
Allocation of mass and stability of soil aggregate in different types of Nei Mongol grasslands
WANGTian通讯作者:
收稿日期:2017-08-16
接受日期:2017-11-14
网络出版日期:2017-11-10
版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。
基金资助:
展开
摘要
关键词:
Abstract
Methods We have established 78 sites with a size of 10 m × 10 m across the transect of Nei Mongol grasslands and collected soil samples from different soil depth up to 1 m. We used wet sieving method to separate different sizes of aggregate partition and used mean mass diameter (MMD) and geometric mean diameter (GMD) in order to evaluate the stability of soil aggregate. The two-way ANOVA was used to test the difference of mass percentage and stability of soil aggregate in different grassland types and soil depths. In addition, a linear regression analysis was used to analyze the correlations of mass percentage and stability of soil aggregate with both mean annual precipitation (MAP) and mean annual temperature (MAT).
Important findings The results showed that the mass percentages of soil aggregate were highest in meadow steppe, while almost equal in typical steppe and desert steppe. However, no significant patterns were found along the soil depth. The mass percentage of soil aggregate fractions were positively correlated with MAP in all soil layers, but negatively correlated with MAT except the layer of 70-100 cm. For the stability of soil aggregate, at 0-10 and 10-20 cm, MMD and GMD of meadow steppe were significantly greater than those of typical and desert steppes, whereas no significant differences among three grassland types were found for other soil layers. Besides, MMD and GMD in meadow steppe and typical steppe gradually decreased along the soil depth.
Keywords:
-->0
PDF (1079KB)元数据多维度评价相关文章收藏文章
本文引用格式导出EndNoteRisBibtex收藏本文-->
土壤团聚体是土壤的重要组成部分, 对维持土壤结构的稳定和土壤养分截留具有重要意义(Six et al., 1998; Cavagnaro et al., 2006)。此外, 土壤团聚体对土壤有机碳具有物理保护作用, 是土壤肥力的基础和评价土壤质量的重要指标(Lal, 2000)。表土中约90%的有机质位于团聚体内(Jastrow, 1996), 团聚体将土壤有机碳与微生物隔离开, 从而保护有机碳难被微生物分解以达到保护作用(Six et al., 2002a), 大团聚体中包裹的小团聚体是土壤有机碳物理保护的必要机制(Denef et al., 2007)。土壤团聚体是通过结合体(如细根, 微生物菌丝, 糖分及黏粒、粉粒等)将土壤颗粒粘结在一起而形成, 因此影响结合体形式和量的因素均可能影响土壤团聚体的形成和分布(Six et al., 2002b)。
草原是受人类活动影响最为严重的区域, 因此草原的碳储蓄能力及其动态变化在全球碳循环中扮演着重要角色(Scurlock & Hall, 1998)。我国内蒙古草原是欧亚草原的一个重要组成部分(Bai et al., 2008), 其经纬度覆盖范围广(98.0°-124.5° E, 37.7°-51.3° N), 导致气温、降雨差异大, 从东到西, 草地类型多样, 分布有温性草甸草原、典型草原和荒漠草原(马文红, 2006)。不同类型草原的气候因子, 包括气温和降水, 地上、地下生物量(Ma et al., 2008; Yang et al., 2010a)、微生物生物量及组成(Li & Chen, 2004)均存在显著差异, 这些差异均可能对土壤团聚体的形成产生影响, 从而导致在不同类型的草原形成不同的团聚体分布格局。另外, 对于不同的土层深度, 其湿度、微生物生物量及组成(Allison et al., 2007)、根系生物量(Ma et al., 2008)以及可溶性有机碳的淋溶(Sanderman & Amundson, 2008)等的不同也可能使团聚体分布存在差异。例如, 在内蒙古半干旱草原耕地向草原过渡的过程中, 由于保护后的耕地草原与天然草原在地上地下生物量以及土壤质地等方面有明显差异, 导致了两者在团聚体组分分配以及稳定性上的差异, 并且这种差异在不同的深度表现不同(Zhang et al., 2013)。
由于团聚体的形成对土壤有机碳具有物理保护作用, 土壤团聚体的稳定性在一定程度上决定了土壤有机碳的稳定性(Bird et al., 2002)。景观尺度的土壤团聚体稳定性由土壤质地和矿物的变异、土壤有机质的输入以及干扰共同决定(Lehrsch et al., 1991; Chenu et al., 2000; Abiven et al., 2009)。一项关于草原生态系统土壤团聚体稳定性多尺度变异的研究发现, 植被覆盖度越高, 土壤团聚体稳定性越高(Bird et al., 2007)。国内对团聚体稳定性的研究主要集中在不同的土地利用方式、耕作以及施肥等情况下对团聚体稳定性的影响(刘恩科等, 2010; 田慎重等, 2013; 李鉴霖等, 2015)。然而, 目前对我国内蒙古草原土壤团聚体的含量以及稳定性在不同类型以及不同土层深度的分布规律还不清楚。
考虑到土壤团聚体对于土壤结构和肥力的重要意义, 以及土壤团聚体稳定性对土壤有机碳的保护作用, 本研究利用呼伦贝尔-锡林浩特-鄂尔多斯样带探讨我国内蒙古3种草原类型不同土层深度的土壤团聚体分布特征及团聚体稳定性分布特征, 具体科学问题如下: (1)土壤团聚体含量的分布是否因草原类型和土层深度而异? (2)草原生态系统土壤团聚体含量的分布与气候因子有何关系? (3)内蒙古3种草原类型的土壤团聚体稳定性呈现什么样的分布格局?
1 材料和方法
1.1 研究区域概况
本研究区域主要集中在蒙古高原上内蒙古境内的天然草原。研究样地位于107.84°-120.06° E, 39.15°-49.62° N, 海拔564-1 546 m, 由东北向西南, 内蒙古草原的年平均气温逐渐增加, 年降水量逐渐减少, 干旱程度逐渐加剧(Wang et al., 2014)。东北半湿润区、中部半干旱区和西部干旱气候区分别形成了草甸草原、典型草原和荒漠草原。草甸草原生长季年降水量277-327 mm, 生长季平均气温11.20-13.60 ℃, 植物种类丰富, 群落以多年生旱生、中旱生植物占优势。主要建群和优势物种有贝加尔针茅(Stipa baicalensis)、羊草(Leymus chinensis)等; 典型草原生长季年降水量174-295 mm, 生长季年平均气温12.60-18.20℃, 是由典型的旱生性多年生草本植物组成的群落类型, 是内蒙古草原的主体,与草甸草原相比, 典型草原的物种丰富程度明显降低, 群落盖度减小, 主要建群和优势物种有大针茅(Stipa grandis)、克氏针茅(S. kryovii)等。荒漠草原生长季年降水量110-219 mm, 生长季年平均气温14.70-17.60 ℃, 荒漠草原是由旱生性更强的多年生矮小草本植物组成的草原群落, 群落生物多样性不高, 主要有小针茅(S. klemenzii)、短花针茅(S. breviflora)等(马文红, 2006; Ma et al., 2010)(表1)。土壤类型从东北向西南分别为黑钙土、栗钙土和棕钙土(Bai et al., 2008)。Table 1
表1
表1三种草原的基本信息
Table 1The basic information of the three types of grassland
| 草原类型 Grassland type | GSP平均值(范围)1) Mean of GSP (range) 1) (mm) | GST平均值(范围) 1) Mean of GSTa(range) 1) (℃) | 氮:磷平均值(范围) 2) Mean of N:P (range) 2) | pH平均值(范围) 3) Mean of pH (range) 3) | 优势种4) Dominant species4) |
|---|---|---|---|---|---|
| 草甸草原 Meadow steppe | 306 (277-327) | 12.50 (11.20-13.60) | 3.83 (3.25-4.37) | 7.10 (7.60-8.00) | 贝加尔针茅 Stipa baicalensis, 羊草 Leymus chinensis |
| 典型草原 Typical steppe | 242 (174-295) | 14.00 (12.60-18.20) | 3.02 (2.93-3.11) | 7.50 (7.26-7.73) | 大针茅 Stipa grandis, 克氏针茅 Stipa kryovii |
| 荒漠草原 Desert steppe | 163 (110-219) | 16.50 (14.70-17.60) | 2.48 (2.41-2.54) | 7.80 (6.65-7.56) | 小针茅 Stipa klemenzii, 短花针茅 Stipa breviflora |
新窗口打开
气象数据的来源与处理: 对全国670个标准气象台站1970-2015年间每月平均气温和降水数据进行克里格插值分析, 然后根据样地的经纬度从插值结果中提取样地的每月平均气温和降水数据, 以45年的平均值作为样地的气温和降水数据。
1.2 土壤样品采集
我们于2015年7-8月沿呼伦贝尔-锡林浩特-鄂尔多斯一线调查了3种草原类型共78个样地(图1): 荒漠草原(20个样方)、典型草原(45个样方)和草甸草原(13个样方)。在每处取样点, 设置10 m × 10 m的大样方, 沿大样方四角及中心位置分别设置了5个1 m × 1 m的小样方。之后, 选取大样方其中一条对角线的3个小样方, 并在每个小样方的对角线位置利用直径50 mm的土钻采集了至少3个剖面的土壤样品, 采样深度依次为0-10 cm、10-20 cm、20-30 cm、30-50 cm、50-70 cm和70-100 cm。采集结束后, 将小样方内同一土层的土壤样品混合成一个混合样品, 收集后用于室内测定。同时记录样地经纬度、海拔、坡度和坡向等信息, 每个点的定位信息以及海拔用GPS测量。 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图1采样点分布图。
-->Fig. 1The distribution of sampling sites.
