 ,2,*1
,2,*1 2
3
4
Changes in hydraulic traits of nine vascular plants from different evolutionary lineages
ZHAO Le-Wen1, CHEN Zi-Yi1, ZOU Ying1, FU Zi-Zhao1, WU Gui-Lin2,3, LIU Xiao-Rong2,3, LUO Qi2,3, LIN Yi-Xue4,2, LI Xiong-Ju1, LIU Zhi-Tong1, LIU Hui,2,*1 2
3
4
通讯作者:
| 基金资助: |
Online:2018-04-19
| Supported by: | SupportedbytheNationalNaturalScienceFoundationofChina.( |

摘要
关键词:
Abstract
Keywords:
PDF (1082KB)摘要页面多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
赵乐文, 陈梓熠, 邹滢, 付子钊, 吴桂林, 刘小容, 罗琦, 林忆雪, 李雄炬, 刘智通, 刘慧. 九种维管植物水力性状的演化趋势. 植物生态学报, 2018, 42(2): 220-228 doi:10.17521/cjpe.2017.0258
ZHAO Le-Wen, CHEN Zi-Yi, ZOU Ying, FU Zi-Zhao, WU Gui-Lin, LIU Xiao-Rong, LUO Qi, LIN Yi-Xue, LI Xiong-Ju, LIU Zhi-Tong, LIU Hui.
19世纪以来, 英国生物学家达尔文提出的生物进化论(现称演化)得到广泛认同。学说的主要观点为: 地球上所有生物起源于同一个祖先, 形成一个分支庞大的系统发育树; 在同一个种群中的个体存在着变异, 那些具有能适应环境的有利变异的个体将存活下来, 并繁殖后代, 不具有有利变异的个体就被淘汰(Darwin, 1859)。维管植物在从海洋到陆地漫长的演化过程中, 植物的生存环境在不断地改变, 因而其水分关系也应表现出逐步形成适应多样化生境如干旱环境的机制, 如提高水分传导效率, 增强抗旱性等(Brodribb et al., 2016)。因此, 研究维管植物在演化过程中水力结构和功能的变化, 有助于我们评估植物在不同水分条件下的适应性, 并据此对现今维管植物的地理分布生境有更加深入的了解。
植物从根系吸收水分, 通过茎干木质部的维管运输至枝条和叶片, 最终通过气孔的蒸腾作用散失到空气中。这一过程中表征植物水分关系的功能性状主要分为导水效率(如枝条比导水率、叶片比导水率、叶片水力导度和木质部边材密度)和失水效率(如叶片蒸腾速率、气孔导度和气孔面积指数)。枝条比导水率是指单位长度和边材面积的枝条在单位压力梯度下的水分传导速率, 是衡量枝条导水能力的最常用参数; 叶片比导水率是导水率除以该枝条上所有叶片的面积, 代表了枝条对叶片的水分供应能力(Santiago et al., 2004)。木质部边材密度的大小直接受到其中维管组织比例的影响, 是衡量枝条导水率的基本结构指标。木材密度越大的植物, 一般具有越小的枝条导水率和更高的耐旱性(Hacke et al., 2001)。叶片导水阻力是植物水分传输的瓶颈, 在整株阻力中具有较高的比重(>30%)(Escalona et al., 1999), 因此叶片水力导度是反映植物水分传输速率和效率的关键指标(Brodribb & Holbrook, 2003a; Scoffoni et al., 2012)。气孔导度是CO2和水气进出气孔的速率, 与光合作用、呼吸作用及蒸腾作用密切相关, 光合速率与气孔导度的比值是潜在水分利用效率(WUEi), 可以指示一个物种在不同水分条件下的适应能力(Nicotra & Davidson, 2010)。气孔面积指数综合了气孔密度和气孔大小的特征, 可以表征植物叶片通过气孔调控水分传导和光合速率的能力(Sack et al., 2003)。
有关植物演化过程中植物生理结构和功能的变化, 国内外有许多基于被子植物内部类群的比较研究(Nicotra et al., 2011; 张亚等, 2014)。研究方向包括水分运输系统的功能在演化过程中的加强(Boyce et al., 2009), 如叶脉密度增加(Brodribb et al., 2007); 以及水分散失调节能力在演化过程中的增强, 如气孔大小和密度的变化(Brodribb & Holbrook, 2003b), 从而影响相应的光合速率的大小(Santiago et al., 2004); 并分析这些指标对生态环境变化的响应(Buckley & Kingsolver, 2012)。此外, 对于从蕨类植物、裸子植物再到被子植物的叶片生理结构和功能变化虽然也有相应研究, 但研究者没有进行茎干或枝条尺度的分析比较(Brodribb & Field, 2010)。然而, 上述研究都只是基于叶脉或导管的形态结构参数或者数学模型来推断其水分传导速率的大小, 并没有实际测定不同类群植物的水分传导速率。对从枝条到叶片, 从水分传导到散失连续过程中水力性状的一致性以及性状间是否存在协同演化的相关研究较少, 因此需要更多有关实验数据的积累和证明。
针对以上前期研究的几点不足, 本研究选取统一生境下的3种蕨类植物、3种裸子植物和3种被子植物, 测定它们和水分传导、水分散失相关的结构和功能性状。本研究目的包括: (1)分析这些功能性状在不同类群间的变化趋势; (2)探讨枝条导水和叶片导水、枝条导水和叶片失水是否具有相关性; (3)进一步检验这些相关性是否存在协同演化关系, 从而评估这9种不同演化类群的植物对水分条件的适应能力。
1 材料和方法
1.1 研究地点与试验材料
本研究在中国科学院华南植物园(广东省广州市, 113.35° E, 23.18° N)开展, 研究地点属南亚热带季风气候, 年降水量1β700 mm, 其中80%的降水来自雨季(4-9月); 年平均气温21.2 ℃, 1月平均气温13.6 ℃, 7月平均气温28.9 ℃。地形为丘陵河谷平原, 海拔20-30 m。基于对华南植物园的前期物种调查, 按照蕨类植物、裸子植物到被子植物的演化顺序, 每个类群选取3个常见且个体数量充足的物种。通过Phylocom数据库找到3个被子植物带有枝长的系统发育树(Webb et al., 2008), 并查阅相关文献获取蕨类植物(Testo & Sundue, 2016)和裸子植物(Quinn et al., 2002)属水平的系统发育树, 从而获知每个类群内物种的原始性和系统发育位置, 并用于系统发育独立差(PIC)分析(表1)。
Table 1
表1
表1本试验中不同演化类群9种维管植物概况
Table 1
| 物种 Species | 缩略词 Abbreviation | 科 Family | |
|---|---|---|---|
| 蕨类植物 Ferns | 芒萁 Dicranopteris pedata | Dp | 里白科 Gleicheniaceae |
| 华南毛蕨 Cyclosorus parasiticus | Cp | 金星蕨科 Thelypteridaceae | |
| 乌毛蕨 Blechnum orientale | Bo | 乌毛蕨科 Blechnaceae | |
| 裸子植物 Gymnosperms | 罗汉松 Podocarpus macrophyllus | Pm | 罗汉松科 Podocarpaceae |
| 竹柏 Podocarpus nagi | Pn | 罗汉松科 Podocarpaceae | |
| 落羽杉 Taxodium distichum | Td | 杉科 Taxodiaceae | |
| 被子植物 Angiosperms | 海南木莲 Manglietia fordiana var. hainanensis | Mh | 木兰科 Magnoliaceae |
| 东京油楠 Sindora tonkinensis | St | 豆科 Fabaceae | |
| 羊蹄甲 Bauhinia purpurea | Bp | 豆科 Fabaceae |
新窗口打开|下载CSV
1.2 研究方法
水力性状测定于2016年8月在中国科学院华南植物园完成, 具体指标和方法如下。1.2.1 枝条比导水率(KS)和叶片比导水率(KL)测定
导水率的测定要求取样枝条的长度超过最大导管长度, 而蕨类和裸子植物的管胞长度在1-2 cm, 本实验室之前测定过3种被子植物的最大导管长度在47-70 cm, 所以本研究为保证所有物种测定结果的可靠性, 3种蕨类植物紧贴地面剪取叶柄长度在30 cm以上的大型羽状复叶(因为蕨类植物的根状茎直接接触土壤水分, 不是限制水分传导的主要因素, 为了和其他物种比较, 本研究用大于20 cm的叶柄来测量导水率), 裸子植物和被子植物分别剪取长度30-80 cm, 直径8-10 mm健康成熟的阳生枝条带回实验室。每个物种选取3-5株成年植株, 每株测量5个重复。
枝条木质部导水率用重力冲水法测定(Sperry et al., 1988)。在较低压力下(0.1 MPa)以20 mmol·L-1 KCl溶液冲洗枝条20 min以完全除去气泡。通过自制的测定装置, 在稳定的水流压力下(5.5 kPa)测定最大导水率(Kh)。枝条比导水率(KS)为单位边材横截面积(AS)的导水率。叶片比导水率(KL)为Kh除以该枝条上所有叶面积(AL)的导水率。由于枝条和叶比导水率(KS和KL)之间具有极强的正相关关系, 本研究中以KS代表枝条尺度的导水效率和其他性状进行相关研究。
1.2.2 叶片水力导度(Kleaf)测定
