Characteristics of normalized difference vegetation index of biological soil crust during the succession process of artificial sand-fixing vegetation in the Tengger Desert, Northern China
ZHAOYun1,2, JIARong-Liang1,*,, GAOYan-Hong1, ZHOUYuan-Yuan1,2, TENGJia-Ling1,2 1Shapotou Desert Research and Experiment Station, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China2University of Chinese Academy of Sciences, Beijing 100049, China 通讯作者:* 通信作者Author for correspondence (E-mail: rongliangjia@163.com) 收稿日期:2016-05-24 修回日期:2017-08-26 网络出版日期:2017-09-30 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(41371099、41671210和41530746)和中国科学院“西部之光”人才培养引进计划
关键词:归一化植被指数;生物土壤结皮;植被演替;降水;温度 Abstract Aims Biological soil crust (hereafter crust) affects normalized difference vegetation index (NDVI) values in arid desert ecosystems. This study aimed to demonstrate the feasibility of combining crust NDVI values with meteorological data to distinguish the crust successional stage at the regional scale. Meanwhile, the characteristics of crust NDVI could provide the basis for the error analysis of NDVI-based surface ecological parameters estimation in desert ecosystems. We also suggested the optimum periods for crust observation based on the multi-temporal remote sensing images.Methods NDVI values of five types of dominant crusts, three typical sand-fixing shrubs and bare sand were collected by spectrometer in the field. Crusts and shrubs were randomly selected in revegetated areas established in 1956, 1964, and 1973 at Shapotou, which is on the southeastern edge of the Tengger Desert. We used the space-for-time method to study the characteristics of crust NDVI values and their responses to precipitation and temperature during the succession process of artificial sand-fixing vegetation. Additionally, we evaluated the contribution of crust NDVI values to the whole ecosystem NDVI values by comparing the NDVI values of crusts, shrubs and bare sand.Important findings 1) With succession process of the artificial sand-fixing vegetation, the crust NDVI values significantly increased. Among different crust types, we found the following order of NDVI values: Didymodon vinealis crust > Bryum argenteum crust > mixed crust > lichen crust > algae crust. 2) Crust NDVI values were significantly affected by precipitation, temperature and their interaction, and the influences showed significant seasonal differences. Furthermore, we found significantly linear correlations between crust NDVI value and precipitation, and between crust NDVI value and the shallow soil moisture content covered by crust. A significantly negative linear correlation between daily mean temperature and crust NDVI value, and a significantly exponential correlation between the surface temperature of crust and its NDVI value. With the succession process of artificial sand-fixing vegetation, the response of crust NDVI value to precipitation and temperature became more sensitive. In addition, the response of crust NDVI value to temperature was more sensitive in spring than in summer, while that to precipitation was less sensitive in spring than in summer. 3) Moss crust NDVI value was significantly higher than that of shrubs and bare sand after the rainfall event in spring, while shrubs NDVI value was significantly higher than that of crust after the rainfall event in summer. Considering the coverage weights of different ground features in sand-fixing areas, crust NDVI values contributed 90.01% and 82.53% in spring and summer, respectively, to the regional NDVI values, which were higher than those of shrubs (9.99% and 17.47% in spring and in summer, respectively). Additionally, with the succession process of artificial sand-fixing vegetation, crust NDVI values contributed more, while shrubs contributed less to regional NDVI values.