-->
1.3 土壤样品分析
所有土壤样品带回实验室风干后, 过8 mm筛, 以去除根系及石块。我们采用湿筛法进行团聚体分级(Six et al., 1998), 具体步骤如下: 称取50.00 g风干土, 将其放置在团聚体分析仪(XY-100, 北京祥宇伟业仪器设备有限公司, 北京)套筛的最上端筛子内, 调整桶内水面的高度, 使水面刚好漫过风干土,将风干土浸泡5 min后, 启动仪器进行分级, 以振幅3 cm振动30 min后, 根据筛孔径的大小分离出< 53 μm、53-250 μm、250-2 000 μm和>2 000 μm四级团聚体组分, 将团聚体套筛小心地从桶内移出, 放置在实验台上, 然后利用药匙、洗瓶和蒸馏水将各级筛中的团聚体组分先转移至烧杯中, 再将烧杯中的团聚体组分和蒸馏水混合物转入相应的铝盒(直径60 mm, 高60 mm), 放入烘箱中进行烘干, 烘箱温度50 ℃ (不超过65 ℃), 烘干后利用药匙将铝盒中的团聚体组分转移至称量纸上进行称量; 称量后, 倒回铝盒中, 再加入浓度为5 g?L-1的六偏磷酸钠((NaPO3)6), 加入的量漫过团聚体1-2 cm, 并放置在摇床上以中速摇动6 min, 以破坏团聚体, 过53 μm筛, 使团聚体与砂砾分离, 将去除砂砾的团聚体重新转入铝盒, 烘干并称量。经上述过程后, 得到相应各级的团聚体及其质量m, 将<53 μm、53-250 μm、250-2 000 μm和>2 000 μm四级团聚体组分的质量分别记为m1、m2、m3和m4。1.4 数据分析
1.4.1 土壤团聚体组分质量百分比的计算将团聚体分级后的每级团聚体组分在烘箱中烘干后称量, 得到每级团聚体组分的质量, 然后将每级团聚体组分的质量除以用于进行团聚体分级的整土的质量, 在我们的研究中指50 g风干土质量, 得到每级团聚体组分的质量百分比(%), 分别记为mp1 (< 53 μm团聚体组分的质量百分比)、mp2 (53- 250 μm团聚体组分的质量百分比)、mp3 (250-2 000 μm团聚体组分的质量百分比)和mp4 (> 2 000 μm团聚体组分的质量百分比), 具体公式如下:
$mp\text{(}i\text{)}=\sum{\frac{m(i)}{M}}\times 100%$ (1)
式中: mp(i)指每级团聚体组分的质量百分比; m(i)指每级团聚体组分的质量; i是各级团聚体组分的编号(1-4分别指<53 μm、53-250 μm、250-2 000 μm和> 2 000 μm团聚体组分); M指用于进行团聚体分级的整土质量, 在我们的研究中M = 50 g。
1.4.2 土壤团聚体稳定性的计算
到目前为止, 测定土壤团聚体稳定性的方法有很多, 包括了水稳性团聚体质量占比、团聚体形成指数、团聚体破坏指数、标准化稳定性指数、稳定性团聚体和稳定性大团聚体指数、平均质量直径(MMD)和几何平均直径(GMD)等(Marquez et al., 2004)。其中, MMD是表征团聚体稳定性的最重要、最常用的指标(Nimmo & Perkins, 2002), 是土壤团聚体组分分布的综合参数, 从而作为表征整土结构的指数(Amezketa, 1999)。GMD同样也能作为团聚体大小分布的一种指数。MMD与GMD作为反映土壤团聚体状况的指标, 其值越大表示土壤的团聚度越高, 团聚体稳定性越强(Nimmo & Perkins, 2002)。其计算方法如下:
MMD = ΣnXiMi (2)
GMD = exp[ΣnMilnXi] (3)
式中: n为粒径分组的组数; Xi为这一粒径组分的平均直径, i是各级团聚体组分的编号(1-4分别指< 53 μm、53-250 μm、250-2 000 μm和> 2 000 μm团聚体组分) (X1 = 26.5 μm; X2 = 151.5 μm; X3 = 1 125 μm、X4 = 2 000 μm); Mi为这一粒径团聚体的质量分数。
1.4.3 统计分析
利用双因素方差分析的方法分析草原类型和土层深度对各层土壤团聚体质量百分比和团聚体稳定性分布的影响。利用一元线性回归分析土壤团聚体质量百分比及团聚体稳定性与年降水量和年平均气温的关系。利用多重比较分析不同草地类型, 及不同土层深度的土壤团聚体质量百分比间的分布差异以及土壤团聚体稳定性的差异。所有的统计分析均由R(3.3.1)软件(R Core Team (2017))完成。柱状图在SigmaPlot 12.0软件中绘制。本研究中空间制图在地理信息系统软件ArcGIS 10.0中完成。
2 结果和分析
2.1 土壤团聚体质量百分比的分配特征
总体而言, 草甸草原、典型草原及荒漠草原在0-1 m中的总团聚体质量百分数分别为(49.52 ± 2.76)%、(32.32 ± 1.08)%和(25.22 ± 1.17)% (表2); 具体每一层中每一级的团聚体质量百分比分布见附录I。双因素方差分析结果显示: 土层深度对团聚体分配效应不显著(p = 0.12), 且两者的交互效应也不显著(p = 0.41), 但草原类型的效应显著(p < 0.01)(附录II)。对3种草原在不同土层深度上的总团聚体质量百分数进行统计分析, 结果表明: 在每一层土层, 草甸草原都显著大于典型草原与荒漠草原(p < 0.01), 而典型草原与荒漠草原之间无显著差异(p > 0.05) (表2)。另外, 3种类型草原的土壤团聚体含量在各个土层深度都没有明显的规律(表2)。Appendix I
附录I
附录I各个土层各级团聚体的质量百分比(平均值±标准误差)
Appendix IThe mass percentage (%) of each aggregate fraction in different soil layers (mean ± SE)
| 土层 Soil layer (cm) | 草甸草原 Meadow steppe (%) | 典型草原 Typical steppe (%) | 荒漠草原 Desert steppe (%) | |
|---|---|---|---|---|
| 0-10 | A1 | 14.15 ± 1.98a | 13.58 ± 1.20a | 14.8 ± 1.49a |
| A2 | 9.33 ± 0.93a | 8.32 ± 0.44a | 5.90 ± 0.45a | |
| A3 | 15.90 ± 2.96a | 10.33 ± 1.24ab | 6.76 ± 0.71b | |
| A4 | 14.15 ± 4.48a | 5.93 ± 0.99b | - | |
| 10-20 | A1 | 18.82 ± 3.41a | 12.89 ± 1.02ab | 12.67 ± 1.30b |
| A2 | 10.37 ± 1.30a | 8.30 ± 0.59a | 6.84 ± 1.09a | |
| A3 | 15.43 ± 3.08a | 9.30 ± 0.93ab | 7.32 ± 0.90b | |
| 20-30 | A4 | 10.22 ± 2.87a | 4.51 ± 0.96b | - |
| A1 | 22.86 ± 3.23a | 13.70 ± 1.39ab | 10.14 ± 1.16b | |
| A2 | 12.93 ± 1.34a | 7.65 ± 0.52ab | 6.64 ± 0.65b | |
| A3 | 12.39 ± 2.54a | 9.24 ± 0.95a | 7.74 ± 1.35a | |
| A4 | 6.95 ± 1.35a | 3.72 ± 1.03a | - | |
| 30-50 | A1 | 18.97 ± 2.43a | 13.55 ± 1.16ab | 10.30 ± 1.20b |
| A2 | 14.70 ± 2.61a | 8.74 ± 0.82ab | 6.93 ± 1.53b | |
| A3 | 13.39 ± 2.83a | 8.02 ± 0.75ab | 6.75 ± 0.66b | |
| A4 | 4.76 ± 0.79a | 3.70 ± 1.17a | - | |
| 50-70 | A1 | 23.89 ± 8.50a | 17.02 ± 3.26b | 9.43 ± 1.50c |
| A2 | 12.34 ± 3.64a | 7.18 ± 1.09ab | 5.32 ± 0.68b | |
| A3 | 17.35 ± 5.94a | 6.84 ± 1.28b | 6.29 ± 0.83b | |
| A4 | - | - | - | |
| 70-100 | A1 | 29.72 ± 9.86a | 14.71 ± 3.01b | 10.09 ± 1.22b |
| A2 | 10.79 ± 2.97a | 6.87 ± 1.08a | 5.15 ± 1.150a | |
| A3 | 14.68 ± 5.49a | 6.65 ± 1.46b | 7.86 ± 2.23b | |
| A4 | - | - | - |
新窗口打开
Appendix II
附录II
附录II土层深度和草原类型对团聚体质量百分数的双因素方差分析
Appendix IIThe two-way ANOVA results of soil depth and grassland type on the mass percentage of aggregate
| 因素 Factor | 团聚体质量百分数 Mass percentage of aggregate |
|---|---|
| 土层深度 Soil depth | 0.12 |
| 草原类型 Grassland type | < 0.01 |
| 土层深度×草原类型 Soil depth × Grassland type | 0.41 |
新窗口打开
Table 2
表2
表2三种草原各层土壤团聚体的质量百分比(平均值±标准误差)
Table 2The mass percentage (%) of total soil aggregate in different soil layers (mean ± SE)
| 土层 Soil layer (cm) | 草甸草原 Meadow steppe (%) | 典型草原 Typical steppe (%) | 荒漠草原 Desert steppe (%) |
|---|---|---|---|
| 0-10 | 49.66 ± 1.12a | 35.07 ± 2.20b | 27.25 ± 1.87b |
| 10-20 | 48.63 ± 2.08a | 32.30 ± 2.19b | 26.84 ± 2.76b |
| 20-30 | 52.15 ± 5.70a | 31.80 ± 2.32b | 24.52 ± 2.68b |
| 30-50 | 48.20 ± 1.79a | 31.53 ± 2.28b | 24.45 ± 3.26b |
| 50-70 | 49.14 ± 14.21a | 31.21 ± 0.82b | 21.04 ± 2.49b |
| 70-100 | 48.40 ± 15.21a | 28.82 ± 1.5b | 23.10 ± 3.73b |
| 0-100 | 49.52 ± 2.76a | 32.32 ± 1.08b | 25.22 ± 1.17b |
新窗口打开
2.2 土壤团聚体含量的分布与气候因子的关系
进一步分析草原生态系统土壤团聚体含量的分布与气候因子之间的关系。结果(表3)表明: 在深度小于50 cm的土层中, 总团聚体的质量百分比与年降水量(MAP)显著正相关(p < 0.01), 而在大于50 cm的土层中, 总团聚体的质量百分比与年降水量几近正相关关系(p = 0.06)(表3)。Table 3