叶片水力导度用蒸腾流法测定(Guyot et al., 2012)。剪取带叶小枝条装入黑色塑料袋带回实验室, 复水24 h, 使植物叶片水分达到饱和状态。在水下切取叶片, 迅速将叶柄连接到充满水的硅胶管-量筒-天平-计算机的装置, 该套自制装置同时配有大功率LED灯、隔热水盆和风扇以加快蒸腾速率(E), 计算机每30 s记录叶片的蒸腾失水情况。当叶片蒸腾约30 min后E趋于稳定, 在已经稳定的记录中选取最后10个点的平均值用于计算Kleaf, 然后将叶片迅速移开, 擦干叶柄水分放入袋中稳定20 min后测定最终水势(Ψfinal)。并用叶面积仪(LI300C, LI-COR, Lincoln, USA)测量相应叶片的叶面积。Kleaf = E/(0 MPa - Ψfinal), 其中E用最后10个点的失水速率除以叶面积计算。每个物种选取3-5株成年植株, 每株测量3个重复。
1.2.3 蒸腾速率(E)和潜在水分利用效率(WUEi)测定
测定在天气晴朗的9:00-11:00进行, 选取充分展开、健康向阳的叶片, 对植物的气体交换指标进行测定。采用LI-6400便携式光合测量系统(LI-COR, Lincoln, USA), CO2浓度设为400 μmol·mol-1, 光强设为1β500 μmol·m-2·s-1, 达到测量植物的饱和光强。每个物种选取5株成年植株, 每株测量3个重复。测量指标包括光合速率(A)、气孔导度(gs)和蒸腾速率(E), 并以A/gs计算WUEi。
1.2.4 气孔面积指数(SPI)测定
选取叶片背面中部, 用纸巾擦净后涂一层透明指甲油, 3 min后用镊子轻轻将指甲油膜揭下, 平铺于载玻片上, 盖上盖玻片于光学显微镜下观察并拍照, 每个物种选取3片健康成熟的叶子, 每个叶片拍摄10个视野。对照片用图形软件ImageJ 1.4.8处理, 直接测量保卫细胞的长度即为气孔长度(SL), 计算视野中单位叶面积的气孔数目即为气孔密度(SD)。计算气孔面积指数, SPI = SL2 × SD (Sack et al., 2003)。
1.2.5 木材密度(WD)测定
首先说明“木材密度”沿用了木本植物测量方法中的名称, 用于描述蕨类植物只是方便比较, 并不一定准确。从几种裸子植物和被子植物取下的大量枝条中, 剪取3-5 cm长的小段新鲜枝条, 剥除树皮和髓质, 在水中浸泡过夜。因为蕨类植物不存在树皮和髓质, 所以不需要剖开去髓, 直接剪取3-5 cm长的一段叶柄在水中浸泡过夜。测量时擦干表面, 用排水法(1/10β000天平上放置盛水烧杯, 物体完全浸入水中时天平读数增加的克数即为该物体的体积毫升数)测定其水分饱和时的体积(V)。样品在70 ℃烘箱中烘干72 h后测量干质量(Mdry), 则木材密度(WD)等于Mdry/V。每个物种选取3-5段枝条进行测量。
1.3 数据统计与分析
用Excel 2013、Origin软件和R软件(R Development Core Team, 2013)进行数据整理及作图。针对本研究提出的3个目的, (1)为了分析水力性状在不同类群间的变化趋势, 对不同性状在不同类群间的差异进行nest ANOVA分析, 并对差异显著的分组进行Turkey HSD多重比较。(2)为了检验枝条导水和叶片导水、枝条导水和叶片失水是否具有相关性, 采用Pearson相关分析获得相关系数。(3)为了检验性状间是否存在协同演化关系, 用系统发育独立差(PIC)对显著相关的性状进行分析。PIC以R软件中ape包的pic功能完成(Paradis et al., 2004)。
2 结果
2.1 KS、KL和Kleaf的测定结果
9种植物的枝条和叶比导水率(KS和KL)从蕨类植物、裸子植物到被子植物逐渐升高, 且蕨类和被子植物之间的差异显著(图1A、1B)。在蕨类植物中, 芒萁的Ks和KL显著高于其他两个接近于0的物种; 裸子植物中, 罗汉松和落羽杉的KS与竹柏相似, 但差异, 羊蹄甲具有最大的Ks和KL (图1A、1B)。叶片水力导度(Kleaf)同样从蕨类植物、裸子植物到被子植它们的KL比竹柏显著更高, 表明它们单位边材面积支持的叶片面积更少; 被子植物也存在较大的种间物逐渐升高, 其中蕨类和裸子植物之间无显著差异,两者和被子植物差异显著, 被子植物中东京油楠和羊蹄甲在9种植物中Kleaf最大(图1C)。图1
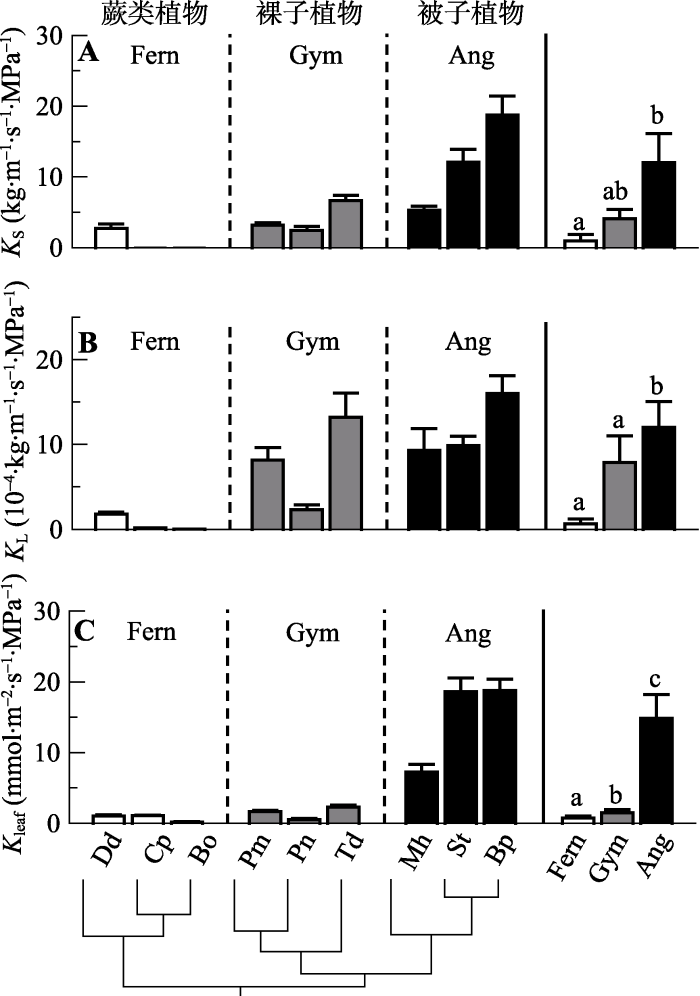 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1不同演化类群9种植物的枝条比导水率(KS)、叶片比导水率(KL)和叶片水力导度(Kleaf)(平均值±标准误差, n = 3-5)。最右图柱上不同字母是HSD多重比较的结果, 9个物种的系统发育树在柱状图最下方, 物种缩写见
Fig. 1Comparison of sapwood-specific hydraulic conductivity (KS), leaf-specific hydraulic conductivity (KL) and leaf hydraulic conductance (Kleaf) among the nine species from three evolutionary lineages (mean ± SE, n = 3-5). Letters on top of each bar in the right column are HSD multiple comparison results, the phylogenetic tree of the nine species is drawn at the bottom. See species abbrivations in
2.2 E和WUEi的测定结果
叶片E虽然表现出从蕨类植物、裸子植物到被子植物稍微升高的趋势, 但在3个演化类群间的差异并不显著(图2A)。与之类似, 叶片WUEi也有从蕨类植物、裸子植物到被子植物升高的趋势, 但在3个演化类群间的差异并不显著(图2B)。在9种植物中, 这两个指标都是被子植物中的羊蹄甲具有最大值, 其他物种之间的差异并无规律。图2
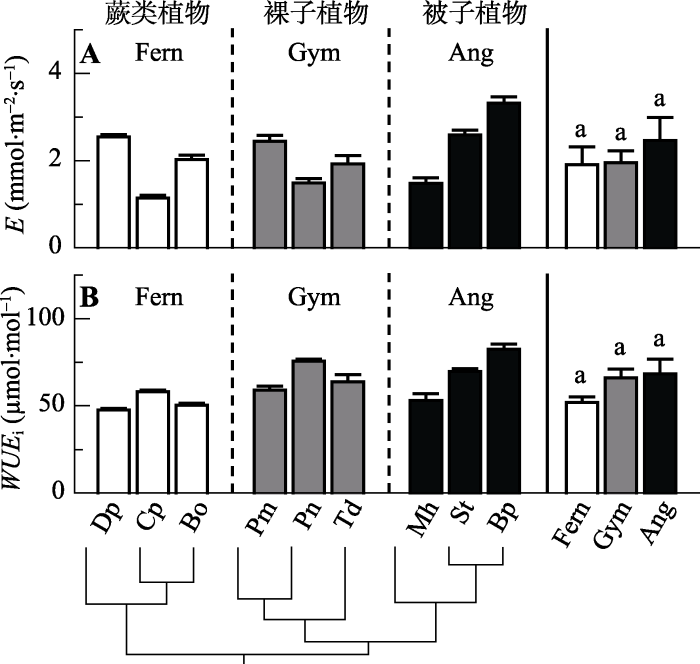 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2不同演化类群9种植物的蒸腾速率(E)和潜在水分利用效率(WUEi)(平均值±标准误差, n = 3-5)。缩写注释同
Fig. 2Comparison of leaf transpiration rate (E) and intrinsic water use efficiency (WUEi) among the nine species from three evolutionary lineages (mean ± SE, n = 3-5). Notes are the same with
2.3 SPI和WD的测定结果
基于气孔密度及气孔长度综合计算得出的SPI在3个演化类群之间并未表现出明显的变化趋势, 且在类群内表现出较大的种间差异, 如被子植物中羊蹄甲具有最小的SPI值, 而裸子植物中竹柏具有最大的SPI值(图3A)。另一个结构指标WD在3个演化类群之间也没有明显的变化趋势, 但在类群内表现出较大的种间差异, 如蕨类植物中芒萁的WD比华南毛蕨和乌毛蕨显著要高(图3B)。图3
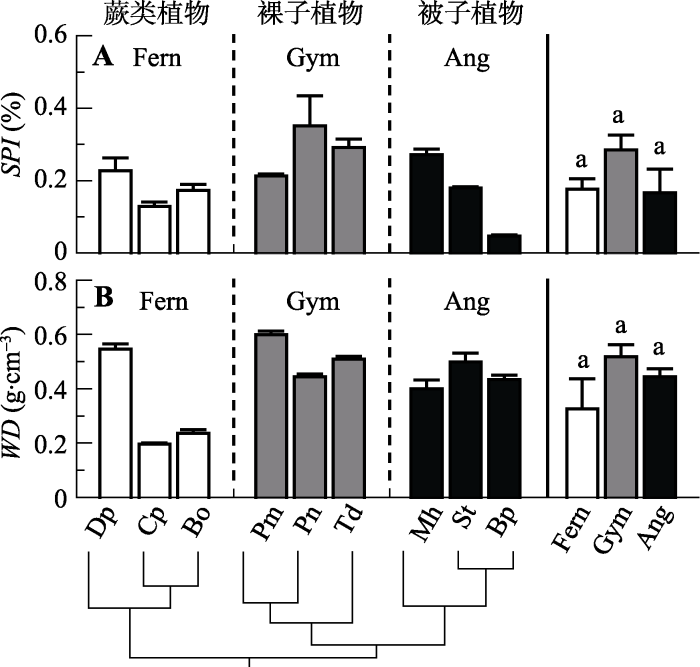 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3不同演化类群9种植物的气孔面积指数(SPI)和木材密度(WD)(平均值±标准误差, n = 3-5)。缩写注释同
Fig. 3Comparison of stomatal area index (SPI) and wood density (WD) among the nine species from three evolutionary lineages (mean ± SE, n = 3-5). Notes are the same with
2.4 水力性状之间的相关性
作为枝条尺度导水效率的衡量指标KS和叶片尺度导水效率的Kleaf, 叶片尺度失水效率的E之间均具有显著的正相关关系, 其中KS和Kleaf的相关系数更是达到0.92 (图4A、4C)。系统发育独立差分析表明KS和Kleaf, KS和E之间的正相关关系不受系统发育背景的影响, 相关性依然显著(图4B、4D)。Ks和其他结构和功能指标中的KL具有显著的正相关关系, 但和WUEi、SPI和WD的相关性不显著。图4
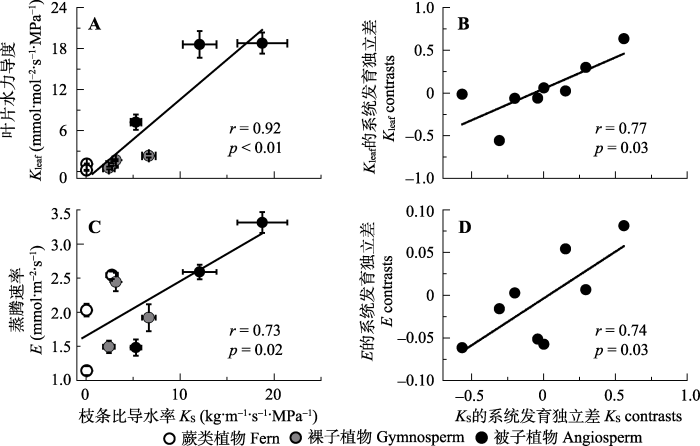 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4不同演化类群9种植物的枝条比导水率(KS)与叶片水力导度(Kleaf)(A, B)和蒸腾速率(E)(C, D)的相关性。A、C是这两对性状的直接相关, B、D是这两对性状的系统发育独立差分析。A, C内数值为平均值±标准误差(n = 3-5), A, B, C, D均报告Pearson相关系数(r)和显著性(p)。
Fig. 4Correlations between sapwood-specific hydraulic conductivity (KS) and leaf hydraulic conductance (Kleaf)(A, B), and leaf transpiration rate (E)(C, D) among the nine species from three evolutionary lineages. A, C, traditional cross-species correlations; B, D, correlations among phylogenetically independent contrasts. In A and C, data are mean ± SE (n = 3-5), Pearson correlation coefficients (r) and p values are reported.
