 ,1,2,*, 陈有军1,2, 魏小星1, 涂卫国31
,1,2,*, 陈有军1,2, 魏小星1, 涂卫国31 2
3
Characteristics of nutrients in two dominant plant species and rhizospheric soils in alpine desert of the Qinghai-Xizang Plateau under contrasting climates
GOU Xiao-Lin1,2, ZHOU Qing-Ping,1,2,*, CHEN You-Jun1,2, WEI Xiao-Xing1, TU Wei-Guo31 2
and 3
通讯作者:
| 基金资助: |
Online:2018-01-20
| Fund supported: |

摘要
关键词:
Abstract
Keywords:
PDF (1182KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
本文引用格式
苟小林, 周青平, 陈有军, 魏小星, 涂卫国. 青藏高原不同气候区高寒沙地两种优势植物及其根际土壤的养分特征. 植物生态学报[J], 2018, 42(1): 133-142 doi:10.17521/cjpe.2017.0120
GOU Xiao-Lin.
干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(Dos Santos et al., 2013; Tomlinson et al., 2013; Gao et al., 2014; Riah et al., 2014)。干旱和湿润环境对植物的生命组成元素碳(C)、氮(N)、磷(P)等产生影响。在对气候变化敏感的贫瘠沙地环境条件下, 植物养分对不同气候的响应情况尚不明确。对气候变化敏感的青藏高原(Jing et al., 2015; Lehnert et al., 2016)是我国重要的生态组成部分(Ding et al., 2017), 其跨度较大, 涵盖了半湿润、半干旱、干旱等多个气候带, 生境类型丰富(林振耀和吴祥定, 1981)。全球气候变暖改变了部分区域降水格局, 降水格局的变化(Polade et al., 2014; Trenberth et al., 2014)对生态系统造成影响(杨元合和朴世龙, 2006; 何奕忻等, 2014; 张宪洲等, 2015), 以青藏高原东南缘的半湿润和东北缘的半干旱气候区为代表的气候区域, 出现了不同面积的草地沙化(李广泳等, 2014; 苟小林等, 2016)。青藏高原东南缘半湿润气候带和东北缘半干旱气候带的沙地同属于高海拔寒冷地区, 生长着相同的优势植物, 但对相同植物的养分特征在半干旱和半湿润环境下的差异研究极为缺乏。首先, 半干旱和半湿润的气候差异可能影响沙地植物对土壤养分的利用过程, 进而影响植物在生命史不同阶段的生长和繁殖, 特别是繁殖策略的选择方向会极大地影响生态系统的更新能力和物种多样性(Aronson & Shmida, 1992; Báez et al., 2013), 但极少的研究展示了高寒半干旱和半湿润条件下沙地植物对土壤养分的利用情况。其次, 植物自身养分情况也可能随着半干旱和半湿润气候差异形成的环境差异表现出不同的响应结果, 导致植物器官养分的分配差异(赵广东等, 2003; 叶鑫等, 2014), 但是并没有相关研究明确解释植物器官养分对气候差异的响应情况。再者, 半干旱和半湿润气候条件可能对土壤造成影响, 土壤养分对半干旱和半湿润气候条件可能有不同的响应情况(Yahdjian & Sala, 2010), 这可能对植物生长造成影响。现阶段的研究对青藏高原半干旱和半湿润气候条件下植物和土壤缺乏较为系统的比较, 特别是在自然生态系统中, 有关研究极少。
川西北和青海湖东分属于青藏高原东南缘和东北缘, 是青藏高原典型的寒冷草地; 川西北属于长江和黄河流域重要的水源地, 属于半湿润气候, 青海湖东是青海湖流域重要的水源涵养地, 属于半干旱气候, 川西北和青海湖东对于水源涵养和生物多样性保护具有非常重要的意义(王莺等, 2015)。两区域具有非常明显的地理气候特征: 1)两区域属于青藏高原寒冷地区, 海拔都在3 000 m以上, 两区域草地生态系统退化较严重, 有不同程度和不同面积的沙化草地(骆成凤等, 2013; 苟小林等, 2016); 2)两区域沙地分布有相同的高寒灌丛和半灌木; 3)川西北每年降水相对丰沛, 较为湿润, 且雨热同季, 而青海湖东年降水相对较少, 较为干旱; 4)两区域每年生长季较短, 每年11月到来年4月为土壤冻结期。本文以川西北和青海湖东高寒沙地上具有代表性的植物中国沙棘(Hippophae rhamnoides subsp. sinensis)和沙蒿(Artemisia desertorum)为优势的灌木和半灌木, 以自然生态系统为基础, 通过调查自然条件下半湿润(川西北)和半干旱(青海湖东)沙地上沙棘和沙蒿整株及其根际土壤碳、氮、磷含量的分布, 分析长期的半湿润和半干旱环境对青藏高原高寒沙地优势植物及根际土壤养分的影响, 为高寒沙地相同植物及其根际土壤养分对不同气候环境的响应研究提供基础。
1 材料和方法
1.1 研究区域概况
研究区域位于四川省阿坝州红原县日干乔生态保护区(31.51°-33.19° N, 101.51°-103.23° E, 海拔3 460 m)和青海省海北州海晏县湖东生态保护区(36.36°-37.05° N, 100.01°-100.86° E, 海拔3 150 m)。日干乔生态保护区属于青藏高原东南缘,为半湿润气候, 年降水量相对充沛; 青海湖湖东生态保护区属于青藏高原东北缘,为半干旱气候, 年降水量相对较少(林振耀和吴祥定, 1981)。两地区为典型的高寒草地, 高温和主要降水集中于每年5到10月, 半干旱气候地区近5年的年平均气温为1.69 ℃, 年降水量440.86 mm; 而半湿润气候地区近5年的年平均气温为2.55 ℃, 年降水量801.76 mm。土壤表层主要为高寒草甸土, 沙化地区以风沙土为主。红原县主要以牦牛为放牧家畜, 而海晏县以藏绵羊为放牧家畜。地表植物主要以适应高寒环境的中国沙棘、沙蒿、鹅绒委陵菜(Potentilla anserina)等为优势植物。1.2 样地设置及样品采集处理
2016年8月中旬在两个研究区内分别随机选择3块100 × 100 m的样地, 在每块样地内分别选取3个中国沙棘和沙蒿的植物群落, 每个群落内随机采集3株中国沙棘和沙蒿的地上枝叶及地下根, 并沿主根采集0-10 cm和10-20 cm的根际土壤, 样品保存于放有冰袋的泡沫箱中, 低温环境下迅速带回实验室进行样品分析。每块样地内的9个样品收集混合为一个样品, 作为一个重复, 半湿润和半干旱高寒沙地上植物和土壤样品各3个重复。植物样品带回实验室初步粉碎后, 在65 ℃下烘干至恒质量, 再利用粉碎机彻底粉碎后进行植物碳(C)、氮(N)、磷(P)含量测定。土壤样品带回后过1 mm筛, 去除沙土中的石子和根系等杂物。将过筛后的土壤分为两份: 一份于室温下风干, 用于土壤pH值、土壤有机碳(SOC)、土壤全氮(STN)、土壤全磷(STP)含量测定; 新鲜土壤存储于4 ℃, 用于土壤硝态氮(SNN)、土壤铵态氮(SAN)、土壤有效磷(SAP)、土壤水分含量测定。
1.3 测定及计算方法
植物样品测定: 植物样品C利用重铬酸钾和浓硫酸氧化, 硫酸亚铁滴定方法进行测定; N利用浓硫酸混合加速剂蒸馏方法进行测定; P利用钼锑抗比色法(鲁如坤, 2000)进行测定。土壤样品测定: 土壤pH值利用土壤pH计测定; 土壤水分在105 ℃下烘干至恒质量测定; SOC利用高温外热重铬酸钾氧化-容量法进行测定; STN利用凯氏法进行测定; STP利用酸溶-钼锑抗比色方法进行测定; SNN利用双波长比色法进行测定; SAN利用靛酚蓝比色方法进行测定; SAP利用钼锑抗比色法进行测定(鲁如坤, 2000)。
计算方法: 植物碳氮比(C:N) = C/N; 植物碳磷比(C:P) = C/P; 植物氮磷比(N:P) = N/P; 土壤碳氮比(SC:SN) = SOC/STN; 土壤碳磷比(SC:SP) = SOC/STP; 土壤氮磷比(SN:SP) = STN/STP。
1.4 统计分析
利用单因素方差分析和最小显著差异法(LSD)分析半干旱和半湿润沙地上植物和土壤不同养分含量的差异, 显著水平为p < 0.05。利用Pearson相关分析法分析土壤和植物养分含量之间的关系。所有数据通过SPSS 19.0计算。2 结果
2.1 不同气候区域植物器官养分含量差异
长期的半干旱和半湿润气候条件下, 沙蒿根和枝叶C、N、P含量差异显著; 中国沙棘根和枝叶C、N含量差异显著, 但P含量差异不显著(图1)。在高寒沙地上, 半湿润条件下的沙蒿根和枝叶C含量显著高于半干旱条件(图1A), 中国沙棘根的C含量规律与沙蒿相似, 但是中国沙棘枝叶的C含量显著低于半干旱条件(图1B)。长期半湿润条件下两种植物根和枝叶的N含量都明显低于半干旱条件(图1C、1D)。高寒沙地上半湿润条件下两种植物根和枝叶的P含量都低于半干旱条件, 但只有沙蒿达到显著水平(图1E、1F)。图1
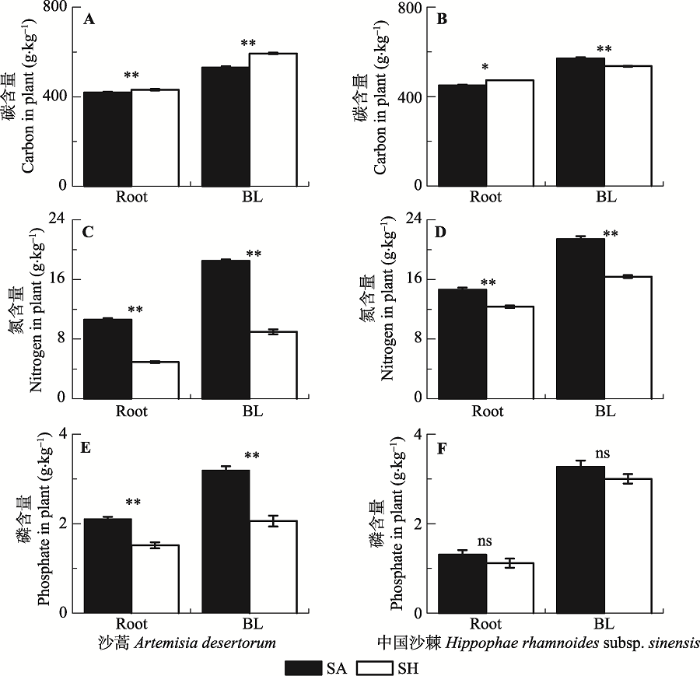 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1两种气候条件下植物器官主要养分含量的比较(平均值±标准偏差, n = 3)。*、**和ns分别代表半干旱和半湿润条件下显著性水平小于0.05、小于0.01和不显著。BL, 枝和叶; Root, 根系。SA, 半干旱; SH, 半湿润。
Fig. 1Comparisons of major nutrients in plant tissues between two different climatic conditions (mean ± SD, n = 3). *, ** and ns indicate that the differences between semi-arid and sub-humid conditions are significant at the levels of 0.05 and 0.01, and not significant, respectively. BL, branches and leaves; Root, roots. SA, semi-arid; SH, semi-humid.