表3
表3各层土壤团聚体总质量百分数与气候因子的一元线性关系
Table 3The linear relationships between total aggregate mass percentage of different soil layers and climate factors
| 土层 Soil layer (cm) | 年降水量 Mean annual precipitation (MAP) (mm) | 年平均气温 Mean annual air temperature (MAT) (℃) | ||||||
|---|---|---|---|---|---|---|---|---|
| a | b | R2 | p | a | b | R2 | p | |
| 0-10 | 0.13 | -2.10 | 0.26 | < 0.01 | -2.78 | 41.06 | 0.20 | < 0.01 |
| 10-20 | 0.12 | 0.17 | 0.18 | < 0.01 | -2.59 | 39.65 | 0.14 | < 0.01 |
| 20-30 | 0.13 | -4.05 | 0.20 | < 0.01 | -3.21 | 10.24 | 0.20 | < 0.01 |
| 30-50 | 0.12 | -1.64 | 0.16 | < 0.01 | -3.21 | 39.26 | 0.21 | < 0.01 |
| 50-70 | 0.13 | -7.19 | 0.11 | 0.06 | -3.32 | 38.21 | 0.15 | 0.03 |
| 70-100 | 0.14 | -11.35 | 0.11 | 0.06 | -2.73 | 36.29 | 0.07 | 0.11 |
新窗口打开
土层深度在70 cm以上的总团聚体的质量百分比与年平均气温呈显著负相关关系(p < 0.01), 而70-100 cm土层(p = 0.11)的总团聚体的质量百分比与年平均气温(MAT)无显著负相关关系(表3)。
2.3 土壤团聚体稳定性的分配特征
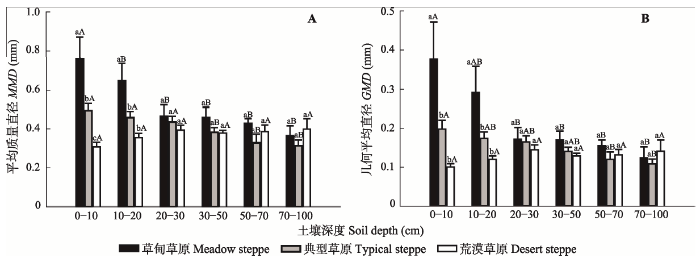
草甸草原与典型草原土壤团聚体的MMD与GMD随着土层深度均呈逐渐降低的趋势, 而荒漠草原并无明显的规律(图2), 具体结果见附录III。统计分析结果表明: 在0-10 cm和10-20 cm土层中, 草甸草原土壤团聚体的MMD与GMD都显著高于典型草原和荒漠草原(p < 0.05), 但在其他土层3种草原土壤团聚体的MMD与GMD都没有显著差异(图2)。对于草甸草原, 0-10 cm土壤团聚体的MMD与GMD都显著高于其他土层(p < 0.05); 对于典型草原, 小于30 cm土层土壤团聚体的MMD与GMD都显著高于大于30 cm; 对于荒漠草原, 各层土壤团聚体的MMD与GMD都无显著差异(图2)。Appendix III
附录III
附录III各个土层团聚体的平均质量直径与几何平均直径(平均值±标准误差)
Appendix IIIThe mean mass diameter and geometric mean diameter of soil aggregates in different soil layers (mean ± SE)
| 土层 Soil layer (cm) | 草甸草原 Meadow steppe | 典型草原 Typical steppe | 荒漠草原 Desert steppe | |||
|---|---|---|---|---|---|---|
| MMD | GMD | MMD | GMD | MMD | GMD | |
| 0-10 | 0.76 ± 0.11 | 0.38 ± 0.10 | 0.49 ± 0.04 | 0.20 ± 0.02 | 0.31 ± 0.02 | 0.04 ± 0.01 |
| 10-20 | 0.65 ± 0.09 | 0.29 ± 0.07 | 0.46 ± 0.03 | 0.17 ± 0.02 | 0.36 ± 0.02 | 0.12 ± 0.01 |
| 20-30 | 0.46 ± 0.06 | 0.17 ± 0.03 | 0.44 ± 0.03 | 0.16 ± 0.02 | 0.39 ± 0.03 | 0.14 ± 0.01 |
| 30-50 | 0.46 ± 0.05 | 0.17 ± 0.02 | 0.38 ± 0.02 | 0.14 ± 0.01 | 0.38 ± 0.01 | 0.13 ± 0.01 |
| 50-70 | 0.43 ± 0.02 | 0.16 ± 0.01 | 0.33 ± 0.04 | 0.12 ± 0.02 | 0.39 ± 0.03 | 0.13 ± 0.01 |
| 70-100 | 0.37 ± 0.05 | 0.12 ± 0.03 | 0.31 ± 0.03 | 0.11 ± 0.01 | 0.40 ± 0.05 | 0.14 ± 0.03 |
新窗口打开
 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT图2三种草原类型在不同土壤深度上的平均质量直径(A)和几何平均直径(B) (平均值±标准误差)。小写字母表示在同一土层中3种草原类型在5%水平上的差异; 大写字母表示同一种草原类型不同土壤深度在5%水平上的差异。
-->Fig. 2Mean mass diameter (MMD) (A) and geometric mean diameter (GMD) (B) of soil aggregates at different soil depths in three types of grassland (mean ± SE). Lowercase letters represent the differences of MMD and GMD in the same soil layer among three types of grassland at the level of 5%. Uppercase letters represent the differences of MMD and GMD among different soil layers in one type of grassland at the level of 5%.
-->
3 讨论
土壤团聚体是土壤有机碳的重要物理保护方式, 其稳定性与土壤有机碳稳定性密切相关(Bird et al., 2002; Denef et al., 2007)。因此, 探讨土壤团聚体形成及稳定的分布格局对于理解生态系统土壤健康及有机碳库稳定性具有重要意义。本研究分析了我国内蒙古不同草原类型土壤团聚体在不同气候梯度及土层深度的分布特征, 发现草原类型对土壤团聚体的形成和稳定具有显著影响, 且土壤团聚体含量随年降水量和年平均气温呈现一定的分布格局。3.1 土壤团聚体的质量分布
土壤团聚体是通过结合体(如细根、微生物菌丝、糖分及黏粒、粉粒等)将土壤颗粒黏结在一起而形成的(Six et al., 2002b), 因此影响结合体的形式和量的因素均可能影响土壤团聚体的形成和分布。有研究发现, 内蒙古草原大部分根系主要分布在0- 30 cm土层(Abrishamkesh et al., 2011); 微生物生物量碳的分布随着土层深度的增加而减少(Blume et al., 2002)。因此, 我们期待团聚体质量百分数随土层深度增加而减少, 而本文的结果显示土层深度对土壤总团聚体的质量百分数并无显著影响。这可能是因为形成大团聚体的有机质较多来源于凋落物、根系和微生物等, 而形成小团聚体的有机质来源并不确定, 导致了大团聚体的含量随着土层深度逐渐减少而小团聚体的含量上升(附录I), 因而总团聚体质量百分数不一定受土层深度的影响。另外, 我们发现草原类型对总团聚体组分的影响显著, 表明内蒙古不同草原类型的土壤团聚体分布具有显著差异, 这可能与不同草原类型的气候因子(包括气温和降雨, 地上、地下生物量(Ma et al., 2008)以及微生物生物量及组成(Li & Chen, 2004))密切相关。在一项关于内蒙古半干旱草原退耕还草的研究中, 在0- 30 cm的土层内, 尽管退耕还草的样地地上和地下生物量随着保护时间推移而增加, 但在研究期间, 天然草原的地上和地下生物量始终都显著高于退耕还草的样地, 导致天然草地在0-30 cm土层中的团聚体含量以及团聚体的稳定性都显著高于退耕还草的样地(Zhang et al., 2013), 本研究与之相似。3.2 土壤团聚体含量的分布与气候因子的关系
降水和气温是影响各个生态系统过程的重要因子, 包括物种分布, 生态系统碳、水及养分循环等(Luo et al., 2008; Austin & van Niel, 2011; Wu et al., 2011)。例如, 中国草原地上、地下生物量及土壤有机碳库的分布格局均受年降水量及年平均气温的影响(Yang et al., 2010a, 2010b), 而内蒙古草原土壤团聚体含量的分布格局与降水和气温的关系并不清楚。本样带调查的结果表明, 总团聚体的质量百分数与年降水量呈正相关关系, 而与年平均气温呈负相关关系(表2), 表明在内蒙古草原生态系统, 降水量高能促进土壤团聚体的形成, 气温高则不利于土壤团聚体的形成。这可能是因为降水量高, 导致地上、地下生物量也高(Yang et al., 2010a), 使得有机碳输入量高, 而促进了土壤团聚体的形成。另外, MAT与MAP对深层(50-70 cm、70-100 cm)土壤的团聚体含量的分布并无显著影响, 这可能是因为气候主要通过影响植物碳输入而影响土壤团聚体的形成, 而深层土壤受植物碳输入的影响较小, 更受土壤质地的影响(Jobbágy & Jackson, 2000)。3.3 土壤团聚体稳定性与草原类型及土层深度的关系
土壤团聚体的MMD和GMD是反映土壤团聚体稳定性的重要指标(Kemper & Chepil, 1965)。本研究结果表明, MMD与GMD在不同草原类型以及不同土壤深度的趋势一致。20 cm以上的土层深度中, 草甸草原土壤团聚体的MMD和GMD均显著高于典型和荒漠草原土壤团聚体的MMD和GMD, 这可能是因为草甸草原较典型草原和荒漠草原湿润, 拥有的物种多样性丰富, 草群盖度较高(李博, 1979) , 从而导致根系发达, 而根系对土壤团聚体的形成和稳定性有很大影响(Pinheiro et al., 2004)。另有研究结果表明, 随着土层深度增加, MMD和GMD都呈现降低的趋势(赵世伟等, 2006; 陈帅和孙涛, 2017), 我们的研究结果与之相似。本研究也发现草甸草原和典型草原土壤团聚体的MMD和GMD随着土层深度逐渐增加而降低, 这可能是因为湿度、微生物生物量及组成随着土层深度逐渐增加而降低, 从而导致形成团聚体的黏合剂含量降低, 进一步导致了团聚体稳定性在深度上的变化(Allison et al., 2007)。然而, 荒漠草原土壤团聚体的MMD和GMD并没有明确的规律, 这可能是因为荒漠草原植被覆盖度低, 其土壤微生物生物量及土壤有机碳含量也低, 使得土壤团聚体稳定性低(Bird et al., 2007), 从而导致在土层深度上差异不明显。The authors have declared that no competing interests exist.
作者声明没有竞争性利益冲突.
