Effects of leachate from understory medicinal plants on litter decomposition and soil enzyme activities of Betula albo-sinensis and Eucommia ulmoides
LUYu-Peng1, XUJi-Yuan2, ZHANGXiao-Xi2, WANGBo-Ya3, XIEBo3, LIUZeng-Wen3,4,*, 1College of Forestry, Northwest A & F University, Yangling, Shaanxi 712100, China;2Institute of Soil and Water Conservation, Northwest A & F University, Yangling, Shaanxi 712100, China;3College of Natural Resources and Environment , Northwest A & F University, Yangling, Shaanxi 712100, China;and 4Key Laboratory for Agricultural Resources and Environmental Remediation in Loess Plateau of Agriculture Ministry of China, Yangling, Shaanxi 712100, China 通讯作者:* 通信作者Author for correspondence (E-mail:zengwenliu2003@aliyun.com) 收稿日期:2017-04-5 接受日期:2016-11-23 网络出版日期:2017-07-28 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:——————————基金项目 国家自然科学基金(30471376)
关键词:林药复合;植物次生代谢物;枯落物分解;养分释放;土壤酶活性 Abstract Aims Forest litter decomposition is an important factor affecting nutrient cycling and ecosystem stability. In a complex system with forest and understory medicinal plants, leachate from the medicinal plants enriched in plant secondary metabolites (PSM) may inhibit litter decomposition and soil enzyme activity of forest trees. Thus, inspection on whether or not this phenomenon exits is one important basis for selecting understory medicinal plants.Methods In this paper, typical forest species Betula albo-sinensis and Eucommia ulmoides and six species of common medicinal plants (Corydalis bungeana, Mentha haplocalyx, Houttuynia cordata, Nepeta cataria, Gynostemma pentaphyllum and Prunella vulgaris) in Qinling Mountains area were taken as objects, and the litter decomposition experiment was carried out. The leachate (water-extraction solution) from the stems and leaves of the medicinal materials were sprayed onto the litter in order to study the effects of leachate from understory plants on forests litter decomposition, nutrient release (carbon, nitrogen and phosphorus) and soil enzyme activity.Important findings For litter of B. albo-sinensis, the decomposition half-life and the turnover period were extended by 76% and 4.3 times, respectively, under H. cordata leachate treatment and the inhibitory effects on the release of carbon and nitrogen were also significant. While under G. pentaphyllum leachate treatment, the half-life of litter decomposition and turnover period were extended by 35% and 2.7 times, respectively, and the inhibitory effects on the release of carbon, nitrogen and phosphorus were all significant. The leachate from these two species of medicinal plants displayed significant inhibitory effects on seven kinds of soil enzymes (invertase, carboxymethyl cellulase, β-glucosidase, dehydrogenase, polyphenol oxidase, protease and phosphatase) activity. For litter of E. ulmoides, the decomposition half-life and the turnover period were extended by 1.7 times and 4.2 times respectively, under H. cordata leachate treatment; while they were extended by 1 times and 9 times respectively, under G. pentaphyllum leachate treatment. The leachate from these two species of medicinal plants displayed significant inhibitory effects on the release of carbon, nitrogen and phosphorus from litter decomposition and the activities of all seven kinds of soil enzymes. Therefore, results suggested that H. cordata and G. pentaphyllum should not be planted under B. albo-sinensis and E. ulmoides forests, or the interplanting density must be low to reduce the inhibitory effects of litter decomposition.