2.2 不同气候条件下植物根际土壤pH值及养分含量的差异
不同气候条件下, 各植物根际土壤pH值均大于7 (图2A、2B)。半湿润条件下的土壤pH值明显低于半干旱条件下的土壤pH值(图2A)。不同气候条件下, 中国沙棘根际0-10 cm与10-20 cm土壤的pH值结果相反, 并且差异显著(图2B)。图2
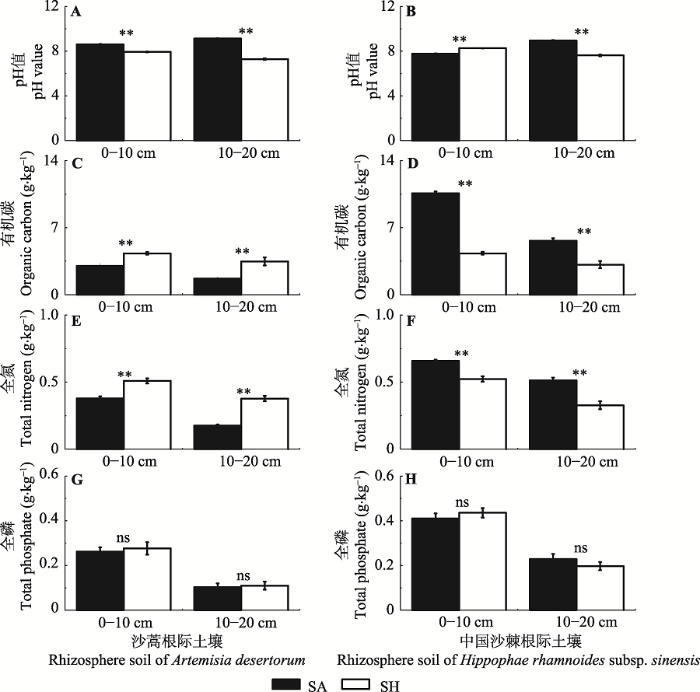 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2两种气候条件下植物根际土壤主要养分含量和pH值的比较(平均值±标准偏差, n = 3)。**和ns分别代表半干旱和半湿润环境下显著性水平小于0.01和不显著。SA, 半干旱; SH, 半湿润。
Fig. 2Comparisons of major nutrients and pH value in rhizospheric soils between two different climatic conditions (mean ± SD, n = 3). ** and ns indicate that the differences between semi-arid and sub-humid conditions are significant at the 0.01 level, and not significant, respectively. SA, semi-arid; SH, sub-humid.
不同气候条件下植物根际土壤C、N含量差异显著(图2C、2D、2E、2F), P含量差异不显著(图2G、2H)。半湿润条件下沙蒿根际土壤SOC和STN含量高于半干旱条件(图2C、2E), 中国沙棘根际土壤SOC和STN含量低于半干旱条件(图2D、2F), 并且差异显著; 而不同气候条件下两种植物根际土壤STP含量间差异不明显(图2G、2H)。
不同气候条件下两种植物根际土壤溶解性N (SAN和SNN)和SAP含量差异显著(图3A、3B、3C、3D、3E、3F)。半湿润条件下沙蒿根际土壤溶解性N 和SAP含量明显高于半干旱条件下(图3A、3C、3E), 半湿润条件下中国沙棘根际土壤溶解性N和SAP含量却明显低于半干旱条件下(图3B、3D、3F)。
图3
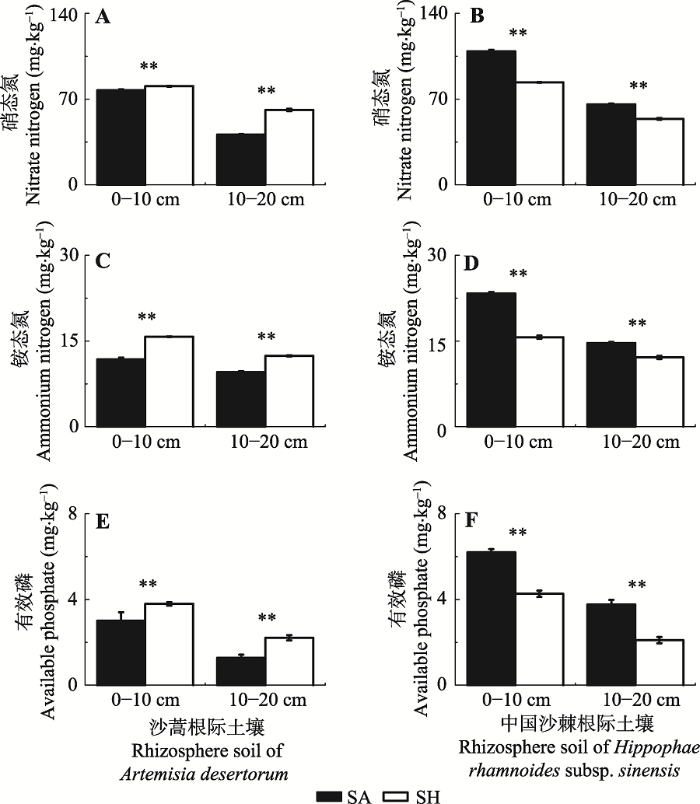 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3两种气候条件下植物根际土壤溶解性养分的比较(平均值±标准偏差, n = 3)。**代表半干旱和半湿润件下显著性水平小于0.01。SA, 半干旱; SH, 半湿润。
Fig. 3Comparisons of dissolved nutrients in rhizospheric soils between two climatic conditions (mean ± SD, n = 3). ** indicate that the differences between semi-arid and sub-humid conditions are significant at the 0.01 level. SA, semi-arid; SH, sub-humid.
2.3 不同气候条件下植物各器官养分比和根际土壤养分比
在半干旱和半湿润气候条件下植物各器官养分比与根际土壤的养分比有显著差异(表1)。在不同气候条件下, 除中国沙棘根的N:P, 中国沙棘枝叶的C:P, 沙蒿根际土壤的SC:SN和中国沙棘根际土壤SN:SP (10-20 cm土层)差异不显著外, 其余养分比值差异都达到了显著水平。半湿润条件下两种植物根及枝叶的C:N和C:P都高于半干旱条件, 而N:P却低于半干旱条件(表1)。半湿润条件下中国沙棘根际土壤SC:SN、SC:SP、SN:SP低于半干旱条件; 而沙蒿根际土壤除去10-20 cm土层的SC:SN外, 其余养分比均表现为半湿润条件下高于半干旱条件下, 跟中国沙棘根际土壤养分比值结果相反(表1)。Table 1
表1
表1两种气候条件下植物器官及土壤养分比的比较
Table 1
| 植物或根际土壤类型 Plant or rhizospheric soil | 器官或土层 Tissue or soil layer | 气候 Climate | 碳氮比 C:N (SC:SN) | 碳磷比 C:P (SC:SP) | 氮磷比 N:P (SN:SP) | |||
|---|---|---|---|---|---|---|---|---|
| M | SD | M | SD | M | SD | |||
| 沙蒿 Artemisia desertorum | 根 Roots | SA | 39.493b | 0.906 | 199.046b | 5.998 | 5.039a | 0.037 |
| SH | 87.596a | 1.527 | 284.528a | 10.402 | 3.247b | 0.063 | ||
| 枝叶 Branches and leaves | SA | 28.666b | 0.531 | 166.050b | 6.180 | 5.791a | 0.120 | |
| SH | 66.201a | 2.762 | 289.091a | 19.047 | 4.366b | 0.175 | ||
| 中国沙棘 Hippophae rhamnoides subsp. sinensis | 根 Roots | SA | 30.730b | 0.514 | 344.732b | 23.832 | 11.213a | 0.634 |
| SH | 38.210a | 0.489 | 425.255a | 38.610 | 11.139a | 1.155 | ||
| 枝叶 Branches and leaves | SA | 26.596b | 0.755 | 174.307a | 6.129 | 6.561a | 0.384 | |
| SH | 32.697a | 0.497 | 178.604a | 7.098 | 5.461b | 0.137 | ||
| 沙蒿根际土壤 Rhizospheric soil of Artemisia desertorum | 0-10 cm | SA | 7.950a | 0.752 | 11.528b | 1.336 | 1.448b | 0.048 |
| SH | 8.481a | 0.519 | 15.761a | 2.008 | 1.860a | 0.215 | ||
| 10-20 cm | SA | 9.694a | 1.384 | 16.668b | 3.595 | 1.709b | 0.180 | |
| SH | 9.233a | 1.247 | 32.617a | 7.558 | 3.540a | 0.787 | ||
| 中国沙棘根际土壤 Rhizospheric soil of Hippophae rhamnoides subsp. sinensis | 0-10 cm | SA | 16.085a | 0.334 | 25.857a | 1.315 | 1.607a | 0.066 |
| SH | 8.266b | 0.440 | 9.920b | 0.242 | 1.202b | 0.061 | ||
| 10-20 cm | SA | 10.999a | 0.486 | 24.849a | 2.788 | 2.265a | 0.305 | |
| SH | 9.567b | 0.303 | 16.118b | 3.423 | 1.679a | 0.309 | ||
新窗口打开|下载CSV
2.4 不同气候条件下植物各器官养分和根际土壤养分的关系
在半干旱和半湿润气候条件下, 沙蒿的枝叶、根和根际土壤C、N、P显著相关, 中国沙棘枝叶、根和根际土壤C、N相关性显著(表2)。在不同气候条件下, 沙蒿枝叶的C与0-10 cm和10-20 cm两个土层的SOC含量显著正相关; 沙蒿枝叶和根的N含量与土壤中的STN、SNN、SAN含量显著负相关; 沙蒿枝叶和根的P含量和土壤中的SAP显著负相关(表2)。在不同气候条件下, 中国沙棘枝叶的C含量与土壤中SOC含量显著正相关, 而中国沙棘根的C含量却与土壤中的SOC含量显著负相关; 中国沙棘根和枝叶的N含量与土壤中的STN、SNN、SAN含量显著正相关(表2)。Table 2
表2
表2两种植物养分含量与土壤养分含量之间的相关系数
Table 2
| 土层 Soil layer | 养分 Nutrient | 沙蒿 Artemisia desertorum | 中国沙棘 Hippophae rhamnoides subsp. sinensis | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 枝叶 Branches and leaves | 根 Roots | 枝叶 Branches and leaves | 根 Roots | ||||||||||
| C | N | P | C | N | P | C | N | P | C | N | P | ||
| 0-10 cm | SOC | 0.96** | -0.975** | -0.953** | 0.827* | -0.976** | -0.949** | 0.983*** | 0.992*** | 0.802ns | -0.972** | 0.986*** | 0.765ns |
| STN | 0.989*** | -0.985*** | -0.991*** | 0.861* | -0.979** | -0.986*** | 0.979** | 0.968** | 0.799ns | -0.954** | 0.994*** | 0.707ns | |
| STP | 0.42ns | -0.348ns | -0.427ns | 0.596ns | -0.333ns | -0.308ns | -0.415ns | -0.654ns | -0.425ns | 0.65ns | -0.481ns | -0.293ns | |
| SNN | 0.915* | -0.942** | -0.908* | 0.77ns | -0.951** | -0.926** | 0.982*** | 0.991*** | 0.816* | -0.967** | 0.985*** | 0.768ns | |
| SAN | 0.997** | -0.996*** | -0.990*** | 0.906* | -0.993*** | -0.979** | 0.974** | 0.996*** | 0.779ns | -0.98** | 0.978** | 0.752ns | |
| SAP | 0.906* | -0.866* | -0.899* | 0.903* | -0.848* | -0.833* | 0.966** | 0.99*** | 0.746ns | -0.974** | 0.968** | 0.712ns | |
| 10-20 cm | SOC | 0.945** | -0.963** | -0.939** | 0.761ns | -0.96** | -0.955** | 0.982*** | 0.964** | 0.781ns | -0.954** | 0.969** | 0.845* |
| STN | 0.995*** | -0.996*** | -0.997*** | 0.867* | -0.994*** | -0.994*** | 0.982*** | 0.958** | 0.813* | -0.931** | 0.963** | 0.849* | |
| STP | 0.239ns | -0.193ns | -0.192ns | 0.615ns | -0.185ns | -0.112ns | 0.576ns | 0.703ns | 0.376ns | -0.77ns | 0.664ns | 0.253ns | |
| SNN | 0.993*** | -0.999*** | -0.987*** | 0.875* | -0.997*** | -0.987*** | 0.991*** | 0.983*** | 0.798ns | -0.97** | 0.995*** | 0.761ns | |
| SAN | 0.994*** | -0.996*** | -0.986*** | 0.9* | -0.995*** | -0.979** | 0.963** | 0.99*** | 0.732ns | -0.98** | 0.961** | 0.738ns | |
| SAP | 0.986*** | -0.98** | -0.989*** | 0.902* | -0.977** | -0.982** | 0.933** | 0.994*** | 0.75ns | -0.993** | 0.956** | 0.714ns | |
新窗口打开|下载CSV
2.5 不同气候条件下植物各器官养分比和根际土壤养分比的相关关系
在半干旱和半湿润气候条件下, 沙蒿枝叶及根的N:P和土壤SN:SP显著相关, 中国沙棘枝叶C:N和土壤SC:SN显著相关, 中国沙棘根C:N、C:P分别和土壤SC:SN、SC:SP显著相关(表3)。在半干旱和半湿润条件下, 沙蒿N:P和土壤SN:SP呈显著负相关关系, 而C:N与土壤SC:SN无明显相关性, C:P只在10-20 cm土壤表现显著正相关性(表3)。在半干旱和半湿润条件下, 中国沙棘枝叶及根的C:N和土壤SC:SN显著负相关, 中国沙棘根的C:P和土壤SC:SP显著负相关, 而中国沙棘根的N:P和土壤SN:SP并没有显著的相关性(表3)。Table 3
表3
表3气候条件下植物器官及根际土壤的养分比的相关系数
Table 3
| 植物 Plant | 土层 Soil layer (cm) | 养分比 Nutrient ratio | 枝叶 Branches and leaves | 根 Roots | ||||
|---|---|---|---|---|---|---|---|---|
| C:N | C:P | N:P | C:N | C:P | N:P | |||
| 沙蒿 Artemisia desertorum | 0-10 | SC:SN | 0.397ns | 0.332ns | -0.505ns | 0.422ns | 0.336ns | -0.482ns |
| SC:SP | 0.826* | 0.758ns | -0.901* | 0.833* | 0.807ns | -0.842* | ||
| SN:SP | 0.864* | 0.814* | -0.902* | 0.86* | 0.87* | -0.843* | ||