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | . Since the beginning of the last century, many studies have reported evidence describing the effects of organic inputs on soil aggregate stability. In 1965, Monnier proposed a conceptual model that considers different patterns of temporal effects on aggregate stability depending on the nature of the organic inputs: easily decomposable products have an intense and transient effect on aggregate stability while more recalcitrant products have a lower but longer term effect. We confronted this conceptual model with a literature review of experimental data from laboratory and field experiments. This literature analysis validated the conceptual model proposed by Monnier and pointed out gaps in our current knowledge concerning the relationships between aggregate stability and organic inputs. Noticeably, the experimental dataset confirmed the biological and temporal effects of organic inputs on aggregate stability as proposed in the model. Monnier's model also related the evolution of aggregate stability to different microbial decomposing agents, but this relationship was not made clear in this literature analysis. No direct or universal relationship was found between the aggregative factors induced by organic input decomposition (binding molecules or decomposers of biomass) and temporal aggregate stability dynamics. This suggests the existence of even more complex relationships. The model can be improved by considering (i) the direct abiotic effect of some organic products immediately after the inputs, (ii) the initial biochemical characteristics of the organic products and (iii) the effects of organic products on the various mechanisms of aggregate breakdown. For now, no trend is evident in the effect of the rate of organic inputs or the effect of the soil characteristics (essentially carbon and clay contents) on aggregate stability. |
| [2] | . |
| [3] | . Simultaneously assessing shifts in microbial community composition along landscape and depth gradients allows us to decouple correlations among environmental variables, thus revealing underlying controls on microbial community composition. We examined how soil microbial community composition changed with depth and along a successional gradient of native prairie restoration. We predicted that carbon would be the primary control on both microbial biomass and community composition, and that deeper, low-carbon soils would be more similar to low-carbon agricultural soils than to high carbon remnant prairie soils. Soil microbial community composition was characterized using phospholipid fatty acid (PLFA) analysis, and explicitly linked to environmental data using structural equations modeling (SEM). We found that total microbial biomass declined strongly with depth, and increased with restoration age, and that changes in microbial biomass were largely attributable to changes in soil C and/or N concentrations, together with both direct and indirect impacts of root biomass and magnesium. Community composition also shifted with depth and age: the relative abundance of sulfate-reducing bacteria increased with both depth and restoration age, while gram-negative bacteria declined with depth and age. In contrast to prediction, deeper, low-C soils were more similar to high-C remnant prairie soils than to low-C agricultural soils, suggesting that carbon is not the primary control on soil microbial community composition. Instead, the effects of depth and restoration age on microbial community composition were mediated via changes in available phosphorus, exchangeable calcium, and soil water, together with a large undetermined effect of depth. Only by examining soil microbial community composition shifts across sites and down the soil column simultaneously were we able to tease apart the impact of these correlates environmental variables. |
| [4] | . Soil aggregate stability is a crucial soil property affecting soil sustainability and crop production. A broad outline of the processes and agents of aggregate formation and aggregate stabilization are presented and discussed in this review. Aggregate stability is difficult to quantify and interpret. The aim of aggregate stability tests is to give a reliable description and ranking of the behavior of soils under the effect of water, wind and management. Numerous methods have been used to determine aggregate stability with varying success. The different methodologies complicate the comparison among aggregate stability data. It is also difficult to obtain a consistent correlation between aggregate stability and other important soil properties such as soil erodibility or crusting potential. This paper reviews the different methods of measurement of soil aggregate stability used in the literature, paying attention to the conditions of sample collection in the field and sample preparation and treatments in the laboratory. A unified methodological framework including the most interesting aspects of existing methods is suggested. The possibility of using aggregate stability data as an estimation of soil erodibility is also discussed. |
| [5] | . Abstract Statistical species distribution models (SDMs) are widely used to predict the potential changes in species distributions under climate change scenarios. We suggest that we need to revisit the conceptual framework and ecological assumptions on which the relationship between species distributions and environment is based. We present a simple conceptual framework to examine the selection of environmental predictors and data resolution scales. These vary widely in recent papers, with light inconsistently included in the models. Focusing on light as a necessary component of plant SDMs, we briefly review its dependence on aspect and slope and existing knowledge of its influence on plant distribution. Differences in light regimes between north- and south-facing aspects in temperate latitudes can produce differences in temperature equivalent to moves 200km polewards. Local topography may create refugia that are not recognized in many climate change SDMs using coarse-scale data. We argue that current assumptions about the selection of predictors and data resolution need further testing. Application of these ideas can clarify many issues of scale, extent and choice of predictors, and potentially improve the use of SDMs for climate change modelling of biodiversity. |
| [6] | . Understanding how the aboveground net primary production (ANPP) of arid and semiarid ecosystems of the world responds to variations in precipitation is crucial for assessing the impacts of climate change on terrestrial ecosystems. Rain-use efficiency (RUE) is an important measure for acquiring this understanding. However, little is known about the response pattern of RUE for the largest contiguous natural grassland region of the world, the Eurasian Steppe. Here we investigated the spatial and temporal patterns of ANPP and RUE and their key driving factors based on a long-term data set from 21 natural arid and semiarid ecosystem sites across the Inner Mongolia steppe region in northern China. Our results showed that, with increasing mean annual precipitation (MAP), (1) ANPP increased while the interannual variability of ANPP declined, (2) plant species richness increased and the relative abundance of key functional groups shifted predictably, and (3) RUE increased in space across different ecosystems but decreased with increasing annual precipitation within a given ecosystem. These results clearly indicate that the patterns of both ANPP and RUE are scale dependent, and the seemingly conflicting patterns of RUE in space vs. time suggest distinctive underlying mechanisms, involving interactions among precipitation, soil N, and biotic factors. Also, while our results supported the existence of a common maximum RUE, they also indicated that its value could be substantially increased by altering resource availability, such as adding nitrogen. Our findings have important implications for understanding and predicting ecological impacts of global climate change and for management practices in arid and semiarid ecosystems in the Inner Mongolia steppe region and beyond. |
| [7] | . To measure and manage for C sequestration in heterogeneous rangeland systems, we need to more fully understand spatial patterns of soil resources. Spatial distributions of aggregate stability and soil carbon were investigated in a semiarid rangeland in New Mexico, USA. Soil was analyzed from plant interspaces, black grama (Bouteloua eriopoda (Torr.) Torr.), and mesquite (Prosopis glandulosa Torr.) in a landscape-replicated study. Aggregate stability at the 250 microm scale, carbonate C, organic C and N, C:N ratio, and glomalin, were all highest under mesquite. Soil C:N ratio was the best predictor of aggregate stability. Estimates of metric tons of C per hectare in the top 10 cm were highly variable at patch and landscape scales, varying from 4.2 to 10.5 under mesquite and from 3.0 to 7.0 in interspaces. High variability of aggregate stability and soil C has important implications for C sequestration. We argue that this multi-scale soil heterogeneity must be considered when measuring and managing for C sequestration. |
| [8] | . Increased soil loss and redistribution are commonly associated with changes in soil structure, yet variability in soil structure in arid ecosystems has been little studied. Soil aggregate stability is a key indicator of soil structure and is correlated with erodibility and water infiltration capacity. In 2000, we compared soil aggregate stability of a complex of Simona (Loamy, mixed, thermic, shallow Typic Paleorthids) and Harrisburg (Coarse-loamy, mixed, thermic, Typic Paleorthids) soils in a Chihuahuan Desert grassland. We examined soil stability at plant and landscape scales by assessing percentage aggregate stability at four sites in two cover classes (plant vs. interspace) located within each of three grass cover and land disturbance classes. To increase measurement sensitivity to changes in soil structure and identify potential early warning indicators for monitoring, we used two different methods for quantifying wet aggregate stability: a laboratory method using a 0.25 mm sieve and a field method using a 1.5 mm sieve. As expected, soil aggregate stability was significantly higher under grass plants than in plant interspaces (44.2 vs. 38.4 for the lab test and 4.4 vs. 3.3 for the field test; P < 0.01). The field test showed higher stability in plots with higher grass cover throughout the top 10 mm soil layer, while disturbance level only affected stability at the soil surface. The laboratory test was insensitive to differences in grass cover and disturbance. |