Keywords:compound system of forest-medicinal;plant secondary metabolites;litter decomposition;nutrients release;soil enzyme activity -->0 PDF (1485KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 卢玉鹏, 许纪元, 张晓曦, 王博雅, 谢博, 刘增文. 林下药用植物淋出物对红桦和杜仲枯落物分解及土壤酶活性的影响. 植物生态学报, 2017, 41(6): 639-649 https://doi.org/10.17521/cjpe.2016.0350 LUYu-Peng, XUJi-Yuan, ZHANGXiao-Xi, WANGBo-Ya, XIEBo, LIUZeng-Wen. Effects of leachate from understory medicinal plants on litter decomposition and soil enzyme activities of Betula albo-sinensis and Eucommia ulmoides. Chinese Journal of Plant Ecology, 2017, 41(6): 639-649 https://doi.org/10.17521/cjpe.2016.0350 红桦(Betula albo-sinensis)和杜仲(Eucommia ulmoides)是秦岭山区以天然林或人工林广泛分布的典型森林树种(郭垚鑫等, 2011; 杨明琰等, 2012), 但是, 许多红桦、杜仲纯林生长后期往往存在衰退、土壤退化等问题(陈竹君等, 2006; 林玥等, 2008), 不但生态效益下降, 而且经济收入减少。为改善这种状况, 利用林药共生原理, 在林下种植药用植物, 发展林药复合种植是一种互利共赢的经营模式(刘晓鹰和程颂, 1992)。但是, 目前的相关研究主要集中在林药复合的经济效益、栽培管理技术和植物生长生理等方面(王继永等, 2003; 吴统贵等, 2011), 很少涉及林药种间关系问题。 在林药复合经营中, 一般遵循的原则是“林木为主、以林护药、以药养林”, 而在林药复合生态系统中, 枯落物分解对整个生态系统中养分循环、土壤肥力恢复以及森林生态系统稳定维持有很大影响(Berg, 2000; Koukol et al., 2006)。因此, 在选择林下药用植物时, 林木枯落物分解是否会受到影响是一个必须要考虑的重要因素。枯落物分解是一个有机质分解的过程, 可分为淋溶作用、自然粉碎作用和代谢作用3个阶段(Tian & Takeishi, 2002), 枯落物分解前期主要是通过腐食动物啃食、细菌真菌的定殖对易分解物质(纤维素、半纤维素和果胶等)的分解, 密切关联的土壤酶包括淀粉酶、蔗糖酶和纤维素酶等; 分解后期主要是真菌对顽拗物(木质素等)的分解, 密切关联的土壤酶包括木聚糖酶和多酚氧化酶等(张瑞清等, 2008; 严海元等, 2010)。进一步研究表明, 影响枯落物分解过程中碳(C)释放的土壤酶主要有淀粉酶、蔗糖酶和纤维素酶等水解酶类以及脱氢酶、过氧化物酶和多酚氧化酶等氧化还原酶类, 而影响枯落物氮(N)、磷(P)释放的关键是微生物自身养分需求及周围环境中N、P的含量(胡霞等, 2012), 发挥主要作用的土壤酶是蛋白酶和磷酸酶。 因此, 在特定气候条件下, 枯落物分解主要与其基质质量和土壤性质有关(Chomel et al., 2016), 而药用植物淋出物所含有的次生代谢物(PSM)可能会对土壤生物和酶活性产生影响, 进而影响林木枯落物分解。有研究表明, PSM中的某些生物碱类可以通过影响土壤动物和微生物活动, 降低枯落物分解速率(Purahong & Hyde, 2011); 单宁可以与蛋白质或金属离子形成不溶性的聚合物, 抑制有机质的进一步分解, 或对土壤生物产生毒害从而影响其取食枯落物碎屑, 并对土壤酶活性也有抑制作用(Hättenschwiler & Vitousek, 2000); 酚类物质可以抑制土壤中真菌的定殖、孢子萌发和菌丝伸长并影响细菌的粘附以及膜蛋白的转运(Mierziak et al., 2014), 进而影响枯落物分解。然而, 目前关于该方面的研究集中在来自植物自身或其枯落物的内源PSM (Chomel et al., 2014)或人工添加某种特定PSM (Hättenschwiler & Vitousek, 2000)对枯落物分解的影响, 而很少关注在自然环境中普遍存在且种类组成更加复杂的外源PSM (Zhang et al., 2016)的影响。 在林药复合生态系统中, 林下药用植物释放出的PSM, 可能会通过影响土壤性质, 改变枯落物分解的外部环境条件, 进而影响枯落物分解与养分释放。为验证该假设, 本文以秦岭红桦和杜仲林以及当地6种常见林下药用植物为对象, 通过以药用植物淋出物(以水浸提法获得)喷浇林木枯落物的室内控制分解试验研究, 分析林下药用植物对红桦和杜仲林木枯落物分解及土壤酶活性的影响, 从而为秦岭山区的林药复合模式(药用植物种类)选择提供科学依据。
1 材料和方法
1.1 样品采集与处理
研究区位于陕西秦岭山区周至县厚畛子林场镇安沟流域境内, 地理位置33.75° N, 107.75° E, 地属秦岭中段北坡, 暖温带湿润气候, 海拔1β500- 2β500 m, 平均气温7.4 ℃, 年降水量900 mm, 该区分布有丰富的天然和人工林植被及大量的药用植物资源, 土壤属山地褐土。于2014年10-11月在研究区采集典型成熟的26年红桦和24年杜仲林当年凋落的完好枯落物, 自然风干后保存备用。于2015年7-8月在研究区采集林下常见药用植物地丁草(Corydalis bungeana)、薄荷(Mentha haplocalyx)、蕺菜(又名鱼腥草)(Houttuynia cordata)、荆芥(Nepeta cataria)、绞股蓝(Gynostemma pentaphyllum)和夏枯草(Prunella vulgaris)的地上茎叶样品, 风干后保存。同时采集当地无林荒地的表层(0-10 cm)土壤, 清除根系、石块等杂物, 直接以鲜土过5 mm土壤筛后充分混合, 适当风干(以便于试验过程中控制水分)备用。