3 讨论
3.1 九种植物水力功能性状的演化趋势
枝条和叶比导水率(KS和KL)、叶片水力导度(Kleaf)反映植物输导组织的水分传导能力, 本研究所取的9个物种, 演化地位越高的物种, 其KS、KL和Kleaf普遍显著高于演化地位低的物种(图1), 这和前人基于解剖结构指标发现的结论一致, 即在演化过程中维管植物输导组织的水分输送能力有大幅度的提高(Brodribb et al., 2005)。本研究同时发现, 蕨类植物中较原始的芒萁, 其KS和KL显著高于另外两种蕨类, 这可能是该物种适应于特殊生境造成的。广东植物志记载乌毛蕨生于较阴湿的水沟旁及坑穴边缘, 华南毛蕨生于山谷密林下或溪边湿地, 而芒萁生于强酸性土的开阔地、荒坡或林缘(吴德邻, 2006)。长期适应于开阔地充足的光环境, 芒萁具有较强的光合速率维持其快速的生长从而占领生态位(宋娟等, 2013), 较强的光合性状往往和较大的水分供给相协同, 由此演化出较强的水分传导能力。而Kleaf的测定由于给予了充分的光照和吸水源, 阴生性的蕨类也可以打开气孔表现出较大的水力导度, 因此并未在3种蕨类Kleaf之间发现显著差异(图1C)。与之类似, 3种华南地区的裸子植物均不受水分来源的限制, 不同的水力性状均是为了适应各自独特的生境演化而来。相对于在阴湿的林下生长的常绿小灌木(罗汉松和竹柏), 落羽杉虽然多生长于水边, 但其为高大的落叶乔木, 强阳生性, 强烈的光照和蒸腾需求是驱动其提高水分运输能力的主要因素(Oren et al., 2001)。在3种被子植物中, 常绿乔木海南木莲是基部被子植物中木兰科物种的典型代表, 而东京油楠和羊蹄甲分别是豆科中甘豆亚科的常绿乔木(豆科中更原始)和蝶形花亚科的落叶乔木(豆科中更演化)的代表(Azani et al., 2017)。前人研究指出落叶现象是对季节性干旱生境的一种适应, 为了补偿落叶带来的物质损耗, 落叶植物一般比常绿植物具有更高的生长速率, 并具有更高的光合效率和水分传导能力(Givnish, 2002)。E反映了植物水分散失的效率。本研究中演化地位高的几种植物的E比演化地位低的几种植物稍有提高, 但差异并不显著(图2A)。因此可以推测, 在演化中维管植物叶片的E有小幅度提高。这可能是因为在演化过程中维管植物叶片光合作用的适应能力提高, 对气孔的调节能力增大, 使得光合作用和蒸腾作用的协同性提高, 在水分充足的前提下E会有所提高(Tuzet et al., 2003)。WUEi反映了植物利用水分、同化CO2的综合能力(Nicotra & Davidson, 2010; 展小云等, 2012)。本研究中9种维管植物的WUEi虽然有提高的趋势, 但差异并不显著(图2B)。虽然可以推测演化地位越高的物种WUEi也越高, 即利用同样的水分, 同化更多的CO2, 更能适应在干旱条件下生存, 但本实验测定的植物都生长在植物园水分充足的生境中, 因此WUEi的差异并不显著, 需要进一步加强对抗旱性指标如叶片的膨压丧失点的测定和比较。
本研究通过比较9种植物的水力性状, 发现不同生境对植物输导组织水分传导能力具有一定的影响, 物种可能采取不同的策略来应对不同的水分条件。然而来自不同演化类群植物水力功能性状的变化趋势是可预测的, 即较原始的物种导水效率较低, 其分布范围更容易被水分条件限制(水分充足时, 导水率高是满足植物生长的前提; 水分不足时,抗旱性高是保证植物生存的因素), 而被子植物具有更快的导水和失水效率, 从而具备对更多样化的水分条件的适应潜力。
3.2 九种植物水力结构性状的演化趋势
SPI由气孔密度及气孔长度综合计算得出, 是叶片结构性状中的重要指标, 一定程度上体现出植物光合作用与蒸腾作用的强度(Sack et al., 2003)。本研究的9种植物, 不同演化地位物种的SPI并未表现出明显的变化趋势, 这并不符合前人发现的规律。前人发现演化过程中, 叶片气孔变小、气孔密度变大, 最终SPI和气孔导度都有变大的趋势, 这是被子植物在古气候CO2降低过程中增强适应性, 从而竞争排除掉蕨类植物和裸子植物, 在全球范围内扩大生境的关键一步(Franks & Beerling, 2009)。同时, 大量前人研究也表明, 不同种类的植物或植物生长的不同时期气孔的指标差异较大(Sack et al., 2003), 并且气孔的各项指标受本世代物种生长地环境的强烈影响。如本实验中9种植物均来自水分充足的生境, 气孔发育过程中不受干旱胁迫, 且本实验在不同演化类群所取的物种数较少, 也会掩盖可能存在的趋势。WD在9种植物中没有明显的变化趋势, 而且在类群内存在较大的种间差异。虽然WD越小表明茎干中的维管比例越多, 植物的导水效率可能会越高(Hacke et al., 2001)。但另一方面, 在演化过程中, 枝干的机械支撑能力也在逐渐增强(如蕨类多是林下草本, 仅有很少的树蕨种类; 但裸子和被子植物则以高大乔木的形态成为很多生境的优势种), 会降低相应的维管组织在枝干中的比例, 两种作用平衡之下, 导致本研究结果变化趋势不明显, 且有较大的种间差异。此外也有前人研究指出, WD并不能很好地衡量维管占比的多少, 因此需要直接测量维管的直径和密度来作为枝条水分传导能力的结构指标(Fan et al., 2012)。在叶片尺度, 除了叶脉密度和叶脉直径如何影响Kleaf的研究, 最新的研究把关注点也投向了叶肉结构的测量, 以及叶脉阻力和叶肉阻力如何共同影响Kleaf (Trifiló et al., 2016)。因此需要通过对更多物种的枝条木质部、叶脉和叶肉组织导水结构如叶肉水力导度的研究, 得出不同类群植物水力性状演化趋势的更清晰结论。
3.3 九种植物枝条和叶片水力性状之间的相关性
本研究中9种植物枝条尺度(KS)和叶片尺度(KL、Kleaf和E)的水力功能性状之间存在显著的协同效应, 这符合植物水分运输和散失的平衡关系, 即更大的水分蒸腾需要更大的枝条和叶片导水来支撑, 和前人在被子植物中的研究结论相一致(Ishida et al., 2008; Liu et al., 2015b)。而且枝条和叶片水力性状的协同性不受系统发育的影响, 即此相关性是协同演化的结果, 是系统发育树基部节点(如科)的分异比末端节点(如属)的分异更大导致(Freckleton et al., 2002)。这意味着枝条和叶片的水力性状的协同性更趋向于一种物理性规律, 而与取样物种来自哪一个演化类群关系不大。本研究中9种植物的KS与SPI和WD的相关性不显著, 和前人研究结果略有不同。在被子植物中基于多个物种的研究中, WD越小, 植物枝条木质部具有越多的导管, 一般和KS呈显著的负相关关系(Liu et al., 2015a)。本研究中的维管植物中同时包含了蕨类植物和裸子植物, 这两类植物并没有演化出成熟的导管, 而利用管胞组织进行导水, 所以即使3种蕨类植物的叶柄具有较低的密度, 它们相应的导水效率依然低下, 造成本研究中KS和WD的脱节。如芒萁具有较高的WD但KS很小, 这提示我们应当加强对维管的解剖研究, 找出影响KS的关键结构性状。另一方面, SPI越大的叶片, 气孔具有更大的开张和蒸腾潜力, 因此也会拉动更大的Kleaf (Sack et al., 2003)。但本研究的9种植物中, 不但没有发现SPI与叶片尺度的KL、E和Kleaf的相关性, 还发现被子植物中的羊蹄甲在9种植物中具有最高的KL、E和Kleaf, 却具有最小最稀疏的气孔, 这一反常的现象值得进一步探讨。推测可能原因是另外几种植物的气孔在阴生环境下并未全部打开(如前所述, 蕨类中的华南毛蕨和乌毛蕨, 裸子植物中的罗汉松和竹柏, 被子植物中的海南木莲都是偏阴生环境), 或者是另外几种植物气孔的大小和密度之间的权衡导致气孔最大导度的调节能力有限(Franks et al., 2009)。最后, 本研究基于9种维管植物发现的水力功能性状之间的协同, 以及水力结构和功能性状不匹配的现象, 提示我们测定更多物种更精细的指标, 如木质部维管大小和密度、叶脉和叶肉导度等来解释功能性状随演化类群的变化趋势。
4 结论
本研究通过测量9种维管植物水力功能和结构相关的性状, 发现水分传导相关的功能性状(KS、KL 和Kleaf)从蕨类植物、裸子植物到被子植物呈现逐渐升高的趋势, 符合植物演化过程中调节能力和适应性更强的假说。但水分散失相关的功能性状(E和WUEi)和结构性状(SPI和WD)并无明显的演化趋势, 这可能是受到物种不同的生境以及取样物种较少的影响。枝条和叶片尺度的导水和失水相关的性状间存在着较好的协同性, 且此协同性受系统发育关系的影响较小。本研究的结论需要通过对更多物种和更精细和直接的水力结构性状的测定, 及物种对不同生境适应性的相关研究来进一步验证。致谢 感谢广州市珠江科技新星(Pearl River S&T Nova Program of Guangzhou)专项(201806010083)资助。感谢广东实验中学钟南山科学人才培养班的石乐义老师在研究中给予的大力支持。

参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.12705/661.3URL [本文引用: 1]

The classification of the legume family proposed here addresses the long-known non-monophyly of the traditionally recognised subfamily Caesalpinioideae, by recognising six robustly supported monophyletic subfamilies. This new classification uses as its framework the most comprehensive phylogenetic analyses of legumes to date, based on plastid matK gene sequences, and including near-complete sampling of genera (698 of the currently recognised 765 genera) and ca. 20% (3696) of known species. The matK gene region has been the most widely sequenced across the legumes, and in most legume lineages, this gene region is sufficiently variable to yield well-supported clades. This analysis resolves the same major clades as in other phylogenies of whole plastid and nuclear gene sets (with much sparser taxon sampling). Our analysis improves upon previous studies that have used large phylogenies of the Leguminosae for addressing evolutionary questions, because it maximises generic sampling and provides a phylogenetic tree that is based on a fully curated set of sequences that are vouchered and taxonomically validated. The phylogenetic trees obtained and the underlying data are available to browse and download, facilitating subsequent analyses that require evolutionary trees. Here we propose a new community-endorsed classification of the family that reflects the phylogenetic structure that is consistently resolved and recognises six subfamilies in Leguminosae: a recircumscribed Caesalpinioideae DC., Cercidoideae Legume Phylogeny Working Group (stat. nov.), Detarioideae Burmeist., Dialioideae Legume Phylogeny Working Group (stat. nov.), Duparquetioideae Legume Phylogeny Working Group (stat. nov.), and Papilionoideae DC. The traditionally recognised subfamily Mimosoideae is a distinct clade nested within the recircum-scribed Caesalpinioideae and is referred to informally as the mimosoid clade pending a forthcoming formal tribal and/or clade-based classification of the new Caesalpinioideae. We provide a key for subfamily identification, descriptions with diagnostic charactertistics for the subfamilies, figures illustrating their floral and fruit diversity, and lists of genera by subfamily. This new classification of Leguminosae represents a consensus view of the international legume systematics community; it invokes both compromise and practicality of use.