| [9] | . Microbial biomass, size and community structure along with an estimate of microbial activity and soil chemical parameters were determined at three depths in two soils (e.g. sandy loam Ultic Hapludalf and silt loam Mollic Hapludalf) replicated three times under one winter and summer season. Microbial biomass and community structure were estimated from phospholipid-PO 4 content and fatty acid methyl ester (FAME) measurements. Microbial activity and assimilative capacity were estimated using a 3 H-acetate incorporation into phospholipids and by incubating the soil samples at the average winter and summer temperatures, 3 and 20 C, respectively. We found that the size of the microbial biomass in both the surface and the subsurface soils was not significantly affected by the seasonal variation but activity increased by as much as 83% at the summer temperatures in the surface soil. We demonstrated using FAME analysis that for both soils seasonal changes in the subsurface microbial community occurred. These findings suggest that winter conditions will shift the population activity level in both the surface and subsurface systems and the biochemical structure of the community in the subsurface. In all cases, the inorganic chemical properties of the soil, as a function of season, remained constant. The greatly increased activity of microbial population at the higher temperature will favor the capacity of the system to utilize nutrients or organic materials that may enter soil. During low temperature seasons the capacity of either surface or subsurface soils to assimilate materials is generally diminished but the reduction reflects changes in metabolism and not a reduced biomass size. |
| [10] | . Effects of arbuscular mycorrhzal (AM) fungi on plant growth and nutrition are well-known, but their effects on the wider soil biota are less clear. This is in part due to difficulties with establishing appropriate non-mycorrhizal controls in the field. Here we present results of a field experiment using a new approach to overcome this problem. A previously well-characterized mycorrhizal defective tomato mutant ( rmc ) and its mycorrhizal wildtype progenitor (76R MYC+) were grown at an organic fresh market tomato farm (Yolo County, CA). At the time of planting, root in-growth cores amended with different levels of N and P, were installed between experimental plants to study localized effects of mycorrhizal and non-mycorrhizal tomato roots on soil ecology. Whilst fruit yield and vegetative production of the two genotypes were very similar at harvest, there were large positive effects of colonization of roots by AM fungi on plant nutrient contents, especially P and Zn. The presence of roots colonized by AM fungi also resulted in improved aggregate stability by increasing the fraction of small macroaggregates, but only when N was added. Effects on the wider soil community including nematodes, fungal biomass as indicated by ergosterol, microbial biomass C, and phospholipid fatty acid (PLFA) profiles were less pronounced. Taken together, these data show that AM fungi provide important ecosystem functions in terms of plant nutrition and aggregate stability, but that a change in this one functional group had only a small effect on the wider soil biota. This indicates a high degree of stability in soil communities of this organic farm. |
| [11] | . . 受人类不合理利用和气候影响的综合作用,松嫩草地退化程度逐渐加剧。本研究选取羊草(Leymus chinensis)群落、虎尾草(Chloris virgata)群落、碱茅(Puccinellia distans)群落和碱蓬(Suaeda glauca)群落作为4个草地退化阶段,通过调查植被群落特征、分析土壤理化性质,测定土壤团聚体稳定性指标,揭示草地植被退化过程中对土壤结构的影响。结果表明,随着退化演替的进行,植被生物量、物种多样性指数和总盖度总体上均呈现为降低的趋势;土壤含水率、有机质含量和全氮均降低,而pH和容重均增加,且4种退化演替阶段的土壤pH均高于8.0;土壤水稳性团聚体的粒径分布中微团聚体(0.25mm)所占比例增加,大团聚体(0.25mm)所占比例和水稳性团聚体的稳定性递减,且在同一退化阶段的0-30cm各土层呈现出水稳性团聚体的稳定性随土层深度增加而稳定性递减的趋势。研究结果对揭示松嫩草地不同退化阶段土壤团聚体稳定性特征以及退化草地恢复重建均有重要指导意义。 |
| [12] | . Soil organic matter is thought to increase aggregate stability by lowering the wettability and increasing the cohesion of aggregates. In SW France, thick humic loamy soils (Vermic Haplumbrepts) have been intensively cropped for 40 years, decreasing the soil organic pool and lowering the soil aggregate stability. This study assessed (i) the contribution of organic matter to aggregate stability b... |
| [13] | . Carbon stabilization by macroaggregate-occluded microaggregates (Mm) has been proposed as a principal mechanism for long-term soil organic carbon (SOC) sequestration in temperate alternative agricultural and (af)forested systems. The aim of this study was to evaluate the importance of the Mm fraction for long-term C stabilization in Oxisols and to validate its diagnostic properties for total SOC changes upon changes in land use. Soil samples were taken from the 0–5 and 5–20 cm soil layers of native forest vegetation (NV), conventional tillage (CT) and no-tillage (NT) systems at an experimental site near Passo Fundo and one near Londrina in Southern Brazil. After aggregate-size separations by wet-sieving, macroaggregate-occluded water-stable microaggregates (53–250 μm) (Mm) were isolated from large (>2000 μm) and small (>250 μm) macroaggregates. Particulate organic matter located inside the Mm (intra-Mm-POM) and the mineral fraction (< 53 μm) associated with the Mm (mineral-Mm) were separated from the POM fraction located outside the Mm (inter-Mm-POM) by density flotation followed by mechanical dispersion. Sand-free Mm-C concentrations on a macroaggregate basis were generally greater under NV and NT compared to CT in the 0–5 cm depth at both sites. Our findings support the importance of Mm (especially the mineral-Mm fraction) as long-term C-stabilization sites in highly weathered tropical soils under sustainable agricultural and natural systems. At both sites, significant differences in total SOC stocks (g C m 612) among different land use systems were always accompanied by parallel Mm-C stock differences. Though total SOC did not differ among land use systems in the 0–20 cm depth at both sites, Mm-C stocks were greater under NT compared to the CT treatment in the 0–20 cm depth at the Londrina site. We concluded that in these highly weathered tropical soils the Mm-C fraction is a more responsive fraction to management changes than total SOC and represents a diagnostic fraction for present as well as potential total SOC changes upon land-use change. |
| [14] | . The of soil aggregates appears to be a primary mechanism in the loss of organic matter caused by long-term cultivation, but little information exists on how the and stabilization of macroaggregates control the process of C aggradation when disturbance is reduced or eliminated. A chronosequence of restored tallgrass prairie was used to investigate the relationships between the of stable macroaggregates (> 212 micrometer dia) and the accrual of particulate and -associated organic matter. Changes in the percentage of macroaggregates and in the accumulation of whole-soil organic C across the chronosequence were both described with a simple exponential model. The rate constant (k) for change in aggregation was more than 35 times the k for total organic C accumulation. Thus, the time required to reach 99% of equilibrium was 10.5 y for macroaggregates and 384 y for whole-soil organic C, providing evidence for the existence of a phased relationship between macroaggregate and C accrual. The input rate for whole-soil organic C to a 10-cm depth was estimated at 1.16 g kg-1 y-1 or 0.133 kg m-2 y-1 (assuming an average bulk density of 1150 kg m-3 for previously cultivated soils in the chronosequence). An increase in macroaggregate-associated C-to-N ratios with time since cultivation suggested that the accumulating organic matter was not "highly processed", but less than 20% of the accrued C occurred in the form of particulate organic matter (density less than or equal to 1.85 g cm-3). Rather, most of the accumulated C occurred in the -associated fraction of macroaggregates, suggesting that inputs of organic debris were rendered relatively rapidly into particles or colloids that are associated with matter and thus are physically protected, slowing decomposition and promoting the development of stable microaggregates within macroaggregates. |
| [15] | . |
| [16] | |
| [17] | . Increasing agronomic productivity and improving the quality of the environment are among the important goals of soil physical management in the tropics. Several perceptions, created by insufficient scientific data and misunderstanding of the basic processes, are being resolved by an improved data base and better understanding of the dynamics of soil physical properties and processes. Principal issues in the tropics for the 21st century include: (i) achieving food security; (ii) curtailing soil degradation and restoring degraded soils; and (iii) improving environment quality. These issues can be addressed by identifying and prioritizing research needs in soil physical management. Important among these are: (i) assessing soil physical constraints at farm scale; (ii) managing soil structure and tilth to minimize the risks of crusting, compaction, and hard-setting; (iii) quantifying and controlling soil erosion by water and wind, developing a cause-effect relationship between soil and climate factors, and evaluating the effects on productivity; (iv) managing soil-water, controlling soil salinity in irrigated agriculture, and developing water harvesting techniques in rain-fed agricultural systems; (v) studying the dynamics of soil physical properties in puddled soils of rice-based cropping systems; (vi) developing conservation tillage and residue management methods to improve soil tilth; (vii) understanding soil moisture retention characteristics in relation to plant available water capacity; and (viii) developing indicators of soil physical quality. Important environmental issues relevant to soil physical management are transport of agricultural chemicals into surface and ground waters, emission of greenhouse gases from soils to the atmosphere, and disposal of urban and industrial wastes. Soil physical management in the tropics must be based on a holistic approach to solve practical problems. It is also important to make the public aware of the contributions of soil science to society's well-being. In addition to enhancing food production, soil physical management needs to address environmental, engineering, social, legal, and archeological issues. Achieving these goals necessitates soil scientists working in close collaboration with engineers, climatologists, geologists, biologists, and specialists in GIS and geostatistics. |
| [18] | . Abstract Aggregate stability, an important property influencing a soil's response to erosive forces, is affected by freezing. The objectives of this laboratory study were to determine how constrainment, number of freeze-thaw cycles, and water content at freezing affect the aggregate stability of six continental USA soils differing in texture, mineralogy, and organic-matter content. Moist aggregates, after being frozen and thawed either zero, one, three, or five times, were vapor wetted to 0.30 kg kg-1 and analyzed by wet sieving. Soils with clay contents of 17% or more and organic-matter contents >3% were the most stable after freezing. Aggregate stability for fine- and medium-textured soils generally decreased linearly with increasing water content at freezing. This linear decrease in stability was more rapid for constrained samples than for unconstrained samples. The stability of field-moist aggregates generally increased from zero to one or three freeze-thaw cycles. For at least one low-organic-matter soil, stability increased from one to three freeze-thaw cycles, but then decreased at five cycles. After thawing, aggregates at water contents of 0.15 kg kg-1 or more that were constrained when frozen were always significantly less stable than aggregates that were un-constrained when frozen. |