Different lowercase letters in same column indicate significant differences (p < 0.05).同列不同小写字母表示显著差异(p < 0.05)。 新窗口打开
2.2 药用植物淋出物对林木枯落物分解速率的影响
根据林木枯落物残留量变化, 建立分解模型并计算出枯落物分解周转期(整体分解)和半衰期(前期分解)(表2)。其中, 对于红桦枯落物, 蕺菜淋出物处理的半衰期和周转期分别延长76%和4.3倍, 绞股蓝淋出物处理的半衰期和周转期分别延长35%和2.7倍, 可见, 二者对红桦枯落物的前期分解和整体分 解均有显著抑制作用; 而地丁草、薄荷、荆芥和夏枯草淋出物处理只对红桦枯落物前期分解有显著抑制作用(半衰期延长16%-36%)。对于杜仲枯落物,蕺菜淋出物处理的半衰期和周转期分别延长1.7倍和4.2倍,绞股蓝淋出物处理的半衰期和周转期分别延长1倍和9倍, 说明二者对杜仲枯落物前期分解和整体分解均有显著抑制作用, 而其他药用植物处理影响不显著。 Table 2 表2 表2不同处理对红桦和杜仲林木枯落物分解速率的影响 Table 2Effects of different treatments on litter decomposition rates of Betula albo-sinensis and Eucommia ulmoides
枯落物 Litters
药用植物 Medicinal plants
枯落物分解模型 Model of litter decomposition
R2
T0.95 (a)
T0.50 (a)
红桦 B. albo-sinensis
地丁草 Corydalis bungeana
R = 0.3194e-7.2672t + 0.6801e-0.2996t
0.965β9
8.712β3bc
1.028β0a
薄荷 Mentha haplocalyx
R = 0.2870e-12.9332t + 0.7200e-0.4001t
0.961β3
6.666β4bc
0.911β4a
蕺菜 Houttuynia cordata
R = 0.4209e-9.7308t + 0.5834e-0.1114t
0.935β1
22.054β3a
1.384β8a
荆芥 Nepeta cataria
R = 0.2370e-11.4407t + 0.7674e-0.3988t
0.952β7
6.848β0bc
1.074β2a
绞股蓝 Gynostemma pentaphyllum
R = 0.4270e-7.2586t + 0.5936e-0.1614t
0.937β9
15.329β5b
1.065β5a
夏枯草 Prunella vulgaris
R = 0.1490e-13.7989t + 0.8500e-0.564t
0.963β3
5.023β4bc
0.940β8a
CK
R = 0.1411e-301.3496t + 0.8589e-0.6864t
0.849β9
4.142β8c
0.788β2b
杜仲 E. ulmoides
地丁草 Corydalis bungeana
R = 0.1973e-558.9289t + 0.8027e-0.8954t
0.911β5
3.100β2c
0.528β7b
薄荷 Mentha haplocalyx
R = 0.1339e-75.016t + 0.8661e-1.2611t
0.960β3
2.261β5c
0.435β6b
蕺菜 Houttuynia cordata
R = 0.3395e-14.7065t + 0.6613e-0.2285t
0.950β1
11.300β6b
1.223β6a
荆芥 Nepeta cataria
R = 0.1409e-14184.4866t + 0.8591e-1.3367t
0.949β7
2.127β5c
0.404β9b
绞股蓝 Gynostemma pentaphyllum
R = 0.4582e-9.5318t + 0.5494e-0.1114t
0.953β8
21.515β3a
0.848β3a
夏枯草 Prunella vulgaris
R = 0.1463e-24.2668t + 0.8542e-1.3354t
0.961β7
2.125β3c
0.401β1b
CK
R = 0.1156e-53.1005t + 0.8844e-1.3324t
0.962β7
2.156β2c
0.428β0b
CK, treatment with distilled water same amount to leachate from medicinal plants; R, residue rate of litter decomposition; t, duration of litter decomposition; R2, coefficient of determination; T0.95, T0.50, turnover period or half-life period of litter decomposition. Different lowercase letters in same column indicate significant differences (p < 0.05).CK, 与药用植物淋出物等量的蒸馏水处理; R, 枯落物分解残余率; t, 分解进行时间; R2, 决定系数; T0.95、T0.50, 分别为枯落物分解周转期和半衰期。同列不同小写字母表示显著差异(p < 0.05)。 新窗口打开
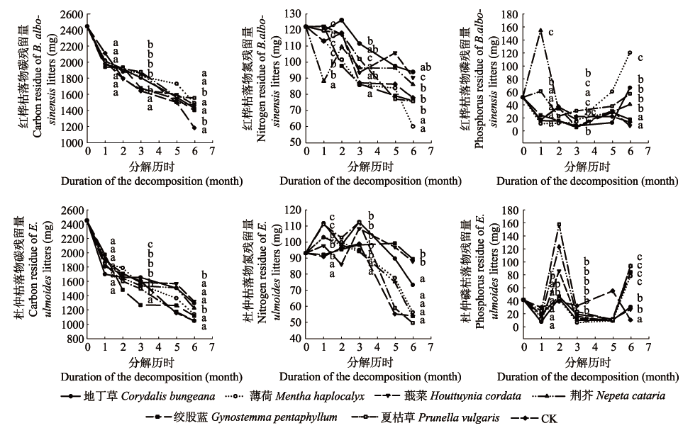
2.3 药用植物淋出物对林木枯落物C、N和P释放的影响
