Diversity of climbing seed plants and their reproductive habit in a karst seasonal rain forest in Nonggang, Guangxi, China
JIANGYu-Liang, LIXian-Kun*,, GUOYi-Li, DINGTao, WANGBin, XIANGWu-Sheng*, Guangxi Key Laboratory of Plant Conservation and Restoration Ecology in Karst Terrain, Guangxi Institute of Botany, Guangxi Zhuang Autonomous Region and Chinese Academy of Sciences, Guilin, Guangxi 541006, China; and Guangxi Youyiguan National Forest Ecosystem Research Station, Pingxiang, Guangxi 532600, China 收稿日期:2016-11-29 接受日期:2017-04-5 网络出版日期:2017-07-10 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家科技基础性工作专项(2015FY210200-14)、国家自然科学基金(NSFC31660130和31760131)、广西自然科学基金(2014GXNFSBA118081)和广西重点研发计划项目(桂科AB16380256) 作者简介: -->* 共同通信作者Co-author for correspondence (E-mail:xiankunli@163.com; xwusheng@qq.com)
关键词:喀斯特;季节性雨林;种子植物;藤本植物;生活型;开花;结实;生境资源 Abstract Aims Diversity of climbing seed plants and their reproductive habits and characteristics are central for the understanding of community structure and dynamics of forests and hence are important for forest protection. However, little is known about the climbing seed plants in northern tropical karst seasonal rain forests. Here, using the data of the species diversity and reproductive habits of climbing seed plants in Nonggang, Guangxi, China, we aim to 1) explore the species diversity and distribution of climbing seed plants in northern tropical karst seasonal rain forests, 2) study the flowering and fruiting phenology and 3) the associations of reproductive characteristics to the environment. Methods Species composition, preferred habitat, flowering time, fruiting time and fruit types of climbing seed plants were surveyed. The seasonality of flowering and fruiting were analyzed by concentration ratio and circular distribution. Climbing seed plants were divided into three groups according to their growth forms and places in spatial forest structure: bush ropes, herbaceous vines and lianas. Monthly flowering ratios, fruiting ratios, fruit types and their ratios in different groups were determined. These relationships of flowering ratio, fruiting ratio, fruit type and its ratio to meteorological factors were investigated using Pearson correlation analysis. Important findings There were a total of 333 species of climbing seed plants in Nonggang karst seasonal rain forest, belonging to 145 genera and 56 families. Bush ropes, herbaceous vines and lianas contained 119, 88 and 126 species, respectively. At species level, herbaceous vines were more abundance in valleys, while bush ropes and lianas were more abundance on slopes. Flowering and fruiting of climbing seed plants occurred seasonally, with flowering peaking in April to September, while fruiting peaking in July to December. The seasonality of flowering and fruiting in bush ropes was weaker than in herbaceous vines and lianas. Flowering ratio was significantly positively correlated with rainfall and air temperature, which suggest that flowering peaks in monsoon season. Peak time for fruiting was about three months later than the peak time of flowering, around the end of monsoon season. The ratio of samara species to all fruiting species in lianas was significantly positively correlated with wind speed, but negatively correlated with rainfall and air temperature. It showed that samara in lianas tended to occur in dry season with high wind speed. In conclusion, species diversity and the seasonal features of reproduction of climbing seed plants in Nonggang karst seasonal rain forest were closely related to the spatial and temporal variations of habitat resources.