| 10-20 | SC:SN | -0.218ns | -0.29ns | 0.114ns | -0.22ns | -0.261ns | 0.189ns | |
| SC:SP | 0.884* | 0.869* | -0.865* | 0.866* | 0.89* | -0.843* | ||
| SN:SP | 0.923** | 0.937* | -0.865* | 0.906* | 0.94** | -0.873* | ||
| 中国沙棘 Hippophae rhamnoides subsp. sinensis | 0-10 | SC:SN | -0.983*** | -0.358ns | 0.919* | -0.983*** | -0.85* | 0.015ns |
| SC:SP | -0.993*** | -0.35ns | 0.938** | -0.981** | -0.815* | 0.076ns | ||
| SN:SP | -0.991*** | -0.369ns | 0.933** | -0.959** | -0.738ns | 0.183ns | ||
| 10-20 | SC:SN | -0.892* | -0.152ns | 0.895* | -0.907* | -0.862* | -0.118ns | |
| SC:SP | -0.8ns | -0.359ns | 0.709ns | -0.843* | -0.909* | -0.316ns | ||
| SN:SP | -0.689ns | -0.373ns | 0.583ns | -0.731ns | -0.843* | -0.374ns | ||
新窗口打开|下载CSV
3 讨论
青藏高原高寒沙地在长期半干旱和半湿润气候条件下, 中国沙棘和沙蒿的养分含量在不同气候条件下差异明显。在半湿润条件下, 沙蒿整株和中国沙棘根系碳的积累增加, 但中国沙棘枝叶碳的含量降低; 而两种植物全株氮含量在半湿润条件下明显降低; 两种植物全株的磷含量也在半湿润条件下降低。部分研究表明气候变暖导致青藏高原降水增加(Wang et al., 2008; You et al., 2015), 半湿润气候区域的降水主要集中在生长季节, 雨热同季, 丰沛的降雨使植物生长迅速(黄占斌等, 2002; 郭忠升和邵明安, 2003), 有机质积累量增加, 特别是沙蒿地上部分生长迅速(Schwinning et al., 2003)。半湿润气候条件下植物的光合作用强烈, 促进有机质的积累, 但半干旱条件下, 植物除了进行光合作用, 还需要增加氮和磷的吸收量, 增加部分糖和蛋白质含量, 用以增加自身抗旱能力(周向睿等, 2014), 因此半湿润条件下植物氮和磷含量低于半干旱条件下。在高寒地区, 部分植物器官氮和磷含量随着环境因子发生变化(杨阔等, 2010), 但是在高寒地区不同气候条件下, 对植物器官碳、氮、磷含量变化机理的认识仍然缺乏, 还需要进行深入研究。各植物根际土壤中养分含量在半干旱和半湿润气候条件下差异显著。半湿润气候条件下沙蒿根系土壤中的碳、氮、磷含量较高, 而中国沙棘根际土壤碳、氮和有效磷的含量却较低。沙蒿为丛生型密集半灌木, 在地上成片分布, 郁闭度极高, 群落中极难分布其他物种, 当年生长迅速, 并且冬季地上部分枯萎, 可以依靠根茎进行无性繁殖。在半湿润条件下, 降水相对充沛, 地下根系快速生长, 地上和地下生物量较大(Wilcox et al., 2004)。在非生长季节, 地上凋落物和地下根能够提供碳、氮、磷, 反馈至土壤中(Jackson & Caldwell, 1989; 吴伊波等, 2014), 更重要的是, 沙蒿地上郁闭度较高(马全林等, 2012), 能有效地减缓降雨造成的土壤淋溶作用, 减少土壤养分损失; 非生长季节地表水分缺乏, 凋落物的分解过程缓慢, 养分归还过程缓慢, 在长期的分解和积累作用下, 半湿润气候沙地上沙蒿根际土壤养分含量变高。而中国沙棘地上部分并不密集, 群落中广泛分布着其他物种, 特别是在半湿润气候沙地上, 禾本科、菊科草本植物是中国沙棘群落中的常见物种, 随着降水条件改善, 草本物种的生长吸收土壤中的养分, 使土壤中的养分含量下降; 并且由于地上郁闭度不高, 降水可能造成的淋溶也减少了土壤中溶解性的养分, 在长期淋溶作用下, 半湿润沙地上中国沙棘根际土壤中溶解性的铵态氮、硝态氮、有效磷含量在半湿润气候条件下显著下降(图3)。而在半干旱条件下, 中国沙棘植株低矮, 相对匮乏的降水可能相对减少了对土壤养分的淋溶作用, 土壤养分损失较少。半湿润条件可能通过对土壤的淋溶作用影响了植物根际土壤养分, 但是其机理需要深入探讨。
不同气候条件下植物和土壤养分比差异显著。在半湿润气候条件下, 相对丰沛的降水使植物快速生长, 植物体碳含量增加, 碳氮比和碳磷比变高(图2; 表3), 但相对丰沛的降水同样可能导致土壤淋溶作用, 淋溶可能导致土壤溶解性氮和有效磷流失(Hu et al., 2008), 土壤氮和磷含量减少。沙蒿群落密集茂盛, 地表郁闭度高, 对淋溶产生的阻碍作用较为良好, 能有效地减缓土壤淋溶作用, 并且地表生物量较大, 形成的凋落物分解后的养分对地下生态系统具有良好的反馈作用(Kemp et al., 2003), 因此沙蒿根际土壤碳氮比、碳磷比、氮磷比在降雨沙地上高于干旱沙地(表3)。而中国沙棘对降水阻碍较小, 可能产生的淋溶作用使中国沙棘根际土壤的养分比值和沙蒿根际土壤养分比值出现了相反的结果(表3)。
在不同的气候条件下, 沙蒿的枝叶、根和根际土壤碳、氮、磷显著相关, 沙蒿枝叶及根的氮磷比和土壤氮磷比显著负相关, 中国沙棘枝叶及根的氮磷比和土壤的氮磷比没有显著相关性。半干旱和半湿润气候差异极可能通过对高寒沙地植物生理以及土壤养分淋溶产生影响(Yahdjian & Sala, 2010), 从而影响高寒沙地植物及其根际土壤的养分分布特征, 特别是青藏高原高寒沙地土壤贫瘠条件下, 土壤养分含量极容易受到外界条件的影响, 半干旱和半湿润气候降水差异显著, 可能对高寒沙地植物及土壤养分产生影响, 但是具体的影响机制和响应方式还需要进一步的研究。
4 结论
青藏高原高寒沙地上, 半干旱和半湿润气候条件下中国沙棘和沙蒿不同器官及其根际土壤养分具有明显差异。半湿润条件沙蒿碳的积累量增加, 半干旱条件下中国沙棘和沙蒿氮和磷积累量增加。半湿润条件下沙蒿根际土壤中的有机碳、全氮、铵态氮、硝态氮、有效磷的含量高于半干旱气候, 而中国沙棘根际土壤的结果却相反。不同气候条件下沙蒿和中国沙棘的养分和根际土壤养分显著相关, 两种植物养分比差异显著, 沙蒿氮磷比与其根际土壤氮磷比以及中国沙棘碳氮比与其根际土壤碳氮比呈负相关关系。有关高寒沙地上不同气候条件下优势植物及其根际土壤养分差异机理需要深入探究。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
URL [本文引用: 1]

Abstract Species diversity was monitored along a gradient from a Mediterranean region to the extreme desert in Israel. Species diversity was highest at Har-Gilo, in the Mediterranean region, with a drastic decline occurring along the transect. Correlations between interannual rainfall and species diversity were highest in the intermediate (steppic) zone, and significant in the desert only in years of relatively high rainfall. -from Authors
DOI:10.1007/s00442-012-2552-0URLPMID:23263528 [本文引用: 1]
Aridland ecosystems are predicted to be responsive to both increases and decreases in precipitation. In addition, chronic droughts may contribute to encroachment of native C 3 shrubs into C 4 -dominated grasslands. We conducted a long-term rainfall manipulation experiment in native grassland, shrubland and the grass hrub ecotone in the northern Chihuahuan Desert, USA. We evaluated the effects of 5 years of experimental drought and 4 years of water addition on plant community structure and dynamics. We assessed the effects of altered rainfall regimes on the abundance of dominant species as well as on species richness and subdominant grasses, forbs and shrubs. Nonmetric multidimensional scaling and MANOVA were used to quantify changes in species composition in response to chronic addition or reduction of rainfall. We found that drought consistently and strongly decreased cover of Bouteloua eriopoda , the dominant C 4 grass in this system, whereas water addition slightly increased cover, with little variation between years. In contrast, neither chronic drought nor increased rainfall had consistent effects on the cover of Larrea tridentata , the dominant C 3 shrub. Species richness declined in shrub-dominated vegetation in response to drought whereas richness increased or was unaffected by water addition or drought in mixed- and grass-dominated vegetation. Cover of subdominant shrubs, grasses and forbs changed significantly over time, primarily in response to interannual rainfall variability more so than to our experimental rainfall treatments. Nevertheless, drought and water addition shifted the species composition of plant communities in all three vegetation types. Overall, we found that B. eriopoda responded strongly to drought and less so to irrigation, whereas L. tridentata showed limited response to either treatment. The strong decline in grass cover and the resistance of shrub cover to rainfall reduction suggest that chronic drought may be a key factor promoting shrub dominance during encroachment into desert grassland.
DOI:10.1038/ngeo2945URL [本文引用: 1]
Climate change is expected to release carbon stored in permafrost soils. Sampling of sites across the Tibetan Plateau in the early 2000s and early 2010s reveals increased carbon stocks in shallow soils, which may offset losses from deeper soils.
[本文引用: 1]
[本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.3321/j.issn:1000-0933.2003.08.023URL [本文引用: 1]
近年来在黄土高原地区多年生林草地,出现了以土壤旱化为主要特征的土壤退 化现象.退化土壤反过来影响植物的生长和发育,最终将导致植物群落衰败和生态系统的退化,从而影响到林草植被的长期稳定,经济效益和生态效益的持续稳定发 挥,这已成为当前林草植被建设的重大问题之一.分析了土壤旱化现象与土壤干层的关系,探讨了土壤干层的划分标准.认为防止土壤旱化的主要措施就是控制林草 地密度和生产力,而控制林草地密度和生产力的理论依据就是土地植被承载力.在黄土高原大部分地区植物吸收和利用的土壤水分主要依靠当地的天然降水,土壤水 分是限制植物生长的决定因子,该类地区土地植被承载力实质上为土壤水分的植被承载力.作者定义土壤水分植被承载力为土壤水分承载植物的最大负荷.它是指在 较长时期内,在现有的条件下,当植物根系可吸收和利用土层范围内土壤水分消耗量等于或小于土壤水分补给量时,所能维持特定植物群落健康生长的最大密度.探 讨了土壤水分植被承载力的确定方法和影响因素,认为凡是影响林草地土壤水分的补给和消耗,植物群落生长发育和植物水分利用效率的因素,包括地理位置、地 形、气候、植被类型及其发育阶段,抚育管理措施都影响土壤水分植被承载力数值.开展土壤水分植被承载力研究对于林草地合理经营与管理具有重要意义.