| [19] | . . |
| [20] | . . 探讨了我国西南地区缙云山亚热带常绿阔叶林(以下简称林地)、果园、坡耕地以及撂荒地4种土地利用方式对土壤有机碳(SOC)组分含量及其分配比例的影响。采用物理分组技术,将SOC分为粗颗粒有机碳(cPOC)、细颗粒有机碳(fPOC)、微团聚体内颗粒有机碳(iPOC)、微团聚体内粉+黏颗粒(s+c_m)有机碳及粉+黏颗粒(s+c)有机碳。研究结果表明:在0—60cm的土壤深度范围内,SOC、cPOC、fPOC、iPOC、s+c_m组分以及s+c组分有机碳平均含量均为林地(9.02、3.14、1.61、0.33、0.42、3.53g/kg)显著高于果园(3.27、0.93、0.27、0.10、0.24、1.73g/kg)和坡耕地(2.58、0.51、0.10、0.12、0.08、1.77g/kg),说明林地开垦会导致SOC及各组分的流失;而撂荒地上述SOC及其各组分含量分别为14.90、5.17、2.36、0.42、0.59和6.36g/kg,均显著高于坡耕地,表明耕地撂荒后SOC及其组分能得到有效的恢复和截存。在SOC的各物理组分中,iPOC的有机碳分配比例最低,4种土地利用方式下均为3%左右;cPOC和fPOC作为活性较强的非保护有机碳库,在林地和撂荒地中所占SOC分配比例最高,达到50%以上;而果园和坡耕地中〈53um的粉+黏颗粒有机碳组成的化学保护有机碳库分配比例最大,分别为65.9%和71.6%,表明林地和撂荒地土壤有机碳的活性远远大于坡耕地及果园,支持更高的土壤肥力。在SOC及其组分中,fPOC可作为评估土地利用变化对土壤有机碳库影响的良好指标。 |
| [21] | . The relationships of soil microbial biomass C (C mic ) or N (N mic ) with mean annual precipitation and temperature were studied along a climatic transect in the Mongolian steppe. Soil organic C (C org ) and total N (N t ), respiration rate, C mic and N mic at depths of 0615 and 5611002cm decreased with increasing aridity. The contents of C org and N t in the 0- to 5-cm soil layers decreased linearly with precipitation reduction along the transect. C mic and N mic changes with precipitation were not linear, with higher changes between 330 and 12802mm mean annual precipitation. C mic /C org and N mic /N t increased with increasing aridity. The metabolic quotient qCO 2 of 0- to 5-cm soil layers was low between 330 and 27302mm precipitation. The relationship between the qCO 2 of the 0- to 5-cm soil layers and the mean annual precipitation was well fitted with a quadratic function y =0.0006 x 2 610.40 x +86.0, where y is the qCO 2 (08mol02CO 2 -C02mmol 611 02C mic ) and x is the mean annual precipitation (mm). C org , N t , C mic , N mic and respiration rate decreased exponentially with increasing mean annual temperature in both the 0- to 5- and 5- to 10-cm soil layers, and change rate was lower when the mean annual temperature was higher than 2.6°C. The close relationships of the mean annual precipitation or temperature with soil C org , N t , C mic , N mic , C mic /C org and qCO 2 indicate that each parameter can be calculated by determining the other parameters in this specific climatic range. |
| [22] | . . |
| [23] | . Abstract Interactive effects of multiple global change factors on ecosystem processes are complex. It is relatively expensive to explore those interactions in manipulative experiments. We conducted a modeling analysis to identify potentially important interactions and to stimulate hypothesis formulation for experimental research. Four models were used to quantify interactive effects of climate warming ( T ), altered precipitation amounts [doubled (DP) and halved (HP)] and seasonality (SP, moving precipitation in July and August to January and February to create summer drought), and elevated [CO 2 ] ( C ) on net primary production (NPP), heterotrophic respiration ( R h ), net ecosystem production (NEP), transpiration, and runoff. We examined those responses in seven ecosystems, including forests, grasslands, and heathlands in different climate zones. The modeling analysis showed that none of the three-way interactions among T , C , and altered precipitation was substantial for either carbon or water processes, nor consistent among the seven ecosystems. However, two-way interactive effects on NPP, R h , and NEP were generally positive (i.e. amplification of one factor's effect by the other factor) between T and C or between T and DP. A negative interaction (i.e. depression of one factor's effect by the other factor) occurred for simulated NPP between T and HP. The interactive effects on runoff were positive between T and HP. Four pairs of two-way interactive effects on plant transpiration were positive and two pairs negative. In addition, wet sites generally had smaller relative changes in NPP, R h , runoff, and transpiration but larger absolute changes in NEP than dry sites in response to the treatments. The modeling results suggest new hypotheses to be tested in multifactor global change experiments. Likewise, more experimental evidence is needed for the further improvement of ecosystem models in order to adequately simulate complex interactive processes. |
| [24] | . . |
| [25] | . Aim Our objective was to document the general relationship between plant species richness (SR) and above-ground net primary productivity (ANPP) at different spatial scales and the environmental influence on this relationship.Location Temperate and alpine grasslands of China.Methods We investigated SR and ANPP at 321 field sites (1355 plots) across the widely distributed temperate and alpine grasslands of China. Ordinary least squares (OLS) regressions were used to test SR NPP relationships among site means. Plot-level data of SR and ANPP were analysed with general linear models (GLMs) and the correlation between SR and ANPP was decomposed into covariance components to test the influence of climatic variables, region, vegetation type and remaining variation among sites on SR, ANPP and their relationship.Results We found positive linear relationships between SR and ANPP among sites in both the alpine and temperate grassland regions and in different grassland vegetation types of these biomes. Environmental gradients such as growing-season precipitation affected both SR and ANPP in parallel. However, after removing the among-site environmental variation, residual SR and ANPP were no longer correlated at the pooled within-site level.Main conclusions The positive SR NPP relationship across large-scale environmental gradients among sites was most likely the result of climatic variables influencing SR and ANPP in parallel. Our results suggest that in China's natural grasslands there is no direct relationship between SR and ANPP, presumably because the pool of available species for local community assembly is large, in contrast to experiments where species pools are artificially reduced. |
| [26] | . Above- and belowground biomasses of grasslands are important parameters for characterizing regional and global carbon cycles in grassland ecosystems. Compared with the relatively detailed information for aboveground biomass (AGB), belowground biomass (BGB) is poorly reported at the regional scales. The present study, based on a total of 113 sampling sites in temperate grassland of the Inner Mongolia, investigated regional distribution patterns of AGB, BGB, vertical distribution of roots, and their relationships with environmental factors. AGB and BGB increased from the southwest to the northeast of the study region. The largest biomass occurred in meadow steppe, with mean AGB and BGB of 196.7 and 1385.2 g/m 2 , respectively; while the lowest biomass occurred in desert steppe, with an AGB of 56.6 g/m 2 and a BGB of 301.0 g/m 2 . In addition, about 47% of root biomass was distributed in the top 10 cm soil. Further statistical analysis indicated that precipitation was the primary determinant factor in shaping these distribution patterns. Vertical distribution of roots was significantly affected by precipitation, while the effects of soil texture and grassland types were weak. |
| [27] | . A new theoretical and experimental framework that permits an accurate determination of aggregate-size stability distribution is presented. The size-stability distribution in addition to estimating aggregate-size distribution distinguishes between amounts of stable and unstable macroaggregates (>250 m). The determination of aggregate-size stability distribution involves the assumptions that soil aggregates can be categorized in terms of their size and water stability (slaking resistance). Experimentally this procedure involves the slaked and capillary-wetted pretreatments; and a subsequent slaking treatment of aggregates >250 m in size. We also propose the stable aggregates index (SAI) and the stable macroaggregates index (SMaI) for studying soil stability based on aggregate resistance to slaking. These indices account for the total weighted average of stable aggregates and the total weighted average of stable macroaggregates, respectively. Both the SAI and the SMaI indices were shown to be sensitive to the effects of vegetation on soil stability under different riparian buffer communities. The SAI and the SMaI indices were higher in surface soils under cool-season grass than any of the other treatments. These soils samples are well aggregated with SAI = 74% and SMaI = 56% followed by SAI = 55% and SMaI = 37% under existing riparian forest, SAI = 40% and SMaI = 21% under 7-yr switchgrass and SAI = 36% and SMaI = 18% under cropped system. |
| [28] | |
| [29] | . Several management systems can improve soil productivity. By studying aggregate stability it is possible to quantify whether or not the management is ameliorating the natural soil properties and the land capability for agriculture. The effect of three tillage systems on the stability of soil aggregates and soil organic carbon was studied in comparison to reference plots with grass and bare soil. Samples were collected at the Pesagro Experimental Research Station in Paty do Alferes, state of Rio de Janeiro, Brazil, from an experiment that has been carried out from 1995 to 2001, on a Dystrophic Red Latosol (Typic Haplorthox). Aggregate size distribution mean weight diameter, geometric mean diameter of the aggregates, and total organic carbon in each aggregate size fraction were determined. The proportion of aggregates with diameter ≥202mm appeared to be a suitable indicator of the influence of tillage systems on aggregation. At a depth of 0–502cm, aggregates ≥202mm were 50% of soil under no-tillage, which was greater than under animal traction (35%) and conventional tillage (30%). Total organic carbon concentration was greater under no-tillage (1902g02kg) than under conventional tillage (1102g02kg) at a depth of 0–502cm, but not significantly different (average 1302g02kg) at a depth of 5–1002cm. Soil exposure with tillage and lack of residue inputs caused declines in aggregation and organic carbon, both of which make soil susceptible to erosion. Adoption of no-tillage led to a decline in aggregation compared with grass reference, but did significantly alter soil organic concentration, suggesting it was a valuable conservation practice for vegetable production on sloping soils. |
| [30] | . For soil carbon to be effectively sequestered beyond a timescale of a few decades, this carbon must become incorporated into passive reservoirs or greater depths, yet the actual mechanisms by which this occurs is at best poorly known. In this study, we quantified the magnitude of dissolved organic carbon (DOC) leaching and subsequent retention in soils of a coniferous forest and a coastal prairie ecosystem. Despite small annual losses of DOC relative to respiratory losses, DOC leaching plays a significant role in transporting C from surface horizons and stabilizing it within the mineral soil. We found that DOC movement into the mineral soil constitutes 22% of the annual C inputs below 4002cm in a coniferous forest, whereas only 2% of the C inputs below 2002cm in a prairie soil could be accounted for by this process. In line with these C input estimates, we calculated advective transport velocities of 1.05 and 0.4502mm02year 611 for the forested and prairie sites, respectively. Radiocarbon measurements of field-collected DOC interpreted with a basic transport-turnover model indicated that DOC which was transported and subsequently absorbed had a mean residence time of 90–15002years. Given these residence times, the process of DOC movement and retention is responsible for 20% of the total mineral soil C stock to 102m in the forest soil and 9% in the prairie soil. These results provide quantitative data confirming differences in C cycles in forests and grasslands, and suggest the need for incorporating a better mechanistic understanding of soil C transport, storage and turnover processes into both local and regional C cycle models. |