Keywords:karst;seasonal rain forest;seed plant;climbing plant;growth form;flowering;fruiting;habitat resource -->0 PDF (4050KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 蒋裕良, 李先琨, 郭屹立, 丁涛, 王斌, 向悟生. 广西弄岗喀斯特季节性雨林藤本种子植物多样性及繁殖习性. 植物生态学报, 2017, 41(7): 716-728 https://doi.org/10.17521/cjpe.2016.0364 JIANGYu-Liang, LIXian-Kun, GUOYi-Li, DINGTao, WANGBin, XIANGWu-Sheng. Diversity of climbing seed plants and their reproductive habit in a karst seasonal rain forest in Nonggang, Guangxi, China. Chinese Journal of Plant Ecology, 2017, 41(7): 716-728 https://doi.org/10.17521/cjpe.2016.0364 藤本植物是森林物种多样性的重要组分, 在热带森林中极为丰富(Gentry, 1991; Schnitzer et al., 2012), 在亚热带森林中也占有十分重要的地位(蔡永立和宋永昌, 2000; 颜立红等, 2006; Hu et al., 2010)。藤本是植物界中较为进化的类群, 在生长、繁殖、适应性和竞争力上具有独特的优势(祁承经等, 2007), 在森林群落中常会降低树木生产力、增加树木死亡率, 延缓林木更新, 对森林结构、动态和演替过程具有重要的影响(Schnitzer & Carson, 2010; Tymen et al., 2016)。 藤本植物多样性和多度与多种生态因子相关, 其中温度和降水量被认为是最关键的非生物因子(Gentry, 1991; Schnitzer & Bongers, 2002; Hu et al., 2010)。藤本植物的导管比较宽大, 其水分运输易受低温和干旱胁迫的影响, 因而比较适宜热量丰富、雨量充沛的气候条件(胡亮, 2011)。但是, 藤本植物通常能够利用比树木更深层的土壤水, 避开旱季的干旱压力, 而且具有更高的水分利用效率(Chen et al., 2015), 因此, 季节性干旱显著的热带森林往往具有非常高的藤本植物丰富度(Gentry, 1991; Schnitzer, 2005)。藤本植物的繁殖物候节律响应季节性森林的干湿季节变化, 例如委内瑞拉热带稀树草原藤本植物的开花结实呈现明显的季节性, 大量的藤本植物在雨季开花、在雨季后期结实, 而熟果期集中在旱季(Ramirez, 2002)。大多数植物的繁殖时间及特性暗示着对环境因子的适应(van Schaik et al., 2003; Batalha & Martins, 2004), 所以掌握森林藤本植物的繁殖时间规律及相关特性有助于揭示藤本植物的生态适应机制, 可为森林生态系统的保护和管理提供科学依据, 但目前这方面的研究还较少。 喀斯特是一种季节性干旱比较明显的生境(肖兴艳等, 2015), 藤本植物分布十分丰富, 例如广西喀斯特地区的植被有较独特的藤刺灌丛类型, 在96个植被群系中, 藤刺灌丛占了28个(李先琨等, 2003), 充分显示了藤本植物在喀斯特植被中的普遍性和重要性。但目前关于喀斯特森林藤本植物的研究很少, 对喀斯特森林藤本植物多样性和生态学特性等仍缺乏深入的了解。喀斯特季节性雨林是中国北热带地区的地域性植被, 其生物多样性丰富, 种类组成不但具有明显的热带性质, 还有许多独特的地方。当前保护较好的喀斯特季节性雨林主要见于广西西南部(孙鸿烈, 2005)。本研究以保存完好的弄岗北热带喀斯特季节性雨林为研究区域, 依托广西友谊关森林生态系统国家定位观测研究站进行了长期的植物调查和环境监测,通过整理和分析藤本种子植物的种类组成、开花和结实时间、种实类型, 以及生境分布等数据, 拟探究以下问题: (1)该区域藤本种子植物多样性及分布特征如何?(2)这些藤本植物开花和结实在时间分布上是否有规律?(3)这些藤本植物开花或结实的发生及种实类型与环境因子的关联。
1 研究方法
1.1 研究区域概况
广西弄岗国家级自然保护区地处中越边境的广西壮族自治区龙州和宁明县境内(106.71°- 107.08° E, 22.23°-22.55° N), 呈东南至西北向, 总面积10 080 hm2。该保护区保存着世界上少有的面积较大、生态系统最为完整的典型北热带喀斯特季节性雨林(李先琨等, 2003)。区内地貌主要为碳酸盐岩上发育的峰丛洼地和峰丛谷地, 石峰密集, 基岩裸露, 陡峻, 地形复杂而破碎, 最高海拔约700 m。 该区属于热带季风气候, 春夏主要受印度洋北部西南季风影响, 风向偏南, 湿热多雨; 夏秋受赤道季风影响, 风向偏东风, 气候炎热、雨量充沛; 冬春受大陆高压东北季风影响, 风向偏东, 气温偏低, 干燥少雨。全年降水量1 150-1 550 mm, 降水量分布不均匀, 其中6、7、8三个月的降水约占全年降水量的50%, 12、1、2三个月的降水仅占全年降水量的6.5% (图1)。由于喀斯特峰丛洼地的特殊地貌, 夏季强降雨会因为排水不畅造成洼地淹水; 而山坡和山脊由于含水层浅薄, 即使在雨季也时常出现地质性干旱。全年日照时数1 500-1 800 h, 年平均日照百分率35%-41%; 9月份日照时数最多, 平均184-208 h, 2-3月份日照时数最少, 平均63-68 h。该区热量丰富, 累年稳定>10 ℃的积温平均为7 433-7 930 ℃, 最冷月平均气温在13 ℃以上, 个别年份极端最低气温可至0 ℃以下, 年最高气温可达39 ℃, 4-10月平均气温在22 ℃以上。风速的地理变化为南部、北部大, 年平均风速1.4-1.5 m·s-1, 西部最小, 平均0.8 m·s-1, 东部大于西部(林区外围数据)。通常林区风速比外围小1.0-2.0 m·s-1, 而风向往往与谷向一致(农绍勤, 1988)。 显示原图|下载原图ZIP|生成PPT 图1弄岗北热带喀斯特区域月平均降水量、风速、气温的变化(改自王斌等, 2014)。 -->Fig. 1Monthly variations of precipitation, wind speed and air temperature in Nonggang northern tropical karst area (adapted from Wang et al., 2014). -->
本次统计藤本种子植物共有333种, 隶属56科145属, 占本研究区种子植物总种数的20.8%; 双子叶藤本植物49科134属299种, 单子叶藤本植物6科10属31种, 裸子藤本植物1科1属3种(表1)。B类群有119种, 隶属27科55属, 木犀科、萝藦科、鼠李科占其种数的30.3%; H类群有88种, 隶属20科51属, 葫芦科、防己科、薯蓣科占其种数的46.6%; L类群有126种, 隶属31科60属, 葡萄科、苏木科、毛茛科占其种数的31.0%。这些藤本种子植物科的分布区有8个类型, 主要为热带广布型(30科); 热带性科占66.1%, 温带性科占10.7%, 其余的23.2%为世界分布性科(表2)。 Table 1 表1 表1弄岗喀斯特季节性雨林藤本种子植物的多样性组成及其占本地区种子植物的比例 Table 1The diversity of climbing seed plants and their proportion in seed plants of Nonggang karst seasonal rain forest
类群 Type
藤本种子植物 Climbing seed plants
种子植物 Seed plants
藤本种子植物占种子植物的比例 Proportions of climbing seed plants in seed plants (%)
科 Family
属 Genus
种 Species
科 Family
属 Genus
种 Species
科 Family
属 Genus
种 Species
裸子植物 Gymnosperm
1
1
3
4
5
10
25.0
20.0
30.0
被子植物 Angiosperm
单子叶植物 Monocotyledon
6
10
31
25
143
255
24.0
7.0
12.2
双子叶植物 Dicotyledon
49
134
299
126
611
1 337
38.9
21.9
22.4
合计 Total
56
145
333
155
759
1 602
36.1
19.1
20.8
新窗口打开 Table 2 表2 表2弄岗喀斯特季节性雨林藤本种子植物科的组成及分布区类型 Table 2The families containing climbing seed plants and their areal-types in Nonggang karst seasonal rain forest
科名 Family
属数 Genus No.
B种数 Species No. of B
H种数 Species No. of H
L种数 Species No. of L
总种数 Total species No.