DOI:10.1098/rspb.2008.1919URL [本文引用: 1]
DOI:10.1073/pnas.1522569113URLPMID:27071104 [本文引用: 1]
Abstract The intricate patterns of veins that adorn the leaves of land plants are among the most important networks in biology. Water flows in these leaf irrigation networks under tension and is vulnerable to embolism-forming cavitations, which cut off water supply, ultimately causing leaf death. Understanding the ways in which plants structure their vein supply network to protect against embolism-induced failure has enormous ecological and evolutionary implications, but until now there has been no way of observing dynamic failure in natural leaf networks. Here we use a new optical method that allows the initiation and spread of embolism bubbles in the leaf network to be visualized. Examining embolism-induced failure of architecturally diverse leaf networks, we found that conservative rules described the progression of hydraulic failure within veins. The most fundamental rule was that within an individual venation network, susceptibility to embolism always increased proportionally with the size of veins, and initial nucleation always occurred in the largest vein. Beyond this general framework, considerable diversity in the pattern of network failure was found between species, related to differences in vein network topology. The highest-risk network was found in a fern species, where single events caused massive disruption to leaf water supply, whereas safer networks in angiosperm leaves contained veins with composite properties, allowing a staged failure of water supply. These results reveal how the size structure of leaf venation is a critical determinant of the spread of embolism damage to leaves during drought.
DOI:10.1111/j.1461-0248.2009.01410.xURLPMID:19968696 [本文引用: 1]
Angiosperm evolution transformed global ecology, and much of this impact derives from the unrivalled vegetative productivity of dominant angiosperm clades. However, the origins of high photosynthetic capacity in angiosperms remain unknown. In this study, we describe the steep trajectory of leaf vein density (Dv) evolution in angiosperms, and predict that this leaf plumbing innovation enabled a major shift in the capacity of leaves to assimilate CO2. Reconstructing leaf vein evolution from an examination of 504 angiosperm species we found a rapid three- to fourfold increase in Dv occurred during the early evolution of angiosperms. We demonstrate how this major shift in leaf vein architecture potentially allowed the maximum photosynthetic capacity in angiosperms to rise above competing groups 140100 Ma. Our data suggest that early terrestrial angiosperms produced leaves with low photosynthetic rates, but that subsequent angiosperm success is linked to a surge in photosynthetic capacity during their early diversification.
DOI:10.1104/pp.107.101352URL [本文引用: 1]
DOI:10.1046/j.1469-8137.2003.00736.xURL
Summary 6168 The hydraulic conductance of leaves ( k leaf ) was examined to determine whether this little understood component of the water transport pathway plays a role in governing leaf phenology of two deciduous dry forest trees ( Calycophyllum candidissimum and Rhedera trinervis ). 6168 k leaf was monitored in parallel with stem hydraulic conductivity (K stem ) during the transition from wet to dry season. The relationships between declining photosynthetic quantum yield during senescence and declining k leaf and K stem were compared. 6168 Divergent patterns were observed in the response of K stem to seasonal drying; however, the behaviour of k leaf was essentially similar in both species. Large (five- to ten-fold) decreases in leaf hydraulic conductance occurred before, and during the later stages of leaf senescence. During senescence, declining k leaf , which continued until leaves were ultimately shed, was associated with a concomitant decline in quantum yield. 6168 We conclude that, in these species, the loss of hydraulic conductance of the leaf vascular system is linked to, and possibly responsible for, the loss of photosynthetic capacity during leaf senescence.
DOI:10.4314/mmj.v21i4.49641URL [本文引用: 2]
The question as to what triggers stomatal closure during leaf desiccation remains controversial. This paper examines characteristics of the vascular and photosynthetic functions of the leaf to determine which responds most similarly to stomata during desiccation. Leaf hydraulic conductance ($\text{K}_{leaf}$) was measured from the relaxation kinetics of leaf water potential ($\Psi _{l}$), and a novel application of this technique allowed the response of$\text{K}_{leaf}$to$\Psi _{l}$to be determined. These "vulnerability curves" show that$\text{K}_{leaf}$is highly sensitive to$\Psi _{l}$and that the response of stomatal conductance to$\Psi _{l}$is closely correlated with the response of$\text{K}_{leaf}$to$\Psi _{l}$. The turgor loss point of leaves was also correlated with$\text{K}_{leaf}$and stomatal closure, whereas the decline in PSII quantum yield during leaf drying occurred at a lower$\Psi _{l}$than stomatal closure. These results indicate that stomatal closure is primarily coordinated with$\text{K}_{leaf}$. However, the close proximity of$\Psi _{l}$at initial stomatal closure and initial loss of$\text{K}_{leaf}$suggest that partial loss of$\text{K}_{leaf}$might occur regularly, presumably necessitating repair of embolisms.
DOI:10.1111/j.1469-8137.2004.01259.xURLPMID:15720695 [本文引用: 1]
Summary 6168 The hydraulic plumbing of vascular plant leaves varies considerably between major plant groups both in the spatial organization of veins, as well as their anatomical structure. 6168 Five conifers, three ferns and 12 angiosperm trees were selected from tropical and temperate forests to investigate whether the profound differences in foliar morphology of these groups lead to correspondingly profound differences in leaf hydraulic efficiency. 6168 We found that angiosperm leaves spanned a range of leaf hydraulic conductance from 3.9 to 36mmol m 2 s 611 MPa 611 , whereas ferns (5.9–11.4mmol m 612 s 611 MPa 611 ) and conifers (1.6–9.0mmol m 612 s 611 MPa 611 ) were uniformly less conductive to liquid water. Leaf hydraulic conductance (K leaf ) correlated strongly with stomatal conductance indicating an internal leaf-level regulation of liquid and vapour conductances. Photosynthetic capacity also increased with K leaf , however, it became saturated at values of K leaf over 20mmol m 612 s 611 MPa 611 . 6168 The data suggest that vessels in the leaves of the angiosperms studied provide them with the flexibility to produce highly conductive leaves with correspondingly high photosynthetic capacities relative to tracheid-bearing species.
DOI:10.1146/annurev-ecolsys-110411-160516URL [本文引用: 1]
Shifts in phenology and distribution in response to both recent and paleontological climate changes vary markedly in both direction and extent among species. These individualistic shifts are inconsistent with common forecasting techniques based on environmental rather than biological niches. What biological details could enhance forecasts? Organismal characteristics such as thermal and hydric limits, seasonal timing and duration of the life cycle, ecological breadth and dispersal capacity, and fitness and evolutionary potential are expected to influence climate change impacts. We review statistical and mechanistic approaches for incorporating traits in predictive models as well as the potential to use phylogeny as a proxy for traits. Traits generally account for a significant but modest fraction of the variation in phenological and range shifts. Further assembly of phenotypic and phylogenetic data coupled with the development of mechanistic approaches is essential to improved forecasts of the ecological consequences of climate change
[本文引用: 1]
DOI:10.1071/PP99019_COURL [本文引用: 1]
Long-term induced water stress in field-grown grapevine leads to a progressive decline of stomatal conductance, accompanied by a decrease in CO 2 assimilation (40%). The apparent quantum yield also decreases (59%), which may reflect a relative increase in alternative processes for electron consumption. There is also a shift to non-stomatal regulation, as judged from significant depletions (37%) in maximum photosynthesis rate at saturating CO 2 related to limited ribulose biphosphate (RuBP) regeneration, whereas small, non-significant effects are observed on carboxylation efficiency. A high correlation (87%) between photosynthesis and stomatal conductance is observed for all experimental data and declines in intercellular CO 2 concentration parallel reductions in stomatal conductance. The data show that field response of grapevines to increasing soil water deficit involves stomatal and non-stomatal effects but, due to gradually induced drought, regulation mechanisms able to adjust mesophyll capacity to the average CO 2 supply. The non-stomatal adjustment seems to be exerted mainly in metabolic pathways related with the RuBP regeneration. Contrasting characteristics were observed for both cultivars. Tempranillo exploited the non-stressful conditions successfully, whereas Manto Negro, responding to its reputation as more drought resistant, showed a higher intrinsic water use efficiency, particularly for low water availability. This advantage seems to be due to lower non-stomatal limitations.