5种结皮NDVI均随固沙植被演替逐渐增大, 且湿结皮NDVI与固沙植被演替时间具有更高的相关性(p < 0.05) (图1, 图2)。同一年代固沙区不同类型结皮NDVI相比, 藓类结皮>混生结皮>地衣结皮> 藻结皮, 其中土生对齿藓结皮NDVI显著高于其他类型结皮(p < 0.05), 真藓结皮NDVI显著高于地衣结皮和藻结皮(p < 0.05), 混生结皮NDVI与地衣结皮NDVI差异不显著。在1965年固沙区混生结皮NDVI与真藓结皮NDVI差异显著(p < 0.05), 而在1964年及1973年固沙区两者差异不显著。此外, 干湿结皮差异显著(p < 0.05) (图2)。 显示原图|下载原图ZIP|生成PPT 图1腾格里沙漠人工固沙植被演替过程中不同地物归一化植被指数(NDVI)的季节变化。1973(42)、1964(51)、1956(59)表示固沙植被建立年份(固沙年限(a))。 -->Fig. 1Seasonal changes in normalized difference vegetation index (NDVI) values with the succession process of the artificial sand-fixing vegetation. 1973(42), 1964(51), 1956(59) represent year of revegetation (history of revegetation (a)). -->
显示原图|下载原图ZIP|生成PPT 图2腾格里沙漠人工固沙植被演替过程生物土壤结皮归一化植被指数(NDVI)的变化特征(平均值+标准偏差)。A, 干结皮。B, 湿结皮。 -->Fig. 2Changes in normalized difference vegetation index (NDVI) values of biological soil crust with the succession process of artificial sand-fixing vegetation (mean + SD). A, Dry crust. B, Wet crust. -->
2.2 腾格里沙漠人工固沙植被演替过程中生物土壤结皮NDVI对降水和温度的响应
结皮NDVI受降水量、气温及两者间的交互作用影响显著且具有明显的季节差异(p < 0.05) (表1, 表2)。结皮NDVI与降水量及其覆盖土壤浅层含水量均呈显著线性正相关关系(p < 0.01), 并且结皮NDVI对水分的敏感性随固沙植被演替而逐渐增加(图3)。结皮NDVI总体与日平均气温呈显著线性负相关关系(p < 0.05)而与结皮表面温度呈显著指数负相关关系(p < 0.05), 并且结皮NDVI对温度的敏感性随固沙植被演替逐渐增加(图4)。结皮NDVI对温度变化的敏感性春季高于夏季, 对水分变化的敏感性夏季高于春季(表2), 逐步回归分析表明结皮覆盖土壤浅层含水量是影响藓类结皮NDVI的最关键因子, 降水是影响藻、地衣和混生结皮NDVI的最关键因子(表1)。 Table 1 表1 表1生物土壤结皮归一化植被指数(NDVI)与固沙植被演替年龄、降水量、气温、结皮表面温度及结皮覆盖土壤浅层含水量的逐步回归方程 Table 1Stepwise regressions of the sand-fixing vegetation successional age, precipitation, daily mean temperature, surface temperature and shallow soil moisture content covered by biological soil crust with their normalized difference vegetation index (NDVI) values
A, sand-fixing vegetation successional age; M, shallow soil moisture content covered by biological soil crust; P, precipitation; ST, surface temperature of biological soil crust; T, daily mean temperature.A, 固沙植被演替年龄; M, 结皮覆盖土壤浅层含水量; P, 降水量; ST, 结皮表面温度; T, 日平均气温。 新窗口打开 Table 2 表2 表2生物土壤结皮归一化植被指数(NDVI)与水分及温度的偏相关系数季节变化 Table 2The partial correlation coefficient of normalized difference vegetation index (NDVI) values of biological soil crust in spring and summer
春季 Spring
夏季 Summer
结皮表面温度 Crust surface temperature
-0.269**
-0.139**
土壤体积含水量 Soil volumetric moisture content
0.146
0.473**
日平均气温 Daily mean temperature
-0.321**
-0.069
降水量 Precipitation
0.388**
0.629**
** represent significant level p < 0.01.**表示回归显著水平p < 0.01。 新窗口打开 显示原图|下载原图ZIP|生成PPT 图3腾格里沙漠人工固沙植被演替过程中生物土壤结皮归一化植被指数(NDVI)对水分的响应。A, F, 藻结皮。B, G, 地衣结皮。C, H, 混生结皮。D, I 真藓结皮。E, J 土生对齿藓结皮。降水量为观测前一日至观测时的累积降水量。1973(42)、1964(51)、1956(59)表示固沙植被建立年份(固沙年限(a))。 -->Fig. 3Responses of normalized difference vegetation index (NDVI) values of biological soil crust to precipitation and shallow soil moisture content covered by biological soil crust during the succession process of artificial sand-fixing vegetation. A, F, Algae crust. B, G, Lichen crust. C, H, Mixed crust. D, I, Bryum argenteum crust. E, J, Didymodon vinealis crust. Precipitation means the cumulative precipitation during the previous 24 hours before the observation time. 1973(42), 1964(51), 1956(59) represent year of revegetation (history of revegetation (a)). -->
2.3 腾格里沙漠人工固沙植被演替过程中生物土壤结皮及灌木对固沙区系统NDVI的贡献率
春季降水后, 藓类结皮NDVI显著高于油蒿、花棒、柠条等灌木及裸沙(p < 0.05); 夏季降水后, 结皮NDVI显著低于灌木(p < 0.05)。若考虑结皮较高的盖度, 春、夏季结皮NDVI对固沙区系统NDVI的贡献率分别为(90.01±2.16)%和(82.53±3.17)%, 均超过灌木((9.99±2.16)%和(17.47±3.17)%), 并且结皮对固沙区系统NDVI的贡献率随着固沙植被演替逐渐增加, 而灌木的贡献率则逐渐降低(图5)。 显示原图|下载原图ZIP|生成PPT 图4腾格里沙漠人工固沙植被演替过程中生物土壤结皮归一化植被指数(NDVI)对温度的响应。A, F, 藻结皮。B, G , 地衣结皮。C, H, 混生结皮。D, I 真藓结皮。E, J 土生对齿藓结皮。1973(42)、1964(51)、1956(59)表示固沙植被建立年份(固沙年限(a))。 -->Fig. 4Responses of normalized difference vegetation index (NDVI) values of biological soil crust to daily mean temperature and surface temperature during the succession process of artificial sand-fixing vegetation. A, F, Algae crust. B, G , Lichen crust. C, H, Mixed crust. D, I, Bryum argenteum crust. E, J, Didymodon vinealis crust. 1973(42), 1964(51), 1956(59) represent year of revegetation (history of revegetation (a)). -->