分布区类型 Areal type
萝藦科 Asclepiadaceae
13
12
5
9
26
2
蝶形花科 Papilionaceae
13
8
6
8
22
1
葫芦科 Cucurbitaceae
13
0
20
2
22
2
葡萄科 Vitaceae
5
0
5
17
22
2
防己科 Menispermaceae
10
0
12
7
19
2
木犀科 Oleaceae
1
13
0
5
18
1
夹竹桃科 Apocynaceae
10
6
0
9
15
2
茜草科 Rubiaceae
4
5
2
5
12
1
苏木科 Caesalpiniaceae
3
0
0
12
12
2 (2-2)
鼠李科 Rhamnaceae
5
11
0
0
11
1
毛茛科 Ranunculaceae
2
0
1
10
11
1
蔷薇科 Rosaceae
2
10
0
0
10
1
旋花科 Convolvulaceae
6
2
5
2
9
1
番荔枝科 Annonaceae
4
8
0
1
9
2
菝葜科 Smilacaceae
2
8
1
0
9
2
薯蓣科 Dioscoreaceae
1
0
9
0
9
2
天南星科 Araceae
4
0
8
0
8
2
桑科 Moraceae
3
6
0
1
7
1
芸香科 Rutaceae
1
2
0
4
6
2
白花菜科 Capparidaceae
1
5
0
0
5
2
马兜铃科 Aristolochiaceae
1
0
2
3
5
2
卫矛科 Celastraceae
2
4
0
0
4
2
西番莲科 Passifloraceae
2
0
4
0
4
2
忍冬科 Caprifoliaceae
1
0
0
4
4
8
翅子藤科 Hippocrateaceae
2
0
0
3
3
3
金虎尾科 Malpighiaceae
2
1
0
2
3
2
菊科 Compositae
1
3
0
0
3
1
买麻藤科 Gnetaceae
1
0
0
3
3
2 (2-2)
五味子科 Schisandraceae
1
0
0
3
3
9
紫金牛科 Myrsinaceae
1
3
0
0
3
2
棕榈科 Arecaceae
1
0
0
3
3
2
使君子科 Combretaceae
2
1
0
1
2
2
远志科 Polygalaceae
2
2
0
0
2
1
胡椒科 Piperaceae
1
0
1
1
2
2
胡颓子科 Elaeagnaceae
1
2
0
0
2
8 (8-4)
桔梗科 Campanulaceae
1
0
2
0
2
1
猕猴桃科 Actinidiaceae
1
0
0
2
2
14
青藤科 Illigeraceae
1
0
0
2
2
2
清风藤科 Sabiaceae
1
0
0
2
2
7
百部科 Stemonaceae
1
0
1
0
1
5
百合科 Liliaceae
1
0
1
0
1
8
大戟科 Euphorbiaceae
1
1
0
0
1
2
大麻科 Cannabaceae
1
0
1
0
1
8
大血藤科 Sargentodoxaceae
1
0
0
1
1
7 (7-4)
含羞草科 Mimosaceae
1
1
0
0
1
2
蓼科 Polygonaceae
1
0
1
0
1
1
落葵科 Basellaceae
1
0
1
0
1
2
马鞭草科 Verbenaceae
1
1
0
0
1
3
牛栓藤科 Connaraceae
1
0
0
1
1
2
漆树科 Anacardiaceae
1
0
0
1
1
2
紫茉莉科 Nyctaginaceae
1
1
0
0
1
3
山柚子科 Opiliaceae
1
1
0
0
1
2
梧桐科 Sterculiaceae
1
0
0
1
1
2
五桠果科 Dilleniaceae
1
0
0
1
1
2 (2-1)
斜翼科 Plagiopteraceae
1
1
0
0
1
7 (7-3)
玄参科 Scrophulariaceae
1
1
0
0
1
1
总计 Total
145
119
88
126
333
B, bushy ropes; H, herbaceous vines; L, lianas. Areal-type: 1, widespread; 2, pantropic; 2-1, tropical Asia-Australasia and tropical America; 2-2, tropical Asia-tropical Africa-tropical America; 3, tropical and subtropical East Asia and tropical South America disjuncted; 5, tropical Asia to tropical Australasia Oceania; 7, tropical Asia; 7-3, Myanmar, Thailand to Southwest China; 7-4, Vietnam or Indochinese Peninsula to South or Southwest China; 8, North Temperate; 8-4, North Temperate and South Temperate disjuncted; 9, East Asia and North America disjuncted; 14, East Asia.B, 藤状灌木; H, 草质藤本; L, 木质藤本。分布区类型: 1, 世界广布; 2, 热带广布; 2-1, 热带亚洲-大洋洲和热带美洲; 2-2, 热带亚洲-热带非洲-热带美洲; 3, 东亚(热带、亚热带)及热带南美洲间断; 5, 热带亚洲至热带大洋洲; 7, 热带亚洲; 7-3, 缅甸、泰国至华西南分布; 7-4, 越南(或中南半岛)至华南或西南分布; 8, 北温带; 8-4, 北温带和南温带间断分布; 9, 东亚及北美间断; 14, 东亚。 新窗口打开 B、H、L类群在山顶、坡地和洼地(谷地)的种数分布统计结果表明: B类群在不同地貌部位的种数分布相对匀称, 种数变异系数为0.49, 而H和L类群分别为0.72和0.53; B、H、L类群分别有54.6%、56.8%、52.4%的物种局限于某一地貌部位分布, 说明了B类群能较大程度地利用不同的生境空间, 多数物种有各自主要的分布生境, 在地形上存在的分异相对较明显(图2)。B类群中于山顶分布的常见种有吊山桃(Secamone sinica)、羽叶金合欢(Acacia pennata)、球兰(Hoya carnosa)等, 于坡地分布的常见种有扭肚藤(Jasminum elongatum)、多花勾儿茶(Berchemia floribunda)、粉背菝葜(Smilax hypoglauca)等, 于洼地分布的常见种有黔桂悬钩子(Rubus feddei)、阔叶瓜馥木(Fissistigma chloroneurum)等。H类群中于洼地分布的物种占较大的组分, 如乌蔹莓(Cayratia japonica)、金钟藤(Merremia boisiana)、凹萼木鳖(Momordica subangulata)、光叶薯蓣(Dioscorea glabra)等, 于山顶分布的物种所占组分较小, 仅调查到9种, 如眼树莲(Dischidia chinensis)、华千金藤(Stephania sinica)、粪箕笃(Stephania longa)、藤桔(Pothos chinensis)等。L类群以于坡地分布的物种为主, 有较多物种于山顶、坡地和洼地均有分布, 如毛青藤(Illigera rhodantha)、暗消藤(Streptocaulon juventas)、二籽扁索藤(Pristimera arborea)、小木通(Clematis armandii)等。 显示原图|下载原图ZIP|生成PPT 图2不同类群藤本种子植物在不同地貌部位的种数分布示意图。B、H和L见表2。 -->Fig. 2Sketch for species numbers of different groups of climbing seed plants in different topography positions. B, H and L see Table 2. -->
2.2 开花结实的季节性和种实类型组成
B、H和L类群的z值均大于相应的z0.05, 表明藤本种子植物开花或结实倾向特定时间集中, 存在高峰日和高峰期。Watson's U2检验表明, B、H、L类群两两之间的开花或结实高峰时间无显著性差别(U2 < U20.05, p > 0.05)。整体的开花高峰期为4月4日至9月10日, 峰值在6月21日前后; 结实高峰期为7月7日至12月19日, 峰值在9月25日前后(表3)。 Table 3 表3 表3不同类群藤本种子植物开花结实的季节性及高峰时间 Table 3Seasonality and peak time of flowering and fruiting in different groups of climbing seed plants