根据红桦和杜仲枯落物分解过程中的C、N、P含量变化测定结果(图1), 2种枯落物分解过程中C表现为持续“释放”, N表现为总体“释放”但存在阶段性“富集”, P表现为“释放-富集”的波动变化。根据枯落物分解“由快到慢”的规律以及土壤酶活性测定时间, 将本试验分为前期(第1个月)、中期(第2-3个月)和后期(第4-6个月)进行分析。 显示原图|下载原图ZIP|生成PPT 图1红桦与杜仲枯落物分解试验前期(第1个月)、中期(第2-3个月)和后期(第4-6个月)不同处理养分释放多重比较。不同小写字母表示表示显著差异(p < 0.05)。 -->Fig. 1Multiple comparisons of nutrient release under different treatments were conducted in the early (first month), middle (second to third months) and late (fourth to sixth months) stages of litter decomposition of Betula albo-sinensis and Eucommia ulmoides. Different lowercase letters indicate significant differences (p < 0.05). -->
根据红桦和杜仲枯落物分解过程中土壤酶活性变化结果(图2, 图3), 一般的规律是蔗糖酶在分 解试验前期和中期活性较高, 羧甲基纤维素酶、β-葡糖苷酶和蛋白酶在中期和后期活性较高, 脱氢酶在中期活性较高, 多酚氧化酶在后期活性较高, 磷酸酶在前期活性较高。 显示原图|下载原图ZIP|生成PPT 图2红桦枯落物分解试验前期(第1个月)、中期(第2-3个月)和后期(第4-6个月)不同处理土壤酶活性多重比较。不同小写字母表示表示显著差异(p < 0.05)。 -->Fig. 2Multiple comparisons of soil enzyme activities under different treatments were conducted in the early (first month), middle (second to third months) and late (fourth to sixth months) stages of litter decomposition of Betula albo-sinensis. Different lowercase letters indicate significant differences (p < 0.05). -->
显示原图|下载原图ZIP|生成PPT 图3杜仲枯落物分解试验前期(第1个月)、中期(第2-3个月)和后期(第4-6个月)不同处理土壤酶活性多重比较。不同小写字母表示表示显著差异(p < 0.05)。 -->Fig. 3Multiple comparisons of soil enzyme activities under different treatments were conducted in the early (first month), middle (second to third months) and late (fourth to sixth months) stages of litter decomposition of Eucommia ulmoides. Different lowercase letters indicate significant differences (p < 0.05). -->
AderiyeBI, OgundanaSK, AdesanyaSA, RobertsMF (1989). The effect of β-sitosterol on spore germination and germ-tube elongation of Aspergillus niger and Botryodiplodia theobromae. International Journal of Food Microbiology, 8, 73-78. [本文引用: 1]
[2]
AggarwalKK, KhanujaSPS, AhmadA, Santha KumarTR, GuptaVK, KumarS (2002). Antimicrobial activity profiles of the two enantiomers of limonene and carvone isolated from the oils of Mentha spicata and Anethum sowa . Flavour and Fragrance Journal, 17, 59-63. [本文引用: 1]
[3]
BatishDR, SinghHP, PandherJK, AroraV, KohliRK (2002). Phytotoxic effect of Parthenium residues on the selected soil properties and growth of chickpea and radish. Weed Biology and Management, 2, 73-78. [本文引用: 1]
[4]
BergB (2000). Litter decomposition and organic matter turnover in northern forest soils. Forest Ecology and Management, 133, 13-22. [本文引用: 2]
[5]
ChenZJ, HeJF, TangDR, LiQX, HengZZ, LiH, LiMS, ChenDW, LuoJM, LiuWY (2002). Nutrients in forest soils of Eucommia ulmoides Oliv and its responses to fertilization in south of Shaanxi Province. Journal of Soil and Water Conservation, 16(4), 94-97. (in Chinese with English abstract)[ 陈竹君, 何景峰, 唐德瑞, 李群学, 衡智州, 李宏, 李孟生, 陈登位, 罗建民, 刘文英 (2006).陕南杜仲林地土壤营养状况及施肥效应 . 水土保持学报, 16(4), 94-97.] [本文引用: 1]
[6]
