。这一研究结果表明, 增施氮肥虽然可以促进草地生产力恢复, 但降低了群落的物种多样性。因此, 在利用施肥治理退化草地时, 不仅要充分考虑草地的退化程度, 还需要进一步权衡由氮添加引起的多样性和生产力的改变对草地生态系统功能造成的影响。
扫码加入读者圈
听语音, 看问答
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOIURL [本文引用: 1]
DOIURL [本文引用: 4]

Nitrogen (N) deposition is widely considered an environmental problem that leads to biodiversity loss and reduced ecosystem resilience; but, N fertilization has also been used as a management tool for enhancing primary production and ground cover, thereby promoting the restoration of degraded lands. However, empirical evaluation of these contrasting impacts is lacking. We tested the dual effects of N enrichment on biodiversity and ecosystem functioning at different organizational levels (i.e., plant species, functional groups, and community) by adding N at 0, 1.75, 5.25, 10.5, 17.5, and 28.0 g N m612 yr611 for four years in two contrasting field sites in Inner Mongolia: an undisturbed mature grassland and a nearby degraded grassland of the same type. N addition had both quantitatively and qualitatively different effects on the two communities. In the mature community, N addition led to a large reduction in species richness, accompanied by increased dominance of early successional annuals and loss of perennial grasses and forbs at all N input rates. In the degraded community, however, N addition increased the productivity and dominance of perennial rhizomatous grasses, with only a slight reduction in species richness and no significant change in annual abundance. The mature grassland was much more sensitive to N-induced changes in community structure, likely as a result of higher soil moisture accentuating limitation by N alone. Our findings suggest that the critical threshold for N-induced species loss to mature Eurasian grasslands is below 1.75 g N m612 yr611, and that changes in aboveground biomass, species richness, and plant functional group composition to both mature and degraded ecosystems saturate at N addition rates of approximately 10.5 g N m612 yr611. This work highlights the tradeoffs that exist in assessing the total impact of N deposition on ecosystem function.
DOIURLPMID [本文引用: 1]
Understanding how the aboveground net primary production (ANPP) of arid and semiarid ecosystems of the world responds to variations in precipitation is crucial for assessing the impacts of climate change on terrestrial ecosystems. Rain-use efficiency (RUE) is an important measure for acquiring this understanding. However, little is known about the response pattern of RUE for the largest contiguous natural grassland region of the world, the Eurasian Steppe. Here we investigated the spatial and temporal patterns of ANPP and RUE and their key driving factors based on a long-term data set from 21 natural arid and semiarid ecosystem sites across the Inner Mongolia steppe region in northern China. Our results showed that, with increasing mean annual precipitation (MAP), (1) ANPP increased while the interannual variability of ANPP declined, (2) plant species richness increased and the relative abundance of key functional groups shifted predictably, and (3) RUE increased in space across different ecosystems but decreased with increasing annual precipitation within a given ecosystem. These results clearly indicate that the patterns of both ANPP and RUE are scale dependent, and the seemingly conflicting patterns of RUE in space vs. time suggest distinctive underlying mechanisms, involving interactions among precipitation, soil N, and biotic factors. Also, while our results supported the existence of a common maximum RUE, they also indicated that its value could be substantially increased by altering resource availability, such as adding nitrogen. Our findings have important implications for understanding and predicting ecological impacts of global climate change and for management practices in arid and semiarid ecosystems in the Inner Mongolia steppe region and beyond.
DOIURL
[本文引用: 1]
DOIURLPMID [本文引用: 1]
Abstract Niche dimensionality provides a general theoretical explanation for biodiversity-more niches, defined by more limiting factors, allow for more ways that species can coexist. Because plant species compete for the same set of limiting resources, theory predicts that addition of a limiting resource eliminates potential trade-offs, reducing the number of species that can coexist. Multiple nutrient limitation of plant production is common and therefore fertilization may reduce diversity by reducing the number or dimensionality of belowground limiting factors. At the same time, nutrient addition, by increasing biomass, should ultimately shift competition from belowground nutrients towards a one-dimensional competitive trade-off for light. Here we show that plant species diversity decreased when a greater number of limiting nutrients were added across 45 grassland sites from a multi-continent experimental network. The number of added nutrients predicted diversity loss, even after controlling for effects of plant biomass, and even where biomass production was not nutrient-limited. We found that elevated resource supply reduced niche dimensionality and diversity and increased both productivity and compositional turnover. Our results point to the importance of understanding dimensionality in ecological systems that are undergoing diversity loss in response to multiple global change factors.
DOIURL [本文引用: 3]
DOIURLPMID [本文引用: 3]
Understanding the response of the plant community to increasing nitrogen (N) deposition is helpful for improving pasture management in semi-arid areas. We implemented a 5-year N addition experiment in aStipa kryloviisteppe of Inner Mongolia, northern China. The aboveground biomass (AGB) and species richness were measured annually. Along with the N addition levels, the species richness declined significantly, and the species composition changed noticeably. However, the total AGB did not exhibit a noticeable increase. We found that compensatory effects of the AGB occurred not only between the grasses and the forbs but also among Gramineae species. The plant responses to N addition, from the community to species level, lessened in dry years compared to wet or normal years. The N addition intensified the reduction of community productivity in dry years. Our study indicated that the compensatory effects of the AGB among the species sustained the stability of grassland productivity. However, biodiversity loss resulting from increasing N deposition might lead the semi-arid grassland ecosystem to be unsustainable, especially in dry years.