显示原图|下载原图ZIP|生成PPT 图5腾格里沙漠人工固沙植被演替过程中生物土壤结皮及灌木对固沙区系统归一化植被指数(NDVI)的贡献率。 -->Fig. 5Changes in the contribution rate of normalized difference vegetation index (NDVI) values of biological soil crust and shrubs to the regional NDVI of sand-fixing system with the succession process of artificial sand-fixing vegetation. -->
BaiXL, WangY, XuJ, LiXR, ZhangJG (2003). Characteristics of reproduction and growth of mosses in the soil crust of fixed dunes in Shapotou area. Journal of Desert Research, 23, 171-173. (in Chinese with English abstract)[白学良, 王瑶, 徐杰, 李新荣, 张景光 (2003). 沙坡头地区固定沙丘结皮层藓类植物的繁殖和生长特性研究 . 中国沙漠, 23, 171-173.] [本文引用: 1]
[2]
ChamizoS, StevensA, CantónY, MirallesI, DomingoF, van WesemaelB (2012). Discriminating soil crust type, development stage and degree of disturbance in semiarid environments from their spectral characteristics. European Journal of Soil Science, 63, 42-53. [本文引用: 2]
[3]
ChengHJ, ZhangYM (2010). Environmental factors affecting soil biocrust distribution. Chinese Journal of Ecology, 29, 133-141. (in Chinese with English abstract)[程军回, 张元明 (2010). 影响生物土壤结皮分布的环境因子 . 生态学杂志, 29, 133-141.] [本文引用: 2]
[4]
ChenJ, ZhangMY, WangL, ShimazakiH, TamuraM (2005). A new index for mapping lichen-dominated biological soil crusts in desert areas. Remote Sensing of Environment, 96, 165-175. [本文引用: 1]
[5]
ColesieC, ScheuS, GreenTGA, WeberB, WirthR, BüdelB (2012). The advantage of growing on moss: Facilitative effects on photosynthetic performance and growth in the cyanobacterial lichen Peltigera rufescens. Oecologia, 169, 599-607. [本文引用: 1]
[6]
Dettweiler-RobinsonE, PonzettiJM, BakkerJD (2013). Long-term changes in biological soil crust cover and composition. Ecological Processes, 2, 1-10. [本文引用: 1]
[7]
DodyA, HakmonR, AsafB, ZaadyE (2011). Indices to monitor biological soil crust growth rate—Lab and field experiments. Natural Science, 3, 478-483. [本文引用: 1]
[8]
EldridgeDJ, TozerME (1997). Environmental factors relating to the distribution of terricolous bryophytes and lichens in semi-arid eastern Australia. Bryologist, 100, 28-39. [本文引用: 1]
[9]
FangSB, YuWG, QiY (2015). Spectra and vegetation index variations in moss soil crust in different seasons, and in wet and dry conditions. International Journal of Applied Earth Observation & Geoinformation, 38, 261-266. [本文引用: 3]
[10]
FangSB, ZhangXS (2011). Impact of moss soil crust on vegetation indexes interpretation. Spectroscopy and Spectral Analysis, 31, 780-783. (in Chinese with English abstract)[房世波, 张新时 (2011). 苔藓结皮影响干旱半干旱植被指数的稳定性 . 光谱学与光谱分析, 31, 780-783.] [本文引用: 2]
[11]
FengL, ZhangJG, ZhangZS, GuoQ, LiXR (2009). Growth and biomass allocation dynamics of Artemisia ordosica in sand-fixing vegetation of the Tengger Desert of China. Journal of Plant Ecology (Chinese Version), 33, 1132-1139. (in Chinese with English abstract)[冯丽, 张景光, 张志山, 郭群, 李新荣 (2009). 腾格里沙漠人工固沙植被中油蒿的生长及生物量分配动态 . 植物生态学报, 33, 1132-1139.] [本文引用: 1]
[12]
FengW (2014). Photosynthetic Carbon Fixation of Biological Soil Crusts in Mu Us Desert and Their Impact on Soil Carbon Emission. PhD dissertation, Beijing Forestry University, Beijing. 61-73. (in Chinese with English abstract)[冯薇 (2014). 毛乌素沙地生物结皮光合固碳过程及对土壤碳排放的影响 . 博士学位论文, 北京林业大学, 北京. 61-73.] [本文引用: 1]
[13]
FengXR, BuCF, HaoHK, YangYZ, ZhangGJ (2015). Research on biological soil crust extraction by spectral analysis in Mu Us Desert, China. Journal of Natural Resources, 30, 1024-1034. (in Chinese with English abstract)[冯秀绒, 卜崇峰, 郝红科, 杨延征, 张广军 (2015). 基于光谱分析的生物结皮提取研究——以毛乌素沙地为例 . 自然资源学报, 30, 1024-1034.] [本文引用: 2]
[14]
FischerT, VesteM, EiseleA, BensO, SpyraW, HüttlRF (2012). Small scale spatial heterogeneity of normalized difference vegetation indices (NDVIs) and hot spots of photosynthesis in biological soil crusts. Flora, 207, 159-167. [本文引用: 1]