DOI:10.3321/j.issn:1000-0933.2003.08.023URL [本文引用: 1]
近年来在黄土高原地区多年生林草地,出现了以土壤旱化为主要特征的土壤退 化现象.退化土壤反过来影响植物的生长和发育,最终将导致植物群落衰败和生态系统的退化,从而影响到林草植被的长期稳定,经济效益和生态效益的持续稳定发 挥,这已成为当前林草植被建设的重大问题之一.分析了土壤旱化现象与土壤干层的关系,探讨了土壤干层的划分标准.认为防止土壤旱化的主要措施就是控制林草 地密度和生产力,而控制林草地密度和生产力的理论依据就是土地植被承载力.在黄土高原大部分地区植物吸收和利用的土壤水分主要依靠当地的天然降水,土壤水 分是限制植物生长的决定因子,该类地区土地植被承载力实质上为土壤水分的植被承载力.作者定义土壤水分植被承载力为土壤水分承载植物的最大负荷.它是指在 较长时期内,在现有的条件下,当植物根系可吸收和利用土层范围内土壤水分消耗量等于或小于土壤水分补给量时,所能维持特定植物群落健康生长的最大密度.探 讨了土壤水分植被承载力的确定方法和影响因素,认为凡是影响林草地土壤水分的补给和消耗,植物群落生长发育和植物水分利用效率的因素,包括地理位置、地 形、气候、植被类型及其发育阶段,抚育管理措施都影响土壤水分植被承载力数值.开展土壤水分植被承载力研究对于林草地合理经营与管理具有重要意义.
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.agwat.2008.05.001URL [本文引用: 1]
Water scarcity and nitrate contamination in groundwater are serious problems in desert oases in Northwest China. Field and 15 N microplot experiments with traditional and improved water and nitrogen management were conducted in a desert oasis in Inner Mongolia Autonomous Region. Water movement, nitrogen transport and crop growth were simulated by the soil–plant system with water and solute transport model (SPWS). The model simulation results, including the water content and nitrate concentration in the soil profile, leaf area index, dry matter weight, crop N uptake and grain yield, were all in good agreement with the field measurements. The water and nitrogen use efficiency of the improved treatment were better than those of the traditional treatment. The water and nitrogen use efficiency under the traditional treatment were 2.002kg02m 613 and 2102kg02kg 611 , respectively, while under the improved treatment, they were 2.202kg02m 613 and 2602kg02kg 611 , respectively. Water drainage accounted for 24–35% of total water input (rainfall and irrigation) for the two treatments. Nitrogen loss by ammonia volatilization and denitrification was less than 5% of the total N input (including the N comes from irrigation). However, 32–61% of total nitrogen input was lost through nitrate leaching, which agreed with the 15 N isotopic result. It is impetrative to improve the water and nitrogen management in the desert oasis.
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/BF00379798URLPMID:28312530 [本文引用: 1]
Abstract Root proliferation in nutrient-rich soil patches is an important mechanism facilitating nutrient capture by plants. Although the phenomenon of root proliferation is well documented, the specific timing of this proliferation has not been investigated. We studied the timing and degree of root proliferation for three perennial species common to the Great Basin region of North America: a shrub, Artemisia tridentata, a native tussock grass, Agropyron spicatum, and an introduced tussock grass, Agropyron desertorum. One day after we applied nutrient solution to small soil patches, the mean relative growth rate of Agropyron desertorum roots in these soil patches was two to four times greater than for roots of the same plants in soil patches reated with distilled water. Most of the increased root growth came from thin, laterally branching roots within the patches. This rapid and striking root proliferation by Agropyron desertorum occurred in response to N-P-K enrichment as well as to P or N enrichment alone. A less competitive bunchgrass, Agrophyron spicatum, showed no tendency to proliferate roots in enriched soil patches during these two-week experiments. The shrub Artemisia tridentata proliferated roots within one day of initial solution injection in the N-enrichment experiment, but root proliferation of this species was more gradual and less consistent in the N-P-K and P-enrichment experiments, respectively. The ability of Agropyron desertorum to proliferate roots rapidly may partly explain both its general competitive success and its superior ability to exploit soil nutrients compared to Agropyron spicatum in Great Basin rangelands of North America.
DOI:10.1038/ncomms9159URLPMID:26328906 [本文引用: 1]
Abstract Plant biodiversity is often correlated with ecosystem functioning in terrestrial ecosystems. However, we know little about the relative and combined effects of above- and belowground biodiversity on multiple ecosystem functions (for example, ecosystem multifunctionality, EMF) or how climate might mediate those relationships. Here we tease apart the effects of biotic and abiotic factors, both above- and belowground, on EMF on the Tibetan Plateau, China. We found that a suite of biotic and abiotic variables account for up to 86% of the variation in EMF, with the combined effects of above- and belowground biodiversity accounting for 45% of the variation in EMF. Our results have two important implications: first, including belowground biodiversity in models can improve the ability to explain and predict EMF. Second, regional-scale variation in climate, and perhaps climate change, can determine, or at least modify, the effects of biodiversity on EMF in natural ecosystems.
DOI:10.1006/jare.2002.1025URL [本文引用: 1]
One of the more uncertain aspects of nutrient cycling in desert ecosystems is the extent to which decomposition is controlled by water availability. Some of this uncertainty may be partly related to the duration of the studies and/or differences in the chemical composition of the decaying litter. We conducted a 3-year field study of rainfall to determine the impact of summer (June September) drought on decomposition of leaf and root litter of two shrub species ( Larrea tridentata and Prosopis glandulosa ) in the northern Chihuahuan Desert (New Mexico, U.S.A.). In both species, leaf litter decayed at a faster rate and was less effected by drought than root litter. Drought had no influence on the rates of decomposition of leaves of either species during the first 18 months, but caused decay rates to decline by about 25% during the latter half of the study. Drought decreased decay of root litter in both species by about 25% throughout the 3 years. There was a general increase in percent nitrogen (%N) of decomposing leaf litter in both species. Root litter %N declined slightly in Larrea but remained relatively constant in Prosopis . Using a modified version of the CENTURY model, we were able to predict most aspects of observed leaf and root litter mass loss and nitrogen dynamics. Overall, the results of this study suggest that relatively large changes in precipitation produce comparatively small changes in rates of decay of both leaf and root litter.
DOI:10.1038/srep24367URLPMID:4829870 [本文引用: 1]
The Tibetan Plateau has been entitled "Third-Pole-Environment" because of its outstanding importance for the climate and the hydrology in East and South-east Asia. Its climatological and hydrological influences are strongly affected by the local grassland vegetation which is supposed to be subject to ongoing degradation. On a local scale, numerous studies focused on grassland degradation of the Tibetan pastures. However, because methods and scales substantially differed among previous studies, the overall pattern of the degradation in the Tibetan Plateau is unknown. Consequently, a satellite based approach was selected to cope with the spatial limitations. Therefore, a MODIS-based vegetation cover product was developed which is fully validated against 600 in situ measurements covering a wide extent of the Tibetan Plateau. The vegetation cover as a proxy for grassland degradation is modelled with low error rates using support vector machine regressions. To identify the changes in the vegetation cover, the trends seen in the new vegetation cover product since the beginning of the new millennium were analysed. The drivers of the vegetation changes were identified by the analysis of trends of climatic variables (precipitation and 2 m air temperature) and land-use (livestock numbers) over the same time. The results reveal that - in contrast to the prevailing opinion - pasture degradation on the Tibetan Plateau is not a generally proceeding process because areas of positive and negative changes are almost equal in extent. The positive and negative vegetation changes have regionally different triggers: While, from 2000 on, the vegetation cover has increased in the north-eastern part of the Tibetan Plateau due to increasing precipitation, it has declined in the central and western parts due to rising air temperature and declining precipitation. Increasing livestock numbers as a result of land use changes exacerbated the negative trends but, contrarily to the assumptions of former studies, were not their exclusive driver. Thus, it can be concluded that climate variability instead of overgrazing has been the primary cause for large scale vegetation cover changes on the Tibetan Plateau since the new millennium.
DOI:10.5846/stxb201211251668URL [本文引用: 1]
植被物候不仅直接受气候变化的影响,还反作用于气候变化.因此,明确植被物候变化的驱动机制对于进一步研 究气候变化与物候的相互作用具有重要的意义.选取位于青藏高原东北部的青海潮流域,利用MODIS 16d增强植被指数(EVI)合成数据,来分析草地物候时空格局特征以及不同EVImax变化趋势下草地物候期(返青期、枯黄期及生长季)的变化趋势.研 究得到以下结果:(1)在气候变化和人类活动等因素的共同作用下,青海湖流域的EVImax变化呈现多元化趋势,EVImax增加、不变、降低趋势并存; (2)1990-2012年期间,流域内温度上升、降水量增加趋势显著,温度上升速率为0.42-0.83℃/10a,降水量增加速率为 43.20-44.68 mm/10a.刚察、天峻气象站草地返青期在2001-2012年期间呈现延迟趋势,枯黄期变化趋势不显著,生长季呈现缩短趋势;(3)流域内草地返青从 4月下旬持续到6月上旬,枯黄期从8月中旬持续到10月上旬,青海湖东岸、南岸、布哈河入湖口区域以及流域西部山坡和平坦的谷底地区牧草最早进入返青期, 返青空间格局呈现由湖岸向四周高海拔地区延伸趋势,草地枯黄空间格局与返青期相反;(4)不同EVImax变化趋势下,草地返青期、枯黄期、生长季变化趋 势表现出差异.草地EVImax降低趋势下,牧草返青期呈现提前趋势,枯黄期延迟,生长季延长;EVImax增加趋势下,牧草返青期延迟,枯黄期变化不明 显,生长季缩短;EVImax不变区、农田的返青与枯黄期变化趋势并不明显,但是农田生长季缩短趋势较明显.
DOI:10.5846/stxb201211251668URL [本文引用: 1]
植被物候不仅直接受气候变化的影响,还反作用于气候变化.因此,明确植被物候变化的驱动机制对于进一步研 究气候变化与物候的相互作用具有重要的意义.选取位于青藏高原东北部的青海潮流域,利用MODIS 16d增强植被指数(EVI)合成数据,来分析草地物候时空格局特征以及不同EVImax变化趋势下草地物候期(返青期、枯黄期及生长季)的变化趋势.研 究得到以下结果:(1)在气候变化和人类活动等因素的共同作用下,青海湖流域的EVImax变化呈现多元化趋势,EVImax增加、不变、降低趋势并存; (2)1990-2012年期间,流域内温度上升、降水量增加趋势显著,温度上升速率为0.42-0.83℃/10a,降水量增加速率为 43.20-44.68 mm/10a.刚察、天峻气象站草地返青期在2001-2012年期间呈现延迟趋势,枯黄期变化趋势不显著,生长季呈现缩短趋势;(3)流域内草地返青从 4月下旬持续到6月上旬,枯黄期从8月中旬持续到10月上旬,青海湖东岸、南岸、布哈河入湖口区域以及流域西部山坡和平坦的谷底地区牧草最早进入返青期, 返青空间格局呈现由湖岸向四周高海拔地区延伸趋势,草地枯黄空间格局与返青期相反;(4)不同EVImax变化趋势下,草地返青期、枯黄期、生长季变化趋 势表现出差异.草地EVImax降低趋势下,牧草返青期呈现提前趋势,枯黄期延迟,生长季延长;EVImax增加趋势下,牧草返青期延迟,枯黄期变化不明 显,生长季缩短;EVImax不变区、农田的返青与枯黄期变化趋势并不明显,但是农田生长季缩短趋势较明显.