DOI:10.1111/j.1365-2745.2011.01939.xURL [本文引用: 1]
1. Stem xylem characteristics have a great impact on growth and adult stature of trees because of their role in mechanical support, long-distance water transport and whole-plant carbon allocation. Yet, despite the potential causal link between xylem traits and plant growth/adult stature, most studies have tried to link wood density, an indirect but easy to measure proxy for wood properties, to tree growth and size.2. To determine whether xylem traits outperform wood density as predictors of tree growth and stature, we evaluated the covariation among wood density, xylem anatomical traits, tree diameter growth rate and adult stature in 40 Asian tropical tree species through principal component analyses and through bivariate correlation, both across species and across phylogenetically independent contrasts.3. Vessel diameter exhibited a tight negative relationship with vessel frequency. Wood density showed a significant correlation with vessel diameter and density, but not with vessel fraction. Most correlations between functional traits indicate adaptive coordination, demonstrated by significant correlations between phylogenetically independent contrasts.4. Across species, diameter growth rate and adult stature were positively correlated with vessel lumen diameter and potential hydraulic conductivity, but not with wood density. Thus, our results suggest that xylem anatomical traits that are linked to hydraulic conductivity are better predictors of tree growth rate and adult stature than wood density.5. Synthesis. We found that xylem anatomical traits have a more significant influence on whole-plant performance due to their direct association with stem hydraulic conductivity, whereas wood density is decoupled from hydraulic function due to complex variations in xylem components.
DOI:10.1073/pnas.0904209106URL [本文引用: 2]
DOI:10.1111/j.1365-3040.2009.002031.xURLPMID:19682293
Abstract Maximum stomatal conductance to water vapour and CO2 (gwmax, gcmax, respectively), which are set at the time of leaf maturity, are determined predominantly by stomatal size (S) and density (D). In theory, many combinations of S and D yield the same gwmax and gcmax, so there is no inherent correlation between S and D, or between S, D and maximum stomatal conductance. However, using basic equations for gas diffusion through stomata of different sizes, we show that a negative correlation between S and D offers several advantages, including plasticity in gwmax and gcmax with minimal change in epidermal area allocation to stomata. Examination of the relationship between S and D in Eucalyptus globulus seedlings and coppice shoots growing in the field under high and low rainfall revealed a strong negative relationship between S and D, whereby S decreased with increasing D according to a negative power function. The results provide evidence that plasticity in maximum stomatal conductance may be constrained by a negative S versus D relationship, with higher maximum stomatal conductance characterized by smaller S and higher D, and a tendency to minimize change in epidermal space allocation to stomata as S and D vary.
DOI:10.1086/343873URLPMID:18707460 [本文引用: 1]
Abstract The question is often raised whether it is statistically necessary to control for phylogenetic associations in comparative studies. To investigate this question, we explore the use of a measure of phylogenetic correlation, lambda, introduced by Pagel (1999), that normally varies between 0 (phylogenetic independence) and 1 (species' traits covary in direct proportion to their shared evolutionary history). Simulations show lambda to be a statistically powerful index for measuring whether data exhibit phylogenetic dependence or not and whether it has low rates of Type I error. Moreover, lambda is robust to incomplete phylogenetic information, which demonstrates that even partial information on phylogeny will improve the accuracy of phylogenetic analyses. To assess whether traits generally show phylogenetic associations, we present a quantitative review of 26 published phylogenetic comparative data sets. The data sets include 103 traits and were chosen from the ecological literature in which debate about the need for phylogenetic correction has been most acute. Eighty-eight percent of data sets contained at least one character that displayed significant phylogenetic dependence, and 60% of characters overall (pooled across studies) showed significant evidence of phylogenetic association. In 16% of tests, phylogenetic correlation could be neither supported nor rejected. However, most of these equivocal results were found in small phylogenies and probably reflect a lack of power. We suggest that the parameter lambda be routinely estimated when analyzing comparative data, since it can also be used simultaneously to adjust the phylogenetic correction in a manner that is optimal for the data set, and we present an example of how this may be done.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s004420100628URL [本文引用: 2]
DOI:10.1007/s00442-008-0965-6URLPMID:18297313 [本文引用: 1]
We examined 15 traits in leaves and stems related to leaf C economy and water use for 32 co-existing angiosperms at ridge sites with shallow soil in the Bonin Islands. Across species, stem density was positively correlated to leaf mass per area (LMA), leaf lifespan (LLS), and total phenolics and condensed tannins per unit leaf N (N-based), and negatively correlated to leaf osmotic potential and saturated water content in leaves. LMA and LLS were negatively correlated to photosynthetic parameters, such as area-, mass-, and N-based assimilation rates. Although stem density and leaf osmotic potential were not associated with photosynthetic parameters, they were associated with some parameters of the leaf C economy, such as LMA and LLS. In the principal component (PCA) analysis, the first three axes accounted for 74.4% of total variation. Axis 1, which explained 41.8% of the total variation, was well associated with parameters for leaf C and N economy. Similarly, axis 2, which explained 22.3% of the total variation, was associated with parameters for water use. Axis 3, which explained 10.3% of the total variation, was associated with chemical defense within leaves. Axes 1 and 2 separated functional types relatively well, i.e., creeping trees, ruderal trees, other woody plants, C(3) shrubs and forbs, palms, and CAM plants, indicating that plant functional types were characterized by similar attributes of traits related to leaf C and N economy and water use. In addition, when the plot was extended by two unrelated traits, leaf mass-based assimilation rates and stem density, it also separated these functional types. These data indicate that differences in the functional types with contrasting plant strategies can be attributed to functional integration among leaf C economy, hydraulics, and leaf longevity, and that both leaf mass-based assimilation rates and stem density are key factors reflecting the different functions of plant species.
DOI:10.1038/srep12246URLPMID:4503962 [本文引用: 1]
The early diverged Magnoliaceae shows a historical temperate-tropical distribution among lineages indicating divergent evolution, yet which ecophysiological traits are phylogenetically conserved, and whether these traits are involved in correlated evolution remain unclear. Integrating phylogeny and 20 ecophysiological traits of 27 species, from the four largest sections of Magnoliaceae, we tested the phylogenetic signals of these traits and the correlated evolution between trait pairs. Phylogenetic niche conservatism (PNC) in water-conducting and nutrient-use related traits was identified, and correlated evolution of several key functional traits was demonstrated. Among the three evergreen sections of tropical origin, Gwillimia had the lowest hydraulic-photosynthetic capacity and the highest drought tolerance compared with Manglietia and Michelia. Contrastingly, the temperate centred deciduous section, Yulania, showed high rates of hydraulic conductivity and photosynthesis at the cost of drought tolerance. This study elucidated the regulation of hydraulic and photosynthetic processes in the temperate-tropical adaptations for Magnoliaceae species, which led to strong phylogenetic signals and PNC in ecophysiological traits across divergent lineages of Magnoliaceae.
DOI:10.1093/treephys/tpv061URLPMID:26209618 [本文引用: 1]
Hydraulic segmentation between proximal and distal organs has been hypothesized to be an important protective mechanism for plants to minimize the detrimental effects of drought-induced hydraulic failure. Uncertainties still exist regarding the degree of segmentation and the role of stomatal regulation in keeping hydraulic integrity of organs at different hierarchies. In the present study, we measured hydraulic conductivity and vulnerability in stems, compound leaf petioles and leaflet laminas of Fraxinus mandshurica Rupr. and Juglans mandshurica Maxim. growing in Changbai Mountain of Northeast China to identify the main locality where hydraulic segmentation occurs along the shoot water transport pathway. Stomatal conductance in response to leaf water potential change was also measured to investigate the role of stomatal regulation in avoiding extensive transpiration-induced embolism. No major contrasts were found between stems and compound leaf petioles in either hydraulic conductivity or vulnerability to drought-induced embolism, whereas a large difference in hydraulic vulnerability exists between compound leaf petioles and leaflet laminas. Furthermore, in contrast to the relatively large safety margins in stems (4.13 and 2.04Pa) and compound leaf petioles (1.33 and 1.93Pa), leaflet lamina hydraulic systems have substantially smaller or even negative safety margins (-0.17 and 0.47Pa) in F. mandshurica and J. mandshurica. Under unstressed water conditions, gas exchange may be better optimized by allowing leaflet vascular system function with small safety margins. In the meantime, hydraulic safety of compound leaf petioles and stems are guaranteed by their large safety margins. In facing severe drought stress, larger safety margins in stems than in compound leaf petioles would allow plants to minimize the risk of catastrophic embolism in stems by sacrificing the whole compound leaves. A strong coordination between hydraulic and stomatal regulation appears to play a critical role in balancing the competing efficiency and safety requirements for xylem water transport and use in plants.
DOI:10.1071/FP09139URL [本文引用: 2]
DOI:10.1071/FP11057URL [本文引用: 1]
DOI:10.1007/s004420000497URLPMID:28547434 [本文引用: 1]
We measured the xylem sap flux in 64-year-old Taxodium distichum (L.) Richard trees growing in a flooded forest using Granier-type sensors to estimate mean canopy stomatal conductance of the stand ($G_{\text{S}}$). Temporal variations in$G_{\text{S}}$were investigated in relation to variation in vapor pressure deficit (D), photosynthetic photon flux density ($Q_{\text{o}}$), and the transpiration rate per unit of leaf area ($E_{\text{L}}$), the latter variable serving as a proxy for plant water potential. We found that$G_{\text{S}}$was only weakly related to$Q_{\text{o}}$below 500 μmol m-2s-1(r2=0.29), but unrelated to$Q_{\text{o}}$above this value. Above$Q_{\text{o}}$=500 μmol m-2s-1and D=0.6 kPa,$G_{\text{S}}$decreased linearly with increasing$E_{\text{L}}$with a poor fit (r2=0.31), and linearly with lnD with a much better fit (r2=0.81). The decrease of$G_{\text{S}}$with lnD was at a rate predicted based on a simple hydraulic model in which stomata regulate the minimum leaf water potential. Based on the hydraulic model, stomatal sensitivity to D is proportional to stomatal conductance at low D. A hurricane caused an 6541% reduction in leaf area. This resulted in a 28% increase in$G_{\text{S}}$at D=1 kPa ($G_{\text{Sref}}$), indicating only partial compensation. As predicted, the increase in$G_{\text{Sref}}$after the hurricane was accompanied by a similar increase in stomatal sensitivity to D (29%). At night,$G_{\text{Sref}}$was 6520% of the daytime value under non-limiting light ($Q_{\text{o}}$>500 μmol m-2s-1). However, stomatal sensitivity to D decreased only to 6546% (both reductions referenced to prehurricane daytime values), thus having more than twice the sensitivity expected based on hydraulic considerations alone. Therefore, non-hydraulic processes must cause heightened nighttime stomatal sensitivity to D.
DOI:10.1093/bioinformatics/btg412URLPMID:14734327 [本文引用: 1]
Analysis of Phylogenetics and Evolution (APE) is a package written in the R language for use in molecular evolution and phylogenetics. APE provides both utility functions for reading and writing data and manipulating phylogenetic trees, as well as several advanced methods for phylogenetic and evolutionary analysis (e.g. comparative and population genetic methods). APE takes advantage of the many R functions for statistics and graphics, and also provides a flexible framework for developing and implementing further statistical methods for the analysis of evolutionary processes. AVAILABILITY: The program is free and available from the official R package archive at http://cran.r-project.org/src/contrib/PACKAGES.html#ape. APE is licensed under the GNU General Public License.