[15]
GroteEE, BelnapJ, HousmanDC, SparksJP (2010). Carbon exchange in biological soil crust communities under differential temperatures and soil water contents: Implications for global change. Global Change Biology, 16, 2763-2774. [本文引用: 1]
[16]
HousmanDC, PowersHH, CollinsAD, BelnapJ (2006). Carbon and nitrogen fixation differ between successional stages of biological soil crusts in the Colorado Plateau and Chihuahuan Desert. Journal of Arid Environments, 66, 620-634. [本文引用: 3]
[17]
JiaRL (2009). Photosynthetic Ecophysiological Characteristics of Moss Crusts in A Revegetated Area of the Tengger Desert, Northern China . PhD dissertation, Cold and Arid Regions Environmental and Engineering Research Institute, Chinese Academy of Sciences, Beijing.(in Chinese with English abstract)[贾荣亮 (2009). 腾格里沙漠人工植被区藓类结皮光合生理生态学研究 . 博士学位论文, 中国科学院寒区旱区环境与工程研究所, 北京.] [本文引用: 4]
[18]
JiaRL, LiXR, LiuLC, GaoYH, LiXJ (2008). Responses of biological soil crusts to sand burial in a revegetated area of the Tengger Desert, Northern China. Soil Biology & Biochemistry, 40, 2827-2834. [本文引用: 2]
[19]
KarnieliA (1997). Development and implementation of spectral crust index over dune sands. International Journal of Remote Sensing, 18, 1207-1220. [本文引用: 1]
[20]
KarnieliA (2003). Natural vegetation phenology assessment by ground spectral measurements in two semi-arid environments. International Journal of Biometeorology, 47, 179-187. [本文引用: 2]
[21]
KarnieliA, KidronGJ, GlaesserC, Ben-DorE (1999). Spectral characteristics of cyanobacteria soil crust in semiarid environments. Remote Sensing of Environment, 69, 67-75. [本文引用: 2]
[22]
KarnieliA, ShachakM, TsoarH, ZaadyE, KaufmanY, DaninA, PorterW (1996) The effect of microphytes on the spectral reflectance of vegetation in semiarid regions. Remote Sensing of Environment, 57, 88-96. [本文引用: 5]
[23]
KleinerEF (1983). Successional trends in an ungrazed, arid grassland over a decade. Journal of Range Management, 36, 114-118. [本文引用: 1]
[24]
LangeOL (2003). Photosynthesis of soil-crust biota as dependent on environmental factors. In: Belnap J, Lange OL eds. Biological Soil Crust: Structure, Function, and Management. Springer, Berlin. 217-240. [本文引用: 3]
[25]
LangeOL, BelnapJ, ReichenbergerH (1998). Photosynthesis of the cyanobacterial soil-crust lichen Collema tenax from arid lands in southern Utah, USA: Role of water content on light and temperature responses of CO2 exchange . Functional Ecology, 12, 195-202. [本文引用: 1]
[26]
LiSZ, ZhengHZ, LiSL, ShenBC (2008). Development characteristics of biotic crusts on Shapotou vegetated sand dunes. Chinese Journal of Ecology, 27, 1675-1679. (in Chinese with English abstract)[李守中, 郑怀舟, 李守丽, 沈宝成 (2008). 沙坡头植被固沙区生物结皮的发育特征 . 生态学杂志, 27, 1675-1679.] [本文引用: 1]
[27]
LiXR (2010).Eco-hydrology of Biological Soil Crusts in Desert Regions of China. Higher Education Pressv, Beijing. (in Chinese)[李新荣 (2010). 荒漠生物土壤结皮生态与水文学研究. 高等教育出版社, 北京.] [本文引用: 3]
[28]
LiXR, HuiR, ZhaoY (2016). Eco-physiology of Biological Soil Crusts in Desert Regions of China. Higher Education Press, Beijing. (in Chinese)[李新荣, 回嵘, 赵洋 (2016). 中国荒漠生物土壤结皮生态生理学研究. 高等教育出版社, 北京.]
[29]
LiXR, JiaRL, ChenYW, HuangL, ZhangP (2011). Association of ant nests with successional stages of biological soil crusts in the Tengger Desert, Northern China. Applied Soil Ecology, 47, 59-66. [本文引用: 1]
[30]
LiXR, ZhangJG, LiuLC, ChenHS, ShiQH (2000). Plant diversity in the process of succession of artificial vegetation type sand environment in an arid desert region of china. Acta Phytoecologica Sinica, 24, 257-261. (in Chinese with English abstract)[李新荣, 张景光, 刘立超, 陈怀顺, 石庆辉 (2000). 我国干旱沙漠地区人工植被与环境演变过程中植物多样性的研究 . 植物生态学报, 24, 257-261.] [本文引用: 1]
[31]
LiXR, ZhangP, SuYG, JiaRL (2012). Carbon fixation by biological soil crusts following revegetation of sand dunes in arid desert regions of China: A four-year field study. Catena, 97, 119-126.