| [31] | . The challenge to identify the biospheric sinks for about half the total carbon emissions from fossil fuels must include a consideration of below-ground ecosystem processes as well as those more easily measured above-ground. Recent studies suggest that tropical grasslands and savannas may contribute more to the 'missing sink' than was previously appreciated, perhaps as much as 0.5 Pg (= 0.5 Gt) carbon per annum. The rapid increase in availability of productivity data facilitated by the Internet will be important for future scaling-up of global change responses, to establish independent lines of evidence about the location and size of carbon sinks. |
| [32] | . The relationship between soil structure and the ability of soil to stabilize soil organic matter (SOM) is a key element in soil C dynamics that has either been overlooked or treated in a cursory fashion when developing SOM models. The purpose of this paper is to review current knowledge of SOM dynamics within the framework of a newly proposed soil C saturation concept. Initially, we distinguish SOM that is protected against decomposition by various mechanisms from that which is not protected from decomposition. Methods of quantification and characteristics of three SOM pools defined as protected are discussed. Soil organic matter can be: (1) physically stabilized, or protected from decomposition, through microaggregation, or (2) intimate association with silt and clay particles, and (3) can be biochemically stabilized through the formation of recalcitrant SOM compounds. In addition to behavior of each SOM pool, we discuss implications of changes in land management on processes by which SOM compounds undergo protection and release. The characteristics and responses to changes in land use or land management are described for the light fraction (LF) and particulate organic matter (POM). We defined the LF and POM not occluded within microaggregates (53 250 m sized aggregates as unprotected. Our conclusions are illustrated in a new conceptual SOM model that differs from most SOM models in that the model state variables are measurable SOM pools. We suggest that physicochemical characteristics inherent to soils define the maximum protective capacity of these pools, which limits increases in SOM (i.e. C sequestration) with increased organic residue inputs. |
| [33] | . This study compares the impacts of no-tillage, conventional tillage, and native grasslands on three particulate organic matter (POM) fractions. |
| [34] | . The long-term stabilization of soil organic matter (SOM) in tropical and temperate regions is mediated by soil biota (e.g. fungi, bacteria, roots and earthworms), soil structure (e.g. aggregation) and their interactions. On average, soil C turnover was twice as fast in tropical compared with temperate regions, but no major differences were observed in SOM quality between the two regions. Probably due to the soil mineralogy dominated by 1:1 clay minerals and oxides in tropical regions, we found a higher aggregate stability, but a lower correlation between C contents and aggregate stability in tropical soils. In addition, a smaller amount of C associated with clay and silt particles was observed in tropical versus temperate soils. In both tropical and temperate soils, a general increase in C levels ($\approx$ 325 $\pm$ 113 kg C$\cdot$ha$^{-1}\cdot$yr$^{-1}$) was observed under no-tillage compared with conventional tillage. On average, in temperate soils under no-tillage, compared with conventional tillage, CH$_4$ uptake ($\approx$ 0.42 $\pm$ 0.10 kg C-CH$_4\cdot$ha$^{-1}\cdot$yr$^{-1}$) increased and N$_2$O emissions increased ($\approx$ 1.95 $\pm$ 0.45 kg N-N$_2$O$\cdot$ha$^{-1}\cdot$yr$^{-1}$). These increased N$_2$O emissions lead to a negative global warming potential when expressed on a CO$_2$ equivalent basis. |
| [35] | . . |
| [36] | . 08 2014 Macmillan Publishers Limited. All rights reserved.Higher aridity and more extreme rainfall events in drylands are predicted due to climate change. Yet, it is unclear how changing precipitation regimes may affect nitrogen (N) cycling, especially in areas with extremely high aridity. Here we investigate soil N isotopic values ( 15 N) along a 3,200 ‰km aridity gradient and reveal a hump-shaped relationship between soil 15 N and aridity index (AI) with a threshold at AI=0.32. Variations of foliar 15 N, the abundance of nitrification and denitrification genes, and metabolic quotient along the gradient provide further evidence for the existence of this threshold. Data support the hypothesis that the increase of gaseous N loss is higher than the increase of net plant N accumulation with increasing AI below AI=0.32, while the opposite is favoured above this threshold. Our results highlight the importance of N-cycling microbes in extremely dry areas and suggest different controlling factors of N-cycling on either side of the threshold. |
| [37] | . Abstract Global mean temperature is predicted to increase by 2 7 C and precipitation to change across the globe by the end of this century. To quantify climate effects on ecosystem processes, a number of climate change experiments have been established around the world in various ecosystems. Despite these efforts, general responses of terrestrial ecosystems to changes in temperature and precipitation, and especially to their combined effects, remain unclear. We used meta-analysis to synthesize ecosystem-level responses to warming, altered precipitation, and their combination. We focused on plant growth and ecosystem carbon (C) balance, including biomass, net primary production (NPP), respiration, net ecosystem exchange (NEE), and ecosystem photosynthesis, synthesizing results from 85 studies. We found that experimental warming and increased precipitation generally stimulated plant growth and ecosystem C fluxes, whereas decreased precipitation had the opposite effects. For example, warming significantly stimulated total NPP, increased ecosystem photosynthesis, and ecosystem respiration. Experimentally reduced precipitation suppressed aboveground NPP (ANPP) and NEE, whereas supplemental precipitation enhanced ANPP and NEE. Plant productivity and ecosystem C fluxes generally showed higher sensitivities to increased precipitation than to decreased precipitation. Interactive effects of warming and altered precipitation tended to be smaller than expected from additive, single-factor effects, though low statistical power limits the strength of these conclusions. New experiments with combined temperature and precipitation manipulations are needed to conclusively determine the importance of temperature recipitation interactions on the C balance of terrestrial ecosystems under future climate conditions. |
| [38] | . AbstractAimThe identification of stoichiometric flexibility is crucial for understanding carbon–nitrogen–phosphorus (C–N–P) interactions and ecosystem dynamics under a changing environment. However, current evidence of stoichiometric flexibility mainly comes from manipulation experiments, with little evidence from large-scale observations.LocationAlpine and temperate grasslands across northern China.MethodsUsing soil profiles derived from a historical national soil inventory and a contemporary regional soil survey across China's grasslands, we examined temporal changes in topsoil C:N:P ratios over recent decades.ResultsTopsoil C:N ratios of five major grassland types exhibited some flexibility but did not show significant changes over the sampling interval. Non-significant changes in topsoil C:N ratios were observed both in alpine grasslands on the Tibetan Plateau and in temperate grasslands on the Inner Mongolian Plateau. Consistent with the relatively stable C:N ratios, the slope of the soil C–N stoichiometric relationship did not differ significantly between the two sampling periods. Soil N:P ratios in the surface layer increased significantly over the sampling interval, however, with an overall increase of 0.60 (95% confidence interval 0.58–0.62). A larger increase in soil N:P ratio was found in temperate grasslands on the Inner Mongolian Plateau than in alpine grasslands on the Tibetan Plateau. Moreover, the slope of the soil N–P stoichiometric relationship in these grassland ecosystems became steeper over the sampling interval.Main conclusionsThese results demonstrate the stability of topsoil C:N stoichiometry but variability in N:P stoichiometry over broad geographical scales, highlighting that soil C and N are tightly coupled, but N and P tend to be decoupled under a changing environment. |
| [39] | . ABSTRACT Aim68 To investigate large-scale patterns of above-ground and below-ground biomass partitioning in grassland ecosystems and to test the isometric theory at the community level. Location68 Northern China, in diverse grassland types spanning temperate grasslands in arid and semi-arid regions to alpine grasslands on the Tibetan Plateau. Methods68 We investigated above-ground and below-ground biomass in China's grasslands by conducting five consecutive sampling campaigns across the northern part of the country during 2001–05. We then documented the root:shoot ratio (R/S) and its relationship with climatic factors for China's grasslands. We further explored relationships between above-ground and below-ground biomass across different grassland types. Results68 Our results indicated that the overall R/S of China's grasslands was larger than the global average (6.3 vs. 3.7). The R/S for China's grasslands did not show any significant trend with either mean annual temperature or mean annual precipitation. Above-ground biomass was nearly proportional to below-ground biomass with a scaling exponent (the slope of log–log linear relationship between above-ground and below-ground biomass) of 1.02 across various grassland types. The slope did not differ significantly between temperate and alpine grasslands or between steppe and meadow. Main conclusions68 Our findings support the isometric theory of above-ground and below-ground biomass partitioning, and suggest that above-ground biomass scales isometrically with below-ground biomass at the community level. |