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURL [本文引用: 1]
[本文引用: 3]
[本文引用: 3]
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 2]
以内蒙古多伦县恢复生态学试验示范研究站弃耕10余年的草地为研究对象,于2006年起分别设置对照、氮素添加、刈割和氮素添加+刈割4种处理,每种处理6次重复,研究弃耕草地氮素添加和刈割对土壤氮矿化的影响,结合土壤理化性质和植被地上生产力的动态变化,分析弃耕草地土壤氮矿化对植被恢复的响应,为当地草地恢复与重建提供理论依据和数据支持。实验结果表明:1氮素添加显著增加了植物地上净初级生产力(ANPP)和土壤无机氮库,与对照相比分别提高115%和196%,同时显著提高了土壤总硝化速率;但是氮素添加对总氨化速率、土壤微生物生物量碳(MBC)、微生物生物量氮(MBN)、微生物生物量碳氮比(MBC/MBN)、微生物呼吸(MR)以及呼吸熵(q CO2)均无显著影响;2总氨化速率和硝化速率对刈割处理的响应均不显著,但是刈割处理显著降低了土壤MR(P0.05);3氮素添加+刈割处理5—7a后,土壤总氨化和硝化速率均无显著变化;但是氮素添加+刈割处理显著增加了ANPP、土壤无机氮库和q CO2,同时显著降低了MBC和MBC/MBN。这说明在弃耕草地适应性管理中,氮素添加可以显著提高草地生产力,但是长期的氮添加对土壤微生物氮的转化是否有利还值得我们进一步研究。
DOIURL [本文引用: 2]
以内蒙古多伦县恢复生态学试验示范研究站弃耕10余年的草地为研究对象,于2006年起分别设置对照、氮素添加、刈割和氮素添加+刈割4种处理,每种处理6次重复,研究弃耕草地氮素添加和刈割对土壤氮矿化的影响,结合土壤理化性质和植被地上生产力的动态变化,分析弃耕草地土壤氮矿化对植被恢复的响应,为当地草地恢复与重建提供理论依据和数据支持。实验结果表明:1氮素添加显著增加了植物地上净初级生产力(ANPP)和土壤无机氮库,与对照相比分别提高115%和196%,同时显著提高了土壤总硝化速率;但是氮素添加对总氨化速率、土壤微生物生物量碳(MBC)、微生物生物量氮(MBN)、微生物生物量碳氮比(MBC/MBN)、微生物呼吸(MR)以及呼吸熵(q CO2)均无显著影响;2总氨化速率和硝化速率对刈割处理的响应均不显著,但是刈割处理显著降低了土壤MR(P0.05);3氮素添加+刈割处理5—7a后,土壤总氨化和硝化速率均无显著变化;但是氮素添加+刈割处理显著增加了ANPP、土壤无机氮库和q CO2,同时显著降低了MBC和MBC/MBN。这说明在弃耕草地适应性管理中,氮素添加可以显著提高草地生产力,但是长期的氮添加对土壤微生物氮的转化是否有利还值得我们进一步研究。
DOIURL [本文引用: 3]
Topsoil characteristics and their determinants in the steppe on the central Inner Mongolian Plateau were investigated. Percentages of different grain size fractions of topsoil samples from 236 plots and corresponding standardized data of four environmental/human-activity factors, namely mean annual temperature (MAT), mean annual precipitation (MAP), human disturbance index (HDI) and land use/land cover (LULC) types were used to perform canonical correspondence analysis (CCA). The roles of both human land uses and climatic factors on topsoil grain size distribution are highlighted. Climatic factors seem to have played a significant role in determining the coarse sand (0.2-1 mm in size) percentages in the steppe and meadow by affecting the vegetation cover, while human disturbance indicated by species composition appears to have significantly influenced the percentages of clay (<0.002 mm in size) and fine-medium silt (0.002-0.016 mm in size) in the typical steppe (P < 0.01). Soil Organic Carbon (SOC) content is negatively correlated with coarse sand percentages (P < 0.01), implying that reduction of soil fertility is linked to soil coarsening. Percentages of fine-medium silts (0.002-0.016 mm in size) are still more abundant in the area where artificial vegetation are planted and remains as the major source of dust storms. This study implies that afforestation in the steppe region has not improved soil conditions and thus reduced dust storms as earlier expected.
DOIURLPMID [本文引用: 1]
Abstract Increased atmospheric nitrogen (N) deposition and altered precipitation regimes have profound impacts on ecosystem functioning in semiarid grasslands. The interactions between those two factors remain largely unknown. A field experiment with N and water additions was conducted in a semiarid grassland in northern China. We examined the responses of aboveground net primary production (ANPP) and plant N use during two contrasting hydrological growing seasons. Nitrogen addition had no impact on ANPP, which may be accounted for by the offset between enhanced plant N uptake and decreased plant nitrogen use efficiency (NUE). Water addition significantly enhanced ANPP, which was largely due to enhanced plant aboveground N uptake. Nitrogen and water additions significantly interacted to affect ANPP, plant N uptake and N concentrations at the community level. Our observations highlight the important role of plant N uptake and use in mediating the effects of N and water addition on ANPP.
DOIURL [本文引用: 1]
1. Grassland on lowland peat has considerable potential for agriculture and conservation. 2. The limited use of fertilizer on the Somerset Moors, designated as an Environmentally Sensitive Area has enabled the impact on vegetation of applying a range of nitrogen levels to be investigated using a randomized block experiment with large plots at Tadham Moor. 3. From 1986 to 1990, the plot measurements each year included the individual percentage cover of some 100 species; these enabled the derivation of a number of measures of plant community structure. 4. The applications of fertilizer encouraged grasses (notably Lolium perenne and Holcus lanatus) which came to dominate the sward at the expense of most other species. All measures of species diversity declined significantly and the sward became taller in plots receiving high levels of nitrogen. Only a very few forbs (e.g. Rumex acetosa) maintained or increased their cover with increased nitrogen. 5. All Carex species, Juncus species and mosses became significantly less common in plots treated with nitrogen, producing an apparently more mesotrophic sward. Short-lived species and low-growing wetland forbs also declined with nitrogen, perhaps due to shading by tall grasses. Legumes were suppressed by high levels of nitrogen, but were common in plots receiving 25 kg ha-1year-1N, where replacement levels of phosphorus and potassium were applied after cutting. 6. Except for grasses, many species common in the experimental site in 1986 declined in cover, particularly in plots receiving high levels of nitrogen. However, the data showed no instances of extinction by 1990. The application of 25 kg ha-1N encouraged the spread of agriculturally productive grasses within 2 years and 50 kg ha-1year-1N significantly reduced species richness in 3 years. 7. Certain detailed findings from this large-scale study of nitrogen application in meadows enable contrasts with previous results to be identified.
[本文引用: 2]
[本文引用: 2]
DOIURLPMID [本文引用: 1]
Abstract Increasing atmospheric nitrogen (N) deposition may affect plant biodiversity, subsequently altering ecosystem stability. While a few studies have explored how simulated N deposition affects community stability and its underlying mechanisms, the experimental levels of N addition used are usually higher than current and future N deposition rates. Thus, their results could produce highly uncertain predictions of ecosystem function, especially if the responses to N deposition are nonlinear. We conducted a manipulative experiment that simulated elevated atmospheric N deposition with several N addition levels to evaluate the effect of N deposition on ecosystem stability and its underlying mechanisms in a semiarid grassland in northern China. Here we show that N addition altered community diversity, reducing species richness, evenness, diversity and dominance. In addition, we found that N addition at current N deposition levels had no significant impact on community stability. In contrast, N addition at levels from 4.6 to 13.8gNm -2 yr -1 significantly decreased community stability, although community stability for the 13.8gNm -2 yr -1 treatment was higher than that for the 4.6gNm -2 yr -1 treatment. These results indicate that the response of community stability to N enrichment is nonlinear. This nonlinear change in community stability was positively correlated with species asynchrony, species richness, and species diversity as well as the stability of dominant species and the stability of the grass functional group. Our data suggest a need to re-evaluate the mechanisms responsible for the effects of N deposition on natural ecosystem stability across multiple levels of N enrichment and that additional experimentation with gradients of N loads more similar to future atmospheric N deposition rates is needed. Copyright 2017 Elsevier B.V. All rights reserved.