[32]
LiXR, ZhangYM, ZhaoYG (2009). A study of biological soil crusts: Recent development trend and prospect. Advances in Earth Science, 24, 11-24. (in Chinese with English abstract)[李新荣, 张元明, 赵允格 (2009). 生物土壤结皮研究: 进展、前沿与展望 . 地球科学进展, 24, 11-24.] [本文引用: 1]
[33]
LiXR, ZhangZS, TanHJ, GaoYH, LiuLC, WangXP (2014). Ecological restoration and recovery in the wind-blown sand hazard areas of northern China: Relationship between soil water and carrying capacity for vegetation in the Tengger Desert. Science China Life Sciences, 57, 539-548. [本文引用: 2]
[34]
LiXR, ZhaoY, HuiR, SuJQ, GaoYH (2014). Progress and trend of development of restoration ecology research in the arid regions of China. Progress in Geography, 33, 1435-1443. (in Chinese with English abstract)[李新荣, 赵洋, 回嵘, 苏洁琼, 高艳红 (2014). 中国干旱区恢复生态学研究进展及趋势评述 . 地理科学进展, 33, 1435-1443.] [本文引用: 2]
[35]
LiXR, ZhouHY, WangXP, ZhuYG, O’connerPJ (2003). The effects of sand stabilization and revegetation on cryptogam species diversity and soil fertility in the Tengger Desert, Northern China. Plant and Soil, 251, 237-245. [本文引用: 1]
[36]
MaestreFT, EscuderoA, MartinezI, GuerreroC, RubioA (2005). Does spatial pattern matter to ecosystem functioning? Insights from biological soil crusts. Functional Ecology, 19, 566-573. [本文引用: 1]
[37]
PettorelliN, VikJO, MysterudA, GaillardJM, TuckerCJ, StensethNC (2005). Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends in Ecology & Evolution, 20, 503-510. [本文引用: 1]
[38]
Rodriguez-CaballeroE, KnerrT, WeberB (2015). Importance of biocrusts in dryland monitoring using spectral indices. Remote Sensing of Environment, 170, 32-39. [本文引用: 2]
[39]
RozensteinO, KarnieliA (2014). Identification and characterization of biological soil crusts in a sand dune desert environment across Israel-Egypt border using LWIR emittance spectroscopy. Journal of Arid Environments, 112, 75-86. [本文引用: 2]
[40]
SchmidtH, KarnieliA (2002). Analysis of the temporal and spatial vegetation patterns in a semi-arid environment observed by NOAA AVHRR imagery and spectral ground measurements. International Journal of Remote Sensing, 23, 3971-3990.
[41]
SuYG, LiXR, ChenYW, ZhangZS (2010). Photosynthetic characteristic of algal crusts in the desert area. Journal of Lanzhou University: Natural Sciences, 46, 1-5. (in Chinese with English abstract)[苏延桂, 李新荣, 陈应武, 张志山 (2010). 不同演替序列的藻结皮净光合速率日变化特征 . 兰州大学学报: 自然科学版, 46, 1-5.] [本文引用: 3]
[42]
TianGQ, BaiXL, XuJ, ZhangJS (2005). Morphological and structural properties as well as adaptation of mosses in microbiotic soil crusts on fixed dunes. Journal of Desert Research, 25, 249-255. (in Chinese with English abstract)[田桂泉, 白学良, 徐杰, 张建升 (2005). 固定沙丘生物结皮层藓类植物形态结构及其适应性研究 . 中国沙漠, 25, 249-255.] [本文引用: 2]
UstinSL, ValkoPG, KefauverSC, SantosMJ, ZimpferJF, SmithSD (2009). Remote sensing of biological soil crust under simulated climate change manipulations in the Mojave Desert. Remote Sensing of Environment, 113, 317-328. [本文引用: 1]
[45]
WangXQ, ZhangYM, WangYC, WanJP, XuM (2006). Eco-environment change of biological crusts on longitudinal dune surface in Gurbantunggut Desert. Journal of Desert Research, 26, 711-716. (in Chinese with English abstract)[王雪芹, 张元明, 王远超, 万金平, 徐曼 (2006). 古尔班通古特沙漠生物结皮小尺度分异的环境特征 . 中国沙漠, 26, 711-716.] [本文引用: 2]
[46]
WangXQ, ZhangYM, ZhangWM, YangDL (2011). The aerodynamic roughness length of biological soil crusts: A case study of Gurbantunggut Desert. Acta Ecologica Sinica, 31, 4153-4160. (in Chinese with English abstract)[王雪芹, 张元明, 张伟民, 杨东亮 (2011). 生物结皮粗糙特征——以古尔班通古特沙漠为例 . 生态学报, 31, 4153-4160.] [本文引用: 2]
[47]
WangY, ZhaoYG, YaoCZ, ZhangPP (2014). Surface roughness characteristics of biological soil crusts and its influencing factors in the Hilly Loess Plateau region, China. Chinese Journal of Applied Ecology, 25, 647-656. (in Chinese with English abstract)[王媛, 赵允格, 姚春竹, 张培培 (2014). 黄土丘陵区生物土壤结皮表面糙度特征及影响因素 . 应用生态学报, 25, 647-656.] [本文引用: 3]
[48]
WeberB, HillJ (2016). Remote sensing of biological soil crusts at different scales. In: Weber B, Büdel B, Belnap J eds. Biological Soil Crusts: An Organizing Principle in Drylands. Springer, Cham,Switzerland. 215-232. [本文引用: 3]
[49]
WeberB, OlehowskiC, KnerrT, HillJ, DeutschewitzK, WesselsDCJ, EitelB, BüdelB (2008). A new approach for mapping of biological soil crusts in semidesert areas with hyperspectral imagery. Remote Sensing of Environment, 112, 2187-2201. [本文引用: 1]