ChomelM, FernandezC, Bousquet-MélouA, GersC, MonnierY, SantonjaM, GauquelinT, GrosR, LecareuxC, BaldyV (2014). Secondary metabolites of Pinus halepensis alter decomposer organisms and litter decomposition during afforestation of abandoned agricultural zones. Journal of Ecology, 102, 411-424. [本文引用: 1]
[7]
ChomelM, Guittonny-LarchevêqueM, FernandezC, GalletC, DesRochersA, ParéD, JacksonBG, BaldyV (2016). Plant secondary metabolites: A key driver of litter decomposition and soil nutrient cycling. Journal of Ecology, 104, 1527-1541. [本文引用: 2]
[8]
GuoYX, KangB, LiG, WangDX, YangGH, WangDW (2011). Species composition and point pattern analysis of standing trees in secondary Betula albo-sinensis forest in Xiaolongshan of west Qinling Mountains. Chinese Journal of Applied Ecology, 22, 2574-2580. (in Chinese with English abstract)[ 郭垚鑫, 康冰, 李刚, 王德祥, 杨改河, 王大伟 (2011). 小陇山红桦次生林物种组成与立木的点格局分析 . 应用生态学报, 22, 2574-2580.] [本文引用: 1]
[9]
HättenschwilerS, VitousekPM (2000). The role of polyphenols in terrestrial ecosystem nutrient cycling. Trends in Ecology & Evolution, 15, 238-243. [本文引用: 2]
[10]
HuX, WuN, WuY, ZuoWQ, GuoHX, WangJN (2012). Effects of snow cover on the decomposition and nutrient dynamics ofSibiraea angustata leaf litter in western Sichuan plateau, Southwest China. Chinese Journal of Applied Ecology, 23, 1226-1232. (in Chinese with English abstract)[ 胡霞, 吴宁, 吴彦, 左万庆, 郭海霞, 王金牛 (2012). 川西高原季节性雪被覆盖对窄叶鲜卑花凋落物分解和养分动态的影响 . 应用生态学报, 23, 1226-1232.] [本文引用: 1]
LinY, RenJY, YueM (2008). Population structure and spatial analysis of Betula albo-sinensis at Taibai Mountain, Northwestern China. Journal of Plant Ecology (Chinese Version), 32, 1335-1345. (in Chinese with English abstract)[ 林玥, 任坚毅, 岳明(2008).太白山红桦种群结构与空间分析 . 植物生态学报, 32, 1335-1345.] [本文引用: 1]
[13]
LiuXY, ChengS (1992). A preliminary study on the artificial forest-medicinal plant complex ecosystem. Journal of Beijing Forestry University, 14(2), 65-71. (in Chinese with English abstract)[ 刘晓鹰, 程颂 (1992). 林药人工复合生态系统的初步研究 . 北京林业大学学报, 14(2), 65-71.] [本文引用: 1]
[14]
ManzoniS, TrofymowJA, JacksonRB, PorporatoA (2010). Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter. Ecological Monographs, 80, 89-106.
[15]
MierziakJ, KostynK, KulmaA (2014). Flavonoids as important molecules of plant interactions with the environment. Molecules, 19, 16240-16265. [本文引用: 1]
[16]
Mimica-DukicN, BozinB, SokovicM, SiminN (2004). Antimicrobial and antioxidant activities of Melissa officinalis L.(Lamiaceae) essential oil. Journal of Agricultural and Food Chemistry, 52, 2485-2489. [本文引用: 1]
[17]
NiuXY, SunXM, ChenDS, ZhangSG (2015). Soil enzyme activities of the litter in Larix kaempferi plantation. Scientia Silvae Sinicae, 51(4), 16-25. (in Chinese with English abstract)[ 牛小云, 孙晓梅, 陈东升, 张守攻 (2015). 日本落叶松人工林枯落物土壤酶活性 . 林业科学, 51(4), 16-25.] [本文引用: 1]
[18]