DOIURL [本文引用: 1]
A rapid increase in grazing intensity since the 1980s has caused large areas of the Inner Mongolian grasslands to become degraded. Increasing atmospheric nitrogen (N) deposition might exert an important influence on vegetation restoration in these degraded grasslands by increasing available N and relieving N limitations on productivity. However, no previous studies have tested the assumption that increasing N deposition promotes vegetation restoration in degraded grasslands. By conducting a 4-year field restoration experiment with four N addition treatments (0, 5, 10, and 20gNm/2 year/1) and two grazing treatments (grazed and ungrazed), we investigated the effects of N enrichment and grazing on the restoration of patches in which vegetation had been degraded. N addition significantly accelerated the restoration of vegetation-degraded grassland patches regarding both plant cover and diversity. Moderate grazing also promoted the restoration of degraded-vegetation patches in term of both plant diversity and species similarity. Importantly, the positive effects of N addition on the restoration of degraded patches may be augmented by grazing. This study demonstrate that low levels of N enrichment (or increasing atmospheric N deposition) positively impact vegetation restoration in degraded grassland patches, particularly under moderate grazing practices. Our findings provide new insights into the management of severely degraded grasslands through the regulation of N inputs and grazing practices.
DOIURL
1 Plant community theory often invokes competition to explain why species diversity declines as productivity increases. Competition for all resources might become more intense and lead to greater competitive exclusion or, alternatively, competition for light only could become more intense and exclude poor light competitors. 2 To test these hypotheses, we constructed communities of seven old-field species using combined monocultures. Constructs experienced no interspecific competition, only shoot competition or only root competition, with and without fertilizer. Diversity in these limited interaction communities was compared to diversity in unfertilized and fertilized mixtures of the seven species. 3 Diversity decreased with fertilization in mixtures and in communities with only root competition. Shoot competition had small effects on the community and did not contribute to changes in diversity with fertilization. 4 Root competition may strongly impact plant community structure in unproductive communities where light never becomes limiting, or under non-equilibrium conditions following human disturbances.
DOIURL [本文引用: 2]
[Aims] Theories based on resource additions indicate that plant species richness is mainly determined by the number of limiting resources. However, the individual effects of various limiting resources on species richness and aboveground net primary productivity (ANPP) are less well understood. Here, we analyzed potential linkages between additions of limiting resources, species loss and ANPP increase and further explored the underlying mechanisms. [Methods] Resources (N, P, K and water) were added in a completely randomized block design to alpine meadow plots in the Qinghai-Tibetan Plateau....
DOIURL [本文引用: 1]
The challenge to identify the biospheric sinks for about half the total carbon emissions from fossil fuels must include a consideration of below-ground ecosystem processes as well as those more easily measured above-ground. Recent studies suggest that tropical grasslands and savannas may contribute more to the 'missing sink' than was previously appreciated, perhaps as much as 0.5 Pg (= 0.5 Gt) carbon per annum. The rapid increase in availability of productivity data facilitated by the Internet will be important for future scaling-up of global change responses, to establish independent lines of evidence about the location and size of carbon sinks.
URL [本文引用: 1]
中国草地面积广阔,自然资源丰富,准确评估草地资源既是合理开发和利用草地的基础,也对生态环境的保护具有重要意义.但是,关于中国草地面积、生产力和承载力等本底数据至今为止尚没有系统的梳理.本文收集、整理了过去几十年来我国草地资源研究的各类数据,并利用1982~2011年的遥感影像(NOAA/AVHRR-NDVI)和气候数据重新估算了我国天然草地生物量和生产力及其近30年的变化.由于草地的定义、数据来源和分析方法不同,现存资料对中国天然草地面积的估算差异很大,变动范围达2倍以上(1.67×10~6~4.31×10~6 km~2),这些资料也表明,我国目前的天然草地面积在2.80×10~6~3.93×10~6 km~2之间比较合适.草地生物量的估算值也存在显著差异,平均地上生物量在79~123 g m~(-2)之间,但本文对最近30年(1982~2011年)天然草地地上生物量的重新估算结果为178 g m~(-2),在此期间平均每年增加0.4 gm~(-2).我国天然草地平均净初级生产力的估算差异更大,为89~320 g Cm~(-2)a~(-1)(平均176 g Cm~(-2) a~(-1)),但许多研究都发现近年来有增加趋势.而基于过去30年平均气候估算,我国天然草地的潜在生产力可高达348 gCm~(-2) a~(-1).另一方面,我国人工草地面积比较小,约为2.09×10~7 hm~2,但生产力高,可达天然草地的2.7~12.1倍.由于我国对天然草地缺乏有效管理,加上人工草地的比例低,目前我国草地对放牧家畜的承载力比较低,很多地方的超载现象较为严重,平均超载率估计为20%.此外,降水的不足始终是影响我国草地生物多样性、生物量和生产力的重要因素,进一步探讨气候变化和过度放牧等人为活动对我国草地资源的数量和质量的影响是十分必要的.