[50]
WuL (2012). On the Formation Process of Lichen Soil Crust and Their Photosynthetic Physiology. PhD dissertation, Institute of Hydrobiology, Chinese Academy of Sciences, Beijing. 23-38. (in Chinese with English abstract)[吴丽 (2012). 地衣结皮形成的生物学过程及其光合生理研究 . 博士学位论文, 中国科学院水生生物研究所, 北京. 23-38.] [本文引用: 1]
[51]
XuJ, BaiXL, TianGQ, HuangJ, ZhangD, FengXH (2005). Ecological function of mosses in biotic crusts on fixed dunes on Tengger Desert and its relation with soil factors. Journal of Desert Research, 25, 234-242. (in Chinese with English abstract)[徐杰, 白学良, 田桂泉, 黄洁, 张镝, 冯晓慧 (2005). 腾格里沙漠固定沙丘结皮层藓类植物的生态功能及与土壤环境因子的关系 . 中国沙漠, 25, 234-242.] [本文引用: 6]
[52]
YaoDL, LiJC, DuY, LiXR, ZhangJG (2002). A land-atmosphere coupling model and mechanism of the crust layer and evolution of canopy in artificial vegetation area of Shapoto. Acta Ecologica Sinica, 22, 452-460. (in Chinese with English abstract)[姚德良, 李家春, 杜岳, 李新荣, 张景光 (2002). 沙坡头人工植被区陆气耦合模式及生物结皮与植被演变的机理研究 . 生态学报, 22, 452-460.] [本文引用: 1]
[53]
ZaadyE, BouskilaA (2002). Lizard burrows association with successional stage of biological soil crusts in an arid sandy region. Journal of Arid Environments, 50, 235-246. [本文引用: 1]
[54]
ZaadyE, KarnieliA, ShachakM (2007). Applying a field spectroscopy technique for assessing successional trends of biological soil crusts in a semi-arid environment. Journal of Arid Environments, 70, 463-477. [本文引用: 1]
[55]
ZaadyE, KuhnU, WilskeB, Sandoval-SotoL, KesselmeierJ (2000). Patterns of CO2 exchange in biological soil crusts of successional age. Soil Biology & Biochemistry, 32, 959-966. [本文引用: 3]
[56]
ZhangJ, ZhangYM (2014). Diurnal variations of chlorophyll fluorescence and CO2 exchange of biological soil crusts in different successional stages in the Gurbantunggut Desert of northwestern China. Ecological Research, 29, 289-298.
[57]
ZhangYM, PanHX, PanBR (2004). Distribution characteristics of biological crust on sand dune surface in Gurbantunggut Desert, Xinjiang. Journal of Soil and Water Conservation, 18(4), 61-64. (in Chinese with English abstract)[张元明, 潘惠霞, 潘伯荣 (2004). 古尔班通古特沙漠不同地貌部位生物结皮的选择性分布 . 水土保持学报, 18(4), 61-64.] [本文引用: 1]
[58]
ZhaoY, JiaRL, TengJL, JiaWX, GaoYH (2017). Response of biological soil crust coverage to wind-blown sand burial during the succession of the artificially sand-fixing vegetation in the Tengger Desert, Northern China. Acta Ecologica Sinica, 37, 6138-6148. (in Chinese with English abstract)[赵芸, 贾荣亮, 滕嘉玲, 贾文雄, 高艳红 (2017). 腾格里沙漠人工固沙植被演替生物土壤结皮盖度对沙埋的响应 . 生态学报, 37, 6138-6148.] [本文引用: 1]
[59]
ZhaoYG, XuMX, BelnapJ (2010). Response of biocrusts’ photosynthesis to environmental factors: A possible explanation of the spatial distribution of biocrusts in the Hilly Loess Plateau region of China. Acta Ecologica Sinica, 17, 4668-4675. (in Chinese with English abstract)[赵允格, 许明祥, JayneBelnap (2010). 生物结皮光合作用对光温水的响应及其对结皮空间分布格局的解译——以黄土丘陵区为例 . 生态学报, 17, 4668-4675.] [本文引用: 2]
[60]
ZhaoYS, ChenDM, YangLM, ZhouXT, LiXM, TangWZ (2003). Principle and Method of Analysis of Remote Sensing Application. Science Press, Beijing. 366-409. (in Chinese)[赵英时, 陈冬梅, 杨立明, 周心铁, 李小文, 唐文周 (2003). 遥感应用分析原理与方法. 科学出版社, 北京. 366-409.] [本文引用: 1]
Small scale spatial heterogeneity of normalized difference vegetation indices (NDVIs) and hot spots of photosynthesis in biological soil crusts. 1 2012
Photosynthesis of the cyanobacterial soil-crust lichen Collema tenax from arid lands in southern Utah, USA: Role of water content on light and temperature responses of CO2 exchange 1 1998
Carbon fixation by biological soil crusts following revegetation of sand dunes in arid desert regions of China: A four-year field study. 2012
生物土壤结皮研究: 进展、前沿与展望 1 2009
... 生物土壤结皮(简称结皮)是干旱沙区重要的地表生物覆盖物, 在沙区土壤生态、水文和地球化学循环等过程中发挥着重要作用, 其发育及演替特征常被作为重要指标来衡量生态系统稳定性和健康程度(李新荣等, 2009, 2014; 李新荣, 2010).因此, 在区域尺度上, 结皮的时空动态对于沙区生态系统管理具有重要的参考价值(李新荣等, 2014; Weber & Hill, 2016).虽然基于实验室测量或地块试验, 针对结皮的空间分布特点(张元明等, 2004; Maestre et al., 2005; 李新荣等, 2016)、拓殖和演替规律(Kleiner, 1983; Eldridge & Tozer, 1997; Zaady & Bouskila, 2002; Dettweiler-Robinson et al., 2013; 李新荣等, 2016)和碳储量估算(贾荣亮, 2009; Grote et al., 2010; 冯薇, 2014)等已有较多报道, 但由于结皮生物自身变水特性且易受到植被、土壤和地形等因素的影响, 其时空分异明显(王雪芹等, 2006; 程军回和张元明, 2010; 赵允格等, 2010), 使得上述研究结果往往难以准确推广到区域尺度.而遥感技术在较大尺度研究方面具有明显的技术优势, 为大尺度结皮时空格局监测、评估其碳储量、固碳/氮潜力和其他生态功能提供了重要机遇(Weber & Hill, 2016). ...