类群 Group
开花 Flowering
结实 Fruiting
集中度 Concentration ratio (M)
高峰日 Peak day (month-day)
高峰期 Peak season (month-day-month-day)
z
集中度 Concentration ratio (M)
高峰日 Peak day (month-day)
高峰期 Peak season (month-day-month-day)
z
B
0.29
6-13
3-15-9-14
31.18
0.23
9-20
6-16-12-30
22.79
H
0.45
7-5
5-24-9-18
68.87
0.43
9-28
7-18-12-14
64.78
L
0.46
6-17
4-7-8-30
69.19
0.44
9-25
7-16-12-11
79.20
B & H & L
0.39
6-21
4-4-9-10
172.62
0.36
9-25
7-7-12-19
169.86
B, H and L see Table 2.B、H和L见表2。 新窗口打开 B类群开花、结实的M值均小于0.3, 表明其开花和结实的时间分布比较均匀, 时间聚集性较差。而H和L类群开花、结实的M值均在0.3-0.5之间, 表明有一定的季节性(表3)。从月开花比率的变化上看, B类群的变化幅度最小(13.6%-53.4%), L类群的变化幅度最大(7.1%- 57.9%); 月结实比率的变化也以B类群的变化幅度最小(21.9%-55.9%), L类群的变化幅度最大(8.7%-69.0%) (图3)。 显示原图|下载原图ZIP|生成PPT 图3弄岗喀斯特季节性雨林各月份藤本种子植物开花和结实比率。B、H和L见表2。 -->Fig. 3Monthly flowering and fruiting ratios in climbing seed plants of Nonggang karst seasonal rain forest. B, H and L see Table 2. -->
本次统计的藤本种子植物种实有瘦果、坚果、蒴果、荚果、蓇葖果、浆果、核果、翅果和裸子藤本植物种子9个类型, 裸子藤本植物种子类型仅限于L类群中存在, 坚果类型于L类群中不存在, 翅果类型于H类群中不存在。B、H和L类群的种实构成及比率月变化呈现一定的差异(图4)。1-12月B类群中肉果丰富度均大于干果; 2-11月H类群中肉果丰富度均大于干果, 1月和12月则相反; 6月L类群中肉果丰富度仅稍大于干果, 其他月份均小于干果(图5)。总体上, B类群的肉果丰富度最大, 而L类群的干果丰富度最大。 显示原图|下载原图ZIP|生成PPT 图4弄岗喀斯特季节性雨林各月份不同类群藤本种子植物的种实构成比率。B、H和L见表2。 -->Fig. 4Monthly variation in the ratios of species with different fruit types in climbing seed plants in Nonggang karst seasonal rain forest. B, H and L see Table 2. -->
显示原图|下载原图ZIP|生成PPT 图5弄岗喀斯特季节性雨林各月份不同类群藤本种子植物肉果与干果的丰富度。B、H和L见表2。 -->Fig. 5Monthly richness of species with fleshy and dry fruits in different groups of climbing seed plants in Nonggang karst seasonal rain forest. B, H and L see Table 2. -->
2.3 开花结实与气象因子的相关性
藤本种子植物的开花、结实比率与气象因子存在一定的相关关系(表4)。B、H、L类群的开花比率与气温和降水量极显著正相关(p < 0.01), 与风速的相关关系不显著(p > 0.05)。B、H、L类群的结实比率与气温和降水量的相关关系不显著(p > 0.05), 与风速呈极显著的负相关关系(p < 0.01)。另外, 在L类群中翅果比率与风速呈极显著正相关关系(p < 0.01), 与气温和降水量呈极显著负相关关系(p < 0.01)。 Table 4 表4 表4不同类群藤本种子植物开花结实比率及木质藤本的翅果比率与气象因子的相关系数 Table 4Coefficients of correlations between meteorological factors and flowering and fruiting ratios in different groups of climbing seed plants and the ratio of samara species to all fruiting species in lianas
气象因子 Meteorological factor
开花 Flowering
结实 Fruiting
B
H
L
B
H
L
气温 Air temperature
0.833**
0.963**
0.854**
0.380
0.269
0.296 (-0.747**)
降水量 Precipitation
0.838**
0.960**
0.880**
0.189
0.042
0.103 (-0.715**)
风速 Wind speed
-0.142
-0.426
-0.217
-0.906**
-0.835**
-0.860** (0.876**)
**, p < 0.01. The numbers in bracket are the coefficients of correlations between meteorological factors and the ratio of samara species to all fruiting species in lianas. B, H and L see Table 2.**, p < 0.01。括号内为木质藤本的翅果比率与气象因子的相关系数。B、H和L见表2。 新窗口打开
BaiB, LiN, LuCH (2011). Influence of habitat fragmentation on frugivores and their seed dispersal. Chinese Journal of Ecology, 30, 2613-2620. (in Chinese with English abstract)[白冰, 李宁, 鲁长虎 (2011). 生境破碎化对食果动物及种子传播的影响 . 生态学杂志, 30, 2613-2620.] [本文引用: 1]
[2]
BatalhaMA, MartinsFR (2004). Reproductive phenology of the cerrado plant community in Emas National Park (central Brazil). Australian Journal of Botany, 52, 149-161. [本文引用: 1]
[3]
CaiYL, SongYC (2000). Diversity of vines in subtropical zone of east China. Journal of Wuhan Botanical Research, 18, 390-396. (in Chinese with English abstract)[蔡永立, 宋永昌 (2000). 中国亚热带东部藤本植物的多样性 . 武汉植物学研究, 18, 390-396.] [本文引用: 1]
[4]
ChenXS (2008). Community Level Plant Reproductive Traits of Tropical Rain Forest in Xishuangbanna, SW China . PhD dissertation, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Xishuangbanna. 30-58. (in Chinese with English abstract)[陈心胜 (2008). 西双版纳热带雨林植物群落的繁殖生物学特性研究 . 博士学位论文, 中国科学院西双版纳热带植物园, 西双版纳. 30-58.] [本文引用: 1]
[5]
ChenYJ, CaoKF, SchnitzerSA, FanZX, ZhangJL, BongersF (2015). Water-use advantage for lianas over trees in tropical seasonal forests. New Phytologist, 205, 128-136. [本文引用: 1]