[本文引用: 2]
[本文引用: 2]
[本文引用: 2]
DOI:10.5846/stxb201204090494URL [本文引用: 1]
青海湖流域属于高寒草地生态系统,草地退化状况是反映该流域生态环境状况的有效指标。在对青海湖流域退化草地分类的基础上,利用遥感手段获得流域退化草地的空间分布和变化信息。采用Markov模型、退化草地动态变化度、转类指数和景观指数对青海湖流域草地退化状况时空变化进行多角度分析。研究表明2010年青海湖流域草地状况良好,未退化草甸与未退化草原在整个流域占绝对优势,是流域面积的38%。2000-2010年期间,流域草地变化不大,退化草地转类指数为-0.0384;草地变化整体上呈极轻微退化趋势,在这11a期间呈现先退化、后改善的状况。流域内不同退化草地类别的空间景观格局表现比较稳定。青海湖流域的优势草地类别是未退化草甸,主要分布在流域中部,2000-2010年期间该类别的动态变化度为1.82%,没有发生大幅度变化,时空变化都比较稳定。中度退化草甸是退化最明显的类别,即使在流域草地呈改善状况的2006-2010年期间,有近7.4万hm2退化为重度退化草甸,占中度退化草甸转变为其他类别的65%。流域中与其他类别相互转换频繁,面积变化幅度最大的类型是轻度、中度退化草甸,及重度退化草原,在流域草地退化发生改善的2006-2010年期间,这3个类别的动态变化度达到15%-21%之间;空间上它们主要分布在流域西北部、青海湖北部以及环青海湖区域。
DOI:10.5846/stxb201204090494URL [本文引用: 1]
青海湖流域属于高寒草地生态系统,草地退化状况是反映该流域生态环境状况的有效指标。在对青海湖流域退化草地分类的基础上,利用遥感手段获得流域退化草地的空间分布和变化信息。采用Markov模型、退化草地动态变化度、转类指数和景观指数对青海湖流域草地退化状况时空变化进行多角度分析。研究表明2010年青海湖流域草地状况良好,未退化草甸与未退化草原在整个流域占绝对优势,是流域面积的38%。2000-2010年期间,流域草地变化不大,退化草地转类指数为-0.0384;草地变化整体上呈极轻微退化趋势,在这11a期间呈现先退化、后改善的状况。流域内不同退化草地类别的空间景观格局表现比较稳定。青海湖流域的优势草地类别是未退化草甸,主要分布在流域中部,2000-2010年期间该类别的动态变化度为1.82%,没有发生大幅度变化,时空变化都比较稳定。中度退化草甸是退化最明显的类别,即使在流域草地呈改善状况的2006-2010年期间,有近7.4万hm2退化为重度退化草甸,占中度退化草甸转变为其他类别的65%。流域中与其他类别相互转换频繁,面积变化幅度最大的类型是轻度、中度退化草甸,及重度退化草原,在流域草地退化发生改善的2006-2010年期间,这3个类别的动态变化度达到15%-21%之间;空间上它们主要分布在流域西北部、青海湖北部以及环青海湖区域。
URL [本文引用: 1]
沙蒿与油蒿广泛分布于我国沙漠地区,是沙地植被的重要建群种和优势种。在腾格里沙漠南缘半流动沙地,实测了两种典型固沙植物沙蒿与油蒿的防风阻沙作用,从灌丛空间构型对比分析了其防风阻沙机制。结果表明,沙蒿与油蒿灌丛均具有明显的降低风速作用,但油蒿灌丛较沙蒿灌丛具有更显著的防风作用,而且对灌丛后不同位置、近地面不同高度层风速的降低程度明显不同。在灌丛后6倍株高范围内,沙蒿灌丛对50cm高度风速降低程度显著大于20cm,而油蒿灌丛对近地面20cm高度层风速降低程度显著大于50cm。在相同风速下,油蒿灌丛后20cm高度平均风速是沙蒿灌丛的1/2,而50cm高度平均风速与沙蒿灌丛相近。同时,沙蒿灌丛阻沙作用弱,而油蒿灌丛具有明显的阻沙作用,单株积沙体积达到45.2±16.1dm3,积沙重量达到72.1±25.7kg,油蒿灌丛积沙量大小与灌丛结构间存在显著的正相关。研究表明,紧密型结构的油蒿灌丛是较松散型结构沙蒿灌丛更为理想的防风固沙植物,其灌丛分枝数多、分枝角度小、生物量大且多分布在近地面层是具有显著防风阻沙作用的根本原因,该结论可为干旱区防风固沙植被建设物种选择提供依据。
URL [本文引用: 1]
沙蒿与油蒿广泛分布于我国沙漠地区,是沙地植被的重要建群种和优势种。在腾格里沙漠南缘半流动沙地,实测了两种典型固沙植物沙蒿与油蒿的防风阻沙作用,从灌丛空间构型对比分析了其防风阻沙机制。结果表明,沙蒿与油蒿灌丛均具有明显的降低风速作用,但油蒿灌丛较沙蒿灌丛具有更显著的防风作用,而且对灌丛后不同位置、近地面不同高度层风速的降低程度明显不同。在灌丛后6倍株高范围内,沙蒿灌丛对50cm高度风速降低程度显著大于20cm,而油蒿灌丛对近地面20cm高度层风速降低程度显著大于50cm。在相同风速下,油蒿灌丛后20cm高度平均风速是沙蒿灌丛的1/2,而50cm高度平均风速与沙蒿灌丛相近。同时,沙蒿灌丛阻沙作用弱,而油蒿灌丛具有明显的阻沙作用,单株积沙体积达到45.2±16.1dm3,积沙重量达到72.1±25.7kg,油蒿灌丛积沙量大小与灌丛结构间存在显著的正相关。研究表明,紧密型结构的油蒿灌丛是较松散型结构沙蒿灌丛更为理想的防风固沙植物,其灌丛分枝数多、分枝角度小、生物量大且多分布在近地面层是具有显著防风阻沙作用的根本原因,该结论可为干旱区防风固沙植被建设物种选择提供依据。
DOI:10.1038/srep04364URLPMID:24621567 [本文引用: 1]
Future changes in the number of dry days per year can either reinforce or counteract projected increases in daily precipitation intensity as the climate warms. We analyze climate model projected changes in the number of dry days using 28 coupled global climate models from the Coupled Model Intercomparison Project, version 5 (CMIP5). We find that the Mediterranean Sea region, parts of Central and South America, and western Indonesia could experience up to 30 more dry days per year by the end of this century. We illustrate how changes in the number of dry days and the precipitation intensity on precipitating days combine to produce changes in annual precipitation, and show that over much of the subtropics the change in number of dry days dominates the annual changes in precipitation and accounts for a large part of the change in interannual precipitation variability.
DOI:10.1016/j.syapm.2013.12.008URLPMID:24582507 [本文引用: 1]
The genetic structure of rhizobia nodulating pea and lentil in Algeria, Northern Africa was determined. A total of 237 isolates were obtained from root nodules collected on lentil (Lens culinaris), proteaginous and forage pea (Pisum sativum) growing in two eco-climatic zones, sub-humid and semi-arid, in Eastern Algeria. They were characterised by PCR-restriction fragment length polymorphism (RFLP) of the 16S 23S rRNA intergenic region (IGS), and the nodD-F symbiotic region. The combination of these haplotypes allowed the isolates to be clustered into 26 distinct genotypes, and all isolates were classified as Rhizobium leguminosarum. Symbiotic marker variation (nodD-F) was low but with the predominance of one nod haplotype (g), which had been recovered previously at a high frequency in Europe. Sequence analysis of the IGS further confirmed its high variability in the studied strains. An AMOVA analysis showed highly significant differentiation in the IGS haplotype distribution between populations from both eco-climatic zones. This differentiation was reflected by differences in dominant genotype frequencies. Conversely, no host plant effect was detected. The nodD gene sequence-based phylogeny suggested that symbiotic gene diversity in pea and lentil nodulating rhizobial populations in Algeria was low compared to that reported elsewhere in the world.
DOI:10.1007/s00442-003-1255-yURLPMID:12695904 [本文引用: 1]
We conducted experiments to examine the quantitative relationships between rainfall event size and rainwater uptake and use by four common native plant species of the Colorado Plateau, including two perennial grasses, Hilaria jamesii (C 4 ) and Oryzopsis hymenoides (C 3 ), and two shrubs, Ceratoides lanata (C 3 ), and Gutierrezia sarothrae (C 3 ). Specifically, we tested the hypothesis that grasses use small rainfall events more efficiently than shrubs and lose this advantage when events are large. Rainfall events between 2 and 20 mm were simulated in spring and summer by applying pulses of deuterium-labeled irrigation water. Afterwards, pulse water fractions in stems and the rates of leaf gas exchange were monitored for 9 days. Cumulative pulse water uptake over this interval (estimated by integrating the product of pulse fraction in stem water and daytime transpiration rate over time) was approximately linearly related to the amount of pulse water added to the ground in all four species. Across species, consistently more pulse water was taken up in summer than in spring. Relative to their leaf areas, the two grass species took up more pulse water than the two shrub species, across all event sizes and in both seasons, thus refuting the initial hypothesis. In spring, pulse water uptake did not significantly increase photosynthetic rates and in summer, pulse water uptake had similar, but relatively small effects on the photosynthetic rates of the three C 3 plants, and a larger effect on the C 4 plant H. jamesii. Based on these data, we introduce an alternative hypothesis for the responses of plant functional types to rainfall events of different sizes, building on cost-benefit considerations for active physiological responses to sudden, unpredictable changes in water availability.
DOI:10.1111/1365-2745.12056URL [本文引用: 1]
1. Drought stress selects for a suite of plant traits at root, stem and leaf level. Two strategies are proposed for trees growing in seasonally water-stressed environments: drought tolerance and drought avoidance. These are respectively associated with evergreen phenology, where plants retain their leaves throughout the year, and deciduous phenology, where plants drop their leaves during dry seasons. Evergreen species are thought to have leaf traits supporting lower photosynthesis and transpiration rates, in order to conserve water during dry periods. 2. We evaluated 18 morphological, chemical and physiological leaf traits of 51 abundant savanna tree species that differed in leaf habit (deciduous and evergreen), selected from two climate types (semi-arid and humid) in three continents (Australia, Africa and South America) (annual rainfall range: 500 1550 mm), and grown in a common garden experiment. We hypothesised that evergreen species have more conservative water use and differ more across climate types than deciduous species because evergreen species are forced to endure extended water deficits during dry seasons. 3. Trait shifts between semi-arid and humid savannas did not differ between evergreen and deciduous species. 4. Evergreen species had similar assimilation rates but lower photosynthetic water-use efficiency (PWUE) than deciduous species, possibly to extend their leaf lifespans by protecting their photosynthetic machinery from overheating through evaporative cooling. 5. Species of humid and semi-arid environments did not differ with respect to assimilation rate or PWUE, but semi-arid species did have smaller leaf sizes and greater leaf potassium and phosphorus concentrations. These traits may enable semi-arid species to maximize growth during episodes of favourable moisture availability. 6. Species from the three continents differed in their leaf traits. These probably reflect the greater proportion of evergreen species in Australia as compared to the other continents and generally infertile soils in the South American sampling sites compared to the wider fertility range in the African sites. 7. Synthesis: Water stress in savannas does not select for more conservative water use, but may select for rapid adjustment to prevailing water conditions and for heat avoidance mechanisms.
DOI:10.1038/nclimate2067URL [本文引用: 1]
Several recently published studies have produced apparently conflicting results of how drought is changing under climate change. The reason is thought to lie in the formulation of the Palmer Drought Severity Index (PDSI) and the data sets used to determine the evapotranspiration component. Here, we make an assessment of the issues with the PDSI in which several other sources of discrepancy emerge, not least how precipitation has changed and is analysed. As well as an improvement in the precipitation data available, accurate attribution of the causes of drought requires accounting for natural variability, especially El Ni o/Southern Oscillation effects, owing to the predilection for wetter land during La Ni a events. Increased heating from global warming may not cause droughts but it is expected that when droughts occur they are likely to set in quicker and be more intense.