ÖzerH, SökmenM, GüllüceM, AdigüzelA, SahinF, SökmenA, KiliçH, BarisÖ (2007). Chemical composition and antimicrobial and antioxidant activities of the essential oil and methanol extract of Hippomarathrum microcarpum (Bieb.) from Turkey. Journal of Agricultural and Food Chemistry, 55, 937-942. [本文引用: 1]
[19]
PurahongW, HydeKD (2011). Effects of fungal endophytes on grass and non-grass litter decomposition rates. Fungal Diversity, 47, 1-7. [本文引用: 1]
[20]
RoyRN, LaskarS, SenSK (2006). Dibutyl phthalate, the bioactive compound produced by Streptomyces albidoflavus 321.2. Microbiological Research, 161, 121-126. [本文引用: 1]
[21]
SellamuthuPS, SivakumarD, SoundyP (2013). Antifungal activity and chemical composition of thyme, peppermint and citronella oils in vapor phase against avocado and peach postharvest pathogens. Journal of Food Safety, 33, 86-93. [本文引用: 1]
[22]
TianXJ, TakeishiT (2002). Relative roles of microorganisms and soil animals on needle litter decomposition in a subalpine coniferous forest. Acta Phytoecologica Sinica, 26, 257-263. [本文引用: 1]
[23]
WangH, WangG, HuangYY, ChenJ, ChenMM (2008). The effects of pH change on the activities of enzymes in an acid soil. Ecology and Environment, 17, 2401-2406. (in Chinese with English abstract)[ 王涵, 王果, 黄颖颖, 陈璟, 陈妹妹 (2008). pH变化对酸性土壤酶活性的影响 . 生态环境, 17, 2401-2406.] [本文引用: 1]
[24]
WangJY, WangWQ, LiuY (2003). Effects of tree and medicinal plant intercropping system on medicinal plants’ yield. Journal of Beijing Forestry University, 25(6), 55-59. (in Chinese with English abstract)[ 王继永, 王文全, 刘勇 (2003). 林药间作系统对药用植物产量的影响 . 北京林业大学学报, 25(6), 55-59.] [本文引用: 1]
[25]
WangZG, ZhaoXS, XuWH, SuYP, YouYM, LiuS, HuY, HuYL, ZhangY (2015). Response of microbial biomass and enzyme activities in black soil to di-n-butyl phthalate contamination. Asian Journal of Ecotoxicology, 10(6), 199-205. (in Chinese with English abstract)[ 王志刚, 赵晓松, 徐伟慧, 苏云鹏, 由义敏, 刘帅, 胡影, 胡云龙, 张颖 (2015). 黑土微生物量和酶活性对邻苯二甲酸二丁酯污染的响应 . 生态毒理学报, 10(6), 199-205.] [本文引用: 1]
[26]
WuCX, LiuSJ, ZhaoGQ (2014). Isolation and identification of the potential allelochemicals in the aqueous extract of yellow sweet clover. Acta Prataculturae Sinica, 23(5), 184-192. (in Chinese with English abstract)[ 邬彩霞, 刘苏娇, 赵国琦 (2014). 黄花草木樨水浸提液中潜在化感物质的分离、鉴定 . 草业学报, 23(5), 184-192.] [本文引用: 1]
[27]
WuTG, YuMK, SunHJ, LiHX, ZhangC, ChengXR (2011).Photosynthetic response to different irradiances of under-growth plants in tree-herb plantation. Chinese Journal of Eco-Agriculture, 19, 338-341. (in Chinese with English abstract)[ 吴统贵, 虞木奎, 孙海菁, 李会欣, 张翠, 成向荣 (2011).林药复合系统林下植物光合特性对生长光强的响应 . 中国生态农业学报, 19, 338-341.] [本文引用: 1]
[28]
YanHY, GuXR, ShenH (2010). Microbial decomposition of forest litter. Chinese Journal of Ecology, 29, 1827-1835. (in Chinese with English abstract)[ 严海元, 辜夕容, 申鸿 (2010). 森林凋落物的微生物分解 . 生态学杂志, 29, 1827-1835.] [本文引用: 4]