[6]
ChenYJ, ChenJW, CaiZQ (2007). Lianas and their functions in tropical forests. Chinese Bulletin of Botany, 24, 240-249. (in Chinese with English abstract)[陈亚军, 陈军文, 蔡志全 (2007). 木质藤本及其在热带森林中的生态学功能 . 植物学通报, 24, 240-249.] [本文引用: 1]
[7]
DengY, JiangZC, QinXM (2012). Water source partitioning among trees growing on carbonate rock in a subtropical region of Guangxi, China. Environmental Earth Sciences, 66, 635-640. [本文引用: 1]
[8]
FangWW, YuSL (2013). Fruit ecology: Its concepts, research advances, and research perspectives. Chinese Journal of Ecology, 32, 2238-2244. (in Chinese with English abstract)[方伟伟, 于顺利 (2013). 果实生态学的概念、研究现状及研究方向 . 生态学杂志, 32, 2238-2244.] [本文引用: 1]
[9]
FranksSJ, SimS, WelsAE (2007). Rapid evolution of flowering time by an annual plant in response to a climate fluctuation. Proceedings of the National Academy of Sciences of the United States of America, 104, 1278-1282. [本文引用: 1]
[10]
GentryAH (1991). The distribution and evolution of climbing plants. In: Putz FE, Mooney HA eds. The Biology of Vines. Cambridge University Press, Cambridge, UK. 3-52. [本文引用: 4]
[11]
GuoK, LiuCC, DongM (2011). Ecological adaptation of plants and control of rocky-desertification on karst region of Southwest China. Chinese Journal of Plant Ecology, 35, 991-999. (in Chinese with English abstract)[郭柯, 刘长成, 董鸣 (2011). 我国西南喀斯特植物生态适应性与石漠化治理 . 植物生态学报, 35, 991-999.] [本文引用: 1]
[12]
GuoYL, WangB, XiangWS, DingT, LuSH, HuangYS, HuangFZ, LiDX, LiXK (2015). Spatial distribution of tree species in a tropical karst seasonal rainforest in Nonggang, Guangxi, southern China. Biodiversity Science, 23, 183-191. (in Chinese with English abstract)[郭屹立, 王斌, 向悟生, 丁涛, 陆树华, 黄俞淞, 黄甫昭, 李冬兴, 李先琨 (2015). 广西弄岗北热带喀斯特季节性雨林监测样地种群空间点格局分析 . 生物多样性, 23, 183-191.] [本文引用: 1]
[13]
HuCG (1988). A report on the investigation of hydrogeology from Nonggang Natural Reserve. Guihaia, 8(Suppl. 1), 17-32. (in Chinese with English abstract)[胡长庚 (1988). 弄岗自然保护区水文地质考察报告 . 广西植物, 8(增刊1), 17-32.] [本文引用: 1]
[14]
HuL (2011). Distribution and diversity of climbing plants in temperate East Asia. Biodiversity Science, 19, 567-573. (in Chinese with English abstract)[胡亮 (2011). 东亚温带藤本植物多样性及其格局 . 生物多样性, 19, 567-573.] [本文引用: 1]
[15]
HuL, LiM, LiZ (2010). Geographical and environmental gradients of lianas and vines in China. Global Ecology and Biogeography, 19, 554-561. [本文引用: 2]
[16]
HuangFZ, DingT, LiXK, GouYL, WangB, XiangWS, WenSJ, LiDX, HeYL (2016). Species diversity for various associations along an altitudinal gradient in the karst seasonal rainforest in Nonggang. Acta Ecologica Sinica, 36, 4509-4517. (in Chinese with English abstract)[黄甫昭, 丁涛, 李先琨, 郭屹立, 王斌, 向悟生, 文淑均, 李冬兴, 何运林 (2016). 弄岗喀斯特季节性雨林不同群丛物种多样性随海拔的变化 . 生态学报, 36, 4509-4517.] [本文引用: 2]
[17]
HuangYS, WuWH, JiangRH, LiuSY, LiuY, LiXK (2013). Primary study on species diversity of plant in Nonggong National Natural Reserve of Guangxi. Guihaia, 33, 346-355. (in Chinese with English abstract)[黄俞淞, 吴望辉, 蒋日红, 刘晟源, 刘演, 李先琨 (2013). 广西弄岗国家级自然保护区植物物种多样性初步研究 . 广西植物, 33, 346-355.] [本文引用: 3]
[18]
HuangZH, HuangCM, ZhouQH, WeiH, MengYJ (2010). Diet and the seasonal changes of the Francois’ langur (Trachypithecus francoisi). Acta Ecologica Sinica, 30, 5501-5508. (in Chinese with English abstract)[黄中豪, 黄乘明, 周岐海, 韦华, 蒙渊君 (2010). 黑叶猴食物组成及其季节性变化 . 生态学报, 30, 5502-5508.] [本文引用: 2]
[19]
KrishnanRM (2002). Reproductive phenology of a wet forest understorey in the Western Ghats, South India. Global Ecology & Biogeography, 11, 179-182. [本文引用: 1]
[20]
LaskyJR, UriarteM, MuscarellaR (2016). Synchrony, compensatory dynamics, and the functional trait basis of phenological diversity in a tropical dry forest tree community: Effects of rainfall seasonality. Environmental Research Letters, 11, 115003. doi: 10.1088/1748-9326/11/11/115003.