DOI:10.1029/2008GL034330URL [本文引用: 1]
Observational evidence presented here indicates that the surface temperatures on the Tibetan Plateau (TP) have increased by about 1.8 C over the past 50 years. The precipitation pattern that is projected as a result of this warming resembles the leading pattern of precipitation variations in East Asia (EA). Numerical experiments with atmospheric general circulation models show that atmospheric heating induced by the rising TP temperatures can enhance East Asian subtropical frontal rainfall. The mechanism of the linkage is found to be through two distinct Rossby wave trains and the isentropic uplift to the east of the TP, which deform the western Pacific Subtropical High and enhance moisture convergence toward the EA subtropical front. The model calculations suggest that the past changes in TP temperatures and EA summer rainfall may be linked, and that projected future increases in TP temperatures may lead to further enhanced summer frontal rainfall in EA region.
DOI:10.11829\j.issn.1001-0629.2014-0190URL [本文引用: 1]
在全球气候变暖和人类活动的共同作用下,黄河源区的生态环境受到严重干扰。目前,气候变化对黄河源生态环境的影响已经得到了很多****的关注。本文在系统总结国内外已有研究成果的基础上,归纳了黄河源区的气候变化特征和已存在的生态环境问题,从径流量、土壤侵蚀、湿地、冻土和植被5个方面综合分析了黄河源区生态环境所面临的主要问题,讨论了气候变化对黄河源区生态环境的影响特点,并在此基础上提出了黄河源生态环境保护的治理对策。结果显示,黄河源区降水略有增加,气温显著升高。黄河源区径流有很大的年际和年代际变化,20世纪60年代中期至80年代后期相对偏丰,90年代初至2008年相对偏枯;源区年径流量距平的年际变化与年降水量距平的年际变化有很好的相关性;降水强度也是影响径流量的原因。黄河源区土壤侵蚀现象严重。湿地面积减少,沼泽和水域的斑块数、破碎度和分维数增加,优势度降低。源区多年冻土已经减薄或消失,多年冻土的边缘地带,垂直方向上形成不衔接冻土和融化夹层,多年冻土分布下界升高50~70 m。源区植被覆盖总体上保持原状,局部出现退化。在黄河源区生态环境保护方面应形成完善的生态补偿机制,加强法制建设,强化社会环保意识,提高执法队伍素质,加强部门联合,实施生态移民,大力发展生态旅游。
DOI:10.11829\j.issn.1001-0629.2014-0190URL [本文引用: 1]
在全球气候变暖和人类活动的共同作用下,黄河源区的生态环境受到严重干扰。目前,气候变化对黄河源生态环境的影响已经得到了很多****的关注。本文在系统总结国内外已有研究成果的基础上,归纳了黄河源区的气候变化特征和已存在的生态环境问题,从径流量、土壤侵蚀、湿地、冻土和植被5个方面综合分析了黄河源区生态环境所面临的主要问题,讨论了气候变化对黄河源区生态环境的影响特点,并在此基础上提出了黄河源生态环境保护的治理对策。结果显示,黄河源区降水略有增加,气温显著升高。黄河源区径流有很大的年际和年代际变化,20世纪60年代中期至80年代后期相对偏丰,90年代初至2008年相对偏枯;源区年径流量距平的年际变化与年降水量距平的年际变化有很好的相关性;降水强度也是影响径流量的原因。黄河源区土壤侵蚀现象严重。湿地面积减少,沼泽和水域的斑块数、破碎度和分维数增加,优势度降低。源区多年冻土已经减薄或消失,多年冻土的边缘地带,垂直方向上形成不衔接冻土和融化夹层,多年冻土分布下界升高50~70 m。源区植被覆盖总体上保持原状,局部出现退化。在黄河源区生态环境保护方面应形成完善的生态补偿机制,加强法制建设,强化社会环保意识,提高执法队伍素质,加强部门联合,实施生态移民,大力发展生态旅游。
DOI:10.1016/S0140-1963(02)00324-5URL [本文引用: 1]
Water is generally considered to be the major limiting factor for perennial shrub growth in the Mojave Desert, USA. However, the responses of active fine roots to soil moisture and microsite differed among Ambrosia dumosa, Ephedra nevadensis, Larrea tridentata, and Lycium pallidum, suggesting differences in root foraging strategies. Ambrosia and Ephedra had a positive linear relationship between active fine root lengths and soil moisture and more roots under the canopy, whereas Larrea had a negative linear relationship and more roots in the interspace. Lycium did not show a significant root/water relationship or significant differences between canopy and interspace microsites.
DOI:10.5846/stxb201307031831URL [本文引用: 1]
植物根系是陆地生态系统重要的碳汇和养分库,细根周转过程是陆地生态系统地下部分碳氮循环的核心环节,在 陆地生态系统如何响应全球变化中起着关键作用.在全球变化敏感地区之一的青藏高原,对该地区的主要植被类型矮嵩草草甸同时采用根钻法、内生长袋法和微根管 法3种观测方法研究细根生产和周转速率,并探讨了极差法、积分法、矩阵法和Kaplan-Meier法等数据处理方法对计算值的影响.研究结果显示:在估 算细根净初级生产力时,根钻法宜采用积分法,内生长袋法宜选用矩阵法;由此进一步以最大细根生物量为基础,根钻法和内生长袋法估测的细根年周转速率分别为 0.36 a-1和0.52 a-1,内生长袋法的估算结果是根钻法的1.44倍.对于微根管法,将其观测得到的细根长度转换为单位面积的生物量值后,采用积分法计算出细根周转速率为 0.84 a-1,远高于传统方法的估算结果;若采用Kaplan-Meier生存分析方法,则计算出的细根周转速率更高达3.41 a-1.
DOI:10.5846/stxb201307031831URL [本文引用: 1]
植物根系是陆地生态系统重要的碳汇和养分库,细根周转过程是陆地生态系统地下部分碳氮循环的核心环节,在 陆地生态系统如何响应全球变化中起着关键作用.在全球变化敏感地区之一的青藏高原,对该地区的主要植被类型矮嵩草草甸同时采用根钻法、内生长袋法和微根管 法3种观测方法研究细根生产和周转速率,并探讨了极差法、积分法、矩阵法和Kaplan-Meier法等数据处理方法对计算值的影响.研究结果显示:在估 算细根净初级生产力时,根钻法宜采用积分法,内生长袋法宜选用矩阵法;由此进一步以最大细根生物量为基础,根钻法和内生长袋法估测的细根年周转速率分别为 0.36 a-1和0.52 a-1,内生长袋法的估算结果是根钻法的1.44倍.对于微根管法,将其观测得到的细根长度转换为单位面积的生物量值后,采用积分法计算出细根周转速率为 0.84 a-1,远高于传统方法的估算结果;若采用Kaplan-Meier生存分析方法,则计算出的细根周转速率更高达3.41 a-1.
DOI:10.1007/s10021-010-9341-6URL [本文引用: 2]
Arid ecosystems receive precipitation pulses of different sizes that may differentially affect nitrogen (N) losses and N turnover during the growing season. We designed a rainfall manipulation experiment in the Patagonian steppe, southern Argentina, where we simulated different precipitation patterns by adding the same amount of water in evenly spaced three-small rainfall events or in one-single large rainfall event, three times during a growing season. We measured the effect of the size of rainfall pulses on N mineralization and N losses by denitrification, ammonia volatilization, and nitrate and ammonia leaching. Irrigation pulses stimulated N mineralization ( P 02<020.05), with small and frequent pulses showing higher responses than large pulses ( P 02<020.10). Irrigation effects were transient and did not result in changes in seasonal net N mineralization suggesting a long-term substrate limitation. Water pulses stimulated gaseous N losses by denitrification, with large pulses showing higher responses than small pulses ( P 02<020.05), but did not stimulate ammonia volatilization. Nitrate leaching also was higher after large than after small precipitation events ( P 02<020.05). Small events produced higher N transformations and lower N losses by denitrification and nitrate leaching than large events, which would produce higher N availability for plant growth. Climate change is expected to increase the frequency of extreme precipitation events and the proportion of large to small rainfall events. Our results suggest that these changes would result in reduced N availability and a competitive advantage for deep-rooted species that prefer nitrate over ammonia. Similarly, the ammonium:nitrate ratio might decrease because large events foster nitrate losses but not ammonium losses.
DOI:10.3773/j.issn.1005-264x.2010.01.004URL [本文引用: 1]
叶片氮(N)和磷(P)的化学计量学研究涉及到植物生态学的众多领域与多个尺度,然而各个尺度上的化学计量学研究并未同步展开.通过对青藏高原47个草地样地连续3年的调查,分析了当地群落水平上的植物叶片N、P含量及其化学计量学特征,并结合温度和降水气候数据研究了N、P含量及N:P比值与这两个气候因子的相关关系.研究结果显示:青藏高原草地群落水平的叶片N含量变化范围为14.8-136.7mg·g~(-1),平均为23.2 mg·g~(-1);P含量变化范围为0.8-2.8mg·g~(-1),平均为1.7 mg·g~(-1);N:P比值变化范围为6.8-25.6,平均为13.5.群落叶片N含量与P含量旱显著正相关关系,叶片的N:P比值与P含量呈显著负相关关系,N:P比值的变化主要由P含量变化决定.另外发现:群落水平叶片N、P含量及N:P比值存在着显著的年际变化,叶片的N、P含量及N:P比值与年平均气温之间存在着极显著的相关关系.通过该研究结果推测:P含量较高的变异系数及其与环境因子表现出的显著相关性,在一定程度上体现了植物群落对当地气候条件的一种适应.
DOI:10.3773/j.issn.1005-264x.2010.01.004URL [本文引用: 1]
叶片氮(N)和磷(P)的化学计量学研究涉及到植物生态学的众多领域与多个尺度,然而各个尺度上的化学计量学研究并未同步展开.通过对青藏高原47个草地样地连续3年的调查,分析了当地群落水平上的植物叶片N、P含量及其化学计量学特征,并结合温度和降水气候数据研究了N、P含量及N:P比值与这两个气候因子的相关关系.研究结果显示:青藏高原草地群落水平的叶片N含量变化范围为14.8-136.7mg·g~(-1),平均为23.2 mg·g~(-1);P含量变化范围为0.8-2.8mg·g~(-1),平均为1.7 mg·g~(-1);N:P比值变化范围为6.8-25.6,平均为13.5.群落叶片N含量与P含量旱显著正相关关系,叶片的N:P比值与P含量呈显著负相关关系,N:P比值的变化主要由P含量变化决定.另外发现:群落水平叶片N、P含量及N:P比值存在着显著的年际变化,叶片的N、P含量及N:P比值与年平均气温之间存在着极显著的相关关系.通过该研究结果推测:P含量较高的变异系数及其与环境因子表现出的显著相关性,在一定程度上体现了植物群落对当地气候条件的一种适应.