Ecological restoration and recovery in the wind-blown sand hazard areas of northern China: Relationship between soil water and carrying capacity for vegetation in the Tengger Desert. 2 2014
... 本试验在中国科学院沙坡头沙漠研究试验站包兰铁路北侧的人工固沙植被区内(37.45° N, 104.95° E, 海拔1 330 m)进行.该区年平均气温10.0 ℃, 低温极值-25.1 ℃, 高温极值38.1 ℃, 年降水量186.2 mm (1956-2012年), 80%的降水集中在5-9月, 5-7月的平均温度为24 ℃, 年潜在蒸发量3 000 mm, 主风向为西北风, 年平均风速2.9 m·s-1, 属于温带荒漠化草原向草原化荒漠的过渡区.该区主要的景观类型是高大密集的格状新型沙丘链, 沙丘由西北向东南倾斜, 呈阶梯状分布, 沙丘间零星散布有沙蓬(Agriophyllum squarrosum)、百花蒿(Stilpnolepis centiflora)等一年生草本植物(Li et al., 2014). ...
Identification and characterization of biological soil crusts in a sand dune desert environment across Israel-Egypt border using LWIR emittance spectroscopy. 2 2014
... 自20世纪90年代开始, 国内外一些****基于对结皮光谱特征的分析研究建立了在遥感影像上提取结皮覆盖度的结皮指数(Karnieli, 1997; Chen et al., 2005; Weber et al., 2008; Rozenstein & Karnieli, 2014), 其中有****认为结合NDVI和结皮指数能够提高结皮覆盖度的提取精度(Rozenstein & Karnieli, 2014; 冯秀绒等, 2015).也有****尝试利用NDVI在遥感影像上区分结皮与维管植物(Karnieli, 2003).但是, 同样由于结皮生物的变水特性及其有别于维管植物的物候节律造成其NDVI的水分变异性和季节变异性(Karnieli et al., 1996; Karnieli, 2003; Fang et al., 2015)较大, 而且不同演替阶段结皮在形态结构和光合生理特征方面差异较大(Zaady et al., 2000; 徐杰等, 2005; Housman et al., 2006; 贾荣亮, 2009; 苏延桂等, 2010; 王媛等, 2014; Zhang & Zhang; 2014), 且其分布随地形变化差异较大(王雪芹等, 2006; 程军回和张元明, 2010; 赵允格等, 2010), 引起干旱半干旱地区NDVI的不稳定性, 增加了遥感图像解译的困难而降低了以NDVI为基本参量进行植被生态参数的遥感估算准确性和精度(Karnieli et al., 1996; 房世波和张新时, 2011; Fang et al., 2015; Rodriguez- Caballero et al., 2015). ...
... 和结皮指数能够提高结皮覆盖度的提取精度(Rozenstein & Karnieli, 2014; 冯秀绒等, 2015).也有****尝试利用NDVI在遥感影像上区分结皮与维管植物(Karnieli, 2003).但是, 同样由于结皮生物的变水特性及其有别于维管植物的物候节律造成其NDVI的水分变异性和季节变异性(Karnieli et al., 1996; Karnieli, 2003; Fang et al., 2015)较大, 而且不同演替阶段结皮在形态结构和光合生理特征方面差异较大(Zaady et al., 2000; 徐杰等, 2005; Housman et al., 2006; 贾荣亮, 2009; 苏延桂等, 2010; 王媛等, 2014; Zhang & Zhang; 2014), 且其分布随地形变化差异较大(王雪芹等, 2006; 程军回和张元明, 2010; 赵允格等, 2010), 引起干旱半干旱地区NDVI的不稳定性, 增加了遥感图像解译的困难而降低了以NDVI为基本参量进行植被生态参数的遥感估算准确性和精度(Karnieli et al., 1996; 房世波和张新时, 2011; Fang et al., 2015; Rodriguez- Caballero et al., 2015). ...
Analysis of the temporal and spatial vegetation patterns in a semi-arid environment observed by NOAA AVHRR imagery and spectral ground measurements. 2002
不同演替序列的藻结皮净光合速率日变化特征 3 2010
... 自20世纪90年代开始, 国内外一些****基于对结皮光谱特征的分析研究建立了在遥感影像上提取结皮覆盖度的结皮指数(Karnieli, 1997; Chen et al., 2005; Weber et al., 2008; Rozenstein & Karnieli, 2014), 其中有****认为结合NDVI和结皮指数能够提高结皮覆盖度的提取精度(Rozenstein & Karnieli, 2014; 冯秀绒等, 2015).也有****尝试利用NDVI在遥感影像上区分结皮与维管植物(Karnieli, 2003).但是, 同样由于结皮生物的变水特性及其有别于维管植物的物候节律造成其NDVI的水分变异性和季节变异性(Karnieli et al., 1996; Karnieli, 2003; Fang et al., 2015)较大, 而且不同演替阶段结皮在形态结构和光合生理特征方面差异较大(Zaady et al., 2000; 徐杰等, 2005; Housman et al., 2006; 贾荣亮, 2009; 苏延桂等, 2010; 王媛等, 2014; Zhang & Zhang; 2014), 且其分布随地形变化差异较大(王雪芹等, 2006; 程军回和张元明, 2010; 赵允格等, 2010), 引起干旱半干旱地区NDVI的不稳定性, 增加了遥感图像解译的困难而降低了以NDVI为基本参量进行植被生态参数的遥感估算准确性和精度(Karnieli et al., 1996; 房世波和张新时, 2011; Fang et al., 2015; Rodriguez- Caballero et al., 2015). ...