[21]
LiWY, LinDQ, LinZS, YanRY (2003). Studies on the diversity of lianas in Tengshan Nature Reserve of Fujian. Journal of Fujian Forestry Science and Technology, 30(1), 28-33. (in Chinese with English abstract)[黎维英, 林道清, 林致盛, 鄢仁杨 (2003). 福建藤山自然保护区藤本植物多样性研究 . 福建林业科技, 30(1), 28-33.] [本文引用: 1]
[22]
LiXK, SuZM, LüSH, OuZL, XiangWS, OuZ, LuSH (2003). The spatial pattern of natural vegetation in the karst regions of Guangxi and the ecological signality for ecosystem rehabilitation and reconstruction. Journal of Mountain Science, 21, 129-139. (in Chinese with English abstract)[李先琨, 苏宗明, 吕仕洪, 欧祖兰, 向悟生, 区智, 陆树华 (2003). 广西岩溶植被自然分布规律及对岩溶生态恢复重建的意义 . 山地学报, 21, 129-139.] [本文引用: 2]
[23]
LiZS, TangJW, ZhengZ, LiQJ, DuanWY, ZhuSZ, GuoXM, ZhaoJW, ZengR (2005). Coenological characteristics of tropical montane rain forest in Xishuangbanna. Journal of Wuhan Botanical Research, 23, 319-326. (in Chinese with English abstract)[李宗善, 唐建维, 郑征, 李庆军, 段文勇, 朱胜忠, 郭贤明, 赵建伟, 曾荣 (2005). 西双版纳热带山地雨林群落学特征分析 . 武汉植物学研究, 23, 319-326.] [本文引用: 1]
[24]
LiangCF, LiangJY, LiuLF, MoXL (1988). A report on the floristic survey on the Nonggang Natural Reserve. Guihaia, 8(Suppl. 1), 83-184. (in Chinese with English abstract)[梁畴芬, 梁健英, 刘兰芳, 莫新礼 (1988). 弄岗自然保护区植物区系考察报告 . 广西植物, 8(增刊1), 83-184.] [本文引用: 2]
[25]
LongGZ (1988). Terrestrial vertebrates in the Nonggang Natural Reserve. Guihaia, 8(Suppl. 1), 267-275. (in Chinese with English abstract)[龙国珍 (1988). 弄岗自然保护区陆栖脊椎动物调查报告 . 广西植物, 8(增刊1), 267-275.] [本文引用: 1]
[26]
NieYP, ChenHS, WangKL (2011). Seasonal variation of water sources for plants growing on continuous rock outcrops in limestone area of Southwest China. Chinese Journal of Plant Ecology, 35, 1029-1037. (in Chinese with English abstract)[聂云鹏, 陈洪松, 王克林 (2011). 石灰岩地区连片出露石丛生境植物水分来源的季节性差异 . 植物生态学报, 35, 1029-1037.] [本文引用: 1]
[27]
NongSQ (1988). A report on the climatological survey from the Nonggang Natural Reserve. Guihaia, 8(Suppl. 1), 74-82. (in Chinese with English abstract)[农绍勤 (1988). 弄岗自然保护区气候资源考察报告 . 广西植物, 8(增刊1), 74-82.] [本文引用: 2]
[28]
PengSJ, HuangZL, PengSL, XuGL (2003). The processes and mechanisms of the dispersal of fleshy-fruited plants at different spatial scales. Acta Ecologica Sinica, 23, 777-786. (in Chinese with English abstract)[彭闪江, 黄忠良, 彭少麟, 徐国良 (2003). 不同空间尺度下的肉果植物扩散过程和机理 . 生态学报, 23, 777-786.] [本文引用: 1]
[29]
PleinM, LängsfeldL, NeuschulzEL, SchultheissC, IngmannL, TöpferT, Böhning-GaeseK, SchleuningM (2013). Constant properties of plant-frugivore networks despite fluctuations in fruit and bird communities in space and time. Ecology, 94, 1296-1306. [本文引用: 1]
QiCJ, YanLH, PengCL (2007). Lianas as a new, suddenly arised force—Description and review of lianas literatures. Journal of Wuhan Botanical Research, 25, 381-395. (in Chinese with English abstract)[祁承经, 颜立红, 彭春良 (2007). 异军突起的藤本植物 . 武汉植物学研究, 25, 381-395.] [本文引用: 2]
[32]
RamirezN (2002). Reproductive phenology, life-forms, and habitats of the Venezuelan Central plain. American Journal of Botany, 85, 836-842. [本文引用: 1]
[33]
RamirezN, BricenoH (2011). Reproductive phenology of 233 species from four herbaceous-shrubby communities in the Gran Sabana Plateau of Venezuela. AoB PLANTS, 3, plr014. doi:10.1093/aobpla/plr014. [本文引用: 1]
[34]
SchnitzerSA (2005). A mechanistic explanation for global patterns of liana abundance and distribution. The American Naturalist, 166, 262-276. [本文引用: 2]
[35]
SchnitzerSA, BongersF (2002). The ecology of lianas and their role in forests. Trends in Ecology & Evolution, 17, 223-230. [本文引用: 1]
[36]
SchnitzerSA, CarsonWP (2010). Lianas suppress tree regeneration and diversity in treefall gaps. Ecology Letters, 13, 849-857. [本文引用: 1]
[37]
SchnitzerSA, ManganSA, DallingJW, BaldeckCA, HubbellSP, LedoA, Muller-LandauH, TobinMF, AguilarS, BrassfieldD, HernandezA, LaoS, PerezR, ValdesO, YorkeSR (2012). Liana abundance, diversity, and distribution on Barro Colorado Island, Panama. PLOS ONE, 7(12), e52114. doi: 10.1371/journal.pone.0052114. [本文引用: 1]
[38]
ShanRQ, XuY, XueDY (2006). The application of the concentration ratio and the circular distribution in the analysis of seasonality of infectious diseases. Disease Surveillance, 21, 589-591. (in Chinese with English abstract)[山若青, 徐毅, 薛大燕 (2006). 应用集中度和圆形分布分析传染病的季节性 . 疾病监控, 21, 589-591.]