[本文引用: 1]
DOI:10.1046/j.0016-8025.2003.01058.xURL [本文引用: 5]
The hydraulic conductance of the leaf lamina ( K lamina ) substantially constrains whole-plant water transport, but little is known of its association with leaf structure and function. K lamina was measured for sun and shade leaves of six woody temperate species growing in moist soil, and tested for correlation with the prevailing leaf irradiance, and with 22 other leaf traits. K lamina varied from 7.40 × 10 615 kg m 612 s 611 MPa 611 for Acer saccharum shade leaves to 2.89 × 10 614 kg m 612 s 611 MPa 611 for Vitis labrusca sun leaves. Tree sun leaves had 15–67% higher K lamina than shade leaves. K lamina was co-ordinated with traits associated with high water flux, including leaf irradiance, petiole hydraulic conductance, guard cell length, and stomatal pore area per lamina area. K lamina was also co-ordinated with lamina thickness, water storage capacitance, 1/mesophyll water transfer resistance, and, in five of the six species, with lamina perimeter/area. However, for the six species, K lamina was independent of inter-related leaf traits including leaf dry mass per area, density, modulus of elasticity, osmotic potential, and cuticular conductance. K lamina was thus co-ordinated with structural and functional traits relating to liquid-phase water transport and to maximum rates of gas exchange, but independent of other traits relating to drought tolerance and to aspects of carbon economy.
DOI:10.1007/s00442-004-1624-1URLPMID:15232729 [本文引用: 2]
We investigated how water transport capacity, wood density and wood anatomy were related to leaf photosynthetic traits in two lowland forests in Panama. Leaf-specific hydraulic conductivity ( k L ) of upper branches was positively correlated with maximum rates of net CO 2 assimilation per unit leaf area ( A area ) and stomatal conductance ( g s ) across 20 species of canopy trees. Maximum k L showed stronger correlation with A area than initial k L suggesting that allocation to photosynthetic potential is proportional to maximum water transport capacity. Terminal branch k L was negatively correlated with A area / g s and positively correlated with photosynthesis per unit N, indicating a trade-off of efficient use of water against efficient use of N in photosynthesis as water transport efficiency varied. Specific hydraulic conductivity calculated from xylem anatomical characteristics ( k theoretical ) was positively related to A area and k L , consistent with relationships among physiological measurements. Branch wood density was negatively correlated with wood water storage at saturation, k L , A area , net CO 2 assimilation per unit leaf mass ( A mass ), and minimum leaf water potential measured on covered leaves, suggesting that wood density constrains physiological function to specific operating ranges. Kinetic and static indices of branch water transport capacity thus exhibit considerable co-ordination with allocation to potential carbon gain. Our results indicate that understanding tree hydraulic architecture provides added insights to comparisons of leaf level measurements among species, and links photosynthetic allocation patterns with branch hydraulic processes.
DOI:10.1093/jxb/err270URLPMID:3254676 [本文引用: 1]
Leaf hydraulic conductance (K(leaf)) is a major determinant of photosynthetic rate in well-watered and drought-stressed plants. Previous work assessed the decline of K(leaf) with decreasing leaf water potential ((leaf)), most typically using rehydration kinetics methods, and found that species varied in the shape of their vulnerability curve, and that hydraulic vulnerability correlated with other leaf functional traits and with drought sensitivity. These findings were tested and extended, using a new steady-state evaporative flux method under high irradiance, and the function for the vulnerability curve of each species was determined individually using maximum likelihood for 10 species varying strongly in drought tolerance. Additionally, the ability of excised leaves to recover in K(leaf) with rehydration was assessed, and a new theoretical framework was developed to estimate how rehydration of measured leaves may affect estimation of hydraulic parameters. As hypothesized, species differed in their vulnerability function. Drought-tolerant species showed shallow linear declines and more negative (leaf) at 80% loss of K(leaf) (P(80)), whereas drought-sensitive species showed steeper, non-linear declines, and less negative P(80). Across species, the maximum K(leaf) was independent of hydraulic vulnerability. Recovery of K(leaf) after 1 h rehydration of leaves dehydrated below their turgor loss point occurred only for four of 10 species. Across species without recovery, a more negative P(80) correlated with the ability to maintain K(leaf) through both dehydration and rehydration. These findings indicate that resistance to K(leaf) decline is important not only in maintaining open stomata during the onset of drought, but also in enabling sustained function during drought recovery.
Journal of Tropical and Subtropical Botany, 21, 489-495.
URL [本文引用: 1]
URL [本文引用: 1]
DOI:10.1111/j.1365-3040.1988.tb01774.xURL [本文引用: 1]
Abstract Abstract Hydraulic conductivity of the xylem is computed as the quotient of mass flow rate and pressure gradient. Measurements on excised plant stems can be difficult to interpret because of time-dependent reductions in flow rate, and because of variable degrees of embolism. Using Acer saccharum Marsh. stems, we found that certain perfusing solutions including dilute fixatives (e.g. 0.05% formaldehyde) and acids with pH below 3 (e.g. 10 mol m 3 oxalic) prevent long-term decline in conductivity. Xylem embolism can be quantified by expressing the initial conductivity as a percentage of the maximum obtained after flow-impeding air emboli have been removed by repeated high-pressure (175 kPa) flushes. Correlation between microbial contamination and declining conductivity suggests that long-term (> 4h) declines are caused by microbial growth within the vessels. Unpredictable trends in short-term (< 4h) measurements may be caused by movements of air emboli in vessels and/or participate matter.
DOI:10.1016/j.ympev.2016.09.003URLPMID:27621129 [本文引用: 1]
Ferns are the second-most diverse lineage of vascular plants on Earth, yet the best-sampled time-calibrated phylogeny of the group to date includes fewer than 5% of global diversity and was published seven years ago. We present a time-calibrated phylogeny that includes nearly half of extant fern diversity. Our results are evaluated in the context of previous studies and the fossil record, and we develop new hypotheses about the radiation of leptosporangiate ferns. We used sequence data from six chloroplast regions for nearly 4000 species of ferns to generate the most comprehensive phylogeny of the group ever published. We calibrate the phylogeny with twenty-six fossils and use an array of phylogenetic methods to resolve phylogenetic relationships, estimate divergence times, and infer speciation, extinction, and net diversification rates. We infer a mid-late Silurian origin for ferns (including horsetails) and an early Carboniferous origin for leptosporangiate ferns. Most derived fern families appeared in the Cretaceous and persisted for millions of years before rapidly diversifying in the Cenozoic. We find no evidence of differential rates of diversification among terrestrial and epiphytic species. Our findings challenge previous hypotheses on the evolutionary history of ferns and present a new paradigm for their Cenozoic radiation. We estimate earlier divergences for most fern lineages than were reported in previous studies and provide evidence of extended persistence of major fern lineages prior to rapid diversification in the last fifty million years.
DOI:10.1093/jxb/erw268URLPMID:27388214 [本文引用: 1]
Abstract Drought stress can impair leaf hydraulic conductance (Kleaf), but the relative contribution of changes in the efficiency of the vein xylem water pathway and in the mesophyll route outside the xylem in driving the decline of Kleaf is still debated. We report direct measurements of dehydration-induced changes in the hydraulic resistance (R=1/K) of whole leaf (Rleaf), as well as of the leaf xylem (Rx) and extra-vascular pathways (Rox) in four Angiosperm species. Rleaf, Rx, and Rox were measured using the vacuum chamber method (VCM). Rleaf values during progressive leaf dehydration were also validated with measurements performed using the rehydration kinetic method (RKM). We analysed correlations between changes in Rx or Rox and Rleaf, as well as between morpho-anatomical traits (including dehydration-induced leaf shrinkage), vulnerability to embolism, and leaf water relation parameters. Measurements revealed that the relative contribution of vascular and extra-vascular hydraulic properties in driving Kleaf decline during dehydration is species-specific. Whilst in two study species the progressive impairment of both vascular and extra-vascular pathways contributed to leaf hydraulic vulnerability, in the other two species the vascular pathway remained substantially unaltered during leaf dehydration, and Kleaf decline was apparently caused only by changes in the hydraulic properties of the extra-vascular compartment. The Author 2016. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com.
DOI:10.1046/j.1365-3040.2003.01035.xURL [本文引用: 1]
A model that couples stomatal conductance, photosynthesis, leaf energy balance and transport of water through the soil lant tmosphere continuum is presented. Stomatal conductance in the model depends on light, temperature and intercellular CO 2 concentration via photosynthesis and on leaf water potential, which in turn is a function of soil water potential, the rate of water flow through the soil and plant, and on xylem hydraulic resistance. Water transport from soil to roots is simulated through solution of Richards' equation. The model captures the observed hysteresis in diurnal variations in stomatal conductance, assimilation rate and transpiration for plant canopies. Hysteresis arises because atmospheric demand for water from the leaves typically peaks in mid-afternoon and because of uneven distribution of soil matric potentials with distance from the roots. Potentials at the root surfaces are lower than in the bulk soil, and once soil water supply starts to limit transpiration, root potentials are substantially less negative in the morning than in the afternoon. This leads to higher stomatal conductances, CO 2 assimilation and transpiration in the morning compared to later in the day. Stomatal conductance is sensitive to soil and plant hydraulic properties and to root length density only after approximately 10 d of soil drying, when supply of water by the soil to the roots becomes limiting. High atmospheric demand causes transpiration rates, LE, to decline at a slightly higher soil water content,s , than at low atmospheric demand, but all curves of LE versus s fall on the same line when soil water supply limits transpiration. Stomatal conductance cannot be modelled in isolation, but must be fully coupled with models of photosynthesis/respiration and the transport of water from soil, through roots, stems and leaves to the atmosphere.
DOI:10.1093/bioinformatics/btn358URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
URL [本文引用: 1]
Based on the measurements of the foliar carbon content (Cmass), nitrogen content (Nmass), isotope abundance (δ13C and δ15N), and light response curve of 10 dominant plant species (Larix gmelinii, Quercus mongolica, Fraxinus mandshurica, Tilia amurensis, Acer mono, Pinus koraiensis, Cunninghamia lanceolata, Schima superba, Pinus massoniana, and Castanea henryi) in the main forests along the North-South Transect of Eastern China (NSTEC), this paper analyzed the differences and the relationships between the foliar water use efficiency and nitrogen use efficiency of the plant species. There existed significant differences in the foliar Nmass and δ15N among the plant species, manifesting as broadleaved species > coniferous species, and deciduous species > evergreen species. The maximum photosynthetic rate (Pn max) was coniferous species > broadleaved species, and deciduous species > evergreen species. Broadleaved and evergreen species tended to have higher foliar instantaneous water use efficiency (WUEi) and long-term water use efficiency (WUE), as compared to coniferous and deciduous species, but an opposite trend was observed in the foliar instantaneous nitrogen use efficiency (NUEi) and long-term nitrogen use efficiency (NUE). Moreover, there was a significant difference in the foliar NUE between evergreen and deciduous species. No significant correlation was observed between WUEi and WUE, but a significant positive correlation existed between NUEi and NUE. There was a significant negative correlation between the foliar water use efficiency and nitrogen use efficiency at instantaneous or long-term time scales. In sum, the foliar water use efficiency and nitrogen use efficiency were dominated by plant life-form, and a trade-off existed between the two resources use efficiencies.