URL [本文引用: 1]
中国草地面积广阔,自然资源丰富,准确评估草地资源既是合理开发和利用草地的基础,也对生态环境的保护具有重要意义.但是,关于中国草地面积、生产力和承载力等本底数据至今为止尚没有系统的梳理.本文收集、整理了过去几十年来我国草地资源研究的各类数据,并利用1982~2011年的遥感影像(NOAA/AVHRR-NDVI)和气候数据重新估算了我国天然草地生物量和生产力及其近30年的变化.由于草地的定义、数据来源和分析方法不同,现存资料对中国天然草地面积的估算差异很大,变动范围达2倍以上(1.67×10~6~4.31×10~6 km~2),这些资料也表明,我国目前的天然草地面积在2.80×10~6~3.93×10~6 km~2之间比较合适.草地生物量的估算值也存在显著差异,平均地上生物量在79~123 g m~(-2)之间,但本文对最近30年(1982~2011年)天然草地地上生物量的重新估算结果为178 g m~(-2),在此期间平均每年增加0.4 gm~(-2).我国天然草地平均净初级生产力的估算差异更大,为89~320 g Cm~(-2)a~(-1)(平均176 g Cm~(-2) a~(-1)),但许多研究都发现近年来有增加趋势.而基于过去30年平均气候估算,我国天然草地的潜在生产力可高达348 gCm~(-2) a~(-1).另一方面,我国人工草地面积比较小,约为2.09×10~7 hm~2,但生产力高,可达天然草地的2.7~12.1倍.由于我国对天然草地缺乏有效管理,加上人工草地的比例低,目前我国草地对放牧家畜的承载力比较低,很多地方的超载现象较为严重,平均超载率估计为20%.此外,降水的不足始终是影响我国草地生物多样性、生物量和生产力的重要因素,进一步探讨气候变化和过度放牧等人为活动对我国草地资源的数量和质量的影响是十分必要的.
DOIURL [本文引用: 3]
Chinese grasslands are extensive natural ecosystems that comprise 40 % of the total land area of the country and are sensitive to N deposition. A field experiment with six N rates (0, 30, 60, 120, 240, and 480 kg N halt;supgt;amp;minus;1lt;/supgt; yrlt;supgt;amp;minus;1lt;/supgt;) was conducted at Duolun, Inner Mongolia, during 2005 and 2010 to identify some effects of N addition on a temperate steppe ecosystem. The dominant plant species in the plots were divided into two categories, grasses and forbs, on the basis of species life forms. Enhanced N deposition, even as little as 30 kg N halt;supgt;amp;minus;1lt;/supgt; yrlt;supgt;amp;minus;1lt;/supgt; above ambient N deposition (16 kg N halt;supgt;amp;minus;1lt;/supgt; yrlt;supgt;amp;minus;1lt;/supgt;), led to a decline in species richness. The cover of grasses increased with N addition rate but their species richness showed a weak change across N treatments. Both species richness and cover of forbs declined strongly with increasing N deposition as shown by linear regression analysis (lt;igt;plt;/igt; lt; 0.05). Increasing N deposition elevated aboveground production of grasses but lowered aboveground biomass of forbs. Plant N concentration, plant amp;delta;lt;supgt;15lt;/supgt;N and soil mineral N increased with N addition, showing positive relationships between plant amp;delta;lt;supgt;15lt;/supgt;N and N concentration, soil mineral N and/or applied N rate. The cessation of N application in the 480 kg N halt;supgt;amp;minus;1lt;/supgt; yrlt;supgt;amp;minus;1lt;/supgt; treatment in 2009 and 2010 led to a slight recovery of the forb species richness relative to total cover and aboveground biomass, coinciding with reduced plant N concentration and soil mineral N. The results show N deposition-induced changes in soil N transformations and plant N assimilation that are closely related to changes in species composition and biomass accumulation in this temperate steppe ecosystem.
DOIURLPMID [本文引用: 1]
Summary Many ecosystems are facing strong perturbations such as nitrogen (N) fertilization, which can greatly alter ecosystem stability via different mechanisms. Understanding such mechanisms is critical for predicting how ecosystems will function in the face of global changes. We examined how 8/yr of N fertilization with different N rates (no N addition or N addition at a low, medium or high rate) and different forms of N (ammonium, nitrate or ammonium nitrate) affected the temporal stability of the aboveground biomass of an alpine meadow on the Tibetan Plateau, and tested four mechanisms (diversity effect, mean-variance scaling, compensatory dynamics and dominance effect) that may alter stability. Compared with the control (no N addition), a high N rate did not affect the diversity effect, the mean-variance scaling or the dominance effect, but significantly decreased compensatory dynamics among species and functional groups, which contributed to the reduction in community stability of the alpine meadow. The form of N did not affect any of the four mechanisms and thus did not affect community stability. A high N rate can change community stability by altering compensatory dynamics, whereas the form of N may not have an effect.
DOIURL [本文引用: 1]
61Soil ammonium and nitrate concentrations frequently respond relatively quickly to reduced N.61The response of plant tissue N concentrations varied between habitats.61Soil processes are often slow to recover from reduced N inputs.61Vegetation species composition is also often slow to recover from reduced N inputs.
DOIURLPMID [本文引用: 1]
A transect of 68 acid grasslands across Great Britain, covering the lower range of ambient annual nitrogen deposition in the industrialized world (5 to$35 kg N ha^{-1} year^{-1}$), indicates that long-term, chronic nitrogen deposition has significantly reduced plant species richness. Species richness declines as a linear function of the rate of inorganic nitrogen deposition, with a reduction of one species per 4-m2quadrat for every$2.5 kg N ha^{-1} year^{-1}$of chronic nitrogen deposition. Species adapted to infertile conditions are systematically reduced at high nitrogen deposition. At the mean chronic nitrogen deposition rate of central Europe ($17 kg N ha^{-1} year^{-1}$), there is a 23% species reduction compared with grasslands receiving the lowest levels of nitrogen deposition.
DOIURL [本文引用: 1]
Abstract Humans dominate many important Earth system processes including the nitrogen (N) cycle. Atmospheric N deposition affects fundamental processes such as carbon cycling, climate regulation, and biodiversity, and could result in changes to fundamental Earth system processes such as primary production. Both modelling and experimentation have suggested a role for anthropogenically altered N deposition in increasing productivity, nevertheless, current understanding of the relative strength of N deposition with respect to other controls on production such as edaphic conditions and climate is limited. Here we use an international multiscale data set to show that atmospheric N deposition is positively correlated to aboveground net primary production (ANPP) observed at the 1-m2 level across a wide range of herbaceous ecosystems. N deposition was a better predictor than climatic drivers and local soil conditions, explaining 16% of observed variation in ANPP globally with an increase of 1 kg N·ha611·yr611 increasing ANPP by 3%. Soil pH explained 8% of observed variation in ANPP while climatic drivers showed no significant relationship. Our results illustrate that the incorporation of global N deposition patterns in Earth system models are likely to substantially improve estimates of primary production in herbaceous systems. In herbaceous systems across the world, humans appear to be partially driving local ANPP through impacts on the N cycle.