[39]
ShiZP (2015). Diversity of Mammals and Birds and Their Ecology in the Nonggang Forest Dynamic Plot, China . Master degree dissertation, Guangxi Normal University, Guilin. (in Chinese with English abstract)[施泽攀 (2015). 基于红外相机对弄岗森林动态监测样地及周边的鸟兽多样性及生态学研究 . 硕士学位论文, 广西师范大学, 桂林.] [本文引用: 1]
[40]
SuZM, ZhaoTL, HuangQC (1988). The vegetation of Longgang Natural Reserve in Guangxi. Guihaia, 8(Suppl. 1), 188-214. (in Chinese with English abstract)[苏宗明, 赵天林, 黄庆昌 (1988). 弄岗自然保护区植被调查报告 . 广西植物, 8(增刊1), 185-214.]
[41]
SunHL (2005). Chinese Ecosystem. Science Press, Beijing. 327-365. (in Chinese)[孙鸿烈 (2005). 中国生态系统. 科学出版社, 北京. 327-365.] [本文引用: 2]
[42]
TangHX (2008). Feeding Ecology of the Rhese Macaque (Macaca mulatta) at Nonggang National Nature Reserve, China . Master degree dissertation, Guangxi Normal University, Guilin. (in Chinese with English abstract)[唐华兴 (2008). 弄岗猕猴(Macaca mulatta)的觅食生态学 . 硕士学位论文, 广西师范大学, 桂林.] [本文引用: 1]
[43]
TymenB, Réjou-MéchainM, DallingJW, FausetS, FeldpauschTR, NordenN, PhillipsOL, TurnerBL, ViersJ, ChaveJ (2016). Evidence for arrested succession in a liana-infested Amazonian forest. Journal of Ecology, 104, 149-159. [本文引用: 1]
[44]
van SchaikCP, TerborghJW, WrightSJ (2003). The phenology of tropical forests: Adaptive significance and consequences for primary consumers. Annual Review of Ecology, Evolution and Systematics, 24, 353-377. [本文引用: 1]
[45]
WangB, HuangYS, LiXK, XiangWS, DingT, HuangFZ, LuSH, HanWH, WenSJ, HeLJ (2014). Species composition and spatial distribution of the 15 ha northern tropical karst seasonal rain forest dynamic plot in Nonggang of Guangxi Southern China. Biodiversity Science, 22, 141-156.[王斌, 黄俞淞, 李先琨, 向悟生, 丁涛, 黄甫昭, 陆树华, 韩文衡, 文淑均, 何兰军 (2014). 弄岗北热带喀斯特季节性雨林15 ha监测样地的树种组成与空间分布 . 生物多样性, 22, 141-156.] [本文引用: 4]
[46]
WuWH (2011). Studies on the Flora and Phytogeography of Nonggang Nature Reserve of Guangxi, China . Master degree dissertation, Guangxi Normal University, Guilin. (in Chinese with English abstract)[吴望辉 (2011). 广西弄岗国家级自然保护区植物区系地理学研究 . 硕士学位论文, 广西师范大学, 桂林.]
[47]
WuZY, ZhouZK, LiDZ, PengH, SunH (2003). The areal- types of the world families of seed plants. Acta Botanica Yunnanica, 25, 245-257. (in Chinese with English abstract)[吴征镒, 周浙昆, 李德铢, 彭华, 孙航 (2003). 世界种子植物科的分布区类型系统 . 云南植物研究, 25, 245-257.] [本文引用: 1]
[48]
XiaoXY, LiuF, YaoB, ShuYG (2015). Research progress of soil moisture in karst area of southwest China. Journal of Yunnan Agricultural University, 30, 958-964. (in Chinese with English abstract)[肖兴艳, 刘方, 姚斌, 舒英格 (2015). 中国西南喀斯特土壤水分研究进展 . 云南农业大学学报, 30, 958-964.] [本文引用: 1]
[49]
XuYL, ZengFP, SongTQ, PengWX, LuLX, YuZ, LiuY (2012). Study on microclimate characteristics of a secondary forest in depression between karst hills. Research of Agricultural Modernization, 33, 239-244. (in Chinese with English abstract)[徐云蕾, 曾馥平, 宋同清, 彭晚霞, 卢凌霄, 俞孜, 刘艳 (2012). 喀斯特峰丛洼地次生林小气候特征研究 . 农业现代化研究, 33, 239-244.] [本文引用: 1]
[50]
YanLH (2007). Sexual system and environmental adaptability of vines in Hupingshan Mountain, Hunan Province. Journal of Northeast Forestry University, 35(7), 35-36, 39. (in Chinese with English abstract)[颜立红 (2007). 湖南壶瓶山藤本植物的有性系统及其环境适应性 . 东北林业大学学报, 35(7), 35-36, 39.] [本文引用: 1]
[51]
YanLH, QiCJ, LiuXX (2006). A study on the flora of the seed vines in central China region. Bulletin of Botanical Research, 26, 497-507. (in Chinese with English abstract)[颜立红, 祁承经, 刘小雄 (2006). 华中地区藤本种子植物区系的研究 . 植物研究, 26, 497-507.] [本文引用: 1]
[52]
YangR, YuLF, AnMT (2008). Analysis on microhabitat characteristic in karst region—Taking Maolan Nature Reserve as an example. Guizhou Agricultural Sciences, 36(6), 168-169. (in Chinese with English abstract)[杨瑞, 喻理飞, 安明态 (2008). 喀斯特区小生境特征现状分析: 以茂兰自然保护区为例 . 贵州农业科学, 36(6), 168-169.] [本文引用: 1]
[53]
YuanTX, ZhangHP, OuZY, TanYB (2014). Effects of topography on the diversity and distribution pattern of ground plants in karst montane forests in southwest Guangxi, China. Chinese Journal of Applied Ecology, 25, 2803-2810. (in Chinese with English abstract)[袁铁象, 张合平, 欧芷阳, 谭一波 (2014). 地形对桂西南喀斯特山地森林地表植物多样性及分布格局的影响 . 应用生态学报, 25, 2803-2810.] [本文引用: 2]
[54]
ZarJH (1999). Biostatistical Analysis. 4th edn . Prentice-Hall, Upper Saddle River, USA. 616-621. [本文引用: 1]
[55]
ZhangYW, YangHP (2001). The studies on flora and ecological features of liana in Fanjing Mountain National Nature Reserve, Guizhou, China. Journal of Wuhan Botanical Research, 19, 269-298. (in Chinese with English abstract)[张玉武, 杨红萍 (2001). 贵州梵净山国家级自然保护区藤本植物的研究 . 武汉植物学研究, 19, 269-298.] [本文引用: 1]
[56]
ZhangZH, HuG, ZhuJD, NiJ (2011). Spatial heterogeneity of soil nutrients and its impact on tree species distribution in a karst forest of Southwest China. Chinese Journal of Plant Ecology, 35, 1038-1049. (in Chinese with English abstract)[张忠华, 胡刚, 祝介东, 倪健 (2011). 喀斯特森林土壤养分的空间异质性及其对树种分布的影响 . 植物生态学报, 35, 1038-1049.] [本文引用: 1]
[57]
ZhouQH, WeiFW, LiM, HuangCM, LuoB (2006). Diet and food choice of Trachypithecus francoisi in the Nonggang Nature Reserve, China. International Journal of Primatology, 27, 1441-1460.