URL [本文引用: 1]
Based on the measurements of the foliar carbon content (Cmass), nitrogen content (Nmass), isotope abundance (δ13C and δ15N), and light response curve of 10 dominant plant species (Larix gmelinii, Quercus mongolica, Fraxinus mandshurica, Tilia amurensis, Acer mono, Pinus koraiensis, Cunninghamia lanceolata, Schima superba, Pinus massoniana, and Castanea henryi) in the main forests along the North-South Transect of Eastern China (NSTEC), this paper analyzed the differences and the relationships between the foliar water use efficiency and nitrogen use efficiency of the plant species. There existed significant differences in the foliar Nmass and δ15N among the plant species, manifesting as broadleaved species > coniferous species, and deciduous species > evergreen species. The maximum photosynthetic rate (Pn max) was coniferous species > broadleaved species, and deciduous species > evergreen species. Broadleaved and evergreen species tended to have higher foliar instantaneous water use efficiency (WUEi) and long-term water use efficiency (WUE), as compared to coniferous and deciduous species, but an opposite trend was observed in the foliar instantaneous nitrogen use efficiency (NUEi) and long-term nitrogen use efficiency (NUE). Moreover, there was a significant difference in the foliar NUE between evergreen and deciduous species. No significant correlation was observed between WUEi and WUE, but a significant positive correlation existed between NUEi and NUE. There was a significant negative correlation between the foliar water use efficiency and nitrogen use efficiency at instantaneous or long-term time scales. In sum, the foliar water use efficiency and nitrogen use efficiency were dominated by plant life-form, and a trade-off existed between the two resources use efficiencies.
DOI:10.3724/SP.J.1142.2014.40320URL [本文引用: 1]
植物叶片通过气孔的水分蒸腾散失和叶脉的水分供应达到水分平衡,而基部被子植物在进化过程中叶片水分供应和蒸腾散失是否达到平衡或关联进化还缺乏了解。本研究以11种基部被子植物为材料,测定了气孔密度、气孔长度、叶脉密度和叶片厚度4个叶片性状,并结合系统发育树,利用系统发育独立对比的方法分析这些性状之间的关联进化。结果显示:沿进化方向,气孔密度和叶脉密度逐渐增加,而气孔长度和叶片厚度有减小的趋势;无论是否考虑系统发育的影响,气孔密度都与叶脉密度呈显著正相关关系,说明二者之间存在关联进化,并证实了基部被子植物叶片水分平衡假说;气孔密度和长度、叶脉密度均与叶片厚度呈显著线性相关,但在去除系统发育的影响后这种线性相关关系不再显著,说明叶片厚度与其它三个叶片性状不存在关联进化。本研究结果还表明,叶片的水分供应和散失乃至CO2通透性的平衡主导着基部被子植物叶片结构和功能的进化。
.
DOI:10.3724/SP.J.1142.2014.40320URL [本文引用: 1]
植物叶片通过气孔的水分蒸腾散失和叶脉的水分供应达到水分平衡,而基部被子植物在进化过程中叶片水分供应和蒸腾散失是否达到平衡或关联进化还缺乏了解。本研究以11种基部被子植物为材料,测定了气孔密度、气孔长度、叶脉密度和叶片厚度4个叶片性状,并结合系统发育树,利用系统发育独立对比的方法分析这些性状之间的关联进化。结果显示:沿进化方向,气孔密度和叶脉密度逐渐增加,而气孔长度和叶片厚度有减小的趋势;无论是否考虑系统发育的影响,气孔密度都与叶脉密度呈显著正相关关系,说明二者之间存在关联进化,并证实了基部被子植物叶片水分平衡假说;气孔密度和长度、叶脉密度均与叶片厚度呈显著线性相关,但在去除系统发育的影响后这种线性相关关系不再显著,说明叶片厚度与其它三个叶片性状不存在关联进化。本研究结果还表明,叶片的水分供应和散失乃至CO2通透性的平衡主导着基部被子植物叶片结构和功能的进化。
A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny, The Legume Phylogeny Working Group (LPWG)
1
2017
... id="C29">枝条和叶比导水率(KS和KL)、叶片水力导度(Kleaf)反映植物输导组织的水分传导能力, 本研究所取的9个物种, 演化地位越高的物种, 其KS、KL和Kleaf普遍显著高于演化地位低的物种(
Angiosperm leaf vein evolution was physiologically and environmentally transformative
1
2009
... id="C5">有关植物演化过程中植物生理结构和功能的变化, 国内外有许多基于被子植物内部类群的比较研究(
Revealing catastrophic failure of leaf networks under stress
1
2016
... id="C3">19世纪以来, 英国生物学家达尔文提出的生物进化论(现称演化)得到广泛认同.学说的主要观点为: 地球上所有生物起源于同一个祖先, 形成一个分支庞大的系统发育树; 在同一个种群中的个体存在着变异, 那些具有能适应环境的有利变异的个体将存活下来, 并繁殖后代, 不具有有利变异的个体就被淘汰(
Leaf hydraulic evolution led a surge in leaf photosynthetic capacity during early angiosperm diversification
1
2010
... id="C5">有关植物演化过程中植物生理结构和功能的变化, 国内外有许多基于被子植物内部类群的比较研究(
Leaf maximum photosynthetic rate and venation are linked by hydraulics
1
2007
... id="C5">有关植物演化过程中植物生理结构和功能的变化, 国内外有许多基于被子植物内部类群的比较研究(
a). Changes in leaf hydraulic conductance during leaf shedding in seasonally dry tropical forest
2003
b). Stomatal closure during leaf dehydration, correlation with other leaf physiological traits
2
2003
... id="C4">植物从根系吸收水分, 通过茎干木质部的维管运输至枝条和叶片, 最终通过气孔的蒸腾作用散失到空气中.这一过程中表征植物水分关系的功能性状主要分为导水效率(如枝条比导水率、叶片比导水率、叶片水力导度和木质部边材密度)和失水效率(如叶片蒸腾速率、气孔导度和气孔面积指数).枝条比导水率是指单位长度和边材面积的枝条在单位压力梯度下的水分传导速率, 是衡量枝条导水能力的最常用参数; 叶片比导水率是导水率除以该枝条上所有叶片的面积, 代表了枝条对叶片的水分供应能力(
... id="C5">有关植物演化过程中植物生理结构和功能的变化, 国内外有许多基于被子植物内部类群的比较研究(
Leaf hydraulic capacity in ferns, conifers and angiosperms, impacts on photosynthetic maxima
1
2005
... id="C29">枝条和叶比导水率(KS和KL)、叶片水力导度(Kleaf)反映植物输导组织的水分传导能力, 本研究所取的9个物种, 演化地位越高的物种, 其KS、KL和Kleaf普遍显著高于演化地位低的物种(
Functional and phylogenetic approaches to forecasting species responses to climate change
1
2012
... id="C5">有关植物演化过程中植物生理结构和功能的变化, 国内外有许多基于被子植物内部类群的比较研究(
On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. John Murray
1
1859
... id="C3">19世纪以来, 英国生物学家达尔文提出的生物进化论(现称演化)得到广泛认同.学说的主要观点为: 地球上所有生物起源于同一个祖先, 形成一个分支庞大的系统发育树; 在同一个种群中的个体存在着变异, 那些具有能适应环境的有利变异的个体将存活下来, 并繁殖后代, 不具有有利变异的个体就被淘汰(
Stomatal and non-stomatal limitations of photosynthesis under water stress in field-grown grapevines
1
1999
... id="C4">植物从根系吸收水分, 通过茎干木质部的维管运输至枝条和叶片, 最终通过气孔的蒸腾作用散失到空气中.这一过程中表征植物水分关系的功能性状主要分为导水效率(如枝条比导水率、叶片比导水率、叶片水力导度和木质部边材密度)和失水效率(如叶片蒸腾速率、气孔导度和气孔面积指数).枝条比导水率是指单位长度和边材面积的枝条在单位压力梯度下的水分传导速率, 是衡量枝条导水能力的最常用参数; 叶片比导水率是导水率除以该枝条上所有叶片的面积, 代表了枝条对叶片的水分供应能力(
Hydraulic conductivity traits predict growth rates and adult stature of 40 Asian tropical tree species better than wood density
1
2012
... id="C33">WD在9种植物中没有明显的变化趋势, 而且在类群内存在较大的种间差异.虽然WD越小表明茎干中的维管比例越多, 植物的导水效率可能会越高(
Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time
2
2009
... id="C32">SPI由气孔密度及气孔长度综合计算得出, 是叶片结构性状中的重要指标, 一定程度上体现出植物光合作用与蒸腾作用的强度(
... id="C35">本研究中9种植物的KS与SPI和WD的相关性不显著, 和前人研究结果略有不同.在被子植物中基于多个物种的研究中, WD越小, 植物枝条木质部具有越多的导管, 一般和KS呈显著的负相关关系(
Plasticity in maximum stomatal conductance constrained by negative correlation between stomatal size and density, an analysis using
2009
Phylogenetic analysis and comparative data, a test and review of evidence
1
2002
... id="C34">本研究中9种植物枝条尺度(KS)和叶片尺度(KL、Kleaf和E)的水力功能性状之间存在显著的协同效应, 这符合植物水分运输和散失的平衡关系, 即更大的水分蒸腾需要更大的枝条和叶片导水来支撑, 和前人在被子植物中的研究结论相一致(
Adaptive significance of evergreen vs. deciduous leaves, solving the triple paradox
1
2002
... id="C29">枝条和叶比导水率(KS和KL)、叶片水力导度(Kleaf)反映植物输导组织的水分传导能力, 本研究所取的9个物种, 演化地位越高的物种, 其KS、KL和Kleaf普遍显著高于演化地位低的物种(
Combined impacts of irradiance and dehydration on leaf hydraulic conductance, insights into vulnerability and stomatal control
1
2012
... id="C16">叶片水力导度用蒸腾流法测定(
Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure
2
2001
... id="C4">植物从根系吸收水分, 通过茎干木质部的维管运输至枝条和叶片, 最终通过气孔的蒸腾作用散失到空气中.这一过程中表征植物水分关系的功能性状主要分为导水效率(如枝条比导水率、叶片比导水率、叶片水力导度和木质部边材密度)和失水效率(如叶片蒸腾速率、气孔导度和气孔面积指数).枝条比导水率是指单位长度和边材面积的枝条在单位压力梯度下的水分传导速率, 是衡量枝条导水能力的最常用参数; 叶片比导水率是导水率除以该枝条上所有叶片的面积, 代表了枝条对叶片的水分供应能力(
... id="C33">WD在9种植物中没有明显的变化趋势, 而且在类群内存在较大的种间差异.虽然WD越小表明茎干中的维管比例越多, 植物的导水效率可能会越高(
Coordination between leaf and stem traits related to leaf carbon gain and hydraulics across 32 drought-tolerant angiosperms
1
2008
... id="C34">本研究中9种植物枝条尺度(KS)和叶片尺度(KL、Kleaf和E)的水力功能性状之间存在显著的协同效应, 这符合植物水分运输和散失的平衡关系, 即更大的水分蒸腾需要更大的枝条和叶片导水来支撑, 和前人在被子植物中的研究结论相一致(
a). Strong phylogenetic signals and phylogenetic niche conservatism in ecophysiological traits across divergent lineages of Magnoliaceae
1
2015
... id="C35">本研究中9种植物的KS与SPI和WD的相关性不显著, 和前人研究结果略有不同.在被子植物中基于多个物种的研究中, WD越小, 植物枝条木质部具有越多的导管, 一般和KS呈显著的负相关关系(
b). Coordination of xylem hydraulics and stomatal regulation in keeping the integrity of xylem water transport in shoots of two compound-leaved tree species
1
2015
... id="C34">本研究中9种植物枝条尺度(KS)和叶片尺度(KL、Kleaf和E)的水力功能性状之间存在显著的协同效应, 这符合植物水分运输和散失的平衡关系, 即更大的水分蒸腾需要更大的枝条和叶片导水来支撑, 和前人在被子植物中的研究结论相一致(
Adaptive phenotypic plasticity and plant water use
2
2010
... id="C4">植物从根系吸收水分, 通过茎干木质部的维管运输至枝条和叶片, 最终通过气孔的蒸腾作用散失到空气中.这一过程中表征植物水分关系的功能性状主要分为导水效率(如枝条比导水率、叶片比导水率、叶片水力导度和木质部边材密度)和失水效率(如叶片蒸腾速率、气孔导度和气孔面积指数).枝条比导水率是指单位长度和边材面积的枝条在单位压力梯度下的水分传导速率, 是衡量枝条导水能力的最常用参数; 叶片比导水率是导水率除以该枝条上所有叶片的面积, 代表了枝条对叶片的水分供应能力(
... id="C30">E反映了植物水分散失的效率.本研究中演化地位高的几种植物的E比演化地位低的几种植物稍有提高, 但差异并不显著(
The evolution and functional significance of leaf shape in the angiosperms
1
2011
... id="C5">有关植物演化过程中植物生理结构和功能的变化, 国内外有许多基于被子植物内部类群的比较研究(
Sensitivity of mean canopy stomatal conductance to vapor pressure deficit in a flooded Taxodium distichum L. forest: Hydraulic and non-hydraulic effects.