Diurnal variations of chlorophyll fluorescence and CO2 exchange of biological soil crusts in different successional stages in the Gurbantunggut Desert of northwestern China. 2014
古尔班通古特沙漠不同地貌部位生物结皮的选择性分布 1 2004
... 生物土壤结皮(简称结皮)是干旱沙区重要的地表生物覆盖物, 在沙区土壤生态、水文和地球化学循环等过程中发挥着重要作用, 其发育及演替特征常被作为重要指标来衡量生态系统稳定性和健康程度(李新荣等, 2009, 2014; 李新荣, 2010).因此, 在区域尺度上, 结皮的时空动态对于沙区生态系统管理具有重要的参考价值(李新荣等, 2014; Weber & Hill, 2016).虽然基于实验室测量或地块试验, 针对结皮的空间分布特点(张元明等, 2004; Maestre et al., 2005; 李新荣等, 2016)、拓殖和演替规律(Kleiner, 1983; Eldridge & Tozer, 1997; Zaady & Bouskila, 2002; Dettweiler-Robinson et al., 2013; 李新荣等, 2016)和碳储量估算(贾荣亮, 2009; Grote et al., 2010; 冯薇, 2014)等已有较多报道, 但由于结皮生物自身变水特性且易受到植被、土壤和地形等因素的影响, 其时空分异明显(王雪芹等, 2006; 程军回和张元明, 2010; 赵允格等, 2010), 使得上述研究结果往往难以准确推广到区域尺度.而遥感技术在较大尺度研究方面具有明显的技术优势, 为大尺度结皮时空格局监测、评估其碳储量、固碳/氮潜力和其他生态功能提供了重要机遇(Weber & Hill, 2016). ...
 , 高艳红
, 高艳红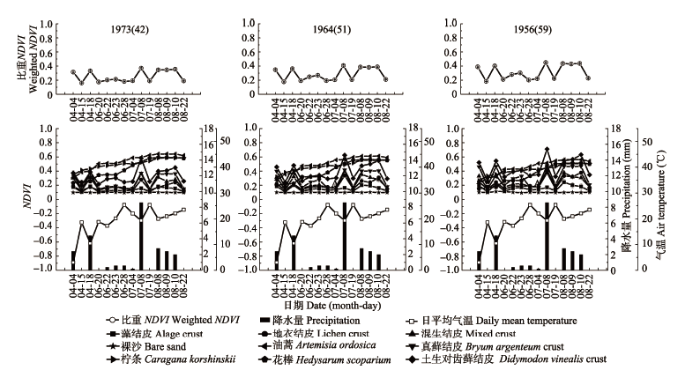 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT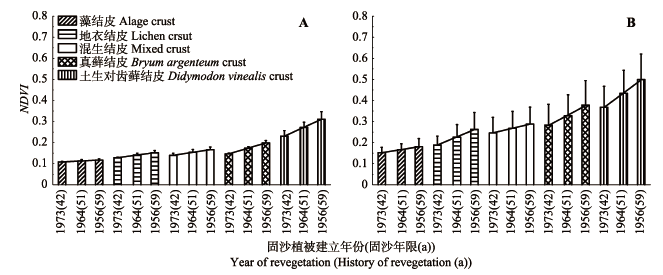 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT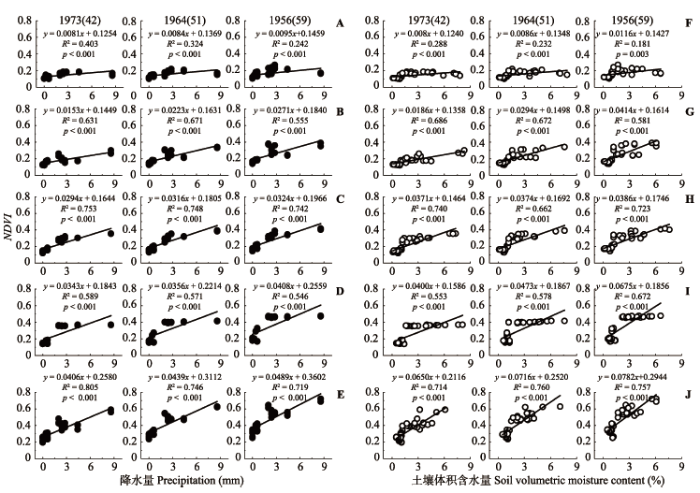 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT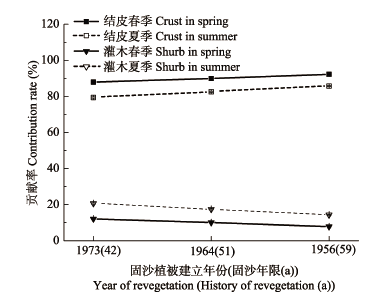 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}