DOI:10.17521/cjpe.2006.0001URL [本文引用: 1]
为揭示气候变化对青藏高原草地生态系统的影响及其生态适应机制,利用1982~1999年间的NOAA/AVHRR NDVI数据和对应的气候资料,研究了近20年来青藏高原草地植被覆盖变化及其与气候因子的关系。结果表明,18年来研究区生长季NDVI显著增加(P=0.015),其增加率和增加量分别为0.41%a^-1和0.0010a^-1。生长季提前和生长季生长加速是青藏高原草地植被生长季NDVI增加的主要原因。春季为NDVI增加率和增加量最大的季节,其增加率和增加量分别为0.92%a^-1和0.0014a^-1;夏季NDVI的增加对生长季NDVI增加的贡献相对较小,其增加率和增加量分别为0.37%a^-1和0.0010a^-1。3种草地(高寒草甸、高寒草原、温性草原)春季NDVI均显著增加(P〈0.01;P=0.001;P=0.002);高寒草甸夏季NDVI显著增加(P=0.027),而高寒草原和温性草原夏季NDVI呈增加趋势,但都不显著(P=0.106;P=0.087);3种草地秋季NDVI则没有明显的变化趋势(P=0.585:P=0.461;P=0.143)。3种草地春季NDVI的增加是由春季温度上升所致。高寒草地(高寒草甸和高寒草原)夏季NDVI的增加是夏季温度和春季降水共同作用的结果。温性草原夏季NDVI变化与气候因子并没有表现出显著的相关关系。高寒草地植被生长对气候变化的响应存在滞后效应。
DOI:10.17521/cjpe.2006.0001URL [本文引用: 1]
为揭示气候变化对青藏高原草地生态系统的影响及其生态适应机制,利用1982~1999年间的NOAA/AVHRR NDVI数据和对应的气候资料,研究了近20年来青藏高原草地植被覆盖变化及其与气候因子的关系。结果表明,18年来研究区生长季NDVI显著增加(P=0.015),其增加率和增加量分别为0.41%a^-1和0.0010a^-1。生长季提前和生长季生长加速是青藏高原草地植被生长季NDVI增加的主要原因。春季为NDVI增加率和增加量最大的季节,其增加率和增加量分别为0.92%a^-1和0.0014a^-1;夏季NDVI的增加对生长季NDVI增加的贡献相对较小,其增加率和增加量分别为0.37%a^-1和0.0010a^-1。3种草地(高寒草甸、高寒草原、温性草原)春季NDVI均显著增加(P〈0.01;P=0.001;P=0.002);高寒草甸夏季NDVI显著增加(P=0.027),而高寒草原和温性草原夏季NDVI呈增加趋势,但都不显著(P=0.106;P=0.087);3种草地秋季NDVI则没有明显的变化趋势(P=0.585:P=0.461;P=0.143)。3种草地春季NDVI的增加是由春季温度上升所致。高寒草地(高寒草甸和高寒草原)夏季NDVI的增加是夏季温度和春季降水共同作用的结果。温性草原夏季NDVI变化与气候因子并没有表现出显著的相关关系。高寒草地植被生长对气候变化的响应存在滞后效应。
[本文引用: 1]
[本文引用: 1]
DOI:10.1007/s00382-014-2310-6URL [本文引用: 1]
Precipitation is a critical component of the water balance, and hence its variability is critical for cryospheric and climate change in the Tibetan Plateau (TP). Mean annual and seasonal precipitation totals are compared between gridded observations interpolated to a high resolution (0.5 0.5) and multiple reanalysis type-datasets during 1979-2001. The latter include two NCEP reanalyses (NCEP1 and NCEP2), two European Centre for Medium-Range Weather Forecasts (ECMWF) reanalyses (ERA-40 and ERA-Interim), three modern reanalyses [the twentieth century reanalysis (20century), MERRA and CFSR] and three merged analysis datasets (CMAP1, CMAP2 and GPCP). Observations show an increase in mean precipitation from the northwestern to the southeastern (SE) regions of the TP which are divided by an isohyet of 400 mm, and overall trends during the studied period are positive. Compared with observations, most of the datasets (NCEP1, NCEP2, CMAP1, CMAP2, ERA-Interim, ERA-40, GPCP, 20century, MERRA and CFSR) can both broadly capture the spatial distributions and identify temporal patterns and variabilities of mean precipitation. However, most multi-datasets overestimate precipitation especially in the SE where summer convection is dominant. There remain substantial disagreements and large discrepancies in precipitation trends due to differences in assimilation systems between datasets. Taylor diagrams are used to show the correlation coefficients, standard deviation, and root-mean-square difference of precipitation totals between interpolated observations and assimilated values on an annual and seasonal basis. Merged analysis data (CMAP1 and CMAP2) agree with observations more closely than reanalyses. Thus not all datasets are equally biased and choice of dataset is important.
[本文引用: 1]
[本文引用: 1]
DOI:10.17521/cjpe.2003.0035URL [本文引用: 1]
我国木蓼属有11种,3变种,其中特有种3个。本文通过对它们的形态,花粉等性状的研究,探 讨了属的系统位置,提出了木蓼族的演化系统。在分析了木蓼属的性状后,认为木蓼组Sect Tragopyrum(Bieb.)Meisn.较为原始,应置于刺木蓼组Sect.Atraphaxis之前。本文还研究了该属的地理分布,初步探讨了它的起源及现代分布格局的形成。
DOI:10.17521/cjpe.2003.0035URL [本文引用: 1]
我国木蓼属有11种,3变种,其中特有种3个。本文通过对它们的形态,花粉等性状的研究,探 讨了属的系统位置,提出了木蓼族的演化系统。在分析了木蓼属的性状后,认为木蓼组Sect Tragopyrum(Bieb.)Meisn.较为原始,应置于刺木蓼组Sect.Atraphaxis之前。本文还研究了该属的地理分布,初步探讨了它的起源及现代分布格局的形成。
DOI:10.11829\j.issn.1001-0629.2013-0342URL [本文引用: 1]
以多浆旱生植物霸王(Zygophyllum xanthoxylum)为材料,采用砂培试验,研究了适量的氮磷钠配方对霸王生长和抗旱性的影响。结果表明,15mmol·L-1 NaNO3和0.5mmol·L-1 NaH2PO4为霸王生长最适浓度。根据最适的氮、磷、钠3种元素的浓度配制成氮磷钠配方(15mmol·L-1 NaNO3+0.5mmol·L-1 NaH2PO4+34.5mmol·L-1 NaCl)。在-1MPa处理下,与对照相比,氮磷钠配方处理的霸王植株的叶干质量、组织含水量、叶面积、叶相对有机干质量和株高分别增加了12%、121%、57%、10%和14%,同时,叶片丙二醛含量和相对质膜透性分别下降了64%和17%。因此,在渗透胁迫下,氮磷钠配方对霸王的生长起到了促进作用,减轻了细胞膜的受损程度,增强了霸王的抗旱能力。
DOI:10.11829\j.issn.1001-0629.2013-0342URL [本文引用: 1]
以多浆旱生植物霸王(Zygophyllum xanthoxylum)为材料,采用砂培试验,研究了适量的氮磷钠配方对霸王生长和抗旱性的影响。结果表明,15mmol·L-1 NaNO3和0.5mmol·L-1 NaH2PO4为霸王生长最适浓度。根据最适的氮、磷、钠3种元素的浓度配制成氮磷钠配方(15mmol·L-1 NaNO3+0.5mmol·L-1 NaH2PO4+34.5mmol·L-1 NaCl)。在-1MPa处理下,与对照相比,氮磷钠配方处理的霸王植株的叶干质量、组织含水量、叶面积、叶相对有机干质量和株高分别增加了12%、121%、57%、10%和14%,同时,叶片丙二醛含量和相对质膜透性分别下降了64%和17%。因此,在渗透胁迫下,氮磷钠配方对霸王的生长起到了促进作用,减轻了细胞膜的受损程度,增强了霸王的抗旱能力。
Plant species diversity along a Mediterranean-desert gradient and its correlation with interannual rainfall fluctuations
1
1992
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
Effects of experimental rainfall manipulations on Chihuahuan Desert grassland and shrubland plant communities
1
2013
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
Decadal soil carbon accumulation across Tibetan permafrost regions
1
2017
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
Seasonal variations of photosynthesis, gas exchange, quantum efficiency of photosystem II and biochemical responses ofJatropha curcas L. grown in semi-humid and semi-arid areas subject to water stress.
1
2013
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
Comparable hydrogen isotopic fractionation of plant leaf wax n-alkanoic acids in arid and humid subtropical ecosystems.
1
2014
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
川西北地区沙化草地特征研究
2
2016
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
... id="C5">川西北和青海湖东分属于青藏高原东南缘和东北缘, 是青藏高原典型的寒冷草地; 川西北属于长江和黄河流域重要的水源地, 属于半湿润气候, 青海湖东是青海湖流域重要的水源涵养地, 属于半干旱气候, 川西北和青海湖东对于水源涵养和生物多样性保护具有非常重要的意义(
川西北地区沙化草地特征研究
2
2016
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
... id="C5">川西北和青海湖东分属于青藏高原东南缘和东北缘, 是青藏高原典型的寒冷草地; 川西北属于长江和黄河流域重要的水源地, 属于半湿润气候, 青海湖东是青海湖流域重要的水源涵养地, 属于半干旱气候, 川西北和青海湖东对于水源涵养和生物多样性保护具有非常重要的意义(
半干旱区人工林草地土壤旱化与土壤水分植被承载力
1
2003
... id="C32">青藏高原高寒沙地在长期半干旱和半湿润气候条件下, 中国沙棘和沙蒿的养分含量在不同气候条件下差异明显.在半湿润条件下, 沙蒿整株和中国沙棘根系碳的积累增加, 但中国沙棘枝叶碳的含量降低; 而两种植物全株氮含量在半湿润条件下明显降低; 两种植物全株的磷含量也在半湿润条件下降低.部分研究表明气候变暖导致青藏高原降水增加(
半干旱区人工林草地土壤旱化与土壤水分植被承载力
1
2003
... id="C32">青藏高原高寒沙地在长期半干旱和半湿润气候条件下, 中国沙棘和沙蒿的养分含量在不同气候条件下差异明显.在半湿润条件下, 沙蒿整株和中国沙棘根系碳的积累增加, 但中国沙棘枝叶碳的含量降低; 而两种植物全株氮含量在半湿润条件下明显降低; 两种植物全株的磷含量也在半湿润条件下降低.部分研究表明气候变暖导致青藏高原降水增加(
青藏高原东北部5000年来气候变化与若尔盖湿地历史生态学研究进展
1
2014
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
青藏高原东北部5000年来气候变化与若尔盖湿地历史生态学研究进展
1
2014
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
Simulation of nitrate leaching under irrigated maize on sandy soil in desert oasis in Inner Mongolia, China
1
2008
... id="C34">不同气候条件下植物和土壤养分比差异显著.在半湿润气候条件下, 相对丰沛的降水使植物快速生长, 植物体碳含量增加, 碳氮比和碳磷比变高(
干湿变化和保水剂对植物生长和水分利用效率的影响
1
2002
... id="C32">青藏高原高寒沙地在长期半干旱和半湿润气候条件下, 中国沙棘和沙蒿的养分含量在不同气候条件下差异明显.在半湿润条件下, 沙蒿整株和中国沙棘根系碳的积累增加, 但中国沙棘枝叶碳的含量降低; 而两种植物全株氮含量在半湿润条件下明显降低; 两种植物全株的磷含量也在半湿润条件下降低.部分研究表明气候变暖导致青藏高原降水增加(
干湿变化和保水剂对植物生长和水分利用效率的影响
1
2002
... id="C32">青藏高原高寒沙地在长期半干旱和半湿润气候条件下, 中国沙棘和沙蒿的养分含量在不同气候条件下差异明显.在半湿润条件下, 沙蒿整株和中国沙棘根系碳的积累增加, 但中国沙棘枝叶碳的含量降低; 而两种植物全株氮含量在半湿润条件下明显降低; 两种植物全株的磷含量也在半湿润条件下降低.部分研究表明气候变暖导致青藏高原降水增加(