DOIURL [本文引用: 2]
Overgrazing is one of the main causes of desertification in the semiarid Horqin sandy grassland of northern China. Excluding grazing livestock is considered as an alternative to restore vegetation in degraded sandy grassland in this region. However, few data are available concerning the impacts of continuous grazing and livestock exclusion on soil properties. In this paper, characteristics of vegetation and soil properties under continuous grazing and exclusion of livestock for 5 and 10 years were examined in representative degraded sandy grassland. Continuous grazing resulted in a considerable decrease in ground cover, which accelerates soil erosion by wind, leading to a further coarseness in surface soil, loss of soil organic C and N, and a decrease in soil biological properties. The grassland under continuous grazing is in the stage of very strong degradation. Excluding livestock grazing enhances vegetation recovery, litter accumulation, and development of annual and perennial grasses. Soil organic C and total N concentrations, soil biological properties including some enzyme activities and basal soil respiration improved following 10-year exclusion of livestock, suggesting that degradation of the grassland is being reversed. The results suggest that excluding grazing livestock on the desertified sandy grassland in the erosion-prone Horqin region has a great potential to restore soil fertility, sequester soil organic carbon and improve biological activity. Soil restoration is a slow process although the vegetation can recover rapidly after removal of livestock. A viable option for sandy grassland management should be to adopt proper exclosure in a rotation grazing system in the initial stage of grassland degradation.
DOIURLPMID [本文引用: 1]
Abstract Loss of plant diversity with increased anthropogenic nitrogen (N) deposition in grasslands has occurred globally. In most cases, competitive exclusion driven by preemption of light or space is invoked as a key mechanism. Here, we provide evidence from a 9-yr N-addition experiment for an alternative mechanism: differential sensitivity of forbs and grasses to increased soil manganese (Mn) levels. In Inner Mongolia steppes, increasing the N supply shifted plant community composition from grassorb codominance (primarily Stipa krylovii and Artemisia frigida , respectively) to exclusive dominance by grass, with associated declines in overall species richness. Reduced abundance of forbs was linked to soil acidification that increased mobilization of soil Mn, with a 10-fold greater accumulation of Mn in forbs than in grasses. The enhanced accumulation of Mn in forbs was correlated with reduced photosynthetic rates and growth, and is consistent with the loss of forb species. Differential accumulation of Mn between forbs and grasses can be linked to fundamental differences between dicots and monocots in the biochemical pathways regulating metal transport. These findings provide a mechanistic explanation for N-induced species loss in temperate grasslands by linking metal mobilization in soil to differential metal acquisition and impacts on key functional groups in these ecosystems.
DOIURL
Enhanced deposition of atmospheric nitrogen (N) resulting from anthropogenic activities has negative impacts on plant diversity in ecosystems. Several mechanisms have been proposed to explain the species loss. Ion toxicity due to N deposition-induced soil acidification has been suggested to be responsible for species loss in acidic grasslands, while few studies have evaluated the role of soil-mediated homeostasis of ions in species loss under elevated N deposition in grasslands with neutral or alkaline soils. To determine whether soil-mediated processes are involved in changes in biodiversity induced by N deposition, the effects of 9-year N addition on soil properties, aboveground biomass (AGB) and species richness were investigated in an Inner Mongolia steppe. Low to moderate N addition rate (2, 4, 8 g N myr) significantly enhanced AGB of graminoids, while high N addition rate ( 16 g N myr) reduced AGB of forbs, leading to an overall increase in AGB of the community under low to moderate N addition rates. Forb richness was significantly reduced by N addition at rates greater than 8 g N myr, while no effect of N addition on graminoid richness was observed, resulting in decline in total species richness. N addition reduced soil pH, depleted base cations (Ca, Mgand K) and mobilized Mn, Fe, Cuand Alions in soils. Soil inorganic-N concentration was negatively correlated with forb richness and biomass, explaining 23.59% variation of forb biomass. The concentrations of base cations (Caand Mg) and metal ions (Mn, Cuand, Fe) showed positively and negatively linear correlation with forb richness, respectively. Changes in the metal ion concentrations accounted for 42.77% variation of forb richness, while reduction of base cations was not associated with the reduction in forb richness. These results reveal that patterns of plant biodiversity in the temperate steppe of Inner Mongolia are primarily driven by increases in metal ion availability, particularly enhanced release of soil Mn.
DOIURL [本文引用: 1]
DOIURLPMID [本文引用: 1]
Abstract It has long been recognized that alternative vegetation states may occur in terrestrial grazing systems. This phenomenon may be of great importance as small environmental fluctuations may lead to relatively sudden and irreversible jumps between vegetation states. Early theoretical studies emphasized saturation of herbivore feeding to explain multiple stable states and catastrophic behaviour. Recent studies on semi-arid grasslands and arctic salt marshes, however, relate catastrophic events in these systems to plant-soil interactions.
[本文引用: 2]
[本文引用: 2]
DOIURL [本文引用: 1]
The dynamics of belowground net primary productivity (BNPP) is of fundamental importance in understanding carbon (C) allocation and storage in grasslands. However, our knowledge of the interannual variability in response of BNPP to ongoing global warming is limited. In this study, we explored temporal responses of BNPP and net primary productivity (NPP) partitioning to warming and clipping in a tallgrass prairie in Oklahoma, USA. Infrared heaters were used to elevate soil temperature by approximately 2 C since November 1999. Annual clipping was to mimic hay harvest. On average from 2005 to 2009, warming increased BNPP by 41.89% in the unclipped subplots and 66.93% in the clipped subplots, with significant increase observed in wet years. Clipping also had significant positive impact on BNPP, which was mostly found under warming. Overall, fBNPP, the fraction of BNPP to NPP, increased under both warming and clipping treatments, more in dry years. Water availability (either precipitation or soil moisture) was the most limiting factor for both BNPP and fBNPP. It strongly dominated the interannual variability in NPP, fBNPP, and their responses to warming and clipping. Our results suggest that water availability regulates tallgrass prairie's responses to warming and land use change, which may eventually influence the global C cycle. With increasing variability in future precipitation patterns, warming effects on the vegetation in this region may become less predictable.
[本文引用: 2]
[本文引用: 2]
DOIURLPMID
Although nitrogen addition and recovery from degradation can both promote production of grassland biomass, these two factors have rarely been investigated in combination. In this study, we established a field experiment with six N-treatment (CK, 10, 20, 30, 40, 5065g65N65m61265yr611) on five fields with different degradation levels in the Inner Mongolian steppe of China from 2011–2013. Our observations showed that while the external nitrogen increased the aboveground biomass in all five grasslands, the magnitude of the effects differed with the severity of degradation. Fields with a higher level of degradation tended to have a higher saturation value (2065g65N65m61265yr611) than those with a lower degradation level (65<651065g65N m61265yr611). After three years of experimentation, species richness showed little change across degradation levels. Among the four functional groups of grasses, sedges, forbs and legumes, grasses shared the most similar response patterns with those of the whole community, demonstrating the predominant role that they play in the restoration of grassland under a stimulus of nitrogen addition.