1
2001
... id="C29">枝条和叶比导水率(KS和KL)、叶片水力导度(Kleaf)反映植物输导组织的水分传导能力, 本研究所取的9个物种, 演化地位越高的物种, 其KS、KL和Kleaf普遍显著高于演化地位低的物种(
APE, analyses of phylogenetics and evolution in R language
1
2004
... id="C24">针对本研究提出的3个目的, (1)为了分析水力性状在不同类群间的变化趋势, 对不同性状在不同类群间的差异进行nest ANOVA分析, 并对差异显著的分组进行Turkey HSD多重比较.(2)为了检验枝条导水和叶片导水、枝条导水和叶片失水是否具有相关性, 采用Pearson相关分析获得相关系数.(3)为了检验性状间是否存在协同演化关系, 用系统发育独立差(PIC)对显著相关的性状进行分析.PIC以R软件中ape包的pic功能完成(
Familial concepts and relationships in the conifer based on
1
2002
... id="C8">基于对华南植物园的前期物种调查, 按照蕨类植物、裸子植物到被子植物的演化顺序, 每个类群选取3个常见且个体数量充足的物种.通过Phylocom数据库找到3个被子植物带有枝长的系统发育树(
The ‘hydrology’ of leaves, co-ordination of structure and function in temperature woody species
5
2003
... id="C4">植物从根系吸收水分, 通过茎干木质部的维管运输至枝条和叶片, 最终通过气孔的蒸腾作用散失到空气中.这一过程中表征植物水分关系的功能性状主要分为导水效率(如枝条比导水率、叶片比导水率、叶片水力导度和木质部边材密度)和失水效率(如叶片蒸腾速率、气孔导度和气孔面积指数).枝条比导水率是指单位长度和边材面积的枝条在单位压力梯度下的水分传导速率, 是衡量枝条导水能力的最常用参数; 叶片比导水率是导水率除以该枝条上所有叶片的面积, 代表了枝条对叶片的水分供应能力(
... id="C20">选取叶片背面中部, 用纸巾擦净后涂一层透明指甲油, 3 min后用镊子轻轻将指甲油膜揭下, 平铺于载玻片上, 盖上盖玻片于光学显微镜下观察并拍照, 每个物种选取3片健康成熟的叶子, 每个叶片拍摄10个视野.对照片用图形软件ImageJ 1.4.8处理, 直接测量保卫细胞的长度即为气孔长度(SL), 计算视野中单位叶面积的气孔数目即为气孔密度(SD).计算气孔面积指数, SPI = SL2 × SD (
... id="C32">SPI由气孔密度及气孔长度综合计算得出, 是叶片结构性状中的重要指标, 一定程度上体现出植物光合作用与蒸腾作用的强度(
... ).同时, 大量前人研究也表明, 不同种类的植物或植物生长的不同时期气孔的指标差异较大(
... id="C35">本研究中9种植物的KS与SPI和WD的相关性不显著, 和前人研究结果略有不同.在被子植物中基于多个物种的研究中, WD越小, 植物枝条木质部具有越多的导管, 一般和KS呈显著的负相关关系(
Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees
2
2004
... id="C4">植物从根系吸收水分, 通过茎干木质部的维管运输至枝条和叶片, 最终通过气孔的蒸腾作用散失到空气中.这一过程中表征植物水分关系的功能性状主要分为导水效率(如枝条比导水率、叶片比导水率、叶片水力导度和木质部边材密度)和失水效率(如叶片蒸腾速率、气孔导度和气孔面积指数).枝条比导水率是指单位长度和边材面积的枝条在单位压力梯度下的水分传导速率, 是衡量枝条导水能力的最常用参数; 叶片比导水率是导水率除以该枝条上所有叶片的面积, 代表了枝条对叶片的水分供应能力(
... id="C5">有关植物演化过程中植物生理结构和功能的变化, 国内外有许多基于被子植物内部类群的比较研究(
Dynamics of leaf hydraulic conductance with water status, quantification and analysis of species differences under steady state
1
2012
... id="C4">植物从根系吸收水分, 通过茎干木质部的维管运输至枝条和叶片, 最终通过气孔的蒸腾作用散失到空气中.这一过程中表征植物水分关系的功能性状主要分为导水效率(如枝条比导水率、叶片比导水率、叶片水力导度和木质部边材密度)和失水效率(如叶片蒸腾速率、气孔导度和气孔面积指数).枝条比导水率是指单位长度和边材面积的枝条在单位压力梯度下的水分传导速率, 是衡量枝条导水能力的最常用参数; 叶片比导水率是导水率除以该枝条上所有叶片的面积, 代表了枝条对叶片的水分供应能力(
鼎湖山季风常绿阔叶林不同生境蕨类植物的叶片功能性状研究
1
2013
... id="C29">枝条和叶比导水率(KS和KL)、叶片水力导度(Kleaf)反映植物输导组织的水分传导能力, 本研究所取的9个物种, 演化地位越高的物种, 其KS、KL和Kleaf普遍显著高于演化地位低的物种(
鼎湖山季风常绿阔叶林不同生境蕨类植物的叶片功能性状研究
1
2013
... id="C29">枝条和叶比导水率(KS和KL)、叶片水力导度(Kleaf)反映植物输导组织的水分传导能力, 本研究所取的9个物种, 演化地位越高的物种, 其KS、KL和Kleaf普遍显著高于演化地位低的物种(
A method for measuring hydraulic conductivity and embolism in xylem
1
1988
... id="C14">枝条木质部导水率用重力冲水法测定(
A 4000-species dataset provides new insight into the evolution of ferns
1
2016
... id="C8">基于对华南植物园的前期物种调查, 按照蕨类植物、裸子植物到被子植物的演化顺序, 每个类群选取3个常见且个体数量充足的物种.通过Phylocom数据库找到3个被子植物带有枝长的系统发育树(
The contribution of vascular and extra-vascular water pathways to drought-induced decline of leaf hydraulic conductance
1
2016
... id="C33">WD在9种植物中没有明显的变化趋势, 而且在类群内存在较大的种间差异.虽然WD越小表明茎干中的维管比例越多, 植物的导水效率可能会越高(
A coupled model of stomatal conductance, photosynthesis and transpiration
1
2003
... id="C30">E反映了植物水分散失的效率.本研究中演化地位高的几种植物的E比演化地位低的几种植物稍有提高, 但差异并不显著(
Phylocom, software for the analysis of phylogenetic community structure trait evolution
1
2008
... id="C8">基于对华南植物园的前期物种调查, 按照蕨类植物、裸子植物到被子植物的演化顺序, 每个类群选取3个常见且个体数量充足的物种.通过Phylocom数据库找到3个被子植物带有枝长的系统发育树(
1
2006
... id="C29">枝条和叶比导水率(KS和KL)、叶片水力导度(Kleaf)反映植物输导组织的水分传导能力, 本研究所取的9个物种, 演化地位越高的物种, 其KS、KL和Kleaf普遍显著高于演化地位低的物种(
1
2006
... id="C29">枝条和叶比导水率(KS和KL)、叶片水力导度(Kleaf)反映植物输导组织的水分传导能力, 本研究所取的9个物种, 演化地位越高的物种, 其KS、KL和Kleaf普遍显著高于演化地位低的物种(
中国东部南北样带森林优势植物叶片的水分利用效率和氮素利用效率
1
2012
... id="C30">E反映了植物水分散失的效率.本研究中演化地位高的几种植物的E比演化地位低的几种植物稍有提高, 但差异并不显著(
中国东部南北样带森林优势植物叶片的水分利用效率和氮素利用效率
1
2012
... id="C30">E反映了植物水分散失的效率.本研究中演化地位高的几种植物的E比演化地位低的几种植物稍有提高, 但差异并不显著(
基部被子植物气孔性状与叶脉密度的关联演化
1
2014
... id="C5">有关植物演化过程中植物生理结构和功能的变化, 国内外有许多基于被子植物内部类群的比较研究(
基部被子植物气孔性状与叶脉密度的关联演化
1
2014
... id="C5">有关植物演化过程中植物生理结构和功能的变化, 国内外有许多基于被子植物内部类群的比较研究(
Copyright © 2021 版权所有 《植物生态学报》编辑部
地址: 北京香山南辛村20号, 邮编: 100093
Tel.: 010-62836134, 62836138; Fax: 010-82599431; E-mail: apes@ibcas.ac.cn, cjpe@ibcas.ac.cn
备案号: 京ICP备16067583号-19

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}