| [40] | . Abstract Climate warming is likely to accelerate the decomposition of soil organic carbon (SOC) which may lead to an increase of carbon release from soils, and thus provide a positive feedback to climate change. However, SOC dynamics in grassland ecosystems over the past two decades remains controversial. In this study, we estimated the magnitude of SOC stock in northern China's grasslands using 981 soil profiles surveyed from 327 sites across the northern part of the country during 2001–2005. We also examined the changes of SOC stock by comparing current measurements with historical records of 275 soil profiles derived from China's National Soil Inventory during the 1980s. Our results showed that, SOC stock in the upper 30cm in northern China's grasslands was estimated to be 10.5PgC (1Pg=10 15 g), with an average density (carbon stock per area) of 5.3kgCm 612 . SOC density (SOCD) did not show significant association with mean annual temperature, but was positively correlated with mean annual precipitation. SOCD increased with soil moisture and reached a plateau when soil moisture was above 30%. Site-level comparison indicated that grassland SOC stock did not change significantly over the past two decades, with a change of 0.08kgCm 612 , ranging from 610.30 to 0.46kgCm 612 at 95% confidence interval. Transect-scale comparison confirmed that grassland SOC stock remained relatively constant from 1980s to 2000s, suggesting that soils in northern China's grasslands have been carbon neutral over the last 20 years. |
| [41] | . Anthropogenic acid deposition may lead to soil acidification, with soil buffering capacity regulating the magnitude of any soil pH change. However, little evidence is available from large-scale observations. Here, we evaluated changes in soil pH across northern China's grasslands over the last two decades using soil profiles obtained from China's Second National Soil Inventory during the 1980s and a more recent regional soil survey during 2001 2005. A transect from the central-southern Tibetan Plateau to the eastern Inner Mongolian Plateau, where Kriging interpolation provided robust predictions of the spatial distribution of soil pH, was then selected to examine pH changes during the survey period. Our results showed that soil pH in the surface layer had declined significantly over the last two decades, with an overall decrease of 0.63 units (95% confidence interval = 0.54 0.73 units). The decline of soil pH was observed in both alpine grasslands on the Tibetan Plateau and temperate grasslands on the Inner Mongolian Plateau. Soil pH decreased more intensively in low soil carbonate regions, while changes of soil pH showed no significant associations with soil cation exchange capacity. These results suggest that grassland soils across northern China have experienced significant acidification from the 1980s to 2000s, with soil carbonates buffering the increase in soil acidity. The buffering process may induce a large loss of carbon from soil carbonates and thus alter the carbon balance in these globally important ecosystems. |
| [42] | . Aims “Grain for Green Program” (GGP), i.e., re-conversion of cropland into forest or grassland, initiated by Chinese government has a profound impact on mitigating environmental degradation. The objectives of this study were to assess the changes of some soil properties during the processes of re-conversion from cropland to grassland over time in the semiarid steppe region of north China. Methods Two sites with different ages of re-conversion were selected for measurements of organic matter (SOM), total nitrogen (TN) and phosphorus (TP), bulk density (BD) and grain size distribution. Saturated hydraulic conductivity was determined by the constant hydraulic head method and unsaturated hydraulic conductivity by disc infiltrometer at tensions of 30, 60 and 15002mm. Soil water content was measured using the gravimetric method. Wetting front depths in the soil after rainfall were also recorded at the study sites. Results Natural grasslands had higher belowground biomass than re-converted grasslands. Re-converted grasslands had lower SOM and TN at depths of 0–2002cm and higher saturated hydraulic conductivity at depths of 0–1002cm than natural grassland. The natural grassland soils had higher soil water contents in the surface soil (0–2002cm) and lower soil water contents at deeper depths than re-converted grassland soils. Soil aggregate stability reached the natural steppe level 1202years after re-conversion. Conclusions The recovery of soil properties after GGP appeared to be slow, and these properties did not return to natural grassland status before cultivation after 1202years of re-conversion. |
| [43] | . . |
1
2009
... 由于团聚体的形成对土壤有机碳具有物理保护作用, 土壤团聚体的稳定性在一定程度上决定了土壤有机碳的稳定性(
Long-term effects of land use on soil aggregate stability
1
2011
... 土壤团聚体是通过结合体(如细根、微生物菌丝、糖分及黏粒、粉粒等)将土壤颗粒黏结在一起而形成的(
Using landscape and depth gradients to decouple the impact of correlated environmental variables on soil microbial community composition
2
2007
... 草原是受人类活动影响最为严重的区域, 因此草原的碳储蓄能力及其动态变化在全球碳循环中扮演着重要角色(
... 土壤团聚体的MMD和GMD是反映土壤团聚体稳定性的重要指标(
Soil aggregate stability: A review
1
1999
... 到目前为止, 测定土壤团聚体稳定性的方法有很多, 包括了水稳性团聚体质量占比、团聚体形成指数、团聚体破坏指数、标准化稳定性指数、稳定性团聚体和稳定性大团聚体指数、平均质量直径(MMD)和几何平均直径(GMD)等(
Improving species distribution models for climate change studies: Variable selection and scale
1
2011
... 降水和气温是影响各个生态系统过程的重要因子, 包括物种分布, 生态系统碳、水及养分循环等(
Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau
2
2008
... 草原是受人类活动影响最为严重的区域, 因此草原的碳储蓄能力及其动态变化在全球碳循环中扮演着重要角色(
... 本研究区域主要集中在蒙古高原上内蒙古境内的天然草原.研究样地位于107.84°-120.06° E, 39.15°-49.62° N, 海拔564-1 546 m, 由东北向西南, 内蒙古草原的年平均气温逐渐增加, 年降水量逐渐减少, 干旱程度逐渐加剧(
Spatial heterogeneity of aggregate stability and soil carbon in semi- arid rangeland
2
2002
... 由于团聚体的形成对土壤有机碳具有物理保护作用, 土壤团聚体的稳定性在一定程度上决定了土壤有机碳的稳定性(
... 土壤团聚体是土壤有机碳的重要物理保护方式, 其稳定性与土壤有机碳稳定性密切相关(
Multi-scale variability in soil aggregate stability: Implications for understanding and predicting semi-arid grassland degradation
2
2007
... 由于团聚体的形成对土壤有机碳具有物理保护作用, 土壤团聚体的稳定性在一定程度上决定了土壤有机碳的稳定性(
... 土壤团聚体的MMD和GMD是反映土壤团聚体稳定性的重要指标(
Surface and subsurface microbial biomass, community structure and metabolic activity as a function of soil depth and season
1
2002
... 土壤团聚体是通过结合体(如细根、微生物菌丝、糖分及黏粒、粉粒等)将土壤颗粒黏结在一起而形成的(
Arbuscular mycorrhizas, microbial communities, nutrient availability, and soil aggregates in organic tomato production
1
2006
... 土壤团聚体是土壤的重要组成部分, 对维持土壤结构的稳定和土壤养分截留具有重要意义(
松嫩草地不同退化阶段的土壤团聚体稳定性
1
2017
... 土壤团聚体的MMD和GMD是反映土壤团聚体稳定性的重要指标(
Organic matter influence on clay wettability and soil aggregate stability
1
2000
... 由于团聚体的形成对土壤有机碳具有物理保护作用, 土壤团聚体的稳定性在一定程度上决定了土壤有机碳的稳定性(
Microaggregate-associated carbon as a diagnostic fraction for management-induced changes in soil organic carbon in two oxisols
2
2007
... 土壤团聚体是土壤的重要组成部分, 对维持土壤结构的稳定和土壤养分截留具有重要意义(
... 土壤团聚体是土壤有机碳的重要物理保护方式, 其稳定性与土壤有机碳稳定性密切相关(
Soil aggregate formation and the accrual of particulate and mineral-associated organic matter
1
1996
... 土壤团聚体是土壤的重要组成部分, 对维持土壤结构的稳定和土壤养分截留具有重要意义(
The vertical distribution of soil organic carbon and its relation to climate and vegetation
1
2000
... 降水和气温是影响各个生态系统过程的重要因子, 包括物种分布, 生态系统碳、水及养分循环等(
1
1965
... 土壤团聚体的MMD和GMD是反映土壤团聚体稳定性的重要指标(
Physical management of soils of the tropics: Priorities for the 21st Century
1
2000
... 土壤团聚体是土壤的重要组成部分, 对维持土壤结构的稳定和土壤养分截留具有重要意义(
Freezing effects on aggregate stability affected by texture, mineralogy, and organic matter
1
1991
... 由于团聚体的形成对土壤有机碳具有物理保护作用, 土壤团聚体的稳定性在一定程度上决定了土壤有机碳的稳定性(
中国草原植被的一般特征
1
1979
... 土壤团聚体的MMD和GMD是反映土壤团聚体稳定性的重要指标(
缙云山不同土地利用方式土壤有机碳组分特征
1
2015
... 由于团聚体的形成对土壤有机碳具有物理保护作用, 土壤团聚体的稳定性在一定程度上决定了土壤有机碳的稳定性(
Soil microbial biomass C and N along a climatic transect in the Mongolian steppe
2
2004
... 草原是受人类活动影响最为严重的区域, 因此草原的碳储蓄能力及其动态变化在全球碳循环中扮演着重要角色(
... 土壤团聚体是通过结合体(如细根、微生物菌丝、糖分及黏粒、粉粒等)将土壤颗粒黏结在一起而形成的(
不同施肥处理对土壤水稳定性团聚体及有机碳分布的影响
1
2010
... 由于团聚体的形成对土壤有机碳具有物理保护作用, 土壤团聚体的稳定性在一定程度上决定了土壤有机碳的稳定性(
Modeled interactive effects of precipitation, temperature, and [CO2] on ecosystem carbon and water dynamics in different climatic zones
1
2008
... 降水和气温是影响各个生态系统过程的重要因子, 包括物种分布, 生态系统碳、水及养分循环等(
内蒙古温带草地的碳储量
3
2006
... 草原是受人类活动影响最为严重的区域, 因此草原的碳储蓄能力及其动态变化在全球碳循环中扮演着重要角色(
... 本研究区域主要集中在蒙古高原上内蒙古境内的天然草原.研究样地位于107.84°-120.06° E, 39.15°-49.62° N, 海拔564-1 546 m, 由东北向西南, 内蒙古草原的年平均气温逐渐增加, 年降水量逐渐减少, 干旱程度逐渐加剧(
... GSP, 生长季降水量; GST, 生长季年平均气温.1), 数据来自
Environmental factors covary with plant diversity-productivity relationships among Chinese grassland sites
2
2010
... 本研究区域主要集中在蒙古高原上内蒙古境内的天然草原.研究样地位于107.84°-120.06° E, 39.15°-49.62° N, 海拔564-1 546 m, 由东北向西南, 内蒙古草原的年平均气温逐渐增加, 年降水量逐渐减少, 干旱程度逐渐加剧(
... GSP, 生长季降水量; GST, 生长季年平均气温.1), 数据来自
Above- and belowground biomass in relation to environmental factors in temperate grasslands, Inner Mongolia
2
2008
... 草原是受人类活动影响最为严重的区域, 因此草原的碳储蓄能力及其动态变化在全球碳循环中扮演着重要角色(
... 土壤团聚体是通过结合体(如细根、微生物菌丝、糖分及黏粒、粉粒等)将土壤颗粒黏结在一起而形成的(
Aggregate-size stability distribution and soil stability
1
2004
... 到目前为止, 测定土壤团聚体稳定性的方法有很多, 包括了水稳性团聚体质量占比、团聚体形成指数、团聚体破坏指数、标准化稳定性指数、稳定性团聚体和稳定性大团聚体指数、平均质量直径(MMD)和几何平均直径(GMD)等(
2
2002
... 到目前为止, 测定土壤团聚体稳定性的方法有很多, 包括了水稳性团聚体质量占比、团聚体形成指数、团聚体破坏指数、标准化稳定性指数、稳定性团聚体和稳定性大团聚体指数、平均质量直径(MMD)和几何平均直径(GMD)等(
... 作为反映土壤团聚体状况的指标, 其值越大表示土壤的团聚度越高, 团聚体稳定性越强(
Aggregate distribution and soil organic matter under different tillage systems for vegetable crops in a red latosol from brazil
1
2004
... 土壤团聚体的MMD和GMD是反映土壤团聚体稳定性的重要指标(
A comparative study of dissolved organic carbon transport and stabilization in california forest and grassland soils
1
2008
... 草原是受人类活动影响最为严重的区域, 因此草原的碳储蓄能力及其动态变化在全球碳循环中扮演着重要角色(
The global carbon sink: A grassland perspective
1
1998
... 草原是受人类活动影响最为严重的区域, 因此草原的碳储蓄能力及其动态变化在全球碳循环中扮演着重要角色(
Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils
1
2002a
... 土壤团聚体是土壤的重要组成部分, 对维持土壤结构的稳定和土壤养分截留具有重要意义(
Aggregation and soil organic matter accumulation in cultivated and native grassland soils
2
1998
... 土壤团聚体是土壤的重要组成部分, 对维持土壤结构的稳定和土壤养分截留具有重要意义(
... 所有土壤样品带回实验室风干后, 过8 mm筛, 以去除根系及石块.我们采用湿筛法进行团聚体分级(
Soil organic matter, biota and aggregation in temperate and tropical soils-effects of no-tillage
2
2002b
... 土壤团聚体是土壤的重要组成部分, 对维持土壤结构的稳定和土壤养分截留具有重要意义(
... 土壤团聚体是通过结合体(如细根、微生物菌丝、糖分及黏粒、粉粒等)将土壤颗粒黏结在一起而形成的(
耕作方式和秸秆还田对华北地区农田土壤水稳性团聚体分布及稳定性的影响
1
2013
... 由于团聚体的形成对土壤有机碳具有物理保护作用, 土壤团聚体的稳定性在一定程度上决定了土壤有机碳的稳定性(
Aridity threshold in controlling ecosystem nitrogen cycling in arid and semi-arid grasslands
1
2014
... 本研究区域主要集中在蒙古高原上内蒙古境内的天然草原.研究样地位于107.84°-120.06° E, 39.15°-49.62° N, 海拔564-1 546 m, 由东北向西南, 内蒙古草原的年平均气温逐渐增加, 年降水量逐渐减少, 干旱程度逐渐加剧(
Responses of terrestrial ecosystems to temperature and precipitation change: A meta-analysis of experimental manipulation
1
2011
... 降水和气温是影响各个生态系统过程的重要因子, 包括物种分布, 生态系统碳、水及养分循环等(
Stoichiometric shifts in surface soils over broad geographical scales: Evidence from China’s grasslands
1
2014
... GSP, 生长季降水量; GST, 生长季年平均气温.1), 数据来自
Large-scale pattern of biomass partitioning across China’s grasslands
3
2010a
... 草原是受人类活动影响最为严重的区域, 因此草原的碳储蓄能力及其动态变化在全球碳循环中扮演着重要角色(
... 降水和气温是影响各个生态系统过程的重要因子, 包括物种分布, 生态系统碳、水及养分循环等(
... ), 表明在内蒙古草原生态系统, 降水量高能促进土壤团聚体的形成, 气温高则不利于土壤团聚体的形成.这可能是因为降水量高, 导致地上、地下生物量也高(
Soil carbon stock and its changes in northern China’s grasslands from 1980s to 2000s
1
2010b
... 降水和气温是影响各个生态系统过程的重要因子, 包括物种分布, 生态系统碳、水及养分循环等(
Significant soil acidification across northern china's grasslands during 1980s-2000s
1
2012
... GSP, 生长季降水量; GST, 生长季年平均气温.1), 数据来自
Changes in some soil properties induced by re-conversion of cropland into grassland in the semiarid steppe zone of Inner Mongolia, China
2
2013
... 草原是受人类活动影响最为严重的区域, 因此草原的碳储蓄能力及其动态变化在全球碳循环中扮演着重要角色(
... 土壤团聚体是通过结合体(如细根、微生物菌丝、糖分及黏粒、粉粒等)将土壤颗粒黏结在一起而形成的(
子午岭植被恢复过程中土壤团聚体有机碳含量的变化
1
2006
... 土壤团聚体的MMD和GMD是反映土壤团聚体稳定性的重要指标(


{kind=link}
{kind=link}
{kind=link}
{kind=link}