DOIURLPMID [本文引用: 2]
Anthropogenic perturbations may affect biodiversity and ecological stability as well as their relationships. However, diversity–stability patterns and associated mechanisms under human disturbances have rarely been explored. We conducted a 7-year field experiment examining the effects of mowing and nutrient addition on the diversity and temporal stability of herbaceous plant communities in a temperate steppe in northern China. Mowing increased population and community stability, whereas nutrient addition had the opposite effects. Stability exhibited positive relationships with species richness at population, functional group and community levels. Treatments did not alter these positive diversity–stability relationships, which were associated with the stabilising effect of species richness on component populations, species asynchrony and portfolio effects. Despite the difficulty of pinpointing causal mechanisms of diversity–stability patterns observed in nature, our results suggest that diversity may still be a useful predictor of the stability of ecosystems confronted with anthropogenic disturbances.
DOIURL
青藏高原正经历着明显的温暖化过程,由此引起的土壤温度的升高促进了土壤中微生物的活性,同时青藏高原东缘地区大气氮沉降十分明显,并呈逐年增加的趋势,这些环境变化均促使土壤中可利用营养元素增加,因此深入了解青藏高原高寒草甸植物生物量对可利用营养元素增加的响应,是准确预测未来全球变化背景下青藏高原高寒草甸碳循环过程的重要基础。该研究基于在青藏高原高寒草甸连续4年(2009-2012年)氮、磷添加后对不同功能群植物地上生物量、群落地上和地下生物量的测定,探讨高寒草甸生态系统碳输入对氮、磷添加的响应。结果表明:(1)氮、磷添加均极显著增加了禾草的地上绝对生物量及其在群落总生物量中所占的比例,同时均显著降低了杂类草在群落总生物量中的比例,此外磷添加极显著降低了莎草地上绝对生物量及其在群落总生物量中所占的比例。(2)氮、磷添加均显著促进了青藏高原高寒草甸的地上生物量增加,分别增加了24%和52%。(3)氮添加对高寒草甸地下生物量无显著影响,而磷添加后地下生物量有增加的趋势。(4)氮添加对高寒草甸植物总生物量无显著影响,而磷添加后植物总生物量显著增加。研究表明,氮、磷添加可缓解青藏高原高寒草甸植物生长的营养限制,促进植物地上部分的生长,然而高寒草甸植物的生长极有可能更受土壤中可利用磷含量的限制。
DOIURL
青藏高原正经历着明显的温暖化过程,由此引起的土壤温度的升高促进了土壤中微生物的活性,同时青藏高原东缘地区大气氮沉降十分明显,并呈逐年增加的趋势,这些环境变化均促使土壤中可利用营养元素增加,因此深入了解青藏高原高寒草甸植物生物量对可利用营养元素增加的响应,是准确预测未来全球变化背景下青藏高原高寒草甸碳循环过程的重要基础。该研究基于在青藏高原高寒草甸连续4年(2009-2012年)氮、磷添加后对不同功能群植物地上生物量、群落地上和地下生物量的测定,探讨高寒草甸生态系统碳输入对氮、磷添加的响应。结果表明:(1)氮、磷添加均极显著增加了禾草的地上绝对生物量及其在群落总生物量中所占的比例,同时均显著降低了杂类草在群落总生物量中的比例,此外磷添加极显著降低了莎草地上绝对生物量及其在群落总生物量中所占的比例。(2)氮、磷添加均显著促进了青藏高原高寒草甸的地上生物量增加,分别增加了24%和52%。(3)氮添加对高寒草甸地下生物量无显著影响,而磷添加后地下生物量有增加的趋势。(4)氮添加对高寒草甸植物总生物量无显著影响,而磷添加后植物总生物量显著增加。研究表明,氮、磷添加可缓解青藏高原高寒草甸植物生长的营养限制,促进植物地上部分的生长,然而高寒草甸植物的生长极有可能更受土壤中可利用磷含量的限制。
[本文引用: 1]
[本文引用: 1]
DOIURL [本文引用: 1]
DOIURLPMID [本文引用: 2]
Abstract Humans are both intentionally (fertilization) and unintentionally (atmospheric nutrient deposition) adding nutrients worldwide. Increasing availability of biologically reactive nitrogen (N) is one of the major drivers of plant species loss. It remains unclear, however, whether plant diversity will be equally reduced by inputs of reactive N coming from either small and frequent N deposition events or large and infrequent N fertilization events. By independently manipulating the rate and frequency of reactive N inputs, our study teases apart these potentially contrasting effects. Plant species richness decreased more quickly at high rates and at low frequency of N addition, which suggests that previous fertilization studies have likely over-estimated the effects of N deposition on plant species loss. N-induced species loss resulted from both acidification and ammonium toxicity. Further study of small and frequent N additions will be necessary to project future rates of plant species loss under increasing aerial N deposition.
Grazing systems, ecosystem response, and global change.
1
2004
... 草地是全球分布最广的一个植被类型(
Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: Evidence from Inner Mongolia grasslands.
4
2010
... 草地是全球分布最广的一个植被类型(
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
... )和生态系统功能(
... 地上生物量响应比率呈先增加后降低的趋势, 重度退化草地呈先增加后基本保持不变的趋势.一些研究表明: 退化草地生态系统受氮限制(
Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau.
1
2008
... 氮添加促进了群落地上生物量增加, 这与他人的研究结果(
Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands.
2008
Long-term nitrogen addition leads to loss of species richness due to litter accumulation and soil acidification in a temperate steppe.
1
2012
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
Addition of multiple limiting resources reduces grassland diversity.
1
2016
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
Competition for light causes plant biodiversity loss after eutrophication.
3
2009
... 覆盖度增加(约60%), 群落下层物种之间光资源竞争加剧(
... 研究结果显示, 不同退化程度的草地群落氮添加均显著增加了禾草的地上生物量及其在群落地上生物量中所占比例, 而对杂类草地上生物量没有显著影响, 但显著降低了其在地上生物量中所占的比例.杨晓霞等(2014)在青藏高原高寒草甸的研究同样发现, 氮添加促进了禾草生物量及比例的增加, 对杂类草地上生物量影响不明显但降低了其比例.Xu等(2015)的研究结果表明, 氮添加对禾草的地上生物量影响比杂类草更为明显.同时该地区之前的研究结果表明, 氮添加促使禾草地上生物量增加, 但降低了杂类草地上生物量(
... 杂类草(如星毛委陵菜等)一般处于群落下层, 在资源竞争中处于劣势.虽然氮添加可以缓解植物生长所受的养分限制, 但是由于处于群落上层的禾草具有更高的水氮利用效率(
Response of aboveground biomass and diversity to nitrogen addition a five-year experiment in semi-arid grassland of Inner Mongolia, China.