ZhouYR (1984). Ecological characteristics of vines in the region of South China. Acta Phytoecologica et Geobotanica Sinica, 8, 199-206. (in Chinese with English abstract)[周远瑞 (1984). 华南藤本植物的生态学特性 . 植物生态学与地植物学丛刊, 8, 199-206.] [本文引用: 2]
[60]
ZhuH (2008). Species composition and diversity of lianas in tropical forests of southern Yunnan (Xishuangbanna), south-western China. Journal of Tropical Forest Science, 20, 111-122. [本文引用: 2]
[61]
ZhuH, WangH, LiBG, SirirugsaP (2003). Biogeography and floristic affinities of the limestone flora in southern Yunnan, China. Annals of the Missouri Botanical Garden, 90, 444-465. [本文引用: 2]
[62]
ZhuH, XuZF, WangH, LiBG, LongBY (2000). Effects of fragmentation on the structure, species composition and diversity of tropical rain forest in Xishuangbanna, Yunnan. Acta Phytoecologica Sinica, 24, 560-568. (in Chinese with English abstract)[朱华, 许再富, 王洪, 李保贵, 龙碧云 (2000). 西双版纳片段热带雨林的结构、物种组成及其变化的研究 . 植物生态学报, 24, 560-568.]
Synchrony, compensatory dynamics, and the functional trait basis of phenological diversity in a tropical dry forest tree community: Effects of rainfall seasonality. 2016
 , 郭屹立, 丁涛, 王斌, 向悟生
, 郭屹立, 丁涛, 王斌, 向悟生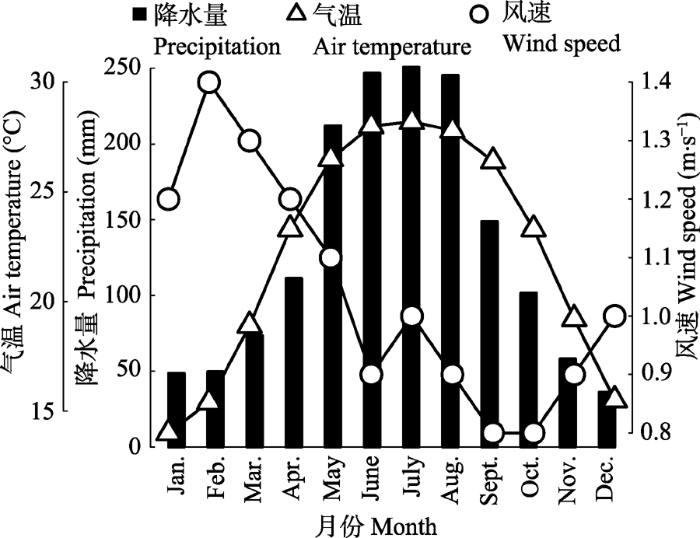 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT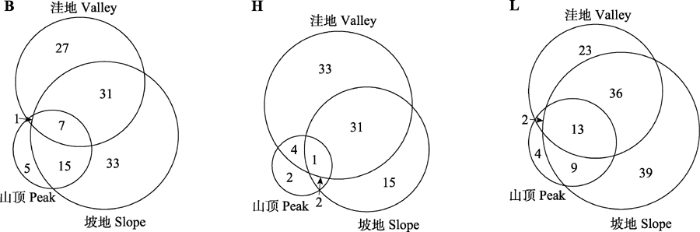 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT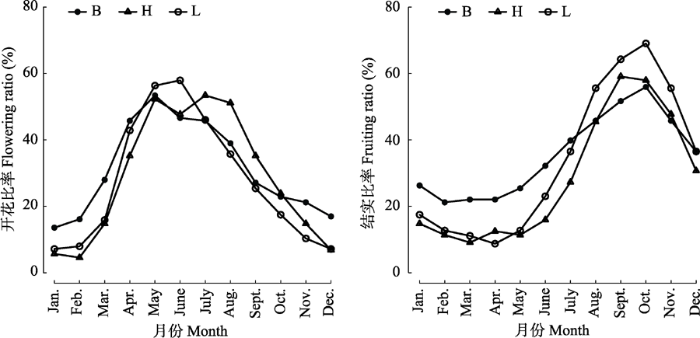 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT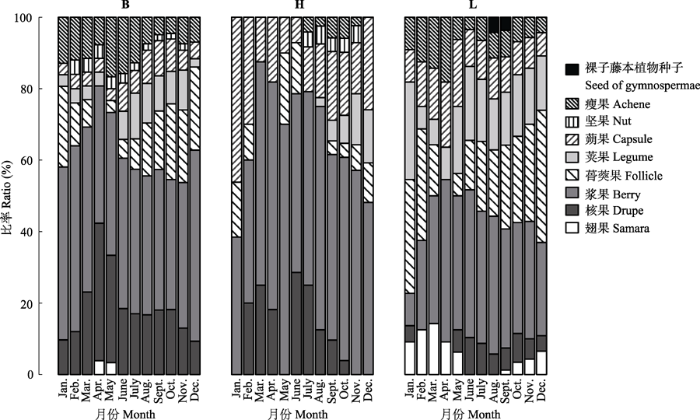 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}