[29]
YangMY, TianJ, MaY, SunC, HuangJH (2012).Isolation, identification and antimicrobial activity of endophyte fungi from medicinal plant Eucommia ulmoides Oliv. in Qinling Mountains. Acta Botanica Boreali-Occidentalia Sinica, 32, 193-198. (in Chinese with English abstract)[ 杨明琰, 田稼, 马瑜, 孙超, 黄继红 (2012).杜仲内生真菌的分离鉴定及抗菌活性研究 . 西北植物学报, 32, 193-198.] [本文引用: 1]
[30]
ZhangJ, Salahuddin, JiL, YangLX, WangHR, YouLX (2016). Effects of larch (Larix gmelinii) phenolic acids on manchurian ash(Fraxinus mandshurica) soil microbial community structure. Allelopathy Journal, 37, 123-135. [本文引用: 1]
[31]
ZhangRQ, SunZJ, WangC, YuanTY (2008). Ecological process of leaf litter decomposition in tropical rainforest in Xishuangbanna, SW China. III. Enzyme dynamics. Journal of Plant Ecology (Chinese Version), 32, 622-631. (in Chinese with English abstract)[ 张瑞清, 孙振钧, 王冲, 袁堂玉 (2008). 西双版纳热带雨林凋落叶分解的生态过程. Ⅲ. 酶活性动态 . 植物生态学报, 32, 622-631.] [本文引用: 3]
[32]
ZhouBL, HanL, YinYL, WuJX, SunCQ, YeXL, BaiLP (2010). Effects of allelochemicals hexadecanoic acid on soil microbial composition and biomass in rhizosphere of eggplant. Journal of Shenyang Agricultural University, 41(3), 275-278. (in Chinese with English abstract)[ 周宝利, 韩琳, 尹玉玲, 武建霞, 孙传奇, 叶雪凌, 白丽萍 (2010). 化感物质棕榈酸对茄子根际土壤微生物组成及微生物量的影响 . 沈阳农业大学学报, 41, 275-278.] [本文引用: 1]
The effect of β-sitosterol on spore germination and germ-tube elongation of Aspergillus niger and Botryodiplodia theobromae. 1 1989
Secondary metabolites of Pinus halepensis alter decomposer organisms and litter decomposition during afforestation of abandoned agricultural zones. 1 2014
Chemical composition and antimicrobial and antioxidant activities of the essential oil and methanol extract of Hippomarathrum microcarpum (Bieb.) from Turkey. 1 2007
... 然而, 地丁草、薄荷、荆芥和夏枯草处理对红桦枯落物前期分解有抑制作用, 对后期分解反而有一定的促进作用, 对杜仲枯落物分解则无影响.分析其原因, 地丁草淋出物中含有β-谷甾醇, 薄荷淋出物中含有薄荷醇、香芹酮、石竹素和邻苯二甲酸二丁酯等, 荆芥淋出物中含有薄荷酮、邻苯二甲酸二丁酯、β-谷甾醇和棕榈酸等, 夏枯草淋出物中含有邻苯二甲酸二丁酯和β-谷甾醇等.其中, 薄荷醇可以抑制桃褐腐病菌、匍枝根霉、青霉菌的菌丝生长(Sellamuthu et al., 2013), 而青霉菌是分解枯落物中纤维素、半纤维素和果胶的主要真菌, 褐腐菌是分解枯落物中木质素的主要真菌(严海元等, 2010); 香芹酮可作为抑菌剂(Aggarwal et al., 2002); 薄荷酮具有抗菌和清除自由基的能力(Mimica-Dukic et al., 2004); 石竹素可以抑制细菌、真菌活性(Özer et al., 2007).但是, 这4种药用植物处理对红桦枯落物后期分解反而有促进作用, 这可能与4种药用植物处理下土壤多酚氧化酶活性(分解木质素的主要酶)均未受到影响和分解试验后期土壤蛋白酶活性的提高有关.而杜仲枯落物分解未受到这4种药用植物处理的影响, 其原因可能是: 首先, 杜仲与红桦枯落物基质质量存在差异, 导致其土壤环境中土壤动物和微生物群落组成与活性有差异(Berg, 2000), 所以对来自药用植物同种PSM的响应不同.其次, 枯落物在分解过程中, 自身也会释放出一些PSM, 对土壤微生物及土壤酶活性产生影响(Batish et al., 2002).同时, PSM作用机制中存在“级联效应” (Chomel et al., 2016), 即某种PSM的作用结果可能对其他PSM的作用产生影响, 因此, 在不同的土壤环境中一种或几种PSM作用结果的差异可能由于“级联效应”而导致最终结果不同.此外, 在红桦枯落物分解过程中, 所有药用植物淋出物对土壤β-葡糖苷酶活性均存在抑制作用, 而在杜仲枯落物分解过程中, 只有蕺菜、荆芥和绞股蓝淋出物存在抑制作用, 这可能也是造成药用植物淋出物对红桦与杜仲枯落物分解速率影响存在差异的原因. ...