3
2016
... 研究结果显示, 不同退化程度的草地群落氮添加均显著增加了禾草的地上生物量及其在群落地上生物量中所占比例, 而对杂类草地上生物量没有显著影响, 但显著降低了其在地上生物量中所占的比例.杨晓霞等(2014)在青藏高原高寒草甸的研究同样发现, 氮添加促进了禾草生物量及比例的增加, 对杂类草地上生物量影响不明显但降低了其比例.Xu等(2015)的研究结果表明, 氮添加对禾草的地上生物量影响比杂类草更为明显.同时该地区之前的研究结果表明, 氮添加促使禾草地上生物量增加, 但降低了杂类草地上生物量(
... ;
... 杂类草(如星毛委陵菜等)一般处于群落下层, 在资源竞争中处于劣势.虽然氮添加可以缓解植物生长所受的养分限制, 但是由于处于群落上层的禾草具有更高的水氮利用效率(
Nitrogen limitation in dry land ecosystems: Responses to geographical and temporal variation in precipitation.
1
1999
... 全球范围内, 有关氮添加对草地生态系统影响以及氮添加降低物种多样性机制的报道中, 只有少量实验是从退化草地恢复的角度出发(
Nutrient enrichment, biodiversity loss, and consequent declines in ecosystem productivity.
1
2013
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
Grassland ecosystems in China: Review of current knowledge and research advancement.
1
2007
... 草地是全球分布最广的一个植被类型(
环青海湖地区天然草地和退耕恢复草地植物群落生物量对氮、磷添加的响应
3
2016
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
... 杂类草(如星毛委陵菜等)一般处于群落下层, 在资源竞争中处于劣势.虽然氮添加可以缓解植物生长所受的养分限制, 但是由于处于群落上层的禾草具有更高的水氮利用效率(
... ), 禾草迅速生长使得其对下层杂类草的生长抑制作用增强, 杂类草在获取水分和养分资源中处于不利位置, 导致其生物量对氮添加响应不明显.另外, 禾草快速生长使其对低矮杂类草遮阴作用增强, 杂类草生长受光限制在一定程度上也抵消了氮添加的积极作用(
环青海湖地区天然草地和退耕恢复草地植物群落生物量对氮、磷添加的响应
3
2016
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
... 杂类草(如星毛委陵菜等)一般处于群落下层, 在资源竞争中处于劣势.虽然氮添加可以缓解植物生长所受的养分限制, 但是由于处于群落上层的禾草具有更高的水氮利用效率(
... ), 禾草迅速生长使得其对下层杂类草的生长抑制作用增强, 杂类草在获取水分和养分资源中处于不利位置, 导致其生物量对氮添加响应不明显.另外, 禾草快速生长使其对低矮杂类草遮阴作用增强, 杂类草生长受光限制在一定程度上也抵消了氮添加的积极作用(
施肥对科尔沁沙质草地群落物种组成和多样性的影响
2
2010
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
... et al., 2015, 2016; Niu et al., 2017)一致, 但其影响在不同程度退化草地有所差异.我们的实验结果表明: 在中度和重度退化草地, 与对照组相比, 氮添加增加了基尼系数; 而在极度退化草地, 氮添加处理下的基尼系数与对照组相比, 没有增加(
施肥对科尔沁沙质草地群落物种组成和多样性的影响
2
2010
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
... et al., 2015, 2016; Niu et al., 2017)一致, 但其影响在不同程度退化草地有所差异.我们的实验结果表明: 在中度和重度退化草地, 与对照组相比, 氮添加增加了基尼系数; 而在极度退化草地, 氮添加处理下的基尼系数与对照组相比, 没有增加(
氮添加和刈割对内蒙古弃耕草地土壤氮矿化的影响
2
2015
... 草地是全球分布最广的一个植被类型(
... 地上生物量响应比率呈先增加后降低的趋势, 重度退化草地呈先增加后基本保持不变的趋势.一些研究表明: 退化草地生态系统受氮限制(
氮添加和刈割对内蒙古弃耕草地土壤氮矿化的影响
2
2015
... 草地是全球分布最广的一个植被类型(
... 地上生物量响应比率呈先增加后降低的趋势, 重度退化草地呈先增加后基本保持不变的趋势.一些研究表明: 退化草地生态系统受氮限制(
Climatic and anthropogenic controls of topsoil features in the semi-arid East Asian steppe.
3
2008
... 本研究实验样地位于内蒙古自治区克什克腾旗南端的乌兰布统草原(
... 以往的研究表明, 植被退化与土壤退化是密切关联的(
... )等植物.根据3个植物类群的相对盖度(
Plant nitrogen uptake drives responses of productivity to nitrogen and water addition in a grassland.
1
2014
... 地上生物量响应比率呈先增加后降低的趋势, 重度退化草地呈先增加后基本保持不变的趋势.一些研究表明: 退化草地生态系统受氮限制(
Experimental assessment of the effects of nitrogen addition under hay-cutting and aftermath grazing on the vegetation of meadows on a Somerset peat moor.
1
1993
... 全球范围内, 有关氮添加对草地生态系统影响以及氮添加降低物种多样性机制的报道中, 只有少量实验是从退化草地恢复的角度出发(
内蒙古草地退化防治对策及碳增汇途径研究
2
2017
... 以往的研究表明, 植被退化与土壤退化是密切关联的(
... ;
内蒙古草地退化防治对策及碳增汇途径研究
2
2017
... 以往的研究表明, 植被退化与土壤退化是密切关联的(
... ;
The impact of nitrogen enrichment on grassland ecosystem stability depends on nitrogen addition level.
1
2017
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
Nitrogen enrichment and grazing accelerate vegetation restoration in degraded grassland patches.
1
2015
... 草地是全球分布最广的一个植被类型(
Root competition can cause a decline in diversity with increased productivity.
2003
Effects of resource additions on species richness and ANPP in an alpine meadow community.
2
2010
... 通过计算生物量响应比率(处理组地上生物量与对照组地上生物量的比值), 来直接比较地上生物量对氮添加处理的响应.植物高度不对称性指数反映了物种对光的竞争能力, 使用株高的基尼系数(Gini)衡量功能群之间大小的不平等(
... 覆盖度增加(约60%), 群落下层物种之间光资源竞争加剧(
The global carbon sink: A grassland perspective.