The timing and degree of root proliferation in fertile-soil microsites for three cold-desert perennials
1
1989
... id="C33">各植物根际土壤中养分含量在半干旱和半湿润气候条件下差异显著.半湿润气候条件下沙蒿根系土壤中的碳、氮、磷含量较高, 而中国沙棘根际土壤碳、氮和有效磷的含量却较低.沙蒿为丛生型密集半灌木, 在地上成片分布, 郁闭度极高, 群落中极难分布其他物种, 当年生长迅速, 并且冬季地上部分枯萎, 可以依靠根茎进行无性繁殖.在半湿润条件下, 降水相对充沛, 地下根系快速生长, 地上和地下生物量较大(
The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate
1
2015
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
Decomposition of leaf and root litter of Chihuahuan desert shrubs: Effects of three years of summer drought
1
2003
... id="C34">不同气候条件下植物和土壤养分比差异显著.在半湿润气候条件下, 相对丰沛的降水使植物快速生长, 植物体碳含量增加, 碳氮比和碳磷比变高(
Climate variability rather than overstocking causes recent large scale cover changes of Tibetan pastures
1
2016
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
青海湖流域草地植被动态变化趋势下的物候时空特征
1
2014
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
青海湖流域草地植被动态变化趋势下的物候时空特征
1
2014
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
青藏高原气候区划
2
1981
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
... id="C6">研究区域位于四川省阿坝州红原县日干乔生态保护区(31.51°-33.19° N, 101.51°-103.23° E, 海拔3 460 m)和青海省海北州海晏县湖东生态保护区(36.36°-37.05° N, 100.01°-100.86° E, 海拔3 150 m).日干乔生态保护区属于青藏高原东南缘,为半湿润气候, 年降水量相对充沛; 青海湖湖东生态保护区属于青藏高原东北缘,为半干旱气候, 年降水量相对较少(
青藏高原气候区划
2
1981
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
... id="C6">研究区域位于四川省阿坝州红原县日干乔生态保护区(31.51°-33.19° N, 101.51°-103.23° E, 海拔3 460 m)和青海省海北州海晏县湖东生态保护区(36.36°-37.05° N, 100.01°-100.86° E, 海拔3 150 m).日干乔生态保护区属于青藏高原东南缘,为半湿润气候, 年降水量相对充沛; 青海湖湖东生态保护区属于青藏高原东北缘,为半干旱气候, 年降水量相对较少(
2
2000
... id="C9">植物样品测定: 植物样品C利用重铬酸钾和浓硫酸氧化, 硫酸亚铁滴定方法进行测定; N利用浓硫酸混合加速剂蒸馏方法进行测定; P利用钼锑抗比色法(
... id="C10">土壤样品测定: 土壤pH值利用土壤pH计测定; 土壤水分在105 ℃下烘干至恒质量测定; SOC利用高温外热重铬酸钾氧化-容量法进行测定; STN利用凯氏法进行测定; STP利用酸溶-钼锑抗比色方法进行测定; SNN利用双波长比色法进行测定; SAN利用靛酚蓝比色方法进行测定; SAP利用钼锑抗比色法进行测定(
2000-2010年青海湖流域草地退化状况时空分析
1
2013
... id="C5">川西北和青海湖东分属于青藏高原东南缘和东北缘, 是青藏高原典型的寒冷草地; 川西北属于长江和黄河流域重要的水源地, 属于半湿润气候, 青海湖东是青海湖流域重要的水源涵养地, 属于半干旱气候, 川西北和青海湖东对于水源涵养和生物多样性保护具有非常重要的意义(
2000-2010年青海湖流域草地退化状况时空分析
1
2013
... id="C5">川西北和青海湖东分属于青藏高原东南缘和东北缘, 是青藏高原典型的寒冷草地; 川西北属于长江和黄河流域重要的水源地, 属于半湿润气候, 青海湖东是青海湖流域重要的水源涵养地, 属于半干旱气候, 川西北和青海湖东对于水源涵养和生物多样性保护具有非常重要的意义(
沙蒿与油蒿灌丛的防风阻沙作用
1
2012
... id="C33">各植物根际土壤中养分含量在半干旱和半湿润气候条件下差异显著.半湿润气候条件下沙蒿根系土壤中的碳、氮、磷含量较高, 而中国沙棘根际土壤碳、氮和有效磷的含量却较低.沙蒿为丛生型密集半灌木, 在地上成片分布, 郁闭度极高, 群落中极难分布其他物种, 当年生长迅速, 并且冬季地上部分枯萎, 可以依靠根茎进行无性繁殖.在半湿润条件下, 降水相对充沛, 地下根系快速生长, 地上和地下生物量较大(
沙蒿与油蒿灌丛的防风阻沙作用
1
2012
... id="C33">各植物根际土壤中养分含量在半干旱和半湿润气候条件下差异显著.半湿润气候条件下沙蒿根系土壤中的碳、氮、磷含量较高, 而中国沙棘根际土壤碳、氮和有效磷的含量却较低.沙蒿为丛生型密集半灌木, 在地上成片分布, 郁闭度极高, 群落中极难分布其他物种, 当年生长迅速, 并且冬季地上部分枯萎, 可以依靠根茎进行无性繁殖.在半湿润条件下, 降水相对充沛, 地下根系快速生长, 地上和地下生物量较大(
The key role of dry days in changing regional climate and precipitation regimes
1
2014
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
Genotypic and symbiotic diversity of Rhizobium populations associated with cultivated lentil and pea in sub-humid and semi-arid regions of Eastern Algeria
1
2014
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
Dominant cold desert plants do not partition warm season precipitation by event size
1
2003
... id="C32">青藏高原高寒沙地在长期半干旱和半湿润气候条件下, 中国沙棘和沙蒿的养分含量在不同气候条件下差异明显.在半湿润条件下, 沙蒿整株和中国沙棘根系碳的积累增加, 但中国沙棘枝叶碳的含量降低; 而两种植物全株氮含量在半湿润条件下明显降低; 两种植物全株的磷含量也在半湿润条件下降低.部分研究表明气候变暖导致青藏高原降水增加(
Leaf adaptations of evergreen and deciduous trees of semi-arid and humid savannas on three continents
1
2013
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
Global warming and changes in drought
1
2014
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
Tibetan Plateau warming and precipitation changes in East Asia
1
2008
... id="C32">青藏高原高寒沙地在长期半干旱和半湿润气候条件下, 中国沙棘和沙蒿的养分含量在不同气候条件下差异明显.在半湿润条件下, 沙蒿整株和中国沙棘根系碳的积累增加, 但中国沙棘枝叶碳的含量降低; 而两种植物全株氮含量在半湿润条件下明显降低; 两种植物全株的磷含量也在半湿润条件下降低.部分研究表明气候变暖导致青藏高原降水增加(
气候变化对黄河源区生态环境的影响
1
2015
... id="C5">川西北和青海湖东分属于青藏高原东南缘和东北缘, 是青藏高原典型的寒冷草地; 川西北属于长江和黄河流域重要的水源地, 属于半湿润气候, 青海湖东是青海湖流域重要的水源涵养地, 属于半干旱气候, 川西北和青海湖东对于水源涵养和生物多样性保护具有非常重要的意义(
气候变化对黄河源区生态环境的影响
1
2015
... id="C5">川西北和青海湖东分属于青藏高原东南缘和东北缘, 是青藏高原典型的寒冷草地; 川西北属于长江和黄河流域重要的水源地, 属于半湿润气候, 青海湖东是青海湖流域重要的水源涵养地, 属于半干旱气候, 川西北和青海湖东对于水源涵养和生物多样性保护具有非常重要的意义(
Fine root growth dynamics of four Mojave Desert shrubs as related to soil moisture and microsite
1
2004
... id="C33">各植物根际土壤中养分含量在半干旱和半湿润气候条件下差异显著.半湿润气候条件下沙蒿根系土壤中的碳、氮、磷含量较高, 而中国沙棘根际土壤碳、氮和有效磷的含量却较低.沙蒿为丛生型密集半灌木, 在地上成片分布, 郁闭度极高, 群落中极难分布其他物种, 当年生长迅速, 并且冬季地上部分枯萎, 可以依靠根茎进行无性繁殖.在半湿润条件下, 降水相对充沛, 地下根系快速生长, 地上和地下生物量较大(
高寒草甸植被细根生产和周转的比较研究
1
2014
... id="C33">各植物根际土壤中养分含量在半干旱和半湿润气候条件下差异显著.半湿润气候条件下沙蒿根系土壤中的碳、氮、磷含量较高, 而中国沙棘根际土壤碳、氮和有效磷的含量却较低.沙蒿为丛生型密集半灌木, 在地上成片分布, 郁闭度极高, 群落中极难分布其他物种, 当年生长迅速, 并且冬季地上部分枯萎, 可以依靠根茎进行无性繁殖.在半湿润条件下, 降水相对充沛, 地下根系快速生长, 地上和地下生物量较大(
高寒草甸植被细根生产和周转的比较研究
1
2014
... id="C33">各植物根际土壤中养分含量在半干旱和半湿润气候条件下差异显著.半湿润气候条件下沙蒿根系土壤中的碳、氮、磷含量较高, 而中国沙棘根际土壤碳、氮和有效磷的含量却较低.沙蒿为丛生型密集半灌木, 在地上成片分布, 郁闭度极高, 群落中极难分布其他物种, 当年生长迅速, 并且冬季地上部分枯萎, 可以依靠根茎进行无性繁殖.在半湿润条件下, 降水相对充沛, 地下根系快速生长, 地上和地下生物量较大(
Size of precipitation pulses controls nitrogen transformation and losses in an arid Patagonian ecosystem
2
2010
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
... id="C35">在不同的气候条件下, 沙蒿的枝叶、根和根际土壤碳、氮、磷显著相关, 沙蒿枝叶及根的氮磷比和土壤氮磷比显著负相关, 中国沙棘枝叶及根的氮磷比和土壤的氮磷比没有显著相关性.半干旱和半湿润气候差异极可能通过对高寒沙地植物生理以及土壤养分淋溶产生影响(
青藏高原草地植物群落冠层叶片氮磷化学计量学分析
1
2010
... id="C32">青藏高原高寒沙地在长期半干旱和半湿润气候条件下, 中国沙棘和沙蒿的养分含量在不同气候条件下差异明显.在半湿润条件下, 沙蒿整株和中国沙棘根系碳的积累增加, 但中国沙棘枝叶碳的含量降低; 而两种植物全株氮含量在半湿润条件下明显降低; 两种植物全株的磷含量也在半湿润条件下降低.部分研究表明气候变暖导致青藏高原降水增加(
青藏高原草地植物群落冠层叶片氮磷化学计量学分析
1
2010
... id="C32">青藏高原高寒沙地在长期半干旱和半湿润气候条件下, 中国沙棘和沙蒿的养分含量在不同气候条件下差异明显.在半湿润条件下, 沙蒿整株和中国沙棘根系碳的积累增加, 但中国沙棘枝叶碳的含量降低; 而两种植物全株氮含量在半湿润条件下明显降低; 两种植物全株的磷含量也在半湿润条件下降低.部分研究表明气候变暖导致青藏高原降水增加(
青藏高原草地植被覆盖变化及其与气候因子的关系
1
2006
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
青藏高原草地植被覆盖变化及其与气候因子的关系
1
2006
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
高寒矮生嵩草草甸主要植物物候特征对养分和水分添加的响应
1
2014
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
高寒矮生嵩草草甸主要植物物候特征对养分和水分添加的响应
1
2014
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
Comparison of multiple datasets with gridded precipitation observations over the Tibetan Plateau
1
2015
... id="C32">青藏高原高寒沙地在长期半干旱和半湿润气候条件下, 中国沙棘和沙蒿的养分含量在不同气候条件下差异明显.在半湿润条件下, 沙蒿整株和中国沙棘根系碳的积累增加, 但中国沙棘枝叶碳的含量降低; 而两种植物全株氮含量在半湿润条件下明显降低; 两种植物全株的磷含量也在半湿润条件下降低.部分研究表明气候变暖导致青藏高原降水增加(
青藏高原生态变化
1
2015
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
青藏高原生态变化
1
2015
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
沙木蓼和沙枣对地下水位变化的生理生态响应I. 叶片养分, 叶绿素, 可溶性糖和淀粉的变化
1
2003
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
沙木蓼和沙枣对地下水位变化的生理生态响应I. 叶片养分, 叶绿素, 可溶性糖和淀粉的变化
1
2003
... id="C4">干旱和湿润的气候环境形成了不同的陆地生态景观, 并极大地影响着植物的生长发育过程(
渗透胁迫下氮磷钠配方对多浆旱生植物霸王生长的影响
1
2014
... id="C32">青藏高原高寒沙地在长期半干旱和半湿润气候条件下, 中国沙棘和沙蒿的养分含量在不同气候条件下差异明显.在半湿润条件下, 沙蒿整株和中国沙棘根系碳的积累增加, 但中国沙棘枝叶碳的含量降低; 而两种植物全株氮含量在半湿润条件下明显降低; 两种植物全株的磷含量也在半湿润条件下降低.部分研究表明气候变暖导致青藏高原降水增加(
渗透胁迫下氮磷钠配方对多浆旱生植物霸王生长的影响
1
2014
... id="C32">青藏高原高寒沙地在长期半干旱和半湿润气候条件下, 中国沙棘和沙蒿的养分含量在不同气候条件下差异明显.在半湿润条件下, 沙蒿整株和中国沙棘根系碳的积累增加, 但中国沙棘枝叶碳的含量降低; 而两种植物全株氮含量在半湿润条件下明显降低; 两种植物全株的磷含量也在半湿润条件下降低.部分研究表明气候变暖导致青藏高原降水增加(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}