Effects of fungal endophytes on grass and non-grass litter decomposition rates. 1 2011
Antifungal activity and chemical composition of thyme, peppermint and citronella oils in vapor phase against avocado and peach postharvest pathogens. 1 2013
... 然而, 地丁草、薄荷、荆芥和夏枯草处理对红桦枯落物前期分解有抑制作用, 对后期分解反而有一定的促进作用, 对杜仲枯落物分解则无影响.分析其原因, 地丁草淋出物中含有β-谷甾醇, 薄荷淋出物中含有薄荷醇、香芹酮、石竹素和邻苯二甲酸二丁酯等, 荆芥淋出物中含有薄荷酮、邻苯二甲酸二丁酯、β-谷甾醇和棕榈酸等, 夏枯草淋出物中含有邻苯二甲酸二丁酯和β-谷甾醇等.其中, 薄荷醇可以抑制桃褐腐病菌、匍枝根霉、青霉菌的菌丝生长(Sellamuthu et al., 2013), 而青霉菌是分解枯落物中纤维素、半纤维素和果胶的主要真菌, 褐腐菌是分解枯落物中木质素的主要真菌(严海元等, 2010); 香芹酮可作为抑菌剂(Aggarwal et al., 2002); 薄荷酮具有抗菌和清除自由基的能力(Mimica-Dukic et al., 2004); 石竹素可以抑制细菌、真菌活性(Özer et al., 2007).但是, 这4种药用植物处理对红桦枯落物后期分解反而有促进作用, 这可能与4种药用植物处理下土壤多酚氧化酶活性(分解木质素的主要酶)均未受到影响和分解试验后期土壤蛋白酶活性的提高有关.而杜仲枯落物分解未受到这4种药用植物处理的影响, 其原因可能是: 首先, 杜仲与红桦枯落物基质质量存在差异, 导致其土壤环境中土壤动物和微生物群落组成与活性有差异(Berg, 2000), 所以对来自药用植物同种PSM的响应不同.其次, 枯落物在分解过程中, 自身也会释放出一些PSM, 对土壤微生物及土壤酶活性产生影响(Batish et al., 2002).同时, PSM作用机制中存在“级联效应” (Chomel et al., 2016), 即某种PSM的作用结果可能对其他PSM的作用产生影响, 因此, 在不同的土壤环境中一种或几种PSM作用结果的差异可能由于“级联效应”而导致最终结果不同.此外, 在红桦枯落物分解过程中, 所有药用植物淋出物对土壤β-葡糖苷酶活性均存在抑制作用, 而在杜仲枯落物分解过程中, 只有蕺菜、荆芥和绞股蓝淋出物存在抑制作用, 这可能也是造成药用植物淋出物对红桦与杜仲枯落物分解速率影响存在差异的原因. ...
Relative roles of microorganisms and soil animals on needle litter decomposition in a subalpine coniferous forest. 1 2002

 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT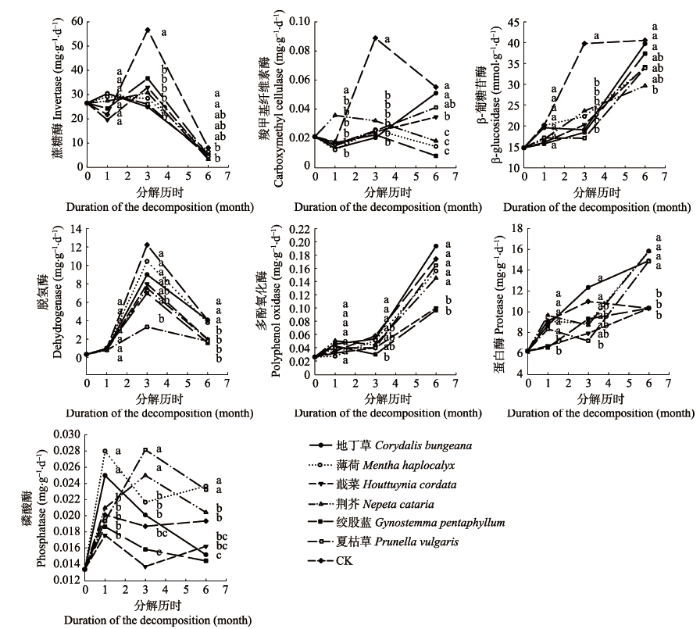 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT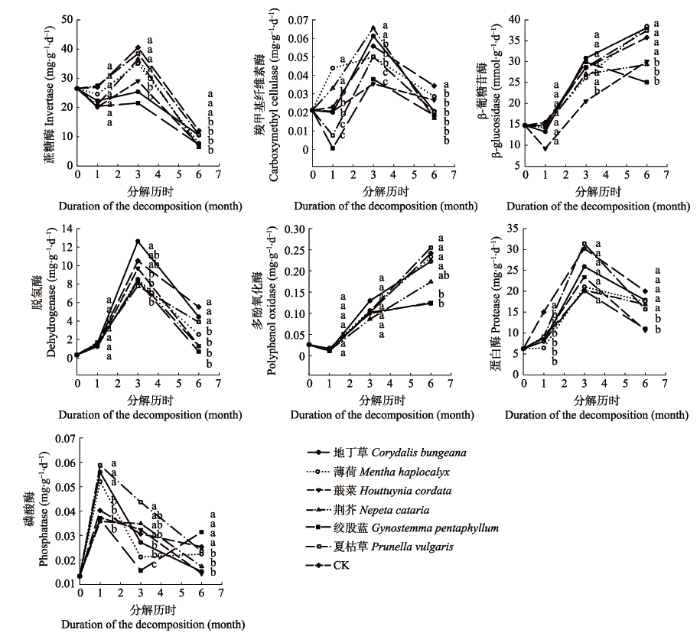 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}