1
1998
... 草地是全球分布最广的一个植被类型(
中国草地资源的现状分析
1
2016
... 草地是全球分布最广的一个植被类型(
中国草地资源的现状分析
1
2016
... 草地是全球分布最广的一个植被类型(
Nitrogen enrichment enhances the dominance of grasses.
3
2011
... 研究结果显示, 不同退化程度的草地群落氮添加均显著增加了禾草的地上生物量及其在群落地上生物量中所占比例, 而对杂类草地上生物量没有显著影响, 但显著降低了其在地上生物量中所占的比例.杨晓霞等(2014)在青藏高原高寒草甸的研究同样发现, 氮添加促进了禾草生物量及比例的增加, 对杂类草地上生物量影响不明显但降低了其比例.Xu等(2015)的研究结果表明, 氮添加对禾草的地上生物量影响比杂类草更为明显.同时该地区之前的研究结果表明, 氮添加促使禾草地上生物量增加, 但降低了杂类草地上生物量(
... ).禾草根系比较发达(例如羊草、根茎型禾草), 具有更高的氮和水分利用效率(
... 杂类草(如星毛委陵菜等)一般处于群落下层, 在资源竞争中处于劣势.虽然氮添加可以缓解植物生长所受的养分限制, 但是由于处于群落上层的禾草具有更高的水氮利用效率(
Reduced compensatory effects explain the nitrogen-mediated reduction in stability of an alpine meadow on the Tibetan Plateau.
1
2015
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
How long do ecosystems take to recover from atmospheric nitrogen deposition?
1
2016
... 全球范围内, 有关氮添加对草地生态系统影响以及氮添加降低物种多样性机制的报道中, 只有少量实验是从退化草地恢复的角度出发(
Impact of nitrogen deposition on the species richness of grasslands.
1
2004
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
Anthropogenic nitrogen deposition predicts local grassland primary production worldwide.
1
2015
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
Influences of continuous grazing and livestock exclusion on soil properties in a degraded sandy grassland, Inner Mongolia, northern China.
2
2005
... 以往的研究表明, 植被退化与土壤退化是密切关联的(
... ).土壤退化主要表现为水土流失、土壤质地逆向改变、土壤肥力下降以及动物和微生物活性下降等(
A novel soil manganese mechanism drives plant species loss with increased nitrogen deposition in a temperate steppe.
1
2016
... 研究结果显示, 不同退化程度的草地群落氮添加均显著增加了禾草的地上生物量及其在群落地上生物量中所占比例, 而对杂类草地上生物量没有显著影响, 但显著降低了其在地上生物量中所占的比例.杨晓霞等(2014)在青藏高原高寒草甸的研究同样发现, 氮添加促进了禾草生物量及比例的增加, 对杂类草地上生物量影响不明显但降低了其比例.Xu等(2015)的研究结果表明, 氮添加对禾草的地上生物量影响比杂类草更为明显.同时该地区之前的研究结果表明, 氮添加促使禾草地上生物量增加, 但降低了杂类草地上生物量(
Disruption of metal ion homeostasis in soils is associated with nitrogen deposition-induced species loss in an Inner Mongolia steppe.
2015
Productivity and sustainability influenced by biodiversity in grassland ecosystems.
1
1996
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
Catastrophic vegetation shifts and soil degradation in terrestrial grazing systems.
1
1997
... 以往的研究表明, 植被退化与土壤退化是密切关联的(
氮添加对内蒙古退化草原生产力的短期影响
2
2016
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
... 全球范围内, 有关氮添加对草地生态系统影响以及氮添加降低物种多样性机制的报道中, 只有少量实验是从退化草地恢复的角度出发(
氮添加对内蒙古退化草原生产力的短期影响
2
2016
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
... 全球范围内, 有关氮添加对草地生态系统影响以及氮添加降低物种多样性机制的报道中, 只有少量实验是从退化草地恢复的角度出发(
Inter-annual variability in responses of belowground net primary productivity (NPP) and NPP partitioning to long-term warming and clipping in a tallgrass prairie.
1
2012
... 草地是全球分布最广的一个植被类型(
养分和水分调控下内蒙古退化草原的生长与恢复
2
2015
... 全球范围内, 有关氮添加对草地生态系统影响以及氮添加降低物种多样性机制的报道中, 只有少量实验是从退化草地恢复的角度出发(
... 地上生物量响应比率呈先增加后降低的趋势, 重度退化草地呈先增加后基本保持不变的趋势.一些研究表明: 退化草地生态系统受氮限制(
养分和水分调控下内蒙古退化草原的生长与恢复
2
2015
... 全球范围内, 有关氮添加对草地生态系统影响以及氮添加降低物种多样性机制的报道中, 只有少量实验是从退化草地恢复的角度出发(
... 地上生物量响应比率呈先增加后降低的趋势, 重度退化草地呈先增加后基本保持不变的趋势.一些研究表明: 退化草地生态系统受氮限制(
Response of aboveground biomass and diversity to nitrogen addition along a degradation gradient in the Inner Mongolian steppe, China.
2015
Diversity-dependent stability under mowing and nutrient addition: Evidence from a 7-year grassland experiment.
2
2012
... 施肥是维持草地生态系统养分平衡及促进生产力提高的重要管理措施(
... ;
青藏高原高寒草甸植物群落生物量对氮、磷添加的响应
2014
青藏高原高寒草甸植物群落生物量对氮、磷添加的响应
2014
科尔沁沙地灌溉与施肥对退化草地生产力的影响
1
2008
... 全球范围内, 有关氮添加对草地生态系统影响以及氮添加降低物种多样性机制的报道中, 只有少量实验是从退化草地恢复的角度出发(
科尔沁沙地灌溉与施肥对退化草地生产力的影响
1
2008
... 全球范围内, 有关氮添加对草地生态系统影响以及氮添加降低物种多样性机制的报道中, 只有少量实验是从退化草地恢复的角度出发(
Productivity depends more on the rate than the frequency of N addition in a temperate grassland.
1
2015
... 氮添加促进了群落地上生物量增加, 这与他人的研究结果(
Rapid plant species loss at high rates and at low frequency of N addition in temperate steppe.
2
2014
... 式中n是样方内物种数, bi是物种i的地上生物量(
... 地上生物量响应比率呈先增加后降低的趋势, 重度退化草地呈先增加后基本保持不变的趋势.一些研究表明: 退化草地生态系统受氮限制(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}